
박시룡 경희대학교 생물학과 졸업 독일 본 대학교 대학원 박사(동물행동학 전공) 현재 한국교원대학교 교수 저서 『 술취한 코끼리가 늘고 있다 』 『 재미있는 동물이야기 』 『 생물학』 (공저) 역서 『 동물의 사회행동 』 『 사고 , 학습 그리고 망각 』 『 사회생물학 1,2 』 (공역) 『 진화의 붉은 실 』 논문 「휘 파람새 노래소리의 방언 」 외 다수
동물행동학의 이해
 동물행동학의 이해
동물행동학의 이해
머리말 과거 60 년 전부터 생물학의 한 분야로 시작된 동물행동학에 대한 관심은 1970 년대 이후부터 이 분야에 관련된 서적 출판이 급증한 데 서 찾아볼 수 있다 . 최근 들어 다른 어떤 생물학 영역보다 동물행동 학과 관련된 전공 서적들이 가장 많이 출판되었다면, 이 분야의 세 계적인 추세를 가히 짐작하게 한다. 그러나 정작 국내의 사정은 너 무 다르다. 우리나라의 대학들은 생물학과에서조차도 강의가 개설되 어 있지 않은 실정이다 . 그리고 이와 관련된 서적이나 전문적 자료 역시 턱없이 부족한 실정에 있다 최근 들어 국내에서도 차츰 이 분 야에 대해 조금씩 관심을 갖기 시작한 것은 그나마 다행한 일로 생 각된다 . 지금까지 국내에서는 몇몇 번역서를 제외하면 제대로 된 동물행동 학 서적이 없는 것이 우리의 현실이다. 무엇보다 이 분야를 처음 공 부하고자 하는 사람들에게는 국내에서 출판된 이 분야의 자료가 절 실히 요구되고 있는 형편이다. 동물행동학 원서들만큼 저자에 따라 다양하고 복잡하게 집필된 분야의 책도 없다. 그렇다 보니 무엇이 중요한 내용인지조차 이 분야를 처음 시작하는 사람들한테는 혼란을 줄 수밖에 없다. 나는 지금까지 10 년 동안 동물행동학을 강의해 오 면서 처음 이 분야를 대하는 사람들이 꼭 알아야 할 내용들을 정리 하여 한 권의 책으로 내게 되었다. 특히 1 장에서는 동물행동학의 접근 방법을 다루는 데 주안점을 두 었다. 그리고 2, 3, 4 장은 과거의 행동연구의 업적을 토대로 행동학
의 기초 지식을 이해할 수 있도록 썼다. 5 장과 6 장에서는 행동생리 에 대해 최신의 분야를 이해하는 데 꼭 필요한 주제가 될 수 있다고 생각되어 따로 장을 마련하였다. 7 장과 8 장은 학습에 관련된 행동영 역으로 행동학에서 모든 환경 조건에 대한 적응 요인과 개체발생 그 리고 개체적응의 과정을 다루었다. 무엇보다 9 장의 사회행동에 많은 지면을 할애했다. 그것은 극소수의 생물을 제의한 대부분의 동물들 여 최소한 짝을 짓기 위해서는 만나야만 하기 때문에 사회행동은 행 동학 분야의 주요한 과제가 아닐 수 없다. 물론 이 분야만 갖고 한 권의 책으로 출판된 것들도 있다. 그러나 여기서는 제한된 지면관계 로 사회행동을 이해하는 데에 필요한 영역들(공격행동, 성적행동, 양 육행동)로 분류하여 제시하였다 . 10 장에서는 행동의 유전을 간략히 다루었고 11 장과 12 장은 이 책의 마지막 내용으로 동물의 가축화를 인위적 적응의 하나로 생각하여 진화의 장에 넣었으며 인간 행동은 비교 행동학의 차원에서 다루었다 . 물론 이 내용들은 각 분야를 완전히 이해하는 데 충분한 것은 못 된댜 다만 각 분야를 보다 깊이 연구하고자 하는 안내서로서의 역 할은 할 수 있으리라 믿는다. 그것을 위해 나는 관련된 내용에 맞추 어 많은 문헌들을 제시하려고 노력했다 . 만일 독자 여러분들이 어떤 분야에 특히 관심을 갖게 되면 그 내용의 문헌을 찾아보는 것이 이 분야를 개척해 나가는 데 주요한 과제가 될 수 있다 . 또 행동학은 같은 종일지라도(비록 어떤 한 종의 행동이 이미 외국에서 취급되었다 하 더라도) 지역에 따라 행동의 차이를 보일 수 있기 때문에 좀더 세심 한 관심이 요구된다. 아마 현명한 독자라면 이 책을 읽으면서 그런 것들을 많이 느낄 수 있으리라 믿는다. 그 밖에도 최근의 학문의 추 세가 모두 첨단을 자랑하면서 고가 장비를 사용하는 것에 비해 행동 학에서는 대개 연구자 스스로 필요에 의해 제작 사용하는 것이 많다 는 점을 감안한다면 이 분야를 그저 이해하는 데서 그쳐서는 안될
것이다 나는 이 책을 통해서 독자들 스스로 이 분야와 관련된 연구를 하고자 하는 동기부여가 될 수 있기를 바란다. 이 책은 생물학도는 물론 축산학, 수의학, 농생물학, 임학 그리고심리학을 공부하는 사람들에게 필요한 내용이 될 수 있다 . 그 동안나는 생물학 이외의 많은 분야에서 동물행동학에 대한 필요성을 강조해 왔지만 제대로 그들에게 추천할 만한 책이 없어 안타까워했다 . 이 책은 야생의 동물들은 물론 가축들을 예로 다루었기 때문에 동물을 대상으로 하는 모든 분야의 사람들에게 많은 도움이 될 수 있을것으로 생각한다 . 한편으로 분야가 한 쪽으로 치우친 감이 없지 않아 있다. 그것은 장을 자료의 양에 따라 나누다 보니 어쩔 수 없었음을 독자들의 이해를 바란다. 끝으로 이 책의 집필을 허락한 대우재단과 출판을 맡아 수고를 아끼지 않은 민음사 여러분께 심심한 감사룰 드린다. 1996.2 저자
차례
머리말 5 제 1 장 행동 연구의 접근 1 행동학의 개념 15 2 행동의 본질 16 3 행동의 연구 18 4 행동학의 분야 22 제 2 장 행동의 기초 1 반사 25 2 역치 변화 29 3 특이적 피로 31 4 갈망행동과 완료행동 34 5 행동의 자발성 37 6 동인 41 7 동인의 분석 43 8 동인의 모델 47 제 3 장 외부자극 1 여과 49 2 해발기구 58 3 신호자극 624 해발인 63
5 해발인과 의사소통 66 6 종간 해발인 69 7 자극의 통합 71 8 초정상자극 73 9 외부자극의 다양성 75 10 외부자극의 종속 76 제 4 장 행동의 순서 1 행동양식의 순서 81 2 갈등행동 83 3 본능 모델 90 제 5 장 행동생리 1 신경계와 행동 95 2 결과 100 3 호르몬과 행동 112 4 호르몬의 효과 범위 114 5 호르몬의 효과 124 6 행동을 조절하는 호르몬 분비 130 제 6 장 행동 리듬과 정위 1 행동 리듬 133 2 정위 140 제 7 장 행동의 발생 1 생득적인가 학습된 것인가? 160 2 행동양식의 성숙 174 3 개체발생학적 기는 변화 178 4 호기심과 놀이행동 182제 1 장 행동 연구의 접근 l 행동학의 개념 동물행동학이란 동물의 행동을 과학적인 방법으로 연구하는 학문 으로 비교행동학 혹은 행동생물학이라고도 부른다 . 동물행동학 e th ol o gy은 그리스어의 eth o s 즉 습성 혹은 행태라는 말에서 유래되었다. 행동은 여러 통합수준에 영향을 미치는 동물의 구성체계에 근거하고 있댜 즉 분자, 생화학적 과정의 수준 혹은 감각기관, 신경계 그리 고 내분비계의 형성 결과에 의해서 동물의 행동은 결정된다. 그러나 이런 행동의 정의를 내린다는 것은 결코 쉬운 일이 아니 댜 행동에 관한 문헌에서조차 정의는 일치하고 있지 않다. 아마 그 것은 취급된 현상이 너무 다양하고 그들의 의미가 비약될 위험소지 가 있기 때문으로 보여진다. 그렇지만 행동은 일반적으로 넓은 의미 에서 자주 사용하고 있다 . 동물들의 이동, 소리내기, 혹은 동작이나 몸짓을 들 수 있으나, 그 밖에도 외부적으로 전혀 알아볼 수 없지만 상호간의 이해에 기여하는 변화(예, 냄새나 체색 변화)까지도 행동에 속한다 그래서 행동이란 우리가 혼히 생각하는 움직임이나 동작에
끝나지 않고, 외견상 부동적일 때도 이런 의미에서 행동이라는 용어 를 사용한다. 한 예로 숫영양이 움직이지 않고 열대의 흰개미가 만 든 언덕위에 서서 어떤 특정 지역을 차지하고 있다든가 암나비가 꽃 에 앉아 냄새를 피워 수컷울 유인하는 것을 들 수있다. 2 행동의 본질 행동은 자극에 대한 반응으로 일어난다. 예를 들면, 배고픔과 같 은 내부 자극이 섭식행동을 유발할 수 있다 . 물론 동물들이 먹이를 찾기 위해 저마다의 독특한 방법을 사용하며, 이것은 각 동물 개체 의 생리적 현상과 밀접한 관계가 있다. 예로부터 인간은 동물의 행동을 관찰하고 설명하는 데 있어서 인 간 자신의 경험에서 얻어진 개념을 사용해 왔다. 예를 들면, 사람이 다가서자 날아가 버리는 곤충을 관찰하면서, 곤충이 사람을 무서워 하여 도망가는 것이라고 해석한다. 지렁이를 바늘로 찔렀을 때 몸을 뒤트는 것을 보고 사람들은 지렁이가 상처를 입고 고통을 참지 못하 기 때문이라고 생각한다. 새가 어린 새끼에게 먹이를 주는 것을 보 고, 어린 새끼를 사랑하기 때문이라고 여겼다 . 그러나 이러한 설명 들은 그 근거가 희박하다 왜냐하면, 위와 같은 상황에서 곤충, 지 렁이 및 새가 공포와 고통과 사랑을 느끼고 있다는 객관적 증거를 관찰할 수 없기 때문이다 . 위와 같은 설명은 인간이 비슷한 상황에 서 경험해오던 것을 다른 동물에 그대로 적용시켜 추측한 것에 지나 지 않는다. 감각 작용은 대뇌 및 신경계의 활동에 의한 결과이며, 곤충과 지 렁이의 뇌는 사람의 뇌와 너무도 다르기 때문에, 이러한 동물의 감 각 작용을 설명하는 데 있어 사람의 감각 작용에 관한 개념을 그대
로 적용시킬 수 없을 것이다. 이와 같은 논리는 정도 차이는 있지만, 새의 행동과 사람의 행동을 비교할 때도 성립한다. 그렇다면 날아가 버리는 곤충과 몸을 뒤트는 지렁이 및 어린 새끼 에게 먹이를 주는 새의 행동을 어떻게 설명할까? 한 가지 사실에 대 해 여러 가지 설명이 가능할 때 가장 간단한 설명을 채택하는 것이 합리적이다. 바꾸어 말하면, 어떤 사실에 관한 설명이 필요 이상으 로 복잡해서는 안 된다. 이와 같은 원리를 염두에 두면, 동물의 행 동을 설명할 때 지나치게 인간 본위적인 해석보다 더 간단한 해석이 가능할 것이다. 곤충은 사람이 다가오는 것을 시각기관으로 받아들 이고, 이 시각적 자극이 반사회로를 통해 날개의 근육을 움직이게 하여 멀리 이동한다. 이러한 설명은 우리의 관찰과 어긋나지 않으 며, 곤충이 공포를 느끼고 도망갈 것을 결정한다고 의식적 행동으로 설명하는 것보다 확실히 더 간단한 설명이다 . 지렁이에 대해서도 마 찬가지이다. 바늘에 의한 자극이 신경계를 통해 전달되어 반사적으 로 몸을 뒤트는 것이며, 지렁이가 고통을 느끼고 있다는 증거는 없 댜 몸을 반사적으로 꿈틀거림으로써 더 이상의 신체손상을 방지하 는데 도움이 되며, 이러한 적응성 행동을 설명하는데 의식이나 의지 와 연관지을 필요가 없다. 어린 새끼에게 먹이를 주는 어미 새의 행 동도 사랑이라는 개념을 개입시키지 않고, 생리적 변화의 자극에 의 한 반응으로써 설명할 수 있다. 그래서 동물행동학의 연구에는 일반적인 논리에서 도출된 최소한 의 가정을 세워야 한다. 그것은 어떤 일에 대해서 단순한 방식과 복 잡한 방식의 두 가지 중에서 하나를 선택해야 한다면, 어떤 행동을 설명할 때 실험 결과를 통해서 단순한 방식의 설명이 부정되지 않는 한, 의식적인 사고보다는 기계적 이유를 가정하는 편이 동물행동학 연구의 접근에 훨씬 더 바람직하다.
3 행동의 연구 • 행동학의 중심 과제는 자연적인 환경에서 방해를 받지 않은 동물 의 적응적 과정으로써의 행동을 이해하는 데 있다. 그래서 행동 연 구는 어떤 행동이 생물학적 의미가 무엇인지롤 아는 것이 중요하다 . 어떤 선택된 행동 양식들이 잘 알려져 있다면 그 기저에 놓여있는 유전적 프로그램이 어떻게 진화되어 왔는지를 이해할 수 있다. 성공 적인 행동 연구에 대한 기본 전제조건은 동물에 관한 광범위한 지식 에 있다. 그 이유는 어떤 동물종의 전체 행동이 잘 알려져 있을 때, 행동의 부분체계가 어떻게 적응되어 왔는지를 정확히 평가되어 분류될 수 있기 때문이다. 이렇게 동물들에 관한 지식을 얻는다는 것은 많은 시간을 들여야 하며, 특히 고도의 사회성을 띤 척추동물 인 경우에는 그들의 복잡한 사회구조가 수년간의 철저한 연구를 통 해서만이 이해될 수 있기 때문에 더욱 많은 시간이 소요된다. 동물 들에 관해 실제 알고 싶어한다면, 어떻게 그들이 자연조건에서 행동 하는지, 즉 그들의 행동이 진화학적으로 적응되어 온 조건들에서 어 떻게 행동하는지를 알아야 한다. 그러므로 동물행동학은 우선 동물 들을 야생 혹은 최소한 자연조건과 유사한 사육조건에서 관찰하여야 한다 이러한 기초위에서만이 계속된 실험실 연구가 의미있다. 이상 적이라면, 야생의 연구와 실험실 연구가 보완적으로 이루어져야만 할것이다. 행동적응의 분석을 위한 기본적인 방법의 하나는 종 비교의 연구 에 있다. 여러 근연 종들의 생태적 지위의 비교를 통해 어떤 특정 종의 특수한 생활조건에 특수한 적응이 무엇인지를 밝히는 것과 여 러 근연 종들의 한 집단내에서 공통적인 행동에 속해 있는 것이 무 엇인가를 알아내는 일이다.
한 동물 종의 전체 행동의 정확한 인식, 자연생활 조건을 위해 일 반화되어 있는 실험실 조건의 의도적 기피 그리고 종들의 비교는 초 기 행동 연구의 중요한 핵심이었다. 1930 년대 초기에 들어서 콘라트 로렌츠 Konrad Lorenz 와 니코 틴버겐 N i ko T i nber gen 은 생명과학분야 의 하나로 동물행동학 e t holo gy울 설 립 하는 데 중추적 인 역 할을 했 댜 그래서 이들은 1973 년에 카를 폰 프리슈 Karl von F ri sch 와 공동 으로 노벨 생리 -의학상을 받게 되었다. 로렌츠는 자기가 취급하는 동물들을 대부분 길러가면서 사람들에게 익숙하게 했다. 그러나 그 동물들을 완전히 자유롭게 돌아다닐 수 있도록 했으며 그 종의 서 식 환경과 매우 가까운 조건에서 살도록 했다. 틴버겐은 사육 조건 을 거의 모두 포기하고 관찰과 실험을 야생으로 옮겨 행했다 . 다른 많은 연구가들(예, Craig , Hein r oth , von Uexkuell, Wh it mann) 과는 달리 로렌츠와 틴버 겐은 행 동생 리 학자인 에 릭 폰 홀스트 Eri ch von Hols t와 함께 최초로 동물행동학의 복합체제를 만들었다. 초창기의 행동학 연구로부터 대략 20 년이 지난 후 틴버겐은 행동학의 최초의 서적인 『본능의 연구 The stu d y of i ns ti nc t』를 출판하게 되었다. 이 책은 그 당 시의 상황에 의거해 분명히 프로그램에 입각한 성격을 띠게 되었다. 행동학을 생물학적으로 연구를 시도한 것은 유럽이었지만 동물의 모든 행동을 다 다룬 것은 아니다. 미국에서는 1920 년대에 이미 동 물 실험을 체계적으로 다루는 심리학 분야가 생겨났다. 원래 행동학 은 여러 동물 종들의 자연스런 행동에 관심을 갖고 있는 학문이라 한다면, 행동주의라 불리는 동물 실험적인 심리학에서는 인간이 관 심의 주체가 되고 있다. 이 동물 실험적 심리학은 오늘날까지 계속 해서 인간의 단순화된 모델로 몇몇 실험동물(예, 쥐, 비둘기, 원숭이) 에 국한하여 연구되고 있다. 이런 연구는 심리학 외에도 의학적으로 관련된 연구 분야에서도 중요한 역할을 해왔다. 동물행동학과 동물 실험적 심리학은 수십년간을 서로 독립적으로 발전해 왔으나, 오늘
날에는 다양하게 상호 영향을 미치고 있다. 이러한 상호 영향은 행 동학이 예전보다 학습 영향에 더 관심을 돌리게 한 반면 학습 실험 에 거의 국한되어 연구해 온 동물실험적 심리학은 동물행동학의 영 향을 받아 행동의 유전적 기초에도 관심을 갖게 되었다. 다른 생물분야와 마찬가지로 행동학은 질적인 관찰, 행동의 명명 그리고 행동의 분류에서 시작하여 양적으로 측정된 행동의 이해로 이어진다 동물의 행동을 관찰을 통해 단순히 서술하는 식의 서술적 행동학은 오늘날 가치가 작은 것으로 평가받기 쉽지만 실제로 그것 은 모든 발전된 행동연구를 위해서 필수적인 전제가 되고 있다. 훌 륭한 동물 관찰자만이 실험적 행동학에서도 성공적인 성과를 거둘 수 있댜 동물의 관찰이 잘 이루어졌을 때만이 실험가설을 세울 수 있고 따라서 실험적 분석에 접근이 가능한 실제 관계를 비로소 인식 하게 된다. 미지의 동물 종에 대한 행동의 연구를 위해 가장 먼저 해야 할 일 은 그 종의 행동목록 e t ho g ram 를 세우는 것이다. 즉 어떤 종의 자연 조건 아래에 일어나는 행동양식을 가능한 한 모두 기술하고 목록을 만드는 일이다. 그것은 행동학에서 꼭 필요한 것으로 예를 들면 행동 의 기능 체계에 따라 다음과 같이 행동목록을 만들어 보는 것이다. ®휴식과잠 ®섭식행동(먹이사냥) ®적(포식자)으로부터의 방어 ®사회행동 이런 큰 분류기능 체계는 좀더 세분화할 수 있다. 예를 들어 사회 행동를 공격행동, 성적 행동, 어미와 새끼 간의 행동으로 나눌 수가 있다 모든 분류와 만찬가지로 기능적 체계에서의 분류 역시 임의로
이루어진다. 행동의 진행과정을 개별적인 행동 요소로 나누는 것은 연구 목적 에 따라 결정된다. 까치의 먹이사냥 행동 범주에서 적응과의 관계를 연구하는 행동생태학자들은 까치가 여러 생활공간에서 하루 얼마나 많은 시간을 보내고 어떤 시간대에 먹이사냥을 하는가를 조사할 수 있다 이때 먹이사냥의 모든 활동들을 요약하는 것으로 충분하다. 그러나 먹이사냥의 각 기술의 효율성을 파악하기 위해 까치는 어떤 개별적 행동양식들을 가지고 지렁이, 달팽이, 산딸기 등을 취하는가 를 매우 상세히 적어야 한다. 무엇보다 전통적인 생물학적 동물행동의 연구는 다음의 네 가지 질문을 통하여 이루어졌다. 첫째, 인과관계 causal 즉 행동의 원인으 로 연구자들은 어떤 행동의 결과를 보고 직접적인 원인을 알려고 했 다. 두번째, 행동의 기능fu nc ti on 으로 이 행동은 무엇을 위한 것인 가? 즉 생물학적 의미가 무엇인가에 관심을 갖게 되었다. 셋째, 개 체발생 on t ogen y으로 발달적 원인을 찾으려 했다. 다시 말하면 행동 의 발생과정을 통해 그 행동의 생성과 변화에 대해 연구를 해왔다. 마지막으로 진화적 원인 evolu ti on 즉 이 행동은 어떻게 진화되어 온 것인가? 하는 물음을 가지고 행동을 연구했다. 근연 종간의 행동 유 형의 비교를 통해 행동의 계통발생학적 관계에 대한 연구는 모두가 이러한 질문에 초점이 맞추어져 있다. 이에 대해 간단한 예로 〈엄지손가락은 왜 다른 손가락들과 다른 방식으로 움직이는가?〉라는 질문을 해보자! 우선 직접적인 원인으로 인과관계의 측면에서 살펴보면, 엄지가 다른 손가락과 골격 구조나 근육 접합에서 차이가 있기 때문으로 보아야 할 것이다. 둘째, 기능 적인 측면에서의 대답은 다른 손가락들과 서로 마주 볼 수 있는 엄 지가 물건을 집어올린다든가 나무를 기어올라가는 것과 같은 행동을 보다 용이하게 해주기 때문이라고 할수 있다. 세번째, 발생학적 측
면에서 손가락이 발생 도중 엄지와 다른 손가락이 다른 구조를 갖도 록 되어 있다는 점에 주목하여 대답할 수 있다. 마지막으로 진화학 적 측면에서 인간은 근연종인 영장류와 같은 동물로부터 진화되어 왔기 때문에 원숭이가 마주보는 엄지손가락을 가지고 있으므로 인간 도 역시 그러하다고 할 수 있을 것이다. 이 네 가지 대답은 모두 옳 울 수도 있지만 그 어느 하나도 완전한 해답은 될 수 없다. 그러나 동물행동학의 연구는 궁극적으로 이러한 의문에 관심을 가질 수 밖 에 없다. 비록 이 네 가지 대답이 서로 독립적이긴 하지만 동물행동 의 연구에 있어서 이런 질문을 도외시하지 않을 때 행동의 적응에 대한 이해가 가능해 진다. 4 행동학의 분야 동물행동학은 실험적으로 연구하는 몇몇 행동학 분야로 나눌 수 있댜 동물행동학에 대한 여러 분야들은 생태학, 생리학, 그리고 유 전학과 같은 생물학의 분야들과 많이 중첩이 되고 있다. 다만 그들 의 방법이나 문제제기를 하는 점에서는 유사하지만 연구 대상이 동 물의 몸구조가 아니고 동물의 행동이라는 점에서 차이가 있다. 물론 동물행동의 연구가 더 행동학적, 더 생태학적 혹은 더 생리학적인가 를 엄밀히 구분할 수가 없다. 이러한 의미에서 행동학 개념에 대한 경계자체가 항상 분명한 것은 아니다. 행동생태학은 비교적 행동학의 최근 분야라고 할 수 있다. 이 분 야는 동물종의 행동과 그들의 생물 및 비생물적 환경 조건과의 관계 롤 연구한다. 이 분야는 동물의 생활공간 혹은 생활공간의 동물개체 군을 다룬다. 생활공간은 특정 서식처에서 예를 들면 사막이나 열대 우림지역에서 근연이 아닌 종들의 경우 관찰될 수 있는 유사한 행동
적응에 관심을 갖고 이때의 행동을 서식처 전형행동으로 나타낸다. 이에 비해 생활주변의 동물개체군에서는 근연종의 개체군으로부터 어떤 식으로 이들 각각의 종들이 서로 차이가 있으며 이런 차이가 다양한 서식처의 적응인지, 혹은 특정 행동의 생물학적 의미는 무엇 인지를 연구하는 것이다. 이러한 근연종들의 개체군 내에서 어떤 한 종들이 완전히 두드러진 생활공간에서 살면서 개체군 전형적인 행동 에서 큰 변이를 보일 때 특히 홍미 있는 결과가 기대될 수 있다. 다 른 행동학 분야에서는 꼭 그렇지 않지만 행동생태학은 어떤 동물종 의 사육 상황에서의 연구와 자연환경에서의 야생 연구가 의미 있는 일치 를 보여 야 한다 (Krebs 와 Davie s 1993) . 행동생태학에서 파생된 중요한 분야로 사회생태학 혹은 사회생물 학이 있다. 사회생물학은 인간을 포함한 종의 환경과 사회구조, 즉 한 종의 개체들의 사회생활(예를 들면 단서성으로, 쌍으로, 혹은 집단으 로써의 생활공간의 분포구성)을 연구하는 학문이다. 여기서 동물의 사 회행동이 중심이 되어 사회행동의 기작 즉 의사소통형태의 도움으로 사회구조를 이루며 유지되는 것들을 다루는 동물사회학과는 완전히 같지는 않다. 행동생리학은 행동의 생리학적 기초를 다루는 학문이다. 이 분야 는 두 개의 행동생리 분야로 구분된다. 이 두 분야는 행동에서 의미 있는 역할을 하는 생물체의 커다란 두 조절계에 집중되어 있다. 즉 신경행동학은 어떤 특정 행동의 기저에 놓여 있는 감각기관과 중추 신경계의 과정에 관심울 갖는다 . 그리고 행동내분비학은 호르몬과 행동사이의 상호관계를 연구하는 분야다. 행동유전학온 행동에 영향을 미치는 유전자들을 밝혀내기 위해서 유전학적 방법으로 행동 양식들의 유전을 연구하는 학문이다. 행동학에는 시간의 변천 과정과 함께 행동의 변화를 다루는 두 분 야가 있다. 행동계통학은 행동의 형질들을 계통발생학적으로 기원과
발달을 연구하는 반면, 행동발생학은 한 개체의 일생 동안의 개체의 행동발달을 연구한다 이 행동발생학은 태아과정에서의 행동발달도 포함된다. 행동학의 비교적 최근의 분야로 인간행동학이 있는데 이것은 행동 학적 방법으로 인간의 행동을 연구하는 것이 기본 목표이며 특히 계 통발생학적으로 내려오는, 또 유전적으로 프로그램화되어 있는 규칙 성에 관심을 갖는다. 그밖에도 순수행동학은 아니지만, 형태학, 생리학 그리고 유전학 과 관련되어 있으면서 행동학에 가까운 생물학적 연구 분야가 있다. 무엇보다 생물음성학과 생물리듬학이 이에 속한다 . 생물음성학은 동 물들의 소리내는 것을 연구의 목표로 삼는다. 이 분야는 녹음방법 및 녹음기의 성능의 발달과 무엇보다 음성분석기의 개발로 동물의 소리를 객관적으로 재생산이 가능하면서 실제로 지난 30 년 전부터 발전하기 시작했다. 청각기관과 발성기관의 연구 및 발성과 소리 지 각에 대한 생리학적 연구나 소리내는 동물의 특징과 그들의 생활공 간의 구조와의 관계 연구도 넓은 의미로 생물음성학에 속한다. 무엇 보다 행동발생학 분야에서 또 음성학적 의사소통(동종간 혹은 이종간 의 소리로써의 상호 이해)과 관련해서 생물음성학온 중요한 지식을 제 공해 주었다 (Busnel 1963, Ehret 1980, Tembrock 1982) . 생물리듬학은 생물의 리듬현상(예, 일주기, 월주기 그리고 연주기적 생체 리듬)과 기저에 놓여 있는 과정을 연구하는 분야다. 이런 주기는 동물의 행동에 있어서 매우 두드러지게 나타날 수 있기 때문에 생물 리듬학과 행동학은 불가분의 관계를 이룬다 (Buen ni n g 1977, Rensin g 1973, Mock 1991).
제 2 장 행동의 기초 1 반사 오랫동안 동물의 행동은 단순히 외부 혹은 내부의 감각자극에 대 한 반응으로만 보아왔다 . 이 반응 행동의 가장 간단한 형태를 반사 re fl ex 라고 한다. 이 반사는 아주 엄밀히 말하면 자극_반응 관계로 표현된다. 죽 같은 조건에서 같은 자극에 대해 반응도 항상 같은 식 으로 이루어진다는 뜻이다 . 이러한 불변성은 반사에 관여한 관련 신경 충격의 전달에 책임을 맡고 있는 신경 요소들간의 고정된 해부학적 관계의 결과로 생겨난 다. 우리는 이것을 반사궁이라고 부른다(그림 2-1) . 반사궁은 수용기 즉 감각세포 혹은 감각기관에서 시작하여 감각신경 혹은 구심성 신 경을 거쳐 중추신경계로 연결되어 다시 중추신경계에서 운동신경 혹 은 원심성 신경을 통해 관련 효과기에 이른다. 가장 간단한 반사궁 온 하나의 구심성 뉴런과 하나의 원심성 뉴런으로 이루어져 있으며 따라서 여기에 하나의 시냅스가 관여한다(단시냅스 반사). 그러나 대 부분의 경우에는 여러 개의 뉴런과 그 뉴런들 사이에 많은 연결 뉴
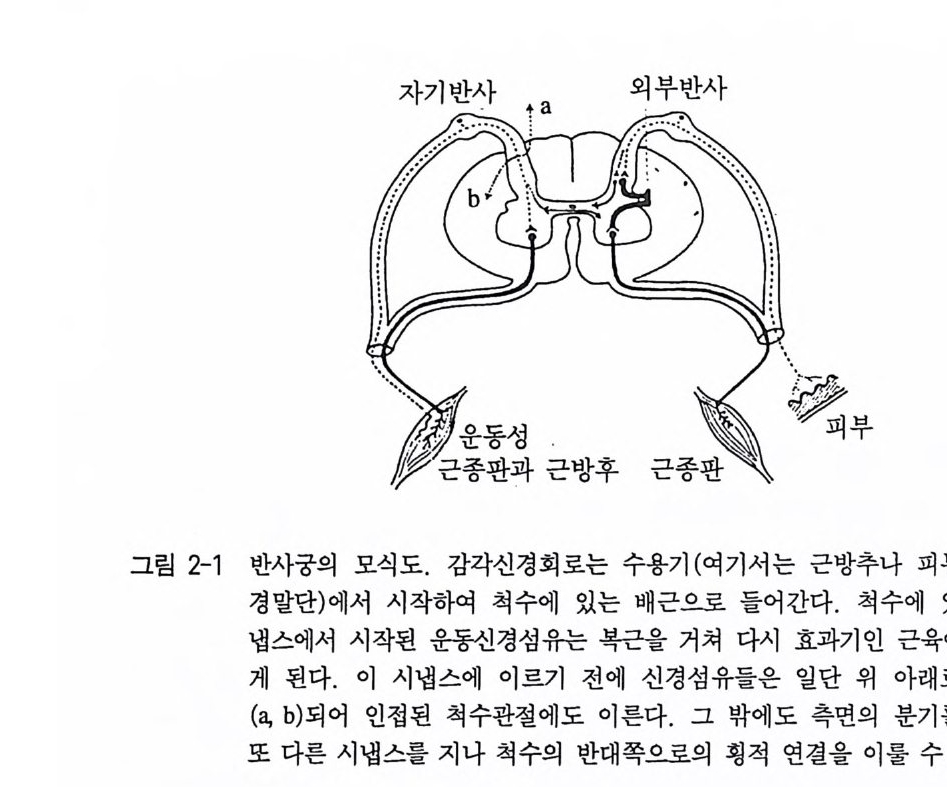 그림 2-1 반사궁의 모식자도 기. 감반]각사동신 성경 회근. 로방는후 수용 기책 (\广여기` 서는· 근呼출방 추나 피부의 신
그림 2-1 반사궁의 모식자도 기. 감반]각사동신 성경 회근. 로방는후 수용 기책 (\广여기` 서는· 근呼출방 추나 피부의 신
런들이 참여하여 여러 개의 시냅스가 생기게 된다(다시넵스 반사). 척 추동물의 경우 많은 반사궁들이 척수만을 지나고 뇌에 이르는 반사 궁은 얼마 안 된다. 그러나 이런 반사궁은 가장 간단하게 형성된 부 위, 소위 능뇌(소뇌, 뇌교, 연수를 가리킴)에 국한되어 남아 있다. 그 래서 반사는 뇌의 최고 중추에 의해 통제받지 않기 때문에 불수의적 으로 작용한다. 이들은 인간의 경우 의식적인 참여 없이도 일어나고 있다. 반사의 반응과정에서 관련 동작의 형태는 항상 생독적이다. 이에 대해 행동 유발을 시키는 자극(해발 자극)과 그에 대한 반응사이의 연결에 관련해서는 커다란 차이가 생겨난다. 일반적으로 이런 연결 도 유전적 요인에 근거하고 있다. 죽 그것은 해당 자극에 대한 과거 의 경험에 의존하고 있지 않다. 우리는 이런 반사의 형태를 무조건
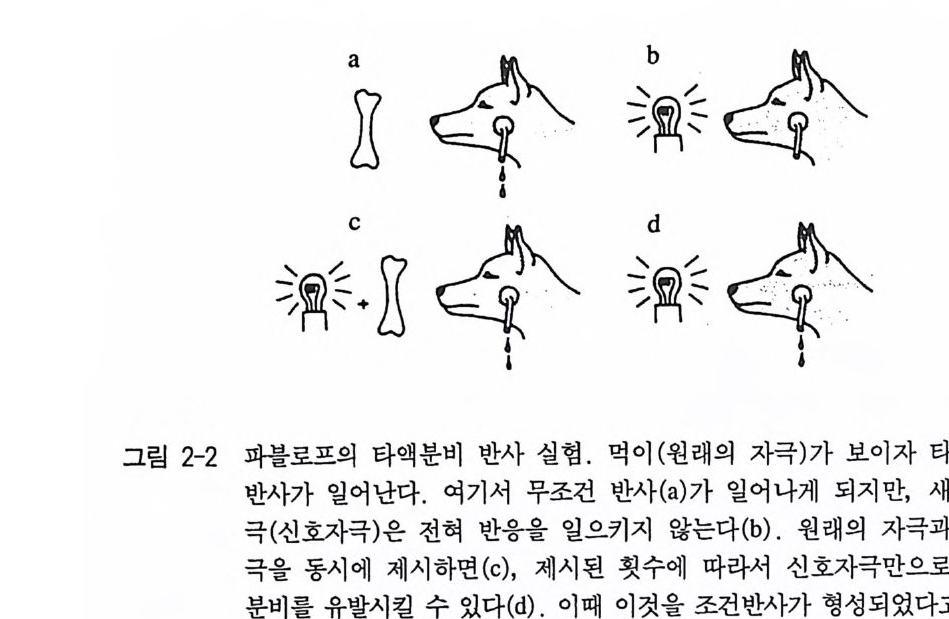
반사 uncond iti oned re fl ex 라고 부른다 . 이것과 구별되는 반사가 있는 데 그런 반사를 조건 반사 condit ion ed re fl ex 라고 한다 . 이 조건 반사 는 해발 자극을 인지하여 자극_반응 관계가 학습과정을 통해서 비 로소 생겨나게 된다. 무조건 반사는 생체가 언제든지 관련 자극에 즉각적으로 또 항상 같은 식으로 반응할 태세가 갖춰져 있어야만 하는 영역에서 일어나 고 있다. 특히 보호 반사(긁기 반사, 닦기 반사, 퇴각 반사, 눈꺼풀 반 사, 동공 반사, 재채기 반사, 기침 반사 , 구토 반사)에서 자주노 나타나 며, 그밖에도 근육의 길이를 유지하는 근육 반사(골지근 반사) 그리 고 몸의 자세와 평형을 유지시키는 신체조절 반사에서 나타난다. 역 사적으로 행동학에서 중요한 역할을 했던 영양학적 영역에서의 반사 는 타액분비 반사가 있다. 대부분의 반사들온 지극히 짧은 지속시간 으로 특징지을 수 있지만 이렇게 길게 지속적으로 반응을 보이는 반 사는 많지 않다. 이런 반사는 타액분비 반사의에도 갓난아이를 포함 한 어린 영장류의 손을 꽉 쥐는 반사(꽉럼 반사)에서 발견할 수 있다 (Prechtl 1 953) . 조건 반사는 러시아의 생리학자 파블로프 (Pawlow 1972) 에 의해 연 구되었다. 그는 개를 대상으로 타액분비 반사의 실험을 했다. 이 반 응은 무조건 반사로, 먹이에서 나온 자극(냄새, 모양)에 의해 반사가 유발되었다. 실험적으로 먹이의 제공을 완전히 다른 신호, 말하자면 불빛 혹은 종소리와 함께 제공해 주면, 얼마간의 실험적인 횟수를 거듭한 후에는 이 타액분비가 원래는 무관심했던 자극만을 통해서 죽 먹이가 없는데도 유발될 수 있다(그림 2-2). 여기서도 자극_반응 관계가 두드러지게 고정적, 말하자면 개가 관련 자극에 틀림없이 반 응한다는 사실은 무조건 반사와 일치하고 있는 셈이다 . 무조건 반사 의 범위 내에서 반응을 처음 일으켰던 외부 자극을 원래 자극이라 하 고, 이런 과제를 2 차적으로 넘겨받은 임의로 선택된 인위적 자극을
 \」戶:
\」戶:
신호자극이라고 한다. 파블로프의 이 실험은 실질적으로 학습과정의 성격에 대해 새로운 인식을 하는 계기가 되었다. 한편 그것은 반사 연구의 급격한 발전 을 가져오게 했으며, 결국 동물의 행동을 오랫동안 단순히 신경계의 반사작용의 결과로 보았던 견해에서 많은 행동양식들이 매우 복잡하 고 반사의 긴 사슬로 이루어졌다는 사실로 인식하게 한 결정적인 계 기가 되었다. 1930 년대 중반에 들어서 홀스트 E ri chvonHols t와 로렌츠 KomadLorenz 는 거의 동시에, 그러나 서로 독립적으로 실험 및 고찰 을 근거로 해서 행동은 단지 자극에 반응할 뿐만 아니라 내부의 자 발적인 요인에 의해서도 유발될 수 있다는 사실을 발표하였다. 그래 서 본능 행동은 개념적으로 반사와 구별할 수 있게 되었으며 그런 자발적인 요소의 존재를 통해서 정의내려지게 됐다 (Hols t 1939, Lorenz 1937). 그후 본능 행동은 반사 사슬에 의해 일어나는 것이 아니라는 사실
들을 제시할 실험들이 계속해서 행해지면서 동물적 행동의 규칙성이 밝혀짐과 동시에 중요한 개념들이 도입될 수 있었다 . 그렇다면 지금 부터 이 본능계의 특성에 대해 알아보기로 하자. 2 역치 변화 간단한 반사와 마찬가지로 복잡한 행동양식들도 외부 자극에 의해 유발된다 여기서 반사와 본능 행동은 근본적인 차이가 생겨난다. 말하자면 반사는 한 자극에 대해 항상 똑 같은 반응이 일어나지만, 본 능 행동은 다양하게 나타날 수 있다. 이런 차이는 역치의 개념을 이용하여 가장 잘 설명할 수 있다. 행 동학에서는 동물의 행동을 유발시킬 수 있는 자극의 최소 크기 혹은 최소의 양을 이해해야만 한다. 이런 최소 크기(양) 이하에 머물러 있 고 따라서 인지할 수 있는 반응을 일으킬 수 없는 자극을 역치하 sub thres hold 자극이라고 부른다 . 반사에서는 이 역치가 언제나 일정하 며, 본능 행동에서는 여러 환경 조건들과 동물의 생리 상태에 따라 심각한 영향을 받을 수 있다. 만일 어떤 행동 양식이 유발되기가 어 렵다 한다면, 우리는 그 행동을 역치상위(역치가 높은 행동)라 할 수 있댜 반대의 개념으로 역치하위(역치가 낮은 행동)라는 말이 종종 사 용된다 . 말하자면, 어떤 행동의 이전 행위 즉 마지막 나타났던 행동과의 시간적 간격은 그 행동의 유발 여부를 결정지울 수 있다. 그 행동이 있은 직후 같은 행동의 유발 정도는 일반적으로 적어진다. 방금 교 미를 끝낸 한 동물은 성적인 자극에 보통 전혀 혹은 경미한 정도의 반응을 일으킬 뿐이다. 또 이와 비슷한 역치상위는 먹이 탐색과 그 밖의 다른 행동영역에서도 나타난다. 반대로 오랜 시간 동안 한 번
도 유발되지 않았던 행동양식은 비교적 쉽게 유발될 수 있고 항상 좀더 간단한 비특정 자극을 통해서도 유발될 수 있다. 이러한 역치 하위에 대한 예들은 이미 많이 알려져 있다. 이런 예들 가운데 가장 잘 알려진 예의 하나는 슬리퍼를 혼드는 개에서 찾아볼 수 있다. 개 과의 육식동물에게 널리 퍼져있는 먹이동물을 혼들어 대는 동작은 그 먹이동물의 목을 부러뜨려 죽이거나 평형유지를 잃게 하고자 하 는 목적으로 행해진다. 그런데 여기서 이런 행동을 해볼 수 있는 기 회를 갖지 못했던 개들은 이 동작들을 비생물적인 물체를 대상으로 하고 있다. 집고양이에게도 공이나 가볍게 움직이는 물체를 주었을 때 먹이잡는 행동을 유발하게 할 수 있다. 자기 동료와 접촉할 기회 가 적은 가축, 동물원의 동물들 그리고 실험실의 동물들은 성적 행 동 영역에 있어서 역치하위가 매우 두드러지게 나타난다. 이 동물들 은 자연상태의 조건하에서 한 번도 본적이 없는 다른 종이나 매우 조잡하게 만든 모형에 대해서도 성적 행동과 교미시도 행위를 보여 준댜 역치가 낮은 조건하에서 자연적인 자극 대신에 한 행동을 유 발시키는 이런 가짜 대상체울 대리 대상체 substit ute obj ec t라고 하고 관련 행동 양식을 대리 대상체 행동 subs titut e ac ti v ity이라고 부른다 (Lorenz 1937) . 극단적인 경우 역치하위는 한 행동이 자발적으로 일어나게 할 수 있다. 죽 외부자극 없이도 행동이 완전히 유발되는 것이다. 이러한 행동을 우리는 진공행동 vacuumac ti v ity이라고 부른다. 이에 대한 연 구는 지금까지 동물의 주변에 있는 어떤 행동 양식이 유발될 수 있 었던 자극들의 여부를 실제로 알아보는 것이 실험적으로 매우 어렵 기 때문에 방법상의 문제가 되고 있다. 그래서 이 진공행동에 대한 실제 뚜렷한 예들은 드문 편이다. 가장 인상적인 것으로 직조새의 매우 복잡한 둥지짓기 행동에서 발견할 수 있다. 직조새는 새장에서 둥지재료로 제공된 지푸라기가 없어도 또 대리 대상체가 없어도 둥
지 짓는 동작을 취 할 수 있다 (Lorenz 1978). 3 특이적 피로 역치의 변화는 행동양식들이 외부 자극에 대해서 일정한 반응뿐만 아니라 각각의 반응 준비에 있어서 차이가 나게 하는 내부적 요인들 이 이 행동양식들의 기저에 있음을 의미한다 . 행동양식의 특이적 피 로현상이 이와 비슷한 상황에서 나타나고 있다. 이것을 행동특이적 피로와 자극특이적 피로로 나누어 살펴보기로 하자 . 3.l 행동특이적 피로 근본적으로 관찰될 수 있는 각 행동들의 유발에 있어서 위에 언급 된 변화는 복잡한 모든 행동양식들에서 같은 정도로 나타나지 않는 다 오히려 많은 행동양식들이 마지막 출현한 직후에 다시 완전히 유발될 수 있는 반면, 일부의 행동들은 시간이 어느 정도 지난 다음 에야 유발이 가능하다. 이 피로가 하나의 전체 특정 행동에 국한하 여 일어날 때 그것울 행동특이적 피로 ac ti on-s p ec ifi c fatig ue 라고 부른 댜 이 차이는 관련 행동양식들에 대한 요구와 관련이 있다. 성적행 동 혹은 섭식행동은-죽 커다란 양의 먹이룰 한번에 먹어야 하는 동물의 경우-그들의 행동 기능을 충족시키기 위해서 단지 간격 을 두고 일어날 필요가 있는 반면, 도피행동이나 방어행동은 계속해 서 유발되어야만 한다. 그러므로 이런 행동들은 행동의 유발에 있어 서 가능한 한 적은 피로가 일어날 때 생물학적으로 의미가 있다 .
3.2 자극특이적 피로 행동 과정의 피로도의 차이는 하나의 동일 행동양식 자체에서 나 타날 수 있다 이 차이는 반응을 유발시키는 자극과 관련이 있다. 어떤 특정 자극을 통해서 여러 번 유발된 다음에 이 자극에 대해 더 이상 반응을 보이지 않는 행동은 다른 자극을 통해서 다시 그 행동 이 유발될 수 있다. 이것은 잠자리 유충의 먹이 잡는 행동에서 아주 분명히 관찰될 수 있다. 이 잠자리 유충의 벌레 참는 채모양의 아랫 입술 lab i um 의 움직임은 정상적으로는 시각 자극(먹이의 크기, 특정 움 직임 등)에 의해 유발된다. 여러번에 걸친 자극의 반복은 잠시 시각 자극에 대한 반응을 사라지게 할 수 있다. 이때 아랫입술에 접촉 자 극을 주면 먹이 잡는 반응은 다시 유발된다. 비슷한 현상이 같은 감각 양상 내에서 예를 들자면 음성학적 영역 내에서 증명할 수 있다 . 짝짓기 소리의 기능을 하는 칠면조의 전형 적인 울음소리는 인공적으로 만든 특정 음을 틀어주어서 유발시킬 수 있다 만일 이 음을 다시 여러 번 제시해 주면 반응이 차츰 떨어 져 결국에는 완전히 사라지게 된다. 그러나 다른 주파수 혹은 다른 세기의 음으로 해서 제공해 주면 곧바로 다시 반응을 불러일으킬 수 있다. 이것은 발성의 일시적 중단이 반응의 반복뿐만 아니라 자극의 반복에 기인된다는 사실을 잘 보여주고 있다. 특정 자극에 대해 반 응도가 이렇게 감소하는 것을 자극익힘, 습관화 혹은 순옹이라고 부 른다 많은 동물행 동학자들은 이 자극특이 적 피 로 stim ulus-spe c if ic fatig ue 를 학습의 가장 간단한 형태의 하나로 보고 있다. 특이적 피로의 두 형태는 우선 반응 정도의 차이가 피로 즉 근육 의 쇠약도 아니고 감각기관의 민감도의 감소에 있지 않다는 것이다. 물론 기관의 관여가 항상 완전히 배제될 수는 없다. 오히려 그것은 중추신경에 자극이 전해지는 과정에서의 신경계의 피로로 보여진다.
여기서 이것을 구심성 전달로 취급이 가능하기 때문에 구심성 억제 라고 부른다. 그러나 감각-운동 영역에서 비슷하게 일어나는 과정 과 이런 현상과의 정확한 구분은 항상 가능한 것은 아니다. Cur i o (1 968) 는 야의실험에서 이 자극익힘 stim ulus hab it ua ti on 이 중 추적인 성질임을 명확히 보여줄 수 있었다 . 그는 둥지를 짓고 있는 알락딱새 Fic e dula h yp oleuca 를 이 용하여 실험 을 하였다. 이 알락딱새 는 무엇보다 올빼미에 대해 경계행동으로 반응하는 많은 작은 조류 가운데 하나다. 이 새가 경계행동을 할 때는 이상한 소리를 내면서 날개와 꼬리깃을 떠는 행동을 보인다. 이 행동은 적합한 모형을 통 해서도 유발될 수 있다. 만일 둥지 구멍 가까이에 있는 이 새에게 박제 올빼미를 보여주면, 강한 경계반응을 보인다. 이에 비해 올빼
 JOO r a . b
JOO r a . b
미와 같게 나무로 만들어 색칠한 모형새는 처음부터 경계반응 즉 혐 오반응을 보이지 않았다. 확실히 이 혐오반응의 유발에서 깃털로 된 몸표면은 중요한 역할을 하고 있다. 이 반응――똑같이 비슷한 반 응행동-은 피로해질 수 있다. 즉 이 박제모형을 반복해서 보여 주면 반응은 차츰 약해서 결국 완전히 사라지게 된다 . 만일 처음에 효과가 없었던 나무모형을 보여준 다음 박제모형을 보여주면, 자극 특이적 피로의 현상과 관련있는 중요한 결과가 발생한다. 그것은 이 제 뒤에 보여준 박제모형에도 혐오반응을 보이지 않는 것이다. 다시 말하면 이 반응능력이 정상적으로는 해발효과가 있는 모형에 대해서 도 사라지게 된것이다(그림 2-3). 이것은 나무모형이 실제로 반응유 발을 시키지 않았으나 그럼에도 불구하고 자극특이적 피로 효과를 지니고 있으며 또 관련 행동양식이 한 번도 나타나지 않을 때, 자극 특이적 피로도 나타날 수 있음을 의미한다. 이로써 자극특이적 피로 가 운동성 피로가 아니라는 사실이 명확해진 셈이다. 4 갈망행동과완료행동 일반적으로 복잡한 행동양식들은 방금 설명한 역치변화와 특이적 피로 현상에 관련하여 서로 성격이 아주 다른 두 가지 단계로 구분 할 수 있다 관련 행동이 끝을 맺는 행위를 완료행동 consumma t o ry ac t이라고 하는데 이것은 보통 비교적 간단하고 짧으며 매우 고형적 인 것이 특징이다. 이 완료행동에 이르게 하는 행동을 갈망행동 app e tit ive behavio r 혹은 탐색 행 동이 라고 하는데 이 행 동은 완료행 동 보다 최소한 길고 다양한 동작과 정위자세로 변하기 쉽다. 이런 차 이는 성적행동과 먹이탐색행동 영역에서 특히 잘 나타나있다. 여기 서 완료행동은 교미행위 내지는 먹이를 잘라서 삼키는 동작을 말한
댜 이에 비해 갈망행동은 포식자를 찾고 낚아채는 긴 연속동작 혹 은 암수 상호간의 동조에 이르게 하는 일련의 동작이 된다. 완료행동은 보통 관련 갈망행동이 오랫동안 더 이상 나타나지 못 하게 한다. 즉 완료행동은 충동을 소진시키는 효과가 있다. 다른 말 로 표현하면, 완료행동은 이미 갈망행동과 완료행동을 유발시켰던 동일 자극에 대해 역치상위를 가짐으로써 일시적으로 같은 행동이 쉽게 유발되지 못하게 한다. 이에 반해 갈망행동의 유발은 다음에 일어나는 행위의 준비에 아무런 영향을 미치지 않는다. 갈망행동은 충동을 소진시키지 않는다. 즉 그것은 행동특이적 피로의 영향을 받 지 않기 때문에 여러번에 걸쳐서 유발될 수 있다 . 갈망행동과 완료행동의 성격상 차이는 여러 기능을 통해서 이해될 수 있다. 한 행동의 궁극적인 목적은 완료행동으로 예를 들면, 한 동물의 먹이요구에 대한 포만과 암컷의 난자를 수정에 이르게 하는 것이다. 이 행동양식은 다음에 생물학적 각 요구를 충족시키는 데 있으며 그래서 이 행동은 각 동물종들의 경우 그리고 각 행동기능 영역에서 아주 긴 시간적 간격을 두고 일어날 수 있다. 갈망행동 영 역에 있어서 행동의 진행과 생물학적 목표 도달 사이의 규칙적인 관 계가 거의 발생하지 않는다. 그 이유는 완료행동까지 이르게 하는 행동양식들의 횟수와 종류가 상황에 따라 매우 다양할 수가 있으며 또 모든 갈망행동 다음에 완료행동이 반드시 오는 것이 아니기 때문 이다(예, 실패한 먹이사냥 행동). 그래서 갈망행동 과정을 통해서 그 갈망행동을 유발시켰던 자극에 대한 조건 반응의 감소는 생물학적으 로 의미가 없을 수도 있다 . 갈망행동은 완료행동을 목표로 삼고자 할 때 목표지향적이 된다. 종종 특정 상황, 예를 들면 번식지 혹은 포란 장소의 도달이 목표가 될 수 있다. 이 경우를 우리는 완료상황이라고 부른다. 공간적인 면 에서도 대부분의 갈망행동들은 완료행동을 유발하게 하는 대상체에
험멕 있기 때문에 종종 목표정위가 성립한다. 물론 가장 간단한 경 우로써 갈망행동은 각 대상체와 만나는 기회를 높여줄 수 있는 이동 활동성이 증가될 때 나타난다. 갈망행동은 아주 복잡하지만 대개 간단한 정위동작(주성요소)만을 다루고 있다. 그러나 일반적으로 갈망행동은 여러 개별적 동작들의 복잡한 연결로 구성되어 있다. 이 경우 시간적으로 매우 길게 나타 날 수 있다. 이러한 예는 철새들의 이동과 많은 회귀성 물고기들의 산란이동을 들 수 있다. 이런 이동들은 번식지나 산란지역의 도착 후에 이 완료상황의 도달과 함께 사라지기 때문에 넓은 의미로 갈망 행동으로 간주할 수 있다. 물론 복잡하고 시간적으로 긴 갈망행동의 경우에 완료행동과 갈망 행동의 구분이 항상 명확한 것은 아니다. 말하자면 완료행동이 일어 나기 전 혹은 완료상황의 도달 전에 행동 연쇄의 일시적 중단으로 완료행동과 비슷한 행동양식이나 상황이 전개될 수 있다. 그러나 여 기서 완료행동은 전체 행동 연쇄의 궁극적인 완성을 의미하지는 않 는다. 이에 대한 예로써 많은 육식동물의 복잡한 사냥행동이 있다. 고양이의 먹이잡는 행동에 대해 레이하우센 Le y hausen(1979) 은 부분 행동들(예, 숨어서 기다리기, 몰래 접근하기, 갑자기 덮치기)이 자빌갓}을 소유하고 있으며, 실제 완료행동처럼 적극적으로 실시되며, 그리고 대리 대상체에 대한 행동으로 진공행동에서 나타날 수 있다는 사실 울 확인한 바 있다. 말하자면 이 부분행동들은 자기 갈망에서 나온 것이다. 비슷한 식으로 암컷과 수컷을 결합시키는 행동양식들은 암 수가 함께 하는 완료상황으로 끝날 수 있지만 이것도 결국 교미로 끝나는 새로운 갈망행동의 영역에 속한다. 이런한 의미에서 동일한 행동이나 동일한 한 가지 행동이 갈망행동뿐만 아니라 완료행동 혹 은 완료상황이 될 수 있다. 완료행동은 종종 생득적 행동의 강한 요소로 표현된다. 갈망행동
에서 이런 생득적 요소가 마찬가지로 나타날 수 있지만 일반적으로 변화된 환경조건을 좀더 잘 이용할 줄 아는 적응적 반응에서 우세하 게 나타난다. 5 행동의 자발성 지금까지 언급된 역치 변화, 특이적 피로 그리고 갈망행동의 현상 들은 간단한 반사와 복잡한 행동양식 사이에서 두 가지 중요한 차이 가 생겨나고 있다. 첫째, 반사는 같은 자극에 같은 방식으로 반응을 하며, 복잡한 행 동양식들은 같은 자극에 대해 전혀 다른 방식으로 반응을 할 수 있다. 둘째, 반사는 수동적으로 일어난다. 즉 동물들은 어떤 반사가 진 행되는 상황을 통제하는 데 직접 관여하지 못한다. 이에 반해 복잡 한 행동양식에서는 완료행동의 진행이 해당 갈망행동에 의해 능동적 으로 바뀌어 질 수 있다. 이런 차이는 복잡한 행동양식의 발생에 직접 자극이 관여할 뿐만 아니라, 동물의 내부에 있는 요인들이 영향을 미칠 수 있기 때문에 나타난다. 내부 요인들이 있다는 사실은 1937 년 로렌츠K. Lorenz 에 의해 확인되었다. 로렌츠는 중추신경계 스스로 본능행동에 특이적 그리고 직접적으로 작동시키는 충격을 발생한다고 생각했다. 내부로 부터, 즉 외부자극의 직접 관여 없이 생기는 행동은 자발적 행동 spo n ta n eous behav i or 이 라고 부른다. 자발적 인 기 초에 만 근거 한 행 동 양식들은 엄밀한 의미에서 모두 진공행동이 된다. 갈망행동도 처음 단계에서는 어쩌면 자발적으로 진행될 수 있다. 그 밖의 복잡한 행동양식들은 서로 다른 부분에서 자발적일 뿐만 아니라 반응적 요소로 이루어졌다. 실제로 어떤 행동이 자발적인지
아닌지 확인은 아주 어렵다. 그 이유는 많은 외부자극들이 특별한 준비없이 관찰자의 눈에 쉽게 띄지 않기 때문이다. 또 다른 이유는 지속적으로 똑같은 환경 조건들도——특정 영향을 일으키는 시간이 지난 후에一一외부자극으로써 영향을 미칠 수 있기 때문이다. 여기 서 서술적 행동연구에 대한 표현 가능성은 한계가 있고, 관련 결과 는 단지 유보될 수밖에 없다. 서술적 행동연구에 반해서 중추자극 상태에 대한 직접적인 증거는 실험적인 방법으로 얻을 수 있다. 로렌츠가 내적 행동충격의 존재에 대한 관찰을 끝내려는 거의 같은 시기에 홀스트는 독립적으로 아주 다른 연구 방향에서 행동요소들의 자발성에 대한 최초의 증거를 제 시 했던 사실은 과학사적으로 홍미 있는 일이었다 (Hols t 1939). 뱀장어의 뱀운동은 홀스트에 의해 연구되었다. 이 당시 널리 퍼졌 던 〈반사학〉에 의하면, 하나의 근섬유의 수축은 그 다음의 고유수용 기를 자극시켜 거기서 수축이 유발된다는 것이다. 이런 견해는 여러 방법에 의한 실험으로 반박되었다. 뱀장어의 중간 부위를 인위적으 로 고정시켜 근수축의 연속을 방해해도 이 뱀운동은 일정한 시간이 지난 뒤에는 몸의 뒤로 연결되었다. 이 부위의 첫번째 근섬유의 수 축은 이전의 근섬유의 수축을 통해서 기계적으로 유발될 수 없는 것 이댜 좀더 직접적으로 척수의 배각, 즉 감각신경 통로을 절단함으 로써 뱀운동은 외부자극과는 무관하게 일어나고 있다는 사실이 증명 되었다. 감각신경을 구심성 신경이라고도 하기 때문에 우리는 이런 외과적 수술을 탈구심화(脫求心化)라고도 부르고 있다. 결국 이런 외 과적 수술은 고유 수용기로부터 오는 모든 정보전달을 완전히 차단 시킨다. 그럼에도 불구하고 탈구심화된 뱀장어에서도 완전한 뱀운동 이 유지되고 있다 . 이것은 이 운동과정이 확실히 중추신경 충격 발 사에 기인하고 있음을 의미한다 . 이 충격 발사는 중추신경계 어딘가 에서 만들어져 감각신경이 완전히 차단되더라도, 다시 말하면 감각
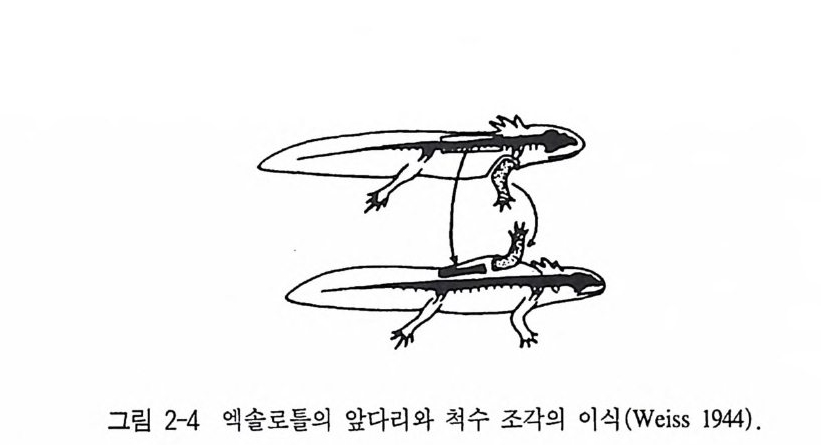 그립 2-4 엑 솔로들의 앞다리 와 척 수 조각의 이 식 (Weis s 1944) .
그립 2-4 엑 솔로들의 앞다리 와 척 수 조각의 이 식 (Weis s 1944) .
세포나 감각기관의 모든 정보전달에 이상이 생겨도 생산될 수 있는 것이다. 그동안 이런 결과를 지지해 주는 일련의 연구들이 있었다. 탈구심 화된 올챙이 역시 종특이적 수영운동을 했고, 금붕어들은 일정한 마 취상태에서도 정상적인 지느러미운동과 호흡운동의 동조성을 보였 다 멕시코산 도롱룡인 엑솔로틀 Amby s t oma mex i canum 의 다리의 배 아세포와 발생과정의 척수의 조각을 떼내 다른 도롱룡 등쪽의 결합 조직에 이식하면, 이식된 척수조각으로부터 차츰 다리의 배아에 있 던 신경섬유가 자라났다(그림 2-4). 이것은 감각성 신경보다 운동성 신경이 더 빨리 명령을 받고 자라나는 것을 의미한다. 이 다리의 배 아에 이미 운동성 신경이 뻗쳐서 감각성 신경의 결여로 인해 감각 자극이 전달될 수는 없는 시기가 있다 . 이 시기에 다리 배아의 최초 의 움직임이 이미 시작하게 된다 . 여기에 있던 충격발사는 그 다음 에 외부자극 없이도 이식된 척수조각에서 발생되어야만 한다 . 이와 비슷한 현상이 행동의 배발생과정에서도 외과적 수술 없이도 일어나 고 있다. 병아리의 배는 이미 포란 3 일째에 최초의 움직임을 보여주 지만, 7 일 혹은 8 일째에는 접촉자극에 반응을 일으키기 시작한다 (Op en heim 1982, Weis s 1941 ) . 그렇다면 중추신경계는 외부 혹은 고유수용기의 감각자극의 도움 없이도 공간적 그리고 시간적으로 완전한 운동을 만들어 내는 능력
 b
b
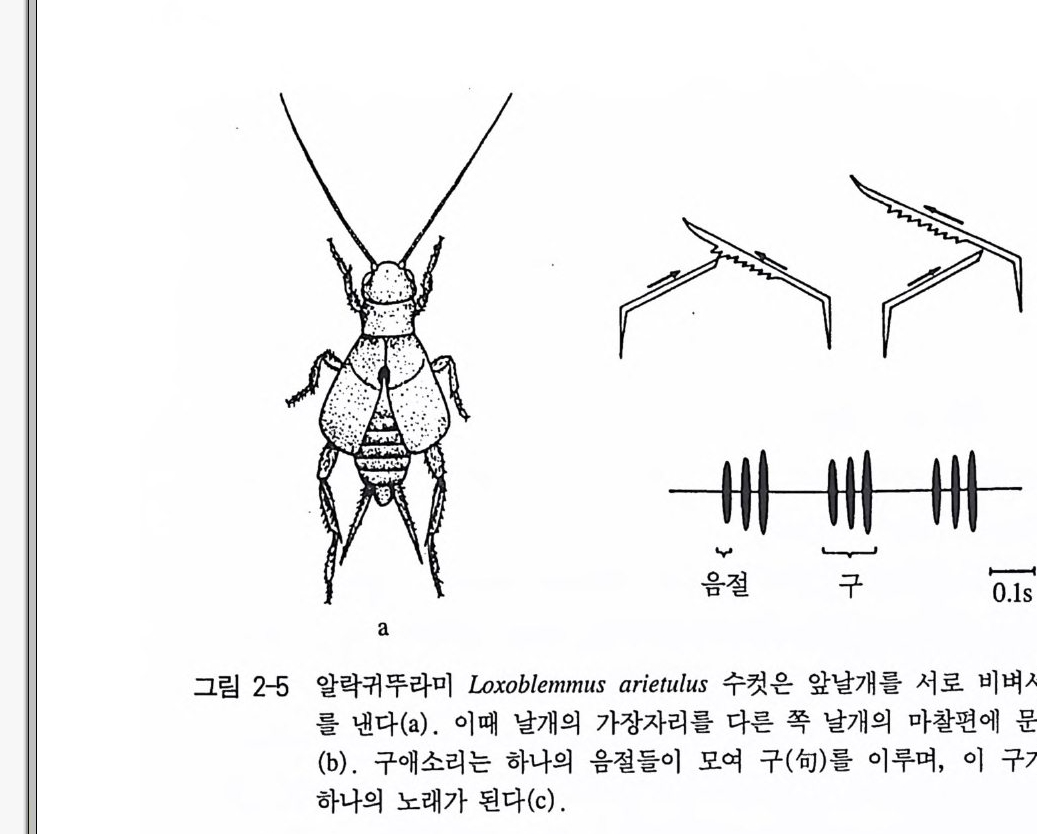
울 갖고 있는 것같이 보인다. 체내의 영양적 영역, 즉 심장 박동이 나 호흡의 자율적 조절의 경우 자율적 -주기적 신경세포군들의 활동 은 이미 오래전부터 알려져 왔다. 위에 언급한 그리고 비슷한 실험 들의 결과가 신체운동들의 연합 영역에서 역시 잘 증명되었다. 이와 같이 행동학에서는 외부의 자극 없이도 일어나는 행동체계를 명 령 체 계 commando s y s t em 라고도 부른다. 귀 뚜라미 의 수컷은 번식 기 에 달하면 종특이적 노래를 부른다. 이들은 한쪽의 앞날개의 가장자 리를 다른 한 날개의 마찰편에 문질러 소리를 낸다. 이렇게 해서 종 특징적 소리를 낼 수 있다(그림 2-5). 이때 암컷은 자기 종의 소리에 게 이끌려오게 된다. 귀뚜라미의 종들의 소리내기는 날개를 리드미 컬하게 움직이는 계통발생에 기인되고 있다. 구애시 이 날개진동수 는 비행 때와 마찬가지로 대략 30Hz 에 달하고 있다. 날개를 마찰시
키는 것이나 나는 것은 신경근육 체계로 일어나고 있다는 점에서 근 본적으로 동일하다. 무엇보다 기본적인 차이는 운동성 충격을 생산 하는 신경성 망상구조인 중추발생기에 있다. 전기적 자극실험을 통 해 숲귀뚜라미 Gr yll us cam p es t r i s 의 노랫소리는 날개진동을 담당하는 가슴신경절뿐만 아니라 뇌에 의해서도 발생될 수 있다 . 뇌와의 연결 지점을 차단해도 이 노랫소리는 가슴신경절에 전기적 자극만을 주어 도 발생하기 때문에, 노래 생산을 조절하는 중추발생기는 날개에 뻗 어 있는 가슴신경절에 놓여 있는 것이 확실하다. 그러나 이 노래발 생기는 정상적인 귀뚜라미의 경우 뇌로부터 오는 충격을 받는다. 이때 뇌는 노래 발생에 관련된 가슴신경절에 소리를 낼 것인가 안 낼 것인가 결정하는 스위치 역할을 하고 있다. 이렇게 중추신경에 위치해 있는 이런 명령체계는 시간적 그리고 공간적 운동성 프로그 램을 구성하는 데에 영향을 미치는 것이 아니고 단지 시동을 거는 작용을 하고 있다. 6 동인 행동에 있어서 자발적 부분들은 사고모델이나 실험에 의해 정확한 설명이 가능할 때, 관찰에 의해 일찍 알아 낼 수 있다. 이것을 우리 는 충동, 열망, 욕구, 혹은 기분이라는 용어를 사용하고 있다. 이런 표현들의 정확한 의미에 대해 또 이런 유사한 용어들의 차이나 정도 에 대해 수많은 논란이 있어 왔다. 그 밖에도 이 용어들은 일상생활 의 대화에서 자주 사용하고 있고 또 이들 용어들간의 관계에 있어서 도 정의가 분명치 않기 때문에 오늘날 행동학에서는 거의 사용하고 있지는 않다 . 그래서 행동학에서는 복잡한 행동양식에 대해 이런 표 현들에 대 한 포괄적 인 의 미 로 동인 (動亂 moti va ti on ) 이 라는 용어 로
자주 사용하고 있다. 그래서 동물행동학에서는 동인을 특정 행동을 취하게 하는 준비로 해석한다. 동인은 모든 행동양식에 있어서 각각 의 시점에 특정값을 가지며 외부와 내부 요인들의 결과가 된다. 이 런 요인들에는 다음의 것들이 해당된다. 내적인 감각자극 내적인 감각자극들은 감각세포(내부 수용기)로 감 지하고, 예를 들면 먹이를 먹거나 물을 마시는 행동영역에서 중요한 역할을 하고 있다. 〈공복〉과 〈갈증〉, 다시 말하면 먹이나 물의 요구 는 원래 혈액속의 포도당의 양(혈당량)이나 염류의 양(삼투압)에 반응 하는 내적인 감각세포의 신호전달에 기인하고 있다. 동인성 신호자극 동인성 신호자극은 다른 자극을 통해 어떤 행동 양식의 유발에 참여할 수 있다. 호르몬 호르몬은 혈액속에 호르몬이 농축되면 여러 행동영역에 서, 무엇보다 성적인 영역에서 행동준비(동인)에 눈에 떨 정도로 영 향을 미칠수 있다. 내적인 리듬 내부 시계는 대부분의 행동양식들이 같은 환경 조건 하에서 진행되는 내적으로 통제되어야만 되는 행동양식들의 빈도나 강도의 편차를 보여주고 있다. 특히 이 내적인 리듬은 24 시간 주기 (이른바 서커디언 리듬, 예를 들면 주행성 동물이나 야행성 동물들의 활동 주기의 분포, 여러 낮 시간대에 명금류들의 노래의 빈도 차이 등)와 연주 기 (예, 1 년중 특정 달에 이동이 한정되어 있거나 번식과 먹이저장 행동양 식이 한정됨)를 말한다. 동물의 성숙 상태 동물의 성숙상태는 어떤 동물이 동일한 자극에
도 나이에 따라 여러 가지 다른 반응을 보일 수 있다 . 이전 행위의 발생 이전 행위의 발생은 방금 일어난 행동전의 시간 적 간격은 규칙적으로 동인의 · 강도와 결부되어 있다(역치 변화 참조). 중추신경계의 자발적 충격발사 충추신경계의 자발적 충격발사는 자 발적 행동 부위와 관련해서 행동의 자발성에서 이미 다루었다 . 위에 언급된 요인들은 제각기 영향을 미치는 것이 아니고 여러 요 인들이 함께 작용하는 것이 보통이다. 예를 들자면, 이전 행위의 발 생 요인은 내부 감각세포로부터 중추신경계로 신호를 보내는데 영향 울 미칠 수 있는 반면, 연중 행동 준비의 변화는 관련 호르몬량에 직 접 적인 결과를 미친다. 7 동인의 분석 동인의 분석은 행동준비를 측정하고 외부 혹은 내부 요인들과 관 련해서 행동준비를 결정해야 하는 과제를 갖고 있다. 어떤 한 동물 의 행동준비는 직접 측정할 수 있는 것이 아니고, 지속적으로 적당 히 유지된 외부 조건하에 행동의 측정도구, 특히 행동진행 과정의 강도, 각각의 동작의 빈도와 지속시간 혹은 잠복기로부터 추론이 가 능하다 . 관찰된 행동양식의 좀더 강한 강도로부터, 말하자면 외부조 건이 동일한데 행동양식의 빈도가 많아졌거나 그 행동의 시간이 길 어졌다면 높은 행동준비, 즉 동인을 갖고 있다고 할 수 있다. 잠복 기는 자극을 가한 시점에서 행동이 시작할 때까지의 시간을 말한다 . 행동준비의 상승은 이런 잠복기의 감소와 관련되어 있다.
이러한 동인 분석은 외견상 가만히 있는 동물일지라도 방금 공격 행동, 성적 행동 혹은 도피행동이 있었는지를 알 수 있다. 물론 이 러한 분석을 위해서는 특정 행동을 지속적으로 반복 관찰해야만 한 댜 그리고 다음과 같은 방법을 가지고 동인분석이 가능하다 (Toa t es 1986, Collga n 1989) . 7. l 수행된 행동 양의 측정 가장 직접적인 방법으로 반응을 일으킬 기회를 주고 얼마나 빈번 히 혹은 얼마나 오랫동안 그 행동을 행하는지 관찰하는 것이다. 마 시는 동인은 마신 양을 측정하고 성적인 동인과 공격적 동인은 좀더 정교한 실험 장치를 통해 측정할 수 있다. 예를 들어, 큰가시고기 한 마리를 시험관 안에 넣고 실험 대상인 큰가시고기가 그 시험관을 향해 무는 수를 세서 공격적 동인을 측정할 수 있다 (Sevens t er 1961 ). 이와 비슷한 방법으로 큰가시고기 암컷을 시험관에 넣어 수컷이 지 그재그춤을 추는 구애동작 수를 세서 성적 동인을 알아볼 수 있다. 그 밖에도 철망바닥에서 모래 목욕을 못 하게 한 암탉울 모래모욕을 할 수 있는 바닥에 옮겨주면 마당에서 기른 암탉보다 목욕하는 시간 이 더 길어진다. 이것은 암탉에게 목욕을 못하게 하면 목욕에 대한 동인이 올라가는 것을 의미한다 (Ves t er g aad 1980). 동인 측정을 통해서 행동양식들의 상호 관계를 알 수 있다. 아프 리 카산인 농어 목 물고기 크리 벤시 스 Pelma t ochrom i s kr i bens i s 는 모든 행동양식들의 행동 준비의 크기나 행동 양식들간의 시간적 관계를 쉽 게 측정 해 볼 수 있다 (He ilig enber g 1963, 1964) . 특히 어 린 이 물고 기는 종종 떼를 지어 다닌다. 그들은 목적지 없이 서로 몸을 가까이 하여 이리 저리 돌아다니며, 각각의 물고기는 자기 이웃과 가능하면 나란히 몸을 유지하려고 한다. 이 무리 짓는 행동의 동인과 공격적

동인은 서로 상반된 관계 를 유지한다 . 이 동인의 크기는 항상 외부 조건을 일정하게 하고 시간 단위 당 관련 행동의 빈도로 측정하였 댜 25 마리의 어린 물고기를 수저에 넣고 30 분 단위로 7 번을 측정하 여 5 분 단위당 무리를 짓는 시간과 공격, 죽 무는 수를 세었다. 결 과 이들은 무리 짓는 빈도가 높을수록 무는 것은 적어지는 것으로 나타났다(그림 2-6). 이것은 무리 짓는 동인과 무는 동인 상호간에 억제역할을 하는 것으로 설명할 수 있다. 7. 2 방해물 설치로 인한 측정 이것은 동물이 행동을 수행하지 못하게 하는 방법으로 이런 방해 에도 불구하고 어느 정도 계속적으로 고집스럽게 행동을 하는지를 관찰함으로써 측정이 가능하다. 예를 들면, 키닌은 사람들에게 매우 쓴 물질로 다른 포유동물들에게도 쓰게 느껴진다. 만일 먹이나 우유 에 이것을 조금씩 넣게 되면 언젠가는 먹이가 너무 써서 먹지 않게 된댜 바로 동물이 참을 수 있는 키닌의 농도는 섭식동인이나 마시 는 동인을 측정하는 기준이 된다. 그래서 동물들은 동인의 크기에 따라 제공된 어려움을 극복하는 정도가 각기 다르다. 그 밖에도 아런 장애물로는 먹이를 먹을 때 전기적 쇼크를 준다거 나 갑자기 바람을 일으키게 할 수 있다. 이런 쇼크의 강도를 변화시 키고 또 먹이에 접근하기 위해 얼마나 그 어려움을 참아내느냐에 따 라 동인을 측정하게 된다. 암쥐는 숫쥐에 도달하기 위해 전기가 흐 르는 철망을 지나가게 한다. 이 암쥐가 멈추게 될 전기 쇼크의 크기 는 에스트로겐 주기에 따라 변화롤 보인다. 그것은 발정기에 가장 높게 나타난다. 이와 비슷한 예로 수탉이 암탉에게 접근하기 위해서 는 추가 달린 문을 열어야만 한다 . 이때 그 문의 무게가 동인 측정 기 구로 사용될 수 있다 (Duncan 과 Kite 1987) .
8 동인의 모델 지금까지 우리는 어떤 행동들은 동인 혹은 내적인 요인에 의해 다 양한 행동양식이 유발되고 있다는 사실을 알게 되었다 . 그렇지만 분 명히 이런 내적 요인만이 행동을 일으키는 원인은 아니다. 동물들은 외부자극에도 반응을 하며 내적 및 외적인 요인들이 함께 작용하여 행동을 유발시키는 과정에 대해서도 이해를 할 필요가 있다 . 이러한 이유로 특히 행동학자들은 행동에 대한 모델을 개발해 왔는데, 그 목적은 실제 동물의 행동을 본따 체계적인 이론을 세우고자 하는데 있다. 적합한 모델의 고안은 동물의 행동에 대한 실험 및 설계를 하는데 좋 은 도움이 된다. 만일 어떤 한 모델이 다양한 조건 아래 동물의 행동을 일관성 있게 설명을 할 수 있다면, 그 동물의 신경계가 작동 하는 원리에 대해 정보를 제공해 줄 수 있다 . 이에 대한 동인의 모 델로 로렌츠(1 937) 의 수력본능모델 psy c h ohy dr auli c model 을 제시할 수 있다. 이 모델에서 로렌츠는 스프링 밸브가 작동하게 한 물탱크 의 경로(물이 완전히 빠져 나간 후 곧바로 물탱크의 물이 채워질 때까지 이 모델은 작동할 수가 없다 . )와 동물행동(먹이룰 먹은 후 곧바로 동인은 더 많은 섭식행동을 유발시키기 어렵다.)을 비교하였다 . 물이 든 이 물탱크를 본능의 중심부라 하고 이것은 동물이 무엇을 하고 싶어하는 열망이라고 하자. 이때 물탱크 위에 위치한 수도꼭지 는 그 본능의 중심부를 자극시켜 주는 내적인 동인성 요인에 해당된 댜 그럼 스프링이 달린 밸브는 행동이 유발되기 위한 준비(행동준 비), 즉 동인이라고 할 수 있다. 이 밸브에 달린 추는 외부자극으로 자극의 크기에 따라 혹은 내적인 동인성 요인, 죽 물탱크에 물이 차 있는 정도에 따라 눈금에 의해 행동의 강도를 알아볼 수 있다(그림 2-7). 본능 중심부에 내적인 요인(물)이 가득 담겨져 있으면 외부자
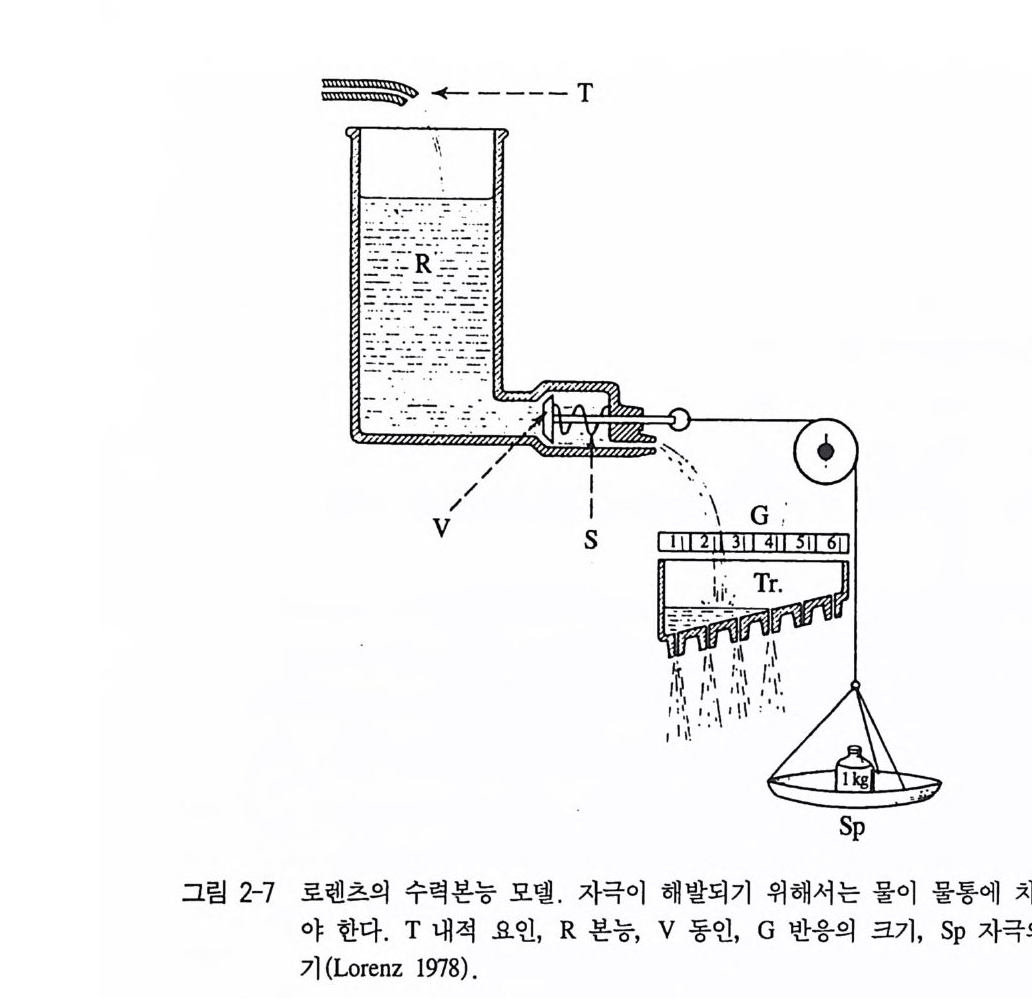 =즉 ~----T
=즉 ~----T
극(추)이 약하더라도 행동은 쉽게 유발(눈금)된다. 물론 이때 동인 (밸브에 작용한 스프링의 힘)은 높다라고 말할 수 있다. 그러나 계속된 자극으로 물탱크 안에 물이 고갈되면, 더 무거운 추를 저울에 올려 놓아야만 물이 나올 수 있다 . 다론 말로 바꾸면 방금 전에 가했던 자극보다 더 큰 자극을 주어야만 행동이 유발된다는 뜻이다 . 극단적 으로 물이 물탱크에 가득차게 되면 추가 없이도, 죽 내적인 동인성 요인들이 너무 많게 되면 자극이 없이도 행동은 유발된다. 이런 행 동을 우리는 이미 진공행동이라고 했다.
제 3 장 의부자극 앞장에서 우리는 반사와 충동행동 간의 차이를 구별하기 위해서 행동의 내적 죽 자발적 영역을 강조한 바 있다 . 충동행동도 다양하 게 의부자극에 의해 영향을 받고 있고(진공행동은 예외) 반사와는 달 리 이 자극에 의해 여러 가지 방식으로 나타나고 있다 . 외부자극은 이런 유발 효과 외에도 움직임의 방향을 결정할 수 있으며(방향성 자 극) 혹은 동물의 행동준비에 영향을 미칠 수 있다(동인성 자극). 1 여과 모든 동물들은 순간마다 환경정보의 홍수 속에 살아가고 있다 . 이 가운데 대부분은 동물들에게 의미가 없고, 단지 일부만이 생물학적 으로 중요한 정보들, 예를 들면 적, 먹이, 혹은 동료에 대한 정보만 을 취한다. 그러므로 동물들이 갖추어야 할 가장 중요한 능력 중의 하나는 반응이 일어나는 자극을 선택하는 데 있다. 이렇게 선택을 위한 여과는 기본적으로 두 기관계인 감각기관과
중추신경계가 맡고 있으며 실제로 이 두 기관계는 자극 선택을 위해 존재하고 있다. 우리는 이것을 각기 말초여과와 중추여과라고 부르 는데, 이런 여과는 각각 감각기관에서 시작해서 중추신경계까지 이 르는 단계로 나누어 취급하려고 한다. 1.1 말초여과 이 말초여과p e rip heral fi l t er i n g는 다음 두 단계로 이루어져 있다. 한 감각기관은 제시된 정보를 전혀 감지하지 못하거나 그것을 감지 한다 하더라도 중추신경계에 전달할 수 없는 것을 말한다. 첫번째 단계는 감각기관의 능력에 의해 결정된다. 그것은 동물의 종마다 매 우 다르다. 박쥐와 많은 나방들은 초음파를 감지할 수 있으며, 벌들 은 자외선을 볼 수 있고, 방울뱀온 적외선에 반응하여 o.oos·c 의 작 은 온도 차이를 느낄 수 있다. 대부분의 포유동물들은 사람보다 훨 씬 더 미세하게 작용하는 후신경을 갖고 있다. 많은 동물들은 인간 을 포함한 대다수의 동물들에게 완전히 차단되어 있는 영역의 주변 자극들에 대해서 반응을 한다 . 많은 곤충들은 복안에 있는 시각세포 의 특이한 미세구조로 편광의 진행 방향을 감지할 수 있다. 이것은 흐린 날에도 태양을 향해 정위를 가능하게 한다. 많은 물고기들은 자기 스스로 생산한 전기자장을 감지할 수 있으며, 꿀벌과 많은 철 새들의 이동에 지구의 자장은 확실히 중요한 역할을 하고 있다. 감지할 수 있는 환경과 행동 실행에 영향을 미칠 수 있는 외부자 극들의 수와 종류는 동물들의 종에 따라 매우 차이가 있을 수 있다. 이런 감각능력에 대한 연구는 감각생리학의 과제가 되고 있다. 감각 생리학은 초기 행동연구에 광범위하게 이용되었다. 그에 대한 고전 적인 예가 프리슈(1 977) 의 꿀벌 훈련 실험을 들 수 있다. 프리슈는 냄새와 시각 능력에 대한 차이룰 알아내기 위해 유리그릇에 냄새물
질과 색깔이 있는 종이판을 놓고 벌을 훈련시켰다. 물론 이런 실험 이 최근의 동물행동 연구에서 전혀 배제되는 것은 아니다 . 어떤 한 종의 상세한 행동연구를 위해서는 필수적인 실험이 되고 있지만, 요 즘은 감각세포에 미세전극을 이용한 전기적 감각생리에도 관심을 갖 고있다. 어떤 한 종의 감각기관의 수용 능력은 그 동물의 생활방식과 결부 된 요구에 적응하는 데 달려 있다 . 이미 언급한 초음파 영역의 청각 능력은 박쥐의 정위음(定位音)을 감지해서 달아날 수 있는 야행성인 나방들에게도 잘 발달되어 있다. 방울뱀의 온도 감각기관은 먹이동 물의 체온 근처에 있는 온도대에 특히 민감하다. 이렇게 감각기관들 의 능력의 한계로 인해 생물학적으로 필요치 않은 환경 정보의 대부 분은 입수되지 않는다 (Burkhard t 1989). 종종 이러한 경로로 종특이적 여과능력이 생겨날 수 있다. 특히 인상적인 예로 먼 거리에서 수컷들을 끌어들일 수 있는 많은 나비류 암컷들의 유인물질이 있다. 감각생리학적 연구에 의하면, 수컷의 더 듬이에 있는 화학적 수용기는 종특이적인 암컷의 유인물질에만 매우 선택적으로 작용하여 화학적으로 매우 근연인 한두 개의 물질을 감 지하는 반면, 다른 물질에는 거의 혹은 전혀 반응을 하지 않는다. 음성학적 영역에서 아열대산 황열모기 Aedesae gypti의 수컷도 종특이 적 여과능력을 갖추고 있다. 음파의 진동을 느끼는 이들 더듬이와 자극을 받아들이는 감각기관인 존스턴 기관Jo hs t on's or g an 은 암컷의 날개 진동수에 가장 민감하게 반응하는 반면, 150Hz 보다 높은 자기 자신의 날개 진동수에는 실제로 거의 영향을 미치지 않도록 만들어 져 있다 이런 식으로 수컷들은 날개 진동소리를 듣고 가까이 지나 가는 암컷들을 인식하여 위치를 알아낼 수 있다. 말초여과에 있어서 암수간의 차이 가 남미 에 서 식 하는 휘 파람개 구리 Eleuth e rodacty lu s co q u i에서도 발견되었다. 이 개구리의 수컷은 두 음절로된 소리를
내는데, lkHz 범위에 놓여 있는 첫번째 음은 다른 수컷이 접근하지 못하게 하는 효과를 갖고 있는 반면, 두번째 음절은 대 략 2kHz 로 암 컷을 끌어들이는 효과를 지니고 있다 . 행동실험에서 확인된 이 두 음절의 기능적 차이는-청신경의 전기생리학적 실험을 통해 확인 된바-내이(內耳)의 민감도에 있어서 암수간의 차이와 일치하고 있다 (N ari ns 과 Capr a nic a 1976). 그 밖에도 말초여과에 대한 예들로 이 미 언급한 밤나방을 들 수 있는데 , 이 밤나방의 청각기관은 사냥을 하는 박쥐의 초음파에 선택적으로 반응을 보이며, 박쥐 스스로도 자 신의 초음파 주파수에 선택적인 민감도를 보이고 있다 (Neuwe i ler 1980) . 지금까지 제시된 예들은 필요하지 않은 동물들의 환경정보는 전혀 취하지 않는다는 사실을 보여주고 있다 . 물론 말초여과의 가능성도 한계가 있다. 무엇보다 이 말초여과는 정보의 요구가 매우 일방적이 고 전문화되어 있는 동물과 행동 영역에 적합하다. 이에 대한 예는 밤나방에서 분명히 알 수 있다. 많은 나방류의 경우에 성체 수컷들 은 번데기에서 나온 후 아주 짧은 시간 동안만을 살면서 더 이상 먹 이를 먹지 않으며 다만 암컷과 교미하는 과제만을 갖고 있다. 그래 서 이 수컷들에게는 생물학적으로 중요한 외부자극인 암컷의 성 유 인물질만이 존재할 뿐이다. 다만 이런 하나의 자극에 대한 후각수용 기의 전문화는 가능하며 생물학적으로 의미가 있다. 비슷한 상황이 다른 전문가 죽, 숙주를 찾는 데 아주 특정한 자극에만 반응할 필요 가 있는 매우 전문화되어 있는 기생동물에서 찾아볼 수 있다. 감각기관들의 능력에 대한 여과는 몇몇 예의의 경우가 있을 수 있 다. 이 경우의 대다수가 감각기관의 능력이 다음 두 가지 이유에서 충분치 못한 데 원인이 있다. 하나는 감각기관들이 적응할 수 있었 던 것보다 생물학적으로 중요한 정보가 종종 너무 복잡한 데 있다 . 많은 동물들은 매우 미세하게 구성된 많은 상세한 자극에 대해서도
반응을 하고 있다. 그래서 이 여과는 감각기관의 능력보다 훨씬 더 뛰어날 것으로 본다. 다른 하나는 같은 수용기가 여러 행동영역에서 보통 매우 다양한 정보들을 취해야만 한다. 마찬가지로 그것은 한 수용기의 감각능력에 대해 한 방향의 여과에 대치되고 있는 것이다. 큰가시고기 수컷은 물벼룩에서 나오는 시각적 자극에는 덥석 무는 행동으로, 녹조류나 다른 식물재료에는 둥지짓기로, 다른 수컷에는 싸움행동으로 그리고 암컷에는 짝짓기로 반응을 보인다. 하나의 행 동영역 내에서조차 종종 비슷한 문제가 발생한다. 예를 들어, 먹이 에서 오는 자극에 대해서는 긍정적인 반응을 보여야 하고, 먹을 수 없는 물질에 오는 자극에 대해서는 부정적인 반응을 보여야만 한다. 그러므로 복잡한 자극들은 복잡한 여과기작을 요구하고 있다. 처 음에는 감각기관에서 여과가 이루어질 수 있고, 그 다음은 일단 감 각기관으로 입수된 모든 정보가 중추신경계에 일부만 전달될 수도 있다 . 이 때 소위 수평 억 제 late r al i nh i b iti on 가 중요한 역 할을 하고 있 댜 한 자극에 의해 일단 흥분되었던 감각세포들은 그 흥분을 동시 에 인접 세포에 보낼 때, 이 홍분된 자극은 수평억제로 인해 약하게 혹은 완전히 억제된다. 이러한 억제는 홍분된 자극의 수렴과 발산에 영향을 미친다. 이것은 감각기관에 위치해 있는 다른 여과기작과 함 께 시간단위 당 정보 양이 감각기관에서 중추신경계까지 여러 번에 걸쳐 감소해 나간다는 사실울 시사하고 있다. 1.2 중추여과 말초여과에서 실행될 수 있는 여러 가능성에도 불구하고 이 말초 여과만으로는 보통 충분치 않은 경우가 발생한다. 그래서 중추신경 계 내에서도 홍분된 자극들의 계속된 선택을 통해 이루어져야만 한 댜 중추여과 centr al filt e ri n g의 기 작과 중추여과가 되 는 정 확한 위 치
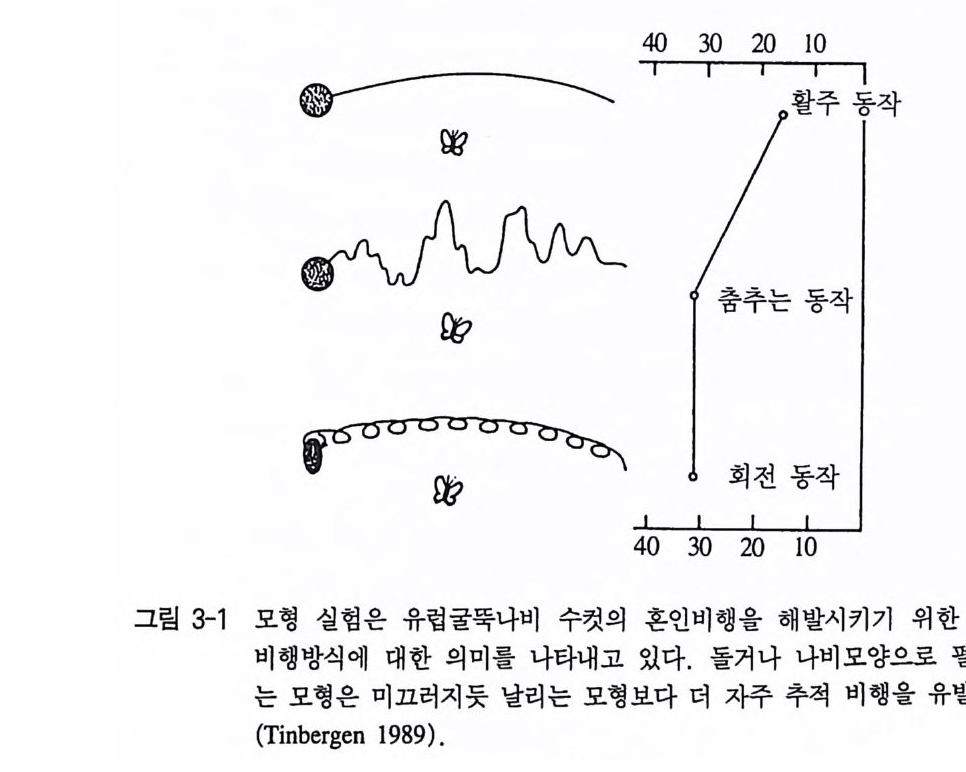
는 아직도 알려져 있지 않다. 이에 대한 연구는 신경생리학에 속한 댜 단지 행동학 분야에서는 이러한 중추적 자극선택이 있는지를 확 인하는 것뿐이다 이것은 모형실험을 실시하면 가장 간단히 나타난 댜 이 실험에서 복잡한 자극 형태의 각 부분들을 나누어 제시하여 필요한 혹은 불필요한 자극구조의 부위를 알 수 있다. 이러한 면에서 매우 인상 깊은 결과는 유럽굴뚝나비 Eumenis semele 에서 연구가 이루어져 있다. 이 나비의 수컷들은 짝짓기 시기 에 공중에 날아가는 암컷들을 추적비행으로 땅에 내려 앉는다. 그런 다음 짧은 구애동작이 있은 후 교미가 이루어진다. 색깔, 모양, 크 기, 명암 그리고 움직임에서 구별되는 모형을 가지고 암컷의 어떤 자극이 수컷의 혼인비행을 유발시키는지를 실험하여 보았다. 실험결 과 색깔, 모양 그리고 크기는 그렇게 중요하지 않았으며 결정적인
 ® 4l0 3l0 210 1l0 |
® 4l0 3l0 210 1l0 |
 10 30
10 30
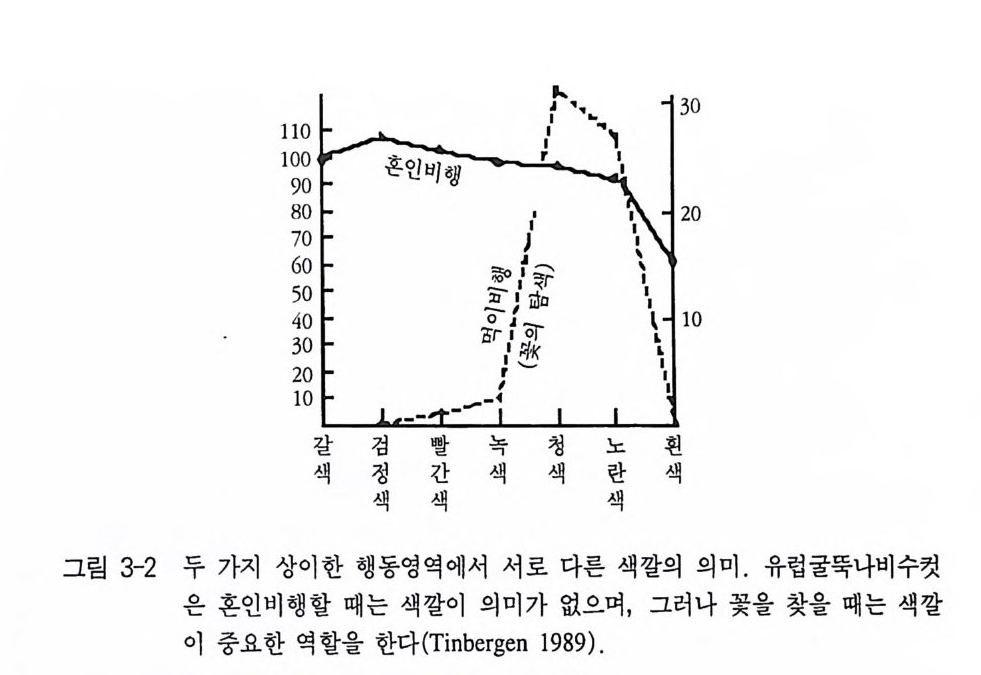
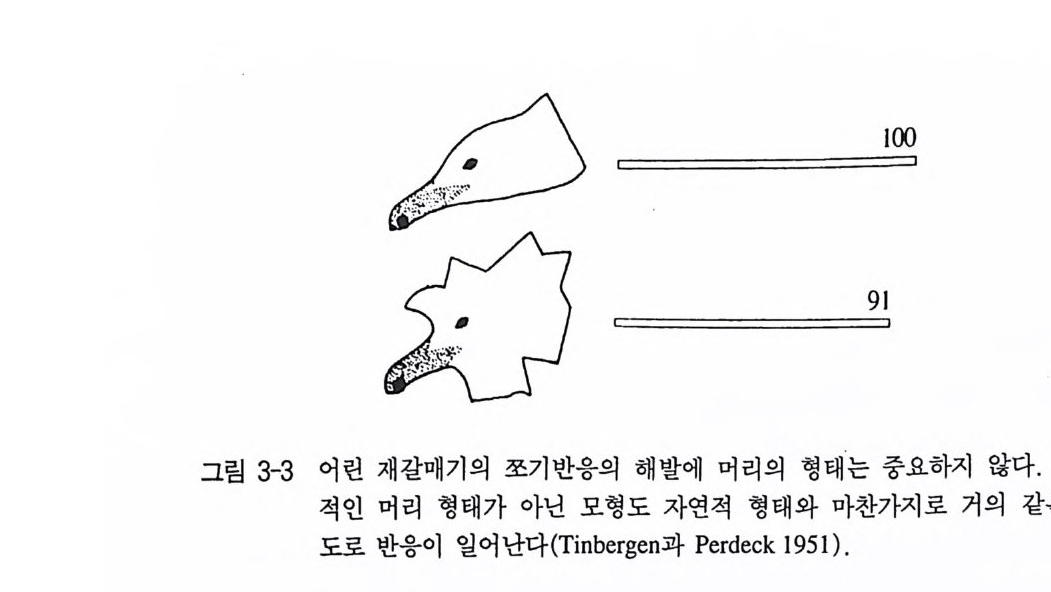
자극은 움직임과 명암이었다. 그래서 가장 최적의 자극은 낚싯줄에 매단 검정 종이모형이 나비처럼 펄럭거리면서 움직였을 때였다(그림 3-1) . 만일 유럽굴뚝나비를 짝짓기 행동영역에서만 조사하면, 이 나 비가 색맹이라는 인상을 받게 된다. 그러나 다른 행동영역 가운데 섭식행동에서는 분명히 파란색과 노란색을 선호한다. 그래서 이 나 비는 색 구별 능력이 매우 뛰어나다는 사실을 알 수 있다 . 바꾸어 말하면, 유럽굴뚝나비는 색맹은 아니나 혼인비행할 때는 마치 색맹 인 것처럼 반응을 보일 뿐이다(그림 3-2, Tin b erge n 1948). 다른 동물종들의 관찰과 실험에 의하면, 이것은 유럽굴뚝나비만이 예외가 아니라는 사실을 알 수 있다. 새끼 재갈매기는 어미의 부리 표시에만 쪼기 반응을 보인다. 그러나 그것은 머리의 모양, 색 혹은 크기가 아니다. 완전히 자연적이지 않은 모형들조차 해당 부리표시 롤 갖고 있기만 하면 쪼기가 유발된다(그림 3-3, T i nber gen 과 Perdeck 1951 ). 번식 경험이 없는 칠면조 암컷은 시각적으로 틀림없이 새끼 를 볼 수 있는 데도 불구하고, 양육행동을 할 때는 새끼의 모습이나
 / 100
/ 100
움직임이 아니고 소리에만 반응을 한다. 그래서 모형에 장치한 스피 커로 새끼소리를 들려주어도 완전히 인위적인 이 모형에 포육반응을 보인다. 반대로 귀를 멀게 한 칠면조 어미는 부화는 시키지만 알에 서 깨어나온 자기 새끼를 죽인다. 비슷한 상황들이 하나의 자극 양상 내에서도 나타날 수 있다. 큰 가시고기 수컷은 공격행동 영역에서 경쟁자의 반짝이는 붉은 복부에 만 반응을 나타내는 반면, 모양, 크기 그리고 표면상태에는 전혀 반 응 하지 않는다. 복부에 회색을 칠한 실제 고기 모양과 비슷한 큰 가시고기 모형은 붉은색을 칠한(물고기 모양을 닮지 않은) 타원형의 나무모형보다 적은 공격행동을 유발시킨다(그림 3-4, Ti nb erge n 1948). 이에 비해 짝짓기시기가 되면 이 수컷은 산란준비가 다된 암 컷의 몸 형태의 특정 표시(불룩한 배)를 인식할 수 있는 상황에 처하 게 된댜 비슷한 방식으로 숫유럽붉은가슴울새 Erith a cus rubecula 는 경쟁자의 전체 모양을 보고 터방어 행동을 취하지 않고, 단지 붉은 가슴 깃털에만 터방어 반응을 보인다 . 그래서 이 수컷의 영역내 나 뭇가지에 고정시킨 붉은 깃털뭉치에는 위협을 하는 반면, 붉은 깃털
 。冬
。冬
을 제거한 붉은가슴울새 박제에는 전혀 반응을 보이지 않는다(그림 3-5, Lack 1943). 쿠바핀치 Tia r is o l i vacea 의 경우 수컷의 색깔 효과는 더욱 크다. 박제 혹은 종이 뭉치를 이용하여 수컷의 노란 목띠를 흉 내만 내도 쿠바핀치 수컷들은 공격을 한다. 몇몇 자극에 대한 이런 제한된 반응을 어떻게 설명할 수 있을까? 아마도 동물들에게는 아주 특정한 자극에만 반응하는 것이 가장 간 단하고 안전할지 모른다. 그 이유는 동물들이 생물학적으로 중요한 대상(적, 먹이, 둥지장소, 은신처)만을 충분히 인식함으로써 필요하지 않은 다른 특징들은 주의를 기울이지 않고 있기 때문이다. 붉은가슴 울새의 터에는 자기 종의 암컷 외에는 가슴에 붉은 색을 가진 다론 새는 존재하지 않는다. 이 터에 붉은 색을 가진 것이 나타났다면, 그 것은 틀림없이 경쟁자가 된다. 큰가시고기 수컷의 붉은 배의 표시도 이 경우와 마찬가지다. 그래서 이 표시는 터주인에게 해당 공격행동
울 맹목적으로 유발시켰을 때 의미가 있다. 이러한 제한은 비슷한 표시 운반자가 한 종의 주변에 적게 나타나면 나타날수록 더 쉽다. 반대로 한 지역에 혼란을 초래할 수있는 종들이 많으면 많을수록 이 제한은 더 어려워진다. 2 해발기구 개구리는 무엇이 파리인가를 어떻게 알까? 이 질문은 다음에 나올 신호자극이 행동을 유발시키는 데 어떻게 작용하는지를 알게 되면 이해가 더욱 쉽다. 이러한 이해는 무엇보다 관련 행동이 일어나기 위해서 여과(선택장치)와 운동중추 사이에 어떻게 연결이 되어 있는 가를 아는 데 있다. 우리는 이 두 장치들간의 능력을 가상적인 장치 로 해 발기 구 releasin g mechan i sm 라고 부른다. 이 것은 특정 반응의 선 택적 해발(자극여과)에 참여하는 모든 구조를 내포하고 있다. 그래서 해발기구를 우리는 신경자극여과 혹은 여과기구라고도 부른다. 생물학적으로 의미 있는 자극 예를 들면, 포획행동을 유발시키는 먹이에서 나오는 자극 혹은 양육행동을 일으키는 새끼에서 나오는 자극들은 해발기구를 통해 동물들의 관련 행동을 가능하게 한다. 유 럽굴뚝나비의 경우 개화된 꽃을 찾는 해발기구는 색깔이 되며, 이에 비해 암컷에게 날아가는 해발기구는 특정한 움직임들과 가능하면 배 경과의 강한 대조다. 해발기구의 위치는 정확히 알 수는 없다. 단지 우리는 중추신경계 뿐만 아니라 감각기관의 어딘가에 있을 것이라고 추측할 뿐이다. 이 들의 여과는 감각기관의 성질에 의해 결정되며, 반면 행동의 해발은 뇌의 운동중추의 어딘가에서 이루어지고 있다. 이 두 제한 내에서 해발기구의 일반적인 성격을 결정하는 구조와 과정이 발견된다. 이
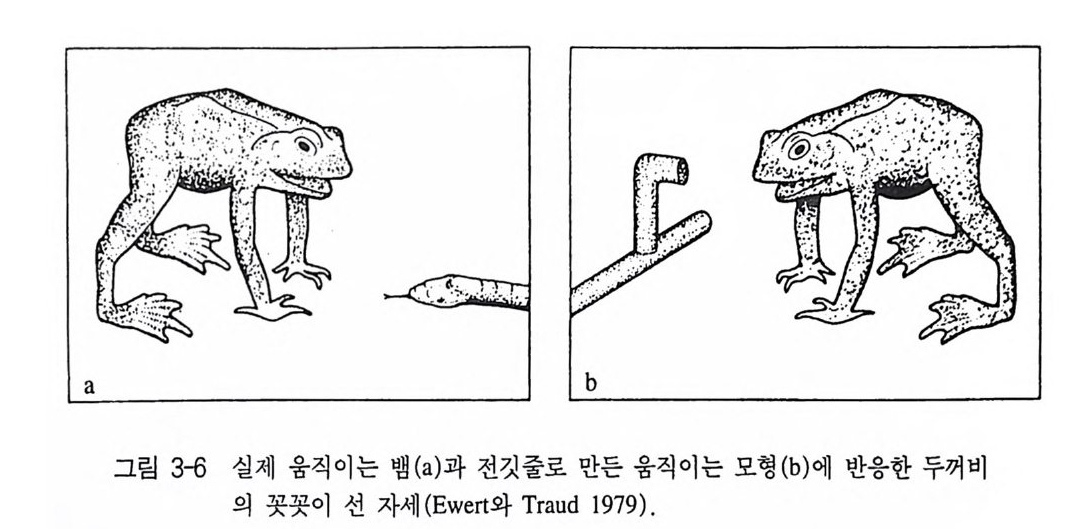 a b
a b
런 모든 기능을 가능하게 만드는 감각기관과 신경계에 있는 연결 스 위치에 관해서 밝혀진 것이 많지는 않다. 그러나 이점에 있어서 눈 의 망막에 있는 시각기의 선택 능력에 관해서 이미 신경생리학적인 연구에 의해 일부 밝혀진 바 있다. 두꺼비는 살아있는 뱀을 만나게 되면 림프주머니를 부풀려 몸을 실제보다 크게 보이려 하고 폿끗이 선 자세를 취한다(그림 3-6a). 살 아있는 뱀 대신에 철사로 뱀 모양을 만들어 실험을 해보아도 마찬가 지의 자세를 취한다(그림 3-6b). 그럼 이 두꺼비가 먹이와 적을 식 별하는 데 사용한 자극(신호자극)은 무엇인가에 대해 좀더 분명히 할 필요가 있다. 실험 결과 두꺼비는 땅바닥을 기는 벌레 모양의 물체 에 몸을 향하는 반응을 보였지만, 수직자세의 물체에는 몸을 다른 쪽으로 돌리는 반응을 보였다(그림 3-7). 이때 먹이와 적에 대한 두 꺼비의 반응은 망막과 더 나아가서 뇌와의 관계에 있음을 알 수 있 댜 실제로 망막에 관련된 모든 신경세포의 활동전위를 측정할 수 있지만, 다양한 시각 자극에 대해 어떻게 반응하는가를 구분하기 위 한 방법으로 신경절세포g an gli on cell 에 전극을 꽂아 활동 전위를 측 정하는 방법이 있다. 지금까지 신경절세포는 여섯 가지 종류로 알려
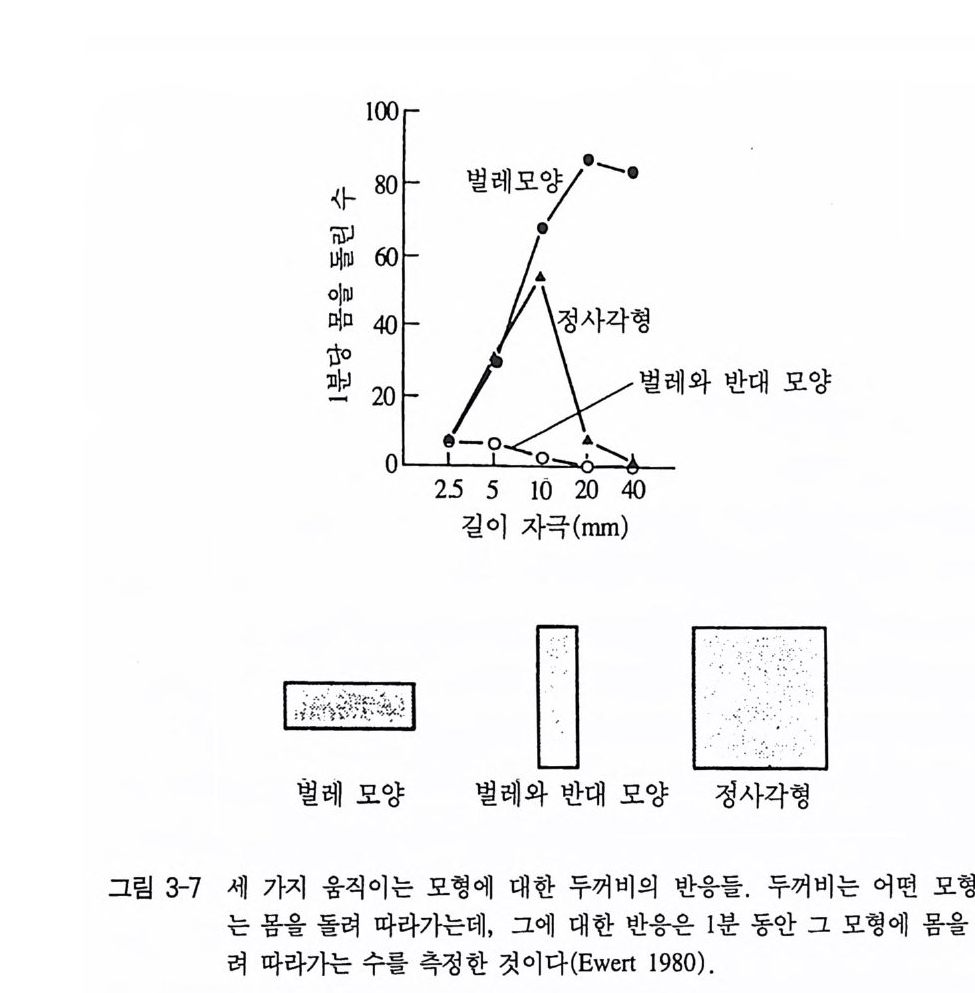 100
100
져 있는데 이 중 네 종류 (Class 1-4) 는 가장 일반적인 신경절세포로 이 두꺼비실험에 이용되었다. 신경절세포 Class 1 과 2 는 동물이 조금 만 움직여도 강하게 반응을 보였다. 이 신경절세포는 작은 물체가 움직일 때 두꺼비가 어떻게 감지할 수 있는가를 설명하는 데에 정밀 한 특성을 지니고 있다. 각각의 신경절세포는 전체 시각 영상의 특 정 부분에만 반응을 보인다. 그러므로 어떤 신경절세포라도 두꺼비 가 물체를 볼 때마다 반응하는 것이 아니고 물체가 망막의 특별한 부위에 도달할 때만 반응을 보인다. 이것을 〈수용역〉이라고 한다. 각 신경절세포의 수용역은 두 부위로 나누어 진다(그림 3-8). 하나
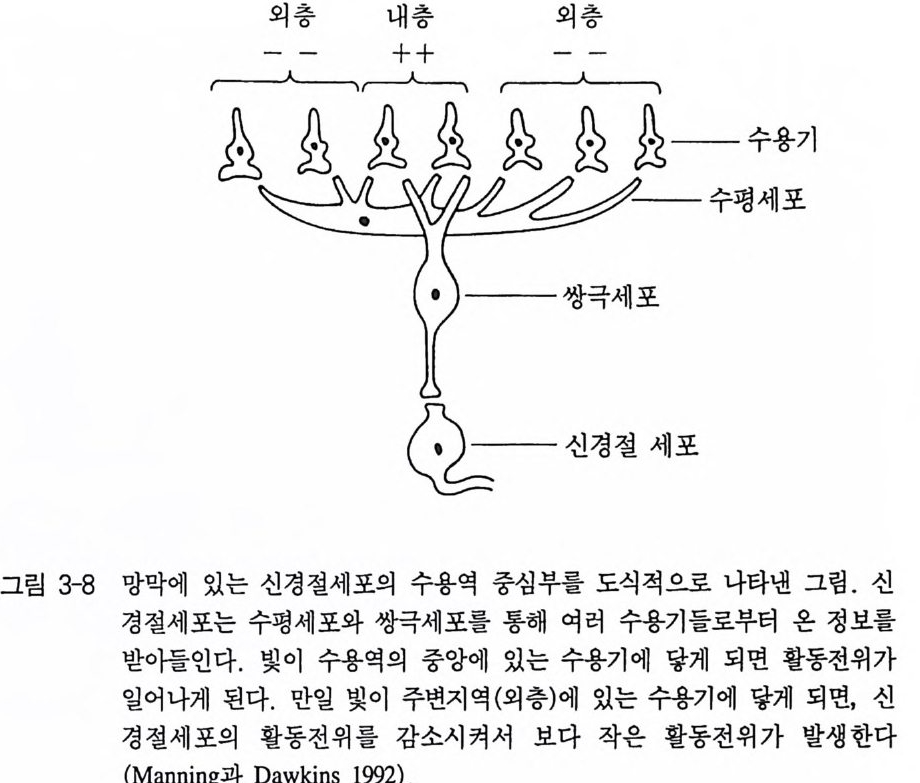 외충 내층 외층
외충 내층 외층
는 내측홍분성중심 다른 하나는 외측억제성둘레라고 부른다 . 이것은 물체가 수용역의 내측 부위에 의해 인식될 때마다 신경절세포는 홍 분되지만, 그 물체가 수용역 외측에 도달하게 되면 이 신경절세포의 활동은 억제된다. 신경절세포의 여러 Class 들은 서로 다른 홍분성 수용역을 갖고 있다. Class 1 과 Class 2 의 신경절세포는 Class 3(6°-8° ) 혹은 Class4 (1 0°-15°) 보다 더 작은 수용역 (20-50) 을 갖 고 있다. 물론 Class 3 과 Class 4 의 신경절세포는 시각이 50 이상이 안 되는 작은 물체에는 전혀 반응을 하지 않는다. 만일 사각형이 두꺼비의 눈 앞에서 움직이게 되면 여러 Class 의 신경절세포에서 충격발사 빈도(활동전위)는 사각형의 크기에 따라 달 라진다 작은 홍분성 중심을 갖는 Class 2 의 세포에서는 아주 작은
모양만을 최고의 반응을 일으키고, 더 큰 사각형은 수용역의 홍분과 억제성 부위를 모두 덮고 있기 때문에 이 신경절세포의 전체 활동전 위는 줄어드는 결과가 된다. 커다란 흥분성 영역을 가진 Class 4 는 억제성 둘레가 자극이 되지 않기 때문에 두꺼비가 훨씬 더 큰 사각 형을 보았을 때 활동을 하게 된다. 결론적으로 Class 1 와 Class 2 의 신경절세포는 먹이자극에 의해서 주로 활동적이며 Class 3 과 Class 4 는 적에 의해 주로 활동한다. 그러나 두꺼비의 망막에 먹이 혹은 적 특이적 탐지기가 있다고 정확하게 말할 수는 없다. 망막에 있는 신 경절세포들의 기능은 눈이 보고 있는 물체의 크기, 각속도(단위 시간 당 물체의 움직이는 속도) 그리고 대비에 대한 기본 정보들을 수집해 서 먹이와 적을 구분하도록 뇌로 보내는 일을 하고 있다. 사실 신호 자극의 분석과 이들에 대한 신경생리학적 기초는 신경행동학의 가장 성공적인 분야들 가운데 하나다 (Camh i 1984, Young 1989). 3 신호자극 반응을 유발시키는 자극들을 열쇠자극 ke y stim ulus 혹은 신호자극 sig n al s ti mulus 이라고 부른다. 해발기구가 중추여과로 자물쇠에 비유 할 수 있는 반면, 이 신호자극은 이 자물쇠에 맞는 자극 형태의 열 쇠가된다. 신호자극은 단일신호(예, 냄새자극, 맛자극, 소리신호, 색깔신호, 형 태신호)가 있는가 하면 복합신호가 있다. 칠면조 암컷은 짧은 목을 보고 비행하는 맹금류를 식별한다. 죽 짧은 목과 긴꼬리를 통해서 생겨나는 날개 모양을 보고 맹금류인지 아닌지를 판단한다(그림 3- 9, 그림 3-10). 새끼 지빠귀는 여러 크기로 만든 마분지 모형 가운데 머리가 무조건 큰것이 아니고, 머리와 몸통 사이의 크기의 비율을 가지고 식별한다.
 4+ t +++ ,+
4+ t +++ ,+
4 해발인 행동학에서는 종종 신호자극과 해발인 releaser 을 동의어로 사용하 고 있다. 그러나 거기에는 아주 작은 차이가 존재한다. 정보전달과 정보입수에 대한 동물들의 적응적 관점에서 두 용어롤 서로 구분해 서 사용하는 다음 두 가지 서로 다른 상황이 있다. 만일 정보입수에 있어서 수신자 한쪽만을 위해 이점이 된다면, 이 적응은 가능한 최 적의 정보를 입수하는 쪽으로 발달할 수 있다. 이에 반해 만일 수신 자는 물론 발신자도 상호 정보교환에 이익이 된다면, 쌍방은 관련 적응을 통해 정보교환의 증진을 위해 기여할 것이다. 이 첫번째 경우는 종간에서 주로 발달해 있고, 두번째 경우는 종
내에서 발달해 있다. 동물들은 식물, 다론 동물종(피식자, 포식자) 그리고 무생물들이 원래 소유하고 있는 표시를 인식할 수 있다. 동 종 내에서는 단지 종내의 의사소통에 기여하며 이 의사소통을 증진 시키는 부수적 구조가 계통발생학적으로 발달되었다. 어떤 나무가 특정 조류에 의해 둥지 장소로 쉽게 인식될 목적으로만 나뭇가지에 색채를 띠지는 않는다. 오히려 새들은 자신들의 둥지 장소를 수관의 형태, 나뭇잎의 형태 등의 표시를 보고 인식하고 있다 . 이것들은 원 래 나무 종 스스로가 주변 환경에 대한 적응으로 소유하고 있는 것 들이다. 포식자와 피식자와의 관계에서 양측은 상대가 쉽게 알아 볼 수 있었던 모든 구조표시와 행동표시들을 피하게 되었다. 포식자는 눈에 띄지 않게 조용히 다가가야 한다. 올빼미는 특별히 발달된 날 개것의 구조로 인해 거의 나는 소리를 내지 않는다 . 이런 식으로 포 식자는 몸을 낮추고 옹크린 채 위장을 한다 . 여기서 정보의 필요성 은 수신자측(포식자)으로 일방적이며, 그래서 정보입수를 증진시키 기 위해 자신의 감각기관의 능력을 높이는 것이다. 이에 비해 종내에서는, 예를 들어 구애행동 혹은 양육행동의 경 우, 정보 요구는 상호적이다. 무엇보다 양측은 상호이해에 관심을 갖는다. 수컷은 암컷을 그리고 암컷의 짝짓기 준비를 인식해야 하며 암컷의 구애행동에 맞추어 반응을 해야만 한다. 반대로 암컷도 수컷 의 접근에 적당한 방식으로 반응을 해야만 한다. 이러한 이유에서 근본적으로 적응은 쌍방에서 일어날 수 있다. 수신자는 자신의 감각 기관들을 증진시킬 수 있으나, 발신자도 좀더 쉽게 인식할 수 있는 자극형태를 발달시켜 자신의 신호를 증진시킬 수있다. 해발인은 상호 의사소통 체계의 구성요소이다 . 그래서 해발인의 진화과정에서 상호이해는 아주 중요하게 작용해 왔다. 정보전달은 이 해발인을 위해서는 주기능이 되며, 신호자극을 위해서는 우연적 인 부기능이 되었다. 재갈매기의 아랫부리에 있는 붉은 반점은 새끼
를 위한 신호로만 발달되었으며, 이에 비해 맹금류의 짧고 긴 꼬리 의 바행 형태는 육상조류에게 공중의 적에 대한 필요한 인식 표시로 단지 우연히 작용해 왔다 . 이런 명백한 차이로 인해 콘라트 로렌츠는 신호자극을 종내의 개 념으로 도입하여 그것을 해발인이라고 불렀다. 그는 해발인을 동종 에 의해 규칙적인 방식으로 반응을 보이는 신호 사용이 종보존의 의 미를 갖는 형태학적 구조 혹은 생득적 동작 형태로 해석하고 있다. 그렇다면 우리는 여기서 해발인에 대해 좀더 간단히 정의내릴 수 있 다. 해발인은 배우자(상대방)에게 반응을 일으키는 임무를 갖고 있는 구조 및 행동의 표시이다 (Lorenz 1935). 그러므로 해발인은 신호자극 의 하위개념에 속해 있다. 모든 해발인은 잠정적인 반응 유발 표시 로 신호자극이 되지만, 모든 신호자극이라고 해서 상호 의사소통에 기여하는 것은 아니다 . 결국 모든 신호자극은 해발인이 될 수 없다. 시각적(색깔, 형태), 음성학적(소리내기) 그리고 화학적(페로몬) 표 시들은 물론 신호 동작들로 이루어진 오히려 매우 눈에 띄고 의식화 된 행동양식과 자세들도 해발인으로 기여할 수 있다. 전달기능을 갖 는 이러한 모든 행동양식들을 표현행동 ex p ress i ve behav i or 이라고 부 른댜 이 행동의 주요 과제인 동종간의 이해를 위해 해발인은 가능 하면 쉽게 인식될 수 있어야 하며 동시에 눈에 띄어야 한다. 그래서 동물계에서 가장 눈에 띄는 구조가 거의 항상 해발인이 되는 것은 우연이 아니다. 대표적인 예들로 숫공작의 부채처럼 펼친 꼬리날개 모습, 숫오리류의 울긋불긋한 깃털 혹은 많은 어린 새의 목구멍 표 시나 부리유두를 들 수 있다. 공격행동의 많은 해발인과 생식격리 작용을 하는 성적 해발인은 동시에 두 가지 성질을 충족시켜야만 한다 . 그들은 가능하면 혼동을 일으켜서는 안 된다 . 다시 말하면 그들은 관련 표시가 다른 종과는 분명히 차이가 있어야 한다. 무엇보다 이것은 같은 장소에 여러 근
연종들이 함께 사는 곳(동소성), 그래서 교잡이 일어날 수 있는 곳에 서 절대적으로 요구되고 있다. 진화과정에서 돌연변이와 자연선택에 의해 형성되어온 해발인의 눈에 띄는 정도는 물론 한계가 있다. 이것은 무엇보다 위장의 요구 와 다른 행동기능의 요구라는 두 요인에 의해 결정된다. 이에 대한 결과로 많은 해발인들은 해발인의 발달에 작용한 다양한 선택압들 사이에 〈타협〉을 이루게 되었다. 해발인들의 이런 타협성은 위장의 요구와 관련해서 아주 분명히 인식될 수 있다. 눈에 띄는 해발인은 종종 집을 수 있는 기관(지느러미의 눈알 무늬 , 오리류의 색깔 있는 날 개 무늬, 공작새의 꼬리날개, 많은 새들의 부풀릴 수 있는 색깔 있는 목주 머니)에서 발달되었다. 이들은 한쪽의 성(대부분 수컷)에서 나타나거 나, 단지 실제 필요한 시기에 한정(조류와 어류에서 발정시기에 혼인색 이 있고 그 밖의 시기에는 보호 혹은 위장색을 띤다)해서 발달한다. 이런 식으로 종내의 정보전달을 위해 필요한 신호는 필요할 때만 눈에 띄 고 위험을 대비하여 시기적 혹은 개체적으로 한정되어 사용된다. 특 히 인상적인 것으로 일부 키클리드 물고기들은 신경조절로 재빨리 색깔을 바꾸어 자기 모습을 변화시킬 수 있는데, 사용하지 않는 시 간에는 눈에 띄는 색깔표시를 지워버릴 뿐만 아니라 여러 행동영역 에 따라서 각기 매우 다양한 신호를 보낼 수 있다. 5 해발인과의사소통 해발인은 위험에 대한 경고신호 혹은 구애행동 둥의 의사소통에 기여하고 있다. 서배너원숭이 Cercop ithe cus ae t h i op s 는 해발인을 통 해 위험의 발생을 예고할 뿐만 아니라 위험의 실체가 무엇인가까지 룰 무리 구성원에게 전해주고 있다. 즉 일련의 비교적 긴 음조로 구
성된 경계음은 표범이 나타났음을 의미하는 소리로 이 소리를 들은 나머지 무리들은 곧바로 나무 꼭대기로 피한다 . 또 짧은 음조로 구 성된 경계음은 매가 나타났음을 알리는 신호로 이 소리를 둘은 무리 들은 하늘을 쳐다보면 아래 덤불 숲으로 피신한다. 짧게 단절된 소 리로 구성된 경계음은 뱀이 나타났다는 신호로 무리는 땅 아래쪽을 살펴보는 행동을 보인다 . 이런 위험에 대한 자세한 정보를 구별하는 것은 이 서배너원숭이뿐만 아니라 닭과 다람쥐 등에서도 관찰되었다. 닭 (Marler 등 1986) 과 꿀벌은 먹이의 종류와 질에 관한 정보를 세분 화하여 전달할 수 있다 . 수탉은 먹이를 발견하게 되면 특이한 소리 를 내는데, 이 소리는 암컷으로 하여금 먹이에 오게 하는 기능을 한 댜 이 먹이음은 좀더 좋은 먹이가 있게 되면 소리의 빈도가 높아진다. 해발인에 대한 다양한 역할은 동물들이 먹이를 취하거나 암컷울 놓고 경쟁할 때 발생하는 갈등상황에서 나타난다. 이러한 갈등상황 에 닥쳤을 때 동물들은 대개 위협 신호 자극을 보내는데 이것은 신 체적 접촉이나 싸움을 통한 상호간의 피해를 최소화하는 기능을 갖 고 있다 붉은사슴의 경우 가을철 발정기가 시작되면 무리중의 암컷 을 차지하기 위해 수컷들끼리 치열한 싸움을 벌인다. 그 결과 치명 적인 부상을 당하는 해를 입는다 . 그러나 대부분의 경우 직접 싸움 을 하기 전에 이들은 위협신호로 포효를 한다. 서로간에 경쟁적으로 포효를 하면서 자신이 더 힘이 셈을 과시한다. 실제로 포효를 하기 위한 근육의 구성이 싸움을 잘하는 것과 관계가 있기 때문에 포효하 는 정도가 센 사슴이 싸움을 더 격렬히 잘할 수 있다고 예측할 수 있댜 따라서 서로간에 상대방의 포효소리를 듣고 자신이 실제로 싸 우면 이길 수 있는지의 여부를 판단할 수 있을 뿐만 아니라 우열관 계도 정할 수 있다. 만약 포효하는 비율이 서로 비슷하다면 이들은 다음 단계의 신호를 만들어 서로간의 전력을 평가한다. 즉 서로 왔 다갔다 하는 식으로 나란히 걸으면서 서로간의 눈을 응시하는 등의
행동으로 전력을 탐색한다 . 이런 과정에서도 우열이 결정되지 않을 경우에는 비로소 싸우는 행동을 나타낸다. 많은 해발인에 의한 의사소통이 구애 행동하는 가운데 일어나고 있다. 대부분의 경우 수컷에 의한 신호작용이 활발한데 어떤 해발인 이 의사소통이 되고 있는지는 밝히기가 쉽지 않다. 분명한 것은 의 사소통을 통해 종과 성을 식별한다는 것이다. 예를 들면 파충류인 이구아나는 종특이적인 머리 움직임을 보이며, 반딧불이는 빛을 내 는 주기가 다르게 나타나고, 농게는 제각기 다론 방식으로 집게를 혼들며, 그리고 개구리와 귀뚜라미는 소리내는 주기를 달리하여 신 호를보낸다. 동물들의 이러한 의사소통을 위한 많은 해발인들이 필요이상으로 공들여져 있는 생각을 하게 한다. 단순히 종과 성을 알리는 신호 역 할만이라면 그러한 과장된 행동양식이 구태여 필요했겠느냐라는 의 문을 제기할 수 있다. 예를 들어, 제비는 오랜 진화과정을 통해 수 컷의 꼬리 깃털이 길어진 것은 암컷의 선택작용에 의해 과장된 장식 으로 발달하게 되었다. 죽 암컷이 꼬리깃이 긴 수컷울 선택하면 할 수록 그 수컷의 자식들은 긴 꼬리깃을 갖고 태어나는 경향을 보이면 암컷의 자식들도 긴 꼬리깃에 대해 더 많은 선호를 나타내게 된다. 최근 해밀턴 H amilt on 과 주크 Z uk:(1 984) 는 성장식과 과시행동의 진 화를 다음과 같이 설명하고 있다 . 건강한 수컷은 자신이 질병이 없 고 기생충이 없음을 나타내야만 한다. 이 질병이 어린 새끼의 치사 와 관계되므로 암컷은 반드시 질병이 없는 우수한 유전자만을 갖고 있는 건강한 수컷을 선택하여 교미를 하여야 한다. 여기서 문제는 암컷이 그러한 우수한 유전자를 어떤 신호를 통해 찾아낼 수 있느냐 댜 그 신호를 암컷은 수컷의 과시행동에서 얻는다고 주장하고 있 다. 죽 과시행동은 많은 에너지를 소모시키는 행위로써 우수한 유전 자를 갖고 있지 않은 개체의 경우는 그러한 소모적인 행위를 제대로
수 59 행할 수 없다는 것이다. 꼬리깃의 경우도 마찬가지의 예라고 볼 수 있다 (Andersson 1982, Grafe n 1990) 어쨌든 이런 장식이나 과장이 신호의 역할을 한다면 누구를 위한 신호인가? 우선 성적 과시는 암컷에 보내는 신호라고 할 수 있겠지 만 또한 다른 수컷에게 보내는 신호일 수도 있다. 즉 구애행동이 수 컷간의 과시행동일 뿐이고 암컷은 단지 그 행동과정에 우세한 수컷 울 선택할 뿐이라고 생각을 해볼 수 있다. 닭, 검은부리까치 그리고 오리의 암컷들은 우세한 수컷들을 선호한다. 우세한 수컷들은 다른 수컷과 경쟁하여 이길 수 있는 능력으로 자신의 건강함과 기질을 보 여줌으로써 이러한 암컷들은 수컷과 수컷의 평가에서 진화되어온 신 호를 사용하여 질적으로 우수한 배우자를 선택할 수 있었을 것이다 (Graves 등 1985, Komers 와 Dh ind sa 1989, Brodsky 등 1988) . 6 종간 해발인 원래 해발인이라는 말은 종내에서만 사용해 왔다. 그러나 그 동안 상호적 정보전달이 가끔 여러 종들에 속해 있는 구성원간에도 일어 나고 있는 것이 알려지게 되었다. 이 경우 종간 해발인이라고 부르 는 것이 옳다고 본다. 한 예로써 많은 명금류(번식기에 노래를 하는 참새목의 종류)의 경계음은 서로 매우 비슷하며 그렇기 때문에 자기 종의 구성원뿐만 아니라 다른 종에게도 위험을 알릴 수 있다(그림 3- 11). 물론 경계음이 아닌 다른 기능의 소리는 매우 다르다. 많은 종 들은 그들이 스스로 이 경계음을 내지 못할 때도 다른 종의 경계음 울 듣고 반응을 보인다 (Marler 1956). 특히 종간 해발인은 다음 두 영역에서 찾아볼 수 있다. 의태는 관 련 신호구조를 발달시킴으로써, 죽 모델의 신호를 모방함으로써 발
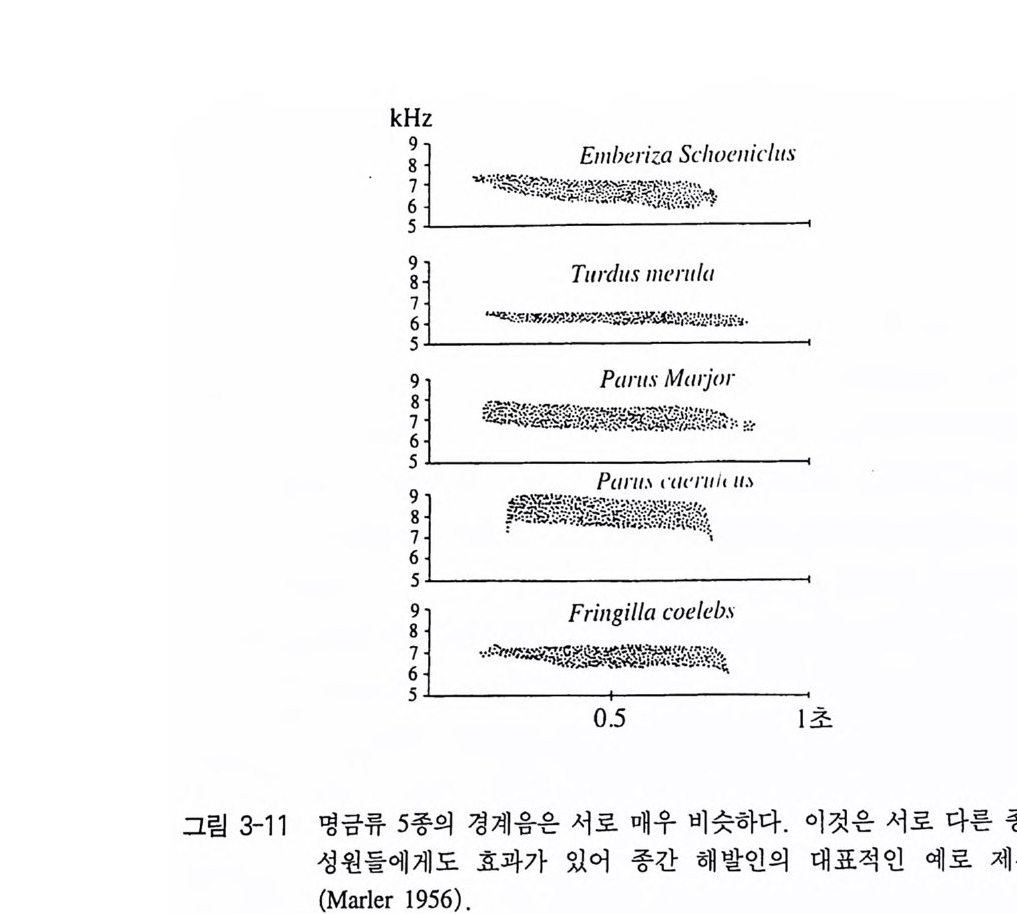 k Hz9 8765
k Hz9 8765
신자가 정보전달의 증진에 기여하며, 공생은 특정 신호가 상호인식 과 공생 파트너의 이해에 기여하고 있다. 이런 이해체계는 예를 들 어 물고기의 청결공생에서 볼 수 있다 . 여러 물고기종들은 공생 파 트너로 하여금 자신을 알아채게 하는데 전문화되어 있다. 대개 이 물고기들은 눈에 띄는 색깔과 마찬가지로 눈에 띄는 수영방식을 보 이고 있다. 이 신호로 인해 이들은 청소물고기가 안전하게 접근할 수 있게 하며 청소물고기는 특이한 몸동작의 신호로, 입을 크게 벌 린다거나 아가미 뚜껑을 짜내는 듯한 시능을 하면서 자신이 먹이동 물이 아니라는 신호를 보낸다. 그러므로 신호자극과 해발인 사이의 결정적인 차이는 정보전달이
동종간 혹은 타종간에 있지 않고 생물학적 이익이 일방적이냐 혹은 상호적이냐에 따라 결정된다 . 이때 다만 종내에 있는 해발인을 우리 는 〈 사회 적 해 발인 socia l releaser 〉 이 라는 말로 좀더 한정 하여 부를 수 있다 이런 차이를 규정짓는 기준과 관련해서 한 가지 예외가 의태 에서 찾아볼 수 있다. 여기서 생물학적 이익은 분명히 일방적이며 그것도 발신자에만 이익이 된다. 그럼에도 불구하고 의태는 상호의 사 소통체계의 모든 특징을 보여주고 있다 . 그러나 모방자가 자신의 신호로 상호 이해의 체계에 적응되었고, 신호수신자 즉 속임을 당하 는 자도 파트너와 함께 발달해 왔으며 이때 상호이익이 될 수 있다 (F it z gi bbon 과 Fanshawe 1988) . 7 자극의 통합 많은 행동양식에는 단 하나의 유발 자극만이 있는 것이 아니고 관 련 행동을 유발시키기 위해 여러 개의 자극들이 함께 작용하기도 한 댜 이 경우 각각의 자극들은 행동의 영향을 미치는 데 서로 촉진시 키는 역할을 한다. 이런 현상은 자이츠 Se itz(1 940) 가 처음으로 키클 리드 물고기의 싸움행동과 위협행동에 대한 실험 결과 〈자극통합 stim ulus summa ti on 〉이 라고 불리 게 되 었다 이 현상온 각각의 자극들 울 분리해서 제공하거나 여러 방법으로 이 자극들을 서로 결합할 수 있기 때문에 모형실험에 의해 가장 훌륭하게 밝혀질 수 있었다 . 재 갈매기가 둥지에서 굴러간 알을 다시 가져오는 반응은 알의 크기는 물론 알의 바탕색과 반점의 중요한 표시들에 의해 일어난다. 큰 알 은 원래의 알보다 더 강한 반응을 보이나, 작지만 반점이 있는 알은 반점이 없는 알보다 더 강한 반응을 보이며, 그리고 녹색 계통의 알 은 갈색 계통인 알보다 더 큰 반응을 보인다 . 가장 효과가 있는 모
형은 양의 반응을 보였던 표시들을 모두 합친 것이었다. 한편 하나 의 표시를 빼면 (예를 들어 반점을 없애면) 다른 표시를 확대시켜서, 예 를 들면 모형을 크게 하여 반응을 원래대로 같게 할 수 있다. 멧비 둘기 암컷의 산란은 수컷으로부터 오는 자극을 통해 유도된다. 만일 암수를 함께 두면 이러한 효과가 가장 빨리 나타난다 . 그러나 번식 집단에서 녹음한 소리를 틀어주는 것만으로도 이 산란을 유도할 수 있고, 반대로 수컷과 함께 있게 했을 때 암컷의 귀를 수술해서 인위 적으로 귀머거리를 만들 수도 있다 . 첫번째의 경우는 암컷이 단지 음성학적으로 감지할 수 있으며 , 두번째의 경우는 시각적 그리고 촉 각적으로 감지할 수 있다. 두 자극군은 효과면에서 서로 종속적이 다 . 다시 말해 두 자극군이 함께 작용하면 가장 강한 반응을 일으킬 수 있다 비슷한 현상이 그 동안 많은 행동영역에서 알려졌다 . 물론 이런 상호강화 현상은 여러 종류의 자극들에만 한정되어 나타나는 것은 아니다 오히려 같은 종류의 자극일지라도 만일 동일 발신자에 의해 여러 번에 걸쳐 연달아 제공되거나 여러 발신자에 의해 동시에 제공되면 그 효과가 훨씬 강화될 수 있다. 아마 많은 동물들의 집단 구애의 효과가 이런 현상에 기인하고 있다. 이렇게 집단구애를 하는 종들은 여러 마리의 수컷들이 공동으로 구애에 참여하여 동종의 암 컷들을 유인하는 데 특히 강력한 효과를 지닌다 . 결론적으로 어떤 한 상황의 전체 자극치는 참여한 각각의 자극 모 두에 의해 구성된다 . 정량적 조사를 통해서 각각의 자극들은 간혹 효과에 있어서 실제로 합쳐져 나타나는 반면, 보통 한 상황의 전체 자극치는 각각의 자극들의 정확한 합에 일치하지 않고 서로 촉진시 킨댜 그래서 〈상호적 자극강화 re cip rocal stim u lus enhancemen t〉라는 말이 생겨나게 되었다 (C uri o 1969).
8 초정상자극 가끔 동물들의 자연적 신호구조는 정보전달의 의미에 있어서 가장 최적의 해발인이 되지 못하고 다른 자극 유형에 의해 그 효과를 능 가한댜 어떤 특정 행동양식이 자연적인 것보다 더 잘 유발시킬 수 있는 자극을 〈초정 상자극 sup er normal s ti mulus 〉이 라고 한다. 이 초정상자극 현상 역시 모형실험에 의해 검토가 가능하다. 육상 에서 번식하는 조류, 예를 들면 검은머리물떼새, 재갈매기 혹은 회 색기러기에게 원래의 알보다 더 눈에 띄는 색깔이나 더 큰 알 혹은 모형알을 제공하면, 원래의 알을 선택하지 않고 눈에 띄는 혹은 크 기가 큰 알을 둥지로 굴린다거나 포란을 한다 . 재갈매기는 정상적인 알색깔보다 인위적으로 푸르고, 노랗고 혹은 붉게 칠한 알에 평균적으 로 더 오래 포란을 한다. 검은머리물떼새는 정상적으로 3 개, 많아야 4 개 알을 낳는데 3 개의 알보다 5 개의 알품기를 선호한다(그림 3-12, Tin b erge n 1948). 유럽굴뚝나비는 자연적인 날개 색보다 배경에 의해
 a
a
더 돋보이는 검정 암컷의 날개모형에 더 높은 빈도의 반응을 보인 다 . 주행성 나비에 속하는 표범나비는 정상보다 높은 날개 진동수를 가진 모형을 선호한다. 숫반딧불이는 자기종 암컷의 불빛보다 더 큰 불빛과 더 노란색의 불빛에 끌린다 . 새끼 재갈매기는 갈매기 머리를 거의 자연에 가깝게 만든 석고 모형보다 3 개의 흰색 띠가 있으면서 끝이 뾰쪽하고 붉고 가는 막대기에 더 자주 쓴다. 검은방울새 Sp inu s s pi nus 는 자연의 소리 (자기 종의 접촉음)를 들려 주었을 때 보다 특정 주파수 대역을 여과해서 들려주었을 때 더 강한 반응울 보인다. 큰 가시고기 수컷은 원래 크기의 암컷의 모형보다 더 크고 더 납작한 모양을 선호한다 (Helb 1973, Rowland 1989) . 초정상자극의 발생은 선택압이 해발인으로 하여금 가능하면 크게 눈에 띄도록 영향을 미쳐온 사실에 주목하게 된다 . 그렇지만 해발인 의 타협이 생겨날 수 있도록 자연조건하에서 다른 선택압이 작용하 게 되었다 . 위장의 요구는 크게 눈에 띄는 쪽으로 관련 신호구조가 계속 발달하는 것과 대립하게 된다. 무엇보다 이것은 육상에서 번식 하는 조류의 알 색깔과 땅에서 사는 데 적응된 나비 암컷의 은폐색 이 해당된다. 그러나 다른 행동의 요구 역시 이러한 발달과 대립할 수 있었다. 갈매기가 가는 막대모양의 부리를 갖고 있었다면 먹이 탐색에 아주 불편했었을 것이며, 나비의 날개 진동수는 동료에 대한 신호보다 비행기작의 요구에 먼저 적응시켜야만 했었을 것이다 (Sta d don 1975) . 가끔 초정상자극은 자연상태에서도 나타나며, 그것도 번식기생과 사회기생에서 나타난다. 새로 태어난 뻐꾸기는 새끼 숙주새의 목구 멍보다 실제 더 광택이 나는 색깔을 띤 목구멍을 갖고 있다. 결국 이것은 강력한 해발 능력을 갖게 되어 숙주 어미는 어린 번식기생자 에게 어쩔수 없이 먹이를 집어 넣을 수 밖에 없다. 화학적 이해의 영역에서 이것과 비교할 만한 것이 딱정벌레목(目)에 속한 반날개과
(科) S t a p h ylini dae 의 애벌레들은 개미집에서 세들어 살고 있다. 그들 은 표피분비선에서 개미에게 양육행동(먹이주기, 핥아주기)을 유발시 키는 물질을 분비한다. 이 물질은 개미 스스로 분비하는 것보다 더 강력한 효과를 지니고 있어 개미가 자기 새끼를 돌보는 대신 반날개 새끼들을 돌보게 된다. 9 의부자극의 다양성 이미 언급했듯이, 외부자극들은 여러 가지 방식으로 행동에 영향 울 미칠 수 있다 . 죽 한 행동양식을 유발시킬 수 있고, 그 행동의 정위를 결정지을 수가 있으며, 혹은 한 동물의 행동준비를 바꿀 수 가 있다. 그리고 다른 자극으로 인한 한 행동양식의 해발 정도에도 영향을 미칠 수 있다. 종종 하나의 자극 혹은 동일 자극에 의해 두 세 가지 과제가 관찰될 수 있다. 이에 대해 분명히 분리된 과제의 예가 있다. 나비에게 방향족 화합물 냄새를 피워 색종이에 내려앉게 할 수 있다 . 이 후각 자극은 탐색행동을 유발시키고, 색종이의 시각 자극은 비행착륙의 방향을 결정짓게 한다. 물벼룩은 물속에 CO2 의 양이 차츰 증가하기 시작하면 산소가 풍부한 곳으로 가기 위해 물표 면으로 모인다. 여기서도 화학적 자극은 수영동작을 가속시키며, 빛 이 오는 방향(시각적 자극)에 의해 정위가 결정된다. 이때 인위적으 로 빛을 수조의 측면 혹은 바닥에서 비춰주면 그곳이 산소가 부족한 곳일지라도 빛이 있는 곳으로만 모인다. 재갈매기 새끼들은 위험에 처하면 풀더미속으로 숨는다. 이러한 행동은 어미의 경계음에 의해 유발되나, 온신처에서 나온 시각적 자극을 통해 방향이 결정된다. 인위적인 상황에서 이런 예가 목격된 적이 있었다. 괭이갈매기들의 둥지 바로 옆에 설치된 위장망 안에서 관찰자의 부주의는 괭이갈매
기 성체들의 경계움을 유발시킨 결과가 되었다. 이때 새끼들은 도피 로 반응을 보였으며, 가장 빨리 다가갈 수 있는 은신처로 숨으려 했 을 때 몇 마리의 새끼가 벌써 이런 식으로 반응을 보인 후 위장막 안으로 들어왔다. 거기서 새끼들은 방금 어미에 의한 주의로 인해 옹크리고 있었다. 수많은 외부자극들은 직접적인 유발효과로 일종의 장시간 효과를 일으킬 수 있으며 동물의 행동준비를 바꿀 수가 있기 때문에 동물들 은 관련 자극을 제시받기 전보다 달리 행동한다. 암집쥐는 생후 1 일 된 죽은 새끼에게 매우 약한 양육행동으로 반응을 보인다. 그러나 이 암컷에게 살아있는 새끼의 형태로 실제 그 전에 더 강력한 자극 울 주게 되면, 죽은 그 새끼에 대해서도 강화된 반응을 보인다. 이 미 철망 안에서 보는 것만으로 행동준비를 높일 수 있기 때문에 그 암컷이 살아 있는 새끼를 실제 접촉시킬 필요는 없다. 10 의부자극의 종속 외부자극의 종속과 관련해서 동물의 행동을 다음 두 가지로 구분 할수있다. 첫째, 단지 처음에는 외부의 자극이 필요하지만 일단 행동이 진행 되면 끝까지 항상 같은 방식으로 진행되는 행동이 있으며, 둘째, 실 제 복잡한 행동으로, 그 형태에 있어서 외부자극에 의해 바꾸어질 수 있으며 또 완전히 그에 종속되어 일어난다. 첫번째 행동양식 집 단을 고정 행 동패 턴 fixe d acti on pat t em (FAP) 이 라고 하며, 두번째 행동양식 집단을 주성 taxi s 이라고 부른다.
10, 1 고정행동 패턴 고정행동 패턴은 코니시 Ko ni sh i(1 966) 가 정의한 바에 의하면 생물 학적으로 의미 있는 움직임의 형태를 표현하는 근수축의 시간적 및 공간적으로 배열된 순서가 된다. 고정행동 패턴은 종특이적인 외부 자극에 의해 해발되어지나, 고정행동 패턴 과정을 유지하는 데는 이 외부자극이 필요치 않다. 고정행동 패턴을 형성하고 있는 각 구성요 소들의 연합은 중추계에서 일어나며 순간적으로 영향을 미치는 외부 자극들의 종류와 방향과는 별개로 일어난다. 그러나 영향을 미칠 수 있는 것은 의부자극의 강도와 속도가 된다. 즉 하나의 고정행동 패턴 은 의부자극의 크기에 따라 빨리 여러 강도로 일어날 수 있으며, 그 러나 형태는 항상 동일하게 진행된다 . 이런 〈형태불변〉 혹은 〈형태 부동〉에 기초 해서 어떤 동물의 고정행동 패턴은 아주 전형적이며 형태학적 형질처럼 정확히 특징지을 수 있다. 그 밖에도 이 형태불 변은 고정행동 패턴이 생득적, 즉 이 행동 패턴의 형질이 유전자에 프로그램화되어 있음을 보여주고 있다. 특히 고정행동 패턴은 화장 (닦기, 굵기, 혼들기), 짝짓기행동, 둥지짓기 혹은 섭식행동 영역에서 두드러지게 나타난다.
10.2 주성* 어떤 행동양식의 정위(定位) 구성분은 형태부동적 고정행동 패턴 에 비해 상당히 변화가 있으며 외부자극에 의해서도 형태가 달라진 다. 여기서 외부자극에 의해서 이 구성분은 실행될 뿐만 아니라 실 행되고 있는 동안에도 계속해서 통제를 받는다. 두 행동성분(고정행 * 자연환경 조건적 요인에 반응을 보이는 동물의 주성은 제 6 장 리듬과 정위에서 따로 다룬다 .
 :一
:一
동 패턴과 주성)간의 차이는 회색기러기의 알굴리기 행동의 예에서 분명히 알 수 있다 . 많은 육상번식성 조류의 경우 둥지 밖에 있는 알을 둥지 안으로 끌어들이는 연속적인 동작에서 찾아볼 수 있다. 이때 목은 길게 앞으로 뻗게 되고 부리의 좌우 균형 유지 동작과 함 께 천천히 목을 안으로 구부리면 알은 둥지 안으로 들어간다(그림 3- 13). 만일 이미 알굴리기가 시작된 기러기로부터 그 알을 치워버리 면 목을 멀리 뻗어 목을 안으로 구부리는 동작은 하지만 좌우 균형 을 유지하고자 하는 수정운동은 일어나지 않는다. 이 알굴리기 자체 는 한번 행해지면 외부자극 없이도 계속 진행되는 고정행동 패턴이라 고 할 수 있다. 이에 비해 균형 유지 동작은 정위반응으로 이 정위반응 이 일어나고 또 그것이 진행되는 것은 외부자극에 종속되어 나타난다. 죽 알이 있을 때는 수정운동이 일어나다가 알이 없을 때는 그 수정운 동이 사라지는 것이다. 이때 우리는 이것을 주성이라고 한다. 주성은 고정행동 패턴과는 대조적으로 중추에서 조절되는 것이 아니고 말초 에 의해 통제를 받고 있기 때문에 나타난다 (Lorenz 와 Tin b erge n 1938) . 회색기러기의 알굴리기 동작은 고정행동 패턴과 주성이 동시에 일 어나고 있는 셈이다. 그러나 종종 이 두 요소가 동시에 일어나지 않 고 연결해서 일어나고 있다. 개구리 한 마리가 먹이인 파리를 발견
하면 몸의 축이 정확히 정면에 설 때까지 몸을 돌린다 . 주성의 요소 로 부를 수 있는 이 방향성 동작이 있은 후에야 고정행동 패턴으로 이어진다 즉 파리를 잡기 위해 혀를 내미는 동작이 이어지는 것이다. 고정행동 패턴과 주성이라는 개념은 일견 비슷한 현상의 개념인 완료행 동 consummato r y ac t과 갈망행 동 app e tit ive behav i or 과는 동일하 게 취급하지 않는다. 완료행동과 갈망행동의 두 개념은 반응 특이적 인 에너지 관점에서 차이가 있지만 의부자극의 종속에 관련해서는 차이가 없다. 일부 개념적으로 일치라는 것은 주성과 갈망행동에는 큰 변이가 존재하고 있는 반면, 고정행동 패턴과 완료행동은 그런 변이가 적다. 두 개념간에 관계를 지어 본다면 완료행동은 확실히 고정행동 패턴인 반면, 갈망행동에는 주성과 고정행동 패턴이 포함 될 수 있다. 예로써 매가 사냥을 위해 자기의 터를 두루 살펴보다가 마침내 한 떼의 물오리를 발견하고 하강비행을 하는 것은 바로 갈망 행동이다. 이것은 확실히 물오리인 먹이를 발견하여 그 쪽을 향하는 과정은 주성에 해당되며 그 물오리를 잡으러 달려드는 행동은 고정 행동 패턴이 된다. 바로 이때의 갈망행동에는 주성과 고정행동 패턴 이 포함되어 있지만, 그 다음 한 마리 물오리를 잡아서 갈기갈기 찢 는 행동은 바로 마지막으로 이루어지는 섭식행동의 완료동작에 해당 된다. 물론 이때의 완료동작은 고정행동 패턴에 속한다. 또 하나의 다른 예에서 이 관계를 보다 명확히 할 수 있다. 새의 경우 자기 집 울 짓기 전에는 집 지을 재료를 주둥이에 물고 있다가 집짓기에 적 당한 장소를 발견하면 집을 짓는, 즉 집짓기 행동의 완료동작으로 들어가는 것이다. 그러나 여기서 새가 둥지 재료를 주둥이에 물고 이리 저리 다니는 행동은 갈망행동으로 이 갈망행동으로 들어가는 데에는 반드시 특정한 자극이 요구되지 않는다. 그래서 이때 우리는 자극의 근원을 향해서 움직이는 주성이라고 할 수 없고 단지 이 갈 망행동은 고정행동 패턴에 속한다.
제꾹J 행동의 순서 행동양식들은 시간적인 순서예 따라 나타날 수 있다. 이들은 하나 의 행동양식이 나타나면 곧바로 이어서 다른 행동양식이 나타날 수 있고 혹은 서로가 영향을 미쳐 행동이 유발되는 데 방해를 일으킬 수있다. l 행동양식의 순서 많은 행동양식들은 서로가 생물학적 목적을 추구하기 위한 기능적 인 관계를 형성하고 있다. 무엇보다 사회적 행동영역에서 예를 들면 짝짓기나 의식화된 싸움에서 관찰될 수 있는 반응연쇄 reac ti onch ain 혹은 동작연쇄 ac t ch ai n 가 여기에 속한다. 연속적으로 이어진 각 동 작(행위)들의 연쇄로 이루어진 행동에서 이 각각의 동작들은 항상 똑 같이 순서에 의해 나타난다. 물론 경우에 따라서는 유연성을 갖고 매우 달리 나타날 수 있다. 일찍이 행동학에서 관찰된 바 있는 큰가 시고기의 짝짓기 행동은 이 동작연쇄를 단적으로 보여주는 좋은 예
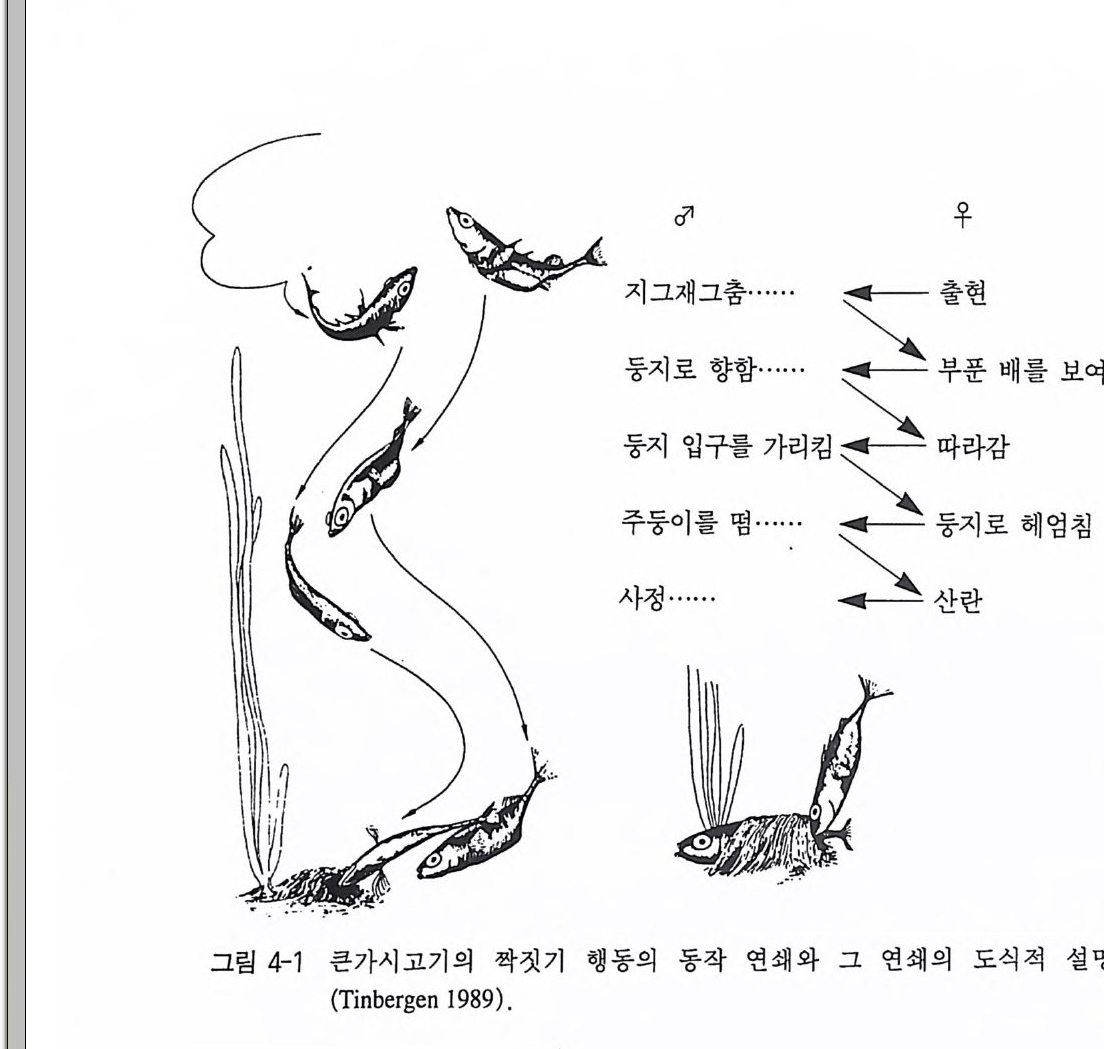 〉二 『呼平約約I 다군노二\〔J[ \}L\L\묘紅回우呼呼 :\감로 H l 헤를 엄 보침 여 줌
〉二 『呼平約約I 다군노二\〔J[ \}L\L\묘紅回우呼呼 :\감로 H l 헤를 엄 보침 여 줌
가 된다. 짝짓기 행동의 진행과정에서 배우자의 이전의 동작이 상대 방의 다음에 이어지는 동작을 유발시킨다(그림 4-1) . 만일 배우자의 반응이 멈추게 되면 상대방은 이전의 동작을 반복하거나 이 동작연 쇄가 해체될 수 있다. 이와 비슷한 동작연쇄는 다른 많은 동물종의 짝짓기 행동에서도 잘 알려져 있다. 동작연쇄는 대개 각 동작들을 시간적 순서로 나타낼 수 있다. 그 렇지만 거기에는 매우 다양한 행동의 완성단계에 이르는 다수의 연 합현상이 존재한다. 이 연합현상은 근육의 개별적 운동 수준에서 시
작된다 물고기 지느러미에 있는 각 지느러미 살(기조)의 리드미컬한 움직임은 두 길항적인 근육이 교대로 수축을 함으로써 일어난다. 전 체 지느러미의 움직임을 위해서 지느러미 기조의 개별적 움직임이 상위에 속한 연합중추에 의해 이루어져야만 한다. 이동행동, 짝짓기 행동, 집짓기 행동 혹은 공격행동에 따라 여러 지느러미의 움직임이 의미 있는 방식으로 연합해야 한다. 일반적으로 관련행동 내에서 개별적 행동양식들이 고도로 연합하 여 시간적으로 나타날 수 있다. 종종 행동기능 영역 내에서 혹은 그 부분영역 내에서 어떤 특정 행동요소들은 그것들이 발생하는 빈도에 서 증감을 보여주거나, 행동요소들 사이에서 특정 빈도로 연속적으 로 나타난다. 번식기 내에서 양육준비(둥지짓기), 구애와 교미 그리 고 양육행동의 행동양식들은 다소 정확한 순서로 연결해서 일어나 며, 또 각각의 이 행동양식 내에서도 개별적인 부분 동작들(예, 조류 의 둥지짓기의 경우 둥지 재료의 수집, 운반 그리고 建造가 될 수 있다)이 어쩌면 시간적 순서로 실행될 수 있다. 이런 식으로 서로 연결된 행 동양식들은 어떤 형태로든 공통의 상위에 속해 있는 중추로부터 종 속되어 있다. 이것은 어떤 동물의 전체 행동이 계층적 구조로 이루 어진 순서가 있음을 뜻한다. 이 순서는 이미 동인을 설명할 때 확인된 바 있으며, 이미 일찍이 확인된 기초적인 의미는 행동 구성에 대한 가상 모델의 개발이 커다란 역할을 했었다(본능 모델 참조). 2 갈등행동 행동 요소들의 상호적인 순서 이외에도 두 행동 요소가 상반되어 발전할 수 있다. 행동양식들은 서로 배타적일 수 있다. 다시 말하면 한 행위의 수행은 동시에 다른 한 행위의 소환을 방해할 수 있다.
이런 상호 억제는 그것이 행동의 혼란을 막아 주고 또 여러 행동양 식들이 순서에 의해 잇달아 일어날 수 있도록 보장하기 때문에 생물 학적으로 의미가 있다. 만일 한 동물이 먹이와 성적 파트너를 놓고 동시에 자극을 받거나, 이미 이런 동인 상태 아래에서 이 두 자극에 반응을 보일 준비가 되어 있다면, 이에 대한 반응이 동시에 이루어 지지 않고 순서대로 일어나거나 또 동시에 홍분된 두 행동양식 가운 데 한 행동양식이 우위에 서서 다른 행동양식을 잠시 억제시키는 것 은 생물학적으로 일종의 이점이 될 수 있다 . 이런 상호억제는 수많 은 행동영역으로부터 잘 알려져 있다. 이것은 여러 행동영역간에 특 히 자주 발생한다. 그것은 공격과 도피의 요소의 길항적인 영역에서 매우 두드러진다. 정상적으로는 외부 상황에 의해 동인 상태에 이르면 어떤 행동양 식의 유발이 정확히 결정되어야 한다. 그러나 가끔 두 개의 서로 일 치할 수 없는 행동 성향이 동시에 그리고 거의 같이 강하게 활성화 되어 유발될 때 그 가운데 하나도 우위에 서지 못하는 경우가 발생할 수 있다 . 이 경우 다움 세 가지로 표현할 수 있는 갈등행동 con fli c t behav i or 을 제시할 수 있다. 죽 상반 감정의 공존행동, 대상 전가행 동, 그리고 전이행동이 갈등행동에서 비롯된다. 2. 1 상반 감정의 공존행동 종종 두개의 상반된 행동양식이 다소 급히 바뀌면서 잇달아 나타 난댜 무엇보다 이것은 동일 대상에게 두 영역에 대한 자극, 예를 들어 경쟁자에게 공격과 도피를 위한 동시적인 자극이나 성적 파트 너에게 구애와 공격을 위한 자극을 제공했을 때 해당된다. 즉 이 상 황은 상반된 반응을 일으키는 두 자극원이 서로 가까이 인접되어 있 울 때 발생한다. 이런 상황에 대해 많이 인용되고 있는 예로 서열이
높은 새 가까이에 배고픈 새 한 마리가 먹이장소를 차지하는 것을 들 수 있다. 이 먹이장소는 배고픈 새가 먹이로 접근하게 하는 자극 이 되며, 이때 이 새는 서열이 높은 새에 의해 도피행동을 유발하게 한댜 이러한 상황에서 불완전한 섭식동작(턱을 서로 비비는 행동)과 도피동작(목빼기, 꼬리움직임)이 교대로 나타날 수 있다. 진화과정에서 두 행동영역의 요소들은 의식화를 통해 시간적으로 서로 긴밀하게 연결되어 왔고 오히려 동시에 나타날 수 있었다. 동 물들의 위협행동은 도피행동과 공격행동의 요소가 결합된 행동임을 쉽게 알 수 있다 . 이러한 몸짓 혹은 동작을 〈상반감정의 공존행동 ambiv a lent behav i or 〉 이라고 부른댜 이런 의미로 구애행동도 성적 행 동 요소로 공격과 도피가 함께 구성되어 있는 상반 감정의 공존 성 격을가질수있다 . 2,2 대상전가행동 상반된 성향을 통해 부정적으로 영향을 미친 하나의 행동양식이 정당한, 즉 자연적인 물체에 향하지 못했을 때 , 다른 형태로 갈등상 황은 표현될 수 있다. 무엇보다 이 경우도 공격성향과 도피성향 간 의 저항에서 관찰될 수 있다. 가장 잘 알려진 한 예는 재갈매기의 싸움행동에서 나타나 있다. 두 경쟁자 가운데 한 마리가 분명히 하 위에 있다면, 하위자는 공격반응으로써 부리를 들어서 상대방을 찍 는 것이 아니고 중간목표물, 즉 폴더미를 향하여 찍는 시늄을 한다 (그림 4-2.2 ). 비슷한 식으로 화가 난 키클리드 물고기의 물기동작은 정상적으로는 경쟁자의 적을 향해 이루어지나 땅바닥을 향해 이루어 질 수 있다. 이 행동의 정위는 대상 물체가 바뀌면서 그에 따라 방 향도 바뀐 것이다. 그래서 우리는 이런 행동을 〈대상 전가행동 re di recte d behav i or 〉이 라고 부른다.
 : L.: 'I, . ~ 2
: L.: 'I, . ~ 2
그림 4-2 동물들의 여러 종류의 전이행동 (T i nber g en 1989) 과 대상전가행동. l 싸 움충동을 배 출 할 때 취하는 재갈매기의 둥지 짓는 동작 . 중간 정도의 강 도 . 2 싸움충동을 배 출 할 때 (강도가 높을 때) 취하는 대상전가 행동. 3 싸움충동을 배 출 할 때 취하는 검은머리물떼새의 잠자는 자세 . 4 싸움충 동을 배 출 할 때 취하는 뒤부리장다리물떼새의 잠자는 동작 . 5 싸움충동 을 배출할 때 취하는 큰가시고기의 모래 파는 자세 . 6 성적 충동을 배출 하고자 할 때 취하는 비오리의 화장행동 . 7 성적 충동을 배 출 할 때 취하 는 발구지의 화장행동 . 8 성적 충동을 배출할 때 취하는 원앙의 화장행 동. 9 성적 충동을 배 출 할 때 취하는 청둥오리의 화장행동. 10 성적 충 동을 배출할 때 취하는 뒤부리장다리물떼새의 화장행동 . 11 성적 충동을 배출할 때 취하는 유럽푸른해오라기의 먹이 잡는 동작 . 12, 13, 14, 싸움 충동을 배출할 때 취하는 가마우지의 구애동작 . 15 성적 충동을 배출할 때 취하는 재갈매기의 구결행동. 16 싸움충동을 배 출 할 때 취하는 수탉 의 먹이 쪼는 행동 .
대상 전가행동들이 사회적 영역에서도 매우 두드러지게 나타나고 있댜 만일 한 동물이 상위자의 동료로부터 위협 혹은 공격을 당했 울 때, 그의 적대적 반응은 종종 공격자 자체에 향하지 않고, 그는 자기보다 서열이 낮은 하위자를 향한다 . 마치 이것은 인간사회에서도 직장의 상관에게 꾸지람을 둘은 회사원이 그의 분노를 그 상관한테 풀지 못하고 자기 부하 직원에게 화를 내는 것과 비교할 수 있다 . 2.3 전이행동 갈등상황 가운데 기대한 행동이 나타나지 않고, 다시 말해 예측된 행동양식의 상황이 전혀 일어나지 않고 완전히 엉뚱한 행동이 나타 나는 경우가 있다. 이에 대해 가장 혼히 인용되는 예는 두 마리의 수탉이 싸움을 하던 중 갑자기 그중에 한 마리가 싸움을 중지하고는 마치 땅바닥에서 먹이를 찾는 것처럼 쪼는 행동을 취한다(그림 4- 2). 이러한 행동을 〈전이행동 d i s p lacemen t behav i or 〉이라고 부른다. 전이행동은 두 개의 서로 모순되는 행동 성향들이 동시에 그리고
대략 같은 강도로 활성화되어 서로 억제상황에 놓여 있을 때에 일어 난다 이런 억제상황에서 억제된 행동양식으로 인한 에너지는 다른 행동영역으로 옮겨가게 되며 이러한 행동영역에서 한 행동이 일어나 게 되는 것으로 볼 수 있다. 만일 이 생각이 옳다면, 시간 지체에 따른 에너지 소모로 인해 원래의 행동은 전이행동의 발생 이후에 약 간 누그러진 형태로 나타나야만 할 것이다. 그렇지만 이에 대한 지 금까지의 증거는 없다. 오히려 수닭의 싸움행동이 〈 전이쪼기 〉 후에 곧바로 완전한 강도로 다시 나타날 수 있다. 이것은 지금까지 전이 행동에 대한 설명과는 대치된다 . 물론 지금까지 전이행동의 현상을 밝히는 데 수많은 연구의 투자 에도 불구하고 아직도 명쾌한 설명과 학자들 간의 궁극적인 의견의 일치는 없었다 . 오히려 이에 대한 설명에 가까운 〈 탈억제가설 dis i n - hibi t ion h yp o th ese 〉이 널리 인정되고 있는 형편이다. 탈억제가설이 란 두 개의 같은 강도로 활성화된 행동영역의 상호억제로 인해-약 하긴 하지만――억제상황 이전에 있었던 제 3 의 행동 성향이 갑자기 나타나는 현상을 말한다 . 어느 정도 지속적으로 활동적인 행동양식으로 섭식행동과 화장행 동, 번식기 동안에는 둥지짓기 혹은 양육행동이 속해 있다. 그래서 전이행동은 이런 행동영역에서 우선적으로 기인되었다. 말하자면 항 온동물인 경우, 깃털 혹은 털을 깨끗이 유지하는 것은 열을 절연하 는 공기층을 확보하기 위해 매우 중요하다. 그래서 닦기동작, 긁기 동작 그리고 혼들기동작들은 종종 다른 일상적인 행위 사이에 나타 나는 것으로 거의 지속적으로 소환될 수 있거나 누적되어 전이행동 으로도 나타난다. 무엇보다 전이행동의 발생과 빈도가 결코 변화무 쌍한 것이 아니고 이 행동의 기저에 있는 동인의 강도에 따라 달라 진다는 사실이 이 탈억제가설을 뒷받침해 주고 있다. 갈등 상황에 있는 배고픈 수탉온 싸움하는 동안 배부른 수탉보다 더 자주 전이쪼
기를 보이며 이때 종종 씨앗을 쪼아먹기도 한다. 부리가 더러워지는 것은 구애 도중 빈도가 증가된 부리닦기로 전이될 수 있으며, 유럽 되새 Fring illa c oelebs 에게 인위적으로 비를 내리게 하면 갈등상황에 서 증가된 깃털닦기로 발전한다 . 이 탈억제가설은 꽃가루를 모으는 꿀벌들에서 정량적으로 증명될 수 있었다. 꿀벌들은 먹이원을 찾는 동안 닦기동작을 수행하며, 이 들이 먹이장소를 다시 떠나기 바로 직전에는 이 닦기동작이 더욱 강 화된다. 만일 벌의 위가 차츰 채워짐으로 인해 줄어드는 흡입성향과 더듬이의 위치로 알 수 있는 이륙성향(날아가는 성향)을 측정하면, 닦 기동작들은 이 두 길항적인 성향이 평형을 유지할 때 항상 발생하고 있음을 알 수 있다. 확실히 닦기는 흡입성향과 이륙성향의 상호억제 로 인해 지속적인 닦기준비를 탈억제시킴으로써 나타난다 . 이 예들 은 전이행동들이 외부에너지를 통해 공급되는 것이 아니고 전이동작 의 빈도와 강도가 해당 행동을 위해 내재해 있는 동인을 통해 결정 된다는 사실을 보여주고 있다. 물론 전이행동은 두 개의 상반된 행동 성향의 상호억제를 통해 발 현되는 갈등상황에서만 나타나는 것은 아니다. 그것은 오히려 일련 의 다른 갈등영역에서도 잘 알려져 있다. 이런 것에는 목표가 너무 빨리 도달되는 상황 혹은 예측된 응답 자극이 결여되는 상황이 해당 된다. 예를 들어 싸움하는 도중에 적대자가 갑자기 도망가거나 또는 연쇄적으로 일어나는 짝짓기 행동 도중에 파트너가 갑자기 행동의 연쇄를 멈추었을 때도 발생한다 . 첫번째 상황의 예는 두 마리의 가 마우지가 싸움하던 중에 한 마리가 갑자기 도망가면 남아있던 다른 한마리는 성적인 전이행동을 보이며, 두번째 상황의 예는 구애 도중 큰가시고기 암컷이 정상적으로 수컷을 따라 둥지로 들어가지 않으면 수컷의 행동은 둥지짓는 동작으로 전이된다. 언급된 두 경우를 살펴 보면 제각기 하나의 행동성향만이 활성화된다. 그럼에도 불구하고
소속 자극이 갑자기 사라져 버리기 때문에 확실히 진정한 억제가 생 겨난다 이 억제를 통하여 방금 활성되었던 다른 행동양식이 탈억제 가 되고 그대로 실행될 수 있다. 전이행동에 대한 설명으로 탈억제가설말고도 또 있다. 그러나 여 러 전이행동들 역시 다양한 기작에 기인되는 것으로 생각할 뿐이다. 의식화를 통해 전이행동은 2 차적인 신호 기능을 가질 수 있으며 사 회적 의사 소통에도 기여할 수 있다. 특히 짝짓기 행동과 위협행동에서 의식화된 전이행동들이 자주 나타나고 있다 (T i nber g en 1940, Sevenste r 1961 ) . 3 본능모델 이미 앞에서 소개된 바 있는 행동의 기본 현상과 규칙성들을 하나 의 도표로 만들어볼 필요가 있다 . 이 도표는 일목요연하게 행동의 구성을 알아볼 수 있고 어느 정도까지 그것의 설명도 가능하다. 행 동의 구성원리는 본능이라는 개념으로 연결시킬 수있으며, 거기서 개발된 모델을 본능 모델이라고 부른다. 틴버겐은 본능을 다음과 같이 정의내리고 있다. 특정 경계, 해발 그리고 방향성 충격에 내부와 외부로 반응하는 계층적으로 구성된 신경기작이며 이것은 완전히 통합된, 생존유지, 그리고 종족유지의 동작으로 반응하게 한다 . 그래서 가장 상세한 본능 모델이 틴버겐 (1 950) 에 의해 만들어졌다. 이것은 이미 언급한 바 있는 행동의 계층 적 구성관계를 말한다. 이 모델은 1 차 본능(상위의 본능)과 2 차 본능 이 연결되어 계속 하위의 본능으로 이어진다. 이것들은 각기 여러 개의 가능한 부분 행동 혹은 그 다음의 상위 본능의 부분행동 복합 체로 구성되어 있다. 최상위의 본능 중심은 주로 내부 요인, 즉 호
르몬에 의해 영향을 받으며, 역시 자발적으로 활동한다. 반면 나머 지 본능 중심들은 외부자극뿐만 아니라 내부자극에 의해서도 영향을 받는다. 상위의 본능 중심에서 그 다음 하위의 중심으로의 에너지 유입은 보통 하나의 블록으로 차단되어 있으며, 이것은 우리가 이미 언급한 바 있는 해발기구가 되며 해당 자극(신호자극 혹은 해발인)에 의해 이 블록이 제거될 수 있다. 각각 지배하고 있는 이 자극 상황 은 다음 하위의 어떤 부분행동들을 소환하는지를 결정한다 . 만일 적 합자극이 없을 경우에 이 볼록은 그대로 머물러 관련 행동양식들이 차단되며, 충격은 해당 단계의 갈망행동을 조절하는 본능 중심으로 흐른다 . 그런 다음 이것은 관련 의부자극을 통해 그 단계의 볼록이 제거되어 그 다음 하위 중심이 홍분될 때까지 지속된다. 상호억제로 인해 순간마다 단 하나의 부분행동만이 수행될 수 있다. 그러므로 동시에 여러 개의 본능 중심들이 상위 중심으로부터 흥분된다면, 다른 영향을 통해 부수적으로 가장 강하게 활성화된 어떤 한 충격만이 전 달된다. 본능 중심의 가장 하위의 수준은 각각의 행동영역의 완료행 · 동에 해당된다. 이 단계 밑에는 물고기 지느러미의 움직임에 대해 이미 말한 바와 같이 상호억제 대신에 각각의 하위 단위들 사이에 통합이 존재한다. 지금까지의 본능 모델을 실제 큰가시고기의 생식 행동을 가지고 설명할 수 있다(그림 4-3). 호르몬인 테스토스테론은 수온의 상승과 함께 최고의 본능 중심에 영향을 미친다. 이 두 요인은 큰가시고기 로 하여금 바닷물(혹은 깊은 민물)에서 수심이 낮은 민물로 이동하게 한댜 〈이동중심〉이라고 하는 이 최상위의 본능 중심은 블록을 갖고 있지 않댜 특별히 신호자극이 필요치 않은 경우에 이 이동은 어느 정도 계속 유지되는데, 이것이 진정한 갈망행동이 된다. 이 행동은 텃세 확보를 위해 적합한 소생활권에 보내지는 신호자극들(수심이 낮 고 따뜻한 민물, 식물이 우거진 곳)이 최초의 생득적 해발기구에 영향
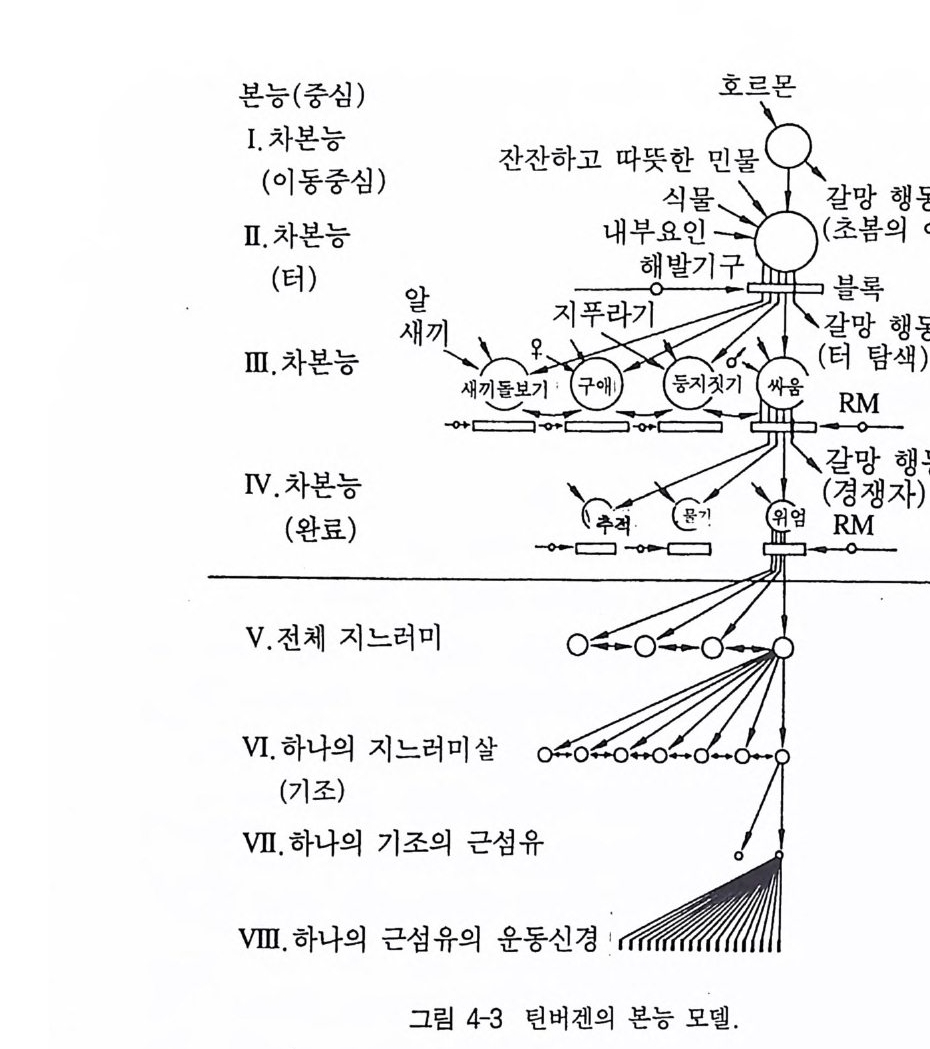 본능(중심) 호르몬
본능(중심) 호르몬
을 미쳐 생식행동의 최고 중심의 블록을 제거하게 된다. 이것을 〈텃 세중심〉이라고 한댜 이때 큰가시고기는 다음 하위의 중심들(싸움중 심, 둥지짓기중심, 짝짓기중심, 부채질중심)이 블록에 의해 차단되어 있 기 때문에 이 중심들이 해당 신호자극들과 만나지 못하고 있는 한, 갈 망행동으로 혼자 있게 된다. 즉 큰가시고기는 싸움을 할 경쟁자나 짝짓기할 암컷이나 둥지를 지울 재료를 발견할 때까지 주변을 이러 저리 돌아다닌다. 예를 들어 터안에서 한 마리의 침입자(수컷)가 싸
움행동을 유발시키게 되면, 터주인은 침입자를 향해 돌진한다(갈망 행동) . 이때 이 침입자는 새롭고 특이한 신호자극을 보내야만 한다. 이 신호자극은 위협(과시), 물기, 추적 등의 완료본능들 가운데 하 나의 본능에 대한 블록을 제거하게 된다. 이로써 이 흥분은 마지막 본능중심으로 전달된다. 틴버겐의 본능 모델과 비교할 수 있는 모델이 베이렌드 Baerends (그림 4-4, 1956) 에 의해 제시되었다 . 이것도 마찬가지로 행동의 계 층구조로 중심의 상호억제가 같은 수준으로 표현되었다. 그러나 하 위구조의 중심들이 종종 여러 개의 상위구조의 중심에 종속되어 있 는 것이 큰 차이를 보여주고 있다. 각 행동양식 가운데 가장 먼저 많 은 기본적 동작 양식이 여러 행동영역 내에 나타날 수 있다. 이것은 주로 도피, 공격, 구애, 혹은 사냥행위의 구성분이 되는 이동 행동양 식이나 다소 원거리 장소 이동(예, 철새의 번식지와 겨울 서식지 간의 이 동)의 목적만을 위한 이동 행동양식에 적용된다. 그러나 파기동작, 쪼 기동작, 물기동작 혹은 덥석물기동작 역시 매우 다양한 행동영역에서 나타날 수 있다. 이런 상호관계는 각기 단 하나의 본능을 하위의 여 러 본능들과 함께 제시하고 있는 틴버겐의 모델과는 차이가 있다.
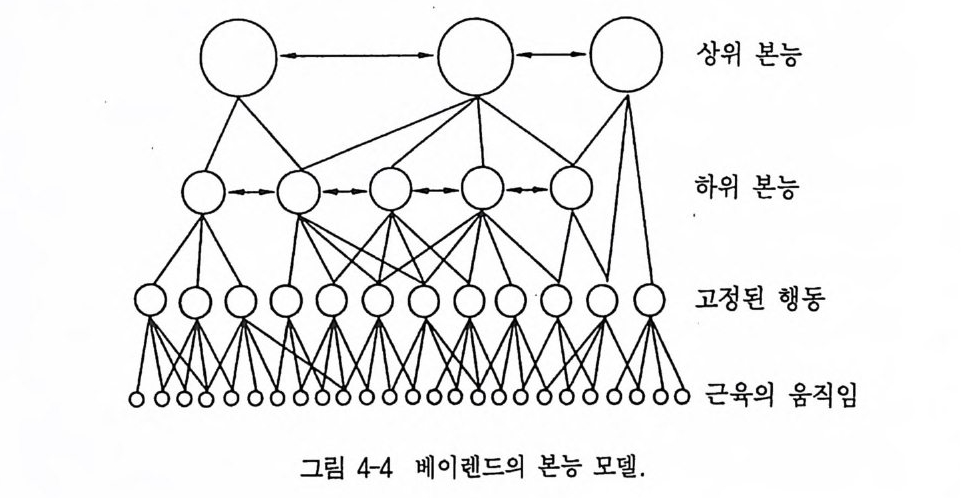 상위 본능
상위 본능
제 5 장 행동생리 지금까지의 살펴본 행동들의 시간적인 순서와 계층적인 구성은 각 각의 행동구성 요소들이 의미 있게 구성되어 작용하는 통합기작에 의해 일어나고 있음을 전제하고 있다. 여기에 근본적으로 신경계와 호르몬의 두 조절기작이 관여하게 된다 . 두 조절기작은 동물의 조절 효과에서 그리고 동물과 환경 사이에서 어느 정도 일치하고 있으 나, 성격면에서 근본적인 차이를 살펴볼 수 있다. 호르몬은 비교적 천천히 효과를 나타내지만, 오래 지속적으로 영향을 미칠 수 있다. 이에 반해 신경계는 매우 빨리 작용을 하며 보통 급속히 짧은 기간 에 조절과정에 관여한다. 두 조절계에 대한 연구는 행동 기저에 놓 여 있는 생리학적 과정을 좀더 접근하여 분석할 수 있게 한다 . l 신경계와 행동 행동의 신경학적 기초, 죽 동물의 행동과 그 동물의 신경계 기능 간의 협력은 이미 오래전부터 관심의 대상이었다. 이에 대한 연구는
인간 행동의 조절에 대한 귀납적 추리를 시도하고자 하는 데서 시작 되었다. 이런 관점에서 인간의 행동을 조절하는 신경계 요소의 정확 한 위치에 대한 의문이 제기되었다. 여기서 국지적 가설과 가소성 가설의 두 가지 주장이 생겨났다 . 국지적 가설은 특정 뇌 일부에 손 상을 입었을 경우에 일어나는 현상으로 모든 행동 형질들은 뇌에 국 지적 위치를 갖는다는 것이다. 이에 비해 가소성 가설은 특정 조절 기능이 뇌의 특정 지역에 엄격하게 배열되어 있지 않고 중추신경계 내에서 구성 요소가 균일하게 분포하고 있으며 더욱이 명확한 구조 적 경계가 없다는 것이다. 이 두 가설간의 논쟁은 결국 신경행동학 적 연구에 의해 뇌 기능의 국지적 가설이 사실로 입증되었다. 그래 서 행동의 신경행동학적 방법을 다음과 같이 제시할 수 있다 . 1.1 절단 특정한 행동 기능을 조절하는 부위를 알아내기 위해 뇌의 일부를 차단하는 것이다 . 이것은 해당 단면을 외과적 수술, 뇌의 신경연결 의 절단 혹은 뇌 조직의 파괴(예, 고주파나 레이저 광선을 이용)를 통 해 해결할 수 있다. 최근에는 뇌 조직을 부분적으로 얼리는 방법도 개발되었다 . 물론 이렇게 물리적인 절단 방법에는 한계가 있다. 한 가지 이유는 보통 더 큰 뇌 부위가 절단되기가 쉽다. 그러나 서로 밀집되어 있는 뇌 구조가 아주 다양한 기능을 소유할 수 있다는 사 실을 다른 연구방법으로 알 수 있다. 또 한 가지 이유로는 모든 의 과적 뇌수술이 동물체의 심각한 영향을 초래할 수 있다. 그래서 수 술후 행동변화의 관찰이 무조건 뇌 조직 절단의 결과라 생각해서는 안되며 일반적인 상해로도 돌릴 수 있다. 뇌 조직의 최초의 제거는 이미 150 년 전에 이루어졌다 . 이로써 절 단방법은 가장 오래된 신경행동학적 방법이라 할 수 있다 . 그러나
위에 지적한 단점으로 인해 이 절단 방법은 그동안 매우 등한시했 다 근래에 와서야 지금까지의 단점을 보완하는 개선된 방법을 통해 이 방법에 대한 의미가 재평가되었으며, 매우 유익한 결과를 가져오 게 되었다. l. 2 뇌자극 특히 세련된 방법은 뇌에 전기적 자극을 주는 것이다. 이 방법은 비교적 오래된 것으로 이 방법이 처음 실시된 것은 대략 100 년전으 로 거슬러올라간다. 이 방법의 실험이 행동생리학 분야에 널리 사용 되면서 헤스 Hess( 그림 5-1, 1954, 1957) 의 고양이 간뇌의 실험과 홀스
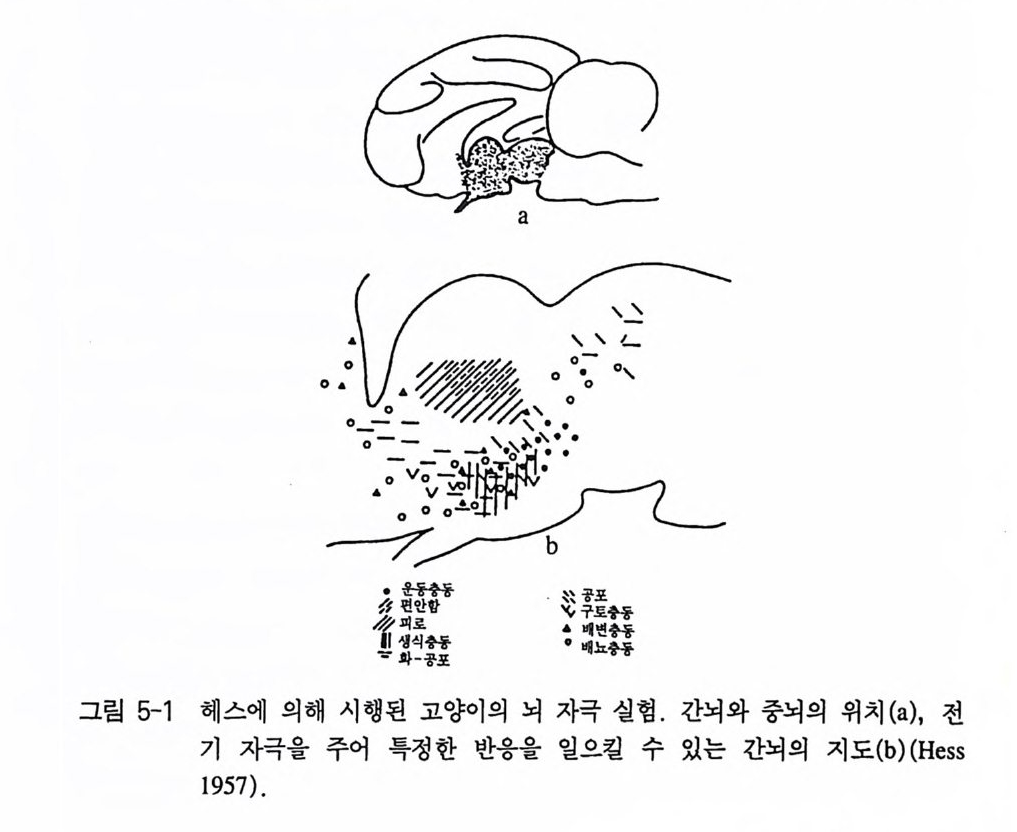 \ \ `느
\ \ `느
트 Hols t와 폴 Paul(l960) 의 닭의 뇌간 실험으로 뇌와 행동과의 관계에 대한 많은 의문이 해결되었다. 오늘날 수많은 척추동물들, 특히 조 류와 포유류 그리고 여러 무척추동물, 무엇보다 곤충과 가재의 뇌 자극 실험의 결과가 나와 있다. 이 뇌 자극을 주기 위해 맨 위쪽에 래커를 칠해 절연시킨 미세한 전극, 대개가 강선( 鋼線 ) 혹은 텅스텐선을 사용한다. 이런 식으로 주 입된 전류는 뇌의 작은 지점, 극히 제한된 부위에 다다를 수 있다. 이 전극은 플라스틱이나 합성수지로 된 전극관에 넣어 나사형 추진 장치를 이용해 천천히 뇌속으로 밀어 넣을 수 있다. 이것을 위해 맨 먼저 미세한 드릴을 이용해 두개골에 작은 구멍을 낸 다음 이 구멍 에 전극관을 꽂는다. 이 전극관은 두개골에 그대로 오랫동안 고정시 켜 놓을 수 있다. 반면 전극관 위에서 여러 개를 뇌안으로 집어 넣 울 수 있게 되어 있는 전극들은 실험할 때마다 삽입된다. 하나의 커 다란 표면 전극은 영점( 零 點)전극으로 두피 아래의 두개골에 고정시 킨다 전극관을 통해 전극들을 천천히 밀어넣음으로써 커다란 뇌 부 위가 접촉될 수 있게 한다. 전극 끝의 정확한 위치탐색은 X- 선이나 혹은 일시적으로 강한 전류을 흐르게 하여 정확한 정보를 얻을 수 있다. 자극발생기로부터 연결된 선은 보통 여유있게 하여 동물이 자 유롭게 움직이게 해야 한다. 하지만 요즘은 무선을 이용해 자극을 줄 수 있는 방법이 개발되어 있기 때문에 그리 큰 문제는 발생하지 않는다. 뇌에 자극을 주려면 표면 전극과 뇌안에 장치한 전극 사이에 교류 전류를 가하거나 뇌안에 깊이 넣은 인접한 두 전극에 교류를 가한 댜 이때 필요한 전류의 크기는 매우 작으며 단지 밀리암페어 정도 의 순간 전류을 가한다. 자극의 크기를 변화시키기 위해 전류의 크 기 혹은 보통 이런 실험에서 0-2 볼트에 해당하는 전압을 변화시킨 댜 일반적으로 자극의 크기에 대한 측정은 전류의 크기로 판단한다.
l.3 활동전위의 유도 이 방법은 전기 화학적 과정의 결과로 인해 활동중인 신경세포가 몸에 정보전달을 할 때 전기적 신호를 생산한다는 사실을 이용하고 있다 . 이 전위는 가장 가는 전극을 이용하여 유도할 수 있으며 또 관련 증폭정도에 따라 측정할 수 있다. 이런 식으로 어느 신경세포 가 어느 정도 활동하는 동안 어떤 식으로 활동적인지 알아낼 수 있 으며, 이것을 통해 뇌신경들의 배열과 작용에 대한 정보를 얻을 수 있다. 물론 실제로 신경 하나 하나의 유도 분리가 쉽지 않다 . 그럼 에도 불구하고 보통 적당한 필터를 통해 각각의 신경들의 반응을 서 로 분리시킬 수 있다 . l.4 방법적 전제조건 모든 신경행동학적 연구에 대한 중요한 전제조건은 적당한 실험동 물을 선택하는것이다. 뇌의 조절기능에 대한 실제 궁극적인 결과는 해당 동물종의 행동이 가능한 한 자연상태에서 많이 알려졌을 때 얻 어질 수 있다 . 그 이유는 이런 정보만이 관찰된 행동을 적절히 의미 를 부여할 수 있기 때문이다. 이때 이 행동이 자연적인가, 죽 이 행 동의 진행이 정상적인 조건에서 일어나는 행동과 일치하는지가 중요 하다. 이러한 면에서 인위적, 즉 뇌 자극을 통해 일어난 행동이 그 실험동물의 동종과 정확하게 일치할 때, 다시 말해 그 종들이 보통 때에도 그런 반응을 보일 때, 보다 좋은 결과를 얻을 수 있다. 실험동물의 뇌에 대한 해부학도 충분히 잘 알려져 있어야 한다. 뇌의 구조와 관련해서 신경행동 연구는 행동양식이 다양하고 변화가 많은 고도로 발달된 척추동물들에서 실험적인 분석의 접근이 어려운 복잡한 뇌의 구조를 갖고 있기 때문에 어느 정도 문제가 발생할 수
있다. 그런 이유에서 많은 경우에 곤충과 다른 무척추동물을 적합한 실험동물로 이용하고 있다. 이들의 중추계는 신경세포의 수가 적기 때문에 많은 방법적 이점을 지니고 있다. 2 결과 2.l 뇌 기능의 위치 실험방법의 선택과는 무관하게 모든 신경행동학적 연구는 한 동물 의 특정 행동 활동성과 그의 중추신경계의 특정 부분 간에 분명한 규칙적인 배열이 성립하며, 즉 뇌의 특정 부위가 각각 특정 행동양 식을 책임지고 있다는 사실을 입증해 주고 있다. 만일 해당 뇌 부위 를 파괴시키면 그 뇌 부위가 맡고 있는 행동양식이 없어져 버린다. 물론 이 배열은 파괴된 뇌 부위의 기능을 첨차 다른 부위가 떠맡을 수 있기 때문에 항상 불변하는 것은 아니다. 뇌 기능의 위치는 무엇보다 절단 실험을 통해 입증되어 왔다. 그 다음에 공간적인 배열은 개별적으로 조사하여 그것을 조합하는 식으 로 이루어졌다. 많은 동물종들은 이렇게 해서 이미 소위 입체적 뇌 지도를 만들 수 있을 정도로 잘 연구되었다. 이 입체의 단면 그림은 대개 기능에 따라 뇌 구역에 이름이 붙여졌다 . 물론 일련의 연구는 이러한 자료를 조심스럽게 해석해야 한다는 사실을 분명히 하고 있 댜 그래서 몇몇 경우에 외상에 의해 없어진 기능이 제 2 의 의상을 통해 다른 뇌 부위에서 다시 복구될 수 있었던 사실이 확인되었다. 분명히 이런 경우에 관련 뇌 부위는 이 기능에 대해 본질적인 일을 맡지 않고 있다는 사실을 보여주는 예가 된다. 그러나 그것은 가끔 특정한 종에서만 국한해서 일어나고 있다 (Nau 와 Deli us 1981 ).
뇌 지도와 뇌에 있는 공간적 기능 분포에 대한 정보는 뇌 자극의 정밀한 실험을 위해 중요한 전제조건이 된다. 이것은 처음부터 목표 지점에 전극을 꽂는 것을 가능하게 할 수 있다. 이 뇌 자극 실험은 절단 실험 결과를 보다 심화시켜 왔으며 보완시켰다. 이것은 중추신 경계의 특정 부위에 전기적 자극을 주어 특정 행동양식을 인위적으 로 불러올 수 있음을 보여주고 있다. 그것은 조류나 곤충의 소리내 기와 같은 동작과 관련되어 있다. 그 동안 많은 동물종들에서 뇌자 극을 통해 거의 모든 행동양식들을 인위적으로 소환시킬 수 있었다. 이런 식으로 소환된 행동요소들의 복잡 정도 차이는 아주 심하다. 이 복잡정도는 아주 간단한 개별 동작(예, 털세우기, 다리의 움직임)에 서부터 비교적 복잡한 행동양식의 연쇄(예, 도피행동, 싸움행동 혹은 사냥행동)까지 해당된다. 행동을 조절하는 뇌 중추의 위치와 관련해서 주로 전기자극을 이 용해 이 조절기능이 모자이크 형태로 분포되어 있다는 사실이 알려 지게 되었다. 뇌의 조절구역은 공간적으로 나뉘어 있으며, 단지 0. 1mm 간격을 두고 서로 분리된 두 전극 끝은 아주 다양한 행동양식 들을 불러낼 수 있으며, 또 하나의 행동이 종종 두 군데 이상의 뇌 부위에 의해 소환될 수 있다. 이 중복된 소환은 실제로 각각의 고유 중추신경들이 관련 동작들의 연결로 이루어진 해당 행동양식에 책임 을 지고 있는지, 이 중추신경들이 각각 중복해서 행동을 하게 하는 지 의문으로 남는다. 실제로 이 두 가능성을 지지하는 많은 자료들 이 보고되었다. 많은 조류의 경우 소리내기와 이 소리내기와 관계된 행동양식들에 대한 관련 뇌 부위의 〈명령체계〉가 발견되었다. 이것 온 전뇌에서 중뇌에까지 퍼져 있으며, 관련 반응들이 전기자극을 통 해 임의의 위치에서 활성화되었다. 귀뚜라미는 다양한 노래 형태(유 인음, 공격음)를 가슴신경절뿐만 아니라 뇌에 전기자극을 가해 소환 시킬 수 있다(그림 5-2). 실험결과에 의하면 이 가슴신경절은 노래
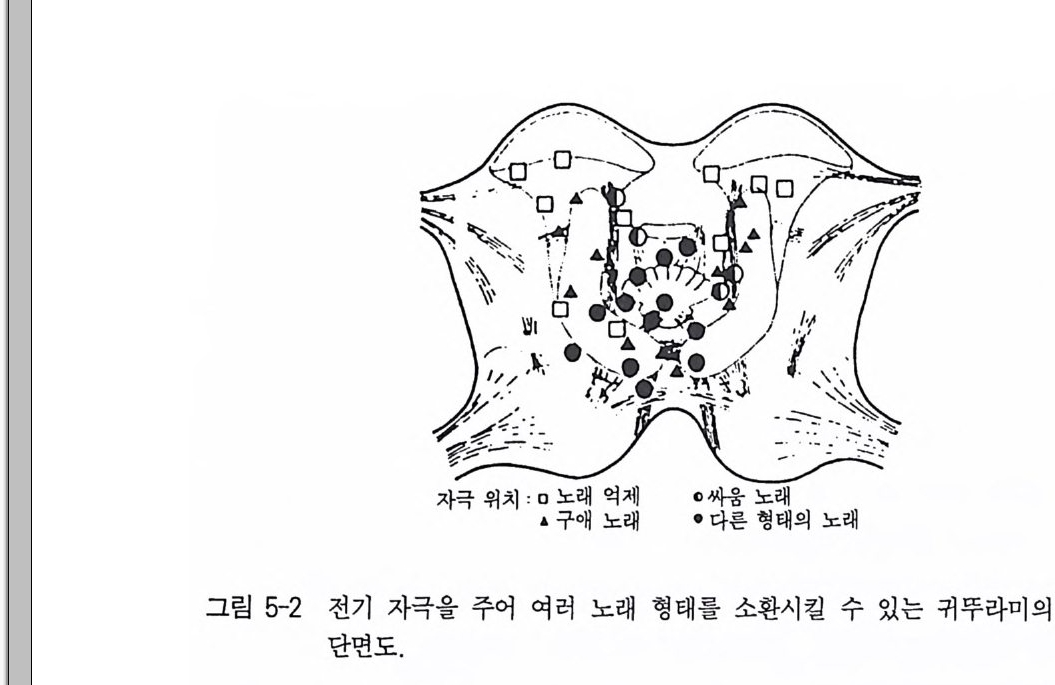 자극 위치 : 0 노래 억제 0 싸움 노래
자극 위치 : 0 노래 억제 0 싸움 노래
형태를 조절하는 데 상당히 독자성을 소유하고 있는 반면, 뇌는 명 령을 내려 단지 소리를 낼 것인지 혹은 어떤 노래유형을 낼 것인지 결정한다 . 귀뚜라미의 유도실험을 통해 명령체계를 한층 더 발전시 킬 수 있다. 이것은 근육에 나타난 활동전위가 중추신경에 나타난 충격과 일 대 일의 비로 매우 정확히 같았음을 보여주고 있다.
뇌에서의 여러 부위에 대한 표현은 무엇보다 같은 동작이 여러 행 동영역에서 나타나 결국 다양한 동인들이 있을 수 있는 경우에 추측 해 볼 수 있다. 닭에서 앉아있기는 어떤 경우에는 잠자기 동인과 포 란 동인 또 다른 경우에는 순수하게 앉아있기가 있을 수 있다. 선 채로 몸을 낮추는 자세는 서열이 높은 경쟁자에 대해 공중적개(空中 敵愼)행동* 혹은 복종행동에 속할 수 있다. 외양적으로 같은 행동요 소의 이런 다양한 기능과 배열은 뇌 자극 실험에 나타난 것처럼 뇌 * 공중적개행동 : 조류에서 공중에 적이 나타나면 몸을 움추려 자신의 몸을 감추 고자 하는 행동에서 유래된 말 .
에서의 다양한 자극영역 위에 있는 활 동과도 일치하고 있다. 2, 2 전기적 뇌 자극에 대한 행동의 반응 중추신경계의 자극을 통해 소환될 수 있는 행동양식들은 의부자극 의 추가적 요구와 관련해서 두 행동군으로 나눌 수 있다. 외부자극 이 없이 전기자극만을 통해서 유발되는 행동이 있고 , 외부자극에 의 해 행동의 발생과 진행이 뇌 자극을 통해 영향을 받는 행동이 있다. 첫번째로 언급한 행동에는 닭의 도피행동이나 귀뚜라미의 노랫소리 가 해당된다. 이 두 행동은 관련 외부자극(공중의 적 혹은 육상의 적 , 경 쟁자, 자기종의 암컷)이 없이도 뇌 자극 후에 수행된다 . 두번째 행동 군은 다시 둘로 나눌 수 있다 . 첫째, 많은 경우에 뇌 자극은 밖에서 알아볼 수 있는 결과로 나타나지 않는다. 그럼에도 불구하고 동물에 게 이전에 반응을 보이지 않았던 자극울 주면, 뇌 자극은 영향을 미 친다. 즉 뇌 자극이 있은 다음에 외부자극에 의해 반응을 보이는 것 이다. 수 닭 은 뇌간의 해당 위치에 자극을 준 후, 전에는 완전히 관 심이 없었던 박제로 만든 수탉을 갑자기 경쟁자로 생각하고 공격을 한다. 이 경우에 뇌 자극을 통해서 직접적으로 행동이 소환되지는 않았지만, 그 뇌 자극은 그 동물의 기분을 바꾸어 놓아 관련 행동이 해당 외부자극을 통해 더 쉽게 유발하게 했다. 여기서 뇌 자극은 효 과면에서 〈 동인성 신호자극〉에 해당된다. 둘째, 첫째의 경우와 차이 가 있는 것은 뇌 자극을 통해 돌아다니거나 혹은 돌아다니면서 찾는 행동이 유발되는데, 이것은 관찰될 수 있지만, 확실히 목표 없는 행 동이다. 종종 이 행동은 보통 동작의 동요로 별 의미가 없었던 것이 었다. 말하자면 이 동물에게 이제 해당 의부자극(예, 먹이 , 배우자 혹 은 경쟁자)을 제공하면, 관련 적합 행동양식이 나타난다 . 여러 개의 다양한 대상 물체를 이용한 선택실험은 그 대상물체들 가운데 하나
에만(예, 경쟁자에만) 각각 반응이 일어남을 보여주고 있다. 명백히 뇌 자극을 통해 〈해발성 상황을 향한 탐색〉, 죽 진정한 갈망행동이 소환될 수 있다. 2.3 뇌 자극과 행동학적 기본 개념 뇌에서 전기적 자극은 행동학적 기본 개념들 가운데 지금까지 관 찰과 순수한 행동실험으로만 정의되었던 몇몇 개념에 새로운 전기를 마련해 주었다. 이 뇌자극에 대한 일반적인 증명은 홀스트와 폴 (1 960) 의 닭 실험에서 가장 잘 나타나 있다. 이들의 실험은 이미 오 래된 일이었으나, 신경생리학과 행동학 사이에 아주 긴밀한 관계가 있음을 입증해준 첫 시도였다. 그 밖에도 이 실험은 서술에 불과했 던 지금까지의 개념들에 대해 근본적인 현상으로 부각되면서 많은 의문들이 입증되는 계기가 되었다. 그러나 그 동안 새로운 연구를 통해 이에 대한 일반 현상들이 좀더 세분화되었고 복잡해졌다. 1) 동인의 측정 한 동물의 기분상태는 행동실험, 예를 들어 모형을 갖고 알아 볼 수 있다 그러나 그것은 전기적 뇌 자극을 통해서도 알 수 있다. 정 량적 방법으로 자극발생기에는 전류의 세기와 전압을 사람이 조작해 서 마음대로 바꿀 수 있어야 한다. 그리고 실험동물에는 뇌 자극을 통해 유발된 반응의 강도, 잠복기 (전기 자극을 가한 시간과 반응의 발 생사이의 시간), 그리고 반응지속기, 무엇보다 소위 과지속기(전기자 극을 멈춘 시간과 반응이 사라진 시간 사이의 길이)가 관찰되야 한다(그 림 5-3). 이 세 척도는 전기자극의 크기에 따라 달라질 뿐만 아니라 (자극이 강하면 반응이 커지고, 잠복기는 짧아지고 그리고 반응지속기는 길 어진다) 그 동물의 동인 상태에 따라서도 달라진다. 어떤 동물이 이
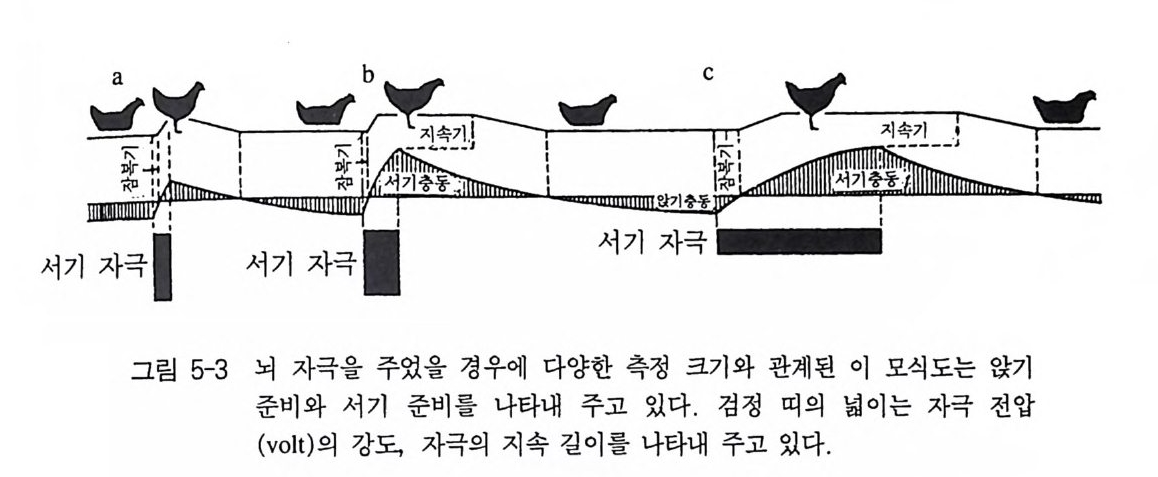 C
C
미 특정한 방향으로 더 강하게 동기부여가 되면 될수록 관련 반응이 유발되기 위해서는 더 작은 자극이 필요하고, 말하자면 같은 크기의 자극에도 더 강하게 반응을 보인다. 이런 실험을 통해 다음의 결과가 밝혀졌다. 외관적으로 같은 행동 도 내부적으로는 매우 다른 강도의 동인이 있을 수 있다. 예를 들어 앉아 있는 닭은_대략 잠의 동인 혹은 포란 동인――매우 단단히 앉아 있을 수 있거나, 혹은 간신히 앉아 있을 수 있는데, 이것은 일 어서기 바로 직전에 발생한 다른 동인에 기초해서 존재한다 . 여기서 그 동물이 앉고자 하는 기분을 전압의 변화에 따라 볼트로 측정할 수 있다. 앉고자 하는 동인이 강하면 강할수록 실제 일어나게 하기 위해서는 더 강한 자극이 필요하다 . 마치 이것은 감각 자극의 발생 효과를 실험할 때와 같이 여기서도 주어진 시점에 특정 행동양식을 유발하는 이때의 자극 크기에 대해 발생역치 혹은 역치자극전류라고 하며, 이때 이 역치 이하의 자극을 역치하 자극이라고 부른다. 그래 서 이미 언급된 결과를 이런 식으로도 말을 바꾸어 보면, 특정 반응 (여기서는 일어서기)을 유발시키는 최소 자극전류는 외관적으로 아주 같은 상황(여기서는 앉아 있는 닭)에서도 완전히 차이가 날 수 있다는 식으로표현할수 있다.
 드色2: 3 ,4〈 뇨5
드色2: 3 ,4〈 뇨5
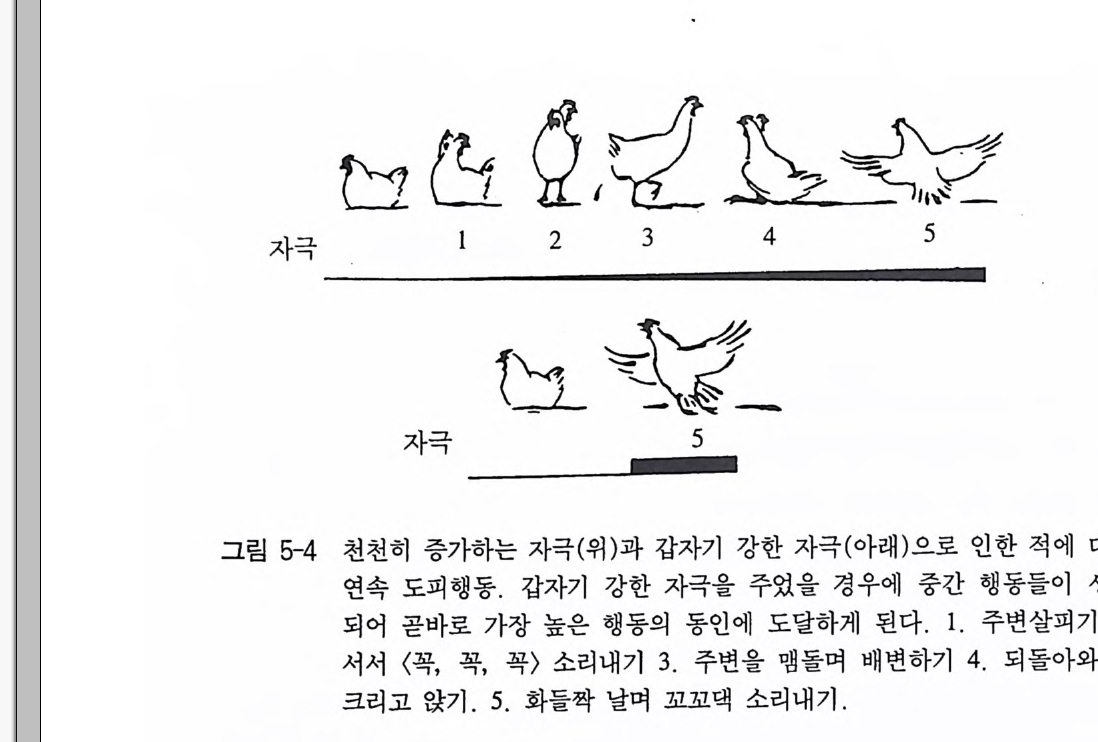
한 행동의 연속에서 개별적인 부분동작들은 하나의 동일 시점에도 여러 발생 역치를 갖는다. 만일 뇌간의 특정 부위에 자극을 주면, 약 한 자극의 경우에 강한 자극보다 다른 부분동작들이 나타난다. 닭의 정상적인 구역질반응의 경우에 부리혼들기, 혀움직이기 그리고 숨가 빠지기로 이어진다. 예를 들어 비슷한 결과가 육상의 적에 대한 도 피행동과 같은 연속동작에서도 나타나 있다(그림 5-4). 행동양식들은 짧은 간격을 두고 반복된 뇌 자극을 통해 피로해질 수 있댜 즉 이 행동양식들은 점차 약해지면서, 이들이 마침내 더 이상 반응을 보이지 않을 때까지, 좀더 긴 잠복기와 짧은 반응지속 기를 보인다(그림 5-5). 일정한 자극 휴식이 있은 다음에는 (혹은 전 보다 더 강한 자극을 줌으로써) 이 행동들은 다시 유발될 수 있다. 이런 반응의 고갈은 물론 개별적 행동영역과 동작 하나 하나에서 아주 다양하게 나타난다 . 이것에 대해 이미 행동양식의 특이적 피로
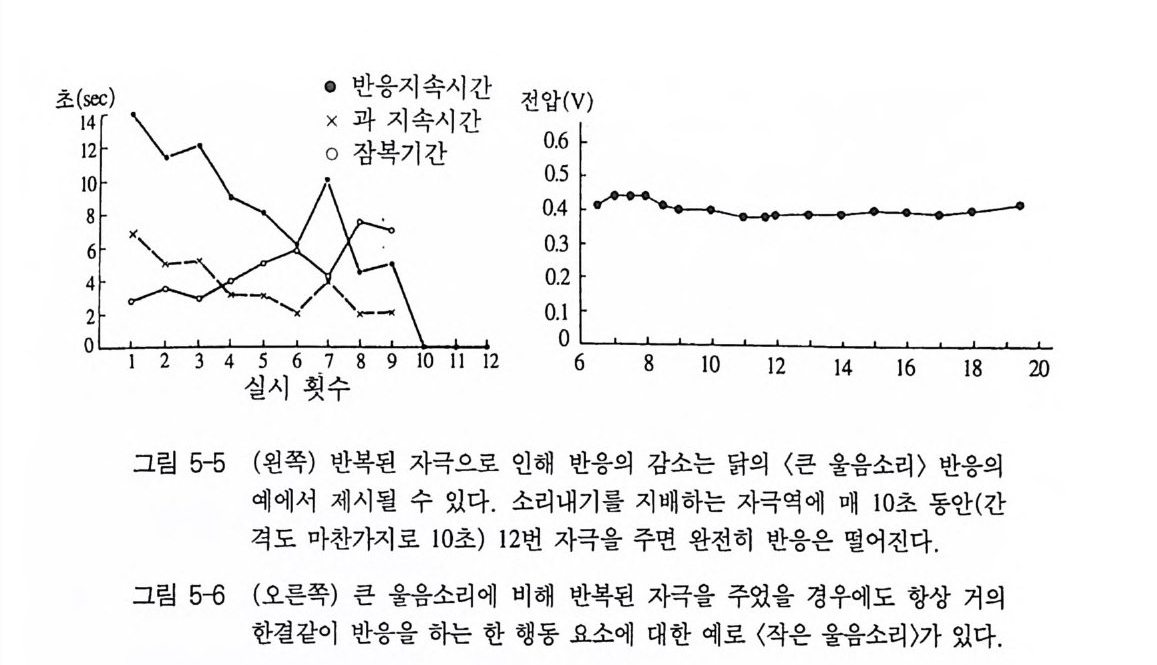 초g間 \ - •0x 잠반과응 복지지기속 간속시시 간 간 전압°.6 ( V )
초g間 \ - •0x 잠반과응 복지지기속 간속시시 간 간 전압°.6 ( V )
현상에서 언급된 바 있다. 여러 동인 체계에 소속 가능한 가장 작은 행동단위, 즉 간단한 동작 형태는 가장 천천히 피로해진다. 이런 행 동단위들은 고갈되지 않고 좀더 오랜 시간 수행될 수 있다(그림 5-6). 2) 행동양식들의 협력 특히 홍미 있는 결과는 동시에 뇌간의 두 지점에 자극을 가한 뇌 자극실험에서 나타났다 . 이때 두 자극장은 동일 행동 혹은 두개의 다른 행동양식을 불러일으킬 수 있다. 여러 행동양식을 담당하는 구 역에 자극을 주었을 경우 다음 두 가지 가능성이 발생한다. 이 두 자극들은 동일한 혹은 서로 차이가 나는 세기를 가질 수 있다. 첫번 째 경우에서 예를 들면, 먹이먹기와 확인 행동의 두 반응이 재빨리 바뀌면서 교대로 나타나거나(소위 진자행동), 혹은 두 개의 소속 행 동양식이 관찰되지 않고, 완전히 새로운 제 3 의 행동이 나타난다. 만일 한 곳을 먼저 강하게 자극하고 다른 한 곳을 약하게 자극하 면, 일반적으로 처음의 자극장에 속한 반응만이 나타난다. 이에 비
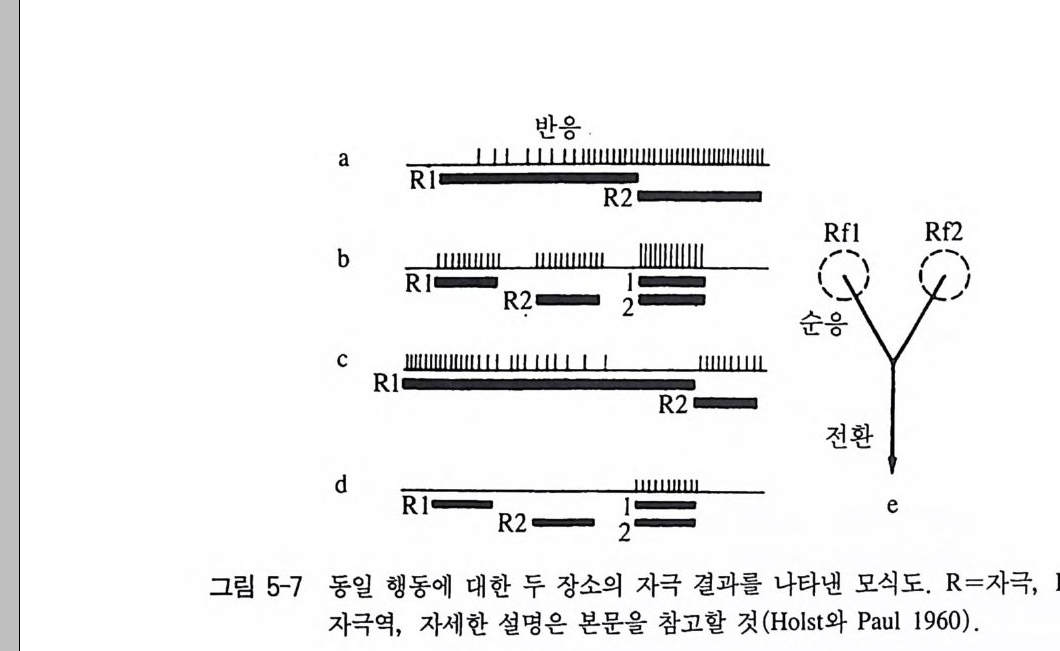 ba l1l1111Il1 1—1I I l 바I|`I I-|I 으II0Ill I lIl 1II1ll1R l12l1—I |IIIl1|I||l|1II||l|I III IIIIIIIIIIII R f l Rf 2
ba l1l1111Il1 1—1I I l 바I|`I I-|I 으II0Ill I lIl 1II1ll1R l12l1—I |IIIl1|I||l|1II||l|I III IIIIIIIIIIII R f l Rf 2
해 약한 자극장의 반응은 억제된다. 이것은 여러 행동양식들을 맡고 있는 신경요소들 간에 억제 교차가 있음을 입증해 주고 있다. 〈재폭 발〉이라고 불리는 홍미 있는 현상온 두 자극장에서 자극 강도의 차 이가 매우 작았을 경우에 일어날 수 있다. 죽 이 경우에 처음에는 강한 자극장에 속한 반응이 일어나지만 자극이 끝난 후에는 갑자기 지금까지 억제되었던 행동양식이 불쑥 튀어나와 진행될 수 있다. 예 를 들어, 〈꼬끼오〉라고 하는 수탉의 울음소리는 관련 자극장의 위에 서 활성화되지만, 동시에 〈앉기〉를 담당하고 있는 억제된 자극장이 좀더 강한 자극으로 인해 갑자기 이 울음소리가 아주 인상적으로 나 타난다. 동일 행동양식을 지배하는 두 뇌 부위를 자극했을 경우, 자극 통 합의 법칙에 일치하는 현상을 관찰할 수 있다. 두 지점을 따로따로 중간 정도의 자극 강도로 자극을 주면, 반응의 강도도 중간 크기로 나타나며(그림 5-7a), 이에 비해 두 지점을 동시에 자극하면, 반응 강도는 분명히 상승한다(그림 5-7b). 이 자극 효과는 역시 부가될 수 있댜 즉 두 자극장으로부터의 홍분이 어디에선가 합쳐서 흐르게
된댜 이런 식으로 따로따로 자극을 주었을 경우에 이 자극이 각각 역치 이하의 크기라 할지라도 이 자극역치를 넘어서게 되어 관련 반 응이 소환될 수 있다(그림 5-7d). 만일 한 곳의 반복된 자극을 통해 피로해진 반응에 이어서 다른 지점에 자극을 주면, 이 반응은 완전 히 다시 일어난다(그림 5_7c). 이것은 행동양식의 특이적 피로현상 이 운동신경의 피로와 관계가 없음을 보여주는 것이다. 오히려 행동 유발은 그 결과가 다른 자극장에도 관련되어 있기 때문에 이미 홍분 이 통합 유출되기 전에 두 자극장으로부터 이루어져야 한다. 따라서 이것을 〈중추적 위치 순응〉이라고 한다(그림 5-7e). 2.4 행동양식의 중추적 프로그램 중추신경계에서의 활동전류의 유도는 전기적 뇌 자극보다 좀더 세 밀하게 연구를 수행할 수 있는 장점이 있다. 그러나 이것은 뇌 신경 세포의 하나 하나를 분리 실시해야 하는 실험상의 어려움도 따른다. 특히 척추동물의 경우 방법상의 어려운 점이 많다. 그럼에도 불구하 고 이들에 대한 몇몇 연구가 지금까지 보고되었다. 쥐의 중뇌와 간 뇌의 시상하부 지역에서 각각의 뉴런들의 기능이 밝혀졌다. 이 뉴런 들은 제각기 특정한 일상적인 활동에서 예를 들면 섭식행동, 냄새맡 기, 혹은 콧수염 움직일 때에 높아진 충격빈도(활동전위)를 보여주고 있댜 물론 관련 행동이나 동작이 멈추면 다시 이 활동전위는 내려 간다 이와 비슷한 활동전위의 상승은 고양이의 간뇌에 있는 특정 신경세포에서 발견되었다. 수컷의 경쟁자가 나타나면 공격유발 자극 에 대한 반응이 일어나는데 이때의 신경세포의 활동전위가 높아진 다. 몇몇 어류에서 예를 들어, 양놀래기과 Lab ri dae 의 어류에서 종특 이적 행동, 예를 들면 먹이동물을 향해 헤엄쳐 오거나, 그 먹이동물 울 씹고 그리고 삼키는 동안 뉴런들이 각각 특이적 활동전위를 보여
주었다. 광범위한 활동전류의 유도 실험은 무척추동물의 중추신경계에서 이루어졌다 특히 이런 실험에는 중추신경계에 비교적 적은 세포수 룰 갖고 있는 가재와 곤충이 적합하다. 그리고 이 동물들의 행동양 식이 일정하게 유지되는 형태를 띠고 있기 때문에 비교적 정량적으 로 측정이 가능한 장점을 갖고 있다. 곤충의 경우 개별적 뉴런의 충 격을 특정 동작과 연결시킬 수 있을 뿐만 아니라 관련 동작 형태와 일치하는 전체 충격 형태도 유도할 수 있다. 이에 대한 연구로써 수 컷 메뚜기에서 비행할 때 날개의 진동 혹은 소리낼 때 날개와 다리 의 마찰동작과 같은 간단한 리듬동작들이 가장 적합하다. 고정장치에 매단 사막메뚜기를 풍동(風洞)에 날리는 실험을 할 수 있다 이 메뚜기가 바닥에 닿는 것을 하지 못하고, 동시에 머리에 나있는 감각모로 공기의 저항을 감지하자마자, 비행동작이 시작되었 댜 이때 한 마디의 좌우 날개는 동조성 진동을 하는데, 이에 비해 몸 한쪽의 앞, 뒤 날개는 시간적으로 서로 바꾸어가며 진동한다. 대 다수의 곤충들에 반해 메뚜기와 잠자리는 소위 직접 날개근육을 갖 고 있다. 다시 말하면, 날개기부에 직접 위치하는 거근( 擧 筋)과 하체 근이 교대로 수축해서 날개의 상하운동의 힘을 전달한다 . 그럼 이렇 게 날고 있는 메뚜기의 날개근육의 운동신경으로부터 활동전위를 측 정하면, ――이 운동신경은 가슴신경절에서 나와 근수축에 명령을 내 린다 -거근과 하체근에 대해서는 교대로, 한 마디의 양 거근 혹 은 양 하체근에 대해 동시에 그리고 몸 한쪽의 두 날개의 거근에 대 해 교대로 연결해서 충격이 발생한다. 이런 리듬형태는 완전히 자연 적인 것으로, 정상적인 날개움직임을 사진을 찍어 정확히 분석해 보 면, 이 리듬형태와 완전히 일치하고 있다 . 신경의 페이스메이커의 기원에 대한 설명은 실험에 의해 가능하 다 . 실험을 통해 날개근육에서 중추신경계로 연결되는 감각성 정보
는 단계적으로 차단이 가능하다 . 즉 이 날개를 인위적으로 고정시켜 서 모든 움직임을 방해하거나, 혹은 신장수용기로부터 날개근육으로 연결된 감각신경을 절단하여 어떤 정보 전달도 차단시킬 수 있다. 이 경우에서도 운동성 신경으로부터 정상적인 리듬 충격 형태가 유 도될 수 있다. 단지 충격의 빈도가 이 경우에는 느려질 뿐이다. 결 국 날개 움직임에 대한 정상적인 운동 형태는 (감각정보가 완전히 차 단된) 가슴신경절에서 유도가 가능한 것이다. 중추신경계에서의 리듬의 기본형태 역시 명백히 가슴신경절에서 형성된다. 이것을 담당하는 발생기는 완전히 독자적으로 작동한다. 다시 말해 메뚜기는 날개의 각각의 자리에 정보전달이 안되도 자신 의 비행 형태를 발생시킬 수 있다(물론 날개로의 정보전달은 이 페이스 메이커에 분명히 조절적안, 정상적일 때는 가속적으로 영향을 미칠 수 있 다). 동일한 방법으로 노래를 하고있는 귀뚜라미의 비행근육으로부터 리듬성 활동전위(충격 형태)를 유도할 수 있다. 이 충격 형태는 여러 노래 유형들이 날개의 마찰로 소환되는 동작 유형과 정확히 일치하 고 있다. 여기서도 충격형태는 감각신경, 죽 청각기관이 들어 있는 앞다리 경절을 절단하여 감각신경의 경로를 차단시켜도 정상적인 리 듬을 유지한다. 이러한 실험들은 중추신경계가 해당 근육들에서 각각의 관련 연속 동작들을 소환하는 완성된 프로그램, 즉 완전한 충격 형태를 독자적 으로 가동시킬 수 있는 상태를 지니고 있음을 입증해 주고 있다. 해 당 감각성 통로 없이, 즉 감각기관으로부터 관련 자극 전달도 없이 이러한 충격 형태의 생산을 〈중추적 조절〉 혹은 〈중추적 프로그램화〉 라고 부른다. 이것은 그동안 많은 동물종에서 많은 행동양식에 대해 의문을 푸는 계기가 될 수 있었다.
3 호르몬과 행동 동물들의 두번째 조절체계인 호르몬계는 천천히 작용하지만, 효과 에 있어서는 지속적이다. 무엇보다 이것은 장기간에 걸쳐 변화를 보 이는 행동영역의 조절에 적합하게 작용하고 있다. 제일 먼저 여기에 는 생식 행동영역이 속한다. 그러므로 특히 이 행동에 대한 뚜렷한 호르몬 영향이 성적인 행동영역과 양육 행동영역에서 잘 알려져 있다. 적은 정도나마 이 호르몬계는 다른 행동영역에서도 나타난다 (Bal t haz art 등 1983, Beach 1961, Guth rie 1981, Nau 와 Deli us 1981, Proeve 와 Sossin k a 1982, Rein b oth 1980, Slate r 1978) . 이 행동학적 내분비 연구는 다음에 소개되는 방법에 의해서 이루어지고 있다. 3. l 분비선과 호르몬 양의 조사 특정 호르몬을 분비하는 내분비선이 연주기에 따라 커다란 활동적 인 차이를 나타낼 때, 그리고 이것과 함께 그 동물의 특정한 행동활 동성에서 차이를 관찰할 수 있을 때 호르몬이 행동을 통제하고 있다 는 암시를 받게 된다. 이것은 우선 성적 행동영역에서 일반적인 현 상이다. 특히 조류와 포유류에서 잘 연구가 되어 있다. 몇몇 열대성 종과 인공적인 조건에서 사육된 동물의 경우는 예외로 하고 동물들 의 분비선은 부분적으로 현저한 정도의 연주기성 차이를 보인다. 이 분비선은 번식기의 끝에 수축했다가 다음의 번식기 초기에 다시성장 한댜 까마귀의 고환의 부피는 연주기에 따라 4-1200mm3 로 변화를 보이며, 유럽지빠귀 Turdus merula 의 경우 1-400mm3 의 편차롤 보인 댜 꿩의 고환은 이른 봄에 1200% 에 달한다. 성적인 활동성의 차이 는 이 크기의 차이와 시간적인 관계를 갖고 있다. 보통 가장 작은 크기의 고환 시점에는 모든 성적인 행동양식들도 결여된다. 많은 조
 (Iull6A 56~
(Iull6A 56~
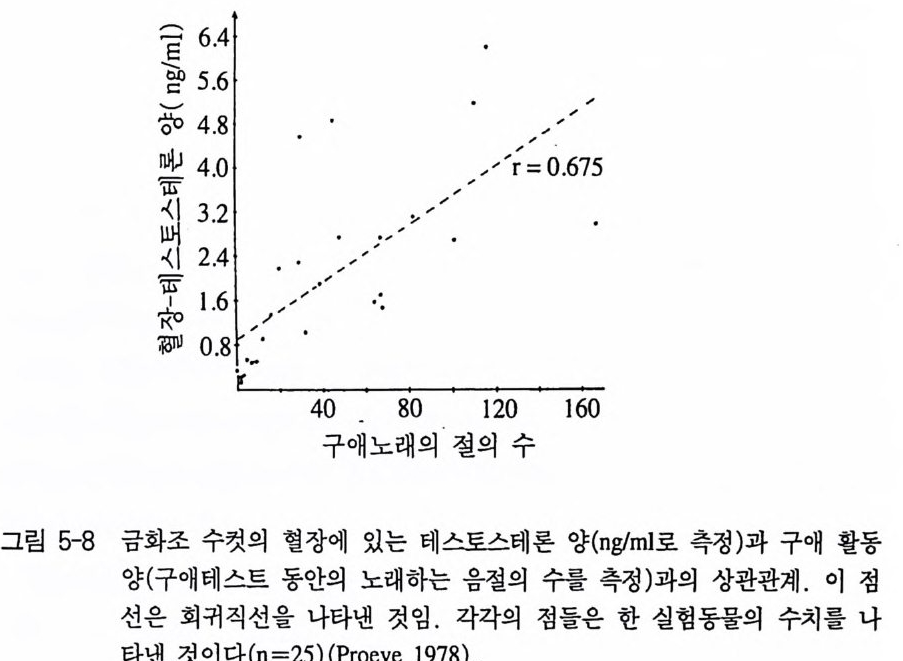
류에서 고환의 크기와 짝짓기 행동의 강도간의 정량적 관계가 조사 되었다 역시 조직학적 생화학적 연구를 통해서 내분비선의 활동성 과 행동 간의 관계가 밝혀질 수 있다. 이러한 관계는 방사선면역측정법에 의해 혈장속의 호르몬 양을 직 접 측정하여 확인할 수 있다 . 이 방사선면역측정법은 방사능에 의해 표시를 한 스테로이드와 특이적 호르몬 항체를 사용하여 각 호르몬 의 농도를 피코그램 (10 - 12 g)까지 측정할 수 있다(그림 5-8). 비교적 일 찍이 소개되었던 이 방법은 구동안 행동-내분비학적 연구에서 보편 화되어 왔다 (P hillips 와 Young ren 1971, Proeve 1978). 이 방사선면역측 정법은 특히 혈장속에 있는 특정 호르몬의 농도에 영향을 미칠 수 있는 특정 행동양식들의 피드백 결과를 알아보는 대 적합하여 성적
행동양식들(구애, 교미)의 결과로써 성호르몬 양의 변화를 추적하는 데 이용되었다. 3.2 수술 행동학을 포함한 생물학 모든 분야에서 호르몬 효과를 알아내기 위한 가장 오래된 방법은 내분비선의 제거다. 제거를 한 후 해당 행 동양식이 없어지면, 관련 분비선의 호르몬이 그 행동을 조절하고 있 다는 사실이 입증된다. 이런 실험은 어느 정도 결함을 수반한다. 이 외과적 수술은 동물체의 일반적인 상해를 초래할 수 있으며 그래서 개개의 행동양식들의 소실을 무조건 특정 호르몬의 결핍으로 돌려서 는 안된다. 여기서 완전히 동일한 조건하에 모의 수술이 항상 병행 되어야 한다. 3.3 호르몬의 대용 이전에 없었거나 혹은 더 이상 발생하지 않는 행동양식이 인위적 인 호르몬 주입 후에 나타나면, 직접적 영향을 가장 확실히 볼 수 있는 방법이 된다. 이 호르몬 주입은 구강 혹은 주사를 통해 할 수 있다. 좀더 장기 지속적 효과는 호르몬을 결정체로 만들어 이식하는 방법인데, 이것은 호르몬이 혈관에 남아 오랜 기간에 걸쳐 지속적으 로 영향을미칠 수 있다. 4 호르몬의 효과 범위 호르몬의 영향에 대한 대부분의 실험적 연구는 척추동물의 성적행
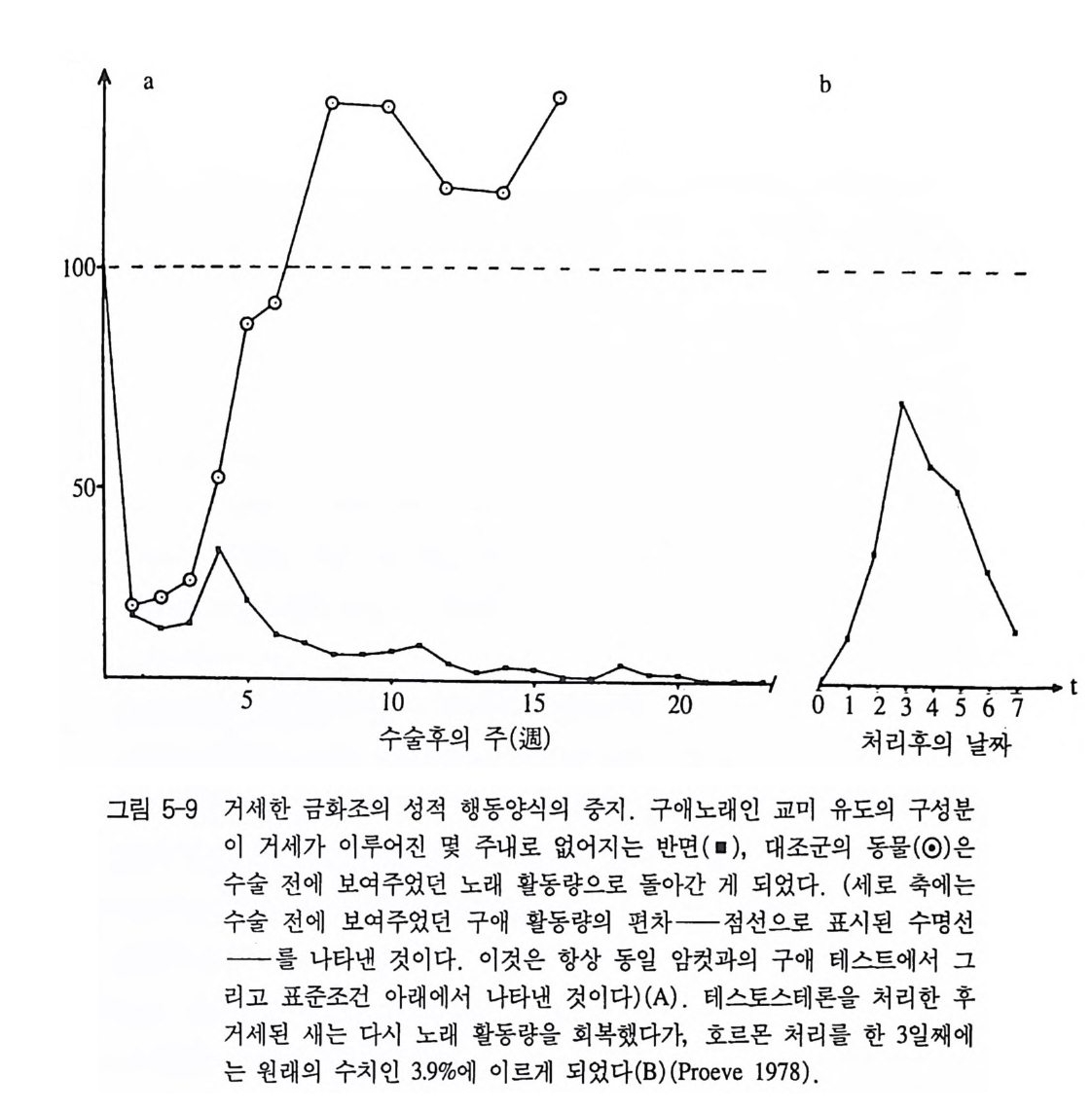
동에서 이루어졌다. 특히 수많은 연구 결과가 포유류에서 나왔으며 그 중에서도 쥐와 마카크원숭이에서 대부분 이루어졌다. 척추동물의 웅성 호르몬을 안드로겐이라고 부른다. 이 호르몬은 주로 고환에서 만들어진다. 가장 중요한 안드로겐은 테스토스테론이 댜 암컷에서는 에스트로겐과 제스타겐의 두 그룹의 성호르몬이 있 댜 우선 에스트로겐은 난소에서 형성된다 . 이것은 난자의 성숙을 조절하며 암컷의 성징 발달에 관여한다. 이 가운데 가장 중요한 대 표적 호르몬은 여포호르몬(에스트리올)이다. 제스타겐도 마찬가지로 성적인 과정에서의 조절에 관여하나, 보통 에스트로겐보다 늦게 작 용하며 결국 교미 후에 이어지는 생식 주기에 더 많이 영향을 미친 다. 예를 들어 포유류는 자궁벽에 난자가 착상할 수 있도록 준비해 야 한다. 이러한 준바를 위해 가장 중요한 제스타겐은 황체호르몬 (프로게스테론)이다 . 호르몬을 통한 행동의 조절은 웅성에 대해 가장 명확히 나타낼 수 있다 그 이유는 정상적으로 한 호르몬 그룹만이 행동 조절에 참여 하고 있기 때문이다. 4,1 수컷의 성행동 거세 지금까지 연구된 모든 척추동물의 고환 제거는 전체 성행동의 완 전한 중단으로 이어졌다. 이때 다음과 같은 규칙성이 나타났다. 대 부분의 경우 성적 행동양식이 일시에 사라지는 것이 아니고 점진적 으로 사라진다 . 성적인 지속기간은 종마다 다르며 쥐와 금화조는 5 개월까지 지속되었으며, 개의 경우에는 5 년까지 지속되었다(그림 5- 9A). 일반적으로 거세의 효과는 나이가 많은 개체보다는 젊은 개체 에서 더 빨리 나타난다. 그 밖에도 성적 영역에서 출발한 모든 행동
양식들이 동시에 사라지는 것은 아니다 . 교미능력은 시간적으로 가 장 먼저 사라진다. 그것은 점차 높은 해발역치를 가진 행동양식으로 이어진댜 비교적 약한 성적 동인을 갖고 있는 구애 요소들은 그에 상응하여 가장 오래 지속된다. 결국 개별적 성적 행동양식들은 거세 를 했어도 종종 지속적으로 활동적일 수 있다. 그러나 여기서 성적 행동양식들이 뇌하수체로부터 직접 통제되는 것인지 혹은 성호르몬 의 역할이 부분적으로 부신 호르몬에서 넘어온 것인지 아직 알려져 있지 않다 . 외과적 거세 대신에 요즘 화학적인 거세가 있다. 이에 대해 소위 〈항안드로겐〉(예, c yp ro t eronace t a t)을 사용한다. 이 합성물질은 안드로 겐 수용기를 차단하여 옹성 호르몬의 효과를 억제한다. 성적 행동에 대한 결과는 근본적으로 고환의 제거와 같다. 방법면에서 화학적 거 세는 그 효과가 가역적인 면에서 커다란 장점이 될 수 있다 . 즉 항 안드로겐의 화학적 분해가 있은 후에는 몇주 혹은 몇달 내에 완전한 성적 활동성이 회복될 수 있다 (Neumann 와 Ste in b eck 1971 ). 또 피를 홀리지 않는 거세 방법이긴 하지만, 가역성이 아닌 방법으로 내분비 선 조직을 레이저로 파괴하는 방법이 최근에 개발되었다 . 대용 반대로 안드로겐의 인공적인 주입으로 옹성 행동에 활발한 영향을 미칠 수 있다. 이러한 실험은 여러 개체군에 실행할 수 있다. 생식 기간 이외의 수컷들, 거세한 수컷들, 어린 개체들 그리고 암컷들로 나눌 수 있다. 겨울철에 새에게 테스토스테론을 주사하여 노래를 부 르게 할 수 있으며 큰가시고기에게 이 호르몬을 주사하면 짝짓기 행 동을 유발시킬 수 있다. 거세된 동물의 경우 없어졌던 행동이 점차 회복된다(그림 5-9B). 이때 개개의 성적 행동요소들이 수술 후에 없 어졌던 것과는 반대의 순서로 다시 나타난다 .
 a b
a b
어린 개체에게 테스토스테론의 효과는 아주 각별하다. 수평아리의 경우 생후 며칠만에 울며, 숫쥐는 생후 14 일에 첫번째의 교미를 실 시한다. 수캉아지는 테스토스테론의 주사 후에 한 쪽 다리를 쳐들고 오줌울 눈다 이것은 개의 경우 실제로 성체 수컷에서만 나타나는 행동이다. 암컷에게도 비슷한 효과가 나타날 수 있다. 암카나리아는 테스토스테론 처리 후 노래하기 시작하며, 암닭은 수컷과 같이 구애 동작을 보이면서 울기도 하며, 그리고 암쥐는 숫쥐의 등을 올라타려 는 행동을 보인다.
4.2 암컷의 성행동 자성 행동의 호르몬 종속과 관련된 연구 결과는 웅성 행동에 비해 서 일관성이 적다. 이것은 두 개의 호르몬 그룹이 함께 관여하여 복 잡하게 작용하는 데 기인한다. 보통 웅성 행동에서와 같이 한 개의 호 르몬에 의해 직접 관련되어 일어나는 현상은 무척 드물게 발생한다 . 그렇지만 여기서도 매우 분명한 결과는 거세에서 나타난다 . 그 효 과는 암컷에서 성행동이 몇주 혹은 몇달 동안에 처음 사라지는 것이 아니고, 보통 거의 급속히 사라지기 때문에 웅성의 행동에서보다 훨 씬 더 강렬하게 작용한다. 재생이 여러 가지 방법으로 가능하다 . 토끼와 고양이의 경우 에스 트로겐 하나만을 통해서 성적 감수성이 완전히 되살아 날 수 있는 반면, 다른 포유류(쥐, 염소)에서는 두 호르몬 그룹이 요구된다. 이 때 에스트리올의 주입은 종종 성행동의 첫 단계가 유발되는 반면, 완 전한 행동의 형성은 추가로 프로게스테론의 주입에 달려있다. 많은 경우에 완전한 효과는 에스트로겐과 제스타겐이 일정한 시간 간격울 두고 그리고 비교적 일정한 양으로 처리하면 기대할 수 있다. 안드 로겐과는 반대로 자성 호르몬의 효과는 물론 자기의 성에 한정되어 작용하고 있다. 수컷의 경우 보통 에스트로겐의 투여로 자성 행동이 유발되지 않는다. 4.3 다른 행동영역에서의 성호르몬의 효과 성호르몬은 실제로 성행동 의예 다론 행동양식에도 영향을 미친 댜 그러나 이런 행동양식으로부터 대부분은 어떤 식으로든 생식과 관계를 맺고 있다 . 공격적 행동에 대한 영향이 매우 명백하게 나타 난다. 둥지짓기도 호르몬에 의해 조절되고, 관련 호르몬의 종류는
어떤 성(암, 수 ) 이 둥지짓는 책임이 있는냐에 따라 다르다 . 만일 홍엽 새 Qu e/ea q uelea 나 큰가시고기와 같이 수컷 혼자 집을 짓는다면, 이 둥지짓기는 테스토스테론을 통해 활성화된다. 이에 반해 암컷이 집 울 짓는다면, 자성 호르몬이 작용한다 . 그러나 이 자성 호르몬의 참 여는 다양하다. 생쥐는 둥지짓기가 프로게스테론에 의해, 멧비둘기 와 A gap om i s 속의 앵무새는 에스트로겐에 의해, 골드햄스터는 두 호 르몬이 함께 작용하여, 토끼는 두 호르몬의 양에 따라 영향을 받는다. 쥐와 병아리에 대한 실험 결과 성호르몬은 학습 능력에도 영향을 미칠 수 있다는 사실이 밝혀졌다 . 이 두 경우에서 학습 능력은 테스 토스테론의 처리 후에 여러 면에서 분명히 향상되었다. 이것은 어느 정도 생물학적으로 의미가 있다. 동물들은 생식기 동안에 주변환경 으로 부터 추가적인 많은 것들을 알아야만 하고(터 경계, 둥지의 상 태, 터 이웃, 배우자, 그리고 새끼의 개별인식) 자·연히 이 시기에 보다 높은 학습 능력 이 요구된다. 마지막으로 어떤 한 동물의 전체 활동 규모가 성호르몬과 직접으 로 연결될 수 있다. 물론 지금까지 연구된 결과의 기본 개념에 근거 해서 이에 대한 일반화는 아직도 불가능하다. 그러나 이 호르몬 양 의 차이는 번식기 동안에 항상 보다 높은 성적 행동과 관련해서 의 미 있게 나타난다. 4.4 그 밖의 호르몬 성호르몬 외에도 특이적 호르몬 효과에 대해 비교적 일관성이 없 는 몇몇 실험 결과가 나와 있다 . 부신피질 호르몬은 성호르몬과 비 슷한 효과를 갖고 있는 것으로 나타났다. 부신피질 호르몬은 화학적 인 조성에서도 성호르몬과 매우 비슷하다. 몸의 가장 중요한 호르몬 분비기관은 뇌하수체다. 뇌하수체는 두
가지 종류의 호르몬을 분비한다 . 표적조직에 직접 도달하는 직접 호 르몬이 있고 다른 호르몬 분비선에 영향을 미치는 간접 호르몬이 있 다. 행동의 영역에서 이 두 부류의 호르몬은 효과 정도도 다르다. 몇몇 뇌하수체 호르몬은 특정 행동영역(예, 새끼돌보기, 공격)에 직접 영향을 미칠 수 있다. 많은 척추동물에서 프로락틴은 양육행동을 조 절하는 일을 맡고 있고 이와 병행하여 영양기관(포유류의 유선, 비둘 기의 소낭유) 발달을 촉진한다. 이에 비해 뇌하수체 호르몬은 다른 호르몬의 분비를 간접적으로 조절하는데, 예를 들어 성호르몬의 분 비에 영향을 미친다 . 물론 이따금 두 기능 양식이 동시에 발생한다. 뇌하수체 두 호르몬인 FSH( 여포자극 호르몬)과 LH( 황체자극 호르몬)은 한편으로 성호르몬 분비에 영향을 미쳐서 행동도 간접적으로 일으키 며, 다른 한편으로는 이 두 호르몬은 많은 경우에 여러 행동양식에 직접적으로도 영향을 미칠 수 있다 . 몇몇 키클리드과의 물고기에서 거세된 수컷들도 완전한 정상적인 성행동을 보여주고 있다. 여기서 뇌하수체 호르몬은 이 행동의 조절을 직접 관장하고 있으며, 반면 안드로겐의 효과는 비교적 적은 것으로 확인되었다. 물론 일부 〈뇌하수체 호르몬〉은 뇌하수체 자체에서 만들어지는 것 이 아니고, 간뇌의 일부인 시상하부에서 만들어져 이 뇌하수체는 단 순히 저장만을 맡고 있을 뿐이다. 4. 5 무척추동물의 호르몬 무척추동물 가운데 가재와 곤충이 가장 잘 연구되었다 . 이들도 성 행동의 영역에서 호르몬의 영향이 가장 분명히 나타나 있다. 물론 척추동물에 비해서 어느 정도 근본적인 차이는 있다. 무척추동물은 성선 자체에서 호르몬을 생산하지 않고 항상 분비선 밖에 분비장소 가 있다 이런 이유에서 무척추동물의 거세는 보통 눈에 띄는 효과
가 나타나지 않는다. 수컷의 게에서 성행동은 수정관 vas de fe rens 에 있는 소위 안드로겐 분비선에서 생산되는 호르몬에 의해 조절된다. 바퀴벌레에서는 뇌 가까이에 위치한 한 쌍의 분비선인 측심체 co rp o ra card i aca 의 호르몬이 성 행 동을 맡고 있다 . 이 호르몬은 하인두 신 경절에서 정상적으로 나오는 교미동작의 억제를 제거하여 교미를 유 발시킨다 암컷의 메뚜기나 초파리는 알라타체 corp o ra allata 호르몬 의 효과하에 수컷과 교미할 수 있다. 성행동 이의에 곤충의 경우에 고치를 짓는 유충의 행동뿐만 아니라 일반적인 동작의 활동성도 호 르몬에 의해 조절될 수 있다 . 4. 6 호르몬들의 연합 작용 지금까지 제시된 예들은 특정 행동양식들의 조절이 보통 하나의 호르몬에 의해 나타나는 것이 아니고 오히려 여러 호르몬들의 연합 작용에 의해 이루어진다는 사실을 보여주고 있다. 물론 수컷의 성행 동은 여기서는 예외가 되고 있다 . 일반적으로 호르몬들은 협력하여 혹은 길항적, 죽 상호 억제하여 작용할 수 있다. 협력관계는 성선 호르몬과 부신피질 호르몬 사이에 서 성립된다. 길항적 호르몬은 일반적으로 웅성과 자성 호르몬이지 만, 항상 그런 것만은 아니다. 두 호르몬은 뇌하수체 호르몬인 프로 락틴과 함께 중요한 길항관계를 갖는다. 그런데 여기서 프로락틴의 분비 증가는 짝짓기 단계에서 새끼돌보기 단계로의 이행을 조절한다. 뇌하수체 호르몬과 뇌하수체 호르몬 가운데 직접 호르몬 사이의 관계가 특히 다양하다. 여기서 한편으로는 다른 호르몬 분비선의 활 성에 뇌하수체 호르몬이 조절기능을 맡고 있으며, 그러나 다른 한편 으로는 뇌하수체의 직접 호르몬과 다른 분비선 호르몬 간에 연합작용 도 존재한다. 버들붕어는 이 종의 전형적인 거품둥지를 짓는데, 호르
몬둘은 각각 협력하여 이 거품둥지를 짓는 데 관여한다. 테스토스테 론은 이 물고기가 물 표면으로 헤엄쳐 공기를 가져 오는 빈도를 높 이며, 뇌하수체 호르몬인 프로락틴은 입에서 점액질 생산울 촉진하 여 둥지 재료로 쓰이는 공기방울의 벽을 만들 수 있게 한다. 웅성과 자성 호르몬 간의 정상적인 길항적 효과에도 불구하고 어 떤 경우에는 완전히 협력작용을 할 수 있다. 멧비둘기의 짝짓기에서 개별적 요소들(사냥, 몸굽히기)이 보통 때에는 테스토스테론에 의해 조절되는 반면, 둥지 장소를 가리킬 때는 자성 호르몬인 에스트리올 에 의해 이루어진다. 여기서 에스트리올은 마찬가지로 고환에서 생 산된다 그럼 하나의 호르몬은 하나의 특정 효과를 일으키는 것이 아니고 거의 항상 여러 호르몬들의 연합 작용으로 행동에 호르몬이 영향을 미치며 그리고 호르몬 양의 비교적 작은 변화라도 효과가 달 라질 수 있다는 사실로 요약될 수 있다. 4.7 경험의 영향 호르몬들간의 다양한 상호작용과는 달리 호르몬계의 영향이 대체 로 적게 나타날 수 있다. 오히려 호르몬과 관련된 행동들이 다른 조 절기작과 더 깊은 관계가 성립된다 . 이것은 동물의 종에 따라 다르 게 나타난다. 종비교는 여러 동물종들에서 호르몬을 통해 활성화된 행동양식들이 완전히 호르몬 수위와는 독립되어 있다는 사실을 보여 주고 있다 여러 가지의 양육 행동들(새끼의 핥기, 감싸주기, 앉기)이 쥐의 경우만 해도 새끼 혹은 거세된 수컷들에서도 나타난다. 많은 조류와 어류에서도 다양한 양육 행동와 둥지짓기 행동들이 호르몬과 무관하게 나타날 수 있다. 아마 이들은 순전히 신경계에 의해 조절 된다고 볼 수 있다. 그러므로 신경과 호르몬의 조절 정도는 명백히 종마다 다르고 행동마다 아주 다르다 .
그 밖에도 호르몬의 종속 정도에 있어서 종내의 차이도 잘 알려져 있다 종내의 차이는 대부분 경험에 의존하고 있다 . 보통 젊은 동물 의 거세가 늙은 동물의 거세보다 더 강하게 영향을 미치며, 이에 반 해 인위적인 호르몬 처리는 더 약하게 작용한다. 암잉꼬의 에스트로 겐은 둥지상자를 찾아오는 행동을 소환시키며, 더욱이 번식 경험이 있는 암컷들은 한 번도 번식 경험을 갖지 않은 개체보다 더 빨리 둥 지상자를 찾는다 . 멧비둘기의 경우 프로게스테론을 통해 번식행동이 촉진되는데 프로락틴을 통해 먹이먹이기 행동도 마찬가지로 경험이 없는 암컷보다 경험이 있는 암컷이 더 빨리 나타난다. 비슷한 연구 결과가 물고기에서도 나타나 있다 . 그래서 원래 호르몬의 지배에 있 던 행동양식이 경험이 풍부해지면서 점차 호르몬과는 독립적으로 유 발될 수 있는 것이다. 포유류 내에서 이런 현상을 비교해 보면, 다음과 같은 규칙성을 인식하게 된다 . 쥐와 다른 설치류에서는 호르몬 영향에서 경험조건 적 차이가 비교적 적은 반면, 고도로 발달한 포유류에서는 이것이 최고에 달하고 있다. 여기서 우리는 몇몇 경우에서 거의 완전한 보 상가능성을 확인할 수 있다 . 거세된 수고양이는 아주 확실히 활동이 떨어진 성행동을 보이나, 거세 전에 이미 여러 번 교미를 했다면 지 속적인 성적인 활동을 보여준다 . 이에 반해서 똑같이 처리한, 경험 이 없는 동물들은 이 성행동이 지속되지 않고 사라진다. 비슷한 결 과가 수캐에서도 나타나며, 몇몇 영장류에서도 성체 수컷의 거세는 성행동에 영향을 미치지 않고 거의 그 행동이 유지된다. 어쩌면 성 행동은 성호르몬과 점차 무관한 쪽으로 진화되온 것으로 보여진다. 이러한 경향은 척추동물 내에서 고등한 동물에서 대뇌가 점차 증대 된 의미와 일치하고 있다 .
5 호르몬의 효과 호르몬 조절이 나타나는 형태나 규모는 호르몬의 종류와 방식에 따라 다르다. 호르몬 수위가 다만 동인에 영향을 미치며 행동이 일 어나는지 일어나지 않는지 혹은 어느 정도 자주 행동이 일어나는지 를 결정한다. 그러나 한 동작이 진행중에는 아무런 영향을 미치지 않는댜 호르몬 수위는 단지 중추신경계와 협력을 이루고 있을 뿐이 댜 이에 대한 현재의 지식 수준에 따라 최소한 네 가지의 호르몬들 의 직간접 효과방식을 열거할 수 있다. 5.l 발신자의 영향 호르몬은 해발인의 효과를 증진시킬 수 있으며 이로써 강화된 반 응을 간접적으로 유발시킬 수 있다. 중요한 공격적 해발인을 나타내 는 큰가시고기 수컷의 배의 붉은 색깔은 옹성 호르몬의 영향을 받아 발달하게 된다. 수탉의 벗, 많은 새들의 현란한 깃털 혹은 암침팬치 의 성기 주변에 발정부품과 같이 공격 내지는 성적인 영역에서 의사 소통의 의미를 갖는 제 2 차 성징들도 성호르몬 혹은 뇌하수체의 성선 자극 호르몬을 통해 영향을 받는다. 음성학적 영역에서 많은 조류의 〈변성〉도 호르몬에 의해 나타난다. 그 밖에도 곤충들의 암컷들에서 수컷을 유인하고 성적으로 자극의 효과를 일으키는 냄새 물질의 형 정도 마찬가지로 호르몬에 의존하고 있으며, 바로 이것은 알라타체 에 의해 조절된다. 해발인의 호르몬 조절 변화는 전 생애 동안 단 한번만 나타나거나 (번식 성숙에 이르기 위한 제 2 차 성징의 분화) 혹은· 규칙적으로 반복해서 연주기로(혼인색) 혹은 몇 주주기(발정부품)의 간격을 두고 나타난다.
5,2 수신자의 영향 반대로 호르몬은 감각기관의 수용능력에 영향을 미치며 이로써 감 각신경으로부터 입수하여 전달된 정보를 변화시킨다 . 숫쥐에서 테스 토스테론은 페니스 표면의 민감도를 증진시키며 접촉자극으로 강화 된 정보가 성행동을 활성화시킨다. 많은 조류의 포란 지점의 민감성 은 프로락틴에 의해 높아진다. 이로써 알과의 접촉은 포란 행동 및 먹이먹이기행동을 활성화시키는 더 강력한 자극을 생산한다 . 5.3 신경계의 활성화 효과 여러 호르몬들의 연합 작용 외에도 호르몬과 각 뇌영역 간의 상호 관계가 일어나고 있다 . 이런 상호작용의 연구에 대한 첫번째 방법은 거세된 동물에 방사능으로 표시된 성호르몬을 주사하는 것이다 . 이 주사 방법은 뇌의 특정 핵심 부위에 국부적으로 매우 눈에 띄는 방 사능 표시 스테로이드의 축적이 생기게 한다. 표시된 스테로이드의 자동 X 선 촬영술을 이용하여 정확한 뇌의 위치를 추적할 수 있다. 숫멧비둘기의 경우 방사능은 시상하부에 테스토스테론으로 처리한 후에 가장 높으며 이에 비해 대뇌에는 현저히 적다. 방사능 표시된 에스트리올로 처리한 암쥐에서는 마찬가지로 시상하부에서는 가장 높은 농도를, 대뇌의 해마상 융기에서는 중간 정도를 그리고 대뇌피 질에서는 가장 약한 농도를 보인다. 이 방사능은 주사후 1- 少`]간에 최고에 달하며 대략 입]간이 지나서는 다시 농도가 낮아진다. 그러 므로 각 뇌부위는 혈류를 타고 온 특정 호르몬을 뇌세포와 세포핵으 로 홉수하여 거기서 한두 시간 농축된다. 두번째 방법은 지금과 반대의 방향에서 이루어지고 있다. 이 방법 은 어느 면에서 전기적 뇌자극 방법과 비슷하나, 전극 대신에 뇌에
직접 이식하는 극소형의 호르몬 결정체를 사용하는 것이다 . 뇌안에 서 호르몬 결정체가 서서히 녹아 이 호르몬은 주변의 구역으로 스며 들어간다. 그렇지만 이런 이식 실험을 통해 행동양식들을 개별적으 로 활성시킬 수 있다. 말하자면 효과가 단지 아주 제한된 자리에 겨 낭된댜 비둘기에서 짝짓기 행동의 개별적 요소들이 테스토스테론 결정체를 이용하여 제각기 분리된 자극 지점에 영향을 미칠 수 있 다 거세된 수탉의 경우 간뇌의 측면에 테스토스테론 이식은 공격행 동을 활성화시키며, 이에 비해 전시각령에 이식하면 교미동작이 활 발해진다 만일 이 두 곳을 동시에 호르몬으로 자극을 주면, 짝짓기 행동의 전체 과정이 쉽게 유발된다 . 이러한 발견은 많은 짝짓기 행 동들이 동시에 성적 -공격적 혼합 동인을 소유하고 있다는 사실을 지지해 주고 있다. 이 두 실험방법을 통해 지금까지 얻은 결과를 비교해 보면, 자동 X 선촬영에 의한 농축 지역과 이식된 호르몬에 대해 민감한 뇌부위 는 분명히 서로 같음을 보여주고 있다. 거세된 멧비둘기의 짝짓기 행동은 정확한 지역(시상하부)으로의 테스토스테론 이식을 통해 활성 화될 수 있다. 이 지역은 주사 후에 방사능 물질의 커다란 농축으로 도 입증될 수 있다. 쥐와 고양이에서도 이러한 일치가 확인된 바 있 댜 결국 이런 것들을 통해 호르몬 종속 행동의 조절이 제각기 뇌안 에 있는 특정 신경과 결합되어 있음이 증명된 셈이다. 5.4 뇌 구조에 대한 결정 효과 물론 성호르몬은 생식가능한 성체에서 뇌에 가장 강한 영향을 미 치지 않고, 오히려 어린 동물에서 뇌에 더 영향을 미친다. 여기서 이런 영향은 무엇보다 호르몬 영향의 지속성으로 판단할 수 있다. 즉 성체에서는 활성 효과가 항상 짧은 시간만 지속되는 반면, 생후
첫날이나 이미 배 발생 도중의 태아에서는 호르몬 효과가 이후의 전 생애의 행동에 역행할 수 없이 영향을 미칠 수 있다 . 무엇보다 이것 은 어떤 동물이 이후의 나이에 웅성이 우위를 두고 행동하느냐 혹은 자성이 행동하느냐를 결정한다. 분명히 호르몬은 이 개체 발생 초기 에 특정 뇌 구조에 결정적인(조직화된) 영향을 미치고 있으며 이 뇌 구조를 어떤 한 쪽으로 고정시킨다. 방법적으로 결정적인 호르몬 영향은 두 가지로 실험을 해볼 수 있 댜 어린 동물을 매우 이른 시기에 거세하여 자체의 호르몬 생산을 차단하거나, 혹은 반대로 홈이 없는 동물이나 또는 거세한 동물에 다른 성의 호르몬을 인공적으로 주입할 수 있다. 이것은 주사와 이 식뿐만 아니라 다른 성의 성선을 이식해도 같은 효과를 나타낼 수 있댜 태아가 산모의 혈액 순환에 연결되어 있는 포유류의 경우 이 러한 호르몬 처리를 임신한 암컷들에 수행할 수 있으며 이로써 새끼 에게 이미 태어나기 전에 호르몬의 영향을 줄 수 있다. 〈화학적 거 세〉 자체는 이러한 식으로 이미 자궁 안에서 가능하다. 조류의 경우 는 예를 들어, 관련 호르몬 용액에 알을 담가서 부화 전에 영향을 미칠 수 있다. 이런 연구는 정상 발생에 대한 호르몬의 효과는 물론 인위적으로 변화시킨 호르몬 대용을 통한 행동 발달의 영향 정도에 대해서도 정 보를 제공해 준다. 첫번째 언급한 영역에서 무엇보다 포유류의 경우 암수간 재미있는 차이를 가져 왔다. 암컷의 경우에 성염색체를 통한 유전적인 성 결정에 따라 성선은 물론 행동을 포함한 2 차 성징(예, 몸 의 크기, 색깔 그리고 형체)도, 다시 말해 호르몬 자극 없이도 암컷 쪽으로 발달한다. 이에 반해 수컷의 경우에는 수컷 쪽으로의 해당 분화과정이 옹성 호르몬에 의해 일어난다. 만일 발생 도중에 안드로 겐이 어떤 단계에서 효과를 일으키지 못하면, 계속된 분화 과정에서 전형적인 암컷의 형질 쪽으로 잘못 발달하게 된다. 어미의 자궁속에
서 호르몬 거세가 일어난 수캐는 다리를 들고 오줌을 누지 못하고 암컷과 같이 앉아서 오줌울 눈다. 태어난 죽시 고환을 제거한 숫쥐 는 나이가 든 후에 홈없는 숫쥐에 비해 암쥐처럼 행동을 한다 . 그러므로 수컷 포유류의 태아의 경우에 이 성행동의 조절중추가 분명히 유전적 행동과는 독립적으로 결정되는 것이 아니고, 발생 초 기에 안드로겐의 영향으로 인해 옹성 쪽으로 기울어진다. 관련 뇌 중추가 암컷으로 분화되는 이러한 1 차적 경향으로 인해 포유류의 이 런 발생 과정을 수컷과는 달리 암컷의 발생을 〈기초 자성화 basic fe maleness 〉라고 부른댜 많은 포유류의 발생 과정을 관찰해 보면, 고 환의 일시적 활동이 아마도 안드로겐의 결정적 작용과 관계가 있는 증거가된다 . 다른 척추동물에서 이런 성별에 따른 차이와 관련된 연구는 얼마 되지 않는다. 그러나 그 가운데 몇몇 조류에서 연구가 이루어져 있 는데, 그 연구에 의하면 그 관계가 포유류와는 정 반대로 나타난다. 죽 조류는 〈기초 응성화 bas i cmaleness 〉의 특징을 띠고 있다. 이것은 뇌에서 행동 조절 중추의 성적 분화와 관련해서 동형접합성(포유류의 암컷둘은 두 개가 같은 성염색체를 갖고 있는 것에 반해, 조류는 수컷들의 성염색체가 같다)은 호르몬에 영향을 덜 받고, 이에 비해 이형집합성 은 호르몬 자극에 의존하고 있다는 사실을 의미한다 (A dki ns-Re g an 1983) . 비자연적인, 다시 말해 상반된 성호르몬 영향에 관련하여 암수는 얼마 차이가 없이 행동을 한다. 물론 여기서도 성체의 나이에 따라 호르몬 영향의 차이가 생겨난다. 이른 시기에 호르몬 처리의 효과가 안전도에서 뛰어나다. 임신한 어미에게 안드로겐을 처리하면, 암쥐 와 암기니피그에서 암컷의 성행동은 지속적으로 억압을 받거나 제거 될 수 있다. 만일 이 처리를 태어난 후 그 새끼에게 계속하게 되 면, 점차 전형적인 수컷의 행동양식으로 나타난다. 성 조절중추의
이러한 호르몬의 결정은 뇌하수체를 통해서 뒤늦게 성선자극 호르몬 분비의 시기와 관련해서 일어날 수 있다. 이것은 응성에서는 지속적 으로 일어나며, 이에 비해 규칙적인 배란기를 갖는 자성에서는 주기 형태로 일어난다. 쥐와 기니피그의 실험결과 암컷의 주기적 기능을 이른 시기 안드로겐 처리로 완전히 억제시킬 수 있었으며, 반대로 태 어난 직후에 웅성에게 거세를 해서 인위적으로 암컷의 성주기를 불 어올 수 있었다 (Faber 와 Ha id 1980, Nau 와 Deliu s 1981 ). 이러한 지속적 이고 뒤늦게는 거의 더 이상 희복될 수 없는 효과는 각인과정을 통 한 장기기억 학습영역에서 비슷하게 나타난다. 이러한 이유에서 〈호 르몬 각인〉 이라는 말이 생겨나게 되었다. 호르몬의 해당 효과가 아주 특정한, 종특이적 시기 동안에만 일어날 수 있기 때문에 원래 의미 의 각인과 일치하는 점이 많다. 물론 여기에서도 개체발생학적 발달 과정에서 영향을 미치는 특정한 시기를 민감기라고 부른다 (Nau 와 Deliu s 1981 ) . 성체 뇌의 호르몬 효과와 어린 개체 뇌의 호르몬 효과 간의 표면 적인 차이에도 불구하고 이 두 과정은 아마도 서로 밀접한 관계를 갖는다. 말하자면, 호르몬 결정의 특성은 뇌의 특정 신경요소들을 민감하게 만들어 장차 각각의 호르몬에 대한 민감도를 높여주는 것 이라 생각해 볼 수 있다. 이런 식으로 발생 임계기간 동안 테스토스 데론이 정확히 존재하게 되면, 분명히 응성 호르몬에 대한 관련 뇌 부위가 민감해지며 자성 호르몬에 대해서는 둔감해진다. 반대로 테 스토스테론이 결여되면 자성 호르몬에 대한 민감도는 높아지고 결국 자옹의 두 성에서 장차 암컷의 성행동으로의 준비를 갖춘다. 물론 어떤 식으로 이러한 호르몬에 의한 민감화가 이루어질 수 있는지는 아직 밝혀져 있지 않다.
6 행동을 조절하는 호르몬 분비 여기서 다움의 질문을 통해 지금까지의 내용을 정리해 보자. 어떤 다양한 방식으로 호르몬은 행동을 조절하고 영향을 미칠 수 있는가? 그러나 이에 대한 해답은 아직 분명하지 않다. 오히려 반대로 특정 한 행동양식들의 유발 혹은 이미 행동생물학적 중요한 자극의 감각 이 호르몬 분비선의 활동에 영향을 미칠 수 있다라는 사실에서 그에 대한 해답을 찾을 수가 있다(그림 5-10). 이러한 영향은 무엇보다 생 식 영역, 죽 양육 혹은 성적 배우자 관계에서 인식할 수 있다. 어린 개체로부터 온 시각적, 청각적 그리고 촉각적 자극들은 뇌하 수체를 제거한 쥐가 30 분 이내에 양육 호르몬인 프로락틴의 급격한 분비를 유도하는 데 충분하다. 즉 새끼의 젖 빠는 자극은 여기서 촉 진적인 호르몬 분비 효과를 일으킨다. 조류에서는 포란할 때 알의 시각적 자극과 피부 접촉은 아주 비슷한 방식으로 프로락틴 분비를 높인다. 역시 짝짓기 행동의 자극적 내지는 동조적 효과는 호르몬의 분비로 이어진다. 멧비둘기의 연구 결과에 의하면 구애하는 수컷에 서 나오는 자극들은 암컷의 뇌와 뇌하수체를 거처 성선에 영향을 미
 0+ 4
0+ 4
치고 다시 이 성선은 행동에 영향을 미친다. 즉 가장 먼저 뇌하수체 는 황체형성 호르몬 (FSH) 의 분비를 증가하도록 영향을 미친다. 이것 은 난소에서 에스트로겐의 분비를 강화시키며, 다른 한편 이 에스트 로겐은 암컷의 둥지짓기 행동에 촉진적인 영향을 미친다. 이 둥지짓 기 행동의 수행은 다시 뇌하수체에서 여포자극 호르몬 (LH) 의 분비를 자극시키고, 이 여포자극 호르몬 (LH) 은 배란을 조절하고 난소에서 프로게스테론의 생산을 촉진시킨다 . 마지막으로 이것은 양육행동의 발달에 책임을 지게 된다 . 비슷한 효과가 동성간에서도 일어날 수 있으며, 무엇보다 집단 번 식하는 동물종에서 발견할 수 있다. 오스트레일리아가 원산지인 사 랑새 Melop si t tac us undula t us 는 수컷의 노랫소리가 암컷의 행동과 생 리에 영향을 미칠 뿐만 아니라 다른 수컷들의 고환의 성장도 자극시 키며 결국 테스토스테론의 생산을 촉진시킨다. 이로써 많은 조류의 집단번식에서 관찰되는 번식 동조성도 상호 자극에 기인되고 있음을 알수 있다. 몇몇 예에서 행동의 진행과 전환은 호르몬과 행동양식 간의 상호 작용을 통해 조절될 수 있음을 보여주고 있다 . 하나의 행동양식은 호르몬의 분비를 촉진하고 , 이 호르몬의 분비는 계속된 호르몬의 분 비에 영향을 미치는 다른 행동을 자극시킨다. 이러한 상호관계 속에 서 중요한 연결고리는 호르몬과 신경자극을 각각 다른 정보 형태로 전환시키도록 하는 중추신경계의 세포들이 된다. 이런 것에는 호르 몬에 민감한 뇌 부위에 있는 세포들이 속한다. 이런 부위는 입수된 호르몬의 정보를 신경성 조절중추에 보내며, 반대로 신경의 자극에 의해 신경분비세포가 신경분비물질을 생산하게 하며, 이것은 다시 호르몬 분비선의 활동에 영향을 미치게 된다.
제 6 장 행동 리듬과 정위 1 행동 리듬 l,1 서커디언 리듬 대부분의 동물들은 하루의 시간에 따라 규칙적으로 행동양상이 달 라지고 있다. 야생 수닭은 밤새 지냈던 나무 위에서 동이 트기 전에 울다가 나무 위에서 내려와 암탉들을 데리고 땅바닥에서 먹이를 찾 는다 이들은 대낮에는 쉬고 오후 늦게 먹이를 찾아다니다가 저녁에 다시 보금자리로 돌아온다 . 일반적으로 야생동물 등의 하루의 활동 시간은 온도의 변화, 포식자의 활동 그리고 먹이의 이용도에 따라 달라진다. 그러나 동물들은 제각기 이런 외부 요인과 무관하게 행동 하는 생물시계 bio l og ica l clock 를 갖고 있다. 우리는 이것울 서커디언 c ir cad i an 이라고 부르는데, 라틴어로 c ir ca 라 함은 〈대략〉이라는 의미 를 갖고 있으며 d i em 이 란 〈하루〉를 뜻한다. 그래서 동물들의 하루의 리듬은 대략 24 시간을 나타낸다. 이 서커디언 리듬은 식물과 동물에 널리 퍼져 있으며 행동뿐만 아니라 물질대사의 모든 측면에도 영향
울 미친다 (Ascho ff I98I, 1989). 야생동물들은 일주기의 활동기에도 아무것도 하지 않은 채 쉬면서 보내는 경우가 많다 . 그러나 이것도 일종의 적응된 행동으로 보인 댜 비활동적인 시간은 소화 (D i amond 등 1986), 에너지 비축 혹은 포 식자로부터의 방어에 필요하다. 실제로 은신처에서 잠자는 행동은 그들 스스로 잠정적인 위험 상황으로부터 피하고 있는 방법일 수 있 댜 꿀벌은 하루의 많은 시간들을 휴식하면서 지낸다 . 일주기의 활동성의 최고치와 최저치의 분포는 어느 정도까지는 종 특이적으로 작용한다. 그러나 정상적으로 주행성인 철새들의 이동시 기는 야간이 될 수 있다 (예, 거의 모든 식충성 작은 새들의 이동은 밤 에 이루어 진다). 대략 동물들의 생활주기에 따라 주행성 종과 야행성 종으로 구분할 수 있다. 일반적으로 이들은 이중점 b ig e mi nus 이라고 부르는 두 정점의 활동 곡선으로 나타난다. 주행성 종의 최고 활동 시기는 이른 아침이며 그 다음 활동 시기는 저녁이 된다. 밤에는 절 대적인 휴식기다. 야행성 종의 최고 활동기는 황혼이 깃든 다음에 도달하며 그 다음 활동기는 이른 아침이다. 그러나 그밖에도 다른 활동 유형, 예를 들면 최고 활동기만 있거나 혹은 낮과 밤이 최고 활동기인 유형들도 있다. 낮과 밤이 최고 활동기를 갖는 동물의 예 는 닷쥐가 있는데, 이 종은 분명히 주행성 혹은 야행성 종이라고 부 롤수없다 . 일주기 는 서 커 디 언 리 듬 cir c adia n rh yt hm 과 주기 적 외 부 요인, 죽 시각 지시자의 협력에 기초를 두고 있다. 이러한 자발적인 리듬의 결정적 증거는 여러 날을 똑같은 외부 조건하에 동물을 사육하는 데 서 알 수 있다. 일주기에서 변화되는 모든 환경조건들은 시각 지시 자로써 생물체에 시각을 알려주고 있기 때문에 이 일주기를 일정하 게 유지시킬 필요가 있다(예, 빛, 온도 습도). 주행성 동물들에게는 지속적으로 밝게 하고, 야행성 동물에게는 지속적으로 어둡게 유지
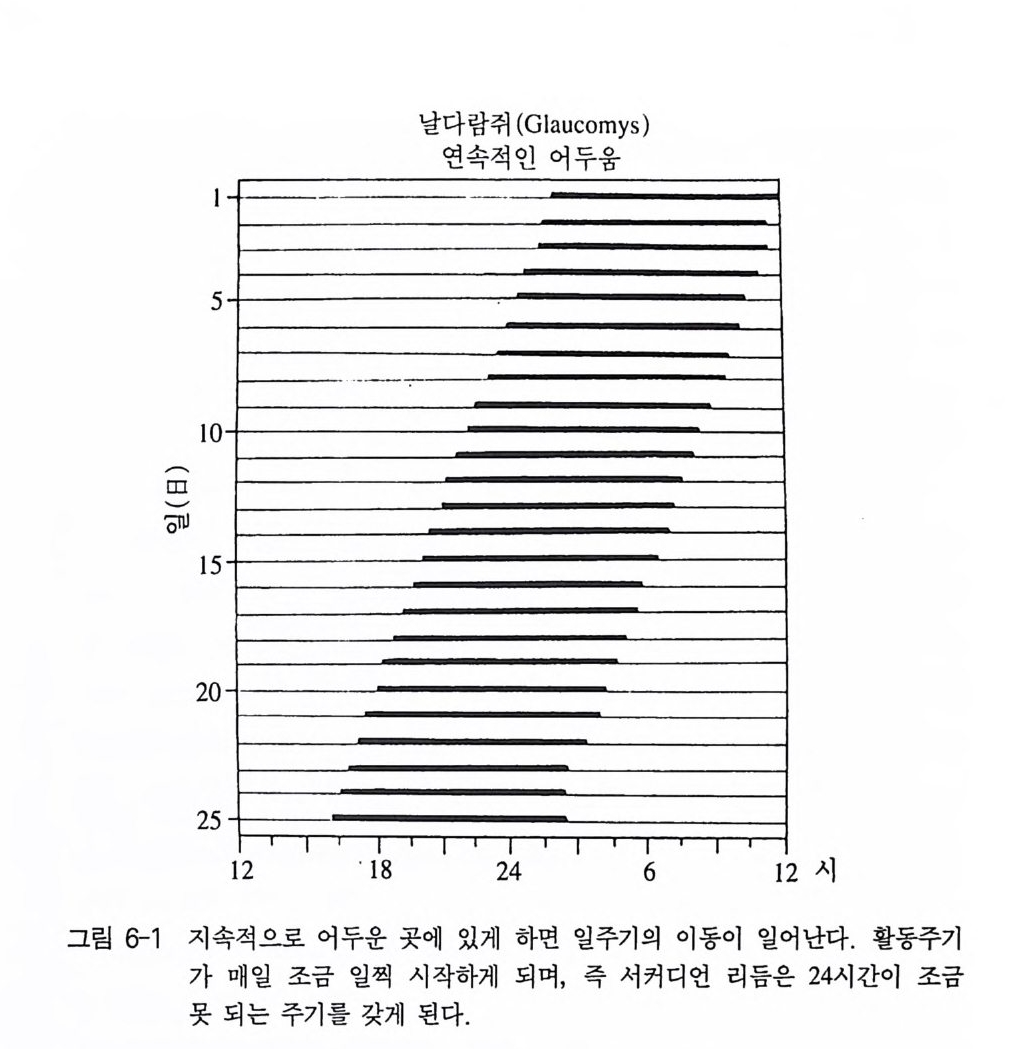 날다람쥐 (Glaucomy s)
날다람쥐 (Glaucomy s)
시켜 주어야 한다. 그럼 순수한 외부 조건적 일일 주기는 이런 조건 하에서 사라지게 된다. 실제로 생체 리듬은 이렇게 일정한 의부 조 건을 유지시키면, 동물이 매일 일찍 혹은 늦게 활동을 시작하는 지 속적인 주기 이동이 일어난다(그림 6-1) . 그래서 내부적 서커디언 리듬은 24 시간보다 대략 더 많거나 더 적은 주기의 길이를 갖는다는 사실을 알 수 있다. 주기적 외부 요인들은 이런 리듬에 대해 주도적 인 역할은 할 수 없다. 그 이유는 어떤 환경 요인들도 관련 주기의 길이룰 완전히 변화시킬 수 없으며 그 밖에도 각 개체는 다른 리듬
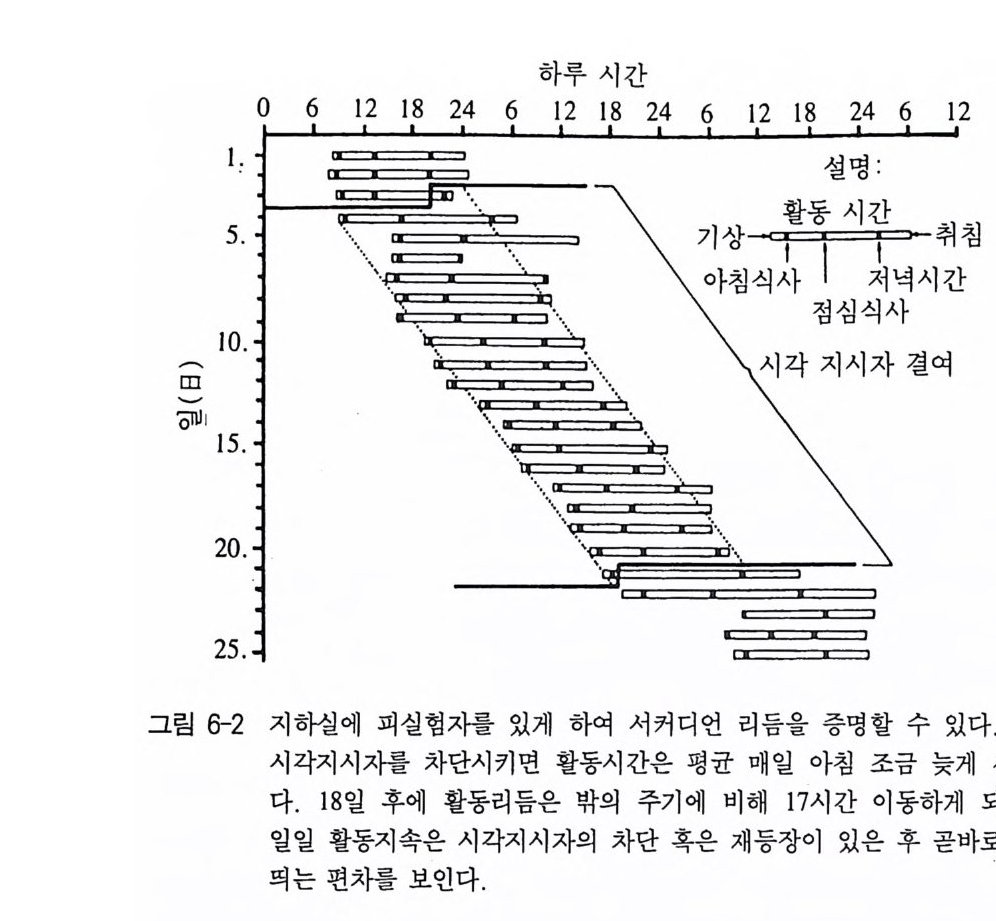 。 6 12 18 24 6 하12루 시18간 2 4 6 12 18 24 6 12
。 6 12 18 24 6 하12루 시18간 2 4 6 12 18 24 6 12
올 보여 주기 때문이다. 우리가 차고 있는 시계가 한 번도 아주 정 확치 않다면 매일 매일 새롭게 시간을 맞추어야 하는 것처럼, 생물 시계도 정확지 않아서 매일 외부요인인 시각지시자에 의해 자연적 인 낮과 밤의 변화와 맞추어져야 한다. 이 시각지시자는 24 시간의 정확한 주기의 길이를 맞추며 주기상태, 즉 정해진 하루의 시간에 최고 및 최저 활동기가 일어나게 한다. 서커디언 리듬은 간단한 단세포 생물에서 인간에 이르기까지 많은 동식물에서 증명되었다. 이를 증명하기 위해 피실험자롤 하루 종일 지하실과 같은 의부와 차단된 곳에 있게 할 수 있다. 불은 임의로 켤 수 있고 끌 수 있으며, 수면시간과 깨어 있는 시간은 피실험자
스스로 결정할 수 있다. 온도와 습도는 일정하게 해야 하며, 실제 시간을 알려 주는 물건들 예를 들면 라디오나 텔레비전은 모두 없앴 다. 이런 조건으로 서커디언 리듬은 24-26 시간, 평균 25 시간의 주 기의 길이로 나타나게 된다(그림 6-2). 이상하게도 피실험자들의 대 략 20% 가 30-33 시간 혹은 50 시간까지의 매우 긴 내부 수면-깽-리 듬이 발견되었다. 그러나 정상적으로 체온과 같은 영양 기능들은 이 런 느린 리듬을 갖게 할 수 있는 것이 아니고 평균 25 시간의 주기의 길이를 갖게 한다. 서커디언 리듬에 대한 생리학적 기작은 지금까지 잘 밝혀져 있지 않다 그러나 여러 조류에서 송과체 e pip h y se 를 의과적으로 제거했을 때 이동성 활동주기가 사라져 버린 사실이 발견되었다. 송과체를 제 거한 참새에다 다른 개체의 송과체를 이식시켰다. 참새는 서커디언 리듬이 다시 회복되었는데, 그것도 송과체를 수여한 개체와 같은 동 일한 주기상태가 관찰되었다. 이러한 발견은 서커디언 조정자 c ir ca di an p acemaker 가 몇몇 조류의 경우 송과체 안에서 찾을 수 있다는 사 실을 암시해 주고 있다. 생물체의 시각지시자의 증명을 위한 다음과 같은 방법이 제시될 수있다 . 첫째, 항상 같은 조건에서 관찰된 자발적 리듬은 시각지시자의 반 복된 출현을 통해서 다시 정확한 24 시간 리듬을 맞출 수 있다(그림 6-3). 둘째, 시각지시자의 주기이동을 통해서 최고의 활동주기는 이동될 수 있다. 예를 들어 동물원에서는 종종 야행성 종의 우리를 낮에 어 둡게 하고 밤에서 완전히 불을 켜둔다. 이에 따라서 이 동물들은 자 신들의 활동성을 과도기가 지난 후 변화되면, 낮에 이 동물들이 활 동하는 것을 관찰할 수 있다. 셋째, 하루의 길이를 인위적으로 짧게 하거나 길게 해서 일일주기
 ........... . I
........... . I
를 일정한 경계지점에서 변화시킬 수 있다. 그러나 이 시각지시자를 내부 리듬으로부터 너무 심하게 빗나가게 해선 안 된다. 21 시간보다 짧은 주기 길이 혹은 26 시간보다 긴 주기 길이는 일반적으로 시각지 시자에 대한 정확한 적용이 못 된다. 만일 생쥐를 인위적으로 16 시 간을 갖는 낮주기에 기르면, 활동 리듬이 파괴되어 활동성이 다소 불규칙적으로 하루 전체에 분포한다.
자연적인 낮-밤 리듬은 주기적 모든 환경 요인들 가운에 변함이 가장 적기 때문에 가장 중요한 시각지시자가 된다. 그러나 예를 들 어, 곤충의 경우 온도와 습도도 함께 작용할 수 있다. 사회적 시각 지시자도 예를 들어 새나 매미가 아침에 노래를 시작하도록 동조하 는 데 있어서 큰 의미를 갖고 있다. l.2 달주기와 연주기 일일주기 이외에도 외부 시각지시자와 내부 리듬의 협력을 통해 일어나는 더 긴 주기의 길이를 갖는 생물학적 리듬이 존재하고 있 댜 많은 무척추동물, 특히 곤충에 있어서 매달 성적 파트너들의 한 두 번의 군집은 특별한 달 리듬에 의해 도달된다. 모기 Clunio mari- nus 의 대부분의 개체군들은 15 일의 주기를 갖는 달 리듬에 의해 군 집을 이룬다. 그 밖에도 일일 리듬에 첨가되어 곤충들은 한달에 두 번 일정한 낮시간대에 알에서 깨어나온다 . 또 다른 곤충들은 30 일의 주기를 가진 달의 번식 리듬을 갖기도 한다. 다모충류인 Eun i ca v i r i d i s 는 특히 복잡한 주기를 갖는다 . 이들은 1 년중 항상 같은 시기 에 며칠간만 번식을 한다 . 이 경우 3 개의 주기가 겹쳐 있게 된다. 연주기는 번식을 10 월 /11 월에 한해서 영향을 미치며, 달주기는 특정 달주기(상현)에 한정시키고 그리고 일일주기는 특정 시간에, 예를 들어 사모아에서는 밤 12 시와 2 시 사이에 한하여 번식을 한다. 번식, 새의 이동 그리고 털갈이의 연주기적 현상을 연주기 cir - cann i an 라고 부른다. 항상 일정한 조건하에 실험동물들을 1 년 내내 가둬 기르면 이러한 입증이 가능하기 때문에 내부적 연 리듬의 연구 는 아직도 그렇게 진척이 많은 것은 아니다. 솔새는 항상 같은 조건 하에 털갈이가 규칙적인 주기를 유지하며, 무게의 증가 그리고 이동 의 동요를 갖게 된다. 다른 경우도 정상적인 주기의 길이에서 분명
한 편차를 갖고 유지된다. 이동 활동성의 시간적 과정은 내부적 프 로그램화가 되어 그 내부 프로그램이 작동되면 새들이 겨울 서식처 로 이동하는 것으로 추측할 수 있다 (Gw i nner 1991).
2 정위 2.l 주성* 정위(定位)는 생물체가 공간적 방향을 정하는 것을 의미한다. 식 물체에서 공간 정위를 우리는 굴성tr o pi sm 이라고 부른다. 이에 반해 자유롭게 움직이는 생물체, 특히 동물의 경우 무엇보다 움직임이 공 간에서 방향을 결정한다. 원래 이 방향성 움직임들을, 예를 들어 광 원에 방향을 정하는 곤충의 움직임을 모두 주성 t ax i s 이라고 부른다. 그러나 움직임의 비방향성 요소를 골라내서 단지 정위적 방향만을 주성이라고 부르게 되었다. 동물들의 거의 모든 움직임이 공간에서 방향을 정하고 있기 때문에 공간적 정위를 동물의 행동의 기본 현상 으로 특징지울 수 있다(예외로 예를 들면 하품이나 재채기와 같은 전형 적인 몇몇 반사가 있다). 만일 어떤 동물이 광원의 방향을 향해 움직이면 정(正)의 광주성 pho to t a xis 혹은 주광성이라고 부르며, 반대로 광원의 반대 방향으로 움직일 때를 부(負)의 광주성이라고 부른다. 자극원에 대한 이런 근 원적 정위를 기본 정위라고도 한다. 이 기본 정위는 종특이적 표시 가 될 수 없다 . 예를 들어 개미는 아침에는 정의 주광성 행동을 하 지만, 저녁에는 부의 주광성 행동을 한다. 물벼룩은 평상시에는 부 * 외부자극과 주성과의 상세한 관계는 제 3 장 외부 자극에서 설명되었다. 여기 서의 주성은 일반적인 자연 환경조건적 정위만을 다룬다.
의 주광성이지만, 이산화탄소가 첨가되면 정의 주광성의 행동으로 옮겨질 수 있다. 주성의 첫번째 분류는 방향성 자극의 성질에 따라 가능하다. 예로 써 만일 폴라나리아가 화학적 먹이 자극을 따라 움직인다면, 우리는 그것을 정의 주화성 chemo t ax i s 라고 부른다. 지구의 자장에 대한 정 위를 주지성 g eo t ax i s 이라고 한다. 예를 들어 소나무행렬나방 Thaume t op oea pity ocamp a 에서 용화가 준비된 유생은 땅에서 용화되 기 위해서 나뭇가지를 타고 아래로 내려온다. 동물들이 수류를 향해 정위를 할 때 주류성 (走流性 rheo t ax i s) 이라고 한다. 예를 들어 연어 는 번식을 위해 강을 따라 거슬러올라가는데 이때 정의 주류성 정위 를한다. 주성에 대한 또 다른 분류는 정위의 기초을 이루는 기작에 의해 가능하다. 가장 간단한 경우에서 어떤 동물은 무정위운동ki nes i s 에 의해 생활공간에서 정위할 수 있다. 그러나 아직 이것을 진정한 주 성이라고 하지 않는다. 그 이유는 이 정위가 자극 방향과 ‘ 관련된 정 위 동작을 통해서 일어나지 않기 때문이다. 오히려 이 동물은 운동 활동성의 변화에 따라 혹은 자극 방향과 관계없는 방향의 변화에 따 라 자극 강도가 변화 한다 . 쥐며느리는 정위 동작으로 습도구배에 따라 정위하여 습기가 많은 생활공간을 찾지 않고, 시행착오 원리에 따라 정위를 한다. 이들이 적당한 생활공간을 발견하게 되면 곧바로 그들은 자신들의 운동활동성을 줄인다. 그래서 통계적으로 보면 대 부분의 동물들은 습기가 있는 지역에서 살게 된다. 굴곡주성 kl i no t ax i s 은 무정위주성과 반대로 진정한 주성에 연결되 어 일어나는 정위 동작에 근거하고 있다. 굴곡주성 정위는 진자성 감각기관의 도움으로 연속된 강도 측정을 통해 가능하다. 금파리 Calliph ora 유충은 빛 에 민감한 몸통의 맨 앞부분이 좌우로 진자운동 을 하여 용화 직전 굴곡주성에 의해 광원으로부터 멀리 향한다. 왼
쪽과 오른쪽의 진동에서 연속적으로 측정한 빛의 세기가 같을 경우 유충의 운동 방향은 항상 일정하게 머문다. 그러나 이 빛의 세기가 왼쪽보다 오른쪽이 더 높으면 이 유충은 왼쪽으로 향하여 빛에서 다 시 멀리 움직인다. 빛이 위에서 비칠 경우 진자 광수용기가 항상 같 은 빛의 세기를 전달받기 때문에 금파리의 유충은 정위하지 않는다. 전향주성( 轉 向走性tr opo t a xi s) 은 좌우 대칭으로 배열된 감각기관의 동시적인 강도 측정에 기초하고 있다. 전향주성에 의해 정위하는 동 물은 두 수용기가 동일한 자극 강도를 감지하게 되면 앞으로 향한 댜 정의 광주성에서 빛이 오른쪽에서 비추면 그 동물은 빛이 비친 오른쪽으로 향한다. 부의 광주성에서는 그 동물은 반대로 왼쪽을 향 해 움직이게 된다. 정위를 하는 방향은 좌우의 자극의 차이로도 결 정되며 목표지점으로 향하는 두 수용기의 홍분의 밸런스를 갖고자 하는 데서 결정된다. 예를 들어, 만일 쥐며느리의 일종인 Armad illi d i um 이 한쪽의 광수용기를 파괴시키면, 자극의 불균형이 계속해서 일어나기 때문에 그 벌레는 원을 그리면서 돌게 된다. 만 일 아셀루스(등각류의 벌레)에 동시에 두 광원을 제시하면, 이 벌레 는 이때 자극의 밸런스가 이루어지기 때문에 중간 지점을 향한다. 목표주성 t elo t ax i s 은 자극의 강도의 측정 에 기 인하지 않고 감각기 관에서 고정된 지점을 유지하는 데 기인한다 . 전향주성과는 반대로 이 목표주성은 한쪽의 감각기관을 없애도 다른 한쪽의 감각기관으로 도 가능하다 잠자리 유충은 한쪽의 눈을 가린 후에도 먹이잡이 채 로 변한 아랫입술로 먹이를 정확히 낚아챈다. 예를 들어 소라게는 이중 광원 실험에서 중앙에 있지 않고 두 광원의 한쪽 광원에 방향 울정한다.
2, 2 지형 표지 정위와 태양 컴퍼스 정위 가장 간단한 경우에만 동물은 한 자극원을 향해 움직이거나 혹은 그 자극원에서 멀리 움직인다. 그러나 종종 일정한 각도를 유지하여 정위하거나, 혹은 그 동물이 여러 개의 정위 표지들의 도움으로 정 위한다. 이러한 복잡한 정위방식은 지형 표지 정위와 태양 컴퍼스 정 위가 있는데, 이런 것들은 보류주성(保留走性 meno t ax i s) 이라고 한다. 잘 알려진 생활공간에서의 정위는 지형표지 정위가 큰 의미가 있 댜 대군락을 이루며 사는 해안에 사는 조류가 정위 표지의 도움으 로 둥지를 다시 찾는 사실을 실험적으로 보여줄 수 있다. 눈에 띄는 돌, 나무, 해조류의 다발 등등을 갖다 놓음으로써 둥지로 돌아오는 어미들의 정위에 혼란을 일으킬 수 있다. 역시 벌들도 먹이장소로 날아갈 때 지형 표지에 의해 정위할 수 있다. 종종 이들은 직선 항 로에서 벗어나 시각적으로 눈에 띄는 나무와 건물과 같은 중간목표 지점을 향한다. 실험에 의해 태양 컴퍼스 정위와 지형 표지 정위가 서로 경쟁해서 나타날 수 있다는 사실이 밝혀졌다 (W il t schko 1981 ). 벌들이 먹이장소에 도착하기 위해 남쪽을 향해 숲 가장자리로 비행 하도록 학습하게 했다. 이 벌들을 동서방향으로 날아가도록 다른 숲 가장자리에다 갖다 놓아 주었다. 대부분의 벌들은 숲 가장자리로 날 아갔으며, 단지 일부만 태양 컴퍼스에 의해 방향을 정한 다음 남쪽 으로 날아갔다. 이 태양 컴퍼스 정위와 관련된 실험으로 태양의 도움으로 고정된 컴퍼스 방향의 억제가 가능하다. 태양이 동에서 서로 움직이기 때문 에 태양과 이룬 각도는 하루의 시간과 밀접한 관계가 있다. 그래서 이 태양 컴퍼스는 내부 시계(서커디언 리듬)를 전제로 하고 있다. 크 레이어 (Kramer 1952) 는 찌르레기로, 프리슈(1 965) 는 꿀벌을 갖고 서로 독립적으로 태양 컴퍼스에 대한 원리를 밝혀냈다.

기의 방향성은 빛의 입사각에 맞추어 일정한 변화를 일으켰다. 결국 이 새들은 태양 컴퍼스에 의해 낮시간이 달라질 때의 태양의 이동을 계산할 수 있는 것으로 보인다. 그래서 태양 컴퍼스는 태양과의 일 정한 각도를 유지하여 내부 시계의 도움으로 이 각도를 변화시킴으 로써 이루어진다. 연주기적 조건의 이동 활동성에 관계없이 훈련방법을 이용해서 태 양 컴퍼스 정위를 연구할 수 있다. 원형 새장의 외각에 뚜껑이 달린 10 개의 급이대를 설치하고 이 급이대로부터 예를 들어 동쪽 급이대 하나만 먹이를 채웠다. 특정한 컴퍼스 방향에 대한 훈련은 맑은 날 씨에 모든 지형 표지를 차단하고도 성공을 했다. 다시 이 찌르레기 들은 태양의 이동을 보상하려고 했다. 이 훈련이 고정된 낮시간대에 실시됐더라도 이 찌르레기들은 각각의 임의의 다른 시간대에서 정확 한 급이대를 선택했다. 이 내부 시계는 예를 들어 아침에는 태양의 방향으로, 정오에는 900 각도로 그리고 저녁에는 태양과 정반대 방 향에서 올바른 급이대를 찾을 수 있게 했다. 계속된 실험에서 이 자연의 태양은 인위적으로 고정해 둔 태양으 로 대치되었다. 원형 새장은 사방이 막힌 방에 위치한 천막 안에 넣 었댜 이 천막의 밖에서 새장을 향해 서치-라이트를 달아 주었다. 물론 이때 불빛이 천막의 천을 통해 통과할 수 있다. 찌르레기 한 마리를 자연의 하늘에서 서쪽을 향하도록 훈련시킨 후 이 새장에 넣 고 관찰했더니, 이 찌르레기는 저녁에 인공 태양의 방향으로 향한 급이대를 선택했으며, 정오에는 이 인공 태양과 900 이룬 각의 급이 대를 그리고 아침에는 이 서치-라이트와 반대 방향의 급이대를 선택 했다(그림 6-6). 이 찌르레기는 마치 이 서치-라이트가 자연의 태양 이 움직이는 것처럼 행동을 한 것이다. 내부시계를 실험적으로 차단시킴으로써 훈련된 컴퍼스 방향으로부 터 편차를 가져올 수 있었다. 예를 들어 찌르레기 한 마리를 맑은
 l二
l二
날에 남쪽 방향에서 먹이를 찾도록 훈련시켰다. 그런 다음 이 새에 게 인위적으로 낮-밤츠 F 기를 6 시간 이동시켜 보았다. 12-18 일 후 에는 이 새에게 정오를 저녁이 되게 한 것이다. 그럼 이 새를 자연 적인 조건하의 정오에 시험을 해보면, 이 새는 태양의 방향에서 급 이대를 찾지 않고 태양으로부터 900 왼쪽에 있는 급이대인 동쪽에서 먹이를 찾았다. 이 모든 실험들을 종합해 보면 찌르레기는 태양을 컴퍼스로 사용하며 하늘에서 움직이고 있는 태양의 각속도를 정확히 인지하고 있는 것으로 나타났다. 이 태양 컴퍼스정위는 완전히 다른 방법을 가지고 꿀벌을 통해 증 명될 수 있었다. 벌통을 밤에 낯선 환경에 옮겨 놓는 실험을 하여 알아 보았다(그림 6-7). 예를 들어 벌들을 하루의 시간대와 관계없 이 벌통에서 200m 서쪽으로 떨어진 먹이장소를 찾는 것을 학습하게 했다. 다음날 아침에 낯선 환경에다 먹이들을 4 개의 모든 먹이장소
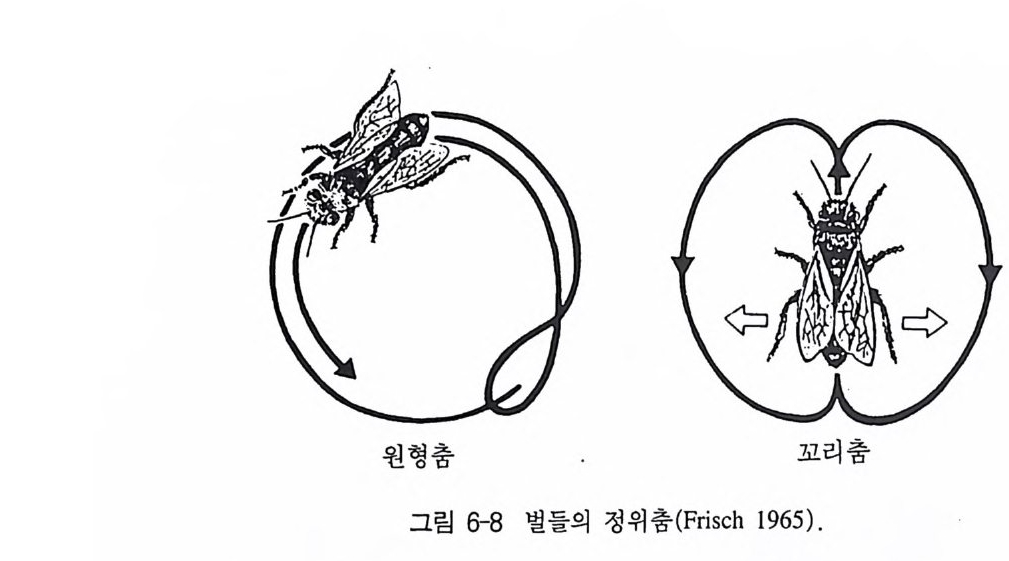 원형춤 꼬리춤
원형춤 꼬리춤
방향에 갖다 놓았다. 이 벌들은 대부분 태양과 반대 방향인 서쪽에 위치한 먹이장소에 도착했다. 따라서 이들은 지형 표지과 관계없이 컴퍼스에 의존해서 정위할 수 있음이 증명되었다. 이 실험에서 이 벌들이 태양울 향해 정위한다는 사실은 배제할 수 없다 이것은 〈꿀벌의 춤〉의 의미가 밝혀지면서 입증되었다. 만일 벌 한 마리가 몇 시간 혹은 며칠이 걸릴 수 있는 인위적으로 갖다 놓은 먹이장소를 발견했다면, 급히 다시 자기의 벌통으로 돌아오게 된다 우리는 벌통에 도착한 이 새로운 일벌에 색깔로 표시를 해주 고 이 벌을 관찰해 보면, 이 벌은 거기서 특이한 춤을 추는 것을 발 견하게 된다(그림 6-8). 만일 벌통의 근처에서 먹이장소를 발견하게 되면, 벌통으로 돌아 오는 벌들은 벌통의 여러 장소를 돌아 다니면 서 원형춤을 춘다. 예를 들어 한두 번 오른쪽으로 원을 그리며 돌다 가 왼쪽으로 돈다. 이 춤추는 일벌 가까이에 있는 벌들은 이 춤추는 벌의 뒤를 따라서 움직인다. 이때 자신들의 더듬이를 춤추는 벌의 꽁무니에 접촉한다. 만일 먹이를 사방 모두에 놓아주면, 원형춤을 통해 신호를 받은 벌들은 벌통의 모든 방향을 돌아다니면서 찾는다. 만일 이 먹이장소가 벌통으로부터 50m 이상 떨어진 곳에 있게 되
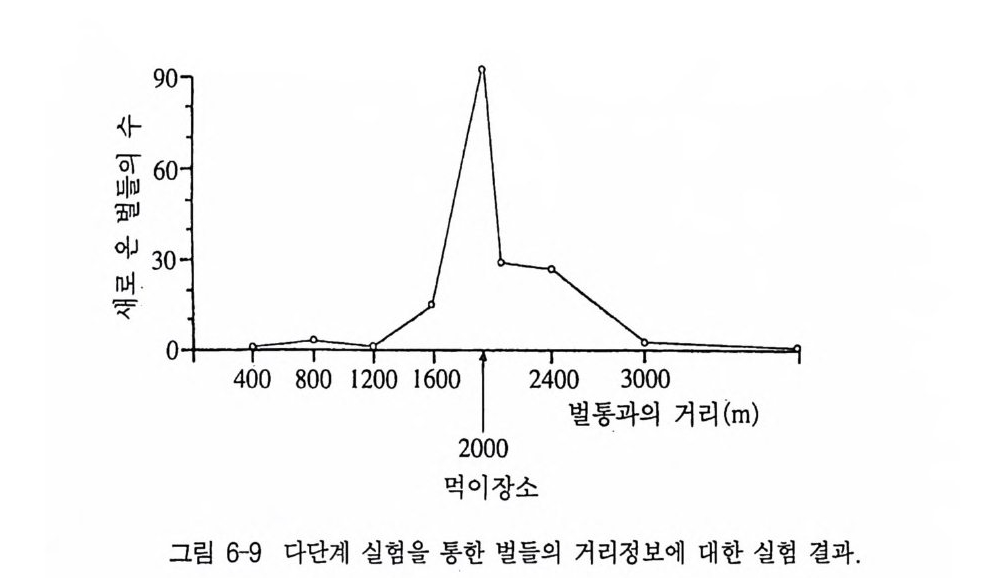 906030
906030
면, 귀가한 이 벌들은 〈꼬리춤〉을 춘다. 이들은 작은 반원을 그린 다음 급히 직선으로 방향을 바꾸어 오던 길로 다시 돌아간다. 이런 식으로 다른 쪽에도 반원이 그려지게 된다. 이 직선의 길에 이들은 복부의 꼬리를 좌우로 혼들면서 움직인다. 원형춤에서처럼 뒤를 따 르는 벌들은 자신들의 더듬이룰 춤추는 벌의 꽁무니에 접촉하여 따 라다닌다 . 꼬리춤을 통해 신호를 받은 벌들은 일정한 거리와 방향에서 먹이 를 찾는다. 이 거리 표시는 다단계적 실험으로 증명될 수 있다(그림 6- 9 ). 표시를 한 적은 무리의 벌들에게 대체로 좋은 먹이장소에서 먹이를 먹도록 했다. 그 다음 이 먹이장소를 농축된 설탕용액으로 갑자기 바꾸어 유혹을 했다. 이때 이 벌들은 벌통에서 열심히 꼬리 춤을 추었다. 그러는 동안 먹이장소의 거리를 다단계로 나누어, 그 러나 항상 같은 방향에다 계속 먹이장소를 설치해 주었다 . 먹이장소 에 도착한 벌들이 자신의 경험이 아니고 꼬리춤을 통해 찾아 오는 가를 확인하기 위해서 새로이 먹이장소에 도착한 벌들은 곧바로 모 두 잡아서 수를 세어 보았다 . 결과 원래의 먹이장소에(꼬리춤을 춘 벌
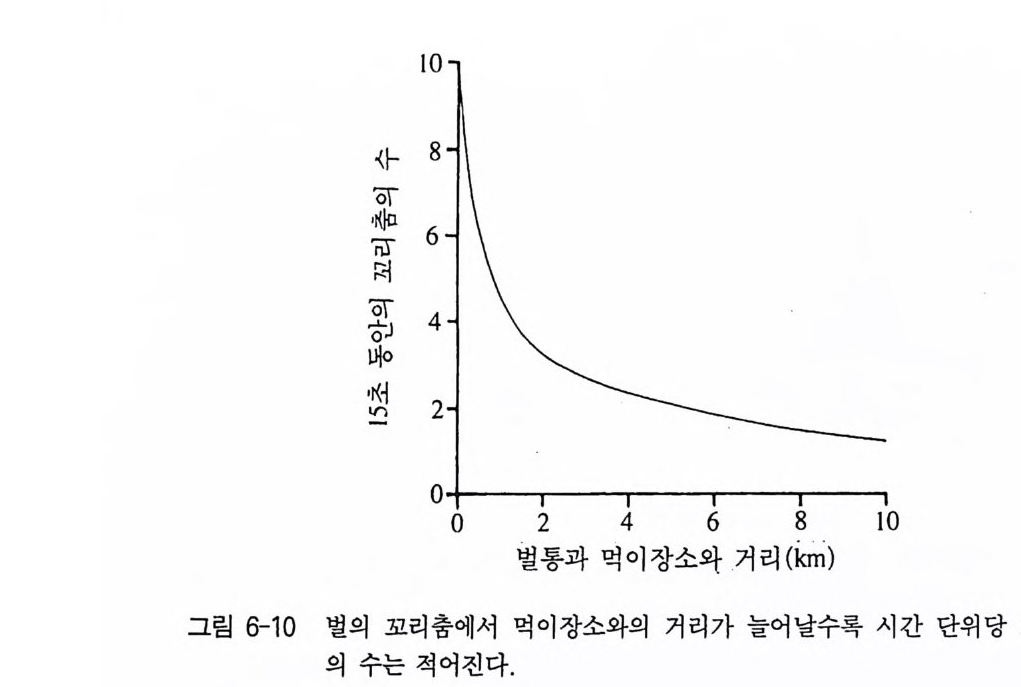 10
10
들이 먹이를 먹었던 곳) 벌들이 제일 많이 찾아 왔다. 이것은 꼬리춤 으로 신호를 받은 벌들이 벌통과의 정확한 거리에서 먹이를 찾고 있 다는 사실을 보여주었다. 결국 이 꼬리춤은 먹이장소의 거리에 따라 서 달라지고 있다. 벌통과 먹이장소가 멀면 멀수록 꼬리춤은 더 느 리게 추게 된다(그림 6-10). 계속된 실험으로 거리 표시로 힘의 소비 가 먹이로 날아갈 때 기초가 된다는 사실을 보여줄 수 있었다. 그리고 바람을 거슬러 날아가야 할 때는 춤을 천천히 추어 정보를 전달한다. 꼬리춤에 내포되어 있는 방향 표시는 부채꼴 실험을 통해 증명될 수 있다(그림 6-11). 다단계 실험에서처럼 벌통에서 꼬리춤이 실시 되면 동시에 새로운 먹이장소들을 부채꼴 모양으로 배치하여 서로 방향을 달리하여 놓는다 . 이것은 꼬리춤으로 신호를 전달받은 대부 분의 벌들이 정확한 거리의 먹이장소를 찾을 뿐만 아니라 정확한 방 향의 먹이장소를 찾는다는 사실을 보여준다. 방향에 대한 정보는 꼬 리춤이 두 개의 반원 사이의 직선(꼬리선)을 그을 때의 정위에 들어
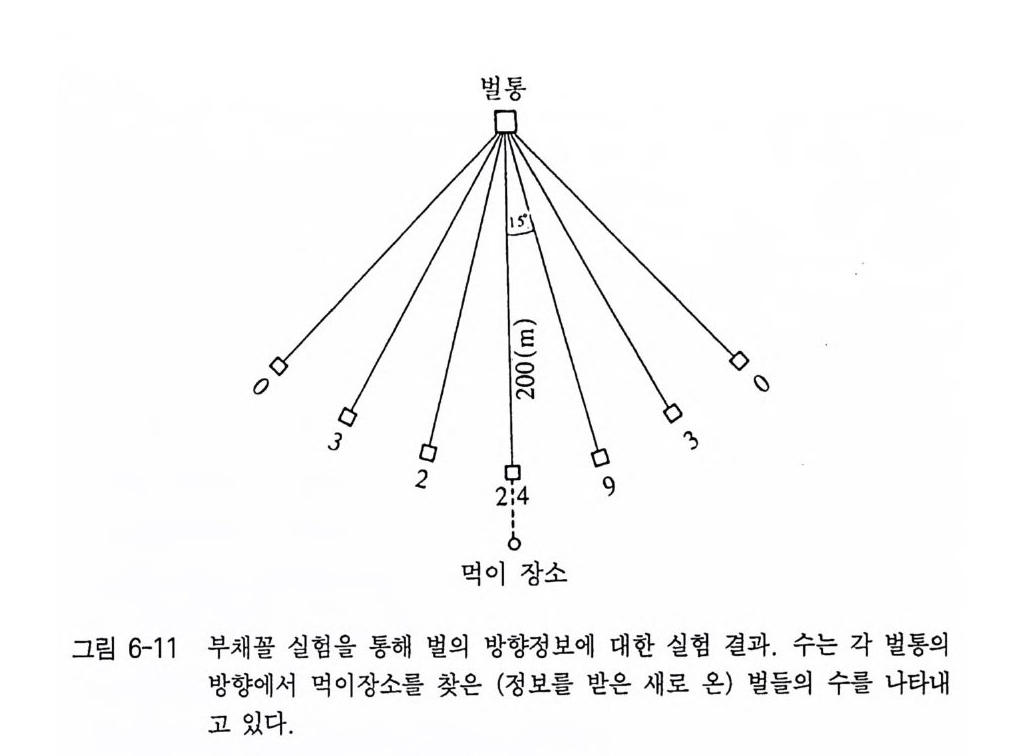 벌통
벌통
있다(그림 6-12). 만일 맑은 날씨에 수평의 면에서, 예를 들어 벌통 의 착륙판에서 춤을 춘다면 꼬리의 진행은 먹이원에 직접 향한다. 이때 아주 흐린 날씨에는 이 춤이 완전히 혼란에 빠져 무정위적이 되는 이유는 반드시 태양이 기준점이 되고 있기 때문이다. 그러나 대부분의 벌들은 맑은 날씨에 수평면에서 춤을 추지 않고, 어두운 벌통 속의 수직 밀랍면에서 춤을 춘다 . 이 경우에도 꼬리 진행은 분 명히 방향을 지시하고 있고, 그것도 중력과 관련해서 정위된다. 이 벌들은 태양과의 꼬리 직선주행 각도를 중력과의 각도로 바꿀 수 있 는 능력을 소유하고 있다. 만일 먹이장소가 태양이 위치한 방향에 놓여있다면 꼬리선은 위로 향하고, 이에 반해 먹이장소가 태양과 반 대 방향에 있다면 꼬리선은 아래를 향한다. 벌들은 태양과 일정한 각도를 이룰 뿐만 아니라 이 각도를 하루의 시간에 맞추어 변화시킬 수 있다. 결국 태양의 궤도가 계산되는 내 부시계가 이 정위에 관여하고 있다. 새로운 둥지장소를 찾아낸 정찰
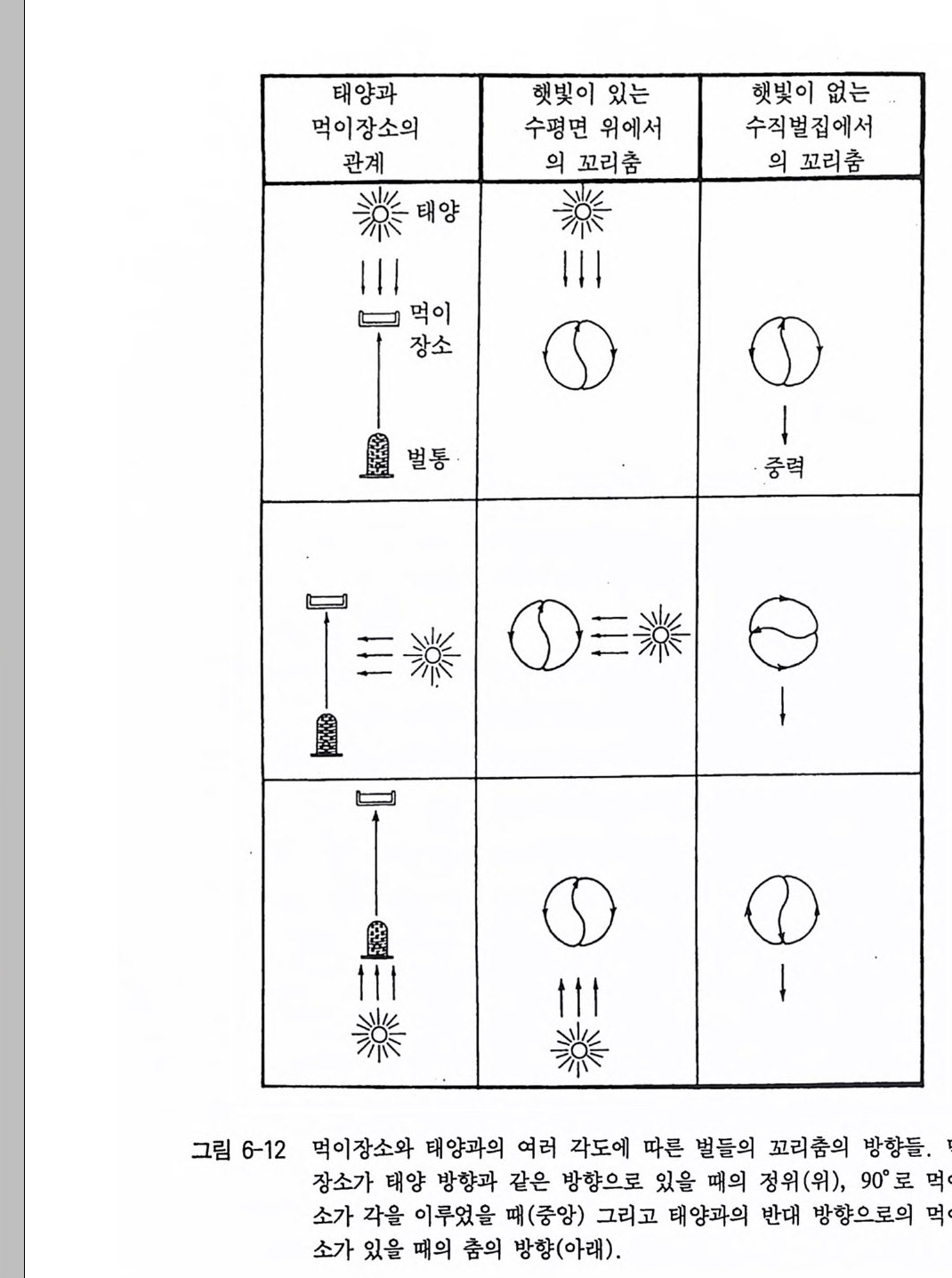
벌들은 어두운 벌통 안에서 여러 시간을 태양을 보지 않고 춤을 춘 댜 이때 이들은 잇달아 변화하는 태양의 방향울 계산하여 여기에 맞추어 중력과의 각도를 수정한다. 항상 일정한 지속적인 빛 조건에 서 벌들을 특정한 먹이시간에 훈련시킬 수 있다. 그렇지만 이것은 대략 24 시간의 리듬이 일어날 때에만 가능하다 . 먹이에 대한 시각지 시자는 서커디언 리듬에서 너무 크게 차이가 나지 않게 한다. 태양 궤도의 계산은 경험에 의존하고 있다. 경험이 없는 젊은 벌 들은 처음에 태양과의 각도에 의지해서만 정위를 한다. 밤에 이전실 험을 해보면 이 벌들은 다음날 아침에 180° 다른 방향으로 정위하게 된다 . 5 일 정도 후에야 이들은 컴퍼스에 의해 정위를 하기 시작한 다 . 이와 같은 현상이 하늘을 보여주지 않은 채 평생 지하실에서 기 른 나이든 벌에서도 나타났다. 이들도 여러날 동안 계속해서 태양 궤도의 단면을 관찰했기 때문에 컴퍼스에 의해 충분히 정위할 수 있 었다 . 먹이장소의 거리와 방향 외에도 춤추는 벌들은 벌통의 동료에게 또 다른 정보를 제공하고 있다 . 먹이원이 풍부하면 풍부할수록 이들 의 춤은 더 오래 지속되며 또 더 열심히 추게 된다. 그 밖에도 꽃냄 새가 이둘의 몸에 붙게 된다. 만일 이 벌들이 꽃가루를 모으면 춤을 따라 추는 동료들에게 그것을 먹게 해서 찾고자 하는 먹이의 냄새에 대해 정보를 전달한다. 찌르레기와는 반대로 벌들의 눈은 자외선에 민감하기 때문에 이들 은 어느 정도 흐린 날씨에서도 정위를 할 수 있다. 아주 흐리거나 먹구름이 꼈을 때라야 이 꼬리춤은 더 이상 정확한 정위를 하지 못 한댜 그 밖에도 이 벌들은 척추동물과는 달리 많은 다른 절지동물 들처럼 태양의 편광에 의해 정위를 할 수 있다. 정상적인 빛은 어떤 면에서도 전달 방향과 비스듬히 진동하는 반면, 편광된 빛에서는 이 진동면이 고정된다. 꿀벌의 눈은 편광에 민감한 특수화된 감각세포
가 존재한다. 푸른 하늘의 빛이 편광이 되면 이 진동 방향이 태양지 점에 따라 일어나기 때문에 이 벌들은 태양이 구름에 가렀을 경우에 도 태양 지점을 판단할 수 있으며 이들의 꼬리춤은 이에 따라서 정 위될 수 있다. 그래서 꿀벌은 태양이 산이나 건물에 가려졌을 때라 도 단 한 점의 푸른 하늘을 볼 수 있는 한 정확하게 정위할 수 있 댜 편광에 따른 정위의 증거로 관찰하고 있는 벌통의 밀람을 수평 으로 바꾸고 원반을 끼워넣어 하늘의 빛을 가렸다 . 이 벌통은 아주 빛이 찰 차단되어 춤추는 벌들이 푸른 하늘의 작은 부위만 볼 수 있 울 뿐이다 . 원판 앞에다 고정시킨 편광 셀로판지는 통과된 빛의 진 행 방향이 하늘의 편광의 진행 방향과 일치하고 있다는 사실에 근거 하여 설치하였다. 이때 꼬리춤은 절대적으로 정확히 정위되었다. 그 러나 빛의 진행 방향이 편광 셀로판지를 다른 데로 돌려 변하면 춤의 정위는 예측 가능한 방식으로 변화될 수 있다 (Wehner 1976, Seely 1985) . 동물계에서 태양 컴퍼스 정위는 널리 퍼져 있다. 옆새우(端脚類)의 일종인 Ta lit rus 는 바다의 모래가 씻기는 곳에서 살며, 만일 파도로 육지로 밀려가거나 혹은 바닷속으로 씻겨들어가게 되면 태양 컴퍼스 를 이용하여 원래의 생활공간을 다시 찾는다. 해안이 남북으로 놓여 있을 경우 이 옆새우는 건조한 땅에서 서쪽으로 도망가고, 이에 비 해 축축한 땅에서는 동쪽으로 도망간다. 이 도피 방향은 각각의 개 체군의 표식이 되며 자연적으로 조성된 해안선에 따라 다르다. 이 태양의 움직임은 방향의 선택에서 보상된다. 바닷가나 개울가에 사 는 다른 절지동물들(단각류, 등각류, 거미류)의 여러 종들은 같은 방 식으로 자신들의 자연 생활공간을 다시 찾는다. 포유류를 제외한 많 은 척추동물들도 커다란 의미에서 태양 컴퍼스 정위를 하는 동물이 라고 할 수 있다 예를 들어 물고기들은 태양의 도움으로 은신처를 찾으며, 도마뱀은 태양이 비칠 수 있는 장소로 다시 찾아 들어 간다.
2.3 그 밖의 컵퍼스 정위 컴퍼스 정위는 태양뿐만 아니라 달이나 별에도 가능하다. Talitr u s 는 낮에 태양을 향해 정위하듯이 밤에는 달을 향해 정확히 정위를 할 수 있다. 이 달의 궤도가 보상(補償)이 되기 때문에 정위는 태양 의 시계뿐만 아니라 달의 시계에도 기초가 되어야만 한다. 혼히 새 들은 밤에 이동을 하며 별의 도움으로 이동 방향을 정할 수 있다. 모형 천체관 실험을 통해 북미산 멧새인 유리멧새 Passer i na cy anea 는 북극성을 기준점으로 삼아 정위하는 것으로 밝혀졌다. 천체를 향한 천문학적 정위 외에도 밤에 이동하는 새들은 자장 컴 퍼스 정위도 발견되었다. 지구 자기장에 대한 정위는 태양 컴퍼스 정위보다 훨씬 더 일반적이다. 이 지각에 대한 생리학적 기작에 대 해서는 지금까지 잘 알려져 있지 않다. 이동성인 유럽붉은가슴울새 Erith a cus rubecula 는 밀폐된 공간에서 지구자장의 총 세기를 변화시 키지 않고 유지시켰을 때 시각적 방향표시자 없이 자신의 자연적인 이동 방향을 유지할 수 있다. 물론 이 새는 이 이동 방향을 별을 갖 고 똑같은 실험을 한 것보다 정확성은 훨씬 떨어졌다. 거의 지구자 장과 같은 세기의 인위적인 자장에서 북쪽 방향의 자석을 변경시켜 이 이동 방향을 다른 쪽으로 돌려놓을 수 있었다. 붉은가슴울새가 별을 향하여 정위할 수 있기 때문에 이 자장 컴퍼스와 별 컴퍼스를 가지고 경쟁시켜 보았다. 이동이 불안한 붉은가슴울새를 자연 별이 있는 하늘의 조건하에 지구자장으로부터 벗어난 북쪽에다 인위적 자 장을 설치하였다. 마침내 이 새로운 이동 방향은 자장의 방향 정보 를 차단했어도 유지되었다. 분명히 별들은 자장의 회전 후에 새로운 방향의 의미를 갖고 있었던 것이다. 이로써 별의 방향에 대한 의미 는 본질적으로 자장 컴퍼스로 인해 달라질 수 있다. 아마도 자장 컴 퍼스가 미리부터 생득적으로 자연적 이동 방향의 유지를 가능하게
한다면, 이 별들은 관련 학습과정 후에 2 차적으로 방향표시자로 사 용될 수 있을 것으로 보인다 (W i l t schko 와 Wi lts c h ko 1988) . 지금까지 이야기한 컴퍼스 정위는 예를 들어 철새의 목적지 발견 을 설명하기 위해서는 충분하다. 철새는 생득적으로 고정된 이동 방 향을 따르며 내부적으로 프로그램화된 이동불안함의 지속과 연결시 키고 있댜 그러나 차량에 실어 운반하는 많은 실험에서 새들을 수 백 킬로미터 떨어진 여러 방향에다 풀어주었을 때 이들은 자신이 태 어난 곳이나 자신이 겨울을 나는 장소를 다시 발견하는 것으로 나타 났댜 전서구(우편배달 비둘기)는 자연에서의 비행 경험이 없는 데다 가 또 아주 짙게 구름이 깔린 날씨에도 다시 돌아온다. 풀어주자 곧 바로 이들은 정확히 최초의 정위를 보여준다. 이러한 귀소실험은 새 들이 진정한 항로 결정 능력을 가지고 있음을 보여주는 것이다. 이 런 능력에 대해 새들은 지도-컴퍼스 개념인 두 단계 절차를 취하고 있는 것으로 추측해 볼 수 있다. 그러므로 새들은 아직 알려지지 않 은 자극 흐름의 기초에 자기가 태어난 곳과 풀어놓은 곳을 비교하여 상황들을 결정할 수 있으며 이로써 정확한 하늘 방향을 찾을 수 있 는 능력을 소유하고 있다. 그래서 이들은 이미 잘 연구된 컴퍼스 정 위의 도움으로 귀소한다. 이 항로 결정 능력의 최초의 단계는 아직 도 다 설명이 불가능 하다. 연어 Oncorhy nchus 는 알을 낳기 위 해 후각을 가지 고 바다에서 모 천으로 다시 돌아온다. 만일 비공을 막으면 이들은 무질서하게 여러 군데의 만물에다 알을 낳는다. 그러나 새들은 후각 능력이 극히 뒤 떨어져 각인되기 때문에 새들에게는 이런 식으로 설명하는 것이 불 가능한 것으로 나타났다 . 그래서 이탈리아에서 행해진 실험에서 전 서구가 후각의 도움으로 항로를 결정할 수 있다는 사실온 아주 뜻밖 의 일이었다. 막혀서 바람에 가려진 큰 새장에서 자란 비둘기는 매 우 엉성한 귀소능력을 보였다. 이들은 생후 일개월 내에 어떤 바람
의 방향에서 특이한 냄새가 나는 것을 배워야만 한다. 만일 이 바람 울 특정한 각도만큼 틀면 성조가 되어 이들은 동일한 각도만큼 툴리 게 정위를 한다. 풀어준 장소에서 이들은 익숙한 냄새를 다시 인식 하여 가려진 컴퍼스 방향을 결정한다. 그 밖의 귀소능력은 태양 컴 퍼스 정위와 자장 컴퍼스 정위를 통해 설명될 수 있다. 많은 실험들 가운데 하나로 어린 전서구에 인위적 올리브기름 냄새의 바람을 복 쪽에서 불도록 했다 . 그런 다음 이 비둘기를 차량에 태워 서쪽으로 이동시켜 이 비둘기의 비공을 올리브기름으로 처리하였다 . 이때 이 비둘기는 마침 북쪽을 향해 운반되어 온 것처럼 정위를 하여 남쪽을 향해 날아갔다. 이런 실험도 새의 항로 결정 능력에 대한 수수께끼 를 아직도 완전히 풀게 하지는 못했다. 지금까지 유럽과 미국에서 전서구의 후각에 의한 항로 결정 능력에 대해 연구를 해왔지만 아직 입증되지 못한 실정이다 (Pap i 1976). 그렇다면 이 항로 결정은 여러 외부조건에 따라 다르며 부분적으로는 아직 알려지지 않은 항로 결 정 기작을 사용하고 있는 것으로 추측할 뿐이다.
제 7 장 행동의 발생 개체발생은 수정된 알세포에서 죽을 때까지의 생물의 발달을 뜻한 다. 행동학 영역에서는 이 가운데 특히 새끼의 행동발달, 즉 태어나 서부터 성숙한 개체로 될 때까지의 과정에서 일어나는 행동에 초점 울 맞춘다. 이러한 제한된 시기 내에서 다음에 열거하는 이유로 인 해 가장 큰 행동의 변화가 일어난다. 첫째, 한 종의 완전한 행동은 점진적인 발달에 의해 이루어진다. 둘째, 어린 개체의 행동에서 계통발생학적 혼적이 남아있을 수 있 으며, 그것은 나중에 다시 소실된다. 셋째, 어린 개체의 행동은 특별히 필요에 의해 이루어진 일련의 전문화된 적응을 나타낼 수 있으며, 마찬가지로 이것은 나중에 사라 진다 이런 행동은 종에 따라 가장 다양하게 나타난다. 그래서 행동의 발생은 발달과정에 있는 여러 종들의 연구에 대해 특별히 유리한 전제조건이 된다. 이에 대한 연구는 두 분야로 나누 어진다. 한 분야는 행동발달 혹은 이때의 성숙과정 및 학습과정에 대해 서술을 하며, 다른 한 분야는 이 발달에 영향을 미치는 의부 및 내부 요인의 분석을 행한다. 최근에는 태아 죽 출산 전의 태아의
행동발달, 예로써 태아의 학습과정에 대해서도 특별한 관심을 갖고 있댜 l 생득적인가 학습된 것인가? 1.1 정보원 행동발달을 위한 유전과 환경의 분담역할론은 실제로 행동학 초창 기 때부터 격렬하게 논의되어 왔다. 이 논의는 인간의 행동에 대해 서도 문제가 되고 있기 때문에 행동학 밖에서도 관심을 갖게 되었 댜 그래서 심리학적 그리고 사회학적 영역에서 이 문제가 논의되어 왔다 이런 과정에서 유전의 의미는 많은 사람들에 의해 확실히 강 조되었으나, 일부에서는 과소평가 내지는 완전히 무시되기도 했다. 최근에는 행동의 유전적 그리고 환경적 역할이 특정 종의 발달에 긴 밀하게 협력하며, 첫째로 두 개의 분리가 실제로 불가능하고 두번째 로 각각의 평가 없이는 발견적 학습에 불과한 것으로 받아들여지고 있댜 게다가 다음의 기본적인 사항이 밝혀지게 되었다. 모든 동물들의 행동은 자신의 환경에 적응되었다. 이것은 사회적 영역의 행동결과 에 적용되는 것처럼 먹이 탐색이나 적의 회피의 동작의 결과에도 적 용된다 행동 목록의 모든 개별적 부분들은 완전한 방식으로 생물 및 무생물 환경의 조건들과 일치하고 있다. 말하자면 새로운 행동생 태학적 연구들은 어느 정도로 이러한 적응들이 사회구조 자체에, 혹은 동물종의 학습 프로그램으로 다가갈 수 있는지를 보여주게 되었다. 모든 적응은 관련 정보를 전제로 하고 있으며, 이것은 근본적으로 두 개의 서로 다른 과정으로 생물체에 도달될 수 있다. 하나는 유전
자를 통해 도달할 수 있고 다른 하나는 감각기관을 통해 도달할 수 있다. 정보는 〈종의 기억〉이라고 하는 게놈에 저장되어 제각기 다음 세대에 계속해서 전달될 수 있고, 혹은 각 세대의 구성원들이 새롭 게 얻어서 각 동물들의 〈개별적 기억〉에 저장될 수 있다. 이것은 근 본적으로 차이가 있다. 이에 대해서도 정보의 출처를 정확히 나타내 는 두 개의 서로 다른 용어를 사용하는 것이 타당한 것으로 보인다. 유전자로부터 기원된 관련 행동형질을 생득적이라고 부르며, 자신의 경험에서 나온 행동을 후천적 혹은 획득된(학습된) 것으로 표현한다. 적응의 출처를 강조하기 위해, 종종 유전적응과 학습적응이라고도 부른다. 행동학 용어 가운데 〈생득적 i nna t e 〉이 라는 말처 럼 일괄성 없게 사 용된 용어도 없다. 그동안 여기에 대해 많은 오해가 있어 왔다. 무 엇보다 종종 이 용어를 관련 행동이 환경과 독립해서 나타나면 의미 를 부여했던 것이다. 물론 여기서 환경의 독립성은 환경이 동물의 행동에 아무런 영향을 미치지 않았을 때를 말한다. 그렇다면 〈환경〉 의 용어와 관련해서 근본적으로 두 가지가 제시되어야 한다. 생물의 발달과정에서 동반되는 일반 환경조건이 있고, 행동형질이 적응해온 특수 환경요인이 있다. 전자의 의미에서는 행동형질이 절대 존재하 지 않는다. 즉 동물의 행동이 완전히 환경과 무관하게 발달할 수 있 는 것을 말한다. 그러나 유전자 자체는 자신의 주변 세계와 끊임없 이 상호작용으로만 영향을 미치고 있으며, 이것은 마찬가지로 조 직, 기관 그리고 생물 전체의 성격에도 적용된다. 이러한 것들의 발 달은 항상 생물체의 내부와 외부의 조건들로부터 영향을 받게 된다. 동물들은 각자 종특이적 적응에 따라 행동하고 있다. 포획행동은 ―번데기가 껍질에서 깨어나오자 마자 먹이를 잡아먹었을 때 세상에 처음 나와서도 정확하게 수행될 수 있으며, 역시 특정 먹이 를 잡는 데 특별한 경험이 요구되지 않는다. 이런 이유로 인해 우리
는 이 경우를 생득적 행동양식이라고 부른다. 이 용어는 여기서 계 통발생과정에서 이미 정보가 그 종의 게놈에 저장되었기 때문에 해 당 환경조건들과 치열한 경쟁을 통해 새로이 얻고자 노력할 필요가 없을 때에만 쓰인다. 그럼에도 불구하고 여기서 환경이 전혀 관여하 지 않는 것은 아니다. 대개의 경우에 일생동안 하나의 생득적 행동도 개인적인 경험을 통해 점차 완전하게 완성된다. 그래서 생득적 개념은 하나의 가설로 쉽게 받아들여지기가 어렵 댜 이것은 환경이 행동형질의 형성에 관여하고 있는 데 그 문제가 있댜 행동의 반응규칙은 각각 유전되어 종종 환경에 노출되어 있게 된댜 그 다음에 어느 정도 수정 범위 내에서 다양한 환경 영향이 어떤 식으로든 유전자에서 나온 정보들이 개별적으로 발현되도록 결 정한댜 이것은 정확히 같은 방식으로 한 생물체의 일반적인 모든 성질에도 적용된다. 무엇보다 학습행동의 여러 영역에서 환경과 유 전자의 끊임없는 상호작용이 분명히 인식될 수 있다. 하나의 예를 통해 이 사실은 좀더 분명해진다. 많은 학습과정은 종마다 매우 차 이가 나는 아주 일정한 연령 기간에만 일어날 수 있다. 여기서도 유 전자는 어떤 나이에 배울 수 있는가를 결정한다. 그러나 이 시간적 한계 내에서 이 학습능력은 언제 개별적으로 실제 학습이 되고, 다 시 말해 그 동물이 관련 경험을 가능한 학습 기간의 처음에, 끝에 혹은 중간에 그의 기억에 저장하는가를 결정한다. ,.2 생득적 행동%모]의 인식 어떤 행동이 생득적이냐의 질문은 실험에 의해서만 결정될 수 있 댜 행동유전학적 연구는 이에 대해 가장 확실한 증거를 제시한다. 그러나 이것은 비교적 적은 동물종들에서만 방법적 기초가 관련되었 을 뿐이다. 정상적으로 영향을 미치는 환경 자극으로부터 어린 동물
울 격리해서 기르면 이런 실험적 방법에 대해 어느 정도 보충을 할 수 있댜 이러한 조건하에 성장한 동물들은 카스퍼-하우저 동물* Kaspa r -Hauser an i mal 이 라고 이름 붙여진다. 실제로 완전히 경험을 빼앗는 것은 불가능하다. 그 이유는 아주 완전히 외부와 차단한다 하더라도 경험을 할 수 있는 여지(예, 자신 의 몸을 가지고)는 항상 남아 있기 때문이다. 설령 완전한 경험박탈이 가능하다 하더라도 발달장애가 분명히 매우 크기 때문에 그 종의 정 상적인 행동과의 비교가 더 이상 불가능해진다. 그래서 카스퍼_하우 저는 우리가 연구하고자 하는 행동영역에서 역할을 하는 자극만을 각각 제거할 수 있기 때문에 문제 발생은 보다 적다. 예를 들어, 우 리가 어떤 종이 생득적 먹이 선택을 하고 있는지를 알려고 한다면, 단 지 그 종의 자연적인 먹이와의 모든 경험을 차단하여 성장한 다음에 도 그 먹이에 대한 자발적 선호를 나타내는지 실험해 볼 수 있다. 이런 실험은 적이나 동료의 인식과 관련되어서도 적용할 수 있다. 이런 경우를 먹이-카스퍼-하우저, 적-카스퍼-하우저, 혹은 사회적 카스퍼 一하우저라고 부른다. 계속해서 감각기관을 차단할 수 있다 . 즉 눈을 가린다든가 혹은 청각기관을 제거해서 귀를 멀게해서 외부 로부터 오는 정보와 격리시킬 수 있으며, 특정 행동양식의 발달은 이러한 자극의 박탈로 억제시킬 수 있다. 〈청각적 카스퍼-하우저〉 의 연구는 동물들의 음성 발달에 유전이 관여하고 있다는 사실을 이 * 카스퍼-하우저 Kaspe r Hauser 라는 이름은 1828 년 독일의 뉘른베르크에 나타난 바 있었던 棄兒의 이름에서 유래되었다. 이 아이의 운명에 대해서는 광범위한 문헌들이 있으나 그의 출신에 대해서 아무도 정확히 아는 사람이 없다. 하우저 가 기억할 수 있었던 정도는 그의 자신의 표현에 의해 어두운 방에 살았다는 것 이 전부다 . 그 아이의 정신적 발달은 생애 동안 극히 제한되어 머물러 있었다. 이런 발달의 장애는 어린아이 시절 충분한 경험을 갖지 못한 데 기인한다 . 그래 서 행동학에서는 어떤 행동이 생득적인가를 알아보기 위해 실험적으로 격리해 서 기른 동물을 카스퍼하우저 동물이라고 부른다.
해하는 계기가 되었다. 물론 모든 카스퍼 -하우저 실험에서 단점도 있다 . 인위적으로 그 리고 다소 격리하여 사육했을 경우-무엇보다 정상적인 사회생활 울 하는 동물의 경우――이 실험동물의 일반적인 상해나 감각기관 에서의 손상이 발생하는 것을 완전히 배제할 수 없는 것이다. 이러 한 상해나 손상은 경험박탈의 특수 효과에 첨가되어 행동발달에도 영향을 미친다. 그래서 이에 대한 실험 결과에 대한 해석은 조심스 럽게 제시되어야 한다. 결과의 의미는 다음 두 가지 가능한 경우에 만 항상 실제로 해석된다. 만일 움직임이나 물체 선택과 같은 행동 형질이 카스퍼_하우저에서도 정상적으로 발달했다면, 확실히 그 행 동은 생득적이라고 추측해도 좋다. 이에 반해 그 행동형질이 없어졌 거나 혹은 비정상적으로 발달되었다면, 반대의 결론으로 단정지을 수는 없는 것이다. 이 결과는 한편으로 학습이 필요한 것으로 결론 울 내릴 수 있고, 다른 한편으로 단순히 사육조건의 결과로 일어난 발달장애가 될 수 있음을 간과해서는 안된다. 많은 경우에 생득적 행동 연구을 위해 경험박탈의 사육이 실행될 수 없도록 되어 있다. 그럼에도 불구하고 구체적인 경우에 생득적 행동을 알아 보기 위해 최소한의 어떤 실마리라도 얻고자 한다면, 이 런 점에서 조심스런 귀납적 추리를 할 수 있는 많은 자료들을 종합 해야만 할 것이다. 실험을 거치지 않고도 다음의 경우는 관찰에 의해 생득적 행동에 의한 것으로 간주할 수 있다. 첫째, 어떤 행동이 형태부동울 통해서 특출하게 나타나거나, 어떤 종의 구성원들이 모두 같은 식으로 그 행동이 수행되며 그리고 의부 자극을 통해서 그 행동의 수행에 영향을 미치지 않을 때, 둘째, 하 나의 복잡한 행동이 첫번 시도에 완전히 나타날 때, 셋째, 관련 형 태적 구조가 완전히 발달하기도 전에 어떤 행동으로 이미 나타날 때
이다. 첫번째 특징은 생득적 행동에 대한 근거로 가장 희박하다. 구애동 작이나 위협동작과 같은 생득적 행동양식들은 실제로 형태부동을 통 해서 특징지어진다. 그러나 학습된 행동도 개체발생 과정에서 이러 한 형태부동에 이룰 수 있다(개체발생학적 의식화). 그래서 생득적 및 형태부동을 동일시 취급하는 것은 금지되어 있다 . 마찬가지로 한 종 내에서 한 형질의 일반적인 분포로 인해 무조건 생득적으로 결론을 내릴 수 없다. 그 이유는 그 형질의 분포가 동일 환경조건에서 같은 방향의 학습과정의 결과가 될 수 있기 때문이다. 무엇보다 이런 현 상은 강제적 학습과정에서 자주 발생한다. 다른 두 개의 분류 기준은 좀 더 근거가 있다. 예를 들어, 한 마 리의 어린 거미가 태어나서 처음으로 이미 수백번 집을 지은 경험이 있는 거미처럼 완전한 집을 지었을 때, 그리고 동시에 복잡한 행동 양식이 연속되어 구체적으로 나타났을 때, 유전적 프로그램을 통해 설명할 수 있는 방법 외에는 달리 표현할 수 없다 . 그리고 움직임이 궁극적인 형태와 크기에 아직 도달하지 못한 기관을 가지고 수행됐을 때, 그래서 행동의 목표에도 도달될 수 없고 연습할 기회도 없었을 때에 이와 똑같이 적용된다. 새끼 거위는 날개 길이가 아직 짧아서 상대방에 전혀 도달할 수 없는 나이에 이미 날개로 공격행동을 취한 댜 형태학적 구조가 형성되기 전에 행동이 아직도 실행되는 않는 경우들은 많지만 이 행동예 관련된 중추신경에는 충격형태가 이미 존재하고 있다. 즉 활동전위의 유도에 의해서 숲귀뚜라미 Gr yll us cam p es t r i s 는 이미 완전 성체로 이르는 마지막 탈피 전에 모든 노래 유형에 대해 완성된 프로그램이 있음이 입증되었다. 의학에서 선천적 병에 사용되는 기준(출산시 어떤 형질의 존재와 결 핍에 의한 기준)은 물론 행동학에서는 완전히 부적절하다. 태어나기 전에 학습과정(태교)이 있다. 이 학습과정에 기초해서 신생아의 행동
은 이미 학습된 요소가 내재되어 있을 수 있으며 , 반대로 생독적 행 동요소들이 성숙과정에서 점차 나타나기도 한다 . 성호르몬에 종속된 행동양식들은 수명이 긴 척추동물과 몇몇 곤충(매미의 경우 최고 17 년 울 산다)에서 비록 그 행동양식들이 부분적으로 혹은 전적으로 유전 적으로 프로그램화되어 있다 하더라도 수 년의 수명 가운데 초기에 나타난다. 1.3 무엇이 생득적인가? 생득적 행동에는 두 개의 가능한 요소가 내포되어 있다. 하나의 운동진행과 이 운동을 유발하고 방향을 정하게 하는 자극의 인식이 유전적으로 이미 프로그램화되어 있을 수 있다 . 첫번째 경우를 유전 성 운동이라고 하며, 두번째 경우를 유전성 감각이라고 한다. 동물 들은 경험에 의해 유전성 감각이 유전성 운동보다 훨씬 자주 발현 되고 있다. 경험이 없이 자란 스컹크는 쥐를 잡고, 던지고, 물고 그리고 혼들 어댈 수 있는 종특이적 먹이 잡는 행동을 보여주지만, 이때 빠져나 가려다가 붙들린 쥐의 몸통 가운데 방금 붙든 그 자리만을 문다. 스 컹크가 점차 어떤 자리를 물면 가장 성공적인가를 배우게 되면 이 물기는 급소를 향한 목덜미 물기로 발전한다. 여기서 행동의 정위도 배워야만 일어난다는 사실을 알게 된다. 어린 오리와 거위는 부화된 직후에 생득적인 행동으로 움직이는 물체를 따라가려는 지향동작을 보이지만, 누구를 따라갈 것인가는 배워야만 한다. 이 경우에 그것 은 습득되어야만 되는 해발자극을 인식하는 것이다 . 이 과정에서 유 전적으로 고정되어 있는 행동양식을 〈무대상 생득적〉이라고 부른다. 물론 이 행동양식은 관련 자극의 인식을 배움으로써 정확히 수행될 수 있댜 많은 간단한 동작들이 생득적이라는 사실온 고정행동 패턴
과 관계를 갖고 있는 데서 알 수 있다. 이것은 여기서 근육의 수축 형태, 죽 개별적 동작의 공간적-시간적 협력이 유전적으로 프로그램 화되었다는 사실을 뜻하고 있다. 그렇게 혼한 것은 아니지만 해발자극과 해발된 행동 간의 관계도 유전적으로 고정되어 있다 . 이 경우에 어떤 한 동물이 이전에 한번 도 만난 적이 없던, 죽 이에 관련해서 학습의 기회가 한 번도 없었 던 자극형태에 정확하게 반응을 한다. 역시 이에 대해서도 일련의 멋진 실험 예들이 있다. 재갈매기 새끼들은 부화 직후에 있는 최초 의 쪼기반응이 이미 여러 날 경험을 한 새끼들과 똑같이 노란색 바탕 에 붉은 반점을 선호한 정위를 보인다. 적을 본 경험이 없이 손에서 자라난 알락딱새 Fic e dula h yp oleuca 는 그럼 에도 불구하고 살아있는 금눈쇠올빼미 A t hene noc t ua 에 대해 전형적인 경계반응을 보이며, 더 욱 부분적으로는 적을 만난 경험이 있는 나이 든 새와 같은 정도의 강한 경계반응을 보인다. 몇몇 북미산 뱀의 경우 먹이선택이 먹이 분포지역에 따라 아주 다양한데, 갓 깨나온 새끼들도 이미 고도의 특이적 먹이선택 능력을 소유하고 있다. 이 모든 경우들을 살펴보면 동물들은 이전에 한 번도 만난 적이 없이도 특정 자극에 대해 어떻 게 행동해야 하는가를 알고 있는 것이다. 유전적으로 프로그램화된 정위반응과 특정 자극에 대한 생독적 인 식에 대한 인상적인 예를 철새와 큰가시고기의 연구에서 발견할 수 있다 북유럽에서 남아프리카까지 이동하는 정원솔새 Sy lv i a bor i n 는 겨울 서식지로의 비행과 번식지로의 귀소비행에 대한 특정한 방향성 을 갖는다. 이것은 실험실에서 손으로 길렀던 한 번도 정위의 경험 이 없었던 동물들에서 증명되었다 (Gw i nner 와 Wi lts c hko 1980). 다른 수컷들에 대해서는 공격행동 신호로 작용하고, 암컷에 대해서는 성 행동의 중요한 신호로 작용하는 큰가시고기 수컷의 붉은 복부의 인 식도 생득적이다. 그 이유는 격리되어 자란 동물들도 정확하게 반응
을 보였기 때문이다 . 이런 표시의 인식에 관련해서 암컷들의 매우 인상적인 자연적 실험이 간단히 이뤄진 적이 있다. 북미의 서부 해 안에 큰가시고기 한 개체군이 살고 있는데 그 개체군 안에는 포식성 물고기에 대해 훌륭한 위장을 하기 위해서 검은색의 단색을 띤 수컷 들이 살고 있다. 이 개체군의 암컷들에게 검은 색깔의 수컷과 정상 적인 붉은 복부의 수컷을 선택하도록 했는데, 완전히 의의의 결과가 나왔댜 암컷들은 검은색의 수컷에 대해 붉은 복부의 수컷들을 5:1 의 비율로 선호하였다. 아마도 이 개체군이 다른 큰가시고기 개체군 으로부터 격리된 이래로 마지막 빙하기에 포식자에 대한 적응이 발 달해 왔기 때문에, 이 검은 수컷의 몸색의 나이가 대략 6000_8000 년으로 추정될 수 있다. 이 암컷들은 이미 수천년 전 다른 방향으로 적응의 이유로 사라져 버린 수컷들의 특징을 오늘날에도 여전히 선 호하여 반응하고 있다. 아직 특이적 자극에 유전적으로 고정된 반응 에 대한 더 좋은 예는 없다. 생득적 신호인식에 있어서 유전자는 응답 자극의 여과를 결정하기 때 문에 , 우리 는 이 것 을 생 득적 해 발기 구 inn ate releasin g mecha- ni sm(IRM) 라고 부른다 해발자극의 성질이 학습되어야 할 때, 생독 적 해 발기 구에 대 해 서 획 득된 해 발기 구 acq ui r ed releasin g mechanis m (ARM) 라고 한다. 이것은 학습을 통해 완전히 새로운 자극 _ 반응 연 결고리가 생겨난다 . 예를 들어 붉은부리갈매기들이 겨울 서식지에서 종종 열린 창문을 통해 먹이를 얻어먹은 경험을 갖게 되었다. 창문 이 열리자 마자 몇몇 갈매기들이 거기서 먹이를 찾기 위해 특이한 소리를 내면서 나는 것이다 . 물속의 폭발 소리도 바닷가 갈매기들을 모여들게 했다 . 그 이유는 이런 폭발이 있자 종종 기절한 물고기나 죽은 물고기들이 물위로 올라오는 것을 보았기 때문이다. IRM 에 대한 표시로 이미 소개된 생독적 행동의 인식 방법 말고도 종종 표시 간소화가 소개된 바 있다. 어떤 동물이 일정한 행동영역
에서 비록 그가 다른 표시들을 여전히 감지하고는 있지만 한 물체의 아주 특정한 표시만을 주의하면, 생득적 반응으로 간주해 볼 수 있 댜 이에 대한 예가 유럽굴뚝나비 수컷의 혼인비행과 큰가시고기 수 컷의 공격행동에 나타나 있다. 그러나 동물들은 제각기 매우 복잡한 IRM 을 갖고 있기 때문에 물론 이 표시방법은 일반적으로 다 적용되 지는 않는다. 유전자가 하등한 동물에서 매우 커다란 역할을 하고 있는 반면, 진 화과정에서 경험은 항상 포괄적인 의미를 갖고 있다. 그래서 순수한 IRM 은 무엇보다 무척추동물에서 찾아볼 수 있으며, 반면 척추동물 에서는 이 IRM 이 보통 적게 나타난다. 즉 개인적인 경험을 통해서 야 척추동물의 IRM 은 완전해질 수 있다. 갓 태어난 유제류의 새끼 들은 처음부터 어미의 앞다리 사이와 뒷다리 사이에서 젖을 찾는 데, 젖꼭지의 정확한 발견은 경험을 통해서 일어난다. 이때 생득적 인 것은 몸의 구석에 해당되는 표시를 인식하는 것이고, 이에 대해 정확한 정위는 시도에 의해 이루어진다. 이 대상 인식과 아주 비슷 한 경우가 추종 각인에서 나타난다.
척추동물의 해발기구는 일반적으로 IRM 보다 더 복잡하다. 그래서 척추동물들은 행동의 개체발생 과정에서 IRM 안에 학습을 첨가시켜 야만 한다. 이러한 해발기구를 〈경험 보충적 생득적 해발기구* EIRM 〉라고 부른다. 학습과정을 통해서 IRM 의 선택력은 증진된다. 경험이 없는 회색기러기 새끼는 움직이는 물체면 무엇이든 따라간 댜 그러나 나중에는 생후 몇 시간 내에 함께 있었던 각인 물체에만 따라간다. 병아리는 처음에는 자기 머리 위로 나는 아무 물체에도 숨는 반응을 일으키지만, 나중에는 자주 나타나는 물체에는 반응을 하지 않는다. 죽 학습을 통해서 병아리의 IRM 은 EIRM 으로 되어 생 * ex경 p e 험 ri e보nc충e 적로 표생현 득할적 수해 발있기는 데구 는여 기영 서어 는로 약in자n a로te rEelIeRasMin 으 g 로 m e사ch용an한is m다 m . od ified by
물학적으로 중요한 의미를 갖는다. EIRM 의 병아리는 하늘에 간혹 나타나는 맹금류에만 숨는 반응을 일으키게 된다. 경험이 없는 까치 는 처음 둥지롤 지울 때는 철사나 텔레비전 안테나와 같은 부적절한 건축재료를 물어오지만, 차츰 어떤 물체가 둥지 재료로 적합한지를 성공과 실패를 통해서 배우게 된다. 이와 비슷한 예로, 청설모는 개 암나무 열매를 보면 처음에는 껍질의 여러 곳을 물어뜯어 상처만 내 지만 여러번 시도를 통해 껍질의 특정 부분에 이빨을 대고 까는 방 법을 습득한다. 마침내 단 한번의 일격으로 열매알을 꺼내먹을 수 있게 된다. 동물 가운데 EIRM 에 대한 가장 연구가 많이 된 예는 명금류의 노 래에서 찾아 볼 수 있다 (Gould 와 Marler 1987, Ba pti s t a 와 Mort on 1988). 노래의 자세한 분석은 사실 대부분의 운동성 행동 패턴에 대한 분석 보다 쉽다. 그 이유는 노래를 녹음하여 소리분석기 sona g ra ph 에다 넣 으면 시간별 주파수로 소리의 파형을 시각적으로 볼 수 있기 때문이 다(그림 7-1) . 실제로 이 소리의 분석은 오히려 언어에 있어서 음절 처럼 설명될 수 있으며 귀로만 구분하기 어려운 소리 유형들을 다양 하게 나타낼 수 있다. 새의 발성은 가장 간단한 음절 no t e 에서 수분 동안 지속되는 정교 한 노래 son g에 이르는 놀라운 다양성을 보여준다. 새의 노래는 조류 중 가장 종수가 많은 참새목 Passe rifo rm 의 참새과 Osc i ne 의 조류 특 징 가운데 하나다. 이 새들의 노래는 그 동안 많은 연구 대상이 되 어 왔다. 이 노래는 많은 다양성을 보여주며 행동 발생의 중요한 기 능적 그리고 진화학적 측면에 대한 연구할 기회를 제공해 주었다 (Slate r 1989). 발생연구에 가장 유용한 것으로 입증된 노래는 되새류 의 중간복합체 가 된다. 푸른되 새 Frin g illa coelebs 는 여 러 유형 의 음 절이 반복해서 이어져 구성된 2-2.5 초의 노래로 되어있다. 각 개체 들은 여러 유형의 노래를 하며, 뿐만 아니라 개체 특이적인 노래를
 __4\c\S I_AB 558288252k-------z H--_ .A\ _Y uh`v _''lh'_ , l\\Aph _u|~L,N_r v \_“\ ,_` lkM ' /A J `,' d세l 시 \ \|\'A사\ ` ,__ J _,,| \ ”w v\ A' u ri_ I.,, ' _hnr '1I '1L_ A\` !' `L,A I\u `` \_ AI,_`I _4A\서、h ’’ 1사 서세 rF' 초 1 7- 1이으 슷한비 살서에은있 3종번고식 지 멧산한국. 림그소나그램 종의새 3 은:붉A. 데 기것으로 시키격리를 하는는 여다 보인식 노래이나, 생가 4). 9Pk 19ar랑과ug : B,새멧(S:쑥새 노, Cn새턱멧뺨
__4\c\S I_AB 558288252k-------z H--_ .A\ _Y uh`v _''lh'_ , l\\Aph _u|~L,N_r v \_“\ ,_` lkM ' /A J `,' d세l 시 \ \|\'A사\ ` ,__ J _,,| \ ”w v\ A' u ri_ I.,, ' _hnr '1I '1L_ A\` !' `L,A I\u `` \_ AI,_`I _4A\서、h ’’ 1사 서세 rF' 초 1 7- 1이으 슷한비 살서에은있 3종번고식 지 멧산한국. 림그소나그램 종의새 3 은:붉A. 데 기것으로 시키격리를 하는는 여다 보인식 노래이나, 생가 4). 9Pk 19ar랑과ug : B,새멧(S:쑥새 노, Cn새턱멧뺨
:[·b wm\\\\\\'\!'/ 642642 N('H _-_一-- l1,\ ),'\[사사사서 MWh' y\^ I~|\ l 『\'i關\\\\\\ )
'古사6 :[ \ NA’ANNAhh- 0.5 1.0 1.5 2.0 시간 (s) 그림 7-2 위로부터 3 개는 푸른되새 노래의 예 . 이 들 은 동일한 기본 형태 를 공유하 고 있으나 세부적으로는 서로 차이가 난다(비록 사람의 귀로는 식별이 어렵지만) . 한 마리의 수컷은 그의 레퍼토리에서 이러한 여러 개의 변이 를 갖게 되며 그들 가운데 어떤 노래는 인접지역에 사는 다 른 새와 공유 하고 있다 . 맨 아래에 있는 소나그램은 유조 시기부터 다른 새와 완전히 격리해서 기론 푸른되새의 노래다 . 이 노래는 푸론되새의 음색과 리듬을 갖고 있지만, 확실히 노래 형태에 있어서 훨 씬 단순하며, 특히 전형적인 푸른되새의 노래인 말단부의 구가 결여되어 있다 (Tho rpe 1961 ).한댜 이렇게 개체 내에서 그리고 종내에서 다양한 노래를 하고 있 지만 이것들은 쉽게 동종으로의 인식이 가능하다(Ki m 와 Park 1993). 이들은 개체 특이성과 종특이성에 의해 이웃에서 노래하는 다른 개 체나 종을 인식하고 있다(그림 7-2). 이런 다양성온 행동발생에 대한 의문을 불러일으키고 있다. 과연 이 런 다양함의 기 원은 어 디 에 있는가? 이 런 의 문은 도프 Tho rp e (1 961)
 매린 카운티
매린 카운티
가 어린 새를 부화하자마자 곧바로 방음장치된 방에서 기른 데서부 터 풀리기 시작했다. 그 이듬해 봄에 수컷들은 격리된 채 노래를 하 기 시작했다. 그들의 노래는 푸른되새로 인식될 수 있었으나 노래의 형태에 있어서 모두가 훨씬 더 단순했으며 개체 내에서 그리고 개체 간에도 변이도 거의 없었다(그림 7-2). 이 실험을 통해 노래의 다양 성은 유전에 의한 것이 아니고 외부 환경의 영향에 기인된다는 사실 이 입증되었다. 이 의적 영향은 어린 새가 둥지에서 그리고 첫 해의 이른 봄에 들었던 성체 새들의 노래였던 것으로 판명되었다. 멧새 과 Embe ri z i dae 의 일종인 북미 산 흰머 리 쑥새 Zonotr i c h ia Leu- cop h ry s 는 이에 대해 좀 더 자세한 정보를 제시하고 있다. 이 흰머 리쑥새는 태평양 연안에 널리 퍼져 살고 있으며, 푸른되새처럼 그들
의 노래도 매우 다양하다. 그러나 이들은 새들이 제한된 구역에서 비슷한 노래를 부르는 경향을 갖는 좀 더 체계적인 다양함을 보여주 고 있다 그래서 이런 지리적 변이를 〈방언 d i alec t 〉 이라고 부른다(그 림 7- 3 ). 만일 어린 수컷들을 부화되자마자 격리시키면 출신 지역과 상관없이 노래를 부른다 . 이들의 노래는 정상적인 노래에 비해 단순 화된 형태로 모두가 매우 유사한 형태를 띠게 된다 . 분명히 이들은 성체 새들의 노랫소리를 듣고 거기에 맞춰 자신들의 간단한 노래 유 형을 변화시킴으로써 지역적인 방언을 하게 되는 것이다 . 이런 학습 과정은 생후 50 일 동안에 일어났다. 수컷들을 그들이 태어난 첫 가 을(생후 50 일이 넘는 시기)에 잡아 혼자 기르면 처음부터 식별가능한 지역적 방언를 시작한다. 이것은 분명히 방언 형성의 민감기가 있음 울 시사하고 있다. 생후 50 일까지의 어린 수컷들을 자신들의 방언 혹은 다른 지역의 방언을 녹음기에 녹음을 해 재생시켜 그 노래를 학습시킬 수 있다. 생후 50 일이 넘으면 이 새들은 녹음된 어떤 소리 에도 학습이 되지 않으며 일단 노래부르는 것이 시작되면 이 새들의 노래는 더 이상 영향을 받지 않는다. 새들은 생후 초기에만 학습에 의해 수정될 수 있는 단순한 노래 형태를 갖는 것으로 나타났다. 어 린 새들은 그들이 듣고 있는 노래를 기억했다가 그들이 처음 노래할 때 그것을 재생산한다. 2 행동양식의 성숙 행동의 개체발생은 이미 언급한 두 정보원들을 두 개의 서로 다른 과정과 일치시켜 왔다. 그것은 유전자에서 넘어온 정보의 실현과 감 각기관에 의한 새로운 정보 입수의 두 과정을 뜻한다. 첫번째 과정 울 성숙이라고 한다면, 두번째 과정을 학습이라고 할 수 있다. 개체
발생 과정에서 이 두 과정은 가까이 맞물려 있다 . 두 과정의 특징과 범위를 밝히기 위해서, 실험적으로 이 둘을 분리할 수 있다. 대부분의 학습과정이 어린 나이에 국한되어 있지 않기 때문에, 행 동의 개체발생과 관련해서 전적으로 학습과정을 설명한다는 것은 그 렇게 바람직한 일이 못 된다. 그래서 학습과정은 다음의 장에서 따 로 다루고 여기서는 성숙과정만울 특징지어 설명할 것이다. 성숙은 행동학적으로 어떤 한 행동양식이 연습없이 완전하게 되는 과정을 말한다 . 어떤 한 행동이 개체발생 과정에서 더 나아져갈 때 그 행동은 성숙을 한다 . 그래서 성숙과정은 어린 동물의 한 행위가 성숙되는 과정에서 완전히 수행하는 것을 방해하여 이 동물의 행동 이 일정한 나이가 된 다음 정상적인 성체의 동료와 비교해 보면 증 명될 수 있다. 새끼 비둘기를 날개를 움직일 수 없는 비좁은 새장에 넣어 기른 다움 그 비둘기가 성체 비둘기처럼 완전한 비행능력을 갖 게 되면 비둘기의 비행능력은 학습이 아닌 성숙과정에 의해 완성된 것임을 입증할 수 있다. 오랫 동안 마취상태로 있게 했던 올챙이가 마취가 풀린 후에 대조구의 올챙이와 똑같이 헤엄을 칠 수 있다면 이것도 성숙과정에 의한 행동이 된다. 카스퍼-하우저로 기른 골든 햄스터 암컷은 생후 80 일이 아직 안된 나이에 처음으로 세상에 나왔 울 때 매우 불규칙하고 너무 작은 둥지를 지었다. 그러나 완전히 똑 같은 조건으로 기른 카스퍼-하우저 암컷들이 첫 임신시 최소 3 개월 이 된 후에는 정상적으로 성장한 동료와 차이가 없는 둥지를 지었 다 . 정위의 능력인 〈목표기구〉는 병아리의 먹이쪼기의 실험에서 보여 주듯이 스스로 성숙될 수 있다. 병아리에게 못이 하나 박힌 점토롤 제시하면, 병아리는 이 가상의 먹이를 향해 쏜다. 첫날에는 못 주변 여러 곳에다 부정확하게 쪼며, 다음날에는 쏜 자국이 점점 좁혀져 결국에는 못대가리를 향해 정확히 쫑 수 있다(그림 7-4). 물론 이것
 a t:. ••반• , . • I I b `닮 I I C• e •• • •, ••. •. ••. • II II d 。 `•\ ••. . ..
a t:. ••반• , . • I I b `닮 I I C• e •• • •, ••. •. ••. • II II d 。 `•\ ••. . ..
은 모두 점토에 찍인 자국으로 판별이 가능하다. 이렇게 쪼기가 나 아져가는 것은 병아리가 못쪼기 실시 사이 사이에 뿌려논 곡식을 쏠 수 있었으므로 정확히 맞힌 쪼기에 대해서는 보상을 받았기 때문 에, 무조건 하나의 학습과정으로 보여진다 . 그러나 명중도가 점차 높아져 가는 것은 7 도 좌측 혹은 우측으로 시야가 옮겨진 프리즘 안 경을 처음부터 씌운 병아리에서도 나타난다. 이 병아리가 존 자국은 못옆 1-2cm 에 놓여졌으며 한 번도 명중시키지 못했다. 이 병아리 는 먹이그릇에서 먹이룰 먹게 했기 때문에 이때 정확하게 맞힌 쪼기 와 맞추지 못한 쪼기 사이에서는 보상의 차이가 생기지 않았다. 그 럼에도 불구하고 이 병아리는 날이갈수록 경험을 가진 대조구 병아 리와 같은 정도로 목표 시야의 범위에 좁혀들어 갔다. 이것은 경험 의존성 및 보상의존성 학습과정이 아니고, 경험과 무관한 성숙과정 이 된댜 즉 증진된 정확도는 쪼기반응에 속한 목표기구의 성숙에 기인하고 있지 학습을 통해 이루어진 것이 아니다 . 이렇게 비록 성숙과정과 학습과정의 기저에 서로 다른 기작이 있 다 하더라도 이 성숙과정과 학습과정은 매우 비슷하게 보일 수 있 댜 비록 이 두 과정이 자연적 관계에서 성숙의 증진에 기여할 수는 있지만, 성숙은 근육의 단련에 있는 것이 아니고, 그렇다고 감각능
력의 증진에 있는 것도 아니다. 바로 성숙은 중추신경의 발달과정에 근거하여 일어난다 . 그래서 성숙과정은 동시에 행동형질에서 일종의 유전적 정보의 전환에 실마리를 제공한다. 물론 하나의 행동양식은 유전자에 의해 한번도 직접 조절된 적이 없다. 오히려 유전자 안에 내포되어 있는 정보는 맨 먼저 중추신경계에서 행동 프로그램을 형 성하게 한다. 그런 다음 이것은 해당 충격 형태에 따라 행동과정을 결정할 수 있다. 이 중추신경의 충격 형태는 많은 경우에서 개인적 경험과 관련해서 더 나아질 수 있다. 즉 환경의 조건에 계속해서 적 응될 수 있는 것이다. 개체발생에서 행동양식이 처음 나타난 것과 병행해서 신경계의 해 당 구조의 분화에 대한 입증이 지금까지 특히 일부의 경우에서만 가 능했다. 멕시코산 도롱뇽의 유생인 엑솔로톨의 행동발생에서 최초의 동작으로 몸을 말아 넣는 반사가 나타난다 . 이 유생의 몸 좌측 전방 에 접촉을 가하면 반대측의 몸통 근육체절들이 수축을 한다. 이 반 사는 척수 안에 들어있는 첫번째 연결 뉴런과 먼저 자리를 잡은 감 각성-운동성 뉴런 간의 결합이 조직학적으로 이루어졌을 때, 처음으 로 정확히 나타난다(그림 7-5). 반사궁이 뒤를 향해 똑같이 발생할 때에, 이미 반사는 계속해서 개체발생학적 발달에서 꼬리 쪽으로 퍼 져나가기 때문에 이 유생은 자극이 한번 가해진 후에 항상 강력하게 몸을 말아 넣는다. 이와 비교할 수 있는 결과가 병아리 배발생에서 나타난다. 병아리 배의 움직임을 달걀의 한 군데에 구멍을 내고 알 껍질에 바셀린을 발라 안을 들여다 볼 수 있게 한다. 첫번째 움직임 은 의부자극 없이도 자발적으로 일어난다. 4 일이 지난후에는 처음으 로 외부자극에 대한 최초의 반응을 확인할 수 있다. 결국 신경계의 운동성 부분은 처음에 발달하고 반응성 동작이 처음 나타남과 함께 신경계의 감각성 부분도 형성된다.
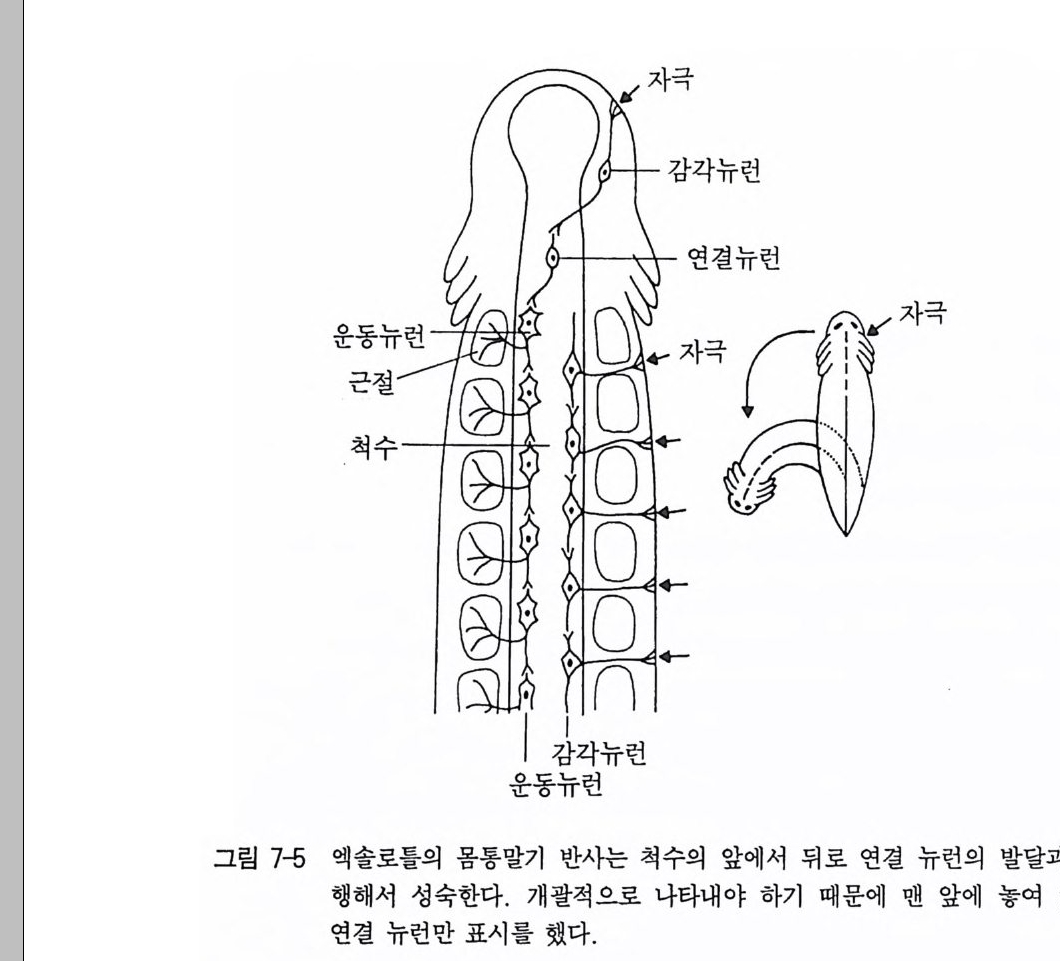 •••K연`r결•극 뉴C런• ( ’L .. 7 • LKt 극
•••K연`r결•극 뉴C런• ( ’L .. 7 • LKt 극
3 개체발생학적 기능 변화 어린 동물들의 행동은 아직 완성되지 않은 성체행동으로 간단히 요약할 수 있는 것이 아니다. 그것은 어린 동물들의 특별한 생활방 식과 요구로부터 설명되는 새끼들의 풍부한 적응을 말한다. 이와 같 은 어린 동물들의 특수 적응을 변형발생 ceno g enes i s 이라고 부른다 . 이것은 완전변태 곤충들의 유충들의 경우 특히 극단적으로 형성된 다. 이 곤충들의 유충들은 종종 성체에 비해서 완전히 다른 생활공
간에서 살며 근본적으로 다른 영양을 섭취한다 . 예를 들어 나비의 번데기는 나뭇잎 위에 살면서 저작성 구기( □ 器 )로 먹이룰 먹는다. 이에 반해 성체 나비는 먼 거리를 이동하면서 꽃을 찾고 홉입성 대 롱 모양의 구기로 꽃꿀을 먹는다 . 이런 차이는 번데기 시절에 신경 조절기작의 기본적 구조 변화에 기인된다. 다른 변형 발생은 그다지 극단적인 것은 아니지만, 어린 동물들의 생존을 위해서는 그 의미가 줄어드는 것은 아니다. 해안 조류의 새끼들은 위험에 처하게 되면 꼼짝않고 조용히 있는다. 보호색으로 자기 몸을 보호하며, 나중에는 적으로 부터 뛰고 혹은 날아서 달아난다. 이에 비해 성체가 되면 이 들은 주변을 경계하면서 소리롤 낸다. 새끼를 돌보는 종의 어린 동 물들은 어미와의 사회성 결합을 발달시켜 왔으며, 이것을 위해 특정 한 신호동작으로 먹이를 구결한다 . 어린 동물들의 많은 행동양식들은 개체발생학적 과정에서 사라진 다 . 그러나 남아 있는 행동도 더러 있지만 그것들은 개체발생학적 기능 변화에 따라 성체 동물들의 경우 새로운 의미로 나타난다. 예 를 들어 어린 동물의 구결행동에서 암컷의 구애행동으로 변할 수 있 댜 괭이갈매기 새끼들의 먹이 구결의 행동은 성체 암컷이 수컷에게 구애할 때와 같다. 암컷이 수컷 앞에서 머리를 들어올리는 행동에서 구결행동이 구애행동으로 변화된 것을 발견할 수 있다. 강아지가 젖 먹기 위해 목을 틀어 위로 향하게 하는 행동으로부터 공격적인 상황 에서 성체의 복종행동으로 상체를 낮추고 앞다리를 들어 올리는 행 동으로 발달한다. 가끔 개체발생학적 기능 변화는 직접 일어날 수 있다. 어린 나무 발발이 Cer t h i a fa m ili ar i s 의 구걸 소리 로부터 어 미 로부터 독립 한 다음 에는 점진적으로 안도음(引 導音 , 다른 개체에게 자기 위치를 알려서 인 도의 역할을 하는 소리)으로 발달한다(그림 7-6). 그렇지만 나이든 개 체의 구걸음을 만드는 신경구조는 그대로 유지하며, 그 소리는 암컷
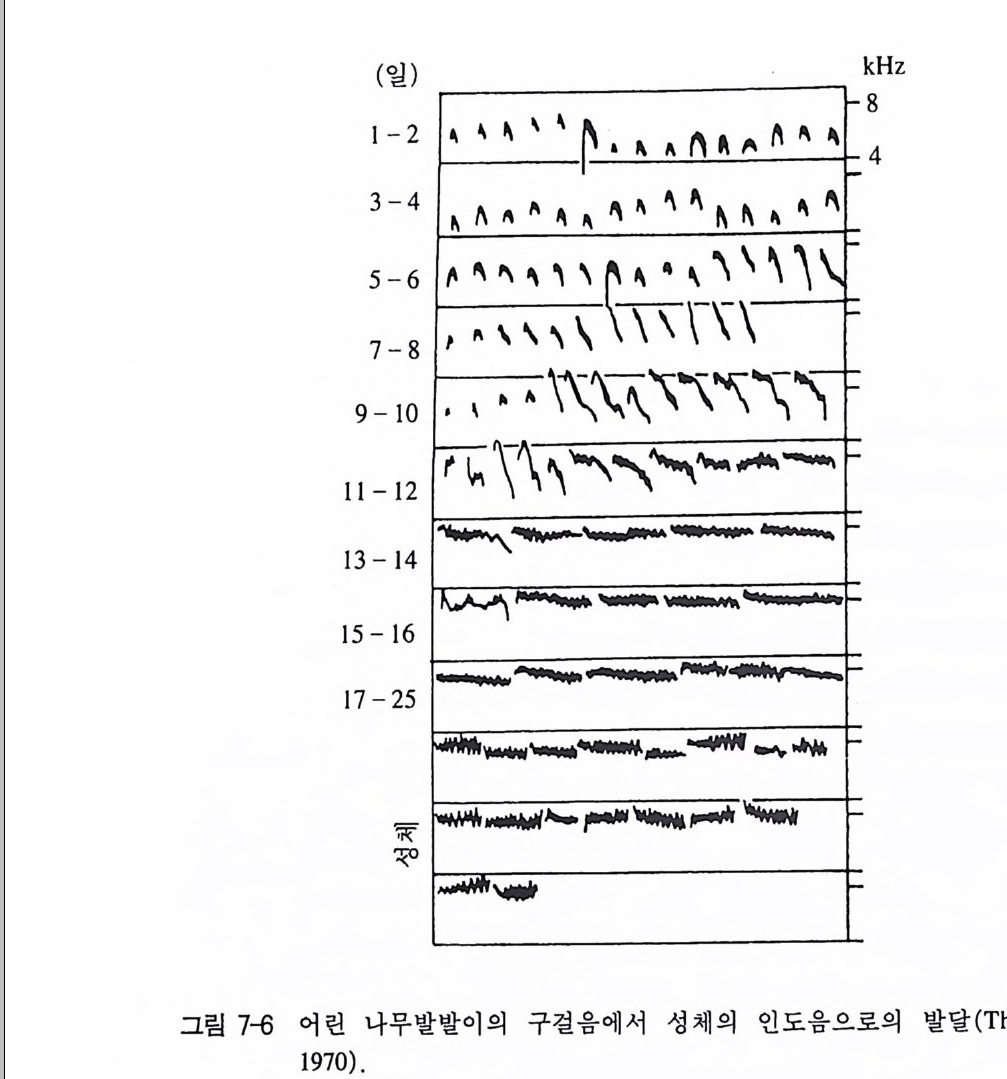 (일) kHz
(일) kHz
이 수컷에게 구결할 때 다시 암컷에게 나타난다. 비슷한 식으로 어 린 정원솔새의 입벌리기에서 점진적으로 멍어리식의 위협으로 발달 한댜 두 마리의 어린 새가 둥지에서 떠나 날기 시작한 후에 뜻밖에 만나게 되면, 비록 그들이 이러한 순간에 양육자로 부터 먹이룰 취 할 준비가 이미 되어 있지 않더라도 그들은 격렬하게 의쳐 서로 입 울 벌린다. 이런 어린 새의 상호 입벌리기에서 점차 부리를 벌린 채
멍어리위협으로 발전한다 . 입벌리기와 멍어리위협은 대략 생후 17- 50 일째의 나이에 같은 의미로 함께 나타난다. 나중에는 단지 멍어리 위협만이 관찰된다. 역시 이런 경우에도 입벌리기를 만드는 신경구 조는 그대로 유지되면서 암컷이 성적인 성숙에 도달한 후에 구애행 동으로 다시 나타난다. 위에서 든 예들의 행동양식에서 기능변화는 형태변화와 관련되어 일어나고 있다. 그러나 하나의 동일 행동양식이 개체발생학적으로 변화되는 것인지 또 개체발생 과정에서 성체의 새로운 행동양식이 발달되어 어린 개체의 행동양식이 새로운 행동양식에 의해 완전히 없어질 때까지 이 행동양식들과 서로 엇갈려 오랫동안 나타나는 것 인지 아직도 완벽하게 확인할 길은 없다. 분명한 것은 행동양식의 형태가 고정되어 있을 때, 개체발생학적 기능은 변화된다는 사실이 다. 아직 완전히 날지 못하는 붉은부리갈매기는 먹이를 가져온 어미 에게 머리를 숙이고 가까이 다가선다. 이 행동 때문에 어미의 회피 경향은 밀고들어 오는 이 새끼에 대해서 억제된다 . 독립된 다음에는 이 행동은 거리를 두고 위협하는 행동으로 변한다. 이 행동으로 어 린 갈매기는 첫번 맞는 겨울철에 먹이를 먹는 동료들을 몰아낸다. 나중에 이 붉은부리갈매기는 다른 위협행동을 선호한다. 그렇지만 머리를 숙이는 자세는 완전히 사라지지 않는다(그림 7-7).
 그림 7-7 성숙한 붉은부리갈매기의 머리 숙이는 자세 . 어미로부터 독립하기 전에
그림 7-7 성숙한 붉은부리갈매기의 머리 숙이는 자세 . 어미로부터 독립하기 전에
4 호기심과 놀이행동 척추동물들은 이미 잘 알려진 물체보다 새로운 물체에 종종 완전 히 달리 반응을 한다. 쥐는 처음에 조심스럽게 새로운 물체를 탐색 울 한다 . 이때 쥐는 살금살금 새로운 물체에 접근한 다음 한발짝 뒤 로 물러선다 . 그 다음 다시 그 물체에 다가가서 쿵쿵대며 냄새를 맡 고 맛을 본다. 이 호기심 행동은 동물들이 주변을 정확히 알게 하여 획득한 학습 경험을 통해서 자신의 생존기회를 확대시켜 준다. 호기 심은 새로운 먹이원을 발견하게 하며, 새로운 위험을 배우게 하고 은신처나 둥지롤 짓는 데 유리한 장소를 인식할 수 있게 한다 . 척추 동물들의 발달과정에서 호기심 행동은 물고기에서 포유류로 발달하 면서 점점 강력하게 발달되어 일어나게 되었다. 포유류는 가장 고도 로 발달된 학습동물일 뿐만 아니라 최소한 어린 시절에 유난히 호기 심을 가진 동물이 된다 . 대부분의 행동양식들이 특정 신호자극에 의 해 유발되고 있는 반면, 호기심 행동에 대한 해발기구는 극히 불특 정적이다 . 반복된 해발인은 습관화로 불리는 반응의 자극 특이적 억 제로 변한다. 이 습관화는 다른 많은 행동양식들에서도 알려져 있지 만, 탐색의 경우에서는 특히 두드러지게 나타난다. 시간이 조금 지 난 다음에는 그 대상 물체를 더 이상 주의하지 않게 되며, 이 탐색 행동은 다시 새로운 물체에 의해서 비로소 해발될 수 있다. 호기심 행동이 연장되어 놀이행동으로 들어갈 수 있거나 혹은 놀이 행동의 구성분으로 포함될 수 있다. 두 행동형태는 성체보다 어린 동 물에서 훨씬 더 두드러지게 나타나 있다 . 놀이 유형의 분류는 많은 과도적 행동과 혼합행동이 있기 때문에 간단하지 않다. 놀이는 종특 이적 행동양식, 예를 들면 공격행동의 요소나 먹이획득의 요소를 더 많이 포함하고 있거나, 혹은 개체적 학습 요소로 대부분 구성되어 있 댜 이것을 종특이적 놀이와 개체특이적 놀이로 분류할 수 있다.
4, 1 종특이적 놀이 종특이적 놀이는 예를 들어 여러 종류의 이동행동과 다른 일반적 몸동작행동을 시험삼아 해보는 동작놀이가 있다. 어린 유제류는 거 의 무진장한 동작충동을 소유하고 있다. 영장류는 자신의 동작 형태 에 있어서 다른 동물들보다 덜 고정되어 있으며 놀이할 때 자신의 몸동작에서 아주 두드러진 독창성을 보여준다(그림 7-8). 싸움놀이
 -.::-,-:. 십.. .——
-.::-,-:. 십.. .——
 그림 7 선 어린 붉은여우의 싸움놀이 .
그림 7 선 어린 붉은여우의 싸움놀이 .
는 식육류에서 특히 잘 나타나 있다(그림 7-9). 강아지들은 눌 서로 맞잡고 싸우는 행동을 벌인다. 놀이적 싸움에 대한 특징으로 절대로 물지 않는 다는 것과 빠른 역할의 전환을 들 수 있다. 하위자는 잠 시후에 상위자가 되며 상위자가 다시 하위자가 되는 것이다 . 식육류 에서뿐만 아니라 많은 다른 어린 포유류에서 이 종특이적 싸움행동 이 놀이에서 일찍이 행해졌다. 어린 염소나 영양들은 예를 들어 앞 다리로 위로 뛰어올라 바닥에 머리를 아래로 해서 떨어지거나 그리 고 머리를 부딪치기도 한다. 싸움놀이와 도피놀이의 경계는 따로 있 지 않고 종종 연결되어 일어난다. 싸움놀이에서처럼 도피놀이는 추 적자가 있고 도망자가 있게 마련이다. 그러나 종특이적 공격행동의 이 가짜 싸움이 도피행동에서도 들어 있다. 근본적인 요소는 도피를 통해서 싸움을 피하는 것이다. 이것은 아이들의 전쟁놀이에 해당된
다 싸움놀이에서처럼 이 도피놀이에서도 놀이룰 하는 동물들의 역 할은 빨리 변한다. 도망자가 추적자가 되고 또 곧바로 추적자가 도 망자가 된다. 포획놀이는 성체의 종특이적 요소를 내포하고 있으나 종종 먹을 수 없는 물체를 대상으로 이 포획행동을 한다. 어린 정원 솔새는 예를 들어 잎이나 가지를 부러뜨리고 곤충과 같은 대상물을 부리로 물어 던지고 그것을 반복해서 혼들어 댄다. 이렇게 해도 이 벌레가 죽지 않으면 그것을 다시 취하여 놀이를 계속한다. 개과 혹 은 고양이과의 식육류는 종종 형제들을 대상으로 먹이포획놀이를 한 댜 그다음 이것은 싸움놀이로 발전하게 되어 이따금 이 두 놀이유 형간의 분명한 경계를 짓는 것이 어려워진다. 생식놀이와 양육놀이 에서 어린 동물들은 종종 성적인 성숙이 이루어지기 오래전부터 말 타기나 둥지짓기와 같은 성체의 행동양식을 놀이하는 식으로 보여준 다 . 어린 정원솔새는 이미 생후 4 주의 나이에 놀이하는 식의 짝짓기 혹은 집짓기를 시작한다. 망아지, 송아지 그리고 강아지들도 교미할 때처럼 말타기를 한다 . 수컷이 하는 교미행동은 놀이 시기에 암컷에 서도 발견된다. 4.2 개체특이적 놀이 동물이 주변으로부터 획득한 경험을 조건으로 한 적응을 위해 개 체특이적 놀이는 특히 의미가 있다. 이 놀이에서 학습을 통한 개별 적 획득 행동요소들이 눈에 띄게 나타난다 . 가장 발달된 놀이가 조 류에서 나타나지만, 이것은 특히 고등한 포유류의 행동 발달을 위해 서 커다란 의미를 갖는다. 창조적인 요소와 시험적 성질은 개체특이 적 놀이에서 특히 분명하게 나타난다. 각 동물들은 새로운 놀이를 찾아 그것을 시험해 본다. 다른 개체는 놀이를 하는 자기 동료를 관 찰하고 그를 모방한다 . 종특이적 놀이는 자연에서의 시험적 놀이가
무시될 수 있다. 손으로 기른 4 마리의 어린 정원솔새의 먹이포획놀 이 도중에 우연히 돌멩이 하나가 빈 유리그릇에 쌩그랑 소리를 내면 서 떨어졌다. 이때 이후로 이 새들은 돌맹이를 위를 향하여 점점 자 주 가지고 가서 그것을 떨어뜨리고, 그 그릇을 홀깃 쳐다본 다음 돌 맹이가 그릇에 부딪치는 맑은 소리를 가만히 듣는 것이었다. 처음 소리가 나자 이 새들에서 다시 움직임이 일어났다 . 2 주가 지난 다음 에 그들은 아주 다양한 물건들을 시험해 보는 놀이로 바꾸기 시작했 다 어린 침팬치들은 가능한 모든 물건들을 조작하면서 연장으로 적 합한지를 시험을 해본다. 이러한 식으로 그들은 예를 들면 상자를 위로 쌓아올리거나, 막대기를 서로 연결시키고 혹은 닫힌 상자를 드 라이버로 여는 것을 배운다. 먹이보상이 요구되는 것은 아니다. 그 러나 나중에 이들은 먹이를 얻기 위해서 자신들이 놀이하면서 획득 한 재주를 활용할 수 있다. 동물들이 놀면서 배운다는 많은 증거들이 있다. 고등한 동물들 죽 포유류나 조류는 성체의 생활에 필요한 경험들이 획득되어야 할 때, 분명한 놀이행동을 보여주고 있으며, 더욱이 어린 시절에 이 놀 이행동이 대부분 나타난다. 자연에서 시험적 놀이의 의미는 잘 알려 져 있다 인간의 아이뿐만 아니라 어린 침팬지들도 진지한 상황에서 보다 놀이에서 많은 것을 더 쉽게 배운다. 종특이적 놀이가 완전히 적응된 종특이적 행동양식의 발달을 위해 중요한 전체조건인지는 지 금까지도 아직 미해결된 과제의 하나로 남아 있다. 4.3 놀이행동의 성격 놀이행동의 성격은 다음 네 가지로 요약될 수 있다. 첫째, 놀이는 진지함이 결여되어 있다. 포획놀이는 종종 대용물이 그 대상이 된다. 예를 들면 새끼 고양이가 실꾸리를 갖고 노는 것이
될 수 있다 도피놀이를 하는 쥐는 집에서 곧바로 다시 나오지만, 정 말 위험할 때는 더 오랜시간 동안 기다리다가 안전하다고 판단되면 조심스럽게 집을 나온다 . 싸움놀이를 할 때 쥐들은 상위자가 하위자 의 역할을 하기도 하고 또 하위자가 상위자의 역할을 하기도 한다. 이것은 승리가 중요한 것이 아니기 때문이다 . 놀이의 활동성은 특정 목적을 갖지 않으며 따라서 진지한 상황에서의 같은 활동성과 구분 된다 가끔 놀이행동은 진지함이 결여된 종특이적 신호행위를 동반 한다 예를 들면 침팬지의 놀이표정을 들 수 있다. 둘째, 개별적 행동 요소들이 진지한 경우보다 놀이에서 더 자유롭 게 결합될 수 있다 . 여러 행동영역에서 나온 요소들은 혼합되어 있 을 수 있다. 예를 들면 싸움놀이와 포획놀이가 혼합될 수 있다. 그 러나 이 놀이행동의 요소들은 우연히 연속되어 일어나지 않는다. 진 지한 경우의 전형적인 행동 연속이 이미 놀이에서도 인식될 수 있다. 셋째, 생물학적으로 중요한 행동준비는 이 놀이행동을 억제시킨 댜 그래서 이 행동 준비가 절대적인 우선 순위를 갖는다 . 배고품과 위험에서 동물들은 놀이를 하지 않다 . 사육 상태에서 그들은 종종 먹이를 향하여 놀이룰 하는데, 이것은 배가 부를 때 먹이를 갖고 놀 이를 하는 안간의 애기들과도 비슷하다. 많은 동물들은 야생 상태보 다 사육 상태에서 훨씬 더 강렬하게 놀이를 한다. 그 이유는 동물들 이 빨리 배불리 먹을 수 있고 야생생활에서의 위험이 사라졌기 때문 에 긴장이 풀리는 상황에 훨씬 자주 처하게 되어 놀이를 시작한다. 넷째, 놀이는 완료행동을 위하여 애쓰지 않는다. 놀이행동은 다른 행동준비가 활성화되어 방해하지 않는 한 거의 무진장한 것이다. 고 양이의 포획놀이는 먹이를 재빨리 붙잡는 것으로 끝나지 않고 다음 순간 다시 그것을 놓았다가 붙잡는 것이다 . 지금까지 놀이준비가 빈 번한 완성을 통해서 어떻게 고갈되는가에 대한 명확한 증거는 없다.
4.4 놀이행동의 기능 놀이행동은 높은 에너지 소모와 위험(예, 상처)과 결부되어 있다. 많은 동물에서 놀이행동이 자주 광범위하게 퍼져 있다는 사실과 고 도로 발달된 행동양식 및 특수한 의사소통 형태가 진화과정에서 나 타났다는 사실은 생물학적 가치를 지니고 있음을 짐작게 한다. 그리 고 진화과정에서 놀이행동으로 발달을 하게 해온 선택압이 작용했음 울 알수 있다. 물론 항상 명확히 분리할 수 없는 두 개의 가능한 의미영역도 근 본적으로 구별하여 설명할 수 있다. 하나는 성장중인 개체를 위한 직접적인 영향이고 다른 하나는 성체를 위한 장기적인 의미다. 첫번 째 언급한 영역에 대해 지금까지 단지 조심스러운 추측이 가능하다. 죽 놀이의 움직임을 통해서 근육의 발달과 놀이할 때 입수한 감각인 상을 통해서 감각기관과 신경계에서의 성숙과정이 긍정적으로 영향 울 받을 수 있다 . 그래서 놀이와 성장의 속도 사이에 직접적인 관계 가 성립된다. 두번째로 놀이행동의 장기적인 의미는 다음의 영역에 까지 확대될 수 있다. 첫째, 운동성 훈련 다시 말해 근육기능의 단련으로까지 확대될 수 있다 둘째, 사회화 과정 죽 사회적 배우자의 개별인식으로의 능력을 포 함한 사회적 역할의 습득, 자기 공격의 통제 그리고 사회적 유대의 발달에까지 확대될 수 있다. 셋째, 인지발달 죽 일반적인 감지능력의 훈련과 증진에까지 확대 될수있다. 물론 이 세 개의 기능영역들은 칼로 자르듯이 서로 분리할 수 있 는 것은 아니다. 모든 세 영역에 해당된 각 놀이행동들이 분명히 같
은 의미를 갖고 있는 것도 아니다. 그러나 대부분 놀이행동은 가장 광범위한 의미에서 경험들의 수집에 기여할 수 있다. 그것은 학습능 력이 전제가 되고 있고, 또 어린 개체의 학습욕구가 놀이기간 동안 특히 크다는 사실을 의미하고 있다 (M arti n 과 Caro 1985).
제 8 장학습 학습은 행동학에서 모든 환경조건에 행동의 개체적응으로 이끄는 전 과정으로 요약될 수 있다. 학습은 감각기관을 통한 정보의 입 수, 기억 그리고 필요할 경우 소환되도록 한다. 학습과정은 단세포 생물에서 척추동물에 이르기까지 모든 동물계에서 밝혀져 있다. 우 리는 일반적으로 어떤 특수한 자극상황에 처해있는 행동이 이전에 만난 결과로 인해 달라지는 사실을 보게 된다. 이렇게 같은 상황에 서의 변화는 바로 학습이 관여하기 때문에 일어난다. 그럼 여기서 동물의 학습행동에 관련해서 다음 네 가지 간단한 질 의로 시작해 보기로 하자! 물론 이에 대한 대답이 이 장에서 주로 다루게 되는 내용들이다. 동물들은 왜 배우는가? 동물들은 무엇을 배우는가? 동물들은 언제 배우는가? 동물들은 어떻게 배우는가?
1 생물학적 의의 획득된 정보전달과 유전적 정보전달의 장단점에 대해 생물학적 평 가를 내리려면, 두 상황간의 근본적인 차이를 구별해야 한다. 각각 정보 전달이라는 한 종류만이 가능하기 때문에 두 상황이 가끔 외부 와 경쟁하는 경우가 있고, 이론적으로 두 상황이 함께 실현되는 경 우 이것은 진화과정에서 서로 경쟁을 해왔다. 첫번째의 상황에 대한 예는 가족 구성원과 무리 구성원의 개별 인식에 기초한 상황을 말한 댜 이 개별 인식은 단지 학습과정으로 가능하다. 두번째의 예는 한 세대가 다음 세대의 출현 전에 이미 죽어버리는 모든 종에 해당된 댜 그래서 이런 종들의 경우 도피반응, 특정 둥지짓는 기술 혹은 동종의 배우자 인식과 같이 각 개체가 경험적 확인을 통해 얻어질 수 없는 모든 성질 등은 유전에 의해서만 전달될 수 있다. 이런 점 에서 많은 곤충류와 다른 무척추동물들은 글자 그대로 사회적 카스 퍼_하우저로 성장한다. 이에 반해서 대개의 경우 정보는 유전 및 학습에 의한 두 경로로 획득될 수 있다. 일반적으로 학습과정은 광범위하게 넓게 퍼져 왔기 때문에 진화과정에서 이 학습과정은 유전적 정보보다 더 최근에 획 득되기 시작했다. 그래서 이 학습과정은 유전적 정보에 대해서 선택 적 장점을 가져야 했다. 이 선택적 장점으로 인해 학습과정은 계통 발생학적 발달과정에서 확고한 자리를 차지하게 되었다. 이 장점은 모든 학습과정에서 똑같이 나타나는 것같이 보인다. 즉 그것은 획득 된 정보가 더 큰 적응성을 나타내고 있다. 어떤 한 종의 환경이 끊 임없이 변화를 하고 있다고 하자. 이것은 무생물적 환경(예, 기후의 변화)과 생물학적 환경(적이나 먹이의 출현과 사라짐)에 같은 식으로 적용되며 외관상 돌연변이를 일으킬 만한 환경 변화룰 의미하고 있 댜 이런 각각의 변화는 학습과정의 범위 안에서 동시대의 세대 내
에서도 일어날 수 있으며, 이에 반해 유전적 프로그램의 범위 안에 서는 새로운 적응이 한쪽 방향을 향해 임의로 나타나는 돌연변이에 근거로만 항상 가능하다. 여기서 검정 복부의 수컷에 반응을 보인 큰가시고기 암컷들의 이야기를 다시 한번 언급할 필요가 있다. 이것 은 생득적 해발기구의 보수성을 보여주는 좋은 예가 된다. 먹이선택 에서 이런 차이는 더 분명하게 나타난다. 어떤 한 종이 한두 개의 먹이 종류에 전문화가 되어 그 종류에 생득적으로 인식을 하게 된다 면, 이 먹이가 사라졌을 경우에 그 종은 멸종하게 될 것이다 . 그러 나 그 종이 관련된 학습능력을 최대로 활용한다면, 언제라도 전환이 가능하게 된다 . 그러므로 계통발생학적 일반 경향이 학습능력의 끊임없는 강력한 발달에 분명히 목표를 두고 있다는 사실은 놀랄 일이 아니다. 죽 이 경향은 유전적 프로그램화를 지배하고 있는 〈폐쇄적 발달 프로그램〉 대신에 개인적 경험을 위해 충분한 놀이공간을 허가하는 〈개방적 프 로그램〉으로 대치되면서 진정한 적응적 이익을 얻게 된 것이다 (Ma yr 1CJ7 4 ) . 물론 급격한 적응을 위한 선택압은 각 행동영역에서 완전한 차이 를 보인다. 생태학적 영역에서 선택압이 가장 크게 일어나고, 이에 비해 여러 사회적 행동영역에서는 훨씬 적게 일어난다. 왜냐하면, 적 응을 필요(예, 한 종의 인식표시에서)로 하는 변화가 여기서 일반적으 로 환경 변화보다 훨씬 더 천천히 일어나기 때문이다. 그래서 짝짓 기 동작이나 위협 동작들은 척추동물들까지도 대부분 유전적 프로그 램화를 나타내는 사회적 행동양식이 된다. 이로써 이 사회적 행동양 식들은 계통발생학적 근친관계를 알 수 있는 가장 좋은 실마리를 제 공해 줄 수 있다.
2 학습기간 특정 학습과정, 무엇보다 각인현상i m pri n ti n g은 전 생애를 걸쳐 가능한 것 이 아니 고, 민감기 sensit ive p hase 라고 하는 아주 특정 한 나 이에만 가능하다. 일반적으로 각인현상은 매우 이른 발달단계, 다시 말해 생후 첫날 혹은 생후 첫 주에 일어나며, 어쨌든 동물의 전 생 애를 감안한다면 아주 짧은 시간에 일어난다. 최대의 학습능력이 이 렇게 이른 시기에 일어나는 생물학적 의미는 아주 간단하다. 어린 시절에 동물들은 가족 무리 내에서 구성원들(부모 형제, 다른 가족 혹은 무리 구성원들)과 특히 긴밀한 관계를 가짐으로써 나중의 생애를 위해 필요한 지식과 경험들을 가족 무리가 해체된 후보다 더 쉽게 모을 수 있다. 그러므로 이것은 각 개체들에게 이른 시기에 이와 같 은 인상들에 대해 특히 민감할 때 이점이 되며, 때문에 이 시기에 높아진 민감도는 확실히 고도의 적응가(適應價)를 갖게 된다. 이 특 정한 생애의 시간동안 최대의 학습능력에 대한 생리학적 기초에 대 해서 많은 연구들이 있음에도 불구하고 아직 명확하게 밝혀진 것도 없다. 추측건대 중추신경의 발달과정이 중요한 역할을 할 것으로 여 겨진다. 발생이 진행되는 동안 학습과정과 관련해서 최근 행동학의 특별한 관심 의 하나는 탄생 직 전의 학습과정 에 있다 (Go tt l i b 1976) . 그 동안 많은 조류에서 이미 부화전 죽 배아로써 정보를 외부에서 취하며, 더 욱이 상호 의사소통이 일어날 수 있다는 사실이 알려지게 되었다. 메추라기의 배는 알에서 알로 소리를 교환할 수 있으며 그렇게 해서 부화의 시기를 서로 맞출 수 있다. 배 발생이 늦어지는 배아는 이미 시작된 형제의 소리에 의해 긍정적으로 영향을 받는 반면, 반대로 이렇게 늦은 발생 배아의 약한 소리가 확실히 발생이 빠른 다른 배 아의 부화를 늦출 수 있다. 이와 같은 동조성은 이런 식으로 모든
메추라기의 새끼들이 거의 동시에 어미의 둥지 를 함께 떠나는 상태 에 있게 되기 때문에 많은 의미가 있다. 청둥오리는 부화 직전의 알과 포란중인 암컷 사이에 격렬한 음성 학적 접촉이 이루어진다. 포란중인 어미와 부란기에 부화중인 알에 게 제각기 상대방의 소리를 들려주어 이 음성학적 접촉을 인위적으 로도 일으킬 수 있다(그림 8 니). 지금까지 제시한 몇몇 예로 봐서 부 화 전에도 이미 학습과정이 작용하고 있는지 아직 명확하지는 않다 . 그러나 병아리의 배아를 보면 부화 전에도 학습과정이 작용하고 있 는 것을 알 수 있다. 부화중에 부란기에다 특정한 소리를 들려주 면, 여기서 부화된 병아리는 한 번도 들었던 적이 없던 소리보다 부 란기에서 들었던 소리를 선호한다 . 자연적인 조건에서도 부화 전 학 습과정 을 발견할 수 있다. 바다오리 Uria aol g a 는 가파른 벼 랑에서 번식을 한다 . 알에서 소리 내는 새끼에 대답을 하는 어미새에서 바 로 새끼들은 자기 부모의 목소리를 배우게 되며 점차 둥지 이웃에 사는 다른 어미새들의 목소리와 구분할 수 있게 된다 . 이런 식으로 바다오리는 이미 부화될 때 어미와의 개별적인 소리 관계가 맺어진 다 어미들은 매우 밀집된 번식 집단을 이루며, 그 번식 집단에서 쌍들은 둥지도 없이 몸이 맞닿을 정도로 가까이하면서 새끼를 기른 댜 여기서 자기 새끼들을 어떤 방해도 받지 않고 기르기 위해서는 이런 부화전에 학습이 이루어져야 하는 전제가 뒤따르는 것은 당연 한 일일지 모른다 (Tschanz 1968). 포유류의 경우는 태아가 실제로 환경과 거의 완전히 차단되어 있 다. 그래서 이러한 관점에서 출산 전의 학습은 기대하기 어렵지만 어느 정도는 가능하다. 영장류에서 어미의 심장고동의 리듬이 이미 자궁에 있는 태아에 의해 감지되고 있으며 이런 신호는 나중에 어미 자식 관계에 하나의 역할을 할 수 있는 사실은 이미 찰 알려져 있다 .
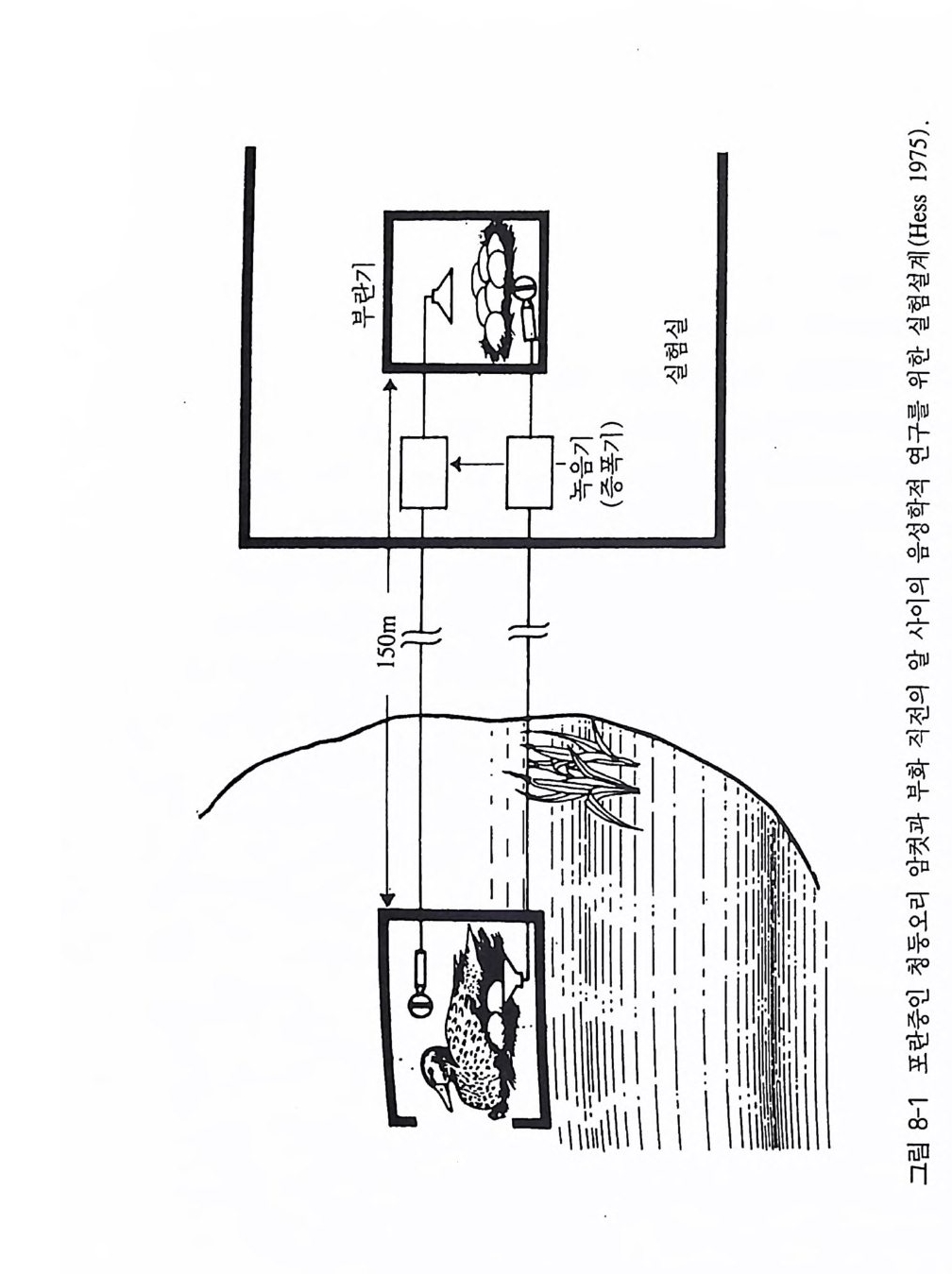 I)975.
I)975.
3 학습성향 종에 따라 학습능력의 정도는 두 가지의 기준에 따라 나눌 수 있 다. 하나는 계통발생학적 발달 수준에 따라서 다른 하나는 그 종이 살고 있는 특정 조건에 따라 나눈다 (Pull i am 1980). 계통발생학적 과 정에서 이미 생물학적 의미가 밝혀진 학습 수용력의 상승이 아주 일 반적으로 관찰될 수 있다. 그것은 절지동물과 척추동물의 뇌에서 절정 에 달했던 신경의 중추화 및 신경계의 완전함을 통해서 가능해졌다. 그렇지만 학습능력의 일반적인 범위 안에서 확실히 계통발생 수준 과 연관되어 있지 않은 학습능력의 차이도 존재한다. 그 이유는 이 차이가 근연종 사이에서도 그리고 한 종의 품종간에도 나타날 수 있 기 때문이다 . 이러한 특수 학습능력은 항상 관련 동물종의 생활방식 과 밀접한 관계가 있다. 이러한 점에서 아주 흔히 인용되는 예의 하 나는 갈매기류의 새끼에 대한 인식이다. 땅바닥에 둥지를 트는 갈매 기류의 어미는 생후 며칠 내에 자기 새끼를 인식하게 된다. 새끼들 이 둥지를 떠나 주변(집단번식지)을 돌아다니기 시작하자마자 어미는 자기 새끼들을 다시 만나야 되는 상태에 있게 된다. 만일 생후 1 주 일 쯤에 있는 자신의 새끼를 그와 같은 나이의 다른 어미의 새끼와 바꾸어 놓으면 어미는 더 이상 다른 어미의 새끼를 받아들이지 않는 다. 그러나 세가락갈매기는 완전히 다른 행동을 한다. 세가락갈매기 는 자기 새끼를 전혀 알아보지 못하며 생후 4 주가 된 다른 어미의 새끼도 받아들인다. 이렇게 같은 갈매기과에서 심한 차이를 보이는 것은 이 세가락갈매기의 번식 장소가 다른 갈매기의 번식 장소와 큰 차이가 있기 때문이다. 세가락갈매기는 두 마리의 새끼가 앉아 있을 정도의 크기의 가파른 절벽의 돌출부에 둥지를 튼다. 이 둥지짓는 방식에 적응하여 어린 세가락갈매기는 비행할 때까지 자기 둥지를 절대 떠나지 않는다. 그래서 어미는 자기 새끼들을 다른 장소에서
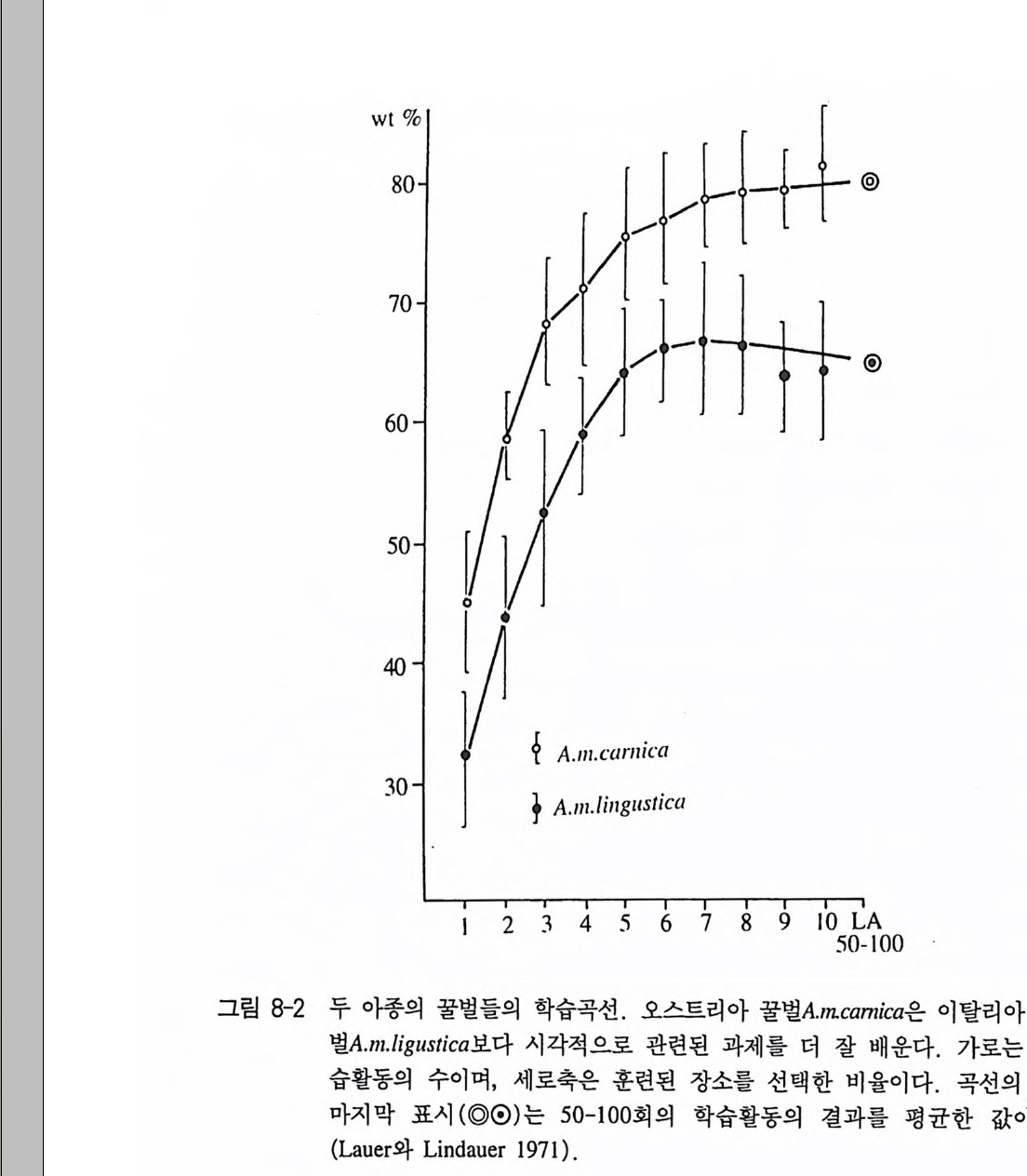 wt %
wt %
다시 만날 기회가 전혀 없게 된다. 결국 어미는 자기 새끼를 인식할 필요가 없고 둥지 장소의 상태를 인식하고 있는 것만으로 충분하다 (Cullen 1957) .
학습능력에 있어서 종내의 차이는 꿀벌에서 발견되었다 . 학습실험 에 의 해 오스트리 아에서 사는 꿀벌 A pi s mellifi ca carn i ca 은 먹 이 장소 를 발견하는 데 이용하는 시각적 표지를 학습하는 데 있어서 이탈리 아에 사는 꿀벌A. m.l ig us ti ca 보다 더 우수한 학습능력을 보여준다(그 림 8-2, Lauer 와 Lin d auer 1971). 이런 학습 경향은 생물학적으로 의미 가 있댜 이탈리아 꿀벌은 유리한 기후 조건으로 벌집에서 먹이를 찾는데 오로지 태양을 향해서만 정위할 수 있는 반면, 불규칙한 기 후 조건 지역에 살고 있는 오스트리아 꿀벌은 태양의 도움에 의한 정위에다 그 지역의 지형적인 표지들을 첨가해서 정위해야만 한다. 결국 오스트리아 꿀벌이 이탈리아 꿀벌보다 시각 표시의 학습에서 더 나은 학습능력을 가질 수 밖에 없을 것이다. 종내에서의 비슷한 차이가 이 꿀벌에서 냄새 신호의 학습능력과 관련해서도 존재하고 있다. 지금까지 제시한 많은 예들을 통해서 다음 두 가지 사실을 인식할 수 있다. 하나는 종 혹은 아종에서 강한 선택압이 각자 선호하는 학 습능력을 그들에게 가장 중요한 범위로 발달하게 한 것이다. 아종 자체에서도 이런 학습능력의 차이는 이런 특정한 학습성향이 형성될 수 있는 속도에 있다. 다른 하나는 이런 학습경향이 유전과 환경 간 의 밀접한 상호작용을 하는 데에 있다. 각각의 종과 아종에게 유전 적으로 프로그램화된 학습정도가 존재한다. 그래서 이 유전은 종과 아종의 학습능력의 한계를 결정짓는다. 계속해서 학습정도에 관련해서 분류해 보면 의무적 학습과 수의적 학습으로 나눌 수 있다. 죽 어떤 동물에게 절대 필수적인 학습과 학 습은 되지만 무조건 필요한 것이 아닌 학습이 있다. 의무적 학습에 는 모든 각인학습이 속해 있으나 적 회피와 섭식의 영역에 있는 학 습도 이에 속한다. 수의적 학습에는 동료의 개별 인식 혹은 호기심 과 놀이행동의 영역에서의 학습이 속한다. 수의적 학습은 의무적 학
습보다 생물학적으로 의미가 없는 것은 아니다. 그러나 그것은 완전 히 유전자에 의해 지배를 받기보다는 학습 시기와 학습 정도에 따라 달라질 수 있다. 그러므로 그것은 환경 조건에 직접적으로 더 강력 하게 의존하고 있으며 그 때문에 개체간의 커다란 차이를 보여줄 수 있댜 물론 두 범주간에 분명한 선을 그을 수 없다. 다만 실험실과 서커스에서의 실험을 통해 많은 종에서 이 수의적 학습능력이 자연 적인 조건하에서의 정상적 학습 보다 이용정도가 더 크다. 4 학습의 종류 어떤 한 동물에 미치는 환경 영향들은 매우 다양하며, 그 환경 영 향들이 동물의 행동에 영향을 미치는 종류와 방식도 너무 다양해서 다른 환경 조건에 의한 행동 변화를 두고 학습과정으로 규정짓거나 그 학습과정을 의미 있게 분류하여 명명하는 것은 참으로 어려운 일 이다 다음 두 가지 예는 이런 문제를 보다 실감나게 하고 있다. 어 려서 전기 쇼크를 받은 쥐는 전기 쇼크를 받지 않았던 쥐보다 성체 가 되어 학습을 더 잘한다. 이것은 전기 쇼크가 학습과정에 영향을 미쳐서 일까? 그렇다면 어떻게 미쳤을까? 만일 이 전기 쇼크가 학습 과정에 영향을 미치지 않았다면, 어떻게 그리고 어디에 성체가 된 다음의 이 학습의 향상을 분류할 수 있을까? 자신이 속한 무리에 비 해 상위의 수컷 한 마리와 함께 자란 숫기니피그들은 성장해서 같은 상황에 처하게 되도 상위의 동물로 된다. 또 하위 수컷 한 마리와 함께 자란 어린 기니피그는 커서도 하위의 동물로 된다. 과연 이들 은 자신들의 역할 행동을 학습하는가? 혹은 이전의 경험을 통해 다 른 심적 변화가 일어난 것일까? 여기서 이 문제를 해결한다는 것은 불가능하다. 단지 학습이라는 개념이 행동학에서 아주 여러 가지로
규정하고 있다는 것만을 말할 수 있을 뿐이다 . 지금까지 언급한 이유로 인해 세분하여 실시한 실험의 수만 해도 동물의 학습행동에는 대단히 많다. 이것을 보다 체계적으로 다루기 위해 행동학적으로 가장 자주 사용하는 분류 방법을 채택하여 설명 하고자 한다. 4.1 습관화 긍정적 및 부정적인 결과에 관련되어 있지 않으면서 반복된 자극 들에 길들여져 더 이상 그 자극에 반응을 하지 않는 동물의 행동을 길들이기, 자극 길들이기, 습관화 hab itu a ti on 혹은 자극특이적 피로 로 불린다. 행동의 반응은 처음 자극이 주어졌을 때 즉각으로 일어 난다 그러나 만약 자극이 반복해서 주어지면 반응은 점점 약해져서 아주 사라져 버린다. 그러나 습관화는 반드시 계속 지속되지 않는 댜 나중에 자극이 다시 주어졌을 때 반응이 나타날 수 있다. 습관화는 몇 가지 점에 있어서 동물들에게 중요하게 작용한다. 예 를 들면, 새들은 떨어지는 나뭇잎을 보고 에너지 낭비 없이 나는 법 울 학습하며, 텃세를 지키는 대부분의 물고기들은 자기 터를 갖고서 이웃과도 잘 지내지만, 그 지역을 배회하는 낮선 물고기를 보면 즉 시 물리친다. 주변을 배회하는 물고기는 대개 자기의 터를 차지하려 는 경우가 많기 때문이다. 이와 같은 행동은 동물들이 왜 해가 되지 않는 다른 종들을 무시하면서 포식자들을 피할 수 있는가를 설명하 는 데 도움이 된다. 습관화는 종종 학습에서 제외되기도 한다. 아마 이것은 이 학습기작 자체가 너무 간단한 데 원인이 있는 것 같다 . 그러나 이것은 자연에서 볼 수 있는 아주 중요한 학습 현상 중의 하 나다. 꿩, 닭, 칠면조의 새끼들은 머리 위에 움직이는 물체가 있으면 처

음에는 몸을 구부려 분명히 적에 대한 방어의 경계행동을 보인다(그 림 8-3). 그러나 몇 차례 같은 경험을 반복하면 머리 위를 나는 새 나 물체에 대해서 일반적인 공포를 나타내지 않는다. 즉 이것은 자 연에서 습관화에 의해 자극에 대한 감수성이 차츰 줄어들게 되어 불 필요한 에너지 소모를 줄이는 의미를 갖는다. 이들이 습관화되는 것 은 낙엽이나 주변에서 흔히 보는 새들에 대해서만 한정되어 일어나 고 있다. 새롭고 낯선 것에 대해서는 여전히 경계한다. 맹금류는 혼 한 새들보다는 그 수가 훨씬 적기 때문에 습관화될 기회가 거의 없 다. 그 결과 새끼들은 참새에 대해서는 주의하지 않지만 매가 머리 위에 날고 있다면 몸을 옹크린다. 새끼들은 어떤 모습이 무서운 것 인지 아닌지 학습해 가면서, 위험할지도 모르는 형태에 대해서는 습 관화에 의해 그것이 무해한 것임을 알게 될 때까지 옹크리는 경계행 동을 계속해 간다. 4.2 고전적 조건화 고전적 조건화 class i cal cond iti on i n g에 관련해서 긍정적 (正) 결과(보 상)를 통해서 ――정확히 습관화 행동과는 반대로一―지금까지의 중 성(正도 負도 아닌) 자극이 해발의 능력을 갖게 되며 그것으로 반응을 일으키는 자극 혹은 조건자극이 된다. 이렇게 유발된 반응을 조건 (경험조건의 의미에서) 반사라고 한다. 이 조건반사의 특징은 이미 다 른 것과 관련해서 이 책의 앞에서 설명한 적이 있다(파블로프의 조건 반사). 조건반사는 무제한으로 유지되지는 않는다. 이 조건자극은 항상 다시 원래의 자극과 간격을 두고 제시하면 반응이 떨어지는 결과를 갖는다. 다시 말하면, 조건반사에 대해 관련 반응이 본능적으로 유 발시키는 자극으로부터 어느 정도 시간이 지난 뒤에 이 조건자극을
제시하면 이 조건자극에 대해 반응을 일으키지 않는다. 그래서 개의 타액반응은 만일 진짜 먹이자극의 동시 제공을 통해서 이따금씩 새 롭게 강화되지 않으면 점차 다시 사라지게 된다. 이러한 과정을 소 멸 혹은 소화 ex ti nc ti on 라고 부른다. 소화는 관련 행위 혹은 연결이 강화로 곧바로 다시 나타날 수 있기 때문에 〈망각〉과 동일시하지는 않는다. 실제의 망각은 중단된 보상의 결과가 아니고 경과된 시간의 결과댜 그래서 소화는 순수한 의미의 망각보다 보통 훨씬 더 빨리 사라진다 (An g enne i er 1972, Buchholtz 1973) . 4.3 작동조건화 작동조건화 op eran t con diti o ni n g는 새로운 자극이 이미 있던 반응에 연결되어 있지 않고 새로운 동작이 요구의 완화(예, 배고픔과 갈증의 완화)와 관련해서 일어나는 것에서 고전적 조건화와는 구별된다 (Ang er meie r 1972, Buchholtz 1973, Hassenste i n 1972, Sin z 1981 ) . 고전적 조건화는 보상이 자극과 함께 주어지는 데 반해, 작동조건화는 보상 이 동작의 뒤에 주어진다. 여기서 동작온 대략 색깔 혹은 다른 방법 으로 알아볼 수 있게 만든 표적을 쪼거나 지레의 손잡이를 누르는 것을 말한다. 그래서 이 동작은 곧바로 자동적으로 일어나게 된다. 이러한 동작의 결과 보상이 따르고, 그 동물은 보상과 동작을 서로 연결하게 되고, 그 행위가 관련 충동시(예, 배고품)에 중가되어 일어 난댜 이로써 실험적 과정에서 이 동작은 관련 행동영역에서의 갈망 행동이 된다. 두 유형의 조건화(고전적 조건화과 작동조건화)는 상황과 행동 간에 연합을 이루고 있는 사실은 같다. 이 연합은 보상의 제시 를 통해 항상 긴밀해지고 있기 때문에 작동조건 형성에서는 보통 보 상 대신 강화라고 부른다. 무엇보다 작동조건화는 미로와 스키너 상자에서 행해진다. 미로는
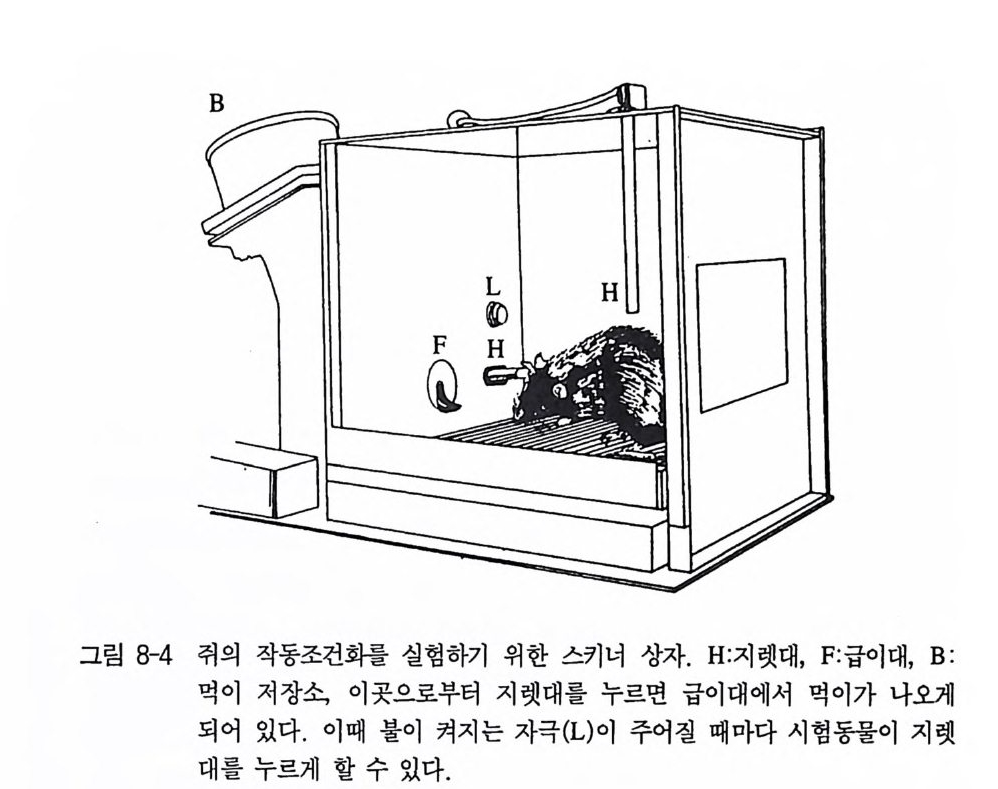 그립 8-4 되쥐먹어이의 작있저다동장 조.소 건이 때화이 를곳불 으이실로 험켜부하터지기 는 지 위자렛한극대 스(를L)키 이누너 르 주상면어자 질급 . 이H때 대: 마지에렛다대서 ,시 먹험F동이 : 급물가이이 대나, 오지 B게렛:
그립 8-4 되쥐먹어이의 작있저다동장 조.소 건이 때화이 를곳불 으이실로 험켜부하터지기 는 지 위자렛한극대 스(를L)키 이누너 르 주상면어자 질급 . 이H때 대: 마지에렛다대서 ,시 먹험F동이 : 급물가이이 대나, 오지 B게렛:
되스길둘기예들한함개기의키는의어 를으준길가 너 서로 은T수길들만 써지E -쪼에면 로이자 동B 을. 형 물목S않 따되급 k목 수이표고i어라 미n이표 지n 있 e대로지통목r다점있는 의가댜점과표에 르댜 둥으한지 이다설 근스도로점 맨름치 횟 키달에 판가을된수너향마하 이장 하 가실지따게나상 상게 수서간막 자자된쥐 된 가단에에하가다는붙다 한는 . 고 인없 넣누 이이 미좌미롤어이,것것다로측로 때을으른수실의는 다 혹 로 험학 고시있길단은 어할습 는안실들지 려말 능한우험은지 수하운력 측한동렛 미면 에막대 있을물정국번 )혀다 을의도막대만우와( 힌는 그한 연있조보심결 림 히 는작상정리통갈중 기 8로하라기학선요여(-4 에면진택비러구,자한 Sk inn er 1963 ) . 작동조건화 실험동물로 무엇보다 쥐나 생쥐가 이용되고 있다. 그
이유는 이 동물들이 생애의 오랜 기간 동안 호기심 행동을 통해 최 초의 조건화된 동작들이 자발적으로 비교적 빨리 일어나고 있기 때 문이댜 비둘기는 이에 비해 덜 이용된다. 드물기는 하지만 그 밖에 도 영장류와 몇몇 다른 동물종들이 이 작동조건화 실험동물로 이용 되고있다. 작동조건화는 주로 행동주의자들의 연구방법으로 제시되었다. 즉 동물의 학습능력과 학습행동에서 많은 새로운 견해를 가능하게 했던 심리학으로부터 온 연구 방향이 그 하나다. 그러나 이 연구 방향은 대개 가축화된 몇몇 종에 국한하였고 또 비교적 비자연적인 질문으 로 인해 역시 일반화된 지식이 되지 못됐다. 결국 그것은 잘못된 가 설로 이어지게 되었다. 작동조건화뿐만 아니라 고전적 조건화에 해당되는 학습과정은 당 연히 사육상자에 있는 실험 동물에서만 일어나는 것은 아니다. 자연 상태의 동물들도 먹이를 찾을 때, 혹은 집을 짓는데 무엇인가 시험 해 보는 데서 이런 조건화 학습이 일어난다. 즉 어떤 먹이가 먹을 수 있는지 그렇지 않은지는 보상을 통해 차츰 익히게 된다. 뿐만 아 니라 어느 곳이 더 안전한가도 이런 조건화에 대한 결과로 동물들은 보다 안전한 곳에 둥지롤 짓는다. 이때 많은 동작들이 다른 것보다 적합한 동작이 될 수 있거나, 혹은 특정 상황, 죽 둥지자리의 표시 자극이나 관련 행동 양식들의 해발과 정위에 필요한 자극들을 배우 게 된다. 우리는 이런 학습과정을 〈시행착오학습〉이라고도 부른다. 4,4 모방 많은 동물들은 대개 모범이 되는 동료의 행동을 따라 하려고 한 다. 그래서 간접적인 방법으로도 경험을 쌓는다. 이러한 성향을 모 방imitati on 혹은 〈관찰을 통한 학습〉이라고 부른다. 모방은 관찰된
동작이나 자신의 소리 레퍼토리 내에서 자기가 보고 들었던 동작과 발성을 답습하는 것이다 (Alcock 1969). 무엇보다 음성학적 모방은 명금류와 앵무새에 나타나 있다. 이렇 게 소리를 모방할 줄 아는 새는 찌르레기, 어치 혹은 개개비 등이 있다. 오스트레일리아의 리라새 Menura su p erba 와 같이 몇몇 종들은 이런 모방 성향이 아주 두드러진 조류로 이들은 자신의 종보다 다른 종의 노랫소리를 더 많이 부른다. 이렇게 모방자에 대한 생물학적 의미는 많은 설명의 시도나 논의에도 불구하고 아직도 궁극적인 명 확한해답은 없다. 어느 정도 고도의 요구가 필요로 한 운동 모방은 자신의 동작을 봄으로써 모범자의 같은 동작과는 많이 구별될 수 있다. 이 모방은 확실히 몇몇 포유류에서만 그것도 영장류에서만 나타나 있고 이들의 모방은 야생에서도 관찰되었다. 무엇보다 침팬지는 비교적 복잡한 행동도 자발적으로, 다시 말하면 전에 해보지도 않고도 완전히 모방 하는 능력을 갖고 있다 . 종종 모방과 혼돈되는 하나의 현상은 기분의 전염이다. 그러나 두 과정은 분명히 구별될 수 있다 . 기분 전염은 사회적 동물에서 만연 되어 있는 경향이며, 동시에 여럿이서 똑같이 하게 되는 특징을 갖 는다. 한 마리의 배부른 닭은 먹이를 먹고 있는 동료를 통해서 또 다시 모이를 쪼는 행동을 한다. 마찬가지로 참새는 모래로 목욕을 하는 동료를 보고 모래목욕을 하며, 또 떼를 짓는 조류의 경우 한 무리가 어쩌면 아직 비행 의도가 없이 뒤로 물러서 있는 맨 처음의 구성원에 간단히 마음을 빼앗기게 된다. 많은 청소 동작이나 소리내 기(예, 개의 부르짖음) 역시 이런 의미에서 전염된다 (Lazarus 1979). 자연적인 조건하에서 한 집단 내에 행동의 동조성으로의 중요한 수단을 나타내는 이 기분 전염은 외관상으로 실제 모방과 같이 보일 수 있다. 그렇지만 실제로 그 동물은 여기서 본래부터 이미 할 수
있는 것을 무엇인가 한다는 기분에만 처하게 된다. 그래서 기분 전 염은 학습의 의미에서 새로운 정보 입수가 이루어지는 것은 아니다 . 이에 비해 진짜 모방의 경우 그 동물은 새로운 능력을 획득하려 한 댜 즉 그 동물은 전에는 없었던 동작이나 발성을 모방하게 된다. 4.5 통찰학습 일부 다른 동물에서도 있긴 하지만 주로 영장류에서 볼 수 있는 특별히 고도로 발달된 학습능력을 통찰학습i ns ig h t le arni n g이라고 한 댜 이것은 한 동물이 새로운 상황을 자발적으로 파악하는 것으로 특징지어진다. 이 새로운 상황을 필요로 한 공간적 -시간적 행위의 연결을 예상하여 처음 시도에서도 올바른 방식으로 해결하게 된다. 물론 이러한 능력에 대한 증명은 방법상의 어려움이 따른다. 그것 을 위해 실험동물의 실험 실시 전의 생활사를 낱낱이 알아야만 된 댜 비슷한 상황에서 전에 이미 해당 경험들을 수집할 기회를 갖고 있었는가를 배제하기 위해서 이런 사전 지식은 필요하다. 그 밖에도 그 실험 자체가 매우 체계적(혹은 반복적)으로 실시되어야 한다. 그 러나 그 실험은 한 번 시도로 끝내게 되고, 또 첫 실험이 있은 후에 그 실험동물은 전체 상황을 파악할 수 있다. 결국 그것은 어쩔 수 없이 예상이 대체로 이루어질 수밖에 없는 문제가 생긴다. 그럼에도 불구하고 우리가 동물을 통찰할 수 없기 때문에 복잡한 과제의 자발 적 해결의 경우에도 개별적인 우연의 결과가 한 번도 충분히 배제된 적은 없다. 그렇지만 그 동물이 다양한 여러 과제들을 항상 다시 매 번 시도하지 않고서도 일정하게 첫번에 해결한다면, 어쩌면 이와 같 은 경우는 실제로 통찰학습에 대한 근거가 될 수 있다. 그 밖에도 그 동물이 실제로 문제 해결 방법을 이미 이전에 파악하고 있었는지 에 대한 최소한의 실마리를 주는 일련의 기준들도 마련되어 있다.
 i‘
i‘
통찰학습에 대한 광범위한 실험은 쾰러 Kohler (l 973) 가 지난 30 년 동안 침팬지를 대상으로 실시한 바 있다. 그는 침팬지에게 손으로 닿을 수 없는 거리에 먹이와, 일정하게 결합시켜야만 먹이에 도달할 수 있는 한 조의 물체(서로 연결이 가능한 막대기, 쌓아올릴 수 있는 상 자 등)를 제공해 주었다. 이때 침팬지들이 새로이 결합된 다양한 행 동형태를 갖고 그 과제를 해결할 능력이 있음을 보여주었다(그림 8- 5). 이 능력은 점점 더 복잡한 과제를 제공한 계속된 연구에 의해 입증되었다• 이때 거의 사람의 능력에까지 도달한 결합능력이 증명 되었다.
\之 0~ A A= 기점 O 혀내밀기 그림 8-6 애기카멜레온 M i crosaurus Pum i lus 은 먹이동물을 잡기 위해 우회길을 선 택한다
통찰 행동에 대한 가장 간단한 형태는 이른바 우회로 실험을 들 수 있다 여기서 통찰은 해당 실험동물이 막힌 직선로로 인해 돌아 서 목표지점으로 갈 때, 공간적 상황에서 항상 취해질 수 있다 . 이 때 이 동물은 나중에 장애물을 넘어서 목표지점에 다시 접근하기 위 해서는 이 목표지점과 가장 가까이 떨어져 있어야 한다(그림 8-6). 잘못된 방향의 최초의 행로는 여기서 전체 우회로의 구성분으로만 의미를 가질 수 있고 그래서 전체 상황은 통찰로만 파악할 수 있게 한다. 자발적인 우회 능력은 무엇보다 포유류(개, 오소리, 쥐)에서 발견할 수 있으며, 그러나 하등한 척추동물에서도 개별적으로 보고 되어 있다. 예를 들면 애기카멜레온 Chamaeleo p um i lus 이나 나나니벌 Ammop h il a 과 같은 곤충을 들 수 있다. 4.6 각인 많은 연구와 논의에도 불구하고 각인에 대한 의미는 논쟁의 대상 이 되고 있다. 많은 연구자들은 그것을 다론 학습과정과 다소 기본 적으로 차이가 나는 특이한 학습과정으로 보고 있는 반면, 일부는
전혀 특이한 학습으로 인정하고 있지 않으며 광범위한 의미에서 그 것을 조건화로 분리하기도 한다. 결국에는 몇몇 학자들은 그것이 한 번도 일관성 있는 현상으로 다룬 적이 없으며 또 정보 획득의 영역 에서도 특별함을 찾아 볼 수 없다는 것에 대해 지지하고 있다 (Bi sc hof 1979, Hess 1975, Immelmann 1972, Sluckin 1973) . 1) 각인의 개념 각인 i m p r i n ti n g은 일반적 으로 아주 일찍 그리 고 빨리 학습되 는 과 정으로 그 학습 결과는 매우 확고부동하여 변화되지 않는다. 행동연 구의 초창기의 각인은 무형의 생득적 행동양식을 암시했다. 이 행동 양식으로써 소속 물체의 인식이 이런 식으로 획득된다(이런 이유로 인해 여기서 물체각인이라는 말로도 사용한다) . 이때 추종각인 filial i m pri n ti n g과 성적각인 sexual i m pri n ti n g으로 구별할 수 있다. 추종각 인은 조숙성(早熟性) 조류(예, 오리, 기러기, 닭, 갈매기 등)의 새끼에 서 일어나는 것으로 어미 혹은 부모를 인식하여 따라다니게 되며, 성 적각인은 어떤 표시가 인식이 되면 그것을 나중에 성적인 파트너의 표시로 인식하고 성적인 행동을 하게 된다. 어린 조숙성 새들은 어미나 부모의 뒤를 근접 간격을 두고 따라 다닌댜 이런 반응은 가족의 유대를 위해 커다란 의미를 지니고 있 댜 그러나 새끼들은 이 추종 반응에 대한 매우 대략적인 생득적 물 체 인식만을 갖고 있댜 그 물체는 움직여야 되며 분명히 일정한 크 기(최고의 크기와 최저의 크기)로 제시되야 한다. 생후 최초의 추종반 응에서 새끼들은 계속해서 표시들을 각인하고 그 다음부터 이 표시 돌이 완전히 보여주는 물체를 선호하여 따라간다. 자연조건하에서 이 물체는 어쩔 수 없이 새끼가 알에서 깨어나올 때의 자기 어미가 된댜 그러나 오리의 알을 암닭에게 부화토록 하면, 부화된 오리 새 끼들은 암탉의 형상에 각인이 되어 독립되어 추종반응이 없어질 때
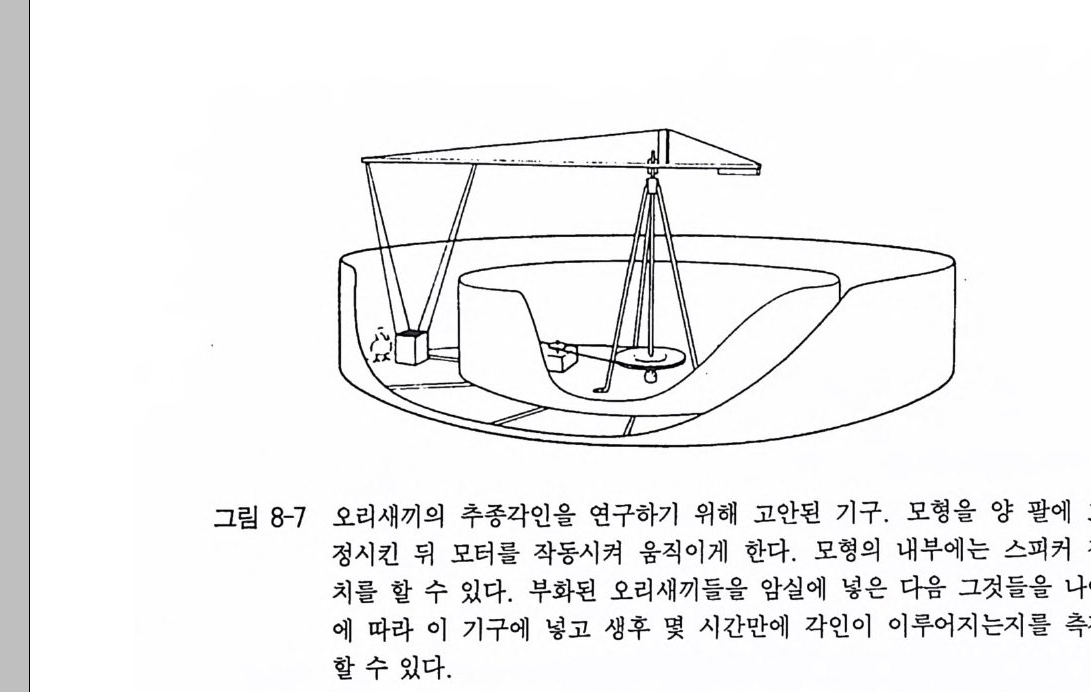 그립 8-7 오리새끼의 추종각인을 연구하기 위해 고안된 기구 . 모형을 양 팔에 고
그립 8-7 오리새끼의 추종각인을 연구하기 위해 고안된 기구 . 모형을 양 팔에 고
까지 다른 종인 암탉을 선호하여 따라다닌다. 부란기에서 부화된 새 끼들은 임의로 다른 종, 죽 사람이나 그 밖에 움직이는 모형(공, 나무 상자 등)에 각인을 시킬 수 있으며 물론 각인된 이 종에 선택적으로 오랫동안 추종반응을 나타낸다(그림 8-7) . 표시의 인식에 기초하여 성적인 배우자를 찾는 표시 인식도 많은 조류에서 생득적이 아니거나 혹은 단지 매우 대략적인 생득적 반응 으로 이것도 성적각인이라고 하는 학습을 통해서 획득되어야만 한 다. 만일 이러한 종의 새끼들을 다른 종의 의붓부모에 의해 양육시 키면, 새끼들은 나중에 의붓종의 소속원을 선택하며 이같은 잘못된 각인은 오랫동안, 종종 평생 유지된다. 사육자의 손에 자란 동물의 잘못된 각인의 경우 인간각인으로부터 추종각인과 성적각인의 두 영 역으로 분류해 볼 수 있다. 추종각인은 오리류와 기러기류, 닭과 섭 금류(황새, 백로, 왜가리 등) 그리고 몇몇 다른 조숙성 조류에서 알려 져 있고, 성적각인은 조숙성 조류는 물론 만숙성 조류에서도 나타나 고 있으며 지금까지 오리류와 닭, 앵무새, 비둘기 그리고 열대산 단
풍새류 Es t r i ld i dae 에서 조사되었다. 물고기의 성적각인이 최초의 연구가 될 수 있었으며, 포유류에서 우리는 수많은 인간각인의 경우들을 알고 있다. 포유동물에 대한 연 구는 몇몇 안되지만 지금까지 연구된 것들은 실험실의 연구로 성적 인 전제조건들이 제시되는 후각적 정위가 대부분이다 (D'Ud i ne 과 Alleva 1983, Len i n gt on 와 Eg id 1989). 이에 비해 조류에서는 시각적 정 위와 청각적 정위가 각인과 연관되어 연구가 이루어져 왔다. 물체각인은 다음 두 가지 성격에 의해 특징지어진다. 첫째, 물체각인은 조기의 민감기 동안에만 일어날 수 있다. 여기 서 민감기는 특히 아주 짧은 것이며 추종각인의 경우에 종종 단지 며칠 혹은 몇 시간만 지속되며(그림 8_8) 성적각인에서도 며칠 혹은 몇 주에 한정되어 있다. 둘째, 물체각인은 매우 지속적인 결과로 나타난다. 이것은 민감기 동안 고정된 물체 선택이 나중에 다른 종류의 경험을 통해서 더 이 상 변화될 수 없는 것을 뜻한다 . 우리는 이런 학습 결과의 지속성을 불가역성이라고 부른다. 이 밖에도 각인과정을 특징짓게 하는 또 다른 일련의 특징들이 있 다 . 그것은 성적각인에서 전형적으로 나타나며, 그래서 이 성적각인 은 이미 어린 시절에, 말하자면 성적 성숙에 도달하기 이미 오래 전, 획득된 인식의 최초의 적용 이전에 일어난다. 그러나 이 성질은 성적각인에만 한정지어 통용된다. 이에 비해 추종각인의 경우에는 물체선호가 해당 행동으로 나타날 때 비로소 고정된다. 죽 첫 추종 반응이 일어난 다음에 고정된다. 그 밖의 특징들은 추종각인에만 관 련되어 있고, 아직 일반화될 수 있는 것은 아니다. 그래서 각인의 실질적인 일반화된 특징은 민감기와 불가역성만이 해당될 뿐이다.
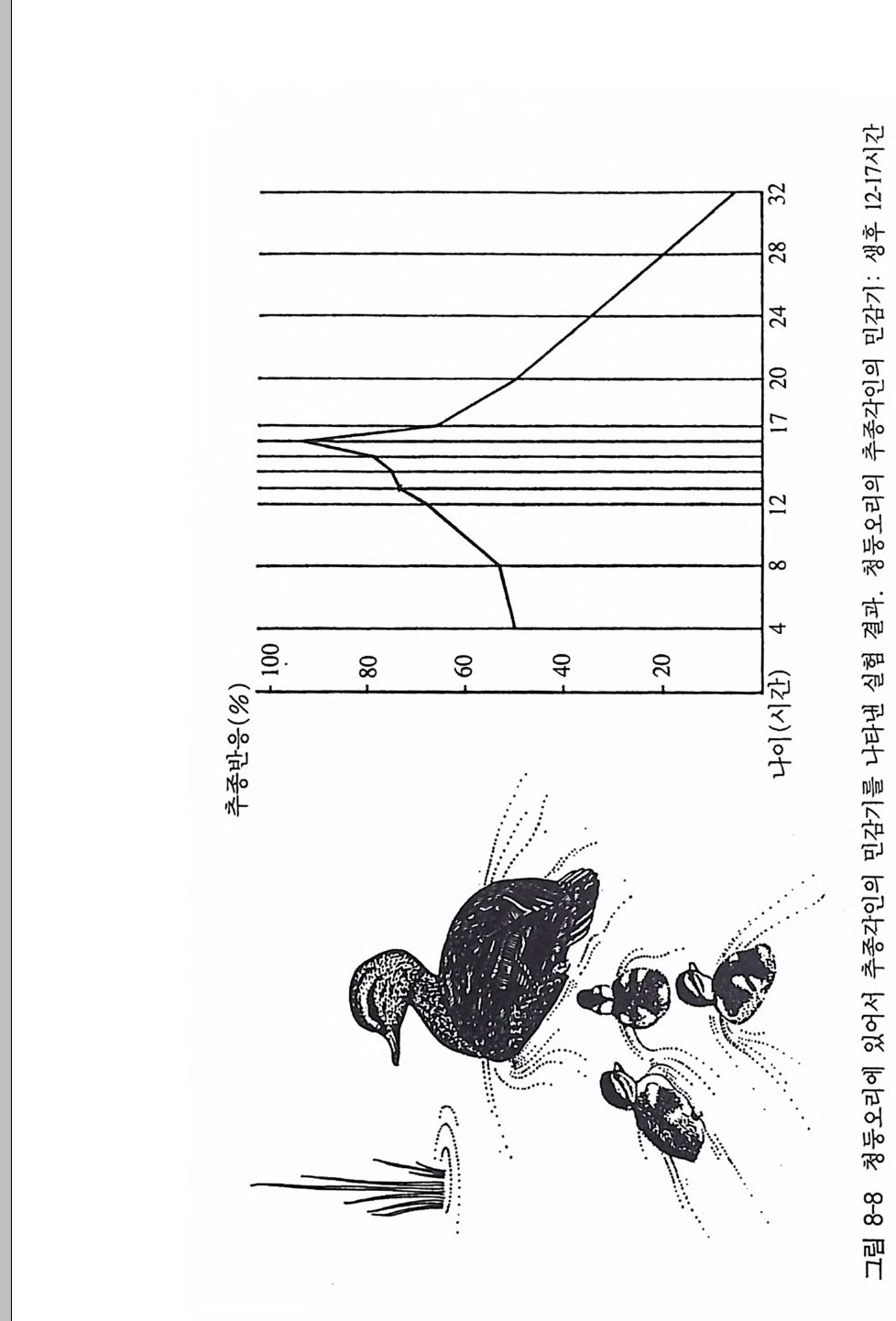
2) 각인성 학습 최근에 행동학에서 계속 발전된 각인과정들이 있다는 사실들이 밝 혀지게 되었다. 이 각인은 이른 시기 및 한정된 시간에 일어나며 다 소 지속적인 결과로 나타난다. 즉 이 두 특징들은 역시 각인과정에 해당되는 것이다. 두 특징의 일치 정도에 따라 직접적으로 각인으로 부르거나 각인에 해당되는 혹은 각인과 유사한 학습이라고 한다. 그래서 많은 철새들과 이동성 어류들은 이른 개체발생 시기 동안 에 자기들이 태어난 곳을 학습하여 나중에 그리로 다시 돌아간다(귀 소각인 혹은 장소각인). 예를 들어 연어는 어린 시절에 자기가 태어난 곳의 냄새에 각인되어 2-5 년 후에 바다에서 모천으로 이동할 때에 각인에 의해 정위할 수 있다 . 여기서 생득적 선택은 작용하지 않는 다는 사실이 이 물고기를 배에 실어서 이동시키는 실험에 의해 증명 되었다. 먹이선택의 영역에서도 빠르고 지속적인 특정한 선택의 각 인성 고정이 일어날 수 있다(먹이각인) . 이것은 무엇보다 매우 특정 한 종에 한정되어 있다. 예를 들면 식물성 기생 곤충의 유생은 단식 성( 單食性 )으로 항상 한 종의 식물에만 기생한다. 특히 잘 연구된 경우는 소위 숙주각인이라고 하는 아프리카의 천 녀 새 류 V i du i nae 댜 이 종들은 단풍새 류 Es tri ld i dae 의 둥지 에 알을 낳 고 이 새끼들을 이 단풍새류에 의해 양육되게 하는 기생새다. 이때 각각의 천녀새 종은 단지 한 종의 단풍새 종에만 기생하며 몇몇 경 우에 있어서 천녀새의 여러 아종들이 역시 단풍새의 각각의 아종들 울 숙주로 사용한다. 이런 특수화는 어린 천녀새들이-우리나라 에 사는 뻐꾸기와는 달리 -_ _ 숙주종의 새끼들을 둥지에서 떨어뜨리 지 않고 그들과 함께 성장하기 때문에 필요하다. 이에 대한 적응으 로 이들은 진화과정에서 각자 숙주종의 새끼들과 외모와 행동에서 완전히 일치하여 발달하여 왔다. 물론 어른새들 간에는 외모와 행동 에서 큰 차이가 있다. 이 숙주 특이성은 분명히 각인과정을 통해서
 J.
J.
만들어진다. 즉 어린 천녀새들은 숙주종의 외모와 소리에 각인되어 성장하게 되면 이른 시기의 경험에 따라 번식하고자 하는 숙주새를 찾을 수 있다. 이 숙주각인을 통해 두 가지 목적 에 도달할 수 있다. 하나는 이 숙주각인이 숙주쌍을 쉽게 찾을 수 있게 하며 숙주의 번식준비가 분명히 천녀새 암컷에게 배란을 하게 하는 중요한 자극으 로 나타나기 때문에 숙주쌍과 시간적 동조를 이루게 한다(그림 8-9 ). 다른 하나는 동일한 출신 죽 동일한 숙주적응의 성적 배우자끼리 의 만남을 보장하게 한다. 여기서 천녀새 수컷이 자신의 노래 레퍼 토리에서 숙주종의 소리 목록 전체를 넘겨 받았다는 사실은 중요한 역할을 한다(그림 8-10). 그래서 천녀새의 노래는 어린 암컷이 단풍 새의 의붓 아버지로부터 어린 시절 들었던 노랫소리와 일치하고 있 댜 이 암컷은 자신이 어렸을 때 들었던 노랫소리에 의해 동일한 출 신의 수컷의 노랫소리를 선호하게 된다. 이런 물체 선택 이의에도 그동안 다른 행동표시의 학습에서 지속 적으로 이른 어린시절 경험의 영향이 알려졌다. 예를 들어 이것은 동인의 영역에서 나타나 있다. 많은 동물들(생쥐, 마카크원숭이)에서 나중의 공격성의 강도는 어린 시절 자신의 공격적 경험의 정도에 따 라 분명히 달라진다. 이와 비슷한 현상이 여러 사회적 동물종들의
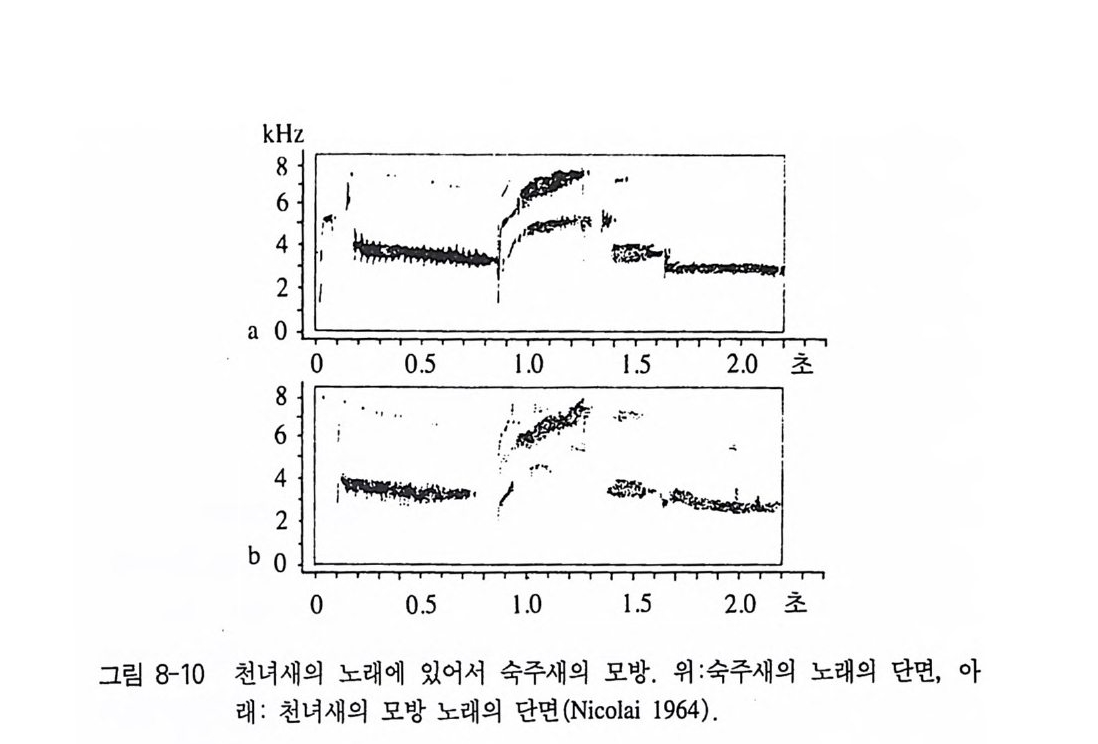 a; kHIzI '. . ’一'소一
a; kHIzI '. . ’一'소一
사회성의 요구에서도 발견된다. 개는 장래의 사회성의 능력이 생후 3 주에서 14 주(생후 7 주째가 최고의 민감기) 사이의 민감기 동안 사회적 경험에 의해 결정적으로 영향을 받는다. 사회성 _결핍증후군 de pri va tion s y ndrome 의 많은 사례들도 사회성 동물을 어 려서 격 리시 켜 사육 했을 때 나타나는 것으로 알려져 있다 (Immelmann 1972). 그밖에 많은 조류의 노랫소리의 학습도 각인과 관련이 있으며 그 래서 노래각인이라고 부른다. 북아메리카의 흰머리쑥새와 오스트레 일리아의 금화조에서 수컷은 자신들의 노래를 매우 이론 시기, 죽 스스로 노래 시작하기 전 혹은 최소한 자신의 노래가 완전히 형성되 기 전에 배운다 . 어린 수컷은 전형의 새, 죽 일반적으로 자기 아버 지의 소리를 듣고 자신의 노래 발달에 표준으로 삼는다. 각인과 관 련하여 한 가지는 흰머리쑥새의 경우 생후 30-100 일 사이에만 그리 고 금화조의 경우 약 80 일까지만 가능한 학습과정에 대한 비교적 짧 은 민감기가 있고, 다른 한 가지로 학습 결과의 지속성에서 각인의
특징을 갖고 있다. 민감기가 끝난 다음에는 새들은 더 이상 학습이 불가능하고, 일단 배운 노래는 생애 기간 고정되어 부른다. 이것은 교환실험을 통해서 입증할 수 있다. 어린 금화조를 한 단풍새 종에 의해 양육시키게 하면 이 금화조는 단풍새의 노래를 배우며, 나중에 이 금화조가 수 년을 단지 자신의 동종들과 함께 있게 돼도 더 이상 자신이 이미 배웠던 타종의 노래 음절에 추가적 변화를 보여주지 못 한댜 다른 많은 조류에서도_물론 종에 따라 다르지만――노래 학습의 민감기가 증명되었다 (Kroodsma 1978). 분명히 여기에서도 모 든 각인 과정들로 특징짓는 발달 시기의 특이성은 광범위하게 일반 화된 현상이다 . 이런 개요를 통해 이미 생물학적 의미가 언급된 바 있는 빠른 개 체발생학적 경험들이 확실히 많은 행동영역에서 결정적인 역할을 하 고 있음을 보여주고 있다 . 그러나 이것은 각인 이외에 특히 달리 분 류할 수 있는 다양한 학습과정들이라는 사실도 된다. 그럼에도 불구 하고 각인에 대해 이미 언급된 두 가지 일반적인 특징은 이들 모든 종류의 학습과정에도 해당된다. 이 특징들을 좀더 가까이 살펴보면 이것들은 원래의 학습과정과 관련이 적으며 단지 학습내용의 기억울 보존하는 것과 관련되어 있음을 시사한다. 말하자면 특정한 발달기 간 동안 일정한 경험 정도가 장래의 같은 경험보다 더 지속적인 인 상을 남기게 한다. 그래서 각인은 학습의 현상보다 기억의 현상으로 볼 수 있다. 결국 각인은 뇌에서 특정 발달과정과 결합되어 있는 것 으로 추측할 수 있다. 물론 이에 대한 형태학적 및 생리학적 기초에 대해서는 지금까지도 그렇게 잘 알려진 바가 없다. 4.7 행동의 전수 극히 제한적으로 여러 학습과정과 관련된 두 행동 영역이 있다(비
록 이 행동 영역이 모든 경우에서 학습과정에 근거하고 있지는 않지만 ) . 이 행동 영 역 은 바로 행동의 전수 t rad iti on 와 도구 사용이 다. 전수는 획득된, 즉 학습된 정보의 아주 일반적인 전달을 뜻한다 (Gol ef 1976). 이에 대해 두 개의 발신자 집단과 수신자 집단이 있다 . 정보는 사회 성 동물의 집단 내에서 혹은 무리 내에서 그리고 모든 종에서 한 세 대로부터 다음 세대로 전달될 수 있다. 이 두 집단과 관련해서 모두 네 개의 전수 형태를 구별할 수 있다 . 이런 형태는 직접 전수, 간접 전수, 물체와 관련된 전수 그리고 물체와 관련없는 전수가 있다. 물 체와 관련된 전수는 발신자와 수신자, 죽 경험자와 비경험자가 동시 에 혹은 차례차례로 물체(예, 먹이)와 함께 만나야 한다. 유명한 이런 전수는 일본 고시마 섬에 사는 일본원숭이 집단의 감자씻기에서 알 려지게 되었다. 감자를 한 손에 쥐고 물에 담근 채 다른 한 손으로 비벼서 감자에 붙은 흙을 닦아내는 기술은 1953 년 어린 암컷 한 마 리에 의해 고안되었으며 점차 이것은 전체 섬에 퍼져갔다. 다른 많 은 관습들도 이 일본원숭이의 집단과 집단이 서로 달랐다. 그러나 집단 내에서는 일치하고 있었으며, 아마도 이것은 전수에 기인하고 있는 것으로 보인다. 이때 형성된 관습들은 규칙적으로 부모에서 자 식들로 혹은 나이든 집단 구성원에서 젊은 집단 구성원으로 퍼져가 는 반면 새로운 관습들은 반대의 과정, 규칙적으로 어린 개체에서 처음 고안된다는 사실은 아주 홍미 있는 일이다 (Kaw ai 1965). 행동 성향에 대한 전수가 전제된 전달을 다른 여러 척추동물에서 도 발견할 수 있다. 쥐는 집단 구성원으로부터 특정한 먹이장소를 선호하고 다른 것을 피하는 것을 배울 수 있다. 박새는 1930 년대 말 영국에서 우유병의 은박지 뚜껑을 여는 방법을 배웠다(그림 8 기1). 이 기술은 분명히 서로가 독립적으로 중복되어 생겨났으며 여러 중 심권에서 급속히 전역에 퍼져갔다. 이런 섭식 전수 혹은 먹이 전수 이의에도 장소 선호도 한 세대에서 다음 세대로 전달될 수 있다. 북
 그림 8-11 우유병의 은박지 뚜껑을 여는 방법을 전수하는 영국 박새.
그림 8-11 우유병의 은박지 뚜껑을 여는 방법을 전수하는 영국 박새.
아메리카의 큰뿔영양은 오늘날까지도 산재된 서식 지역 사이의 좁은 길을 통해 이동한다. 이 길은 이 영양들이 이미 빙하기 말기에 알고 있었댜 지금 이런 많은 길들은 그 동안의 결과로 생긴 삼림상태로 인해 이 영양들에게 완전히 적합지 않은 지역을 가로질러져 있다. 많은 철새 및 이동성 어류의 〈장소신뢰성〉도 역시 이러한 장소 전수 에 대한 예로써 제시할 수 있다. 그 밖에도 자기 종의 소리의 인식도 어린 동물들이 제각기 아버지 로부터 배울 때 전수를 통해서 한 세대에서 다음 세대로 전파될 수 있다 무엇보다 많은 명금류들은 이러한 〈노래 전수〉가 알려져 있 댜 이런 전수의 안정성에 대해 하나의 인상적인 예가 약 100 년전 뉴질랜드로 귀화한 유럽산 몇몇 명금류에서 찾아 볼 수 있다. 최근 에 녹음기와 음성분석기(소나그램)를 이용한 비교 연구는 뉴질랜드의 개체군의 노랫소리가 유럽 원래의 개체군의 노랫소리와 오늘날에도 일치하고 있음을 보여주고 있다. 연구된 종들 가운데 최소한 2 종은 노랫소리의 대부분이 학습됐음이 알려졌다. 여기서 학습된 정보는 완전한 지리적 격리에도 불구하고 원래의 개체군으로부터 수백년 이 상을 변화되지 않고 전달되고 있다(Thi elcke 1974). 그래서 새 노래의
방언의 형성과 전달에서 전수는 중요한 역할을 하고 있다. 물체 관련 정보 전달에 비해 물체와 관련이 없는 전수의 정보 전 달은 상당한 차이를 보인다. 이 전수의 정보 전달의 경우 정보는 특 정 물체인 먹이원의 상태와 확보에 대해, 또 먹이원의 부재에 대해 서도 전달될 수 있다. 물론 이것은 관련 신호들이 정보 전달의 인식 표시에 사용되는 것을 전제로 한다. 이런 가능성은 꿀벌의 〈춤언어〉 에서 실현되었다. 춤언어를 통해서 꽃을 방문했다가 집으로 돌아오 는 일벌들은 자신들의 동료들에게 특정한 동작의 연결에 의해 먹이 원의 상태, 거리 그리고 풍부함에 대한 정보를 줄 수 있다. 먹이원 이 벌집에서 가까이 있을 때 나타나는 〈원형춤〉은 벌집 위를 재빨리 그리고 눈에 띄게 빙빙 돌아서 단지 다른 일벌들이 이 춤추는 일벌 에 주의를 돌리게 하여 이 춤추는 벌 뒤를 따라서 춤을 추도록 유발 한다. 이들이 벌집을 나갈 때 방향을 향할 수 있는 먹이원에 대한 원래의 정보는 집에 돌아온 일벌의 몸에 묻은 꽃냄새에 함유되어 있 다 . 일벌들은 더듬이로 뒤를 따라 다니면서 냄새의 정보를 감지한 댜 이에 비해 좀 더 멀리 떨어져 있는 먹이원을 나타내는 〈꼬리춤〉 은 필요한 정보를 전달할 수 있다. 이때 벌들은 수직으로 놓여있는 봉방 위에서 직접 표현할 수 없는 태양의 방향에 대해 먹이 위치의 상태를 중력의 도움을 빌려서 나타내고 있는 놀라운 전환능력을 시 행하고 있다. 춤추는 벌은 OO자 를 그리면서 돈다. 그들은 복부로 재 빨리 꼬리를 움직이게 한다. 꼬리춤을 위로 .향 하여 추면 먹이는 직 접 태양의 방향에 있게 되고, 만일 벌들이 수평에서 오른쪽 혹은 왼 쪽으로 기울어진 각도로 춤을 추면, 먹이원으로 날아갈 때 태양 위 치와 해당 각도를 이루게 된다. 그 밖에도 꼬리춤의 선을 그리면서 움직이는 속도를 통해서 먹이가 위치한 거리를 나타낸다. 벌집에서 먹이원이 멀면 멀 수록 꼬리춤은 보다 천천히 그리고 보다 힘있게 추게 된다. 원형춤에서 꼬리춤으로 넘어갈 때는 집에서 먹이장소의
거리가 50-lOOm 에 달한다 (Fr i sch 1965). 물체와 관련이 없는 비슷한 정보전달이 침팬지 실험에서 행해질 수 있다 . 침팬지들은 음절을 끊어서 정확히 발음할 수 있는 능력이 없기 때문에 구두의 의사소통을 하지 못하다. 그러나 그들온 청각장 애자가 수화를 사용하는 것처럼 손신호롤 학습하며, 이러한 방법으 로 자기의 사육사와 의사소통을 할 수 있다. 침팬지의 이런 · 수화와 꿀벌의 춤언어 사이의 공통점은 정보가 관련 물체가 없이 전달될 수 있는신호사용에 있다. 전수를 분류할 수 있는 두번째 분류 기준은 참여 배우자의 이해와 서로 관련되어 있다. 직접적 형태에 있어서 경험이 없는 배우자는 부모나 혹은 그 집단의 소속원으로부터 직접 배운다. 간접적인 형태 는 많은 기생성 곤충에서 일어나는 것으로, 정보 전달이 암컷이 특 정 숙주 식물이나 혹은 숙주 동물에게 자신의 알을 낳아 알에서 깨 나온 유충이 장차 성체가 되어 산란할 때 다시 찾기 때문에 해당 숙 주종에 매우 견고하게 고정되어 이루어져 있다. 이런 식으로 이런 선택은 연속된 두 세대의 소속원들이 서로 협력하지 않고도 한 세대 에서 다음 세대로 전해지게 된다. 이런 전수의 형태에 대한 전제 조 건은 단순히 새끼(알)에 대한 어미의 배려가 있어야 한다. 직접적 전 수는 물체와 관련되어 있는 경우와 물체와 관련이 없는 경우로 두 가지 가능성이 실질적으로 나타날 수 있는 반면, 간접적 전수는 항 상 물체와 관련되어 있다. 지금까지 열거한 모든 전수는 한 종의 소속원들간에 일어나고 있 댜 그렇지만 어떤 경우에는 소위 종간의 전수도 있다. 가장 찰 알 려진 예는 생득적 자기 노래 성분 이외에 숙주종의 소리를 학습하여 자신의 노래로 재구성하는 천녀새의 수컷들이 있다. 여기서 각 세대 의 정보는 다른 종에서 새로이 넘겨 받게 된다. 전수는 기본 구성이 꿀벌의 춤언어처럼 생독적인 고정된 암호 시
 ``` : 룬零뺑혼 際誕 효』 :쇼, 奪-i , '
``` : 룬零뺑혼 際誕 효』 :쇼, 奪-i , '
스템으로 구성되어 있을 수 있고, 많은 장소전수와 섭식전수와 같이 각인에서 기인되어 있을 수 있으며, 그 밖에도 일본원숭이의 감자씻 기처럼 모방을 통해 학습의 기초에 근거해서 발전될 수 있다 . 4,8 도구사용 많은 종들은 화장할 때 , 먹이를 손에 넣을 때 그리고 먹이를 조리 할 때 혹은 기타 다른 것과 관련해서 물체를 도구로 사용하는 능력 을 갖고 있다 (B i scho f 1979, Guil m et 1977). 코끼리는 코로 막대기를 들 고 등을 긁는다. 민머리독수리는 타조의 알을 돌멩이로 깨뜨려 먹는 다(그림 8-12). 침팬지는 표범의 박제를 막대기로 치거나 높이 매달 아 놓은 바나나에 도달하기 위해 상자를 위로 쌓아올린다. 도구 사 용에 대해 특별히 많이 인용하고 있는 예는 갈라파고스의 딱따구리
핀치가 된다. 이 새는 부리로 나뭇가지나 선인장의 가시를 집어 벌 레를 파낸다. 재봉개미는 분비선에서 실을 뽑는 자신의 유충을 턱사 이로 물고 두 개의 인접된 나무잎 사이룰 이리 저리 오간다. 이렇게 해서 나뭇잎 가장자리는 실로 밀집되게 짜서 서로 붙게 만든다. 이 개미들도 살아 있는 물체를 도구로 사용한다 . 이들은 성체가 되면 더 이상 실을 생산할 능력을 갖지 못한다 (Holldobler 와 Wi lso n 1977). 일부 동물들은 도구를 스스로 만들거나 개선시킬 수 있다. 침팬지 는 나뭇가지를 꺾어 그것의 껍질을 벗기고 옆에 달려있는 잔가지를 없앤 다음 흰개미집의 구멍을 쑤신다. 딱따구리핀치도 자신이 가시 를적당히 자를수 있다. 도구 사용은-재봉개미에서 있는 것처럼-생득적 기초에 근 거할 수 있다. 그러나 대개는 이 도구 사용이 학습에 의해 이루어지 고 있댜 이것은 민머리독수리가 알을 돌로 깨는 것처럼 시행착오를 통해 일어날 수 있고, 그러나 그것은-무엇보다 도구가 시험해 보면서 만들어지는 것이 아니고 미리 설계되어 사용될 때-철저 히 현명한 능력의 표현으로 볼 수 있다. 종종 도구 사용의 특정한 종류는 전수를 통하여 계속해서 전달되고 있다.
제 9 장 사회행동 동물들은 단지 몇몇 종만이 사회성에 전혀 관심 없이 살아가고 있 댜 우선 이것에 속하는 바다 밑에 고착생활을 하는 하등 바다생물 체들은 생식세포를 바닷물에 방사할 뿐 동종이 함께 만나는 일은 거 의 없다. 그러나 이들 스스로가 화학물질을 방출하여 이웃과의 사이 에 시간적 동조성에 도달할 수 있어야 하기 때문에 이것으로 비로소 사회적 접촉을 이룬 셈이다. 이에 비해 대부분의 동물의 종들은 주 로 생식(짝짓기, 새끼기르기)을 위해 일생을 통해 여러 번 동료와의 만남을갖는다. 사회성이란 말은 행동학에서 중요한 용어로 사용되고 있으나 인간 사회의 범주와는 동일하게 취급하지 않는다. 동물의 사회성이란 관 련된 행동들이 종내에서 의사소통의 관계를 이루었을 때를 의미한 댜 이러한 의미에서 사회행동은 동료들 사이에서 일어나는 제반 행 동을 말한다. 결국 생물학적으로 중요한 의미를 갖는 동종내 공격행 동 역시 사회행동의 범주로 다룬다. 이 장에서는 동물들의 싸움행동 울 시작으로 성적행동, 양육행동(새끼돌보기), 집단행동으로 나누어 동물들의 사회행동을 살펴보기로 한다.
l 싸움행동 l.l 싸움행동이란? 싸움행동은 기능적으로 공격과 도피로 분류된다 . 가장 광범위한 의미로 공격행동은 공격적인 모든 행동을 내포하고 있다. 여기서 공 격적인 행동이란 공격 준비, 공격성 혹은 공격 충동으로써 한 동물 에 내재되어 있는 동인을 가리킨다. 영어의 투기적 행동 a g on i s tic behav i or 과 동일하게 사용하지는 않는다. 오히려 투기적 행동은 공격 과 도피가 교차(예, 위협행동)되어 생기는 행동을 말한다. 공격행동은 종내에는 물론 종간의 개체들에서 나타난다. 종간 공 격 (種間攻擊i n t ers p ec ifi c a ggr ess i on) 은 마실 물, 먹 이 혹은 둥지 자리 를 놓고 방어행동과 경쟁행동을 포함하고 있다. 많은 연구자들은 종간 공격으로 육식성 종의 먹이포획 행동을 들고 있는 반면, 일부 연구 자들은 섭식 행동 그 자체를 꼽고 있다. 이것에 비해 종내 공격(種內 攻擊i n tr as p e cifi c a gg ress i on) 은 같은 종의 개 체 가 먹 이 나 배 우자와 같은 경쟁 대상물을 놓고 일어나는 경쟁의 결과에 한정하고 있다. 종간 공격과 종내 공격 사이에 인과관계가 성립하는지, 말하자면 종내 공격은 포획행동이나 포식성 동물에 대항하여 벌이는 방어행동 에서 나왔는지, 아니면 종간에 벌이는 다른 행동 영역에서 나왔는지 의견이 분분하다. 무엇보다 먹이동물을 사냥하고 죽이는 것은 계통 분류학적으로 종내 공격에서 기원된 것으로 보인다. 그렇지만 많은 사실들이 직접적인 관계를 갖고 있지는 않은 것 같다. 그래서 종종 육식성 동물에 비해 온순하다고 하는 많은 초식성 동물도 동종간의 높은 강도의 공격을 보이기도 한다. 그밖에도 이 공격행동은 종내 혹은 종간의 싸움에서 서로 독립적으로 나타나고 있다. 이 두 종류 의 공격은 신경 및 호르몬 생리 연구에서 뇌 자극을 통해 각기 다른
자극 부위에서 출발하고 있으며 호르몬을 통한 다양한 방법에도 이 두 종류의 공격에 달리 영향이 미칠 수 있다는 사실을 보여 주고 있 댜 물론 반대의 주장도 가능하다 . 개코원숭이 Pa pi o 에서 두 가지가 긍정적인 관계가 있음을 발견할 수 있다 . 한 수컷이 암컷을 차지하 기 위해 싸우는 공격 정도가 포식성 적에 대해 그 수컷이 자기 무리 를 방어하려는 공격 정도보다 더 강도가 높다 . 종내 공격 혹은 종간 공격 행동을 통합해서 생각하느냐 혹은 분리해서 생각하느냐에 대한 궁극적 인 이 론을 제시 하기 에는 아직 이 른 감이 없지 않다 (Hun ti ngfo rd 1976) . 보통 행동학적 관점에서 종내 공격 행동이 종간 공격 행동보다 더 큰 홍미를 보여 주는 것은 사실이다 . 그 이유는 종내 공격이 아주 고도로 발달된 행동을 여러 유형으로 보여 주고 있기 때문이다. 그 러므로 동종의 싸움행동에 한정지어서 그 행동의 형태와 규칙성에 대해 다루기로 한다. l.2 생물학적 의의 종내 공격에 대한 생물학적 의의는 매우 다양하다. 우선 공격행동 울 통해 한 종의 소속 개체들은 거의 같은 정도로 생활공간을 방어 하며 살고 있고 또 각기 요구되는 먹이자원을 놓고서 그 공간을 최 적으로 사용하고 있다. 이러한 사실로 공격행동은 생활공간의 요구 에 의해 최고의 집단밀도를 넘어섰을 때 그 집단에 먹이 부족이 오 기 전 과잉의 개체들에게 이동을 강요당하게 한다. 동시에 지금까지 살지 않았던 지역으로의 서식을 유도할 수 있다. 경쟁적인 동료가 공격행동으로 밀려나감으로써 공격은 번식을 위해 공간적 확장을 보 장해 주는 역할을 하며, 따라서 자연히 개체가 솜아짐으로 해서 전 염병이 퍼지는 것을 저지할 수도 있고 보호색을 지닌 종의 경우 포
식성 적에 대해 더 나은 보호의 제공을 받을 수 있다. 공격행동은 성적 선택(性的 選擇 sexual selec ti on) 을 위해 기여를 한다. 성적 선택 과정에서 공격은 가장 강하고 가장 건강한 개체가 번식을 성공하도 록 선택하게 한다. 그리고 집단 서열이 발달된 사회성 종에서는 가 장 경험이 풍부한 개체로 하여금 우두머리 역할을 하게 한다. 물론 모든 동물의 종에서 모두가 같은 기능을 하는 것은 아니다. 각 종들 은 각기 또다른 종내 공격의 기능을 갖는다. 이렇게 여러 가지의 생물학적 기능에 반하여 개체와 그 종에게 몇 몇 결정적인 손실도 있다. 즉 싸움을 하는 동물들은 쇠약해지거나 부상을 당할 수 있다 . 그리고 그들은 좀더 남의 눈에 띄거나 아니면 덜 관심을 끌 수밖에 없기 때문에 위험에 처할 가능성은 더 높다. 게다가 싸움을 하는 동물들은 다른 활동을 하지 못하기 때문에 시간 을 소비할 수 있다. 이론적으로 또 위험이 될 수 있는 것이 있다. 육체적으로 힘이 약하거나 비교적 경험이 많지 않은 어린 개체들은 동종의 공격에 의해 심한 침해를 받게 되어 자기 자신의 번식 능력 올 상실할 수도 있다. 이러한 이유에서 한 동물의 종내 공격 정도와 형태가 개체에 대해서 생존을 위한 요구, 그 종 전체에 대해서는 밀 도조절 요구에 근거 를 둔 선택 압 selecti on p ressure 에 대 항하여 발전 하여 왔다. 이런 타협 형태는 한편으론 종마다 공격의 동인 크기가 다양함을 보여 주고 있는 것이고 다른 한편으론 많은 종들에서 공격의 이점이 완전 보장되나 손실을 줄이는 행동으로 발전해 왔다는 사실이다. 여 기에다 위협행동, 위엄행동, 복종행동, 달램행동, 텃세행동, 그리고 개체 간격은 위험하지 않은 싸움 형태에 속한다(Ki ng 1973, Lorenz 1983, Tin b erge n 1968, Hu ni n gfo rd 와 Turner 1987, Archer 1988) .
l.3 위협행동과 위엄행동 위협행동t hrea t behav i or 은 실제 싸움과 비슷한 기능을 하면서 위 협행동 관여자는 보다 작게 해를 입고, 결국 공격으로 인한 상해를 줄일 수 있다. 이 행동의 목적은 경쟁자를 노려봄으로써 실제 싸움 이 일어나기 전 물러나게 하는 데 있다. 여기에는 특히 두 가지 위협행동의 형태가 자주 일어난다. 한가지 는 위협 가해자는 대개 자기 몸을 크게 만든다. 다시 말하면 털이나 깃털을 세운다든지, 피부 주름이나 아가미 뚜껑을 밖으로 들어나게 한다든가, 아니면 성대를 부풀려서 몸을 크게 만드는 것이다(그림 9-1). 또 위협 가해자는 의도 동작을 연결시킬 수 있도록 무기(이 빨, 부리, 뿔 동)를 눈에 띄게 만든다. 실제 싸움 행위는 이러한 동작 울 취해 특이한 몸색깔을 첨가시키는 경우가 많다. 동인 분석을 해보면 위협행동은 두 개의 혼합된 충동이 내재되어 있는 것을 발견할 수 있다. 죽 공격하려는 성향과 도피하려는 성향 이 함께 관여하여 위협행동을 나타낸다. 많은 경우 이미 이에 관련된 행동들이 상반감정의 공존 형태로 나타나고 있다. 하나의 예로, 붉은 부리 갈매기 Larus r i d i bundus 의 아주 가장 두터 운 목은 목근육을 함께 긴장시켜서 만드는데, 이 때의 근육 상태는 상대방에게 부리로 찍을
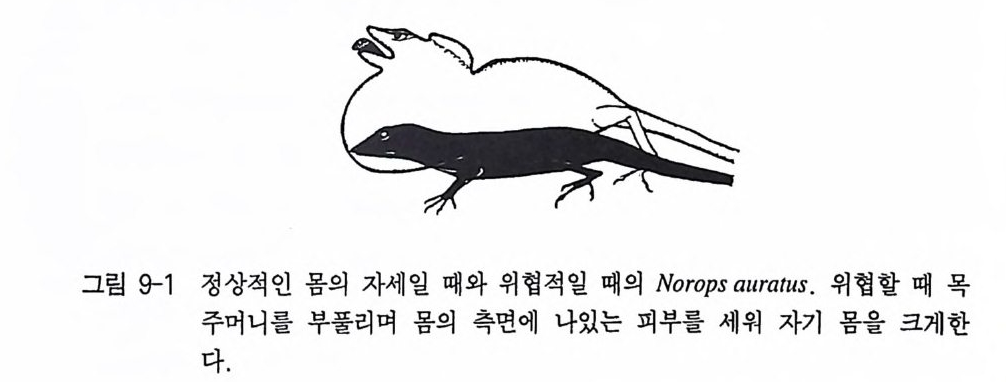 그림 9-1 정상적인 몸의 자세일 때와 위협적일 때의 Norop sa uratu s . 위협할 때 목
그림 9-1 정상적인 몸의 자세일 때와 위협적일 때의 Norop sa uratu s . 위협할 때 목
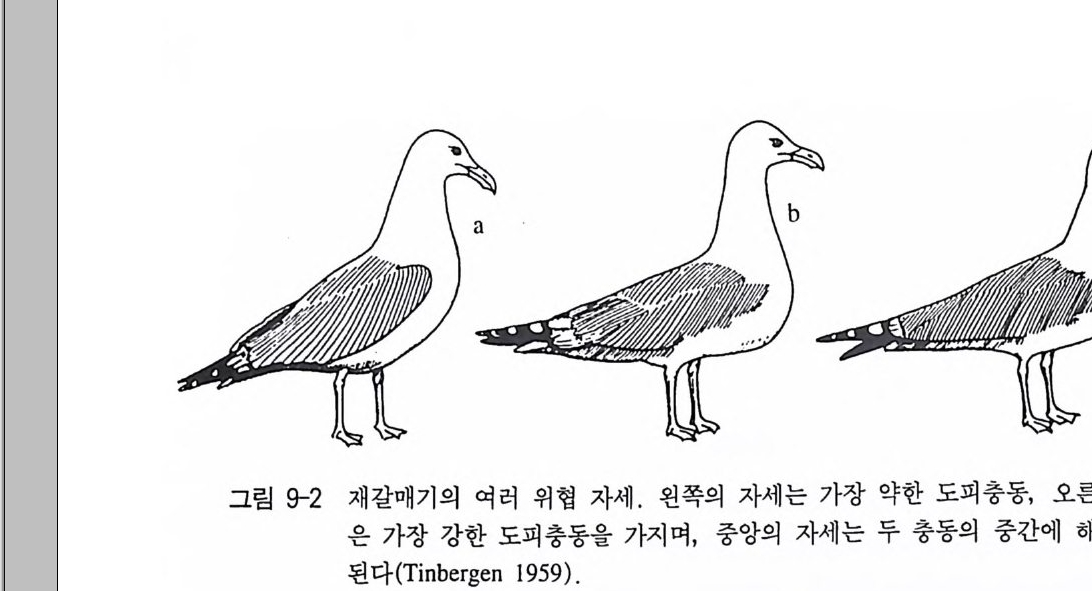 그림 9-2 재갈매기의 여러 위협 자세. 왼쪽의 자세는 가장 약한 도피충동, 오른쪽
그림 9-2 재갈매기의 여러 위협 자세. 왼쪽의 자세는 가장 약한 도피충동, 오른쪽
수도 있고 경우에 따라서는 머리를 뒤로 쉽게 이동시킬 수 있는 가 장 적합한 자세다 . 그 밖에도 많은 동물들은 여러 단계의 위협행동 울 갖는 것이 일반적이다(그림 9-2). 위협행동 영역에서 나온 많은 동작 유형과 몸의 자세는 이중의 기 능을 갖는다. 동성인 경쟁자에 대해서는 물리치거나 위축당하게 하 는 기능과 이성에 대해서는 유인 역할을 한다. 종종 구애동작에 한 정 된 행 동 요소들이 위 엄 행 동 epi ga m ic d i s p la y으로도 나타난다. 이 위엄행동은 주로 수컷에서 나타난다. 물론 위엄행동이냐 위협행동이 냐의 구분은 행동의 기능이 정확히 알려진 경우에만 가능하다. 지금 까지 이 두 의미가 항상 정확하게 구분되어 있지는 않으나 위엄이라 는 말은 종종 위협행동의 동의어로 쓰여왔다. 위협행동이 경쟁자 격퇴나 배우자 유인이 결합된 것이라는 의미는 분명하다 위협행동은 번식 초기 즉 터를 확보하고 방어하는 데에서 다발적으로 나타난다. 그것은 아마 터주인에게는 번식 준비로 표현 되며, 짝이 없는 암컷에 대해서는 긍정적 반응을 하게 하는 자극행 동으로 표현된다. 그러므로 위엄행동은 구애할 때 특히 많은 동물들 이 짝울 이 룰 때 하나의 중요한 역 할을 하고 있다 (E i ble-E i bes f eld t
1957, Lorenz 1963 ) . ,.4 복종행동과 달램행동 종내 공격의 부정적인 결과를 피하고자 하는 위협행동과 같은 기 능을 할 수 있는 또다른 행 동들이 복종행 동 submi ss iv e g es t ure 과 달 램 행 동 app e asement g es t ure 이 댜 이 두 행 동은 공격 을 누그러 뜨리 게 하는 목적이 있지만 형태나 효과면에서는 완전히 다르다. 복종행동 은 종특이적 위협행동과 완전히 대치되는 것으로 표현된다. 몸크기 는 몸의 부속체를 접거나 구부려서 작게 만든 다음 위험스런 무기나 다른 공격적 해발인으로부터 보호를 받는다. 또는 반대 방향으로 몸 울 돌려 특히 급소가 될 만한 몸 부위(목, 목덜미)를 적으로부터 피하 는 것이다 . 몸 색깔의 급속한 조절이 가능한 동물들(예, 오징어, 일부 파충류)은 특별한 움직임없이도 해당 색깔을 바꿈으로써 공격적 해발 인으로 작용하는 색깔 신호를 사라지게 할 수 있다. 복종행동은 종 내싸움에서 하위자에 의해 취해지며 그것은 매우 효과적으로 공격을 저지시킬 수 있는 힘을 갖고 있다. 결국 최소한 그 싸움은 잠시동안 정지된다. 그래서 하위자가 상해를 입거나 죽움을 당하는 것을 저지 시키는 물기 억제 기능을 갖는다 (Da th e 1972). 복종행동은 싸움을 잘 하는 동물에서 잘 발달되어 있고, 무엇보다 도피가 불가능한 상황에 서 잘 나타난다(그림 9-3). 달램행동의 기능은 복종행동과 반대로 이전에 있던 공격 신호를 저지하는 데 있는 것이 아니고 일반적으로 어미와 새끼 관계 혹은 성적 영역에서 나오는 것으로 단지 불화를 해소하는 데 있다. 이 달 램행동은 짝을 짓거나 무리를 지어 사는 동물의 종에서 인사gr ee ti n g 로 동료들이 좀더 가까이 접근하게 하거나 함께 머물 수 있도록 한다. 복종과 달램 행동 사이의 아주 정확한 구분은 두 요소가 서로 결
 • -一 --· -
• -一 --· -
합될 수 있기 때문에 항상 가능한 것은 아니다. 그러므로 많은 연구 자들은 서로 구별하여 사용하지 않고 두 개념을 동의어로 사용하고 있다 (Lorenz 1983, Schenkel 1967, Walth e r 1979, Wi ck ler 1966) . 1.5 텃세 위협행동 그 자체가 시간적으로 제약을 받는다면 보다 효과적인 방법으로 공격에 의한 상해를 없앨 수 있는 것이 있다. 여기서 어떤 구역 설정이 전제되어야 한다. 이 구역의 설정 초기에는 주로 싸움 행동과 위협행동을 통해 방어가 이루어지지만 나중에는 싸움없이도 싸움에 해당되는 효과를 볼 수 있다. 종내 공격의 주 목적은 스스로 의 손실을 줄이면서 공간을 서로 분배하는 데 있다. 이렇게 구역과 관련해서 일어나는 일련의 경쟁적 행동들을 우리는 텃세행동t e rrit o rial behav i or 이 라 한댜 그렇지만 이 텃세행동이라고 해서 전체의 다툼 횟수만을 줄이지는
못하고 오히려 시간이나 에너지 소비를 많이 줄이는 일로 바뀔 수 있다. 결국 이것만으로도 개체 혹은 집단에게 부수적인 이점을 가져 다 준댜 일반적으로 구역은 어떤 동물이 새끼기르는 시기 훨씬 전 부터, 심지어는 짝짓기 이전에 형성되는 것이 보통이다 . 그러므로 짝짓기나 새끼기르기 행동들이 표현되기 전부터 구역 설정과 관련된 공격 영역의 행동들이 이미 최고의 수준을 넘을 수 있다. 어떤 지역에서 한 개체가 동성인 동종의 경쟁자를 내쫓음으로써 선택적 방어를 하는 구역이 존재하는데 우리는 이것을 터 t e rrit o ry라 부른다. 비록 이 터가 그 구역의 터 주인에게 특정한 생활공간을 보 장해 주는 일을 하지만, 동물들의 생활방식이나 식성에 따라서 터의 기능은 다소 차이가 있을 수 있다. 그러나 다음과 같이 터의 일반적 인 주요 기능을 요약할 수 있다. ® 터는 터 주인에게 충분한 먹이를 보장해 주는 역할을 한다. ® 터는 공동의 만남의 장소로 짝짓기를 용이하게 해준다 . ® 터는 주변의 익숙함에 의해 적이 나타났을 때 도망갈 장소에 신속 히 도달하게 해준다. ® 터는 같은 이유로 먹이를 찾는 데도 쉽게 해준다. ® 터는 동종에 의해 침해받는 것을 막아 주며 그것을 통해 번식하는 데 불필요한 방해를 없애 준다. 대부분의 경우에 터는 여러 개의 기능이 동시에 작용한다. 구체적 으로 어떤 기능이 종특이적으로 발달하게 한 선택의 이점이 되었는 지, 또 오늘날의 기능 가운데 어떤 것이 좀더 진화되어 나타났는지 항상식별할수는 없다. 텃세행동은 척추동물의 모든 강(綱) 수준에서 나타나며 무척추동 물에서는 거미불가사리, 게, 거미 그리고 특히 곤충(귀뚜라미, 잠자
리, 사마귀 등)에서 나타난다 . 용어상 터와 구별되는 것으로 행동권 home ran g e 과 총생 활권 tot a l ran g e 이 있다 행 동권과 총생 활권 이 자 주 서로 비슷한 의미로 사용되기도 하지만 용어상 엄밀하게 구별할 경우 미세한 차이를 발견할 수 있다. 총생활권이란 한 개체 혹은 조 직적으로 구성된 집단이 일생 동안 지내는 전체 영역을 말한다. 이 경우 총생활권은 다니는 길과 행동권을 포함하고 있다 . 이에 반해 행동권은 한개체, 한 쌍 혹은 한 집단이 규칙적으로 머무는 곳으로 터와 구별되는 점은 동종의 개체에 의해 추방당하지 않는다는 것이 다. 종종 행동권은 두 개 이상의 터 사이에 위치해 있는 중간지역을 지칭하기도 하고 또 자연자원(물이 있는 장소, 휴식처, 잠자리)을 갖는 지역을 말한다. 이런 공간적 경계는 단순히 개체들 혹은 집단들이 서로가 만나는 것을 꺼리는 곳에 의해 결정될 수도 있다. 물론 연중 서로가 회피하는 지역경계 혹은 실제 방어하는 구역이 바뀔 수 있기 때문에 두 개념사이의 경계가 항상 명확한 것은 아니다(그림 9- 4) (Mit an i와 Rodman 1979, Walth e r 1967, 1979) 1) 터의 분류 동물계에 존재하는 터는 여러 관점에서 분류될 수 있다. 각 분류 체계에 따라 다음에서 전형적인 몇몇 예를 제시하고자 한다. 기능에따른분류 이미 제시한 터 기능의 대부분이 섭식과 번식의 관계에서 설명되 었다 그래서 대략 먹이터, 번식터 그리고 두 개의 기능을 모두 갖 고 있는 터로 구분할 수 있다. 그러나 그 경계는 항상 명확한 것은 아니다. 〈먹이-번식터〉 이것은 가장 혼한 터의 형태로 어미 및 새끼 모 두에 의해 요구되는 번식지역과 먹이지역을 동시에 만족시킬 수 있
 A 。방없 어는구 배역회이구 역(터)
A 。방없 어는구 배역회이구 역(터)
어야 한다. 그래서 먹이와 번식에 대한 구분이 거의 안 된다.(그림 9-5A) 〈먹이터〉 조류는 번식 기간 외에도 터를 갖는 경우가 많다. 특히 겨울 철새의 경우 겨울 서식처가 순수한 번식터와는 대조적으로 사 용된다. 이때의 터란 개체들이 먹이를 구하는 먹이터로만의 기능을 갖는다. 포유류와 어류에서도 먹이터를 따로 갖는 경우가 많다. 〈번식터〉 터가 번식에 비중을 두어 기능을 할 때가 많은데, 이런 터 형태는 주로 짝짓기의 장소 그리고 새끼기르는 장소로 국한하여 쓰인다 무엇보다 번식터는 번식집단을 이루는 바다조류에서 발견되 며 이들은 둥지의 가까운 주변만을 방어하고 먹이터는 따로 갖지 않
 `.`.·..i i`, . .얗經 ...... \ I 際홉......,.·. .. ... ..
`.`.·..i i`, . .얗經 ...... \ I 際홉......,.·. .. ... ..
는다 이때 이 터는 단순히 새끼를 기르는 장소로만 기능을 한다(그 림 9-SB). 많은 조류의 짝짓기터는 번식 영역 내에서 또다른 목적을 갖는댜 많은 뇌조류의 수컷들은 동종의 경쟁자에 대항하여 구애장 소를 방어하여 그 터를 교미장소로 사용한다 . 이 터는 경쟁자로부터 방해를 받지 않고 안전하게 교미하도록 보장해 주고 있다. 이들은 지속적으로 짝 형성을 하지않고 교미하는 그 시간만 암수가 함께 지 내게 된다 이런 경우 우리는 이 터를 구애장 lek 이라고 표현한다. 수 컷들은 구애장을 하나나(큰뇌조) 여러 개(검은멧닭)를 갖기도 한다 (Vos 1979). 기제류(예, 얼룩말), 코뿔소 그리고 하마와 같은 일부 포 유류들은 수컷의 텃세권이 주로 짝짓기 행동할 때 나타난다. 이들은 터 크기와 먹이 사이에 직접적인 관계는 발견되지 않는다(그림 9-6).
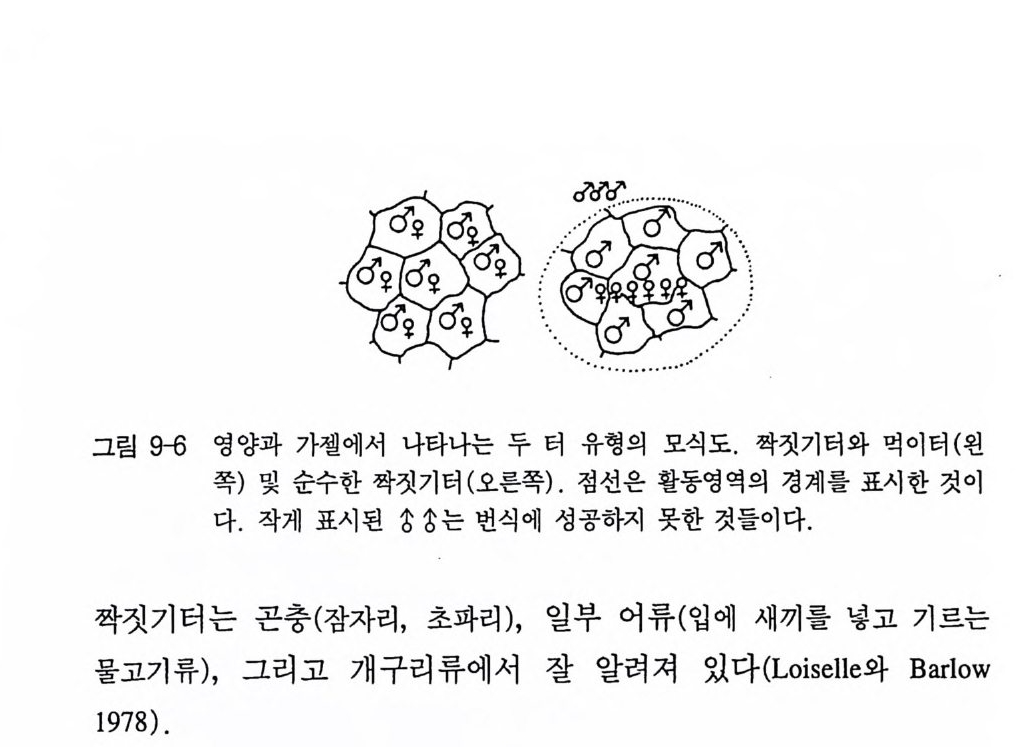 ••••····· ·· ••• •• •
••••····· ·· ••• •• •
터 소유에 따른분류 〈단독터〉 먹이터와 번식터는 보통 한 개체에 의해서만 차지되고 방어된다. 새끼기르기는 암수 중 한쪽만이 그 책임을 맡는 종(벌새, 큰 가시고기)에서 이런 단독터를 갖는다. 〈짝터〉 번식을 목적으로만 존재하는 터는 암수 두 마리에 의해 차지되는 경우가 많다. 이때 방어의 형태는 다양하다. 두 마리가 함 께 터를 방어하는 경우, 수컷 한 마리에 의한 방어, 아주 예외적으 로 암컷(일부 키클리드, 세가락메추라기)에 의해서만 방어되는 경우가 있다. 단독터와 짝터 사이의 중간 위치에 있는 경우는.일부다처제인 키클리드과의 Lamp ro log as con g olens i s 의 터에서 볼 수 있다. 이 물고 기는 한 마리의 수컷 터내에서 각각의 암컷들이 자기소유의 버금터 를차지한다. 〈집단터 > 많은 포유류(영장류, 식육류, 설치류)와 조류(코뿔새, 피 리새)는 하나의 터를 한 집단이 공동으로 소유하고 있다. 여기에서
암수 그리고 그 밑에 있는 자손들이 터 방어에 참여할 수 있다. 터 소유기간에 따른분류 많은 동물들은 일년중 특정 기간에만 텃세를 갖는다. 즉 그것은 그 터가 번식 영역 내에서 기능을 수행하는 경우가 그렇다. 그리고 이것은 온대성 기후에 사는 대부분의 동물에서와같이 시간적으로 번 식이 한정되어 일어나는 동물에게서 두드러지게 나타난다. 반대로 겨울터는 이런 번식기 이의에서만 존재한다. 단지 일부 종(예, 여 우, 오소리)들은 일년내내 자기 터에서 살기도 한다. 이에 비해 열대 성 기후에 사는 동물들은 온대성 동물보다 터를 더 오래 지키는 경 향이 있다. 무엇보다 조류와 포유류의 많은 수가 이에 해당된다. 그 밖에도 극히 짧은 기간의 터도 존재한다. 아프리카에 사는 검 은꼬리 누 Connochaetes t aur i nus 의 이 동 무리 의 경 우 수컷 이 수분 이 내에 짝터를 확보하여 그 터를 몇 시간 혹은 며칠 동안만 갖고 그 무리가 다시 이동할 때 터는 없어지게 된다. 철새들도 여름-겨울 서식처를 오가는 도중 중간지역에서 잠시 텃세 행동을 보여줄 수 있 댜 왕잠자리과인 Aeschna c y anea 는 시간적 텃세 행동을 보여 주는 좋은 예가 된다. 수컷들은 각기 짝짓기 장소에서 1 0-40분 정도 머물 고 그곳에서 암컷과 교미를 시도한다. 이 과정에서 터 주인은 비행 공격으로 다른 수컷들을 몰아낸다. 이 시간의 끝무렵에는 싸움하려 는 충동이 사라지고 그 터주인은 새로운 한 마리 수컷에 의해 추방 당하거나, 결국에는 외부의 압력없이도 이 짝짓기터를 떠난다. 포유 류 가운데 이와 비슷한 〈시간 터〉가 있는데, 치타와 야생화된 집고 양이에서 나타나고 있다. 이들은 여러 개체 혹은 여러 무리가 불규 칙적인 간격으로 어느 정도 짜여진 시간표에 의해 각기 차례로 같은 장소에 머문다. 그렇지만 이들은 동시에 만나지는 않고 시간대를 달 리하여 같은 터를 사용한다. 그 이유는 이들이 선행자가 남겨 논 냄
 그림 9-7 한 마리의 포유류의 터 내부의 모식도 . 굵은 선은 터 경계, 가는 선은 각
그림 9-7 한 마리의 포유류의 터 내부의 모식도 . 굵은 선은 터 경계, 가는 선은 각
새 흔적의 신선하고 오래된 정도에 따라 특정 터 부위가 비어 있는 가를 알 수 있기 때문이다 (Le y hausen 1975). 2) 터 구조 여러 동물 종들 가운데에, 특히 포유류의 경우에는 하루의 일과가 터 내부의 일정한 자리와 관련되어 일어나고 있다. 주로 다니는 길 을 통해 상호연결되어 있는 이런 자리에는 잠자리, 먹이장소, 오줌 과 똥을 누는 자리, 물 마시는 곳, 목욕장소, 웅덩이, 나무들, 감시 용 언덕, 그리고 은신처 등이 있다. 터 내부에서 특히 중요한 중심 자리는 집 혹은 둥지가 된다. 공간적으로 매우 확장된 터는 목적지 와 길 연결 체제의 형태를 갖고 있다. 이런 목적지 사이에 위치한 지역들은 거의 (혹은 전혀) 방어되지 않는다. 엄밀히 말하면 이런 지
역은 더 이상 자신의 터에 속하지 않는 것이다(그림 9-7). 3) 터 경계 터 크기는 어느 정도 종특이적이라고 할 수 있다. 그래서 그 종의 몸 크기와 식성에 따라 터 크기가 달라진다. 일반적으로 번식터가 먹이터보다 작다. 몸 크기가 큰 종은 작은 종보다 큰 터를 갖고, 육 식동물이 초식동물보다 큰 터를 갖는다. 터 크기는 다양한 생활방식 으로 사는 근연종일 경우에도 분명한 차이를 보여 준다 . 초식동물 가운데 가장 큰 터는 그레비얼룩말과 들당나귀이고 육식동물 가운데 는 호랑이와 사자가 가장 큰 터를 갖는 것으로 알려져 있다 . 이들의 터면적은 대략 수백 제곱미터에 달한다. 종내에서도 터 크기는 개체간 큰 차이를 보이고 있다. 나이가 많 거나 경험이 풍부한 수컷이 젊은 수컷보다 흔히 더 큰 터를 사용한 댜 그리고 터는 최적의 생활조건일 때가 나쁜 생활조건일 때보다 작을 수 있다. 특히 먹이나 적의 위협과 같은 요인들은 집단밀도에 큰 영향을 미친다. 따라서 어떤 한 지역에 개체가 밀집하여 서식하 게 되면 될수록 각각의 터는 어쩔 수 없이 더 작아져야만 한다. 이 런 경향은 터의 기능이 완전히 보장되고 항상 계통발생학적으로 형 성된 최소의 일정 크기까지만 가능하다. 이런 최소 크기의 유지는 터 주인의 방어충동에 의해서 보장된다. 방어충동은 터의 중앙에서 부터 외각으로 향해감에 따라 줄어드는 반면 도피충동은 증가한다. 그래서 터 외각 지역은 터 중앙에서보다 훨씬 덜 방어를 하며 경쟁 자의 공격의 빈도가 높거나 강할 때는 그 지역을 포기하고 떠난다. 이에 비해 터 중앙에서는 공격충동이 매우 커서 어떤 상황에서도 결 코 포기하려 하지 않는다.
 \:'--틀-----鉉------~----:-~- -,--_\-___\ __\___二__二 .,. 〈-_크’/ / I
\:'--틀-----鉉------~----:-~- -,--_\-___\ __\___二__二 .,. 〈-_크’/ / I
4) 터 표시 행동 터가 기능을 하기 위해서는 이미 확보된 지역에 표시를 해두어야 만 한다. 이것은 다음과 같이 감각형식에 따라 분류할수 있다. 〈시각적 터 표시〉 하나의 터는 터 주인이 그안에서 시각적으로 모여줌으로써 표시될 수 있다. 이때 노출된 자리를 찾아서 경쟁자에 게 특이한 몸의 움직임이나 자세를 보여 주어 눈에 띄는 신호를 보 낼 수 있다. 광택의 색을 띠는 물잠자리 Colop ter y x 수컷은 정해진 자리에 앉았다가 터의 경계선을 따라 눈에 띄는 원형비행을 하며 난 다(그림 9-8). 영양 수컷은 다른 개체의 눈에 띄도록 흰개미집 언덕
이나 다른 높은 곳에 올라감으로써 터 표시를 한다. 〈청각적 터 표시〉 소리를 낼 수 있는 동물들은 소리로써 터 표시 를 한댜 가장 잘 알려진 예는 명금류의 노랫소리로 이 새들은 터 설정시기에 특히 자주 울어대며 이미 확보된 지역으로부터 새로이 도착한 개체들을 물리친다 . 기각류(바다표범, 바다코끼리), 영장류(짖 는원숭이, 긴팔원숭이, 오랑우탄), 무미류 그리고 도마뱀(도마뱀붙이)들 도 소리로 터룰 주장한다. 이런 청각적 터 표시 가운데 직접 성대에서 소리를 내지 않는 동 물들도 있다. 딱따구리는 부리로 나무를 두드려 소리를 내고 깍도요 는 꼬리깃을 펼쳐 공기중에 진동시켜 소리를 낸다. 그밖에 게들도 두드리는 신호나 진동 신호로 터 표시를 하는 것으로 알려져 있지만 아직 이에 대한 해석은 정확하지 않다. 〈후각적 터 표시〉 포유류는 흔히 냄새물질에 의해서 터 표시를 한 댜 이들은 원래 몸에서 분비한 물질을 사용한다. 물론 이 물질분비 가 터 표시 자체를 위해서는 부수적인 기능을 갖기도 하지만 (오줌, 똥, 침), 터 표시 목적으로만 특별히 분비선에서 만들어지기도 한다. 하마는 똥울 뿌려서, 유대류와 설치류는 침을 사용하여, 개, 고양이 과 동물들, 코뿔소, 원원류, 그리고 많은 설치류가 오줌으로 터를 표시한다. 터 표시 목적으로만 분비되는 분비선은 혼히 항문 근처에 있거나(족제비과 동물의 항문선) 머리(有踊類의 前眼脈)에 위치해 있다. 그밖에 다른 몸 부위에서도 분비물질이 생성된다. 그러나 많은 동물 둘이 여러 분비선을 함께 사용한다. 터 표시 참여에 따라서 암수가 함께하기도 하고 수컷 혼자서 터 표시를 하기도 한다. 혼히 이런 터 표시는 번식기에만 한정하여 완전한 기능을 발휘하기도 한다. 이때 의 분비물은 노출된 부위(나뭇가지의 끝, 풀잎의 상단 부분)에 주로 바 른다(그림 9-9). 많은 포유류의 경우 냄새표시는 이중의 목적을 갖 는다 즉 경쟁자에 대한 터 표시뿐만 아니라 동시에 터내에서 터 주
 취선
취선
인이 정위케 하는 목적으로도 쓰인다. 특히 터의 면적이 상당히 클 경 우에는 정위를 위한 표시 기능이 중요하게 작용한다 (Brown 과 Macdonald 1985, Duvall 등 1986) . 포유류 이외의 동물들은 후각적 터 표시가 지금까지 드물게 보고 되었다. 뒤영벌 수컷은 하악선에서 냄새 물질을 분비하여 자기의 구 애터에 표시를 한다. 역시 단서성 벌에 속하는 꿀벌과의 Eug lo ssin e 벌도 일종의 냄새로 표시를 하나, 스스로 분비한 냄새가 아닌 꽃으 로 부터 모은 냄새를 이용하여 터 표시를 한다 (Pa yne 등 1986). 〈전기적 터 표시〉 물고기의 일부가 특수하게 발달된 발전기관으 로 전기적 터 표시를 한다. 아 발전기관은 전기를 일으킬 수 있는 변형된 근조직으로 되어 있다. 이 기관은 먹이를 잡고, 적에 대한 방어 그리고 진탕의 물에서 정위하는 데도 쓰인다. 적어도 이런 물 고기류가운데 아프리카에 사는 전기뱀장어는 동종에 대항하여 터의 경계선에서도 발전을 한다. 〈결합된 터 표시〉 터 표시를 하는 데에 있어서 많은 경우 동시에
여러 감각형식이 요구된다. 많은 새들은 노출된 장소에서 터 노래를 부르거나 날면서 노래를 하여 청각적 표시와 시각적 표시가 결합되 어 일어난다 후각적 터 표시 경우에서도 오줌, 똥 혹은 선분비에다 눈에 띄는 몸의 자세나 움직임이 결합하여 시각적 신호가 참여하게 되는 셈이다. 〈터 표시 형식에 따른 고찰 〉 터 표시를 위해 사용되는 각각의 감 각형식은 그 나름대로 이점을 갖고 있다. 이 이점은 각 감각형식의 성격에서뿐만 아니라 이 감각형식을 사용하는 종의 생활방식(주행성 혹은 야행성, 앞이 터진 생활공간 혹은 밀집된 생활공간)에서 생겨났다. 이미 언급된 공격 상해를 없앤다는 의미에서 냄새물질은 가장 효과 적인 해결책을 마련해 주었다. 그 이유는 이 물질이 터 주인이 부재 시에도 기능을 충족시켜 주기 때문에 시각적 혹은 청각적 터 표시보 다 덜 위험스러운 데 있다. 시각적 영역에서도 이와 비슷한 하나의 예가 있다. 달랑게는 은신처, 구애장소 그리고 교미장소로 사용하기 위해 터를 표시한다. 즉 달랑게는 집을 짓기 위해 땅을 팔 때 쌓아 놓은 모래성 흙더미로 다른 수컷들에게 지속적인 신호를 보낸다(그 림 9-10). 이것은 냄새표시와 마찬가지로 터 주인이 공격적 상해롤 입는 직접적인 위험성을 제거시키는 이점을 지니고 있다. 후각적 터 표시는 다른 형식의 터 표시보다 더 지속적이라는 이점 을 갖고 있울 뿐만 아니라 동시에 여러 개의 정보를 함께 제공할 수 있다 즉 냄새신호의 화학적 조성에 의해 성별, 나이 그리고 개체간 차이가 각각 달리 나타나기 때문에 개체의 크기, 나이, 성별, 생리 적 상태, 이동 방향 그리고 체류시간의 정보를 준다 (Gosl i n g 1982). 청각적 신호 역시 개체 특이성에 대한 폭넓은 장점을 갖고 있으며 이것으로 터 이웃 개체의 상호인식을 가능하게 해준다. 그리고 서로 이웃한 개체들간의 싸움을 지속적으로 피할 수 있으며 터 소유를 위 해 위험스럽게 새로이 접근해 오는 것을 막을 수 있다 . 마지막으로
 1!! ! 1 !1 ! l ! l !l ! ! i !i !! ! l ! i i i i! l ! l ll !l l1 ifn1T I
1!! ! 1 !1 ! l ! l !l ! ! i !i !! ! l ! i i i i! l ! l ll !l l1 ifn1T I
시각적 신호의 이점은 무엇보다 상대방으로 하여금 터 주인의 위치 를 쉽게 알릴 수 있는 데 있다. 그러므로 시각적 신호는 특히 근접 거 리 에서 의 신호로 아주 적 합하다 (Hed ig er 1949, Ralls 1971, Walth e r 1969) . 이러한 근본적인 차이 의에도 생활공간은 특정 신호 형태의 사용 에 영향을 미친다. 그래서 노출된 지역에 사는 많은 종들의 경우 시 각적 터 표시가 지배적인 반면 숲이 우거진 생활공간에 사는 종들은 청각 및 후각적 신호를 더 많이 사용한다. 이러한 규칙의 예를 일반 적으로 시각적 정위가 지배적인 영장류에서 찾아 볼 수 있다. 이들 가운데 어떤 종들은 빽빽이 들어찬 숲의 상단부에서 살고 있는데, 이 들은 시각적 신호가 밀도가 높은 식물로 인해 짧은 거리에서만이 효 과적으로 작용한다. 그리고 냄새로 인한 터 표시는 공기 흐름으로 지속적이지 못하다. 그렇기 때문에 이런 생활공간에 사는 영장류는
터 표시를 위해 청각적 표시로 하게 되고 결국 이 들 에게 음성 표현 이 발달한 것은 결코 우연이 아니다 . 이러한 발달은 근연 관계가 아닌 네 개의 영장류 집단(여우원숭이과의 인드리, 꼬리감기원숭이과의 짖는원 숭이, 긴팔원숭이과의 기번, 성성이과의 오랑우탄)에서 서로 독립적으로 일어났기 때문에, 한 행동의 수렴에 따른 실례로 제시될 수 있다. 몇몇 경우에 있어서는 터 표시 행동이 분명히 두 가지 과제를 동 시에 만족시킬 수 있다. 그것은 다른 수컷들을 물리치면서 동종의 암컷들을 유인하는 것이다. 이러한 이중의 기능은 무엇보다 명금류 의 노랫소리에서 찾아볼 수 있다. 비록 암컷을 유인하는 효과가 아 직 분명하게 밝혀지지는 않았지만, 그것에 대해 많은 증거가 있다 . 죽 여름 철새들은 이른 봄에 수컷들이 먼저 터를 장악하고 암컷과 짝을 이룰 때까지 아주 자주 울어댄다. 그러나 짝짓기가 이루어진 이후에는 이 노랫소리가 눈에 띄게 감소한다. 그렇지만 짝짓기 이후 에도 어떤 유인의 기능이 남아 있을 수 있다. 많은 종에서 짝을 이 룬 수컷들이 암컷과 접촉관계를 잃었을 때 특히 더 울어댄다. 여기 서 노래는 짝 형성에 기여하는 것같이 보인다. 역시 무미류 수컷들의 짝짓기 소리는 틀림없이 암컷들을 유인하고 다른 수컷들에게는 간격을 유지하게 한다. 지금까지는 위협행동과 터 표시행동을 서로 분리하여 취급되었다 . 그러나 모든 터 표시는 동시에 위협이 되며 터 표시로 생기는 행동 들은 서로 뒤섞여 구분할 수 없게 되거나 부분적으로 서로가 같아져 그 다음에는 이 터 표시행동은 위협행동의 낮은 강도에 해당될 수 있댜 위협행동과 터 표시행동의 차이는 다음 상황에서 나타나고 있 댜 터는 방향성이 없이 표시되며 또 경쟁자가 없이도 경쟁자를 목 표로 위협하게 된다. 그러나 이런 차이는 매우 미미하여 일부 학자 들은 실질적으로 표시하는 경우, 즉 터 주인이 부재시 터의 효과를 발전시킬(냄새표시, 달랑게의 모래성) 때에만 터 표시의 개념을 사용
 그림 9-11 북미산 붉은등꾀꼬리 수컷이 날개를 옆으로 부풀리며 노래 를 한다 . 이
그림 9-11 북미산 붉은등꾀꼬리 수컷이 날개를 옆으로 부풀리며 노래 를 한다 . 이
하고, 한편 터 표시와 관련해서 일어나는 그밖의 모든 행동들을 위 협행동으로 보고 있다 . 사실 터 표시에 대한 대부분의 연구가 관찰에 근거해서만 이루어 졌다 그렇다고 해서 이에 대한 실험적 연구가 전혀 없는 것도 아니 댜 북미산 붉은등꾀꼬리 A g ela i us p hoen i ceus 는 조밀하지 않게 무리 를 지어 번식하는 새로 수컷들은 각자 작은 터를 방어한다 . 터 방어 를 할 때 옆으로 펼친 날개와 함께 노래를 부른다. 이때 광택이 나 는 붉은 날개 무늬가 더욱 더 돋보이게 된다(그림 9-11). 만일 이 새 의 성대의 신경을 절단하여 벙어리로 만들거나 날개 무늬를 검게 칠 해 버리면 실제로 이 행동이 경쟁자를 멀리하게 한다는 사실을 알 수 있댜 그럼 이 새는 자기 터를 잃게 되고 최소한 경계선에서 급 격 히 잦은 부상을 감당해 야만 한다 (Peek 1972).
5) 종간 텃세 터는 직접적인 경쟁을 피하는 데 기여를 하며 그래서 보통 동종에 대해서 방어가 된다. 그렇지만 한 지역에서 두 종이 언제나 같은 생 태학적 요구를 필요로 하고 마찬가지로 직접 서로가 경쟁을 해야 할 때에는 다른 종의 소속 개체라도 자기 터에 살지 못하게 한다 . 이런 경우를 우리는 종간 텃세라고 부르는데 딱따구리, 벌새, 개개비, 때 까치, 검은딱새, 상모솔새 그리고 까마귀 등에서 잘 알려져 있다. 이에 관련된 종들은 종간 상호이해하는 터 표시가 가능하고 그것으 로 종간 해발인의 기능을 갖는 소리내기나 특정한 움직임에 있어서 외관상 종내의 터 행동과 일치하고 있다. 많은 개미 종들 역시 그들 소유의 진딧물의 체류지를 방어할 때 동종에 대해서 뿐만 아니라 타 종에 대해서도 텃세를 한다 . .1 ,6 개체 간격 하이데거 Hed ig er (l 941) 는 동종 개체간의 공간 관계에 의해 접촉 형태와 간격형태로 분류한 바 있다. 접촉동물들은 육체적인 접촉을 갖는 반면 간격동물들은 일정 거리를 유지하게 되며 이때 일정 거리 죽 개체 간격으로 표현되는 거리안으로 들어오게 되면 개체들은 공 격하거나도망간다. 이 두 형태는 여러 근연종 집단에서 눈에 뛸 정도로 무분별하게 나타나고 있다. 우선 접촉 형태의 동물들로는 돼지, 하마, 대개의 설치류, 원숭이류, 앵무새류, 동박새류, 거북, 일부 도마뱀류, 도룡 뇽, 뱀장어, 알락곰치, 대개의 메기류가 있다. 그리고 간격동물로는 반추류, 홍학, 갈매기, 찌르레기, 제비(그림 9-12), 대부분의 맹금 류, 강꼬치고기, 송어, 연어 등이 있다. 이 간격동물들은 무리지어 번 식하거나 무리를 지어다니는 동물에서 많이 발견할 수 있다. 그래서
 ’
’
두형태의 구분은 사회적 요구 정도에 의해 나타나는 것으로 보인다. 물론 두 형태가 항상 동물에 따라 구분되는 것만은 아니다. 많은 접촉 종에서 대부분의 시간을 육체적 접촉을 유지하면서 살지만 일 부 종에서는 주로 휴식기간 동안 활동이 가능한 커다란 면적을 가지 려 한다. 또 이 접촉행동은 특정상황(위험, 불리한 기후조건), 특정 개체 (배우자 사이, 어미 -새끼 사이), 특정 연령층(어린 동물들), 혹은 암수 중 한쪽(성적 이형의 금화조는 암컷간에는 몸의 접촉을 유지하지만 수컷간에는 서로 접촉을 갖지 않는다)에 한정지을 수 있다. 실제로 극 단적인 간격동물의 경우에도 교미 및 번식행동과 관련된 상황에서 육체적 접촉을 갖는다. 그래서 간격 형태를 갖는 동물에서도 많은 차이가 생겨난다. 개체 간격의 크기는 전깃줄에 앉아 있는 제비나 찌르레기에서 처럼 종에 따라 비교적 일정한 것으로 나타나 있는 반 면, 일부에서는 개체 혹은 성특이적 차이룰 갖기도 한다. 예를 들어 수컷 되새는 18-25cm 의 거리를 유지하고 암컷은 7-12cm 의 거리를 갖는다. 진짜 접촉행동은 외부 위험에 대한 보호본능으로 발달해 왔다. 사 향소의 밀집된 원형대열은 포식동물로부터의 방어에 기여하며, 황제 펭귄의 대군집 형성이나 박쥐의 포도송이 같은 무리 형성은 체온이
내려가는 것을 막아 준다. 이에 비해 다른 종들은 환경요인과 직접 적인 관계가 발견되지 않았다. 이에 대한 극단적인 예로 오스트레일 리아산 숲비둘기 Ar t amus 는 기온이 50·C 이상일 때도 분명히 자발적 으로 길게 늘어서거나 따닥따닥 몸을 대고 무리를 형성한다. 이 경 우 원래 추위 방어로 기여했던 접촉행동이 2 차적인 짝 형성과 무리 형성 기능으로 옮겨온 것으로 추측된다. 한편 동물들이 이런 개체간격을 갖는 것은 중요한 의미를 갖는다. 이 개체간격은 특정 개인 구역으로써 예를 들면 풀을 뜯는 무리 혹 은 먹이를 먹으러 가는 밀집된 무리에서 먹이의 어떤 일부를 보장하 게 하고 서로간의 방해를 억제시키는 역할을 한다. 이런 관점에서 개체간격은 오히려 〈움직이는 터〉로써 개체발생학적으로 터 소유의 이 전 단계로 볼 수 있다. 개체간격은 한 종이 일단 정주하게 되면 자동적으로 고정된 공간부위로 바뀐다. 그래서 터와 개체간격의 분 리에 대한 의문이 제기되어 왔다. 터는 이렇게 움직이는 터의 개념을 포함하고 있기 때문에 장소 종 속성의 기준으로 통용되는 것은 아니다. 야생닭과 오리류에서 수컷 들은 번식 초기에 암컷 주변의 특정 지역을 방어한다. 결국 이미 언 급한 바 있는 〈시간 터〉는 움직이는 터와 고정된 터의 중간 위치에 해당된다. 여기서 시간터가 절대적으로 공간 개념을 갖는 것이 아니 고 실제로 후각적 터 표시 이용으로 확대된 개념으로 사용되기 때문 에 개체간격 표시에 영향을 미친다. 그렇지만 방어행동이 장소와 관련되어 있는지 혹은 그렇지 않은지 의 차이는 분명히 나타난다. 그래서 간격행동에서 동물들은 자기 자신 의 상해에 대해서만 방어하는 반면 터 행동에서는 주변의 대상물 죽 둥지나 새끼 주변에 대해서도 방어를 한다. 그렇기 때문에 터 방어는 개체간격으로 제한하는 것보다 번식과 훨씬 더 연관되어 있다. 결국 두 행동은 항상 분명히 구분되는 것은 아니다 (Hall1966, Hedig e r 1941 ).
l.7 의식화싸움 위협, 터 그리고 간격행동들은 실제 싸움행동처럼 많은 경우 같은 기능을 만족시키나 이런 것들이 서로 대신하여 그 역할을 할 수는 없다. 즉 터를 정할 때, 서열을 정할 때, 혹은 성적 파트너를 두고 다툴 때 진짜 싸움이 일어날 수 있다 . 이들 기능이 단지 상대방을 밀어내는 데 있는 것이지 상해를 입힌다거나 죽이는 데 있는 것이 아니기 때문에 종내 싸움에서는 다른 행동이 일어나게 된다. 죽 이 런 행동은 위험한 무기를 사용하는 종에서 주로 통용되고 있다. 뱀 은 옆으로 머리를 누르고 많은 도마뱀류들은 정면에서 머리를 밀어 싸움을 한다(그림 9-1 3 ). 많은 경우에 날카로운 이빨 사용을 자제한 다 . 키클리드과의 물고기들은 꼬리로 물을 쳐서 싸움을 하는데 상대 방의 머리를 향해 압력 파장을 보내거나 턱을 물어 이리저리 끌어당 기지만 진짜로 물지는 않는다. 많은 유제류들은 앞다리를 치켜들어 상대를 칠 수가 있다. 역시 무기는 경쟁자와의 싸움에서는 사용하지 않는다 오히려 이들은 이마에 난 부수체를 사용하여 싸움을 하는데
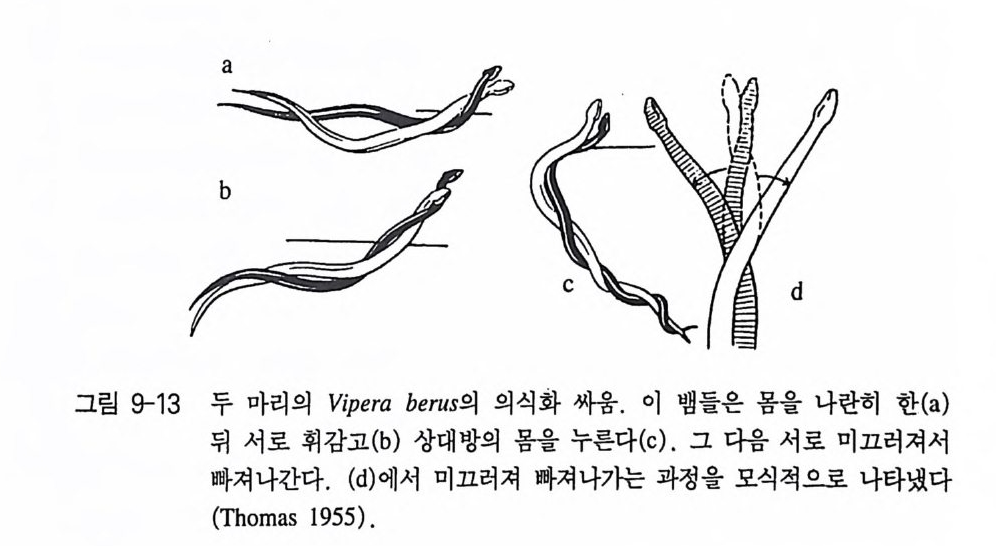 `
`
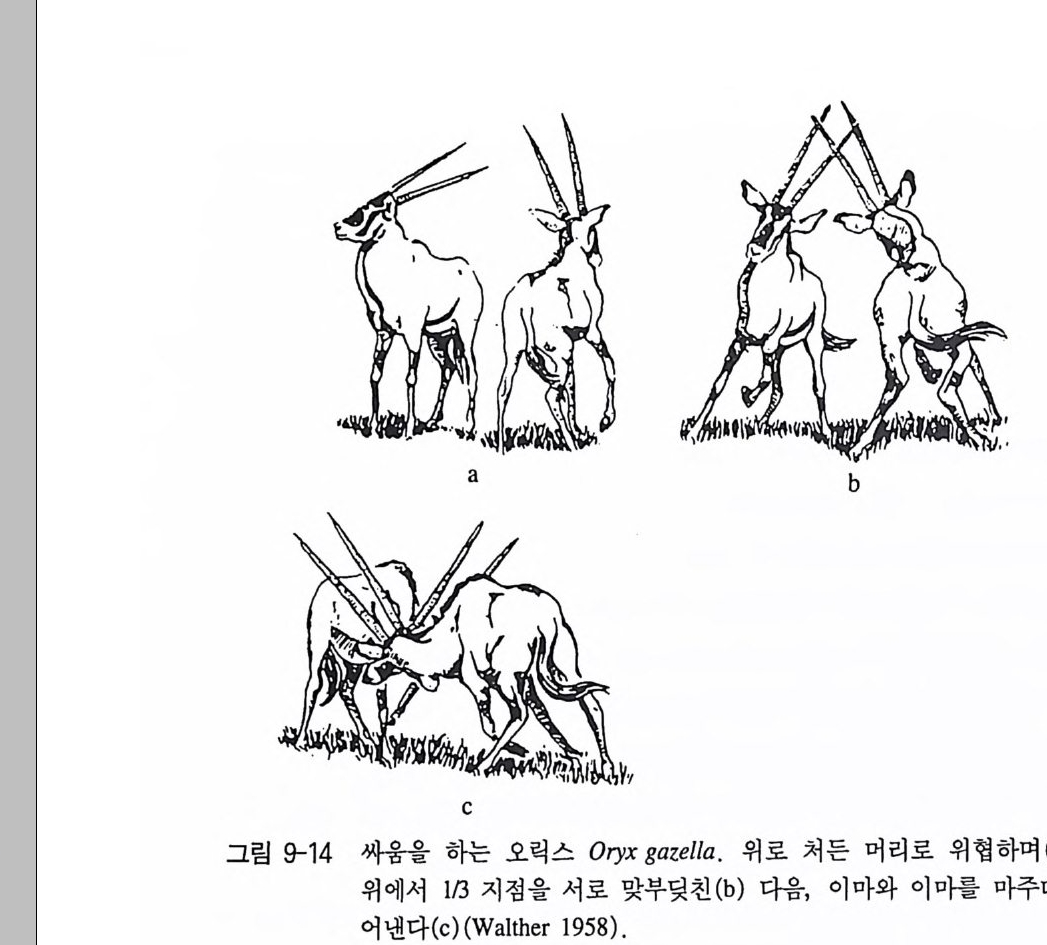 a b
a b
이 부수체는 관련 종들의 싸움 형태에 적응된 형태로 일반적으로 심 각한 상해를 입히지는 않게 되어있다. 어떤 경우에는 싸움행동이 이 두 영역과 비슷하지만 그 결과를 보면 다르다. 즉 뿔울 가진 영양종 들의 싸움 형태를 보면(그림 9-14 ) 뿔과 뿔울 서로 엇갈리게 끼고 밀 거나 이마와 이마를 마주대고 미는데 이것은 그렇게 위험스러운 싸 움 형태는 아니다. 이에 반해 이와 같은 공격성 부수체를 갖지 못한 다른 종에서는 적에 대해 이런 공격으로 몸에 심각한 상해를 줄 수 가있다 . 이런 싸움들은 오히려 고정된 행동 요소들로 이루어져 있으면서 또 이들 행동 요소들이 일정한 규칙에 의해 진행되고 있기 때문에
우리 는 그 싸움 형 태 를 경 기 (競技)싸움 혹은 의 식 화 싸움 ritua liz e d fig h t이라고 부른다. 이렇게 상해를 입히지 않는 싸움 형태로 발달하 게 한 선택압이 강력하게 작용할 수 있었던 것은 승부가 결정날 때 까지 진행되는 많은 경기싸움이 상해를 입히거나 죽이는 종간 싸움 보다 더 많은 에너지와 시간 소모를 요구한다는 사실로부터 분명히 알 수있다 (E i bl-E i bes feld t 1963, Lorenz 1943, 1983). l.8 종내의 상해싸움과 죽임 종내 공격 가운데 진짜 상해를 입히는 싸움은 드물게 발견된다. 많은 동물들은 경쟁자를 심하게 상해를 입힐 수 있는 무기를 사용하 지 않고 오히려 그런 것의 사용을 자제하려고 한다. 그러나 상해를 입힐 수 있는 무기를 가진 동물들은 스스로 상위자 공격을 복종행동 으로 누그러뜨리거나 자기 몸의 표면적을 가능한 한 작게 줄여서 보 호를 한다. 싸움을 하는 숫하마나 숫바다코끼리의 이빨이 실제 피부 아래의 지방층으로 뚫고 들어가기는 어렵다. 그리고 숫사자의 경우 가장 취약한 몸 부위가 두터운 갈기로 되어 있어 상대방의 이빨이나 발톱에 안전하게 보호될 수 있다. 종내의 다툼으로 생길 수 있는 상 해를 막아주는 이러한 적응은 종내의 특이적 공격이 죽이기보다는 먼저 밀어내는 데 있다는 데서 찾아볼 수 있다 . 행동학에서는 오랫동안 동종간에는 죽이는 행위가 사육 상태를 제 외하고는 거의 일어나지 않는 것으로 믿고 있었다. 사육 상태에서 관찰된 싸움이 죽음으로 몰고 가게 된 것은 하위자가 충분히 숨거나 도망할 곳이 없었을 때만 일어나는 것으로 보고되었다. 이런 견해는 오늘날에는 더이상 받아들여지고 있지는 않다. 그 이유는 자연상태 에 사는 동물들도 동료에 의해 죽임을 당하는 사례들이 많이 관찰되 고 있기 때문이다. 또 특정 가정 아래 생물학적 의미를 충족시킬 수
있는 설명이 가능하기 때문에 동종간에도 죽임이 생겨날 수 있는 것 은 일종의 자연선택의 결과로도 볼 수 있다 (Be rtr am 등 1982, Goodall 1979, Lorenz 1983) . 죽음으로 이끄는 이런 싸움은 침팬지, 사자, 하이에나, 사향소, 여 러 설치류, 갈매기, 그리고 큰회색머리아비 Gavia arc ti ca 에서 이미 보고된 바 있다. 이곳에서는 대개의 경우 무리 생활을 하는 종을 취 급하면서도 일반적으로 죽이는 사례는 같은 무리의 소속 개체가 아 닌 다른 무리의 개체들을 대상으로 생겨나고 있다. 종내의 살해억제 기작은 확실히 다른 무리의 구성원들간에 적용되지 않거나 제한적으 로만 적용되어 왔다. 이러한 관점에서 상대 무리에 대한 적대적 싸 움충동은 종간 공격에서의 싸움과 어느 정도 유사하다. 자기 무리 소속 개체에 대해 억제기작이 제한되는 것은 생물학적 인 의미를 갖는다. 보통 다른 무리간의 구성원들은 혈연관계에 있어 서 서로 가깝지 않다. 그래서 이들은 혈연 선택에 있어서 총집단유 전자에 자기 몫을 확산시키는 데 기여할 수가 없다. 이런 제한으로 확실히 동종간의 죽임이 많은 종에서 자연스런 행동요인으로 인정받 게 된 것은 최근에 와서다. 예전의 연구논 이런 억제기작의 효과가 완 전히 보장되는 사회성 집단 내부에서의 행동에 우선 관심을 가졌다. 어린 새끼의 살해(영아살해)는 두 가지 서로 다른 상황에서 나타나 는 것으로 자기 어미에 의해 살해될 수 있고 또 외부 수컷들에 의해 일어날 수 있다. 첫번째 경우는 병들었거나 발육 상태가 나쁜 새끼 • 들에서 일어나지만 건강한 새끼에서도 관찰되었다. 이것은 환경조건 이 양육을 안정되게 보장할 수 없는 경우에 일어난다. 여러 마리의 새끼들을 동시에 길러내는 것이 불가능할 때 나이가 먹은 새끼를 위 해 새로 태어난 개체는 희생을 당한다. 자연 상태에서 영아 살해가 이따금 멧돼지, 붉은여우, 황새 그리고 여러 맹금류에서 관찰된 바 있다 사육 상태에서는 사회적 스트레스와 같은 방해의 결과 실제
자주 생겨난다. 이 경우 심지어는 어미로부터 죽임을 당할 뿐만 아 니라 잡아먹히기도 한다 . 외부 수컷들에 의한 영아 살해는 지금까지 언급한 불리한 환경조 건으로 생겨난 종내의 살해보다 훨씬 다른 뜻을 내포하고 있다 . 무 엇보다 하렘을 형성하는 사자나 인도산 랑구르(영장류)에서 이 영아 살해가 관찰되었다. 이들 동물의 경우 한 수컷이 지금까지의 하렘 수컷울 굴복시켜 그 수컷을 내쫓고 하렘 자리를 차지하게 되면 아직 독립하지 않은 암컷들의 새끼들을 죽인다. 물론 이때 이 새끼들을 잡아먹지는 않는다. 이 영아 살해의 궁극적인 목적은 암컷들이 수유 기간 동안에는 발정이 일어나지 못하기 때문에 새끼를 죽여버려 좀 더 빨리 새로운 짝짓기 준비에 도달케 하는 데 있다. 이 새로운 하 렘 수컷은 자기와 혈연이 아닌 전임자의 새끼들을 희생하여 가능한 한 빨리 자기 자식을 만들 해산 시점을 앞당기므로 해서 분명히 그 다음 세대에 자신의 유전자를 최대로 전하게 된다(Rij ksen 1981 ). l.9 동인에 대한 제문제 종내 공격의 광범위한 분포로 인하여 생물학적인 중요한 과제는 동물의 공격적 행동충동이 어느 식으로 조절되느냐가 문제가 되고 있다. 여기서 다양하게 문제를 제기할 수 있다. 공격성에 대해 유전 적 단서가 있는가? 공격성은 환경요인에 얼마나 영향을 받을까? 이 공격성에 생물학적 보충 기작이 작용할까? 자기 공격충동이란 존재 할까? 인간의 행동과 관련해서 매우 중요한 이런 질문들은 오늘날까지 궁극적으로 완전히 설명되지 못하고 있으며 부분적으로는 여전히 논 쟁이 되고 있다. 그럼에도 불구하고 일부 설명이 가능한 많은 연구 의 결과도 있다.
1) 종특이적 공격 공격성이 유전적이라는 사실은 어떤 종의 종내 공격성의 범위가 확실히 어느 정도 한정되어 지켜지고 있으며 그러므로 종마다 매우 다양할 수 있다는데 근거한다. 때때로 근연종간에도 분명한 차이가 나타난다 무리를 이루어 번식하는 바다 조류 가운데 가파른 벼랑에 둥지를 트는 북부부비새 Marus bassanus 는 실제로 땅바닥에 둥지를 트는 남부부비새M. cap ens i s 보다 더 공격적이다. 이런 차이는 둥지를 트는 방식과 관계가 있다. 즉 북부부비새는 적당한 바위의 생태지위 를 방어하는 데 높은 공격충동을 필요로 하고 있다. 이에 비해 둥지 트는 면적을 넉넉히 이용하고 있는 남부부바새는 이러한 공격충동이 요구되지 않는다. 남부부비새의 경우 무리의 중앙에서 둥지를 튼 새 들은 적당한 이륙 장소에 도달하기까지 빈번히 다른 쌍들의 터를 지 나쳐야 하는 불리함도 있을 수 있다. 만일 남부부비새가 북부부비새 와 똑같이 공격적이었다면 이웃한 쌍들간에 경계 싸움이 끊임없이 있었을 것이다. 결국 비슷한 많은 사례들을 살펴보면 모든 종들이 계통발생 과정에서 생태학적 조건에 가장 잘 적응한 종특이적 종내 공격 범위로 발달하게 되었음울 알 수 있다 . 2) 인위적인 공격 선택 생태적 적응의 결과로 종마다 공격성의 평균치가 유전자에 자리잡 고 있을거라고 짐작할 수 있다. 이것은 관련 개체의 낮은 번식 성공 을 가져오는 높은 공격충동이나 낮은 공격충동은 자연선택에 의해 계속 일정하게 유지될 수 있다. 이런식으로 변화된 환경조건들은 새 로운 적응으로 이르게 할 수 있다. 만일 자연선택이 실제로 결과가 빨리 나타나는 인위적 도태에 의 해 생겨났다면 이러한 상상은 가능하다. 그래서 여러 근연집단에서 나온 몇몇 종들의 경우에 가축화 과정에서의 공격성이 가장 공격적
인 개체의 선택을 통해 아주 두드러지게 상승시킬 수 있다(鬪魚 투 우, 싸움닭). 집쥐의 경우 공격충동이 다양한 여러 품종들을 선택하 여 사육된 바 있다. 여러 계열의 개체들을 서로 교잡한 결과 E 의 공격성은 양친의 공격성의 중간위치에 놓이게 되는 유전적 기초를 제시할 수 있다. 3) 환경에 의한 영향 공격성이 유전적 근거에 의해 결정된다고 해서 그것이 단지 생득 적이고 환경 영향이 종내 공격성의 형성에 아무런 역할을 하지 않는 다고는 말할 수 없다. 오히려 많은 경우에서 사회적 경험이 공격 형 성에 매우 심각한 영향을 미친다는 사실이 증명되고 있다. 죽 생후 초기에 이러한 영향은 아주 지속적인 인상으로 계속 남아있게 된다. 그래서 수컷 생쥐를 어렸을 때부터 규칙적으로 손으로 쓰다듬어 주 면 커서도 온순한 행동을 보인다. 반대로 생후 초기부터 먹이룰 먹 는데 싸움을 하게 하면 커서도 그 쥐는 매우 공격적이 되며, 또 배 가 전혀 고프지 않는 데도 먹이 앞에서 싸움을 하는 것을 볼 수 있 댜 생쥐의 경우 보통 싸움에서 일단 승리를 거두면 계속 공격충동 이 늘어나는 반면 지게 되면 차츰 그 싸움충동은 사그라들게 된다. 이때 이런 성향이 명백하게 드러나면 날수록 생애의 더 이른 시기에 관련 경험들이 모아져야 된다. 생후 2_3 개월을 친어미 없이 매우 공격적인 계모한테서 자란 어린 히말라야원숭이는 커서 공격충동이 엄청난 과잉상태를 나타내는데, 그것은 온순한 친어미에 의해 양육 된 개체보다 2 배 이상의 공격적인 행동을 보여준다. 비슷한 결과가 다른 종에서도 많이 관찰되었다. 전기적 쇼크를 가해도 공격성을 높 일 수 있지만, 여기서도 그 결과는 가한 쇼크 정도에 따라 달라진다. 동물의 공격성에 관해 현재까지 연구된 자료를 토대로 종합해 보 면, 종특이적 공격 크기에 대한 유전자들은 명백히 특정 형질을 발
현시킬 수 있고 개체 특성은 종내의 일정 범위 내에서 경험에 의해 결정된다. 그러므로 이런 범위 내에서 유전과 환경의 밀접한 상호관 계가 작용한다. 이에 대해 지금까지 행동학에서는 거의 등한시해 온 것이 사실이다. 오히려 공격성온 생득적이거나 혹은 환경에 의해서 달라지는 것으로만 의미를 부여해 왔다. 4) 공격과 호르몬 동물의 공격성을 조절하는 관점에서 보았을 때 종내 공격은 다른 행동기능과 구별할 수 있다. 죽 그것은 호르몬 조절과 중추신경 조 절에 대한 실험적 근거에 의해 구별된다. 연주기에 따라 종내 공격 의 높이가 달라지기 때문에 호르몬 조절에 의해 유리하게 증명될 수 있다. 이런 경우 호르몬은 높은 공격성(터 확보 텃세 싸움)이 요구되 는 시점으로 인해 번식 초기에 최고에 이른다. 터를 소유하고 있는 큰가시고기 수컷은 봄에 배가 붉은 모형에 강한 공격성을 유발하지 만 늦가을에는 그 붉은 모형에 전혀 관심을 갖지 않는다. 이러한 두 행동체계의 시간적 평행관계로 성호르몬에 대한 조절이 생물학적으 로 가장 의미있는 것으로 나타난다. 거세실험과 호르몬 처리는 이러 한 관계를 실제로 증명해 주고 있다. 물론 호르몬 조절에 의해 나타난 공격행동은 아주 다양하다. 안드 로겐은 보통 공격을 촉진시키는 데 작용하는 반면, 자성 호르몬의 역할은 아직도 일정하게 나타나지 않고 있다. 그밖에 부신 호르몬과 뇌하수체의 성선자극 호르몬은 동물들의 공격성에 다양한 영향을 미 칠 수 있다. 그러나 여기에 대해서는 아직 정확하게 밝혀진 게 없다. 안드로겐 죽 테스토스테론은 무엇보다 번식과 직접적인 관계를 가 지면서 공격(예, 텃세 싸움, 암컷이나 둥지 재료를 빼앗기 위한 싸움)에 영향을 미치는 많은 실험결과가 보고되었다. 그리고 수컷에 있는 호 르몬이 암컷에게도 작용하는 사례도 있다. 흰머리쑥새 Zono t r i ch i a
leucop hry s 는 암컷 이 터 방어 에 능동적 으로 참여 하는데 번식 기 에 암 컷에서도 높은 양의 테스토스테론이 발견되었다. 그밖에도 효과가 암컷에서도 나타날 수 있었던 뇌하수체 호르몬들은 공격적 성질을 조절한다. 5) 공격욕구 공격적 동인 조절에 대한 논의는 자기공격 충동의 존재와 본질에 초점이 맞추어지고 있다. 근본적으로 여기서 두 가지가 문제가 제기 될 수 있다. 어떤 한 동물에서 시간적으로 변화하는 공격행동 준비 는 이전에 입력된 환경 정보들(예, 경쟁자를 본 정보, 터에서 나온 자 극)에 따라 변하든지, 자발적 죽 환경과 무관한 요인들이 이 행동 준비에 관여한다. 이 행동 준비가 자극에 의한 반응으로 볼 것인지 아니면 행동 그 자체인지는 공격성이 유전적인 근거에 의해서 나타 난다라는 사실과는 아무런 관계가 없다. 그 이유는 이런 동인이 자 발적 충격발사의 형태로만 표현되는 것이 아니고 외부 자극에 의한 어떤 특정한 여러 반응 형태로 나타날 수 있기 때문이다. 공격이 자발적으로 일어난다라는 사실은 지금까지 잘 연구된 바는 없다 실제 진공一공격 vaccum-a gg ress i on 은 아직 확인된 바 없지 만, 진공행동 vaccum ac ti v ity은 이미 알려져 있다. 그러나 일종의 싸 움준비의 정체성(停淸性)은 많은 자료가 있다. 키클리드과의 많은 물 고기류와 꿩류에서 수컷들은 암컷을 공격하고 그들을 짝지어 놓으면 심지어 죽이기까지 한다. 이것은 수컷들로부터 터 이웃과의 경쟁자 싸움을 하고자 하는 욕구를 빼앗아 버렸기 때문으로 풀이된다. 만일 이 수컷들을 다른 수컷들과 싸움을 하게 해주면 암컷들을 전혀 해치 지 않고 성공적인 짝짓기가 일어난다. 이때 다른 수컷들을 유리 칸 막이나 철망을 통해 이 수컷들에게 보여주기만 해도 효과는 대단히 크다.
이렇게 경쟁자 없는 수컷의 싸움 준비의 상승은 공격적 동인이 쌓 일 수 있다는 데서 찾아볼 수 있지만, 이런 정체가 오로지 체내 조 건에 의한 것으로만 볼 수는 없다. 키클리드과 물고기에서 최소한 암수가 서로 외형적으로 비슷하게 생겼기 때문에 암컷에 의해서도 수컷의 싸움 준비를 상승시키는 자극이 나올 가능성이 대두된다 . 이 런 방면의 연구가 다른 키클리드과의 물고기에서도 확대된 바 있다. 연구결과 성체의 동료가 지속적으로 눈에 뛸 때나 그와 비슷한 모형 이 있을 때 이런 공격성이 상승되고 있다는 사실이 밝혀지게 되었 댜 이에 비해 꿩류에서는 암수의 극단적인 색깔 차이로 인해 외부 에서 기인된 욕구의 축적이라는 가능성은 조금 적다. 동물들을 완전히 개별적으로 기르면 마찬가지로 공격적 행동준비 가 상승된다. 참집 게 Pa g urus samuel i s 의 경 우 혼자 있게 하면 공격 행 동 준비가 점차로 상승되는데 처음에는 공격행동이 높은 해발역치를 갖다가 나중에는 낮은 해발역치로 공격행동의 빈도가 상승한다. 병 아리나 생쥐도 새끼때부터 격리해서 홀로 키우면 공격적인 개체가 된댜 쌍을 격리시키는 것에 비해 한 마리씩 기르는 것이 방법면에서 장 점이 될 수도 있다. 그 이유는 동료로부터 나오는 자극을 통하여 개 별적 영향들을 확실히 제거할 수 있기 때문이다. 그러나 이런 장점 에 대해서 결정적인 단점도 있을 수 있다. 말하자면 한 마리씩 가둔 동물들은 오히려 모든 환경자극에 대해 상승된 민감도가 높아져 여 러 종류의 발생학적 장해를 가져올 수 있기 때문이다. 이런 이유로 인해 공격적 해발인에 대한 강한 반응들이 실제 공격적 동인의 상승 에서 종특이적으로 기인된 것인지 혹은 이 반응들이 단순히 그 동물 의 일반적인 민감성의 표현인지 아직 모른다. 결국 공격적 행동준비 의 정체성에 대해 지금까지 나온 자료들도 자발적 공격 요소들이 있 느냐의 질문에는 아직 궁극적인 대답을 할 수 없다는 것이다 .
끝으로 자율적 싸움욕구와 관련해서 똑같이 적용되는 것이 있다. 자율적 싸움욕구의 존재는 싸움할 수 있게 보상되는 작동조건화 실 험을 통해 입증할 수 있다 . 많은 물고기나 새들을 대상으로 이 실험 이 가능하다. 동물이 어떤 특정 장소에 오면 그곳에서 경쟁자와 싸 움을 할 수 있게 하거나 위협적인 모형 혹은 거울에 비친 자기 자신 의 모습을 보여주어 그장소에 오도록 훈련시킬 수 있다. 역시 햄스 터나 생쥐를 가지고도 T 자형 미로의 해당 출구에 경쟁자와 싸움을 하도록 학습이 가능하다 . 경쟁싸움이 있은 직후 격리된 쥐는 그 경 쟁자에게 다시 접근하기 위해 전류가 호르는 철망도 건너간다. 이 경우 그곳에 전류가 흐르는 전압의 크기에 따라 각 싸움충동은 양적 으로 측정이 가능하다. 이런 사실은 자율적 싸움욕구가 있다는 중요한 단서가 되고 있다 . 물론 이것으로 싸움욕구의 자발적 성질에 대해 다 설명할 수는 없다. 그 이유는 완전 격리했을 경우 그 동물은 아마도 민감한 환경자극으 로도 공격적으로 될 수 있다는 사실을 완전 배제할 수가 없기 때문이 다 결국 공격적 동인에 대한 외부적 관여가 있을 것으로 추측된다. 2 성적 행동 2.l 생물학적 의의 성적 행동은 생물의 기본 현상 가운데 하나다. 몇몇 하동한 생물 울 제외하고는 성분화와 생식에 대해 거의 모든 생물체에서 잘 알려 져 있댜 한 세대 혹은 여러 세대를 단지 무성생식 죽 이분법이나 출아법으로 번식할 수 있는 종이 있지만 이런 종에서도 규칙적 내지 는 불규칙적 간격으로 세대교번이 일어난다.
성적 행동의 생물학적 의미는 모든 생물체에서 일치하고 있다. 생 식세포의 형성 과정에서 일어나는 감수분열과 정자와 난자가 만나 수정된 세포는 염색체 위에 자리잡고 있는 부계와 모계의 유전자~의 새로운 분열로 이어진다. 그러므로 성적 행동은 그 생물체 변화에 대 한 전제 조건이 되며, 이것은 다시 환경에 대한 적응의 결과이자 생 물체 자체에게는 고도로 발달을 가능하게 한 기본적 원동력이 된다. 이러한 의미에서 가장 중요한 과정은 난자와 정자의 핵융합인 수 정인 것이다. 이것은 교미가 전제된다. 즉 수정은 정자가 난자의 세 포막을 뚫고 들어가는 것이고 교미는 수컷의 생식기와 암컷의 생식 기가 합치는 것을 의미한다. 그러나 이런 개념은 명확하게 항상 구 별되는 것은 아니다. 교미행위에 이르게하는 제반 행동에는 구애 cou rt s hip와 짝짓기 ma ti n g가 있다. 짝짓기는 짝형성 pair fo rma ti on 과 짝유대p a i rbond i n g로 구분될 수 있는데, 짝형성은 짝유대를 위한 암 수의 만남이라고 한다면 짝유대는 암수의 지속적인 짝유지를 내포하 고 있댜 그러나 이러한 분류는 항상 가능한 것은 아니다 . 그 이유 는 수많은 종들의 구애행동이 실제로 교미에까지 이르는 것이 아니 고 짝형성 혹은 단순히 암수가 함께 머무는 정도로만 끝날 수 있기 때문이다. 동물들은 성공적안 교미를 위해 형태적, 생리적 그리고 행동적 일 련의 조건이 전제된다 . 이 가운데 다음과 같은 행동영역의 요구가 전제될 수 있다. ® 배우자는 종과 성을 서로 인식해야 한댜 ® 배우자는 서로 만나 자신들의 생식기룰 합쳐야 한다 . ® 배우자는 동시에 교미 준비 죽 정확한 동조가 일어나야 된다. ® 배우자는 종내의 공격성을 극복해야 한다 (Bas t ock 1969, Hutc h iso n 1978).
2,2 성적 행동의 기능 성적 행동은 이미 제시한 전제 조건을 충족해야 되는 과제를 갖고 있댜 여기서 이런 각각의 과제에는 서로 다양한 행동 양식들이 나 타날 수 있다. 그래서 공격행동과 양육행동을 제외하고는 이 성적 영역은 특별히 다양하고 고도로 발달된 행동으로 특징지어진다. 이 것은 다음의 각 영역에서의 규칙성을 통해 알 수 있다 . 1) 암수의 만남 지속적인 쌍이나 집단으로 살지 않는 종의 경우에 있어서 짝짓기 를 위하여 암컷과 수컷은 간격을 좁혀 서로 접근해야만 한다. 바로 서로의 접근을 위하여 배우자의 유인은 종의 대부분에서 수컷이 주 도권을 갖는다. 물론 예외가 곤충에서 주로 나타나기도 한다. 종종 이 유인 기작은 터 표시 기작 즉 동성인 경쟁자를 물리치는 기작과 일치하고 있다. 그래서 여기에서는 암수의 만남에서 주로 나타나는 사항만을 제시하고자 한다 . 여기에서도 터 표시처럼 하나의 자극 양 상 혹은 여러 개의 자극 양상이 동시에 또는 연속적으로 결합하여 효과를 나타내는 자극 양상들이 있을 수 있다. 시각적 유인 수많은 동물종들은 색깔 표시로 성적 파트너를 유인하고 있다. 그 색깔 표시를 움직이게 한다든가, 그것을 크게 한다든가 혹은 펼쳤다 접었다 하여 눈에 띄게 한다 . 열대 혹은 아열대 바닷가에 사는 농게 의 수컷들은 두 집게발 가운데 큰 집게발 하나를 이런 목적으로 리 드미컬하게 이리저리로 움직인다. 이것은 대개의 종에서 눈에 띄는 색깔로 되어 있다(그림 9-15). 이 집게발의 움직임이 두드러지게 되 는 것은 몸의 움직임이 동반되어 더욱 강화될 수 있다. 열대지방의
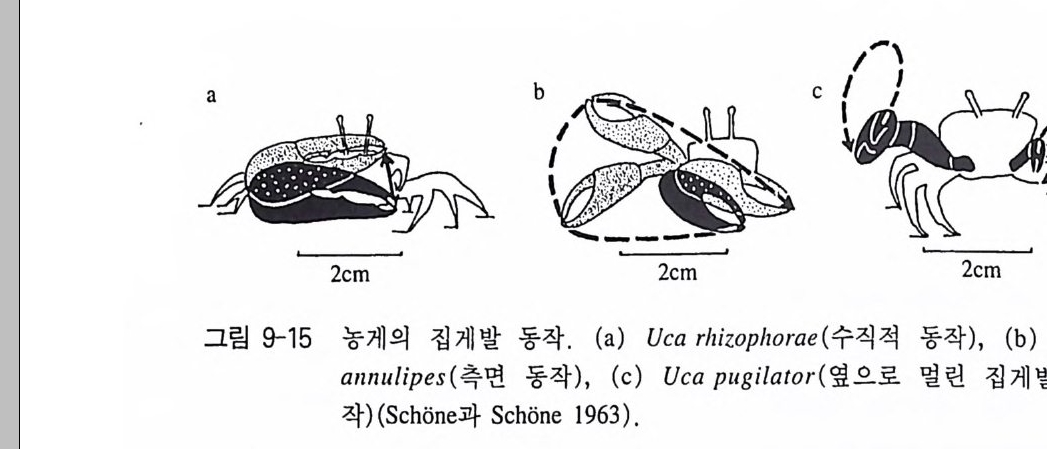 a b
a b
군함새는 수컷이 번식 초기에 붉은 목주머니를 부풀려 보여주거나 또 수많은 조류에서 색깔을 띤 깃털 부위를 발달시켜 눈에 띄게 한 다. 하나의 대표적인 예가 공작 수컷의 부채처럼 펼친 꼬리깃을 들 수 있다 시각적 유인 신호는 역시 지속적인 자극원이 될 수도 있고 반딧불이의 불빛처럼 여러번에 걸쳐 반복되기도 한다. 반딧불이의 경우 날지 못하는 암컷들은 파트너를 유인하는 신호로 사용한다 . 이 들은 밤에 특정 자리, 종종 높은 위치의 발광 장소를 찾아 복부를 지지하고는 복부 끝에 있는 발광기관을 위로 치켜올린다. 많은 종에 서 복부의 움직임이 신호효과를 강화시킬 수 있다. 이에비해 날 수 있는 수컷들은 이 빛신호로 배우자가 될 암컷울 향해 날아갈 수 있 댜 만일 수컷이 암컷 위로 날아오게 되면, 날개를 접고 아주 정확 히 암컷 옆에 착륙하게 된다. 이 빛신호에 대한 유인 및 조절효과는 거울이나 인공적인 빛을 가지고 증명될 수 있다 (Llo y d 1966). 음성학적 유인 음성학적 신호는 특히 조류, 양서류 그리고 곤충에서 성적 파트너 를 찾는 것과 관련해서 사용하고 있다. 새들의 노랫소리의 유인 기 능과 무미류의 짝짓기 소리가 이미 증명되었다. 많은 곤충(예, 매
미, 메뚜기, 귀뚜라미)에서도 암컷들이 수컷의 소리에 의해 유인된다. 귀뚜라미의 수컷은 두 개의 서로 다른 노래 형태 외에도 단지 암컷 만을 유인하는 목적으로 쓰이는 구애노래를 갖고 있다 (Tembrock 1982) . 후각적 유인 특정 냄새물질로 성적 파트너의 유인은 몇몇 동물군에서 광범위하 게 나타나 있다. 그러나 지금까지 단지 일부만 정확히 연구되어 있 울 뿐이다. 이러한 연구는 주로 포유동물과 곤충에 집중되어 있다. 햄스터 수컷으로부터 암컷을 유인하는 냄새물질이 옆구리에 위치한 분비선에서 만들어지고 있다는 사실이 알려졌으며, 암집쥐의 오줌 속에서는 쥐의 성적인 상태를 알려주는 정보가 들어 있다 (Hurs t 1988). 특히 멀리까지 퍼지는 냄새신호가 많은 밤나방의 암컷에서 만들어지고 있다. 이들은 복부의 끝에 있는 분비샘에서 수컷의 더듬 이에 있는 감각세포로 감지할 수 있는 냄새물질을 분비한다. 이 세 포들의 민감성은 놀라울만큼 예민하다. 누에나방의 경우 수컷의 더 듬이에 유인물질인 몇 개의 봄비콜 분자만 닿아도 충분히 홍분된다. 이런 식으로 수컷은 수 킬로미터 떨어진 거리에서 유인될 수 있다. 산누에나방의 일종인 Acti as selene 의 수컷은 46 킬로미터 정도 떨어진 지점에서도 방금 탈피한 암컷이 있는 장소를 찾아낼 수 있다. 집단구애를동한유인 많은 동물들은 여러 마리의 수컷들이 함께 암컷울 유인할 수 있는 행동을 공동으로 펼치기도 한다. 이것은 수 많은 종에서 많은 수컷 들이 공동으로 소리를 내는 메뚜기류와 매미류의 노래에서 찾아볼 수 있다 이것은 열대지방에 사는 반딧불이의 동시에 번쩍거리는 신 호나 북미에 사는 색깔이 밀집되어 보이는 잉어류의 수컷들의 집단
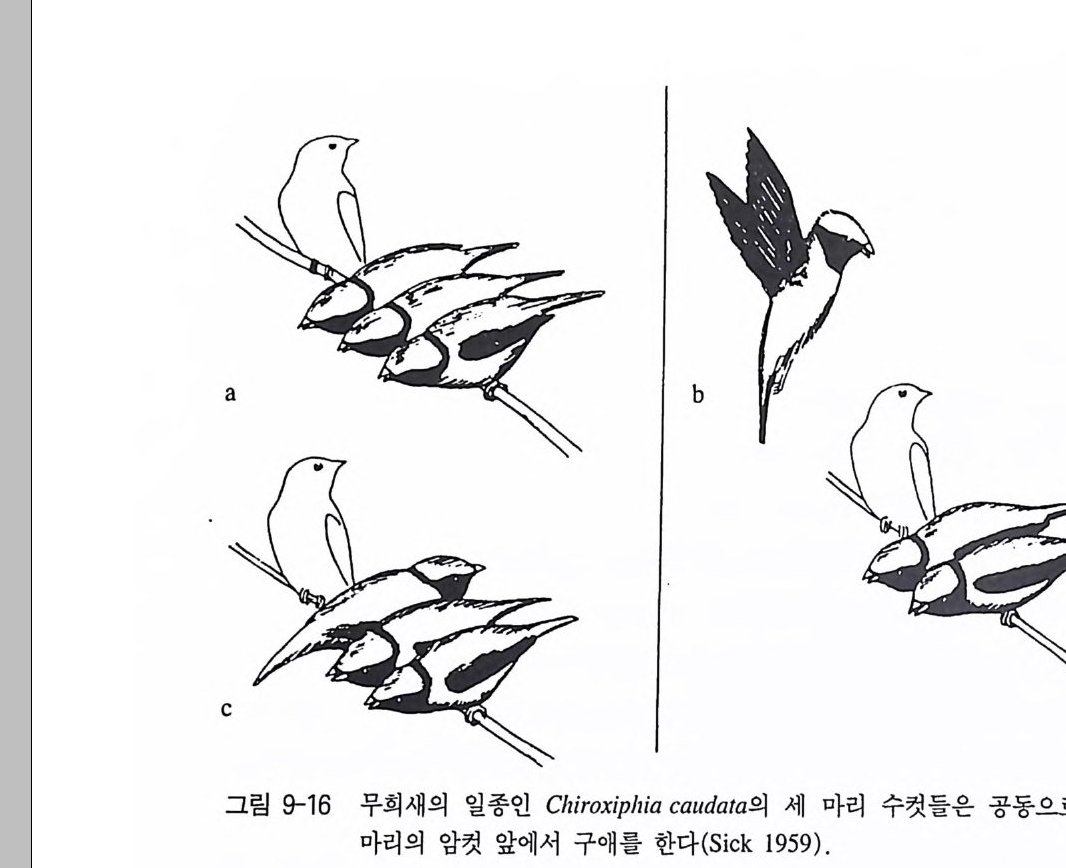 a b
a b
에서도 같다. 몇몇 벌새류의 수컷들은 합창이라고 할 수 있는 공동 의 노래를 위해 서로 만난다. 이때 최고 60 마리가 하나의 노래 장소 에서 관찰된 적이 있었으며 흰극락조의 경우 규칙적으로 두 마리의 수컷들이 서로 대칭자세로 〈이중구애〉를 한다. 이때 이들은 각기 한 나뭇가지에서 교대해 가면서 머리를 아래로 하고 매달리는 행동을 취한다. 역시 또 다른 극락조 Parad i sae i dae 에서 여러 마리의 수컷들 이 공동의 구애를 위해 서로가 모인다. 이런 현상온 남미에 사는 많 은 조류에서 예를 들면 무희새 P ipri dae( 그림 9-16) 나 바위자고(꿩과) 에서 알려져있다. 이들 종들은 모두가 공통점이 있다. 성적 이형이 매우 분화되어 있으며 수컷들은 매우 화려한 색깔과 매혹적인 깃털 울 갖고 있다. 그래서 아마도 집단 구애의 이런 형태는 자극의 통합
원리에 의해 동종의 암컷에게 특히 강렬한 유인효과를 발휘하며 진 화과정에서 분명한 신호효과를 지닌 선택압을 통해 발전해 왔다. 2) 종내 공격성의 극복 암수의 교미에 필수적인 상호접촉온 성적인 욕구 이의에도 공격성 을 활성화시킨다. 이것은 단독으로 생활하며 각자의 텃세를 방어하 거나 개체 간격을 유지하는 종에서 일반적이다 . 이런 성향을 누그러 트리는 것은 구애행동의 중요한 과제가 된다. 실제 많은 동물에서 구애과정이나 구애 후에 아주 두드러진 공격 성이 나타난다. 구애하는 큰가시고기의 지그재그춤은 암컷의 공격을 저지하게 하며, 구애하는 금화조의 경우에는 수컷이 부리를 암컷을 향해 반복해서 찌르는 듯한 전후운동을 하여 암컷의 공격성을 누그 러뜨린다 이러한 의미의 행동과 관련해서 몇몇 거미류는 종종 실제 몸집이 작은 수컷들이 암컷에게 먹이동물로 보여서 죽임을 당하는 위험에 있기도 한다. 이에 대한 적응에 있어서 많은 종들의 수컷들 은 고도로 발달된 신호동작과 동작의 연쇄성을 발달시켜 왔다. 이 연쇄성온 동종의 수컷으로써 암컷에게 식별이 가능할 뿐만 아니라 먹이동물과는 차이가 나도록 발전하여 왔다. 그럼에도 불구하고 많 은 종에서 수컷은 교미 후에 혹은 교미하는 동안 암컷으로부터 잡혀 먹히기도 한다 . 그러나 외견상 공격성이 결여된 종에서도 구애과정에서나 때때로 구애동작의 상반 감정의 공존 형태에서도 성적 그리고 투기적인 요 소들의 상호작용으로부터 여러 번에 걸처 개체발생학적 기원을 찾아 볼 수 있다. 이때 규칙적으로 수컷들에서 성적 그리고 공격적 요소 들이 결합하고 이에 비해 암컷들에서는 성적 그리고 도피요소의 행 동결합이 생겨나지만 때때로 거미류나 조류에서처럼 암컷이 번식기 초에 상위의 서열을 차지할 때도 있다. 물론 반대의 서열도 존재한
다. 그래서 동물들의 매우 많은 연속적 구애동작들은 한편으로는 성 적 동인과 다른 한편으로는 공격과 도피의 동인 사이의 타협행동으 로 볼 수 있다. 가장 잘 알려진 예가 이미 언급된 큰가시고기의 지 그재그 춤이다. 여기서 암컷에 향하는 지그는 공격적 요소이고 이에 반해 암컷에서 멀어져 둥지로 향하는 재그는 성적인 요소가 내포되어 있다 물론 많은 경우 이러한 의미 부여는 도식화되는 심각한 위험에 빠 질 수도 있다. 위협신호와 같이 폭넓게 쓰이는 의미는 구애 혹은 짝 짓기 행동에서 이미 지적된 바 있다(위엄행동). 암수가 단지 교미의 목적뿐만 아니라 지속적으로 함께하는 모든 종에서는 역시 공격성을 억제하는 메커니즘이 오랜 기간 동안 효과 적 이어야만 한다. 이 경우에 교미 전단계의 행동 성분뿐 아니라 짝 유대를 강화하는 중요한 수단으로 교미전 행동과 같은 행동양식들이 표현된다. 그러므로 짝짓기, 구애 그리고 짝형성 행동 사이의 구별 은 이런 점에서 특히 어렵다. 짝짓기 행동에서 공격 억제 메커니즘에 대한 두 가지 방법이 자주 나타난다. 고의적으로 무기를 은익하는 것과 공격을 가장한 행동성 향을 활성화하는 것이 있다 . 첫번째에서 가장 잘 알려진 예는 황새 가 머리를 등뒤로 돌려 윗부리와 아랫부리를 서로 맞부딪치는 것과 붉은부리갈매기가 머리를 돌리는 것이 이에 속한다. 붉은부리갈매기 의 머리돌리기는 상대가 흰 뒤통수를 마주보게 하는데 이렇게 되면 위협행동에 중요한 역할을 하는 검은 얼굴을 감추게 되는 것이다. 까마귀의 일종인 Corvus cora.x 역시 비슷한 목돌리기를 하는데 이 목돌리기를 통해 상대로부터 강한 부리를 다른 쪽으로 향하는 것이 다(그림 9-17). 이런 많은 행동들은 복종행동의 형태와 닮은 점이 있 고 실제 싸움 전에 오랫동안 이용되고 있기 때문에 가짜복종행동이 라고한다.
 그림 9--17 까마귀의 일종인 Corvuscorax 의 복종적인 목돌리기 (Gw i nner 1964).
그림 9--17 까마귀의 일종인 Corvuscorax 의 복종적인 목돌리기 (Gw i nner 1964).
공격성을 억제시키는 다른 행동기능 범주에 대해 무엇보다 계통발 생학적으로 둥지짓기와 새끼돌보기가 제시되고 있다. 이 두 범주에 서 나온 행동요소들은 수많은 종의 암수간 행동에서 발견할 수 있 다. 특히 조류에서 자주 동장한다(예, 단풍새류 Es tri l di dae 의 수컷) . 뿔 논병아리 Podic e ps cr i s t a t us 의 암수는 구애의 특정 시점에 둥지 재료 가 될만한 것들을 부리에 문다. 그리고 그것을 짝에게 건네주거나 그렇지 않으면 부리를 움직이거나 머리를 움직인다거나 혹은 몸을 움직여 눈에 띄게 한다. 새끼돌보기 행동을 살펴보면 무엇보다 먹이전달 행동이 구애에서 넘어왔음을 알 수 있다. 즉 먹이를 주는 행동 요소(먹이 제공과 건네 줌)뿐만 아니라 먹이를 받는 상대의 행동 요소(구걸)들이 이 행동의 영역에 속한다. 일반적으로 수컷의 바다제비나 갈매기는 마치 새끼 가 어미로부터 먹이를 받아먹듯 자기 암컷을 먹인다. 바다제비는 사 냥해 온 물고기를 부리로 직접 건네주는 반면 갈매기는 반쯤 소화된 먹이를 게워내어 암컷에게 먹게 하는데 이때 암컷은 약간 벌린 수컷 의 부리에서 그것을 꺼내 먹는다. 역시 여러 단풍새류와 무엇보다 앵무새에서도 이런 구애급이가 일반화 되어있다(그림 9-18). 여기서
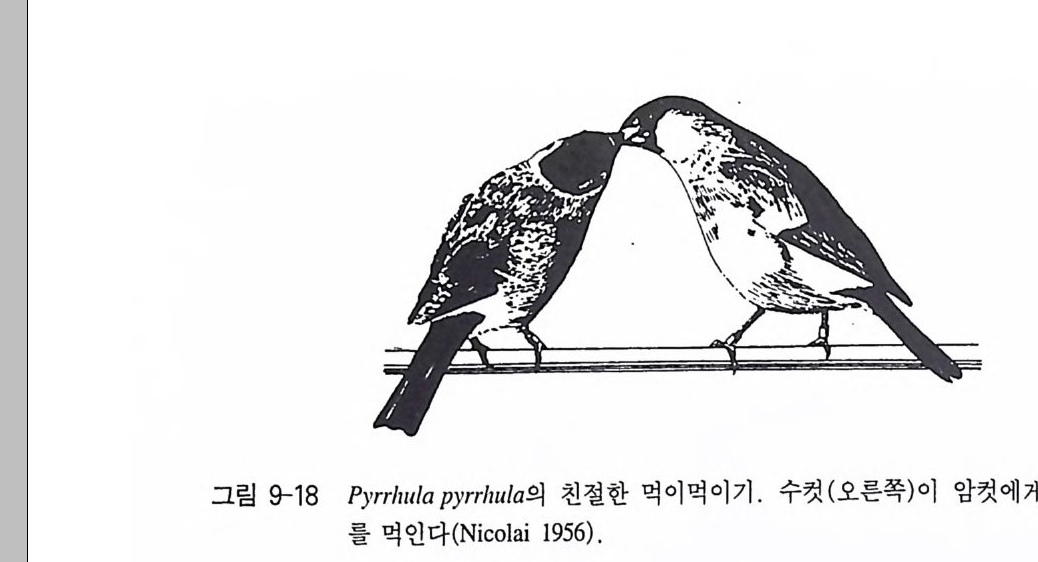 그림 9-18 P y rrhula p y rr/111/a 의 친절한 먹이먹이기 . 수컷(오른쪽)이 암컷에게 먹이
그림 9-18 P y rrhula p y rr/111/a 의 친절한 먹이먹이기 . 수컷(오른쪽)이 암컷에게 먹이
많은 종의 수컷이 암컷을 먹이는 반면 일부 종에서는 상호 먹이교환 이 이루어지기도 한다. 원래 수컷이 암컷에게 먹이를 먹이는 행동은 영양학적인 의미를 갖고 있다 이 행동은 알을 만드는 과정에서 암컷에게 먹이룰 부가 적으로 공급해 주는 역할을 한다. 그러므로 이 행동은 영양학적 임 계기간 동안 암컷의 최적 영양공급을 하게 하는 유리한 선택압에 의 해 발달해 왔으며 결국 기능 확대 혹은 기능 변화를 통해서 비로소 짝짓기 행동에서 제 2 의 공격억제 효과를 갖게 되었다고 가정할 수 있댜 이러한 기능 변화와 관련해서 먹이먹이기는 자주 상징적인 특 징만을 갖기도 한다 . 이 행동은 많은 조류에서 〈부리 내밀기〉와 같 은 행동이 아직 행해지고 있는데 이때 먹이는 더 이상 건네주지는 않는댜 여기서 많은 근연집단으로부터 두 개의 가능성이 이미 발달 되었음을 확인할 수 있다. 종비교를 통해 이 행동의 가상적 계통발 생학적 발달이 뒤늦게 이루어졌음을 알 수 있다. 닭목(目)의 조류의 수컷들은 구애시 암컷에게 먹이룰 제공하는 반면 일부에서는 전혀 먹이가 없는 데도 불구하고 부리로 바닥을 쪼거나 아래를 가리키는 동작을취한다.
먹이먹이기 행동의 두번째 범주인 구걸 행동에서도 짝짓기 행동에 서의 동작, 몸짓 그리고 소리내기를 넘겨받았다 . 그래서 많은 명금 류의 새끼들이 어미에게 날개를 떠는 동작을 취하여 먹이를 달라고 구걸하는 것처럼 암컷은 수컷에게 이런 동작을 취해 교미를 요구한 댜 포유류(햄스터, 영양, 사슴)에서도 교미 도입 과정에서 수컷들이 소리를 내는데 이것은 성체의 행동 레퍼토리 가운데 어린애 같은 행 동 요소로 이것을 유아행동i n fa n ti li sm 이라고 부른다 . 물론 공격성 억제효과를 지닌 짝짓기 행동 영역 가운데 교미 후의 행동인 짝짓기 이후의 행동(후짝짓기 행동)도 포함된다. 이것은 교미 후에 곧바로 일어나며 일반적으로 원래의 짝짓기 행동과는 구별되는 매우 복잡한 고도로 발달된 행동을 말한다. 오스트레일리아 되새류의 일종인 Poep h i laacu ti dauda 의 수컷은 교 미후에 암컷 곁에 바싹 다가선다. 그런 다음 상체를 아래로 구부리 면서 머리를 돌려 이 새의 전형적인 검정 목부위를 암컷에게 낮은 자세에서 보여준다. 이때 이 검정 목은 상호간의 깃청소를 하게 하 는 중요한 해발인으로 작용한다. 많은 아프리카산 되새류의 경우 수 컷들은 교미를 성공시킨 뒤에 암컷 곁에 앉아서 암컷의 교미 자세를 취한다 . 이로써 암컷은 날아오르는 듯한 약한 의도동작을 취한다. 청둥오리 수컷도 교미 후에 분명히 암컷 짝짓기 동작인 자맥질 동작 을 보여준다 그리고 이 동작을 취하면서 암컷의 주변을 돈다. 혹고니 Cy gn u~ olor 의 경 우 교미 후 암수는 가슴과 가슴울 마주 대 고 위로 일어선 다음 부리를 좌우로 2-3 번 정도 혼든다. 이러한 짝 짓기 이후의 행동에 대한 생물학적 의미는 교미 후 배우자와의 계속 된 동거를 위해서 다시 발생할 수 있는 공격 내지는 도피 성향을 극 복하는 데 있다. 그러므로 이 후짝짓기는 동시에 짝유대을 위해 행 해지고 있다. 이 후짝짓기 행동의 또 다른 의미는 교미시에 쓰고 남은 여분의
정력을 소비하는 데 있다 . 이런 견해는 최소한 선택압이 어느 정도 작용하는 데서 찾아볼 수 있다. 즉 이 선택압은 단순히 과잉의 에너 지 소비 목적을 갖고 복잡한 행동으로 발전하게 할 수 있었던 것이 다 (Bas t ock 1969) . 3) 동조성 인간, 가금류, 열대성 척추동물 그리고 많은 기생동물을 제외하고 는 대부분의 동물들의 번식은 특정 연주기에 한정해서 일어나고 있 댜 이렇게 번식주기가 일정한 데에는 두 가지 이유가 있다. 한 가 지는 성숙한 배아세포의 생산으로 생식과 결부된 다양한 양육 행동 들이 특히 생리적으로 요구되기 때문이며, 다른 하나는 새끼가―― 최대로 생존 기회를 보장해 주는――먹이 상태가 질과 양적으로 유 리하고 그 먹이는 쉽게 접근이 가능한 정해진 시기에 태어나야 하기 때문이다. 동물의 종 혹은 집단의 생식시기는 계통발생과정에서 자연선택에 의해 일년 중 특정한 달에 한정되어 왔다 . 이 특정한 달은 선택요인 으로 먹이제공 이외에 다른 환경 조건들이 최소한 부수적인 영향으 로 작용할 수 있었다. 예를 들면 온도, 공기중의 습도, 토양의 습 도, 산란이나 새끼 돌볼 가능성 그리고 다른 미세한 요인들을 말한 댜 많은 열대성 동물들과 같이 생식 준비기간이 매우 길 수도 있고 어쩌면 수 개월이 걸릴 수 있거나, 남태평야의 팔로로웜(산호초에 서 식하는 털갯지네과의 다모충)과 같이 일년 가운데 며칠 혹은 몇 시간 동안 국한될 수 있다. 생식과 관련된 생리과정의 시기에 맞추어 생 식행동은 규칙적으로 일년중 일정한 시간에만 나타나고 있다. 바로 성호르몬이 이에 대한 조절역할을 맡고 있다. 하나의 생물학적 중요한 문제는 생물체와 생물체 주변의 주기적 변화 과정 사이의 동조성인 것이다. 이것은 생식주기에서 특히 명확
해졌다. 출산은 발생주기 그리고 새끼의 독립은 성장주기의 마지막 단계에 나타난다 . 생식선의 성장, 둥지짓기 그리고 여타의 다른 양 육 행동형태 및 배발생이 이런 발생 혹은 성장주기 과정에서 일어난 댜 이것은 종의 크기에 따라 몇 주 혹은 몇 달이 걸릴 수 있다. 대 개가 이 과정은 가장 유리한 생존조건의 시점 이전에 이미 진행되고 있기 때문에 어느 정도 그 전에 일찍 끝나게 된다 . 그러므로 생물체 는 아주 이른 시기에 앞으로의 더 나은 주변 환경에 대한 정보를 입 수하는 것이 전제조건이 된다. 출생 전 다소 긴 시간 간격으로 생식 주기를 작동시키는 주변의 환경자극과 또 생식에 필요한 생리적 성 숙과정을 위해 시간적으로 계획된 이 환경자극들은 규칙적인 것도 아니고 우선 먹이 제공이나 새끼의 성공적인 양육을 위한 환경요인 들도 아니다 오히려 이것들은 생식을 위한 유리한 조건들이 나타날 것을 미리 알려주는 경보성 요인들인 것이다. 지리적으로 높은 위도 에 사는 많은 동물들에게 낮의 길이가 1 년주기로 차이가 나는 것이 이에 속한다. 즉 동지가 지나면서 낮이 점점 길어짐은 많은 조류에 서 생식선의 성숙과 생식행동의 재유발을 일으킨다. 생물학적 용어 로, 계통발생 과정에서 각각의 생식시기의 발달을 가져오게 하는 환 경요인들에 대해 직접요인 ultim ate fa c t or 이라 하고 실제로 이러한 적 응을 조절하는 자극을 간접 요인 pro xim ate fa c t or 이 라고 부른다. ** ** 이 두 용어는 원래 조류학에서 기원되었으며, 맨 처음 연주기의 범위에 한해 서 사용되었다. 그러나 이것이 처음 소개된 이후로 계통발생학적 선택 요인과 실제적 조절 요인 간의 이러한 차이가 다른 행동의 범주에서도 관찰되었다. 그 래서 오늘날 간접요인으로써 환경에 특히 유리한 성질을 보이는 개체의 좀더 높은 생활 기대와 번식 성공에 대해 이러한 성질이 한 종내에서 혹은 한 개체군 내에서 점차 확산시켜 온 환경조건들을 가리킨다 . 예로써 생활공간의 선택 특 히 번식을 위한 적합한 장소의 선택을 들 수 있다. 이때 직접요인들은 여러 생 활공간에서 생존 내지는 번식하는 개체의 생존율과 번식성공률에 영향을 미치 는 것들을 말한다(예 , 먹이상태, 위장의 가능성). 간접요인은 예를 들면 땅바닥 의 색깔과 구조, 먹이의 밀도와 생김새, 잎의 모양과 크기로써 이것들은 적합
한 생활공간을 인식하고 다시 찾을 수 있게 하는 어떤 개체에게 필요한 신호가 된다.
낮의 길이나 그밖의 환경조건들 이외에도 이 성행동은 중요한 간 접요인이 될 수 있다. 이것은 다음의 이유에서 생물학적 의미를 갖 는댜 일반적인 환경요인들을 통해, 무엇보다 이 환경요인들이 연중 매우 눈에 띄는 변화를 보일 때, 한 개체군 내에서 생식주기들이 일 제히 가동된다. 그렇지만 연령이나 경험이 다른 개체의 차이가 항상 존재하게 된다. 성공적인 교미는, 무엇보다 물고기들의 체외수정은 물속에 정액과 난자를 뿌려놓지만 암수간의 정확한 시간적 일치를 요구하고 있다 . 이런 정확성은 일반적인 환경요인들에 의해 가능해 질 수는 없다. 그것은 단지 이들의 상호작용이 일치할 때만이 도달 될 수 있다. 그러므로 성행동의 또다른 과제가 암수의 동조성에 있 게 된다. 난혼을 하는 동물의 경우(암수가 실제 짝을 짓지 않고 교미만 을 위해 만나는 종) 이 과제가 교미로 들어가므로 해서 충족되는 반 면, 오랜 기간 동안 결합을 유지하는 동물의 경우 이 과제가 짝유대 행 동에서 넘어오기 때문에 오히려 눈에 덜 띈다 (Ascho ff 1955, Immelmann 1962) . 이 시간적 일치는 일반적으로 암수가 환경의 자극에 강하게 반응 한 다음 한 쪽이 다른 쪽의 성적 발달을 자극하므로 해서 도달된다 . 일반적으로 수컷은 환경개방형인 성을 갖고 있다. 명금류의 연구에 의하면 수컷이 일조량의 상승 죽(인공적인) 낮의 길이의 증가를 통해 고환의 성숙, 노래부르기 그리고 성적 행동양식들의 유발에 이를 수 있다는 사실이 확인되었다. 이에 비해 암컷은 같은 조건에서 난소의 부분적 성숙이나 관련된 성행동의 불완전한 발달을 가져온 반면 행 동의 근본적인 역할에서 이런 환경요인에 의해 비로소 완전한 성숙 에 도달된다. 여기서 수컷의 생식 준비기간이 암컷의 생식준비기간 보다 종종 더 길어지게 된다. 그래서 많은 조류에서 수컷들의 고환
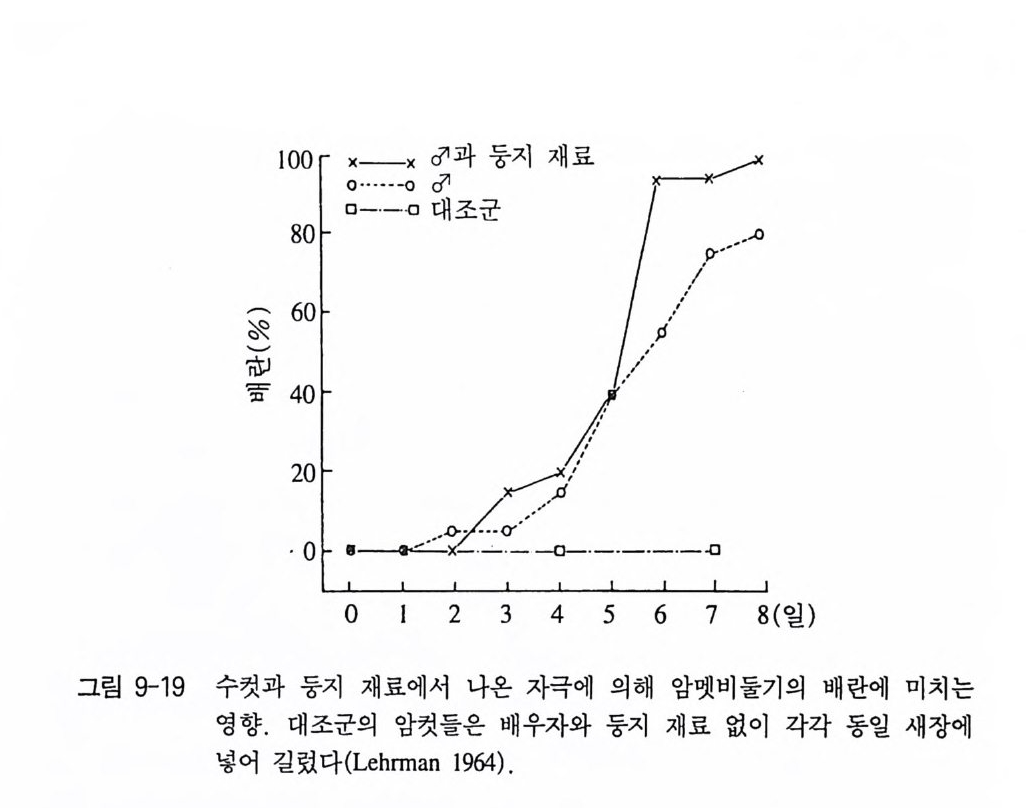 100
100
이 암컷들의 난소보다 더 이른 시기에 발달하게 된다. 철새의 경우 수컷들이 며칠 일찍 성선이 발달된 채 번식지에 먼저 도착한다. 암 컷에 대한 수컷의 구애행동의 자극효과는 이미 많은 관찰과 실험들 에 의해 입증되었다. 닷쥐, 토끼, 땅다람쥐, 밍크 그리고 족제비와 같은 포유류의 암컷들은 항상 성공적인 교미가 이루어져야만 배란을 한댜 예를 들어 토끼의 경우 교미 후 10 시간 정도 지나야 배란을 한다 소위 이런 유도배란은 난세포들이 수컷의 정자가 있을 때만이 만들어지고 있음을 뜻한다. 여기서 교미행위가 시간을 일치시키는 역할을 하고 있다는 사실을 알 수 있다. 성행동과 관련된 다른 유사 한 행동들도 이런 효과를 갖고 있다. 예를 들면 한 마리씩 기른 멧 비둘기 암컷은 새장에 있는 둥지에는 관심을 보이지 않을 뿐만 아니 라 산란도 하지 않지만 수컷을 넣어주면 며칠 이내로 알을 낳기 시 작한다(그림 9 一 19). 이때 유리창 밖으로 비둘기를 놓아 주어도 같은
결과가 일어나지만 만일 구애행동을 하지 못하는 거세된 수컷을 새 장에 넣어주면 산란을 유도할 수가 없다. 잉꼬의 경우 음성학적 자 극이 중요한 역할을 한다. 수컷의 소리를 듣고 자란 암컷둘은 적어 도 18 일 이내에 산란을 시작했지만 수컷의 접촉을 갖지 못한 암컷들 은 실험이 시작된 지 30 일이 지나도 산란을 하지 못했다 . 이와 비슷 한 결과가 다른 비둘기 종과 많은 명금류에서도 발견되었다. 포유류 의 경우 후각적 자극의 영향에 대해 연구된 바 있다 . 생쥐의 경우 수컷을 그물망으로 된 우리안에 가두었을 때 이 수컷은 암컷의 생식 선 발달에 긍정적인 영향을 미칠 수가 있다. 또 전에 수컷이 살던 우리에 암컷을 넣어도 마찬가지의 결과가 일어난다. 멧비둘기를 포함한 일부 조류에서 암컷의 호르몬 생성이 구애를 하는 수컷의 자극을 통해 일어난다는 사실이 잘 알려져 있다. 즉 수 컷의 구애행동은 암컷의 생식선의 발달, 산란 그리고 둥지짓는 행동 을 하게 한다 . 암수의 동조효과가 많은 동물들의 구애행동에 내재되 어 있다고 추측할 수 있다 . 즉 암수의 연속 동작이 오랫동안 이루어 질 때 보면 암수가 똑같이 아니면 거의 비슷한 행동을 보여준다. 그 것은 많은 벌새의 혼인비행 (그림 9-20) 이나 큰가시고기의 혼인춤과 같이 하나의 행위가 상대방의 그 다음 동작을 유발시킨다. 구애행동의 자극 및 동조 효과가 배우자에게만 항상 한정해서 일 어나는 것은 아니다. 오히려 그것은 어떤 경우에 다른 쌍에도 전염 의 효과를 가질 수 있고 그래서 행동과 호르몬 간의 상호작용으로 인해 생식준비에 영향을 미칠 수 있다. 결국 동종의 전체 집단 내의 생식발생이 동시에 작용할 수 있게 한다. 여기에도 수많은 관찰과 실험적 증거들이 있다. 집단번식을 하는 조류의 경우 집단내 혹은 집단 지역내의 쌍들은 거의 동시에 산란을 하고 포란을 시작하는 반면 지역적으로 떨어져 있는 다른 집단과는 커다란 차이가 생겨난다. 역시 대군집에서의 산란 시작은 소집단에
 /> '.,,./
/> '.,,./
서보다 오히려 일찍 나타나며 중심부에 밀집되어 있는 개체들이 의 각의 개체들보다 일찍 산란을 한다. 이러한 차이는 번식장소에서의 일반적인 환경 요인보다는 오히려 상호간의 영향에 원인이 있다 . 사 실 잉꼬의 경우 암수의 생식선의 발달이 구애하는 이웃쌍에 의해 촉 진될 수 있으며, 멧비둘기의 경우 한 쌍씩 새장에 넣어 독립해서 키 운 것보다 쌍이 든 새장들을 함께 해서 키웠을 때 암컷들의 생식선 성숙을 수컷의 소리가 촉진시킬 수 있다. 몇몇 종들은 구애동작이 쌍의 배우자 사이에서만 일어나는 것이 아니고 많은 개체들의 합동 구애로의 발전을 통해 집단동조 효과를 한층 증대시켜 왔다. 이에 대한 좋은 예로써 날개를 펼쳐서 몸 치장
하는 홍학의 의식행동을 들 수 있다 . 바로 수십 혹은 수백 마리의 홍학들이 이 의식에 참여하여 집단동조 효과를 이룬다** .
** 여러 마리의 수컷들이 암컷들을 끌어들이기 위해 합동구애를 할 경우 여기서 사회적 구애라는 용어 사용은 피한다.
암수간의 상호 동조성에 대한 생물학적 의미가 수정을 보장하는 데 보다 중요한 반면 그것이 짝 이외의 시간적 동조성에는 수정과 관계가 없이도 작용한다 . 많은 수서동물들은 수많은 수컷들과 암컷 들이 동시에 물속에다 성세포를 방출할 때 알의 수정 기회를 높여 주고 있기 때문에 이 목적 역시 같다. 그러나 집단번식하는 조류의 경우 다른 이유가 작용할 수 있다. 여기서 산란 시작의 정확한 동조 성은 다음의 두 가지 점에서 포식자로부터 지키는 데 역할을 한다. 한 가지는 모든 집단 구성원들이 동시에 있다는 것은 포식자에게 공 동의 공격효과를 높여줄 수 있고, 다른 한 가지는 집단번식 시기에 먹이탈취자(포식자)들에게 먹이 과잉공급이 생겨날 수 있다. 죽 번식 기간이 오래 지속되는 것보다 알이나 새끼로서의 기간이 짧을 때 이 런 과잉공급 효과가 일어난다. 알과 새끼가 있는 동안은 이 집단의 번식기에 포식자에게 먹이의 공급과잉이 생겨난다. 그렇지만 이 기 간은 번식의 전체 시간을 감안하면 아주 짧은 기간으로 이 집단에게 는 전체적인 손실이 줄어드는 셈이다. 그밖에도 집단번식은 한 쌍이 둥지를 짓고 이웃 쌍은 포란을 하는 것보다 모든 쌍이 번식시기(둥지 짓기-포란-새끼양육)를 같이 했을 때 이웃간의 방해의 횟수를 확실 히 줄일 수 있다 . 열대성 기후와 같이 연주기가 비교적 불규칙적이 어서 생식선 발달을 일으키는 환경자극이 결여된 기후지역에서는 사 회적 자극이 부수적인 의미를 갖고 있다. 죽 이 지역에 사는 쌍들은 구애행동으로 서로를 홍분시켜 유리한 생활 조건이 오면 생식이 급 속이 시작될 수 있다. 이런 식으로 이미 언급된 바 있는 홍학의 집 단구애도 생식 시기가 같은 개체들이 함께 모여 공동의 번식집단을
이루게 하는 데 기여하고 있다(I mmelmann 1971). 4) 생식 격리 타종과의 교잡은 생물학적으로 단점이 된다. 일반적으로 그것은 생존 혹은 생식가능한 자손을 갖지 못하고 그 종의 번식 잠재력을 소멸시키는 결과를 가져온다. 대개 매우 가까운 근친인 경우에 수정 가능한 교잡을 이룰 수 있는 종일지라도 유전자의 혼합은 유전자복 합체의 불균형으로 행동의 원인이 되는 호르몬에 자주 방해를 일으 켜서 환경적응에 장해를 가져온다 . 그러므로 종간 교배의 방해가 강 한 선택압으로 작용하여 자연적인 조건하에서의 교잡은 대개의 동물 들에게 거의 일어나지 않는다 (Ma y r 1967). 생식 격리로 모든 형질들은 교잡가능한 종들을 구분하는 도구가 된댜 예를 들면 곤충, 거미, 그리고 많은 달팽이류에서 수컷과 암 컷의 생식기관이 종특이적인 자물쇠-열쇠 시스템으로 되어 있어 단 지 동종의 배우자에게만 교미가 가능하다. 효과적인 장벽은 무엇보 다 많은 수서동물들에서 일어나는 것처럼 짝짓기 시간대의 연중 차 이다 또 초파리나 아주 근연의 물고기 종들에서 알려진 것처럼 짝 짓기 활동이 하루중 여러 시간대에 나뉘어 있다. 그러나 종간 교잡 을 막는 가장 중요한 역할은 행동에 있다. 여기서 성적 행동의 중요 한 과제가 제시될 수 있다. 암수를 결합하게 하는 신호들이 일반적 으로 매우 종특이적으로 나타나기 때문에 자기 종의 소속 개체만이 그 신호에 반응을 할 수 있다. 그것은 다음의 두 가지 방법에 의해 도달될 수 있다. 신호 자체가 그 성격에 있어서 혹은 신호를 내는 종에 있어서 차이가 나거나, 그 렇지 않으면 신호의 조합에서 오차가 생겨날 가능성을 최대로 줄일 수 있는 여러 신호들이 구애 순서의 형태로 연결되어 제시된다. 그 래서 배우자 동조성 효과 외에 좀더 복잡한 구애순서의 기능이 종격
리의 지역에서 고려될 수 있다. 보통 교잡의 위험성이 크면 클수록 두 종은 서로 더 가까운 근친 일 가능성이 높다. 그래서 두 종은 분류학적으로 서로 가까운 위치 에 놓여 있게 된다. 따라서 종격리의 특별한 문제가 여러 근연종들 이 동일 지역에서 함께 사는 곳에서 생겨나게 된다. 여기서 성적인 해발인이 특히 정확해야 하며 혼란을 초래해서는 절대 안된다. 많은 오리류가 함께 사는 지역에서 숫오리들의 화려한 깃털은 눈에 띄게 매우 다양하다. 이에 비해 몇몇 오리류가 살고 있는 지역 특히 서로 바뀔 위험이 적은 지역에서는 이 숫오리들도 종종 우중충한 색깔 다 시 말해 해발인이 빈약하게 발달되어 있다. 이러한 차이는 한 종내 에서도 나타날 수 있다. 하와이 섬에서 서식하는 수컷 청동오리들은 본토에 사는 아종에 비해 일년 내내 눈에 띄지 않는 암컷과 비슷한 깃털을 갖는다. 많은 조류들은 근연종들이 여럿이 살고 있는 지역에 서 각 종마다 매우 특징적이고 잘 분간할 수 있는 노래를 부르는 반 면 작은 섬지역처럼 동소종의 수가 적은 곳에서는 종내의 노랫소리 가 훨씬 더 큰 변이를 보여준다. 이에 대한 예는 카나리아 제도의 테네리페섬의 푸른박새 Parus caeruleus 와 대서양에 위치한 아조레스 제도의 상모솔새 Re g ulus re g ulus 가 있다. 특히 동소종들의 정 확하고 종특이적인 신호에 대한 예는 반딧불이의 불빛(그림 9 一 21), 메뚜기 과의 곤충(그림 9-22) 과 무미류의 소리, 밤나방들의 냄새물질 혹은 농게류의 집게발의 움직임이 해당된다. 개체군간의 생식 격리 자체가 한 종을 형성한다는 연구가 이미 알 려진 바 있다. 생식 격리는 특정 환경에 개체군 특이적인 미세한 적 옹이 유전적으로 작용하거나 혹은 학습과정의 기초에 의해 성립되며 그리고 전수로 인해 전달되어 여러 적응 형태의 개체들의 교잡을 통 해 다시 그 적응이 없어져 가는 것을 차단한다 . 이런 종내의 격리에 있어서 행동형질들은 중요한 역할을 할 수 있
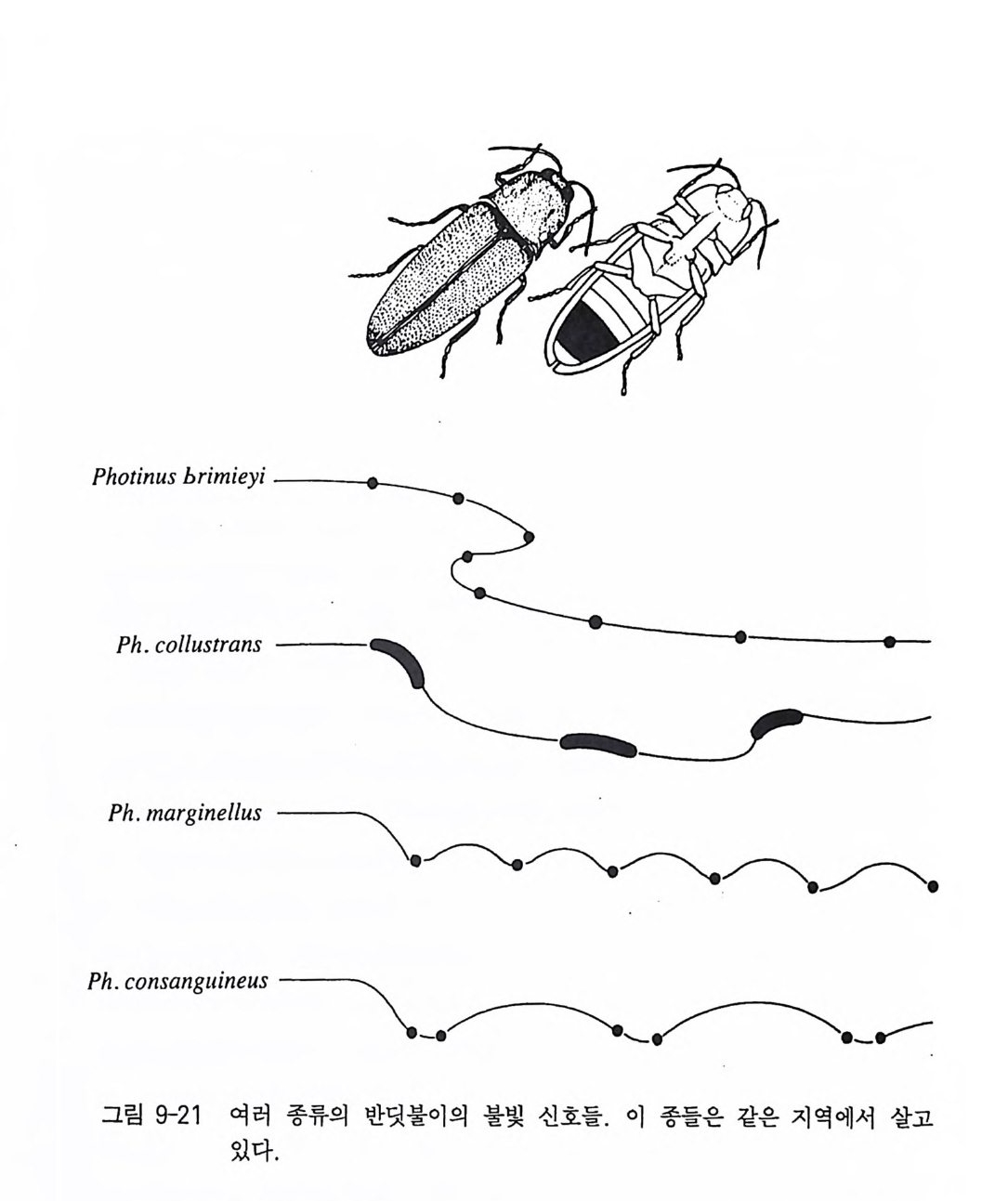 Photi nu s brim i ey i -._____
Photi nu s brim i ey i -._____
댜 조류들의 방언이 근래에 많이 연구되고 있다. 방언이란 같은 종 의 노랫소리가 지역에 따라 서로 다른 소리를 내는 것을 뜻한다. 비 록 방언의 정확한 기능에 대해 아직 완전히 밝혀지지 않았지만 최소 한 몇몇 경우에서 방언이 종내의 특이적 적응을 확보하는 데 기여할
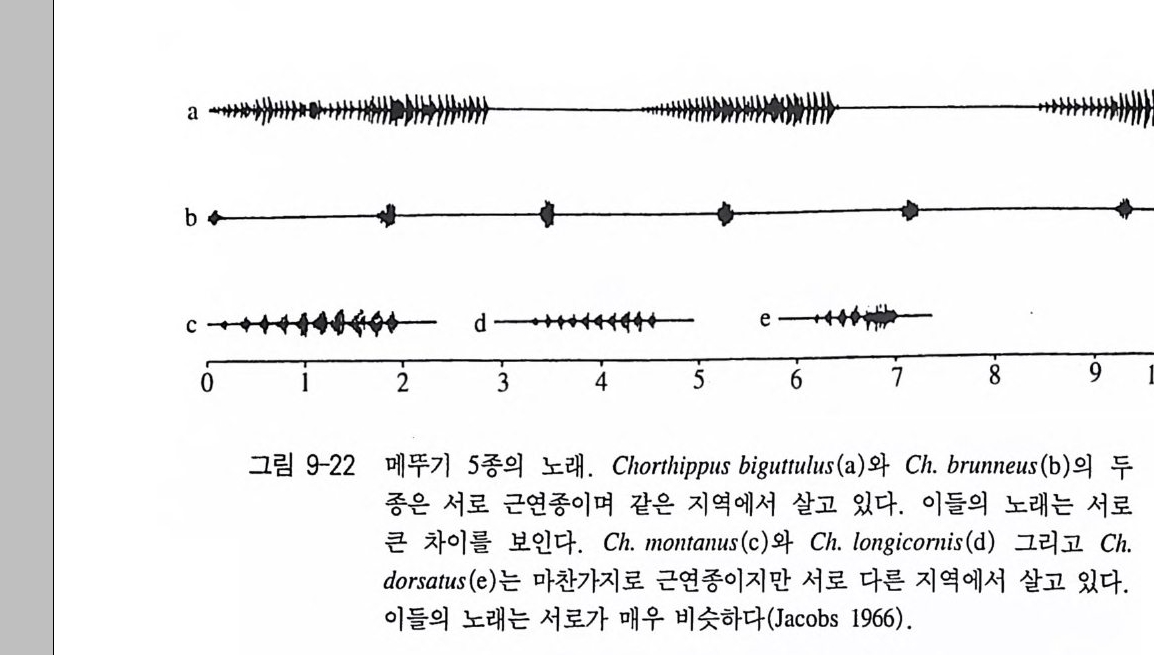 ba ♦께 '메'"사 1 패 )읊'))h))h i h) ` nIIIIIH)l'•HHl) 삐 WllI) • 패卜빠,빠一牌
ba ♦께 '메'"사 1 패 )읊'))h))h i h) ` nIIIIIH)l'•HHl) 삐 WllI) • 패卜빠,빠一牌
수 있다고 추측하고 있댜 즉 이것은 다양한 생태형을 만들어 온 종, 말하자면 각각의 개체군들이 자신들의 환경 요구(예를 들면 특정 먹이 선호나 생태 지위 선호)에서 다소 서로 큰 차이가 나는 종에서 적 용될 수 있댜 여기서 방언은 암컷들이 어렸을 때 부친이나 이웃 경 쟁자들로부터 들은 노래 유형과 유사하게 노래를 하는 수컷들과 짝 을 이룰 때 올바른 짝 형성, 다시 말하면 동일 적응 형태의 배우자 들과의 만남을 촉진시킬 수 있다. 결국 방언은 암컷과 수컷들이 겨 울 서식처에서 다시 돌아왔을 때 각각의 개체군 지역을 발견토록 하 는 중요한 길안내자 역할을 할 수 있다. 이런 면에 대한 증거가 멧 새류에 속한 Emberiz a hor t ulana 에서 명확히 제시되었다. 이 새는 여 러 생태형들이 분명히 자기 지역 방언을 사용하고 있었으며 그 밖에 도 방언을 하는 다른 조류의 연구에서 방언이 ·모자이크식 격리 효과 가 있음이 밝혀 졌다 (Conrads 와 Conrads 1971 ).
2,3 짝유대의 행동 동물들은 암수가 교미만을 위해 만나기도 하고 일부 종에서는 좀 더 오랜 기간을 암수가 함께 머물면서 짝을 형성하기도 한다. 장기 적 짝유대는 모든 척추동물문과 게와 곤충에서 각각 밝혀 졌다 (Hendri ch s 1978, W ick ler 1966) . 암수가 함께 있음으로 해서 생기는 생물학적 이점은 많다. 우선 새끼를 양육하는 데 암수가 함께 참여하는 곳에서는 필수 요건이다. 그러므로 이것은 번식이 일어날 때에 계속 중요한 역할을 담당하고 있다 그것은 새로운 짝짓기와 암수를 결합하게 하는 행동양식들을 많이 만들 수 있다. 뿐만 아니라 배우자의 동조성을 촉진시킬 수 있 고 종간 교잡을 어렵게 하여 생식 격리에 기여할 수 있다. 만일 암수가 이미 번식기가 시작되기 전부터 함께 살았다면 그들 의 생식선의 연주기 발생은 이들을 조절하는 의부요인들이 실제로 같을 뿐만 아니라 상호간의 행동학적 영향을 통해 그들이 번식기 직 전에 처음 만났을 때보다 더 일치하게 된다. 이러한 동조성은 번식 의 시점에 직접적으로 효과가 나타날 수 있다. 세가락갈매기는 이미 전년도에 함께 번식을 경험했던 쌍은 새로이 짝을 이룬 쌍보다 일찍 산란을 시작한다. 마지막 부화 단계에서 파트너를 바꾼 암컷은 역시 번식성공도가 낮다. 다시 말하면 이런 암컷은 적은 수의 알을 낳고 그 알에서 새끼가 깨어나와 날 수 있을 정도로 성장하는 비율도 떨 어진다. 그러므로 한 쌍내에서 장기적으로 일어나는 이 동조성은 중요한 과제들 가운데 하나를 떠맡게 된다. 비둘기에서 원래의 구애행동 죽 교미의 도입과정은 이미 보다 오래 함께 산 쌍들의 경우 새로운 쌍 보다 더 짧고 덜 눈에 띄며 그래서 좀더 세련되게 진행된다. 짝유대와 생식 격리간의 관계가 무엇보다 암수가 장기적으로 함께
사는 종과 살지 않는 종이 존재하는 동물 집단에서 확인되었다. 예 를 들면 뉴기니의 극락조 Parad i saedae 의 조류가 그런 경 우다. 이 조 류의 집단에서 잡종들이 비교적 자주 나오고 지금까지 속(屬)과 종 (種)의 수가 많이 기록되었다. 그들은 생식 준비가 된 암컷이 구애장 에서 수컷을 찾고, 교미 후엔 혼자서 둥지를 짓고, 포란을 하며, 새끼 를 양육하는 그런 종들이다. 이에 비해 암수가 견고한 쌍을 이루어 수컷이 새끼 양육에 참여하는 종에서는 하나도 잡종이 발견되지 않 았다 높은 교잡의 빈도는 일반적으로 결함 있는 짝형성을 하는 조 류에서 발견되는데 무엇보다 벌새, 마나킨새, 뇌조가 대표적이다. 그러므로 암수간의 장기적인 유대는 확실히 결정적으로 잘못된 짝짓 기를 피하는 데 기여할 수 있다. 2.4 유대의 형태 일반적으로 쌍내에서만이 죽 한 마리의 수컷과 한 마리의 암컷 사 이에서 만이 이성결합(異性結合)이 존재한다. 이러한 일부일처제는 일년 내내 혹은 거의 평생 유지되기도 하는데 이것은 모든 척추동물 에서 나타나며 부분적으로 무척추동물 가운데 사막에 사는 등각류와 일부 게 종류에서 알려져 있다. 조류의 경우 일부일처제가 특히 잘 발달되어 있는 반면 포유류에서는 그것이 드물기는 하지만 개과의 식육류, 설치류 그리고 유제류의 일부와 여러 영장류(예, 긴팔원숭이 Hyl o bati da e, 티 티 원숭이 Brachy tel es arachnoid e s 마모셋 Call ithr ic ida e) 에 서 알려져 있다 (Emlen 과 Or ing 1977, Lins enma ir 1979, W itt enber g er 와 Til so n 1980). 그러나 하나의 쌍을 넘어 지속적인 유대가 여러 마리의 이성의 동 료에 확대될 수 있다 . 만일 한 마리의 수컷이 여러 마리의 암컷들과 짝을 이룬다면 그것을 우리는 하렘이라고 부른다. 이 하렘은 포유류
(예, 많은 영장류, 얼룩말, 일부 영양), 어류(키클리드과의 물고기)에서 잘 알려져 있다 (Emeln 과 Orin g 1977, W itt enber g er 와 Til so n 1980). 이것 과 반대 의 관계 인 일처 다부제 는 태 스마니 아의 물닭 Trib o ny x mort ier i 이 유명하다. 이 물닭은 규칙적으로 두 마리의 수컷들이 단지 한 마 리의 암컷과 짝을 이룬다(Ri d p a th 1972). 그러나 최근까지 일부일처제라고 알려졌던 조류에서 그렇지 않다 는 결과가 나와 관심을 끌고 있다. 야의 관찰을 통해 오스트레일리 아의 금화조는 한 암컷이 교미하는 전체 횟수 가운데 평균 5% 가 자 신의 배우자가 아닌 다른 수컷과 교미하는 것으로 관찰되었다. 최근 개발된 유전자 지문 DNA fi ng e rpri n ti n g에 의해 한 배에서 나온 금화 조 새끼들의 8% 가 자신의 아버지가 아닌 다른 수컷의 새끼들로 밝 혀졌다 이런 유대형태를 〈쌍이의자 짝짓기 ex tr a- pai rma ti n g〉라고 한 댜 그 밖에도 유전자 지문을 이용해 많은 조류에서 이런 쌍이의자 짝짓기가 알려지게 되었는데, 지금까지 유럽산 122 종의 참새목 조류 의 연구에서 39% 가 이런 유대 형태를 갖고 있는 것으로 나타났다 (B i rkhead 와 Molle~ 1992) . 2.5 유대의 기작 한 쌍의 장기적 유대는 기본적으로 다음 두 가지 방법에 의해 보 장을 받을 수 있다. 하나는 환경과의 관계 그리고 다른 하나는 배우 자와의 상호결합을 통해서 이루어진다. 환경과의 관계에 대한 좋은 예는 많은 게 종류에서 발견할 수 있는데 이들은 어느 정도 강요된 부부관계를 형성한다. 수컷과 암컷이 함께 이미 어린 시기에 해면이 나 산호초 구멍으로 기어 들어가 그들의 여생을 그곳에서 보낸다. 이들은 이 구멍에서 더 이상 빠져나갈 수 없을 정도의 크기까지 자 라기 때문에 특별한 유대 기작이 요구될 필요가 없게 된다. 많은 철
새들의 경우 암수가 각기 서로 독립적으로 살았던 겨울 서식처에서 이른 봄에 다시 전년도의 번식지에 도착한다. 그래서 이들의 짝유대 는 바로 장소 의존적이 된다. 이 짝유대는 암수가 단지 번식지에서 강한 결합을 갖고 다음해에 자동적으로 다시 서로 만나 쌍이 유지될 수 있다 쌍이외의 범주에서 일종의 집합과 비슷한 성질을 갖고 있 는 이런 짝유대 형태는 장소부부 혹은 경우에 따라 배우자가 개별적 으로 서로를 모르고 있기 때문에 익명의 일부일처제라고도 부른다. 그러나 지속적인 짝유대에서 대개 배우자간의 진짜 결합이 생겨난 댜 이것은 한 마리의 수컷이 자기 짝의 암컷에게 낯선 암컷들을 대 하는 것보다 달리 행동하기 때문에 개별적 인식이 전제된다 . 이 경 우에 특정 행동양식들이 이 유대를 유지시키는 데 기여할 수 있다 . 이 행동양식들의 형태와 기원이 각 종들에 따라 매우 다양하다. 특 히 조류에 있어서 일부일처제가 넓게 자리잡고 있기 때문에 잘 연구 되어 있는 실정이다 . 여기서 두 가지 규칙성이 발견된다. 종종 짝유 대의 행위들 역시 구애행동에서 출발하여 쌍의 배우자들-쌍 가 운데 하나 혹은 양자-로부터 만들어지기도 한다 . 이 가운데 원래의 구애 즉 교미 준비에서 생기는 일련의 똑같은 행동들이 짝유대를 하게 한다. 그래서 오스트레일리아의 금화조는 번식기 이외에도 자주 구애행동을 취한다 . 아 행동은 짝유대의 전 단계로 보통 교미에까지 이르지는 않는다. 영장류 가운데 개코원숭 이 Pa pi o 나 히말라야원숭이 Macaca mula tta 는 교미행위 자체가 여러 암수 개체들간의 유대를 ‘ 촉진시킨다. 연결된 교미행위들은 간헐적인 멈춤동작에 의해 나누어져 실제로 사정에까지 이르기 때문에 이 개 별적 교미동작은 유대기작으로 취급될 수 있다. 이와 같은 현상이 인도산 과일박쥐 P t ero pi dae 에서 잘 알려 져 있다. 이 과일박쥐 는 사정 까지 3-7 번의 교미동작들이 필요하다. 이런 연속 동작은 번식기에만 존재하지만 개별적 교미동작은 모든 시기에 걸쳐서 일어나고 있다.
심지어는 이 개별 교미동작은 임신중인 암컷과도 일어난다 . 또 어떤 경우에는 구애행동에서 이탈된 부분 행위들이 개별적으로 나타난다 . 이것은 번식기 이외에도 나타나는 것으로 보아 확실히 짝유대에 기 여하고 있다 . 예를 들면 많은 앵무새류 그리고 까마귀과의 Corvus cora x 에서 일년 내내 관찰되는 구애 먹이먹이기가 이에 속한다 . 암수가 함께 하는 짝유대 행동 양식들에는 이중창 혹은 노래교환 이 속한다. 이것은 무엇보다 열대성 조류에서 흔히 나타나지만 긴팔 원숭이 H y loba ti dae 와 같이 일부일처제의 포유류에서도 가끔 나타나 기도 한다 이들 종에서 한 쌍의 배우자들은 자기들의 노래를 동시 에 혹은 서로 바꿔가면서 부른다 . 아마 원래는 암컷과 수컷의 노랫 소리가 같다가 점차 노래를 구성하는 절과 절의 부분들이 암수간 서 로 다른 차이를 가져왔을 것으로 보인다. 몇몇 아프리카산 때까치는 특히 고도로 발달된 공동의 노랫소리를 갖고 있다. 이 노래는 각자 서로의 극히 짧은 반응 시간을 두고 불려지는데 붉은배숲때까치 Lania r iu s barbarus 의 경 우 단지 약 125ms 의 반응 시 간 기 록을 갖고 있댜 이 이중창의 노래는 번식기 이외에도 불려지고 있기 때문에 이 노래는 쌍의 지속적인 유대에 기여하고 있음을 알 수 있다. 그 밖에도 야행성 혹은 밀집된 삼림에서 사는 동물들의 경우 암수의 소 리내기는 중요한 장소를 가르켜 주는 역할을 하며 상대방울 서로 발 견할 수 있게 한다 (Helversen 1980, Hin d e 1978) . 포유류와 조류에 있어서 짝짓기 행동의 좀더 확대된 행동양식으로 써 상호간의 털이나 깃털다듬기 (=화장gr oo mi n g)가 보편화되어 있다 (Cullen 1953). 화장을 위해 새들은 부리를 사용하고 영장류는 손을 사용하며 그 밖의 포유류는 혀나 이빨울 사용한다. 사회적 화장 soci al g roo mi n g이 화장을 받는 자 스스로 닿지 못하는 곳에 우선적 으로 이루어져야 하기 때문에 화장은 상호간 청소를 해주고 기생충 울 제거해 주는 데 기여하고 있다. 그러나 이런 청소 기능의 실질적
기능은 아직 잘 알려지지 않았다. 어쩌면 이 실질적 기능이 상호간 털과 깃털을 다듬는 원래의 기능이 될 수도 있겠지만 계통발생 과정 에서 짝을 결합해 주는 과제가 기능변화를 통해 첨가되어 더 중요하 게 되었을 것으로 추측된다. 적극적으로 몸을 서로 접근시키는 배우 자간의 접촉행동 역시 유대를 촉진시킬 수 있다. 게다가 둥지짓기 동작들이나 둥지와 관련해서 일어나는 다른 행동양식들도 원래의 둥 지짓는기간 이외에도 일어난다면 같은 목적에 기여할 수 있다. 몇몇 오스트레일리아의 단풍새과 Estr ild i da e 새들의 수컷들은 둥지 장소를 찾고 있는 과정에서 특이한 몸 동작과 소리를 질러 자기 짝이 될 암 컷에게 둥지 장소를 알려 준다. 여기서 많은 종들이 둥지의식이 발 달되었다. 이것은 부분적으로 암수의 완전한 동조동작에서 나왔으며 둥지짓기 시작 전에만 일어나는 것이 아니고 번식기 이외에도 나타 난다 많은 행동양식들이 짝유대 기능과 관련해서 특히 자주 일어나고 있기 때문에, 만일 배우자들이 예를 들어 먹이를 찾으러 나갈 경우 와 같이 잠시 분리되었다가 다시 만나게 되면 이 행동을 짝유대와 관련된 인사행동이라 부른다 . 2.6 짝유대의 동인 구애행위로 짝이 결합하면 이 행동양식들은 성적인 동인이 될 수 있댜 그러나 짝유대 행위들이 번식기 이외에도 일어나고 있다면 순 수한 성적 동인으로 보기 어렵다. 그래서 보통 유대충동으로 표현되 는 자율동인 auto n omous mo ti v ati on 은 이 짝유대 행 동이 기 초가 될 수 있다. 이에 대한 실제 증거는 회색기러기의 트럼펫 소리와 열대산 새우의 〈짝앉기〉에서 찾아 볼 수 있댜 이 두 경우는 그 행동양식의 형태에 있어서 다른 행동양식들과는 분명히 다르고 공격적, 성적, 그
리고 양육 행동들과도 시간적으로 관계없이 나타날 뿐만 아니라 네 개의 고전적 사회충동(공격, 도피, 성, 양육)으로도 간주되지 않는다 . 이들의 배우자들을 서로 떼어 놓자마자 그들은 서로를 찾는데 이것 은 자기 갈망행동에 해당된다. 유대충동이 이 두 개의 완전히 다른 동물군에서 발견되었기 때문에 자율동인이 지속적인 결합을 하는 다 른 동물종의 짝유대에도 기초가 되고 있을 것으로 보인다. 3 양육행동 3.l 생물학적 의의 양육 혹은 새끼돌보기의 생물학적 목적은 자손의 생존 기회를 높 여주고 그렇게 하여 다음 세대에 자신의 유전자 몫을 남기거나 높이 고자하는 데 있다. 양육 혹은 새끼돌보는 데에 들이는 비용은 매우 다양할 수가 있 댜 포식자에 대한 심각한 위험에 처해 있는 종이나 아주 단기간 사 용할 수 있는 장소, 예를 들면 금방 말라버릴 물가에 의존하여 새끼 를 기르는 종들은 나타나게 될 손실을 메우기 위해 많은 수의 자손 울 생산해야만 한다. 그렇기 때문에 그들은 개개의 새끼들에게 양질 의 양육행동을 투자할 수가 없다. 이러한 경우를 r_ 전략이라고 하며 이때 해당 종을 기회주의자라고 한다 (P i anka 1970). r- 전략 종에 대 해 자손이 비교적 큰 생존 기회를 갖는 종들과 비교되는데 이런 종 들은 대개가 몸이 크고 방어적이며 지속적인 서식처에서 산다. 이들 온 비교적 적은 번식률과 높은 양육능률로 나타난다 (K- 전략). 요컨대 K_ 전략적인 종들은 자신의 유전자의 확산에 관련해서 어 미 _새끼 간의 갈등 상황이 일어난다 (T ri vers 1974). 한 마리의 새끼에
게 들이는 양육 능력은 동시에 다론 자손에 대한 양육 가능성을 줄 이게 된다 결국 이런 양육 능력은 새끼를 기르는 데 쓰이는 총비용 과 그 비용을 갖고 돌볼 수 있는 새끼 수가 한정되어 일어나거나 장 기적인 새끼양육이 새로운 자식을 낳을 수 있는 시점을 지연시킴으 로써 결정된다. 여기서 근친가설을 살펴보면 새끼와 어미 간의 〈관심갈등〉이 생겨 난다. 새끼는 가능하면 어미의 높은 양육비용을 가지려고 노력하는 반면 어미는 자신의 최대 번식 성공에 필요한 만큼만을 위해 새끼들 의 관심에서 이런 비용을 줄이려고 노력한다. 이때 이런 갈등 정도 에 있어서 계속 새끼들을 낳을 확률이 비교적 큰 젊은 어미와 어쩌 면 마지막 새끼들에게 더 투자할 수 있는 나이든 어미 사이에서 차 이가 생겨난다. 새끼의 나이도 이런 점을 고려한다면 하나의 역할을 하는 것이 틀림없다. 새끼가 나이들면 들수록 더 많이 스스로를 돌 볼 수 있으며, 그래서 어미 보살핌을 통해 얻게 되는 장점은 더 출 어든다. 한편 어미 쪽에서 보면, 예를 들면 새끼가 더 커지기 때문 에 반대로 비용이 더 커지게 된다. 사실 수많은 관찰들은 이런 생각을 확인시켜 주었다. 고양이나 개 에서 새끼양육의 기간을 3 단계로 나눌 수 있다. 첫단계는 새끼와의 접촉의 주도권이 무엇보다 어미에게서 나오고 두번째 단계는 그 주 도권이 양측에 있으며 마지막 단계인 이유기 직전, 즉 어미로부터 새끼를 떼놓으려 하는 시기에는 자기 새끼에 대해 공격적이고 회피 로 반응하는 어미에 대해 새끼는 어미가 주려고 하는 것보다 더 많 은 양육 비용을 가지려고 한다. 이런 〈이유 갈등 wean i n g con fli c t〉으 로 발달해가는 비슷한 행동들이 쥐, 영장류 그리고 여러 종류의 조 류에서 관찰된 바 있다 (Ma y nard-Sm it h 1977, Triv e rs 1974, Elwood 1983, Cohen-Salmon 등 1985) .
3,2 양육행동의 과제 양육행동 즉 새끼돌보기의 기본 과제는 새끼의 보호와 영양 섭취 에 있다 . 그 밖에도 기능들(예 , 정온동물의 경우 체온 유지, 장기적인 어미-새끼 결합을 하는 종의 경우 각종 정보 전달)이 더 첨가될 수 있 댜 위험으로 부터의 경계, 숨기기, 지키기 , 그리고 적극적인 방어 는 자식을 보호하는 데 기여를 한다. 땅바닥에 알을 낳는 많은 조류 들은 적을 유혹하는 방법으로 매우 눈에 띄게 방어를 한다 . 어미새 는 포식자에게 다리를 절거나 두 날개를 늘어뜨리고 걷거나 또 한쪽 날개를 부러진 것처럼 속여서 회피한다. 만일 이 가짜 부상자를 따 르던 포식자가 새끼나 알에서 충분히 멀리 떨어져 있게 되면, 이 어 미는 이 유인 자세를 중지하고 둥지 혹은 새끼에게 다시 날아간다. 많은 식육류와 설치류들은 위험시 자기 새끼들을 이빨로 조심스럽 게 물고 안전한 장소로 운반한다. 새끼들이 둥지 밖에 나와 있게 되 면 그들은 다시 이런 방법으로 옮겨다 놓는다. 이런 새끼운반행동은 전혀 피로해지지 않는 몇몇 안되는 행동들의 한 예로 제시된다는 점 에서 일반적인 홍미를 끌 수 있다. 새끼운반은 다른 동물군에서도 일어난다. 대부분의 유대류에서처럼 특별히 발달된 몸의 주머니에 의해 운반되고 혹은 많은 종류의 거미, 전갈, 뿔논병아리 그리고 수 많은 포유류(예, 코알라, 개미핥기, 나무늘보, 박쥐, 영장류)에서처럼 어미(부모)의 몸위에서 운반된다 . 이때 포유류 새끼들은 손과 발로 어미몸에 꽉 달라붙는다. 이것은 손가락과 발가락의 굴곡반사 작용 으로 일어나고 있다. 박쥐와 영장류들은 여기에다 입술로 어미의 젖 꼭지롤 꽉 무는 것이 첨가된다 . 특별히 인상적인 방법에 의해서 어미와의 신체접촉을 하는 동물은 키클리드과의 물고기 Til ap i a 로 어미는 알을 입속에 넣어 보호하는데 이때 알들은 입속에서 지속적인 수류로 산소가 풍부한 물이 공급된
다 부화 후에도 새끼들은 얼마 동안 어미의 입속에서 머무는데, 탐 색을 위해 잠시 나왔다가 위험이 닥치면 다시 어미의 입속으로 들어 간댜 이런 행동을 하는 종들은 키클리드과의 물고기에서 두드러지게 나타나지만 다른 물고기류에서도 간혹 나타난다 (S i lur ifo rmes, Cy pri - nodonto i d e i, Anabanto i d e i) (Pe t ers 와 Berns 1982) . 새끼의 영양 섭취에는 능동적인 먹이먹이기와 부모가 새끼를 적당 한 장소로 안내하여 먹이를 먹이는 유도가 있다. 이러한 먹이 유도 는 무엇보다 몇몇 조숙성 조류에서 보편화되어 있다. 능동적 먹이먹 이기는 부모가 먹이를 가져와 새끼에게 그대로 먹여주는 것이 있는 가 하면 새끼의 영양 공급을 위해 어미몸에서 물질을 생산하는-것이 있다 이런 것에는 포유류의 젖, 비둘기의 소낭유, 홍학의 전위액, 디 스커스 (Sy mp h y sodon dis c us, 키클리드과의 물고기)의 피부 점액, 그리고 꿀벌(일벌)의 선분바물이 속한다. 항온동물(조류와 포유류)은 부모로부터 알이나 새끼에게 열의 전달 이 하나의 중요한 양육 행동으로 표현된다. 이 행동은 새끼가 스스 로 체온 조절 능력을 갖출 때까지 행해진다. 조류의 경우 새끼의 열 공급이 모래목욕으로 나타난다. 어류의 많은 경우 알의 산소공급이 부모 혹은 부모 중 한쪽의 지느러미 움직임(부채질하기)을 통해 촉진 된다. 역시 새끼를 화장해 주는 것도 양육행동에 속한다. 즉 기생충 제거나 둥지 청결 유지가 그 예가 된다. 이러한 직접적인 과제 외에도 양육행동은 새끼에게 정보 전달 면 에서 중요한 의미가 될 수 있다. 중요한 예가 새들의 노랫소리가 될 수 있다. 노랫소리는 대개가 학습이 되어야만 부를 수 있고 이런 학 습과정은 매우 이른 시기, 다시 말하면 종종 어미로부터 독립되기 전이나, 어떤 종은 이미 둥지에 있을 때 일어날 수 있다. 그러므로 부친의 노래가 새끼들에게 하나의 모델 역할을 하고 있다는 사실이 밝혀졌다. 이런 식으로 노래의 특정 형태가 세대를 거듭해서 전해지
게 된다. 먹이 인식, 먹이 취득기술 혹은 표적지 인식 그리고 다른 위험들을 여러 방법으로 양친을 통해 정보를 취할 수 있다. 여기서 고등한 포유류들은 모방이 중요한 역 할을 하고 있다. 결론적으로 양육행동은 두 세대간의 확고한 유대와 전수를 형성해 가는 중요한 수단이 된다. 이런 기능은 계통발생 과정에서 확실히 이차적으로 생겨난 것이다. 그렇지만 오늘날 몇몇 포유류의 경우에 확실히 이 기능이 매우 중요해져서 새끼들은 영양학적 독립기가 넘 어서도 여전히 얼마간은 부모 곁에 머물러 있게 된다. 예를 들어 세 가락나무늘보의 새끼는 단지 1 개월 동안 젖을 먹지만 계속 5 개월을 어미 등에 업혀다닌다. 이 기간에 새끼는 먹이 식물, 휴식장소 그리 고 다른 환경 조건들에 대한 경험을 수집한다. 부모와 새끼의 긴 동 거 생활은 몇몇 다른 포유류나 많은 장수하는 조류에서 잘 알려져 있는데 이런 생활을 통해 마찬가지로 정보 취득에 기여할 수 있음을 알수있다. 3.3 가족 형성 양육행동은 거의 모든 동물문(門)에서 나타나고 있다. 가장 하등 한 다세포 동물인 해면동물이나 강장동물 스스로 간단한 형태의 양 육행동이 알려져 있다. 물론 하등한 무척추동물 내에서 양육행동은 하나의 예외적인 현상으로 나타난다. 두 개의 동물문(門), 죽 절지 동물(게, 거미, 곤충)과 척추동물에서 양육행동은 일반적으로 퍼져 있고 그 가운데 곤충강(綱)의 흰개미류나 척추동물문의 조류와 포유 류에서는 예의없이 모든 종에서 나타나 있다. 물론 이 양육행동에 성체의 참여는 다양하다. 양친 중 하나만 참여할 수 있고 양친 모두 혹은 가족을 구성하는 성체 모두가 함께 참여할 수가 있다. 따라서 가족형성도 다양하다. 가족이란 말은 행동학에서 부모와 새끼가 함
께 살때에 순수한 의미로 쓰인다 . 부모가 새끼돌보기 참여에 따라 양친가족, 모친가족 그리고 부친가족으로 구분하며 이러한 구분에서 좀더 세분할 수 있다. 암수가 둥지에 동시에 혹은 교대로 같은 일을 하여 새끼를 돌보는 양친가족은 많은 조류에서 있지만 어류(예, 키클리드과의 많은 물고기) 와 몇몇 포유류(예, 긴팔원숭이 , 티티원숭이, 그리고 개과의 식육류)에서 도 존재하고 있다 . 양친 사이에 일의 분담을 맡는 가족을 부모가족 이라고도 한다. 이런 가족은 많은 맹금류와 코뿔새 Bucero ti dae 가 대 표적이다. 이들은 수컷이 먹이를 사냥해 운반해 오면 암컷이 그것을 새끼에게 공급한다. 만일 일의 분담이 커서 원래의 새끼돌보기를 암 컷만이 하고 수컷은 단지 터를 방어하거나 자기 가족을 지키는 경우 가 있는데 이것을 수컷-어미 가족이라고 한다. 이런 가족의 예는 많은 명금류와 포유류에서 잘 알려져 있다 . 이것은 순수한 모친가족 으로의 과도기적 단계에 해당된다. 모친가족은 대부분 포유류의 전형적인 가족구조다. 이들은 어미의 새끼돌보는 역할이 모유의 공급을 통해 의무적이다 . 그러나 이것은 조류(예, 닭, 벌새, 극락조), 어류, 거미, 그리고 전갈에서도 나타난 댜 부친가족의 경우 단지 수컷만이 새끼를 돌보는데 이에 대한 예 는 큰가시고기, 해마, 그리고 조류 가운데 도요류 Phalaro p od i dae, 세 가락메추라기, 에뮤, 그리고 타조가 있다 . 곤충의 사회도 가족 형태롤 갖추고 있는데 이 중에 말벌, 뒤영 벌, 꿀벌, 그리고 개미는 모친가족이고 수컷이 혼인비행 후에도 여 왕과 함께 사는 흰개미는 양친가족에 속한다. 몇몇 동물종의 경우 부모 이외에도 다른 가족 구성원들이 양육행동에 참여하기도 한다. 이 러 한 집 단가족은 뒤 에 논하기 로 하겠다 (Pe t ers 1948) .
3, 4 %낵행동의 행동 적응 다른 사회 행동과 같이 이 양육행동은 동료간의 의사소통에 필요 한 수단을 요구한다 . 이로써 다양하고 매우 고도로 발달된 행동양식 들이 생겨나게 되었다. 이들은 두 가지 주요한 목적과 일치하고 있 댜 종내의 공격성을 누그러뜨리는 것과 먹이 인도의 개시 및 실행 에 있다. 공격적 동인은 부모간에는 물론 부모와 새끼 간에서도 일어날 수 있다 부모라는 쌍의 관계에서 특별히 위험이 생겨날 수 있으며 이 때 새끼돌보기 행위들은 배우자간의 육체적 접촉이 요구된다. 적응 의 관점에서 많은 종들은 구애와 짝유대 행동의 기능과 같은 공격억 제 행동양식을 취한다. 우선 많은 조류의 교대의식이 이에 속한다. 이것은 둥지에 돌아온 새와 포란중인 배우자 간의 포란 교대시 일어 난다 어떤 종에서는 이 교대시 둥지 재료를 부리에 문다 . 구애 중 에 둥지 상징물 이용에 비슷한 것을 보여 주는 이런 행동의 공격억 제 효과는 바로 날지 못하는 갈라파고스의 가마우지에서 아주 분명 히 나타난다. 둥지로 돌아오는 새는 둥지 교대를 위해 불가사리 혹 은 조류( 羅類 )를 가져오는데 이것은 분명히 배우자로부터 공격적 움 직임을 줄이는 역할을 한다 . 만일 돌아오는 이 새(귀소자)에게서 둥 지 상징물을 빼앗아 버리면, 공격은 이 귀소자를 향해 즉각적으로 일어나며, 곧바로 아주 격렬한 싸움으로 변해 결국 이 귀소자는 도 망가고 만다. 새끼들도 어미에게 공격적 성향을 일으킬 수 있다. 이 공격 성향 은 두 가지 방법에 의해 억제될 수 있다. 새끼들은 복종적 효과가 있는 행동양식을 소유하고 있거나 공격신호의 발사를 어느 정도 피 할 수 있다. 하나의 복종적 기능은 부모의 먹이주기 반응을 일으키 는 직접적인 효과 이외에 새끼의 구결행동에 내재되어 있을 수 있
댜 이 구걸행동은 이런 성질 때문에 공격성향을 억제시키는 다른 사회적 행동 범위 가운데 2 차적 행동으로 발달하게 되었다. 공격신호의 회피는 새끼들이 특별한 깃털(털) 혹은 얼굴 모양을 갖고 있으므로 가능할 수 있다. 이것은 이중의 기능을 갖는다. 하나 는 일반적으로 매우 눈에 띄지 않으며 좀더 나은 은폐로써 극도의 보호를 제공한다 . 또 다른 하나는 새끼들의 깃털이나 얼굴 모양이 성체의 공격 대상 표적으로써 완전히 결여되어 나타나기 때문에 거 의 공격성을 유발시키지 않는다. 양육행동이나 가족 행동 범주로부터 새끼 인식은 새끼들의 특별한 표시를 통해 무엇보다 집단생활을 하는 동물종에서 의미가 있다. 여 기서 집단 내 새끼들은 특별한 지위를 차지한다. 그들은 안하무인격 으로 자유분망함을 누리며 또 집단 서열에 아직 소속되지 않아 오히 려 그들에게 좀더 큰 학습기회가 열려져 있다. 그러나 그들이 모든 집단구성원으로부터 새끼로써 인정받는다는 것이 중요하다. 이에 대 해 혹고니에 대한 실험적인 증거가 있다. 이 종의 경우 대개의 새끼 들이 암갈색의 깃털을 갖고 있는데 반해 일부가 성체처럼 흰색의 체 색을 갖고 있다. 갈색 모델과 흰색 모델의 모형 실험에서 흰색 모델 온 갈색 모델보다 터 소유 고니쌍으로부터 훨씬 자주 위협과 공격을 받았다. 이것은 자연 상태의 관찰에 의하면 흰색의 새끼에 대한 자 기 부모의 공격성이 갈색의 새끼에 대한 공격성보다 높았다. 반대로 부모가 새끼에게 공격성향을 불러일으킬 수도 있다. 이런 경우에 어미들은 공격을 억제시키는 행동을 사용한다. 예를 들면 해 오라기의 경우 어미가 먹이를 먹이기 위해 둥지에 도착하면 우선 깊 이 머리를 숙이고 세 개의 흰 머리깃을 세운다. 물론 그것은 평상시 에는 접혀져 있다. 이 깃은 녹청색의 금속 광택이 나는 검은색 머리 바탕과 하나의 분명한 대조를 이루며 그것은 새끼에게 매우 눈에 띄 는 신호로 전달된다(그림 9-23). 어미는 새끼에게 이 깃을 보인 다음
 •! , '.1 ;,· ,'. .
•! , '.1 ;,· ,'. .
에야 둥지 가까이로 다가갈 수 있다. 만일 이 어미에게 머리숙이는 동작을 못하도록 방해를 하면 그는 자기 새끼들에게 공격을 받는다. 종내(종 특 이적) 공격 이외에 포식성동물들의 경우 그들의 새끼들을 먹이로 착각하게 될 위험성이 생겨난다. 이런 관계에서도 어떤 특수 한 적응이 발달해 왔다. 그것은 몇몇 거미류에서 알과 새끼를 돌보 는 시간에는 전혀 먹이를 취하지 않는다든지, 또 많은 입속에 새끼 를 기르는(구강 양육) 물고기 Til ap i a 도 비슷한 억제 현상이 생겨났다. 새끼에게 먹이주기는 참여 배우자들간의 고도의 협동심과 시간적 동조성을 요구하며, 이것은 교미에 이르게 하는 일련의 행동과도 비 슷하다 이처럼 먹이주기 행동에 있어서도 행동 연쇄와 눈에 띄는 해발인의 발달이 다양해졌다. 이것들은 부리와 부리로 먹이를 건네 주는 조류에서 특히 잘 발달되어 있다 . 여기서 해발인에 속하는 것 은 종종 광택이 나는 새끼새의 입안(그림 9-24) 과 구걸음 그리고 구 걸행동으로 표현할 수 있는 새끼의 모든 동작들을 말한다. 이것들의 궁극적인 목적은 어미의 먹이주기 반응을 일으키는 데 있다. 반대로 역시 부모들도 형태적인 특징이나 행동 양식으로 새끼로 하여금 구 걸하도록 하는 신호를 보낸다. 이것은 새끼의 미경험으로 인해 특히 생득적 해발기구의 분석에 적합하기 때문에 이런 연구는 행동학 발
 ;.. 響二:리:〔:::반점
;.. 響二:리:〔:::반점
달 초기에 이루어졌다 . 가장 잘 알려진 예가 구걸 해발인으로 작용 하는 재갈매기의 부리의 붉은 반점이다 . 4 집단행동 많은 동물종들은 쌍의 개념을 넘어서 성적으로 성숙한 동종의 개 체들이 일정 기간 혹은 지속적으로 집단을 이루며 산다. 이런 사회 집단의 기능과 연령별 그리고 성별 조성은 동물의 종마다 매우 다양 하댜 그렇지만 몇몇 일반적인 규칙성이 있음을 알 수 있다 (Kummer 1975, Remane 1976) .
4. l 사회적 유대의 생물학적 기능 집단 형성이 많은 종에서 가장 다양한 분류학적 위치를 나타내고 있는 사실온 이 집단형성에 많은 생물학적 의미가 생겨날 수 있음을 시사해 주고 있다. 집단 유대로 얻는 이점 및 계통발생학적으로 사 희 형성의 발달을 촉진시켜 온 선택압의 이점들은 매우 다양하다. 1) 적의 회피 집단 유대는 다음 세 가지 방법으로 각 동물들에게 포식자로부터 더 나은 보호를 제공해 준다 . ®상호경계 가장 널리 퍼져 실행되고 있는 것이 이 상호경계다. 집단은 항상 경계를 하는데 그것은 반응이 가장 빠른 집단구성원에 의해 행해진 다 . 집단의 보호에 있어서 각 개체들은 다른 일들에 전념하기 위해 잠시 동안 방심할 처지에 놓여 있을 수 있다 . 이 상호경계에는 여러 감각 양상이 이용된다. 음성및 화학적 신호가 특히 자주 사용된다. 예를 들면 새들의 경계음과 물고기의 경고 및 놀램물질이 있다. 이 것 가운데 경계 신호는 종의 한계를 넘어서 상호경계의 의미 이상의 효과가 있을 수 있다. 일반적으로 가장 잘 알려진 예는 여러 명금류 의 유사한 경계음에서 알 수 있다. ®혼란효과 많은 종들(예, 새, 물고기)은 포식자가 나타나자마자 떼를 지어 연 합한다. 이것은 포식자에게 분명히 한마리의 먹이동물에 대한 조준 의 어려움을 높여서 아주 배고프지 않은 포식자가 가능하면 방향 전 환을 하게 하여 그 집단의 각 구성원들에게 생존의 기회를 높여 준
다. 반대로 먹이동물의 이런 행동에 포식자들도 적응해 왔다. 포식 자들은 집단의 각 개체들을 밀어젖히려고 한다. 실험에 의하면 큰가 시고기는 무리에서 가장 멀리 떨어져 있는 물벼룩을 우선적으로 잡 아먹는다 그리고 이 큰가시고기뿐만 아니라 강꼬치고기에서도 외견 상 먹이동물들이 밀집되어 뭉쳐 있는 경우 전형적인 갈등행동이 관 찰된다. ®협동방어 무장하지 않은 많은 동물들 스스로 방어를 할 수 있다. 그러므로 많은 종에서 집단 구성원들이 방어의 목적으로 능동적인 유대를 이 룬다 물소, 영양, 그리고 들소 들은 방어형 원형대열을 형성하는데 이때 성체들의 뿔은 밖으로 향하게 하고 새끼들을 보호하기 위해 대 열 한쪽으로 가둔다(그림 9-25). 개코원숭이의 경우 다자란 수컷들 이 포식자와 자기 자신의 집단 사이에 서서 공동의 위협과 위장 공 격으로 자칼, 들개 혹은 표범 들을 물리칠 수 있다. 꿀벌들은 자기 들의 집에 쳐들어온 말벌들을 떼를 지어 방어한다. 2) 먹이 획득 반대로 사회적 결합 역시 먹이 획득 즉 포식자에게 장점이 될 수 있는데 그것은 다음의 두 가지 관점에서 나타난다. ®협동사냥 많은 동물들은 무리를 지어 사냥을 하며 그렇게 하여 보다 높은 사냥 성공을 가져온다. 사자들은 무리를 지어 사냥을 할 때 혼자서 사냥할 때보다 평균 두배의 성공을 거둔다. 아프리카 들개의 경우 각각의 무리 구성원들은 그 구성원으로부터 가장 가까운 가젤영양 뒤를 쫓아 사냥을 한다. 이때 거리가 좁혀지지 않게 되면 그 구성원
 _--- - --
_--- - --
들은 다른 가젤영양의 사냥 기회를 엿보아 사냥을 성공시킨다. 물고 기를 잡아 먹는 사다새(그림 9-26), 비오리 그리고 뱀가마우지 Anh i n gi dae 는 물위에서 먹잇감이 되는 물고기들을 수영을 해서 원형 으로 둘러싸거나 물이 얕은 곳으로 그 고기떼를 몰아서 사냥을 한 다. 범고래, 바다사자 혹은 바다코끼리들도 무리를 지어 에워싸는 방식으로사냥을 한다.
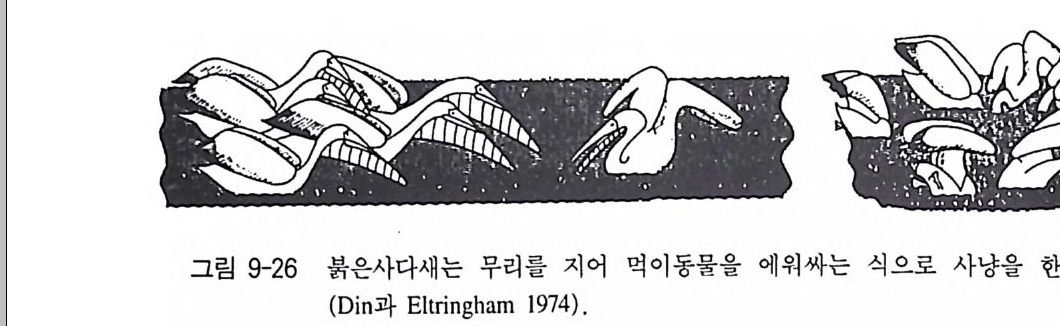 그림 9-26 붉은사다새는 무리 를 지어 먹이동물을 에워싸는 식으로 사냥을 한다
그림 9-26 붉은사다새는 무리 를 지어 먹이동물을 에워싸는 식으로 사냥을 한다
®정보교환 비육식성이나 개별적으로 먹이 사냥을 하는 종에 있어서도 효율적 인 이점은 상호간의 정보 교환에 있다. 많은 조류의 공동 야간 잠자 리는 보호를 상승시킬 뿐만 아니라 실제 정보센터 역할을 할 수 있 다는 사실이 잘 밝혀져 있다 . 그 전날 풍부한 먹이를 노획했던 동물 들은 그 다음날 아침에 목표지점에 곧바로 날아가서 먹이를 찾는 데 성공적이지 못한 개체나 그로 인해 망설이는 개체들로 하여금 먹이 를 쉽게 찾게 한다. 영장류의 무리에서도 정보 교환은 중요한 역할 울 하는데, 무엇보다 물먹는 장소나 먹이 장소――대략 고갈되는 시기에-가 부족하여 그런 장소가 멀리 떨어져 있게 되면 그것을 발견하기 위한 각 개체들의 많은 경험이 요구된다. 3) 번식 번식에 대해서도 사회적 결합은 생물학적 이점을 가져올 수 있다. 이 잇점은 역시 적 회피 지역에서 나타난다. 그래서 집단 번식 조류 들은 각 번식 쌍이 다소 자기들의 둥지 자리만을 방어할지라도 포식 자를 공격하여 몰아내는 데 단독 번식 조류보다 더 효과적이다. 또 다른 이점들은, 공동 구애를 하는 수컷들에 의한 암컷들의 유인에 있어서 번식 초기에 쌍의 상호자극과 동조성 그리고 공동의 양육행 동에서도 나타난다. 생쥐의 경우 어린 새끼들은 여러 마리의 암컷들
이 그 새끼들을 협동해서 기 를 때 더 빨리 성장한다는 사실이 잘 알 려져 있다 . 무엇보다 사회성 곤충, 많은 포유류 그리고 조류의 경우 유충 및 새끼들은 자기 어미 혹은 자기 부모한테만 양육되는 것이 아니고 다른 사회계급, 다른 집단 구성원 혹은 다른 가족 구성원에 의해서도 보살핌을 받는다(조력자). 비록 조력자의 정확한 의미가 아 직도 모든 경우에 완전히 설명되지는 못하지만 확실히 이 조력자가 번식 성공의 확대에 기여할 수 있다는 사실은 이미 증명되었다. 즉 정확한 연구결과에 의하면 조력자가 있는 가족에서 성공적으로 길러 진 새끼의 수는 단독 쌍에서 길러진 새끼 수보다 거의 항상 많았다 는것이다 . 4) 집짓기 집단서식 동물들은 개별적으로 할 수 없는 협동 건축 능력을 갖는 댜 사회성 곤충의 집, 비버의 댐, 남미산 푸른앵무새 My iops it ta monachus 와 아프리 카산 집 단직 조새 Phil e ta i r u s so ci us 의 거 대 한 공동 둥지들이 이에 속한다. 경관을 변화시키는 비버의 댐은 일정한 수면 유지를 가능하게 해준다. 이렇게 해서 집의 입구는 물밑에서 항상 잘 보호된다(그림 9-27). 집단직조새들은 번식기 이외에도 이 거대 한 둥지에서 잠을 자는데 둥지 안의 온도는 밖의 온도보다 높은 23°C 까지 유지된다. 이런식으로 이 새들은 그들의 분포지역에서 매우 추 운겨울 밤을 이겨낸다. 5) 분업 사회적 협동에서 각각의 일들은 다양한 방법으로 사회구성원들에 게 분담되어질 수 있다. 이로써 동물 각자에게 개체의 생존을 위한 모든 일을 해야하므로 그 종에서 요구되는 행동들이 발달해·왔다. 사회성 곤충에서 이 분업이 특히 잘 발달되어 있다. 여기서 동물들
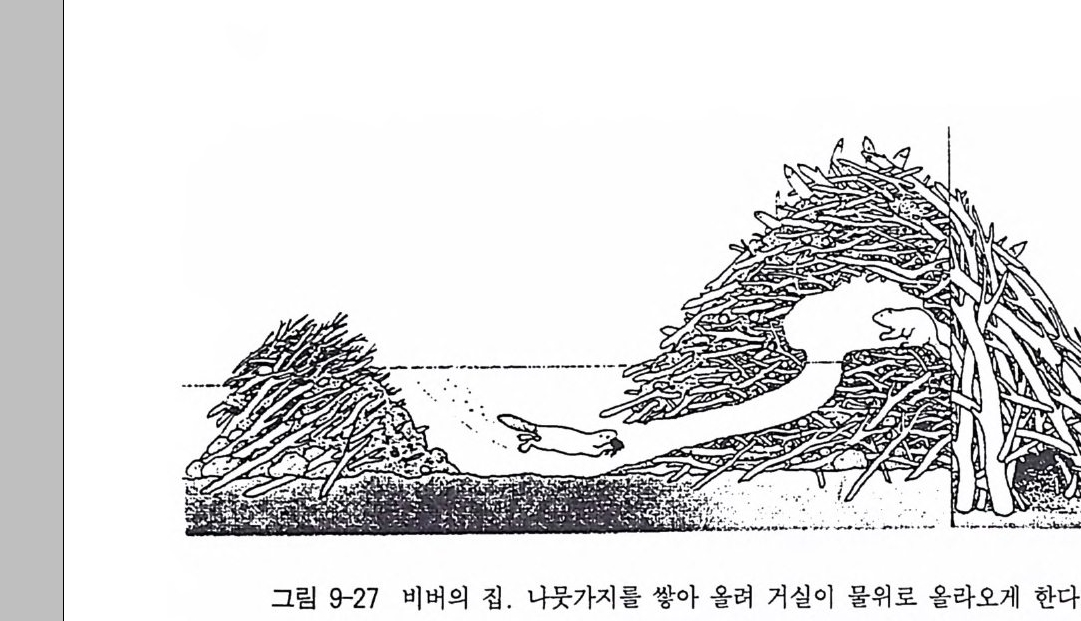 그립 9-27 비버의 집 . 나뭇가지 를 쌓 아 올려 거실이 물위로 올 라오게 한다.
그립 9-27 비버의 집 . 나뭇가지 를 쌓 아 올려 거실이 물위로 올 라오게 한다.
의 일은 여러 계급(카스트)간 매우 차이가 날 수 있는 몸의 구조, 성 별, 혹은 연령에 따라 다르다 꿀벌에 있는 연령의 경우, 같은 개체 가 그의 생애 동안 여러 역할을 맡는다. 척추동물에서는 이 분업이 지도자나 감시자의 형태로 주로 집단서식 포유류에서 나타난다. 새 끼를 돌보는 많은 척추동물의 경우, 암수간에 일종의 분업이 존재하 는데, 예를 들면 새끼를 지키는 역할과 먹이 사냥이나 먹이 운반과 먹이 분배 역할이 있다. 특히 여기서 중요한 기능 역할은 사회적 생활양식이 집단 구성원 각자에게 가져올 수 있는 여러 이점들이 된다 . 집단은 그 집단의 개 체가 개별적으로 일을 하는 것보다 항상 더 많은 일을 할 수 있다 . 그래서 많은 동물들은 의무적 사회성이 되었고, 사회성 곤충에서처 럼 개별적으로는 생존의 기회가 매우 희박하거나 거의 없을 정도다. 종종 이미 열거한 집단의 여러 기능들이 동시에 충족되기도 하는 데, 원래 집단의 형성이 어떤 선택압에 의해 발달해 왔는가 그리고 어떤 생물학적 이점들이 이차적으로 생겨났는지 결정한다는 것은 매 우어렵다.
4.2 사회 구조 종내의 경쟁은 사회 생활의 이점들이 단점으로도 작용할 수 있다. 예를 들면 작은 면적에 개체 수가 너무 많을 경우 먹이원이 부족하 여 먹이 경쟁이 생길 수 있다. 은폐의 요구 역시 개체의 운집으로 숨을 곳이 없어 경쟁이 생겨난다. 그러므로 각 종들의 사회 구조는 각기 〈타협 모색〉, 즉 계통발생 과정에서 사회적 유대의 요구를 방 해하는 선택압과 연합하여 발달해 왔다. 그것은 각 종에서 어떤 한 계 내에 특징적으로 나타나지만, 연령, 성별, 연주기 그리고 지리적 차이를 보여줄 수 있다. 무엇보다 조류와 포유류에 대한 오랜 야외 연구를 통해 특정 생활 조건에 대한 사회 구조는 매우 다양하고 섬 세한 적응과 정착된 환경 조건에 고도의 융통성을 갖고 있음이 밝혀 졌다 (Crook 1970, Emlen 와 Or ing 1977, Schaller 와 Crawshaw 1981). 이 연 구의 결과는 조심스럽게 다음과 같이 일반화될 수 있다. 공간적으로 그리고 연주기적으로 비교적 균일하게 분포된 먹이 제공을 이용할 줄 아는 원시림에 사는 종들은 종종 쌍으로 혹은 작은 무리를 지어 사는 반면, 매우 많은 개체로 구성된 무리들은 먹이가 시간과 공간 을 초월해 많은 양으로 존재하는 넓고 건조한 생활 공간에서 더 많 이 생긴다. 그러나 이런 규칙으로부터 많은 예의가 있을 수 있으며 한 종의 사회 구성은 그 종이 살고 있는 환경 조건들이 정확히 분석 될 때만이 실제로 이해될 수 있다. 4.3 이타주의 1) 발생 동물들의 사회 형성에 대개 〈서로 함께〉만이 존재한다. 실제 〈서 로 위하기〉는 아주 드문 편이다. 대개의 종에서 집단 구성원들은 개
별적으로 하고 싶은 것을 하며 이미 열거한 결합의 생물학적 이점들 은 경우에 따라서 여러 동료들이 동시에 같이 일을 함으로써 부작용 이 될 수 있다 . 물고기를 잡는 사다새, 구애를 하는 홍학 혹은 잠자 리에서 먹이원으로 날아가는 새는 집단에서 혼자 혹은 쌍으로 행동 하는 것과 다를 바 없으나, 그들의 행동의 결과는 집단으로 일할 때 자신과 자기 동료들에게 더 큰 것이다. 진정한 〈이타주의 al trui sm 〉는 한 동물 스스로가 자기 자신의 생존 기회를 줄이고서 동료를 도와주는 행동으로 일반적으로 새끼돌보기 범주의 행동(양육행동)에서 나타난다. 성체간 즉 사회성 집단의 구성 원간에는 이 이타행동이 극히 드물게 나타난다. 그렇지만 최근 관찰 과 실험에 의해 그런 행동에 대한 보고된 실례가 많아졌다. 이타행 동의 발생은 무엇보다 다음 세 가지 행동 범주에서 나타난다. 상호 경계 및 방어, 새끼돌보기의 조력, 상호 영양교환이 있다 . 협동방어의 경우, 개코원숭이에서처럼 그 무리의 공격자와 그 무 리 나머지 구성원간에 개코원숭이들이 서 있을 때 이타주의적 요소 가 항상 관여한다. 많은 사회성 곤충에서 그 사회 집단의 방어는 주 로 혹은 오로지 특정 개체군의 과제가 된다 . 이런 개체군은 형태적 으로는 다른 카스트와 차이가 있으면서 오히려 〈병정〉으로 표현된 댜 상해자나 병자에 대한 행동에서도 이타주의 요소들이 나타날 수 있지만, 그것이 확실히 일부의 경우에 나타나 있다. 돌고래는 상처 난 동료를 호흡하도록 물표면으로 밀어 주고(그림 9 一 28), 향고래와 바다코끼리는 상해자를 중심으로 때를 짓고, 코끼리는 일어날 수 있 도록 도와준다. 몽구스는 아픈 동료를 따뜻하게 해주며 몸을 닦아주 고 먹이에 접근할 수 있는 유리한 곳으로 인도한다. 그밖에도 많은 비슷한 관찰들이 검증 확인되었다. 수많은 포유류, 조류 그리고 어류에서 집단 혹은 가족의 구성원들 이 새끼 를 양육할 때 어미나 그들의 부모를 도와준다. (Rey er 1980,
 •:. 晃· · 《` ... \
•:. 晃· · 《` ... \
Taborsk y와 Lim berge r 1981). 아주 다양한 근연 집단을 이루는 쇠물닭 과 그 밖의 수많은 열대 아열대 새들의 경우, 어미(부모)의 새끼양육 을 도와주는 개체는 지난해에 태어난 조력자의 역할을 하는 젊은 새 들이다 일부 영장류에서 새끼를 가지지 않은 무리 구성원들은 일시 적으로 자기 어미의 새끼들을 넘겨받을 수 있으며 사회성 곤충들의 양육행동은 어미 혹은 부모 대신 완전히 나이 먹은 자매가 맡는다. 성체간 상호 영양교환도 이타 행동의 범주에 넣을 수 있다. 이것 은 꿀벌에서 실험적으로 증명되었다. 방사능 물질을 먹이에 표시하 여 벌집 내에서 먹이 분배가 필요에 따라 일어나고 있다는 사실이 확인되었다 (W i c k.l er 1967). 말하자면 가진 자는 필요로 하는 자에게 주는 것이다. 분업으로 다른 일을 하기 때문에 먹이룰 얻을 수 없거 나 충분히 먹이룰 먹을 수 없는 벌들 역시 이런 먹이 전달을 통해 먹이를 공급받는다 . 비슷한 먹이 분배체제가 아프리카의 하이에나에 서 잘 알려져 있다. 많은 고등 영장류 즉 긴팔원숭이와 침팬지에 의 해 영양교환이 관찰된 바 있다 . 마찬가지로 최근에 흡혈박쥐에서도 이 이타행동이 관찰되었다 (Wi lkins on 1990) . 흡혈박쥐 는 예 리 한 이 빨로 가축에 게 3mm 정 도의 작은 상처를 입힌 다음 피가 나오는 곳에 혀를 내고 피를 먹는다. 이 흡혈박쥐들은 밤의 어두운 시간에 활동적이며 낮 동안은 속이 빈 나무 구멍 또는 동굴에서 산다. 때때로 사냥 경험이 미숙한 젊은 박 쥐들은 피를 먹지 못하고 되돌아 오는데 이 박쥐들에게 다른 동료 박쥐가 자신이 먹은 피를 토해 내어 종종 먹인다. 또한 그들은 친족 이 아닌 박쥐에게도 피를 먹이는데 특히 과거에 자신에게 피를 먹인 박쥐에게 피를 토해 주는 것으로 관찰되었다. 2) 생물학적 의의 비교_계통발생학적 행동 고찰에 의한 이타행동의 이행와 의미는
초기까지만 해도 밝혀지지 않았다. 이 행동이 생겨난 것은 이기적 행동만을 위해 있는 자연선택의 결과와 확실히 일치하고 있지 않기 때문에 혼란을 일으킬 수 있다. 최근 사회생물학의 등장과 고찰이 이러한 이해의 결점을 보완할 수 있게 되었다. 이 새로운 인식은 다 음 두 가지 면에서 특징지어진다. 하나는 많은 이타행동 양식들이 수혜자뿐만 아니라 공여자에게도 이점을 가져올 수 있고, 또 하나는 집단 생물학적 고찰로 순수한 이타행동은 특정한 근연관계 아래 자 연선택에 의해 생겨날 수 있다는 사실을 인식하게 되었다. ®이타주의와 이기주의 흔히 언급된 일련의 이타행동 양식들을 좀 자세히 살펴보면, 원래 보다 분명히 덜 이타적이다. 그것은 다음 세 가지 실례를 통해 알 수 있다. 가장 간단한 이타행동의 형태 가운데 하나로서 경계음을 이용한 상호경계나 그 밖의 신호는 확실히 집단 구성원들에게 위험 에 대한 정보를 전달하는 것으로 받아들여질 수 있다 . 그러나 경고 자 역시 이점을 누리고 있음을 생각해 보아야 할 것이다. 예를 들어 상호경계의 결과로 포식자의 성공률이 줄어든 이점은 그 포식자를 그 지역에서 떠나게 할 수 있다. 비슷한 예로 사회적 털(혹은 깃털) 다듬기(화장)도 기생충이 제거되는 수혜자에게 기여를 한다. 이것은 공여자에게 이점이 배제되는 것은 아니다. 사회적 화장이 관찰되는 대부분의 종들은 접촉성 동물들이다. 그들은 공동의 둥지나 잠자리 에서 서로를 화장해 주면서 밤을 보낸다. 여기서 항상 기생충의 전 달 가능성 때문에 배우자의 기생충 제거는 자신의 일처럼 중요하다. 한편 이미 언급된 새끼돌보기 조력자에게도 여러 가지 이점이 생 겨날 수 있다. 무엇보다 그들은 자기 스스로의 새끼를 갖기 전에 새 끼돌보는 데 따른 제반 경험을 하게 된다. 부모의 텃세권에 계속 머 문다는 것 역시 아마도 긍정적인 일로 나타나며 그 밖의 이점들도
최소한 생겨날 수 있다. ®이타주의와 혈연도 많은 경우에 이타적 요소가 완전히 알려지지 않았지만, 그런 요소 들이 동물 행동에 실제 존재하고 있다는 사실은 부정하기 어렵다 . 그러므로 생물들의 모든 형질들처럼 역시 행동에 있어서도 선택압이 작용함이 틀림없다. 결국 이 선택압의 영향 아래 관련 행동 능력둘 이 발전해 온 것이다. 그렇다면 이타적 행동이 생물학적 기능을 만 족시킬 수 있으며 어떤 행동 범위에서 그것이 나타날 수 있는지를 질문해 볼 필요가 있다. 이러한 질문은 분명히 긍정적인 대답이 나 올 수 있다 그러므로 그 대답은 수혜자가 공여자와 근친일 때 이타 적 행동은 항상 이점이 된다는 사실을 엄밀히 규정하고 있다. 즉 수 혜자가 새끼를 낳으면 근친인 공여자의 유전자가 그 다음 세대에 전 달되기 때문에 항상 이 행동은 공여자와 수혜자에게 이점이 된다는 것이댜 사회생물학적 이론에 따르면, 두 개체간의 혈연도가 높으면 높을 수록 생물학적 이점은 높다 . 그러므로 이타적 행동은 특히 양육행동 의 기능 범위에서 잘 나타나 있다는 사실은 당연한 것이다 . 여기서 새끼들의 생존 기회를 높이는 각각의 행동양식(먹이 전달, 경계, 방어 등등)에 대한 번식 성공률은 자신의 유전자를 전달하여 그것을 극대 화한다는 의미애서 가장 큰 것이다 . 그러나 고도로 발달한 사회 집 단과 사회성 곤충 내에서도 구성원들은 가족으로부터 이러한 사회 구조가 생겨났기 때문에 일반적으로 아주 가까운 혈연으로 이타행동 이 생물학적 이점을 가져올 수 있다. 이것은 막시류(개미, 벌 따위)의 사회에서 행해지고 있는데 이들 곤충 사회에서 일꾼들은 스스로 자 신의 유전자를 더 이상 전달할 수 없는 상황에 있다. 사회 전체가 자매들로 구성되어 있기 때문에 그들은 자신들의 유충의 양육을 통
해서 직접적으로 그리고 여왕(왕)과 일꾼의 유충의 양육을 통해서 간접적으로 이러한 전달에 기여하고 있다. 이런한 맥락에서 이타적 행동은 폐쇄적 사회에서 특히 잘 발달되었다. 이에 비해 군집간의 관점에서 극단적으로 다른 군집의 동종을 죽음에까지 이르게 하는 공격적 행동들이 생겨날 수 있다. 결국 이타행동도 바로 〈집단 이기 주의 〉 와 연결되어 있을 수 있다. 물론 이타행동의 이점들이 집단 안에서 근연이 아닌 개체를 향해 발생할 수 있다 . 이것은 이타행동 양식들이 두 개체 상호간에서 나 타나거나(소위 상호이타주의) 사회 단위의 모든 구성원들에 거의 똑같 아 분포되어 있을 때만이 가능하다. 죽 경계를 하여 스스로 위험에 처할 수 있는 그 개체는 다음 번에 다른 개체의 경계행동으로 수혜 자가 될 수 있는 것이다 . 대체로, 표현형에 의해 이타주의적이라고 할 수 있는 이러한 행동 양식 자체들이 상호관계로 인해 혹은 포괄적응도의 의미에서 생겨난 것이 아니고 자연선택의 결과로 생겨난 것으로 설명될 수 있다. 결 국 모든 종들의 이타주의의 정도와 형태는 자신의 생존과 다음 세대 에 자신의 유전자를 전달하고자 하는 서로 상반된 요구의 타협에 의 해 일어 난다 (Ma y r 1967, Wi lso n 1975) . 4.4 집단의 종류 동물들의 집단 형성은 환경 조건에 의해 나타나거나 혹은 부수적 으로 사회적 유인, 즉 동종을 서로 찾고 함께 머물려고 하는 힘에 의해 기인될 수 있다. 여기서 전자~ 집합이라고 한다면 후자를 사 회라고 할 수 있다.
1) 집합 모임이라고 부를 수 있는 이 집합은 많은 개체들이 개별적으로 같 은 장소 예를 들면, 물먹는 곳, 유리한 은신처, 혹은 적당한 잠자리 나 동면 장소를 찾아 모임을 통해 일어난다 . 여기서 동물들은 서로 모이고자 의도된 모임이 아니고 모두가 자기 스스로를 위해 관련된 장소를 찾는다. 그래서 이런 모임은 사회적 의미에서 우연적이다. 이런 〈위선적 사회〉는 모든 동물 집단에서 잘 알려져 있다 . 2) 사회 특정한 환경 조건만을 가지고 설명할 수 없는 진짜 집단 형성은 여러 형태로 나타난다. 비클러 W i ckler (l 976) 는 이것을 분류의 원칙으 로 정했다. 그는 개방사회와 폐쇄사회로 나누며 개방사회는 구성원 들이 끊임없이 교환되는 것을 통해 특징지었다. 집단 전체의 행동은 구성원의 이탈을 통해서도 그리고 이입자의 출현을 통해서도 근본적 으로 방해나 변화를 받지 않는다. 이에 비해 폐쇄사회는 이러한 구 성원의 교환이 불가능하다. 이 사회 형태는 서로 상반된 행동을 취 하는 두 부류의 구성원들로 구분된다. 다양한 사회관계를 유지하는 자기 집단 구성원과 무관심 혹은 공격적인 행동을 취하기도 하고 극 단적인 경우에는 죽이기까지 하는 타집단 구성원이 있다 . 자기 집단 소속 개체와 타집단 소속 개체에 대한 양극적인 행동은 폐쇄집단에서도 개체의 일부 교환이 이루어지고 있기 때문에 물론 항상 아주 절대적인 것은 아니다. 그래서 사회가 폐쇄되어 있다라는 범위는 각 종마다 그리고 더욱이 한 종내의 여러 개체군에 따라서도 완전히 다룰 수 있다. 그럼 이 두 사회 형태를 다음과 같이 특징적인 형태에 따라 분류 할수있다.
®개방된 익명사회 떼 혹은 외형상 군집이라고 하는 이 집단에는 곤충의 떼, 새떼 그 리고 포유동물의 무리가 있다. 집합과는 반대로 이 집단은 보통 장 소 의존적이 아니다. 물론 우연한 모임과의 분명한 한계는 모든 경 우에 가능한 것은 아니다 . ®개방된 실명사회 사회적 의미에 있어서 고도의 발달을 가져온 것은 조류와 많은 어 류의 번식집단을 말한다 . 그들은 지속적으로 함께 머물면서 최소 한 도로 이웃 개체와의 개인적인 인지가 발달될 수 있다. 여기서도 만 일 구성원이 사라졌을 때 그 집단의 행동은 변하지 않으며, 이입자 들은 빈 자리나 가장자리를 차지할 수 있다. 폐쇄사회 체계는 집단 구성원과 집단의 개체와의 구별을 가능하게 하는 인지 기작을 요구하고 있다. ® 익명 폐쇄사회 친족으로나 곤충 사회의 국가와 같은 집단으로서의 이런 사회는 집단 구성원들이 개체를 초월한 특징을 인식하고 있다. 이런 특징은 집단내에서 똑같이 형성되어 있을 때 다른 집단과는 다르다. 이런 구성원을 나타내는 소위 신분증 기능을 하는 것은 보통 공동집단의 냄새 혹은 친족들의 냄새가 된다. 이 냄새는 두 가지 방법으로 생성 된다. 죽 몸 자체의 분비샘에서 혹은 빌려온 냄새물질을 통해서 갖 게 된다 많은 설치류, 페카리 Taya s su (남, 북미산 멧돼지), 그리고 주머니 하늘다람쥐 Pe t au ri dae 의 경 우 집 단구성 원들은 오줌이나 분비 물질로 몸의 접촉 혹은 실제 배우자 표시를 하여 같은 집단냄새롤 피운댜 이에 비해 꿀벌의 경우, 각각의 무리는 벌통냄새라고 하는
무리 냄새를 소유하고 있는데, 같은 소속 무리가 금방 찾았던 꽃을 몸에 발라 그들 무리의 특유의 냄새를 나타낸다. 상호 인식에 대한 집단 냄새의 의미는 실험적으로 쉽게 검토가 가 능하댜 타집단 개체에게 자기 친족의 특유한 냄새를 발라 놓으면 이 개체를 자기 집단 개체로 받아들이지만, 몸에 타집단 구성원의 냄새 물질을 발라 놓으면 자기 구성원일지라도 타집단 개체 취급을 한다. 구성원 신분증에 근거한 폐쇄사회의 이점은 구성원의 가능한 수에 있댜죽 개별 인식에는 한계가 있는 반면 집단 냄새로 많은 구성원 들을 확대할 수 있다 . 꿀벌의 무리는 무려 7 만 마리까지 구성될 수 있다. ®실명 폐쇄사회 단지 고등 척추동물에서 알려져 있는 고도의 사회성 결합에서 그 구성원들은 서로 개별적으로 인식하고 있다. 여기서 만일 구성원 중 한 마리가 없어지면 그 집단의 행동에 변화가 생긴다. 예를 들면, 탐 색행동이 유발될 수 있다. 이 실명의 사회는 원원류, 유인원, 개과 의 식육류, 사자, 그리고 사향고양이 Vi verr i nae 에서 찾아볼 수 있다. 아프리카 들개는 여러 마리의 수컷과 암컷 그리고 새끼들의 무리를 지어 살면서 새끼양육에 모든 성체들이 참여한다. 그들은 새끼들을 서로 돌보려고 훔쳐가기도 하며, 또 새끼의 어미가 죽어도 그 새끼 는 무리에 의해 양육된다. 무리가 사냥에 나가면 한두 마리의 성체 들은 새끼를 위한 감시자로 그대로 머물러 있게 된다. 사냥 나간 개 체들은 대략 손바닥만한 먹이 조각을 물고 무리에 다시 돌아온 뒤에 그것을 다시 목을 졸라 죽이는 시늄을 한다. 그렇게 해서 새끼들과 함께 머물러 있던 성체들은 그 먹이룰 취한다. 배고픈 성체들은 각 기 동료에게 먹이를 달라고 구걸을 할 수 있다 . 이러한 먹이 분배
기작으로 별로 사냥이 성공적이지 못한 이 사냥꾼들은 무리 전체를 먹여 기를 수 있다. 이 실명의 사회 성립에 근본적인 전제가 되는 것은 동료간의 개별 인식에 있다. 이 개별 인식은 배우자, 부모, 새 끼, 터 이웃 개체나 둥지 이웃 개체에 한정되며 이것은 냄새, 외형 혹은 목소리로 가능하다. 4.5 개별 인식 개별화된 사회구성에 대한 기본 전제가 되는 동료들간의 개별 인 식은 물론 이런 사회 형태의 종들에 한정되어 일어나는 것은 아니 댜 오히려 개별 인식 능력은 대부분이 척추동물에 있고 일부 무척 추동물에서도 보고된 바 있다 . 그것은 집단 형성 외에도 쌍의 배우 자간, 터 이웃간, 어미와 새끼들간 혹은 자매 집단 내에서도 나타난 다 (Beer 1970, Breed 와 Bekoff 1981, Lins enma ir 1979) . 개별인식은 사회적 결합과 순위제를 유지하는 모든 종에게는 하나 의 전제 조건이 되고 있다. 무엇보다 그것은 다론 개체의 인식이 장 소와 관계없이 일어나야 하는 모든 곳에서 의미가 있다. 죽 어떤 동 물이 위치해 있는 장소의 표시가 보조 수단으로 사용될 수 없는 곳 에서 개별 인식은 중요하게 작용한다. 개별 인식은 시각적 표시(예, 많은 조류에서 인식되는 머리 무늬나 생 김새), 소리내기(조류와 영장류는 소리를 통해 개별 인식이 가능하다) 그 리고 냄새 신호(무엇보다 포유류, 어류 그리고 무척추동물에서 잘 알려져 있다)에 의해 이루어질 수 있다. 그것은 각인 과정에서 형성될 수 있 거나 개체군의 소속원 사이에서 장기간의 상호접촉 결과로 나타날 수있다. 최근에는 동물들의 개별 인식 현상이 사회생물학적 근친관계와 관 련해서 많은 홍미를 불러일으키고 있다 (He pp er 1991). 그 이유는 개
별 인식이 근친 인식에도 기여하고 있으며 그것으로 말미암아 비슷 한 유전자형의 개체와의 짝짓기로 자신의 유전자의 확대를 극대화시 킬 수 있기 때문으로 생각하고 있다 . 반대로 개별 인식 역시 근친상 간의 단점이 될 수 있는 근연 개체와의 짝짓기를 억제시킬 수 있다. 그래서 최근에 개별 인식이 중요한 전제가 되는 근친상간의 실질적 인 회피 사례가 보고되었다. 몇몇 조류의 경우, 형제 자매 간에 서 로 짝짓는 것을 피한다. 오랫동안 그들을 함께 가둬 길러도 전혀 성 적인 활동을 보여주지 않는다. 야생의 침팬지의 경우에도 젊은 암컷 들에게 형제가 성적인 접근을 해오면 더욱 거센 방어행동이 관찰되 었다. 4.6 집단의 기원 집단 형성에 대한 두 가지 원리가 가능하다. 동물들은 모여들거나 함께 머물 수 있다 (Mar iel 1971 ). 첫번째 경우, 이전에 집단으로 산 적이 없던 개체들이 어떤 특정 시간대에 예를 들면 번식이나 이동을 위해 서로 모이는 것을 말한다. 두번째 경우는 태어날 때부터 함께 있었던 개체들이 지속적으로 같이 머물러 있는 것을 의미한다. 이런 사회 형성의 두 부류는 모든 동물계에서 매우 일정하게 나타난다. 개방사회는 첫번째 방식에 의해 생겨나며 계통발생학적으로 집합에 서 발전해 왔다. 이에 비해 폐쇄사회는 가족에서 출발했으며 독립한 새끼가 부모와 함께 머무는 것에서 기인된다. 이것은 친척으로 이루 어진 포유류나 조류의 비교적 작은 무리 혹은 수천 마리로 이루어진 곤충사회에도 적용된다. 이러한 이유에서 실제 고도로 발달된 사회 체제가 새끼양육을 하는 동물군에서 압도적으로 존재한다. 폐쇄사회가 가족에서 기원했다는 근거는 두 가지 중요한 새끼양육 의 특징과 결부되어 있다. 하나는 이타주의적 요소이고 다른 하나는
보통 다극성 즉 한 마리가 여러 마리의 동종의 개체와 상대하는 것 에서 근거를 찾아볼 수 있다 . 그래서 새끼양육은 유대기작의 발달을 위해 아주 좋은 전제조건을 내포하고 있다. 정상적으로는 하나의 가족이 시간적인 사회형성을 나타낸다. 그에 대한 생물학적 의미는 새끼들의 양육에 있는 것이다. 독립시기에 달 한 뒤에는 새끼들은 부모와 이별을 한다 . 폐쇄사회를 이루는 종에서 즉각적인 가족의 해체는 일어나지 않는다. 물론 가족이 함께 머무는 기간은 매우 다양할 수 있다 . 많은 경우에 새끼가 성적인 성숙과 함 께 분리가 되기도 하고 일부의 경우에는 그 유대가 그 이후에도 지 속될 수 있으며 경우에 따라서는 평생 지속되기도 한다. 이것에는 곤충 사회뿐만 아니라 몇몇 설치류, 육식동물, 유제류 그리고 영장 류가있다. 4.7 유대기작 짝형성뿐만 아니라 집단생활하는 동물에서 특히 문제가 되어 타협 이 요구되는 행동은 종내 공격성이다. 이미 언급된 종내 경쟁의 원 인으로부터 공격성온 사회성 종에서도 근본적으로 존재하기는 하지 만 단지 타집단의 구성원에 대해서 유발된다 . 이에 비해 자기 집단 내에서 공격성은 유대에 불리하게 작용할 수 있다. 그러므로 종내 공격을 일으키는 긍정적 및 부정적 선택압의 영향하에서 대부분의 사회성 종들은 계통발생학적 과정에서 집단 내 공격성을 억제시켜서 밀착된 공동생활을 가능하게 하는 기작을 발달시켜 왔다. 순위제 형 성과 달램 기능을 갖는 행동양식의 발달이 이에 속한다. 1) 순위제 일반적으로 폐쇄사회는 서열에 의해 구성된다. 순위제hi erarch y의
형태는 종마다 다양하다. 그것은 수직적인 순위제로 구성원 모두를 지배하는 서열이 가장 높은 알파 개체에서 복종적인 가장 서열이 낮 은 오메가 개체에 이른다. 이러한 수직적 순위제에서 여러 가지의 파생된 순위제가 존재한다. 그중 하나로 종종 삼각관계의 순위제가 있는데 이것은 베타는 알파에 대해 상위, 알파는 감마에 대해서 상 위 그러나 감마는 다시 베타에 대해 상위 서열을 갖는다. 순위제는 성혼합형 혹은 성분리형이 존재한다. 성분리형인 경우, 수 컷과 암컷의 서열 순서가 서로가 독립적(예, 토끼, 마카크원숭이, 개코 원숭이)이거나, 암컷의 서열이 짝인 수컷의 서열에 따라 달라진다 (예, 까마귀) . 이런 종속 서열은 새끼에서도 잘 알려져 있는데 이 새 끼의 서열은 많은 종에서(예, 마카크원숭이) 어미의 서열에 의해 결정 된다. 순위제는 오랜 기간 유지될 수 있다. 그러나 그것은 보통 최소한 이웃 서열간에 밀어내기가 일어날 수 있기 때문에 성장한 개체가 차 츰 승진되거나 혹은 상위 서열자가 강등함으로 해서 극적인 상황이 전개되기도 한다. 흡혈박쥐는 무리를 지어 오래된 나무굴이나 동굴 에서 사는데, 서열에 따라 매달리는 곳이 다르다. 하렘 수컷은 암컷 집단과 늘 몇 센티미터 떨어져 생활하는데, 암컷 가운데 누가 발정 기에 이르면 짝짓기를 하기 위해 암컷 집단 내로 들어간다. 그러나 이 하렘 수컷 바로 아래 서열의 수컷은 암컷들이 모여 있는 곳 아래 쪽에 홀로 떨어져 있으면서, 언제든지 하렘 수컷과 한판의 승부를 겨뤄 하렘의 자리를 차지할 기회를 노린다. 이보다 서열이 더 낮은 수컷은 동굴 벽면 제일 아래쪽에 자리를 잡고 있는데(그림 9-29), 수 컷들은 암컷들과 짝짓기하는 데 보다 유리한 위치를 차지하기 위해 끊임없이 자리 다툼을 벌인다. 사육 조건에서 흡혈박쥐는 성장한 젊 온 수컷 개체에 의해 하렘 자리를 내주는 것이 관찰되었는데 이 하 렘의 자리는 1 개월에서 20 개월을 두고 새로 성장한 수컷들에 의해
 7
7
바뀌어져 갔다 (Park 1991). 개코원숭이 암컷들이 발정주기에 따라 서 열이 달라지듯이 많은 순위제는 주기적인 변화를 보여준다 . 종속 순 위제의 경우 암컷의 위치는 짝형성에 따라 극적인 변화가 일어날 수 있다 . 서열이 높은 동물들은 보통 어떤 특권 (먹이나 물먹는 장소에 먼저 접근, 유리한 잠자리나 휴식처 사용, 짝짓기 지역에서의 특권)을 누린다. 그러나 그들은 집단 내에서 특정한 의무가 지워질 수 있다(지도자 역 할, 감시기능, 방어). 종종 매우 서열이 높은 개체는 집단 구성원들 사이의 싸움에서 중재자로 관여한다. 물론 사회적 서열의 현상은 집단생활을 하는 동물들에만 한정되어 나타나는 것은 아니다. 쌍으로 생활하는 종에서도 무엇보다 짝형성 이 지속적일 때 서열 차이는 있을 수 있다. 적어도 수컷이 상위를 차지하지만 많은 종에서 번식 초기에 잠시 역전된 순위가 일어나기 도 한다. 이 역순위는 새끼양육을 위해 주책임을 맡고 있는 암컷을 사나운 수컷의 공격으로부터 지켜준다. 그래서 이런 우열의 변화는
특히 매우 공격적인 종들(예, 인도산 지빠귀 Cop s y c husmalabar i cus) 에서 잘 알려져 있으나 역시 많은 되새과의 조류(멋쟁이새)에서도 일어나 고있다. 순위제는 지금까지 어류, 파충류, 조류 그리고 포유류에서 잘 알 려져 있댜 자연 상태에서는 단독으로 사는 동물일지라도 사육 상황 에서는 많은 경우에 사회적 서열을 띤다. 먹이원(예, 썩은 고기를 먹 는 종의 경우 시체)이나 물먹는 장소에서 먼저 먹는 여러 특권은 집단 생활을 하는 동물의 경우, 여러 집단간에서도 그리고 여러 종간에서 도 일반적으로 나타날 수 있다. 종간에 나타나는 이런 현상을 헤디 거 Hed ig er (1 941) 는 생물학적 순위제라는 말로 부르고 있다. 순위제는 어느 정도 집단 내의 결속을 강화시키는 데 기여하여 왔 댜 다툰다는 것은 순위제가 새로 형성될 때 혹은 변화(예, 성장중인 새끼들의 순위제 가담)할 때만 국한되어 있고, 반면 시간대에 따라 다 양한 권리는 다소 싸움없이도 존중되거나 다툼이 이웃 서열(비슷한 서열) 개체에 한정되어 일어난다. 이러한 관점에서 순위제는 텃세제 와 기능적으로 유사성을 갖는다. 순위제의 존재에 대해 행동학 연구의 초기에는 닭의 관찰을 통해 주의를 갖게 되었다. 닭에서 순위 다툼은 부리를 찍어 일어난다. 이 러한 이유에서 행동 연구의 초기에는 순위제의 개념을 쪼기 순위 p eckorder 라는 말로 사용했지만, 오늘날에는 더 이상 사용하지 않 는다. 2) 집단 유대의 행동 집단 내 공격성을 누그러뜨리는 두 가지 가능성은 공격과 도피가 계속해서 억제되어야 하고, 그렇게 하여 집단 구성원 상호간의 접근 과 동거가 가능하게 하는 행동 성향이 활성화되어야 한다. 이런 목 적을 충족시키는 행동 형태는 달램행동 혹은 인사행동으로 표현된
댜 이런 행동들은 구성원들이 잠시 떨어져 있은 후에 다시 만남이 있게되면 특히 자주 일어난다. 유대기작을 위해 계통발생학적으로 이미 공격억제와 배우자 유대 성향을 띠는 두 가지 주요 행동이 이용된다. 그것은 양육행동와 성 적 행동이다. 사실 대개의 집단형성 행동양식은 이런 범주의 행동에 서 출발했다. @양육행동 가족에서 나온 폐쇄사회의 기원에 따라 양육 행동은 집단 형성에 널리 퍼져 있다. 사회성 곤충에서 성체들은 입과 입으로 먹이를 건 네준다. 그것은 마치 그들의 단서성인 조상종에서 유충이 성체로 부 터 먹이를 받아먹는 것과 같은 식의 먹이공급 방식이다. 역시 척추 동물들도 짝짓기 행동과 짝유대 행동 범주에서 이미 중요한 역할을 하는 먹이전달(영양교환) 행동양식들이 확실히 집단 유대에서도 생겨 나게 되었다. 이 성체들간의 상호 먹이먹이기는 이미 언급된 바 있 댜 역시 오스트레일리아의 숲비둘기 Ar t amus 에서 서로 짝이 아닌 성체 집단 구성원들에게도 먹이를 먹일 수 있다. 더 이상 실제로 먹 이 전달이 일어나지 않는 종의 경우에서조차 이런 행위의 흔적 형태 로 보아 그 기원이 새끼양육의 기능 영역에서 나온 것임을 어렵지 않 게 짐작할 수 있다. 집단 구성원들은 새끼의 구결행동과 똑같이 혹 은 이 행동에서 출발한 듯한 동작으로 자주 서로 인사를 한다(조류의 경우 구결행동과 구걸소리, 포유류의 경우 주둥이로 찌르기와 입술 핥기). 좀더 넓은 범주의 새끼양육에 사회적 화장과 둥지짓기가 속한다. 많은 종에서 짝유대에 중요한 역할을 하는 서로 긁어주기, 서로 핥 기, 혹은 서로 털고르기가 나타난다. 물론 이것은 짝이 아닌 집단 구성원간에도 일어난다. 또 많은 조류에서 둥지짓기 동작이 인사행 동으로써 사회적 접촉을 하게 하는 데 기여할 수 있다.
 그림 9-30 모든 단풍새류에서 암컷의 교미 요구로써 기여하는 꼬리깃 진동은
그림 9-30 모든 단풍새류에서 암컷의 교미 요구로써 기여하는 꼬리깃 진동은
®성적 행동 유대기작의 계통발생학적 발달에 대한 양육행동외에도 단서성인 종과 집단생활을 하는 동물들의 조상종에서도 한 쌍의 배우자간의 상호결합이 존재한다. 이런 배우자간의 상호결합은 보통 짝짓기 행 동에서 기원된 행동양식들을 통해 유지된다 . 역시 이런 것들에서 일 부는 집단 내 사회적 접촉에 기여할 수 있다. 그래서 많은 키클리드 과의 물고기들의 경우 집단 내 공격억제 동작이 수컷의 구애동작과 일치하고 있다. 단풍새류에 속한 오스트레일리아의 Poeph il ap e rson- a ta 의 경우 집단 구성원들은 꼬리깃울 위 아래로 진동시켜 인사를 한댜 이것은 암컷이 교미를 요구할 때도 이런 행동을 취한다(그림 9-30). 역시 많은 영장류들도 암컷의 교미 요구 행동이 사회적 화해 행동으로 작용한다. 이 사회적 화해 행동은 상위자에게 하위자가 자 기의 성기를 제시함으로써 표현된다. 이렇게 두 경우처럼 성적 행동 영역에서 암컷에게 한정되어 있는 관련 행동양식들이 암수간의 사회 적 행동에서 나타나고 있다.
제 10 장 행동의 유전 행동유전학 분야는 그 출발에 따라 두 가지로 분류된다 . 유전학자 들은 유전적 문제를 해결하기 위해 매우 간단하고 쉽게 관찰할 수 있는 행동양식들을 대상으로 연구한다. 무엇보다 이들은 특정한 행 동 요소들을 발현시키는 유전자가 무엇인가를 밝혀내는 데 행동을 하나의 형질로 이용하고 있다. 지금까지는 거의가 초파리를 대상으 로 이런 연구가 이루어졌다. 그 이유는 초파리가 이런 연구를 위한 유전적 지식을 얻는 데 실제 충분하며 이런 유전자의 제거를 통한 유전적 지식과 이렇게 해서 나타난 행동을 유전자를 처리하지 않은 다른 개체들의 행동과 비교하는 연구가 가능하기 때문이다 . 유전학 자에 비해 행동 자체가 관심의 대상이 되는 행동학자들은 행동의 유 전적 기초를 설명하기 위해 고전적 유전적 방법, 죽 교배실험을 해 왔다. 말하자면 행동유전학은 지금까지 논쟁의 대상이 되었던 생득 적 영역과 학습의 영역의 주제에 대해 행동 성향의 유전적 근거를 갖고 궁극적인 증거로 제시할 수 있게 되었다 . 그러나 안타깝게도 지금까지 이 두 행동유전학적 연구 분야 사이에 실제적인 역할이 이 루진 바가 없다 .
l 연구방법 행동학적 측면에서 행동유전의 연구는 다른 행동학 분야 혹은 일 반 유전학 분야보다 더 많이 이루어지지 않았다. 행동 형질을 대상 으로 한 교배실험이 쉽지 않은 데 원인이 있으며, 무엇보다 다형질 교배를 위해 개체 수가 많이 요구되는 데에 문제가 있다. 그래서 이 런 실험은 원래 실험실 조건 혹은 최소한 우리 안에서 사육이 가능 하고 게다가 세대가 짧은 종에 한정되어 실시될 수밖에 없다. 그리 고 이런 종들은 행동을 쉽게 알아볼 수 있고 측정이 가능한 행동 차 이를 보여야 한다. 그러나 이것을 통해서 서로가 매우 가까운 근연 관계로써 서로 교배가 가능해야 되며 많은 자손을 낳을 수 있어야 한다 물론 이 경우 E 에서만 한정되어서는 안 된다. 그러나 실제로 이러한 전제 조건들을 만족시킬 만한 행동 양식은 매우 드문 편이다. 행동유전학적 연구들은 근본적으로 다음과 같은 결과를 가져올 수 있다. 교배한 두 종의 잡종의 행동을 서술 및 정량화하여 그 양친의 행동과 비교할 수 있다. 그렇게 함으로써 유전법칙에 대해 밝혀낼 수 있으며 무엇보다 E 의 행동이 중간유전 혹은 양친 종의 행동만 나타나는지, 다시 말해 그것이 우성(우열의 법칙)으로 작용하는지에 대해 알아볼 수 있다. E 간의 계속된 교배실험을 통해서 하나의 행 동양식이 유전자에 의해 지배를 받고 있는지(단성교배) 혹은 여러 유 전자 쌍들이 행동의 발현에 참여하는지(양성교배 혹은 다성교배)를 알 아볼 수 있다. 그러나 지금까지 이런 두 번째 단계는 이미 언급된 방법적 이유로 말미암아 아주 적게 실시될 수밖에 없었다. 행동형질 에 대한 유전적인 고정의 중요한 설명들은 인위적인 선택에 의해 이 루어진다. 다시 말해 이 유전적 고정은 특정 형질을 띤 개체의 선택 을 통해서 또 목적을 갖고 어느 정도 동종교배에 근거한 계속된 교 배실험을 통해 이루어졌다(그림 10-1) . 수백년 거듭되는 이런 실험
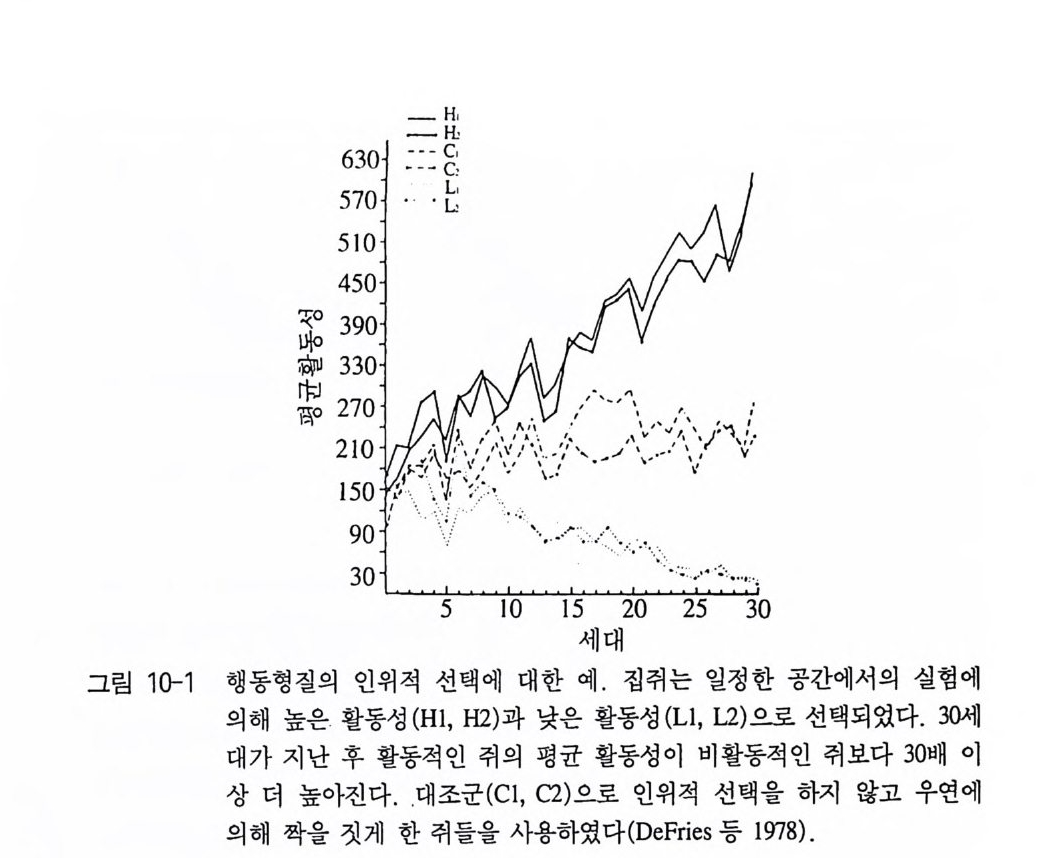 63----05 70.一국- 5 L 1 0450390330270210150
63----05 70.一국- 5 L 1 0450390330270210150
은 가축화의 과정에서 가능하다. 이때 형태적인 형질뿐만 아니라 행 동학적 형질도 인공 선택을 통해 점차 변화될 수 있는 것이다 . 2 잡종의 행동 잡종의 행동에 대해 이미 여러 종에서 많이 연구되어 왔다(오리 류, 비둘기, 앵무새류, 참새목의 일부, 어류 및 곤충) . 이들은 기본적으 로 서로 매우 비슷한 결과를 가져왔다. 양천종간에 단지 정량적인 차이를 보여준 행동양식인 경우 그 잡종의 행동들은 보통 불완전우 성의 유전 성질을 갖는다. 꿩과 닭 사이의 잡종 수컷들은 정확히 두
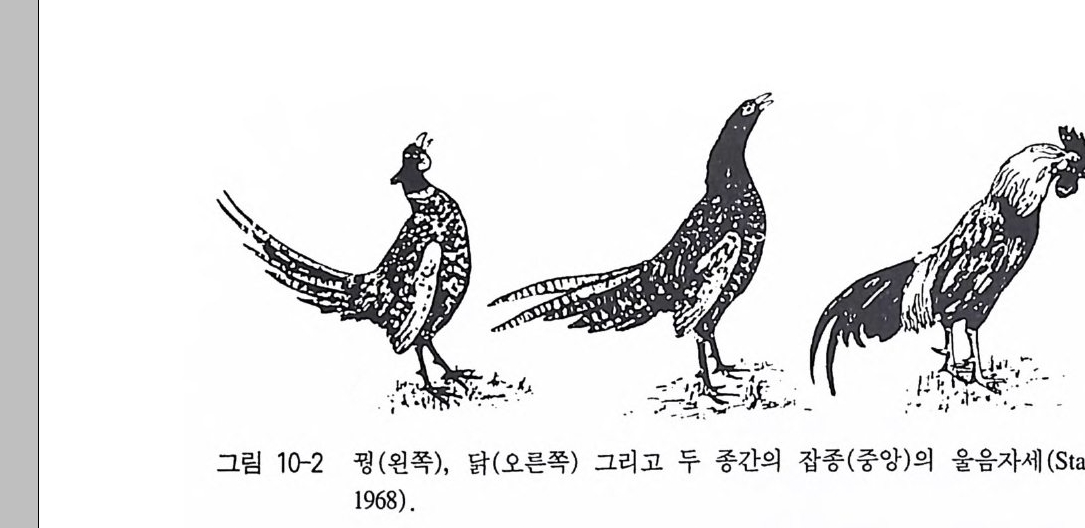 --. I \ ./! ' ~:'.. • '.
--. I \ ./! ' ~:'.. • '.
양친종의 중간에 놓여 있는 울음자세를 띤다(그림 10-2). (이 두 종은 속이 다른 먼 친척 관계로 인해 매우 드문 진정한 속과 속 사이의 잡종이 된다) 물을 빨아먹는 단풍새류와 물을 찍어먹는 단풍새류 사이의 잡 종은 물을 빨아먹지만, 부리를 물에 담그는 깊이는 대략 양친종의 중간 자세를 보인다. 이러한 불완전우성의 유전은 메뚜기과의 두 종 간의 잡종의 생득적 울음소리에서도 잘 나타나 있다(그림 10-3). 만일 이 양친종 가운데 한 종이 다른 종이 갖고 있지 않은 행동형 질을 소유하고 있다면 이 형질은 우열의 법칙에 따라 교잡종에서 보 통 완전히 나타나거나 혹은 전혀 나타나지 않게 된다 . 그러나 복잡 한 행동양식의 경우에 있어서 이 잡종들은 두 양친종들의 행동 요소 들울 부분적으로 모자이크식의 혼합을 갖게 된다. 예를 들어 그것은 Tila p i a 속의 2 종의 키 클리 드인 알을 바닥에 낳는 T. t hollon i종과 알을 낳아 곧바로 입 에 넣는 T. n i lo ti ca 종의 교배 에서 나타났다. 이 두 종 의 잡종은 불규칙적인 교환을 하여 양친종들의 행동 요소들을 모두 보여준다. 이들은 알을 바닥에 부착시키고, 지느러미로 부채질하 며, 알의 일부를 입안에 넣는 반면 나머지 알을 계속 부채질한 다음 입에 넣은 알을 다시 뱉는 동, 다양한 행동을 나타냈다. 그러나 이 때 성공적인 새끼의 양육은 이루어지지 않았다 (He inri ch 1967). 이렇
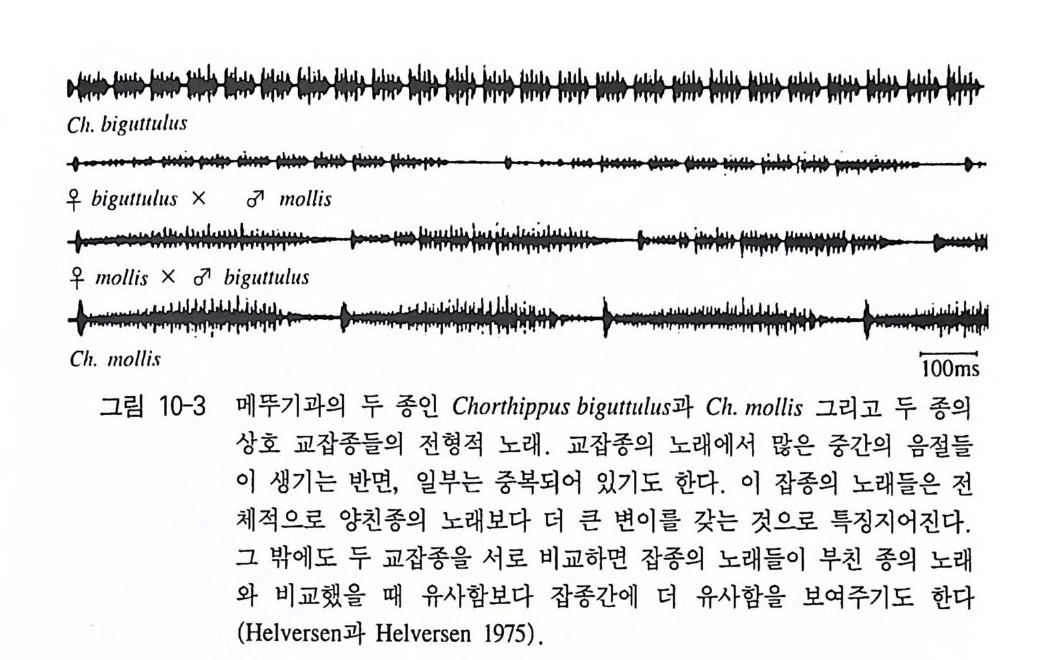 庫圖細軸細細細州 I 伽胤 I 卽險神빼軸빼禪圖勘圈卽曲i柚伽
庫圖細軸細細細州 I 伽胤 I 卽險神빼軸빼禪圖勘圈卽曲i柚伽
게 행동의 불협화에 대해 최근 자연 상태에서도 보고되었다. 이것은 인위적인 교배실험이 아니라는 데서 더 큰 관심을 모으고 있다. 이 것은 사회구조 형태가 서로 다른 2 종의 개코원숭이에서 발견되었다. 이 종들의 분포지역의 경계지점에서 관찰되었던 잡종둘은 사회조직 에 있어서 2 종의 혼합 형질을 갖고 있었다 (Kummer1975). A g ap orn i s 속의 작은앵무새의 교배실험을 통해 매우 재미있는 결과 가 나타났다. A.rose i co lli s 는 둥지 재료를 번식 구역으로 운반하는데 아주 희안한 방법으로 나른다. 이 종은 처음에 부리로 둥지 재료를 찢어 그것을 미저골것과 꼬리깃 사이에 꽂는다. 물론 이 둥지 재료 는 깃털에 있는 갈고리 때문에 운반 도중에 쉽게 떨어지지 않는다 (그림 10 국). A .fi she ri는 이 재료를 정상적인 방식, 죽 부리로 운반 한댜 이 두 종과의 잡종은 처음에는 깃털로 운반하려 시도하지만 그것을 성공시키지 못한다. 말하자면 이 잡종은 둥지 재료를 깃 아 래에 꽂지만 그것을 다시 곧바로 부리에 문다. 그 밖에도 이 잡종은
 그림 10- 4 미저골것과 꼬리깃 사이에 둥지 재료를 꽂고 그것을 둥지로 운반하는
그림 10- 4 미저골것과 꼬리깃 사이에 둥지 재료를 꽂고 그것을 둥지로 운반하는
원래부터 깃의 깊숙한 곳에다 그 재료를 밀어 넣지 못하고 또 가슴 깃털속과 같은 잘못된 몸 부위에 끼우려고 시도한다. 역시 이 잡종 은 원래 진정한 중간유전도 아니고 양친종의 행동을 갖고 있지 않다. 오히려 이 종은 양친종의 행동 요소들이 분산되어 나타난 경우다. A g ap orn i s 를 대상으로 한 교배실험에서 다음 2 가지 결과가 나타났 댜 잡종은 모두 양친종보다 더 길고 많은 둥지 재료를 잘랐으며 이 로써 잡종강세라고 불리는 형태유전(예, 잡종이 종종 양천종보다 크기 가 크다)에서 발생하는 성향을 보여주고 있다. 그 밖에도 이런 여러 몸 부위에 재료꽂는 행동은 A gap orn i s 속에 소속된 원시종인 다른 종 둘 (A . cana, A. ta ran t a) 의 정상행동에 해당된다. 그럼 여기서 행동의 아 타비즘 A t av i sm, 죽 양친형의 행동보다 속(屬)의 계열 행동과 더 닮 은 행동의 조상형을 의미하고 있다. 역시 이런 아타비즘은 무엇보다 형태유전에서도 나타난다. 3 교배실험 지금까지 연구는 잡종의 생식 능력의 부재로 인해 항상 F, 까지만 행해져 왔다. F2 까지 계속 연장된 몇몇 실험들은 행동양식의 유전
방식에 대해 확대된 분석을 가능하게 했다. 원충류인 Rhabdit is i ne r- m i s( 線形動物門)의 두 아종을 교배시키면 숙주 곤충과의 접촉 기회를 높이기 위해 머리를 흔드는 잡종만 나온다. 이것들을 자가교배시키 면 F 2 에서는 머리를 혼드는 개체와 머리를 혼들지 않는 개체가 3:I 의 비로 나타난다 . 이것은 머리를 혼드는 형질이 우성이자 단성교배 로 유전된다는 사실을 나타내고 있다. 이것에 비해 행동양식의 양성교배는 꿀벌의 두 품종간에 서로 차 이가 나는 양육 행동에서 찾아 볼 수 있다. 이것은 그들의 봉방에서 죽은 유충을 제거하는 행동으로, 소위 〈위생벌〉은 이러한 방들의 뚜 껑을 열고 죽은 애벌레를 꺼낸다. 이에 비해 〈비위생벌〉에게는 이런 행동이 결여되어 있다 . 이 두 계열을 교배시키면 비위생벌만 나온 댜 다시 이것을 위생벌과 역교배시키면 4 개의 서로 다른 유형이 나 타난다. 이 가운데 두 유형은 정상적이 아닌, 즉 위생벌도 비위생벌 도 아닌 봉방의 뚜껑울 열지만 죽은 유충을 제거하지 못하는 품종과 스스로는 뚜껑은 열지 못하지만 죽은 유충을 꺼낼 수 있는 품종이 태어난다. 이 네 행동 유형들은 대략 동수의 비로 나타났다. 이러한 결과로 두 가지의 부분적 행위가 두 개의 서로 다른 유전자에 의해 조절되고 있으며 두 개의 유전자가 열성인 것을 알 수 있다. 결국 완전한 위생행동은 두 유전자가 동형 유전자일 때만 생겨난다 (Roth e nbuhler 1964) . 물론 지금까지 알려진 사실들은 이런 단성교배나 양성교배의 유전 방식이 예외를 갖고 있다는 사실을 지지해 주고 있다. 죽 대부분의 행동형질들은 폴리진에 의해 발현되고 있는 것이다. 이 폴리진은 하 나의 형질을 발현시키는 데 항상 단순하게 서로 작용하지는 않는다. 즉 한 개체군에서 개체간의 변이는 연속적인 경향을 띠며, 그래서 3:1 혹은 1 : 1 의 규칙적인 비율로 결과가 나오지 않는다. 그래서 생 쥐의 공격성을 공격적이다 비공격적이다라고 단순히 분류할 수 없
댜 오히려 공격성의 정도가 광범위한 다양성을 보여 준다. 이 형질 의 연속적인 변이는 정량적인 유전분석 방법을 필요로 하며 그렇게 할 경우에 행동학적 형질이 어느 정도 유전적으로 구성되어 있는지 에 대한 일반적 개요만을 보여 줄 뿐이다. 이런 폴리진의 조절은 실무율식의 영향에 비해 개별적인 유전자를 통해서 자연선택이 단계적 성질을 갖고 있기 때문에 가장 작은 단계의 변화된 환경조건에 적응할 수 있게 해주는 생물학적 이점도 된다. 물론 유전자들은 간접적 방식으로도, 즉 수용기(예, 감각기관의 감 각능력), 중추신경계(예, 기억 형성의 능력), 호르몬 분비, 호르몬의 반응 준비 혹은 다른 형태학적 특징과 생리학적 성질의 변화를 통해 서도 행동에 영향을 미칠 수 있다. 4 선택실험 많은 실험들에서 인위적 선택을 통해 한 개체군 내에서 유전적 다 양성이 행동양식에도 적용되고 있음이 층명되었다. 양 극단적 표현 형의 선택적 장기 사육을 통해 선택된 계열 유지가 거의 항상 가능 하댜 선택실험을 실시하는 데 최소한 4-6 세대가 요구되고 있기 때 문에 초파리, 쥐 혹은 생쥐와 같이 세대가 짧은 종에 관심이 집중되어 있댜 그 밖에도 최근 귀뚜라미를 대상으로 선택실험이 이루어졌다. 귀뚜라미 G ry llus 는 두 종류의 수컷이 있다. 〈소리꾼 call 〉이라고 부 르는 수컷은 노래를 많이 하며 그 소리로 암컷을 유인한다. 이에 비 해 〈들러리꾼 sa t e llit e 〉이라고 부르는 수컷은 비교적 조용히 울며 소 리꾼의 소리를 듣고 이끌려 가는 암컷을 도중에 가로챈다. 물론 소 리꾼은 들러리꾼보다 더 많이 암컷을 유인하나 이 소리꾼의 소리에 의해 기생충인 파리가 모여 들어 더 괴로움을 겪는다. 이 수컷들을
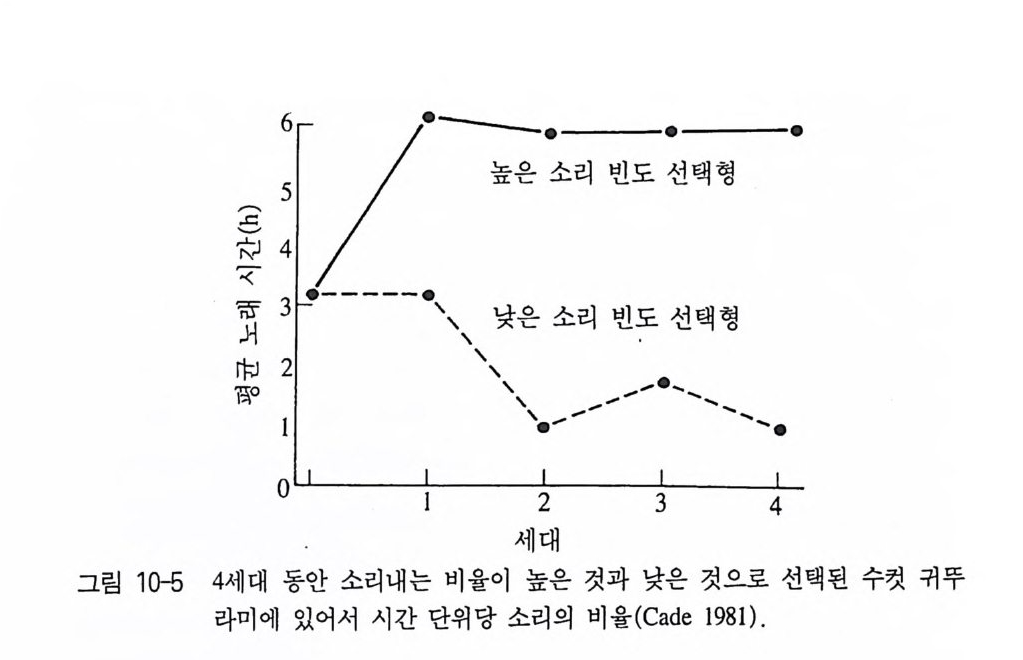 ; 5:6 r〔 /.\\/\•\-_높,-은.• \소 \리 .빈`도- •-선 택택형- -\ •
; 5:6 r〔 /.\\/\•\-_높,-은.• \소 \리 .빈`도- •-선 택택형- -\ •
실험실에서 격리시킨 디옴- 사육상자에 마이크를 장치하여 각각의 수 컷들이 부론 노래의 양을 측정할 수 있다 . 생후 7 일에서 16 일까지 하루에 노래한 평균 양을 측정해 보았더니 노래하는 수컷들의 수는 두 개의 정점을 지닌 분포 곡선을 보였다. 두 분포 곡선의 각각의 끝에 해당된 2 -4마리의 수컷들을 선택하여 그것들을 자매간이 아닌 처녀들과 짝을 짓게 했다. 이런 선택실험을 4 세대 동안 계속했더니 원래의 개체군에서 상당한 변이를 보이는 노래하는 두 유전적 군집 으로 나뉘어 갔다(그림 10-5, Cade 1981 ). 이 선택실험들은 자연 조건하에서도 비슷한 형태로 발생하는 행동 양식과 관련되어 있다. 주지성 행동은 자연적인 생활공간에서 곤충 이 정위를 하는 데 큰 의미를 갖고 있다. 의미 있게 배열된 T 자 미 로에 많은 수의 초파리들을 주지성 행동에 따라 분류할 수 있다(그림 10-6). 이것은 이 행동이 자연에서 출발한 개체군에서 특히 변화가 심했음을 보여 주었다. 많은 파리들은 강한 정의 주지성 반응을 보 인 반면 다른 파리들은 부의 반응을 보였다. 선택을 통해 주지성 정
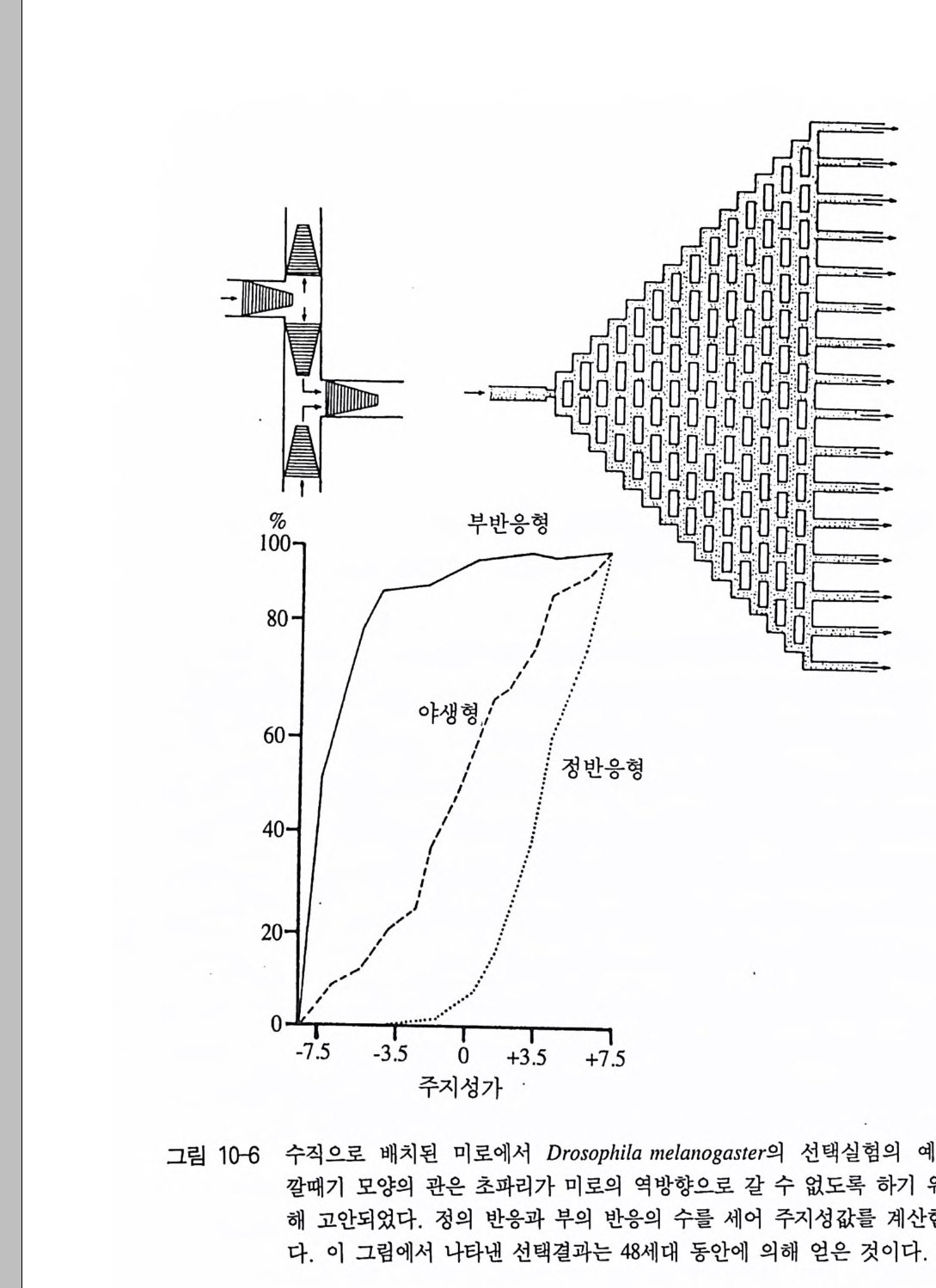 8%0 100 므I
8%0 100 므I
과 주지성 부의 계열을 사육할 수 있다. 특별한 교배기술을 이용해 이 주지성 행동에 관여한 초파리 염색체를 찾아내는 데 성공했다. 선택실험에서 파악된 유전자들은 최소한 초파리의 3 개의 큰 염색체 위에 흩어져 있음이 확인되었다. 이러한 종류의 선택실험의 결과는 행동양식의 진화를 이해하는 데 큰 의미를 갖고 있다. 자연 개체군의 유전자 풀에 유전자가 저장이 되어 있는 것으로 가정한다면, 이 유전자들은 한쪽 방향으로 정량적 으로 발현시켜며 또 개체군의 모든 개체에 존재하는 것이 아니고 단 지 개체에 따라 유전자 빈도가 달라진다(예, 모든 개체의 50%). 만일 환경조건들이 변화하면 자연 선택을 통해 개체군 내에서 어떤 행동 에 관여하는 행동유전자들의 빈도의 이동이 생길 수 있으며, 이로써 정량적 행동 변화가 일어날 수 있다. 그래서 행동을 근연종들간에 비교를 하면 이 근연종들은 근본적으로 동일한 행동양식을 취하고 있으나, 정량적 행동양식에서 많은 차이를 보여 주고 있다. 예를 들 어 어떤 행동양식의 빈도가 차이 난다거나, 어떤 기본 동작에서의 세부적인 차이를 보인다. 행동의 최초의 진화학적 변화는 그 행동 성향의 영역 내의 정량적 변화와 관련되어 있다. 예를 들어 근연종 들에 있어서 전체 고정 행동 패턴들의 있고 없고의 차이는 훨씬 드 물며 아마도 대부분 소진화의 단계들이 합쳐서 비로소 눈에 띄는 행 동 차이를 가져오게 된 것이다. 특정 행동 성향에 대한 선택은 동물의 다른 많은 변화를 거의 항 상 초래한다. 그 이유는 유전자들이 원래의 행동양식에 영향을 미치 지 않고, 동물의 여러 영향과 관련된 생화학적 요인에 영향을 미치 기 때문이다. 생쥐는 옹성에 있어서 높고 낮은 공격성으로 선택될 수 있다. 그러나 이 선택된 계열은 싸움행동에서만 차이가 나는 것 은 아니다. 공격성의 중가는 운동성 활동의 증가, 수많은 반사의 낮 온 역치, 성적 충동과 호기심 그리고 학습능력의 감소, 뇌세포의
RNA 의 감소를 초래한다. 여기서 개별적인 원인과 결과가 무엇인가 는 당연히 계속된 생리학적 분석에서만이 밝혀질 수 있다 . 5 행동의 돌연변이 모든 개체군에서 끊임없이 새로운 행동의 돌연변이가 일어난다고 생각해야 할 것이다. 그러나 이것은 행동의 돌연변이가 일반적으로 행동에 있어서 단지 미미한 정량적인 변화에만 영향을 미치고 있기 때문에 대부분 알려지지 않고 있다. 눈으로 확인이 안되는 돌연변이 의 유전자들은 아마도 자연선택을 위해 가장 중요한 물질을 나타내 고 있거나 근본적으로 종의 계통발생학적인 적응을 하게 하는 것들 이댜 그러나 생리학적 행동유전학은 방법적 이유로 우선 눈에 더 띄는 돌연변이들을 다루어 왔다. 유전학적으로 잘 연구된 몇몇 실험 동물로부터 비교적 크고 눈에 띄는 행동 변화를 수반하는 수많은 행 동 돌연변이들이 알려져 있다. 집쥐에서 괴상한 비정상 행동에 기인 하는 250 개 이상의 행동 돌연변이들이 보고되었다(영어의 예, ree-!er, weaver, waltz e r, shalcer, zig z ag 등등). 이 비정상 행동의 원인은 가 끔 비교적 쉽게 발견될 수 있다. 예를 들어 조직학적인 연구로 몇몇 돌연변이체들이 뇌량(腦梁 Corp u s callosum) 이 없거나, 신경세포에서 수초 혹은 내이(內耳)에서 이석(耳石)이 결여되어 있는 것으로 나타 났다. 대개 이런 눈에 띄는 돌연변이들은 동종교배 계열에서 나타난 다. 이 부정적인 선택치는 돌연변이들이 대개 생식능력이 없거나 동 형 집합자로 죽는 결과로 나타난다. 초파리의 경우 수많은 행동 돌연변이들이 방사능 물질이나 돌연변 이 유발 물질로 만들어 낼 수 있다. 행동 돌연변이들은 돌연변이 유 발 수술로도 만들어 낼 수 있긴 하지만 극히 드물기 때문에 세련된
기술이 행동 차이를 다소 자동적인 방법으로 분리시킬 수 있도록 발 달되어 왔다(예, 주지성 반응 실험의 미로). 만일 이런 식으로 비교적 조잡하고 선택적 단점을 가진 돌연변이들만이 압도적으로 많이 나올 수 있다면 비교적 미세한 행동변화들에 영향을 미친 몇몇 돌연변이 들도 그 중에 항상 존재하게 된다. 예를 들면 편차를 보이거나 결 여된 서커디언 리듬을 갖는 돌연변이들이 발견되었다(예, 19 시간, 28 시간, 무리듬). 또 짧아진 혹은 길어진 교미시간, 변화된 주지성 혹 은 주광성 반응이나 다른 학습행동을 갖는 돌연변이들이 생겨났다 . 어떤 돌연변이체는 야생형과 똑같이 학습을 하지만 야생형보다 4 배 빨리 학습한 것을 잊어버린다 (Franck 1985). 한편 지금까지 수많은 돌연변이가 초파리에서 기록되어 왔다. bar 라고 불리는 한 유전자가 복안에 있는 개안의 수를 감소시키는 반 면, wh it e 라고 불려지는 다른 유전자는 복안의 색소화를 감소시킨 댜 bar 와 wh it e 는 모두 초파리의 시각에 영향을 미친다. 이런 유전 자를 갖는 초파리 수컷은 볼 수가 없어 구애과정에서 암컷으로부터 시각적인 자극을 받지 못해 암컷에 접근하는 데 어려움이 있다. ves- tigial 과 dump y 유전자는 초파리의 날개를 기형적으로 만든다. 구애 과정에서 초파리 수컷의 행동의 주요 성분은 암컷에 접근하여 날개 를 진동시키 는 것인데, 가끔 ve rtigi al 과 dump y 유전자를 갖는 수컷 들은 적합한 행동을 할 수가 없다. 그래서 이런 모든 유전자들은 눈 이나 날개라는 중간 매체를 통해 구애행동에 영향을 미친다. 이와 같이 구애행동을 발현시키는 유전자라고 직접 말하기보다는, 실제로 많은 유전자들은 행동과 관련해서 일어나고 있다고 해야 할 것이다. 모자이크형 돌연변이 초파리를 실험실에서 만들어 유전자가 행동 을 발현시키는 원리가 연구되었다. 이 모자이크 초파리는 그들의 몸 에 유전적으로 다른 여러 세포를 갖고 있다. 어떤 세포는 수컷인 유 전자를 갖고 있으며 또 어떤 세포는 암컷인 유전자를 갖는다 . 만일
모자이크 초파리가 남성적 행동을 보여준다면 어떤 부위가 그 행동 을 지배하는지를 성모자이크 현상gy nandromo rp h 울 이용할 수 있다. 이에 대한 연구가 최근에 초파리의 구애행동에서 이루어졌다 . 초파리의 구애의식은 소위 정위라고 하는 스탭을 밟으면서 시작된 다 이 구애 수컷은 암컷과 약 0.2 m m 떨어진 곳에서 마주보며 있게 된댜 그런 다음 수컷은 앞다리로 복부를 가볍게 두드리며 암컷이 달아나면 따라간다. 다음 순간 수컷은 한쪽 날개를 들어 보이며 날 개를 진동시켜 〈사랑의 노래〉를 구사한다. 잠시후 수컷은 자신의 주 둥이를 내밀고 암컷의 생식기를 핥는다. 그런 다음 수컷은 암컷의 등 위로 올라가서 암컷과 교미를 한다. 홀 Hall (l 994) 은 최초로 초파리의 구애과정과 관련한 유전자들을 발견하였다 . 그는 초파리 한 마리의 체내에서 수컷의 세포와 암컷의 세포가 혼합되어 운반되는 성모자이크 초파리를 만들었다. 이 기술은 초파리의 생식발생에 대한 이해에 기초를 두고 있다. 초파리의 배에서 이러한 발생은 각 세포안에 들어 있는 X- 염색체에 의해 조절된다. 하나의 X- 염색체를 갖고 있는 세포들은 수컷의 해부 학적 구조와 행동을 일으키며, 두 개의 X- 염색체를 갖고 있는 세포 들은 암컷의 구조와 행동을 나타낸다. 만일 초파리 한 마리가 주로 암컷의 세포들을 갖고 있으나 뇌의 특정 부분에 수컷의 세포들이 들 어 있다면, 전형적인 수컷의 구애행동을 나타낸다 . 이 모자이크의 많은 실험이 있은 후 홀은 구애행동의 첫 과정(암 컷울 향한 정위, 복부의 두드림, 따라다니고 날개펴기)이 초파리 뇌 상단 과 후뇌 가까운 비교적 작은 지점에 있는 수컷세포를 필요로 하고 있다는 사실을 결론내렸다. 이 지점은 초파리의 여러 감각계로 부터 신호를 통합하는 곳이다. 달리 말하면 이 지역에 있는 수컷세포들은 수컷에 나타나는 구애행동을 통제한다. 이 구애행동의 다음 단계는 특히 정확한 운동성 협력을 하기 위해서는 신경계에 첨가된 수컷세
포를 필요로 한다 . 예를 들어 적당한 구애노래를 실행하기 위해 초 파리들은 이 통제 부위뿐만 아니라 가슴신경절에도 수컷세포울 갖고 있어야 한다. 그러나 이런 연구는 척추동물에서는 그리 간단하지는 않다. 그 이유는 척추동물의 성적 행동은 몸 전체에 영향을 미치는 생식선에 의한 호르몬의 결과로 발달하기 때문이다 (Greens p an 1995).
제 11 장 행동의 전화 생물체의 형태, 기능 그리고 생활방식에 대한 다양성은 생물이 생 겨난 이후부터 끊임없는 변화 과정의 결과로 생겨났다. 이것은 점진 적인 생명체로의 발전, 간단한 단계의 구조에서 보다 고등하고 특수 화된 형태로의 발전을 가져왔다 . 이런 과정을 진화라고 부른다. 이 진화과정에서 가장 중요한 추진력이 된 것은 돌연변이와 선택이었 다. 돌연변이는 유전 물질의 비약적인 변화를 가져왔다. 이것은 진 화과정과 관련해서 우연의 사건이며 , 말하자면 자연선택의 〈원료〉만 을 만들어 냈다. 자연선택은 생물체의 적합도에 따라서 생존과 번식의 기회를 결정 한다 즉 유전에 의해 환경조건에 최적의 적응력을 보이는 개체들은 덜 유리하게 적응된 개체보다 평균적으로 더 높은 생존능력과 번식 능력을 갖춘 자손들을 생산한다. 이것은 개체군 내에서 특정 유전자 들이 높은 빈도로 증가하는 반면, 다른 유전자들은 감소하게 되어 한 종의 전체 유전자는 점차 최적의 환경 적응의 방향으로 닙評]어 가는 것을 말한다. 그러므로 돌연변이에 바해 자연선택은 항상 진화 에 올바른 상태를 부여하고 있다.
모든 동물종들의 행동도 진화의 테두리 안에 놓여 있으며, 이전 단계의 다른 종류의 행동에서 발전해 왔다 . 그래서 행동은 계통발생 학적 발달과정이 알려지게 되면, 마지막 단계의 행동을 정확히 이해 할 수 있게 된다. l 연구방법 물론 행동 진화의 연구 방법은 어려움이 많다 . 우선 계통발생학적 연구의 가장 중요한 단서가 되는 화석이 행동진화의 연구에서는 거 의 찾아볼 수 없다. 있다면 이미 멸종된 종의 이동 방법에 의존하여 어떤 정보를 줄 화석화된 발자국 혹은 식성과 섭식 방법에 대한 실 마리를 제공할 화석화된 위 내용물과 먹이 흔적과 같은 몇몇 행동화 석이 있을 뿐이다 (Ber g I978). 그 밖에도 현존하는 동물종의 몸구조 로부터 추측 가능한 이동 방법에 대해 어떤 실마리를 끌어낼 수 있 고, 화석의 배열 상태를 보고 관련 종의 집단 구조에 대해 어떤 의 미를 찾을 수 있다. 물론 이런 간접적인 행동 증거의 범위와 폭이 매우 제한되어 있어서 이런 것들로부터 어떤 결정적인 정보를 기대 할수는 없다. 형태학에서 계통발생학적 과정의 재구성에 매우 본질적으로 기여 하고 있는 멸종된 동물 형태에 대한 정보는 행동 진화학에서는 그만 큼 이용되고 있지는 못하고 있는 실정이다. 오히려 행동양식들의 진 화에 대한 실마리가 현존하는 동물종의 행동으로부터 풀어야만 된 댜 이에 대해서 다음 두 가지 가능성을 제시할 수 있다. 첫째는 행 동의 개체발생을 연구할 수 있고 둘째는 여러 근연종들의 행동을 서 로 비교할 수 있다. 여기서 근연종과의 행동 비교는 이미 형태학적 인 계통분류에서 많은 연구가 이루어져 있기 때문에 그만큼 유리할
수있다. 2 상동관계의 행동 행동에 대한 계통발생학적 연구로 서로 비교될 만한 행동형질들이 상동인지 상사인지 밝히는 것은 중요하다. 그러나 여기서 행동학과 다른 생물학적 부분 원리와 비교해서 특히 새로운 문제에 직면하게 된다. 한편으로는 방법상의 문제에 있어서, 다른 한편으로는 정의를 내리는 데 있어서 어려움이 생긴다. 상동이란 일반적으로 말해 동일한 조상에서 유래된 몸의 구조와 다른 모든 몸의 형태적 특징에 있어서 일치하고 있을 때 이 말을 사 용하고 있다. 상사란 같은 기능의 적응으로 각각 계통발생학적 근연 관계와는 완전히 무관하게 형성되었을 때를 일컫는다. 형태학에서 상사에 대해 가장 잘 알려진 예로 새의 날개와 곤충의 날개 혹은 척 추동물의 눈과 두족류의 눈이 있다. 상동이 될 수 있는 기본적인 기준은 관련 형질들의 형성을 지시하 는 일반적인 정보 저장소의 존재에 의한다. 여기서 정보저장소란 유 전적 구조인 게놈g enome 을 말한다. 그러나 행동학에서는 다르다. 즉 행동의 형질에 있어서는 기본적으로 기억의 저장소인 제 2 의 정보 저장소가 필요하다. 이런 사실은 행동에 있어서 근연이 아닌 두 종 에서도 같은 형질이 나타날 수 있는 것으로 기본적 결론에 이른다. 그리고 또 다시 상동의 행동형질들의 운반자들은――상동이 아닌 그밖의 형질들에 비해서_하나의 공통의 조상형에서 유래가 필수 적인 것이 아니며 그래서 상동이란 이미 계통발생학적 근연에 대한 기준으로부터 표현될 문제가 아니라는 결론에 이르게 된다. 예들 들 어 한 종의 새가 다른 종으로부터 노래를 배운다면 이 소리내기는
두 종이 매우 다론 종(근연이 아닌 종)에 속해 있다 하더라도 서로 상 동이 될 수 있다. 그래서 행동학에서는 형태학적 계통발생과 집단적 계통발생과의 사이를 분명히 구분해야 한다. 형태학적 계통발생은 형태의 계통발생과 근연관계를 다루고 있으며 집단적 계통발생은 관 련 행동형질을 갖는 그 형질의 운반자 즉 그 종의 계통발생 및 근연 관계를 다루고 있다 . 마치 정보가 한 세대에서 다른 세대로 전달되는 종 역시 상동형질 이 이 두 세대 집단에서 차이가 난다 . 즉 생득적 형질인 경우에 정 보는 유전에 의해 전달되며, 획득된 형질인 경우는 전수에 의해 전 달된다 특히 계통발생학적으로 홍미가 있는 것에는 〈 전수 상동 〉 이 댜 이것은 다른 종의 발성을 배우는 것이 종의 학습 프로그램의 고 정된 구성분일 때 나타난다. 예를 들면 기생성 조류인 천녀새에서처 럼 이 종이 자신의 숙주 종의 발성을 넘겨받게 되어 이 천녀새의 소 리는 숙주새의 소리와는 전수 상동의 관계를 갖게 된다. 방법적인 면에 있어서도 이미 언급된 화석이 없다는 것은 상동관 계 연구에도 커다란 단점이 되고 있다. 여기서 상동은 상동형질의 수와 상동이 일어나는 규칙성에 따라 실제 겨우 설명되고 있는 실정 이다 그래서 상동의 존재를 추측게 하는 일련의 상동 기준이 마련 될 수밖에 없다. 두 종에서 한 행동양식의 진행이 가능한 한 다수의 형질에서 일치할 때(특별한 질적 기준) 혹은 한 행동양식이 두 종에서 사용체계가 비교적 같은 상황일 때 (상황 기준) (예로써 두 종의 한 행 동연쇄 내의 동일 상황에서 한 행동양식이 나타날 때) 상동으로 인정할 수 있댜 두 종에서 서로 비슷한 행동이 적다 하더라도 두 종 사이 에 단계적 변화가 있다면 상동이 될 수 있다 . 이러한 과도형태는 관 련 종의 개체발생 과정 혹은 근연종들간에 생겨날 수 있다(과도형태 로 인한 결합 기준) . 이런 기준들이 충족되면 될수록 그리고 각각의 기준들이 서로 더 잘 일치하면 할수록 실제 상동관계가 될 가능성은
훨씬 더 높다 (Remane 1971, W ick ler 1965, Wi ck ler 1967) . 3 종간비교 근연종간의 행동 목록들을 비교함으로써 각 행동양식의 계통발생 학적 발달과정을 비교적 정확히 재구성할 수 있다. 이따금 근연집단 내에서 단계적 변화를 띤 유사한 일련의 전체 형태가 제시될 수 있 으며 많은 경우에 고도로 진화된 종의 행동양식을 이런 관계에서 원 시 종과 비교를 통해서도 알 수가 있다 (Ma y r1967). 많은 열대성 단 풍새 류 Es tri ld i dae 의 수컷들은 구애시 부리 에 풀줄기를 물고 둥지 짓 는 행동에서 유래된 동작을 취한다 . 이렇게 둥지짓기 요소를 넘겨받 은 구애행동은 이 단풍새류에서 똑같이 나타나고 있다. 그 이유는 이 단풍새들이 많은 명금류에 비해 둥지를 짓는 데 암컷뿐만 아니라 수컷들도 참여하고 있기 때문이다 . 많은 다른 행동형질들을 기초해 서 더 원시적인 단풍새류로 생각하는 많은 종들은 구애시에도 둥지 짓기와 같은 종류의 풀줄기를 부리에 무는 반면, 오스트레일리아의 태양새 Neochm i a p hae t on 와 같은 종은 구애할 때, 둥지 지을 때와는 다른 재료를 사용한다. 여기서 풀줄기는 단지 상징적인 의미만을 갖 는댜 그래서 많은 경우에 구애시 순수한 둥지 재료의 사용은 완전 히 사라져 갔다. 이런 발달은 여러 과도 단계를 거처 이루어졌다. 많은 종에서 구애의 도입 단계에서만 풀줄기를 물고, 실제의 구애춤 에서는 풀줄기를 사용하지 않는다. 또 다른 종에서 풀줄기 사용이 임 의로 나타난다. 즉 수컷이 풀줄기를 물고 혹은 풀줄기 없이도 구애 를 할 수 있다. 결국 몇 종은 풀줄기춤이 드문 예외로만 관찰되고 있 댜 풀줄기 구애를 하지 않는 종들조차도 그들의 구애행위의 형태가 오히려 둥지짓기 행동에서 유래되었음을 인식할 수 있다(그림 11-
 그림 11- 1 단풍새 Est r il daas t r il d( 오른쪽)과 같은 많은 단풍새류들은 수컷이 구애시
그림 11- 1 단풍새 Est r il daas t r il d( 오른쪽)과 같은 많은 단풍새류들은 수컷이 구애시
1, Irnmelmann 1962) . 4 행동의 개체발생 계통발생학적 발달에 대한 특히 많은 연구결과가 행동 개체발생에 대한 연구로 관심을 돌리게 했다. 그것은 새끼의 행동에서 원시 종
의 행동 즉 관련된 조상형과 비슷하거나 거의 같은 행동의 형질이 나타나는데 그 원인을 찾아볼 수 있다. 정상적으로 더 이상 부리에 풀줄기를 물고 구애를 하지 않는 오스 트레일리아의 단풍새들의 어린 수컷들은 가끔 풀줄기를 입에 무는 행동을 한다. 땅바닥에다 알을 낳는 종달새는 성체가 되면 양다리를 하나씩 움직여 걷는데, 둥지에 나온 지 하루만 지나면 두 다리를 동 시에 땅에 대고 껑충껑충 뛴다. 이것은 근친종인 덤불에 알을 낳는 명금류의 행동과 같다. 이 명금류는 성체가 되도 양다리를 껑충껑충 뛰어 이동한다. 어린 바다코끼리들은 작은 물체를 향해 붙잡거나, 물 고, 혼들어 대는 행동을 보여 주는데, 이것은 육식성 포유동물의 먹 이동물울 잡는 행동을 연상시키고 있으며, 아마도 이들의 조상이 포 식성 기각류가 될 수 있다는 사실을 암시해 주고 있다. 육상생활형 으로 변한 열대성 게 종류의 하나인 야자집게 B i rg us la t ro 의 새끼들 은 일정한 기간 동안 소라 껍질을 찾는 행동을 취한다. 이 행동은 근연종인 소라게가 성체의 나이에 이르면 전형적으로 나타난다. 그 렇다면 새끼의 행동에서 원시종의 행동양식에 대해 조심스런 판단울 해볼 수 있다 (Rasa 1971, Rich ard 1979, Sudhaus 1974). 5 행동의 흔적 과거의 적응으로 조상으로부터 물려받아 오늘날까지 유지해 온 행 동양식 들을 행 동의 혼적 behavio u ral rel i c t이 라고 부른다. 비 록 이 행 동양식들이 그동안 아무런 기능도 없게 되었거나 최소한 원래의 행 동양식의 기능을 상실해 버렸을 수도 있다. 지금까지 소개된 예는 계통발생학적 흔적들이 특히 자주 발견될 수 있는 어린 동물들에서 만 이루어졌다 . 그러나 어떤 동물에서는 그 혼적이 성체에서도 나타
나며, 더욱이 하나의 운동 방식이 행동학적 기능을 충족시켰던 소속 기관을 계통발생학적으로 잔존하게 한 경우에서 특히 잘 나타난다. 따라서 많은 원숭이류(마카크)는 꼬리가 기형적으로 짧고 무게 중심 을 유지하는 데 별로 적당치 않아도 이들은 오늘에도 여전히 꼬리로 균형을 잡는 행동을 취한다 . 예를 들어, 고라니나 문작 Mun ti acus mun tj ac 과 같은 많은 사슴의 원시종들은 위협할 때 입술을 위로 치 켜올려 잇몸과 함께 송곳니를 들어내놓는다. 이와 같은 위협 동작은 몇몇 고등한 사슴류에서도 보여 준다 . 예를 들어, 붉은사슴 Cervus ela p hus 의 경우 뿔을 길게 형성함으로 해서 이 송곳니는 크게 퇴화 되었으며 그래서 싸울 때는 이 송곳니는 더 이상 사용하지 않아도 아직도 이런 위협 동작은 찾아볼 수 있다. 이들은 오늘날에는 더 이 상 갖고 있지 않지만 이런 흔적 무기를 보여줌으로써 위협을 한다. 수상성(樹上性) 뜸부기는 한번 둥지에서 벗어난 알은 더 이상 굴리지 않지만 실험상태에서는 자연상태에서 없었던 전형적인 알굴리기 행 동을 보인다. 그리고 몇몇 바다조류(예, 바다오리)는 원래의 둥지를 더 이상 짓지 않고 맨바닥에 알을 낳기는 하지만 구애 때는 둥지짓 는동작을보인다 . 종종 전형적 흔적으로 대표되는 행동양식은 많은 새들이 발로 몸 을 굵는 행동이 될 수 있다. 이때 새들은 발이 머리에 닿게 하기 위 해 날개를 아래로 내리고 같은 쪽의 다리를 날개 위로 쳐든다 . 이렇 게 힘들게 보이는 동작은 어린 새에서는 무게 중심을 잃게 하는 것 으로, 원래 네 개의 다리를 갖고 있는 몸 균형을 유지할 수 있는 동 물들에서만이 찾아볼 수 있는 것이다. 사지를 갖는 동물들은 현저히 앞에 위치한 앞다리 위로 뒷다리를 쭉 뻗어 머리에까지 닿게 할 수 있다. 그럼에도 불구하고 이 동작의 연속은 비록 사지 동물들이 이 동작이 운동의 수행능력과 관련해서 직접 실행되고 있다고 하더라도 놀라울 만큼 고정된 방법으로 나타난다. 즉 많은 앵무새류는 먹이룰
취할 때 발로 직접 먹이를 취해 날개 아래로 해서 부리로 가져온다. 그러나 굵고자 할 때는 날개 위로 해서 머리를 굵는다 . 만일에 부리 기부 근처를 긁고자 한다면 먹이룰 취하는 것 처럼 거의 같은 위치 에 발이 도달해야만 된다 (Brere t on 과 Immelmann 1962). 그래서 이 동 작이 실제로 파충류 시대에서 온 유산으로 보는 주장도 일리는 있 댜 그러나 지금은 일부가 이러한 의미와는 대립되고 있다. 죽 한편 으로 많은 종에서 개체발생학적 규칙에 따라 어린 동물들은 항상 다 리를 날개 아래로 해서 긁지만 성체가 되면 날개 위로 해서 긁는다. 다른 한편으로는 이 두 굵는 방법의 분포 역시 오늘날과 원시 조류 집단에서 분명히 임의로 나타나 어떤 것이 더 오래된 것인가를 알 수는 없다. 결국 많은 근연 집단(예, 앵무새, 구세계솔새 Parul i dae) 을 살펴보면 그들 가운데 일부가 날개 아래로 굵고 또 다른 일부가 위 로 긁는 서로 가까운 혈연종들이 있다(그림 11-2). 그러므로 이러한
 그림 11-2 머리깃을 청소할 때 발로 머리에 이르게 하는 종류와 방식은 근연종인
그림 11-2 머리깃을 청소할 때 발로 머리에 이르게 하는 종류와 방식은 근연종인
행동의 궁극적인 평가는 아직 완전한 것은 아니다 (Bur tt과 Ha ilm an 1978) . 6 의식화 종간 비교나 행동의 개체발생의 연구를 통해서 진화학적 발달의 규칙성은 개체간 의사소통에 기여하는 또 전체적으로 표정 행동으로 불리는 행동양식에서 분명히 나타날 수 있다. 의사소통에서 가장 효 과적이고 거의 실수가 없는 정보교환은 획득할 수 있기 때문에, 이 런 많은 행동양식들은 확실히 신호전달의 증진에 기여해 온 진화학 적 변화 과정에서, 무엇보다 공격행동 및 성적 행동에서 기원된 것 들에서 발견될 수 있다. 이런 과정을 인간적인 관습형성이라는 말을 본따 의식화( 儀 式化rituari za ti on) 라고 부른다. 의식화 행동은 여러 종들간의 비교 그리고 의사소통 과정에서 쉽 게 인식될 수 있는 일련의 특징에 있어서 비의식화 행동과 구별된 다. 실제로 이 의식화는 동작과정을 간소화하고, 과장하며 그리고 격식화함으로써 리드미컬하게 반복되어 나타난다. 게다가 눈에 띄는 몸의 표시(예, 체색)를 발달시켜 동작의 신호효과를 훨씬 더 증대시 킬수있다. 과장은 수많은 구애와 위협의 행동양식에서 관찰된다. 이런 행동 양식들은 정상적인 동작보다 훨씬 더 많은 에너지 소모가 따른다. 예들 들면, 구애하는 칠면조 수컷은 날개춤을 추듯이 이리저리 걸으 며 붉은사슴 수컷은 상투적인 싸움 직전 서로가 옆으로 걸으면서 아 주 천천히 부자연스런 걸음거리를 취한다. 많은 의도동작들도 과장 을 통해서야 그들의 신호효과를 갖게 된다. 과장에 이어 두번째로 자주 달라지는 것은 행동의 격식화다. 의식
화 동작의 과정은 그의 형태에 있어서 일반적으로 상당히 고정되어 있는 반면 비의식화의 경우 그 과정은 심하게 변한다. 그래서 이 동, 화장(몸치장) 혹은 먹이탐색은 그들의 강도와 완성도에 있어서 필요에 따라 달라진다. 이에 반해 의식화된 행동형태 죽 사회적 의 사소통에 기여하는 행동형태에 있어서 항상 계속 같은 속도와 똑같 은 강도로 진행된다 . 예로써 구애에서 넘어온 많은 화장행동이나 부 리로 두드리는 딱따구리의 소리가 있다. 이 딱따구리의 두드리는 소 리는 먹이탐색시에는 심하게 변화할 수 있지만 구애할 때는 아주 특 정한 종특이적 리듬을 갖추고 있다. 행동의 신호기능은 계속된 리듬 의 반복으로 향상된다 . 이것은 무엇보다 음성학적 영역에서, 예를 들면 많은 곤충과 양서류의 소리, 어린 새의 구걸소리 혹은 많은 조 류의 구애노래 son g와 구애소리 call 에서 찾아볼 수 있다. 수비둘기의 단조롭게 연결된 쿡쿡…… 소리도 여기에 속해 있다. 고형화된 시각 신호에는 농게의 집게발을 반복적으로 움직이는 것과 반딧불이의 빛 신호가 이에 해당된다. 물론 의식화는 계통발생학적 과정에서만이 생기는 것이 아니라 성 체의 고정된 행동양식들이 흔히 매우 가변적인 새끼의 행동에서 만 들어졌을 때 행동의 개체발생 과정에서도 일어날 수 있다. 이에 대 해 하나의 뚜렷한 예는 많은 명금류에서 관찰되듯이 매우 변화가 많 은 어린 새의 노래는 다소 같은 형태의 성체 수컷의 노래로부터 형 성된 과도기에 해당된다. 이런 경우를 개체발생학적 의식화라고 부 른다(그림 11-3, Tho rpe 와 Pil c her 1958). 이 의식화와 비슷한 행동양식의 형식화는 비자연상태의 성체의 동 물에서도 나타날 수 있다. 동물원에서는 움직일 수 있는 공간의 제 한으로 동물들이 소위 운동의 고정화 현상이 일어나고 있다. 말하자 면 강요된 결과로 몇 시간을 같은 동작을 취하는 것으로써, 예를 들 면 목을 돌리는 동작, 이리저리 혼드는 동작, 몸 전체를 돌리는 동
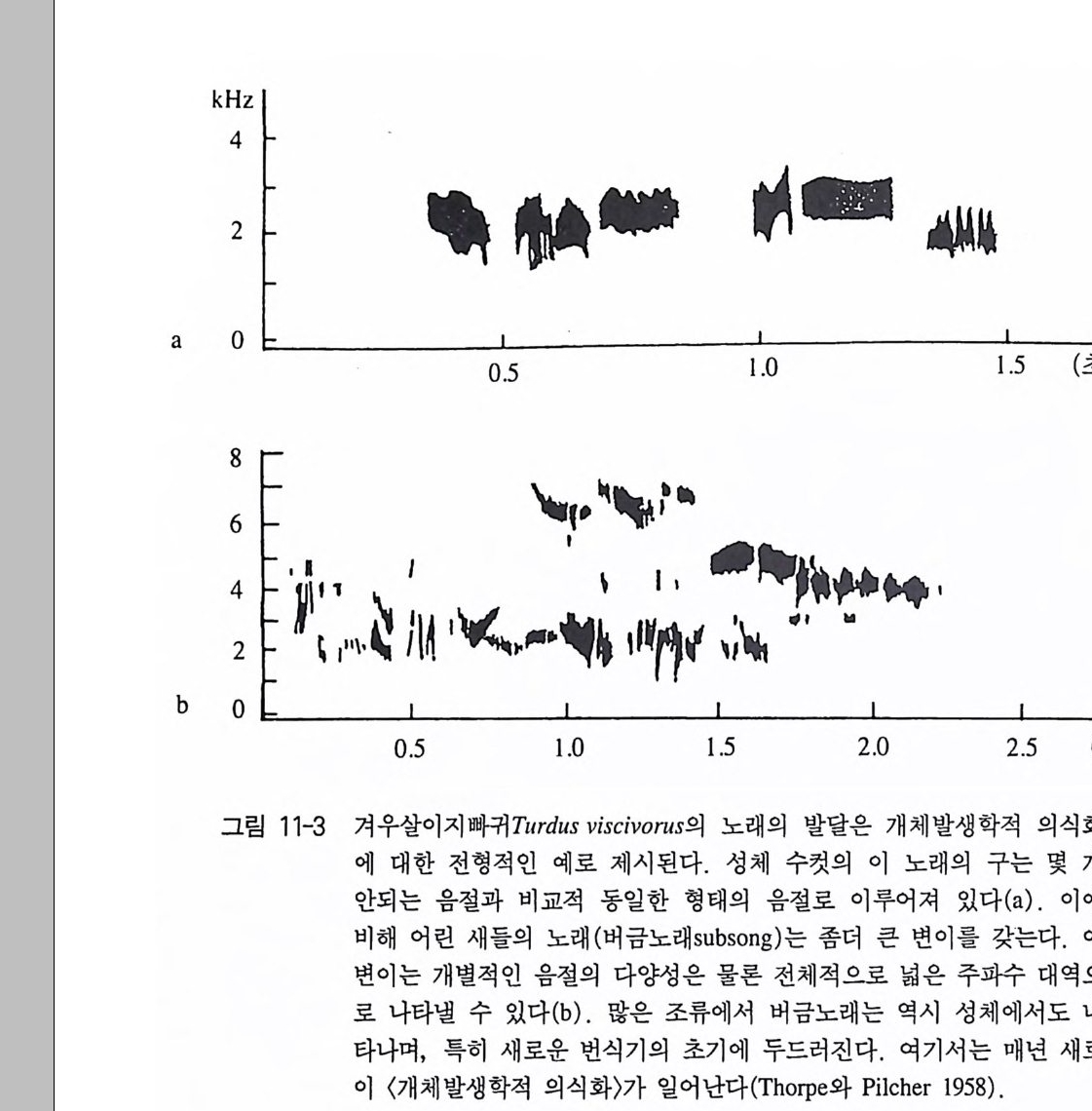 kHz
kHz
작, 그리고 정해진 거리를 왔다갔다하는 동작들이 있다 . 동물원의 많은 동물들의 구걸동작들 역시 거의 의식화된 형태를 갖는다. 의식화된 행동양식에 대한 출처로 실제 모든 행동 영역들이 문제 가 되고 있다. 그러나 이런 의식 행동양식들이 의도동작i n t en ti on movemen t과 전이 동작 dis p l a cement ac ti v ity의 행 동 범 주에 서 출발했
 그림 11-4 가마우지의 날아가고자 하는 의도동작 . 가마우지는 날기 직전에 볼 수
그림 11-4 가마우지의 날아가고자 하는 의도동작 . 가마우지는 날기 직전에 볼 수
댜 의도동작은 아직도 완전한 행동을 미리 알려 주지는 못한다. 그 것은 마치 둥지짓기 전에 둥지 재료를 쪼거나 많은 조류에서 날기 직전에 볼 수 있는 무릎굽히기와 같은 것이다(그림 11-4). 이 의도동 작은 관련 행동양식울 일으키는 자극이 필요한 역치가에 아직 도달 하지 못했을 때 나타난다. 의도동작은 의식화된 형태로 중요한 사회 적 신호로 표현될 수 있다. 많은 위협동작(예, 이빨 드러내기)들은 싸 움의 의도동작이며, 단풍새류의 풀줄기 구애는 둥지짓기의 의도동작 에서 기원된 것이며, 또 게논(긴꼬리원숭이屬)의 입술로 쩝쩝거리는 인사동작은 아마도 사회적 화장의 의도동작인 것이다. 이미 언급한 무릎굽히기는 떼를 짓는 조류에서 특히 과장되어 나타날 수 있는 데, 이것은 아마도 동료들의 이륙을 동조시키는 효과를 지닌다. 이 새들은 비행하고자 하는 의도에서도 몸을 상하로 크게 혼든다 (Andrew 1957, Daanje 1 950) . 전이동작 역시 의식화를 통해 이차 신호기능을 갖고 있으며 사회 적 의사소통에도 기여하고 있다. 전이동작은 특별히 구애행동과 위
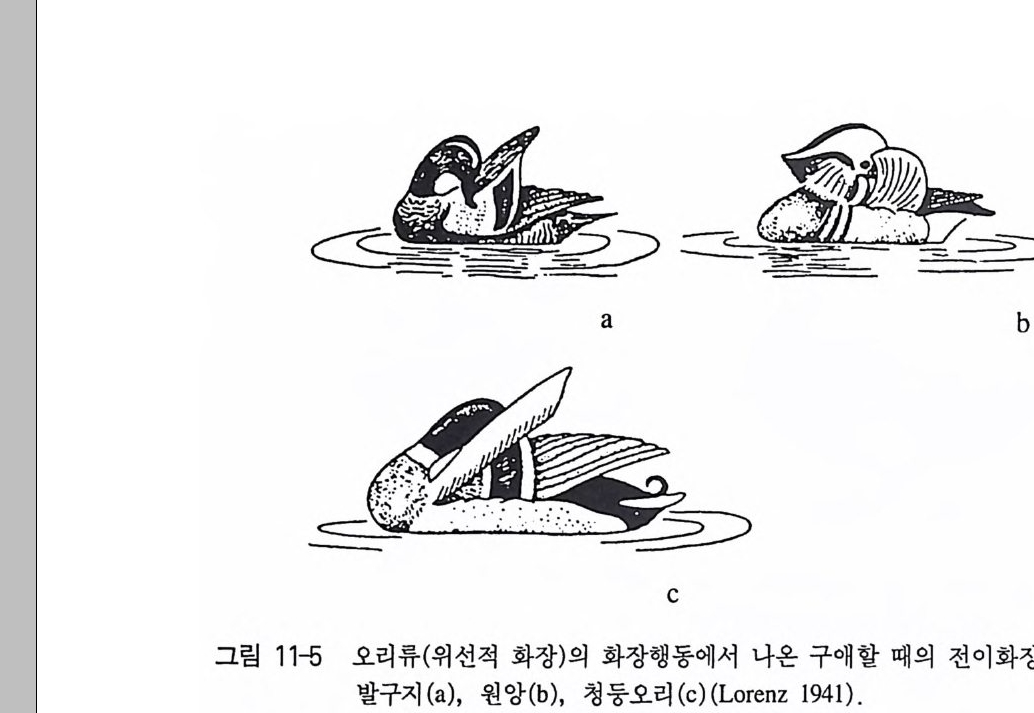 ‘三〈-
‘三〈-
협행동에서 넘어온 것으로 생각된다. 즉 이들 행동에 내재하고 있는 반발적인 동인의 결과로 이 두 행동영역에서 특히 많은 전이동작들 이 나타나고 있는 데서 근거를 찾아볼 수 있다. 이런 면에 대해 가 장 일찍이 연구된 예 가운데 하나는 화장 영역에서 나온 여러 종류 의 오리류의 구애행동 때 〈가짜 화장p seudo-clean i n g〉울 들 수 있다 (그림 11-5, Lorenz 1941 ) . 행동의 형태변화와 기능변화 의에도 의식화가 되는 동안에 동인에 서도 하나의 변화가 나타날 수 있다. 죽 단풍새류의 구애에서 둥지 짓기 의도동작은 더 이상 둥지짓기 의도가 없고 성적인 동인으로 대 체되었으며 따라서 이것은 구애행위의 고정된 구성 성분인 의식화된 화장동작에 해당된다.
7 행동의 수렵 지금까지 취급해 온 계통발생학적 행동발달의 형태는 집단의 계통 발생학적 문제와 관련되어 있으며 또 상동의 의미를 지닌 계통발생 학적 혈연관계에 의해서 모든 개연성으로 볼 수 있는 형질과도 각각 관련되어 있다. 발달 가능성에서 마찬가지로 홍미를 끄는 측면은 행 동의 수렴 behavio r nl convergence 에 대 한 발달사가 된다. 우선 수렴에 대한 개념과 이 수렴과 상동을 구별하는 것이 필요하 댜 상동은 공통의 정보저장소의 존재와 관련되어 있다. 이것은 특 수한 환경 여건과 경쟁하기 전에 한 세대에서 이런 환경에 적응할 수 있는 다음 세대로의 정보전달을 보장하고 있다. 이런 환경의 정 보는 역시 간접적이다 . 이에 반해 수렴은 정보가 직접 획득되어 진다. 같은 혹은 비슷한 환경조건으로 인해 다양한 개체나 종들의 특정 형질들이 서로 독립 적으로 즉 공통의 정보저장소가 없이 형성되어 이런 환경여건에 이 들 개체나 종들이 서로 항상 더 비슷하게 적응될 때 수렴이라고 부 론댜 형태학 분야에 대한 예로써 지느러미가 2 차적으로 물속 생활 에 적응된 척추동물은 파충류(어룡), 조류(펭귄) 그리고 포유류(물 개, 고래)가 있다. 이들은 척추동물의 다리로는 상동이지만 지느러미 로는 물고기와는 상사 그리고 수렴적 발달에 대한 이유에서 매우 비 슷하게 형성된 것이다. 행동에 있어서의 수렴은 종내에서뿐만 아니라 종간에서도 발달되 었으며 생득적 형질뿐만 아니라 획득된 형질에서도 나타나고 있다. 만일 한 마리의 어린 개체가 특정 행동 즉 적을 피하거나 먹이를 취 하는 것을 전수에 의해 배우게 된다면 모든 동물들의 그런 행동은 상동이 되며 그 행동은 관련 물체에 대한 자신의 경험과는 무관하게 발달되어 왔다. 예를 들어, 그 동물은 어떤 먹이동물이 먹을 수 있
는 건지 먹을 수 없는 것인지를 매번 그것을 직접 먹어 보지 않고도 동료들의 반응을 보고도 알 수 있다. 이에 반해 각 개체들이 이런 경험들을 스스로 직접 터득해야 될 때 관련행동은 수렴되어 일어난 댜 이와 같은 것이 종간에서도 일어난다. 두 종이 가까운 혈연관계 로 인해 같은 행동을 갖고 있거나 한 종이 다른 종으로부터 배워서 익혔다면 그것은 상동이 되며, 만일 그것이 독립적으로 형성되어 왔 다면 그것은 수렴적 발달에 근거하고 있다. 예를 들어, 여기서 상동 이란 여러 종들의 벌새의 정지비행이고 수렴은 벌새의 정지비행과 꿀을 먹는 그밖의 조류나 물을 먹는 동물들의 정지비행이 된다. 이 들의 비행은 벌새와 비슷하지만 완전히 고도로 발달하지는 못했다. 이 세 집단에서 꽃 위에서 머무는 비행 능력은 먹이를 취하기 위해 꽃 방문의 적응에 있어서 서로가 독립적으로 발달해 온 것이다. 수렴적 발달은 같은 생활조건에 같은 방향성 적응에서 항상 기인 한다 그래서 대개의 지상성 조류들은 수상성 조류들이 대부분 껑충 껑충 뛰어서 이동하는 계통분류학적 자세와는 달리 걷듯이 이동한 댜 알굴리기는 지상성 번식 조류의 경우 가장 다양한 근연집단(타조 류, 오리류, 닭류, 섭금류)에서 발달되어 왔으며, 확실히 자신의 둥지 에서 알이 없어지는 것에 대한 적응과 그래서 알을 멀리서 둥지로 끌어들여야 하는 위험을 내포하고 있다. 몇몇 오스트레일리아산 단 풍새류들은 비둘기처럼 물을 빨아 마시고, 일부는 닭처럼 물을 찍어 마신댜 이 두 분류의 비교를 통해 시간절약성 빨기가 특히 건조한 생활 환경에 사는 종들의 여러 근연집단에서 서로 독립적으로 발달 해 온 것임을 알 수 있다. 반면 습지의 생활 환경에 사는 근연종들 은 물을 찍어 먹는다. 분명히 빨아서 물 마시는 방법은 앞이 툭 트 인 환경에서 몸을 드러내 놓음으로써 포식자가 쉽게 접근 가능한 물 마시는 장소에서 머무는 시간을 줄이고자 하는 적응의 표현으로 볼 수 있다 (Cade 둥 1966, Goodwi n 196, Immelmann 과 Immelmann 1967) .
수렴적 행동발달에 대한 좋은 예는 이미 언급된 작은 조류의 경계음 으로, 이들의 소리는 서로가 비슷해서 종간 특이적 신호로 작용할 수있다. 행동 수렴, 생활공간 및 생활 형태의 전형적인 행동에 대한 연구 는 동물종들 혹은 근연집단의 적응 가능성에 대해 일반화할 수 있으 며 따라서 진화 연구에도 중요한 기여를 할 수 있다. 8 행동의 공전화 행동학에서 포식자와 피식자 간의 행동은 일종의 두 종간의 경쟁 을 통해 진화되어 왔다. 종족 보존을 위해 피식자는 포식자에게 먹 히지 않아야 하며 포식자는 피식자를 잡아먹어야 한다. 이런 상호 경쟁을 통해 두 종은 함께 진화해 왔다. 우리는 이것을 공진화(共進 化 co-evolu ti on) 라고 부른다. 이러한 공진화는 뻐꾸기와 숙주새에서도 잘 나타나 있다 . 뻐꾸기는 자신의 알을 부화시키기 위해 숙주새를 속여야 하며 숙주새는 뻐꾸기에 속지 않기 위해 알의 식별력을 발달 시켜 왔다. 그럼 동물들의 공진화를 다음과 같이 포식자와 피식자 간의 경쟁 그리고 기생새와 숙주새 간의 경쟁으로 나누어 살펴보기 로하자 . 8. l 포식자와 피식자 간의 경쟁 종간 경쟁은 포식자와 피식자 간의 경쟁에서 비롯된다. 포식자는 생존을 위해 먹이를 잡고 피식자는 먹이로 희생당하지 않기 위해서 스스로 몸을 지키지 않으면 안 된다. 그래서 동물들의 먹이 잡는 기 술도 아주 다양하다. 또 자기 몸을 지키기 위한 전략을 갖는다. 이
러한 기술과 전략은 상호 경쟁을 통해 오늘날까지 진화되어 왔다. 왜 포식자는 자신들의 포식자를 멸종으로 몰고가지도 않고 또 피 식자는 스스로 멸종되지 않고 진화하였는가 ? 이에 대한 가설로 첫 째, 신중한 포식자는 피식자를 멸종시키거나 과잉번식하는 것을 피 한다 이것은 군선택 gro up selec ti on 에 의해서 고려될 수 있다 . 신중 한 포식자의 집단에서 이기적으로 다른 개체보다 더 많이 먹는 어떤 개체가 있다고 하자 . 이 개체는 신중한 개체보다는 장래의 세대에 더 많은 유전자를 남길 수 있다. 그러나 신중한 포식은 한 개체가 자원을 독점적으로 사용하는 집단의 이익보다는 개안의 미래울 위해 식량을 비축하는 쪽으로 진화할 수 있다. 두번째 가설로 만일 집단 멸종이 일반적일 경우, 자연에서 안정된 포식자-피식자 체계가 있 다면 불안정한 모든 체계는 정말 멸종되었을 것이다. 세번째 가설은 피식자는 군비 경쟁에서 앞선다는 것이다. 포식자-피식자 체계는 피식자가 군비 경쟁에서 항상 한 발짝 앞서기 때문에 안정적이다. 토끼는 여우보다 더 빨리 달린다. 왜냐하면 토끼는 자신의 생존을 위해 달리는 반면 여우는 단지 한끼의 식사를 위해 달리고 있기 때 문이댜 즉 여우는 토끼와의 경쟁에서 실패해도 번식하지만 여우와 의 경쟁에서 실패한 토끼는 번식에 실패하여 결국 죽고 만다. 그래 서 선택압은 여우가 성공적으로 사냥하는 능력보다 토끼가 도망가는 능력의 향상 쪽으로 작용해 왔다. 많은 경우 피식자는 포식자보다 세대가 더 짧기 때문에 더 빠른 속도로 진화할 수 있다 . 그래서 피식자는 군비 경쟁에서 한 걸음 앞 설 수 있다. 이것은 족제비-쥐, 새-곤충과 같은 경쟁의 예가 있 댜 그러나 가끔은 포식자가 더 빨리 번식할 수 있는 능력을 갖기도 한다(진딧물-장미). 왜 피식자는 자신의 포식자를 멸종시킬 정도의 강력한 도피를 발달시키지 않았는가? 이에 대한 가설로 피식자의 능 력이 향상하여 포식자의 숫자가 줄어들기 때문에 그 포식자들은 피
식자의 선택에 좀 더 향상된 노력을 기울이지 않게 된다는 것이다. 포식자는 피식자를 멸종에 이르도록 하지 않는 또 한 가지 가설은 피식자가 줄어들면 포식자는 다른 먹이를 찾아야만 한다. 그래서 여 러 종류의 먹이에 대한 포식의 적응이 보통 다르기 때문에 포식자는 어떤 한 종만을 멸종에 이르도록 국한하지 않는다 . 8.2 기생새와 숙주새의 공진화 우리 나라에 번식하는 뻐꾸기 종류는 자신의 알과 유사한 알을 갖 고 있는 숙주새에 탁란을 한다. 뻐꾸기는 같은 종내에서도 다양한 색깔의 알을 낳는다. 같은 뻐꾸기라도 어떤 놈들은 휘파람새의 둥지 를 찾아 알을 낳는 놈들이 있는가 하면 어떤 놈은 개개비의 둥지를 찾아 알을 낳는 놈들이 있다. 그러나 어떻게 해서 같은 종의 뻐꾸기 들이 자신들의 알에 맞추어 숙주를 선택하고 있는지는 아직도 잘 알 려져 있지 않았다. 다만 하나의 가능성은 뻐꾸기 새끼(암컷)가 각인 을 통해서 숙주의 특징을 학습하여 자신의 어미와 같은 형태의 알을 낳으면 그 알과 어울리는 숙주새에 탁란하게 된다는 것이다. 뻐꾸기 암컷은 숙주새의 둥지에 탁란을 할 때 특별한 절차를 밟는 댜 먼저 뻐꾸기 암컷은 숙주새가 둥지 짓는 것을 지켜 봄으로써 둥 지를 찾게 된다. 그런 다음 숙주새가 알을 낳기 시작할 때까지 기다 린다. 뻐꾸기 암컷은 숙주새가 알을 낳는 기간을 이용해 오후 시간 대에 숙주새 둥지 하나에 한 개의 알을 낳는다. 알을 낳기 전 뻐꾸 기 암컷은 숙주새의 둥지 가까운 곳에 위치한 나뭇가지에서 조용히 기다린다 갑자기 둥지로 내려 앉아 숙주새의 알 하나를 부리로 물 고 자신의 알 하나를 낳는다. 그리고 날아가서 훔친 알을 먹어치운 다. 숙주새의 둥지에서 일을 치르는 시간은 10 초도 채 못된다. 뻐꾸 기 암컷에 의해 맡겨진 알은 이젠 숙주새가 돌보게 된다. 숙주새는
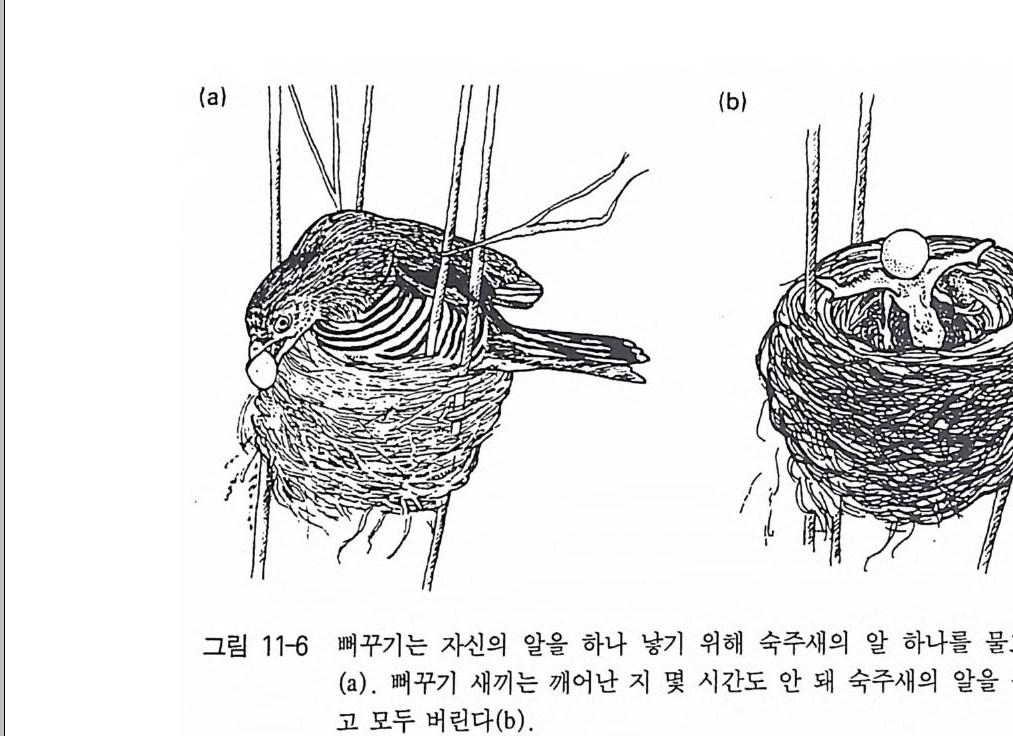 (a) (bl
(a) (bl
가끔 뻐꾸기 알을 거절하지만, 많은 경우가 그것을 받아들인다. 보 통 숙주새의 알보다 먼저 깨어나온 뻐꾸기 새끼는 부화한 지 몇 시 간 만에 숙주새의 알을 등에 업고 하니씩 둥지밖으로 버린다(그림 11-6). 결국 뻐꾸기 새끼는 혼자서 숙주새의 둥지를 점령하고 숙주 새의 헌신적인 보살핌에 의해 길러진다. 기생새와 숙주새는 서로 선택압에 반응하여 어느 정도까지 진화되 어 왔는가? 이것은 다양한 색깔의 모형 알에 대한 숙주새의 반응을 통해 실험적으로 연구되었다. 뻐꾸기가 숙주새로부터 선택되기 위해 행동한다는 증거가 다음과 같이 제시되었다. 첫째, 뻐꾸기가 알을 낳는 순서는 숙주새의 방어를 앞서도록 특별히 고안되었다. 모형 뻐 꾸기 알에 대한 숙주새 둥지의 탁란 실험은 모형 알이 숙주새 자신 의 알과 잘 조화되지 않거나, 너무 일찍 알을 낳게 되면 숙주새에 의해 거절하는 것으로 밝혀졌다 (Dav i es 와 Brooke 1988). 그러므로 뻐
꾸기의 알 모방과 산란의 타이밍은 탁란의 성공을 증가시키기 위해 적응되어 왔다. 둘째, 다양한 뻐꾸기 종에서 나타나는 숙주 알의 모 방 정도는 그들의 숙주새에 의해 구별되는 정도를 반영해 주고 있 다. 예를 들어 유럽억새풀새에 탁란울 하는 뻐꾸기는 유럽억새풀새 가 다른 대부분의 숙주새와는 달리 그들의 둥지에서 낯선 알을 버리 지 않기 때문에 모방 알을 낳지 않는다. 숙주새 역시 뻐꾸기 알을 무조건 받아들이지는 않는다. 그 증거로 는 첫째, 뻐꾸기가 선호하는 숙주새들은 낮선 알을 버리는 습성 및 뻐꾸기에 대해 강하게 반발하는 행동은 뻐꾸기 탁란에 대한 반응으 로 특별히 진화되어 왔다. 그러나 종자를 먹거나 나무 구멍에 둥지 를 트는 종들은 뻐꾸기 탁란 대상 숙주새로 적합하지 않다 . 이 종들 은 모두가 자신이 낳은 알과 낯선 알을 식별하지도 못할 뿐만 아니 라 뻐꾸기가 선호하는 숙주새들과는 달리 둥지 근처에 성체 뻐꾸기 가 와도 공격하지도 않는다 (Moksnes 등 1991 ). 둘째, 뻐꾸기로부터 격리되어 있는 숙주종의 군집은 비모방성 알을 거의 버리지 않는다 (Soler 와 Moller 1990). 예를 들면 뻐꾸기가 살지 않는 아이슬란드에 서 서식하는 유럽밭종다리 군집은 영국에 서식하는 유럽밭종다리 군 집보다 알 식별 능력이 떨어진다 (Dav i es 와 Brooke 1989). 데이비스 Dav i es 와 브룩 Brooke (l 991) 은 오랜 야외 실험 결과 영국 에서 숙주새의 둥지가 뻐꾸기에 의해 탁란되는 비율은 유럽밭종다리 Anth u s p ra t ens i s 의 둥지 가 2.7 % , 유럽 개 개 비 Acrocep h alus scir pa ceus 의 둥지 가 5.5 %, 알락할미새 Mota c il la a lba 둥지 가 4%, 그리 고 유럽 억 새 풀새 Prune /l a modular i s 의 둥지 가 1. 9% 라고 조사하였다. 유럽 억 새풀새를 제외하고는 모두 뻐꾸기의 알이 숙주새의 알과 비슷한 것 에 비해 유럽억새풀새에 탁란을 하는 뻐꾸기의 알은 이 유럽억새풀 새의 알과 현저한 대조를 이루고 있었다. 그럼에도 불구하고 유럽억 새풀새가 뻐꾸기의 알을 구별해 내지 못하는 것은 유럽억새풀새의
둥지가 최근에 뻐꾸기에 의해 탁란의 대상으로 선택되었기 때문인 것으로 추측하고 있댜 그렇다면 거의 알을 버리지 않는 것으로 보 이는 종들(예, 유럽억새풀새)은 아직 뻐꾸기의 탁란에 대해 역적응을 진화시킬 만한 충분한 시간을 갖지 못한 최근의 숙주새인 반면, 강 하게 알을 거부하는 종들(예, 유럽개개비, 알락할미새)은 상호 경쟁의 후기에 있는 오래된 숙주새들일 가능성이 크다. 결국 기생새는 숙주 새를 속이기 위한 전략으로 진화되어 왔으며, 이러한 전략은 기생새 로 하여금 새로운 숙주새를 선택하도록 진화될 수 있다. 이에 비해 숙주새들은 기생새로 부터 속지 않기 위해 알 식별 능력을 증진하여 왔다. 9 행동의 가축화 계통발생이 종의 변화로 이어지는 과정과 진화에 대한 이해를 돕 는 데 가축화 현상은 한 가지 모델이 되고 있다. 이러한 과정을 거 쳐온 동물(그리고 식물)을 가축화되었다고 부른다. 가축들은 구조 적, 생리학적 그리고 행동학적 특징에 있어서 야생 고유의 형태와 차이가 나타난다. 이러한 특징들은 가축화 형질이라 한다. 고유 형 태와의 차이가 유전자 상태로 고정되어 있고 다음 세대로 이어질 때 진정한 가축화 형질이라고 정의할 수 있다. 그러나 여기서 실제 가 축화의 한계를 규정짓는 데 자주 어려움이 발생하고 있다. 많은 구 조적 및 생리학적 특징의 형성이 많은 경우 환경요인에 의해서도 이 루어지기 때문에 사육조건에 의한 변화된 환경은 진정한 가축화 형 질들을 축적시키거나 이런 가축 현상으로 나타나게 수정시킨다. 그 리고 이런 수정은 야생동물을 사육했을 경우-즉 어려서부터 사 육했을 경우――일어날 수 있다. 여기서 호르몬 분비의 변화 혹은
자극이 빈약한 환경에서의 갑각기관과 중추신경계의 축소를 가져온 다 (Herr 와 Rohrs 1973, Nach t sche i m 과 Ste n g el 1977, Sossin k a 1982). 이런 수정적 축적이 보다 많아지면 많아질수록, 행동영역에서 가 축화 현상이 진짜 일어났는지는 알 수가 없다. 이러한 이유로 인해 행동학적 가축화 연구는 아직 시작 단계에 머물러 있으며 지금까지 이에 대한 많은 연구가 일반화되고 있지는 못하다. 그럼에도 불구하 고 이런 분야가 오랜 시간 동안 큰 홍미를 끌어 왔다. 그것은 인간 역시 스스로 가축화되어 있으며 그래서 동물 행동에서 가축화에 대 한 설명으로 가능한 인간에게도 같은 현상에 대한 증거를 기대하기 때문인 것으로 보인다 (Lorenz 1940, Sauer 1966, Sossin k a 1970). 10 가축화 형질 가축화된 동물들이 그들의 조상형과 비교해서 구별되는 변화는 다 음 두 가지 일반 규칙에 있다. 한 가지는 가축화된 동물들이 야생형 에 비해 그들의 구조와 생리 그리고 행동 특성에 있어서 커다란 종 내의 근본적인 변화로 인해 특징지어진다. 다른 하나는 전형적인 많 은 가축화 특징들의 경우에 눈에 띄는 수렴이 나타난다. 물론 이 수 렴온 근연이 아닌 종들의 형태에서 종종 관찰될 수 있다. 그래서 여 러 가축화된 종에서 매우 비슷한 특징적 형태 변화가 존재한다. 개, 돼지 그리고 염소에서 곱슬거리는 털과 몸이 길고 짧은 다리가 이 에 대한 예가 된다 (Herre 1980, Herre 과 Rohrs 1973). 특히 행동과 관련되어 있는 중추신경계도 같은 변화를 보여 준다. 이런 변화는 뇌 크기의 일반적 감소 외에도 계통발생학적으로 가장 최근에 발달된 뇌 부위의 무게 및 분화의 감소를 가져왔다. 많은 경 우에서 이런 특징들의 변화가 매우 두드러져서 가축화 조건 변화의
고유한 규칙성에 대한 추측으로 받아들여지고 있다 (Kruska 1970). 가축화된 동물들의 행동은 야생형의 행동과 구분될 수 있으며―― 물론 아주 적은 정도이긴 하지만_또 많은 곳에서는 일치하기도 한댜 K. Lorenz 는 이미 오래전에 가축화 조건이 되는 행동 변화의 몇몇 규칙성에 대해 주목을 했다. 이들 가운데 가장 중요한 것을 행 동준비(충동의 높이)의 변화, 즉 행동양식의 활동 특이적 에너지 상 승과 감소, 그리고 생득적 해발기구의 변화로 보았다 (Lorenz 1940). 10. l 행동준비의 변화 가축의 많은 행동양식들은 야생형보다 분명히 더 빈도가 낮든지 혹은 더 빈도가 높게 나타난다. 더 빈도가 낮게 나타나는 현상을 위 축 a tr oph y으로 표현할 수 있는데 이런 것에는 공격, 경계, 도피, 방 어, 새끼돌보기 그리고 보통 자발적으로 이동하는 활동들이 속한다 (그림 11-7). 극단적인 경우에 어떤 행동양식들은 완전히 사라져 버 릴 수가 있다. 이런 위축은 오랫동안 가둬 기른 카나리아의 노랫소 리에서 증명된 바 있다. 이들의 노래가 야생 카나리아의 노래보다 한두 개의 음절이 적다 (N i col ai 1959) . 이 에 비해 비 대 h yp e rtr o p h y로 표현되 는 변화는 무엇보다 성 적 행 동 영역에서 확인되었다. 이것은 보통 성적 반응의 빈도 상승으로 야생형의 성행동과 비교해서 특정 연주기와 관계 없이 일어나고 있 댜 특정 연주기는 완전히 무관한 연주기로 발전될 수 있으며, 그것 온 확실히 높은 성적 동인으로 이런 행동양식들의 빈도가 증가한 데 서 그 원인이 있다. 그 밖에도 이런 변화는 짝짓기 행동의 전체 과정의 이동에서 찾아 볼 수 있다. 야생형에서 구애연쇄 가운데 아주 중요한 부분을 하게 하는 도입 요소들이 가축에서 시간적으로 매우 단축되었거나 완전히
 5000-, 매 12 시간 동안의 비행
5000-, 매 12 시간 동안의 비행
없어져 전체 짝짓기 행동이 교미 자체가 된다. 이러한 부분 행위의 분화된 빈도 변화는 가축들에서 정상적으로 도입되는 혹은 뒤에 연 결되는 요소가 없이 행동연쇄의 각 부분들 가운데 하나만이-고 양이나 개는 먹이를 먹기 전 먹이동물을 혼들어 기절시키는 행동의 도입이 없어진 것처럼――일어날 수 있다.
10. 2 생득적 해발기구의 변화 야생 동물들은 사회적 영역에서 아주 특정한 자극 형태에 규칙적 으로 매우 민감하게 반응을 하며, 그리고 긴 행동의 연쇄 과정에서 다음에 연결된 부분 행위의 출현은 관련 해발인의 존재 여부에 달려 있다 가축에서는 이런 선택이 종종 심하게 축소되어 버렸다 . 그것 온 첫째 성적 반응과 새끼돌보기나 사회행동의 영역에서 일어나고 있다 이러한 이유에서 교잡이나 대리모에 의한 사육이 야생 동물보 다 훨씬 쉽다. 그래서 닭의 조상인 붉은야생닭은 자신의 새끼만을 알아 볼 수 있는 머리 표시에 의해 자신의 새끼를 데리고 다니는 반 면, 가금의 암닭은 자신과 근친이 아닌 다른 종의 새끼들도 받아들 인댜 이런 행동은 가금의 품종에 넓게 퍼져 있다 . 물론 여기서 사 회적 경험의 정도를 고려한 비교 및 정량적 연구가 지금까지 이루어 져 있지 않다는 사실을 지적해 두고 싶다 . 적어도 그것은 생득적 해 발 기구로 취급될 것인가 아니면 이렇게 빈약해진 선택력이 축소된 학습능력의 결과로 표현될 것인가는 앞으로 설명이 될 수 있어야 할 것이다. 가축화 조건적인 행동변화에 대한 행동준비와 생득적 해발기구의 에도 거의 모든 행동 분야에서 많은 개별적 현상들이 나타나 있다. 특히 학습행동의 전체 분야가 일반적으로 관련되어 있다. 여기서 많 은 가축화된 동물들은 두드러진 학습능력의 축소를 보여주고 있다. 가축화 과정에서 행동의 변화에 대해 다음과 같이 설명될 수 있 댜 많은 경우에 하나의 새로운 행동양식이 생겨났으며(그림 11-8) 일부 경우에서는 야생동물의 행동 목록에서 행동의 하나가 완전히 사라지기도 했다. 생득적 행동양식의 과정도, 즉 유전적으로 고정된 형태는 일반적으로 유지되며, 그 과정이 변화되는 경우에는 이 변화 가 항상 단순화 다시 말해 분화의 소실 성격을 갖고 있지만, 긍정적
 그림 11-8 가축화 과정에서一 새로운 행동양식이 생겨나는 것에 대한 몇몇 예들 가
그림 11-8 가축화 과정에서一 새로운 행동양식이 생겨나는 것에 대한 몇몇 예들 가
인 새로운 발생의 성격은 갖고 있지 않다 . 그래서 대부분의 가축화 조건 행동 변화는 단순히 정량적 성격을 띠며 이미 언급된 빈도와 선택력의 변화에서 생겨난다. 이로써 가~홉陸] 과정은 정량적 변화가 일어날 수 있는 많은 효과에 대한 좋은 예를 제공해 주고 있다.
제 12 장 행동학과 심리학 과학적으로 ` 동물 행동에 대한 연구가 시작되면서부터 이에 대한 지식이 인간에게도 적용될 수 있는지가 관심의 대상이 되어 왔다 . 이런 주제에 대한 논의는 항상 오해의 소지를 불러일으켜 왔다. 이 런 논의는 동물행동학 밖에서도 있었지만, 행동학 안에서도 매우 다 양하게 논의가 되어 왔다. 이런 논의는 동물행동학적 결과와 인간심 리학에서의 원리를 그대로 적용 설명하는 것에서부터 이런 비교 자 체를 전면 부인하는 것에 이르고 있다. 전자의 생각은 인간의 계통 발생학적 기원이 인간이 아닌 조상에서 출발했다는 명백한 사실에 근거하여 많은 신체의 형태와 기능에서 일치 내지는 유사함에 기인 하고 있다. 이에 대해 완전히 상반된 생각은 인간은 그의 행동에 있 어서 인간이 아닌 생물체와는 거리가 먼 종에서 진화되어 왔다는 사 실을 인정하려 하지 않는다 . 그래서 이런 비교가 허락될 수 없거나 최소한 이론적으로나 실제적으로 사실을 인정할 수 없다는 견해에 뿌리를 두고 있다.
] 두 분야의 비교 이런 관점에서 이미 언급된 다양한 생각들 가운데 하나는 행동학 적 지식을 적용하는 데 고려될 만한 측면들을 매우 다양하게 찾으려 고 하는 데 있다. 물론 한 종에서 얻은 결과를 인간에 적용할 수는 없댜 마치 그것은 각 동물들의 종마다 행동 목록의 특성이 다르기 때문에 각각의 종에다 직접 적용할 수 없는 것과 같다. 그러나 행동 학은 비교과학으로서 (인간)심리학보다는 더 큰 방법적 이점을 지니 고 있을 뿐만 아니라 여러 종들 혹은 종들의 집단을 연구하고 비교 하여 일정한 공통점, 예를 들어 여러 행동영역들의 발달에 있어서 특정 동물 집단을 특징지울 수 있는 규칙성을 밝힐 수 있는 가능성 이 개방되어 있다 . 개개의 근연종의 수준에서 시작한 이런 비교는 어느 정도 충분히 관련 정보가 이용될 수 있는 한에서 점차로 높은 체계적 단위(과, 목, 강)로 확대될 수 있으며 구세계원숭이의 공통의 행동 특성, 즉 영장류를 일반화시키거나 혹은 좀더 확장시켜 포유류 전체의 특성을 발견하는 데 시도될 수 있다. 인간도 동물로 분류한 분류체계(영장류, 포유류)측면에서 보면 비 교는 근본적으로 오히려 가능하며 허용될 수 있다. 그러나 여기서도 인간은 직접적 적용의 형태로 인정되지 못한다. 오히려 동물행동학 에서 얻어진 지식이 항상 가설로만 인정되고 있다. 물론 이 가설은 동물과 비슷한 현상을 인간 행동에서 찾을 수 있도록 심리학에 자극 을줄수 있다. 비교하는 것과 관련된 이런 한계 의에도 심리학 내의 여러 세부 분야에 관련해서도 차이가 발생한다. 어쩌면 생리학적 기초와 아주 긴밀한 행동 특성에서 동물 행동과 가장 먼저 일치하는 것으로 볼 수 있댜 그러므로 이런 견해가 일반적으로 행동발달의 초기에 가장 큰 것으로 추측될 수 있다. 왜냐하면 발생 초기의 생물학적 유사성
이 완전히 성장한 생물체에서의 유사성보다 일반적으로 항상 많기 때문이다. 그래서 비교는 무엇보다 생리심리학(감각심리학, 신경학 ) , 임 상심리학(재활심리학, 약물학) 그리고 발달심리 및 학습심리학에서 가 능성을 찾을 수 있다. 이에 비해 인간심리학의 다른 영역들은 매우 〈전형적 인간적인〉, 죽 생물학적 기초에서 매우 동떨어져 발달해 온, 말하자면 이러한 기초를 더 이상 인정할 수 없기 때문에 최소한 의 비교도 허용되지 않는다. 이런 것에는 개인심리학, 조직심리학, 분 석심리학, 검사심리학 그리고 언어심리학을 들 수 있다. 2 응용범위 만일 이미 지적된 면에서 그리고 이미 언급된 한계들로 동물-인 간 비교가 가능하다고 인정된다면, 동물과 인간의 비교를 가능하게 하는 지식의 이론적 발전과 이해 외에도 실질적으로 가능한 결과에 대한 질문도 관심을 갖게 된다 . 동물과 인간의 비교는 다음 두 영역 내에 있다. 하나의 영역은 동물에 대한 정량적, 실험적 연구의 수행 이고 다른 하나의 영역은 행동학적 연구방법을 인간에게 적용하는 데 있다. 동물에 대한 연구의 방법적 이점들을 이용하는 전자의 영 역에서, 이 이점들을 대표하는 질문이 연구되었다. 이런 질문의 대 답은 가능한 한 인간의 행동의 더 발전된 이해에도 기여할 수 있다. 그러나 인간에 대한 행동학적 연구 가능성은 윤리적 이유에서 한계 가 있다 (Cranach 등 l
 그림 12-1 어린 히말라야원숭이가 두 개의 철 망 모형에 의해 길러졌다. 그 중에
그림 12-1 어린 히말라야원숭이가 두 개의 철 망 모형에 의해 길러졌다. 그 중에
대한 연구가 되고 있댜 이 연구는 할로우H.F. Harlow 가 히말라야원 숭이 Macaca mula tt a 를 대상으로 시 작하였으며 그 동안 여 러 종에서 많은 연구가 계속되었다 (Harlow 1958, Harlow 와 Harlow 1962, Suom i 등 1973). 할로우는 나이가 다른 여러 어린 히말라야원숭이들을 기간을 달리해서 어미와 격리시켜 놓은 다음 그들을 다시 철망으로 만든 어 미 모형과 접촉을 하게 했다(그림 12-1) . 이 실험의 결과 다음의 사 실이 알려지게 되었다.
® 어미와 새끼 간의 결합은 장차의 사회화 과정을 위한 전제 조건이 된다 이 결합이 결여되면 심각한 발달장애를 가져올 수 있다. ® 어미로부터 일시적 격리에 따른 결과는 나이에 따라 상당한 차이가 생긴다 ® 결합이 이루어지기 위해서는 긴밀한 육체적 접촉을 통한 보상이 먹 이 (젖)를 통한 보상보다 더 결정적인 작용을 한다. 설사 각인 과정의 민감기를 기억하는 가장 민감한 연령층의 상태 나 종들마다 격리로 인한 장애 정도가 매우 많은 차이가 있을 수 있 다고 하더라도 비교 연구는 영장류로부터 이미 얻어낸 원리들이 일 반화되어 있는 실정이다(그림 12-2, Harlow 1958, Harlow 와 Harlow 1962) . 지금까지 인간을 제외한 영장류에 대한 이런 연구는 인간 발달심 리학적 연구에도 적잖은 영향을 미쳐 왔다. 이것은 이른 시기에 엄 마와의 격리로 인한 결과가 행동에서 전형적인 발달장애인 애정 및 정 서결핍증, 즉 고아로 자라난 아이의 정신장애증으로 나타난다 (Bl urt on- Jon es 1972, Blurto n -Jo h es 1976, Bowlby 1973, Bowlby 1975, Scha ffer 1977) . 이미 발견된 행동 특성, 발달기의 특이성와 초기 경험적 영향의 지속성에 있어서 위에서 이미 언급된 행동의 일반 규칙성에 대한 예 가 다루어지고 있다. 확실히 이런 행동의 규칙성에 대한 연구가 인 간심리학에도 실마리를 제공할 수 있다. 인간과 가까운 동물이 아 닌, 즉 영장류에 속하지 않는 동물에 대한 연구에서 결과는 좀 더 일반화될 수 있어야 하며 단지 좀더 신중하게 적용될 수 있어야 할 것이다 그럼에도 불구하고 이런 결과들은 제각기 인간과의 비슷한 과정에 대한 좀더 나은 이해 쪽으로 혹은 새로운 연구방법 상의 개 발 쪽으로 최소한의 사고를 불러일으켜 왔다. 이에 대한 예로 소리의 발달, 놀이행동, 사회적 장애(예, 근친상간회피), 혹은 영아살해와 같
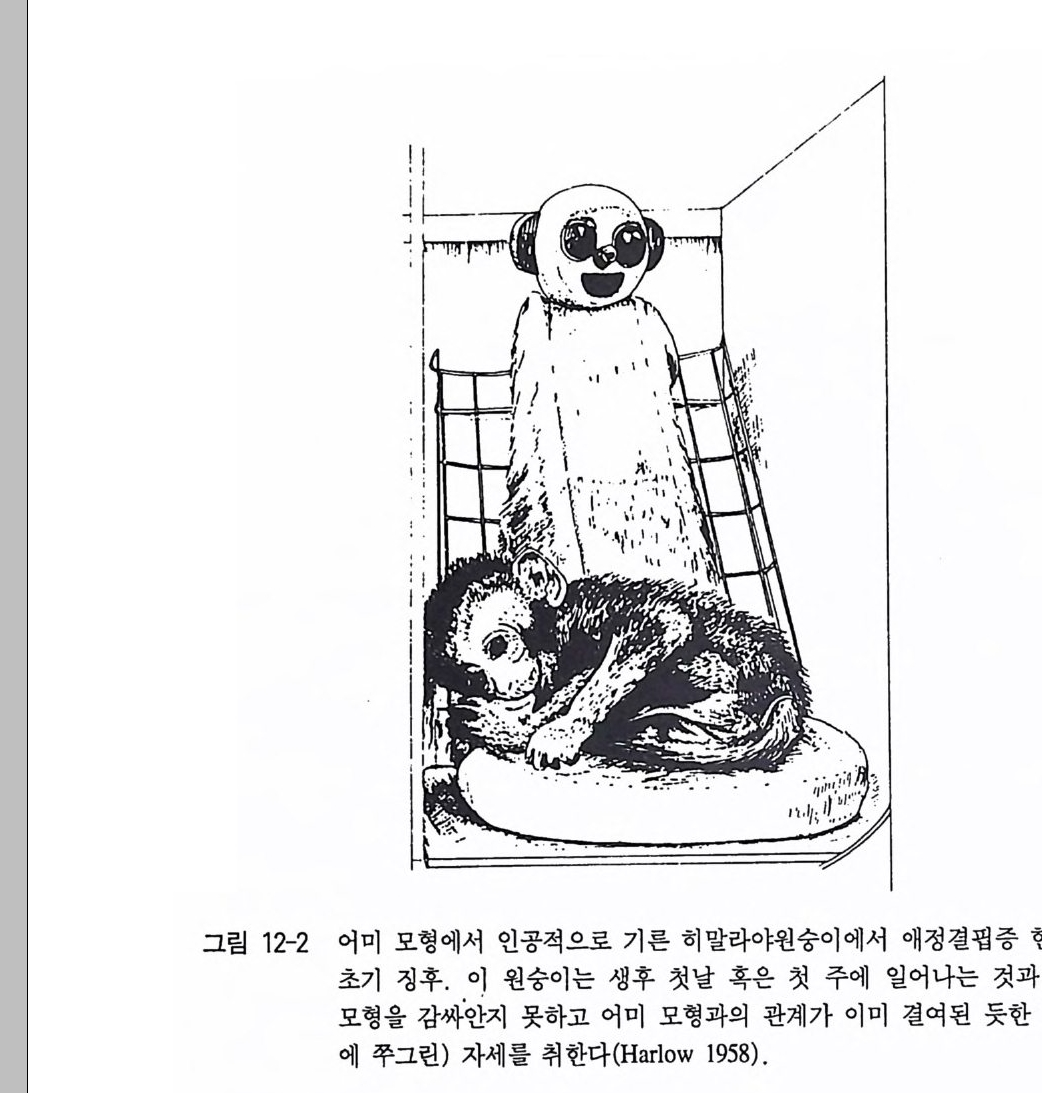 /,
/,
온 여 러 종류의 연구 대상들을 들 수 있다 (B i scho f 1972, Lenneberg 1973) . 이미 앞의 내용에서 이야기한 비교 연구처럼 이런 실험에서 종의 올바른 선택도 아주 중요하다 (Harlow 등 l'l7 2 ). 여기서 인간과 근친 인 영장류가 가장 적합하지만, 이런 동물군 내에서도 차이가 있게 마련이댜 그래서 침팬지는 오늘날 생존하고 있는 동물들 가운데 인
간 다음에 위치해 있는 종으로 많은 학습능력에 대한 연구에 가장 적합한 것으로 증명되어 왔다 . 반면 침팬지는 사회구조에 대한 연구 에는 적합지 못한 것으로 나타났다. 왜냐하면 이들은 인간의 사회구 조에 비해 사회체계의 형성에서 거의 인간과 모순된 행동을 하기 때 문이다 . 침팬지말고도 남미산 마모셋 Call ithri c i dae 은 사회구조에 대 한 좀 더 나온 예가 되고 있다. 이 예는 근연 정도에 있어서 인간과 는 아주 가깝지 못하지만 사회체계에 있어서는, 영장류 가운데 예외 적으로 지속적인 일부일처제를 유지하면서 어린 개체를 돌보는 데 부친도 참여하는 데서, 인간과 좀 더 일치하는 점이 많다. 만일 우 리가 인간의 언어 발달에 관해서 발성의 발달에 관한 규칙성을 연구 하려고 한다면(예, 민감기의 존재 여부), 포유류 이외의 동물, 무엇보 다 명금류가 최적의 연구 대상 동물임을 깨닫게 된다. 2. 2 인간행동학의 연구 비교연구의 두번째 가능성은 행동학적 연구방법의 적용과 인간 행 동 연구에 대한 문제 제기를 들 수 있다. 다시 말하면 심리학에서 비교 및 계통발생학적인 면을 도입하는 것이다. 인간행동학이라고 하는 이 연구 분야는 현대인도 진화의 법칙에 따라 계통발생 과정에 서 생겨난 존재라는 사실에서 출발했다. 인간도 다른 생물체와 마찬 가지로 자연 환경에 많은 특이적 적응을 통해 진화되어 왔다. 결국 인간 행동에서 공통되고 관습적인 특성을 찾고, 그들의 유전적 기본 성질을 발견하여, 문명화된 인간으로의 발달과정의 변화를 추적하고 그리고 인간 행동에 있어서 지나간 과거의 적응을 찾고자 하는 과제 룰 갖는다. 이러한 적응은 문명화된 인간의 변화된 생활방식으로 단 점이 될 수 있기 때문에 더 이상 필요한 것이 아니다 . 그러나 이 단 점들은 계통발생학적인 흔적으로 오늘날까지 잔존해 왔다.
이런 견해는 지금까지의 인간 행동에 대한 많은 오해 를 해결해 가 는 데 기여할 수 있었다 . 그 이유는 그것이 생물학적으로 그러나 심 리학적으로 보통의 방식으로 직간접적인 요인 사이에, 즉 계통발생 학적 원인과 실제로 규정된 메커니즘 사이에서 차이가 나지 않기 때 문이댜 이것은 이미 소개된 어미 -새끼 결합에 대한 육체적 접촉에 대한 의미에서 알 수 있다. 즉 무리가 이동할 때 스스로 따라 다닐 상태에 아직 이르지 못해 어미 몸에 의지하여 운반되는 어린 영장류 를 어미로부터 격리시키면, 그 새끼에게는 일단 위험한 일이 될 수 있다. 그래서 이것은 확실히 끊임없는 육체적 접근을 보장해 주는 메커니즘의 발달에 고도의 선택압을 부과시켜 온 것이다 . 어린 동물 의 행동(따라다니는 행동, 꽉 붙드는 행동)은 물론 유대충동이라고 할 수 있는 동인도 이것에 속한다. 이 동인은 선택조건들 아래 원래 특 정 생활 방식의 적응으로 발달해 왔다. 비록 이 선택조건들이 더 이 상 존재하지 않았다 하더라도 이 동인은 아주 확실히 오늘날까지 유 지되어 온 것이다. 만일 이것이 실현되지 않았다면, 무엇보다 어떤 발달장애가 일어났을 것이다 . 그렇지만 이 발달장애는 생물학적으로 살펴보면 그저 미리 계획된 행동양식의 보존적 형질에 대한 예에 불 과한 것이다 . 연구방법으로써 인간행동학에 대해一~ 종간 비교 혹은 개체군 간의 비교와는 반대로-첫째로 문화의 비교가 제시 된다. 이 문화의 비교는 비슷하거나 같은 방식으로 여러 문화가 형 성된 행동 특징들을 조사할 수 있다. 그렇기 때문에 인간의 행동도 동물의 종 특징과 관련된 성질들을 취급할 수 있다. 이러한 방법의 도입은 인간의 행동 특징들이 비교적 원시 상태로 남아 있거나 지리 적으로 서로 멀리 떨어져 있어서 의사소통이 일어나지 않은 문화에 서 나타날 때 특별히 올바르게 평 가되 고 있다 (Le i derman 등 l
능성이 전혀 없거나 단지 미미한 정도로 갖고 있는 신생아, 혹은 어 떤 특정 감각기관의 결여로 특정 경험들을 오랫동안 할 수 없었던 사람들, 예를 들면 눈이 멀고 듣지도 못하는 그리고 말도 못하는 아 이들 대상으로 한 연구를 들 수 있다. 여기서 어떤 행동양식들이 정 상적으로 그리고 자발적으로 나타나는 것에 대한 질문에 특히 관심 을 갖게 된다 . 말하자면 어떤 행동양식의 실행이 특정 경험들을 요 구하며 그래서 동물에서 이미 제시된 방법인 생득적인 행동 특징들 로 분류할 것인가를 연구하는 것이다. 3 인간 행동의 계통발생 이제는 계통발생학적 적응의 결과로 유전적 기초의 참여에 대한 예로써 검토되고 있는 몇몇 행동형질이 언급되어야 할 것이다. 이 행동형질들은 동작의 과정, 자극의 인식, 특별한 학습성향 그리고 동인 및 충동과 관련이 있다. 몇몇 동작의 과정에서 유전적 프로그 램화에 대한 언급이 가장 분명하다. 여기서 이 동작과정은 최소한 이미 갓난아이에서 있는 매우 간단한 동작과 관계가 있다. 엄마의 젖가슴을 찾을 때 자동적으로 작용하는 머리를 혼드는 동작, 마실 때 호홉 동작과 삼키는 동작을 일치시키는 것 혹은 선 자세로 걸을 때 아장아장 걷는 동작이 이에 속해 있다. 가장 잘 알려진 예는 갓 난아이의 손바닥에 손가락으로 자극을 주면 꽉럼반사룰 일으키는 동 작이다(그림 12-3). 이것은 신생아일수록 두드러진다 (Prech tl 1953). 갓난아이의 행동 이외에도 첫째, 조건적일 수 없는 그리고 둘째, 경 험가능성이 없이도, 다시 말해 귀가 먹고 눈이 먼 아이에서 나타나 는 행동양식이 있다 . 이 때문에 이것은 확실히 유전적으로 미리 계 획된 행동양식이다. 즉 미소지울 때 표정 동작은 아마도 보고된 모
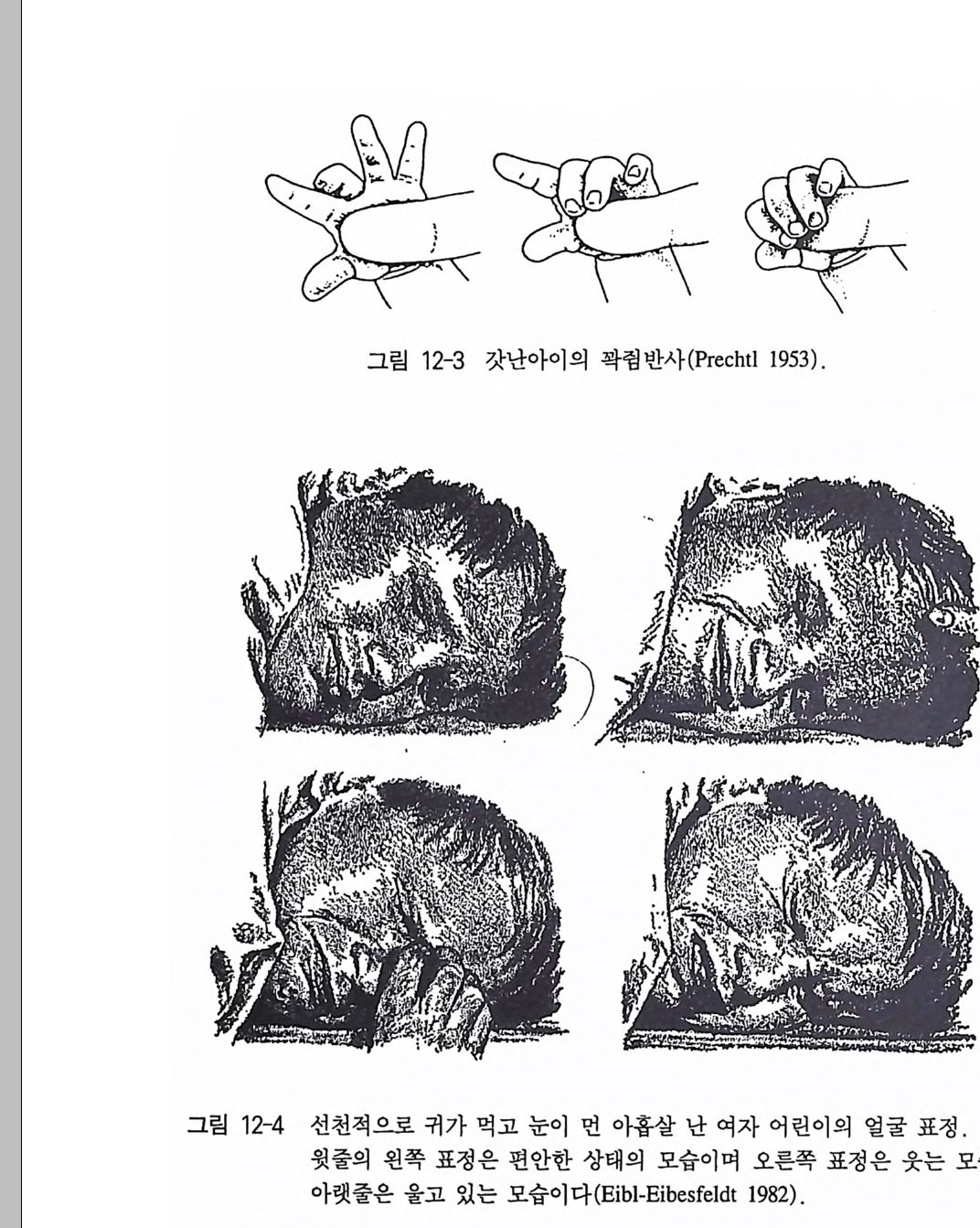 그림 12-3 갓난아이의 꽉림반사 (Prech t l I953).
그림 12-3 갓난아이의 꽉림반사 (Prech t l I953).
든 예들 가운에 가장 일찍 상황에 따른 동작과정으로 분류될 수 있 었다(그림 12-4). 그 다음은 사회적 행동양식으로, 눈썹을 재빨리
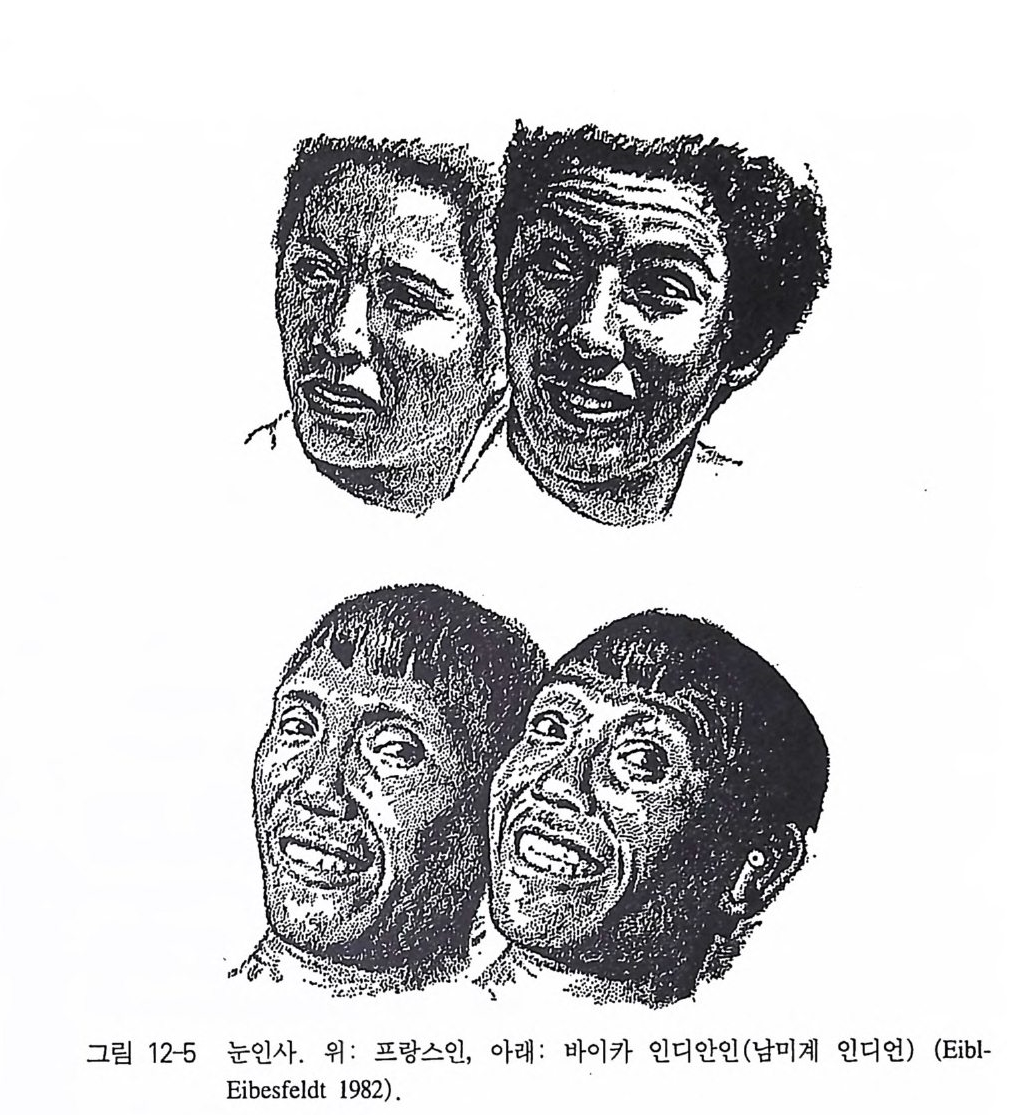 그림 12-5 눈인사 . 위 : 프랑스인, 아래 : 바이카 인디안인(남미계 인디언) (Eib l -
그림 12-5 눈인사 . 위 : 프랑스인, 아래 : 바이카 인디안인(남미계 인디언) (Eib l -
들어올림으로 생기는 〈눈인사〉는 이 행동이 넓게 퍼져 있기 때문에 비슷한 프로그램화로 생각하지 않을 수 없게 되었다(그림 12-5). 이 에 대한 증거로 갓난아이의 행동양식들이 일정하게 연결해서 나타나 는 것은 우연한 것이 아니고 측정할 수 있는 특정 규칙들에 따라 일 어나며 그레서 동물행동학에서 이미 밝혀진 행동연쇄와 비슷하다는 점이다.
 \\广
\\广
동물행동학의 생득적 해발기구라는 의미에서 유전적으로 미리 계 획된 인식 메커니즘에 대한 지식이 원래 결정적인 것이 아니다. 여 러 번에 걸쳐 인용된 예는 육체적 형질들의 결합인 어린애 행동이 있다 이것은 인간의 경우 보살핌의 반응을 불러일으킨다(그림 12- 6). 그러나 이런 반응이 일어나는 것에 대한 연구가 아직도 지지부 진하고 있는 한, 유전적 기초에 관련된 이 행동이 경험에 의해 생겨 난다는 궁극적인 평가는 불가능하다. 이와 같은 것이 남녀의 이차
성징에 반응하여 이따금 경험에 의한 것으로 여기는 것도 유전적인 프로그램에 해당되기 때문이다 (Blu rt on- Jo ne s 1972). 동물의 학습 성향은 그것이 여러 종에서 조사된 것처럼, 인간에게 도 마찬가지로 반복되어 추측되고 있다. 이것은 특히 언어학습의 재 능을 거론할 수 있다 . 이 언어는 언어학습보다 시기적으로 더 늦게 나타나는 수많은 다른 동작보다 근본적으로 더 복잡한 근육의 협동 을 요구하고 있다. 유전적으로 미리 결정될 수 있는 학습능력의 한 계에 더 분명한 실마리가 시간적인 측면에 있다 . 특정 경험들은 여 러 연령 정도에 따라 확실히 매우 다양하게 지속적인 기억으로 남긴 다 . 이런 시기 특이성에 대해 이미 사회화 능력과 관련해서 지적한 바있다 . 인간에게 공격충동이 어느 정도 유전적인가는 현재의 지식으로는 단정지을 수가 없다 . 공격적 동인의 생물학적 기능에 대해 이미 이 책에서 설명했기 때문에 여기서는 이 문제에 관해 더 이상 깊게 다 루지 않겠다 . 생득적 행동 요소를 알아내는 것에 대한 어려움은 이미 이 책에서 소개된 바 있다. 인간행동학 영역에 서로 이런 가능성들은 물론 아 직도 더 많은 한계가 있다. 여기서는 실제 적절한 형질로 특정 행동 성질들의 유전성만을 취급한다. 그래서 인간의 행동유전학은 큰 관 심을 일으키게 되었다. 가계 조사와 쌍생아연구는 입양 조사와 가족 특이적 행동 성질들을 통해 인지적 영역에서 조사되어 왔다 . 이런 연구의 가장 큰 부분은 심리병리학에서 예측, 인식도 그리고 치료도 로 인해 일시적으로 행해졌다. 여기서 염색체 이상(예 , 남자의 성염색 체가 두 배인 것)과 행동의 특이한 편차(예, 범죄 성향을 띠는 것) 간의 관계가 거론될 수 있다. 성과 관련된 유전(정신분열중과 알코을중독충) 역시 여러 번에 걸쳐 입증되어 왔다. 전체적으로 행동유전학은 치료 학 영역에 대한 이 학문의 직접적인 의미 외에도 계속해서 일반적
지식의 축적에도 기여하고 있다. 이런 지식은 자연과학적인 증명에 서 인간의 유전적 행동을 부각시켜 왔으며 ――우선 최소한 심리병 리학적 영역에서_一그리고 유전이냐 아니냐의 오래된 논쟁을 불식 시 키 게 되 었다 (Blu rt on- Jones 1976) .
참고문헌 Adk ins -Reg an , E., 1983: Sex ste r oid s and the dif fere nti at i on of avia n rep ro ducti ve behavio u r. In: J. Balth a zart, R. Prove und R. Gi lles (Hrsg .), Honnones and Behavio u r in Hi gh er Vert eb rate s . Sp ring er , Berlin . Andersson, M., 1982: Sexual selecti on , natu r al selecti on and qua li ty advert ise ment. Bio l . J. Linn . Soc. 17, 375-393. Andrew, R. J., 1957: The agg re ssiv e and court sh ip behavio u r of cert ain Emberi zi n a e. Behavio u r 10, 255-308. Ang e rmeie r , W. F., 1972: Kontr ol le des Verhalte n s. das Lemen am Er folg . Sp ring er , Berlin , Heid e lberg, New York. Archer, J., 1988: The Behavio u ral Bi ol og y of Ag gre ssio n . Cambri dg e Univ e rsit y Press, Cambri dg e. Aschoff , J., 1955: Jah respe r i od ik der Fort pflan zung bei Warmbliit em . Stu d iu m gen . 8, 742-776. Aschoff , J., 1981: Bio l og ica l Rhy thm s. Handbook of Behavio u ral Neurobio l og y 4. Phenum, New York. Aschoff , J., 1989: Temp o ral orie n ta t i on : cir c adie n clocks in anim als and humans. An im . Behav. 37, 881-896 Baerends, G. P., 1956: Aufb a u tier is c hen Verhalte n s. Handbuch der Zoolog ie, 8. Band, 7. Lie f e ru ng , Walte r de Gruy ter , Berlin , New York, 1-32. Balt ha zart, J., Prove, E. & Gi lles , R., 1983: Hormones and Behavio u r in Hi gh er Vert eb rate s . Sp ring er , Berlin . Bap tista , L. F. & Mort on , M. L. 1988: Song lea rning in monta n e wh ite- crowed spa r-rows : from whom and when. An im . Behav. 30, 1753-1764. Basto c k, M., 1969: Das Lieb eswerben der Tie r e. Fis c her, Stu ttgart. Beach, F. A., 1961: Honnones and Behavio r . Hoeber, New York.
Beer, C. G. , 1970: Indiv i d u al recog n iti on of voic e in the socia l behavio u r of bir d s. Adv. Stu d y Beh., 27-74. Berg, C. J., 1978: Develop m ent and evoluti on of behavio r in mollusks, wit h emp h asis . on chang e s in ste r eoty pe . In: Burgh ardt, G. M. & Bekoff , M. (e d.): The Develop m ent of Behavio r : Comp a rati ve and evoluti on ary asp e cts . Garland STPM Press, New York, London, 3-17. Bert ram , B., Clutt on -B rock, T., Dunbar, F., Rubenste i n , D. & Wrang h am, R.,, 1982: Current Problems in Socio b io l og y. Cambri dg e Univ e rsity Press, C ambri dg e . Bir k head, T. R, & M! illl er, A. P. 1992. Sp er m Comp e tit ion in Bir d s : Evoluti on ary Causes and Conseq ue nces. A cademi c Press, London. Bis c hof, H. J., 1979: A model of im p ri n t i ng evolved from neuroP. h y si o l og ica l con- cept s. Z. Tie r p s yc h ol. 51, 126-139. Bis c hof, N., 1972: Inzuchtb a r rier en in Saug et ie r sozie t a ten . Homo 23, 330-351 . Blurt on -Jo n es, N., 1972: Eth o log ica l Stu d ie s of C hil d Behavio r . Cambrid g e Univ e rsity Press, Cambri dg e . Blurt on -Jo n es, N. G., 1976: Growi ng po in t s i n human eth o log y: Anoth e r link betw een eth o log y and the soc ial scie n ces? In: Bate s on, P. P. G. & Hin d e, R. A. (ed.): Growi ng Poin t s i n Eth o log y. Cambri dg e Un ive rsity Press, Cambri dg e . 427-450. Bowlby , J., 1973: Mi itter li ch e Zuwendung und ge is t ig e Gesundheit . Ki nd ler, Mi inc hen. Bowlby, J., 1975: Bin d ung . Ein e Analys e der Mutt er -K i nd -Bezie h ung . Ki nd ler, Mi inc hen. Breed, M. D. & Bekoff , M., 1981: Indiv i d u al recog n it ion and socia l relati on ship s. J. Theor. Bio l. 88, 589-593. . Brereto n , J. LE G. & Immelmann, K., 1962: Head-scratc h in g in the Psit tac if orm es. Ibis 104, 169-175. Brodsky, L. M., Ankney, C. D. & Dennis , D. G. 1988: The inf l ue nce of male domi - nace on socia l int e r actio n in black ducks and mallards. Anim . Behav. 36, 1371- 1378. Brown, R. E. & Macdonald, D. W.(eds) 1985: Socia l odours in Mammals, Vols. I and 2. Clarendon Press, Oxfo r d.
Buchholtz , C h., 1 973: Da s Lemen bei Tie r en. Fis c her, S tu t t ga rt. Burkhardt, 0. , 1 989: UV vis io n : a bir d 's vie w of fea th e rs. J. Comp . Phy si o l . A. 164, 787-796. Burtt, E. H. & Hail m an, J. P., 1978: Head-scratc h in g among Nort h Americ a n wood-warblers (Parulid a e). Ibis 120, 152-1 7 0. Busnel, R. G., 1963: Acoustic Behavio u r of Anim als. Elsevie r , Amste r dam, London, New York. Bi. in ni ng , E., 1977: Di e ph y s io l og isc he Uhr. Ci rc adia n e Rhy thm i k und Bi o- chronometr ie. 3. Aufl ag e . Sp ring er , Berlin , Heid e lberg, New York. Cade, T. J., E . J. W illou g h by & G. L. Maclean, 1966: Drin k in g behavio r of sand- gro use in the Nami b and Kalaha ri desert s, Africa . A uk 83, 124-126. Cade, W . H. , 1981: Alte r nati ve male str a te g ies : gen eti c dif fere nces in cric k ets . Scie n ce 212, 563-4. Camhi, J . M. 1984: Neuroeth o log y: nerve cells and the natu r al behavio r of anim al. Sin a uer Associa t e s , Sunderland, MA . Cohen-S a lmon, C., Carlie r , M., Robert ou x, M., Jou haneu, J., Semal, C. & Pa ille tt e, M ., 1 985: Di ffere nces in pa tt er ns of pu p care in mi ce . V. Pup ultr as on ic emi s- sio n s and pu p care behavio u r. Phy si o l . and Behav. 35, 167-174. Colga n , P. 1989. Anim al Moti va ti on . Chap m an and Hall, London. Conrads, K. & Conrads, W ., 197l: Reg ion aldia l ekte des Orto l ans (Emberiz a hortu l ana) in deuts c hland. Vog el welt 9 2, 81-100. Cranach, M. v., F op pa , W. Lep e nie s , W. & Ploog , D., 1979: Human Eth o log y - Claim s and Li m i ts of a New Di sc ip l in e . Cambrid g e Univ e rsit y Press, Cambri dg e. Crook, J. H. , 1 970: Socia l Behavio u r in Bi rd s and Mammals. Academi c Press, London, New York. Cullen, E., 1 957: Adap tation s in the kittiw ake to cliff -ne sti ng . Ibis 9 9, 275-302. Cullen, J. M. , 1 953: Allo-, a uto - , and hete r op re en ing . I bis 105, 121 . Curio , E., 1968: Di e Adap tat i on ein e r Handlung ohne den zug e horig e n Beweg un g sa blauf. Verh. Dt sc h. Zoo!. Ges. 1967, 153-163. Akad. Verlag sg e s., Leip z ig .
Curio , E., I969: fun kti on sweis e und Sta m mesg e schic h te des Flug feind erkennens ein ig e r Darwi nf i nk en(Geosp izi n a e). Z. Tie r p s yc h ol. 26, 394-487. Daanje , A., 1950: On locomoto r y movements in bir d s and the int e n ti on movements deri ve d from the m. Behavio u r 3, 48-98. Dath e , H., 1972: Bei8 h emmung ? W iss . Z. Humboldt- U niv . Berlin , Math . Nat. R. XXI, 445-4 48 . Davie s , N. B. & Brooke, M. de L. 1988: Cuckoos versus reed warblers: Adap tat i on s and counte r -adapt at i on . Anim . Behav. 36, 262-284. Davie s , N. B. & Brooke, M. de L. 1989: An exp e rim enta l stu d y of co-evoluti on betw e en the cuckoo Cuculus canorus and its h osts . I. Host egg dis c ri m i na ti on . J. Anim . Ecol. 58, 207-224. Davie s , N. B. & Brooke, M . de L. 1991: Coevoluti o n of the Cuckoo and Its H osts . Sci. Amer. 223, 66-73. De Fri es , J . C., Gerva is, M. C. und E. A. Thomas, 1978: Respo nse to 30 ge nerati on s of selecti on for op en fiel d acti vi t y in laborato r y mice . Behavio r Geneti cs 8, 3-13 . Diam ond, J. N., K arasov, W. H., Phan, D. & Ca rpe nte r , F. L. 1986: Hummi ng bi r d diges tiv e ph y si o l og y: a dete r m ina nt of fora g ing bout freq ue ncy . Natu r e (Lond.) 320, 62-63. Dil ge r, W., 1962: Meth o ds and obje c ti ve s of eth o log y. Liv i n g Bi rd I , 83-92. Din , N. A. & Eltri ng h am, K., 1974: Ecolog ica l sep a ratio n betw een Whit e and Pin k -backed Pelic a ns in the Ruwunzori Nati on al Park. Ug an da. Ibis 116, 28-43. D'Udin e , B. & Alleva, E. 1983: Early exp er i en ce and sexual pre fe r ences in rodents . I n Bate s on, P.P.G. (ed .) , Mate choic e , pp. 311-327. Cambrid g e Univ e rsit y Press, Cambri dg e. Duncan, I. J. H. & Ki te, V. G., 1987: Some inv estig a ti on s int o moti va ti on in the domesti c fow l. Ap pl. Anim . Behav. Sci. 18, 387-3 8 8. Duvall, W . D., M ueller-Scharze, D. & Sil ve rste in, R. M. 1986: Chem ica l Sig n als in Vert eb rate s , 4 . Plenum Press, New York. Ehret, G., 1 980: Develop m ent of sound communic a ti on in mammals. A dv. Stu d y Beh. 11, 179-225. Ei bl -Eib e sfe l dt, I., 1957: Di e Ausdrucksfo r men der Saug e ti er e. Handbuch der
Zoolog ie. 8. B and, 6. Lie f e r ung. Walte r de Gruy ter , Berlin , N ew York, 1-26. Eib l -Eib e sf e l dt, I ., 1963: Ang e borenes und Erworbenes im Verhalte n ein i g e r Saug er . Z. Tie r p s yc h ol. 20, 705-7 5 4. Eib l -Eib e sfe ld t, I., 1982: Lie b e und Hass . R . Pip e r & Co. Verlag, Mi inc hen. Elwood, R. W. (ed.) , 1 983: Parenta l Behavio u r of R odents . Joh n Wi le y , Chic h este r. Emlen, S. T. & Orin g , L. W. , 1977: Ecolog y, sexual selecti on , and the evoluti on of mati ng sys t e m s. S cie n ce I97, 215-223. Ewert, J . P., 1980: Neuroeth o log y-A n Intr o ducti on to the Neurop h y s io l og ica l Fundamenta l s of Behavio r . Sp ring e r -V erlag, B erlin . Ewert , J. P. & Traud, R., 1 979: Releasin g stim uli for anti -pre dato r behavio u r in the common toa d Bufo b ufo L . Behavio u r 68, 170-180. Fa be r, H. v. & Haid , H. , 1 980: Endokrin o log ie, Bio c h e mi e und Phy s io l og ie der Hormone. 3 . A uf! . Ulmer, Stu ttgart. Fit zg i b b on, C. D. & Fansh a we, J. H. 1988: Sto t t ing in Thomson's ga zelles: an hoest sig n al of con dit ion . Behav. Ecol. Socio b io l . 23, 67-74. Frank, D., 1985: Verhalte n sbio l og ie- Ein f i ihr ung in die Eth o log ie. 2. Aufl . Thie m e, Stu t t gart. Frisc h, K. V., 1965: Tanzspr a che und Orie n ti er ung der Bie n en. Sp ri n g er , Berlin , Heid e lberg. Fri sc h, K. V., 1 977: Aus dem Leben der Bie n en. Sp ring e r , Berlin . Goodall, J ., 1979: Lif e and death at G ombe. Nati on al Geog rap h i c 155, 592-621 . Goodwi n, D . , 1 966: Austr al ia n Pig e ons: t he ir a ff ini t ies and sta tus . E mu 66, 319-336. Goslin g , L. M., 1982: A reassessment of the fun cti on of scent marki ng in ter r itor i es . z. Tie r p s yc h ol. 60, 89-118. Gott lie b , G., 1976: Concep tion s of pre nata l develop m ent: Behavio r al embry o log y. Psyc h o!. Rev. 83, 215-2 3 4. Gould, J . L. & Marler, P. 1987: Leami ng by ins ti nc t. Sci. Amer. 2 56, 62-73. Grafe n , A., 1990: Bio l og ica l sig n als as hand ica ps . J. Theor. Bio l . 144, 517-525. Graves, H . B., H able, C. P. & Jen k ins , T. H., 1 985: Sexual selecti on in Gallus. Eff ec ts of morp h olog y and domi na nce on fem ale spa tia l behavio u r. B ehav. Proc. 11, 189-1 9 7.
Greensp a n , R. J., 1995: Underst a n din g the Gene ti c Cons tr u cti on of Behav io r . Sci. Amer. 2 72, 74-79. Guil m et, G. M. , 1977: The ev oluti o n of too l-us in g and too l-m akin g behavio u r. Ma n 12, 33-4 7 . Guth rie, D . M. , 1 981 : Neuroeth o log y: an Intr od ucti o n . Blac k well, O xfo r d. Gwi nn er, E. & W ilts c hko, W ., 1980: Endog en ous ly contr o lled chang e s in mi gra to r y dir ec ti on in the gar den warbler, S y lv ia borin . J. Comp . Phy si o l . 125, 267-273. Gwi nn er, E., 1964: Unte r suchung en Uber das Ausdrucks-u n d Sozia l verhalte n des Kolkraben (Corvus corax corax L.) Z. Tie r p s yc h ol. 21, 657-748. Gwi nn er, E., 1991: Inte r nal Rhy thm s in Bi rd Mi gr ati on In : D. W . Mock(e d .) : Behavio r and Evoluti on of Bir d s. W. H. Freeman and Comp a ny New York, 20- 31. Hail m an, J., 1977: Op tica l Sig n als: Anim al Communic a ti on and Lig h t. India n a Univ e rsit y Press, Bloomi ng ton , India n a. Hall, E. T., 1 966: The Hi dd en dim ensio n . Doubleday, New York. Hall, J . C. 1994: The mati ng of a fly. Sc ien ce 264, 1702-1714. Ha milto n , W. D. & Zuk, M. , 1 984: Heri tab le true fitne ss and brig h t bir d s: a role for par asite s ? Scie n ce(NY) 218, 384-387. Harlow, H. F., 1 958: The natu r e of love. Am. Psyc h o!. 13, 673-6 8 5. Harlow, H. F., G luck, J. P. & Suom i, S . J., 1 972: Generaliz a ti on of behavio r al data betw e en nonhuman and human anim als. A m. Psy ch ol. 27, 709-716. Harlow, H. F. & Harlow, H. K., 1 962: Socia l dep ri v a ti on in monkey . Sci. A mer. 207, 136-146. Hassenste i n , B., 1972: Bedin g u ng en filr Lemp ro zesse-te l eonomi sc h ge sehen. Nova acta Leop ol din a , No. 206, p.2 89-320. Hedig e r, H., 1941 : Bi ol og isc he Gesetz m l iBi g k eit en im Verhalte n von W irb eltie r en. Mi tt. Natu rf. Ges. Bem( l94 0). Hedig e r, H., 1949: Sli ug et i er t er r itor i en und ihr e Markie r ung . Bi jdr . Di er k. 28, 172-184. Hedi ger , H., 1954: Sk izz en zu ein e r Tie r p s yc holog ie im zoo und im Zir ku s. Europ a Verlag, Stu ttgart.
Hein r ic h , W. , 1 967: Unte r suchung en zum Sexualverhalte n in der Gatt un g Til ap ia (Ci ch lid a e, Teleost e i ) u nd bei Art ba sta r den. Z. Tie r p s yc h ol. 24, 684-754. Helb, H. W ., 1973: Analys e der arti so lie r enden pa ramete r im Gesang des Fit is(P hyl l oscop u s t. tro chil us ) Mi l U nte r suchung en zur Obje k ti vi e r ung der ana- lyt isc hen Meth o de. J. Om . 114, 145-2 0 6. Helversen, D. V., 1980: Str u ctu r e and fun cti on of anti ph onal duets . Acta XVII Cong r. Int. Orn., Berlin 1978, 682-688. Helversen, D. v. & Helversen, 0. v., 1975: Verhalten sge n eti sc he unte r suchung en am akusti sc hen Kommunik a ti on ssy s te m der Feldheuschrecken (Orth o p the ra, Acrid i d a e). Z . clmp . Phy si o l . 104, 273-299. Hendric h s, H., 1978: Di e sozia l e Orga nis a ti on von Sii ug e ti er po p u lati on en. Siiu g e t ie rk undl. Mi tt. 26, 81-116. Hepp e r, P ., 1991: Kin Recogn i t ion . Cambrid g e Univ e rsity Press, Cambri dg e . Herre, W. & Rohrs, M. , 1973: Haustie r e -zo olog isc h ges ehen. Fis c her, Stu ttgart. Herre, W., 1980: Grundlage n zoolog isc her Domesti ka ti on sfo r schung . Nova acta Leop ol din a No. 241, 3-1 6 . Hess, E., 1956: Sp ac e per cept ion in chi ck s. Sci. Amer. 195, 71-80. Hess, E., 1975: Priig u ng . Die friihk i nd lic h e Entw ic k lung von Verhalte n smuste m bei Tie r und Mensch. Kind ler, Mi inc hen. Hess, W. R., 1954: Das Zwi sc henhim . Sy n drome, Loka lisa ti on , Funkti on . Schwabe, Basel. Hess, W. R., 1957: Die Formatio reti cu laris d es Him sta m mes im verhalte n s-ph ys i o l o- gisc hen Aspe k t. Arch. Psyc h ia t r. Neurol. 196, 329-336. Hey m er, A., 1973: Verhalt en sstu d ie n an Prachtl ibe llen. Forts c hrit te der Verhalten sfo rs chung Nr. 1 1. Parey, Berlin . Hin d e, R. A., 1978: Domi na nce and role: Two concept s wit h dual meanin g s . J . of Socia l & Bio l og ica l Str uc tu re s 1, 27-38. Holldobler, B. K. & Wi lso n, E. 0., 1977: Weaver ants . S ci. Amer. 237, 146-154. Holst, E. V., 1939: Die relativ e Koordin a ti on als Phanomen und als Meth o de zentr al- nervoser Funkti on sanalys e . Erg. P hy sio l . 42, 228-306. Holst, E. v. & Paul, U. v. St., 1960: Vom Wi rlc ung sg e f ii ge der Tri eb e. Natu rwiss . 18,
409-422. Hunti ng f ord , F. A., 1976: T he relati on ship betw een int e r -and int r a -spe c ific agg re ssio n . Anim . Behav. 24, 485-4 9 7. Hunti gford , F. A. & Turner, A. 1987: Anim al Confl ict . Chap m an and Hall, L ondon. Hurst, J., 1989: The comp le x netw o rk of olfa c to r y communic a ti on in po p u lati on s of wi ld house mi ce Mus domestic u s. R utt y: Marki ng s and inv estig a ti on wi thi n fam - ily grou p s. Anim . Behav. 37, 705-725. Hutc h is o n, J, B., 1978: Bi ol og ica l Dete r mi na nts of Sexual Behavio r . Joh n W ile y, London. Immelmann, K. & Immelmann, G., 1967: Verhalt en sokolog isc he Stu d ie n an afr ika nis - chen und austr al is c hen Estr ild id e n, Zool. Jb. 94, 609-684. Immelmann, K., 1962: Beit ra g e zu ein e r vergl e ic h enden Bi ol og ie austr a lis c her Prachtf ink en (Sp e rmesti da e). Zool. Jb. Sy st. 90, 1-196. Immelmann, K., 1967: Perio d is c he Vorga ng e in der Fortp flan zung tier is c her Orga n is m en. Stu d . gen . 20, 15-33. Immelmann, K., 1 971: Ecolog ica l asp e cts of per i od ic rep ro ducti on . In: Farner, D. S. und J. R. King ( e d.) : A via n Bio l og y. Band I. Academ ic Press, New York, 341-389. Immelmann, K., 1972: Sexual and oth e r long -ter m asp e cts o f im p rint i ng in bir d s and oth e r spe c ie s . Adv. Stu d y Behav. 4, 147-174. Jac obs, W . , 1966: Di e Gesang e der Heuschrecke. In: Burkhardt, D . , Schleid t , W . und H. Altn e r: S ig na le in der Tie r welt. Moos, M i inc hen. Kawa i, M . , 1 965; Newly acq ui r ed pre cultu r al behavio r of the natu r al troo p of Japa nese monkeys on Koshim a isl et. Prim ate s 6, 1-30. Kim , K. W . & Park, S. R. ,19 93: lntr aind iv i d u al and Inte r in d iv i d u al Varia t i on s of Ste r eoty pe d Song s in Gray -h eaded Bunti ng (E mberi za fuc ata ) . Korean J. Zool. 36, 476-4 8 6. King , J . A., 1973: The ecolohy of ag gres siv e behavio r . Ann. Rev. Ecol. Sy st . 4, 117- 138. Kohler, W., 1973: Inte l l ige nzp r iif un g e n an Menschenaff en . Sp ri n g e r, Berlin , Heid e lberg. New York.
Kommers, P. E. & Dhin d sa, M. S. 1989: Infl ue nce of domi na nce and age on mate choic e in black-bil led mag pies : an exp er im enta l stu d y . Anim . Behav. 37, 645- 655. Konis h i, M ., 1966: The attr i b u te s of ins ti nc t. Behavio u r 27, 316-328. Kramer, G., 1952: Sonnenori en ti er ung der Vog el . Zoo!. Anz. S up pl. 16, 72-81. Krebs, J. R. & Davie s , N. B., 1993: An Intr od ucti on to Behavio u ral Ecolog y. 3th ed. Blackwell, Oxfo rd . Kroodsma, D. E., 1978: Aspe c ts o f learnin g in the onto g e n y of bir d song : where, from whom, when, how many , whic h , and how accurate l y. In: Burgh a rdt, H. M. und M. Bekoff (H rsg .) . The Develop m ent of Behavio r : Comp a rati ve and Evoluti on a ry Aspe c ts . G arland STPM Press, New York, London. Kruska, D., 1970: Vergl e ic h end cyt oa rchit ek to n is c he Unte r suchung en an Gehir n en von Wi ldu nd Hausschwein e n. Z. Anat. Entw ic k l. - Gesch. 131, 291-324. Kummer, H., 1975: Sozia lv erhalt en der Pri m ate n . Sp ring e r , Berlin , Heid e lberg, New York. Lack, D., 1 943: The Lif e of the Robin . Wi the rby, London. Lauer, J. & Lin d auer, M. , 1 971: Geneti sc h fixi e r te Lerndis p o sit ion en bei der Honig b ie n e. A bh. Akad. Wi ss . Lit . Main z . Inf. O rg. 1, 1-87. Lazarus, J., 1979: Flock siz e and behavio u r in capt ive Red-bil le d Weaverbir d s (Qu elea que lea): I mp li ca ti on s for socia l fac ili tation and the fun cti on s of floc k- ing . Behavio u r71, 127-145. Lehrman, D. S., 1 964: The repr o ducti ve behavio r of Ring Doves. Sci. Amer. 211, 5, 48-54. Lenin g t on , S. & Eg id, K., 1989: Envir o nmenta l inf l ue nces on the pre fe re nces of wil d fem ale house mi ce for males of dif feri n g t-c omp le x ge noty pe s. Behavio r Genecti cs 19, 257-266. Leid e rman, P. H., Tulkin , S. R. & Rosenfe ld , A., 1977: Cult ur e and Infa n cy . Va riat io n s in the Human Exp er ie n ce. Academ ic P ress, New York. Lennerberg, E. H., 1973: Bio l og isc he Grundlage n der Spr a che. Suhrkamp , Fra nkfurt. Ley ha usen, P., 1 975: Verhalten sstu d ie n an Katz e n. Parey, Berlin , Hamburg. Ley ha usen, P., 1979: Katz e n-ein e Verhalte n skunde. 5. Aufl ag e . par ey, Berlin ,
Hamburg. Lin s enmair , K. E., 1967: Konstr u kti on und Sig n alfu n kti on der Sandp yra mi de der Reit er krabbe Ocyp o de sarata n Forsk. Z. Tie r p s yc hol. 24, 403-456. Lins enmair , K. E. , I 979: Unte r suchung e n zur Sozio b olog ie der W tist e n assel Hemi lep istu s reaumuri und verwandte r Isop o denarte n (lso p o da , Onis c oid e a) : Paarbin d ung und Evoluti on der Isog am ie. V erh. Dt sc h. Zool. Ges. 1979, 60-72. Lloy d , J. E., I 966: Stu d ie s on the flas h communic a ti on sig n als in Photi nu s fire fl ies . Mi sc . Publ. Zool. Univ . Mi ch . No. 130. Lois e lle, P. U. & Barlow, G. W ., 1 978: Do fish es lek lik e bir d s? In: Reese, E. S. und F. J. L ig h te r (Hrsg. ), Contr as ts i n Behavio r . j oh n Wi le y, New York, 31-75. Lorenz, K., 1935: Der Kump an in der Umwelt d es Vog el s. J. Om . 83, 137-213 und 289-4 1 3. Lorenz, K., 1 937: Uber den Beg riff der Insti nk th a ndlung. Folia bio t h e oreti ca 2, 17-50. Lorenz, K., 1 940: Durch Domesti ka ti on verursachte Sto r ung en artei g e nen Verhalt en s. Z. ang ew . Psyc h ol. Charakte r k. 59, 2-81 . Lorenz, K., 1941: Vergl e ic h ende Beweg u ng s stu d ie n an Anati ne n. J. Orn, 89, Ergi inz ung sb and, 194-293. Lorenz, K., 1943: Die ang eb orenen Fonnen mog li c h er Er fah rung . Z. Tie r p s yc hol. 5, 235-409. Lorenz, K., 1983: Das sog en annte Bose.Deuts c her Taschenbuch Verlag, Mt inc hen. Lorenz, K., 1978: Vergl e ic h ende Verhalte n sfo r schung . Grundlag e n der Eth o log ie. Sp ring e r, W i en , New York. Lorenz, K. & Tin b erge n , N. , 1 938: Ta xis und Insti nk th a ndlung in der Eir o llbeweg u ng der Graug an s. Z . Tie r p s yc h ol. 2, 1-2 9 . Mannin g , A. & Dawk ins , M. S., 1992: An Intr od ucti on to An im al Behavio u r. 4th ed. Cambri dg e Unversity Press. Marki , H ., 1971: Yorn Eig e nnutz des Uneig e nnti tzige n. Natu r wi ss . Rdsch. 24, 281-289. Marler, P., 1956: Uber die Eig e nscha ften ein ige r tier lic h er Rufe . J. Om. 97, 220-227. Marler, P ., Duft y, A. & Pic k ert , R . A 1986: Vocal communic a ti on in the domesti c chi ck en. I. Does a sender commun ica te infon nati on about the qua li ty of a foo d
refe re nt to a receiv e r? Anim . Behav. 34, 188-193. Marler, P. & Tamura, M. , 1964: Cultu r ally tra nsmi tted pat te r ns of vocal behavio r in spa r rows. Sci en ce 146, 1483-1486. Ma rtin, P. & Curo, Y. M. 1985: On the fun cti on s of pla y and its role in behavio u ral develop m ent. Adv. Stu d y Behav. 15, 59-103. Mayn ard Smi th, J., 1977: Parenta l inv estm ent: A pro spe c ti ve analys i s. Anim . Behav. 25, 1-9 . Mayr , E., 1 967: Artbe g riff und Evoluti on . Parey, B erlin , H amburg. Mayr , E., 1974: Behavio r pro g ram s and evoluti on ary stra te g i es . Amer. Scie n ti st 62, 650-659. McBr ide , G., 1971: Anim al Fam ilie s . Reader's Dig es t Associa t io n , Pleasantv ille, N ew York. Mi tan i, J. C. & Rodman, P. S ., 1 979: Terr itor i al ity : The relati on of rang ing pat te r n and home rang e siz e to defe n dabil it y , wit h an analys i s o f ter r itor i al ity among prim ate spe c ie s . S ocio b io l .2 4 1-251 . Mock, D. W. , 1 991: Behavio r and Evoluti on of Bir d s. W. H. Freemand & Comp an y , New York. Moksnes, A., Roskaft, E., B raa, A. T., K orsnes, L., Lamp e, H. M. & Pederson, H. Ch., 19 91 : Behavio u al respo nses of po te n ti al hosts t ow ards artifici a l cuckoo egg s and dumm ies . Behvio u r 116, 64-89. Nachts h eim , H. & Ste n g el , H., 1977: Vom Wi ld ti er zum Haustie r. 3. Aufl ag e . Parey, Berlin, Hamburg. Na rins , P. M. & Capr a nic a , R. R., 1976: Sexual dif fere nces in the audit or y sys t e m of the tree frog Eleuth e rodacty lu s coq ui . Sci en ce 192, 378-380. Nau, F. & Deliu s , J. D., 1981: Disc repa n t eff ec ts of unil a te r al and bila te r al fore bra in lesio n s on the vis u al per f on nance ofp ige ons. Behav. Brai n Res. 2, 119-124. Neumann, F. & Ste in b eck, H., 1971: Honnonale Beein f l us sung des Verhalte n s Klin. Wochenschr. 49, 790-806. Neuweil er, G., 1980: Audit or y pro cessin g of echoes: pe rip h eral pro cessin g . In: Busnel, R. G. und J. F. Fis h (ed.), Anim al Sonar Sys te m s. Plenum, New York, 519-548.
Ni co lai, J., 1956: Zur Bi ol og ie und Eth o log ie des Gi m p e ls. Z. Tie r p s yc h ol. 13, 93- 132. Ni co lai, J., 1959: Verhalte n sstu d ie n an ein i g e n afr ika nis c hen und pa laarkti sc hen Girli tz en . Zool. Jb. Sy st. 87, 317-362. Ni co lai , J ., 1964: Der Brutp a rasit ism us der Vi du in a e als eth o log isc hes Problem. Z. Tie rp s yc h ol. 21, 129-204. Nic o lai , J., 1976: Evoluti ve Neuerung en in der Baiz von Hausta u benrassen (Columba livia var. domesti ca ) als Erge b nis menschlic h er Zuchtw ahl. Z. Tie r p s yc h ol. 40, 225-243. Op pe nheim , R. W., 1982: Di e Neuroembryo log ie des Verhalte n s: Forts c hrit te, Probleme und Perspe kti ve n. In: lmmelmann, K., G. W. Barlow, L. Petr ino vic h und M. Main (Hrsg .): Verhalte n sentw i ck lung bei Mensch und Ti er . Das Bie le fe ld -Proje k t. Parey, Berlin , Hamburg. Pap i, F., 1976: The olfa c to r y navig a ti on sy s te m of the homi ng pige on. In: Verhandlung en der Deuts c hen Zoolog isc hen Gesellschaft , R ath m ay er , W(ed.) . Fis c her, Stu ttgart. Park, S. R. , 1991 : Develop m ent of Socia l Str u ctu r e in a Cap tive Colony of the Common Vamp ire Bat, Desmodus rotu n dus. E th o log y 89, 335-341 . Pawlow, J. P., 1972: Die B edin g ten Refl ex e. Kind ler, Mi lnc hen. Payn e , T. L., Bir ch , M. C., & Kennedy, C. E. J. (eds), 1986: Mechanis m s in Insects Olfa c ti on . Clarendon Press, Oxfo rd . Peek, F. W., 1972: An exp er i m enta l stu d y of the ter r itor i al fun cti on of vocal and vis u - al dis p l a y in the male Redwi ng e d Blackbir d (Ag e laiu s ph oenic e us). Anim . Behav. 10, 112-118. Pete r s, H. M. & Berns, S., 1982: Die Maulbrutp fleg e der Cic h li de n. Unte r suchung en zur Evoluti on ein e s Verhalte n smuste r s. Z. f. zool. Sy st . u. Evol. -fors chung 20, 18-52. Pete r s, H. M ., 1948: Grundfr ag e n der Ti er ps y c holog ie: Ordnung s - und Gesta l tp ro bleme. Enke, Stu ttgart. Ph illips, R. E. & Young ren , 0. M., 1971: Brai n stim ulati on and spe ci es -ty pica l behav- iou r: a cti vi t ies evoked by electr ica l stim ulati on of the brai ns of ch ick ens (Gallus
gal lus). A nim . Behav. 19, 757-779. Pia n ka, E. R., 1970: On r-a nd K-selecti on . Am. Natu r ali st 1 04, 592-597. Pil leri , G. & Knuckey , J., 1969: Behavio u r pat te r ns of some Delph in i d a e observed in the Weste r n Medit er ranean. Z. Tie r p s yc h ol. 26, 48-72. Pia n ka, E. R., 1970: On r-und K-selecti on . am. Natu ra lis t 1 04, 592-597. Prechtl , H. F. R., 1953: Sta m mesge s chic h tl ich e Reste im Verhalte n des Saug li n g s . Umschau 53, 656-658. Prechtl , H. F. R., 1 953: Zur Phy si o l og ie der ang eb orenen auslosenden Mechanis m en. Behavio u r 5, 32-50. Prove, E. & Sossin k a, R ., 1982: Radio i m munoassay of pla sma hormones and its use in inv estig a tio n s of hormone and behavio r correlatio n s in bir d s. In: Scanes, C. G. et al. (Hrsg .) : Asp e cts of Avia n Endocrin o log y: Practi ca l and Theoreti ca l Imp li ca ti on s. G rad. Stu d ie s , T exas Tech Univ . 26, 97-103. Prove, E., 1974: Der Ein f l uB von Kastr a ti on und Hormonsubstu i o n auf das Sexualverhalten mannli ch er Ze,b raf ink en(Taenio p y gia g u tta t a casta n oti s G ould). J. O m. 115, 338-347. Prove, E., 1 978: Qu anti tat iv e Unte r suchung en zu Wechselbezie h ung en zwi sc hen Balzakti vi ta t und Testo s te r onti tem bei mannlic h en Zebrafi nk en (Taenio p y gia gu tta t a casta n oti s G ould). Z . Tie r p s yc h ol. 48, 47-67. Pull iam , H. R., 1980: Prog ra mmed to Learn. Columbia U niv e rsity Press, New York. Ralls, K ., 1971: Mammalia n scent marki ng . sci en ce 171, 443-4 4 9. Rasa, 0. A. E., 1 971: Socia l Inte r acti on and Obje c t Ma nipu lati on in Weaned Pup s of the Nort he rn Eleph ant Seal Mi ro unga ang u sti ro str is. Z. Tie r p s yc h ol. 29, 82-102. Rein b oth , R., 1 980: Vergl e ic h ende Endok rino log ie. T hiem e, Stu ttgart, New York. Remane, A., 1 971 : Die Grundlage n des Nati irli c h en Sy st e m s, der vergl e ic h enden Anato m ie u nd der Phy lo g en eti k. Koltz, Konig st e i n . . Remane, A., 1 976: Sozia l leben der Tie r e. Fis c her, Stu ttgart. Rensin g , L., 1973: Bio l og isc he Rhy thm en und Reg u latio n . Fis c her, Stu ttgart. Reye r , H.- U ., 1980: Flexib l e helpe r stru c tu re as an ecologi ca l adapt ation in the Pie d King tish er(Cery le rudis r udis L .). B ehav. Ecol. Socio b io l . 6, 219-227. Ric h ard, G., 1 979: Onto g e n esis a nd ph yl o g en esis: mutu a l consta int s . Adv. Stu d y Beh.
9, 229-178. Ridp at h , M. G., 1972: The Tasmania n Nati ve Hen, Trib o ny x mortie r i. CSIRO W ild l. Res. 17, 53-90. Rijks en, H. D., 1981: Infa n t killing : a pos sib l e conseq ue nce of a dis p u t e d leader role. Behavio u r 78, 138-168. Roth e nbuhler, W. C., 1964: Behavio r gen eti cs of nest cleanin g in honey b ees. IV. Respo nses of Fl and backcross gen erati on s to dis e ase kille d brood. Am. Zoo!. 4, 111-123. Rowland, W. J., 1989: Mate choic e and the sup e momality eff ec t in fem ale sti ck le-backs (Gaste r oste u s aculeutu s ). Behav. Ecol. Socio b io l . 24, 433-438. Sauer, E. G. F., 1966: Zwi sc henartl ich es Verhalte n und das Problem der Domesti ka ti on . Jou m. S. W. A Wi ss ensch. Gesellsch. 20, 5-22. Scha ffer, H. R., 1977: Stu d ie s on Moth e r-Infa n t Inte r acti on . Academ ic Press. London. Schaller, G. B. & Crawshaw, F. G. , 1 981: Soc ial orga n iz a ti on in a Capy b ara po p u la- tion . Saug et ie r kundl. M i tt. 29, 3-16. Schenkel, R., 1967: Submi ss io n : Its fea tu r es and fun cti on in the wolf and dog . Am. Zoolog ist 7 , 319-329. Schone, H. & Schone, H., 1963: Balz und andere Verhalte n sweis e n der Mang rov ekrabbe Gonio p s is cruenta t a Latr . und das W ink verhalt en der euli- tor alen Brachy ur en. Z. Tie rp s yc h ol. 20, 641-656. Seely, T., 1985: Honey b ee Ecolog y. Princ eto n Un ive rsity Press, N. J. Seit z, A., 1940:. D i e Paarbil du ng bei ein i g e n Ci ch li de n. I. Di e Paarbil du ng bei Asta t o t i lap ia s tr ig ige na Pfe ffer . Z . Tie r p s yc h ol. 4, 40-84. Sevenste r , P., 1961: A causal analys is of a dis p la cdment acti vi t y(fan nin g in Gaste r oste u s aculeatu s L.). Behavio u r Sup pl. 9, 1-170. Sic k , H., 1959: Di e B alz der Schmuckvog el. J. Om. 100, 269-302. Sin z , R., 1981: Lemen und Gedachtn is, 3. Aufl . Fis c her, Stu t t gart, New York. Sk inn er, B. F., 1 963: Op er ant Behavio r . Amer. Psyc h ol. 18, 503-515. Slate r, P. J. B., 1978: Sex Hormones and Behavio u r. The Instit ute of Bi ol og y's Stu d ie s in Bio l og y No. 103. Arnold, London. Slate r , P. J. B., 1989: Bi rd Song learnin g ; causes and conseq ue neds. Eth o log y,
Ecolog y and Evoluti on 1, 19-46. Sluckin , W., 1 973: Imp ri n t i ng and Early Learnin g . Aldin e Publis h in g Comp . Chic a g o . Soler, M . & Moller, A. P., 1 990: Durati on of sym p a tr y and co-e v oluti on betw een the gre at spo tt ed cuckoo and its m ag pie host. Natu r e 343, 748-750. Sossin k a, R., 1970: Domesti ka ti on serschein u ng en beim Zebrafi nk en Taenio p ygia gut- tat a casta n oti s ( Gould). Zoo!. Jb. Sy st . 97, 455-521 . Sossin k a, R., 1982: Domesti ca ti on in bir d s. In: Farner, D. S. and J. R. Ki ng (eds.) : Avia n Bio l og y, Band VI, Academi c Press, N ew York, 373-403. Sta d don, J. E. R., 1975; A note on the evoluti on a ry sig n if ica nce of sup e rnormal stim - uli. Am. Nat. 109, 541-545. Sta d ie . Chr. , 1968: Verhalte n sweis e n von Gatt un g sb asta r den Phasia n us colchic u s Gallus ga llus f. domesti ca im Vergl e ic h mi t denen der Ausg an g sa rt en . Verh. Dt sc h. Zoo!. Ges. 1967, 493-519, Akad. Verlag sg e s ., L eip z ig . Sudhaus, W., 1974: Rekap itul ati on en in der Eth o kolog ie der Wasseramsel (Cin c lus cin c lus). Beit r. V og el kd. 20, 461-466. Suom i, S. J., Coll ins M. L., & Harlow, H. F., 1973: Ef fec ts of per manent sep ar ati on from moth e r on inf a n t monkey s. Dev. Psy ch o!. 376-384. Taborsky, M. & Lim berge r , D., 1981: Helpe rs in fish . Behav. Ecol. Socio b io l 8, 143- 145. Tembrock, G., 1982: Ti er sti m menfo r schung . Ei ne Ein f t ihr ung in die Bi oa lcusti k. 3. Aufl ag e. Zie m sen, W itten berg- L uth e rsta d t. Thie l cke, G., 1974: Sta b il itat erlernte r Sin g v og e l-Gesag e tro tz vollsta n dig e r ge o- gra p h is c her Isolati on . Vog el wa rte 27, 209-215. Thomas, E., 1955: Der Kommentk am p f der Ireuzott er (Vip e ra berus L.). Natu r wi ss . 42, 539. Thorp e , W . H., 1961: Bir d Song . Cambri dg e Univ e rsity Press, Cambri dg e. Thorp e , W. H. & Pil c her, P . M., 1 958: The natu r e and characte r i st ic s of s ubsong . Br it. Bir d s 51, 509-514. Tin b erge n, N., 1 940: Di e Uberspr u ng b eweg u ng . Z. Tie r p s yc h ol. 4, 1-4 0 . Tin b erge n , N. , 1 948: Socia l releasers and the exp er i m enta l meth o d req ui r ed for the ir stu d y. W ils . B ull. 60, 6-51 .
Tin b erge n , N., 1950: The hie r archic a l orga n iz a ti on of nervous mechanis m s underluin g ins tin c ti ve behavio u r. Sy m p . Soc. Exp . Bio l . 4., 305-312. Tin b erge n, N., 1 959: Comp a rati ve stu d ie s of the behavio u r of gu lls (Larid a e): a pro g res s repo r t . B ehavio u r 15, 1-70. Tin b erge n , N., 1 968: On war and pea ce in anim al, and man. Scie n ce 160, 1411-1418. Tin b erge n , N., 1989: The Stu d y of Insti nc t. Clarendon Press. O xfo rd . Tin b erge n , N., 1 990: Socia l Behavio u r in Anim als. C hapm an & Hall, London. Tin b erge n , N., M eeuse, B. J. D., B oerma, L. K. & Varossie a u, W. W., 1943: Die Ba iz des Samt falt e r s (Eumenis s emele L.) Z. Tie r p s yc h ol. 5, 182-226. Tin b erge n , N. & Perdeck, A. C., 1 951 : On the stim ulus situ a tio n releasin g the beg ging respo nse in the newly- h atc h ed Herr ing Gull chi ck (Larus aug en ta t u s ). Behavio u r 3, 1-38. Toate s , R., 1986: Moti va ti on al sys t e m . Cambri dg e Univ e rsity Press. C ambri dg e . Tri ve rs, R .L . , 1 974: Parent- o ff sp r ing con flict. Ameri ca n Zoolog ist 1 4, 249-264. Tschanz, B., 1968: Trott el lummen. Beih e ft 4 zur Zeit sc hrift fiir Tie r ps yc holog ie. Parey, Berlin , H amburg. Veste r ga a d, K., 1980: The regu lati on of dustb a th ing and oth e r pa tte r ns in the lay ing hen: a Lorenzia n app r oach. In Moss, R. (ed.), The Lay ing Hen and its Envir or unent. Matt inu s Nij ho ff , The Hag ue . Vos, G. J. de, 1979: Adapt ed ness of arena behavio u r in Black Grouse (Tetr a o tet r ix ) and oth e r grou se spe c ie s (Tetr ao nin a e). Behavio u r 68, 277-314. Wagn e r, H. 0., 1 954: Versuch ein e r Analys e der Kolib r i ba lz. Z . Tie r p s yc h ol. 11, 182 - 212. Walth e r, F., 1958: Zum Kamp f-u nd Paarung s verhalte n ein i g e r Anti lo p e n. Z. Tie rp s yc h ol. 15, 340-380. Walth e r, F., 1967: Huft ier te r it or i en und ihr e Marki er ung . In: Hedig e r, H . (Hrsg. ): Die Stra 8e n der Tie r e. Vie w eg, Braunschweig , 26-4 5 . Walth e r, F., 1 979: Das Verhalte n der Homt rag er (Bov_id a e). H andbuch der Zoolog ie, 8. Band, 54. Liefe ru ng . W alte r de Gruy ter , Berlin , New York, 1-1 8 4. Wehner, R ., 1976: Polar ize d-lig h navig a tio n by ins ects . Sci. Amer. 235, 106-121. Weis s , P ., 1941 : Auto n omous versus refl ex og en ous acti vi t y of the centr al nervous sys -
ter n. Proc. Amer. Phil o s. Soc. 84, 53-64. W ick ler, W ., 1 965: Uber den tax onomi sc hen Wert homolog er Verhalte n smerkmale. Natu r wi ss . 5 2, 269-270. Wi ck ler, W., 1966: Ursp ru ng und bio l og isc he Deutu n g des Genit al pr i is e nti er ens mi inn lic h er Pri m ate n . Z. Tie r p s yc h ol. . 23, 422-4 3 7. W ick ler, W ., 1967: Vergl e ic h ende Verhalte n sfo r schung und Phy lo g e neti k. In: Heberer, G . (Hrsg. ) : Di e Evoluti on der Orga n is m en. 3. At rla ge , Band 1. Fis c her, Stu t t gart. Wi ck ler, W., 1 969: Sin d wi r S under -Natu r ge s etz e der Ehe. Droemer, M t inc hen. W ilkins on, G., 1990: Food sha ring in vamp ire bats . Sci. A mer. , 2 72,6 4 -70 . Wi lso n, E. 0., 1 975: Socio b io l og y. The New Sy n th e sis . Belknap Press, Cambri dg e, Mass. Wi lts c hko, R., 1981: Die Sonnenori en ti er ung der Vog el I und II. J. Ornit ho l. 122, 1- 27. Wi lts c hko, W . & Wi ltsc hko, R. , 1 988: Mag ne ti c versus celesti al ori en ta tion in migra- tion bir d s. Trend in Ecol. and Evol. 3, 13-15. W itten berge r , J. F. & Til so n, R. L., 1980: The evoluti on of monog am y : hyp o th e se and evid e nce. Ann. Rev. Ecol. Sy st. 11, 197-232. Young , D. , 1989: Nerve Cells and An im al Behavio u r. Univ e rsit y of Cambri dg e Press, Cambri dg e.
기 가젤영양 301 강꼬치고기 300 개 115 개개비 2(JJ , 꼬 8 개구리 78 개코원숭이 227, 286, 300, 306, 319 거미 233 거미불가사리 233 검은꼬리누 238 검은딱새 %8 검은머리물떼새 73 검은멧닭 236 검은방울새 74 검은부리까치 69 겨우살이지빠귀 350 고양이 97 골드햄 스터 119, 175 공작 65 과일박쥐 286 괭이갈매기 179 구세계솔새 347 귀뚜라미 68 그랜드가젤 232 그레바얼룩말 240 극락조 284 금눈쇠올빼미 167 금파리 141 금화조 115, 117, 217, 218, 267, 286
기니피그 128, 129 기번 2A6 긴팔원숭이 2A2, 284, '2K7 까마귀 112, 248 꿀벌 50, 67, 143, 198, 19() 꿩 112 L 나나니벌 210 나무발발이 179, 180 나방 50 나비 16 노랑턱멧새 171 농게 68, 263 뇌조 284 C: 다람쥐 67 다모충류 139 단풍새 213. 215, 288, 343, 344 달랑게 244 닭 67, 69 도마뱀붙이 242 돌고래 306 두꺼비 59 들당나귀 240 디스커스 292 딱따구리 %8
딱따구리핀치 223 닷쥐 134, 275 땅다람쥐 刃 5 때까치 248 2 랑구르 255 리라새 21J7 마나킨새 284 마모셋 284, 373 마카크원승이 115, 216 메뚜기 110 메추라기 195 멧돼지 2S4 멧비둘기 72, 19( ), 122, 123, 130 모기 139 무희새 %6 문작 346 물닭 285 물벼룩 53, 140 물잠자리 241 민머리독수리 223 밍크 Z/5 닌 바다사자 301 바다오리 195 바다제비 %9
바다코끼 리 242, 253 , 301, 306 바다표법 242 바위자고 266 바퀴벌레 121 박새 219 박쥐 50, 52 반날개과 74 반딧불이 68, 74, 264 방울뱀 50, 51 뱀가 O H--지 301 뱀장어 38 버들붕어 121 벌 50 벌새 248, T76, 284 범고래 301 붉은등꾀꼬리 247 붉은배숲때까치 287 붉은부리갈매기 168, 181, 229, 268 붉은빵멧새 171 붉은사다새 302 붉은사슴 67, 346 붉은여우 254 비버 304 뻐꾸기 74, 357 뿔논병 아리 269, 291 人 사랑새 131 사자 240, 254
사향고양이 314 사향소 254 산누에나방 265 상모솔새 248, 280 생쥐 119 서배너원숭이 66 세가락갈매기 I'll , 2 83 세가락나무늘보 293 세가락매추라기 237, 294 소나무행렬나방 141 솔새 139 、 숲귀뚜라미 41, 165 숲비 둘기 250, 321 스컹크 166 쑥새 171 。 알락귀뚜라미 40 알락딱새 33, 167 알락할미새 359 애기카멜레온 210 앵무새 119 야자집게 345 양놀래기과 1()C ) 어치 幻 7 얼룩말 236 에뮤 294 엑솔로틀 39, 177 연어 156, 215
영양 16 옆새우 154 오랑우탄 242, 246 오리 69 오릭스 252 올빼미 33, 64 왕잠자리 238 유럽개개비 359 유럽굴뚝나비 54 55, 73, 1(:B 유럽되새 89 유럽밭종다리 359 유럽붉은가슴울새 56, 57, 155 유럽억새풀새 359 유럽지빠귀 112 유리멧새 155 이구아나 68 인드리 246 일본원숭이 219, 223 x: 잠자리 32 재갈매기 56, 64, 73, 85, 167 재봉개미 224 전갈 291 전서구 156 정원솔새 167, 180, 185, 186 재비 68 족제비 刀 5 주머니하늘다람쥐 313
쥐며느리 142 지렁이 16 직조새 30 집단직조새 303 집쥐 76 짖는원숭이 242, 246 찌르레기 143, 144, 145, 2If7 -K 참새 137 참새과 170 참새목 170 참집게 %0 천녀새 216, 342 천녀새류 215 청둥오리 195, 271 청소물고기 70 칠면조 32, 55, 62 침 팬지 124, 186, 224, 254 구 카나리 아 117, 362 코끼리 223 코뿔새 294 코뿔소 236, 242 쿠바핀치 57 크리벤시스 44 45 큰가시고기 44, 53, 56, 57, 81, 91,
116, 119, 124, 167, 168, 169, 267 . 268 . 276 큰뇌조 236 큰뿔영양 220 큰회색머리아비 254 키클리드 85, 120, 237 E 타조 294 태양새 343 토끼 275 티티원숭이 284 工 팔로로원 272 페카리 313 표범나비 74 푸른되 새 170, 172 푸른박새 280 푸른앵무새 303 -o_ 하마 236, 253 하이에나 254 해오라기 296, 2
혹고니 271 홍엽새 119 홍학 278, 292 황새 254, 268 황열모기 51 회색기러기 73, 78, 169 휘파람개구리 51
홉혈박쥐 308, 318 흰개미 16 흰귀벌새 277 흰극락조 266 흰머리쑥새 173, 217, 258 흰발생쥐 138 히말라야원숭이 257. 286. 370
-I 가슴신경절 41, 101, llO, lll 가짜 복종행동 268 가짜 화장 352 가축화 3fi0 , 365 각인 129, 210, 2ll, 223, 357, 371 각인현상 194 간격행동 251 간접요인 273 갈등상황 67, 89 갈등행동 83, 84, 300 갈망행동 34, 35, 37, 79, 289 감수분열 262 감자씻기 219 강제적 학습과정 165 개방된 실명사회 313 개방된 익명사회 313 개방사회 312 개별동작 101 개별 인식 188, 192, 199, 315, 316 개별적 기억 161 개인심리학 369 개체간격 228, 248, 250, 267 개체발생 21, 159, 174, 340 개체발생학적 의식화 165 거세 115, 120, 123, 258 검사심리학 369 게놈 340 경계음 67, 69, 76
경계행동 33, 203 경고신호 66 경험 123 경험박탈 163, 164 고유수용기 38 고전적 조건화 203 고정 행동패턴 76, 77, 78, 166 공격억제 321,322 공격억제 메커니즘 268 공격억제 효과 270 공격적 동인 261 공격적 해발인 124 공격충동 240, 255, 256, 257, 379 공격행동 20, 44, 56, 65, 83, 167, 182, 226, 227, 263 공동구애 302 공동둥지 303 공중적개행동 102 공진화 355, 357 과시행동 68, 69 과지속기 104 관심갈등 290 광수용기 142 광주성 140 교대의식 295 교미행동 185 교배실험 32,3 , 324, 327, 328 구걸음 179, 180 구걸 해발인 298 구결행동 179, 295, 2 이, 321
구성원 신분증 314 구심성 신경 25 구애급이 269 구애노래 117, 265, 349 구애동작 44,54 구애소리 40, 349 구애의식 336 구애장 236, 284 구애행동 64, 66, 69, 165, 267, 275, 276, 283, 287, 335, 336, 351 구역질 반응 106 군선택 356 굴곡반사 291 굴곡주성 141 굴성 140 귀소 실험 156 근육 반사 27 근친상간 316 근친 인식 316 기분전염 '}JJ7 기초 옹성화 128 기초 자성화 128 깃털다듬기 287 꼬리춤 149, 221 꽉럽 반사 27, 375 꿀벌의 춤 148 L 내부시계 145, 151
내부요인 90 내측흥분성중심 61 노래 170 노래각인 217 노래전수 220 놀램물질 29< ) 놀이표정 l'if/ 놀이행동 182, 185, 188, 19< J, 371 뇌자극 101, 104 뇌하수체 119, 129, 130 뇌 하수체 호르몬 120, 121 눈인사 377 C 다시냅스 반사 26 다형질 교배 324 단독터 237 단성 교배 324, 329 단시냅스 반사 25 단일신호 62 달램행동 228, 231, 320 달주기 139 대리대상체 30 대리대상체행동 30 대상전가행동 85 대용 116 도구사용 219, 223 도피 놀이 184, 187 도피행동 31, 44, 103
돌연변이 193, 334, 339 동물사회학 23 동물행동 17, 18 동물행동학 15, 19, 367, 374 동인 41, 43, 47, 83, 104, 124, 255 동인분석 44, 229 동인상태 104 동인성 신호자극 42, 103 동인성 자극 49 동인의 분석 43 동인체계 107 동인측정기구 46 동작연쇄 81, 82 동조효과 276 동종교배 324 동형집합성 128 둥지짓기 77, 88, 118, 119, 122, 269, 273, 321, 351 a 말초여과 50, 51, 52 먹이각인 215 먹이먹이기 125, 'X!O, 'XII, 292 먹이-번식터 234 먹이전달 %9 먹이-카스퍼-하우저 163 먹이탐색행동 34 먹이터 234, 235 먹이포획놀이 185
멍어리위협 181 명령체계 40, 41, 101 모래모욕 44, 2ITT 모방 206 모자이크형 돌연변이 335 모친가족 294 목표기구 175 176 목표정위 36 목표주성 142 무대상 생득적 166 무정위운동 141 무조건반사 27, 28 물체각인 211, 213 민감기 194, 214, 218, 2fi0, 371 l::t 반사 25 반사궁 25, % 반사사슬 28 반사학 38 반사회로 17 반응연쇄 81 반응의 강도 104 반응지속기 104 발달장애 164, 371 발산 53 발생역치 105 발신자 124, 219 방사선 면역측정법 113
방어충동 2AO 방어행동 31, 226, 316 방언 174, 280 방향성 자극 49 배회구역 235 버금노래 350 번식기생 74 번식터 234, 235 번식행동 123 변형발생 178 보호반사 27 복종행동 102, 228, 231, 268 복합신호 62 본능모델 90,92 본능 행동 28 부리내밀기 270 부신피질 호르몬 119, 121 부채꼴 실험 150 부친가족 294 분석심리학 369 분업 303 불완전우성 325, 326 비대 362 비위생벌 329 人 사회 312 사회구조 305 사회기생 74
사회생물학 23 사회생태학 23 사회성-결핍증후군 217 사회적 결합 315 사회적 서열 319 사회적 유인 311 사회적 의사소통 90, 351 사회적장애 371 사회 적 카스퍼-하우저 163, 192 사회적 해발인 71 사회적 화장 287. 321 사회적 화해행동 322 사회충동 289 사회행동 20, 225 상동 340 상반감정의 공존행동 84, 85, 229 상사 340 상호강화현상 72 상호경 계 29'J . 306. 30' ) 상호먹이교환 270 상호방어 306 상호억 제 84, 89, 121 상호영 양교환 306 308 상호의사소통 체계 71 상호이타주의 311 상호인식 314 상호입벌리기 180 상호적 자극강화 72 새끼돌보기 121, '2ff). 289. 294. 306 새끼돌보기조력자 꼬)
새까운반행동 291 생득적 해발기구 36, 91, 168, 193, 2'!/, 378 생득적 행동 162, 164, 165, l(i6 생리심리학 369 생물리듬학 24 생물시계 133 생물음성학 24 생식격리 65, 279, 280 생식놀이 185 생식행동 92 서커디언 리듬 134, 135, 136 서커디언 조정자 137 선택실험 330, 333 선택압 (i6, 74, 188, 228, 267, 270, 272, 279, 299, 304, 305, 310, 374 섭식동인 46 섭식전수 223 섭식행동 16, 20, 31, 77, 79, 88 성모자이크 현상 336 성선자극 호르몬 129 성숙 174, 176 성염색체 127 성장식 68 성적각인 211, 212, 213 성적과시 69 성적동인 44 성적선택 228 성적행동 20, 31, 34, 44, 225, 262,
263, 322 성행동 118, 274, 275 성호르몬 114, 115, 118, 119, 123, 126, l(i6 , 272 소낭유 292 소리내기 15, 101, 207, 271, 315, 340 소화 204 송과체 137 수력본능 모델 47 수렴 53 수술 114 수신자 125, 219 수용역 (i() 수정관 121 수정운동 78 수직적 순위제 317 수컷-어미 가족 294 수평억제 53 숙주각인 215 216 순위제 315, 317, 319, 320 스키너 상자 204 습관화 201 시각적 유인 263 시각적 표시 315 시각지시자 134, 136, 137, 138 시간터 238, 250 시냅스 25 시상하부 125 시행착오학습 206 신경계 95
신경절세포 59, 60 신경행동학 23, 62 신장수용기 111 신호자극 28, 58, 62, 65, 70, 91 신호전달 42 심리학 19, 368, 373 싸움놀이 183, 184 싸움행동 53, 71, 93, 225, 226, 227, 251, 333 쌍이외자 짝짓기 285 。 아타비즘 328 안드로겐 115, 127, 129 알굴리기 354 알굴리기 행동 78 알라타체 121, 124 r- 전략 289 양성교배 324, 329 양육놀이 185 양육준바 83 양육행동 55, 58, 64, 75, 76, 88, 120, 122, 225, 263, 272, 289, 291, 293, 294, 295, 296, 310, 321 양친가족 294 어마새끼 결합 291, 374 언어심리학 369 에스트로겐 46, 115, 118
에스트리올 122 X- 염색체 336 여과 49 여과기구 58 여과기작 53 여포자극호르몬 (LH) 131 여포호르몬 115 역교배 329 역치변화 29 역치상위 29 역치자극전류 105 역치하위 29 역치하 자극 29, 105 연주기 124, 139 연합현상 82 열쇠자극 62 염색체 333 염색체 이상 379 영아살해 254, 255, 371 영양교환 321 완료동작 79 완료상황 35 완료행동 34, 35, 79, lF:7 외부자극 38, 47, 49, 50, 75, 78, 10'3 외측억제성둘레 61 우열의 법칙 324 운동성훈련 188 운동신경 25 울음자세 326 움직이는 터 250
옹성호르몬 115, 124, 127, 129 원심성신경 25 원형춤 148, 221 위생벌 329 위선적 사회 312 위엄행동 228, 230. 268 위장 74 위축 362 위협동작 165, 193 위협행동 71, 90, 181, 228, 229, 230, 246, 352 유대 285 유대기작 317, 321 유대충동 289, 374 유아행동 271 유전성 감각 .1(, 6 유전성 운동 lf,6 유전자지문 285 유전적응 161 은폐색 74 음성발달 163 음성 분석 기 24, 220 음성학적 유인 264 음절 170 의도동작 271, 350, 351 의무적 사회성 304 의사소통 64, f,6, 68, 188, 295, 374 의식화 348, 349 의식화 싸움 253 의태 69
이기주의 3()C ) 이동중심 91 이동행동 83 이유갈등 290 이중구애 266 이중점 134 이타주의 305, 306, 3()C ), 310 이타행동 308, 311 이형접합성 128 익명 폐쇄사회 313 인간각인 212 인간행동학 24, 373, 379 인과관계 21 인도음 179, 180 인사 231 인사행동 288, 320, 321 인지발달 188 일부일처제 284. 285, 286. 287. 373 일주기 134 일처다부제 285 임상심리학 369 츠 자극역치 10') 자극익힘 33 자극통합 71 자극특이적 피로 32, 201 자동 X 선 촬영술 125 자발적 충격발사 43
자발적 행동 37 자성 호르몬 119, 122 자연선택 254, 256, 272, 339 자율동인 288, 289 자율적 싸움욕구 261 자장 컴퍼스 155 작동조건화 204, 206, 261 잠복기 43, 1 야 장소 신뢰성 220 장소전수 223 적-카스퍼 -하우저 163 전수 219 전수 상동 342 전이 동작 350, 351 전이쪼기 88 전이행동 86, 89 전향주성 142 전형행동 22 절단 % 접촉행동 249 정보교환 302 정보원 l(i() 정 보저 장소 340, 353 정위동작 36 정위음 51 제스타겐 115, 118 조건반사 27, 28, 203 조건자극 203 조력자 303 조직심리학 369
존스턴 기관 51 종간 공격 226 종간 해발인 69 종내공격 226 종의 기억 161 종특이적 놀이 183 종특이적 싸움행동 184 주광성 140 주류성 141 주성 76, 77, 78, 140 주주기 124 주지성 141 주지성 행동 333 주화성 141 중간유전 324 중추여과 50, 53 중추적 위치순응 1()C ) 중추적 자극선택 54 중추적 조절 111 중추적 프로그램화 111 지그재그춤 44, 267, 268 지도-컴퍼스 개념 156 지향동작 l(>( i 직접요인 刃 3 진공-공격 259 진공행동 30, 36, 37, 48, 49, lfJ l, 259 진화 339 진화적 원인 21 집단구애 72, 刃 8
집단냄새 313 집단동조 277 집단번식 131, 276 집단이기주의 311 집단터 237 집단행동 225, 298 집짓기 83, 303 집합 311, 312 짝유대 262, 268, 271, 274, 283, 286, 288, 289, 295, 321 짝짓기 81, 193, 262 짝짓기소리 32 짝짓기행동 77, 81, 83, 90, 321 짝터 237 짝형성 262, 268, 284, 317, 319 쪼기순위 320 大 청각적 카스퍼-하우저 163 청결공생 70 초정상 자극 73, 74 총생활권 234 추적비행 54 추종각인 211, 212 축소 364 춤언어 221 충격형태 111 충동행동 49 측심체 121
충격발사 빈도 61 구 카스트 304 카스퍼-하우저 175 카스퍼-하우저 동물 163 카스퍼-하우저 실험 164 컴퍼스 정위 155 K- 전략 289 E` 타액분비 반사 27 타협모색 305 탈구심화 38 탈억제가설 88, 89 탐색행동 34, 314 태교 165 태양컴퍼스 145, 155 터 233, 234 터 경계 240 터구조 239 터 표시행동 2A6 텃세권 309 텃세중심 92 텃세행동 228, 232, 233 테스토스테론 91, 115, 116, 117, 119, 122, 125, 129, 131, 258 테스토스테론 결정체 126
통찰학습 208 갔 D 통합기작 95 투기적 행동 226 특이적 피로 31 특이 적 피 로현상 1()() 포 파불로프의 조건반사 203 페쇄적 발달 프로그램 193 폐쇄사회 312, 313, 316, 317 포괄적응도 311 포란행동 125 포육반응 56 포획행동 58, 185, 161, 226 폴리진 330 표현행동 65 풀줄기 구애 344, 351 풀기춤 343 프로게스테론 115, 118, 119, 123, 131 프로락틴 120, 121, 122, 123, 125, 130 -o_ 하렘 284 학습 174, 191 학습과정 165 학습기간 194 학습성 향 1 이, 199, 375
학습심리학 369 학습적응 161 학습행동 162 항로결정 기작 157 항안드로겐 116 해마상 융기 125 해발기구 58, 62, 91 해발역치 116 해발인 63, 64, 65, 66, 68, 70, 74, 124, 182, 刀 1, 280, 364 해발자극 26, 167 행동계통학 23 행동권 234 행동 내분비학 23 행동 돌연변이 334 행동목록 20, l(i( ) 행동발달 159, 164 행동발생학 23 행동생리학 23 행동생태학 22,23 행동수렴 355 행동양식의 연쇄 101 행동연쇄 363, 377 행동유전학 23, 379 행동의 공진화 355 행동의 기능 21 행동의 수렴 353 행동의 전수 219 행동의 흔적 345 행동주의 19
행동준비 42, 43, 49, 362 행동충격 38 행동특이적 피로 31 혈연도 310 혈연 선택 254 혐오 반응 34 협동방어 300 협동사냥 300 형태부동 77, 165 형태불변 77 호기심 행동 182, 206 호르몬 42, 90, 95, 113, 114, 121, 124, 258 호르몬 결정체 126
호르몬계 112 호르몬 대용 127 혼란효과 299 혼인비행 54, 55, 169, 276, 294 혼인춤 276 화장 3()(), 349 화장행동 88 화학적 거세 127 활동전위 1()(), 111 황체형성 호르몬 (FSH) 131 황체호르몬 115 획득된 해발기구 168 후각적 유인 265 후짝짓기 T/1
Acroceph alus sci rp a ceus 359 Acti as selene 265 Aedes aeg ypti 51 Aeschna cya n ea 238 Ag ap o mi s cana 328 Ag ap o mi sf i sh eri 327 Ag ap o mi s 119, 327, 328 Ag ap o mi s roseic o ll is 327, 328 Ag ap o mi s tar anta 328 Ag el aiu s pho enic e us 247 Ambys t o n ia mexic a numi 39 Ammop h il a 210 Anhin g ida e 301 Anth u s pra te n sis 359 Ap is m ellifi ca cami ca 198, 199 Ap is m ellifi ca ligu sti ca 198, 199 Arta m us 250, 321 Ath e ne noctu a 167 Aue/ea que lea 119 Bi rg u s latr o 345 Brachyt el es arachnoid e s 284 Buceroti da e 294 Calli ph ora 141 Ca llithricida e 284, 373 Cercop ithe cus aeth i o p s 66 Cert hi a f am i liar is 179 Cervus elaph us 346 Chamaeleo dorsatu s 282
Chamaeleo long ico mi s 282 Chamaeleo pum i lus 210 Chir o xip h ia caudata 266 Chort hi p pu s big u tt ul us 282 Chort hi p pu s brunneus 282 Chort hi p pu s monta n us 282 Clunio marin u s 139 Colop tery x 241 Connonchaete s tau rin u s 238 Cops ych us malabari cu s 320 Corvus corax 268, 287 Cry ll us camp es tr is 41 Cy gn us olor 271 Drosop hi l a m elanog as te r 332 Eleuth e rodacty lu s coq ui 51 Emberiza hort ul ana 282 Erith a cus rubecula 56, 155 Estr i ld a astr i ld 344 Estr ildida e 213, 215, 269, 288, 343 Eug lo ssin e 243 Eumenis s emele 54 Eunic a vir i d i s 139 Fic e dula hyp o leuca 33, 167 Fr ing illa c oelebs 89, 170 Gavia arctic a 254 Gr yll us camp es tr is 165
Hy lo bati da e 284, 287 Hy lo charis l eucoti s 277 Labri da e 109 Lamp ro log as cong ol ensis 2 37 Lania r i us barbarus 287 Larus rid i b u ndus 229 Loxoblemmus ari en tu lu s 40 Macaca mulatta 286, 370 Melops itt ac us undulatu s 13 I Menura sup er ba 207 Mota c il la a lba 359 Munti ac us muntj ac 346 My iop s it ta m onachus 303 Neochmi a p ha eto n 343 Norop s auratu s 229 Oncorhyn c hus 156 Ory x gaz ella 252 Oscin e 170 Pag ur us samueli s 2 60 Pap io 227, 286 Paradi sa edae 266, 284 Pa rulida e 347 Parus caeruleus 280 Passeri for m 170 Passeri na cyan ea 155
Peromy sc us 138 Peta u ri da e 313 Phalarop od ida e 294 Phil e ta i r u s soc ius 303 Pip rida e 266 Plemato c hromi s k rib e nsis 44, 45 Podic e ps cri sta t u s 269 Poeph il a acuti da uda 271, 322 Prunella modularis 359 Pter op ida e 286 Pyrr hula pyrr hula 270 Regu l us regu l us 280 Rhabdit is i ne nnis 329 Sp inu s spi nu s 74 Sta ph y linida e 75 Sy lv ia bori n 167 Sym p hy s o don dis c us 292 Tali tru s 154, 155 Ta ya ssu 313 Thaumeto p o ea pityo camp a 141 Tia r i s o li va cea 51 Tilap ia 291, 297, 326 Tilap ia n il o ti ca 326 Til ap i a t ho lloni 326 Tri bo nyx mortie r i 285 Turdus merula 112 Turdus vis c i vo rus 350
Uca amnuli pe s 264 Vea pu g ilato r 264 Vea rhiz a p h orae 264 Ur ia a olga 195 Vermi vo ra chry so p ter a 347
Vermi vo ra pe reg rina 347 Vi du in a e 215 Vip e ra berus 251 Vi ve rr ina e 314 7.on otr ich ia leucop h ry s 173, 258
acq ui r ed releasin g mechanis m (ARM) 168 act cha in 81 acti on -spe cific f atigu e 31 ago n isti c b ehavio r 226 altru ism 306 ambiv a lent behavio r 85 app e asement ges tu re 231 app etitive behav ior 34, 79 atr op hy 362 auto n omous moti va ti on 288 basic fem aleness 128 basic m aleness 128 behavio ra l converge n ce 353 behavio u ral reli ct 345 big em inu s 134 bio l og ica l clock 133 call 349 causal 21 cenog en esis 178 centr al filtering 53 chemota xis 141 cir ca d ian 133 cir ca d ian pac emaker 137 cir ca d ian rhy thm 134 cir ca n nian 139 classic a l cond ition ing 203 co -ev oluti on 355
commando sys t e m 40 cond ition ed refl ex 27 con flict behavio r 84 consummato r y act 34, 79 corp o ra allata 121 corp o ra cardi ac a 121 cou rtship 262 cyp ro te r onaceta t 116 dep riva ti on syn d onne 217 diale ct 174 disin hibi t ion hy po th e se 88 disp a c ement acti vi t y 350 disp l a cement behavio r 87 DNA finge rp rinting 285 EIRM 169, 170 epi gamic d isp l a y 230 epi ph y se 137 eth o g ram 20 eth o log y 15, 19 evoluti on 21 exp re ssiv e behavio r 65 exti nc ti on 204 extr a- pa ir matin g 285 fillial im p rint i ng 211 fIX ed acti on pat t er n(FAP) 76 func ti on 21
gan g lion cell 59 gen ome 340 geo ta xis 141 gre etin g 231 groo m ing 287 grou p selecti on 356 gyn andormorp h 336 habitu a tio n 201 home rang e 234 hyp e rtr op h y 362 imitat io n 206 im p rinting 194, 211 infan ti lism 271 inn ate 161 inn ate releasin g mecha nism (IRM) 168 ins ig h t lear ning 208 int e n ti on movement 350 inter spe c if ic a gg res sio n 226 int r as pe c if ic a gg res sio n 226 IRM 169 Joh sto n 's orga n 51 Kaspa r -Hauser anima l 163 key stim u lus 62 kine sis 141 klino ta xis 141
late r al ing h ibi t ion 53 lek 236 mati ng 262 moti va ti on 41 note 170 onto g e n y 21 ope r ant cond ition i ng 204 pair bond ing 262 pair fon natio n 262 pec k order 320 per ip h eral filtering 50 pho to t a xis 140 pro x ima te f ac to r 273 pse udo-clean ing 352 psy c h ohy dr aul ic m ode 147 reacti on chai n 81 recip ro cal stit nu lus enhancement 72 redi rected behavi or 85 refl ex 25 releaser 63 releasin g mecha nism 58 rheota xis 141 ritualize d fight 253 rituarization 348
selectio n pre ssure 228 sensit ive pha se 194 sexual im p rint in g 211 sexual selecti on 228 sig n al stim u lus 62 socia l groo m ing 287 soci al releaser 71 song 170,349 spo n ta ne ous behavio r 37 stim u lus habit ua tio n 33 stim u lus-spe c if ic f atigu e 32 subm issi v e ges tu re 231 subsong 350 substit ute acti vi ty 30 substit ute obje c t 30 subth res hold 29 sup em onnal sim u lus 73
taxis 76, 140 tel ota x is 142 ter r itor i al behavio r 232 ter r itor y 223 threa t b ehavio r 229 tot a l rang e 234 trad i tion 219 trop i sm 140 trop o t a x is 142 ultim ate fac to r 273 uncond ition ed refl ex 27 vaccum acti vi ty 30, 259 vaccum-agg res sio n 259 wean ing con flict 290
박시룡 경희대학교 생물학과 졸업 독일 본 대학교 대학원(동물행동학 전공) 현재 한국교원대학교 생물교육과 교수 저서 『술취한 코끼리가 늘고 있다 』 『재미있는 동물이야기』 역서 『동물의 사회행동』 『사고. 학습 그리고 망각』 『사회생물학 I,2 』(공역) 『진화의 붉은 실』 논문 「휘파람새 노래소리의 방언」 의 다수 동물행동학의 이해 대우학술총서 • 자연과학 105 1 판 1 쇄 찍음 • 1996 년 2 월 15 일 1 판 1 쇄 펴 냄 • 1996 년 2 월 25 일 지은이 • 朴是龍 펴낸이 • 朴孟浩 펴 낸곳 • (주) 인옵^t 출판등록 1991 . i2. 2 0. 제 1 6-4 90 호 135-120 서 울시 강남구 신사동 506 ·강남출판문화센터 5 층 대표전화 515-2000 팩시밀리 515-2 0 07 값 21,000 원 © 박시룡, 1996 자연과학, 생물학 KDC /49 1 .5 Pri nte d in Seoul, Korea ISBN 89-374 -35 93-4 94470 89-374 -30 0 0-2 (세트) 지은이와 합의하여 인지를 불이지 않습니다.
대우학술총서 (자연과학) 1 소립자와 게이지 상호작용 김진의 55 양자광학 이상수 2 동력학특론 어병호 56 효소반응 속도론 서정현 3 질소고정 송승달 57 화성암 성인론 이민성 4 상전이와 임계현상 김두철 58 확률론 구자홍 5 촉매작용 진종식 59 분자 분광학 소현수 6 뫼스 바 우어 분광학 옥항납 60 벡터속 이론 양재현 7 극 미 량원소의 영양 승정자 61 곤충신경 생리학 부경생 8 수소화붕소와 유기붕소 화합물 윤농민 62 에너지띠 이론 모혜정 9 항생물질의 전합성 강석구 63 수학 기초론 김상문 10 국소적 형태의 Aliy a h- si n g er 64 신경과학 김승업·박찬웅 지표이론 지동표 65 BCH 부호와 Reed- 11 Mucop ol y sacchar i des 의 Solomon 부호 이만영 생화학 및 생물리학 박준우 66 양자 전기역학 김영덕 12 천체물리학 홍승수 67 군환론 박재걸 13 프 로 스타글라딘 합성 김성각 68 대수기하학 조영현 14 천연물화학연구법 우원식 69 양자장이론 이재형 15 지 방영양 김숙희 70 해양오염과 생태계 심재형 16 결정화유리 김병호 71 비기체 연소합성 ( SHS ) 여철현 17 고 분자에 의한 화학반응 조의환 72 크 로마토그래피 이대운 18 과학혁명 김영식 73 곤충의 사회 행동 추종길 19 한국지질론 장기홍 74 동위원소 지질학 김규한 20 정 보 이론 한영열 75 X- 선 결정학 김양 21 원자핵반응론 정운혁 77 통계역학 조순탁 22 파괴역학 김상철 78 고분자의 구조와 형태학 아석현 23 분자궤도 이론 이익춘 79 LC 에 의한 광학 이성질체의 24 반응속 도 론 정경훈 분리 현명호 25 미 분위상수학 이현구 80 신 경 전달물질 T서 유현 26 자기공명방법 조성호 81 발생과 유전자 발현 이양림 27 플라스마 물리학과 핵융합 최덕인 82 스테로이드 화학 김완주김득준 28 천문관측과 분석 이시우 83 다체계론 엄정인 29 석탄에너지변환기술 김상돈 84 중핵반응론 김병택 30 해양미고생물학 백광호 85 비가역 열역학 이철수 31 편미분방정식론 김종식 86 등각장론 임채호 32 대통일 이론 소광섭 87 방사선생물학 남상열 33 금속전자계의 다체이론 김덕주 88 석유지질학 이용일 34 액정중합체 진정일 89 베르누이 시행의 통계적 분석 35 복합재료 권숙인 배도선 · 김성인 36 단백질 생합성 박인원 90 신경세포생리학 강만식 37 한국의 광물종 김수진 91 생리활성을 가진 C-P 화합물의 38 일반상대론 이철훈 화학 김용준·강익중 39 레이저 광산란 분광학 김종진 92 생물유기화학 서정헌 40 복소다양체론 김상문 93 조직배양 김승업 41 역학적 연구방법 김일순 94 유기전이금속화합물 조남숙 외 42 핵구조물리학 민동필 95 실내환경 과학 김윤신 43 후리에 해석과 의미분 작용소 김도한 96 유한요소법 정상권 44 한국의 고생물 이 하영 97 대수적 위상수학 우무하 • 김재룡 45 질량분석학 김명수 98 파인만 적분론 장건수 46 금변론 박대현 99 응용 미생물학 박무영 47 생체에너지 주충노 100 리보플라빈 이상선 48 리이만 기하학 박을룡 101 노화 김숙희 • 김화영 49 군표현론 박승안 102 매트릭스 격리분광학 정기호 50 비선형 편미분 방정식론 하기식 103 신경계 조직배양 김승업 51 생체막 김형만 104 지구화학 김규한 52 수리분류학 고철환 105 은하계의 형성과 화학적 진화 53 찰스 다윈 정용재 이시우 외 54 금속부식 박용수