 신경과학
신경과학 박초냉 서울대학교 의과대학 졸 업 현재 서울대학교 의과대학 교수 및 서울대학교 의과대학 신경과학연구소 소장 김승업 서울대학교 의과대학 졸업 현재 캐나다브리티쉬 콜롬비아대학 교수
신경과학
 ` 드}-·l:~ L · m • T 『 •
` 드}-·l:~ L · m • T 『 •
 신경과학
신경과학
책 머리에 우리 몸은 약 6~1S 의 각기 기능을 달리하는 세포 들 로 이 루 어졌 다. 따라서 생체의 생명과 기능이 유지된다 함은 이 들 세포의 기능이 원활하게 이루어짐을 말한다 하겠다. 이 같 이 세포의 기능 을 원활하게 함으로써 생체 총체로서의 기능이 조화 를 이루게 하기 위하여 우리 생체에는 크게 두 가지 조절계 를 갖는다. 즉 신경계와 내분비(홀몬)계이다 . 내분비계는 생체에 대한 외래자극 또는 조직 장기로부터의 요구에 부응하여 내분비선으로부터 분비된 홀몬이 혈액을 통하여 원격 부위의 목표장기에 도달함으로써 해당 장기의 기능을 조절 하게 되는 것으로 우리의 일상생활의 통신방법으로 비유한다면 무선통신에 해당한다 할 수 있다 . 신경계는 주위 환경으로부터 각종 자극을 수용하여 신경섬유 를 통하여 중추에 전달하고 이를 감지, 분석하고 기억되었던 경험을 토대로 주어진 자극에 대처하는 적절한 명령(신호)을 주요 장기 에 신경섬유를 통하여 전달함으로써 해당 장기의 기능을 조절하 는 유선통신에 비유할 수 있는 조절계이다. 이같은 신경계는 1010_1011 개의 신경세포 (Neuron) 로 구성되어 있는 것으로 신경계의 성질을 알아보고자 하는 생명과학의 한 분야인 신경과학은 그동안 많은 학자들의 노력으로 물리학, 수 학, 화학 동 기초 학문의 발전과 더불어 세포생물학을 비롯한 면역학, 유전학, 분자생물학 동의 생명과학 관련 분야의 지식과 실험기법의 획기적인 발전으로 신경계의 형태학적 및 기능학적인 성질들이 많이 알려져 있으나 아직도 생명과학 분야 중 그 어느
분야보다도 풀어야 할 많은 문제들을 가지고 있는 미개척 분야라 하겠댜 따라서 이 책은 신경계 기능의 구성단위인 신경세포에서 일어 나는 현상을 간추려 소개하여서 생명과학을 공부하는 사람들의 신경계에 관한 이해를 돕고 나아가 유능한 젊은 과학도들로 하여 금 신경의 신비를 개척하는 데 관심을 기울여 주기를 기대하는 마음으로 신경과학의 입문서로서 마련하였다. 따라서 신경세포의 특 정기능을 깊이 있게 다루기보다는 신경세포에서 일어나는 현상 을 포괄적으로 다루었다. 제 1 장에서는 신경세포의 기본 구조와 뇌의 조직발생 그리고 신경전도 및 시납스의 성질들을 다루었고 제 2 장에서는 뇌신경계 의 발생과정과 뇌신경계의 형태학적, 전기생리학적, 신경화학적 및 조직배양적 연구방법을 개괄하였다. 제 3 장은 신경계의 전달물 질에 의한 자극전달 과정을 살펴보았고 제 4 장에서는 수용체의 성질과 그 연구방법 그리고 세포막에서의 자극도입 과정을 다루 었댜 이것이 우리나라에서의 신경과학 발전에 조그마한 도움이라 도 될 수 있다면 더없는 다행으로 생각한다. 그리고 이 책을 마련 하도록 도와주신 대우재단에 감사드린다. 1990 년 3 월 저자
신경과학
■ 차례 제1장 신경계의 기본구조와 기능 ―― 김승업 15 • 서론 16 • 신경계의 기본구조 18 • 뇌의 조직발생 20 • 신경세포(뉴론) 22 • 말초신경계 24 • 신경재생 24 • 신경흥분 전달 29 • 시납스 39 • 신경교세포 (그리아) 43 • 혈액-뇌 장벽 44 • 척수의 운동뉴론 46 • 소뇌 46 • 전뇌 코린계 뉴론 48 • 탈수질환제2장 뇌신경발생과 연구방법론 ―― 김승업
서론 • 51 신경계의 발생 • 52 중추신경계에서의 모노아민(카테콜아민)의 발현 • 55 신경성장인자 • 55 신경전달물질의 선택 • 58 시납스 형성 • 59 뇌신경연구법-형태학적 방법 • 61 전기 생리학 • 65 신경화학 • 65 신경 조직배양 • 66제3장 신경전달물질과 신경계의 생화학적 기능 ―― 박찬옹
69 • 서론 70 • 신경전달물질 71 • 신경전달물질의 동태 78 • 아세틸코린 86 • 카테콜아민 97 • 세로토닌 103 • 감마 아미노브티릴산 (GABA) 107 • 뉴로펩타이드 111 • 섭스탄스피 (SP) 114 • 오피오이드 펩타이드 116 • 프로스타그란딘 118 • 신경계의 생화학적 연구방법제4장 신경전달물질 수용체 ―― 박찬옹
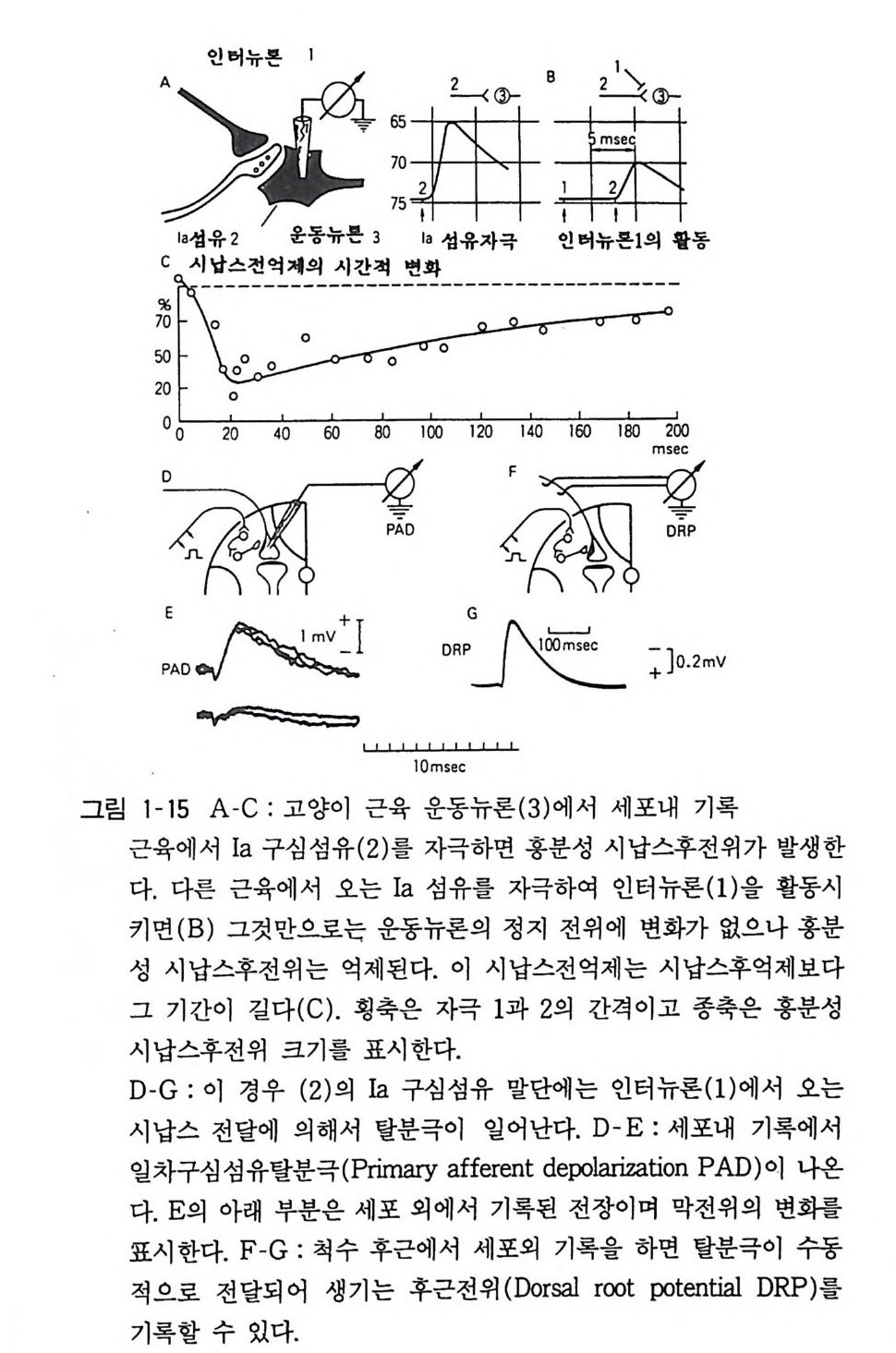
수용체의 분자량 측정 • 123 단크론 항체의 수용체 연구에의 응용 • 126 아세틸코린 수용체의 분자구조 • 128 수용체의 합성과 대사 • 131 이온통로 단백의 분자적 성질 • 133 자극도입 • 134 Ca++ -칼모듀린계 • 140 아데닐레이트 사이크라제 • 141 참고서적 • 147 용어해설 • 149 찾아보기 • 157그림목록 16• 그림 1-1 신경관의 발생 17• 그림 1-2 뇌의 발달 19• 그림 1-3 대뇌피질이 발달하는 시기에 보는 뉴론의 이동 19• 그림 1-4 대뇌피질의 구조 20• 그림 1-5 배양 인간 뉴론에서 보는 신경섬유 성장 21 •구림 1-6 여러가지 종류의 뉴론 23 •그림 1-7 25• 그림 1-8 세포 내외의 이온조성과 정지막전위의 발생 26• 그림 1-9 뉴론의 활동전위의 여러가지 27• 그림 1-10 활동전위가 발생할 때 보는 이온채널의 변화 28 •그림 1-11 시각계 뉴론의 옹답 30• 그림 1-12 개구리 신경-근접합부에서 본 시납스 전달 32• 그림 1-13 홍분성 시납스후전위 34• 그림 1-14 억제성 시납스후전위 37• 그림 1-15 39• 그림 1-16 반복자극 후 증강과 이종시납스간 촉진
40• 그림 1-1 7 해마 (H ippoca m p us) 뉴론회로와 장기적 증강 41 •그림 1-18 은염색 표본에서 보는 쥐의 오리고덴드로사이트와 아스트로사이트 42• 그림 1- 19 아스트로사이트 (A) 와 오리고덴드로사이트 (B) 모형도 45• 그림 1-2 0 운동뉴론이 어느 근육을 지배하는가에 대한 실험 47• 그림 1-2 1 소뇌의 조직구조 50• 그림 1-2 2 성인(사람) 뇌에서 분리 배양한 오리고덴드로사이트 53• 그림 2- 1 병아리 뇌의 구조와 기능이 발현하는 시기 54• 그림 2- 2 마우스 태아 뇌 각 부분에서의 도파민 (DA), 노르아드레나린 (NA) 세로토닌 (5 - HT) 형광이 발현하는 시기를 형광현미경으로 조사 56• 그림 2- 3 인간 태아 뉴론에 대한 신경성장인자 (NGF) 의 효과 57 • 그림 2-4 마우스 악하선에서 추출한 신경성장인자 (NGF) /3체인의 일차구조 58• 그림 2- 5 신생 쥐(랫트) 교감신경절 뉴론의 화학전달물질 분화에 대한 신경성장촉진인자의 영향 60• 그림 2-6 중추신경계 조직에서 보는 각종 시납스의 형상 62• 그림 2-7 신생 쥐(랫트) 교감신경절 뉴론의 카테콜아민 형광 63• 그림 2-8 병아리 척수 운동뉴론의 아세틸코린에스테라제 효소염색 64• 그림 2-9 인간뉴론을 단크론 항체로 형광염색한 뒤 관찰 71 •그림 3- 1 Loew i의 실험 개념도 73• 그림 3-2 신경전달물질의 동태 76 • 그림 3- 3 홍분성 또는 억제성 신경전달 과정 78 • 그림 3-4 축색류 79• 그림 3- 5 아세틸코린의 합성과 대사 80• 그림 3-6 흰쥐 뇌의 코린성 신경제 83• 그림 3-7 자율신경계 및 운동신경의 코린성 신경전달 85 • 그림 3-8 자율신경 코린성신경 전달과정에 작용하는 약물 87 • 그림 3-9 카테콜아민의 합성과 대사 88• 그림 3-10 흰쥐 뇌의 노르아드레나린 뉴론 88• 그림 3-11 흰쥐 뇌의 도파민뉴론 92• 그림 3-12 도파민뉴론의 생리학적 기능 95• 그림 3-13 아드레나린성약물의 작용부위 97• 그림 3-14 도파민뉴론에 작용하는 약물 98• 그림 3-15 세로토닌의 합성과 대사
98· 그림 3-16 흰쥐 뇌의 세로토닌뉴론 101 •그림 3-17 세로토닌의 생리적 작용 103 •그림 3- 18 세로토닌뉴론에 작용하는 약 물 104 · . 그림 3-19 GABA 대사와 GABA Shunt 105• 그림 3-2 0 GABA 뉴론의 합성과 대사 106· 그림 3-21 중추신경계에서의 GABA 의 억제작용 106• 그림 3-22 GABA 뉴론과 약물 115 •그림 3- 23 세 가지 오피오이드 펩타이드계의 전구단백과 각 오피오이드 펩타이드의 관계 ll7 • 그림 3-24 프로스타그란딘의 합성 경로 120• 그림 3-25 밀도차원심분리에 의한 뇌조직의 세포하분획의 제작 122• 그림 3-26 유전자 조작법에 의한 펩타이드 아미노산 배열 결정 124• 그림 4-1 광친화성 표식법과 SDS - PAGE 의 결과 125• 그림 4- 2 세포막단백 정제과정의 개념도 127• 그림 4-3 단크론 항체생성의 개념도 129• 그림 4- 4 세포막 아세틸코린 수용체의 모형도 130• 그림 4-5 리간드결합에 의한 수용체의 활성화와 탈감작의 원리 131 •그림 4-6 수용체의 세포내 운명 133• 그림 4 구 소디움 이온통로의 모형 134• 그림 4-8 세포막단백의 용해화와 재구성 원리 135• 그림 4-9 신호도입과정의 개념도 136• 그림 4-10 면역단백 자극에 의한 인지질메틸화와 히스타민 유리과정 137• 그림 4-11 포스파티딜이노시톨의 대사과정 138 • 그림 4- 12 포스파티딜이노시톨과 Ca++ 이동관계 139• 그림 4-13 포스파티덜이노시톨계를 통한 외래신호의 세포내 반응으로의 변화과정 140• 그림 4-14 칼모듀린의 작용기전 141 •그림 4- 15 칼모듀린의 여러가지 기능 142• 그림 4-16 칼모듀린 분자 143• 그림 4-17 수용체, GTP 결합단백, 아데닐산사이크라제 작용 144• 그림 4- 18 홍분성 또는 억제성 수용체와 GTP 결합단백 및 아데닐산사이크라제 상호관계
표목록 72• 표 3- 1 신경전달 물 질 및 후보물질 82• 표 3-2 아세틸코린 수용체의 분류 83 •표 3-3 부 교감신경 주효장기의 반옹 89• 표 3-4 카테콜아민 수용체의 분류와 리간드 91 •표 3- 5 교감신경 주효장기의 반응 94• 표 3-6 아드레나린성 약물 95• 표 3-7 아드레나린성 베타수용체 차단제 100• 표 3-8 세로토닌 수용체 109 • 표 3-9 전달물질후보 뉴로펩타이드 109• 표 3-10 기존 신경전달물질과 동시에 존재하는 뉴로펩타이드 111 •표 3- 11 섭스탄스피 및 유사 펩타이드 126• 표 4- 1 신경전달물질 수용체의 분자량 130• 표 4- 2 니코틴성 아세틸코린 수용체의 분자량 146• 표 4-3 아데닐레이트 사이크라제 촉진계와 억제계
 신경과학
신경과학
제 l 장 신경계의 기본구조와 기능 l 서론 신경계 를 구성하는 세포는 크게 나누어서 신경세포 (neuron) 와 신경교세포(gli a) 가 있다. 신경세포 혹은 뉴론에는 여러가지 종류가 있으며 이들은 서로 신경종말 혹은 시납스 (s yn a pse)에 의해서 교통을 하며 그 연결 패턴은 정확하게 결정되어 있어서 신경계 전체가 하나의 커다란 네트웍을 이루고 있다. 시납스에서 의 조절, 통합, 가소적 변화(p las ti c ity)가 있어서 신경 계의 모든 기능의 기본이 된다. 신경교세포 혹은 그리아(glia)의 기능에 대해서는 아직 불분명 한 점이 많은데, 최근에 뉴론-그리아 상호관계란 점에서 새로운 지견이 알려지고 이에 따라 그리아가 신경계 기능 유지에 큰 역할을 하고 있음을 알게 되었다 . 신경계의 병에는 선천적인 것 그리고 변성질환인 것이 많아서 그 진단과 치료는 현대 의학에서
다론 진료분과에 비하여 가장 뒤떨어졌다 할 수 있다 . 최 근 에는 이러한 신경계 질환의 원인을 세포 수준이나 분자 수준에서 분 석, 이해할 수 있게 되어 새로운 시대가 열리고 있다. 2 신경계의 기본구조 신경계의 구조를 이해함에 있어서 하동동물에서 고동동 물 에 이르는 계통발생학적인 뇌의 비교분석이 중요하다. 또한 뇌의 발생 과정을 개체발생의 면에서 분석함이 중요하다. 발생 초기에 수정란이 분열을 계속하여 초기배(신경배)가 되 고, 그 표면(외배엽)의 일부가 변화하여서 신경판 (neural pla te ) 이 된다. 신경계 조직은 모두 신경판에서 유래하며 신경관이 형성 되는 시점에서 신경계와 다른 조직과의 구별, 죽 분화가 이루어진 다. 이어서 신경구 (neural gr oove) 가 생기고 이것이 다시 분화하 여서 신경관 (neural t ube) 이 되며 신경판의 양단이 신경관에 들어 가지 않고 따로 독립하여 신경융 (neural cres t)이 된다. 신경융의 세포는 분열과 이동을 계속하여 감각 말초신경계나 자율신경계 를 만든다(그림 1-1) .
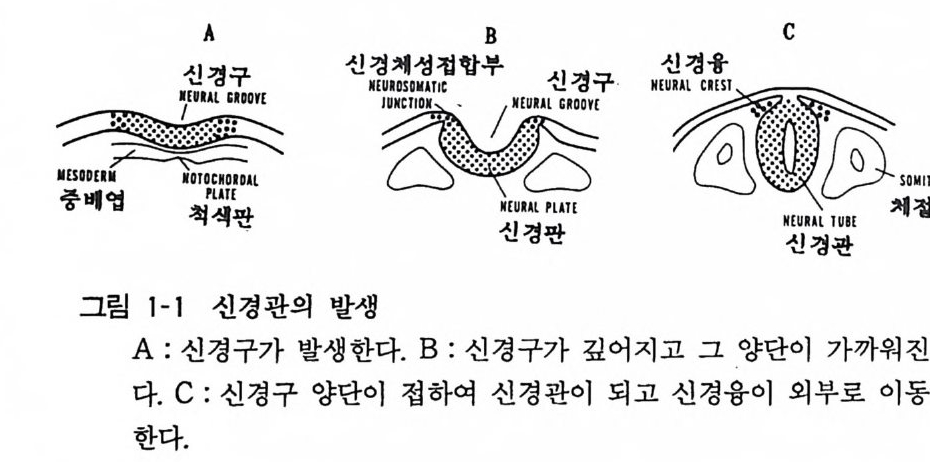 A
A
신경관이 발달하여 중추신경계 를 이루게 된다. 신경관 후부는 그 형태가 대체로 보존되어 척수 (s pin al cord) 가 되는데, 전부는 많은 변화를 계속하여 뇌의 각 부분을 구성한다(그림 1-2). 신경 관의 튜브는 뒤에 척수에서 중심관이 되고, 뇌에서는 뇌실이 된 댜 척추동물 말초신경계의 신경절이나 무척추동물의 뇌인 신경절 에서는 신경세포가 그룹지어 모여 있음에 비해서 척추동물 중추 신경계는 신경세포충 (shee t)이 여러가지 변화를 보이고 있다고 생각하면 된다. 신경관의 형성 이상이 있으면 척추이분 (s pin a b ifi da) 이라는 선천성 이상이 된다.
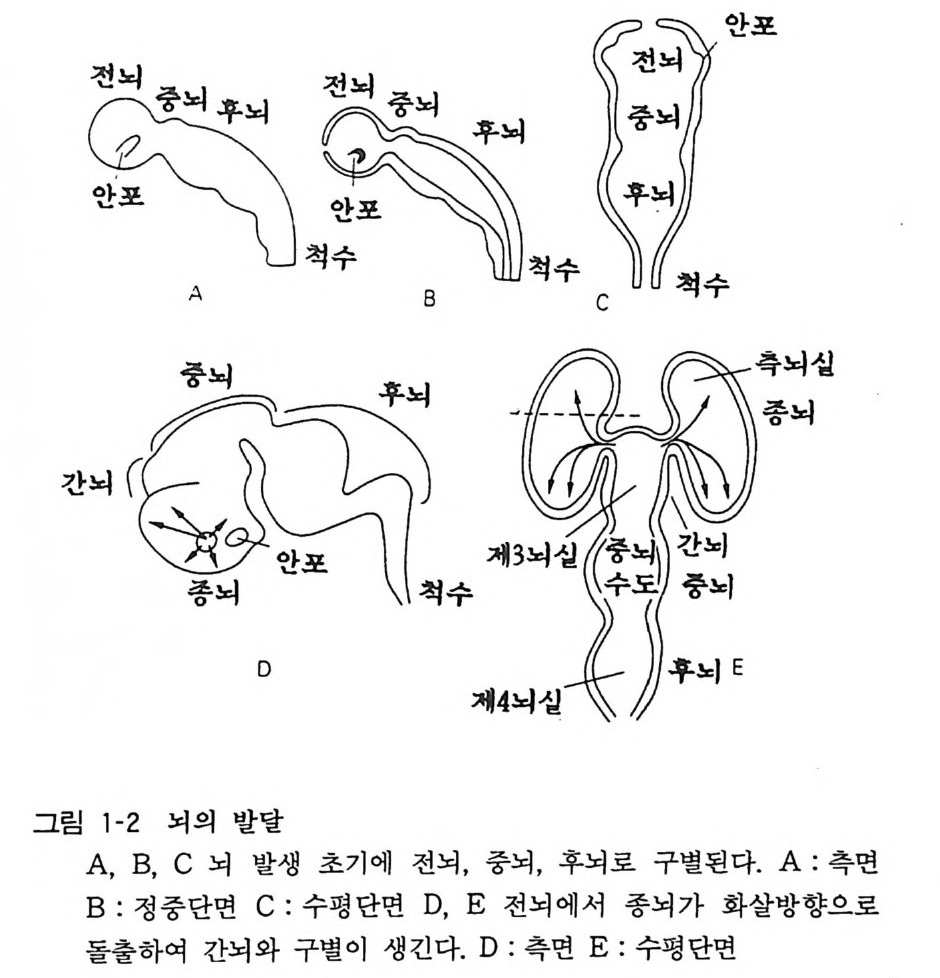 전뇌
전뇌
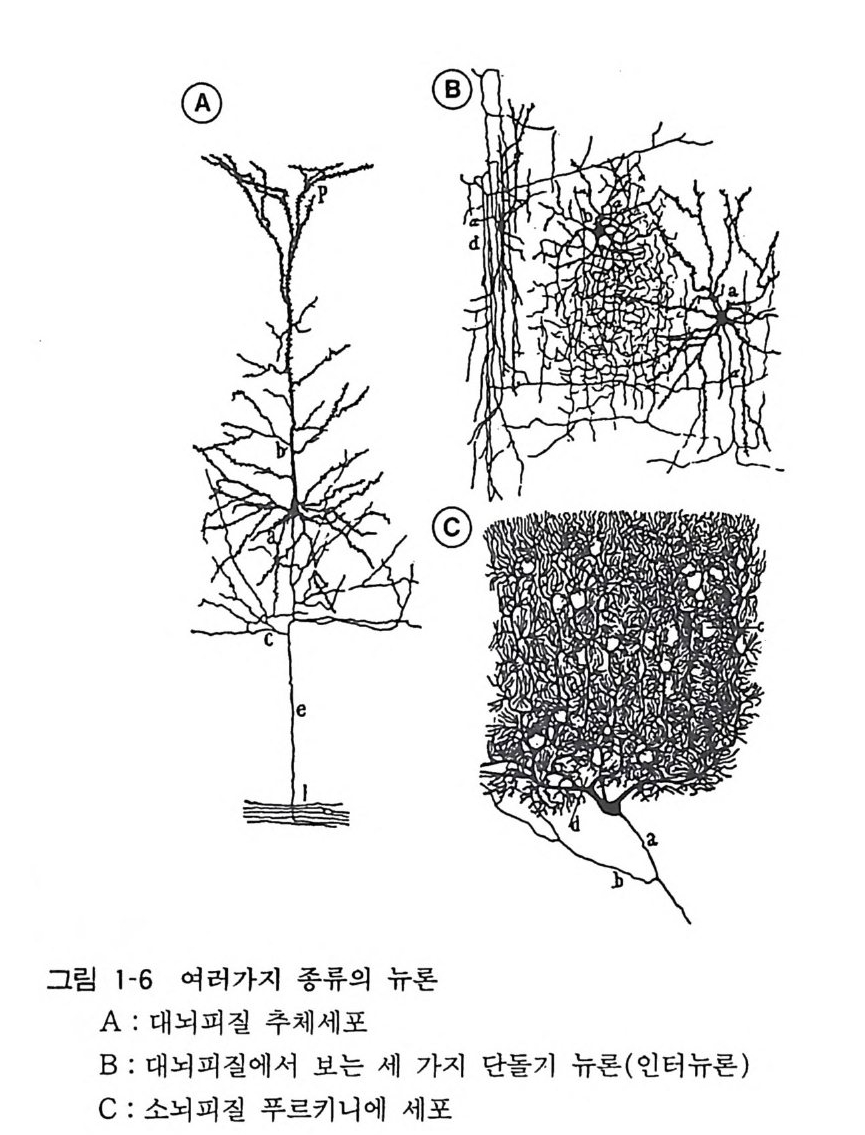
3 뇌의 조직발생 중추신경계의 세포는 신경세포와 신경교세포 혹 은 그리아의 두 종류가 있는데 이들은 모두 신경관 (neural t ube) 의 관벽의 세포(신경상피 세포)가 분열 증식하고 차례로 이동하여 구성된 다. 사람의 뇌신경 세포의 증가 동안에 DNA 의 양 을 측 정하여 보면, 임신 5 주에서 20 주까지 그리고 생 후 2 개월 정 도 에서 두 개의 피크가 있는데 앞의 피크 때 주로 뉴론의 증식이 있 고 , 뒤의 피크는 그리아의 증식시기에 해당된다. 대부분의 뉴 론 은 임신 초 8 주일 안에 분열을 마치는데, 소뇌 과립세포(gr anule cell) 와 같 이 생후 일년까지 분열하는 예외가 있기는 하다. 증식한 뉴론은 그자리에 눌러앉아서 성숙하는 것이 아니고 각기 일정한 장소로 이동한 다음 그곳에서 분화, 성숙한다. 그 결과 대뇌피질에서는 대충 6 층의 층 구조가 구성되고, 뇌간부에서 는 신경핵 (nucleus) 이 구성된다. 대뇌피질에서는 뇌실벽에서 형성 된 뉴론이 방사성 그리아 (ra di al gli a) 를 따라서 사닥다리 를 올라 가듯 그 돌기를 따라 이동하여 대뇌 표면을 향하여 움직인다 (그림 1-3). 뒤에 이동하는 뉴론은 먼저 이동한 세포 를 추월하여 표충으로 이동한다. 이 초기 발생기에 뉴론 이동에 이상이 있으면 뇌의 선천성 이상이 일어난다. 마우스의 선천성 이상에서 관찰되 는 리러 (Reeler) 나 위버 (Weaver) 는 대뇌나 소뇌의 충상 구조에 이상이 있어서 그에 수반하여 운동장애가 있고 오래 살지 못한다 (그림 1-4)•• 발생의 둘째 단계는 뉴론이 돌기를 신장하고, 다른 뉴론과의 사이에서 시납스 (s yn a p se) 에 의한 결합을 만드는 것이 다(그림 1-5). 뉴론 돌기에는 축색 (axon) 과 수상돌기 (den drit e) 의 두 가지가 있다(그림 1-6). 뇌 각 부분의 뉴론이 특유한 형태 의 수상돌기를 갖고, 축색을 신장하여 상대방의 뉴론을 정확하게 가려내고 그와 결합을 가지게 되는 메카니즘은 아직도 알려지지 않고있다.
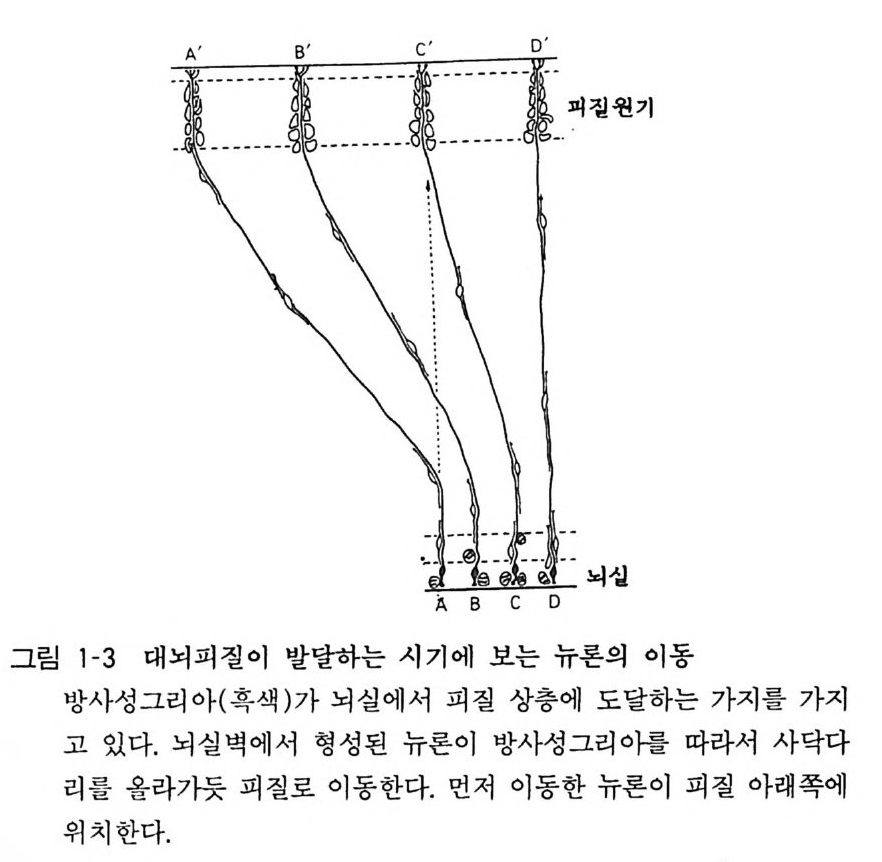 , 0’ F ' D'
, 0’ F ' D'
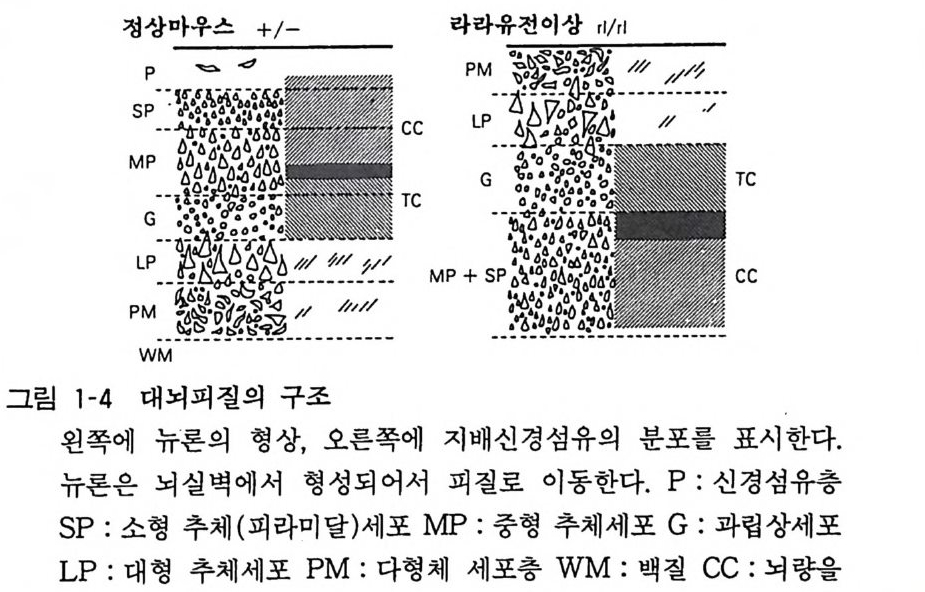 정상마우스 +/ - 라라유전
정상마우스 +/ - 라라유전
통하여 반대측에서 오는 섬유층 TC : 시상에서 오는 섬유 라라유전 이상에서는 충구조가 역전하고 있다.
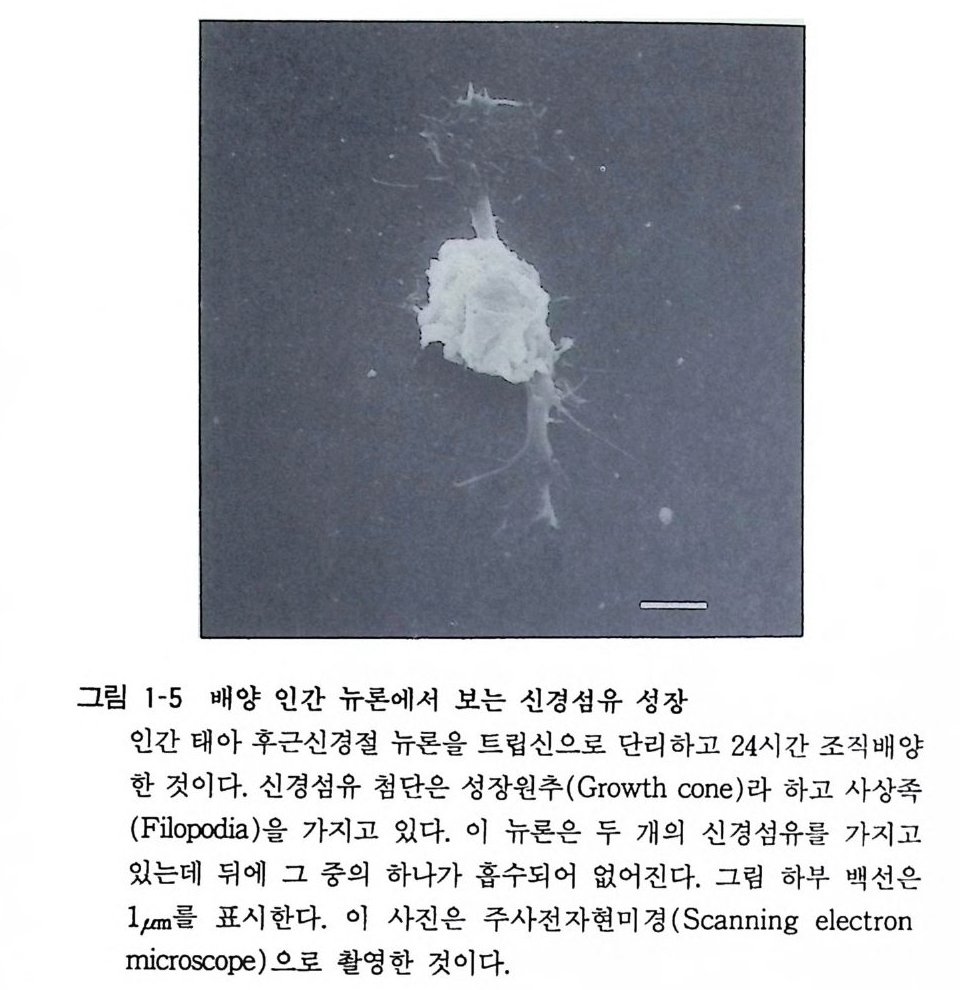 그림 1-5 배양 인간 뉴론에서 보는 신경섭유 성장
그림 1-5 배양 인간 뉴론에서 보는 신경섭유 성장
4 신경세포(뉴론) 뉴론의 기본적 형태는 세포체, 축색 그리고 수상돌기의 셋으로 구성되는데, 그림과 같이 다른 세포의 축색에서 . 신호를 받아들이
 ®
®
는 수상돌기, 자기 자신의 신호를 만드는 세포체 그리고 신호를 보내는 축색이 있다(그림 1-5). 소뇌의 푸르키니에 세포(p ur kinje cell) 의 수상돌기와 세포체에는 25 만 개 이상의 세포에서 축색이 와서 시납스를 만들고 있다. 척수전각의 운동뉴론에는 약 1 만 개의 시납스가 형성되어 있 다. 시납스전신경의 활동전위 (acti on pot en ti al) 는 시납스 전달의
결과 시 납스후전위 (post s y n ap tic po te n ti al, PSP) 로 바뀌 고 그 전체가 어느 수준을 넘으면 축색 기시부 (axon h ill ock) 에서 활동 전위가 발생하고 이것이 축색을 통해 전도가 이루어진다. 200 억이 넘는다고 추산되는 뉴론에는 많은 종류가 있는데 그 분류에 의하 면 멀리 떨어진 뇌 부분에 장거리 축색을 보내는 전달뉴론 (rela y neuron) 과 축색이 한 장소에 머물러서 그곳에서 정보변환이나 통합을 하는 국소회로 뉴론 (local cir c u it neuron) 이 있다. 후자는 개재뉴론 혹은 인터뉴론(i n t emeuron) 이라 불리기도 한다. 대뇌피 질의 전달뉴론은 주로 제 5 충에 있으며, 소뇌피질의 전달뉴론은 푸르키니에 세포가 그 역할을 한다. 소뇌피질에는 네 종류의 인터 뉴론이 있는데 그 수가 푸르키니에 세포의 100 배가 된다. • 축색의 종말이 만드는 시납스에는 흥분성 혹은 억제성의 두 종류가 있다. 예컨대 척수의 운동뉴론은 흥분성이고, 소뇌의 푸르 키니에 세포는 억제성이다. 뉴론은 신경전달물질에 따라서 분류할 수 있는데 척수 운동뉴 론은 아세틸코린을 방출하는 코린 작동성이고 푸르키니에 세포는 GABA 작동성이다. 한 개의 뉴론이 한 가지 신경전달물질을 방출한다는 학설이 과거에는 정설이었는데 이것을 데일 (Dale) 의 원리라 칭한다. 최근에 와서 한 개의 뉴론이 두 종류의 전달물 질을 가지는 경우와, 경우에 따라서는 한 가지 전달물질에서 다 른 전달물질로 바뀌는 수가 있다는 것이 알려져 있다. 이를 시납스의 가소성 (pla sti cit y) 이 라 한다. 5 말초신경계 척수 운동뉴론의 축색은 척수 전근 (ven tral roo t)에서 나와서 골격근육을 지배하고 그 밖에도 감각신경, 부교감신경이 온몸에 퍼져 분포하고 있어서 이들이 말초신경계를 구성한다. 감각신경,
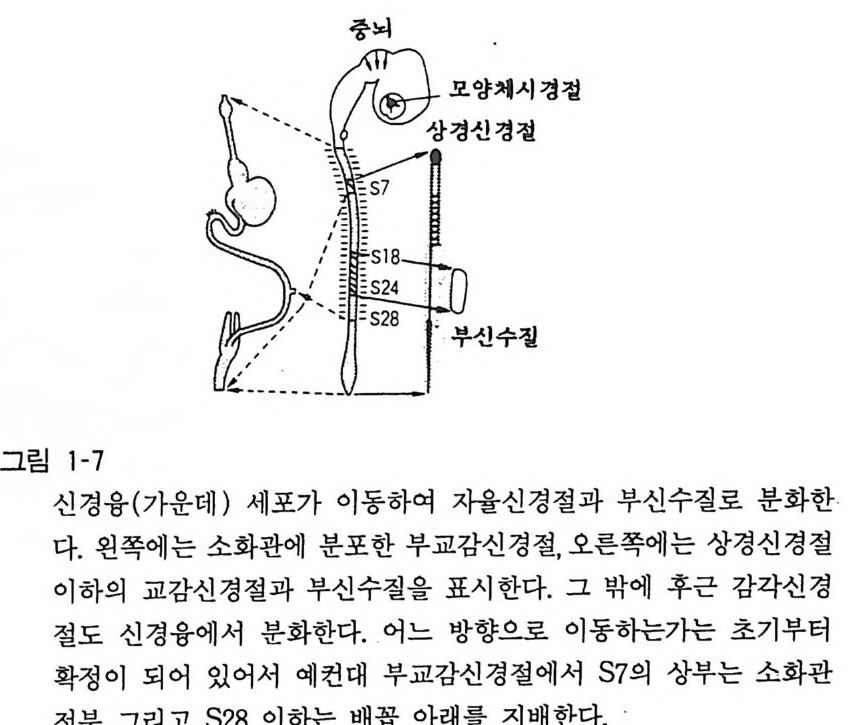
교감신경, 부 교감절후신경을 만드는 뉴론 들 이 접합하여 그룹이 되면 이 를 신경절(g an gli on) 이라 하며, 이것은 발생 초기에 신경 융 (neural crest) 세포가 이동하여 발달한 것이다(그림 1-7). 어떠 한 기전으로 인해서 이러한 이동이 정확하고 세밀하게 행하여지 는지 확실치가 않다. 그러나 최근 세포생물학적인 연구에 의해서 화이브로넥틴(fi bronec ti on), 라미닌 (l aminin), 코라젠 (colla g en) 그리 고 프로테오그라이칸(p ro t eo g l y can) 등의 세포간 물질 (extr a cellular ma t r i x) 이 이러한 세포이동예 중요한 역할을 한다는 것 이 알 려 지고 있다. 신경융의 세포는 발달 초기어 r는 무슨 종류의 세 포로 발전 하 는 지 결 정되어 있지 않고 이동중에 통과하는 조직 이나 종착 조직의 영향 을 받아서 분화한다는 것 이 확실하게 되었 다 (2 장 4 절 참 고 ).
 중뇌
중뇌
척수 전근에서 운동성 신경이 출발하고, 후근에서 감각성 신경 이 들어오고 있음이 정설로 되어 있다.
6 신경재생 온혈동물의 중추신경계에서는 외상 및 혈관장 애로 인하여 신경 섬유 (nerve fi ber) 가 절단되면, 이들이 재생하여 다시 결합 이 생기는 신경재생 (nerve re g enera ti on) 이 일어나지 않는다는 것 이 정설이다. 교통사고로 척수 손상이 있으면 일생 월 체어 생 활을 하게 마련이다. 최근 카나다의 아가요 (A gu a y o) 와 그 밖의 학자 들이 시신경과 척수에서 부분적이나마 재생이 일어남을 발표하 여 그 성과가 더욱 발전되기 를 기대하는 바 크다. 이 렇게 재생에 필요한 조건과 요소가 규명되면 이들이 치료에 적용되는 날이 가까운 줄 안다. 한편 말초신경계에서는 재생이 용이하게 일어나는 것이 특징 이 다. 상처에서 말단부의 신경섬유는 죽어버리나, 남은 슈완세포 (Schwarm cell) 와 그가 만드는 튜브를 따라서 상처의 중추부에서 신경섬유가 재생하여 말단으로 자라난다. 때로는 먼저대로의 길을 가지 않고 다른 방향으로 재생하여 나가서 그로 인하여 불완전한 기능회복이 올 수도 있다. 외상뿐만 아니라 신경염이나 당뇨병성 신경염에서도 신경재생이 필요한데 이룰 촉진하기 위하여 비타민 B 복합체, 스테로이드 홀몬, 갑상선 홀몬 등이 사용되어 부분적인 효과가 있음이 실증되었다. 최근에는 당지질인 강그리오사이드 (g an gli os i de) 가 신경재생에 효력이 있다 하여 주목받고 있다. 7 신경 흥분전달 세포의 안팎으로 이온 농도가 다르고 세포막의 이온통과가
제한되어 있으므로 세포 안팎으로 전위차 혹은 막전위 (membrane pot en ti al) 가 일어난다(그림 1-8). 양이온의 경우 세포내에는 밖에 비하여 포타쉬움 (K 기이 많고 소디움 (Na 기이 적다. 세포막은 K+ 은 통과시키나 N 하의 통과를 막는다. 그 결과 농도구배 혹은 농도경사(gr a di en t)로 인하여 K 저 나가고 전기적 중성이 된다. 이때 전위차가 발생하여 K + 를 세포 안으로 보내려는 움직임이 일어난다.
 A B
A B
B : 세포막 O.OOlµmXlµm 에서 본다면 K 가 농도구배에 의해서 안에 서 밖으로 나간다. 이에 따라 전기적 중성이 깨져서 전위차가 생기 고 K 를 세포 안으로 돌려보내려는 움직임이 생긴다. K· 6 이온이 세포 밖으로 나오면 평형이 이루어진다. 이때의 전위차는 정지막전 위 -90 mV 이다(막의 전기용량은 1 F/ c m2). 막에서 1 µm 이내에 있는 이온의 수는 그림에 표시되어 있다 . 0 . 00lµm 2 의 막에서 세포 밖으로 나가는 K 이온의 농도는 직 경 lOµm 에 서 50µm 에 달하는 뉴론에 있는 이온 총수에 비하면 보잘것없으나 그래도 이 6 개의 K 이온이 O.lV 나 되는 큰 막전위를 발생시킨다.
 A B
A B
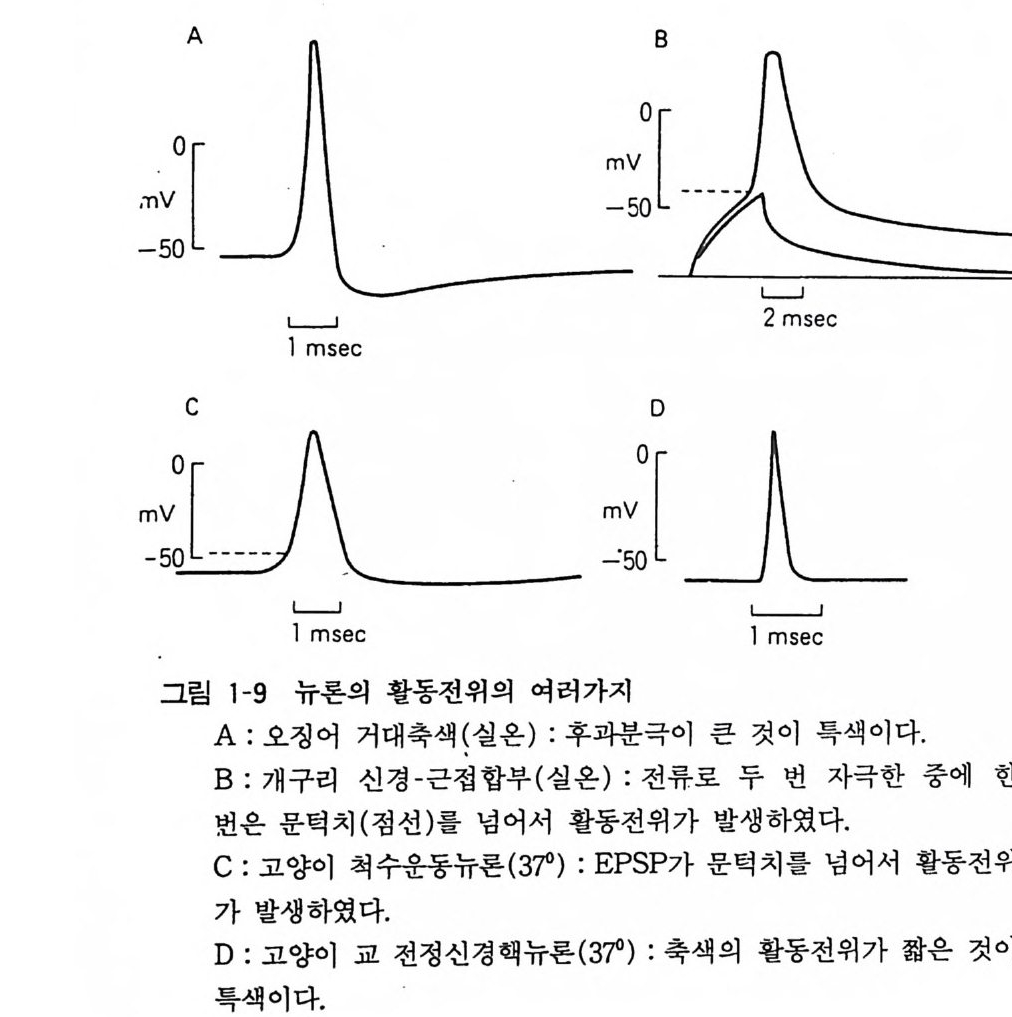
뉴론과 근육세포의 경우 이 정지막전위 (restin g pot en ti al) 가 (여ac러ti 가on지 p o자t e극n 에ti a l)의 이하다.여 정일지시막적전으위로는 변세화포한내다가. ―이 5것0이m V 활에동서전 —위9 OmV 인데 활동전위는 20mV 에서 50mV 이며 lmsec 정도 계속된 다(그림 1-9). 이것은 N 안에 대한 투과성이 일시적으로 올라가서 그때까지 통과 못하던 N 하가 한 번에 유입하기 때문이고 이어서 투 과성은 원상복귀하고 K+ 투과성이 증가함으로 막전위가 다시 정지 레벨로 돌아간다. 세포막에는 지질충 속에 단백질로 이루어진 이온채널(i on channel) 이 혼재하여 있고, 투과성이라 함은 이들 이온채널이 열려서 이온이 통과함을 의미한다. 활동전위에 관련된 Na+ 채널과
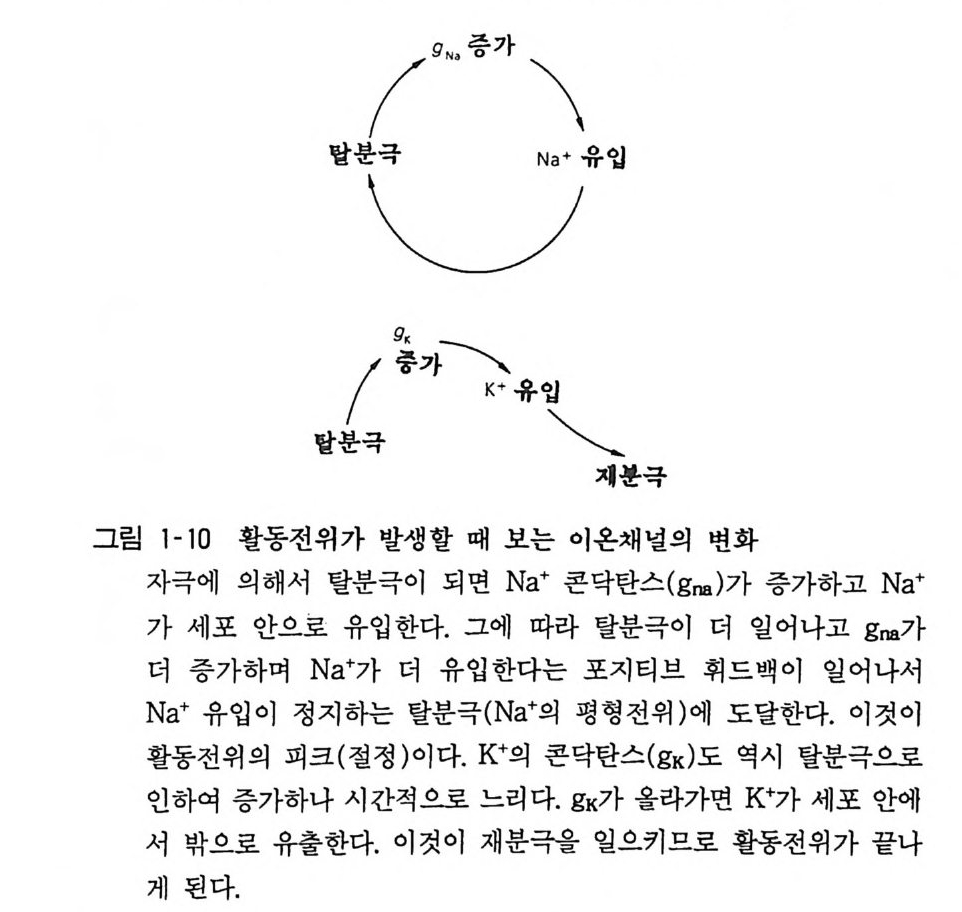 \( g N. 증가 \
\( g N. 증가 \
K 채널은 최근에 그 본질이 분자학적으로 규명이 되었다. N 하 투 과성은 막전위가 감소되면(탈분극) 증가한다. 이어서 N 하이 유입 하여 탈분극이 촉진되고 다시 더 N 하 투과성이 증가하는 포지티 브 휘드백 (po sit ive fee d-back) 현상이 일어난다(그림 1-1 0 ). 막전위가 어느 수준에 달하여 문턱치 이상으로 탈분극하면 자동 적으로 같은 레벨에 달하여 같은 크기의 활동전위가 된다.
 1 ) 중십부조사 巨 21 )) 毛둔뀝룹二
1 ) 중십부조사 巨 21 )) 毛둔뀝룹二
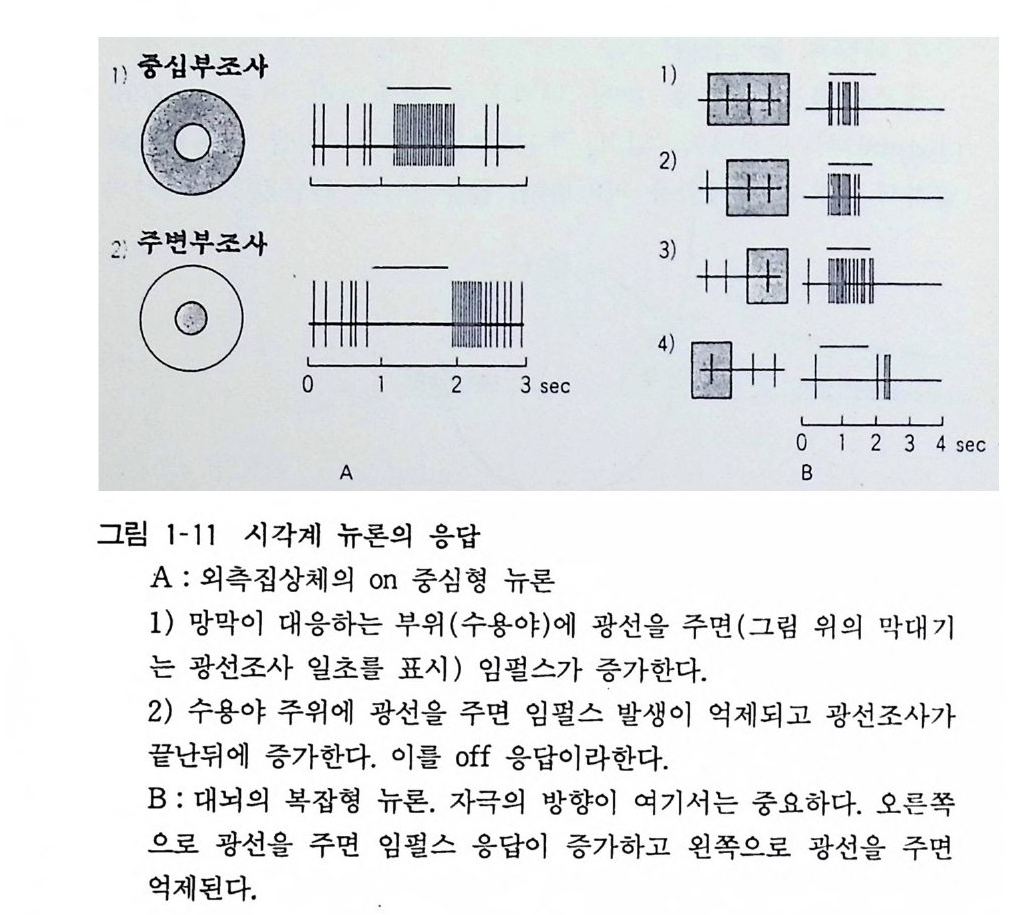
감각신경의 첨단에서는 자극에 의하여 기동전위(g ener~ t or pot en tial)가 일어나고 이것이 문턱치를 지나면 활동전위가 발생 한다. 뉴론에서는 시납스에서 생긴 시납스후전위 (PSP) 가 문턱치 이상이라면 활동전위가 발생한다. 축색에서는 그 축색의 일부에서
발생 한 활 동 전 위가 수동 적 으 로 전달 되어 문턱치 이상으 로 탈 분극 하면 그곳 에서 활 동 전 위 가 발 생 한 다. 이와 같 이 하여 길 다란 축색 에서 는 활 동전위가 장 거 리 전 도 를 이룩한다. 전 도 속 도는 축색의 직경 과 유수섬유냐 무 수섬유냐에 따라 결정되는데 일반적으로 1-1 2 0m / 초이다 . 활 동전위의 빈도에 의해서 신경계의 정보가 전달 되는데 이 를 임 펄스 (imp ulse) 라 한다. 눈에 빛 을 주면 시각뉴 론 에서 임 펄 스 빈도가 20 헬츠 (Herz) (희 /초)로 올라간다(그림 1- 1 1). 피 부 나 근 육 을 강하게 자극하면 그 부분의 감각신경이 수 백 헬츠 의 높은 임 펄스를 발생시킨다. 8 시납스 8. I 시납스전달 두 뉴론 사이의 접촉 부 인 시납스에서는 어떠한 정보전달이 이 루 어지는가. 이 메카니즘은 1950 년에서 1960 년대에 걸쳐서 신경 생리 학 의 가장 큰 중심문제였다. 영국의 캇츠 (Ka t z), 미렐디 (Miled i ), 미국의 쿠 훌 러 (Ku ffl er) 등은 신경 - 근육접합부 를 중심으 로 그리고 호주의 엑클스 (Eccles) 가 중추신경계를 써서 연구한 결 과 시납스 전달의 기구가 해명되었다 . 엑클스가 1964 년에 쓴 시납스의 생리학에는 그 연구가 상세하게 기재되어 있다. 시납스전 신경의 신경세포체 안에서 신경전달물질이 합성되 고, 축색에 의해서 수송되어 말단부로 가서 시납스소포 (s yn a pti c ves i cle) 에 저장이 된다. 이 시납스소포 안의 전달물질 농도는 극히 높고, 그 전달물질이 세포 밖으로 방출되는 것은 개구분비 (Exoc yt os i s) 에 의한다. 소포막과 세포막이 융합하여 시납스소포 내부와 세포 외부가 접속되고 소포 안의 전달물질이 밖으로 방출 되는 것이다. 이 융합과 방출은 신경의 정지 상태에서도 자발적으
로 일어난다. 운동신경의 경우 그 말단에서 자발 적 으 로 방 출 된 전달물질인 아세틸코린에 의해 근육섬유에 미소종판전위 (m ini atu r e endp la te poten ti al, MEPP) 가 발생한다(그림 1-1 2 ). 활 동 전위가 운동신경을 전도하여서 말단부까지 오면 대량의 아세틸코 린이 방출되고 근육에 종판전위 (endp la te pot en ti al) 가 발생 한다 (그림 1-12). 이때 대량의 시납스소포의 전달 물 질이 한꺼번에 방출된다. 이같이 방출되는 전달물질의 최소량은 일정하고 그
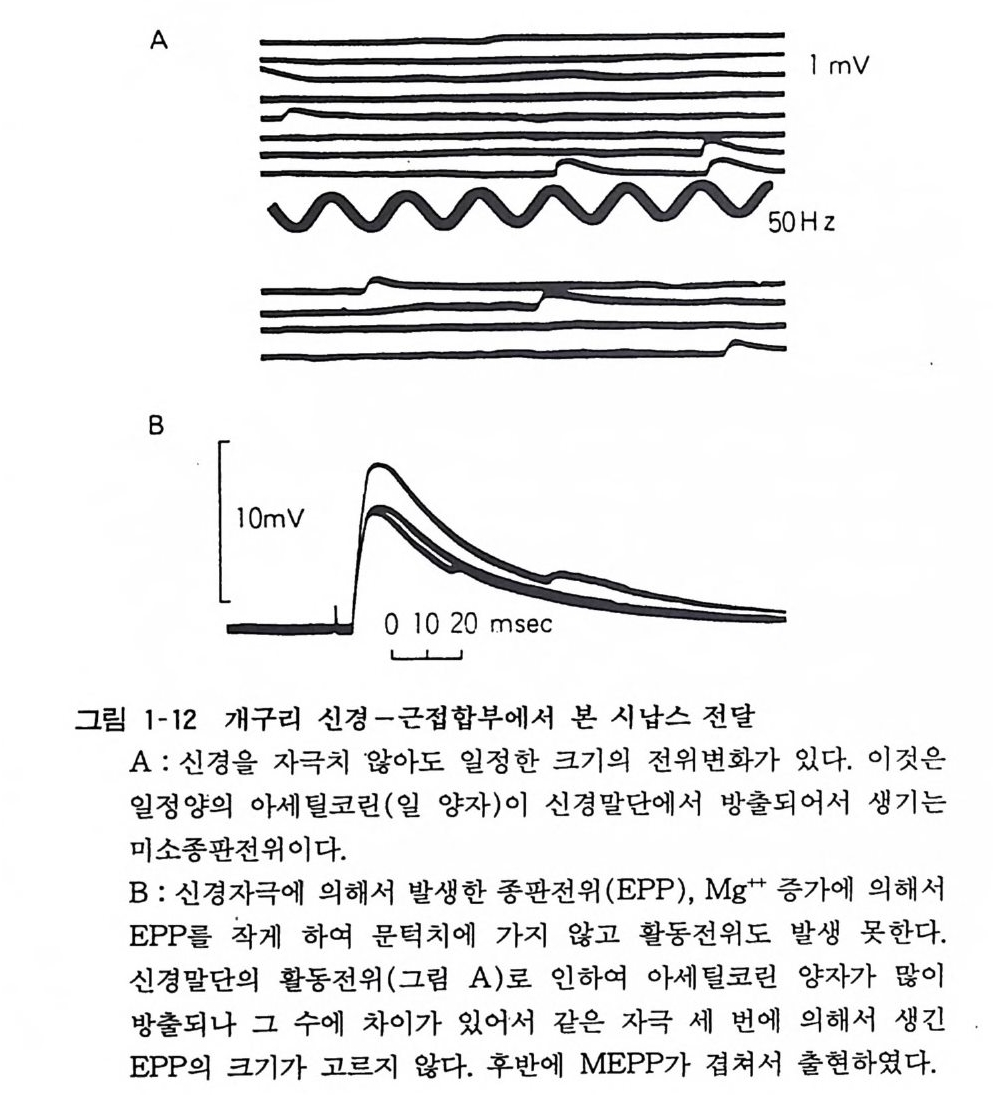 A
A
최 소 량의 배 수 의 율 로 서 방 출 량의 증가 가 일어난다. 이 같 은 방출 방 법을 양자 적 방 출 (Q uan t um release) 이라 하고 최소단위 를 양자(Q uan t um) 라 한다. 운동신 경 에 있어서 일 양자는 아세틸코 린 일만 분 자 가량이다. 활 동 전 위가 시납스에 도 달 하면 세포 밖에 는 칼 슘이 유입되어 그것이 전달물 질 방출의 방아쇠가 되어 방출 확률 이 올라간다. 칼슘 (Ca ++ )의 농도 를 십분의 일로 하고 마그네 슘 (M g 기 농 도를 올 리면 활동전위에는 영향 없이 전달물질 방출 을 억제 할 수가 있다. 시 납스 에서 방 출 된 전 달 물질 은 200-SOOA 의 좁은 시납스 간격 (sy n ap tic cle ft) 을 지나서 시납스후 세포에 있는 수용체 (rece p t or) 에 작 용한다. 전달물질 과 수용체가 결합하여 그 작용으로 이온채널이 열리 고 이온이 홀 러서 전위 변화가 일어난다 . 신경 근 육 접 합 부 (neuromscular j unc ti on) 의 경우 이 룰 종판전위라 하나 뇌에서는 일반 적 으로 시납스후전위라 하다. 8. 2 흥분성 시납스 흥분 성 시납스막에 있는 이온채널은 전달물질이 작용하면 열려 서 Na+ 과 K 지온을 통과시키고 Cl- 은 통과시키지 아니한댜 양 이온 을 무차별하게 투과시키면 막전위가 소실하게 된다. 죽 흥분 성 시납스후전위 (Excit ator y pos ts y n ap tic poten ti al, EPSP) 는 0 전위로 가까워지려는 변화로서 활동전위 발생의 문턱치를 지나 면 활동전위를 발생시킨다(그림 1-13). 신 경-근접합부나 부교감신 경절에서는 시납스전섬유 한 개의 활동으로 인한 시납스후세포에 생기는 흥분성 시납스후전위가 커서 그것만으로 활동전위가 발생 한댜 일반적으로 중추 시납스에서는 한 개의 시납스전섬유의 활동으로 조그만 홍분성 시납스후전위밖에 발쟁치 않고 활동전위 가 연속적으로 보내어지든가 각종의 시납스전신경이 동시에 활동 하여 흥분성 시납스후전위가 겹쳐서 그제야 시납스후세포가 홍분 한다. 전자를 홍분성 시납스후전위의 시간적 가중(t em p oral
summati on ), 후자는 공간 적 가중 (s pati al summa ti on) 이라 불 린다 (그림 1-13).
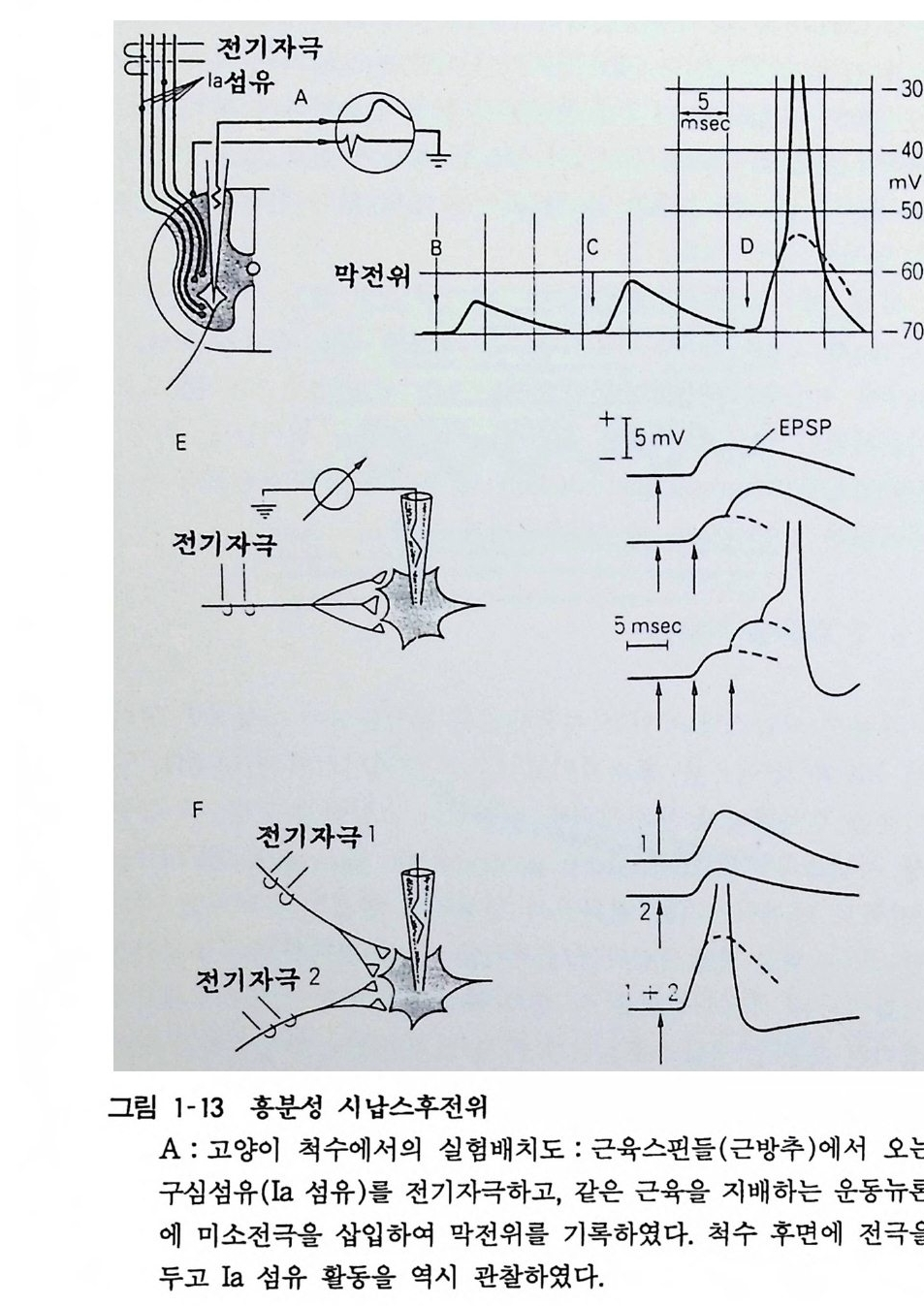 -30
-30
B-D : 자극을 크게 하면 활동하는 Ia 섬유수가 증가하고 흥분성 시납스후전위가 크게 되며 문턱치 를 지나면 (D) 활동전위가 발생하 여 척수전근에서 나오는 축색을 전도하여 나간다. E : 한 번의 자극으로 나오는 흥분성 시납스후전위가 작더라도 되풀 이하여 자극하면 흥분성 시납스후전위가 겹쳐서 커진다(이를 시간적 가중이라 한다) . F : 따로따로의 신경자극으로 나오는 흥분성 시납스후전위가 작더라 도 양쪽을 같이 자극하면 흥분성 시납스후전위가 겹쳐서 커진다. 이 를 공간적 가중이라 한다 . 8. 3 억제성 시납스 호주의 뢰위 (Loew i)가 1921 년에 시납스 전달은 화학전달물질 의 작용을 통해서 일어난다는 신경의 화학적 전달설을 입증하였 다. 이것은 부교감신경과 심장 사이에 있는 억제성 시납스에 관한 실험이었다. 중추신경계에서는 좀더 일찍이 금세기초에 셰링톤 (Sherr ingt on) 이 척수반사를 연구하여 이것이 억제현상임을 밝힌 바 있다. 그 뒤로 여러가지 학설이 나와서 시납스전신경이 둘로 나누어져서 하나는 흥분성 시납스 또 하나는 억제성 시납스를 만든다는 설, 신경말단에 도달한 활동전위의 전류가 직접 시납스 후세포에 들어가서 억제현상이 일어난다는 설 둥 여러 모로 혼란 한 시대가 있었다. 1951 년 호주의 엑클스 (Eccles) 가 고양이 척수 운동뉴론에 미소전극을 넣어서 억제성 시납스후전위를 처음으로 기록하였다(그림 1-14). 억제성 시납스는 홍분성 시납스나 마찬가 지로 시납스전 종말에서 전달물질이 방출되어 이에 의해서 시납 스후막의 이온 투과성이 올라가 억제성 시납스후전위가 발생한 다. 억제성 시납스만을 만드는 억제성 뉴론의 존재를 밝힌 것은 이갈이 엑클스의 업적이다. 엑클스는 1927 년 셰링톤의 연구실에 들어가서 그동안 흥분성 및 억제성 시납스의 연구를 계속한 바 있다. 갑각류(새우 종류)
에서는 척추동물과 달라서 홍분성 신경과 억제성 신경이 동시에 근육에 와서 시납스를 만들고 있어 이 를 이용하여 캇츠 (Ka tz), 쿠훌러 (Ku ffl er) , 타케우치 (Takeuc hi) 등의 여러 신경생리학자가 억제성 시납스의 연구를 하여 그 생리기구 를 해명한 바 있다.
 Sm v l :
Sm v l :
초기에는 억제성 뉴론은 척수전각에서 운동뉴론에 시납스를 만드는 Ia 개제 뉴론(그림 1-15) 이나 그 주위에 있는 렌쇼우 세포 (Renshaw cell) 와 같은 작은 뉴론이라 규정되었는데 소뇌의 푸르키니에 세포 (Pur kinj e cell) 와 같은 대형 뉴론이 역시 억제성 뉴론임이 밝혀지면서 억제성 뉴론은 흥분성 뉴론이나 마찬가지로 그 중요성이 인식되게 되었다. 억제성 시납스의 전달기구를 요약 하면 다음과 같다. 활동전위가 신경종말에 도달하여 전달물질이 방출되는 것은 홍분성이나 억제성 시납스에서나 마찬가지이다. 파상풍 (Te tan us) 독소는 척수에서 억제성 전달물질인 그리신 (Gl y c i ne) 의 방출을 저지한댜 이에 따라 억제성 시납스 작용이 저하되어 전신의 근육이 강하게 수축한다. 즉 과도의 흥분상태가 되는 것이다. 억제성 전달물질은 시납스후의 수용체에 결합하여 이온채널을 연다 . 홍분성 시납스에서는 양이온 투과성이 일어남에 반하여 억제성 시납스에서는 음이온채널이 열려서 음이온이 투과 한다. 8. 4 시납스전억제 시납스 기구에는 앞에서 언급한 것 이외에도 시납스전억제 (pre sy n ap tic i nh i b iti on) 가 있다. 이것은 흥분성 시납스의 시납스전 섬유 말단에 다른 신경섬유가 와서 시납스를 만들어서(축색-축색 간 시납스) 그 작용으로 흥분성 전달물질의 방출이 방지되고 흥분전달이 억제된다(그림 1-15). 앞서 말한 억제성 시납스는 시납스후억제(po s tsyn a pti c inhi b iti on) 라 하여 그 뉴론에 들어오 는 흥분성 시납스의 작용을 모두 억제한다. 그 반면 시납스전억제 는 일부의 흥분성 시납스의 활동만을 억제하는 것이다. 시납스전억제는 척수나 시상과 같은 감각경로에 많고 감각 정보를 세밀하게 처리하는 데 중요한 역할을 한다고 생각된다. 이 종류의 시납스에서는 흥분성 시납스전말단에 작용하여 그 부분을 탈분국한다. 그 부분에 활동전위가 전도해 오면 탈분극과
겹쳐서 활동전위의 높이가 작아진다. 전달물질의 방출량은 활동전 위의 크기에 따라 달라짐으로 방출은 억제된다. 이 경우 방출 양자 한 개 크기는 변치 않고 양자의 수가 감소한다. 8. 5 특수한 시납스기구 닭, 생선 그리고 무척추동물에서는 시납스전신경이 흥분하여 발생한 전류가 시납스후세포에 직접 홀러서 그 세포 를 흥분시키 거나 억제하는 수가 있다. 앞서 기술한 화학시납스와 구 별 해서 이들을 전기성 시납스라 한다 (2 장 그림 2-6 참고). 전기성 시납스를 전자현미경으로 보면 시납스전막과 시납스후 막 사이에 갭결합(g a p jun c ti on) 이 있어서 이 를 통하여 저분자 물질이 두 세포 사이를 이동하게 된다. 포유동물의 신경계에서도 가끔 1 갭결합을 볼 수가 있는데 이들이 전기성 시납스의 작용이 있는지는 분명하지 않다. 시납스전말단은 작고 그에 비해서 시납스후세포는 크므로 그 사이에 갭결합이 있어도 이것이 시납스후 세포의 흥분성을 바꿀 정도로 대량의 전류를 보낼 수가 없다. 그러므로 시납스전말단의 작용을 증폭하기 위해서 전달물질의 방출, 수용체와의 결합 등의 복잡한 과정이 필요한 것이다. 신경정보를 속히 전달하기 위해서 는 앞서 기술한세 가지 화학시납스(홍분성 시납스, 억제성 시납 스, 시납스전억제)가 중요한 역할을 하는데 그 밖에도 다른 시납 스 기구가 알려져 있다. 교감신경절 세포의 속도가 느린 시납스후 전위에서나 대뇌 뉴론의 아세틸코린 반응에서는 막전위가 변화하 고 있음에도 불구하고 이온 투과성이 저하되어 있다. 정지상태에 서 열려 있던 이온채널이 전달물질의 작용으로 오히려 닫혀 버리 게 되는 것이다. 사이크릭 AMP 를 포함한 이차 전달자 (second messen g er) 에 의해서 대사적 변화를 받는 시납스가 존재한다고도 생각된다. 엑클스는 이들을 메타보트로픽 (me t abo t ro pi c) 시납스라 하고
 7%C>0 I-시 \-납인o- 터-스-뉴건-튼-억- 계-t。-의l - -시--간적 번화三 :
7%C>0 I-시 \-납인o- 터-스-뉴건-튼-억- 계-t。-의l - -시--간적 번화三 :
이 온 투과성 에 인한 시 납스 를 이오노트로픽 (ion otr o p ic ) 시 납스라 분류하고 있다. 8. 6 시 납스의 가소성 (Plas ti c ity) 기억, 학습, 훈련 등의 고등 신경중추에서 일어나는 활동이 어떠한 기구로 이루어지는지 그 전체상은 확실치가 않다. 그러나 세포 수준에서의 연구에서는 시납스의 가소성, 즉 시납스전달의 장기적 변화가 기본이 된다는 시납스설이 가장 유력시된다. 경험 이 계속되고 반복되면 그러한 활동을 많이 한 신경에서는 그 시납스 사이즈가 커지거나 시납스의 수가 증가하리라 생각되는데 실제로 그러한 구조적인 변화가 오는지는 현재까지 알려진 바 없다. 이에 반해서 시납스를 되풀이하여 활동시킴으로써 기능적인 변화를 일으킬 수 있다는 증거는 있다. 척수 반응에서 자극을 단시간 안에 되풀이해서 주면 그 뒤에 반사가 크게 되는 현상은 반복자극 후 증강(po s t-t e tani c pot en ti a ti on) 이라 하며 그 일례라 하겠다. 이는 시납스전섬유 말단에서 막전위, 세포내 Ca++ 혹은 이차 전달자의 변화가 일어나서 다음에 오는 반사에 영향을 주는 것인데 수분 이내에 종결되므로 기억이나 학습을 이것으로 설명 할 수는 없다고 생각된다. 되풀이되는 자극이 그 뉴론 옆에 위치 한 뉴론에서도 증강효과를 가져오는 수가 있는데 이 를 이종시납 스간 촉진 (heter osy n ati c fac il itation ) 이 라 한다. 이 는 ' 활동을 되 풀 이하는 시납스전 말단에서 조절물질이 세포 외로 방출되어 그 옆의 시납스전 말단이 이를 섭취하고 시납스전달에 변화가 오는 것이라 생각된다(그림 1-16). 반복효과는 척수에서 수분의 단위로 일어남에 비하여 뇌에서는 수일의 단위로 일어나는 현상이 최근에 알려지게 되었다. 해마 (Hippocamp us) 의 구심섬유와 과립세포 사이의 시납스에서 이러 한 현상이 발견되었다. 이를 장기적 증강(l on g-t erm poten ti ation ) 이라 하는데 시납스후 성인 과립세포에서 Ca++ 농도가 증가하고
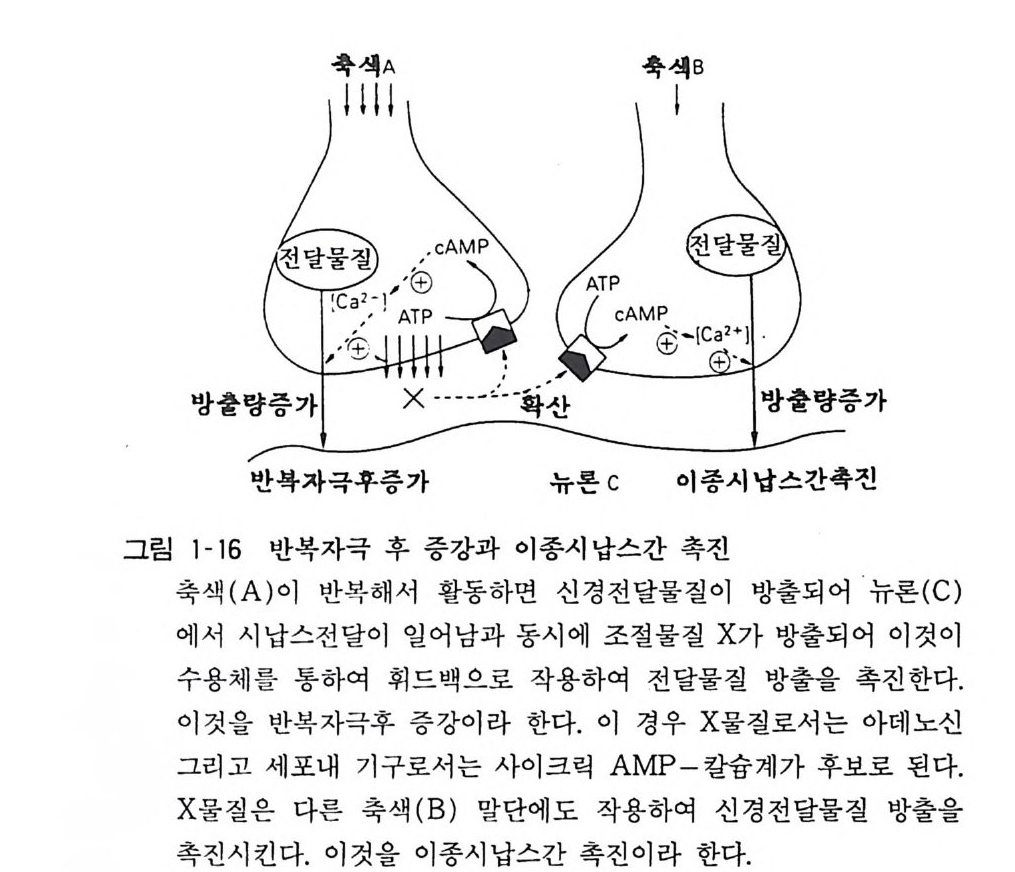 _축색lA I 축?
_축색lA I 축?
칼모듀린계가 매개하여 전달물질인 구루타만산 (Glu tami c acid ) 의 수용체 수가 증가하는 현상이다(그림 1-17). 장기적 억제 (lon g -ter m de p ress i on) 라는 현상도 알려져 있는데 이는 소뇌의 평행섬유-푸르키니에 세포간 시납스전달이 등상섬유 (cl im b ing fi ber) 에 의해서 장기간 억제를 받는 현상이다. 9 신경교세포(그리아) 중추신경계는 뉴론과 그리아의 두 가지 세포군으로 구성되어 있는데 그리아가 뉴론의 열 배 혹은 그 이상의 숫자가 있다고
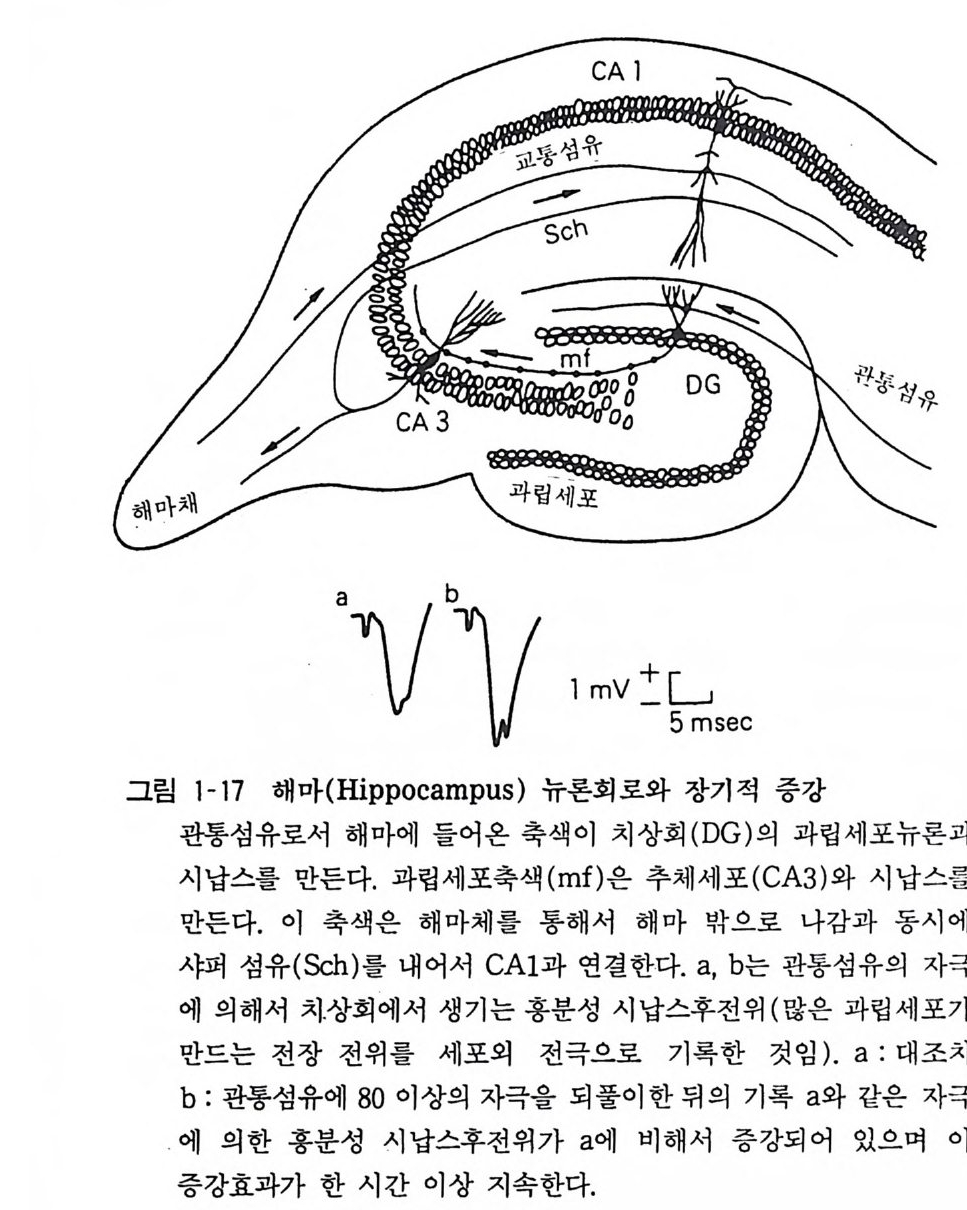 av bv l mV:~sec
av bv l mV:~sec
추산되고 있다 . 이들이 모두 의배엽에서 분화되어 나오는데 그리 아는 활동전위를 갖지 아니하고 정보전달에는 관여하지 않는 것으로 인식되고 있다. 뇌가 병에 걸리거나 상처를 입는 경우 그리아는 뉴론과 달리 분열증식하게 된다. 이렇게 분열증식하는 그리 아는 성상교세포 혹은 아스트로사이트 (As tr oc yt e) 가 대부분
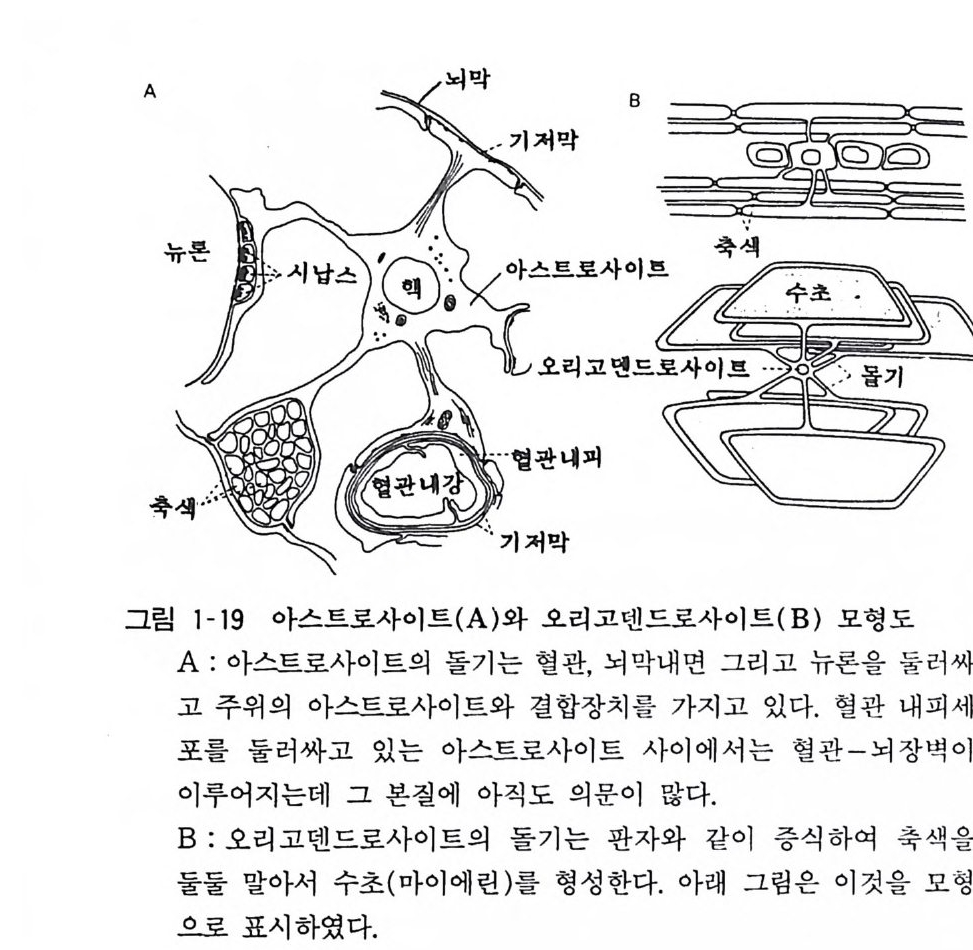
이다. 중추신경계의 그리아는 아스트로사이트와 오리고덴드로사이 트 (Ol ig odendroc yt e) 혹은 희돌기 교세포로 이분되고(그림 1- 18) 말초신경계에서는 슈완세포 (Schwarm cell) 가 있다. 아스트로 사이트 혹은 성상교세포는 그 이름이 말하듯 여러 개의 돌기를 방사성으로 가지고 있고 그 돌기의 말단은 손발같이 퍼져서 뉴 론, 혈관, 뇌 표면의 뇌막에 접해서 기저막 (basemen t membrane) 이라는 접착면을 가진다(그림 1-19). 오리고덴드로사이트와 슈완세포는 중추신경계와 말초신경계에 서 각각 신경섬유 축색을 감아서 수초 (m y e lin shea th)를 형성한 다. 수초는 신경막에서의 절연도를 높이고 활동전위의 전달속도를 높이는 역할을 하고 있다. 오리고덴드로사이트는 그이름 그대로 돌기 수가 적은 세포라 하여 명명되었는데 그 이름의 현실성은 없으나 오랫동안 사용해 온 관습으로 그대로 쓰이고 있다. 하나의 오리고덴드로사이트가 삼십에서 사십 개의 신경섬유를 감을 수
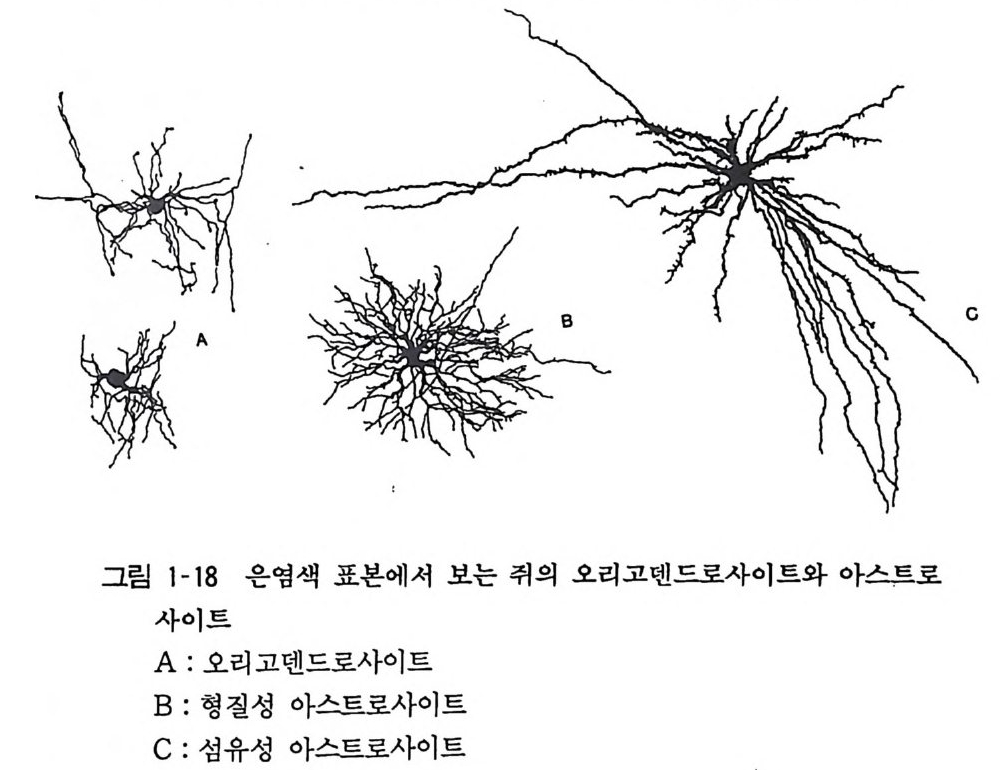 결
결
 A
A
있다는 것이 현재 알려져 있다(그림 1-19). 독일의 병리학자 빌효 우 (V ir chow) 가 1846 년에 처음 그리아 세포에 대하여 기술하였는 데 그는 이 세포들이 뉴론 사이를 집착하는 세포들이라 하여 그리아라 명명하였다. 그 뒤 1883 년에 이탈리아 골지 (Gol gi)가 은염색법이란 특이한 조직염색법을 고안하여 그리아 돌기가 혈관 과 뉴론 사이를 연결하고 있음을 발견하였다• 그래서 그리아는 피 속의 영양물질을 뉴론에 운반하고 그 노폐물을 혈중으로 운반 하는 것이라 생각하였다(골지가 기재한 세포는 지금의 아스트로 사이트롤 의미한다). 현재로서는 영양물질이 아스트로사이트를 통하지 않고도 세포간격을 확산하여 전달될 수 있어서 이러한
역할은 필요치 않다고 본다. 과거에는 아스트로사이트가 신경전달 의 혼선을 막는다고도 생각되었는데 전류가 저항이 높은 세포막 을 지나서 다음 세포로 전달될 수 없으므로 이 학설도 통용되지 않는다. 최근의 연구에 의하면 아스트로사이트가 신경전달물질을 섭취 제거하고 성장영양물질을 생산하는 두 가지 중요한 역할을 하고 있다고 생각된다. 시납스에서 방출된 전달물질을 제거하는 기전에는 분해효소에 의한 제거, 시납스전 말단에 의한 재홉취 그리고 확산에 의한 희석의 세 가지 기전이 있는데 이에 더하여 GABA 와 같은 아미 노산계 전달물질과 카테콜아민은 주위에 있는 아스트로사이토가 섭취 제거한다고 생각된다. 과거에는 그리아가 분비기관이란 가설 이 있었는데, 최근의 조직 배양실험에 의하면 뉴론의 생존이나 재생을 촉진하는 성장물질이 아스트로사이트에 의해서 생산됨을 알게 되었다. 그 성장물질을 순수하게 분리하고 그 본태를 구명하 는 연구가 여러 곳에서 수행되고 있다. 10 혈액여 장벽 혈액-뇌 장벽 (blood-br ain ba rri er) 은 혈액에서 뇌 실질에 가는 물질이동을 조절하는 장벽이고 이것은 뇌조직의 방어 기구라 생각할 수 있다. 중추신경계에 작용하는 약물, 독물의 작용발현에 중요한 의미가 있다. 많은 물질이 이 장벽에서 투과 제한을 받아 서 뇌에 들어가지 못하가 때문이다. 현재로서는 모세혈관을 구성 하는 혈관 내피세포와 그를 둘러싸고 있는 아스트로사이트 그리 고 이 두 가지 세포 사이에 존재하는 기저막이 혈관 ;1 장벽에서 중요한 역할을 한다고 생각되는데 그 각자의 기능에 대하여서는 명백하지 않다(그림 1-19). 포유동물의 초생기(사람에서는 생후 두 달까지)에는 이 장벽이
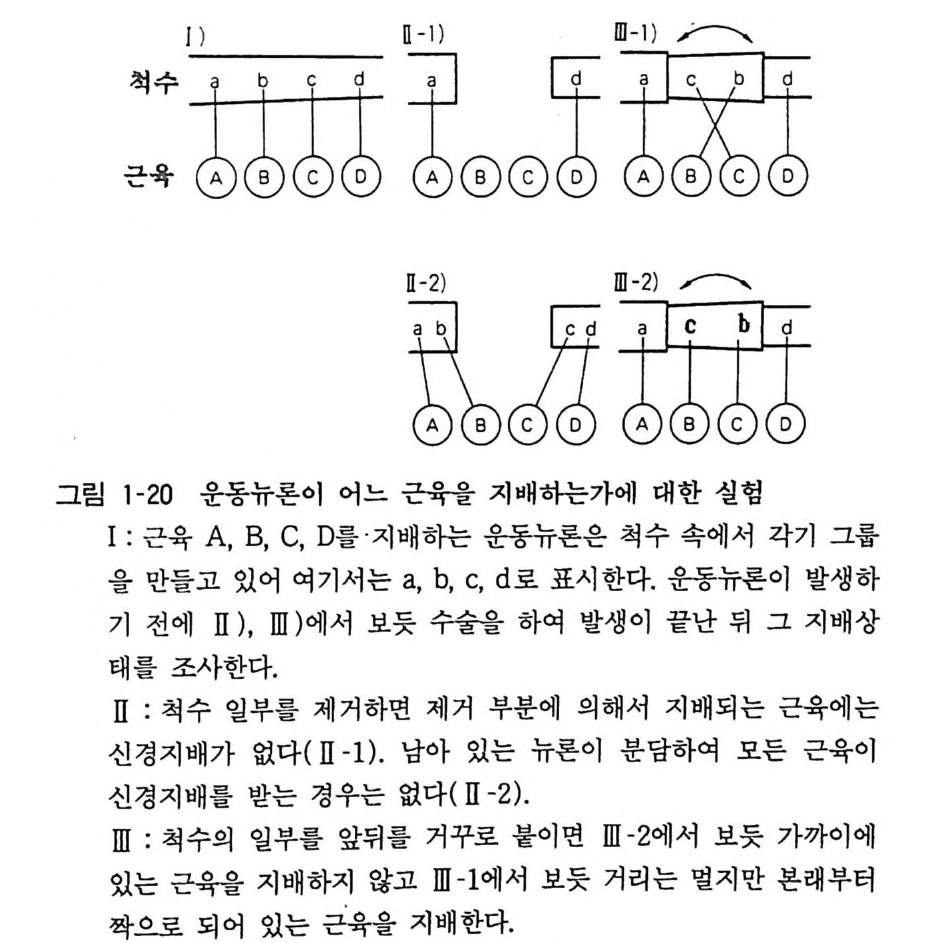
발달하지 않아서 대부분의 물질에 대하여 뇌가 무방 비상태에 있댜 신생아 소아과 환자의 특수성이 여기에 있다. 11 척수의 운동뉴론 여기서는 척수의 운동뉴론의 예를 통하여 중추신경계의 구조와 기능에 대하여 고찰하고자 한다. 골격근을 지배하는 운동뉴론은 척수의 전각에 있다. 같은 근육에 신경섬유 를 보내는 뉴론은 대체 로 한 곳에 모여서 다른 근육을 지배하는 뉴론군과 따로 있어서 이를 운동뉴론 그룹이라 한다. 운동뉴론의 수는 발생 초기에는 성숙기보다 50% 가량 더 많은 데 신경섬유가 근육에 도착하여 신경-근접합부 를 만드는 시기에 많은 뉴론이 퇴화 사멸하여 그 수가 감소한다. 이 룰 계획적 세포 사(p ro gr ammed cell dea th)라 한다. 운동뉴론이 어떻게 발생 분화하는가라는 문제는 극히 홍미있는 일인데 미국의 랜드맷서 (Landmesser) 는 초기의 병아리 배에 미소수술을 하여 실험을 하였다(그림 1- 2 0). 운동뉴론이 겨우 나오기 시작한 이틀 반 정도의 이른 시기에 척수의 일부 를 제거 하고 근육지배를 조사하였다. 만일 이 시기에 어느 뉴론이 어느 근육을 지배하는가 정해지지 않았다면 남아 있는 척수뉴론이 전근육을 지배한다고 생각할 수 있다. 결과적으로 보면 제거된 척수신경에 대응된 근육에는 신경지배가 없어서 발달 초기부터 운동뉴론의 분화가 결정되어 있음을 알 수가 있다. 잘라낸 척수의 위 아래를 뒤집어서 이식하여 그 결과를 보니 척수신경의 섬유는 먼 길을 돌고 돌아서 본래의 근육을 지배함을 알았다. 신경발생이 유전적으로 결정되는가 혹은 환경에 의해서 조정되 는가 하는 두 가지 기본입장이 있는데 앞서 말한 실험은 유전인 자를 지지하는 입장이겠다. 계획적 세포사가 일어나는 것은 정확
한 신경회로가 구성됨에 있어서 중요한 뜻이 있겠는데, 신경회로 가 처음에는 극히 우발적으로 구성되고 그릇된 회로는 제거되는 과정을 거쳐서 신경지배계가 완성되는 것이라 한다면 세포사는 잘못된 회로를 없애는 것이라 하겠다. 그러나 신경-근차단제를 써서 세포사를 방지한 동물에 있어서도 운동뉴론 지배가 정확하 게 구성됨을 볼 때 세포사는 그릇된 뉴론을 제거하는 목적이라기 보다 모든 근육섬유가 빠짐없이 신경지배를 받도록 보장하는 메카니즘이라 볼 수가 있겠다. 즉 필요 이상의 다수의 뉴론이 존재하여 신경지배를 완성한 뒤 불필요한 뉴론이 제거되는 기전 이라 생각하면 된다.
 :I ) :二Il-二1) 〔〔ID- 1) /广一
:I ) :二Il-二1) 〔〔ID- 1) /广一
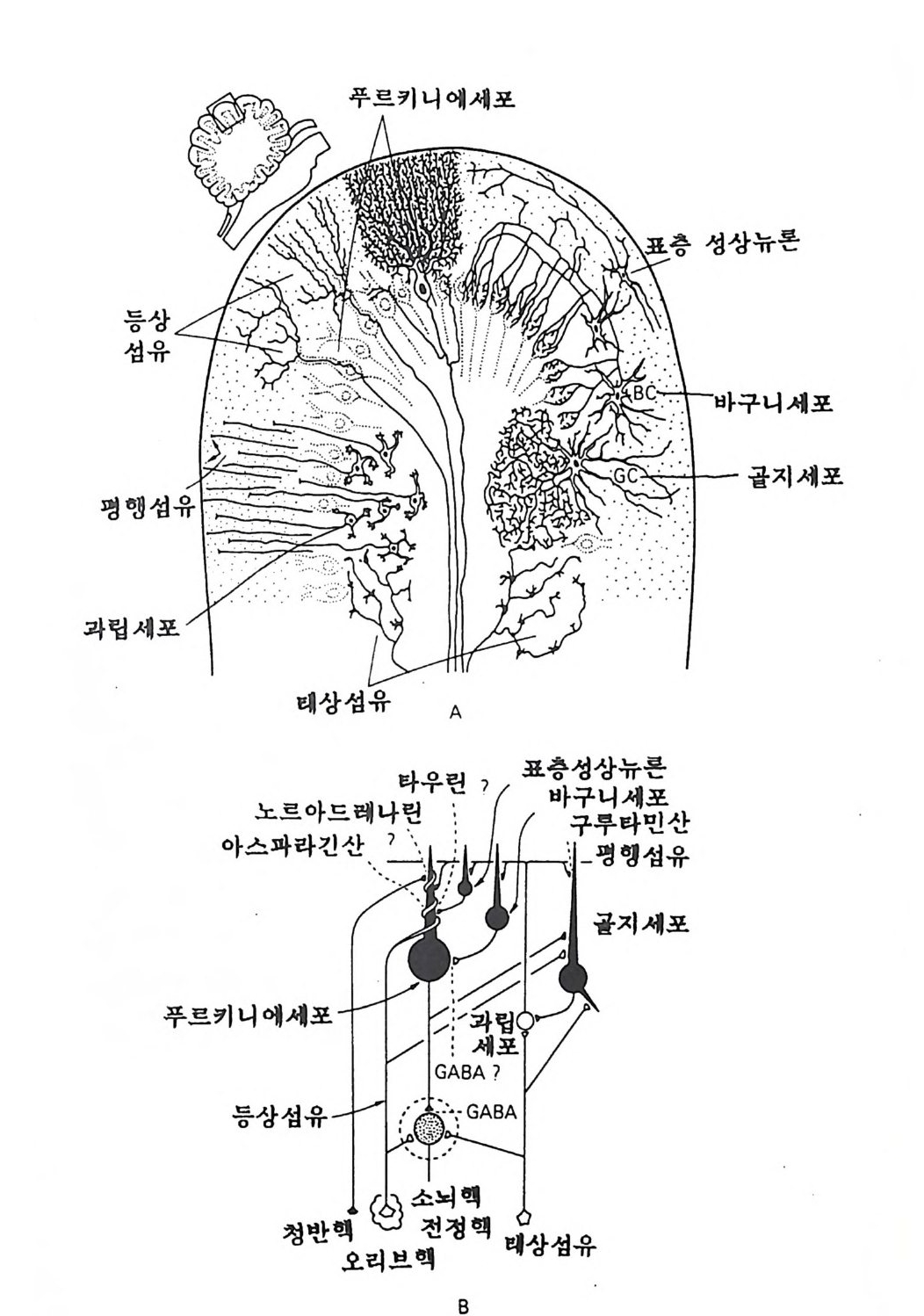
12 소뇌 소뇌에 종양이 있다든가 소뇌외상이 있는 경우에 운동과 자세 에 장애가 와서 운동협조가 없어진다. 소뇌피질의 다섯 가지 뉴론 이 각각 정확한 층 구조를 이루고 있음은 금 세기초 은염색법을 이용한 카할 (Ca j al) 의 연구에 의해서 상세히 기 록 되었다(그림 1-21 ). 카할은 이러한 신경조직학의 기초 를 쌓아서 1913 년에 전기 한 이탈리아의 골지와 함께 노벨의학상을 수여받았다. 소뇌 각 뉴론의 기능과 그 결합 형식은 1960 년대에 엑콜스 (Eccles) 일파가 행한 전기생리학실험의 결과로 명백하게 되었다 (그림 1-21). 이러한 소뇌의 구조와 기능에 관한 기초적인 성과에 더하여 소뇌 전체의 시스템으로서의 기능에 대한 연구가 계속되 고 있댜 13 전뇌 코린계 뉴론 최근에는 인구의 고령화가 사회적으로 그리고 의학적으로 중요 한 문제가 되고 있다. 노년층의 65 세 이상 인구 중에서 10- 15 % 가량이 치매(노망)의 증세를 갖는다. 기억상실, 인지불능, 계산불 능에 더하여 장소와 시간의 관념이 흐려지는(지남력 상실) 증세 가 있게 된다. 이들 치매 증세를 가진 고령자의 60% 정도가 알츠 하이머병 (Alzheim e r's dise ase) 환자라 진단되는데 그 병인은 불명이며 현재 생화학적 분자학적인 연구가 각국에서 성행하고 있다. 알츠하이머 환자의 뇌는 육안적으로 확실히 위축되어 있으며, 광학현미경에서는 노인반 (se nil e p la q ue) 과 신경원섬유의 변화 ( neurofi br ill ar y tan g le s) 와 같은 병 리 적 변화가 있다. 전 자는
 등상
등상
그림 1-21 소뇌의 조직구조 A : 이십세기초 스페인의 노벨상 수상자 카할 (Ca ja l) 이 은염색 표본 을 중심으로 만든 모식도. 소뇌를 정중선에서 잘라서 만들어 보면(왼쪽 위) 소뇌는 다수의 소엽으로 구성되어 있다. 그 소엽 하나 를 확대하면 아래와 같 다. 표면에서부터 분자충, 푸르키니에 세포층, 과립세포충 그리고 안에 백질로 구성된다. B : 현재 알려지고 있는 소뇌 뉴론회로의 상세 를 보이는 모식도. 흥분성 뉴론은 백색, 억제성 뉴론은 흑색 으로 표시하였고 그들 이 사용하는 신경전달 물질도 표시하였다. 아미로이드섬유 (am y lo i d) 가 세포 외에 축적되고 그것을 변성 퇴화한 신경돌기가 신경종말을 포위하고 있는 것이며, 후자는 뉴론 속에 뉴로피라멘트 (neuro fil amen t)와 그에 관련된 포 리 펩타 이드가 집합한 것이라 생각된다. 최근에는 이러한 알츠하이머병 환자의 뇌에서 전뇌 기저부의 코린계 뉴론이 변성 소멸하고 있다 는 소견이 있어서 아세틸코린계의 사멸이 알츠하이머병의 배경 병인이라 설명하는 경향이 있다. 파킨손병 (Parki ns on's di sease) 에서 엘 -도파 (L - DOPA) 를 쓰는 치료법과 마찬가지로 알츠하이머병 환자에서 아세틸코린을 보충 하자는 관점에서 아세틸코린의 전구체인 코린이나 그 복합체인 레시친을 대량 투여하는 치료법이 시험적으로 쓰여지고 있으며 저하된 아세틸코린성 전달을 개선하기 위하여 항아세틸코린에스 테라제 약인 피소스티그민 (Ph y sos tigmin e) 의 시험적 투여가 역시 사용되고 있다. 그러나 이러한 시험적 치료법이 별로 효과를 못 본다는 임상보고가 최근에 발표되어 있다. 14 탈수질환 수초 혹은 마이에린 (m y e lin shea th)이 파괴되는 중추신경 병이
있는데 이들을 탈수질환이라 한다 . 이 경우 수초와 그것을 만드는 오리고덴드로사이트는 파괴 소실되나 그 속의 축색은 보존된다. 이때 수초의 소실로 신경정보 전달이 저해되는 것이다. 탈수질환 중에서 다발성 경화증 (mul tip le scleros i s) 은 미국에서 인구 천 명에 하나, 전국에 20 만 명 이상의 환자가 있는 것으로 판명되고 있다. 이 병은 우리나라에서는 아직 그 환자 케이스의 발표가 없으나 이것은 진단법이 확립되지 아니한 탓이라 생각되며 일본 의 경우 를 미루어 보아서 10 만 명에 하나 정도의 발생률이 있지 않은가 생각된다. 이 병은 40 세 이상에서 발병하며 악화, 경쾌가 되풀이된다. 국소적인 탈수 (dem y e li na ti on) 가 뇌의 각 부분에 발생하여 그 부위에 따라 운동장애, 장님의 증세가 나오게 된다. 그 병인에 대하여서는 바이러스 감염, 면역 이상, 영양 이상, 유전 적 소인 둥의 학설이 있으나 아직 정설이 없다. 탈수 부위에서 수초 를 만드는 오리고덴드로사이트가 변성 소실하고 있어서 그로 인해서 수초 재생이 불가능한 것이라 생각된다. 탈수질환의 동물 모델로서는 실험성 알러지성 뇌척수염이 있는데 이는 실험동물에 수초 를 포함한 중추신경조직의 호모지네이트를 투여하여 중추신 경계에 탈수를 발생시키는 모델이다. 최근에는 뉴론-그리아 상관이란 관점에서 뉴론-오리고덴드로사 이트의 상호관계, 수초 형성, 탈수의 연구가 성행하고 있다. 필자 는 수백만 개의 오리고덴드로사이트를 사후 10 시간 이내의 사람 뇌에서 순수 분리하고 이를 6 개월 이상 조직 배양하여 그 구조와 기능에 대한 연구를 계속하고 있다. 이 연구과제에서 인간 뉴론과 인간 오리고덴드로사이트의 공동 배양계에서 수초 발생이 가능한 가 연구중이다(그림 1-22).
 그림 1-22 성인(사람) 뇌에서 분리 배양한 오리고덴드로사이트
그림 1-22 성인(사람) 뇌에서 분리 배양한 오리고덴드로사이트
제 2 장 뇌신경 발생과 연구 방법론 l 서론 뇌신경계는 정보전달, 처리, 제어에 있어서 중심적 역할을 하는 뉴론과 그것을 형태적, 기능적으로 지지하는 그리아로서 구성되는 데 뇌의 발생, 발육 시기에는 뉴론-뉴론간, 뉴론-그리아간에 상호 작용이 진행되고 있다. 뉴론-뉴론 사이에서 시납스가 언제, 어떻 게 형성되기 시작하는가, 정보전달 이외에도 시납스 혹은 다른 구조를 통해서 서로 어떠한 영향을 주고 있는가 하는 문제는 뇌신경계를 이해함에 있어서 중요한 과제의 하나이다. 따라서 신경계의 발생, 발육을 생각할 때 발육기의 신경전달물 질의 동태, 죽 생합성효소, 흡입, 분해효소 그리고 수용체 작용 둥이 그 중심문제가 되겠고 그 밖에도 신경성장인자, 영양물질의 구조, 작용이 중요하다. 어떠 한 종류의 시납스인가, 죽 시납스가 사용하는 신경전달물질이 무엇인가, 처음부터 결정이 되어 있는
가, 혹은 시납스가 놓인 환경에 의해서 결정이 되는가는 홍미로운 문제가 아닐 수 없다. 여기에서는 위에 말한 문제를 간단하게 기술하고 그에 이어서 뇌신경계를 연구함에 있어서 어떠한 방법론이 현재 널리 사용되 고 있는지 살펴보고자 한다. 2 신경계의 발생 언제 어떻게 중추신경계 조직이 발생하고 그 기능이 발현하는 가에 대해서는 제 1 장의 신경계의 기본구조에서 상술한 바 있는데 여기에서는 개체 발생 초기에 일어나는 여러가지 구조기능의 전모를 시간적으로 분석하고자 한다. 병아리-닭의 뇌신경계는 개체 발생의 연구에 잘 쓰여지는 조직 이다. 그림 2-1 은 병아리에 있어서 언제 어떻게 중추신경계 조직 이 형성되고 그 기능이 발현되는가를 모식적으로 보인 것이다. 부란 직후부터 약 3 일간 신경상피세포 (neuroe pit he li al cell) 는 분열증식하고 증식이 정지되면, 외방으로 이동을 하여 종착지점에 도착하면 분화를 시작하여 뉴론이 된다. 이 시기에 축색이나 수상 돌기가 방사상으로 퍼지게 된다. 부란 5 일경부터 초기 뉴론에서는 축색말단으로 향하여 물질이동이 일어나는데 이룰 축색이동이라 한다. 이 시기에 축색이동은 속도가 늦기 때문에 지연성 축색류 (slow axop la sm ic fl ow) 라 한다. 하나의 뉴론의 축색 이 발육하여 다른 뉴론에 접촉하면 그곳에 시납스가 형성된다. 이를 시납스 발생 (s ynapt o g enes i s) 이라 하고 부란 6 일경부터 부화 직후까지 계속된다. 이때까지는 염기적 대사가 주로 일어나는데 그 뒤로 신경계의 산소 소비가 크게 증가하여 산화적 당대사가 활발하여 진다. 부란 10 일경부터 속도가 빠른 축색류, 죽 속행성 축색류 (fas t axop lasmic fl ow) 에 의해서 신경전달물질과 그것을 포함하
고 있는 소포가 신경세포체에서 시납스까지 운반된다. 신경상피세 포의 증식을 거쳐서 뉴론의 이동, 분화가 끝나면 같은 부위의 신경상피세포에서 그리아의 발생 분화가 일어난다. 이것은 부란 10 일경에 시작된댜 부란 15 일경부터 뉴론은 흥분성을 가지며 뇌에서는 뇌의 전기활동을 표현하는 뇌파가 발생한다. 이와 평행 하여 오리고덴드로사이트가 신경섬유 주위에 수초를 형성하고 또한 혈관이 분포하게 된댜 혈관의 내피세포와 아스트로사이트가 협조하여 혈액土 1 장벽이 이 시기에 형성되어 뇌는 그 기본적인 형상과 기능 을 갖게 된다. 위에서는 병아리 닭의 뇌의 예를 들어 서 그 구조와 기능에 대하여 기술하였는데 다른 종류의 동물에 있어서도 비슷한 과정으로 발달을 하게 된다. 발생, 발육기의 뇌에서는 그 시기에 따라서 핵산 (deox yri bonule i c acid DNA 와
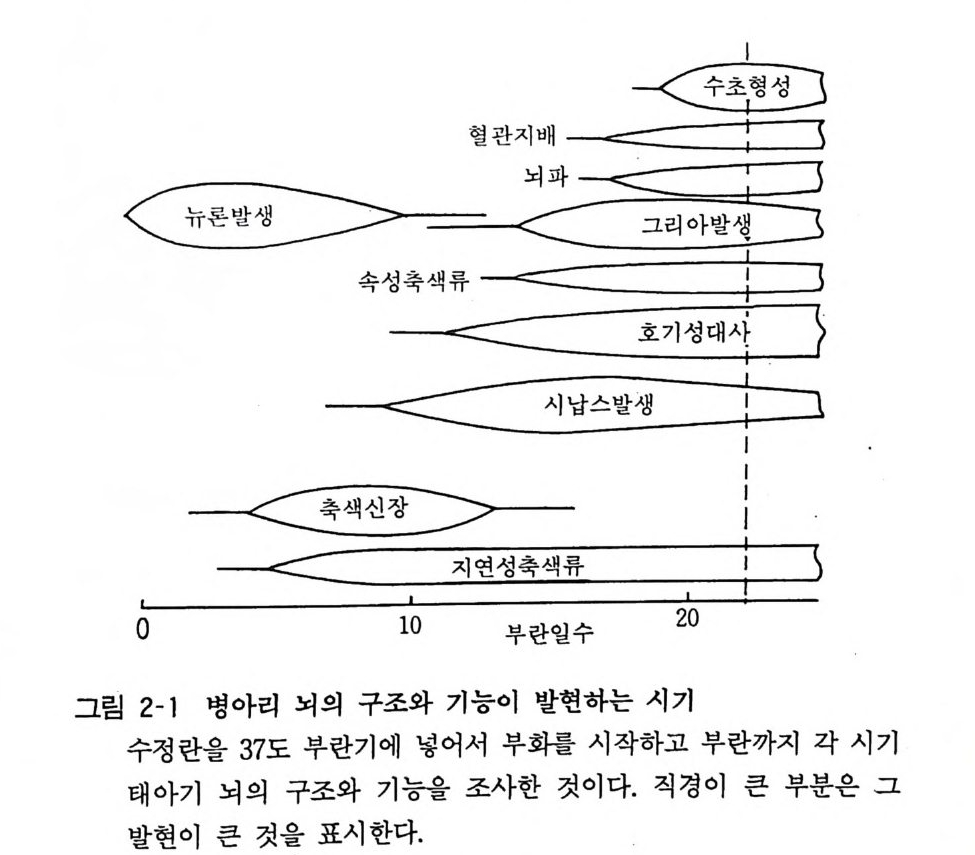 혈 관지배 __< =<二二 7 一 _IF _~
혈 관지배 __< =<二二 7 一 _IF _~
rib o nucleic acid RNA), 단백질, 지질의 양이 많이 달 라진다. 핵산은 세포의 분열과 증식, 단백질은 세포기능에 관련된 효소와 그 밖의 물질, 지질은 세포막 성분 특히 수초 형성에 관련되어서 변화한다. 그 밖에도 전해질 (Na+, K+, Cl - ) 의 뇌 농도가 뇌 발육 기에 변화한다. 쥐의 경우 신생기에 Na+ 와 c1- 농도가 높고 그 뒤 차차 감소한다. 한편 K+ 는 신생기에 낮고 성숙함에 따라 세포 내 농도가 중가한다 .
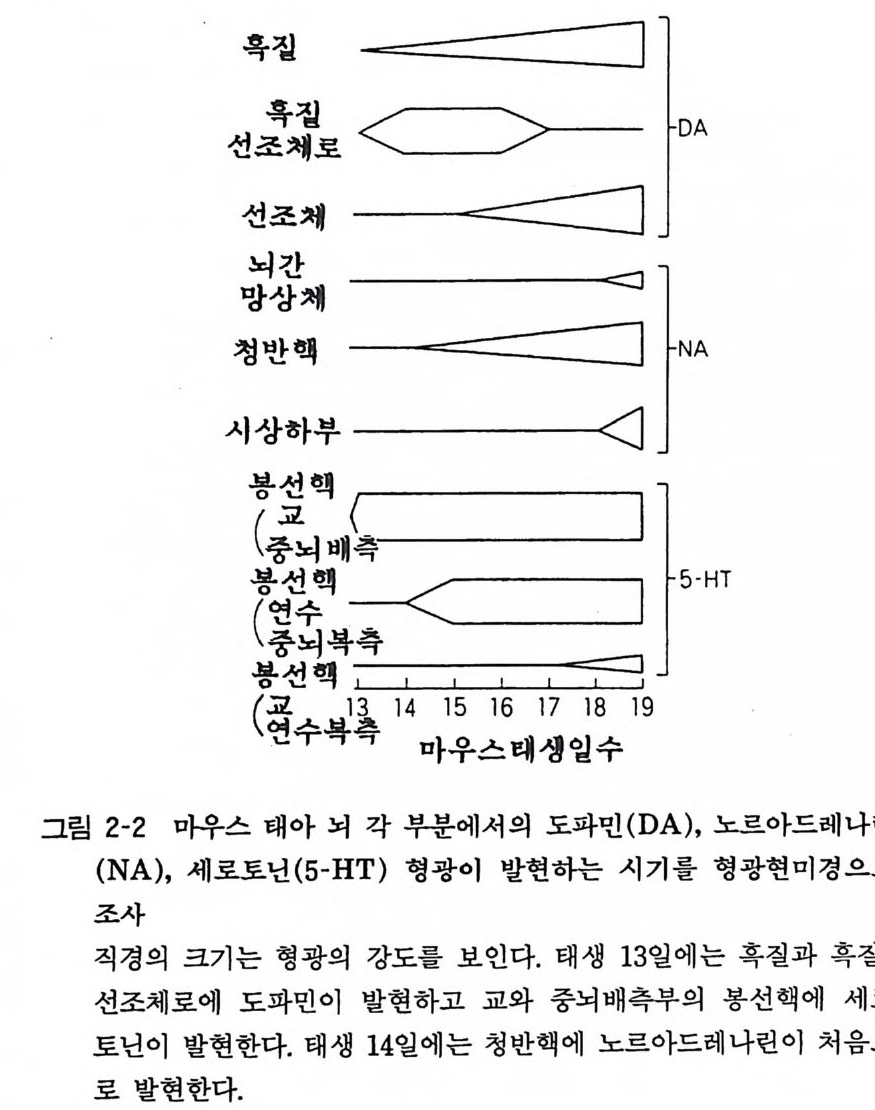 혹질 _===二二二二二〕
혹질 _===二二二二二〕
3 중추신경계에서의 모노아민(카테콜아민)의 발현 스웨덴의 학자들이 1960 년 초기에 노르아드레나린 (noradren alin e) 이나 도파민 (do pami ne) , 세로토닌 (sero t o nin)에 홀마린을 작용시키면 형광을 가지는 물질로 바꾸어짐을 발견하고 뇌 안에 서의 모노아민 신경계의 대략을 연구 기술하였다(제 3 장 3-11, 3-1 2 , 3-19). 태생기 마우스 뇌의 각 부위에서 노르아드레나린, 도파민, 세로토닌의 발현 시기를 조사하면 그림 2-2 와 같다. 태생 13 일에 도파민이 혹질-선조체에 출현하고 세로토닌은 봉선핵에 출현한다. 한편 노르아드레나린은 태생 14 일 청반핵에 출현한다. 그 뒤로 뇌 각부에 위에 말한 모노아민이 점차로 증가한다. 생화학적으로 이들 모노아민의 뇌내 함량을 측정하면 마우스 13-15 일에 출현하며, 노르아드레나린과 도파민의 생합성효소인 타이로신 수산화효소(ty ros i ne hy drox y lase) 도 뇌 성숙기의 5- 10% 가량이 측정되었다. 이렇게 태생기에 출현하는 모노아민은 어떠한 기능을 가지고 있는가. 모노아민은 하나의 뉴론에서 다른 뉴론으로 정보전달을 하는 신경전달물질의 기능을 하고 있는데, 그 밖에도 영양물질의 작용을 하고 있다는 가능성이 있다. 이를 영양인자(t ro ph i c fa c t or) 라 한다. 4 신경성장인자 (Nerve Growt h Facto r ) 레 비 몬탈치니 (Lev i -Mon tal c ini)가 1950 년 초기에 뱀독이나 마우스 육종의 추출액 속에 신경성장인자 (NGF) 가 있어서 병아리 교감신경절 뉴론의 조직 배양에서 뉴론축색의 성장이 수배나 촉진된다는 보고를 하였다(그림 2-3). 이 NGF 가 교감신경절뿐 아니라 지각신경절에서도 효과적으로 뉴론축색 성장을 촉진시킴
 그림 2-3 인간 태아 뉴론에 대한 신경성장인자 (NGF) 의 효과
그림 2-3 인간 태아 뉴론에 대한 신경성장인자 (NGF) 의 효과
을 알게 되었다. 그 뒤로 뱀독뿐 아니고 숫마우스 악하선에서 대량으로 NGF 가 추출 정제되었으며 그 화학성분이 분명하게 되었다. 이 NGF 는 a, /3, r 의 세 유니트가 결합한 형태이며, 성장 촉진 작용은 /3에 있음을 알게 되었다. 이 f3부분은 두 개의 체인 으로 되어 있고 하나의 체인은 118 개 아미노산이 펩타이드 결합 을 하고 있으며 세 개의 S-S 결합을 가진 분자량 13,000 의 펩타 이드임이 밝혀졌다(그림 2-4). 과거에 이 NGF 는 뉴론의 분열을 촉진하는 것이라 생각되었는데 현재 알려진 바에 의하면 그보다 뉴론의 퇴화 사멸을 억제함으로써 뉴론의 숫적 감소를 방지하고 있다고 한다. NGF 의 수용체는 후근 지각신경절, 뇌, 교감신경 지배장기(예컨대 심장)에 존재한다. 저자의 연구실에서는 NGF 수용체에 특이한 단크론 항체를 써서 NGF 수용체의 존재를 형광
 20
20
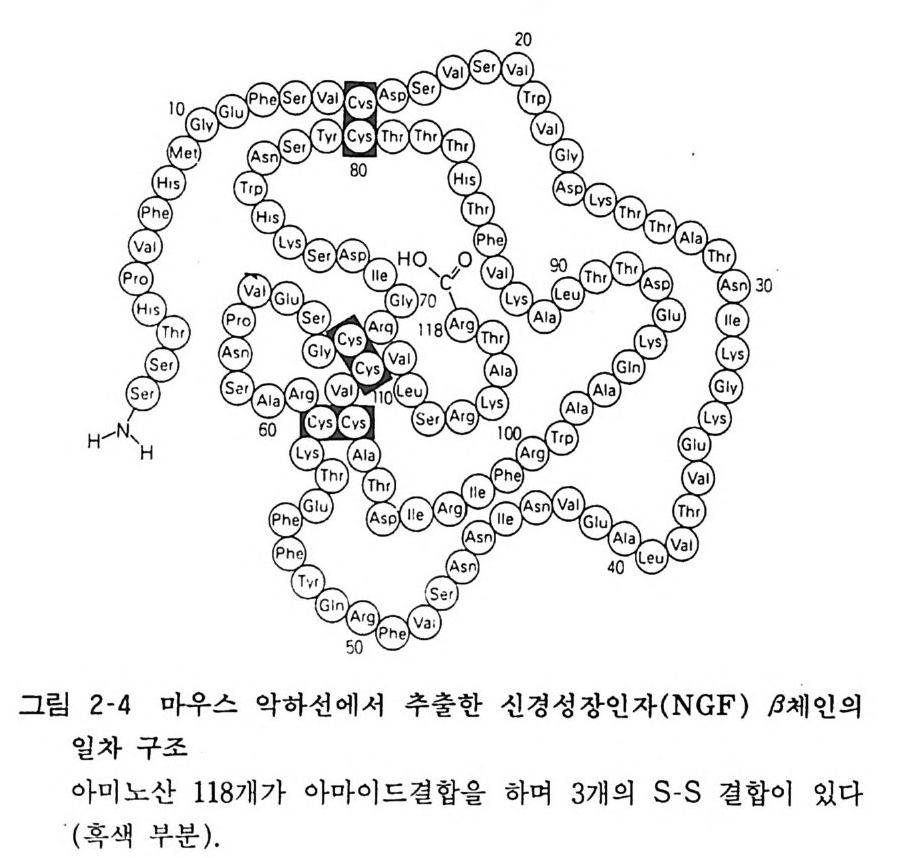
현미 경 으로 연구하고 있는데 뉴론 이의에도 슈완세포 (Schwarm cell), 흑 색종 세포 (melanoma) 와 같이 신경융 (neural crest) 유래 의 정상 세포나 암세포에도 NGF 수용체가 다량 존재함을 알게 되었다 . 생체내에서는 심장과 같은 교감신경 지배장기에서는 NGF 를 생합성하고 이것이 신경종말에서 흡수되고 축색에서 역방향으로 수송되어 뉴론 세포에 도달하면 이곳에서 단백질합성을 촉진한다 고 생각된다. 조직배양에서 보면 NGF 가 수용체에 결합한 뒤에 축색내의 미소관(mi cro t ubule) 이나 악틴 (ac tin) 형성이 촉진되고 이들 세포골격 성분은 세포내 물질의 운반이나 세포구조 유지에 중요한 역할을 하고 있다. 마우스 악하선에서의 NGF 함량은 암컷보다 수컷에서 10 배 가량 많고 발육기보다 성숙기에 많다.
그 이유에 대해서는 분명치가 않다. NGF 수용체의 수는 병아리 뇌와 심장에서 보면 부란 7-8 일에 최고이고 그 뒤에 감소한다. 5 신경전달물질의 선택 뉴론의 신경전달물질이 선천적으로 처음부터 결정이 되어 있는 가 혹은 뉴론의 주위 환경에 따라서 선택되는가라는 문제점에 관하여 현재에는 신경전달물질이 환경에 따라서 후천적으로 결정 된다는 것이 일반적으로 믿어지고 있다. 교감신경절 뉴론의 조직 배양에서 그들이 노르아드레나린과 아세틸코린 어느 것을 선택하 는가라는 실험이 있어서 이룰 소개한다. 그림 2-5 에서 보듯 신생 쥐(랫트) 교감신경 뉴론을 분리하고
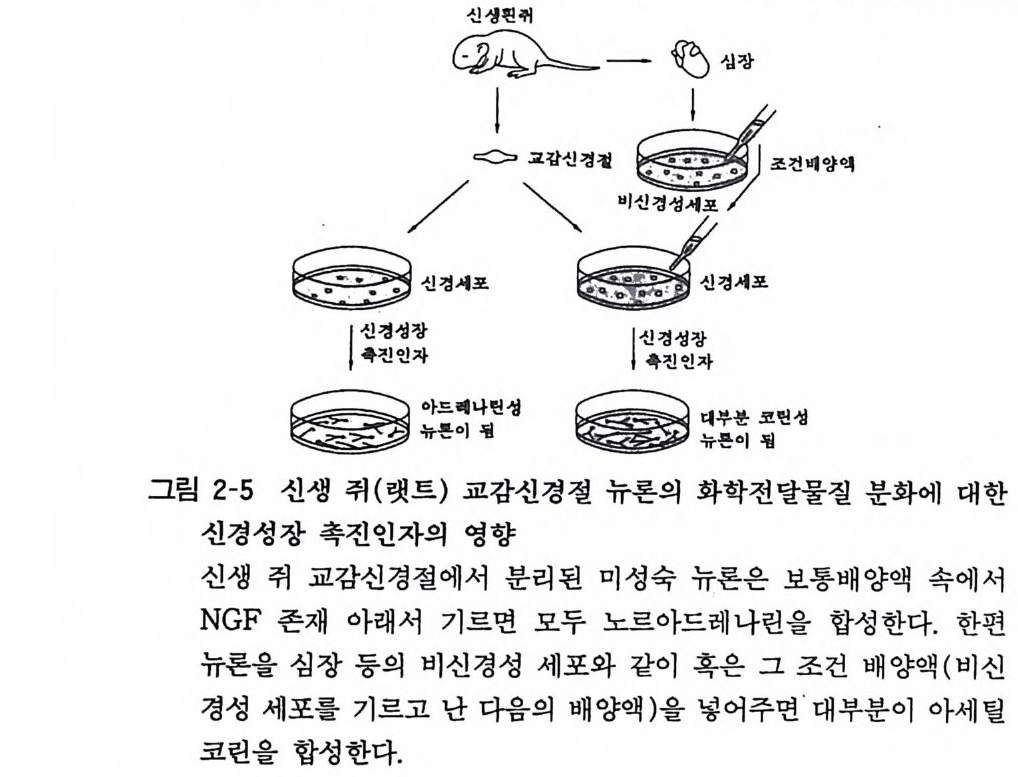 도신생핀一취 炳십강
도신생핀一취 炳십강
NGF 존재 아래 20 일간 조직 배양하면 노르아드레나린을 합성하 는 뉴론이 나온다. 한편 신생 쥐의 심장 근육 세포 를 조직 배양하 고 난 뒤 그 조직 배양액을 따로 꺼내어 그 속에서 신생 쥐 교감 신경 뉴론을 배양하면, 20 일 뒤에는 노르아드레나린 양이 감소하 고 아세틸코린 양이 대폭 증가하였다. 이 심장 배양액에서의 전체 뉴론 수는 NGF 존재 아래의 뉴론 수와 같았 다. 이러한 결과에서 보면 신생기 쥐 교감신경절 뉴론은 NGF 존재 아래 노르아드레나 린 뉴론으로 분화되는 데 이 시기에 외 부환경을 바꾸면 전달물질 이 변환되어 아세 틸코린성 뉴론이 된다. 심장 배양액 속의 미지분 자가 아세 틸코린으로 바꾸 는 작용을 가지고 있다고 생각된다. 신경전달물질은 노르 아드레나린이나 아세틸코린 등 처음부터 결정되어 있는 것 이 아니고, 주위의 뉴론 그리아 그리고 비신경성 세포 등의 영향을 받고 결정됨을 알 수가 있다. 6 시납스 형성 중추신경계 조직에는 100 조에서 200 조 (2X10) 의 시납스, 그리 고 100 억에서 200 억 (2X1010) 의 뉴론이 존재한다는 계산이 있다. 하나의 뉴론이 어떻게 다른 상대방 뉴론을 찾아내어서 시납스를 형성하고 그들이 모두 아무런 사고 없아 정확한 신경회로를 어떻 게 만들어 내는지 아직 그 정체가 미지로 남아 있다. 현재 우리가 가지고 있는 가설로서 다음과 같은 것이 있다. 첫째가 화학적 유인설로서 표적세포에서 각기 특이한 화학적 유인물질이 분비되어서 그에 끌려서 뉴론이 상대방을 찾아낸다는 것이고, 둘째가 화학적 촉진설로서 뉴론섬유의 첨단이 되는 성장 원추가 그 접촉하는 표면을 식별하면서 이동하다가 정확한 상대 방울 찾아낸다는 것이다. 이에 관련하여 표적뉴론의 구조물질이 시납스전부 뉴론의 발육을 촉진시킨다는 발표가 있다. 죽 태생마
우스 혹질 뉴론의 조직배양에 표적 신경 부위가 되 는 선조체 뉴론 의 시납스막 성분을 가하여 주면 도파민 생합성이 크게 증가하였 다. 다른 중추신경계 막성분으로는 그 촉진 효과가 없었다고 한 댜 이것은 하나의 뉴론이 그 표적뉴론을 인지함에 있어서 특수한 물질이 관여함을 말하고 있는 것이다. 시납스의 종류에는 여러가 지가 있는데 신경종말이 뉴론의 어느 부분과 접속하고 있는지에 따라 분류가 된다(그림 2-6).
 \ HJ lrL||
\ HJ lrL||
7 뇌신경 연구법-형태학적 방법 뇌의 두 부위를 맺는 신경경로를 밝히는 것은 신경해부학의 기본이 된다. 과거에는 신경세포체를 파괴하고 이로 인해서 변성 한 축색말단을 탐색하는 방법으로 그 경로를 검색한 바 있다. 1972 년에 과산화효소 (Horsera di sh pe ro xi dase) 를 사용하여 신경 경로를 검색하는 조직화학적 방법이 확립되어 그 이후로 이 방법 을 써서 중추신경계의 배선도가 정확하게 기록될 수가 있게 되었 다 . 이 과산화효소가 뇌의 어떤 부위에 주사되면 그곳에 있는 축색말단에서 홉수되어 역행성 수송 (re t ro gr ade t rans port)에 의해 서 그 모체가 되는 뉴론세포체를 검색하게 되는 방법이다. 이와 반대로 순행성 수송 (an t ero gr ade tr ans port)에 의한 방법에서는 방사성 아미노산을 사용한다. 아미노산이 뉴론세포체에 홉입되어 단백질합성에 쓰여진다. 이 단백질이 순행성 축색수송으로 신경말 단에 가는 것이다. 시납스를 넘어서 이차적인 뉴론을 마-크할 수가 있는데 이룰 트란스시 납스성 표식 (tran ssy n ap tic labeli ng ) 이라 한다. 최근에는 형광색소, 렉틴(l ec tin) 동을 사용하는 연구 업적이 나오게 되었다. 미소전극으로 전기활동을 조사한 뒤 그 뉴론의 전체 형태를 보기 위해서 미소전극으로 형광색소나 과산화효소를 주입한 뒤 현미경으로 관찰하는 수가 있다. 최근에는 배양 뉴론을 살아 있는 채 장시간 관찰하기 위해 뉴론을 형광색소로 표식하고 고감도 TV 카메라로 관측하는 방법이 있다. 필자의 연구실에서는 쥐 해마 뉴론의 칼슘 취득의 과정을 Fura-2 라는 칼슘국흡착 색소를 써서 연구중이다. 이 경우 칼슘 취득이 여러가지 신경전달물질에 의해서 촉진 혹은 억제됨을 알 수 있었다. 조직화학도 역시 유익한 연구법의 하나인데 최근에는 항원 항체반응을 사용하는 면역조직화학이 성행하고 있다. 고유한 조직 화학에서는 카테콜아민 조직형광법이 쓰인다. 이것은 뉴론 중의
카테콜아민이나 세 로토 닌이 호름알 데 하 이 드 증 기 에 의 해서 형광 물질이 되어 이 것을 형광현미 경 으 로 관찰하는 것 이다 (그림 2- 7). 한편 아세 틸 코린에 스 타라제 (ace ty l c ho li n es t era s e) 를 조직화학 적 으로 검 출 하는 방법이 자주 쓰 이 는 데 이 효소는 코린성 뉴론과 시납스 외에도 적혈 구 막에도 존 재하여 그 신 빙성에 약간 문 제가 있다(그림 2-8). 1970 년대 이 후로 섭 스탄스 피 (su bsta n ce P) 나 소마토스타틴 (soma t os tatin) 등 의 신 경펩 타이 드 의 중 요 성 이 인 식 되어 이에 대한 항체가 제 작 되었다. 이 를 써서 면역조직화학의 수법으로 이들 펩타이드의 뇌내 분 포 가 확실 하 게 되 었다 . 그 밖 에 도 GABA 합성효소, 아세틸코린 합성 효 소, 카 테 콜 아 민 합성효소 에 대한 항체가 정제되어서 GABA 성, 코린 성, 카 테 콜 아 민성 뉴론 의 뇌내 분포가 상세히 알려지게 되었다. 더 나 가 서 는 아세 틸코
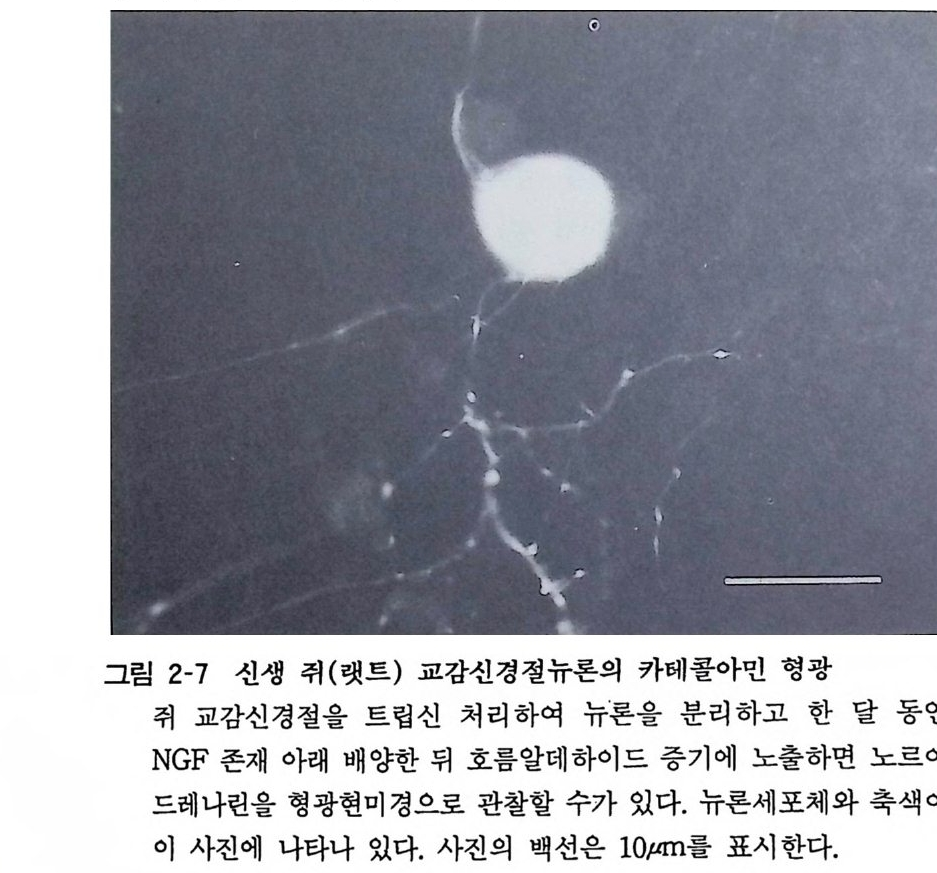 그림 2-7 신생 쥐(랫트) 교감신경전뉴론의 카테콜아민 형광
그림 2-7 신생 쥐(랫트) 교감신경전뉴론의 카테콜아민 형광
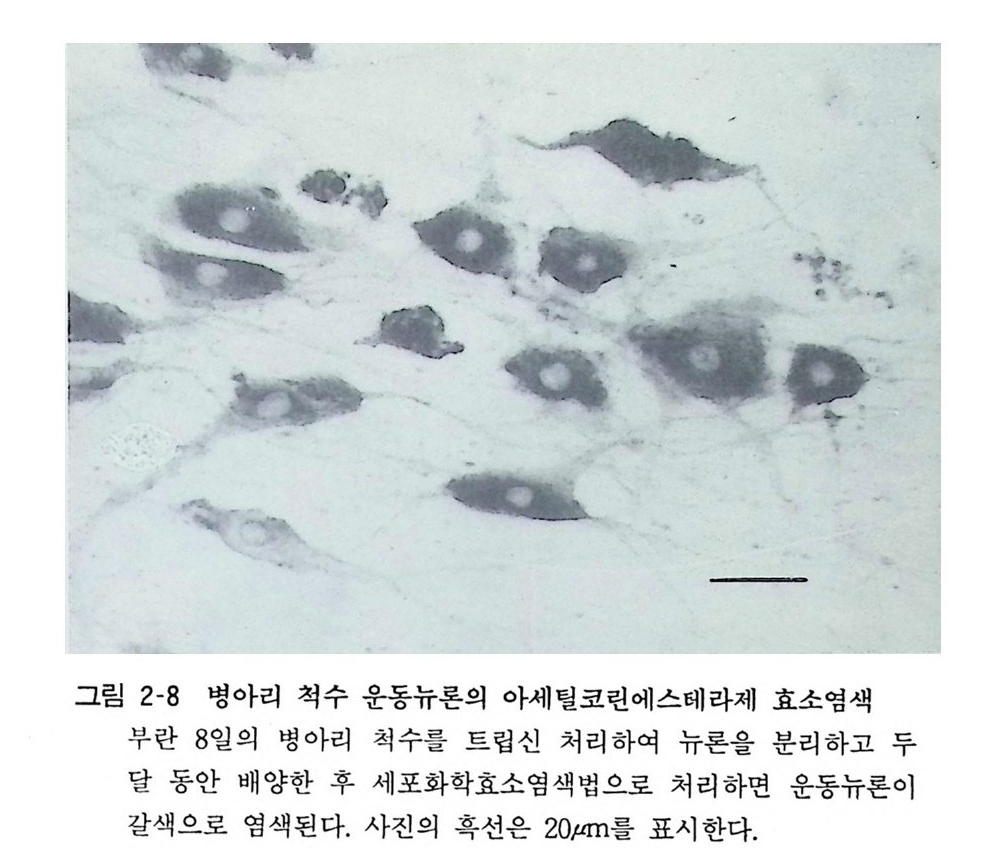 그림 2-8 병아리 척수 운동뉴론의 아세틸코린에스테라제 효소염색
그림 2-8 병아리 척수 운동뉴론의 아세틸코린에스테라제 효소염색
린 , 도 파 민, 노르아드레나란 GABA, 그라이신과 같은 신경 전달 물질 의 특 수한 항체가 정제되어 뇌신경 연구에 커다란 진보가 있게 되었다. 1975 년에 개발된 단크론항체법 (monoclonal an ti bod y)에 의해서 정제된 항체는 앞으로 면역조직화학수법을 이용한 뇌신경 연구의 발전에 커다란 기여를 할 것이다. 위에 말한 신경전달물질에 관한 연구 이외에도 뉴론이나 그리아에 특유한 세포골격 단백질, 죽 뉴론에 특이한 뉴로필라멘트 (neuro fil amen t), 아스트로사이트에 특이 한 그라이오필라멘트(gli o fil amen t)에 대한 항체를 사용하여 뉴론과 아스트로사이트를 구별할 수가 있다(그림 2-9 ) . 오리고덴 드로사이트를 위해서는 수초 염기성 단백질 (m y e lin basi c pro t- ein )이 나 가락토세 레 브로사이 드 (ga lacto c:er ebrosid e ) 에 대 한 항체
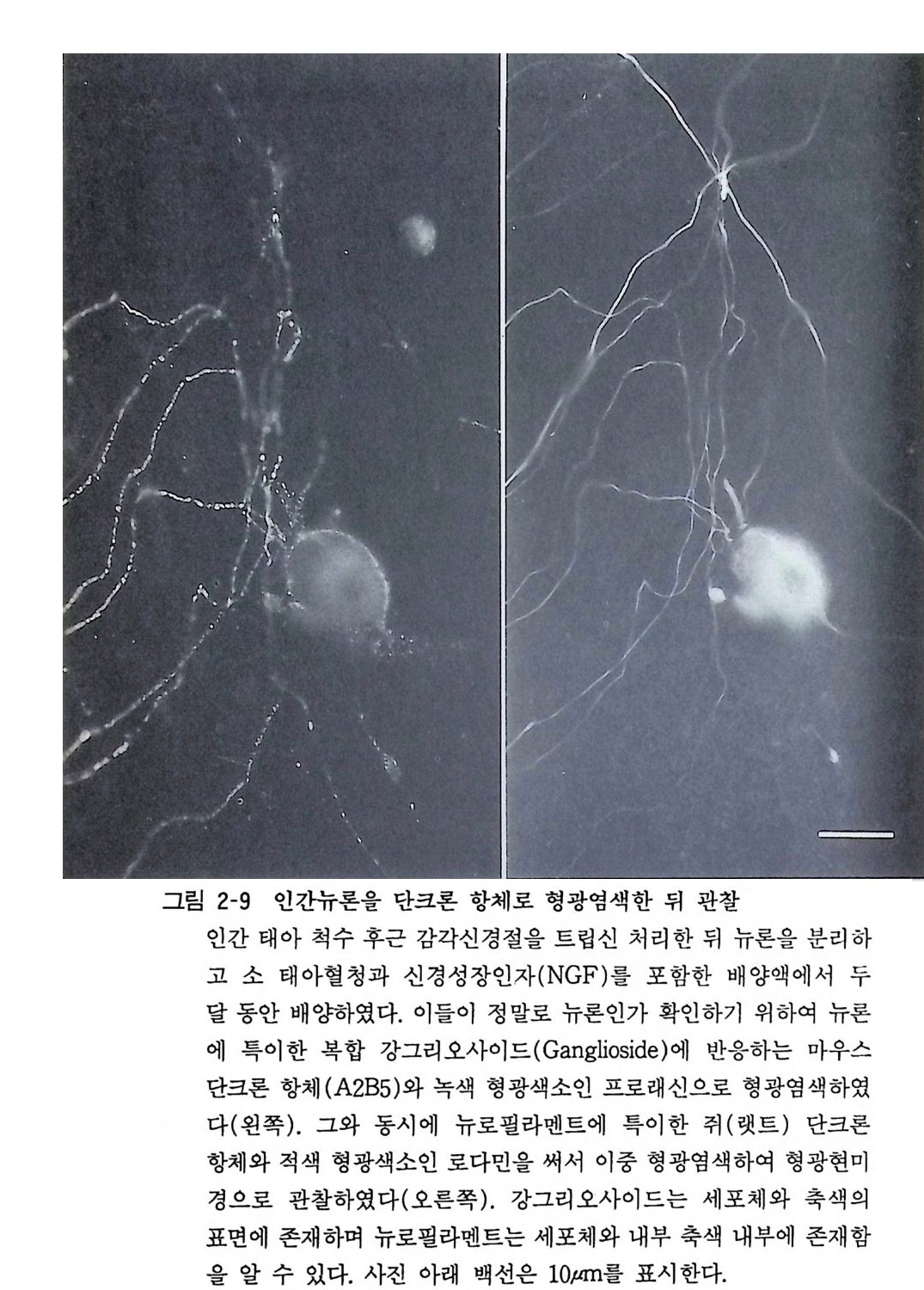 그림 2-9 인간뉴론을 단크론 항체로 형광영색한 뒤 관찰
그림 2-9 인간뉴론을 단크론 항체로 형광영색한 뒤 관찰
나 단크론 항체 를 사용하여 이들의 세포 를 표식 구별할 수 있 다. 전자현미경과 면역조직화학의 수법을 병용하여서 미세구조상 으로 어느 특이물질의 분포 를 살피는 등 금후 면역조직화학의 장래가 크게 주목된다. 8 전기 생리학 1950 년대에 시작된 생리학적 방법으로서 세포내에 전극을 삽입 하고 막전위 를 기록하고 분석함으로써 신경과 근육의 흥분전도, 시납스진달의 매카니즘을 밝혀낸 미소전극법의 공로는 참으로 크댜 한 개의 뇌신경 뉴론이 내는 임펄스를 세포 외에서 기록하 여 얻어진 정보처리 를 분석하는 연구방법은 마운트캣슬 (Moun t cas t le) 의 체성감각 분야, 휴벨 (Hubel) 과 위젤 (W i esel) 의 시각 분야에 있어서의 연구 등을 비롯해서 뇌 연구 각 분야에서 사용 되고 있다. 한편으로는 분자적 분석의 방향으로 내어 (Neher) 등에 의해서 개발된 팻치크램프법 (pa tc h clam p)이 있다. 이것은 미세한 유리 파이팻을 세포 표면에 접속시켜서 유리관 안에 국한된 세포막의 일 부 분인 팻치 안에 있는 이온채널의 활동을 기록하는 연구법이 다. 아세틸코린을 투여하면 아세틸코린 분자와 그 수용체가 결합 하고 이 수용체 하나의 활성화에 의한 이온전류를 기록할 수가 있다. 이 밖에도 전세포 크램프법 (whole cell clam p)도 이와 비슷 한 분자적 수법을 사용한다. 9 신경화학 이 분야에서는 대사 일반, 신경전달물질, 수용체, 신경펩타이
드, 신경계 특이 단백질, 특이 당지질 그리고 유전자 분리, 유전자 교환 등의 활발한 연구과제가 포합된다. 간이나 근육조직을 균질화하여 대상으로 쓰듯 뇌조직 전체를 사용하는 연구방법은 불가능하므로 뇌의 일정 부위 혹은 같은 종류의 뉴론만 분리하는 등의 기술이 필요하다. 뇌의 동결 스라이스를 만들어서 그 국소된 일정 부위 를 펀치 아우트하는 방법이 있다. 필자의 연구실에서는 단백분해효소로 뇌를 분해하여 단일세포로 만든 다음에 이를 밀도차 원심분리법 울 써서 밀도가 큰 세포(뉴론)와 밀도가 작은 세포(오리고덴드로 사이트)로 분리하여 이들의 생화학적 기능을 검색하고 있다. 미국 과 캐나다에서는 특정 신경질환으로 사망한 환자의 뇌조직을 동결보관 혹은 고정보관하여 조직을 필요로 하는 연구자에게 배부하여 특정 뇌 연구를 촉진시킨다는 취지로 운영되는 뇌 은행 (Brai n bank) 이 전국에 세 곳 있다. 최근에는 측정기술이 개량, 향상되어 있어서 극소량의 아미노 산, 단백질 지질 그 밖의 물질의 측정이 가능하게 되었다. 이에는 페이퍼 크로마토그래피, 젤 크로마토그래피, 가스 크로마토그래 피, 매스스팩트로메트리의 사용에 의해서 정성 정량이 가능하게 되었다• 사이크로트론으로 생성된 방사성 불소를 사용하여 파킨슨 병의 진단을 할 수가 있는 팻트 혹은 포지트론 에미손 토모그래 피(po s itr on em iss io n t omo gra ph y)의 사용은 질환 진단뿐이 아니 고 뇌신경 연구에도 위력을 발휘할 것이다. 또 핵자기공명 (nucleo magn et i c resonance) 법과 더불어 뇌신경 연구 분야의 많은 진보 가 기대된다. 10 신경 조직배양 1907 년 미국 예일 대학의 해리슨 (Ha rri son) 이 올챙이 척추를
개구리 임파액 속에 보존하면 신경섬유가 성장한다는 논문을 발표한 것이 신경 조직배양의 시조가 되었다. 그 뒤 이차대전이 끝나가까지 별로 진전이 없다가 1950 년대에 미국의 머레이 (Mu rra y)와 포메라트 (Pomera t)가 뉴론과 그리아의 조직배양 센터 를 만들어 신경발달과 신경의 영양 요구 그리고 신경독성학 의 분야에서 많은 연구가 이루어졌다. 필자는 1965 년부터 컬럼비아 대학 머레이 교수 밑에서 삼년간 신경 조직배양을 연구하였는데 그 중심과제는 시험관 속에서 자라나는 쥐나 병아리의 초기 신경세포가 언제 어떠한 기전으로 시납스나 수초를 구성하게 되는가 하는 것이었다. 1970 년대까지 뉴론의 신경생리학적인 연구와 신경독이나 뉴론 에 영향 을 주는 약제 를 투여한 뒤 배양조직을 전자현미경으로 검색하는 신경병리학적인 연구가 주종을 이루었다. 그러나 197 0 년 후반으로 들 어서면서부터는 뉴론에 대한 성장인자 검색, 뉴 론 -그 리아 상관관계, 그리아에 대한 증식물질 분석 등 세포생물학 적인 연구가 성행하게 되었다. 1984 년 이후로는 유전공학의 발전 으로 배양 신경조직을 사용하여 유전자 유입(g ene tra nsfe c ti on ) 수법으로 분자생물학적인 방법이 또한 사용되게 되었다. 필자의 연구실에서는 현재 사망한 뒤 20 시간 이내의 인간 뇌(성인과 태아)에서 뉴론과 그리아(아스트로사이트, 오리고덴드로사이트, 슈완세포) 를 순수 분리한 뒤 이들을 6 개월에서 1 년간 조직배양하 고 뉴론-그리아 상관관계 그리고 인간 레트로바이러스 감염 등의 연구를 계속하고 있다. 특기할 것은 필자의 연구실이 인간 뉴론과 오리고덴드로사이트의 조직배양을 세계에서 처음으로 성공하였다 는 사실이다(그림 2-9).
제 3 장 신경전달물질과 신경계의 생화학적 기능 l 서론 말초신경계에서 아세틸코린 (ace tylcho lin e) 과 노르아드레나린 (noradrena lin e) 이 신경전달물질이라는 것이 밝혀지면서 중추신경 계에서는 이들에 더하여 새로이 도파민 (do p a mi ne), 세로토닌 (seroto n in), 감마아미노부티릴산 (GABA) , 섭스탄스피 (subs ta nce-P) 동이 신경전달물질로서 또는 그 후보물질로 동장하게 되었다. 이들 물질의 신경계에서의 정보전달 역할에 관한 세포, 세포하 또는 분자 수준에서의 중요한 사실들이 최근 20 여 년 동안에 속속 밝혀져 가고 있다• 또한 이들 성과는 임상적으로 신경계의 질환, 예를 들면 파킨손씨병, 우울증, 분열병 둥의 병인 울 밝히는 데 크게 공헌하고 있으며 약물요법에 있어서도 적지않 게 공헌하고 있는 것이다. 이제 신경계의 기능에 관하여 신경전달 물질의 신경접합부에서의 동태라는 관점에서 설명하고자 한다.
특히 신경전달물질의 합성, 저장, 유리, 수용체와의 결합 그리고 불활성화(흡입 또는 대사)에 관한 기본적인 지견과 이들 각 단계 의 조절에 관하여 알아보고자 한다. 나아가 아세틸코린, 카테콜아 민 (ca t echol amin e), 세로토닌, 감마아미노부티릴산, 섭스탄스피 동 각 전달물질의 생리적 역할과 질환의 연관성 그리고 약물작용 에 대 하여 알아보고자 한다. 2 신경전달물질 뉴론과 뉴론 또는 뉴론과 주효장기세포 (e ff ec t or cell) 사이에는 약 200A 정도의 간격이 있다. 이같은 연결 부위를 접합부 (s y n a p&킵 라고 한다. 접합부에서의 정보전달은 특정 화학물질의 중개 로 이루어진다. 이같은 물질을 화학적 전달물질 (che mi cal tra n- smitter ) 또는 신 경 전달물질 ( neurotr a nsm itter ) 이 라고 한다. 화학 물질에 의하여 정보가 전달될 것이라는 생각을 처음 주창한 사람 은 엘리오트 (E lli o t)이고 이것을 실험적으로 증명한 사람은 뢰위 (Loe wi)라 하겠다. 개구리의 적출심장을 링겔액으로 관류하면서 심장의 미주신경 울 자극하여 심장의 기능이 억제되었을 때 이 심장을 관류한 링겔액으로 별개의 적출심장을 관류함으로써 제 2 심장의 기능이 억제되는 것을 관찰하고 뢰위는 미주신경을 전기자극하였을 때 신경말단으로부터 심장의 기능을 억제하는 화학물질이 유리된 결과라고 생각하고 이 물질을 미주신경물질 (va gu s s t o ff)이라 하였다(그림 3-1). 이것은 뒤에 아세틸코린인 것으로 밝혀졌다• 미주신경처럼 신경말단으로부터 아세틸코린을 신경전달물질로 서 유리하는 신경을 코린성 신경 (cho lin er gi c nerve) 이라고 한 다. 이와 같이 기타 신경들도 전달물질에 따라 같은 방법으로 이름붙여진다.
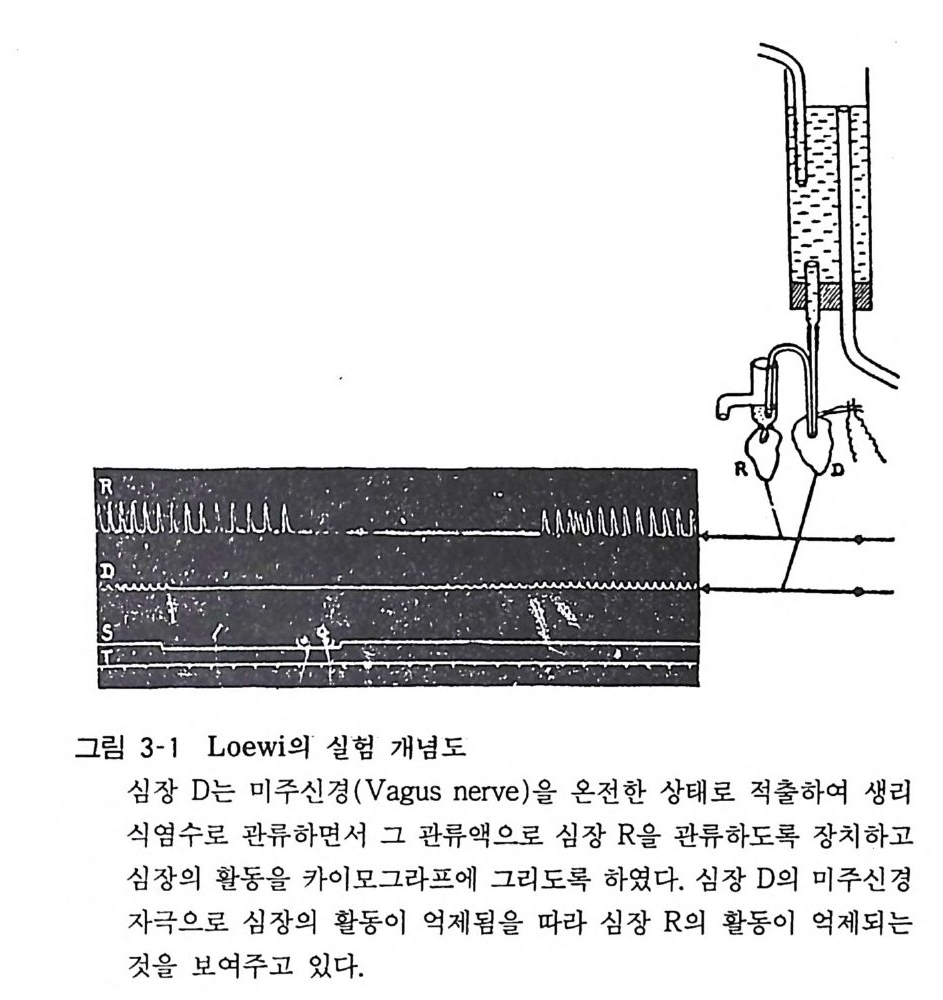 그림 3-1 Loew i의 실험 개념도
그림 3-1 Loew i의 실험 개념도
현재까지 신경전달물질이라고 생각되는 화학물질은 수십 종에 이르고 있으나 그 중 몇 가지 유력한 전달물질은 다음과 같다 (표 3-1). 3 신경전달물질의 동태 신경전달물질에 의하여 자극전달이 이루어지기 위하여는 무엇
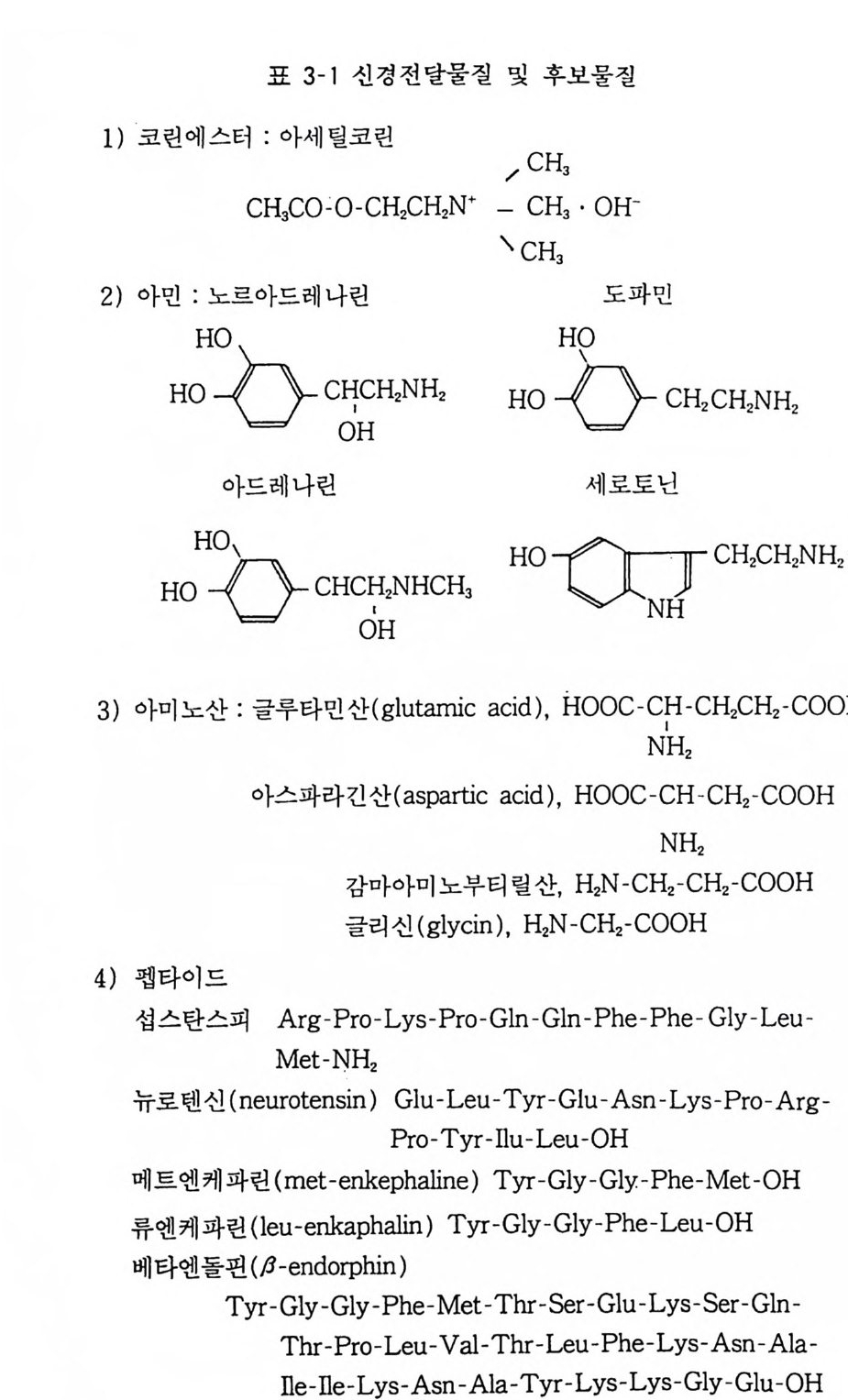 표 3 기 신경전달물질 및 후보물질
표 3 기 신경전달물질 및 후보물질
보다 우선 자극전달이 일어나는 장소에 해당 신경전달물질이 존재하여야 하고 이를 합성하는 효소가 · 해당하는 장소에 존재하 여야 한다. 합성된 전달물질은 그 역할이 주어질 때까지 일정 부위에 저장되고 이는 신경자극에 부응하여 유리되면 접합후막에 작용하여 자극을 전달하게 된다. 신경자극의 종식은 유리되어 접합후막에 작용한 전달물질의 불활성화에 의한다(그림 3-2).
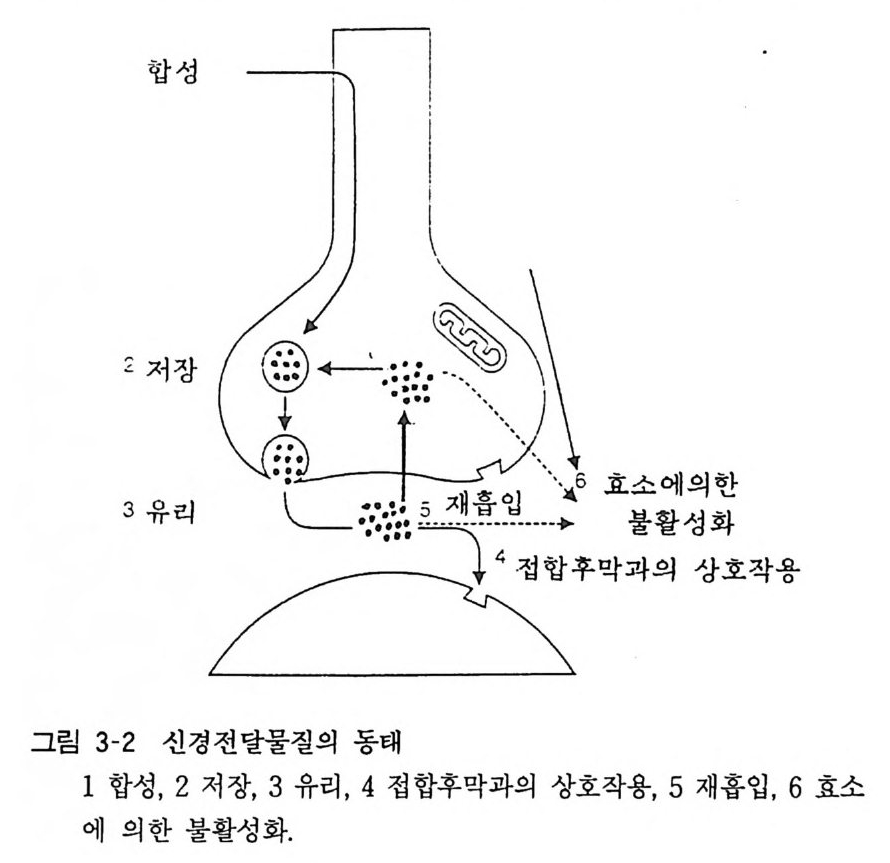 합성
합성
3. 1 합성 (Sy n th e sis ) 신경전달물질의 합성 경로는 각각의 신경계에서 자세하게 설명 될 것이나 일반적으로 아민류의 전달물질은 그 전구물질이 신경 말단에서 흡입 (u ptak e) 되어 그곳에서 합성된다고 생각된다. 펩타
이드(pepti de) 류의 경우는 우선 고 분자 의 단백 질 이 세 포 체에서 합성되고 이것이 분해되어 전달물질인 펩타이드가 되어 축색류 (axonal fl ow) 를 따라 신경말단에 운반된다고 생각되 고 있으나 자세한 것은 아직 밝혀져 있지 않다. 3. 2 저 장 (S t ora g e) 뉴론에서 합성된 전달물질은 그것이 전달 물 질로서의 역 할을 할 때까지 효소의 작용을 받지 못하도록 안정한 상태로 저장된 다. 뉴론의 접합부를 전자현미경으로 관찰하면 접 합 부 소 포 (s yn ap tic ves i cle) 라고 하는 직경이 500A 정도의 소포가 있다 . 전달 물질은 이 접합부소포내에 저장된다. 접합부소포는 속이 비어 있는 것도 있고 내부에 높은 전자밀도 를 나타내는 것도 있다. 후자를 유심소포 (cored vesic l e) 또는 과립소포(g ranula t ed vesic l e) 라고 하며 카테콜아민류의 저장부에서 잘 볼 수 있으며 이같은 소포는 신경계 외에도 신경분비계 세포, 장크롬친화성 세포 (ente r ochromaff ine cell ) 등에 서 도 볼 수 있다. 3. 3 유리 (Release) 전기적 충격이 뉴론 말단에 도달하면 접합부소포 속에 저장되 어 있던 전달물질이 접합부간격 (s yna pti c g a p)내로 유리된다. 이같은 유리현상이 일어나기 위해서는 신경세포 외부로부터 대량 의 Ca++ 이온이 세포내로 유입될 필요가 있다• 이같은 Ca++ 유입 은 전달물질의 저장부인 접합부소포가 뉴론의 접합전막(p res y napt ic membrane) 까지 이동하여 융합함으로써 개구하여 내부의 전달물질을 세포 밖으로 유출시키는 개구분비 (exoc yt os i s) 라는 현상을 유발하는 것으로 생각되고 있다. 그러나 현재까지 이같은 개구분비만으로 신경말단으로부터 전달물질이 유리될 것이라는 충분한중거는 없다.
뉴론 말단으로부터 유리 된 전달물질은 접 합후막(po s tsy na pti c membrane) 의 수용체에 결합하여 반응을 나타낼 것이나 일부는 접합전막에 존재하는 수용체에 작용하여 되먹이 기전(f eed-back mecha ni sm) 에 의하여 자신의 유리량을 조절한다. 이같은 기전에 는 집합전막에서의 Ca+t 이온 이용도의 저하가 관계하는 것같 댜 이 기전에 관여하는 접합전막 수용체를 자가수용체 (au t ore ce pt or) 라고 한다. 자가수용체에 의한 전달물질 유리조절 기전 외에 내인성 물질 (endo g enous subs tan ce) 의 작용으로 전달물질의 유리가 조절되는 수용체가 접합전막에 존재한다고 알려져 있다. 이 같 은 내인성 물질로는 프로스타그란딘(p ros tag lan din), 아데노신 (adenosin e ), 안지오텐신 (an gi o t ens in) 등이 알려져 있다. 3. 4 수용체와의 결합 유리된 전달물질이 결합함으로써 작용을 나타내게 하는 접합후 막에 존재하는 수용체는 최근까지 개념적인 것에 불과하였다. 최근에 이르러 여러가지 연구방법과 기술의 발달로 수용체의 본태와 성질들이 밝혀지고 있다. 수용체와 결합하여 반응을 유발하는 물질을 효능제 (A g o nis t) 라고 한다. 전달물질을 포함하는 효능제가 수용체와 결합하여 반응을 유발하는 모든 기전이 소상하게 밝혀져 있지는 않으나 소위 접합후전위(po s t s y na pti c pot en ti al) 를 발생시키는 기전은 다음과 같은 현상일 것으로 생각한다. 효능제가 수용체와 · 결합하면 수용체와 연관된 이온통로(i on channel) 가 열리고 막의 이온투과성이 증대하여 막전위의 변화가 일어난다(그림 3-3). 한편 효능제가 수용체와 결합하면 세포내 효소활성이 상승함으로써 세포내의 제 2 정보전달체 (second messeng e r) 가 생 성 되 고 이 것 이 단백 키 나제 (pro te i n kina se) 를 활성화하고 이 효소가 막단백을 인산화함으로써 이온통로가 열린 다고한다•
최근 캘리포니아산 전기뱀장 어의 전기기관의 아세 틸코린 수용 체를 구성하는 서브타입과 그 아미 노산 조성 이 밝혀졌다 . 멀지 않아 포유동물의 기타 뉴론 수용체의 구조도 밝혀질 것으로 기대 된다.
 E,j It. A P -- _2. ErPS-P -__ - 仁3. ^_P - -
E,j It. A P -- _2. ErPS-P -__ - 仁3. ^_P - -
3. 5 불활성 화(I nac ti va ti on) 접합부 간격 으로 유리되어 수용체와의 상호작용 을 일으킨 전달 물질은 다음의 신경충 격(신 호)에 부응하여 유리되는 전달물질이 작용할 수 있도록 신속하게 소실되지 않으면 안 된다 . 유리된 노르아드레나린의 약 80% 는 신경말단부로 재흡입된 다. 이 같은 재홉입 (reu ptak e) 은 전달물질의 접합부 간격 으로부터 의 소실 기전 중 가장 중요한 것 이다. 재 홉 입 기전은 능동수송 기전으로서 온도와 N 하 이온 의존성인 것으로 알려져 있다. 재홉입 이외의 불활 성화 기전으로는 효소적 분해 를 들 수 있 다. 아세 틸코린은 접합후막에 존재하는 분해효소인 아세틸코린 에 스 테라제 (ace ty lcho lin es t erase) 에 의하여 신속하게 가수분해된 다 . 분해산물 인 코린은 접합전막 에서 홉 입되어 아세틸코린 합성에 다시 사용된다. 펩타이드류 전달물질의 불활성 화에 대하여는 분명치 않은 점이 많으나 최근 펩타이드 전달물질 후보의 하나인 섭스탄스피가 신경말단부에서 유리된 후 효소적으로 저분자의 펩타이드로 분해 되 고 이 분 해산 물 이 신경 말단 또는 교세포(gli al cell) 로 재홉인되 는 현상이 발견되었다. 3. 6 축색 류 (Axonal flow ) 신경전달물질을 비롯하여 신경말단에서 관찰되는 물질이나 구조물의 대부분은 세포체 또는 축색내에서 형성되어 말단부로 수송된다. 이같은 역할을 하는 운반기전으로 축색류 또는 축색내 수송 (axonal t rans port)이 있다. 축색류에는 위에서 말한 것같이 세포체에서 말단부로 향하는 하행성 수송 외에도 말단부에서 세포체로 향하는 상행성 또는 역행성 수송도 있다(그림 3-4). 축색류의 속도는 수송되는 물질에 따라 다르나 하루에 1mm 에서 lm 까지의 넓은 범위를 가지고 있다. 운반되는 것의 주종은 단백
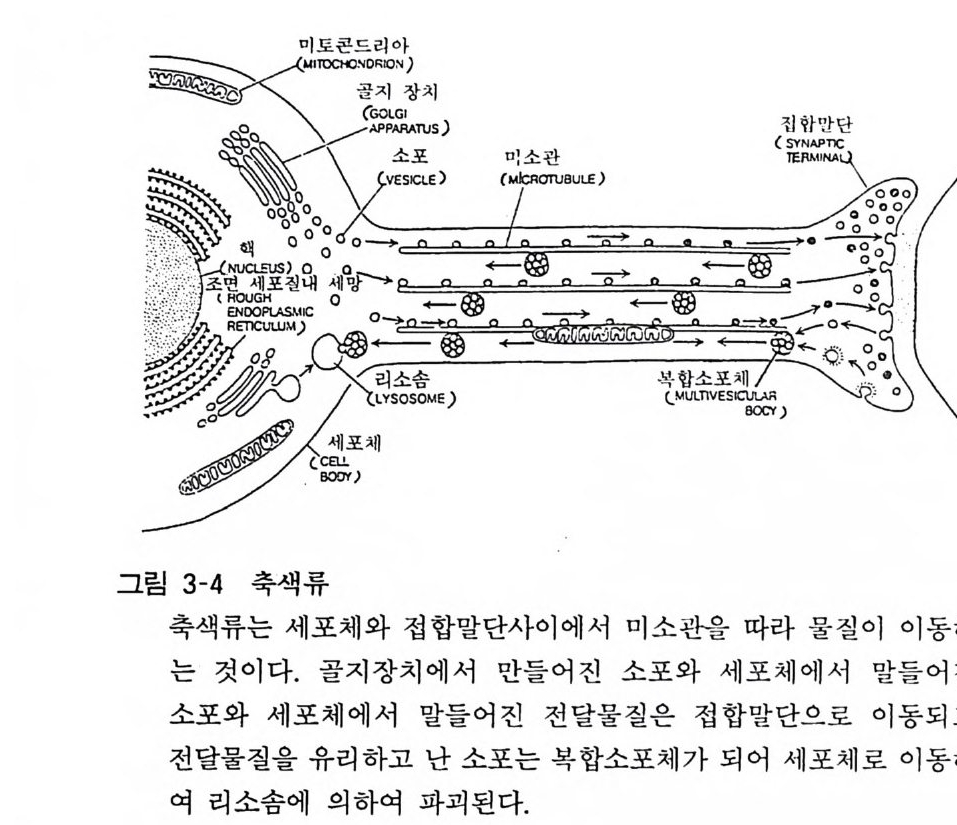 미 토콘드I리SIO아N)
미 토콘드I리SIO아N)
질, 지질, 아미노산, 세포내 과립 등이다. 단백질 중에는 전달물질 의 합성, 분해에 관여하는 효소, 미소관(mi cro t ubule) 구성단백이 포함된다. 또 과립으로는 미토콘드리아, 신경분비과립 (neurosec- reto r y gran ule), 접 합부소포 등이 있다. 축색류는 콜키 친 (colch- icin e ), 빈브라스틴(vin blas tin e) 동에 의해 억제된다• 이것은 축색 류에 미소관이 관여한다는 것을 나타내는 것이다. 축색류가 신경 ' 활성에 직접적으로 관계되지는 않으며 신경에 전기자극을 주더라 도 유속은 크게 영향을 받지 않는다. 4 아세 틸코린 (Acety le holi ne ) 아세틸코린은 금세기초에 신경생리적 활성이 알려진 것으로
미주신경 말단에서 유리됨이 처음으로 발견된 전달물질이다. 그러 나 이것의 정량법이 확립되지 못한 관계로 그 후의 연구는 큰 발전을 보지 못하였다. 최근 아세틸코린 수용체의 여러가지 성질 이 밝혀짐에 따라 생리학적 기능을 밝힐 수 있게 되었다. 4. 1 합성과 대사 아세틸코린 (Ach) 은 아세틸코엔자임에이 (Ace ty l CoA) 로부터 코린아세틸전이효소 (ChAc) 에 의하여 합성된다. 그리고 Acety l CoA 는 미토콘드리아에서 생성된다. 합성된 Ach 은 신경말단부에서 접합부소포내에 저장되고 신경 충격에 부응하여 접합부 간격으로 유리되어 수용체에 작용한 다움 수용체 가까이에 존재하는 분해효소인 아세틸코린에스테라 제 (AchE) 에 의하여 코린과 초산으로 신속하게 가수분해된댜 이들 분해산물은 신경말단부로 재흡입되어 Ach 으로 다시 합성되 어 접합부소포에 저장됨으로써 전달물질로 재사용되는 것이다
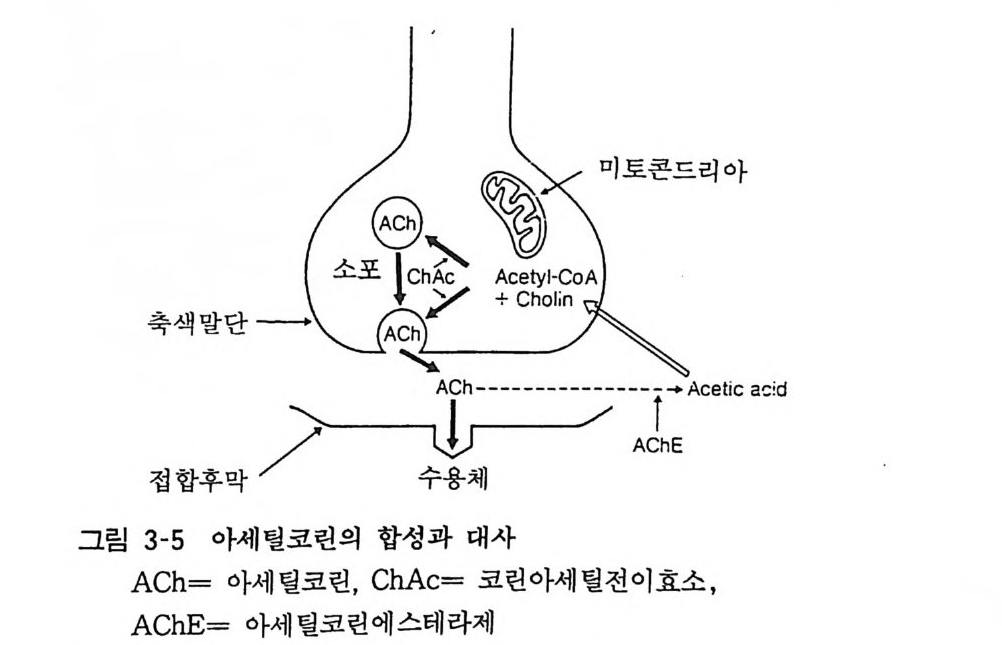 먀@〉電 零g\ / oA 미토콘드리아
먀@〉電 零g\ / oA 미토콘드리아
세포내에서의 아세틸코린 합성은 세 가지 인자에 의하여 조절 된다고 생각한다. 즉 ChAc 에 대 한 Ach 의 저 해, 기 질 량, Acety l CoA 와 코린의 이용도이다. 이중 고친화성인 코린 흡입이 가장 중요한 인자로 생각되고 있다. 4. 2 코린성 신경 (Chol i ner gi c neuron) 아세틸코린을 전달물질로 하는 신경을 51..린성 신경이라고 한 다. 중추신경계내 코린성 신경의 주행은 최끈에 이르러 서서히 밝혀지고 있어 두 가지 중추신경내 신경로가 아세틸코린에스테라 제를 함유하는 것으로 알려져 있다. 한 가지는 중뇌의 설상핵 (Nucleus cune iformi s) 에서 기시하여 시개 (Tec t um) , 술상핵 (Ge nicu late bod y)과 시상 (Thalamus) 에 섬유를 보내고, 피개로 (Tractu s t e g men t a li s) 는 혹질 (Subs t an ti a n ig ra) 과 복측피개 ( Teg m enta l is ventr a lis ) 에 서 기 시 하고 후자는 기 저 핵 ( Basal ga ng li a) 과 더 나아가 대뇌피질 (Cor t ex) 및 후구 (Ol fac t o ry bulb) 에 이른다. 기타 변연계 (L im b i c s y s t em) 의 코린성 뉴론은 해마(Hippocamp us) 에서 끝난다(그림 3-6). 그러나 극히 일부를 제외하고는 아직도 그 생리적 역할은 분명하지가 않다. 말초신경
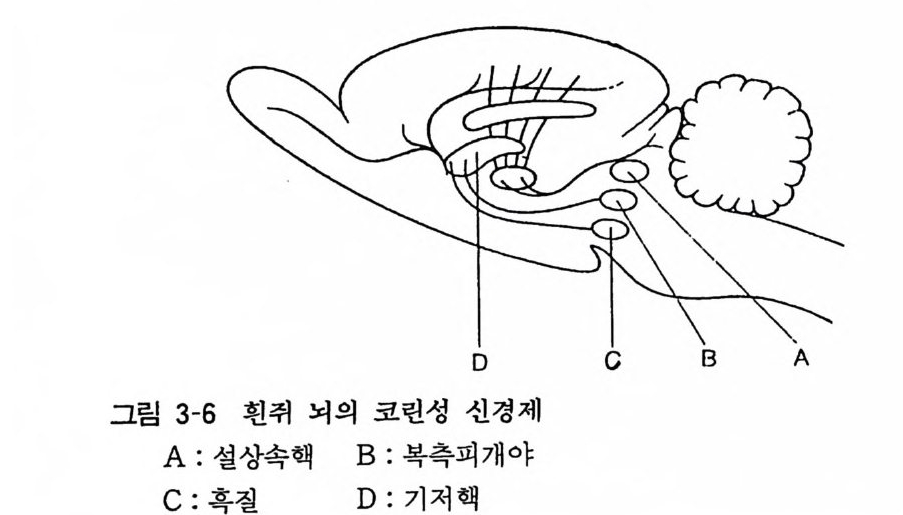 D c B
D c B
계에서는 자율신경 절전섬유, 부교감신경 절후섬유 그리고 운동신 경이 코린성 신경이다. 4. 3 코린성 수용체 (Chol i ner gi c recep tor ) 아세틸코린 수용체에는 니코틴성 수용체 (n i co tini c recep tor ) 와 무스카린성 수용체 (muscarin ic rece pt or) 의 두 가지가 있댜 니코틴성 수용체는 주로 운동신경과 골격근의 접합부인 근-신경 접 합부 ( neuromuscular jun c tion ) 와 자율신 경 철 ( auto n om ic ga n- gli o n ) 에 존재하고 중추신경에도 일부 존재한다. 뱀독의 일종인 알파붕가로톡신 (a-bun g aro t o xin)은 이 수용체와 거의 비가역적으 로 결합합으로 이 수용체의 분리 동정 실험에 이용된다. 이같은 성질을 가진 물질을 리간드(lig and) 라 하고 앞서 말한 전기뱀장어 의 전기기관에서 이같은 성질을 이용하여 니코틴성 수용체를 분리 동정하였다. 아세틸코린 수용체 중에서 가장 잘 알려진 것으 로 분자량은 약 28 만이고 당을 함유하는 단백질로서 5 개의 서브 유닐으로 이루어졌다. 무스카린성 수용체는 말초신경에서는 부교감신경의 절후섬유가 지배하는 모든 주효장기 (eff ec to r or g an) 에 분포하고 자율신경절 에도 존재한다고 한다. 그러나 자율신경질에서는 앞서 말한 바와 같이 니코틴성 수용체가 더 중요한 역할을 한다. 중추신경계의 아세틸코린 수용체는 대부분 무스카린성 수용체이다. 무스카린성 수용체의 본태는 아직 밝혀져 있지 않으나 분자량 7-11 만일 것으로 추정되고 있다. 표 3-2 에 리간드(길항제 또는 효능제)에 대한 반응의 특이성을 이용한 코린성 수용체의 분류를 정리하였다• 무스카린성 수용체가 자극되면 주효장기 세포내의 cGMP 가 중가한댜 이것에는 Ca++ 이온을 필요로 한다. 또 무스카린성 수용체의 자극은 세포막 포스파티딜 이노시톨의 대사회전이 빨라 진다. 이것에도 Ca++ 이온을 필요로 한다. 이들 반응은 모두가
전달물질로서의 아세틸코린의 정보전 달 과정과 관계되는 것으로 생각된다. 4. 4 코린성 신경의 생리적 역할 말초 코린성 신경인 운동신경이 자극되면 골 격근 수축을 일으 킨다. 한편 부교감신경 절후섬유의 무스카린성 수용체는 무스카 린 투여로 나타내는 반응과 유사한 작용을 나타낸다(표 3- 3) . 일반적 으로 말하면 생체가 안정상태에 있을 때의 반웅이라 할 수 있 다 . 이것에 비하여 뒤에 다시 언급되겠으나 아드레나린성 신경은 생체가 활동적이거나 홍분상태 때의 반응이라 하겠다.
표 3-2 아세틸코린 수용체의 분류 수용체형 존재부위 효능제 길항 제 쿠라레 (d-T c) 근-신경접합부 PTMA ( d- tu b ocurarin e ) 니코틴성 , 니코틴 헥사메토니움 신경절 (Nic o ti ne ) ( Hexameth o n iur n ) 카바콜 쿠라레 중추신경 ( Carbachol ) 부교감신경 무스카린 아트로핀 절후섬유 (Musc arine ) (At rop ine ) 무스카린성 중추신경 무스카린 아트로핀 옥소트레모린 (Oxotr em ori ne ) PTMA : Pheny lt ri me th yla mrnon iur n.
이와 같이 일반적으로 하나의 장기는 교감신경과 부교감신경의 길항적인 이중 지배를 받는다. 따라서 이들 두 가지 신경의 긴장 도의 변화에 따라 장기의 기능이 유지된다. 그림 3-7 은 아세틸코 린에 의하여 자극전달이 일어나는 부위를 나타낸 것이다. 죽 부교 감신경 절후섬유 외에도 자율신경절(교감신경, 부교감신경 공히) 과 부신수질 그리고 운동신경이 아세틸코린으로 자극된다.
표 3-3 부교감신경 주효장기 의 반응 주효장기 반응 홍채괄약근 수축(축동) 모양체근 수축 심장 심박동수 감소 수축력 감소 전도속도 증가 혈관 확장 기관지근 수축 기관지분비 촉진 위장관 운동 촉진 위장분비 촉진 담낭, 담관 수축 방광 기저부 수축 괄약근 이완 땀샘 분비 촉진 침샘 분바 촉진 부신 아드레나린 분비 촉진 간 그리코겐 합성 촉진
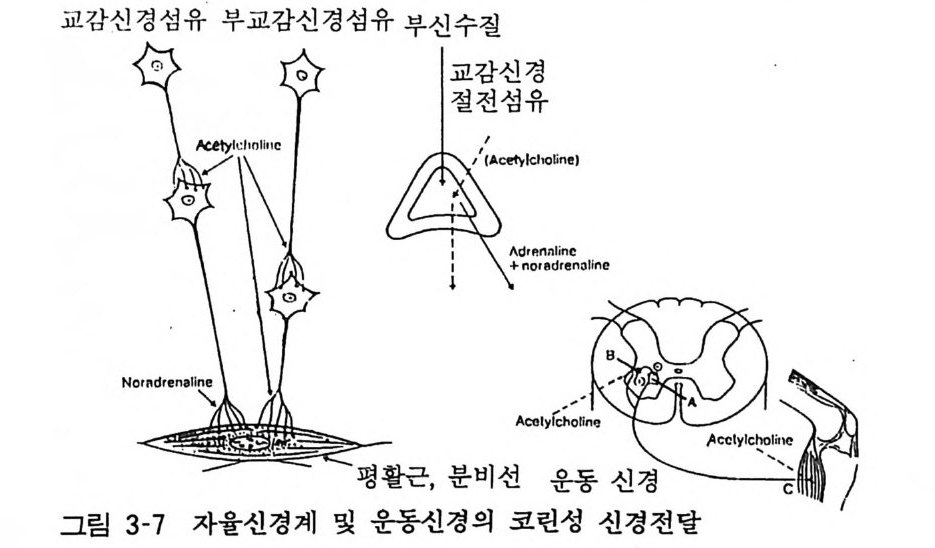 교감신경섬유 부교감신경섬유:
교감신경섬유 부교감신경섬유:
4. 5 코린성 신경과 질병 골격근의 운동시 급속하게 피로하고 운동불능 상태가 되는 증상을 중증근무력증 (M y asth e ni a gr av i s) 이라고 한다. 이 질환은 니코틴성 아세틸코린 수용체에 대한 항체가 생성되는 일종의 자가면역질환이다. 운동장애와 정신증상을 나타내는 헌팅톤 무도 병 (Hun tingt on's chorea) 이라는 질병이 있다. 이것은 중추신경계 의 일부 무스카린성 수용체가 현저하게 감소된 것에 기인하는 것으로 생각된다. 4. 6 코린성 신경과 약물 1) 운동신경 네오스티그민 (Neos tigmin e) 은 항 ChE 작용을 가지고 있으나 이것과는 별개로 근-신경접합부에 대하여 소량으로는 자극작용을 대량으로는 억제작용을 나타낸다. 헤미코리니움 (He mi cho lini urn) 온 신경말단부에서의 코린의 홉입을 억제함으로써 아세틸코린 합성을 저해하여 골격근의 이완을 일으킨다• 항생물질인 네오마이 신 (Neom y c i n), 스트렙토마이신 (S t re pt om y c i n), 가나마이신 (Kanarny c in ) 등으로 환자를 치료하는 도중 때때로 근무력증을 일으키는 것이 알려져 있다. 이것은 항생물질이 신경말단에서 접합부소포의 이동을 방해함으로써 아세틸코린 유리를 저해하여 근육이완을 일으키는 것으로 해석하고 있다. 염기성 세균인 보투 리누스균이 생성하는 보투리누스독 (Bo tulin us t o xin)은 극히 소량 으로도 운동장애 둥 중독증상을 나타낸다. 이것은 신경말단부에서 아세틸코린의 유리를 억제하는 작용을 가지고 있다. 남아메리카 인디언이 사용하는 화살독의 성분인 투보쿠라린(쿠라레 : Tub- ocura rin e) 은 니코틴성 수용체에서 아세틸코린과 상경적으로 길항하여 호홉근을 비롯한 골격근 마비를 나타낸다. 데카메토니움 (Decameth o n ium ) 이 나 석 시 닐코린 (Succin yl~ holi ne ) 은 아세 틸코
린과 같이 수용체 를 탈분극시키는 것이나 그 작용이 지속적이므 로 결과적으로 근육이완작용을 나타내게 된다(그림 3-8).
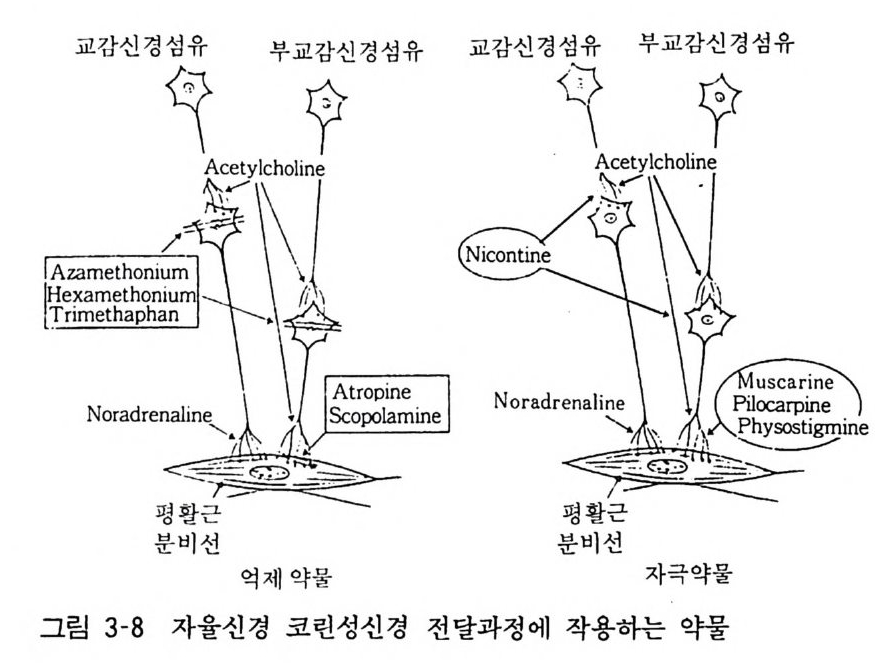 교감신경섬유 부교亨감신 경섬유 교감신경<섬유 부교 감亨신경 섬유
교감신경섬유 부교亨감신 경섬유 교감신경<섬유 부교 감亨신경 섬유
2) 자율신경 헥사메토니움 (Hexame th o ni um) 은 신경계의 니코틴성 수용체에 서 아세틸코린과 상경적으로 길항하여 신경절에서의 자극전달을 차단한다• 그 결과로 혈압하강, 소화관운동의 감소, 긴장저하, 분비억제 등의 효과가 나타난다. 한때 의약품으로 사용되었으나 작용발현이 급속하고 작용이 격렬하여 여러가지 부작용을 나타내 므로 현재는 별로 사용되지 않는다. 아트로핀 (A tr o pin e), 스코폴 아민 (Sco po l amin e) 등의 식물성분은 부교감신경의 절후섬유가 지배하는 무스카린성 수용체에 작용하여 아세틸코린과 상경적으 로 길항한다. 그 결과 아세틸코린이 나타내는 여러가지\간옹에 길항하게 된다. 이들은 의약품으로도 널리 사용되고 있다. 한편 메 타코린 (Meth a choli ne ) , 카바콜 (Carbachol) , 베타네 콜 (Be tha nechol) 동은 일종의 코린에스터로서 코린성 순용체에 대하여
효능제로 작용한다. 천연 알카로이드인 필로칼핀 (P i locar pi ne), 아레코린 (Arecho li ne), 무스카린 (Musca ri ne) 도 아세틸코린 같은 효능제이다(그림 3-8). 5 카테콜아민 (Ca t echolam i nes) 카테콜아민은 카테콜핵을 가진 생리활성 아민으로서 통상 도파 민 (Do pamin e), 아드레나린 (Adren ali ne), 노르아드레나린 (Nora dren alin e) 을 일컫는다. 카테콜아민의 연구는 19 세기말과 20 세기초에 걸쳐 시작되어 중추신경계와 말초신경계의 전달물질이 라는 것이 밝혀지게 되었다. 5. 1 합성과 대사 카테콜아민은 일련의 효소가 관여하는 반응에 의하여 타이로신 (T yr os in e) 으로부터 합성된다. 죽 전구물질인 타이로신이 신경말 단에서 흡입되면 이것은 타이로신 수산화효소 (TH) 에 의하여 DOPA 가 생성되고 이것은 다시 DOPA 탈탄산효소의 작용을 받아 도파민이 만들어진다. 뒤에도 다시 언급되겠으나 도파민은 그 자체가 신경전달물질이기도 하다. 도파민은 다시 도파민 베타 수산화효소 (DBH) 의 작용으로 노르아드레나린이 만들어진다. 말초신경계에서는 이렇게 합성된 노르아드레나린이 접합부소포내 에 저장되는 것이다. 중추신경계 또는 부신수질에서는 노르아드레나린이 페닐에타놀 아민 -N- 메틸전이효소 (PNMT) 에 의하여 아드레나린으로 변환된 다. 이같은 카테콜아민 합성과정의 속도를 제어하는 속도제한 효소는 타이로신 수산화효소로서 이것의 활성이 억제되면 카테콜 아민 함량이 감소한다.
신경충격으로 신경말단으로부터 유리된 카테콜아민은 수용체에 작용하고 앞서 말한 바와 같이 대부분이 신속하게 신경말단으로 재흡입된다. 재흡입된 카테콜아민과 일부 세포질내로 유리된 카테콜아민은 미토콘드리아의 모노아민 옥시다제 (MAO) 에 의하여 대사된다. 또 한 가지 카테콜아민 대사효소로는 카테콜 - 0 - 메칠전이효소 (COMT) 가 있다. 결국 카테콜아민은 이들 두 가지 대사효소의 작용으로 불활성화된다. 이들 대사산물은 뇌척수액, 혈액을 거쳐 소변으로 배설되므로 중추신경계 카테콜아민 뉴론의 활성을 반영 하는 임상검사로 이용된다(그림 3-9 ) . 유리된 카테콜아민의 재흡입은 신경계에서뿐 아니라 부신수질 에서도 일어나는 현상이다.
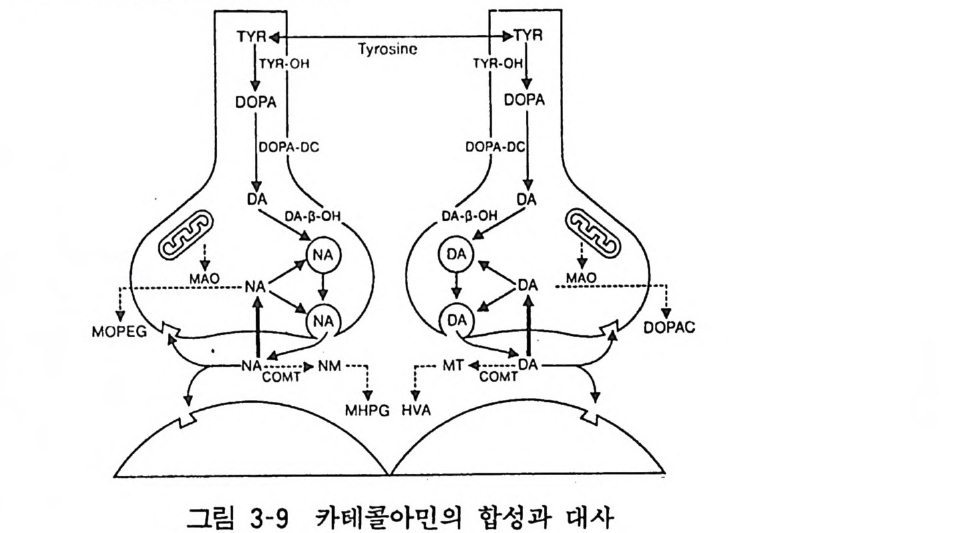 二 DTlYAO RPA: Ty ro s m c TYR
二 DTlYAO RPA: Ty ro s m c TYR
5. 2 카테콜아민성 신경 중추신경내 카테콜아민성 뉴론의 주행은 옹게르스테트 (Ung erste dt ) 둥의 연구자들에 의하여 밝혀져 있다. 노르아드레나린 뉴론의 세포체는 미측중뇌 (Caudal midb rai n), 뇌교 (Pons), 연수 (Medulla oblon g a ta)에 위치한다. 연수에서는 척수로 하행성 섬유
를 내고 한편 뇌교, 소뇌, 시상하부, 피질에 상행성 섬유를 뻗찬 다. 또 청반핵 (Locus ceruleus) 에서 복측속 (Ven t ral bundle) 또는 배측속 (Dorsal bundle) 을 거쳐 소뇌, 피질, 해마 등 뇌 여러 부위 에 축색을 보내고 뇌교에서 시작되는 뉴론은 시상하부에 미친다 (그림 3-10). 도파민 뉴론은 혹질 (Subs tanti a nigra) 을 출발하여 미상핵 (Cauda t e nucleus), 담창구 (P alli durn) 에 이르는 혹질-선조 체계 (N igr o-s tri a tal path wa y)와 복측피개핵 부근에서 출발하여 중격 (Se ptal nuclei) , 편도핵 (Am yg dala) 에 이르는 중뇌-변연계 (Meso-lim b ic sys te m ) 그리고 시상하부에 존재하고 정중융기 (Med ian em ine nce ) 를 지 배 하는 융기 -누 두 계 ( Tubero-in fun di- bular sys te m ) 둥이 있다. 중추신경계의 아드레나린 뉴론에 대하 여는 분명치 않은 점이 많다(그림 3-11). 말초신경계의 카테콜아 민 뉴론은 자율신경의 교감신경 절후섬유가 노르아드레나린 뉴론 이다.
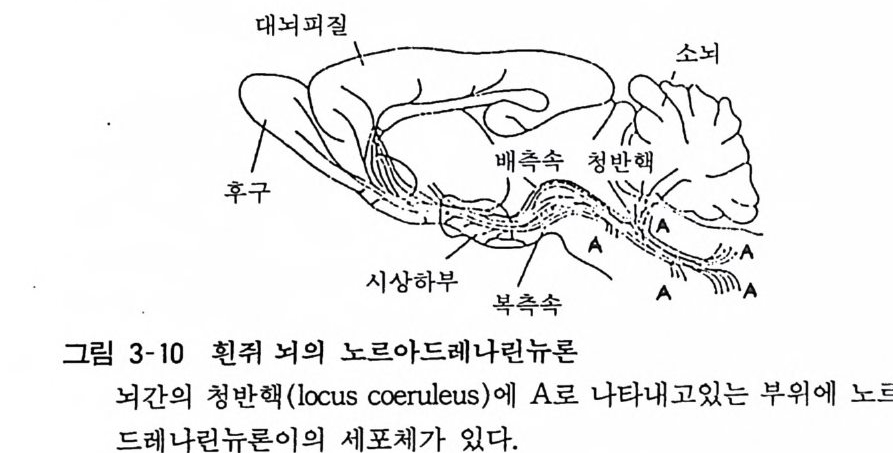 그림 3-10 흰쥐 뇌의 노르아드레나린뉴론
그림 3-10 흰쥐 뇌의 노르아드레나린뉴론
 후
후
5,. 3 카테콜아민 수용체 카테콜아민 뉴론의 수용체는 여러가지로 세분되어 있다. 알파 (a) 수용체의 경우 접합후부의 어 수용체와 접합전부에 존재하는 언 수용체로 나누어진다. a2 수용체는 자가수용체 (au t orece pt or) 로서 신경말단으로부터 의 노르아드레나린 유리를 조절하는 역할 을 한다. 그러나 최근의 결합실험에 의하면 언 수용체가 접합후부 에도 존재한다고 한다. 수용체 검정에 이용되는 리간드들은 다음 표에 정리되어 있다 (표 3-4).
표 3-4 카테콜아민 수용체의 분류와 리간드 수용체 효능제 리간드 길항제 «1 노르아드레나린 알( Noradrena line ) 용파수아( A드dr레en나a린 lin e ) 프(P라ra조zo신sin ) 아체
드레나 a2 크(C 로lo니n i딘din e ) 요( Y힘oh빈 im b in e ) 린성 타베/3I 도( D부o타bu민ta mine ) 메(M 토et프o p로 r o 롤lo l ) 용체수/32 테(T 르er부bu타ta린 lin e ) 부(B톡ut사o x 민 am ine ) 도파D1 도(D파op민 a mine ) 스( S피pi p페e론ro n e } 민성D2 아( A포p o몰m핀 o rp hine ) 할( H로al페op리e r돌ido l )한편 노르아드레나린 수용체에는 베타(fl) 수용체라 하는 것이 있고 이것은 아데닐레이트 사이크라제 (Aden y lat e c y clase) 와 함께 신경 이외의 조직에서도 여러가지 활성을 나타내는 것으로
생화학적 연구에 많은 관심의 대상이 되는 수용체이다. 최근의 연구에 의하면 f]수용체는 다시 fl 1 과 [J 2 수용체로 나누어지고 흰쥐 의 대뇌에는 fl 1 수용체가 대부분이고 소장에는 fl 2 수용체가 대 부 분 이라는 것이 리간드 결합실험을 통하여 알려졌다. 도파민 수용체도 세분되는 것이 알려졌다. D1 수용체는 아데닐레 이트 사이크라제와 함께 활성을 나타내는 도파민 수용체이고 D2 수용체는 아데닐레이트 사이크라제 활성을 억제하는 것으로 최근 알려졌다. 그 외에도 효능제 또는 길항제와의 결합 특 성에 따라 D3, D4 수용체로 분류하고 있다. 5.4 카테콜아민 신경의 생리적 역할 1) 말초신경 노르아드레나린은 교감신경 절후섬유의 전달물질이다. 그것의 생리작용은 교감신경이 자극되었을 때 각 장기가 나타내는 반응 이라 하겠다. 교감신경 홍분시에 나타나는 각 주효장기의 반응과 그것에 관여하는 수용체를 정리하면 다음과 같다. 아드레나린도 노르아드 레나린과 거의 비슷하게 작용하나 아드레나린은 노르아드레나린 에 비하여 /3수용체에 대한 효과가 더 크다. 앞서 말했듯이 코린 성 신경이 생체의 안정시 장기 기능유지에 관여하는 것에 바하여 아드레나린성 신경은 생체가 활동적이거나 흥분상태인 경우 장기 기능에 관여하는 것이다. 교감신경 홍분시에는 현저한 일과성 혈압상승을 나타낸다. 이것 은 심장기능 항진과 말초혈관 수축 등의 작용에 의하는 것이며 한편 소화관의 운동이나 긴장도는 억제된다(표 3-5). 카테콜아민은 아데니레이트 사이크라제를 활성화함으로써 세포 내 제 2 정보전달체인 cAMP 의 생성을 증가시키고 이것에 의하여 당대사 및 지방대사의 증가를 나타내게 된다.
표 3-5 교감신경 주효장기의 반응 주효장기 수용체 닌'--O} 으 홍채산대근 a( 수축(동공산대) 모양체근 (3 이완 심장 /31 심박수증가 수축력 증가 전도속도 증가 관상혈관 f12 확장 피부, 점막 혈관 a 수축 골격근 혈관 fJ2 확장 내장 혈관 a 수축 기관지근 /32 화-1 자0 장관 평활근 aI, /32 운동, 긴장도 억제 담낭, 담관 f] 이완 방광 기저부 fl 이완 방광괄약근 a’ 수축 땀샘 ar 경도의 분비 촉진 타액선 a, /3 분비 촉진(농축) 가'--0자 a, B 그라이코겐 분해 촉진
도파민은 부갑상선에 존재하는 도파민 수용체에 작용하여 cAMP 를 층가시킴으로써 부갑상선 홀몬분비를 촉진한다. 또 뇌 하수체 전엽에는 도파민 수용체가 존재하여 프로락틴 (Prolac tin)이라고 하는 유즙분바 촉진홀몬의 분비를 억제한다. 뇌하수 체 중엽의 알파메라닌 (a-mel anin e) 자극홀몬의 분비에 대하여 /3수용체 자극은 촉진적으로, D2 수용체 자극은 억제적으로 반응한 다고 한다(그림 3-12). 2) 중추신경계 중추신경계의 카테콜아민 뉴론의 생리적 역할에 관하여는 아직 분명치 않은 점이 많다. 쥬베(J ouve t) 둥은 생체의 수면이 중추
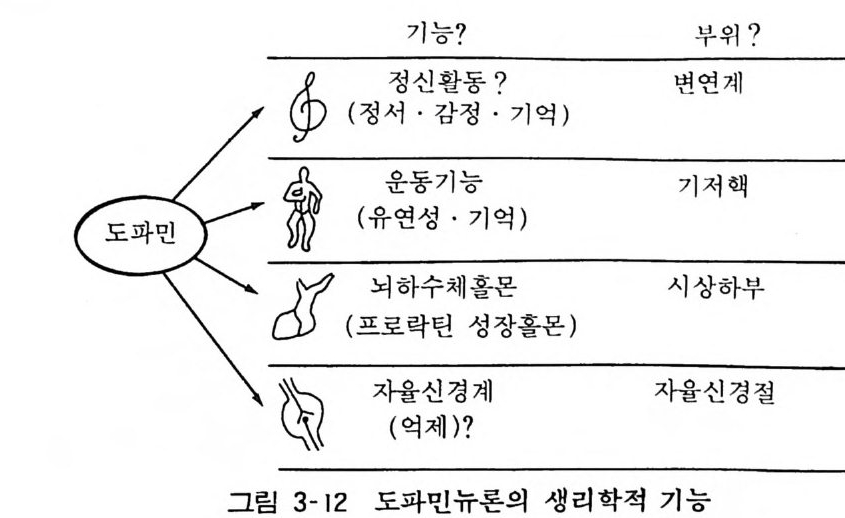 기능? 부위?
기능? 부위?
카테콜아민 뉴론에 의하여 통제된다고 주장하고 있다. 이들에 따르면 수면중에는 뇌파 소견이 각성상태와 비슷하고 안구운동이 기록되는 역설적 수면(p arado xi cal slee p)이라는 수면상태가 있는 데 카테콜아민 뉴론은 이같은 수면상태 발현과 관계된다고 한 다. 생체의 식욕을 조절하는 중추가 시상 하부에 있다. 이 부위에 서 a 수용체 자극은 식욕항진을, /3수용체 자극은 식욕억제를 나타 낸다. 동물 뇌의 내측 전뇌속(inn er fore brai n bundle) 에 전극을 꽂고 동물이 페달을 밟으면 자극이 가해지도록 장치하면 동물은 쾌락을 느끼는 것처럼 페달을 계속 밟는 것을 볼 수 있다. 이같은 뇌 부위를 보수계 (reward s y s t em) 라고 하는대 이 부위도 노르아 드레나린성 뉴론의 주행과 일치한다. 그 외에도 중추 노르아드레나린성 뉴론은 혈압조절, 체온조절, 정신활동, 운동기능의 통합에 관계된다. 중추 도파민 뉴론은 운동 기능과 관계가 깊다. 이 신경을 자극하면 동물의 운동량이 증가하 고 상동행동 (s t ereo type beha vi or) 이 관찰된다. 5. 5 카테콜아민성 신경과 질병 중년 남녀에서 흔히 발생하는 운동장애를 주 중상으로 하는
질병은 파킨손씨병 (Parki ns on's di sease) 을 들 수 있다. 이 병은 진전(t remor), 근경축, 무동 등 증상으로 극도의 운동곤란에 빠지 게 되는 질병이다. 환자의 뇌를 조사해 보면 추체외로계 (ex t ra pyra m ida l s y s t em) 라고 하는 부위 특히 혹질, 미상핵의 도파민 뉴론이 변성을 보인다. 헌팅톤 무도병은 파킨손씨병과는 반대로 운동기능이 과도하게 되는 질병으로 안면, 경부, 사지 동의 불수의적 운동이 계속적으 로 나타나는 것이다. 이 질병 때는 미상핵에 있어 도파민에 대한 감수성의 증가를 볼 수 있다. 정신분열병은 환각, 망상, 행동이상을 주 증상으로 하는 정신질 환이다. 분열병 치료약의 대부분은 공통적으로 중추 도파민성 신경전달에 길항한다. 이같은 사실에서 분열병은 중추 도파민 뉴론의 기능항진에 의할 것이라고 추정되고 있다. 이 경우는 특히 중뇌-변연계 도파민 뉴론이 밀접하게 관계되는 것 같다. 중추의 카테콜아민 함량과 뒤에 설명될 세로토닌의 함량을 감소시키는 약물인 레세르핀 (rese rpi ne) 은 사람을 포함하여 동물 을 우울궁중에 빠지 게 한다. 이같은 사실로 우울증의 노르아드레나린 가설 또는 세로토닌 가설을 널리 받아들이게 되었다. 즉 우울병은 중추 카테콜아민 또는 세로토닌 함량이 저하되어 발생된다는 가설이다. 그러나 뒤에 다시 언급되겠으나 우울병 환자에서 카테콜아민 또는 세로 토닌 수용체의 수가 증가되어 있으므로 우울병은 뉴론의 활성도 와 더불어 수용체의 감수성 변화가 미묘하게 관련되고 있을 것으 로 생각된다. 5. 6 카테콜아민 신경과 약물 1) 말초신경 말초신경의 카테콜아민 뉴론(주로 교감신경)을 자극했을 때의 반응과 유사한 반응을 유발하는 약물을 아드레나린성 약물이라고
한다. 중요한 아드레나린성 약물의 화학구조와 작용 수용체 그리 고 나타내는 반응을 표 3-6 에 정리하였다. 표 3-6 아드레나린성 약물 약물명 R1 R2 R3 R, R5 수용체 주요반응 노르아드레나린 OH OH OH H H a’ 혈 압상승 ( Noradrenalin e ) 아드레나린 OH OH OH H CH3 a, /32 혈압상승 (A drenalin e ) (이 Is소op프 ro로t e 테r en레o놀l ) OH OH OH H CH(CH3)2 /3h /32 기관지확장 도파민 OH OH H H H /3I, D1 승압, 이 뇨 (Dop amine ) 훼닐에프린 H OH OH H CH3 a’ 승압 (Phenyl e p hrine ) 시네프린 OH H OH H CH3 a’ 승압 (S yn e ph rine ) 예훼드린 H H OH CH3 CH3 «1, /32 기관지확장 ( Ep he d rine ) 메타라미놀 H OH OH CH3 H a’ 승압 ( Meta ramino l ) 살부타몰 OH CH20H H H C(CH3)3 f31 기관지확장 (Salbuta w ol )
 RI:-CH-CH-N/ 凡
RI:-CH-CH-N/ 凡
a 수용체에 작용하여 혈압을 상승시키는 약물은 속 둥 혈압이 저하된 환자에 사용된다. 기관지 확장약은 천식에 사용될 수 있 다. 아드레나린성 a- 수용체 차단제인 디벤아민(Di ben amin e), 펜옥 시 벤자민 (Phenoxy b en zamine ) , 펜톨아민 (Phento l am ine ). 톨라조
린 (Tolazo li ne) 동은 혈압하강제로 사용되었으나 부작용의 문제로 현재는 잘 쓰이지 않는다. 현재 임상적으로는 쓰이지 않는 요힘빈은 접합부전막의 a2- 수용 체 를 선택적으로 차단시키는 약물이다. 아드레나린성 /3-수용체 차단제는 /3-수용체 효능제인 이소프로 테레놀의 화학구조를 변화시킨 것이 많다. 현재 수많은 유도체가 합성되어 심장의 부정맥이나 고혈압치료제로서 임상적으로 사용 되고 있다. 중요한 몇 가지를 그림에 나타내었다(그림 3 - 13)( 표 3-7 ) .
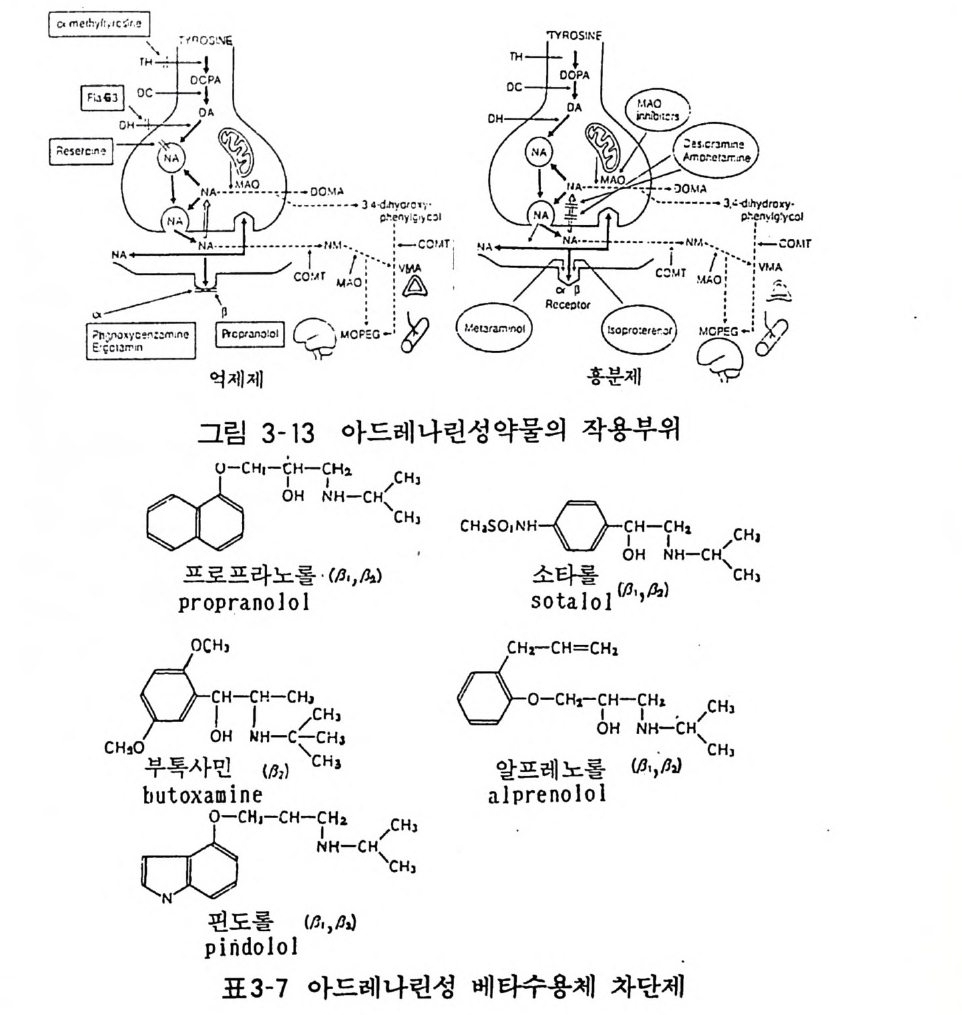 •-•- _ O• •O• M-A- ·- 3 .,,,CCW ·
•-•- _ O• •O• M-A- ·- 3 .,,,CCW ·
교감신경 아드레나린성 뉴론 말단에 작용하는 일련의 약물이 있다. 구안에티딘 (Guane thidin e) 은 신경말단에 작용하여 신경충격 에 따르는 노르아드레나린 유리를 억제한다. 그러나 시간이 경과 하면 노르아드레나린 저장을 고갈시킨다. 보레티리움 (Bre tyli um) 도 비슷하게 노르아드레나린 유리를 억제하는 약물이다. 이들 두 약물은 모두 중추신경계에는 도달하지 못하므로 말초신경계에 대한 작용으로 혈압하강 작용을 나타낸다• 레세르핀도 신경말단부 에 작용하여 노르아드레나린 저장을 고갈시키는 약물이다. 그러나 이것은 중추신경계에도 작용하여 진정작용도 나타낸다. 2) 중추신경 크로니딘 (Clo nidin e) 은 «2 - 수용체에 작용하여 노르아드레나린 유리를 억제하는 작용이 있으므로 이 약물은 혈압하강약으로 사용된다. 이것은 중추신경계 아드레나린성 신경에 대한 작용일 것으로 생각되고 있다. 알파메 틸도파 (a-me ty ldo p a) 도 혈압하강 작용 약물이다• 이것의 작용기전은 여러가지 가설이 있으나 대사 에 의하여 알파메틸노르아드레나린 («-me t h y lnoradrenal i ne) 을 생성하여 중추의 «2- 수용체를 자극함으로써 노르아드레나린 유리 를 억제하는 것이 주 작용기전일 것으로 생각된다. 정신분열 치료약은, 앞서도 언급한 바와 같이 중추신경계 도파 민수용체 길항제들이다• 이것에는 클로르프로마진 (Chlo rp romaz ine ), 할로페리돌 (Halo peri dol) 동이 있다. 정신병 의 일종인 우울 중 치료약에는 중추 카테콜아민 뉴론의 카테콜아민 재흡입 저해 작용이 있다. 그 대표적인 약물이 이미프라민(Imip r amin e) 이다. 각성제인 암페타민 (Am p he tamin e) 은 정신분열병과 유사한 증상 인 망상, 행동이상 둥을 일으킬 수 있다. 이 작용은 중추 카테콜 아민 뉴론으로부터 카테콜아민 유리를 촉진하는 것에 관계된다고 생각된다(그림 3-14).
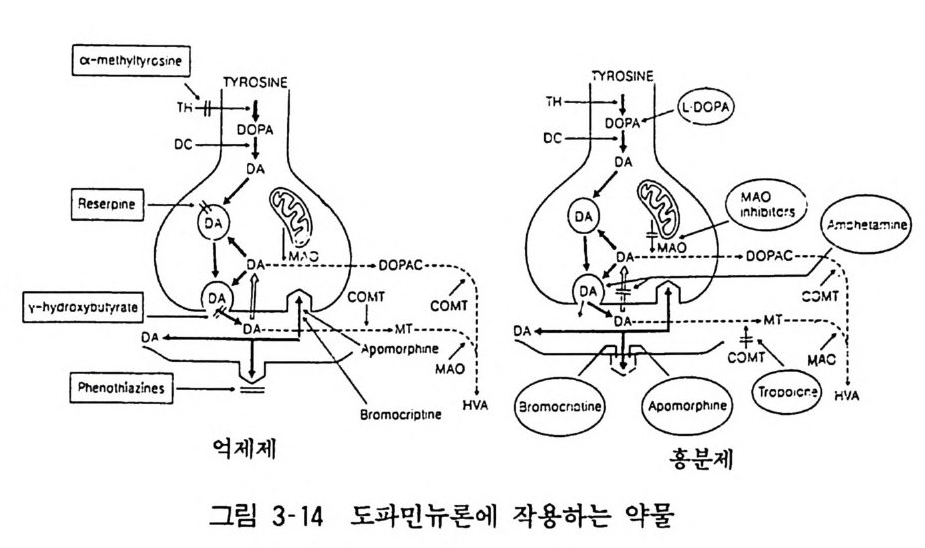 ~R i :\\
~R i :\\
6 세로토닌 (Sero t on i n, 5-HT) 세로토닌은 인돌핵(I ndole) 을 가진 생리활성 아민의 하나로 1940 년경에 발견되었다. 소화관이나 혈소판에 다량 함유되어 있고 최근 중추신경계에 세로토닌 뉴론이 존재하는 것이 밝혀져 중추 신경계의 전달물질일 것으로 생각되는 것이다. 6. I 합성과 대사 세로토닌은 생체내에서 트리프토판(trypt o p han) 으로부터 합성 된다. 이 것은 트리 프토판 - 5 - 수산화효소(trypt op han-5-h y dro xy lase) 에 의하여 5 번 탄소 위치가 수산화됨으로써 생성된다. 이 효소는 세포질내에 존재하고 합성된 세로토닌은 접합부소포내에 저장된다고 생각된다. 이 단계가 세로토닌 합성의 속도 조절단계 라고 생각된다. 기질인 트리프토판의 양은 적어서 이 전구물질의 이용도가 합성속도 조절단계로 생각되기도 한다. 신경충격으로 유리된 세로토닌은 모노아민 옥시다제에 의하여 5- 하이드록시인
돌아세틸산 (5-HIAA) 으로 대사되어 배 설된 댜 다 른 전달물질 에서 와 마찬가지로 유리된 세로토닌의 대부분은 신경말단에서 재흡입 된다(그림 3-15).
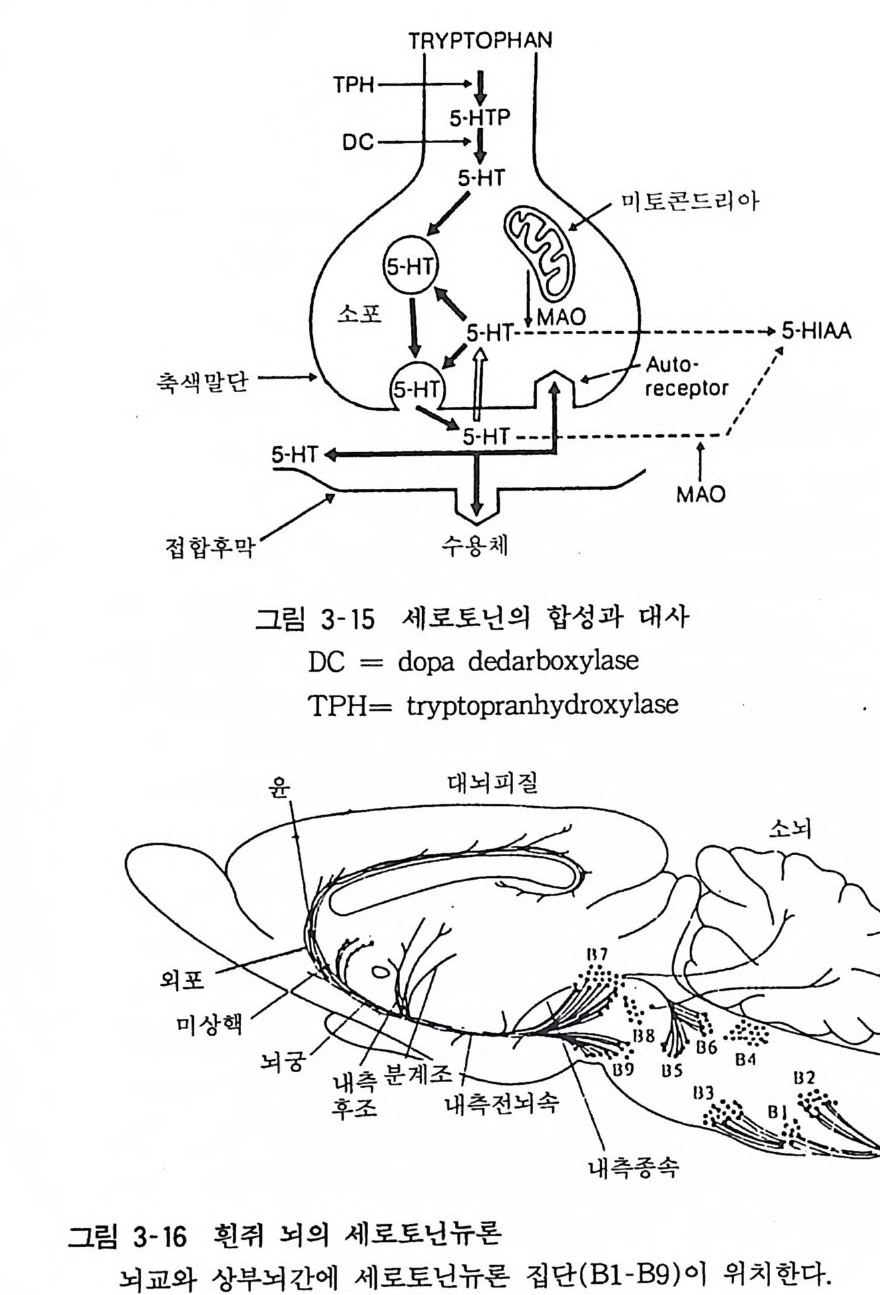 TRYPTOPHAN
TRYPTOPHAN
6. 2 세로토닌성 신경 중추신경계 세로토닌 뉴론의 주행은 카테콜아민 뉴론과 함께 스웨덴 학자들에 의하여 밝혀졌다. 세포체는 하위 뇌간 (Bra i n s t em) 의 편도핵 (Am yg dalo i d nucleus) 이라는 곳에 있고 중뇌나 뇌교 등에서 출발한 섬유가 상행하여 대뇌피질, 대뇌변연계, 시 상, 시항하부, 선조체 등에 뻗고 있다. 한편 연수에서 나오는 섬유 는 하행하여 척수의 일정 부위에서 끝나고 있다(그림 3-16). 6. 3 수용체 세로토닌 수용체에는 두 종류가 있다. 죽 5-HT1, 5-HT2 수용체 라고 부른다 5-HT2 수용체는 중추신경계에서는 전두엽 (Fron tal lobe) 에 많고 5-HTI 수용체는 해마(Hippocamp us) 에 많다. 정신 병 치료약인 스피페론 (S pipe rone) 은 5-HT2 수용체에 대한 친화 성이 강하여 이 수용체의 표식에 이용된다. 전부터 알려져 있던 효능제와 길항제들은 대부분 양 수용체에 대하여 유사한 친화성 을 가지나 최근에 개발된 케탄세린 (Ke tan se rin e) 이라는 약물은 5-HT2 수용체에 대하여 선택적인 친화성을 나타낸다. 5-HT 에 대한 결합친화성이 큰 수용체를 5-HT1 이라 한다. 5-HT1 수용체는 아데닐레이트 사이크라제와 연계되어 주로 중추신경계에서 세로토닌에 의하여 나타나는 억제성 반응들에 관계하고 한편 5-HT2 수용체는 이 효소와 연계되지 않고 주로 홍분성 반응에 관계된다고 한다(표 3-8). 6. 4 세로토닌성 신경과 생리적 역할 현재까지 말초신경계에서는 세로토닌 뉴론이 알려져 있지 않 다. 그러나 세로토닌은 소화관이나 혈소판에 다량 함유되어 있고 일정한 생리적 역할을 나타낸다. 예를 들면 장관운동의 항진,
표 3-8 세로토닌 수용체 리간드 수용체 효능제 길항제 S-HT1 LSD 메테르고린 5-HT, ( Mete r go l in e ) 메티서르지드 (M eth y se rgi de ) 5-HT2 LSD 스피로페리 돌 (S p iro p eri do l ) 케탄세린 (K eta ns eri ne )
지각신경 자극(통증), 혈압조절 동이다. 중추신경계의 세로토닌 뉴론은 생체의 정신기능, 행동, 수면 등과 깊은 관계 를 갖는다 . 동물읊 장기간 격리하여 한 마리씩 사육하면 투쟁적 행동 을 나타 내게 되며 이때 세로토닌 뉴론의 활성이 저하되어 있다. 또 세로 토닌뉴론을 인공적으로 파괴하면 동물의 자발적 운동의 증가, 상동행동의 중가가 관찰된다. 역으로 세로토닌 뉴론을 전기적으로 자극하면 동물의 자발 적 운동이 감소하고 진정 내지 수면 상태가 된다. 이같은 사실로 세로토닌 뉴론은 본래 동물의 일반행동에 대하여 억제적 역 할을 하는 것으로 생각된다. 카테콜아민이 역설적 수면과 관계 있다는 것은 이미 설명하였 으나 세로토닌은 이와는 달리 본태의 수면과 관계가 있다. 세로토닌은 말초에서 지각신경을 자극하여 통증을 일으키고 중추신경에서는 통각에 대한 감수성을 저하시키는 방향으로 작용 한다. 온혈동물의 체온조절은 시상하부의 체온조절 중추에서 이루 어지는데 세로토닌 뉴론의 말단부가 시상하부에 뻗어 있어 체온 조절에 관계될 것으로 생각할 수 있다. 여러 연구결과에 의하면 세로토닌 뉴론은 산욕열 (Pue rpe ral fe ver) 의 중추기전을 돕는 방향으로 작용하는 것이 밝혀졌다(그림 3-17).
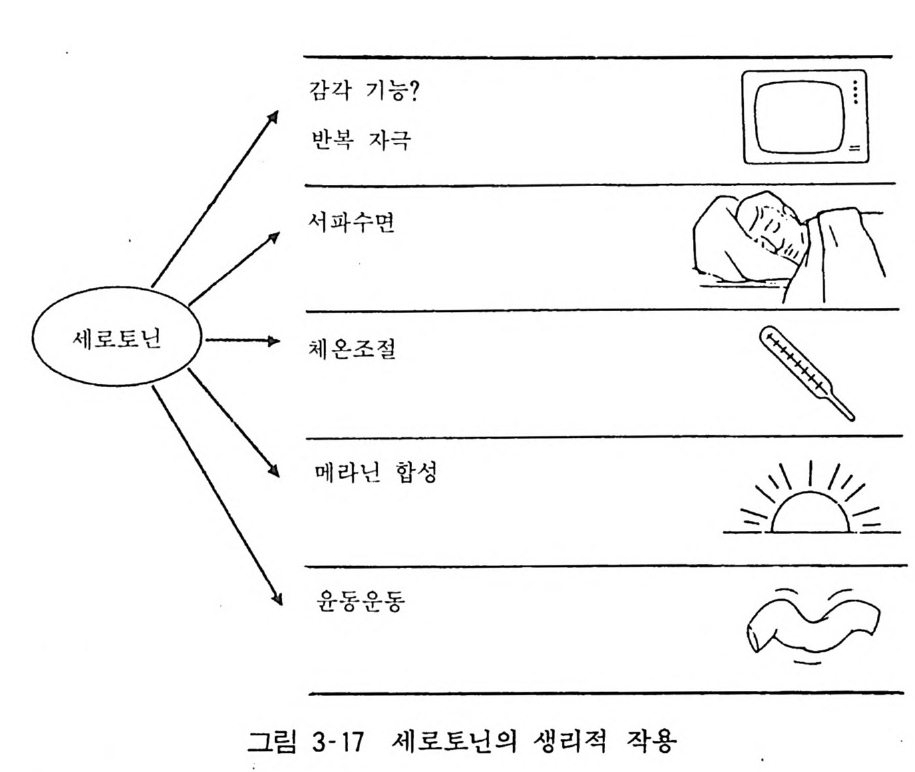 감각 기 능 ? 二
감각 기 능 ? 二
6. 5 세로토닌성 신경과 질병 세로토닌이 발견되었을 당시 세로토닌 수용체 효능제인 LSD- 25 가 정 신 분열병 적 증상을 나타낸다는 사실에서 중추 세로토닌 뉴 론 의 활 성도와 정신분열병의 관계 를 주목하게 되었다 . LSD- 25 뿐 아니라 정신이상을 일으키는 화학물질들 중에는 세로토닌 화 학 구조의 골격을 이루는 인돌핵을 가진 것이 많다 . 또 정신분열 병 환자의 소변 중에는 정상인의 소변에서는 발견되지 않는 세로 토닌의 이상 대사물이 발견된다. 또한 정신분열병 치료제인 스피 페론은 세로토닌 수용체를 차단한다. 이같은 사실들로부터 정신분 열병의 병인으로 중추 세로토닌 뉴론의 활성도와의 관계 를 강조 하는 설이 있으나 앞서도 말한 바와 같이 현재까지는 도파민 뉴론의 활성도가 보다 깊이 관계된다고 생각되고 있다. 중추 세로토닌 함량의 저하가 우울증을 초래한다는 우울병의
세로토닌 가설은 이미 설명했다. 최근의 연구에 의하면 뉴론의 활성도에 더하여 세로토닌 수용체에 대하여도 고려해야 한다는 결론에 이르고 있다. 죽 우울증에는 세로토닌 뉴론의 활성도가 낮고 그것에 상웅하 여 세로토닌 수용체의 감수성이 증가되어 있다. 이같은 상태에서 스트레스에 의하여 세로토닌이 유리되면 정상인에 비하여 중추 세로토닌의 작용이 강하게 나타남으로 우울증이 유발된다는 생각 이다. 6. 6 세로토닌 신경과 약물 카테콜아민 뉴론에 작용하는 약물의 대부분은 세로토닌 뉴론에 영향을 미친다. 레세르핀은 중추 카테콜아민뿐 아니라 세로토닌도 고갈f l 킨다. 따라서 이들 두 가지 아민 중 어느 쪽이 레세르핀의 진정작용과 연관이 있을지는 확실치 않다. 항우울제인 이미프라민 도 카테콜아민과 마찬가지로 세로토닌의 신경말단에서의 재홉입 올 억제한다. 항정신병약인 스피페론이 세로토닌 수용체 를 차단한 다는 것은 이미 언급했으나 길항제인 메티테핀 (Me tit e pi ne) 등도 분열병 치료제로 사용된다. 암페타민이나 LSD-25 동은 소량으로 생체에서 정신이상이나 이상행동을 일으키는 물질로서 정신이상 발현물질 또는 환각제라고 한다. 이같은 약물 중에는 카테콜아민 과 유사한 구조나 인돌핵을 가진 것들이 많다. 인돌핵을 가진 것은 세로토닌 수용체에 대한 친화성이 큰 것이라 생각된다. 따라 서 환각 발현에는 세로토닌 뉴론이 관여할 것이 예상된다• 그러나 현재까지 자세한 기전에 대하여는 밝혀져 있지 않다. 대마의 유효 성분인 마리화나는 테트라하이드로칸나비놀 (THC) 의 화학구조를 갖는다. THC 를 흰쥐에 투여하면 정상적인 흰쥐에서는 볼 수 없는 이상행동을 나타낸다. 그 중에서도 생쥐를 교살하는 행동이 특징적이다. 중추 세로토닌 뉴론의 활동도가 저하하면 이같은 행동을 나타내는 것을 알게 되 었다(그림 3-18).
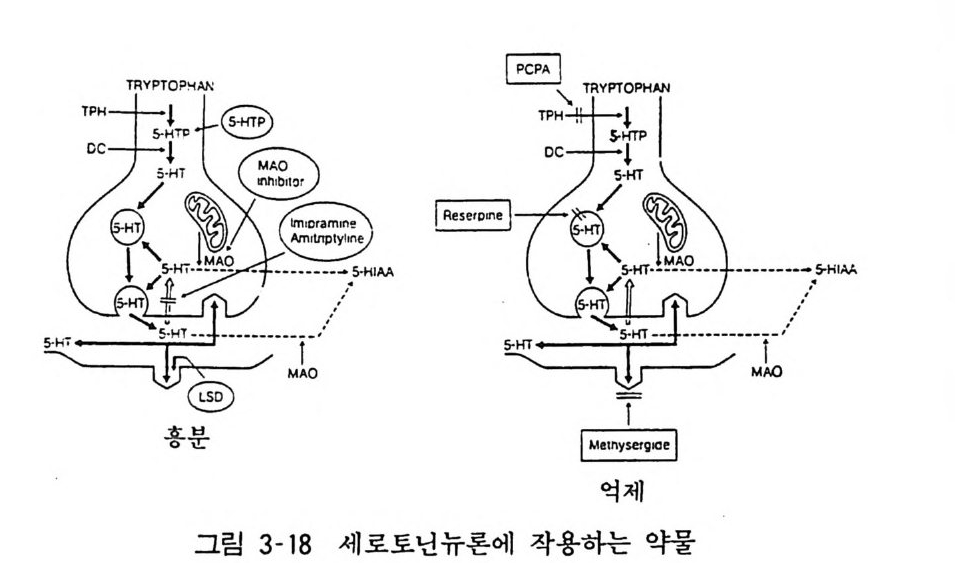 IB Y PTO ?>< 사 : 孛 TR Y P訂T/
IB Y PTO ?>< 사 : 孛 TR Y P訂T/
7 감마아미노브티릴산 (GABA) 척추동물의 신경계에는 여러가지 아미노산이 존재한다. 그 중에 서 어떤 종류의 아미노산은 특히 뇌내에서 생산되는 것으로 생각 된다. 따라서 일반적인 아미노산과는 달리 대사적 의미나 세포의 생리 적 기능 유지의 역할 이외에 신경전달물질로서의 역할을 가질 것으로 생각되는 것이다. 이 같 은 아미노산계열의 전달물질로는 글루타민산, 아스파라긴 산 , 감마아미노브티릴산, 그리신 둥이 있다. 이중에서도 GABA 는 1950 년 처음으로 포유동물 뇌내에 다량 존재한다는 것이 알려 지고 나서 중추신경계 억제성 전달물질로서의 의의가 있어서 연구되어 온 아미노산이다• 7. 1 합성과 대사 GABA 는 글루타민산에서 글루타민산 탈탄산효소 (GAD) 에
의하여 생성된다. 또 GABA 아민전이효소 (GABA-T) 에 의하여 알파케토글루탈산 (a-ke t o g lu tari c acid ) 사이에 가역적으로 아미 노 기의 전이를 받아 석시닐세미알데하이드 (succ iny lse rni aldeh y de) 와 글루타민산(g lu tami c ac i d) 으로 바뀐다. 석시닐세미알데하이드 는 탈수소효소 (SSADH) 에 의하여 석시닐산이 된다. 이것은 TCA 회로에 들어가 산화된다 . 이 같은 반응계 를
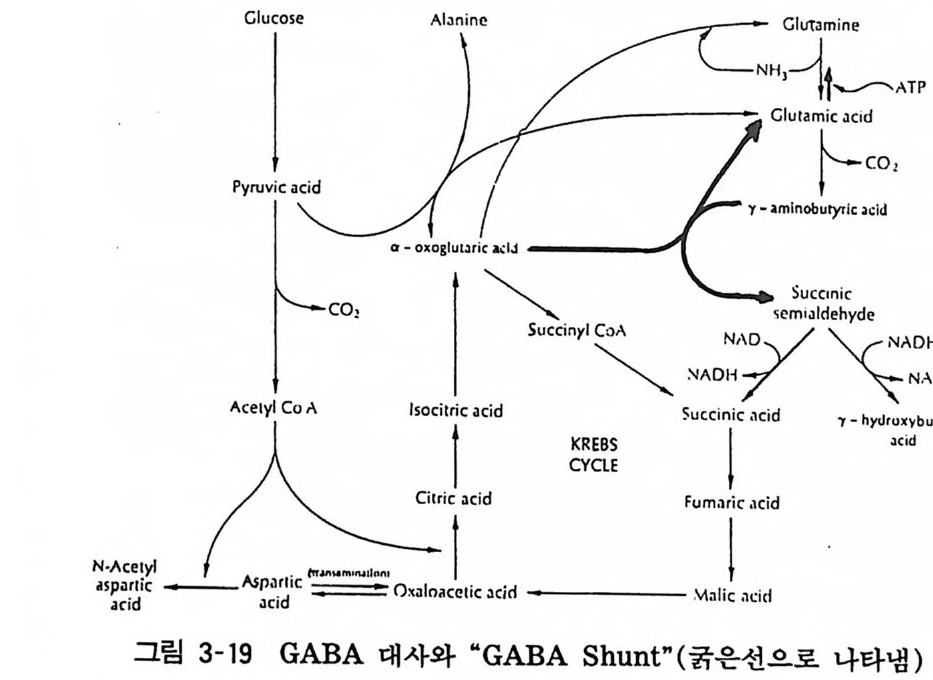 Glucose Ala nin e GIUlam i ne
Glucose Ala nin e GIUlam i ne
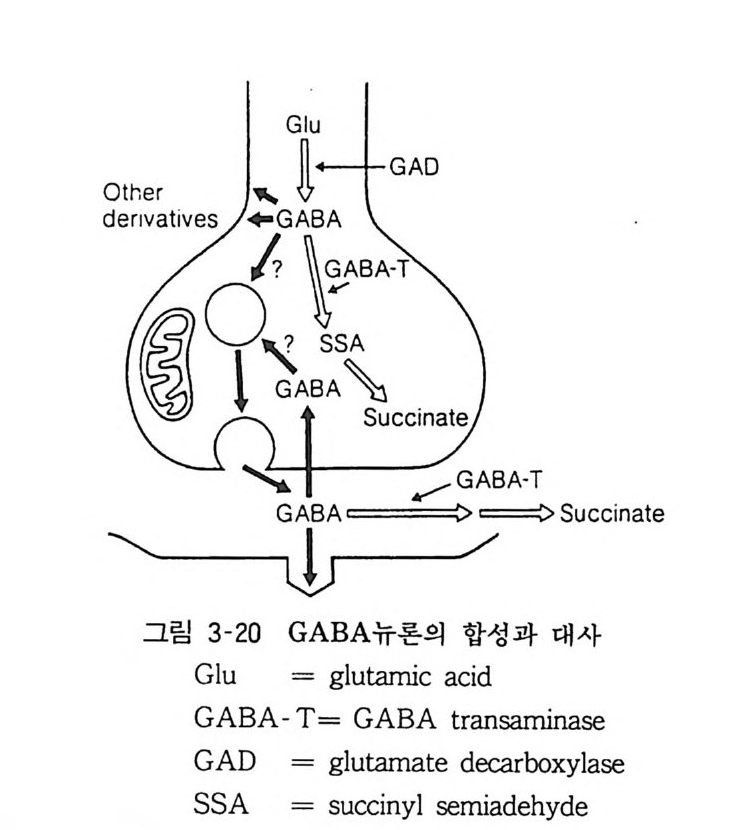 ,') _s`"u�A G
,') _s`"u�A G
�X��� ������ (h yperpo l arizati on) ��. ����X
Հ� x�!�|��Ŕ� GABA h�ɷt� ι� t� ����Ŕ� -��� ɔ�� ��<���\� GABA t�` �t� p�� ��`�D� `� ���1�t� l�� (���� 3-21). 7. 3 GABA ©ƴ� GABA ©ƴ��� �\� ��l��� t�� ���� ����X��. ��8�\� ��@� L�$����� J�X�<ǘ� GABA �� 8���ɹX� �Ō� (CI) t�(� ,���1� �� ��t�� �i���ɹ����� ������D � |�<Ǩ��. 0�|�� ©ƴ̔� t��@� ����� ��XՌ� �(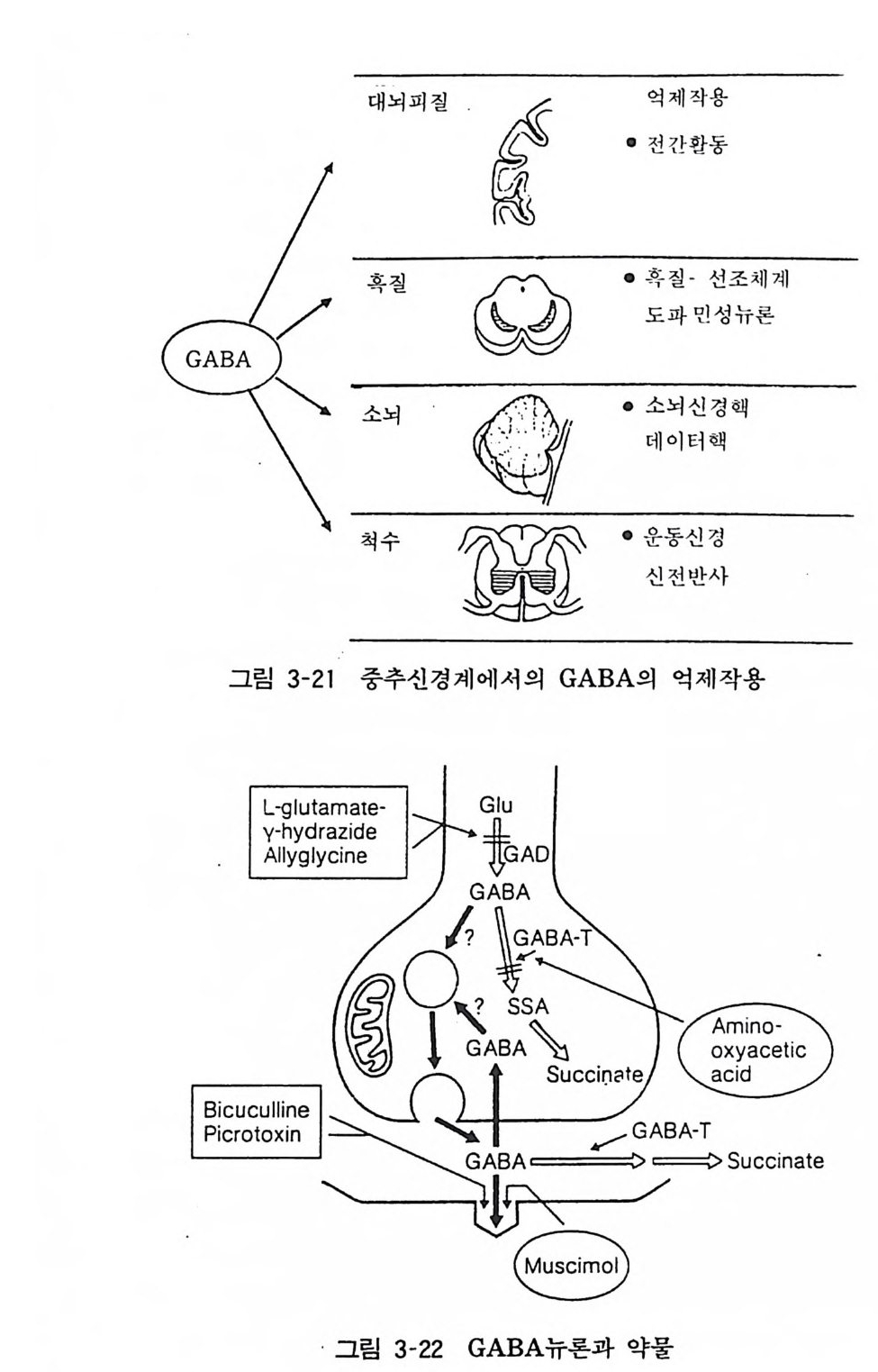 -- :
-- :
이용한 연구로서 중추신경계 GABA 수용체는 소뇌, 대뇌피질, 중뇌, 해마, 선조체, 연수, 시상하부, 척수의 순으로 존재한다고 알려졌다(그림 3-22). 신경병의 치료약으로 잘 사용되는 벤조디아 제핀 (Be nzodi aze pi nes) 은 GABA 뉴론의 활성을 높이는 작용이 있다. 최근의 연구에 의하면 GABA 수용체와 벤조디아제핀 수용 체는 세포막 위에 대단히 근접하여 존재하고 서로 상호작용을 나타내는 것으로 알려졌다. 7. 4 GABA 성 신경과 질병 파킨손씨병 때 혹질, 미상핵의 도파민 뉴론의 변성이 관계된다 는 것은 이미 언급했다. 이 질병 때 GABA 뉴론의 활성이 저하 된댜 한편 헌팅톤 무도병 환자에서는 선조체 뉴론의 변성과 GABA 함량 저하, GAD 활성 저하 및 GABA 와 수용체 결합의 증가를 볼 수 있다. 또 이 증상 때 소뇌의 GABA 함량도 증가한 다. 따라서 이들 질환에서는 GABA 뉴론의 활성, 특히 수용체 성상의 변화가 중요한 의미를 갖는다고 한다. 8 뉴로펩 타이드(N euro p e pti de) 신경계 조직에 존재하여 생리활성을 갖는 펩타이드에 관한 연구가 활발하게 이루어지고 있다. 1975 년 몰핀과 · 유사한 활성을 가진 엔케파린이 뇌내에서 발견된 후 뉴로펩타이드의 연구는 매력적 중요 연구대상으로 부각되어 현저한 발전을 보았다. 새로운 생리활성 펩타이드가 분리되어 구조가 밝혀지면 그 생리적 작용 규명에 노력이 집중된다. 새롭게 발견된 펩타이드가 신경전달물질이라고 하려면 몇 가지 조건을 만족시켜야 한다.
1) 접합전부(신경말단)에 존재하는가 2) 뉴론에서 합성되고 그곳에 저장되는가 3) 뉴론에 대하여 홍분 또는 억제 작용이 있는가 4) 뉴론으로부터 유리되는가 5) 접합전섬유의 자극으로 접합 후 뉴론에 전위변화 를 유발하 는 7} 현재까지 발견된 뉴로펩타이드 중에서 위의 조건을 완전하게 만족시키는 것은 없다. SP, LH-RH, 엔케파린 (enke p ha li ns) 등이 가장 가능성이 큰 것이다. 이같은 기준으로 볼 때 전달물질이라 하기에는 미흡한 물질들은 뉴로모듀레이타 (neuromodula t or) 라고 한다. 이들은 자극전달에 관여하기는 하나 직접적이 아니고 간접 적으로 조절역할을 한다는 것을 의미한다. 그러나 뉴로펩타이드 를 전달물질이라 할 것인지 뉴로모듀레이타라 할 것인지가 분명하게 되어 있지 않다. 어떻게 분류할 것인지는 그리 중요한 것이 아니 고 보다 중요한 것은 각 펩타이드 신경계에 있어서 구체 적 인 기능적 역할과 그 기전을 밝히는 것이다. 표 3 - 9 에 전달물질로서의 가능성이 있는 뉴로펩타이드 를 정리 하였다. 종래 한 개의 뉴론은 한 가지 신경전달물질 을 갖는다는 데일 (Dale) 의 원리가 있었으나 뉴로펩타이드의 등장으로 종래에 전달물질(후보)이라고 알려진 아세틸코린, 노르아드레나린, 도파 민, 세로토닌 둥과 함께 뉴로펩타이드가 공존하는 것이 여러 예에 서 발견되었다(표 3-10). 아민과 펩타이드의 공존의 생리적 의 미, 아민 유리와 뉴로펩타이드의 기능적 관련성에 관한 금후의 연구 결과가 주목된다. 어찌되었건 뉴로펩타이드가 신경계에 존재 하는 것이 알려짐에 따라 신경전달 과정이 수정되어야 하지 않을 까 생각된다. 예를 들면 코레치스토키닌 (CCK) 또는 VIP(vas- oact ive intes tin al pepti de) 는 장관에서 신경내분비 홀몬으로 작용 하나 세포와 세포 사이의 정보전달을 담당하는 물질이라는 점과 뇌에도 이들 물질이 존재한다는 점을 고려하면 뇌에서도 이들
표 3-9 전달물질후보 뉴로펩타이드 엔갑돌상핀선 자( E극nd홀or몬p 펩h i유n타 )리이 홀드몬 (TRH) 뉴론에의+ 존+재+ +뉴론+에+서++ + 유?+리+ + +++ 안지오텐신 II(Ang iot e n sin II) 바옥섭난 소시포스토프탄자레신스극 피홀신(O 몬((x SV y uat유b soosc리t pian r 홀n)e sc몬s ei n (P )L )H -RH) +++ ?+ 뉴로텐 신 ( Neurote n sin ) 소마토스타틴 ( Somato s ta tin) + 코가혈스레관치트활 스성린 토장( G키관as닌펩tr i타(nC 이)C K드) ( VIP) ++ ? 브라디 키 닌 ( Brady kinin ) +
표 3-10 기존신경전달물질과 동시에 존재하는 뉴로펩타이드 기존전달물질 뉴로펩타이드 부위
세로토닌 섭스탄스피 연수(흰쥐) TRH 연수(흰쥐) 섭스탄스피, TRH 연수(흰쥐) 노르아드레나린 소마토스타틴 교감신경절(기니픽) 부신수질(사람) 엔케파린 상경교감신경절(흰쥐 ) 부신수질(각종동물) 뉴로텐신 부신수질 아드레나린 엔케파린 부신수질(각종동물) 도파민 엔케파린 경동맥구(고양이, 개) CCK 뇌 복측피개(흰쥐, 사람) 아세틸코린 VIP 자율신경절(고양이)물질이 신경홀몬으로 작용할지도 모르겠다. 이 경우 신경홀몬 작용이라 함은 표적부위가 유리부위로부터 멀리 떨어져 있고 효과가 강하고 효과의 지속 시간이 길다는 것이 특징이다. 따라서 펩타이드의 뇌 또는 신경절에 있어서의 작용 시간 경과와 역치 (세포를 홍분시키는 데 필요한 최소 자극량)가 밝혀지고 종래의 전달물질과의 차이가 밝혀지면 펩타이드의 뇌에 있어서의 기능과 작용기전이 분명해질 것이다. 새로운 펩타이드가 발견되고 그 구조(아미노산 조성)가 밝혀지 면 펩타이드가 합성에 의하여 대량 생산되고 그 생리적 역할과 약리작용이 검토된다. 또 항체가 만들어지면 그 펩타이드의 미량 분석을 위한 방사성 동위원소가 개발되고 형광표식 항체의 분포 로 조직내 분포를 추정하게 된다. 이같은 면역조직화학적 방법에 의하여 소화관에서 분리된 펩타이드인 SP, VIP, 가스트린 (Gas trin),코레치스토키닌이 뇌내에 존재한다는 것이 알려졌다. 이같이 대부분의 생리활성 펩타이드가 신경조직과 위장관에 존재 한다는 것은 홍미있는 사실이나 그 생리적 의미는 명확치가 않 다. 뉴로펩타이드 연구는 최근 10 년 동안에 많은 진전을 보아 알 파-, 베타-네오엔돌핀 (a-, /3- neoendorp hin), 교돌핀 (K y o t orr p h in), 뉴로키닌 (Neuro kinin), 카세이몰핀 (Case irn o rp h in) 등이 새로 이 발견되었으며 금후에도 많은 중요한 과제들이 남아 있다. 예 를 들면 1) 작용기전, 죽 신경전달 과정에서의 역할 2) 생리적 약리 적 작용 3) 종래부터 알려져 있는 신경전달물질을 함유하는 뉴론 과의 기능적 해부적 관련성 4) 특이한 펩타이드 단백분해효소의 존재 여부와 이것의 신경활성 펩타이드 합성, 분해와의 관계 5) 신경질환과의 관련성 6) 의약품으로서의 유용성 7) 구조 유사성 을 이용한 길항제 발견 가능성 8) 나아가 많은 종류의 신경활성 펩타이드가 존재하는가, 존재한다면 무엇 때문에 많은 종류가 존재하는가둥이다.
9 섭 스탄스피 (Substa n ce P) 중추신경계에 존재하고 뉴 론 에 대하여 활성을 나타내는 펩타이 드 를 신경활성 펩타이드라고 한다. 최근 신경활성 펩타이드에 관한 연구가 크게 관심을 끌어 급속한 진전을 보게 되어 속속 새로운 물 질이 발견되고 그 성질이 밝혀지고 있다. 그 결과 몇 가지 펩타이드는 아민이나 아미노산처럼 신경전달물질로서의 역 할을 나타낼 것으로 생각되고 있다. 그중에서 섭스탄스피 (SP) 는 그 가능성이 가장 높은 것이라 하겠다. 섭스탄스피 (SP) 는 1931 년 폰 율러 (von Euler) 와 갇덤 (Gad dum) 이 처음 뇌와 장관에서 발견한 활성물질로서 타액분비 촉진작용 (s i alo g o gi c ac ti v ity)과 평활근 수축작용 (s p asmo g e ni c ac ti v ity)을 나타내는 펩타이드이다. 기타 신경활성 펩타이드의 대부분도 장관에 존재한다. SP 와 거의 똑같은 활성을 나타내는 피사레민(p h y sale rni n) (남미의 개구리 피부 펩타이드) 에레도이신 (eledois i n ) (지중해 두족류의 타액선 펩타이드)이 SP 와 대단히 유사한 구조 를 가진다는 것은 홍미있는 일이다(표 3-11).
표 3-11 섭스탄스피 및 유사 펩타이드 섭 스탄스피 Arg- Pro-Ly s- Pro-Gln-Gln-P h e-Phe-Gly- Leu-(Substa nc e P) Met- N H2 피사레 민
9. 1 합성과 대사 SP 의 뉴론내에서의 합성 과정에 대해서는 현재까지 전혀 알려 져 있지 않다• 최근의 연구에 의하면 세포체에서 우선 고분자
단백질의 전구물질이 합성되고 SP 로 분해된 다음 접합부소포내에 저장되어 축색류를 따라 신경말단부로 운반되는 것으로 생각된 다. 이와 같은 과정은 다른 펩타이드 전달물질에도 해당하는 것으 로 생각된다. 후근신경절 (Dorsal root g an gli a) 을 이용한 시험관적 연구에서 방사성 메티오닌의 SP 로의 결합이 사이크로핵사미드 (c y clohex ami de) 에 의해서 차단됨이 알려졌다. 이것은 SP 합성이 리보솜에서 일어난다는 것을 말하는 하나의 근거가 되는 것이 다. 중추신경내에서 SP 의 분해 과정에 대해서도 분명치 않은 점이 많다. 가토 (Ka t o) 둥의 연구에 의하면 우선 N 말단의 아미노 산 2-4 개가 떨어지고 아미노산 7 개의 펩타이드가 된다 [(5-11) SP]. 이어서 아미노펩티다제 (am i no pepti dase) 와 엔도펩티다제 (endo pepti dase) 에 의하여 잘게 끊어져 불활성화된다. SP 는 아민이나 아미노산 신경전달물질에서와 같은 신경말단으 로의 재흡입 기전은 존재하지 않는다. 그러나 최근 연구로 (5- ll)SP 까지 분해된 다음 신경말단이나 그리아세포로 흡입되는 현상의 존재 가능성이 알려졌다. 이같은 과정은 아세틸코린의 재흡입 과정과 유사하나 아세틸코린과는 달라 분해산물인 (5- ll)SP 는 그 자체가 불활성 펩타이드가 아니고 오히려 SP 보다 생리적 활성이 높은 것이다. 9. 2 SP 성 신경 SP 는 생체 여러 장기 조직에 널리 분포하고 있어 방사성면역검 정법이나 면역조직화학적 연구로 그 분포가 알려져 중추신경계에 는 혹질에 가장 높은 농도가 함유되어 있다고 하며 추체외로계인 내측담창구 (Inner glo bus palli dus) 에도 높은 농도로 존재한다. 그리고 시상하부, 구심성 감각계, 척수의 후각 (Dorsal horn) 에 풍부하게 존재한다. 또 연수의 삼차신경핵(trig e min al nucleus) 에도 존재한다. 이같이 SP 가 여러가지 중추신경계 기능에 관계되 는 부위인 내분비-변연계, 추체외로계, 감각체신경계에 존재한다
는 것은 이 펩타이드가 중추신경계에서 복합적인 생리 기능에 관계될 것임을 나타내는 것이다. 9. 3 SP 성 신경의 생리적 역할 SP 는 소화관을 비롯하여 말초장기에도 존재한다. 이들 부위에 서도 생리활성을 나타낸다. SP 는 혈관 평활근을 이완시켜 혈압을 하강시킨다. 한편 포유동물의 위장관 평활근에 대해서는 강력한 수축을 일으킨다. SP 는 대부분의 포유동물에서 타액분비를 촉진 한댜 이들 작용은 모두가 효과장기에 대한 SP 의 직접작용의 결과로 나타나는 것이며 SP 뉴론에 대한 작용에 의한 것은 아니 다. SP 는 대부분의 신경세포에서 흥분작용을 나타낸다. 척수 운동 뉴론에 대한 작용은 강력하다. SP 는 일차 지각뉴론의 신경전달물 질로 생각되고 있어서 말초로부터의 지각의 전달 특히 통각의 전달에 관여하는 것으로 생각되고 있다. 그러나 한편으로는 SP 가 진통작용을 갖는다는 보고도 있어서 일정치가 않다. SP 를 동물의 중추신경내에 주입하면 동물 종에 따라서는 운동량이나 상동행동의 증가를 보이기도 하고 반대로 진정작용을 나타내기도 한다. 체온에 대한 영향도 동물에 따라 다르다. 이상과 같이 SP 의 작용은 아직 분명치가 않으나 동물의 정신, 운동 기능에 어떤 영향을 미치는 것은 사실이다. 9. 4 SP 성 신경과 질병 이미 언급한 헌팅톤 무도병은 미상핵의 진행성 위축에 의하여 발생하는 질병으로 우성유전을 나타낸다. 병태생리학적으로는 선조체의 GABA, 아세틸코린 감소와 도파민의 증가가 알려져 있으나 최근 이 질병 때 혹질의 SP 가 현저하게 감소되어 있는 것이 알려졌다. 아마도 헌팅톤 무도병 때는 선조체의 SP 뉴론의
변성이 일어났을 것이라 생각된다. 9. 5 SP 신경의 효능제와 길항제 SP 신경의 기능 연구를 어렵게 하는 요인 중의 하나는 SP 신경 에 대한 효과적이고 특이한 효능제 또는 길항제가 없다는 것이 다. 최근 D- 아미노산 대체로 SP 유도체를 얻었으며 D-Pro 또는 D - T rp가 그 예로서 이들은 중추신경계 및 말초장기에서 SP 의 여러 작용을 차단한다고 한다. IO 오피 오이 드 펩 타이 드 (O pt o i d pe p tide ) 오피오이드 펩타이드의 발견은 몰핀 수용체에 대한 결합특성 연구로부터 이루어졌다. 일반적으로 생체 조직에 함유되어 있는 전달물질이 증명되고 이것에 작용하는 수용체의 증명을 위하여 여러가지 리간드들이 이용되고 그 결과 수용체의 특성이 밝혀지 는 것이 순서이나 오피오이드 펩타이드의 경우는 몰핀 또는 나록 손 (Naloxone) 둥의 리간드 결합특성으로 몰핀 수용체가 알려지고 이어서 이것에 상응하는 전달물질로서 오피오이드 펩타이드의 존재가 알려지게 된 것이다. 몰핀 수용체에 작용하는 물질을 뇌 추출물에서 분리하고 그 구조가 밝혀져 엔케파린 (enke p hal i n) 은 메티오닌-엔케파린과 류신-엔케파린의 두 가지가 있음이 밝혀졌다. 유전공학적 방법으로 베타엔돌핀(/3 -endo rphin)의 모체단백질이 부신피질자극홀몬 (ACTH) 의 모체단백질인 프로오피오메라노코틴 (pro -op iom elanocort in) 이 라는 것을 밝혔다. 프로오피오메 라노코틴 이 효소적으로 분해되어 a-MSH, ACTH, 베타리포트로핀(이것이 더 분해되어 감마와 베타엔돌핀이 된다)이 생성된다. 현재 오피오
이드 펩타이드 (o pi o i d pepti de) 의 전구체는 세 종류로 분류한다. 앞에 말한 프로오피오메라노코틴 (POMC) 외에 프로엔케파린 에이(p roenke p ha lin e A) 는 메티오닌엔케파린과 류신엔케파린을 생성하고 프로다이놀핀은 다이놀핀 (d yn o rp h in e), 알파 - , 베타 네오엔돌핀을 생성한다(그림 3- 23 ).
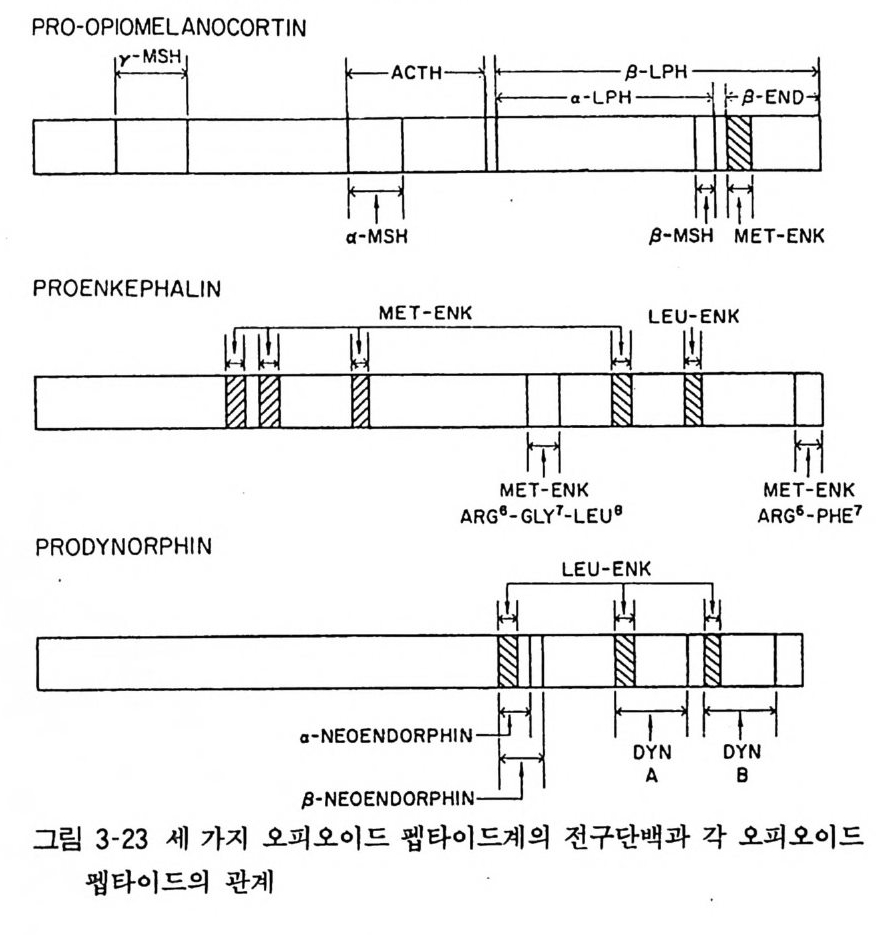 rAICTHl 尸三〕广
rAICTHl 尸三〕广
IO. I 수용체와 리간드 몰핀 (Morp h i ne) 유사약물들의 결합특성에 따라 뇌와 기타 조직 장기에는 여덟 가지의 수용체가 존재할 것으로 알려지고 있다. 이중에서 중추신경계에는 뮤(µ), 카파(,c), 델타 (IS), 시그마
((1) 둥이 있음이 많은 증거들로 증명되었다. 이들의 여러 장기 조직의 분포나 리간드에 대한 결합특성에 대한 연구결과들은 다양하나 진통효과는 뮤와 카파 수용체에 연관되고 불쾌감 (dy sp h o ri a ) 이 나 향정 신성 (psych oto m ime ti c ) 효과는 시 그마 수용 체와 관련된다고 생각되고 있다. 델타 수용체는 주로 대뇌변연계 에 분포하는 것으로 미루어 정동적 행동 (a ff ec ti ve behav i or) 과 관련될 것으로 생각된다. 시그마 수용체를 제외한 모든 타입의 몰핀 수용체에 대하여 길항제인 나록손 (Naloxone) 은 결합친화성이 높다 . 일반적으로 뮤 수용체에 대한 친화성이 카파나 델타 수용체보다 열 배 크 다. 엔케파린 결합특성은 델타 수용체를 구별하는 데에 절대적인 역할을 하나 이와 유사한 결합특성으로 델타 수용체에 작용하는 약물은 잘 알려져 있지 않다. 뮤, 카파 그리고 시그마 수용체에 대한 약물의 결합특성은 표 3-11 에 정리하였다. 11 프로스타그란먼(P ros t ag land i ns) 1982 년 노벨 의학 생리학상은 프로스타그란딘 (PGs) 에 대한 연구업적으로 베인 (Vane, 영국), 베르그스트롬 (Ber g s t rom, 스웨덴), 사무엘손 (Samuelson, 스웨덴)들에게 주어졌다. 1930 년대에 시작된 PGs 의 연구는 1970 년을 중심으로 급속 한 진전을 보았다. 트롬복산(thr omboxane), 프로스타싸이크린 (p ros t ac y cl i n) 의 발견을 위시하여 1980 년대의 류코트리엔 (l euko tri ene) 의 발견에 이르렀다. PGs, 트롬복산 (TX), 프로스타싸이크린 (PG i s), 류코트리엔 은 모두가 불포화 지방산인 아라키돈산 (arac hi do ni c ac i d) 에서 효소에 의하여 합성된다(그림 3-24). 이들 중 뇌에서 생합성
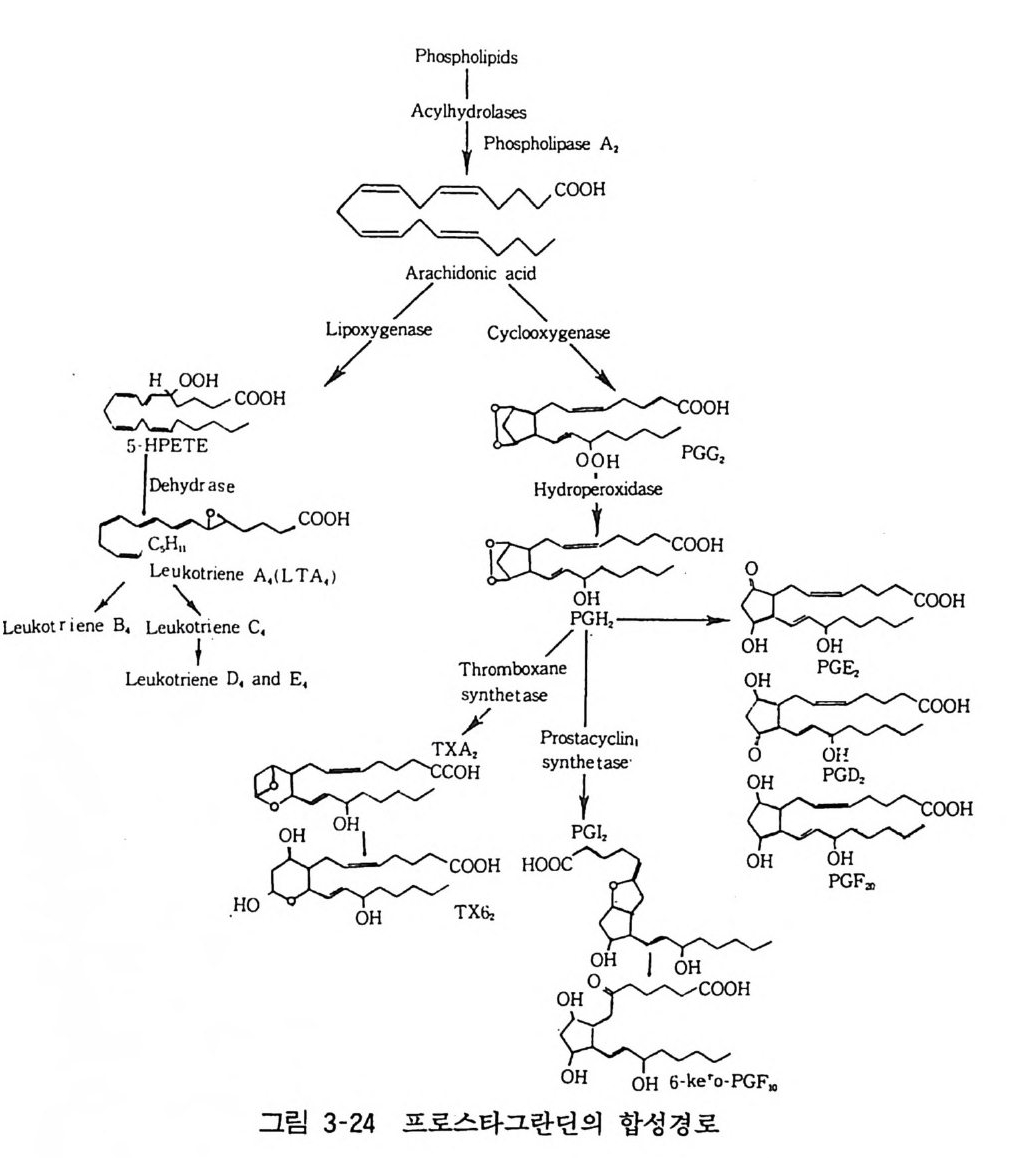 APchyo slhp yth I d or l oiP p lhaidsoses s p h olip a se A,
APchyo slhp yth I d or l oiP p lhaidsoses s p h olip a se A,
활성 이 알려 진 PGE2, PGF2, PGD2, TXA 파 있다. PGs 는 강력한 평활근작용, 혈소판 응집작용, 염증작용을 갖는 것으로 알려졌다. 또 교감신경계에 대한 작용으로 PG E:i는 교감신경 억제효과를 나타내며 이것은 교감신경 전달물질 유리의 감소에 기인하는 것으로 생각된다. 또 PG 는 접합후막의 노르에피테프 린에 대한 반응성에도 변화를 일으킬 것으로 생각된다. 뇌에서
는 PGE :i, PGF2,,, PGD 가 존재하나 이중 PGD 마 가장 많댜 PGD2 의 합성효소와 분해효소도 뇌에 존재한댜 뇌에서의 생리 적 역할로서는 혈압, 체온, 체액, 섭식 둥의 조절에 관여하고 시상하부나 뇌하수체 홀몬분비에 대한 작용이 있다. 가장 주목 을 끄는 것은 자연수면에 가까운 수면을 일으킨다는 점이다. 따라서 내인성 수면물질로서 작용할 가능성이 있는 것이다. 중추신경계에서 신경전달물질일 것인지는 아직 분명치 않다. PG 는 뇌에 거의 균등하게 널리 분포한댜 따라서 PG 가 어떤 특정 기능의 전달물질로 작용할 가능성은 생각하기 힘들다. 오히려 모듀레이터일 가능성을 생각할 수 있다. 신경전달물질 이 접합후막에 작용하여 세포막 아라키돈산으로 부 터 PG 가 생성되고 이것이 신경전달물질의 유리 또는 접합후막 수용체에 대한 감수성의 변화를 일으켜 신경전달 과정을 변화시킬 수 있을 것으로 생각하는 것이다. PGD2 의 접합후막에 대한 특이한 결합과 cAMP 증가작용이 알려져 있다. 12 신경계의 생화학적 연구방법 신경계 특히 뇌의 활성을 연구하기 위하여 여러가지 표본이나 기법이 이용되고 있다. 여기서 현재 자주 이용되고 있는 생화학적 연구방법을 간략하게 살펴보기로 한다. 12. I In v i vo 에서 의 방법 In vi vo 에서의 뇌의 활성을 해석하는 것은 생리적 조건에 가까 운 상태에서의 검토가 가능하다는 점에서 우수한 방법이다. 그러 나 뇌의 특정 부위에서 활성을 조사하는 것은 매우 힘든다. 일반
적으로는 뇌 척수액, 혈액, 소변에서 전달물질이나 이것의 대사물 의 양적 변동을 조사함으로써 뇌의 활성을 추정하고 있다. 사람의 정신질환 진단에 이용되는 방법이다. 이것은 기술적으로 용이하기는 하나 얻어진 결과가 뇌 이외의 말초장기 활성 변화에 의 할 수도 있는 것이다. 이것을 피하기 위해서 동물 실험에서 뇌순환과 전신순환을 분리하고 뇌순환만으로 해석하는 방법도 있댜 이 것을 뇌관류법이라 한다. 머리 를 완전히 분리하여 관류하 는 방법과 혈류 의 일부 를 분리하는 방법이 있다. 관류액으로는 신선한 혈액 이나 크렙스-링겔액 (Kreb's-R ing er) 같은 영양액을 사용할 수 있다. 간접적으로는 앞서 언급되었던 각종 뉴론에 작용 하는 약물들을 투여하여 나타나는 기능적 또는 행동적 변화로 미 루 어 뇌의 특정 활 성 을 추정하는 방법이 있다. 12. 2 In v it ro 에서의 방법 In v itr o 에서 의 방법 으로는 오래 전부터 뇌 절편이 이 용되 었다. 뇌 절편은 뉴론을 주위 조직으로부터 분리하지 않고 비교적 생리 적 조건에 가까운 환경에서 실험한다는 이점이 있는 표본이다. 때로는 뉴론의 주행을 따라 절편을 만들어야 한다는 주장도 있었 으나 최근에는 0.5mm3 정도의 블록 모양의 절편이 자주 이용된 다. 이같은 절편에서는 뉴론이 부분적으로 절단되는 표본이므로 in v i vo 에서의 기능을 어느 정도나 반영할 것이냐는 문제가 있기 는 하나 비교적 일정한 실험성적을 얻을 수 있고 in vi vo 의 실험 성적과 평행하므로 흔히 이용된다 . 뇌절편을 포함하여 in vitro 에서의 실험은 조작이 간단하고, 영양액이나 실험약물의 농도를 자유롭게 변화시킬 수 있고, 뇌조직으로부터의 유리물질(전달물 질, 대사물, 효소 등)의 양적 변동을 정확하게 잡아낼 수 있고 뇌-혈액장벽 (bl ocxi -br ain b arri er) 을 고려하지 않아도 되고, 말초 장기에 대한 영향을 고려하지 않아도 되는 등의 이점을 들 수 있다.
특정 뉴론이나 그리아세포를 분리하여 실험에 이용할 수도 있다. 이것은 직접 뇌조직으로부터 목적하는 세포를 분리하는 방법과 분리세포의 모양으로 목적하는 특정 세포집단을 뇌조직에 서 모으는 방법이 있다. 전자는 입체현미경에서 단일 뉴론을 분리 해내는 방법이고 후자는 세포들간의 밀도차를 이용하여 밀도차 원심분리법으로 분리하는 방법이다(그림 3-25) . 배양세포를 이용하 는 방법도 있다.
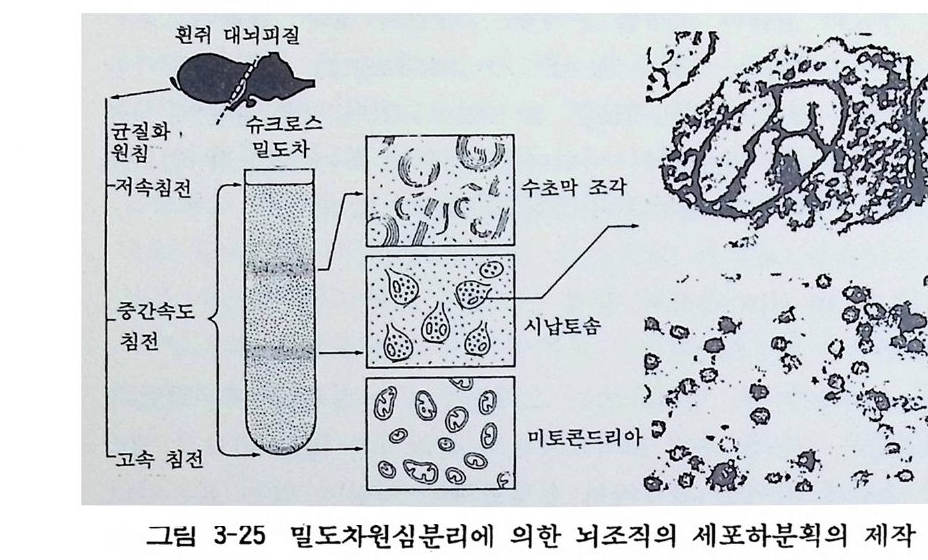 - . 흰쥐 대뇌피질 .- Z 도 ’ - ... ~-. ? 、 _ . 츠 ·
- . 흰쥐 대뇌피질 .- Z 도 ’ - ... ~-. ? 、 _ . 츠 ·
원심분리법은 세포분획의 분리에도 이용된다. 뇌 세포를 일정조 건에서 균질화 (homo g e nize)하고 분획 입자의 크기와 밀도차 를 이용하여 초원심분리로 각 분획을 분리하는 방법이다. 이같은 방법으로 세포핵, 마이에린, 신경말단부(시납토좀 s y na pt osome), 마이크로좀, 접합부소포, 라이소좀, 리보좀, 미토콘드리아 등이 분리된다. 이중 시납토좀은 신경말단부가 축색이나 그리아세포로 부터 분리되어 만들어지는 과립으로서 in vi vo 에 있어서의 신경말 단부(접합부)의 생화학적 여러 성질을 비교적 안정하게 유지하고 있는 것이다. 따라서 in vitro 실험에 흔히 이용된다. 특히 접합부 의 기능 연구에 없어서는 안 될 표본이라 하겠다. 또 이 표본을
저장액으로 처리하면 내부에 함유되어 있던 접합부소포나 수용성 분획을 분리할 수 있다 . 또 이 표본에는 접합후막도 포함되므로 수용체 분리에도 이용된다. 12. 3 분자생물학적 방법 최근의 분자생물학적 연구 기법의 발전은 획기적이다. 이제까지 의 펩타이드 발견과 구조결정 연구는 대량의 조직 장기로부터 각종의 정제 방법을 조합하여 펩타이드를 순수 분리하는 방법이 시행되었으나 최근에는 전혀 새로운 방법으로 핵산에 초점을 맞춘 유전공학적 방법이 각광을 받고 있다• 이 방법을 간단하게 설명하면 소량의 조직으로부터 rnRNA 를 추출하고 이 rnRNA 또는 이 mRNA 에 대하여 역전사하여 만들어진 DNA(cDNA) 를 프라스미드에 삽입시켜 대장균 등의 세포내로 도입한다. 이 cDNA 를 전사하여 mRNA 를 만드는 세포집락 (colon y)을 뽑아내 어 이 세포를 배양함으로써 대량의 rnRNA 를 만들고 그 염기 배열을 쉽게 규명할 수 있게 된다. 이것으로부터 대응하는 아미노 산 배열을 규명할 수도 있다. 단백질 합성 기전을 교묘히 이용하 여 소량의 출발 조직에서 비교적 간단하고 신속하게 새로운 펩타 이드 를 발견하고 그 구조를 결정할 수 있는 방법이다• 이 방법은 이제부터 새로운 생리활성 펩타이드의 발견, 합성, 분해의 연구에 큰 역할을 할 것으로 기대된다(그림 3-26).
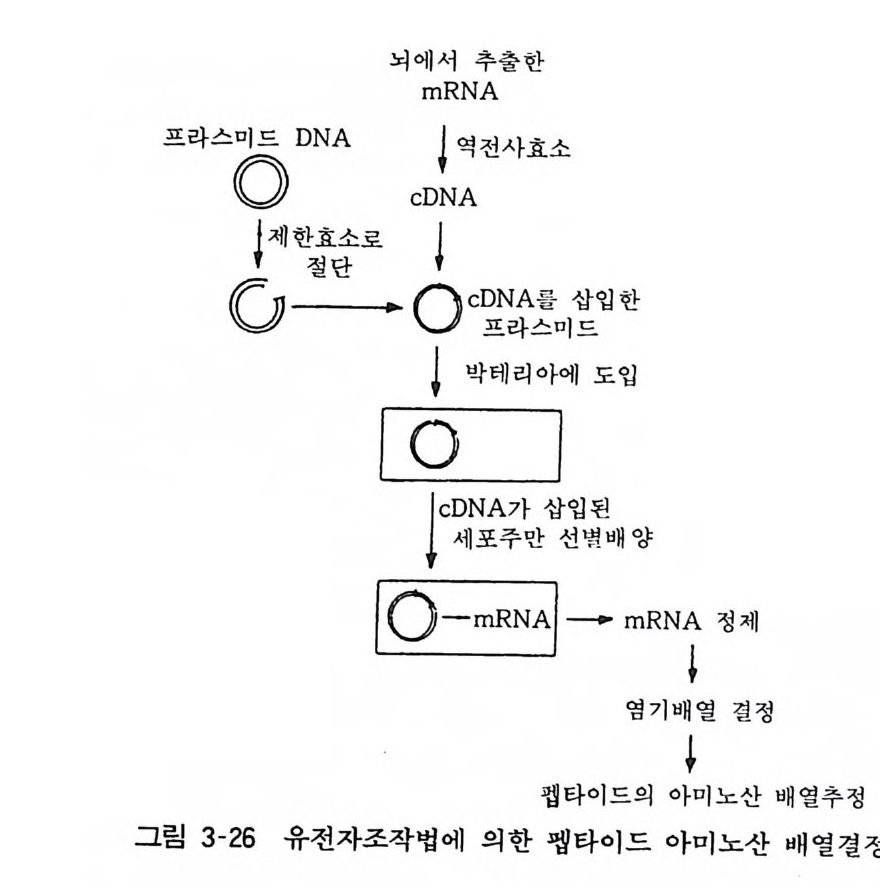 뇌에m서R N추A출 한
뇌에m서R N추A출 한
제 4 장 신경전달물질 수용체 방사능표식 리간드를 세포막분획에 결합시켜 수용체를 표식하 는 소위 수용체결합법의 확립으로 신경전달물질 수용체, 약물 수용체의 분자 수준의 연구가 최근 10 년 동안에 큰 진전을 보았 다. 이 방법으로 리간드의 수용체에 대한 친화성, 수용체의 밀 도, 협동성(수용체 분자간의 상호작용) 등을 정량적으로 분석할 수 있게 되었다. 나아가 병적 상태의 수용체, 약물 연속투여 후 수용체의 분자수준의 변화 유무 등을 알아볼 수 있게 되었다. l 수용체의 분자량 측정 원심분리법은 단백질 분자량 측정에 널리 이용되어 왔다. 이 중 최근에 개발된 공기작동 초원심분리법 (air drive n ultra c ent- rifug e) 은 세포막 단백의 성질과 이것의 반응액내에서의 상호작 용 연구에 유용하게 이용되는 것이다. 두 가지 이용법이 있다.
하나는 단백질과 작은 분자간의 결합반응을 분석하는 것이고 또 한 가지는 분자량을 측정하기 위한 침강평형법이다. 후자는 대단히 민감한 방법으로 125I 표식단백을 이용하여 나노그람 정도 의 시료로도 분자량을 측정할 수 있다. 수용체결합법에 있어 리간 드의 결합은 물리적인 것으로 가역적이고 쉽게 해리된다. 이같은 성질을 이용하여 수용체의 분자적 여러 성질을 자세하게 알아 보기 위하여 친화성 표식 (a ffinity label) 을 이용한 광친화성표식 법(p ho t oa ffinity labe ling)이 이용된다(그림 4-1). 즉 반응성이 큰
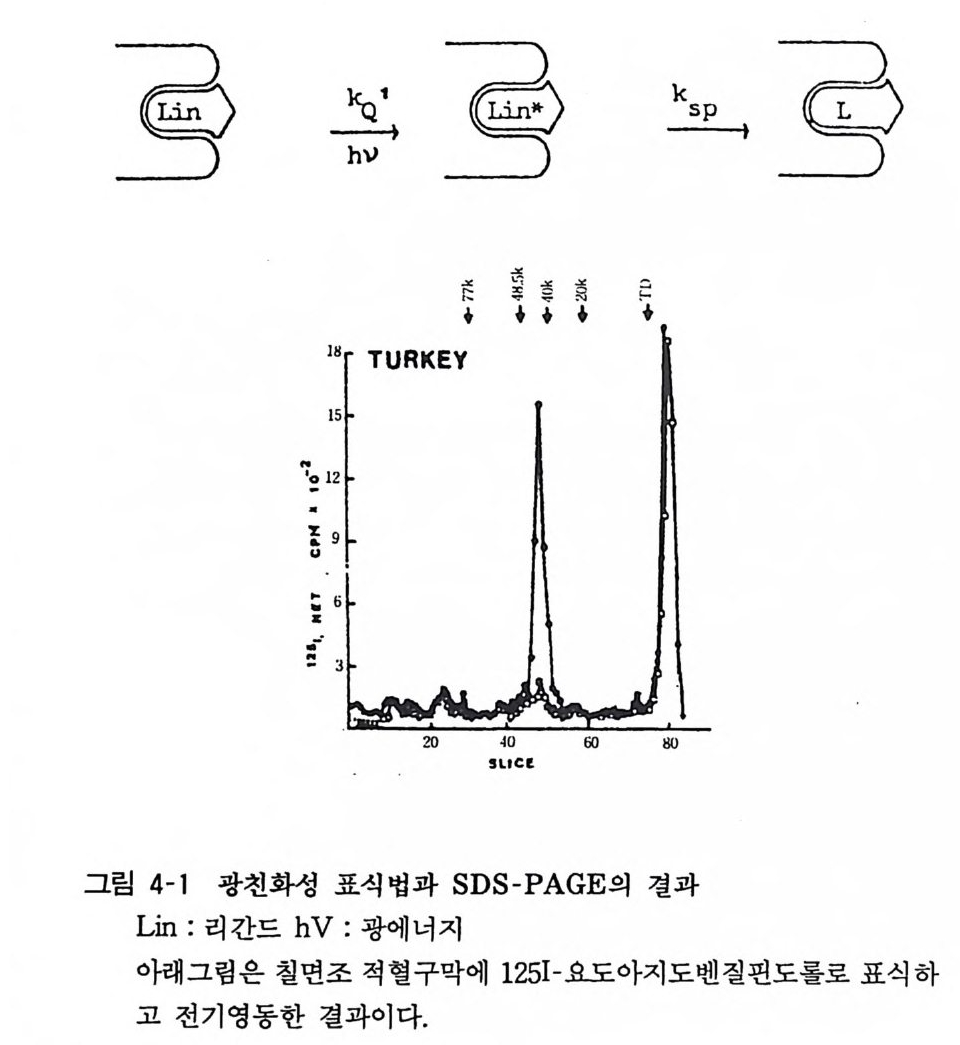 글18 .. 릅TURKEYf‘ 己三준‘ 호* 칭‘ 已 * k sp一 己〕
글18 .. 릅TURKEYf‘ 己三준‘ 호* 칭‘ 已 * k sp一 己〕
반응기를 갖는 리간드(방사성 동위원소로 표식해 놓는다)를 세포 막분획과 반응시키고 자외선 또는 레이저 광선을 쪼여 리간드와 수용체 사이에 안정적인 공유결합을 이루도록 하고 방사능 활성 을 이용하여 수용체 분자의 여러 성질을 조사하는 것이다. 일반적 으로는 세포막 분획을 용액화하고 SDS 포리아크릴아마이드겔 전기영동 (PAGE) 에 걸어 분자량을 추정하는 방법이 흔히 이용된 다(그림 4-2). 수용체의 분자구조(서브유닐 등), 기능(이온통로
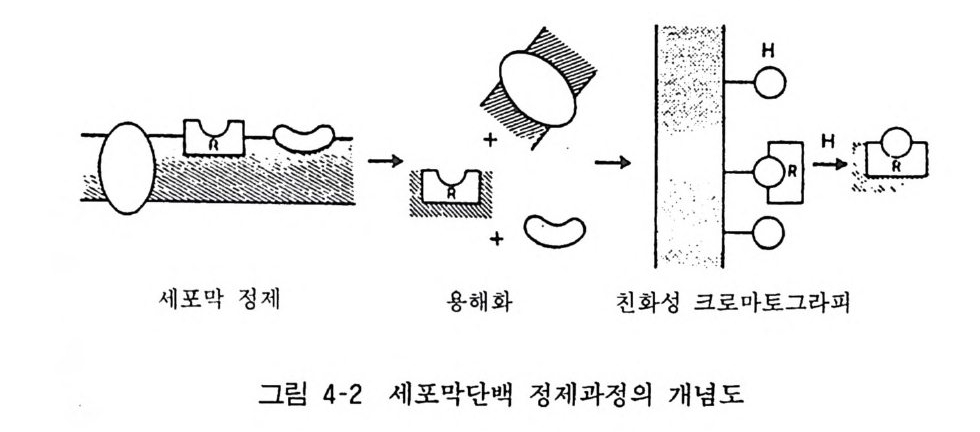 기 :노 :@
기 :노 :@
등)과의 연관성도 추정할 수 있다. 현재 니코틴성 아세틸코린 수용체 와 떤-트리 페 닐메 틸포스포리 움 (3H- trip hen y lme thy l p hos ph ori um ), 무스카린성 아세틸코린 수용체와 3H-N- 메 틸 -4- 피페리 딜 벤 질레 이 트 (3H -N-meth yl- 4-pi peridy lb enz yla te ) , «1-아 드레 나린 수용체와 떤-펜옥시벤자민(3 H- p heno x.y b enz amine ), /J 1- 아드레나 린 수용체 와 125I- 요도아지도벤질핀도를 (1251-io d oa zido benz ylp i nd - olol) 둥에 의한 결과가 보고되어 있다(표 4-1). Vente r 동은 수용체 크기를 추정하는 데에 방사선 조사/실활법 (aed i a ti on in ac ti va ti on) (표적 크기 분석법)을 사용하여 뇌를 비롯한 각종 장기세포의 분자 크기를 결정하고 있다. 이 방법의 원리는 수용체 에 방사선을 조사하면 수용체의 결합활성을 잃게 되는데 수용체 의 분자량이 클수록 활성을 잃는 정도가 크다고 하는 사실에 의한 것으로 대량의 각종 선량을 세포막분획에 조사하고 선량-
실활 관계의 직선의 기울기로부터 기지 분자량 의 그것과 비교하 여 수용체의 분자량을 추정하는 것이다. 이것의 특 징은 수용체를 용액화하거나 분리 정제할 필요가 없이 세포막분획자체에 방사선 을 조사하여 간단한 조작으로 분자량을 추정할 수 있다는 이점이 있다. 한편 대량의 방사선원을 필요로 하며 이 같 은 장치는 쉽게 얻어지는 것은 아니다. 그러나 이 방법으로 분자량을 측정함으로 써 막내에서의 수용체 존재 상태, 즉 단량체인지 2 량체인지 를 알아볼 수 있다.
표 4-1 신경전달물질 수용체의 분자량 무스카린성 aI- 아드레나 수용체 Ach 수용체 린성 수용체 /3 1 - 수용체 /3 2 · 수용체 방사선조사 80,000 160,000 90,000 109,000 /실활법 (흰쥐 뇌 등) (흰쥐 간) (칠면조 적혈구) (게 폐) SDS- 80,0 0 0 85,000 70,000 58,000 PAGE (흰쥐, 사람. 개뇌) (흰쥐 간) ( 칠면조 적혈구) (서브유닐) 초원심법 (8흰6쥐,0 0뇌0 ) (흰83쥐,0 00간 ) (개65 ,0심0장0 ) (개9 0간,0,0 0폐 ) 중수합용체상의태 단량체 이량체 단량체 이량제
2 단크론 항체 (Monoclonal an ti bod y)의 수용체 연구에의 응용 또 한 가지 수용체 연구의 분자 수준 접근 방법으로 수용체에 대한 단크론 항체를 생성하고 타수용체와 면역학적으로 교차 반응이 있는지를 조사하여 수용체 분자의 유사성을 추정하는 방법이 벤터 둥에 의하여 행하여졌다. 프레이저 (Fraser) 와 벤터 (Vente r , 1980) 는 부분적으로 정제한 칠면조 적혈구 /3 1- 수용체에
대한 단크론항체 를 만들었다. 이 항체는 칠면조 적혈구 /3-수용 체에 대한 리간드 결합을 억제하였을 뿐 아니라 기타 여러 조직 의 /3 - 수용체에 대한 리간드 결합도 억제하였다. 이것은 /3-수용체 결합 부위에 리간드들과 항체에 대한 공통적인 결정기가 존재합 울 뜻하는 것이다. 그러나 일반적으로 항원은 순수하지 않으므로 설혹 한 종류의 항원이라도 항원 결정기는 복수이므로 단일의 항원 결정기에 대한 항체 분자만을 얻기는 힘들다. 그리고 무스카 린성 Ach 수용체와 a:1- AD 수용체의 유사성, /31 -, /3 2 - 수용체 사 이, 또 D2- 수용체와 5-H T 수용체 사이에 유사성이 있는 것 같고 이는 수용체의 분자진화라는 관점에서 홍미있는 사실이다. 여기서 사용하고 있는 단크론 항체는 신경 연구에서뿐 아니라 의학 생물학 연구에서 널리 이용되는 유용성이 큰 항체로서 여기 에서는 간략하게 설명하기로 한다(그림 4-3 ) .
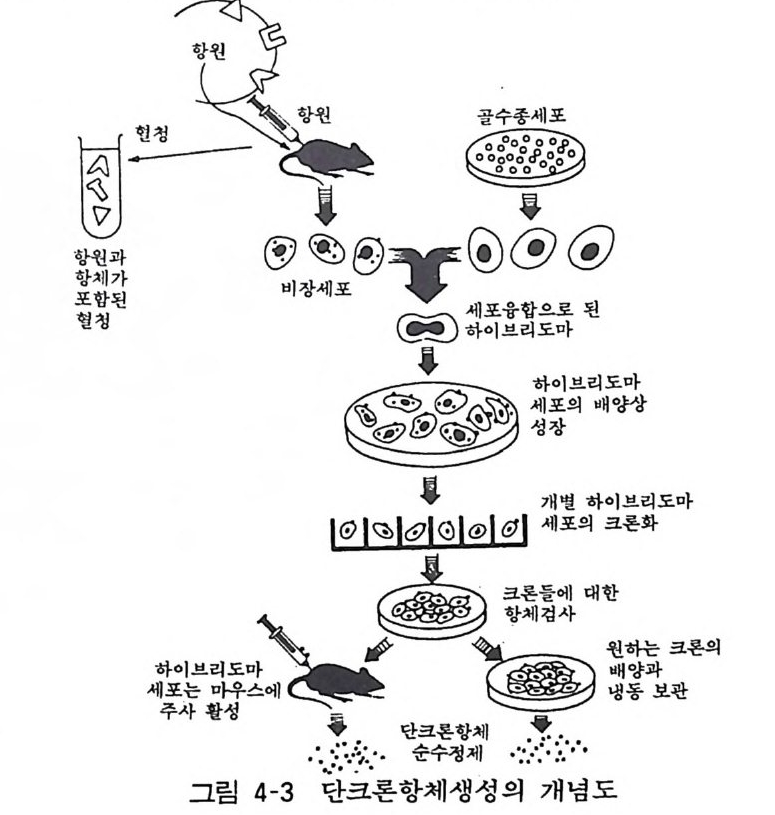 麟포현 함청: 된 언 5포 @g T@;‘t- ; 하세~軍 이포브융~의리합도으-마로배 ~된 ° EJ -
麟포현 함청: 된 언 5포 @g T@;‘t- ; 하세~軍 이포브융~의리합도으-마로배 ~된 ° EJ -
항원이 생체내에 들어오면 그 항원에 대하여 특이적인 림프구 (크론)가 자극되어 항체가 합성, 분비된다. 골수종 세포 (m y eloma cell) 는 암화된 형질세포로 단일 크론의 항체 를 합성 분비하고 증식을 계속할 수 있다. 이 골수종 세포에 항원으로 자극된 림프 구 (B 세포)를 세포융합시켜 림프구에서 유래하는 항체를 합성 분비하는 세포주를 얻을 수 있는 것이다. 이같은 방법으로 단일 항원 결정기에 대응하는 항체를 다량 얻을 수 있게 된다. 실제로 는 그림에 나타난 것처럼 항원을 주사한 마우스의 비장 림프구와 골수종 세포를 포리에티렌그라이콜로 세포융합시켜 배양하고 그 중에서 하이브리도마(잡종세포)만을 골라내어 이것을 다시 배양하여 목적하는 항체를 생산하는 하이브리도마 를 골 라내어 하이브리도마 크론을 확립하고 다시 반복 배양하여 몇 대에 걸쳐 강하고 확실한 단크론 항체를 생성하는 하이브리도마를 얻는다. 이것을 대량 배양하여 서로 다른 수용체간의 상동성 , 면역침전법 또는 친화성 칼람에 의한 수용체의 분리정제, 세포 또는 조직에 있어 수용체의 분포 등 수용체 연구에 획기적 진전을 가져왔다. 3 아세틸코린 수용체의 분자구조 수용체 분자의 여러 성질이 가장 잘 알려진 것은 무어라 해도 니코틴성 아세틸코린 수용체라 하겠다. 니코틴성 아세틸코린 수용 체는 골격근에 존재하는 것으로 압도적으로 다량 존재하는 곳은 전기뱀장어 (To rpedo calif o rni ca) 의 발전기관이고 이것의 수용체 가 가장 손쉬운 연구의 대상이므로 연구가 잘 되어 있다. 이 전기 • 기관을 계면활성제를 사용하여 용해하고 이를 아피니티칼람크로 마토그라피에 의하여 정제한다(그림 4-4). 이것의 분자량은 25 만인 당단백이고 a, /3, Y, 6 4 종의 서브타입으로 되어 있고 각각 의 분자량도 결정되어 있다(표 4-2).
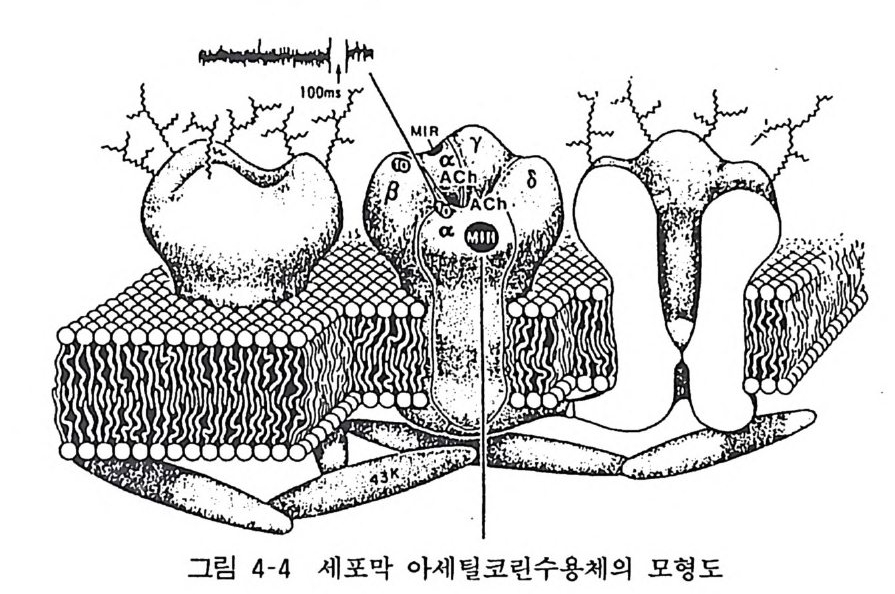 그림 4-4 세포막 아세틸코린수용체의 모형도
그림 4-4 세포막 아세틸코린수용체의 모형도
표 4-2 니코틴성 아세틸코린 수용체의 분자량 서브타입 아미노산수 분자량 a 437 50,116 /3 469 53.681 . Y 489 56,275 8 501 57,565
두 개의 a- 서브유닐 위에 아세틸코린 결합 부위가 있다(부근에 S-S 결합이 있다). 아세틸코린 결합에 의하여 % g, Y, 8 서브유 닐으로 둘러싸인 이온통로가 열리고 N 하 이온이 세포내로 유입됨 으로써 세포가 흥분하게 된다. 현재 a- 서브유닐에 아세틸코린이 결합하면 어떻게 /3, r, cS 서브유닐에 영향을 미치고 이온통로를 열리 게 하는지에 대하여는 상세히 밝혀져 있지 않다(그림 4-5). 최근 각 서브유닐에 대한 DNA(cDNA) 가 크로닝되어 그 염기 배열로부터 아미노산 조성이 결정되었다. 이 결과에 의하면 각 서브유닐의 cDNA 를 아프리카 개구리 난세포에 도입하면 각 서브 유닐이 생합성되고 이것을 세포막에 함입시키면 막 표면에 나타
나며 이것에 아세틸코린을 작용시키면 세포막의 탈분극이 일어남 을 관찰하였다. 이것은 각 서브유님이 핵산으로부터 생합성되어 완전한 니코틴성 아세틸코린 수용체의 기능을 세포막에 나타냄을 보여준 것이다. a, f3, % 8 서브유닐 중 어느 하나라도 결여되면 수용체는 불안정하고 8 서브유닐은 없더라도 아세틸코린에 대한 반응성은 있고 8 서브유닐은 어느 정도 Y - 서브유닐으로 대치될 수 있다는 것도 알려졌다. 외래성의 유전정보에 따르는 효율적인 기능단백의 합성과 그 기능의 발현기전에 관한 연구에 아프리카 개구리의 난세포가 유용하게 이용되는 것으로 금후 이 세포 를 이용한 연구가 크게 전개될 것으로 생각되며 바나드 (Barnard) 둥은 GABA/ 벤조디아제핀 수용체 생합성과 세포내 기능발현 양상 연구에 이 세포를 이용하곡 있으며 최근 사람의 mRNA 로부 터 기능을 가진 N 하 이온통로를 생합성하였다고 한다. 사람의 골격근 니코틴성 아세틸코린 수용체도 정제되어 a, f3, Y, 8 서브 유님으로 이루어지고 a- 서브유님이 아세틸코린 결합 부위이고 전기기관의 a- 서브유닐 아미노산 조성과 매우 높은 상동성을 보인다라는 것을 알았고 이는 중증근무력증의 발현기전과 그 치료에 크게 기여할 중요한 연구로서 기대된다.
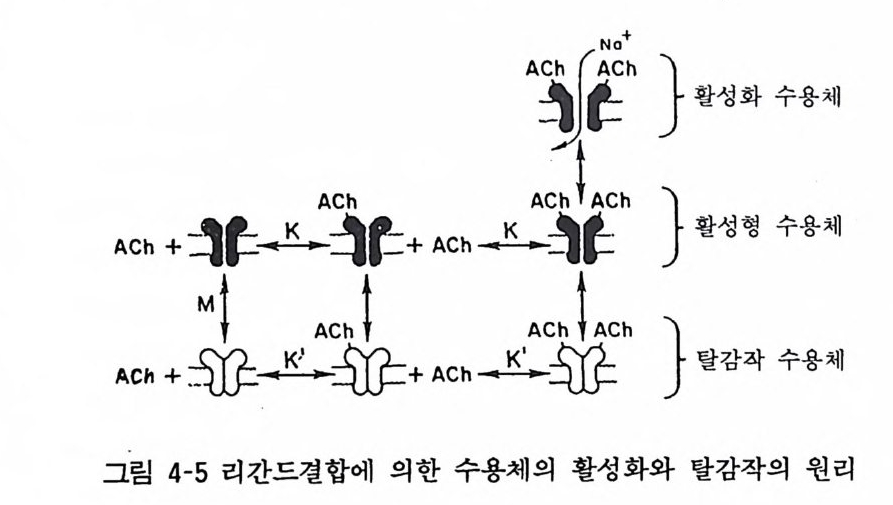 A갑 }紅화수용체
A갑 }紅화수용체
4 수용체의 합성과 대사 신경전달물질을 인식하고 결합시켜 특정 생체반응을 유도하는 수용체는 세포막에 존재하는 것이나 어떻게 세포내에서 합성되고 분자적인 수식을 받아 운반되어 세포막에 조립되는가 하는 것은 중요한 연구과제이다. 특히 어떤 효능제에 의하여 세포나 조직이 장기간 노출되면 그 효능제에 대한 수용체의 감수성이 저하하는 탈감작 (desens itizati on) 현상이 잘 알려져 있다. 그러나 그 기전에 관해서는 잘 모른다. 탈감작 현상은 1) 수용체의 효능제 결합부위가 세포막내 또는 세포내로 묻혀 버린다. 2) 수용체 분자의 합성이 저하한다. 3) 수용체의 세포내 대사가 촉진된다. 둥의 가능성을 생각할 수 있다. 이 점에 관하여 골격근의 근-신경 접합부에서 니코틴성 아세틸코린 수용체에 관한 연구가 이루어져 있다.
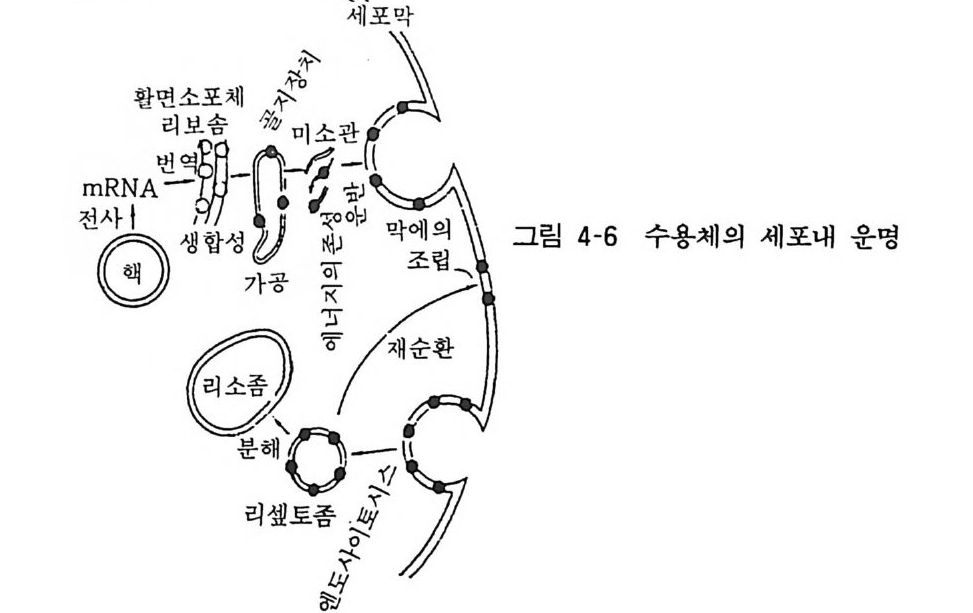 홍 그림 4 6 습 체의 세 포 내 운 명
홍 그림 4 6 습 체의 세 포 내 운 명
수용체는 일반적인 단백질과 마찬가지로 활면리보좀 (smoo t h ri bosome) 에서 합성되어 소포체막을 통과할 때 당쇄가 첨가되고 골지기구에서 a, fl, y, o 의 서브유닐이 조립된다. 이 것은 에너지 를 사용하는 과정으로 미소관에 의해 세포내 를 이동하여 세포 막 에 조립되면서 막 표면에 나타나게 된다. 한편 막 표면의 수용체는 항상 엔도사이토시스 (endoc ytos i s) 에 의하여 세포내로 들 어가 소포(리셉토좀이라 함)가 되어 리소좀 (l y sosome) 에 의하여 분해 된다(그림 4-6). 막의 수용체는 끊임없이 합성과 분해가 반복되는 동적인 평형상태를 이룬다. 골격근 의 아세틸코린 수용체의 반감기 는 황브로우 (Fambrou g h) 의 연구 보고에 의하면 근- 신경 접합부 에 서는 5-10 일이나 접합부 이외의 부위에서는 20 시간 정도라고 한다. 운동신경을 절제하면 접합부 이외의 부 위에서 아세 틸콜린 수용체 밀도가 증가하며 이것은 새로이 합성된 것으로 대사속도 도 빠르다고 한다. 발육단계에서 신경이 근육과 접합부를 형성하 면 이제까지 근육 여기저기에 산재해 있던 수용체가 접합부로 집합한다고 한다. 이같은 수용체의 분포에 대해서 신경이 어떤 기전으로 영향을 미치는 것인지는 아직 모른다. 수용체 를 집합시 키는 인자가 신경으로부터 분비되는지도 모르겠다. 앞으로의 홍미 있는 연구과제의 하나이다. 아세틸코린 수용체 이외에도 베타아 드 레나린 수용체의탈감작시 숫적인 감소 (down re gul a ti on) 를 가져 오는 기전도 적혈구막을 사용하여 레프코비츠 (Le fk ow itz) 등이 연구 보고하고 있다. 수용체가 세포내로 함입되면(i n t erna lizati on), 일부는 리소좀에 의해 파괴되고 일부는 재차 세포막에 끼어들어 (recyc lin g) 재이용된다고 한다. 탈감작시에도 수용체는 세포내로 함입되나 수용체와 함께 역할을 나타내는 인자인 GTP 결합조절 단백, 아데닐사이크라제 촉매 유닐온 그대로 세포막에 잔존한다는 사실은 홍미있는 것이다. 신경세포막에서의 기타 수용체들의 탈감 작시에도 수용체의 감소가 같은 기전으로 일어날 것인지는 앞으 로 밝혀져야 할 것이다. 베타아드레나린 수용체가 축색내 미소관 에 의해 운반되어 신경말단에 도달하는 것도 알려져 있다.
5 이온통로 단백의 분자적 성질 신 경 세포막 을 경계로 한 Na+, K+ , Ca ++ 의 이동은 신경홍분, 신경전 달물 질의 유리에 중요한 관련이 있다. 이 들 이온통로 단백 은 최 근 캇테랄 (Ca tt erall) 등에 의하여 흰쥐의 뇌에서 분 리 정제 되어 그 분 자구조가 밝혀졌다. N 하 이온통로 단백은 3H- 삭시톡신 (Sax it o xin)의 결합 활 성을 지 표 로 하여 용액화 정제되고 , 분자량 3 1. 6 만 의 당단백으로 a - ( 분자 량 27 만), /3 1-(3 . 9 만), /3 2 -(3.7 만) 의 서 브 유님으로 되어 있고(그림 4- 7 ), 이 단백질을 레시틴 (lecit hin) 등 인지 질 의 인공막(lipo some) 에 함입시키면 신경독에 반응하여 N 하가 유입하 는 현 상 을 볼 수 있었다고 한다.
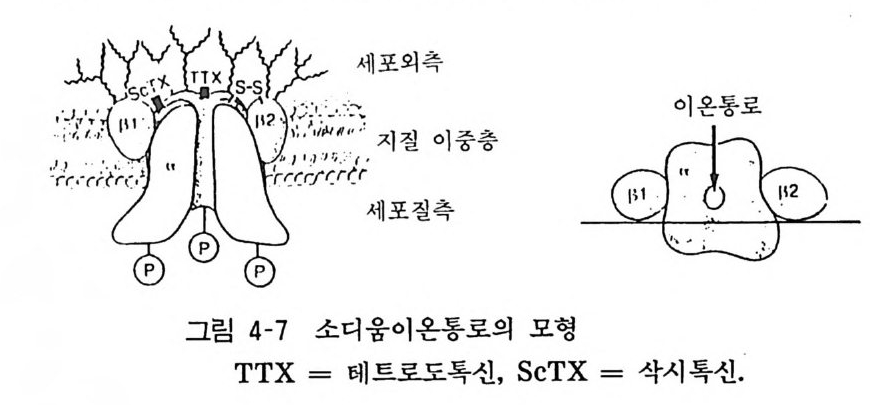 세포 외 측
세포 외 측
이같이 정제한 막내 기능단백(수용체 동)을 인공막에 함입시켜 재구성막 (recons tit u t ed membrane) 을 만들어 기능을 재현토록 하는 연구방법(그림 4 - 8) 은 중요한 것으로 수용체-효과기계 연구 롤 단순화한 실험계이다. 또 캇테랄 (Ca tt erall) 등은 Ca++ 이온통 로 단백도 정제하여 분자량이 21-27.8 만이고 a- (1 3 만), /J -(5 만), r-(3.3 만)의 서브유닐으로 되 어 있음을 밝혔다. 이들 각 서브유닐 의 단크론 항체를 얻을 수 있으면 각 서브유닐의 기능과 서브유 닐 사이의 상호작용도 밝혀질 것으로 생각된다 . 현재 Ca++ 이온통 로는 탈분극에 관련하여 기능을 나타내는 것과 수용체(아마도
«2- 아드레나린수용체)와 관련하여 기능을 나타내는 적어도 두 종류가 존재할 것으로 생각되고 있다. 이들 두 종류의 분자 수준 의 차이, 그 기능의 차이 등도 이같은 접근방법으로 멀지 않아 밝혀질 것이다. 특히 최근 Ca++ 길항제라고 하는 것으로 혈압강하 제, 항협심층약으로 쓰이는 베라파밀 (Vera p am i l) 을 위시하여 니페디핀 (N ifedipin), 딜티아젭 (D ilti azem) 동이 개발되어 이들 약물의 혈관 평활근, 심근 동에서의 작용기전의 규명을 위해서도 Ca++ 이온통로의 분자 수준의 접근방법은 중요한 과제이다.
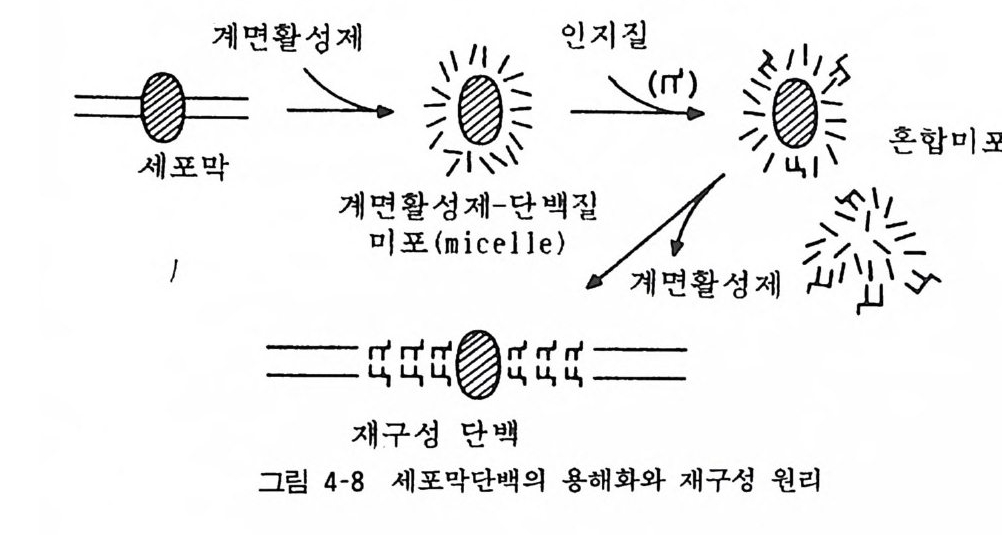 텔세포一막계 면활圈성제멸 二麟7問1\隊\정' /인지혼4질계 면l활합성~ h l제Il i~미,l L\ J\맙\堯 .포:: r
텔세포一막계 면활圈성제멸 二麟7問1\隊\정' /인지혼4질계 면l활합성~ h l제Il i~미,l L\ J\맙\堯 .포:: r
6 자극도입 (Im p ul se tra nsducti on ) 이제까지 수용체 자체의 성질에 대하여 알아보았다. 세포막에 존재하는 수용체가 약물 등의 외래 자극으로 자극되었을 때 세포 막과 세포내에서의 일련의 과정을 거쳐 세포기능의 변화를 일으 키게 되는 것으로 이같은 일련의 과정은 이온통로를 통한 이온의 이동을 비롯하여 소위 제 2 전달체 (second messen g er) 라고 하는 cAMP, 포스파티딜이노시톨 및 칼슘이온 동에 의하여 수용체에서
받아들인 정보를 세포내로 도입 (impu lse t ransduc ti on) 하게 되는 것이다(그림 4-9).
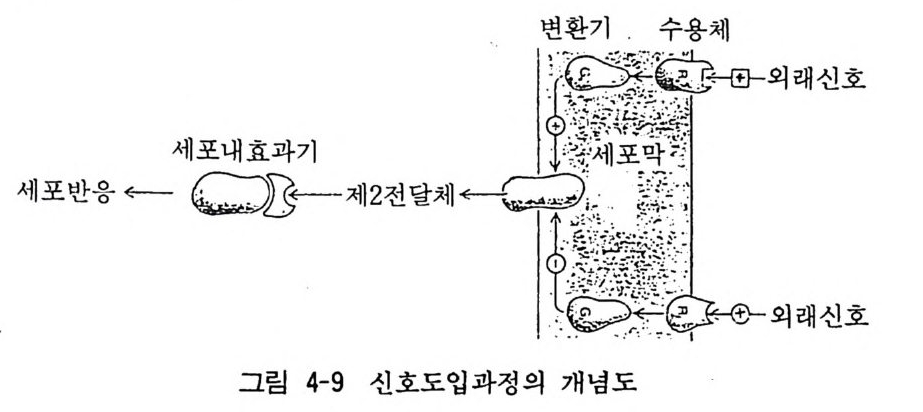 I 변, · 환?_ 기` 후 .후 수 >용체
I 변, · 환?_ 기` 후 .후 수 >용체
6. 1 인지질 (Phos p hol ipi d) 의 메틸화 반응 수용체가 세포막내에 존재하여 세포막 표면에서 효능제를 인식 하고 결합하면 이온통로가 열리거나 효소가 활성화되어 생체반옹 을 유도하게 되나 이 수용체는 단순히 세포막내에 고정되어 정적 인 상태로 있는 것은 아니라고 생각한다. 지질 이중층 사이룰 이동한다거나 막 내부로 함몰되거나 막 표면으로 튀어오르거나 하는 둥 동적인 상태에 있다• 이같은 막의 유동성의 크기가 수용 체뿐 아니라 막내에 존재하는 각종 효소나 기능인자의 활성 발현 과 관계가 깊다는 것은 쉽게 생각할 수 있다. 히라다(Hir a ta) 등은 적혈구막의 베타 수용체에 효능제가 결합 하면 막내의 인지질이 메틸화하여 막 유동성이 커지고 아데닐레 이트 사이크라제가 활성화된다는 것을 보고하였다. 막에 존재하는 두 종류의 인지 질 메 틸전 이 효소(p hos p ho lipi d meth ylt ra nsf e ra se) I 과 II 에 의해 S- 아데노실메티오닌의 메틸기가 포스파티딜에타 놀아민에 전이되어 포스파티딜 N- 모노메틸에타놀아민과포스파티 딜코린이 된다. 인지질의 메틸화에 의해 막내의 미세환경에서의 유동성이 증가하여 베타아드레나린 수용체와 GTP 결합조절단백
과 아데닐레이트 사이크라제의 촉매 부위가 상호작용하여 활성화 된다고 생각한다(그림 4-10).
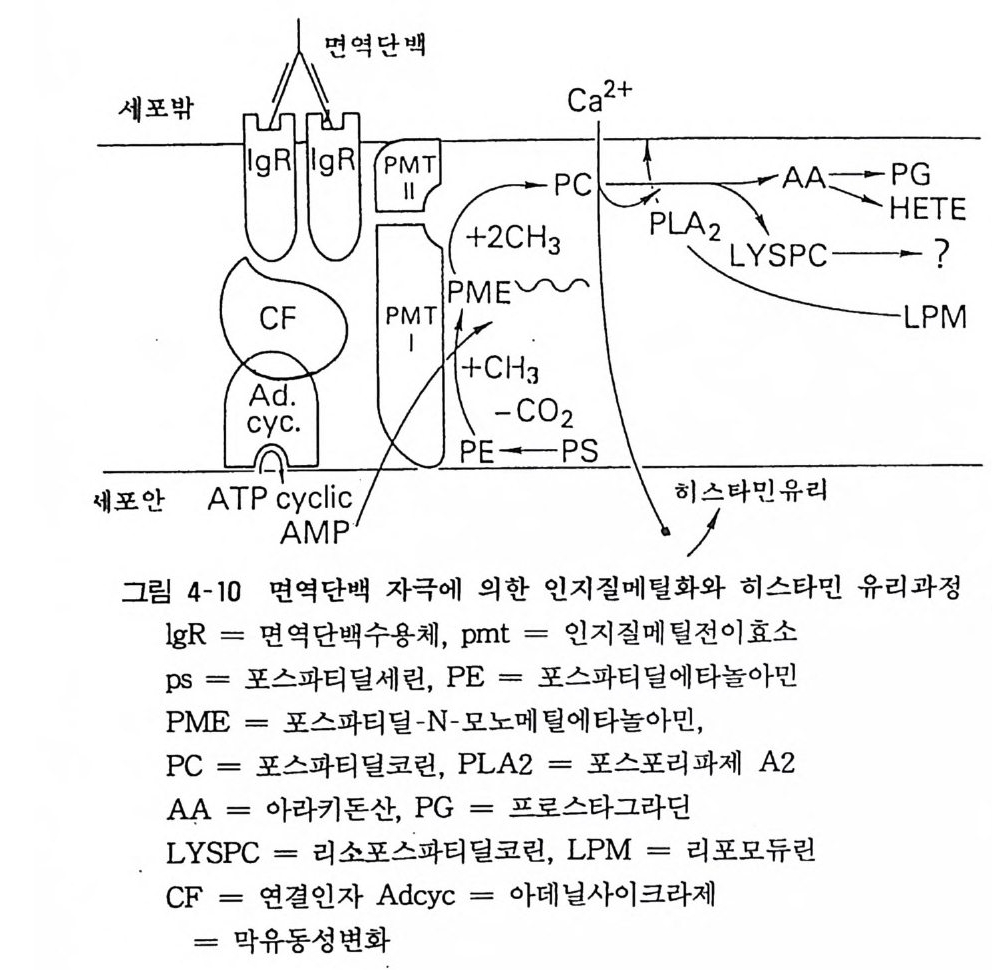 면역단백
면역단백
최근 뇌의 신경세포막에서도 신경전달물질이 수용체에 작용하 여 이같은 반응이 일어난다는 것이 알려졌다(도파민, 히스타민). 또 최근 인지질이 메틸화하면 Ca -t+이 세포내로 유입한다는 것도 알려졌다. 막 유동성의 증가가 어떻게 Ca -t+유입을 일으키는지는 찰 모른다. Ca -t+은 세포기능에 있어 중요한 역할을 나타내는 것으 로 앞으로 밝혀져야 할 과제이다.
6. 2 포스파티덜이노시톨의 대사 포스파티딜이노시톨(p hos p ha ti dy l in os it ol) 의 대사경로는 포스 파티딜이노시톨에 포스포리파제 C(p h osph olip a se C) 가 작용하면 디아씰그리세롤(di ac y l g l y cerol) 과 이노시톨 1, 2- 사이크릭포스페 이트가 되고 다음 단계를 거쳐 포스파티딜이노시톨이 재합성된다 (그림 4-11). 세포막에 존재하는 포스포리파제 C 는 아세틸코린 (무스카린성 수용체를 거쳐), 노르아드레나린 («1~ 수용체를 거쳐), 바소프레신, 안지오텐신 등에 의하여 활성화되고 포스파티딜이노
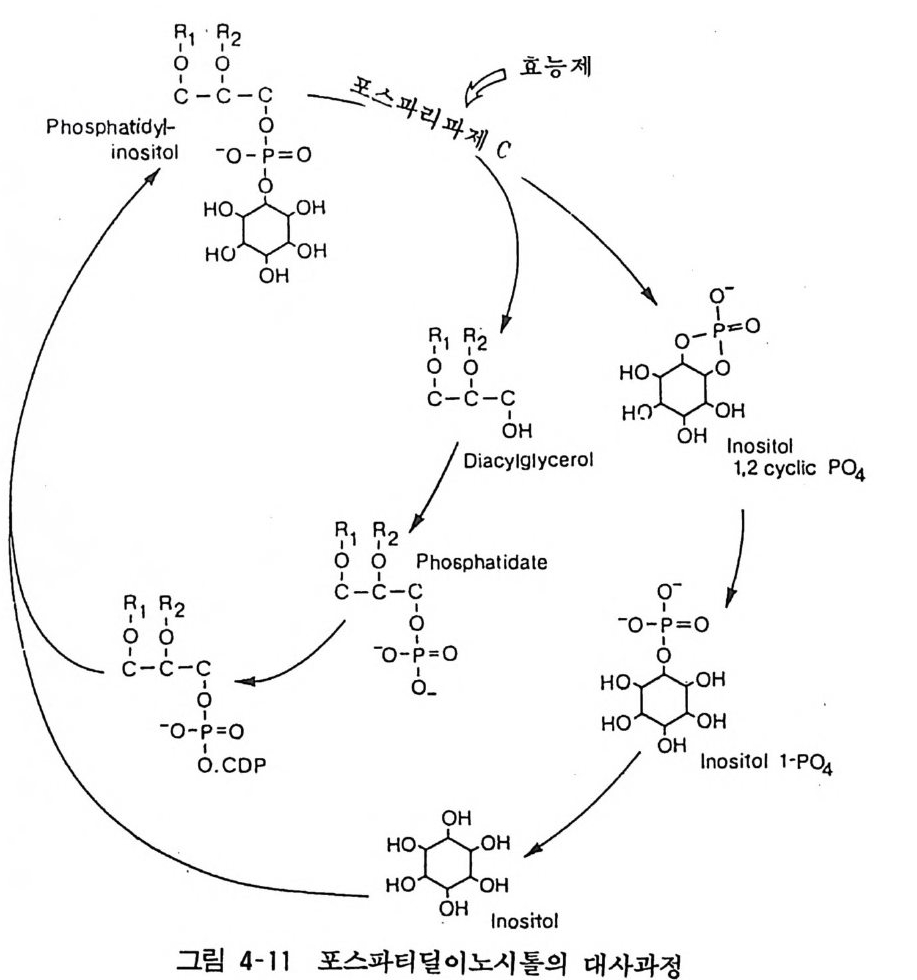 ~1 • 면 2
~1 • 면 2
시톨의 대사회전이 촉진된다. 여기서 포스파티딜산이 Ca ++ 을 세포 밖에서 세포내로 유입시킨다고 생각하고 있다(그림 4-12). 그 자세한 기전은 아직 알려져 있지 않으나 세포내 Ca++ 농도가 증가하는 것은 사실이고 그 Ca++ 이 다채로운 기능을 나타내는 데에 관여한다. 죽 1) 프로스타그란딘 생성 2) C- 키나제의 활성 화 3) 구아닐릭 싸이크라제의 활성화 둥이다. 또 포스파티딜이노 시톨 분해에 의하여 생성되는 산물의 하나인 디아씰그리세롤 (DG) 은 C- 키나제 (C- kin ase) 의 활성화에 중요하다는 사실도 알려
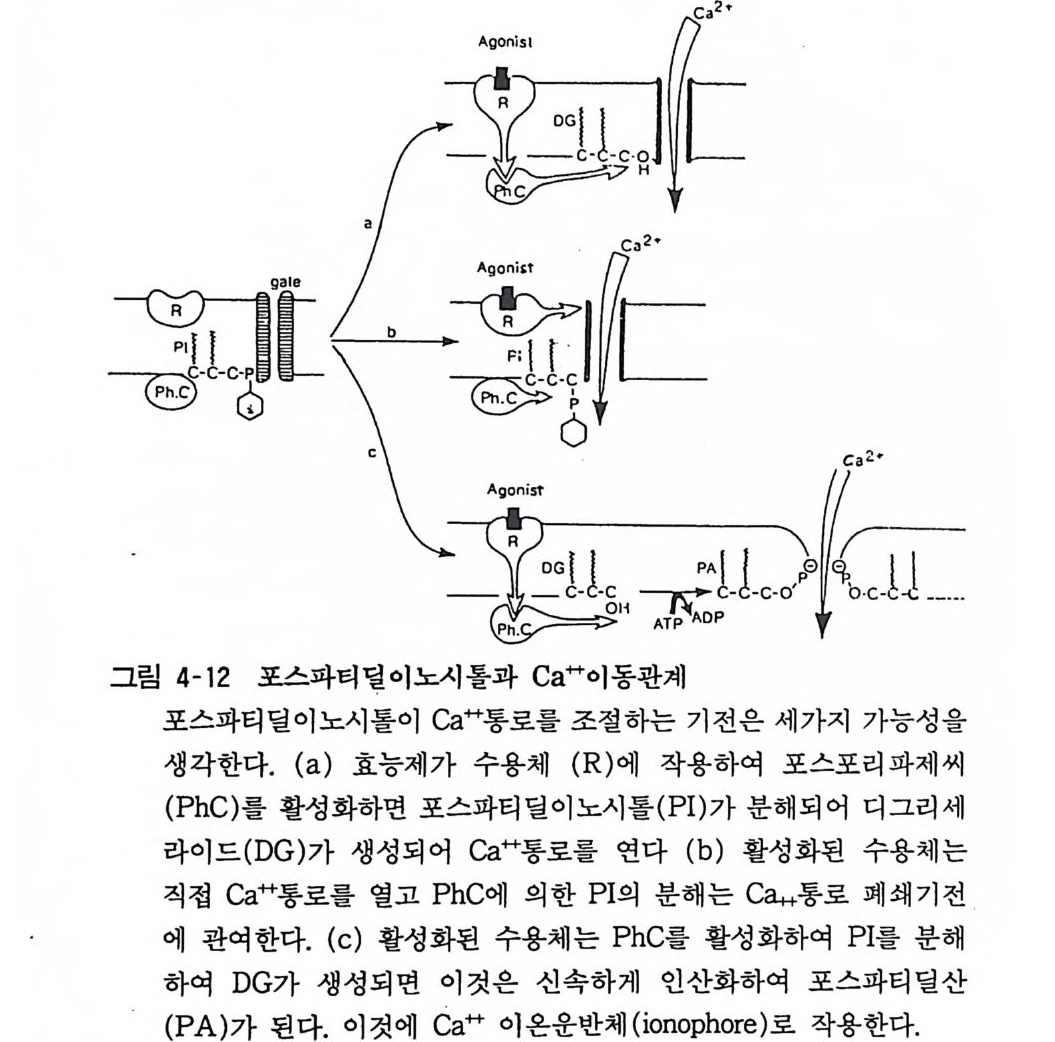 g[
g[
져 있댜 그리고 최근에는 포 스파 티딜이노시 톨 - 3, 4 - 바스포스페이 트 (PIP 기가 분해되어 생기는 이노시톨트리포스페이트 (IP3) 에 의한 세포내 Ca ++ 의 유리작용이 중요시되고 있다 . 이같이 전달물질의 자극에 의하여 포스파티딜이노시톨의 대사가 촉진되고 그 결과 Ca++ 이 세포내로 유입되어 생체반응이 일어나게 된다는 매력적인 생각은 1975 년 밋첼 (M it chell) 이 주장하였으나 지금까지도 자세한 전모는 밝혀지지 않고 있다(그림 4-13). 그러나 수용체를 중개로 하여 인지질 메틸화 그리고 포스포이노시톨 대사회전의 촉진 등 세포막 인지질의 대사가 2 차 전달자 (second messen g er) 일 가능성은 홍 미 를 끄는 사실이나 또 Ca++ 은 포스포리파제 A2 라는 효소를 활 성화하여 아라키돈산을 생성하여 이것이 프로스타그란 딘을 생성한다. 이 포 스포 리파제 A2 는 세포막내에 있는 리포모듀 린(lipo mod ulin)에 의하여 활성이 억제되고 이 리포모듀린의 작용 은 Ca++ 에 의하여 조절된다. 인지질 대사와 Ca++ 의 동태 사이의 관계와 신경계 기능에서의 역 할 은 앞으로의 연구 대상으로 새로 운 전개가 기대된다.
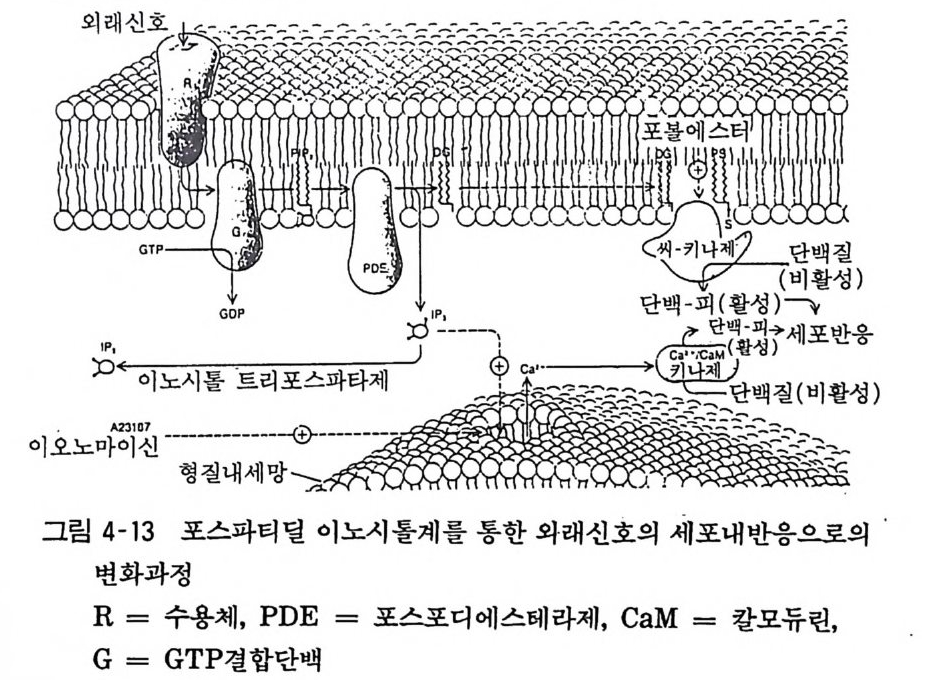 다〉g:전麟)
다〉g:전麟)
7 Ca++ -칼모듀린계 2 차 전달자로서 Ca++이 세포내로 유입되어 각종의 생체반응을 일으키게 할 가능성에 대하여 언급하였으나 자극에 의하여 유리 된 세포내 유리 Ca++ 농도가 세포반응을 조절하는 생화학적 기전 은 극히 최근에 밝혀졌다. 이 경우 중요한 역할을 담당하는 것으 로 칼슘수용단백 (calmod ulin)이 있다. 칼모듀린은 여러 장기조직 세포에 광범위하게 존재하며 다중 기능을 내어(그림 4-14) 세포 내 칼모듀린은 근육의 수축, 세포분열뿐 아니라 신경전달물질의 합성과 유리, 및 수용체 기능에도 관여한다고 한다. 칼모듀린은 분자량 16,000 의 단백질로서 4 개의 Ca++ 결합 부위를 갖는다고 하며 그 아미노산 조성도 밝혀졌다(그림 4-15). Ca++ 이 칼모듀린 에 결합하면 이 복합체가 불활성인 효소에 결합하여 효소를 활성 화한다고 한다(그림 4-16). 이같은 기전으로 활성화되는 효소로서 는 아데닐레이트 싸이크라제와 포스포디에스테라제, 트리프토판과 타이로신의 수산화효소, C- 키나제 등으로 접합전부 또는 후부에 서 칼모듀린이 중요한 조절역할을 할 가능성을 강하게 나타내는 것이다.
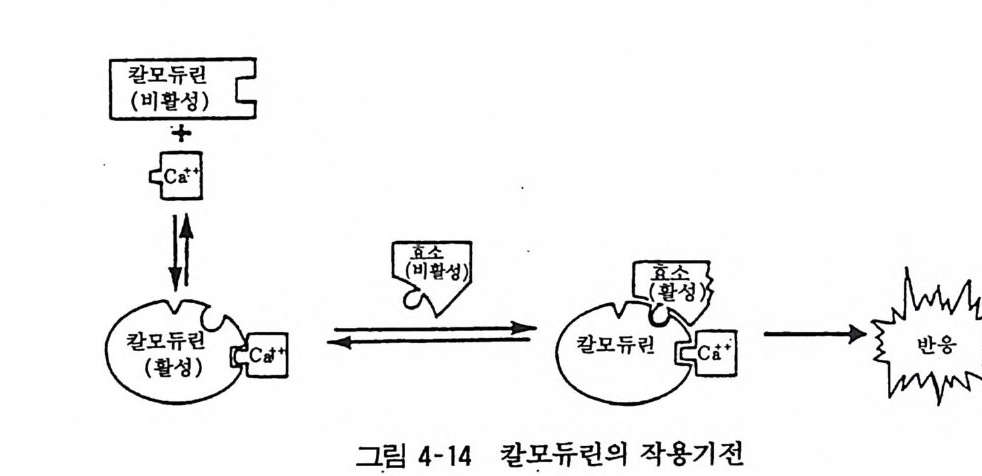 +
+
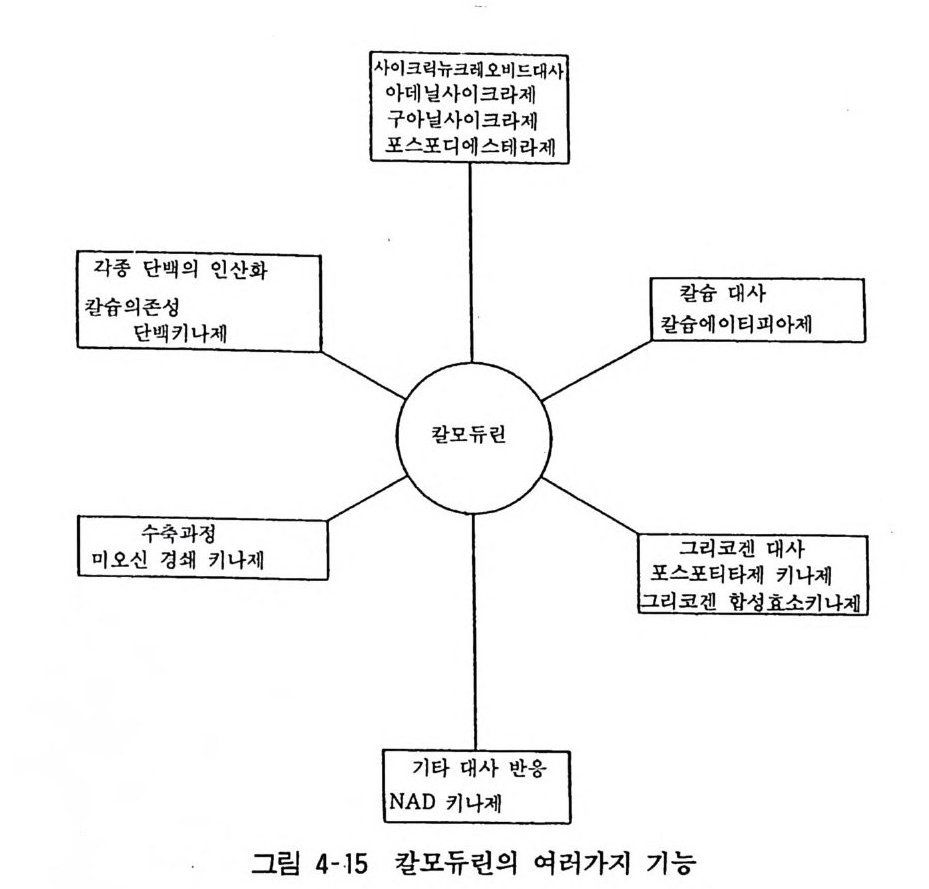 사이크릭뉴크레오비드대사
사이크릭뉴크레오비드대사
8 아데 닐레이트 사이크라제 (Aden y la t e cyc l ase) 수용체에 전달물질이 결합하면 Na+ 둥의 이온통로가 열려 신경 세포가 흥분한다거나 억제된다거나 하는데 신경흥분에 있어 직접 또는 간접적으로 신경세포의 대사면에서 영향을 미치는 것이 아데닐레이트 사이크라제이다. 수용체에서 효과기 (e ff ec t or) 까지 어떤 기전으로 세포막의 정보 가 전달되어 각종의 세포기능을 나타내게 하는가에 대한 연구의 진전은 아데닐레이트 사이크라제와 연계되는 기전에 관한 것이
 rn'm ' o
rn'm ' o
다. 앞서 인지질의 대사와 관련하여 2 차 전달자로서 Ca++ 을 생각 하였으나 여기서 2 차 전달자로서 환형 AMP(cAMP) 가 중요한 것이다. ATP 에서 cAMP 를 생성하는 효소인 아데닐레이트 사이 크라제의 활성 조절기전에 관한 연구는 간이나 적혈구에서 이루 어졌다. 여러가지 새로운 사실들이 알려져 있고 신경계에서도 똑같은 기전일 것으로 알려져 있다. 이것에 대한 최근의 새로운 지견을 살펴보기로 한다• 8. 1 아데닐레이트 사이크라제 촉진계 노르아드레나린(/J-수용체), 도파민 (D1- 수용체), 세로토닌 (S1- 수용체), 히스타민 (H1- 수용체), 그루카곤, 부신피질 자극홀몬(
ACTH), 프로스타그란딘, 아데노신 (A2- 수용체) 등이 각각의 수용 체에 결합하면 아데닐레이트 싸이크라제 활성이 촉진된다. 이 반응에는 구아닌 뉴크레오티드(예를 들면 GTP) 가 필수적인 것으 로 이 GTP 는 수용체나 아데닐레이트 사이크라제 촉매단백과는 상이한 특이적 단백에 결합하여 수용체에 대한 전달물질의 결합 친화성이나 사이크라제의 활성을 조절한다. 이를 GTP 결합조절 단백이라고 한다. 신경전달물질 (T), 수용체 (R), GTP 결합조절단백 (G) 그리고 아데닐레이트 사이크라제 (C) 가 어떻게 상호작용하여 cAMP 를 생성하는가 를 개략적으로 살펴보기로 한다. T 가 R 에 결합하면 T - R 에 G 가 결합하여 T-R-G 복합체가 된댜 T-R-G 때에 R 은 T 에 대한 친화성이 높고 T-R 때는 친화 성이 낮다. G 에는 GDP 가 결합되어 있으나 T-R-G 가 형성되면 GDP 는 T-R-G 에서 유리되고 대신 GTP 가 결합한다. 이렇게 하여 생성된 G-GTP 는 T-R 에서 해리되어 C 와 결합하여 C 를 활성화함으로서 ATP 로부터 cAMP 를 생성하게 된다. G 는 GTP 를 가수분해하여 GDP 를 만들고 이는 G-GDP 가 되므로 원상태로 되돌아간다(그림 4-17). 이같은 수용체-아데닐레이트 사이크라제 계의 활성조절에 있어 중심이 되는 GTP 결합조절단백은 두 개 또는 세 개 서브유닐으로 되어 있고 촉진성 GTP 결합조절단백은 a- 서브유닐이 GTP 결합 부위이고 GTP 가수분해 효소활성을 가지고 있다. /3-서브유님은 조절단백이고 a- 서브유닐의 활성을
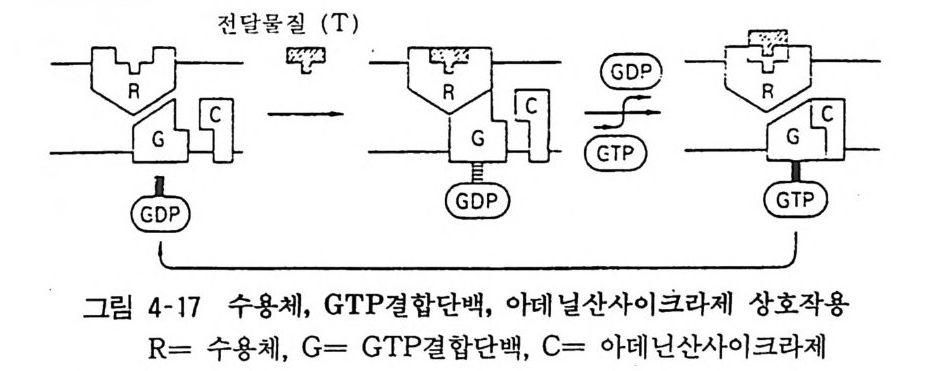 〔中전달물질 (T ) 言\
〔中전달물질 (T ) 言\
억제하는 기능을 갖는다. T 서브유닐의 기능은 잘 알려져 있지 않다(그림 4-18). 한편 콜레라 독소는 조직내의 cAMP 를 증가시 키는 것으로 이것의 작용부위는 촉진성 GTP 결합조절단백의 a- 서브유닐이다. 니코틴 아데닌디뉴크레오티드 (NAD) 의 ADP- 라이보실 부분이 콜레라독이 갖는 효소활성 (ADP- 라이보실 전이 효소)에 의하여 a- 서브유닐에 옮겨져서 a- 서브유닐의 GTP 가수 분해 효소활성을 잃게 된다. 따라서 GTP 가 계속하여 a- 서브유닐 에 남아 있게 되므로 아데닐레이트 싸이크라제의 활성이 지속함 으로 cAMP 가 증가하게 된다. 이 콜레라독은 GTP 결합조절단백 연구에 있어 대단히 중요한 역할을 하는 것이다.
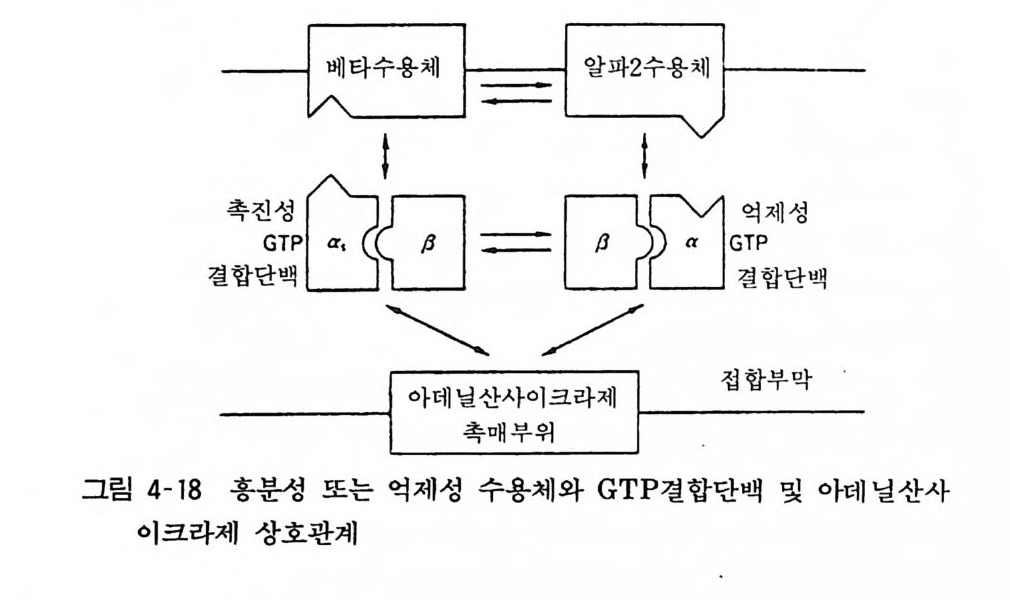 결 초합 「진 단G 성백 PT 曰아曰!\데닐 산 사一―이 크라제三:/ 曰접합G>부T P 막성단 백
결 초합 「진 단G 성백 PT 曰아曰!\데닐 산 사一―이 크라제三:/ 曰접합G>부T P 막성단 백
8. 2 아데닐레이트 싸이크라제 억제계 노르아드레나린 (a2- 수용체), 도파민 (D2- 수용체), 아데노신 (Al -수용체), 오피에이트 (o piat e), 바소프레신 (vaso p ress in), 소마토스 타틴 (soma to s tatin) 동이 각각의 수용체에 결합하면 아데닐레이트 싸이크라제 활성이 억제된다. 이같은 수용체-아데닐레이트 싸이크 라제 억제계에도 앞서 말한 촉진계처럼 GTP 결합조절단백이
관여하는 것으로 알려졌다. 이 조절단백은 효능제의 수용체에 대한 친화성을 조절하여 아데닐레이트 사이크라제 활성이 억제된 댜 이 조절단백은 효능제의 수용체에 대한 친화성을 조절하여 아데닐레이트 싸이크라제 활성 억제를 유발한다. 죽 억제성 GTP 결합조절단백도 a, B, Y- 서브유닐으로 되어 있고 a- 서브유닐이 촉진성 GTP 결합조절단백의 경우에 비하여 분자량이 작아 4 1 , 000 이다. 억제성 GTP 결합조절단백에 특이적으로 작용하여 ADP - 라이보실화에 의하여 이 조절단백의 활성을 잃게 하는 백일 해균 독소가 알려졌다. 아이렐액티베이팅 프로테인(I AP) 이라고 하는 이 독소의 발견으로 수용체-아데닐레이트 싸이크라제억제계 기전의 홍미있는 지견을 얻게 되었다. IAP 와 억제성 GTP 결합 조절단백 관계는 우이 (U i, 1984) 등이 상세하게 설명하고 있다. 최근의 연구로 뇌 접합부막에서도 1) 억제성 GTP 결합조절단 백이 존재하고 2) 적어도 분자량 41,000 인 서브유닐이 있고 3) 언-수용체 효능제 (Clon idine ) 또는 길항제 (Yo him bin)의 막결합 친화성을 조절하고 4) «2- 수용체 자극에 의한 아데닐레이트 사이 크라제 활성 억제 발현에 관여하는 것이 알려졌다. 또 접합후막의 B- 수용체를 이소프로테레놀로 자극하면 a2- 수용체의 수가 증가하 는 기전에도 이 촉진성 GTP 결합조절단백과 억제성 GTP 결합 조절단백의 상호작용이 관계될 것으로 생각하는 연구들이 진행되 고 있다. 최근 촉진성 GTP 결합조절단백과 함께 억제성 GTP 결합조절 단백도 사이에서 정제되어 분자 수준에서의 이들의 실체가 상세 하게 밝혀지고 있다. 특히 이들의 신경계에서의 기능, 질병 그리 고 약작용 등의 관계에 관한 연구도 크게 기대되고 있는 것이 다.
표 4-3 아데닐레이트 사이크라제 촉진계와 억제계 촉진성 외래자극 억제성 외래자극 조직 조직반옹 골격극 그리코겐 분해 지방세포 지질 분해 촉진 (아베드타레수나용린체) 심장 수축력 및 수축속도증가 장관 수분 분비 평활근 이완 갑상선 자극홀몬 갑상선 타이록신 분비 바소프레신 코o 파e 수분 재흡수 (V2 수용체) 그루카곤 간 그리코겐 분해 세로토닌 타액선 수분 분비 프로스타사이크린 혈소판 응집, 분비 억제 아드레나린 혈소판 응집, 분비 촉진 (알파 2 수용체) 지방세포 지질 분해 저하 아데노신 지방세포 지질 분해 저하
참고서적 1. G. M. Sheph erd : Ne ttro bio l og y. Oxfo rd Un ive rsit y Press; Ox for d, 1983. 2. P.L. McGeer, JC . Eccles, E.G. McGeer : Molecular Neurobio l og y of the Mammalia n Brain . 2nd Ed. Plenum, New York, 1987. 3. G. Guroff : Molecular Neurobio l og y. M. Dekker, New York, 1980. 4. J. R. Coope r, F.E. Bloom , R.H . Roth : The Bi oc hemi ca l Basis of Neurop ha rmacology 5th Ed. Oxfo rd : Un ive rsit y Press, Oxfo rd , l986. 5. M. Jac obson : Develop me nta l neurobio l og y. 2nd Ed. Plenum, New York, 1978. 6. G.J . Sie g e l , P.W. Albers, B.W. Ag ran oft , RT. Katz ma n : Basic Neurochemi str y . 3rd Ed. Lit tle, Brown, Bosto n , 1981. 7. A.J. Turner, H.S. Bachelard : Neurochem ist r y : A Practi ca l Ap p- roach. IRL Press, Oxfo rd , 1987. 8. A. Pete r s, S. Palay, H. Webste r : The Fin e Str u ctu r e of the Ner- vous Sy s te m : The Neurons and Supp o rtin g Cells, W.B. Saunders, Ph ilad elph ia, 1976. 9. A. Asbu ry, G. McKhann, W. McDonald : Di se ases of the Nervous Sy s te m : Clin i c a l Neit rob io l og y. W.B. Saunders, Ph ilad elph ia, 1986. 10. B. Katz : Nerve, Muscle and Sy n ap se. McGraw~ H ill, New York, 1966. 11. J. Eccles : The Phys io l og y of Nerve Cells. Joh ns Hop kins Un ive rs ity Press, 1957.
12. J. Eccles : Tlze Plzy s io lo gJ • of Sy n ap s es. Sp ringe r, Heid e lberg, 1964. 13. S. Ramon Y. Caja l : Recollecti on s of My Lif e. Un ive rsit y of Pen- nsyl v a nia Press, Ph ilad elph ia, 1937. 14. J.D . Wats o n : The Molecular bio l og y of tlze Genes. W. Benja min, Menlo Park, 1978. 15. R.D . Lund : Develop m ent and Plasti cit y of the Brain . Oxfo r d Un ive rsit y Press, Oxfo rd , 1978. 16. S.W . Kuf fler, J.G . Nic h olls : From Neruon to Brain . Sin a ner, Sunderland, M 琴 ., 1976. 17. S. Ramon Y. Caja l : Stu d ie s on Verte b rate Neurog e1 1esis . C.C. Thomas, Sp ringfiel d, III , 1960. 18. E. Kendall : Cellular Basis of Behavio r . Freeman, San Francis c o, 1976. 19. E.R. Kandel & J.H . Schwa rtz : Prin c ip le s of Neural Scie 1 1ce 2nd ed., Elsev ier Scie n ce Publis h i ng Co., Inc. 1985. 20. G.A . Robis o n and J.W . Lambie : Towards Understa n din g Recept ors . Elsev ier , Nort h- Holland, 1981. 21. J. R. Coope r, E.E. Bloom, R.H . Roth : The Bi oc hemi ca l Basis of Neurop ha rmacology , Oxfo r d Un ive rsi ty Press, Oxfo rd , 1982. 22. D.T. Krieg e r, M. J. Brownste i n , J.B . Ma rtin : Brain Pep tide s, Jo hn wiley & sons, New York, 1983. 23. J.C. Vente r , L.C. Ha rriso n : Recept or Bi oc hemi st ry and Metl zo dolog_ v , Alan a. Lis s , Inc., New York, 1984.
용어해설 강그리오시드(g an g li os i de) : 세포막에 결합된 지질당단백으로서 임상적으 로는 퇴행성 신경질환 (de g enera ti ve neural d i sease) 과 관계가 있을 것으 로 알려져 관심의 대상이 되고 있다. 잽접합부(g a pj unc ti on) : 세포들 사이에 작은 수분통로 (a q ueous channel) 로 서로 교통하고 있는 장소를 말한다. 공동전달 (co t ransm i ss i on) : 단일 뉴론자극으로 두 가지 전달물질이 동시 에 유리되어 동일한 주효세포에 영향을 미치는 현상이다. 아직까지 실험 적으로는 단일 뉴론에 두 가지 전달물질이 공존 (coe xi s tan ce) 하고 있음 이 많은 예에서 알려져 있으나, 생체에서의 의의에 대하여는 논란의 대상이 되고 있다. 교세포(그리아세포, gli al cell) : 신경계는 뉴론과 그리아세포의 두 가지 세포로 구성된다. 그리아세포에는 성상세포, 오리고덴드로사이트 그리고 또 한 가지 마이크로그리아(mi cro gli a) 가 있고·뇌실을 싸고 있는 상의 (epe n dy ma), 망막의 뮬러 (Muller) 세포, 소뇌피질의 베르그만 (Ber gman) 세포 등도 특이한 그리아의 일종으로 간주된다. 그리아세포는 뉴론을 지지하여 신경계의 형태를 유지하고, 수초를 형성하는 중추신경계 오리 고덴드로사이트는 말초에서는 슈완세포 (Schwarm cell) 라 한다. 국소순환뉴론(1 ocal cir c uit neuron) : 축색이나 수상돌기가 특정 구조에만 국한하는 뉴론을 말한다. 특수신경핵, 신경절 또는 대뇌피질 둥에
존재한다. 나트륨콩 H! 펌프 (sod i um- p o t ass i um pu mp ) : 뉴론을 포함하여 모든 살아 있는 세포는 세포내에 칼륨을 세포 밖에는 나트륨을 높은 농도로 유지하 고 있다. 이같은 세포막을 경계로 한 이온들의 농도경사는 나트륨과 칼륨에 의하여 활성화되는 Na-K-ATPase 인 나트륨 - 칼륨펌프에 의해 유지되는 것이다. 뇌파 (elec t roence p halo g ra ph y) : 머리 표면에서 감지하여 기록되는 뇌의 전기적 활동이다. 정상인에서 파형의 빈도에 따라 a, fl 등 네 가지 파형 이 있으며 질병에 따라 빈도와 폭이 달라진다. 뉴로필라멘트 (neuro fi lamen t) : 신경세포 골격 구성성분의 하나로 세포체 에서 돌기(축색, 수상돌기)에 걸쳐 길이로 뻗어 있는 섬유 모양의 구조 물이다. 이것은 세포막 및 소포, 미토콘드리아 둥 세포내 구조물과 연관 되어 있고 그들 상호간에도 연결이 있다. 뉴론 (neuron) : 신경세포, 죽 신경계의 구성단위이다. 이것은 세포체 (cell bcxly, soma perikary on) 와두 개의 돌기인 수상돌기와 축색으로 이루어 진다. 막전위 (membrane po te n ti al) : 세포막을 경계로 세포 밖에는 나트륨 이온 이 많고 세포 안쪽에는 칼륨이온이 많다. 이같은 세포 내외의 이온 농도 경사와 세포막의 이온들에 대한 투과성의 선택성으로 세포막 내외의 전위차를 이루게 된다. 정지상태의 막전위를 정지막전위 (resti ng mem-brane poten ti al) 라 하고 활동상태 에 는 활동전위 (acti on poten ti al ), 접 합 후전위 (postsynaptic poten ti al), 종판전위 (end pla te poten ti al ), 수용체전 위 (recep tor poten ti al ) 둥이 생 긴다. 면역형 광(i mmuno fl uorescence) : 항체 를 형 광물질로 표식 (lab eli ng) 하고 이를 조직과 반응시켜 형광현미경에서 항체의 위치를 알아보는 것이다. 미세관 (m i cro t ubule) : 세포골격 구성성분의 하나로서 속이 비어 있는
미세한 관으로 거의 모든 세포에 존재한다. 신경축색에서는 신경필라멘 트와 평행으로 다발을 이루고 뻗어 있으며 소포의 이동, 축색수송 둥의 기능을 담당한다. 성상세포 (as t roc yt es) : 그리아세포(gli al cell) 는 그 형태에 따라 성상세포 와 오리고덴드로사이트 (ol ig odendroc yt e) 로 나뉘고 성상세포는 다시 섬유성(fi brous) 과 원형질성(p ro t o p las mi c) 세포로 나누어진다. 성장추(g row t h cone) : 뉴론도 다른 세포에서와 마찬가지로 배양 초기에 는 둥 근 모양의 세포로 시작된다. 축색 (axon) 은 세포체에서 돋아나는 성장추(gr o wth cone) 라는 아메바양 돌기가 자라서 되는 것이다. 세포골격 (c yt oskele t on) : 각종 세포 특히 뉴론은 여러가지 세포골격 단백 들의 협동으로 그 기능과 형태가 유지된다. 세 가지 종류로 대별된다 . 액틴 (ac tin)과 그 관련 단백, 미세관(mi cro t ubule) 과 그 관련 단백 그리 고 뉴로필라멘트 (neuro fil amen t)가 그것이다. 세포간 물질 (ex t racellular matr ix) : 뉴론과 세포들 간에는 질서 있는 세포 간 물질 분자와 밀접한 관계가 있다. 이들 세포간 물질은 뉴론의 기능, 성장과 발육 및 재생과 관계가 있다. 크게 두 가지 부류가 있다. 프로테 오그리 칸(p ro t eo g l y can) 으로는 헤파린 (he parin)과 히아루로닌산 (h y al uron ic ac i d) 이 있고 당단백(g l y co p ro t e in)으로는 라미닌(larninin)' 화이 브로넥틴(fi bronec tin), 엔트악틴 (en ta c tin) 둥이 있다. 세포막 통로 (membrane channel) : 세포막을 관통하는 단백질로 이루어진 통로로서 이것을 통하여 이온(i on) 이나 체액에 녹아 있는 물질들의 세포 내외로의 이동이 일어나고 조절된다. 소포 (ves i cle) : 뉴론으로부터 유리되는 물질을 저장하는 작은 주머니로서 신경분비소포 (neurosecre t o ry v esi cle) 는 펩타이드 홀몬을 분비하는 신경 분비세포에 존재하는 것이고 시납스소포 (s ynapti c v esi cle) 는 시납스전 신경말단에서 자극전달물질을 저장하고 있는 것이다.
수상돌기 (dendr it e) : 뉴론에서 신호의 접수처라 할 수 있는 돌기로서 많은 가지를 가졌다는 것에서 붙여진 이름이다. 이것에서 접수된 신호는 축색을 통하여 전달된다. 그러나 수상돌기간의 신호교환 (dendrcxlendr iti c inter acti on )도 일 어 난다고 한다. 수용체 (rece pt or) : 신경접합후막에 존재하여 신경자극전달 물질과 반응함 으로써 신경자극에 부응하는 세포내 반응을 중개하는 것으로 노르에피네 프린과 반응하는 수용체는 아드레나린성 수용체 아세틸콜린에 대한 콜린 성 수용체, 오피움 펩타이드에 대한 수용체는 오피움 수용체, 세로토닌에 대한 세로토닌 수용체 둥 자극전달물질 특유의 수용체가 각기 존재한 다. 수초 (rn y el i n) : 축색을 여러 겹으로 둘러싸고 있는 막으로 중추신경계에서 는 오리고덴드로사이트에서 만들어지고 말초신경에서 슈완세포가 만든 다. 이는 전기적 절연체의 역할을 하여 신경전도를 빠르게 하는 데에 관계 된다 (sal tat o ry conducti on ) . 슈완세포 (schwann cell) : 축색으로 이루어진 말초신경섬유는 슈완세포로 둘러싸여 있다. 말초신경에는 유수섬유와 무수섬유가 있으며 유수섬유는 한 개의 축색을 수초형성을 이루는 슈완세포가 둘러싸고 무수섬유는 한 개 또는 그 이상의 축색들이 슈완세포 돌기에 둘러싸여 있으나 수초 형성을 이루지 않는다. 시납스 (s y na p se) : 뉴론과 뉴론 사이의 기능적 연결부위이다. 이 연결부위 를 통하여 뉴론간의 신호전달이 이루어지며 화학적 전달물질로 신호전달 이 일어나는 것을 화학적 시납스 (che mi cal s y na pse)라 하고 전기적으로 신호전달이 이루어지는 것을 전기 적 시납스 (elec tri cal s y na pse)라 한 다. 시 납스 가소성 (sy n ap tic pla sti cit y) : 신 경 계 에 서 상황 변화에 적 응하는 반옹이 이루어지려면 신경회로의 가변성 또는 가소성이 요구된다. 죽 시납스 가소성은 그 기능의 변화가 일어날 수 있도록 대치되거나 수의 중감이 일어나는 것을 말한다.
시납스 억제 (s y na pti c inh ib i t ion ) : 신경 흥분성의 조절은 시납스 억제를 통한 억제적 또는 홍분적 영향의 평형으로、 이루어진다 . 이같은 시납스 억 제 는 특이 적 인 억 제 성 인 터 뉴론 ( inh ib i t or y inter neuron ) 에 의 하며 억 제 적 전달물질로는 그리신(g l y c in e), GABA 가 있다. 시납스 억제에는 시납 스후 억 제 ( pos ts y n ap tic inh ib i t ion ) 와 시 납스전 억 제 ( pre syn ap tic inh- i b iti on) 의 두 가지 억제 방식이 있다. 시납스 전달 (s y na pti c tra nsmi ss io n ) : 시납스를 통한 뉴론 상호간의 신호 전달 과정을 말하며 화학적 전달물질에 의한 자극전달을 화학적 전달 (chem ica l t ransm i ss i on) 이라 하고 전기적으로 신호전달이 이루어지는 것을 전기 적 전달을 화확적 전달 (elec tri c t rans mi ss i on) 이라 한다. 신경 성장인자 (nerve gro wt h fac to r ) : 신경의 성장촉진을 유발하는 거대 분자로서 독사의 뱀독과 생쥐 악하선에서 발견된 것을 시작으로 여러 동물 조직에서 얻을 수 있다. 신경롱 (neural crest) : 척추동물 태생기의 초가 신경원기 (neural primo - r di wn) 인 신경융기 (neural fo ld) 에서 유래하는 잠정적인 구조이다. 신경 융세포는 정확한 시기에 정확하게 정해진 장소로 이동하여 각종 세포로 분화한다. 신경세포 이동 (neural mi gr ati on ) : 척추동물의 신경세포는 성숙한 뇌에서 영주하는 장소와 별개의 장소에서 발생된다. 신경세포 이동이란 세포가 기원하는 장소에서 종착지인 성숙 뇌의 영주 장소로 이동하는 현상을 말한다. 이는 축색의 성장과는 다른 것이다. 신경전달물질 (neuro t ransm itt er) : 뉴론과 뉴론 사이 또는 뉴론과 주효장기 사이의 접합부에서 신경자극을 전달하는 화학적 물질이다. 말초신경에서 잘 알려진 아세틸코린, 노르에피네프린을 비롯하여 중추신경계에서 GABA, 그리신, 구루타민산 둥 아미노산과 도파민, 세로토닌, 히스타민 둥의 아민류 그리고 엔돌핀, 엔케파린, VIP, 뉴로텐신 둥의 펩타이드 둥 약 50 종이 거론되고 있다.
신경펩타이드 (neuro p e pti de) : 오피움양 펩타이드 (o pi od pepti de) 인 엔돌 핀, 엔케파란 다이놀핀을 비롯하여 최근에 중추신경계에서 자극전달 내지 자극전달의 조절물질로 주목을 끌고 있는 펩타이드로서 섭스탄스 피, 뉴로텐신, VIP, 콜레치스토키닌 동 약 30 가지의 펩타이드가 거론되 고 있다. 신호변환 (s ig nal tra nsducti on ) : 전기적 신경신호(활동 전압)는 화학적 자극전달물질을 유리하고 이것이 세포막 수용체에 작용하면 일련의 신호 변환 반응을 일으켜 세폰기능의 변화를 초래한다. 이같은 신호변환에 는 아데닐레이트 싸이크라제 -cAMP 계, G - 단백 GTP 계, 칼슘 - 칼모듀린계 둥이 관여하여 세포내 반응을 나타내게 한다. 아드레나린성 신경계 (adrener gi c nervous sys t e m ) : 말초 자율신경계는 생리학적(기능적)으로는 교감신경계와 부교감신경계로 분류되나 신경전 달물질에 따라서는 노르아드레나린(또는 노르에피네프린)에 의하여 자극 전달이 이루어지는 신경계를 아드레나린성 신경계라 하며 말초 교감신경 계가 이에 해당하나 중추신경계 중에서도 노르아드레나린에 의하여 자극 전달이 이루어지는 신경계가 있다. 인터뉴론(i n t emeuron) : 개재뉴론(int ernunc i al neuron) 또는 경계뉴론 (inter calate d neuron) 이라고 하며 비교적 짧은 축색을 갖고 국소적으로 작용하는 것이다. 그 기능에 따라 억제성 인터뉴론(inhi b it o ry int e r ne- uron) 과 홍분성 인터뉴론 (exc itat o ry int erneuron) 이 있다. 자율신경계 (au t onom i c nervous sys t e m ) : 신경계 중에서 내장의 평활근 또는 분비선을 지배하여 그것들의 불수의적 운동과 기능을 조절하는 신경계이다. 교감신경계와 부교감신경계로 구분된다. 전압 크램프 (vol t a g e clamp ) : 세포막을 통한 이온 전류를 측정하기 위한 전기생리학적 기법이다. 세포내의 미세전극을 통하여 세포내 전위를 인위적으로 임의의 수준으로 일정하게 유지토록 하는 것이다. 카테콜아민 (ca t echola mi ne) : 벤젠 환 (benzen ring)에 두 개의 ― OH 기와
또 하나의 에틸아민 측쇄를 가진 화합물로서 도파민, 노르에피네프린, 에피네프린을 일컫는다 . 콜린성 뉴론 (chol i ner gi c neuron) : 아세틸콜린에 의하여 자극전달이 이루 어지는 뉴론을 말하며 말초 부교감신경, 운동신경 그리고 자율신경절전 섬유(p re gan gli o ni c fi ber) 가 이것에 속하고 중추신경계에도 코린성 뉴론 이 존재한다 . 축색 (axon) : 뉴론의 일부분으로 신경자극전도 (conduc ti on) 를 담당하는 특이적 부분이다. 수 밀리미터에서 수 미터에 이르는 다양한 길이룰 갖고 있으며 수초 (m y e lin)의 유무에 따라 유수섬유 (m y e lin a t ed fibe r) 또는 무수섬유 (norun y e linat ed fi ber) 가 있다. 축색수송 (axonal tra nsp o rt ) : 신경세포에서 단백질 합성은 주로 세포체 (perik a ry on) 에서 이루어진댜 이같이 합성된 단백질은 축색류 (axonal fl ow) 로 신경 말단 (nerve t e rminal)으로 수송된다. 반대 방향의 수송도 일어날 수 있어 이룰 역행성 축색수송 (re tr o gr ade axonal t rans port)이라 한댜 팻치 크램프(p a t ch clamp ) : 세포막을 통한 이온전류를 측정하는 최신의 신경생리학적 방법이다. 미세유리피펫을 세포막에 대고 홉인하여 피펫 끝을 세포막으로 막히 게 하는 것으로 on-cell patch , inside -out patch , whole-c e ll pa tc h , outs id e -out patch 둥 네 가지 방법 이 있다. 혈관여 장벽 (Blood-bra i n barrie r ) : 뇌의 세포외액 (ex tr acellular fluid) 조성은 혈장의 조성과 많은 차이가 있다. 이같은 차이는 타조직의 모세 혈관과는 구조적으로나 기능적으로 다른 모세혈관벽에 의한다.
찾아보기
-1 가나마이신 (Kanam y c in) 84 가락토세 레 브로사이드 (ga)actocereb - rosid e ) 63 GABA 션트 (Shun t) 104 GABA 아민전이효소 (GABA-T) 104 가스트린 (Gas trin) 109 가토 (Ka t o) 112 갇덤 (Gaddurn) 111 감마아미노브티릴산 (GABA) 103 감마 엔돌핀 114 갑상선자극홀몬 유리 홀 몬 (TRH) 109 강그리 오사이 드 (ga ng lios id e ) 24, 64 개 구분비 (exocy tos is ) 29 개제뉴론 22, 35 갭 결합(g a p junc tio n ) 36 결합조 절 단백 145 계획적 세포사(p ro gr ammedcelldea t h) 44 골수종 세포 (m y eloma cell) 128 골지 (Golgi ) 42, 46 공간적 가중 (s p a ti al summatio n ) 33 교돌핀 ( Ky ot o r r phin) 110 과산화효소 (Horsera di sh pero x ida se) 61
광친 화성 표식 법 (ph oto a ff inity labe- ling) 124 구아닌 뉴크레오티드 (GTP) 143 구아닐릭 사이크라제 138 구안에티딘 (Guane thidin e) 96 국소회로 뉴론(l ocal circ uit neuron) 22 그라이 오필 라멘트 (gli of i lam ent ) 63 그리 신 (gly c i n e ) 35 글루타민산 103 글루타민산 탈탄산효소 (GAD) 103 기동전위 (ge nerato r poten tia l) 28 기저막 43 L 나록손 (Naloxone) 114 난포자극홀몬 유리홀몬 (LH-RH) 109 내어 (Neher) 65 네오마이신 (Neom y c in) 84 네 오스티 그민 (Neostig mine ) 84 노르아드레 나린 (Noradrer ialine ) 86, 94 노르아드레 나린 뉴론 88 노인반(senil e pla q u e) 46 뇌 관류법 11.9 뇌신경 발생 51
뇌 은행 (Br ain bank) 66 뇌의 조직발생 18 뇌절편 119 뉴로모듀레 이타 (neuromodula t or) 108 뉴로키 닌 (Neuroki nin ) 110 뉴로텐신 (Neuro t ens in) 109 뉴로펩타이드 (Neuro pepti de) 107 뉴로피라멘트 (neuro filam en t) 48, 63 뉴론-그리아상호관계 15 뉴론 돌기 18 니코틴성 아세틸코린 수용체 128 니코틴성 수용체 (nico ti nic recep tor ) 81 니코틴 아데닌디뉴크레오티드 (NAD) 144 C: 다발성 경 화중 (multip le scleros is ) 49 다이놀핀 (d yn o rphin e) 1I5 단크론 항체 (Monoclonal antil _xxiy) 126 단크론 항체 법 (monoclonal anti bo dy ) 63 데 일 (Dale) 의 원리 22, 108 데 카메 토니 움 (Decame tho ni wn)
84 도파민 (Do pamin e) 86, 94 도파민 베타수산화효소 (DB H) 86 DOPA 탈탄산 효소 (TH) 86 디벤아민 (D i be namin e) 94 디 아씰그리 세 롤 (di ac y l g l y cerol ) 137 N 하 이온 통로 133 D1 수용체 90 다 수용체 90 DNA(cDNA) 121 2 라미 닌(laminin) 23 랜드맷서 (Landmesser) 44 레비 몬탈치니 (Le vi -Mon tal c ini) 55 레세르핀 (rese rpin e) 93 렌쇼우 세포 (Renshaw cell) 35 뢰위 (Loew i) 33, 70 류신 엔케파린 115 류코트리 엔(l euko tri ene) 116 리 러 (Reeler) 18 리 포모듀린 ( lipom odu lin ) 139 Lo 축색류 (Axonal flow ) 77
□ 마리화나 102 마운트 캣슬 (Moun t castle ) 65 마이 에 린 (my el in sheath ) 48 말초 신경계 23 면역조직화학 61 메타라미놀 (Me t aram i nol) 94 매 타보트로픽 (meta b otr o p ic) 시 납 스 36 매티오닌 엔케파린 115 메 타코린 (Meth a cholin e ) 85 메 티 테 핀 ( Meti tep i ne ) I0 2 모노아민 옥시다제 (MAO) 87 무스카린 (Muscarin e ) 86 무스카린성 수용체 (musca rini c re-cep tor ) 81 뮤시몰 (musc im ol) 105 미 레 이 (Murray ) 67 미 렐디 (Mi led i) 29 미소전극 61 미소종판전위 (mi niat u r e endp la te JX)ten ti al, MEPP ) 30 마상핵 93 미주신경물질 (va gu s sto f f ) 70 밀도차원심분리법 66 밋 첼 ( Mi tch ell ) 139 CX:1 수용체 89 CX:2 수용체 89 cx:- MSH 114
닌 바소프레 신 ( Vasop re ssin ) I 09 반복자극 후 증강(po s t-t e tani c po-ten ti at i on ) 38 방사성 그리 아 (ra dia l gli a) 18 방사선실활법 (rad iation inac ti va ti on ) 125 백일해균 독소 145 베르그스트롬 (Ber g s t rom) 116 베인 (Vane) 116 베타리포트로핀 114 베타네콜 (Be th anechol) 85 베타(/J)수용체 89 베 타 엔돌핀 (/J-e ndorp hin) 114 베타 엔돌핀 114 벤조디 아제 핀 (Ben zodiazepine s) 107 벤터 (Vente r ) 126 브레티리움 (Bre tyli wn) 96 보수계 (reward sys t e m ) 92 보투리 누스독 (Botu linu s tox in) 84 본태의 수면 100 부신피질 자극홀몬 (ACTH) 114 부톡사민 (buto x a mine ) 95 불쾌 감 (d y s p ho ria ) 116 브라디키닌 (Brad ykinin) 109 비쿠쿠린 (b i cuc ullin e) 105 빈브라스틴 (vinb lastin e ) 78 빌효우 (V ir chow) 42
VIP(vasoacti ve intes ti nal petide ) 108 人 사무엘손 (Samuelson) 116 삭시독신 (S axit o xin) 133 산욕열 (Puerp era l fev er) 100 살부타몰 (Salbu ta wol) 94 상동행동 (s t ereo type behav ior ) 92 석시닐세미알데하이드 (s uccin yls em ial dehy d e ) I 04 석 시 닐코린 (Succin ylc holin e ) 84 섭스탄스피 (Substa nc e P) 109, 111 성상교세포 혹은 아스트로사이트 ( Astr ocyte ) 40 성장원추 59 셰 링 톤 (Sherr ington ) 33 세로토닌 (Sero t o nin, 5-HT) 97 세로토닌 뉴론 98, 103 세로토닌의 합성과 대사 98 세포간 물질 (e xtra cellular matr ix)
23 세포체 20 소뇌과립 세포(gr anule cell) 18 소마토스타틴 (Soma t os tatin) 109 소타롤 (sota lol ) 95 소포(리 셀토좀) 132 잎 속행성 축색 류(fast axop lasmic f low )52 수상돌기 (dend rite) 18, 20 수용체 123 수초 ( my e lin sheath ) 41 수초 염 기 성 단백 질 (my e lin basic pro te i n ) 63 숫적 인 감소 (down reg ula ti on ) 132 슈완 세 포 ( Schwann cell ) 41 시 간적 가중 ( tem p ora l summati on ) 31 시납스 (s yn a pse) 15, 29 시 납스 간격 ( syn a p tic cleft ) 31 시납스 발생 (s ynapt o g enes i s) 52 시납스 소포 (s yn a pti c vesic l e) 29 시납스의 가소성(p las ti c ity) 22, 38 시 납스전 억 제 (pre sy n ap tic inhibi - tion ) 35 시 납스후전위 (postsy n a p tic poten - tiaL PSP) 22 시 납스후억 제 (postsy n aptic inhibi - tion ) 35 시납스 형성 59 시네프린 (S yn e phrin e) 94 신경관 (neural tub e) 16 신 경 구 (neural groo ve) 16 신경교세포(glia) 15 신경말단부(시납토좀 Sy naptosom e) 120 신경상피 세포 18
신경성장인자 (NGF) 55 신경세포 (neuron) 15, 20 신경융 (neural crest) 16 신 경 원 섬 유 변 화( neurofi br i llary tan g le s) 46 신경재생 24 신 경 절 (ga ng li on ) 23 신경종말 15 신경판 (neural pla te ) 16 신경핵 (nucleus) 18 스코폴아민 (Scop: >la mi ne ) 85 스트렙 토마이 신 (St rep tom y c in ) 84 스피페론 (S pipe rone) 99 5-HT1 수용체 99 5-HT2 수용체 99 ^) 아데 닐레 이트 사이크라제 (Adeny - late cyc l ase) 89, 135, 141 아데닐사이크라제 촉매 유닐 132 아드레나린 (Adre nalin e) 86, 94 아라키돈산 (arac hi do ni c aci d) 116 아레코린 (Arecho lin e) 86 아미로이드 섬유 (am y lo i d) 48 아스트로사이트 43 아세 틸코린 ( Acety lc holin e ) 78 아이텔액티베이팅프로테인 (IAP) 145
아트로핀 (A tr o pin e) 85 안지오텐신 II (Ang iot e n sin ) 109 알츠하이 머 병 (Alzheim e r's d is eas e) 46 알파메라닌 (a-mel anin e) 91 알파메틸노르아드레나린 (a-meth yln oradren aline ) 96 알파메 틸도파 (a-me ty ldo pa) 96 알파, 베타-네오엔돌핀 (a, /3- neoendorp hin) llO, 115 알프레 노롤 (al p renolol) 95 암페타민 (Am p he tamin e) 97, 102 양자적 방출(q uan t um release) 31 억제성 GTP 145 억제성 시납스 33 에레도이신 (eledo isin) 111 에 페 드린 (Ep h e drine ) 94 엑클스 (Eccles) 29, 33, 46 엔도사이토시스 (endoc ytosis) 132 엔돌핀 (Endorp hin) 109 엔케파린 (enke p迫lin) 114 엘리오트 (E lli o t) 70 역설적 수면 100 오리 고덴드로사이 트 (O lig odendro cy te) 41 오피오이드 펩타이드(Opi o i d pep- tide ) 114 옥시토신 (O xytocin) 109 요힘빈 95 우울중 93
우이 (U i) 145 운동뉴론 그룹 42 옹게르스테트 (U ng ers t ed t) 87 위 (membrane poten ti al ) 25 위버 (Weaver) 18 위 젤 (W ies el) 65 은염색법 42 이노시톨트리포스페이트(!P 3) 139 이미프라민(Imip ram in e) 97 이 소프로테 레 놀 ( Isop ro te r enol ) 94, 95 이오38 노’ 트로픽 (ion otr op ic) 시 납스 이온 채널(i on channel) 27 이종시납스간 촉진 (he t eros yn a ti c fac ili tation ) 38 이차 전달자 (second messen ge r) 36 인돌핵 (Indole) 97 인지질 메틸전이효소(p hos p ho lipi d meth ylt ra n sfe ra se ) 135 인지질 (Phos p ho lipi d) 메 틸화 반응 135 인지질의 인공막(liposo me) 133 인터 뉴론 (intem euron) 22 임펄스(imp ulse) 29 GTP 결합조절단백 132, 143 internalization 132 LSD 33, 101, 102 mRNA 121
ACTH 114 cAMP 134 7 자가수용체 (au t orece pt or) 87 자극도입 (impu lse tra nsducti on ) 134, 135 장기 적 억 제 (lon g- te r m dep re ssio n ) 39 장기 적 증강(l o ng - t erm poten ti ation ) 38 재 구성 막 ( reconsti tut e d membrane ) 133 재 이 용 ( recy cl in g ) 132 전 기 뱅 장어 ( Torp edo calif om i ca ) 128 전기성 시납스 36 전뉴론 (rela y neuron) 22 전세포 크램 프법 (whole cell clamp ) 65 정신분열 치료약 96 정 지 막전위 (restin g poten ti al ) 27 제 2 전달체 (second messen ge r) 134 중중근 무력증 (M y as th e nia grav is ) 84, 130 중추신경계 17 쥬베 (Jo uvet) 91 지 연성 축색류 (slow axopl asmic f low ) 52
GTP 결 합조 절 단백 132, 143 天 척수 (s pi nal cord) 17 척 추 이 분 ( spi na bif ida ) 17 체온 조절 100 촉진 성 GTP 결 합조 절 단백 145 추체 외 로 계 ( extr a p yra m ida l sys t e m ) 93 축 색 (axon) 18, 20 축 색 기시 부 (axon hil lo ck) 22 치매 (노망) 46 E 타이로신 수산화효소 (TH) 86 타케우치 (Takeuchi) 34 탈 감작 (desens itizati on) 131 탈수소효소 (SSADH) 104 탈수질환 49 테트라하이드로 칸나비놀 (THC) 102 톨라졸린 (Tolazoli ne ) 94, 95 트란스시 납스성 표식 (tra nssyn a pt ic labelin g ) 61 트롬복산(t hromboxane) I 16 토리프토판(trypt o p han) 97 트리프토판 -5- 수산화효소(trypt o -
ph an- 5- hy d roxy la se) 97 더 카바콜 (Carbachol) 85 카세 이 몰핀 ( Caseim o rph in ) 110 카테콜 - O- 메틸전이효소 (COMT) 87 카테 콜아민 ( Cate c holami ne s ) 86 카할 (Ca ja l) 46 칼슘수용단백 (Calmodul i n) 140 칼슘 이온 134 캇츠 (Ka tz) 29, 34 캇탄세 린 (Keta n seri ne ) 99 캇테 랄 (Ca tt erall) 133 코라젠 (colla g en) 23 코레치스토키닌 (CCK) 108, 109 코린성 수용체 (Cho lin er gi c recep tor ) 81 콜레라 독소 144 콜키친 (colco i c in e) 78 쿠훌러 (Kuff ler ) 29, 34 크로니 딘 (clon idine ) 96 클로르프로마진 ( Chlorp ro ma zine ) 97 c- 키 나제 (C-ki na se) 138 cAMP 134 Ca++ 이 온통로 134 Ca++ 길항제 134
工 파상풍(t e tan us) 35 파킨손씨 병 (Parkin s on's dis e ase) 93, 107 팻치크램프법 (patch clamp } 65 팻토 (PET) 66 페닐에타놀아민 -N - 메틸전이효소 (PNMT) 86 펜옥시벤자민 ( Phenoxy b enzami ne ) 94 펜톨아민 (Phento l ami ne ) 94 포메라트 (Pomera t) 67 포스파티 딜 이 노시 톨 (ph osp h ati dy l ino si tol ) 134, 137 포스포리 파제 A2 139 포스포리파제 C(p h osph olip as e C) 137 포지트론 에미손 토모그래피(po s itron em ission tom og rap h y ) 66 포지티브 휘드백 (po sit ive fee d- back) 27 폰 율러 (von Euler) 111 푸르키니에 세포 (Pur kinje cell) 22, 35 프로다이놀핀 115 프로락틴 (Prolacti on ) 91 프로스타그란딘 (pro sta gla nd ins ) 116, 138 프로스타싸이 크린 (pro sta c y cl i n) 116
프로엔 케 파린 에 이 ( pro enkep h alin e A) 115 프로오피오메라노코틴(p ro opi om elanocortin ) 114 프로테오그라이 칸(p ro t eo g l y can) 23 프로프라노롤( pro p ra nolol ) 95 피 사레 민 ( ph y sa lemi n ) 111 핀도롤(pin dolol) 95 필로칼핀 (P i locar pi ne) 86 Fura-2 61 -_o 하이브리도마(잡종세포) 128 5- 하이 드록시 인돌 아세 틸산 (5- HIAA) 97 할로페 리 돌 ( Halop erido l ) 97 해 리 슨 ( Ha rriso n ) 66 핵자기공명 (nucleomag ne ti c reson-ance) 66 향정 신 성 (psyc hoto m ime ti c ) 116 헌팅톤 무도병 (Hunti ngton 's chorea) 84, 93, 107, 113 혈관 내피세포 43 혈관 활성장관 펩타이드 (VIP) 109 혈액 -뇌 장벽 (blood-brai n ba rrier ) 43
헤 미 코리 니 움( Hemi ch olin ium ) 84 헥 사메 토니 움 ( Hexameth o n ium ) 85 화이 브로넥 틴 ( fibr onecti n ) 23 화학적 유인설 59 화학적 촉진설 59 환형 AMP(cAMP) 142 활동전위 (acti on poten ti al ) 22, 27 효능제 130 훼 닐 에 프린 ( Pheny le p h rin e ) 94 휴벨 (Hubel) 65 혹질 93 흥분성 시납스 31 흥분성 시 납스후전위 (EPSP) 31 희돌기 교세포 41
1897.1
박찬웅 서울대학교 의과대학 졸업 현재 서울대학교 의과대학 교수 및 서울대학교 의과대학 신경과학연구소 소장 김승업 서울대학교 의과대학 졸업 현재 캐나다 브리티쉬 콜롬비아대학 교수 신경과학 대 우학술총서 • 자연과학 64 찍은날―-— 1990 년 3 월 30 일 펴낸날―― -)990 년 4 월 IO 일 • 지은이―-_박찬옹 • 검승업 펴낸이-~朴孟浩 펴낸곳_民音社 ■ 출판등록 1966. 5. 19. 제 1 - 142 호 우편대체번호 010041-31-0 5 23282 은행지로번호 3007783 110-111 서울 종로구 관철동 44-1 7344234 • 6110 (편집부) 734 -20 00 • 2101, 735-8524 (영 업부) 733-7010 (팩시밀리) Prin ted in Seoul, Korea • 값 5 , 000 원 © 박찬옹 • 김승업, 1990 자연과학 • 생물학 KDC/472 • 193
대우학술총서 ( 자 연과 학 ) I 소립자와 게이지 상호작용 검진으1/ 4,800 원 2 동력학특론 이병호 /5,400 원 3 질소고정 每중달 /2 , 800 원 4 相轉移와 임계현상 검두절 /2,800 원 5 촉매작용 진종식 /2,800 원 6 뫼스 H 유어 분광학 옥랑남 /2 , 印원 7 극미량원소의 영양 승정자 /6, 았)원 8 수소화봉소와 유기붕소화합물 윤능민 /5 , 0CIJ원 9 항생물질의 전합성 강석구 /9 , 0CIJ원 10 국소적 형태의 At iya h-Si ng er 지표이론 지동표 /2, 印)원 11 Muco p ol y sacchar i des 의 생화학 및 생물리학 박준우 /3,800 원 12 ASTROPHYSICS 홍승수 /4 ,7 00 원 13 프로스타 글란딘 합성 김성각 /3,600 원 14 천연물화학 연구법 우원식 /9, 0CIJ원 15 脂防營養 김숙호 1/6 , 300 원 16 결정화 유리 김병호 /4, :xXl원 17 고분자의 화학반응 조의환 /4 , 0CIJ원 18 과학혁명 김영식 /4,200 원 19 한국지질론 정기홍 /4, 0CIJ원 20 정보이론 한엽열 /4, :xXl원 21 원자핵반응론 정운력 /8, :xXJ원 22 파괴역학 김상절 /4 ,7 00 원 23 분자궤도이론 이익준 /3,300 원 24 반응속도론 정경훈 /4, 0CIJ원 25 미분위상수학 이현구 /2, :xXJ원 26 磁氣共鳴방법 조성호 /4, 紅)원 27 퓰 a 巳마 물리학과 핵융합 죄덕인 /4 , 邸원 28 천문관측과 분석 01 시우 /4,800 원 29 석탄에너지 번환기술 김상돈 /6 , :xXJ원 30 해양籠古生物學 백광호 /6,400 원 31 편미분방정식론 김종식 /4, 0CIJ원 32 대통일이론 소광섭 /6, 0CIJ원 33 금속전자계의 다체이론 김덕주 /6,600 원 34 액정중합체 진정일 /6,200 원 35 복합재료 권숙인 /5,200 원 36 탠백질 생합성 박인원 /9, 0CIJ원 37 한국의 광물종 김수진 /7, 0CIJ원 38 일반상대론 01 절훈 /6, 0CIJ원 39 레이저광산란분광학 김종진 /5, 0CIJ원 40 복소다양체론 김상문 /7, 0CIJ원 41 役學的 연구방법 김일순 /4,200 원 42 핵구조 물리학 민등팥 /5,300 원 43 후라에 해석과 의마분 작용소 김도한 /5,800 원 44 한국의 고생물 이하영 /9, 0CIJ원 45 질량분석학 김명수 /6,0C 迎