
백광호 저자는 서울대 문리대 지질학과 및 동대학원을 졸업하고, 서독 키일대학교에서 이학박사학위를 취득했다. 동력자원연구소 선임연구원과 스탠포드대학 객원교수 를 거쳐, 현재는 고려대학교 이과대학 지질학과교수 로 재직중이다 . 저서로는 『 지구과학 』 I·Il 가 있으며, 「페르시아만의 현대개형류에 대한 연구」 외 15 편의 논문이 있다.
 해양미고생물학
해양미고생물학
 해양미고생물학
해양미고생물학
머리말 바다는 인류의 마지막 남은 寶庫라고 한다• 세계 각국은 겅쟁적으로 해 양의 연구와 개발에 정력적인 관심을 쏟고 있다. 바다 속에는 막대한 양의 식량 및 광물자원이 숨겨져 있기 때문이다. 이러한 자원경제적인 면뿐만 아니라 바다는 아직도 그 生成과 歷史에 관한 깊은 신비 속에 잠겨 있는 것이다. 古海洋의 旅程온 바로 지구라는 행성의 역사를 대변한다고 해도 과언이 아니다. 현재 육지 . 는 많은 부분이 옛날 지질시대 고해양의 생성과 소멸의 역사로 짜여져 있기 때문이다. 따라서 현재 바다에서 일어나고 있 는 지질작용을 이해하고 그 원리를 추출하는 일은 곧 지구의 과거 역사를 복원하는 데 첫걸음이 되는 것이다. 海洋微古生物學은 현미경적 크기의 해양미고생물, 즉 微化石울 연구하 는 학문이다. 연구 대상이 되는 미화석의 종류는 대단히 다양해서 분류학 적으로 전혀 관련이 없는 동뭉 및 식물군을 광범위하게 포함한다. 대부분 의 미화석군은 原生動物에 속하나, 어떤 것은 다세포 혹은 큰 생물의 작은 부분인 것도 있다. 소속의 다양성에도 불구하고 이들을 모두 마화석군으로 묶어 다루는 것이 실용적이고 유용하기 때문에 微古生物學이란 분야가 생 긴 것이다. 이 책은 지구과학 분야중 특히 지질학, 해양학 그리고 자원탐사를 전공 으로 하는 대학원이나 학부의 고학년 과정에 알맞도록 해양미고생물학의 원리와 해양 미화석의 각론을 종합 정리한 것이다. 그림 및 도표의 자료들 을 많이 삽입하고 되도록 평이한 설명을 평으로써 비전공인 분들을 위한 배려를 했다.
제 1 장에서는 해양미고생물학의 의의, 연구사, 연구의 목적 그리고 응용 에 관한 전반적인 서론부를 섣고, 제 2 장에서는 해양미고생물학의 기본원 리들을 다루었다. 제 3 장은 현재 가장 널리 연구 • 활용되고 있는 미화석 10 종류를 선별하여 고생물학적 독성과 지질학적 응용에 중점을 둔 微化石 各論편이다. 이들은 有孔蟲, 介形蟲, 석회질 난노플랑크톤, 石灰漢, 放散 蟲 바다규조, 규질편모충과 에브리디안, 코노돈트, 쌍편모조류, 花粉세包 子화석 등이다. 각 종류별로 그들의 생물사, 형태, 분류, 생태 및 고생태, 生地理, 生層序 그리고 진화의 순서로 고찰하였다. 우리나라의 실정을 감 안하여 有孔蟲과 介形蟲은 특별히 많은 설명을 가하였다. 한반도는 삼면이 바다로 둘러싸여 있고 최근 일어나기 시작한 대륙붕 개 발 및 고해양학적인 연구열은 해양학 전반의 도약을 예고하고 있다. 이러 한 때 이 책의 탄생을 . 다행히 생각하며 앞으로 더 훌륭한 책이 많이 출판 되리라 믿는다. 많은 부족한 점은 앞으로 보완되리라고 믿으며 끝으로 원 고 정리, 교정 등의 일을 도와주신 동력자원연구소 최계림박사, 고려대학 교 지질학과의 고생물학 연구실의 이의형, 정혜경, 유인창 석사, 김용기 학사 등에 심심한 사의를 표한다. 1986 년 5 월 白光浩
해양미고생물학·차례
머리말 5제1장 序論1 海洋微古生物學 112 硏究史 143 연구의 목적과 응용 16제2장 海洋微古生物學의 기본원리1 生物層序學 191.1 생존대와 분대 191.2 충서대비 212 古生態學 252.1 해양생태학 252.2 고생물지리 283 進化學 323.1 진화이론의 발전 323.2 진화와 미고생물학 384 海洋微古生物學 40제3장 海洋微化石 各論1 有孔虫 451.1 서론 451.2 생활사 471.3 형태 및 분류 521.4 생태 및 고생태 781.5 생지리, 생층서 및 진화 862 介形虫 104
2.1 서언 1042.2 생활사 1062.3 형태 및 분류 1122.4 생태 및 고생태 1422.5 생지리 및 생층서 1463 石灰質 난노플랑크톤 1563.1 서언 1563.2 생활사 1583.3 형태와 분류 1613.4 생태 1773.5 고생지리와 고기후 1793.6 생물층서 및 진화 1814 石灰藻 1894.1 서언 1894.2 생활사 1904.3 형태와 분류 1914.4 생태와 고생태 2024.5 생물층서 2045 放散虫 2105.1 서언 2105.2 생활사 2125.3 형태와 분류 2145.4 생태 2185.5 고생태 및 고해양학 2195.6 생물층서 2226 바다 硅藻 2316.1 서언 2316.2 생활사 2316.3 형태와 분류 2336.4 생태와 고생태 2346.5 생물층서 및 진화 2407 珪質靴毛虫 및 에브리디안 247
7.1 서언 247 7.2 생활사 2487.3 형태 및 분류 2507.4 생태 및 고생태 2507.5 생물층서 및 진화 2517.6 에브리디안 2S28 코노논트 2578.1 서언 2578.2 형태 및 분류 2588.3 고생태 2658.4 생물층서 및 진화 2659 双鞭毛 藻類 2699.1 서언 2699.2 형태 및 분류 2719.3 생태 2799.4 생지리 고생태 2809.5 생물층서 및 진화 28310 花粉과 胞子 29110.1 서언 29110.2 형태 및 분류 292 10.3 군집의 형성 296 10.4 생물층서 및 고생태 300 한글색인 311 영문색인 314제 1 장 序 論 (Intr o ducti on ) 1 海洋微古生物學 바다에서 생활하는 대부분의 부유성 및 저서성 생물들은 경질부를 갖고 있 지 않기 때문에 화석으로 보존되기가 어렵다. 그래서 소수의 그룹만이 화 석화되며 이들은 성해처 퇴적물의 가장 중요한 성분을 이룬다. 화석으로서 지층에 보존되는 미생물 그룹은 해양환경의 발달사를 암시하는 칭보를 간 직하고 있으므로 해양 지질학자들의 큰 관심을 끈다. 정의에 따르면 미고생물학(mi cro p aleon t ol ogy)은 현미경적인 크기의 화석 울 연구하는 학문이다. 연구 대상이 되는 미화석의 종류는 대단히 다양해 서 분류학적으로 서로 전혀 관련이 없는 동물 및 식물군을 모두 포괄한다• 단지 현미경으로 관찰해야 되는 화석그룹들이 미화석에 포함되며 현미경 의 도움이 필요없는 그룹은 거화석군으로 거고생물학의 연구 분야에 포함된다. 따라서 미고생물학에는 어떤 통일적이고 동질한 원리가 통용되 지 않는 것으로 보이며 오히려 고생물학 전반에 걸친 나아가서는 지질학 의 원리가 이들 세부 분야의 연구를 관장한다고 볼 수 있다. 대부분의 해양 미화석군은 원생생물에 속하지만 어떤 것들은 다세포생 뭄이거나 혹은 큰 생물의 작은 부분인 경우도 있다. 소속의 다양성에도 불 구하고 이들을 모두 미화석으로 묶어 다루는 것이 실용적이고유용하기 때 문에 미고생물학이 탄생된 것이다. 대다수의 해양 미화석군은 그들의 골격이 방해석 (혹은 애래거나이트) 또
는 오팔린 실리카 성분으로 이루어져 있다. 때로 는 종류에 따라서 복잡한 유기질이나 모래룰 시멘트화한 형태의 각을 갖는 경우도 있으나 이 들은 해 저 퇴적물의 일부 룰 이룰 뿐이며 또한 고해양 환경의 지시자로서 역할이 미미하다. 석회질 해양 플랑크론으로는 유공충 (Foram i n if e r a) 과 코코 리 드 ( C occo l i- th op r id a ) 가 대표적 이며 규질 플랑크 돈 으 로는 규조 (D i a t oms) 와 방산 층 (Ra d i olar i a) 이 중요한 몫을 한다. 한편 대 륙붕보 다 깊은 지역의 해저에서는 유공충과 더불어 개형충 (Os t racoda) 이 대 표적 인 저생 미화석이다. 이밖에 도 석회조 (Calcareous alga e), 규질 편모충 (S i li co fl a g ella t a ) , 쌍편모 조류 ( D i n o flag e llata ) ;, 화분과 포자 (Pollen and S p ore) 등이 해양 지 잘학자등의 콴신윤 보
 CALCAREOUS
CALCAREOUS
으고 있다. 역사적 지질학 분야에 있어서 해양 미화석의 가치는 그들이 모든 지질 시대룽 대변하는 다양한 해양 환경의 퇴적물 내에 분포한다는 접 외에도 다음과 같은 강점을 지닌다. 그들의 미세한 크기, 개체수가 풍부한 집단, 광범위한 지리적 분포는 소량의 시료만으로도 충분히 칭량적인 분석이 가 능하다. 대부분의 플랑크론과 다수의 저생 미화석의 광역적인 수평 분포 는 지층의 대바 및 고지리의 복원에 대단히 유용하다. 환언하면 해양미화 석은 시준화석 및 시상화석으로서 높은 가치를 갖는다. 해양 미화석은 선캄브리아대 지층으로부터 현세에 이르는 퇴적물에 함 유되어 있으며, 모든 충서 단면에서 한두 그룹의 미화석군이 생물 충서 및 고생태 복원에 일조한다. 그림 1 은 주요 해양 미화석군의 시대적 생존 상 황을 도시한 것이다. 고생태 환경을 연구하기 위해서도 해양 미화석은 대단히 유용한데 이들 이 현존하는 다양한 해양 환경에 모두 서식하기 때문이다. 그림 2 에서 주 요 해양 환경의 범위를 볼 수 있다. 그림 1 에서 방산층, 규질 편모충, 석회질 난노플랑크론 (nannop la nkto n ), 프테로포드(pt ero p ods), 유공충의 일부 및 규조는 0~200m 의 수심에서 생 황하는 부유성 생물이며 육지로 가까와질수록 그 수가 격감한다. 이들은 과 거 지 질시대의 해양 환경의 변화, 특히 수온의 변화를 추적하게 해준다. 반
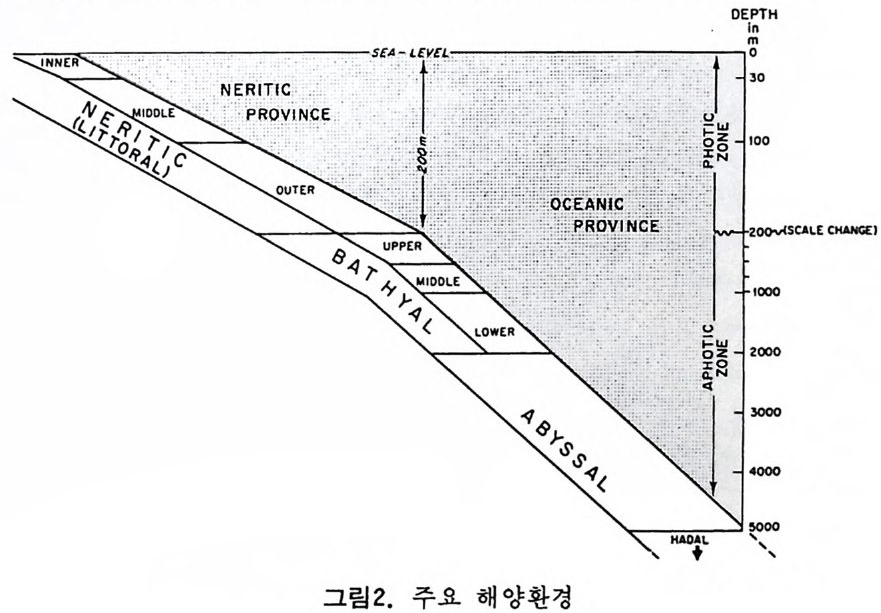 OEPTH
OEPTH
면에 개형충, 선태 동물 (Bry ozoa), 유공충이나 규조의 일부는 이동성이거 나 고착성인 저서성 생물로서 이들은 특히 수심, 저질물 및 기타 물리 • 화 학적인 여러 가지 환경요소들에 의해 결정되는 생태학적 분포 양상을 보여 줌으로써 해양 처부의 환경 변화를 추정하는 도구로 이용된다. 쌍편모 조 류 등은 그들의 생활사 중에 부유 단계와 저생 단계가 있어 특히 홍해나 흑해와 갇이 내해나 연근해의 고생태 연구에 적합하다. 코노돈트는 고생대와 초기 중생대의 지층 대비와 고해양 환경 복원에 중 요한 역할을 하며 화분과 포자는 육상 식물로부터 분리 이동된 것이지만 古氣侯의 강력한 지시자로 이용된다. 천해 지역에서 이들의 분포 양상은 육상의 기후 변화뿐만 아니라 해수의 이동에 관한 정보를 주기도 한다. 2 硏究史 인류 최초의 미화석에 관한 언급은 기원전 Herodo t us 나 St ra bo 등의 피 라밋을 만든 석회암 중의 화폐석 (Nummul it es) 에 대한 언급에서 비롯된다. 그후 화석의 유기물 기원을 밝힌 사람으로는 동양의 Chu Hs i (l227) 와 그 보다 3 세기 후에 서양의 Leonardo da V in c i(1 452-1519) 를 들수 있다 .1660 년 Anto n ie van Leeuwenhoek 에 의 해 현미 정 이 발명 된 후 비 로소 체 계 적 인 미고생물학이 태동되었다. 16 세기에 이미 유공충은 박물학자들의 관심을 끌었으며 후에 Alcid e d'Orb ig n y(l 802-1875) 는 이들을 두족류에 넣어 자세 히 분류하고 많은 연구 업적을 남겼다. 그 중에는 유공충을 생물충서학적 도 구로 이용한 것도 포함된다. 1835 년 Felix Du j ar di n 은 유공충을 두족류로부 터 분리하여 Rh i zo p여요그룹으로 새로운 분류륭 시도하였으며 많은 사람들 이 이룰 인정 하였다. C.G.Ehrenber g(l 795-1876) 는 규질 편모충. 에브리 디 아. 코코리드. 디스코스타. 쌍편모 조류 등의 원생생물들과 더불어 다 수의 방산충, 규조. 유공충 등을 새로이 발전. 기재한 탁월한 학자였다. 19 세기 후반에는 많은 미화석 그룹에 대한 기재학이 크게 발전하였으며 특 히 N.C.W illiam son, W. K. Parker, T. R. Jon es, W. B. Carpe n te r , H. B. Brady, C. D.Sherborne 등 소위 〈영국 학파〉의 공헌이 괄목할 만하다. 같은 시기에 방산충은 E.Haeckel(1862-1887) 에 의해서. 개형충은 Sars (1866) 에 의해 기본 분류학적 연구가 시행되었으며. 1875 년부터 Schmi dt Dia t o m A t las 가 발간되가시작하는 동 미화석류의 활발한 기재 0~ 구가 진행 되었다. 앞서 영국 학파에 의해 충서학적인 가치가 무시되었던 유공충도 이 시
기에 다시 재연구가 시작되였다. 19 세 기 후반의 이 러 한 왕성 한 미 화석 연구 붕은 H.M S. Challeng e r 호의 1873 년에서 1876 년에 걷찬 항해가 정정적인 자국제가 되었던 것이다 .1884 년 H.B.Brad y는 찰렌저호의 항해 기간 동안 채집한 유공충의 모노그라프 룰 발간했으며 이 책은 현재도 신제삼계와 현생 유공충 연구에 기본 자료 가 되고 있다. 미고생물학 연구의 두번째 혹은 분석적 발전 단계는 생물의 구조 멍 형 태학적 연구로부터 분류의 natu ral s y s t em 으로의 접근과 또한 생물 층서 학적인 적용에 관한 연구로서 대변된다. 해양 미고생울학에 대한 관심의 부황과 심해저에 대한 시추작업의 시작 이 미고생물학·연구의 세번째 또는 개관적인 미화석의 연구단계로 비약시 킨 주원인이 되었다. 제 2 차세계대전 이후 특히 1950 년과 1960 년대의 해 양 탐사와 피스톤 코어링 기술의 발달은 해양 미고생물학의 발전에 크게 기여를 하였다 . 해양 탐사는 1965 년의 JO IDES Pro g ram 과 그 후계 사업인 Deep Sea Drill in g Proje c t (DS DP) 가 1968 년 시 작됨으로써 해양학 전반에 혁 신적인 영향을 끼치게 되었다. DSDP 의 일환으로 해양 시추선인 D/V Glomar Challen g er 호는 최근까지
 그림 3. D/V Glomar Challen g er 號
그림 3. D/V Glomar Challen g er 號
북국해륭 제외한 전 해양의 수백사 1 지섬에서 사 추륭 실시하 었 나. 그 상 과 수심 1~ 4km 인 해처로부터 수만 m 에 당하는 중기 쥬라계 이후의 시추 코 아를 얻음으로써 1 억 5 천만년 동안의 생물의 진화와 해양 환경의 변천사 롤 연구할 수 있게 되었다. 이로써 해양 미고생물학은 해저의 신비룽풀수 있는 우한한 지삭의 보고가 되었다. 3 연구의 목적과응용 해양 퇴적물내에는 대개의 경우 여러 종류의 미화석군이 함유되어 있는 데 그 종류는 퇴적물의 생성시기와 퇴적 환경에 따라 다양하게 경정된다. 따라서 미화석군은 지질학자들이 암석의 연대와 그것이 생성될 당시의 수 십 수온, 영도 동음 알려고 할 때 신속하고도 신빙성 있는 대답음 주게 된 다. 각종의 지질 연구소, 석유회사나 석탄회사와 같은 기관에서는 한 팀의 미고생물학자들을 고용하여 그들이 다루게 되는 지층과 암석에 대한 정보 륭 얻는다. 이러한 미고생물학의 상업적 측면이 이 분야를 크게 발전시키 는 경정적인 역할을 한 것은 물론이다. 그러나 다소 철학적이고 사회학적 인 측면이 있어서 현재 우리 지구상의 생태계의 발달과 안정에 관한 이해 는 미화석의 기록으로부터 추론해 낸 것이다. 특히 많은 마화석 종류는 먹 이 사슬의 가장 기저가 되거나 그 부근에 속하는 것들아 많가 때문에 더욱 그러하다. 이 밖에 생물 진화에 대한 비일도 미화석군의 연구를 통하여 벗겨지고 있으며 특히 지구 역사의 3/4 이상을 차지하는 선캄브리아대의 초기 유기 물의 진화에 관한 주요 정보를 얻을 수 있다는 접에서 주목을 받고 있다. 미고생물학의 상업 적 응용은 1 차세계대전을 즈음하여 발생한 석유 수 요의 급증으로 인하여 크게 자국되었으며 유공층 군집이 지층의 대비를 위 한 도구로서 사용되기 시작하였다. 이로써 분류학이 주가 되던 미화석 연 구가 충서학적으로 방향 전환아 시작된 것이다. 석유 탐사를 위하여 전세계적으로 지표나 지하의 지질 조사에 미고생물 학이 보편적으로 환용되었으며 이에 맞추어 대학에서도 교과과정을조정하 여 지구과학의 한 연구 분야로 발전하였다. 1911 년 Au g us t ana 대학의 J. A. Udden 교수에 의해서 미화석이 유정의 시추공간의 지층 대비와 연대 측정 에서의 중요성이 강조된 이후 1 차 세계대전 기간중 미국의 미주리 광산대
학. 컹럼비아대학 및 텍사 스 대학 등에서 이고생물학이 정규 과목으로 개 설되었다. 1924 년 하바드대학에 부설된 유공충 연구룽 위한 Cuslunan Labora t or y가 설립되면서 새로운 전기룽 마련한 미고생물학은 1924 년 스텐퍼드대학에서 처음으로 개형충 연구가 시작되었다 . 이어서 대공황시대에 시카고대학에 미고생뭄학이 도입되었다 . 2 차세계대전까지는 주로 석유 탐사와 관련된 생뭄 층서학적 대비가 주요 목적이었다고 할 수 있으며 반연에 대전이 끝 난 후의 미고생물학은 고생태와 고수심도의 복원으로 그 연구 방향이 바뀌 었다 .
제 2 장 海洋微古生物學의 기본원리 (Basic C oncep ts in Marin e Mi cr op a leonto l og y) 1 生物層序學 (Bio s tr a ti gr aph y ) 1 • 1 생존대 (Bio z ones) 와 분대 (Zonati on ) 층서학의 한 분과인 생물충서학은 좀온 의미로서는 화석과 화석을 포함 하는 암석의 공간적 분포와 시간적 선후관계를 다루는 연구분야이다. 충서 학은 매우 주관적인 연구 방법을 채택하고 있으므로 층서학적 단위의 명명 과 분류의 통일을 기하기 위해서 많은 학자들의 의견을 최대한 반영하는 규약이 필요하게 되었다. 1961 년 발표된 “미국 충서학 명명위원회 (Ame ri can Commi ss io n on St ra tig r aph i c Nomencla t ure)” 의 충서학적인 명명에 관한 규약 (Code on St ra tig r aph ic Nomencla t ure) 이 대표적인 것으로 여기에는 명명규약 뿐만 아니라 중요한 과학적 개념들을 포함하고 있다. 충서학의 기본이 되는 원리는 바로 “지층누중의 법칙 (Su pe r po s iti on)” 과 地史的 사건의 `' 연속성 (ordi na l p ro g ress i on) ” 으로서 가장 널리 이용되는 것 이다. 이 원리에 입각하여 지층의 충서 (s t ra tig ra ph y)와地年代(g eochronol o gy)의 골격이 형성된다. 충서기록을 구분하는 데는 암석단위와 화석의 산출에 의한 충서단위가 있다. 이룽 생물충서 단위 (bios tr a ti gr aph i c unit ) 라고 하며 이는 암체를 화석 군의 출현, 최대빈도 또는 소멸 등 고생물학적인 기준으로 세분한 것이다. 생물의 진화현상온 특정한 시간내에 집적된 퇴적물의 양으로 대변되는 지 층 분대의 툴을 세울 수 있게 해 준다. 생물충서단위의 기본적인 단위는
帶 (zone) 이며, 추상적인 대의 하나로 생존대 (bi ozone) 가 있는데 이 생존대 는 어느 화석종이 생존했던 기간 중 퇴적된 범세계적인 지층을 가리킨다. 생존대는 특성에 따라서 보통 세 가지 종류로구분된다. CD assemblage zone : 지층내에 함유된 전체 화석군 중에서 특정한 그룹 에 의해 정의된 지층군. ® rang e zone : 지층내에 함유된 전체 화 석군 중에서 선별된 멸 종류의 충서 범위로 결정된 지층군. @acme zone : 화석군의 상호 관련이나 충서 범위에 관계없이 어떤 종류 둘의 정량적인 존재량에 의해 정의된 지층군. acme zone 은 다른 두 종류보다는 정 성 적 으로 중요성 이 낮으며 대부분의 생충서 연구를 위해 이용되는 부유성 미화석군의 생존대는 rang e zone 으로 서 다음과 갇온 종류가 있다 @ 1axon rang e zone- 어느 종이나 속의 화석의 수평적 및 수직적 전체 적인 존재 범위를 대표하는 지층군. @ concurrent rang e zone- 두 가지 이상의 분류군이 겹쳐져서 분포하 는 범위로서 결정된 지층군. © opp e l zone- 특별히 선정된 화석군에 의해서 결정된 rar ge wne 의 일 종. @ lin e ag e zone (ph y lo zone) -어떤 종류의 계통 발생이나 계통선을 대표 하는 그룹을 포함하는 지층군. @ acme zone- 어떤 분류 단위들(종, 속 등)의 최대 분포로서 정해진 지층군. CD Inte r val zone- 두 개의 뚜렷한 생충서 층준 사이로 정해진 zone. 이상의 생충서 생존대에 관한 깊은 논의는 1972 년 Mon t real 에서 열렸던 제 24 차 국제지 질학총회 (I. G.C.) 의 Repo r t 5 인 “Prelim ina ry Repo r t on Bio - str a tig r aph i c Un it s” 를 참고하면 된다. 고생물군은 시공간적으로 결코 동일 성 있게 지층내에 분포되지 않기 때문에 생충서적 생존대 역시 지속적, 광 역적으로 설정되지 않는다. 해양의 생물충서 분야에서 가장 잘 알려진 분대 (zona ti on) 의 예는 석회 질 및 규질 플랑크론을 이용한 중생대와 신생대 지충일 것이다. 2 차대전 을 전후로 한 남서부 소련과 칼리브 해역 그리고 1960 년 후반의 DSDP 팀 사의 철과 적어도 新제 3 기예 속하는 지층에 대한 지구 전역의 광역적인 생충서 분대가 가농하게 되었다. 이로써 방사성 동위원소의 자연 붕괴로
서 계산되는 절대 시간표와 더불어 해양 영역의 ti m e - sca le 이 확립되었다. 현재로 부터 50. 0 만년 전 사이의 시 간표는 고지 자기의 시간표 (Cox, 1969) 에 방사성 절대 연령 자료가 정량적인 값들을 주 고 이러한 값들응 해저 코 아로부터 얻어 낸 생충서학적 변화가 조정함으로저 안 들 어진다 . 500 만년 전부터 2 천 5 백만 년 전 사이의 시간표는 생충서학적 지사와 K-Ar 연대 자료 , 고 지자기의 해저 자료들이 적절하게 비교 대비됨으로 써 보완된다. 생층서에서는 특히 방산층이 중요하여 이 로부터 다 른 미화 석군과의 간접적 인 비교가 가능하다. 2 천 5 백만년 이전은 생충서 자료와 지자기의 반전 자료가 K-Ar 연대 값 에 조절되어 시간표가 작성된다 . K-Ar 자료의 큰 오차 때문에 정확성은 오 래된 시대일수록 떨어지지안 중생대의 시간표의 작성을 눈앞에 보고 있다. 1 • 2 충서 대 비 (St ra ti gra p hi c Correlati on i 서로 격리된 충서 단면이나 시추 코아들간의 대비륭 위해서 가장 널리 쓰이는 방법은 화석군의 수직 분포에 따라 절정된 충서 범위 (st ra t igr ap h ic ran g es) 와 생존대 (zone) 라고 할 수 있다. 충서 단면들은 암층의 구조적 양상, 지형, 암상 및 화석의 내용 등이 서로 다르므로 지층에 대한 세밀 한 기재와 더불어 각 화석종의 층서 범위가 정밀하게 관찰 기록되어야 한 다. 단일종의 충서 범위가 여러 개의 충서 단면에서 일치한다면 이러한 화 석종의 최초나 최후의 출현이 대략적인 지질시대의 동일성을 지시한다고 보기 때문에 대비된다고 할 수 있다. 그러나 이러한 화석종이 장소에 따 라 멸종시기가 다르거나 이주현상 때문에 반드시 같은 시기를 지시한다고 볼 수 없는 경우도 있기 때문에 주의가 필요하다. 따라서 균일한 환경조건이 오래 계속된 특수한 조건에서는 전성 기 (acme) 에 근거해서 대비하는 것이 정확을 기하는데 큰 도움을 주기도 한다. 또한 단일종보다는 둘 이상의 종들에 근거한 대비가 훨씬 안전하며 대부분의 생충서 적 단위가 생물군집대 (assemblag e zone) 인 까닭이 여기에 있다. 한 지역에서 산출된 화석군집은 충서관계가 잘 연구된 지역의 표준군집 (s t라 1dard assemblag es) 과 종 대비를 할 수 있다. 두 군집의 비교는 공통종 의 %를 쓰거나 특별히 고안된 지표를 이용한다. 표준화석 (ind ex fo ss i ls) 은 넓은 지 리 적분포, 생태학적으로 강한 내성, 풍부 한 산출 급속한 진화속도, 독특한 형태 둥의 조건을 갖춘 화석으로 특히 미화석에는 많은 종류가 포함된다 . 어떤 경우에는 형태적으로 나타난 변
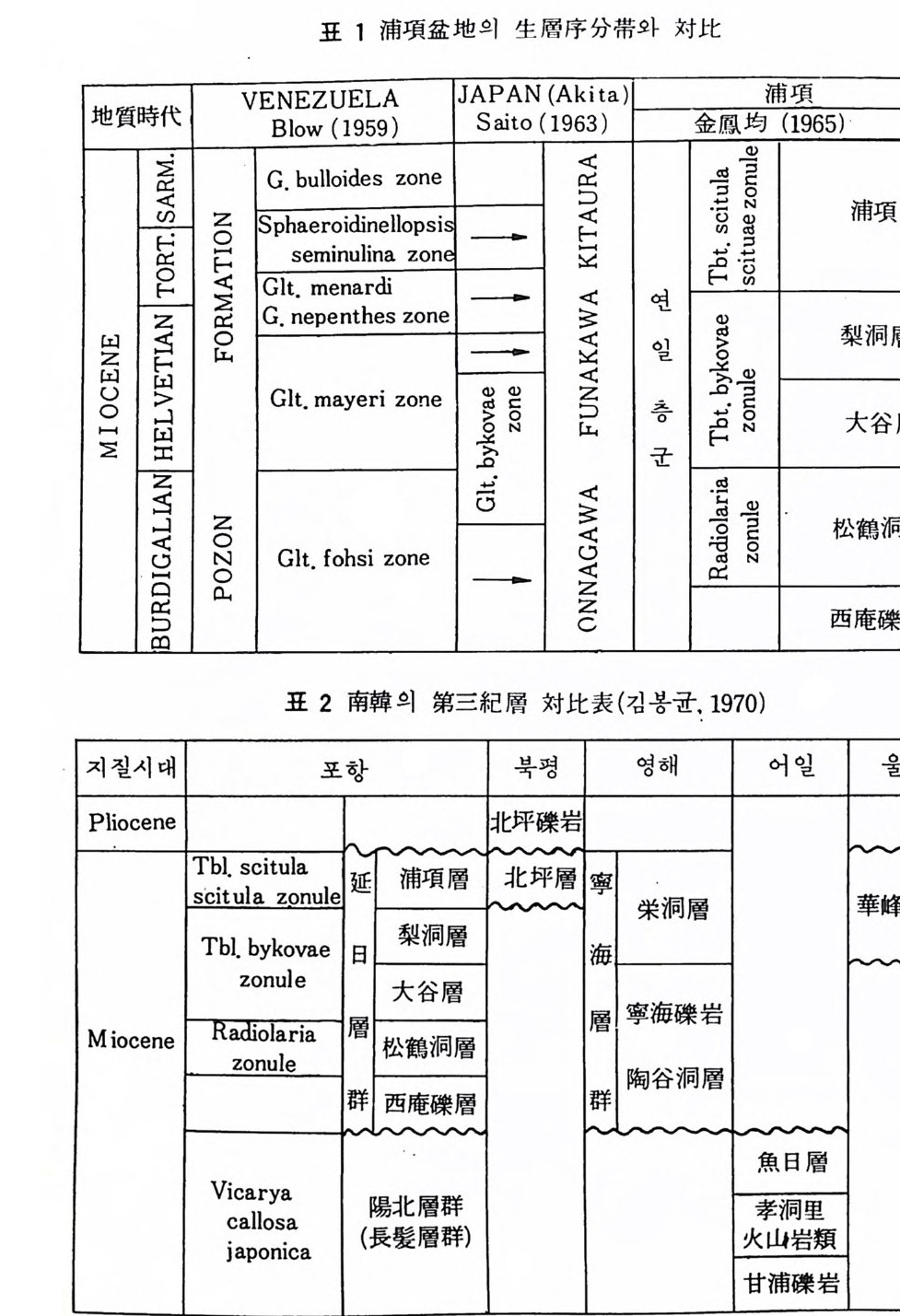 표 1 浦項盆地의 生層序分帝와 챠.I:t
표 1 浦項盆地의 生層序分帝와 챠.I:t
화가 시간적인 동시성을 보여주어 대비에 이용되는 경우가 있다.예컨대 부 유성 유공충의 일종인 Globorot a lia mena 油.i의 말린 방향의 변화 (dex t ral s i n i s t ral) 는 해저의 시추 코아에서 훌륭한 대비의 도구로 사용될 수도 있다. 이 밖에도 생태학적인 해석에 의해 지층 대비가 가능한 경우도 있다.즉 일정한 고환경 아래에서는 특징적인 암상과 화석상이 형성된다는 원리를 이용하여 한 지역의 충서 내용을 면밀하게 관찰한다. 그림 1 에는 하나의 가상적인 해침과 그 뒤에 잇달아 해퇴가 일어난 지역의 충서단면이 도시 되어 있다. 이 가상적인 지층단위가 퇴적되는 동안 서쪽이 육지이고 동쪽
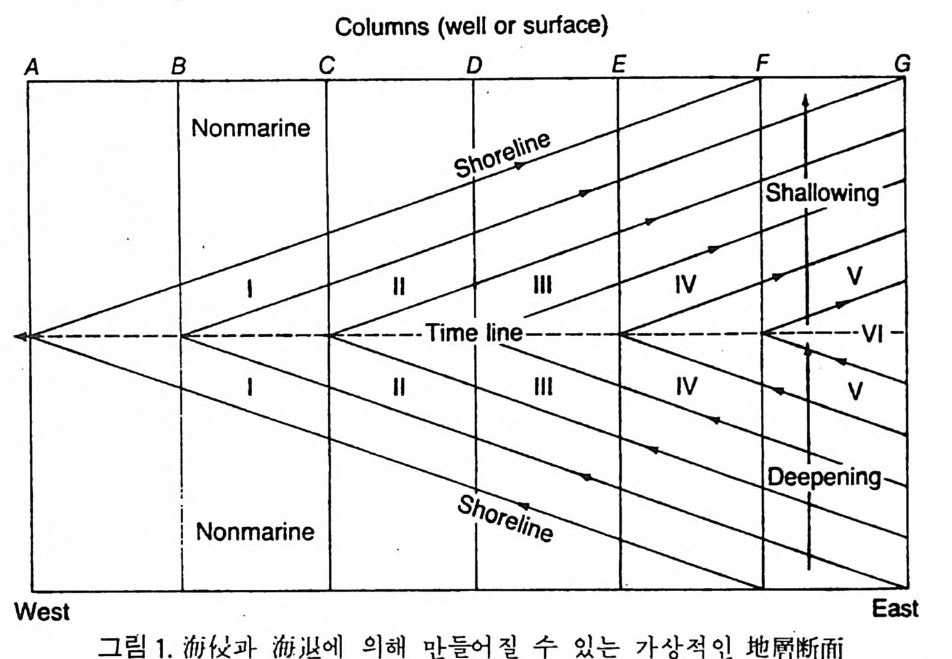 Columns (well or surf ac e)
Columns (well or surf ac e)
에 바다가 있었다면 해안선은 처음에는(아랫부분) 서쪽으로 이동하다가 나 중에는(윗부분) 다시 동쪽으로 후퇴하였다. 지충의 각 부분은 수심이나 기 타 다른 환경요소의 차이로 각각 상이한 암상과 생물상을 형성하여 6 개의 해성 상들은 각각 해안선에 나란하게 발달된다. 각 암상에서 서쪽 한계를 나타내는 지점이 바로 바다가 서쪽으로 이동한 최대 해침기물 지시한다. 모든 상에 서 나타난 이들 접을 연결한 선이 단일 시간을 의미하는 대비의 선이 된다. Shaw(1964) 는 여러 지역의 충서단면에서 발견되는 화석군집과 기타 다 론 충서자료들을 종합적으로 통계처리하여 이들울 대비하는 방안을 재시 하였다. 두 지역을 대비하려 할 때 각각을 X-Y 축에 단면의 길이를 수직
 • • - -- -- 노。& ' ••, ••• ♦• , • , • r-~~++: .♦-
• • - -- -- 노。& ' ••, ••• ♦• , • , • r-~~++: .♦-
거리로 표시하고 산출하는 화석군의 출현과 소멸의 위치를 표시함으로써 여러 가지 지질학적인 문제들을 추정할 수 있다. (Raup & St an ley (1977 ) 와 Mi ller (1977) 참조) . 이때 대비에 표준이 되는 단연을 Comp os it e · S t an dard Refe r ence Sect ion (CSRS) 이라고 하며 이와 대비될 새로운 단면들과 직각좌표에 도시함으 로써 Lin e of Correla ti on(LOC) 룽 구할 수 있게 된다. LOC 는 시간의 동일 성, 퇴적룹, 화석군의 분포범위, 결층의 존재 여부를 밝히는 데 이용된다 . 이 방법은 두껍고 연속적인 노두가 발달한 지역에서 유리하며 최근에는 油 井 탐사에 많이 이용되고 있다. 이들 자료로부터 우리는 시간一충서 관계 룰 파악할 수 있을 뿐만 아니라 석유지질학에 중요한 과제인 고생물, 중서 및 고환경에 관한 정보를 얻을 수 있다(그림 2, 그림 3 참조). 2 古生態孝 ( Paleoecolog y) 2 • 1 해양생태학 (Marin e Ecosy st e m) 현 재 바다의 생태계는 대부분의 고생물학자들이 과거 지질시대의 퇴적 몽 의 앙상과 생물상을 해석하는 데 모델로 생각하고 있다. 이러한 동일과 정성에 입각한 비교는 중요한 장애 요소도 포함하고 있는데 과거 지질시 대 중 캄브리아기 이래로 기후가 현재보다는 대개 온난했다는 접과 과거 의 수륙분포가 오늘과는 달리 많은 부분이 양온 해수에 잠겨 있었다는 점 이다.
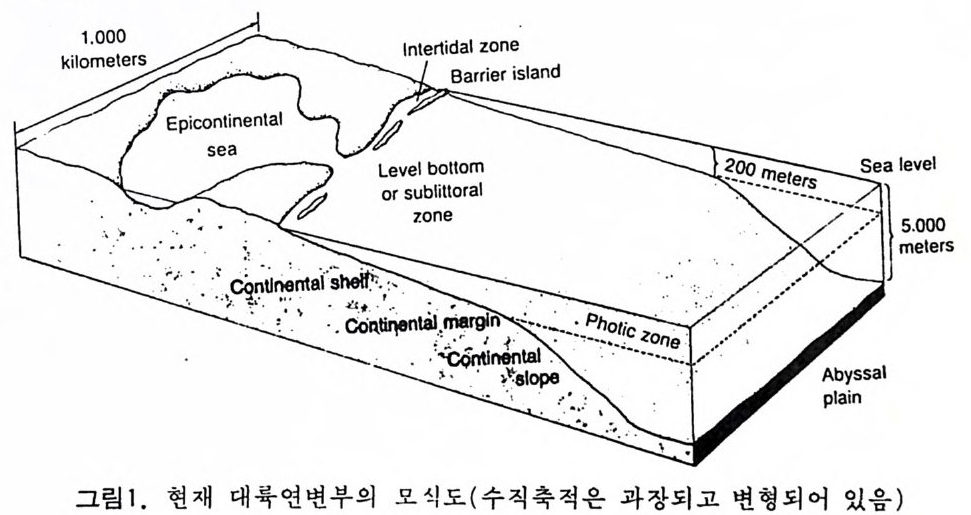 1.0 0 0
1.0 0 0
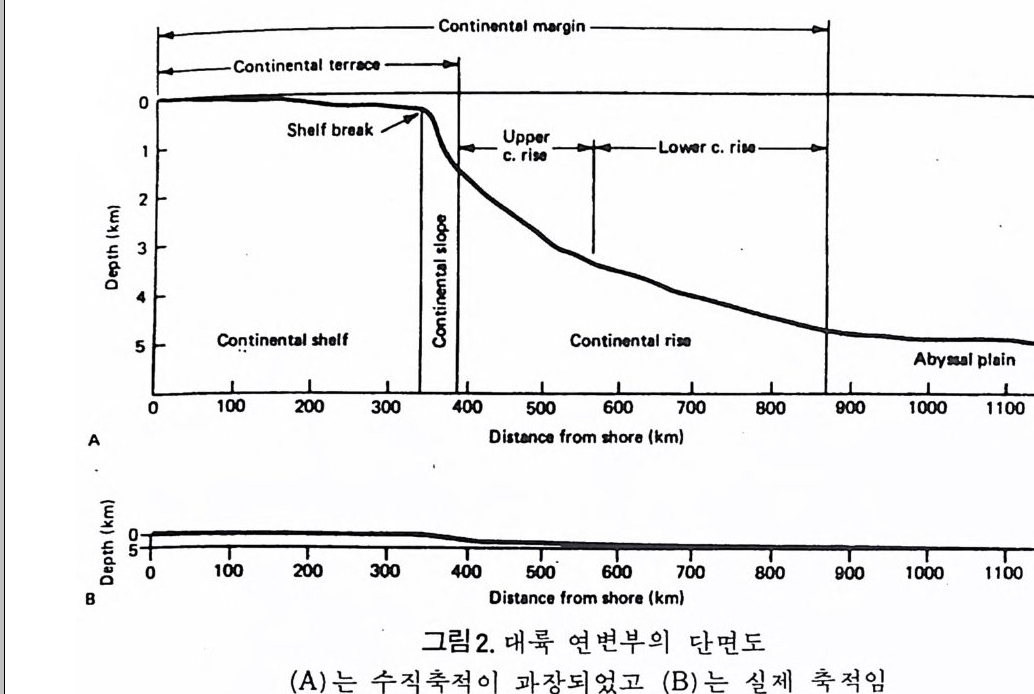 仁: ~n ti nen t ■ I te rr..,. Con1in o n111 margi n
仁: ~n ti nen t ■ I te rr..,. Con1in o n111 margi n
고생태학에서는 특히 대륙주변부의 고환경이 중요한 지사의 열쇠를 쥐 고 있으므로 현재 이 부분에 대한 연구에 큰 비중을 두고 있다. 지역적인 동물군을 연구하는 고생태학에서는 일차적으로 저서생물에 관 심을 둔다. 까닭은 이들이 화석으로서 퇴적물내에 안전하게 보존될 수 있 으며 사후의 이동(p os t mor t em t rans po r t)이 적으므로 고생물의 생태환경을 그대로 추정하는데 용이하기 때문이다. 반면 회유성 종들은 죽은 뒤 상당 히 이동하여 바닥에 낙하하므로 보존도 어렵고 원래 그들이 살던 환경과 는 상이한 곳에 퇴적될 수도 있으므로 시상화석으로서의 가치가 적다. 따라서 미화석 군집울 고생태학적으로 바르게 해석하기 위해서는 이들 이 어떤 경로의 사후 운반을 거친 것인가를 먼저 확실하게 추적해야 할 펄 요가있다. 화석군의 사후운반은그림 3 에서와같이 나누어 추론할수있 다. 단 이들의 식별은 상당히 숙달된 안목이 필요하다. 그람 4 는 바다에서의 먹이순환(f ood c y cle) 을 단순화해서 나타낸 것이다. 현재 바다에서의 주요 생산자는 단세포부유성 식물인 규조(di a t oms) 와 쌍 편모조류(dinoflag ella t es) 등이 포함된다. 원양에서는 부유성 식물이 광합 성을 하는 유일한 생산자이지만 두광대(p ho ti c zone) 에 속하는 대륙봉상에서
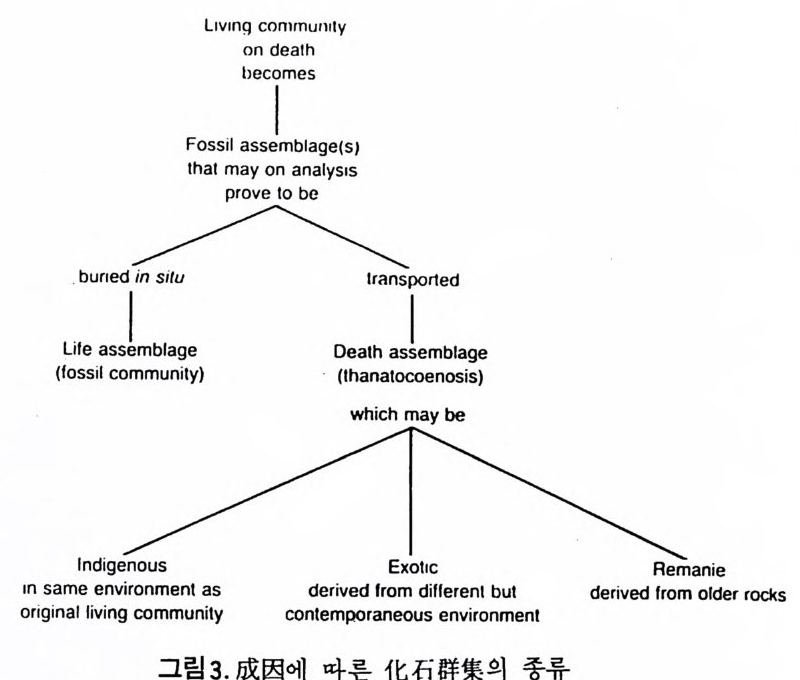 Llvrn~ cIomm urnty
Llvrn~ cIomm urnty
는 해초등이 생산자에 가담한다. 바다에서의 생물량 피라밋 (bio m ass pyra m- i d) 은 대개 뒤집힌 모습이다. 해양 미고생물학에서는 이들 일차 생산자들 중 각질인 그룹들이 화석 기록에 남기 때문에 연구 대상이 된다. 해양 생태계는 한 개체의생리기구와 같이 海樓생물과 주변 환경인물리· 화학적 인자들이 만들어내는 하나의 계를 말한다. 해양 생태계의 형성은 수괴의 공간적 분포, 시간적 제어, 먹이 사술과 갇은 종간의 에너지 교환, 그리고 화학적 순환과 갇은 작용이 얽혀서 이루어진다. 생태계의 구성은 오랜 시간을 거친 생물의 환경에 대한 적응의 결과인 진화의 산물이다. 또한 동시에 구성원들의 공존상태는 각 종들의 생활습 성이 환경에 적옹하여 살아남게 된 공간적인 배열을 결정한 것이다. · 바다 의 두광대에서 각 생물군들의 수직적 충상분포가 좋은 예이다. 해양 생물 은 활동시간이 지속적이라기보다는 어떤 독특한 주기성이 있으며 또한종 둘 간에는 상호 밀접한 시간의 틀로 짜여진 생태계를 이루고 있다.또한 생 태계는 에너지와 물질의 교환이 환경 요소와 생물계 사이에서 일어나고 있 다. 예를 들어 海樓 미생물들의 각의 성분인 CaCOs 는 바다에서의 C 아의 순 환에 가장 주요한 역할을 한다. 이처럼 해양 생태계에는 많은 연구과제가 될 복잡한 생태현상이 일어나고 있다. 앞서 말한대로 현재 해양의 생태계
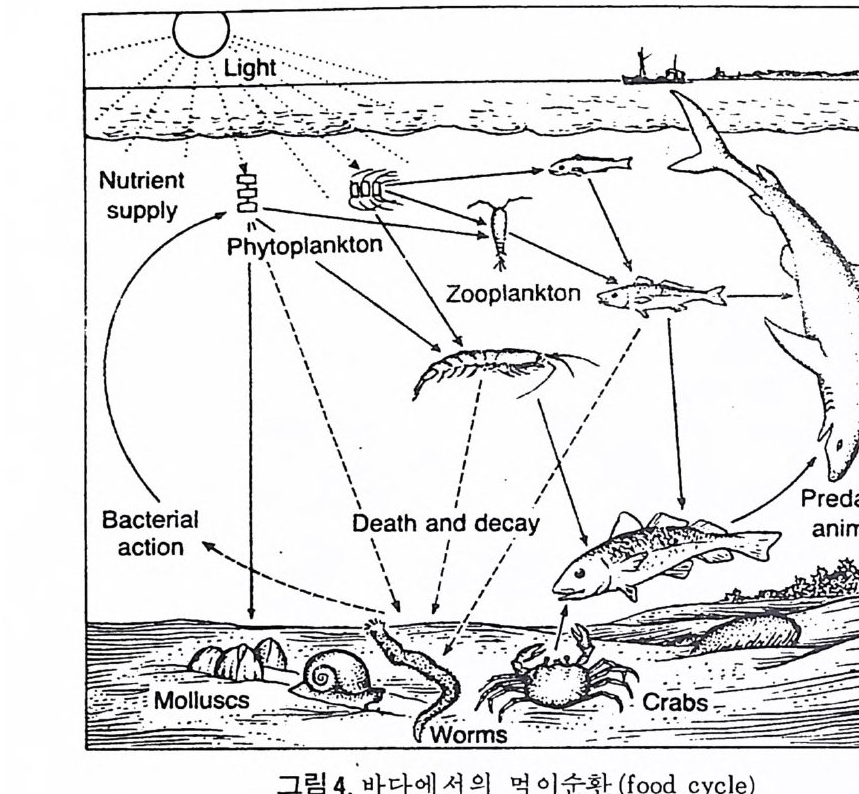 ····•••.•· •. -• .-•-· •. : .. \-- .-. ~一 . ./-- . L 혼ig .? .h•• t. .·.< . . ::·· ... .. .
····•••.•· •. -• .-•-· •. : .. \-- .-. ~一 . ./-- . L 혼ig .? .h•• t. .·.< . . ::·· ... .. .
에 관한 모든 연구자료는 고생태학에 필수 불가결한 것이다. 그러나 〈현 재는 과거를 아는 열쇠〉임에는 큰 오류가 없으나 현재 해양환경과 현생생 물과의 관계가 그대로 과거의 해양과 고생물의 관계로 비교 적용되는 것 은 아님이 속속 드러나고 있다. 이유는 알려진 대로 고해양의 제한요소둘 과 수륙분포 등이 지구가 진화함에 따라 변화해 왔으며 생태학의 기본논 리인 원인-결과론에서 적절한 원인을 충분하게 탐지할 수 없는 지층과 화 석의 기록의 불완전성 때문이라고 하겠다. 그럼에도 불구하고 해양 고생태학 혹은 고해양학(p alrooceano g ra p h y)은 해 양 미고생물학 분야에서 가장 독창적이고 빠른 속도로 발전해 가는 분과 중의 하나임에는 의문이 없다. 2 • 2 고생물지리 (Paleobio g eo g ra ph y ) 해수의 순환은 동력학적으로 해저지형과 기후에 의해 영향을 받는다. 또 한 오랜 지질시대를 통하여 암권(Iit hos p here) 은 끊임없이 판구조론 (Pla t e
T ec t on i cs) 에 입각한 운동을 해왔으므로 해저의 지형변화는 해수의 운동뿐 안 아니라 해양 생태계에도 변화의 역사를 부여하였다. 생물학과 해양학에 대한 지질학의 중요성이 가장 뚜렷하게 부각되는 분 야가 생물지리학이란 점은 충분히 타당하다. 최근 지구과학계의 혁신으로 불리는 판구조론의 대두는 오랫동안 알려져 왔던 생물지리에 관한 많은동 설들을 재고케 하였다. 현재와 같은 해양의 윤곽이 처음으로 형성되기 시 작한 시간으로 추정되는 2 억년전 이후의 해양의 역사는 절대연대가 아직 도 불 확실 함에도 불구하고 상대적인 충서와 상호관계는 찰 알려져 있다. Pan g a ea 를 거쳐 Gondwana 대륙의 분할로 이어지는 암권의 운동은 현재 지역 벌 시간별로 그 규모와 함께 잘 알려져 있다 . 생지리학적 분포양상을 해저의 미화석의 시공간적 분포에 적용하는 데는 사후이동과 보존 조건이 주요 제한요소이 다. 석회질과 규질의 미화석 골격의 차별적인 용해현상은 현재 중요한 연구 테마로 등장했으며 (Berge r,1974) 심해저 퇴적물에 포함된 미화석군집의 성 분과 양이 어떤 경로를 동해 형성된 것인가 하는 문제들이 연구되고 있다. 미화석군의 해저분포는 주로 유공충, 방산층, 코코리드 등에 의해 기본적 인 자료가 모아지고 이로부터 해양의 발달사가 고생태, 고지리, 고해양학 적으로 추정된다. 산생대 석회질 난노플랑크톤이나 부유성 유공층군집의 분포는 시대별로 대서양의 고환경 변화를 추적하는 도구로 사용되었다. (Ha q & 外, 1977). 해양의 생태요소들의 변화 중 위도에 따론 해수의 고수온의 복원은 이들 미생물군집의 생태학적 선호가 신생대 기간 중 비교적 안정했었다는 가정 울 전제로 한다. 古제 3 기의 생물군집에 의한 고수온의 변화 경향은 또한 이 시대의 부유성 화석들의 산소 동위원소의 분석 결과 및 육상의 식물의 변화와 함께 대비되고 있으며 이로써 좀 더 신빙성있는 결과가 얻어지고 있다. 미화석군이 퇴적물 내에 퇴적된 후 받게 되는 diag enes i s 에 의한 변질과 심해성 석회암 및 쳐어트의 생성에 관한 연구도 활발하다. 이러한 변화는 미화석의 보존상태를 다양하게 하며 심해처의 음향파 충서학 (acous ti c str a tig ra p h y)을 가능케 하는 반사면들을 만들기도 한다. 이 분야에서는 미고 생물학자와 지구물리학자의 공동 연구가 요구된다. 고해양학에서 미고생물학의 역할은 지구물리학과의 결합을 유도하였다. 미화석군은 고환경의 지시자일 뿐만 아니라 생물충서의 도구이기도 하다.
 LRaki f_et_ . ,V.B ,.a-a-llziek /ya -l -?· • ., I I ’ ’一-·----- ,J>I禪曺 益 ch Gorda Plat e
LRaki f_et_ . ,V.B ,.a-a-llziek /ya -l -?· • ., I I ’ ’一-·----- ,J>I禪曺 益 ch Gorda Plat e
 집
집
과거 지질시대의 분포양상에 대한 해석은 궁극적으로 고지리와 고심도의 복원에 의존한다. 따라서 판구조 운동이라는 공간적인 변화 자료가 미화석 연구에 필요불 가결하게 된다. 현재 여러 가지 미화석군의 분포양상에 의해서 해수의 순 환이나 퇴적작용의 역사가 밝혀지고 있다. 3 進化學 (Evolu ti on) 3· 1 진화이론의 발전 18 세기 말경 지질학자들은 지층을 분대하고 선후관계를 정하는 데 화석 이 대단히 유용하다는 사실을 알게 되었다 0/v. Smi th) . 그 후 C.L y ell 은 1830 년 『地質學原理』에서지층은·연속적으로 형성되었으므로. 그 속에 함유 되어 있는 고생물군도 접전적으로 이행한다는 사실을 명백히 했다. 이러한 생각들은 생물의 종이 불변이라고 믿는 18 세기까지의 고칭관념을 바꾸는 데 중요한 역 할을 하여 1859 년 C. Da rwi n 의 자연도태설을 . 낳게 하였다. 과거 지질시대의 생뭉의 역사는 지층에 함유되어 있는 고생물의 유해나 그 흔적, 죽 화석을 직접 연구함으로써 규명이 가능하다. 지각의 표층부에 노출되어 있는 지층으로부터 인류는 많은 종류의 화석을 발견하였으며 앞 으로도 새로운 화석의 발굴을 위한 노력은 계속될 것이다. 그러나 이들이 한때 지구상에 서식하던 고생물이었다는 사실을 인식하 게 된 것은 그리 오래지 않다. 화석은 고대로부터 다양하게 이해되어 왔 는데 창조주의 실패작. 자연의 장난, 악마의 흉계라는 생각에서 노아의 홍 수와 갇은 주기적인 재앙의 산물이라는 생각까지 당시 우세하던 법칙이나 사상에 따라 편리하게 해석되었다. 지질학의 궁극적 목표는 지구의 역사(地史)불 복원하는 일이므로 많은 학자들이 지구의 물리적 역사와 더불어 생물의 변천에 관한 신비를 풀기 에 여념이 없다. 역사학이란 과거의 사실을 구체적으로 추적해야 되기 때 문에 무엇이, 언제, 어디서라는 명제를 해결해야 하며 또한자연법칙은우 리에게 어떤 현상의 가능성을 제시해 준다. 따라서 생물학의 법칙과 역사 학의 과제들이 고생몽학과 같은 자연사학을 탄생시켜서 생물이 〈어떻게〉 변천해 왔는가륭 풀어 주는 임무룰 갖게 되었다. 방사성 물질의 자연 붕괴 현상과 생웅의 진화 현상은 지질학적 시간표를 작성하는데 기본이 되는두 가지 자연 현상이다. 전자는 칙접적으로시간을 측정하는데 이용되며 후자
는 상대적인 시대의 순서를 정해 주는 원리이다. 원자력 시계의 기본원리 는 19 세기 말에 이미 발견되었으나 이것을 암석과 지층의 연대 측정에 응 용한 것은 20 세기 초 (1907) 에 불과하다. 반면 화석을 지층의 상대적인 층 서 를 확립 하는데 활용한 것 은 18 세 기 말 (W illi a m Smi th : 1796) 로 거슬러 울 라간다. 지질학적 자료들이 쌓여감에 따라 생물의 변천에 관한 설마리가 풀리기 시작했고 생물의 진화이론을 발전시키는데 적지 않은 공헌을 하게 되었다. 그러면 화석기록을 읽음으로써 얻을 수 있는 생물의 진화설을 지지하는 여 러 가지 현상들을 살펴보기로 하자. 생물의 진화 연구에 있어 서 고생물학의 역 할은 큰 규모의 증거를 제시 하는 것이다. 여기에는 계통발생의 관계와 그 속도, 전체적인 변화의 경 향과 양상에 관한 것들이 포함된다. 이러한 현상들은 대전화 (macroevolu tion ) 또는 종간전화 (tra nssp e cif ic evoluti on ) 라고도 불린다. 화석기록의 불완전성 때문에 고생물학자들은 개체집단이나 종내에서 일 어나는 소규모의 변화물 발견하기가 매우 어렵다. 반면에 생물학에서는 소 진화를 다루기가 훨씬 쉬운 대신 관찰할 수 있는 시간이 매우 짧은 결점이 있다. 따라서 생물학적 관찰은 생물역사의 일부, 즉 현세와 제 4 기 정도 까지 거슬러 울라갈 수 있을 뿐이다. 종에 대한 화석의 기록이 매우 불완전하기 때문에 대진화의 연구는대부 분이 속이나과의 수준에서 이루어진다. 속이나 과에 대한 화석의 자료 는 대진화적으로 의미있는 분석을 하기에 충분하지만 어느 강이나문에 속 하는 속과 과가 다른 강이나 문예 속하는 속과 과와 같은 정도로 비교될 수 없다는 문제점을 안고 있다. 화석 기록에서 얻어지는 생물의 진화와 관련되는 대진화적 현상들을 알 아보자. 많은 고생물들은 긴 지구의 역사동안 변천되어 오는 과정에서 멸 종 (ex ti nc ti on) 이라는 운명을 감수해야만 했다. 고생물이 급격한 환경 변화 를 충분히 극복하여 잔존하기 위해서는 그 종내에 충분한 유전인자의 변이 가 존재하여 새로운 환경에 적응할 개체를 산출할 수 있든가 아니면 그런 인자의 개체가 죽시 나타나야 한다. 그렇지 않으면 적응변화에 성공하지 못하고 멸종하게 될 것이다. 이러한 변화를 일으키는 외부적 요인은 경쟁, 포식, 물리적 환경변화 밋 군집내의 기회 변화 등을 들 수 있겠는데 실제 지층의 기록으로 나타나는 여러가지 지질학적 환경 변화들과 멸종을 포함 한 생물의 변화 현상과는 상당히 밀접한 관계가 있음이 밝혀졌다.
자연도태와 멸종의 강도는 시공간적인 면에서 매우 다양하게 관찰된다. 적 응방산 (adap tive r adi a ti on) 과 폭발적 진화 (exp lo siv e evoluti on ) 라는 용어 들은 새로운 종이 갑자기 증가하는 현상들을 말하며 집단소멸 (mass exti nc ti on ) 온 분류 단위들이 갑자기 큰 규모로 사멸되는 현상을 말한다. 화석 기 록은 전화의 속도를 연구하는데 중요한 자료를 제공하는데 이러한 종들의 장시간에 걸친 변화에 의해 여러가지 방법으로 측정할 수 있다. 변화가 매우 늦은 계통진화(p h y le ti c evolu ti on) 의 경우 그 속도는 형태학 적 또는 분류학적 기준을 사용하여 측정하며 시간에 따른 변화로부터 진 화경향 (evolu ti onar y t rend) 을 알아낼 수 있다. 실제로는 때때로 지층의 층 서상의 갭 (gap)이 보편적으로 나타나기 때문에 연속적인 전화경향과 속도 룰 측정하는데 난접이 있으며\분류학적 기준을 사용하는 경우에는 種이나 亞種 간의 분류학적 경계의 인위성 · 때문에 다소의 문제가 있다. 이와는 달리 계통발생 속도(p hy lo g ene ti c ra t es) 는 상세한 계통발생은 고 려하지 않고 屬이나 科와 같은 고차 분류단위의 계동발생에서 누진적인 진 화의 속도를 관측하는 것이다. 이는 실제로 밀접한 유연관계의 생물군내 에 평균적인 형태학적 변화를 사간축에 도시하는 것이다. 이 방법은 불충 분한 화석의 기록을 분석하고 지질시대의 큰 간격을 재는 데 편리하다. 한 예로서 약 4 억년 전에 출현하여 현재까지 살고 있는 페어 0mg fish ) 屬둘의 진화를 추적하기 위해서 20 개 이상의 형태학적 변수를 분석해 보 면 전체적인 변화 속도는 초기에 높고 2 억년 전 이후는 급격히 낮게 감소 함을 알 수 있었다. 화석기록은 또한 전체적인 진화에 있어서 새로운 진화방향들이 나타나 는 속도, 즉 다양화(di vers ifi ca ti on) 의 속도를 측정할 수 있다. 새로운 방향 이란 새로운 분류단위의 출현을 의미하므로 예컨대 100 만년 동안에 출현한 신속 또는 신과의 수 또는 그들의 %를 계산할 수 있다. 이룰 기초로 해서 분류학적 빈도율, 소멸속도, 평균수명 등을 계산할 수 있다. 예를 들어 500 만년 전의 조개류의 약 50% 가 현재 생존하고 있으며 종 의 평균수명은 약 1,000 만년임이 밝혀졌다. 갇은 방법으로 포유류종의 평 균수명 이 100~200 만년 정도로 매우 짧다는 것을 알 수 있다. 계동발생에 있어서 진화속도의 변화는 생물역사를 이해하는 데 가장 극 적인 현상들 중의 하나이다. 그러나 아직까지는 진화속도의 분포에 관해 서는 자료가 불충분하다. 만일 계동진화와 분기의 종분화가 전체적으로 거 의 비슷한 비율을 보인다면 어느 형태의 진화가 더 큰 비중을 차지하는가
는 문제가 안될 것이다. 그러나 어느 형태의 진화가 분명하게 우세하다연 두 가지 형태 중에 택일이 가능할 것이다. 진화의 50% 이상이 종의 증식을 동해서 일어난다고 보는 의견을 진화 의 단점 식 모델 (pu nct ua te d equ i li br iu m model of evoluti on ) 이 라고 하며 이는 계통발생 모델의 성질이 계단상 또는 구독접식인 특징을 가진다. 반연에 50 % 이상의 진화가 설정된 종 내의 계통진화로서 나타나는 것을 계통적 접 이 성 모델 (ph yl e ti c gra dualism or gra :d u aii st i c model) 이 라고 하며 이는 Dar- w i n 이 제창한 재래적인 관접이다. 특히 전자는 최근 Eldr idg e 와 Gould(1972) 에 의해 발표된 것으로 큰 주목을 받고 있다. 지난 1980 년 10 월 Ch i c ago 의 야외 • 자연사 박물관에서 열렀던 학회에서도 대부분의 고생물학자, 해부학
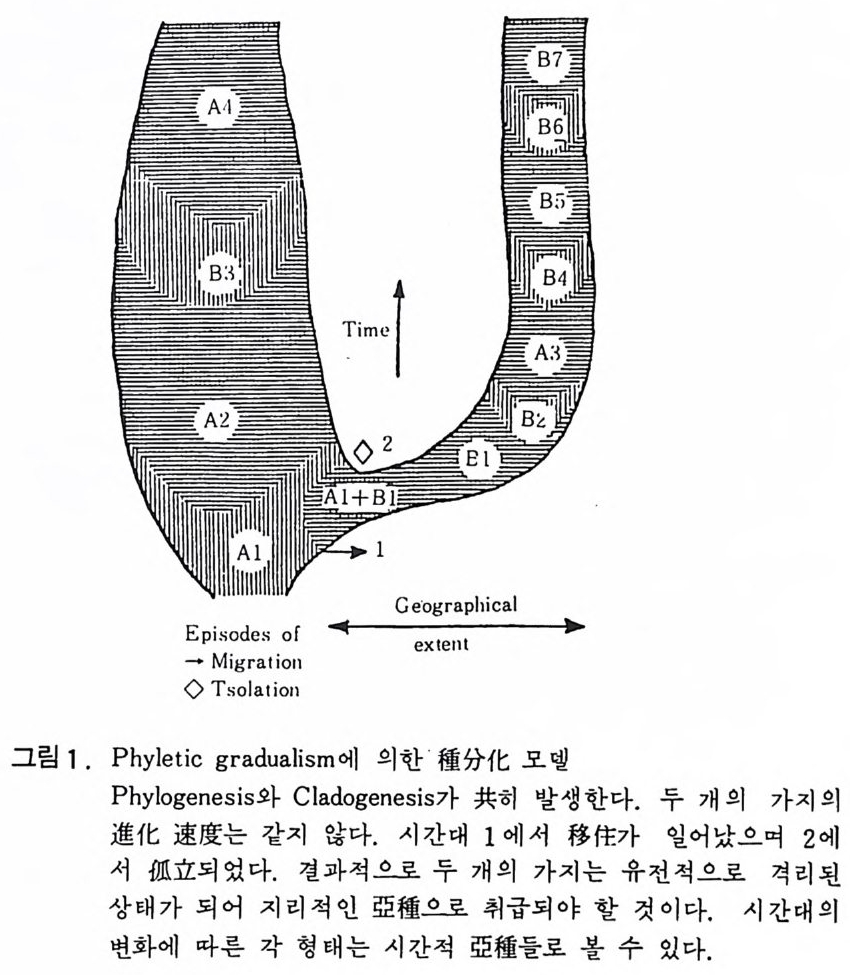 Ge·og r a ph ic a l
Ge·og r a ph ic a l
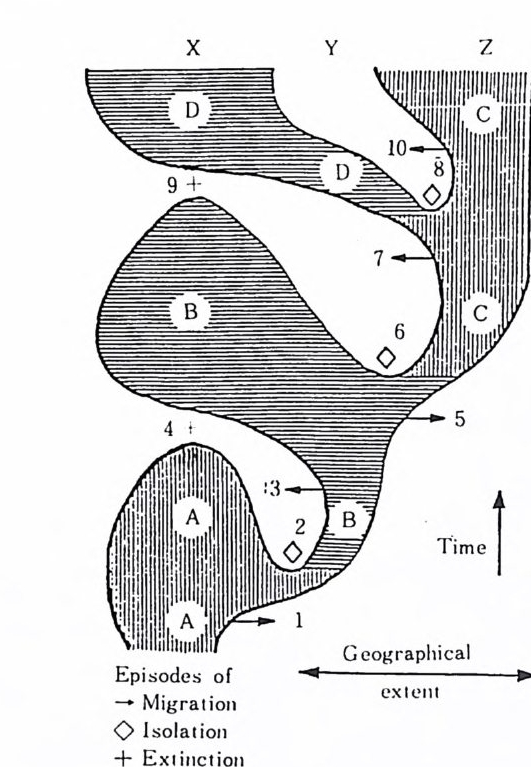 x Y z
x Y z
그림 2 . Punct u at e d e q u il i br i a 에 의한 種分 化 모 델 X 지역의 種 A 는 시간대 1 에서 Y 지역으로 이주하여 分枝하고 2 에서 고립되면 形態#)으로 種 B 를 만든다. 3 의 시간대에서 이들 이 다시 X 지역으로 이주하여 種 A 를 代置하면 4 에서 絶滅이 일 어난다. 種 B 는 5 에서 Z 지역으로 이주하여 6 에서 고립이 다 생긴 후 種 C 로 전화한다. 유사하게 種 C 쇼 1 한 가지는 7 에서 Y 지역 으로 이동하여 8 에서 격리되어 種 D 로 번하고 다시 X 지역 을 침 법하여 種 B 를 대치한다. 따라서 X 지역은 세 가지의 연속적인 A, B 그리고 D 種들이 급격하게 서로를 대치하는 기록으로 지 충에 남게 된다. 자, 진화유전학자 및 발생학자들로 구성된 참석자들로부터 지지를 받은 것 도 이 모델이다. 상기 두 모델에 대한 검증이 계속되고 있으며 화석의 종 류에 따라서 각기 다른 모델에 맞추어지는 경우가 많다. 만일 pu nct u at e d e q u ili br i a 가 유효하다면 분류단위들은 오랜 기간의 변화를 겪지 않고 종분 화가 거의 없는 상태로 지속될 것이다. 찰 알려진 〈살아 있는 화석〉 생물
군이 좋 은 예이다. 앞 서 언 급한 페 어의 경우 급속한 종분화가 있은 뒤 진 화가 중지 된 사 설음 보았 다. Gi nk g o, Me ra s eq uo ia , Ne o p ilin a , S p henodon 의 경 우도 이와 비 슷 한 양상 을 보인다 . 또한 진화가 대부분 종분화 현상으로 일 어난다 연 다 앙성 이 낮은 상 태로 잔존한 생물군은 오랜 시간이 흉러도 크 게 번 화하지 않 을 것이다. 이와는 달리 계동발생의 속도가 계통적 변화에 의 해 주 로 영 향을 받 는 다면 이 들 〈 살아 있는 화석군 〉 도 수천만 혹은 수억 년 동안 새 로운 특징 을 갖는 屬이나 科로 진화했어야 한다 . 目이나 網에 는 살 아 있는 화석이 많 으며 이러 한 변화가 전혀 보이지 않는다. 다시 말 하여 〈 살 아 있 는 화 석군 〉 의 진화사는 새로운 진화모델을 지시하는 것으로 보 인다. 대다수의 생 물 의 진 화 사에서 관측이 되는 적응방산 현상은 이 모 델을 적 용하 는 데 유리하며 적응방산의 검증이 계속 실시되고 있다. 다음 은 화석 기록에 나타난 전화경 향(t rend) 에 관해 알아보기로 하자 . 금 세 기 초 까지만 해도 진화론자들은 화석기록에 나타나는 변화가 직선적인 계 통 전 화 륭 보 인다 고 믿 었 다 . 정 향전화 (or t ho g enes i s) 설에 따르면 극단적 으로 어떤 동 기 에 의 해 서 진화가 시작되면 그 생물이 적응을 더 이상못해 서 멸종할 때까지 한 방 향 으로 계속된다고 생각했다. 이러한 예는 명 종류 의 기형적 인 형 태의 생 물 군에 적용되는데 이 논리는 자연도태도부정했다. 대 표적이 예 로 십하게 말란 굴 종류 , 아일렌드 사슴의 뿔. T it ano t heres 의 코뿡 ,군 도 이 빨 고양이의 송곳니 등이 있는데 이들은 최근에 잘보존된 화 석유 연일히 조사한 후 모두 적응에 평요한 현상으로 해석하게 되었다.또 한 전 화를 한 방향으로 이끌어 가려고 하는 신비한 힘을 믿지 않게 되었 다. 말의 계통발생수 를 살펴 보면 많은 역 전 (reversal) 과 침 체 (sta gi1a t ion ) 의 기간이 포함되어 직선상의 진 화경로 현상도 거부되었다. 화석의 연구에서 계동진화의 경향을 추적하기는 매우 어렵다 . 대부분의 계통진화는 대단히 느리고 진화경향은 그 성질상 추정통계학적이기 때문 이다 . 실제로 관찰되는 변화는 계동발생의 경향인 경우가 많다 . 계통진화의 경향이 우리가 쉽게 복원할 수 없는 국지적인 환경변화와 단 기간의 洞洙壓을 반영하는 대신 계통발생의 경향은 주요 형태의 변화룹추 적함으로써 진화경향의 관찰이 홍F 신 용이하다. 알의 진화는 여러 가지의 형질들 가운데 특히 말굽의 수가 적은 방향으로, 목이 비교적 긴 쪽으로. 그리고 어금니가 좀더 복잡한 쪽으로 계통발생의 경향이 나타난다. 이러한 경향은 나뭇잎을 뜯어 먹는 상태에서 풀 을 뜯어 먹는 상태로의 변환이 일
부 관련이 있어 보인다. 발톱수가 감소하 고 대신 중 간 발톱 이 커져 말발 굽을 만든 이유는 초원의 단단한 잔디 룹 멀리 달릴 수 있 는 능 력을 얻기 위해서이며 긴 목은 땅바다의 풀을 뜯을 수 있고 어금니는 규 질 의 풀 을씹 는 데 효과적으로 복잡화되었다. 체구의 증대는 아마도 적을 방어하고인 내력을 증가시키는 기능이였을 것이다. 이러한 추론은 현생종의 기능형태 에 관한 지식으로부터 얻어진 것이며 화석기록에 나타난 계통발생 경향이 어떤 적응의 의미룰 갖는지를 해석하는 데 이러한 지식이 대단히 중요하다. 분지된 계열들 안에서의 순수한 변화나 일반적인 변화 룽 나타내는 계통 발생의 경향은 하나의 양상(p a tt ern) 으로 생각할 수 있으며 이는 바로 계 동발생의 윤곽 (con figu ra ti on of p h y lo g en y )이 된다. 전화의 양상에서 가장 특징적인 것은 적응방산이다. 이는 주로 진화학적 비약이나 적응쇄신이 일 어난 직후에 나타나서 새로운 고차분류의 기원을 이룬다. 이러한 예로서 가장 국적인 것은 파충류의 각질 양모란 (amn i o t e eg g), 식물계의 씨와 화 분의 발달 등을 들 수 있겠다. 과거 지질시대에는 가끔 급격한 환경변화 에 따른 비약적인 적응이 생물군에 나타나서 비슷한 생활습성을 갖는 다 른 생물군과의 경쟁을 동해 우세한-서식지를 쟁취하고 열세에 있던 생물군 을 절멸시키는 생태학적 치환현상을 초래하였다. 이 밖에도 전혀 관계없 는 두 생물군이 바슷한 생활습성을 갖게 됨에 따라서 밀접한 형태학적 유 사성 을 갖는 적 응수 령 (adap tive converge nce) 현 상 또는 병 렬 진 화 (pa rallelis m ) 도 관찰이 된다. 이상 화석기록을 자료로 해서 얻어지는 대진화 현상들을 개괄적으로 살 펴보고 또 생물학적으로 해석을 시도해 보았다. 지질학은 생물학에 생물 사에 대한 정보를 주고 대신 현생 생물의 여러가지 법칙을 연구에 응용한 다. 생물의 진화는 현재 주장되고 있는 어떤 모델에 의하든지 실재한다. 고생물학의 자료들은 생물들이 뚜렷이 진화해 왔다는 사실을 밝혀 준다. 단지 그것이 어떤 형식을 취했는지는 미해결이다. 현존하는 여러가지 진 화의 모델에 대한 지질학적 검증도 황발히 계속될 것이다. 3 • 2 진화와 미고생물학 (Evoluti on and Mi cr op al eont o log y) 미화석군집에 응용이 되는 진화의 논리들은 현생물의 생물학 및 생태학 에 기초를 두고 있으므로 진화학과 생태학의 밀접한 관련을 알 수 있다.진 화론은 대신 미고생물학자들이 손대지 않았던 유전학에 논지의 바탕을 얻 고 있다. 그러나 전화와 유전의 기본적 이해는 고생태학과 생충서학 분야
에 필수적인 것이다. 전화학에서 種의 개념이 중요함은 고생물학에서도 마찬가지이지안 실제 화석의 기목은 생물학적 種의 특성을 갖추기가 어렵다. 그 대신 화석종은 주관적이며 실용적인 관접에서 정의된다. 화석종은 외부의 보존가능한 형 태학적인 구조 등 다른 전문가들도 인정할 수 있는 증좌들에 의하여 기술 된다. 또한 선조-후손관계나 충서학적 범위 둥은 화석분류에 대단히 중 요하다. 간혹 무성생식을 하는 종류들이 형태적으로 다양하거나 개제발생 과정에서의 상이한 형태변화로 독특한 명명과 분류가 행해지는 경우도 있 다. 이밖에도 어떤 계통내에서도 생태학적인 원인에 의한 種간의 변이. 큰 규모의 적응방산, 개체 혹은 개체발생 중에 일어나는 변이 등은 미고생뭉 학자의 해석에 영향을 끼친다. 따라서 한 지역의 집단을 대표하는 한 그 룹이 묶여서 한 종으로 인식되야 한다는 H ypodigm이라는 종의 개념이 제 안되었다 (S i m p son, 1940). 또한 오늘날 분류는 계동발생 (p hy lo g en y)에 근거 해야 한다는 견해도 널리 받아들여지고 있다 (Si m p so n, 1961). 해양 미고생 뭉학자들은 해처에서 얻는 퇴적물로부터 사체군집(t hana t ocoenoses) 을 얻 어 연구륭 하므로 제 3 기부터 계통발생적으로 진화해온 제 3 기의 플랑크 돈 그룹의 분류를 계통발생에 기초하여 행할 수 있다. 생물지리학적 자료 도 분류에 대단히 중요하다. 이리한 자료는 수령현상이나 병렬진화와 같 은 현상을 설명할 수도 있을 것이다. 또한 g eocl i ne 에 속하는 형태변이를 설정할 수 있으며 이는 특히 고생태 복원에 중요하다. 미화석군의 진화를 이해하는 열쇠는 현생종과 화석종을 야외와 실험실 에서 관찰하여 그들의 생물학을 이해하는 일이다. 성장을 지배하는 조전 을 이해하연 화석종들이 살던 당시의 고환경을 추적할 수 있다. 최근 실 험실에서 미생물들을 배양할 수 있게 되면서 새로운 장의 문이 열린 셈이 며 이러한 연구로부터 분류에 중요한 미세 표면구조나 생체광물질 조성이 환경요소의 변화에 칙접 영향을 받는다는 사실을 알게 되었다. 이로써 고 환경, 고기후의 복원을 위한 형태학적 응용이 새로운 분야로 대두되였다. 해저의 충서기록에 풍부하게 산층되는 플랑크톤 미화석류는 생물의 진 화연구에 이상적이다. 최근의 학자들에 의하면 동·식물 플랑크돈의 종 다 양도, 전화속도 및 절멸속도를 조절하는 중요한 환경요소 중 고기후가 일 차적인 역할을 했다는 결과가 “0/160 의 고수온 곡선연구로 밝혀졌다 (Be rgg re n, 1969 ; Haq , 1973). 한편 생물연대학은 계통선상에서 계통발생의 경향을 형태학적 변화로
써 추적할 수 있게 되었으며 여기에다 최근에는 아미노산 생화학적 연구 가 種 내의 혹은 種간의 유전적 관계를 규명하고 있다 (K i n g, 1975). 이러한 연구는 앞으로 〈 na t ural 〉분류에 한 걸음 더 접근시킬 것이며 진화연구에도 크게 공헌할 것이다. 해양 지구화학과.미고생물학은 공동으로 ·탄산영의 순환과 같은 분야에 서 협력하고 있다. C 야의 대기와 해양에서의 순환과 함량에 관한 연구는 지표에서 살고 있는 생물뿐만 아니라 해수에서는 생산성, 퇴적작용, 플랑 크톤 골격의 형성 등에 영향을 주며 이는 또한 해수의 모든 물리화학적 현 상에 칙접적으로 관여하기 때문에 앞으로 이에 관한 연구가 중요하게 대 두되고 있다. 4 미래의 海洋微古生物孛 지난 십수년간 해양미고생물학은 괄목할 만한 변혁이 생층서, 생물연대 학. 고해양학 및 플랑크톤 진화학 분야에서 있었으며 당분간 현재의 연구 경향은 지속될 것으로 보인다. 이러한 발전은 해처의 시추기술, 컴퓨터, 주 사전자현미경 (SEM) 등의 개발에 힘입은 것이다. 결과적으로 오늘날 해양 미고생물학은 해양의 생물권의 진화역사를 해석하는데 근본적인 역할을 하 게 되었으며 해양지구물리학자와 해양지질학자들과의 공동연구가 증가일로 에 있다. 앞으로 21 세기를 앞두고 해양미고생물학이 갈 방향을 예전해 본 다. 생물충서학과 생물연대학 분야에서 우리는 고위도 지역의 플랑크돈 생 충서 분대, 이들 자료를 천수 지역의 대형 유공충에 의한 분대와 대바하 는 작업 생물충서와 지자기층서 (m agn e t os t ra tig ra p h y)와의 대비 및 이 두 가지를 방사연대 (r adi ochronolo gy)에 퀘어 맞추는 작업 동에 관한 수정 및 보완을 예견할 수 있다. 특히 중생대와 초기 신생대의 연대가 충실해져서 지구 전체의 자사 및 생물사에 크게 공헌할 것이다. 고해양학 분야에서는 고생물지리와 고기후 복원에 기대룽 한다• 제 4 기 기후학에서 우리는 과거 160 만년간의 기온과 해수의 순환을 밝힘으로써 장 래의 변화를 예측할 수 있을 것이다. 제 4 기 이전의 고해양학에서는 산소 와 탄소의 동위원소의 분석 값이 생지리, 생화학, 키후 및 해양순환의 역 사를 밝혀 줄 것이다. 규질 및 석회질 미화석의 상대적 분포 변화는 당시 의 생산성, 용해 및 해양의 CCD 에 관한 정보를 주어 지구의 퇴적작용과
침석작용의 종합화가 기대된다. 따라서 마고생물학의 퇴적학 및 지구화학 과의 협력이 요구된다. 대륙봉과 대륙사면 지역의 미고생물학을 이용한 지사연구는 대륙연변부 에 매장되어 있는 석유 등 지하자원을 탐사하려는 계획에 큰 도움을 중 것 이다. 이러한 연구와 발맞추어 저서성 유공충을 위시한 미화석군은 심해 의 해수의 순환이나 해저분지들의 생성과 전화, 地構造的 발달사를 구명 하는데 이용될 것이고 해양에 떨어져 나온 대륙지각의 침강역사 등도 밝 힐 것으로 기대된다. 보충교재 Be rg e r, W .H . and Rot h, P.H., 1975. Oceanic mi cr op a leonrolog y:pr og r ess a,rl. pr os- pc c l. Rev. Geop h y s . Sp a ce Phy s ., 13(3) : 561-63 5. [C omp r ehensiv e survey of res ea rch in oce a nic mi cr o p a lco n t ol og y durin g the pe rio d 1970-1975.] Be rg g r en , W.A., 1971 . Oce a nog r ap h ic mi cr op a leont ol og y. EOS, 52 : 249-256. [A su mmary of research in oc ea nic mi cr op a leonrolog y bet w een 1967 and 19 70. ] Ei ch e r , D.L ., 1968. Geolog ic Ti m e. Prenti ce -Hall, Eng le wood Clif fs, N. J., 141 pp. [A concis e tre a t m ent of the concep t and gr owt h of ·ge olog ic tim e-scale and its app li c· a ti on .] Kummell, B. and Raup , D. (Edit o rs) , 1965. Handbook of Paleont ol og ica l Techn- iq,u es. Freeman an
and the his t o ry of cir c ulati on in the At la nti c Oc ean. In : W.W.Hay ( Edit o r), St ud ie s in Paleooceanog r aph y. Soc. Econ. Paleo n to l ., Mi ne ral . . Sp e c. Pub!., 20 : 126 -1 86. Be rg g r en, W.A. and Hollis t e r, C.D., 1977. Plate tec t on ic s and pa leocir c ulati on : commot ion in the ocean. Tec ton op h y s ic s , 38 0 -2 ) : 11-48 . Cox. A., 1969. Ge om ag n eti c reversa ls. Scie n ce, 163 (386 4) : 237-~ 45. DeBe er, G. R. (1958) Embryo s and Ance s to rs. London, Oxf o rd Un iv e rsi t y Pr- Haellsasm, , 19A7., pe. d(. A( 19th7o 7r) ouPga ht t reer vnise w o fo fE vhoeltue t ri eocn h. rAonmys , tew rid tahm m, aEnyls e evx iae mr ,p l5e 9s 1.) p. (A collecti on of evolut ion ary cont ri b u t ion s by severa l pa leont ol og ist s . ) Haq, 8., 1973. Trans g r es sio n s , clim ati c chang e and the div e r si t y of calcareous nannop la nkto n . Mar. Geol., 15 (1973) : M25-M 30. Haq , 8., Premoli- S il v a , I. a nd Lohmann, G.P., 1977. Calcareo u s pla nkt o n pa le- obio g e og r ap h ic e,id ence for majo r clim ati c fluc t u ati o, ;s in the earl y Ce n ozo ic At la nti c Ocean. J. Geop h y s . Res., 82( 27 ) : 3861 一 3876. Honjo , S., 1976. Coccolit h s : pr oducti on , t ranspo rta ti on and sed im ent a t i on . Mar. Mi cr op a leont ol ., 1(1) : 65-70 . Kenne tt , J.P . and oth ers, 1975. Cenozoic pa leooceanog r ap h y in the sou th w e st Pacif ic Oc ean, Ant a rct! c gla cia l io n , and the de v e lo p m e n t of the cir c 11m-Ant - areti c current s. In : J. P. Ke n nett , R. E . Ho u lz and oth ers, Init ial Re p o rts of the Deep - Sea Dril l in g Proje c t, 19. U.S. Gove rn me n t Prin t i ng Of fice , Wa-shin g t on, D. C., pp. 1155 기 169. Kin g Jr. , K.,1975. Ami no acid comp o sit ion of the sil ic i f ied mat ri x in fos sil po lyc - ys t i ne i:-a dio l aria . Mi c rop a leo n t o l., 21 (2 ) : 215 -226. Kin g Jr. , K. and Hare, P.E., 1972a. Ami no acid comp o sit ion of pla nkt on ic fo r a- ami ni f er a . A pa leobio c hemi ca l app r oach t o evo lut i on . Scie n ce, 1975 (4029) : 14 61-1463. Kin g Jr. , K., and Hare, P.E., 19726. Ami no acid comp o s it i on of th e tes ts as a tax onomi c charact e r for liv i n g and fos sil pla nkto n ic fo rami ni f er a, Mi cr op a le- ont ol og y, 18(3) : 285-29 3. May r , E., 1966. Anim al Sp e cie s and Evolut ion . Belknap Pres s of Harvard Univ e rsit y, Cambrid g e , Mass., 797 pp, May r, E. (19 71 ) Pop u lati on s, Sp e c ies and Evolut ion . Cambrid g e , Mass., Harv- ard Univ l ' rs it y Press. 453 p. (An abrid g e d and re a dable versio n : o f May r's . 19 63 book,l'mp h asiz i n g the sp e cie s as the unit of large -sc al e evoluti on .) Olson, E. C. (1965) The Evolut ion of Li fe. Loudon, Weid e n fe l d and Nic o ls on, 300 p. (A dis c ussio n of the _ma jo r aspe ct s of evoluti on , espe cia l ly tho se evi- dent in the verte brate fos sil record) . Packham, C.H. and Van der Li ng e n, G. J., 1973. Prog r essiv e carbonate dia g e n- esis at Deep Sea Drill ing Sit e s 206, 207, 208, and 210 in the Sout h west Pacif ic and its relati on ship to sedim ent pr op e rti es and seis m i c refl ec to rs.
In : J.E . Andrews, R.E. Burns and oth ers, Init ial Rep o rts of the Deep Sea Drill in g Proje c t, 21. U. S. Government Prin t i ng Of fice , Washin g ton , D. C., pp. 495 -507. Raup D.M.& St an ley S.M. (19 71) Prin c ip le s of Paleont o log y. Freeman & Co. 481 p. Rensch, 8. (19 60) Evoluti on above the Sp e cie s Level. New York. Columbia Univ e rsit y Press, 419 p. (A t reati se on the evolut ion of hig h er tax a wit h re- fer ence·s to many German arti cl es.) Schlang e r, S, 0. and Doug la s, R.G., 1974. The pe lag ic- ooze-chalk-lim esto n e tr- ansit ion and its im p li ca ti on for marin e str a ti gr aph y. In : K.J. Hsu and H.Je - nky n s (Edit o rs) , Pelag ic Sedim ent s : On Land and Under the Sea. Sp e c. Pu- bl. Int. Assoc. Sedim ento l., 1 : 117-148. Schop f, T.J. M ., ed. (1972) Models in Paleobio l og y. San Francis c o, Freeman, Coop e r, and Co. 250 p. (A collecti on of pa leobio l og ica l pa p e rs, many of wh- ich are of an evoluti on ary natu re.) Sim p s on, G.G., 1940. Ty pe s in modern tax onomy . Am. J. Sc i., 238 : 413-431. Si m p so n, G.G., 1953. The Majo r Featu res of Evoluti on. Columbia Univ e rsit y Press, New York., N.Y., 434 pp. Sim p s on, G.G., 1961 . Prin c ip le s of Anim al Taxonomy . Columbia Univ e rsit y Pr- ess, New York, N.Y., 247 pp. S tan ley, ste v en M. (19 79) Macroevoluti on , pa tt er n and pr oc·e s sess. W. H. Fre- eman & Co. 332 p. Sy lv este r -Bradley P. C. (1977) Bi os tr a ti gr aph ic a l Tests of Evolut ion ary Th-
eory (in Concep ts and Meth o ds of Bio s tr a ti gr ap h y edit ed by Kauf fm an & Hazel) . Dowdon, Hut ch is o n & Ross 658 p. Valent ine , J. W. (1973) Evoluti on ary Paleoecolog y of the Marin e Bi os ph ere. Eng le wood Cl iffs, N.J. , Prenti ce -Hall. 511 p. (A ge neral rev iew of ecolog ica l ~nd evoluti on ary th eory as app li e d to marin e life .) VaInn t.H i nPt le a, n kJ.t E on .,i c 1 C9 o6n9(. ., TGheen envaat ,u r2e :o 2f 6b7 i-o s 2 t7 r 2a. ti gr aph ic zones. Proc. Fi rs t제 3 장 海洋微化石 各論 (Mar in e Mi cr ofo s sils ) 1 有孔虫 (Forami ni f er a) 1 • 1 서론 유공충은 원생동물 (Pro t ozoa) 중 위족충류 (Sarc odi na) 에 속하는 것으로 서 캄브리아기에 충현, 오랜지질지대를 거쳐 오늘에 이른다. 현재의 해양 에 넓게 분포되고 있는 이 유공충은 대부분이 底生生活울 하고 있으며 일 부는 浮遊生活울 하고 있다 . 고생대 이래 주로 해성층내에서 화석으로 풍 부히 산출되고 있으며 표준화석 또는 시상화석으로 이용되고 있는 種이 많아 지질학적으로 매우 중요한 역할을 담당하는 미화석이다. 유공충의 분류는 처 음으로 De Blain v i lle (1825) 에 의 해 시 도되 었 다. 그후 d'Orb ig n y (1826) 에 의해 대규모적인 분류가 본격적으로 행해졌으며,유공충 ( Forami ni f er a ) 이 란 이 름도 d' Orb ig n y에 의 해 명 명 되 어 졌 다. 어 원을 간단히 보면 라틴어의 fo ramen( 孔)과 f ero( 有)의 복합어이다. 명명 당시 유공충의 각이 외관상 연체동물 중 두족류와 유사하여 유공충을 두족류에 포함시켰 다. 그러나 두족류의 경우 각내에 s ip hon 이라는 管을 갖는데 반하여 유공 충은 내부를 구분짓는 격벽 (se pt a) 에 구멍을 갖는 정도에 지나지 않으므로 ceph a lop o des for am ini f er es 라 하여 전 자인 ceph a lop odes s ip hon if eres 와 구별 하였다. 죽 유공충 체부는 두족류와 같은 분화기관을 갖지 못하며 단지 원 형 질 (pr oto p l a sm) 로 구성 되 어 있 다. 유공충은 10cm 이상에 달하는 것도 있지만 대부분의 종은 1mm 이하의 각
에 싸여 진 단핵 내지는 다 5식 성 원형 질체로서 허족(p s eudo podi a) 을 갖는다. 유공충의 허족은 음식 섭취, 소화. 배층, 포낭 (c y s t)의 형성, 각의 부착, 이동 동의 기능을 갖는다. 유공충은 세대교번 (alte r nati on of ga, era ti ons) 을 하며. 따라서 동종이형 (di mor p h i sm) 현상을 보인다. 유성의 단핵성 세대는 편모상의 배우자를 생산하며 이것이 합체하여 무성세대를 만든다. 무성세 대는 다핵의 소구형으로 이는 다시 감수분열에 의해 유성세대를 만든다. 보통 소구형 내지 미구형 (mi cr osph eric ) 개체는 초실(p roloculus) 이 작고 각 이 크며 무성세대에서 볼 수 있으며 대구형 (me g alos p her i c) 의 개체는 초실 이 크고 각이 작으며 유성세대에서 볼 수 있다 . 유공충이 인간의 역사에 처음으로 등장하는 것은 BC 5 세기경 Herodotu s 가 이집트의 피라밋 석재 중에 포함된 대형 유공충을 화폐석 (Nummul it es) 이라 칭한 데서부터 비롯된다. 그후 · AD 1 세기경 Caiu s Plin i u s Secundus 에 의해 Da p hn i a 라 불러졌는데 이는 조로아스터 교도가 사용하던 월계수 잎과 담았다는 점에 기인한다. 17 세 기의 현미 경 발명과 더불어 Beccariu s (1731) 에 의해 소형 유공충이 처음으로 기재되었다. 1758 년 L i nne 에 의해 발간된 Sy s te m a Na t ura 라는 책에서 동물분류 입장으로 서술된 것을 보면 두족류 혹은 전혀 다른 동물군 속에 포함시켜 취급하고 있는 정도에 지나 지 않았다. 그러던 것이 d'Orb ig n y (1826) 에 와서 대규모적인 분류작업을 하 게 되었다. 그뿐만 아니라 많은 지역의 여러 시대에 걸친 지층으로부터 화 석종을 기재하고 유공충군의 지리적 변화 맞 지질작 산상에 대한 관심은 오늘날의 유공충을 이용한 생충서학(bi os t ra tig ra ph y)연구의 선구적 역할을 한 셈이다. Schu lt ze(1854) 에 의해 현생종에 대한 각과 원형질의 연구가 이 루어졌다. 유공충에 대한 고생물학적 연구와 생물학적 연구는 19 세기 중엽 부터 본격적으로 행해졌으며 20 세기에 들어서면서 경제적 응용면 및 석유 개발과 관련되어서 급격한 발전상을 보이고 있다. 지층의 연대결정 ·대비 • 생성환경 추정 등 여러 측면의 연구가 유공충을 통해 이루어지고 있으며 최근 부유성 (pla nkto n ic ) 유공충에 대 한 연구가 황발하다. 유공충은 보통 5,OOOm 심도에까지 컬쳐 서석한다. 저서성 종은 캄브리 아기 이래, 부유성 종은 중생대부터 발전해 왔다. 대체로 유공충은 크게 3 g rou p;부유성 (pla nkto n ic s ), 소형 저 서 성 (smaller benth i e s ), 대 형 저 서 성 (lar ge r ben t h i cs) 으로 구분되 며 특히 대 형 저 서 성 인 Fusul i nacea 계 통은 고생 대 후기에 전성기를 갖는다.
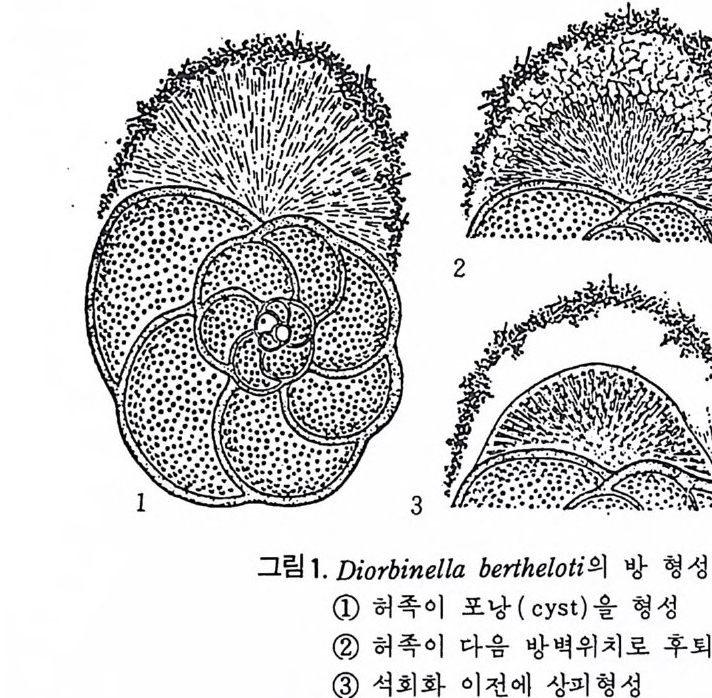
1 • 2 생활사 화석으로 잔존하는 부분은 각과 같은 경조직으로. 연체부는보존되지 않 기 때문에 고생뭉학을 연구하는 많은 사람들이 연체부를 우시하고 각만을 연구하는 경향이 있는데. 사실상 연체부와 경조칙과의 관계물 따로 떼어 놓은 고생물은 생각할 수조차 없다. 따라서 현생 생물에 대한 충분한 지 식을 가지고 화석을 통한 고생물을 연구하며 고생태를 밝히기 위해 우선 현생 유공충에 대해 알아둘 필요가 있다. 1) 원형질체 유공충은 원형질과 이를 보호하는 각(t es t)으로 이루어졌다.원형질은 아 에바와 같이 바교적 어두운 색으로 된 내질 (endo p lasm) 과 밝은 색의 외질 (ecto p la sm) 2 층으로 구성 된다. 내질과 여기서 나온 허족은 외부에 접하게 되며 각을 분비한다. 내질은 한개 혹은 그 이상의 핵 (nucleus) 을 가지며 핵의 수는 種이나 세대에 따라 차이가 있다. 내질로부터 돌출해 나온 허족을 통해 먹이 섭취, 소화,배출, 각벽 구축, 포낭형성, 각의 부착 및 이동을 한다. 허족은 과립질망상 (g ranulore ti cula t e) 으로 가늘고 길게 떨어져 나가거나 합해지므로 그 모습을 딴 Rhiz o p odia 혹은 M yx o podi a 라고도 불리어진다. 허족의 운동은 활발하며 종류에 따라. 혹은 같은 종이 라도 세대에 따라 운동의 다양성을 보여준다. 허족을 이용하여 규조류 • 해조류 • 편모조류 • 갑각류 동의 미소생물이나 유 생 혹은 유기울을 먹이로 섭취하는 holozoic 영양양식을 갖는다. 허족 자 체에는 먹이 선택능력이 있다. 각의 성질에 대해서는 형태에 관한 부분에서 자세히 다루기로 한다. 2) 성장 새로운 방 (chamber) 을 만드는 데 있어 허족은 중요한 역할을 하게 된다. 새로 만들어진 방은 이전의 방과 동형이며 크기만 커지는 것이 보통이다. 새로 형성되는 방의 크기는 그전 방의 원형질 양에 비례하며 . 방의 증대 율은 유공충의 분류에 도움되는 주된 요소 중의 하나이다 . 석 회 질 성 분의 多宰形 (po ]vth a lamous) 유공충의 성 장을 보면 우선 각의 최종 방의 口-f l(a p er t ure) 으로부터 허족이 나오게 되며. 이 허족 외측의 얇은 막과 외부로부터 모은 재료로서 포낭을 형성한다. 허족은 다시 후퇴 하게 되고 포낭내부에 새로운 방 공간을 만들게 되며 표면에는 새로이 석 회질 벽이 분비된다. 일반적으로 허족이 입을 통해 나와 포낭울 형성한 뒤 이 포낭 외면이 석회화하기까지는 8 시간 정도를 요하며, 생리작용 및 환
동기능을 하기까지는 약 24 시간을 요한다고 한다(그림 1 참고).
 3
3
유공충은 어느 정도 재생능력이 있어 원형질에 각을 포함한 파편은 새 로운 각을 만들어가며 비교적 불규칙한 형태로 재생한다. 때로는 둘 이상 의 유공충 각이 하나로 합해지는 경우가 있는데, 주로 같은 세대의 크기 가 비슷한 유체가 합해지는 것으로 이룰 쌍생 (twi n) 이라 한다. 쌍생의 경 우 세포질 (c yt op lasm) 은 유합하나 핵은 유합되지 않으므로 생식에 의한 개 체 간의 융합 (pla sto g a my) 하고는 다르다. 3) 생활사 유공충의 생물학적 측면에 대해서는 그다지 자세하게 연구되어 있지 못 한 실정이다. 더우기 유공충의 생활사를 연구하고자 했던 동기마저도 현 생 유공충이 아니라 화석인 화폐석이었다. De La Har p e(1879) 의 화폐석 연구 결과에 의하면 穀의'크기가 큰 것은 初宰이 육안으로 볼 수 없을 만 큼 미소하고, 穀이 소형인 경우 초실이 크다는 사실이다. Munie r -Chai- mas 는 이 현상을 동종이형성으로 설명하였으나 그에 대한 생물학적 의의 는 알 수 없었다. 그후 화폐석 이외에도 많은 유공충으로부터 동종이형성
현상이 발견되 었으며, 초실이 큰 것을 대구형 (meg a losp h eric ) , 작은 것은 소구형 (m i cros p her i c) 이 라 하였다. L i s t er 와 Schaud i n(1894) 에 의 해 동종아 형성 현상은 세대교번에 의해 생기는 것으로 증명되었다. 소구형 개체가 분영하연 단핵응 갖는 개체군을 만든다. 이 분열현상을 sch i zo g o11 _v라 하는데 이렇게 하여 생겨난 개제는 대구형을 갖는다. 개제군 음 만든 모 체 유 schiz o1 1t 혹은 aga mont 라 하며 생 겨 난 개 제 륭 ga mont 라 한 다. 대구형의 g amon t는 성숙기에 들어서연서 배우자(ga me t e) 를 만들며 이 매우자가 안나 형성된 접합자 (z yg o t e) 는 작은 초실을 만들며 이것이 성장 하여 소구형의 sch i zon t가 된다. schiz o nt 세대의 핵의 염색체 수는 2l( 이 매 체 . dip lo id ) 이 며 gam o11t 세 대 는 n (반수 체 , hap lo id ) 으로 접 합자에 의 해 2n 상 태 가 된다(그랑 2 참고).
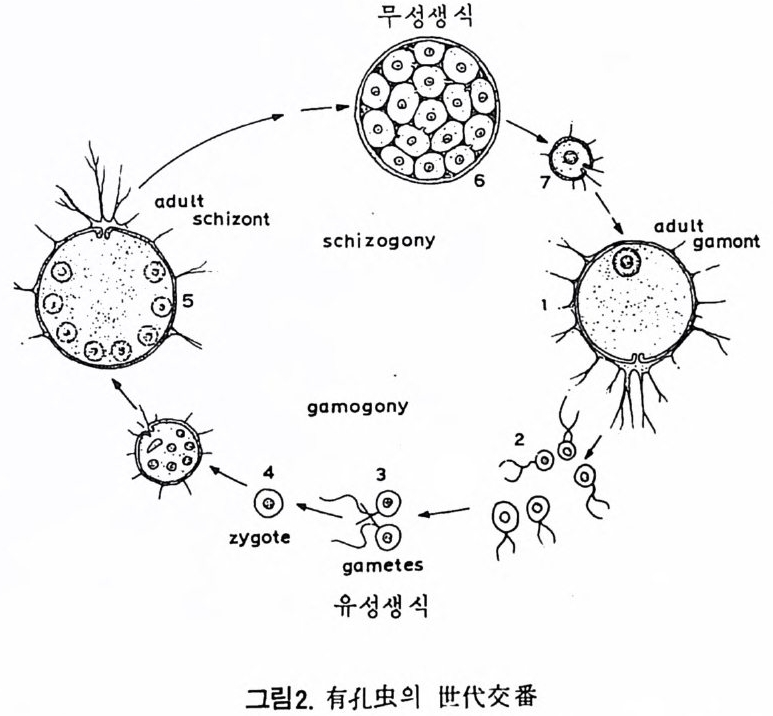 무성생식
무성생식
유공충의 배우자는 동형배우자(i so gam e t e) 로서 대다수의 것이 편모름 가 지며 극소가 아메바상을 보여주고 있다. 최 근 My e rs 나 Le Calvez 에 의 하연 이 형 성 생 환환 Oife cy cl e) 은 일부에 지 나지 않으며 이외에도 다양한 모습을 보여주고 있음이 밝혀졌다.
그림 3 은 종류가 다른 유공충 3 종의 생활사 를 보여주는 그림으로서 단히 5 단계로 구분해 볼 수 있다.
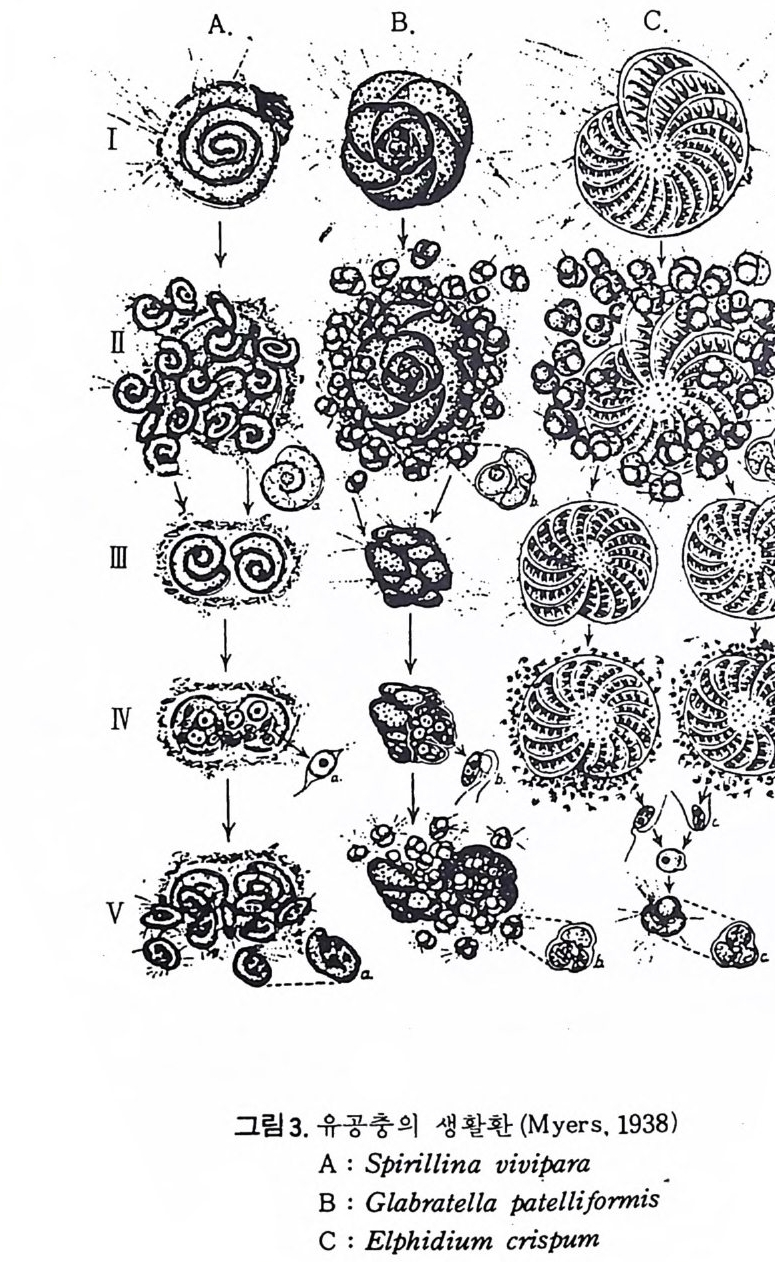 그림 3. 유공충의 생 활환 (M ye rs, 1938)
그림 3. 유공충의 생 활환 (M ye rs, 1938)
a 를 g amon t세대, B 륭 sch i zon t세대라 할 때 이형성 생활환이란 a-/3- -a -fl 의 반복을 말한다. Ho f ker(1930) 는 어떤 종에서 대구형의 초실의 크기 변화에 착안, garno nt 에는 탄핵을 갖는 것 (aI) 과 다핵 (a2) 을 갖는 것이 있음을 발견하였으며 그 결과 aI-a2-/3- -a I-a2-B 같은 순환주기물 생각해 냈다. 이러한 생활환을· 그는 상형성 (t r i mor ph i sm) 이라 정의하였다. 그러나 현재 상형성온 대구형 중에 다 8식 성도 있다는 의미 정도로 사용되고 있다. 유공충의 생 활사 해 명 에 공헌을 한 Le Calvez 의 연구를 Arnold(1956) 가 하나의 생 활환의 도표로 종합한 것 이 그림 4 에 나타나 있다. 유공충의 생석범으로는 여기에 기재된 것 이외에도 출아법 (b u dding)이나 분 리 법 (fra g me nta t i on ) 을 갖는 종 이 있 다. Ho f ker 에 의해 처음으로 세대의 변화가 계절변화와 관계가 있음이 밝혀 졌다.
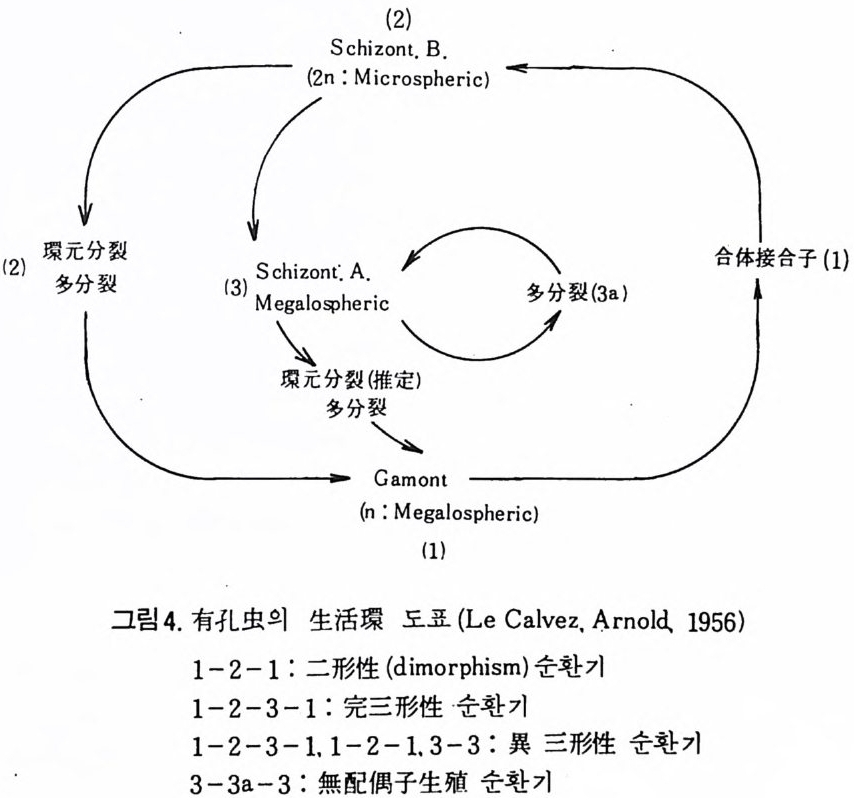 (2)
(2)
유공충의 세대에 따른 다형성 현상은 각의 형태에 기초한 분류에 문제 점을 제시한다. 그외에도 유공충의 방의 배열 역시 성장단계에서 변화되 는 경우가 있다. 4) 이동 (M ovement) 저서성 유공충은 고착성 혹은 이동성의 성질을 가지며 허족을 이용하여 움직인다. 평균속도는 1 시간당 1cm 정도에 지나지 않는다. 부유성 유공충의 경우는 해수의 움직임을 통해 이동되며 허족과는 거의 무관하다• 5) 공생 • 기생생물 유공충은 조류와 공생을 한다. 저서성 • 부유성에 관계없이 단세포의 남 조류, 녹조류 혹은 편모조류 등과 공생을 하는데 이들과의 공생관계에 대 해 자세히 연구된 것은 아니다 . Lee(1965) 에 의하연 공생조류 등이 각 방 안의 3/4~4/5 정도를 차지하고 있다고 한다 . 그 외에도 유공충에 균류 및 선충류 등이 기생하고 있는데 화석상에서는 발견하기가 극히 힘들다 . 반대 로 유공충이 다른 종에 기생하는 예가 있다. 유공충의 기생은 숙주의 사 멸의 원인이 된다. 1 • 3 형태 및 분류 1·3·1 형태 유공충은 주로 穀(t es t), 般의 외 형 적 내부구조, 穀 벽 (wall) 의 조성 맞 구조, 방의 형 태 및 배 열, 장식 (ornamenta ti on ) 등울 기 초로 분류되 고 있 다. 유공충의 각은 거의 모든 부분에서 변화가 매우 다양하다 . 각의 크기 는 보통 0.02mm~190mm 까지 꽤 변폭이 심하다. 그 실예로 각의 크기에 대 한 기록을 살펴보면 백악기의 Loft us ia elon g a ta가 118mm, 제 3 기의 Numm- ulite s giz ehens is가 110mm, 현생종 중 Sta n nop hy ll um zonar i um이 190mm 를 보 여준다. 유공충의 생체에 관한 형태학적 지식은 분류의 기준이 될 수 있 을 정도로 충분하지는 않다. 화석의 경우는 제외하고라도 현생종에 있어 서도 각의 형태 및 구성이 분류의 기초가 되고 있다 . I) 각벽의 구성 유공충의 각벽은 성인상 2 종류로 구분된다. 하나는 유공충 자체가 분비 하는 것이고, 다른 하나는 외부물질을 모아 분비물에 교착시키는 것으로 이때 분비물은 화학적 · 광물학적 특성의 차이에 의해 몇 g rou p으로 구분된 다.
W i ll i am 이 19 세기 후반 처음으로 각벽의 성질의 중요성에 대해 발표하였 으나 그 당시는 분류에 관한 많은 이론으로 혼란한 시기였다. 현재 식별되고 있는 각벽의 조성 및 구조 동의 대부분을 밝힌 Wo od.(1 949) 의 연구 철과에 의하면 이들은 적응에 의해 변화하는 것이 아니며 분류상 가장 중요한 역 할을 하는 것으로 강조되고 있다. Loebl i ch 와 Tapp a n (1964) 은 Wood 의 연구절과를 기초로 다음과 같은 종류로 구분하였다. A) Pseudochit ino us tes ts (의 키 틴 질 각) B) Ag glu t ina te d tes ts (교착질 각) C) Porcelaneous te sts (자기 질 각) D) Mi cr og ra nular te sts (마 립 질 각) E) Hy al in e calcareous tes ts (유리 석 회 질 각) a) Perfo r ate gra nular walls ( 다공 입 상 각 벽 ) b) Monocrys ta l lin e walls (단 결 정 질 각벽 ) c) Perfo r ate radi al walls (다공 방사상 각벽 ) F) Sp icu lar te sts ( 침 상 체 질 각) A) Pseu.d o chit ino us tes ts ( 의 키 던 질 각) ; 얇고 탄력 성 있는 투명 한 각으로 당단백질로 이뤄졌다. 종래엔 이 성분으로 이뤄진 각을 키틴질, 젤라틴질, 게라탄질 등으로 불러왔으나 어느 것도 화학성분을 정확히 표현하지는 못 하고 있다 . La gyn acea 상과에 속하는 유공충은 의키틴 성분을 갖는다. 이 러 한 성 분은 단순형 (unil oc ular) 혹은 단실 형 (monoth a lamous) 종류에 서 도 볼 수 있다 . 간혹 화분분석을 위해 산에 처리한 시료 중 미소유공충(Mi cro fo rami ni f er a) 이 라고 불려 졌던 작은 유공충이 간혹 발견되는데 이 유공충의 석회질각의 내막을 이루는 의키탄질이 남겨지기도 한다. B) Ag glu ti na te d tes t (교착질각) ; 석영, 장석, 운모 및 여러 중광물, 접토 나 생물의 유해 등을 주재료로 하여 유기성 cemen t질로 고결된 각을 말하 며 사질각 (arenaceous t es t s) 이라고도 한다. 대부분의 종이 생활장소에 산 재한 재료를 환용하는데 반해 특정 종은 재료룽 선택하는 능력을 갖는다. 이런 선택능력은 원형질의 화학적 성질과 관련이 있다.그 외에도 cemen t와 교착물질 사이의 양적 비움은 주변 환경이나 종류에 의해 번화한다. cement 로는 유기 cement, 탄산 석 회 , 수산화물, 그 외 에 여 러 종류 의 cemen t가 있는데 그 중 규질 (sili c e ous cement) 가 많은 경우는 규질각 (sili ce ous tes ts ) 을 형 성 한 다. C) Porcelaneous t es t s( 자기질각);외각이 도자기 표면 같은 느낌을 주는
불투명한 석회질각으로 벽공(p ore) 은 거의 보이지 않는다. 방해석 미립자 로 구성되었으며 경칭의 배열은 불규칙한 것으로 알려져 왔으나 최근 Towe 와 C if e lli (1967) 에 의해 대부분은 불규칙하나 외표면의 일부에 방향성을 갖 는 막상의 층이 존재함이 밝혀졌다 이 자기 질 각을 갖는 유공충은 모두 무공질 (i m per fo ra t e) 로 알려 져 있었 으나 일부 종은 벽공을 갖는다. 표면에 사질의 피막을 쓰고 있는 종류도 있다. D) Mi cr og ra nular t es t s( 미립질각);고생대의 대표적유공충인 Fusulin i n a 亞目의 각벽의 특징으로 작은 방해석 결정이 밀집하여 구성되었다. 결정의 집합상태는 규칙적이며 산지가 다른 시료일지라도 같은 상태를 보이는데, 이는 분비현상에 의해 형성된 각임을 시사한다. 입자에 의한 규칙적인 배 열을 보여주는 외에도 섭유상을 보이며, 때론 미립자 층과 섬유상 층이 합 해진 복합벽을 만든다. 복합벽을 갖는 유공충은 고생대 후기에 충현하였 다. 미립질 각을 갖는 각벽의 구조는 그림 5 와 같다.
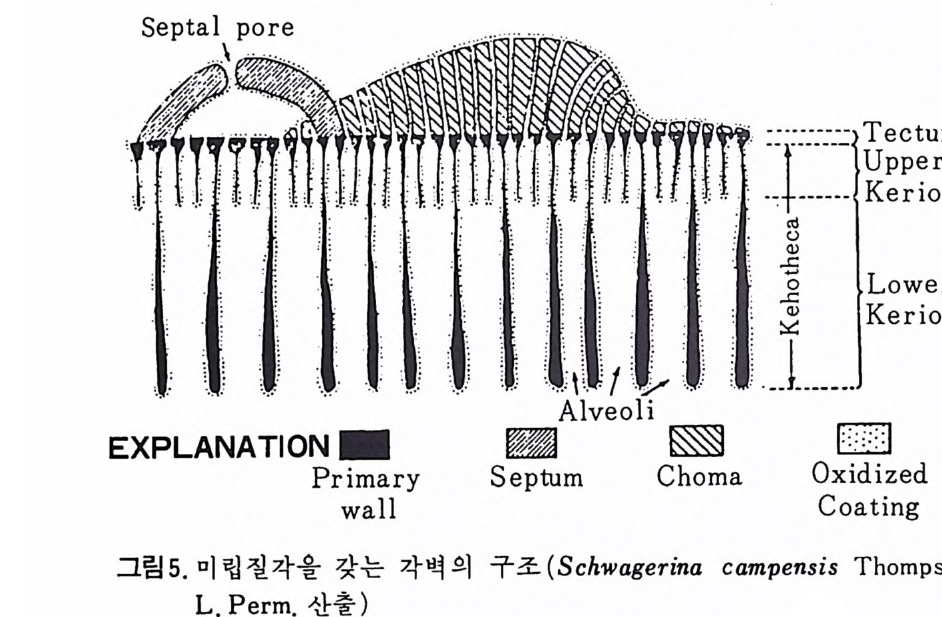 二Siep tall 幅 p o| re 'l II틀 Hr l, lml |m | :1:i기〕 〔 눕:;:;l u er0m: heca
二Siep tall 幅 p o| re 'l II틀 Hr l, lml |m | :1:i기〕 〔 눕:;:;l u er0m: heca
E) H:v a lin e calcareous tes t (유리 석 회 질 각) ; 이 성 분을 갖는 유공충 각을 빛이 통과할 때 유리속을 통과하는 것과 같은 느낌이 둥어 유리질 01 :v al i 11c) 이라 한다. 대부분의 것이 다공질(p e rf ora t e) 로 방사상 (ra di al) 내지는 입상 {g ranular) 의 각벽을 갖는다. 방사상의 경우 각벽을구성하는 석회질 결칭
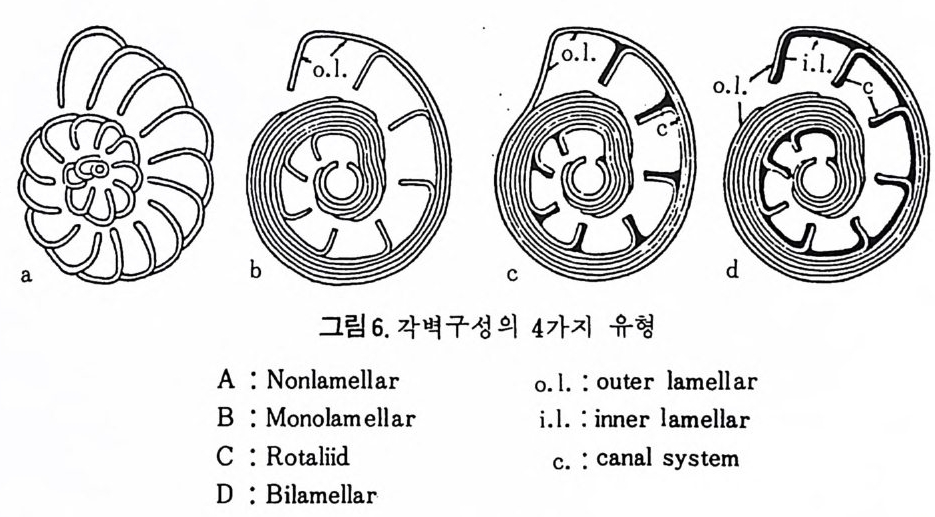
의 광축은 벽면에 수직이다. 여기에 속하는 종은 주로 방해석으로 이루어 졌으며 간혹은 ar agon it e 로 구성 되 었으며 예로 Roberti na cea 상과 물 둘 수 있다. 입상 각벽의 경우는 벽면에 경사물 보이며 발달한다. 단 겅정질 각 벽 (monocrys t a l lin e walls) 의 각은 단일의 방해석 결정으로 구성되였으나 방 사상 구조를 갖지 않는 것으로 S pir i ll i n i dae 과에 속하는 종류를 말한다. F) Sp icu lar tes ts (침 상체 질 각) ; 침 상의 방해 석 절 정 의 집 합체 상으로, 유 공충 자체로부터 분바된 것 이며 Car t er i na 속에서만 봉 수 있다. 현생 유공충을 대상으로 연구한 절과에 의하면 각을 . 구성하는 주된 성 분은 교착질 각의 정우 S i 02 가 75% 이상을 차지하며 그외에 CaO, Fe20J, Al20J 등의 성분을 들 수 있다. 석회질 각의 경우는 85% 이상이 CaCO J이 며 Mg C OJ ,F e20 J , S i아 등이 보조성분으로 존재한다. 원소로 보연 Ca,Si, Mg 등 17 원소가 밝혀졌으며, Mn 의 경우 유리질보다는 자기질 유공충 내 에서 높은 함량을 보인다. 또한 수온의 저하에 따라 감소 경향을 보이는 Mn 은 스트론튬 맛 베륨과 함께 중요한 원소이다. 2) 각벽의 층상구조 유공충 각의 단연에 나타나는 각벽의 구성을 보면 크게 4 가지 유형으로 구분된다(그림 6 참고).
 o.
o.
A) Nonlamellar( 무충상)각;전 방의 구공을 따라 새로 생기는 방의 각벽 이 부가되는 유형으로 교착질, 미립질 혹은 자기질 각을 갖는 유공충에 보 편적으로 발달한다. 이에 반해 새로운 방의 각벽을 형성하는 층 Oamella) 이
방의 형성과 동시에 기존 방의 외표면을 싸 는 경우 점접 기존 벽이 두꺼 워지면서 층상구조가 발달한다. 이러한 층상구조 를 갖는 각을 층상각이라 하며 양식에 따라 3 가지 유형으로 구분한다. B) Monolame li ar( 단층상)각; 새로운 방울 만드는 충이 방을 형성하는 것과 동시에 기존 穀의 표면을 덮는다. C) Ro t a lii d 상 ;층이 기존 穀의 표면은 물론 최종 방의 전면, 죽 성 장방 향에 향한 면까지 덮어 싸기 때문에 방 사이의 격벽은 2 중이 된다. D) B i lamellar( 복충상)각 ; 충이 원래 2 중으로 되어 있어 외측 층은 기존 각
 5
5
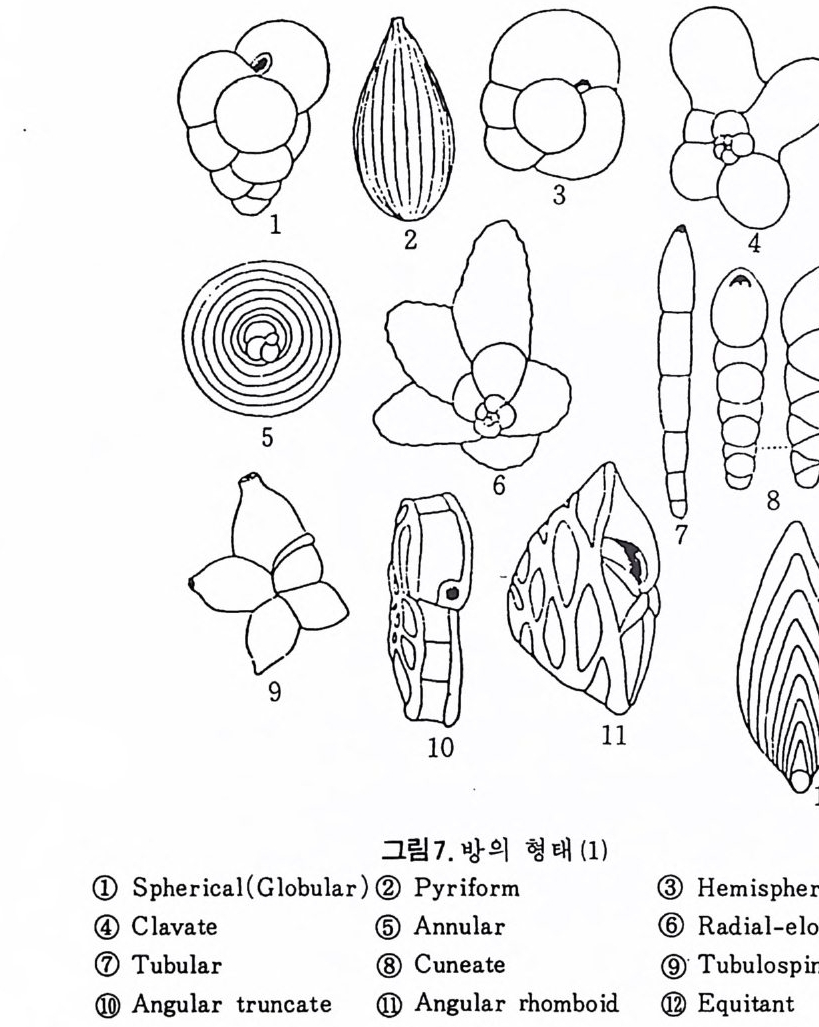
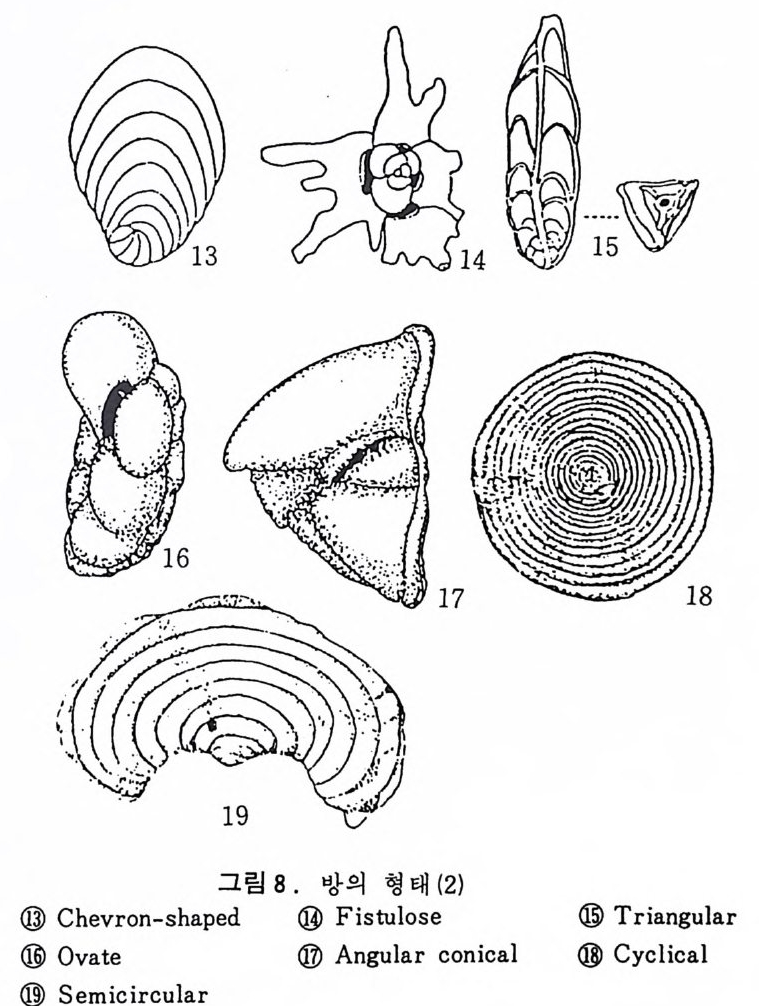
의 표떤음 싸여 새로운 방을 형성하는데 반해 내측 층은 새로 형성되는 방 내에 한정되어 발달한다. 부유성 유공충 중 일부는 성장이 끝난 뒤 각의 표면이 피각 (crus t)에 의 해 둘러싸이는 것을 분 수 있다. 3) 방 (chamber) 의 형 태 와 배 열 유공충의 각은 한 개 이상의 방으로 구성되어 있으며 다양한 형태를 갖 는다. 형태학적 다양성온 이룰 구성하는 방의 형태와 배영의 다양성에 기 인한다(그림 7,8 참고).
 14 일,...'YI l
14 일,...'YI l
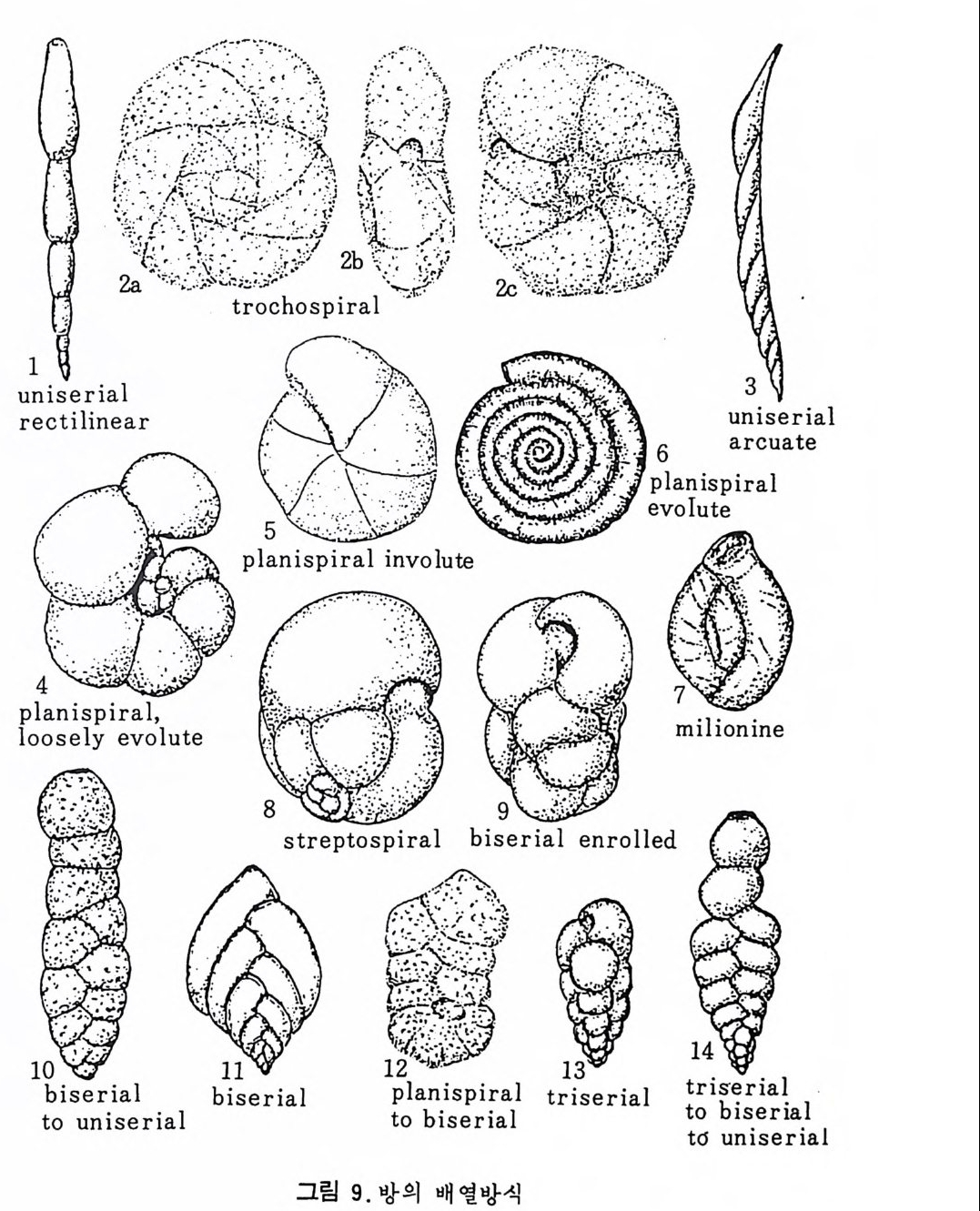
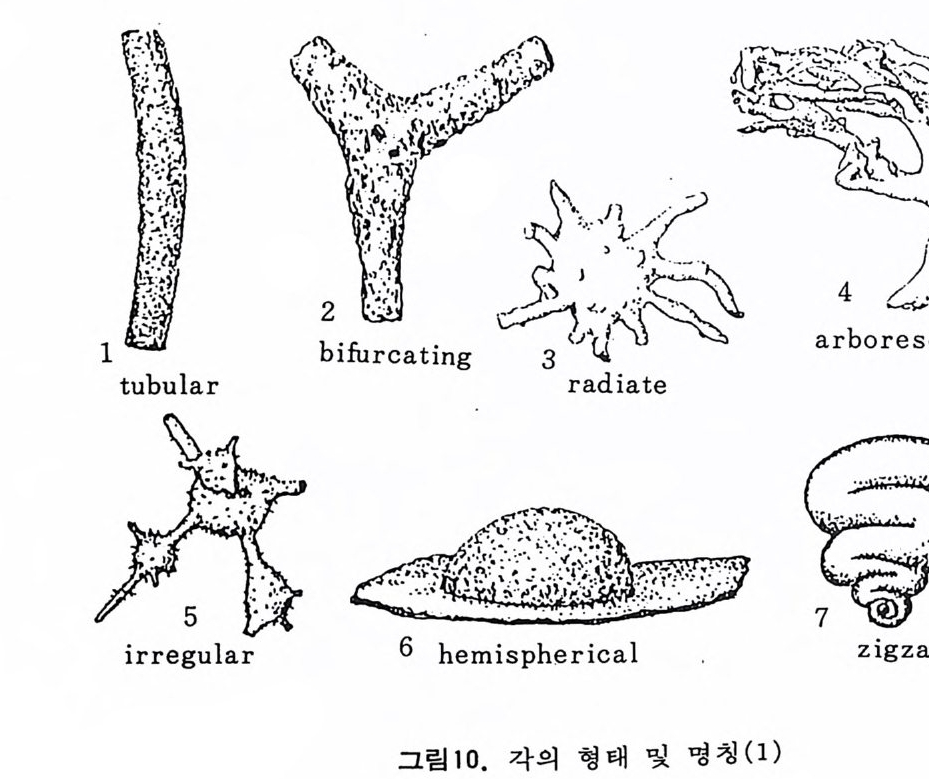
그림 7,8 에서 보듯 방 형태를 칭하는 이름이 언제나 3 차원적 표현 만은 아 니 다. 예로 sph eric a l (구형 ), pyr ifo rm (서 양배 형), clavate (곤봉형) , fist u l os e (부정 형관상) 둥은 입 체 적 형 태를 가리 키는데 반해 annular (年輪狀), rad ial - elo ngat e( 放射管狀), cunea t e( 쐐기 형). equ it an t (逆 V 형)등은 한 측면에 서 관찰 된 형태를 표현한 것이다. 방의 배열은 개체의 성장에 관계없이 같은 양식으로 배열되는 것과, 성 장에 따라 변화를 보이며 배열되는 것 동 2 가지 방법으로 구분할 수 있다. 이같은 방 배열의 차이는 유공충의 종족 유연관계를 밝히는데 큰 도움을 준다. 방의 경우 성장 축 (ax i s of g ro wt h) 이 직선이거나, 한 평면상에서 선 회, 혹은 선회면으로부터 선상으로 변화하며 성장하는 것이 보통이다. 성 장시 방의 배열방식은 그림 9 와 같다. 방의 배열양식에 대해 간단히 설명하면 다음과 같다. A) 선상배열; a) Un ise rial recti line ar (단열 직 선상) ; 성 장축 이 1 열로 직 선 . b) Un ise rial arcua t e( 단열호상) ; 성장축이 1 열로 곡선. c) Bis e ria l (2 열상) ; 좌우 교대로 방이 발달. B) 평면선회상 배 열; a) Plani sp i ra l, evolute ; 初室에 서 최 종 방까지 의 선회 가 보 이 며 좌우대 칭. b) Plan isp i ra l, inv olute ; 최 종 방의 선회 만 보 이 며 좌우대 칭 . C) 나선상 배열; a) Trochos pi ral( 소라나선상) ; 한면에서 보면 최종 선회만이 보이고 반 대 면에서는 전부 보인다. 전자를 spi ra l sid e , 후자를 umbil ica l s i de 라 하며 중십부의 우묵히 들어간 곳을 umb i l i cus 라 한다. b) Trise ria l (3 열상); 소라나선형의 일종으로 한 선회는 3 방으로 구성 D) M i l i ol i ne 상 배 열 ; 1 선회가 2 방으로 구성되는데 이 선회연 자체가 연속적으로 120:14 4: 때론 180° 등 각도로 회전한다. M ili olacea 에 속하는 종류들이 이 배열 을 갖는다. E) St re p tos pi ra l (연쇄 나선상) ; 성장도중 평면선회상(p lan i s pi ral) 혹은 소라나선상의 선회축이 꼬인 것을 말한다. 그외 전혀 규격성 없이 배열되거나, 복합된 형태를 보여주는 것이 있다.
 i\!`t:‘:: ,;-l\g'多::~ ;令F.· 广 ·· \갑I...?i \군, ·: ’·'}. ., ..은• \:.e•\ . -: i;; ,::-:::•훈:\'.. .! .-·,-\ . :. t동 .’ •.\ • ,•묘...; ·국:'.:1, •·?\✓. ' '집15 ; · 2b8\riE ..,/ .·':\:~. .• :·‘ g,..,: .. 1켰 . ? ;',\. ’':.1. i > ...· .' ,, •;.. '.••-:,. ,•. . . ?. . .... ?.. -.; ·: ·^·
i\!`t:‘:: ,;-l\g'多::~ ;令F.· 广 ·· \갑I...?i \군, ·: ’·'}. ., ..은• \:.e•\ . -: i;; ,::-:::•훈:\'.. .! .-·,-\ . :. t동 .’ •.\ • ,•묘...; ·국:'.:1, •·?\✓. ' '집15 ; · 2b8\riE ..,/ .·':\:~. .• :·‘ g,..,: .. 1켰 . ? ;',\. ’':.1. i > ...· .' ,, •;.. '.••-:,. ,•. . . ?. . .... ?.. -.; ·: ·^·
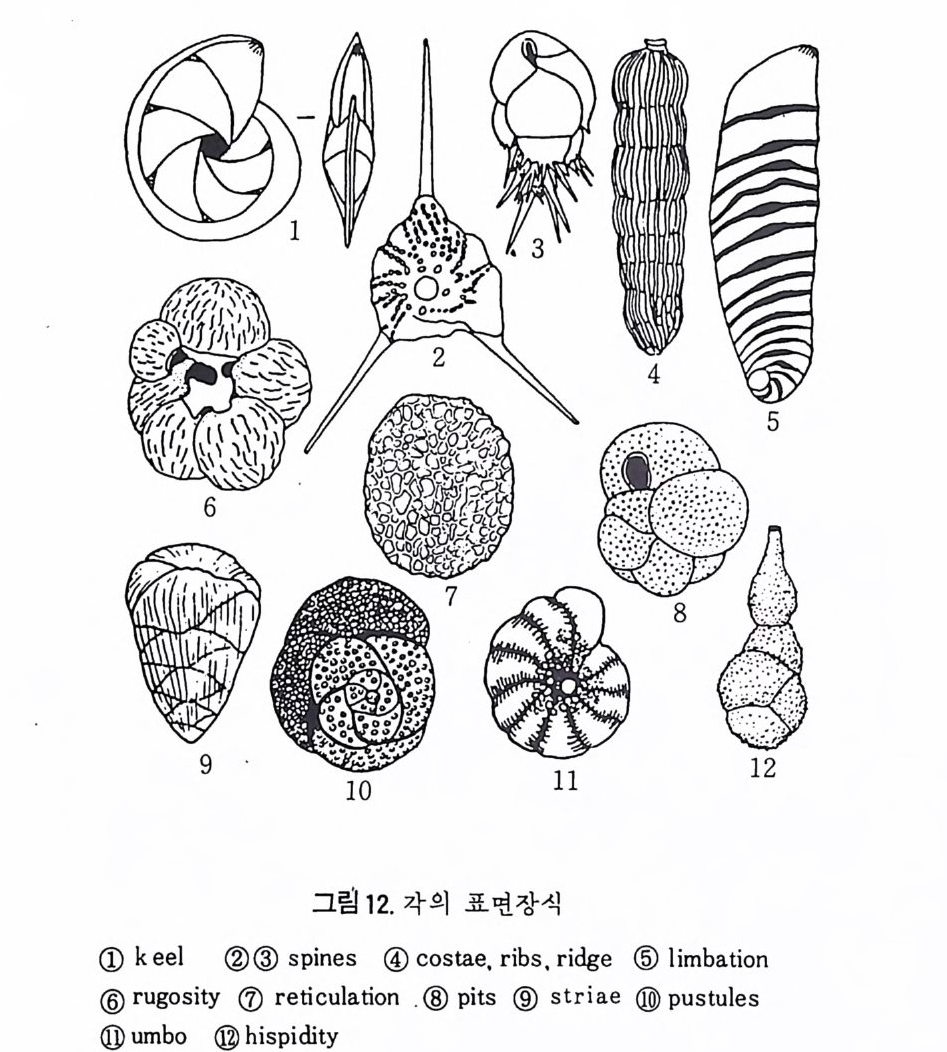
4) 穀의 형 태 및 표면 장식 (Ornamenta tion ) 각 방의 형태와는 별도로, 유공충이 성장하여 감에 따라새로운 방이 생 겨나면서 고유의 배열규칙을 따라 고유의 개체모습을 띠게 된다.이들의 형 태학적 명칭은 그림 10,11 과 같다. 석회질의 유공충은 각 표면에 다양한 장식이 발달하는 예가 많다. 그링 12 에서 보는 바와 같이 이들 표면장식은 모양이나 발달위치에 따라 여러 가지로 구별된다. 유리 질 유공충은 주로 rib s , rid ge , fur rows, spi ne s 등의 표면 장식 을 가 지며, 자기질 유공충은 str i a e , r idg es 를 갖는 경향이 있다. 표면장식의 생 태 표현형적 변이룰 같은 종내에서 볼 수 있다. 5) 벽 공 구조 ( P erfo r ati on ) 벽공구조란각벽에발달하는구멍을총칭하는것으로般내의의 세포질의
 1 bif ur cati ng 3
1 bif ur cati ng 3
 ,4‘
,4‘
동로로서의 기능을 갖는다. 호흡이나 배출작용과도 관계가 있으며 주로유 리 석회질 穀에 현처히 발달되며 각 구멍 하나하나를 벽공이라 한다. 벽공 의 크기는 穀내의 장소 및 성 장단계에 따라 변화한다. 저서성 -부유성 유 공충 모두 종에 따라 벽공의 크기나 단위면적당의 수가 틀리다. 벽공의 분 포양상(p ore fa br i c) 은 분류학적으로 중요시되어 가는 경향이 있다 . 층상벽 구조를 갖는 유공충의 경우 층의 두께가 증가함에 따라 벽공이 외부로 연
장되 어 canal i culus 를 형 성 한다. 6) 도관계 (Canal sy s te m ) 와 Alveoli 도관계란 유리 석회질 각의 穀 구성물내에 발달하는 가느다란管狀의 복 합체를 의미한다. 층내에 발달하는 int r a lamellar canal 과 층간에 형성되는 int e r lamellar pa sa gf' s 도 노로』 d 포함시킬 수 있다. 도관제는 分岐 또는 합류하며 복잡한 연락망을 구성한다(그림 13 참고).
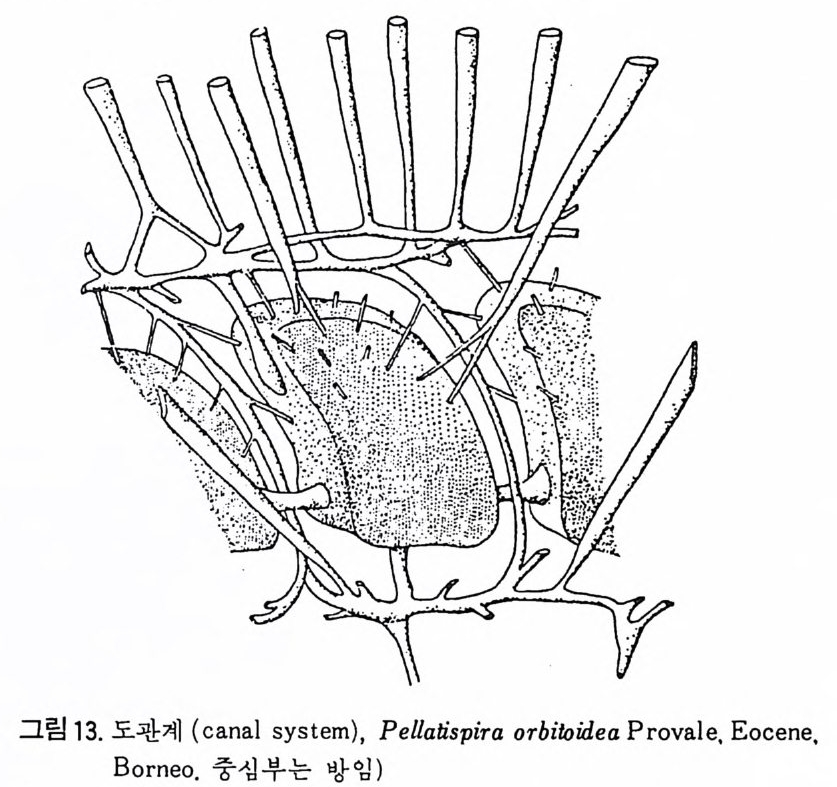 그림 13. 도관계 (canal sys te m ), Pellatis p ira orbito ide a Provale, Eocene,
그림 13. 도관계 (canal sys te m ), Pellatis p ira orbito ide a Provale, Eocene,
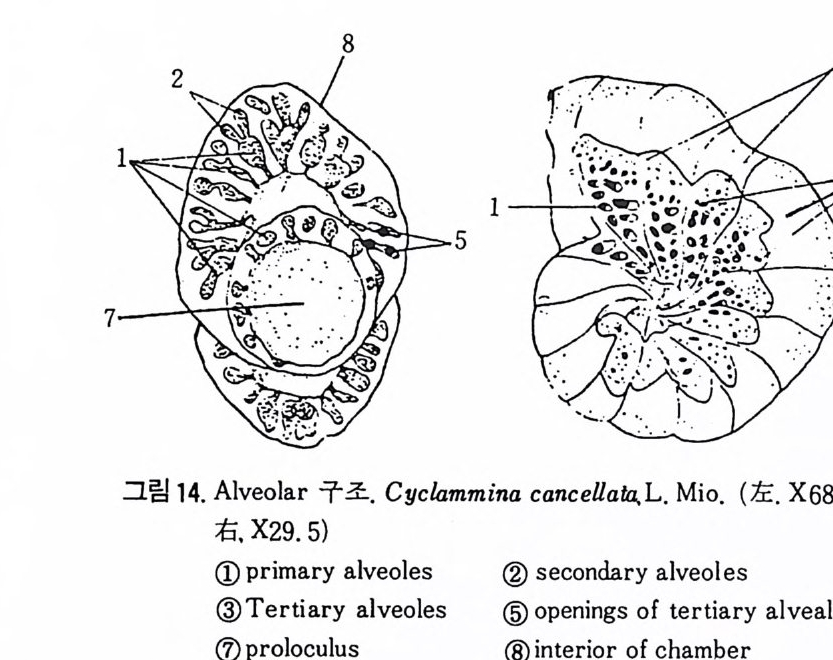
미립질, 자기질 및 교착질 각을 갖는 유공충의 일부는 alveol i라는 기포 상의 공동을 각벽 내부에 가지며 이룰 alveolar 구조라 한다. (그림 14 참고). 교착질각을 갖는 것 중 일부는 성 장에 따라 alveol i가 중가하며 서로 연 결되어 수지상을 형성하는데 이를 미로상(l ab y r i n t h i c) 구조라 한다. Alveoli 는 여러 방향으로 발달하나 보통은 각벽면에 대해 수직인 경향이 있으며 이에 대한 기능적 의의에 대해서는 아칙 불확실하다. 7) 口 .fl (Ap e rtu re) 구공이란 유공충 각 내외부로 세포질이 이동하도록 연결된 동로로 방의
 8
8
형성과 동시에 형태, 위치, 수 등의 존재양식이 정해지는 유전적 특성을 갖는다. 따라서 구공의 형태학적 특성은 종 • 속 등의 계통 분류상 중요시 되고 있다. 구공에는 기본적인 pr im ary ap er t ure 와 supp le menta r y 혹은 accessory a p er t ure 라고 불리는 부차적 구공이 있다. 구공 위치는 일반적으 로 방의 형태나 배열 등에 의해 결정되므로 단열상 (un i ser i al) 의 배열을 갖 는 경우는 마지막 방의 말단부 부근에, 평면선회상의 경우는 성장방향에 향한 면의 기저부나 면상 혹은 주변부에 발달한다. pr im ary ap er t ure 의 형 태, 위치는 그림 15 와 같다. 이들은 위치에 따라, te rmi na l, subte rmi na l,areal, int e r io m argi na l, equ ato r ial , umbil ic a l, spi ro umbili c a l, extr a umbili c a l-umbili c a l 등으로 구분되며, 형 태에 따라 slit, cir c ular, sic k le-shape d, pe ar-shape d, bott le neck-lik e (ph ia l in e ) , dendriti c, radi al , crescenti c, hooded, bop -sh ape d, crucifo rm , sie v e-lik e (crib r ate ) , arched 및 이 들이 복합된 mult iple -ape rtu r e 동으로 구분된다. 그림 16 은 sup ple menta r y ape rtu r e, accessory ape rtu r e 및 구공의 변형구조 룽 보여주는 것으로 그림에서 보이는 바와 같이 relic t ap er t ure 란 pr im ary ap er t ure 가 새로 형성되는 방에 의해 덮이지 않고 남겨져 표면에 노출된 것 을 말한다. 구공 혹은 umb i l i cus 를 덮는 것으로는 bulla 나 t e gi lla 가 있다.
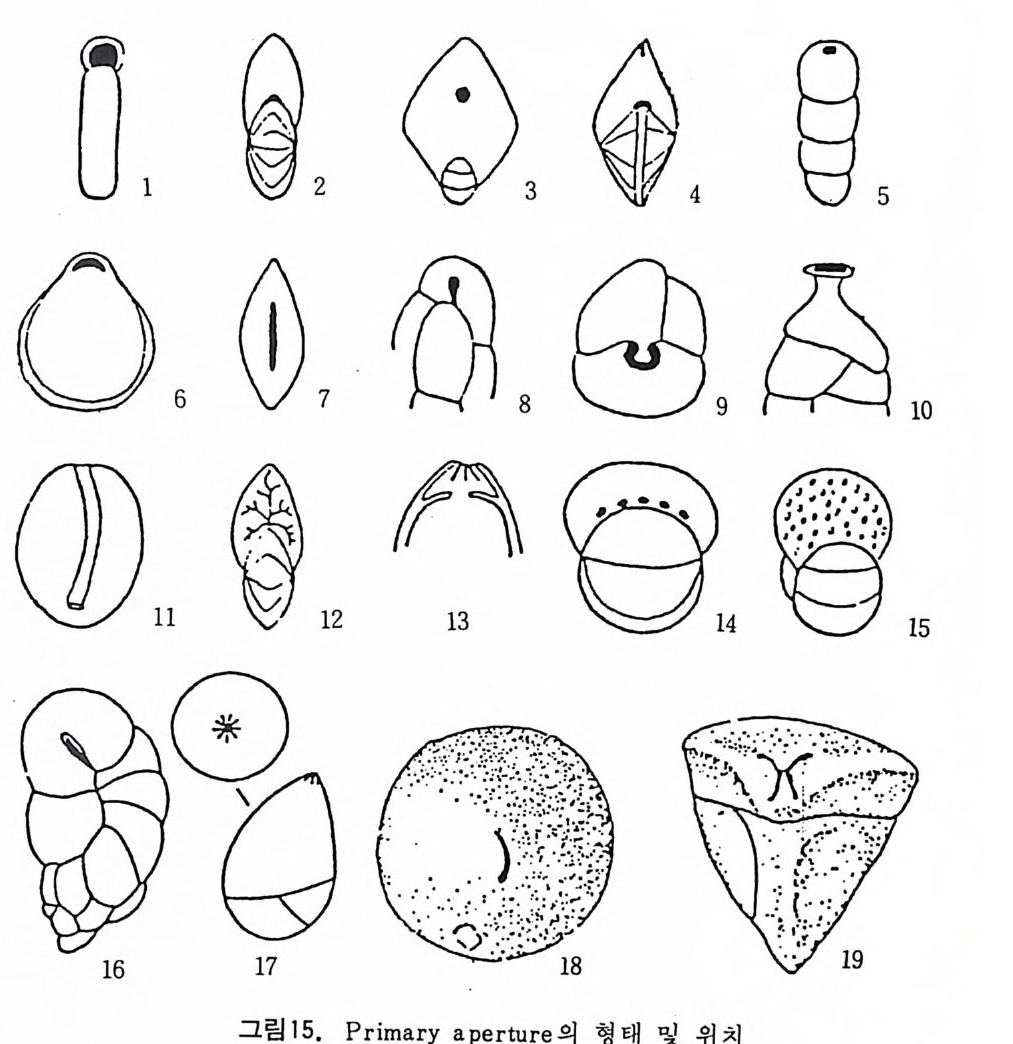 「 1 \ :3 ` :5
「 1 \ :3 ` :5
CD fre e end of a spi ra l tub e ;@basal, slit- s hap e d ape rtu r e ;@ape rtu r e in cent - ®re coifr c ufrloanr ,t a l tepr lma ni en,a ls oa-pc ea rllteu dr e a ;r@ea ls iac pk e l er-tsu hr aep ; e@d , basusablt e ar nmdi npa el raipp he errtau lr ea p; ~ e r tsul r ite- s ; lik e , ter mi na l ap e rtu r e seen from above ; @pe arshape d ap e rtu r e ;@ semi circ ul- ar ap e rtu r e wi th too th ;@ bot tle neck-lik e ap e rtu r e ;@ape rtu r e wi th ento s olenia l tub e ;@dendriti c ap e rtu r e ;@ secti on thr oug h a radia l ter mi na l ap e rtu r e wi th ap e rtu r al chamber whic h is connecte d to the orig ina l chamber by a cir c ular orifi ce ;@ multip le ap e rtu r e at the base of the fron ta l area ;@ sie v e-lik e (crib r - ose) ap e rtu r es.@) loop - shape d ap e rtu r e @ radia l ap e rtu r e @ crescenti c ap e rtu r e @) crucif or m ape rtu r e
 2홀 큐
2홀 큐
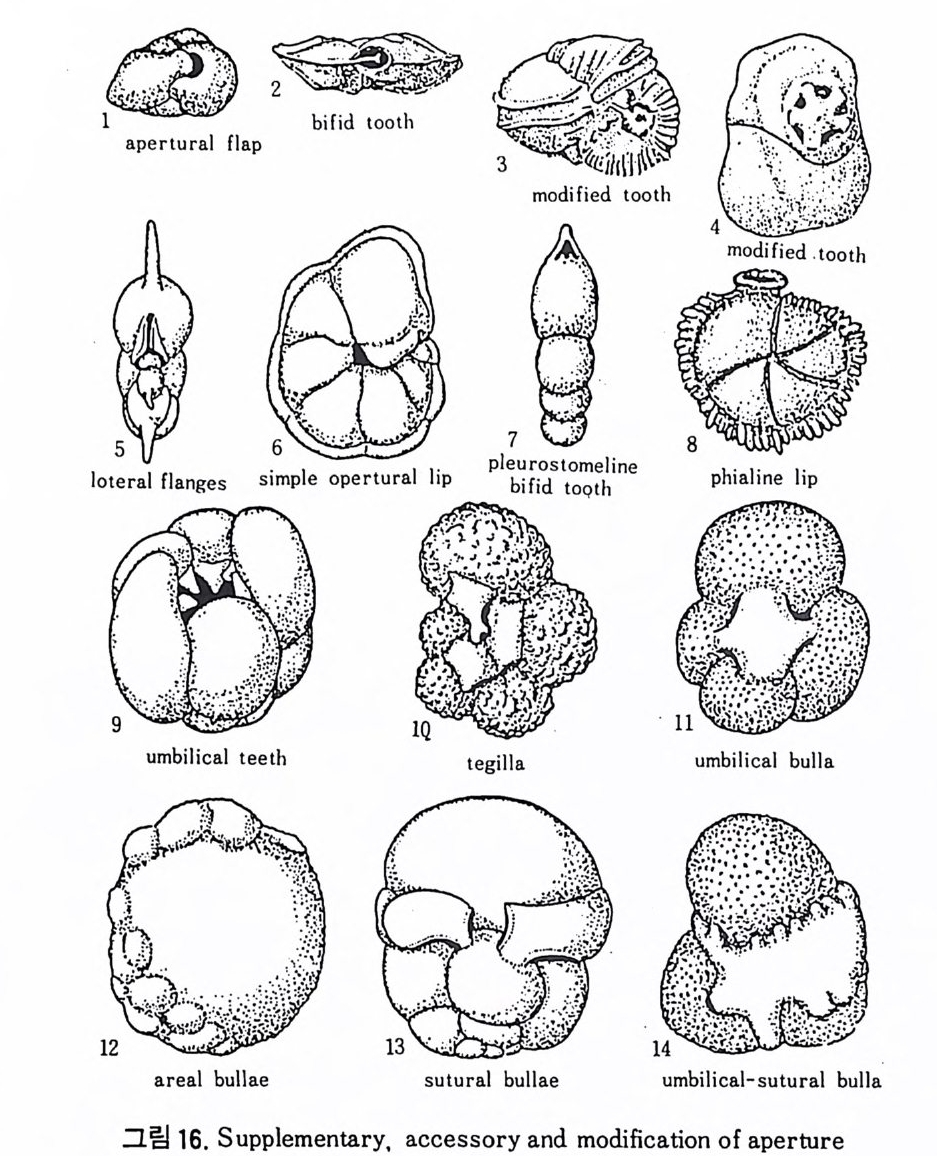
구공의 변형은 크게 외부변형과 내부변형 둘로 구분할 수 있으며 내부 변형에 대해서는 좀더 뒤에서 다루기로 한다. 외부변형으로는 그림 16 에 서 보는 바와 같이 ap e rtu ral lip , flan g e, bulla, teg illa, ap e rtu ral to ot h, ap e - rt ur al flap , umbil ica l t ee t h 동을 들 수 있다. 치아는 형태에 따라 sin g le too th , bif id too t h, comp le x to ot h 동으로 구별되며 다른 특징들과 함께 種 의 구분에 큰 역할을 한다.
8) 치 판 (Tooth Plate ) 치 판은 ent o s oleman tub e, hemi cy l in d ric a l sip h on, secondary par titi ons 와 함께 구공의 내부 변형에 의한 것으로 유공충의 방내에 존재하는 특수구조라 할 수 있다. 부분적으로는 방의 내벽이나 구공 혹은 Iamella 에 부착된다 . 구공 에 일부분이 나와 구공치아 (a per t ural t oo t h) 처럼 보이기도 한다. Hog lu nd (1947) 에 의해 처음으로 발전되였으며 Ho fk er 에 의해 명명되였다. 이는 방 과 동시예 형성되며 시간이 흐름에 따라 복잡한 형태로 변하는 경향이 있 다. 기능에 대해서는 아직 밝혀지지 않았으나 형태 잇 위치는 분류상 중 요시되고 있다. 의키틴질 각을 갖는 유공충의 경우 ento s olenia n tub e 라고 하는 관상구조가 구공에서 방 내부로 연결되고 있으며 치판과는 구별되고 있 다(그림 17 참고). 9) 회전방향 소라나선상의 배열을 갖는 경우, 방은 오른쪽 혹은 왼쪽 방향으로 감아 울라가는데, 유공충에 있어서 이 방향은 시간적 내지는 지리적 변화에 따 라 재미있는 회전경향을 보여준다. 한 예로 지리적 변화에 따른 회전경 향을 보자(그림 18 참고). 일반적으로 회전방향은 수온과 관계가 깊으며,난 류성인 경우 오른감기, 한류성 해수에는 왼감기가 우세하다 . 회전방향을 달 리하는 개체의 비율을 이용하여 古 기후의 변화 및 지층의 층위를 알아낼
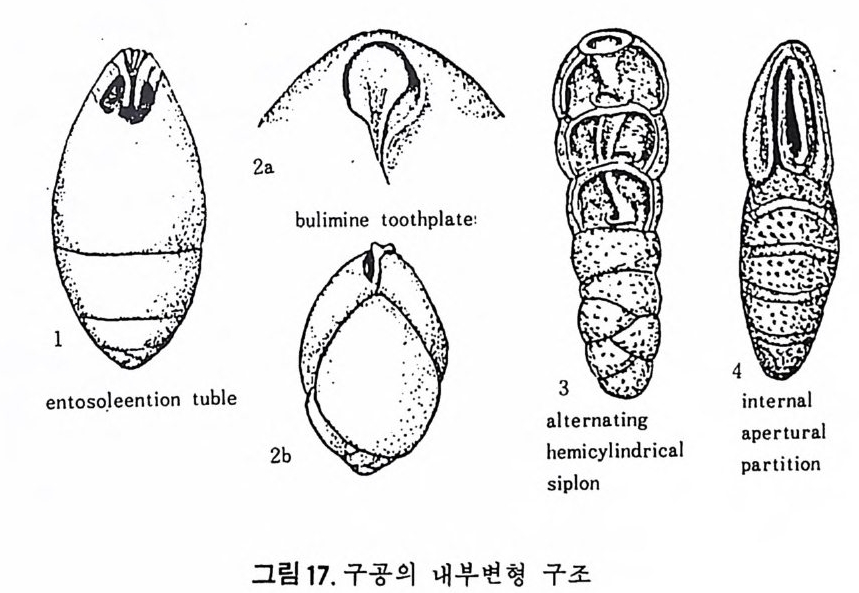 buli mi n e too th p la te ·
buli mi n e too th p la te ·
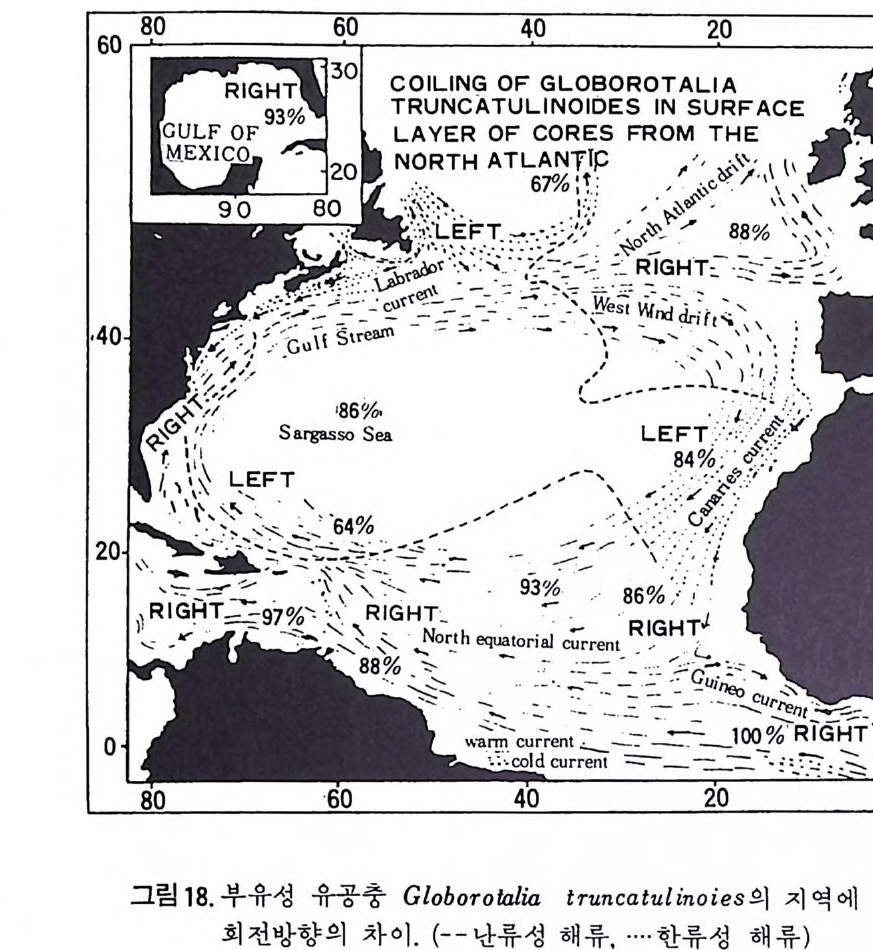 60 80 60 40 20 。 60
60 80 60 40 20 。 60
순가 있다. 한류성 종으로 널리 알려 진 Globig e rin a p ach y derma 의 경우 극 해 근처에서는 거의 100% 가 왼감기 를 보이나 중위도로 오연 오른감기를 보이는 개체가 증가한다. 1 • 3 • 2 분류 유공충의 분류는 다른 생물체들과 달라 연체부의 형질과는 무관하게 거 의 전적으로 穀에 의해 분류되고 있다. 이러한 각에 의한 분류는 유공충의 생활사나 배우자 및 핵의 성질 동 생물학적 특성에 의한 분류와도 찰 일치되고 있으므로 화석을 대상으로 연구하는 고생물학자에게 있어 다행스러운 일이라 할 수 있다.
지금까지 많은 학자에 의해 유공충의 분류가 이루어졌으나 그 들 은 분류 의 기본적 개념 및 기 준 에 차이가 있으므로 분류 역시 다소의 차이룽 보 이 고 있 다. 현 재 Loeblich and Tapp a n (1958) 에 의 한 분류 가 가 장 널 리 사 용되고 있다고 할 수 있으며 그외에도 금세기 후 반에 들어 Pokorny (l95 8), Reis s 0958, 1963) , Hofk e r (195i) , Sig a l (1952) , Raus er- Chernous ov a and Furse nk o 0 959) 등에 의한 새로운 분류가 시도되였다. 유공충의 실제적인 분류에 앞서 간단히 분류의 역사를 살펴보기로 한다 봉란서의 d ' Orb ig n y ( 1826) 에 의해 처음으로 유공충을 포항한 대규모의 분 류가 이루어 졌으며, 유공충을 目 (order) 단위로 놓고, 5 科(f am il y ),52 屬(g e nus ), 544 種 (spe cie s ) 으로 구분하였다. 그는 방의 배영에 기초하여 科룽 구분하였다. 영국의 W illi am so n (1 858) 은 穀 및 각벽의 형태학적 성 질을 중요시하여 분 류하였으며 자기 질, 유리 석 회 질 잇 교착질각 등 3 그룹을 구분하기도 하 였다. 독일 Schul tz e (1 854) 는 각벽의 유우에 의해 2 분할 것을 제창하였다. Reuss(l862) 는 이의 전을 따랐으며 동시에 W i ll i amson 의 3 구분융 합해 재구 분하였으며, 한편 방의 모양 및 배열을 고려하여 많은 科(f am il y)를 설정하 였다. 2 0 세기에 들어서면서 석유개발을 위한 지층연대 결정에 유공충 화석이 널리 이용되면서 유공충에 대한 연구 여시 후[발해졌다. Cuslunan 은 당시의 지도적 연구자로서 연구 초기에는 Brad y를 따라 10 과로 구분하였다. 그 후 1927 년에 45 과로, 1948 년에는 50 과로 구분하였다. 한편 Glaessner(1945) 는 穀의 일반적인 형태와 방의 배열을 가장 중요한 분류기준으로 삼았으며, 각벽은 같은 군 내에서도 다양한 변이룽 보이는 것 으로 해석, 37 과와 7 上科 (su p er fa m i l y)로구분하였다 ;As t rorh i z i de a, L it u olid e a, Endoth y ri d e a, Mi lio l i d e a, La ge nid e a, Bulim in id e a, 밋 Ro t aJ ii dea 로 Astr o r- liiz i d e a , Li t uol idi ea 는 교 착 질 각, M il i ol i dea 는 자 기 질 각, La ge nid e a, Bulim i n id e a, Ro t al ii d e a 는 유리 석회질의 穀을 갖는다. Hofk e r (1951) 의 경 우, 지 금까지 의 目 (order) 위 치 에 서 亞網 (sub class) 으로 유공충의 분류위 치를 바꾸었으며, 그 亞綱 중 Denta ta 目울 신선하 여 이 물 다시 Proto f o r ami na ta , Bif ora mi na ta , De~t er ofo r ami na ta 등 3 아목으 로 나누었다. 그는 치판 및 구공을 가장 중요시하였으나. 종래의 분류 기 준이었던 각벽의 격벽의 구조나 방의 배열 등을 소흥히 생각한 비교적 독
독한 분류물 보이고 있다. 그의 분류는 지질시대에 따 른 산출상황 맛 명 명규약 등을 무시하고 있어 많은 비판 을 받고 있으나 반면에 殺의 내부구 조 연구에 공헌한 바가 크다. Sig a l (1952) , Pokorny 0958) 의 체 계 는 Glaessner 의 분류 기 준을 일 부 개 량 했다고 할 수 있다. Smout (1954) 는 Glaessner 에 의 한 Ro t al ii dea 중 2 중격 벽 으로 도관계 가 발 달한 것을 Rota l iid e a, 격 벽 이 한 겹 으로 된 것을 D i scorb i dea 라 하여 둘로 나누었다. 부유성 유공충의 경우, 1950 년대에 들어와 분류작업이 시작되었으며, 층 서적 및 지리적 분포가 조사되고 진화계열 연구가 시작되었다 . 1964 년 Loebl i ch 와 Ta ppan 에 의한 분류는 지금까지의 것 중 가장 종합적 체계를 갖추고 있다. 분류기준에 대해서는 좀더 뒤에서 자세히 설명키로 하겠으며 우선 간단히 살펴보면 각벽의 구성과 미세구조에 의해 유공충목 울 亞 n 이나 上科로 분류하고 있다. 각벽의 광물학적 조성과 미세구조, 각 벽의 층상구조, 구공. 치판.… 등의 각의 형질에 의한 새로운 분류기준이 라 할 수 있으며 17 상과 95 과 약 1, 200 屬 27, 000 종으로 구분되고 있다. 최근 들어 좀더 많은 특성, 예를 들어 각벽의 조성 및 미세구조, 방의 배 열ha b口i4t a t형) ,태 원, 형구 질공, 의개 체형 변태이 및(o n t변 o g이 e성ne. ti c 생ch활an습g e성 s) , 맞생 식서 (식re지p r o (dliufec t i ohn a)b.i t지 질an학d 적 분포범 위 (ge olog ic rang e s) 등을 고려 하여 분류하는 경 향이 있다. 유공충의 분류에 있어서. 각 군의 분류 기준으로 사용되는 형질, 즉 형 태학적 특성이 모든 분류군에 공통되는 것이 아니라 각 분류단위마다 다 르기 때문에 많은 곤란이 따르고 있으나 일반적인 가준이 되고 있는 분류 형질은 다음과 같다. 亞目 및 上科는 각벽의 구성과 미세구조에 의해 구분되고 있는데. 이는 각을 분비하는 원형질의 생화학적 성질에 의해 결정되므로 중요시되고 있다. 교착질각 혹은 미립짐각을 갖는 유공충의 경우 방의 형태에 의해 상과 가 구별되나 유리 석회질각을 갖는 유공충에는 적용되지 못하고 있다 . 과 는 방 및 격벽의 형태. 방의 배열, 口 1L 의 기본위치와 치판 등을 포함한 장식 등에 의해 구분되며, 속의 경우 방의 형태 및 배열, 생황양식 등에 의해. 종의 경우 각의 크기, 각과 방 및 구공 등의 상대적 비율, 표면장식 등에 의해 구분되는 경향이 있다.
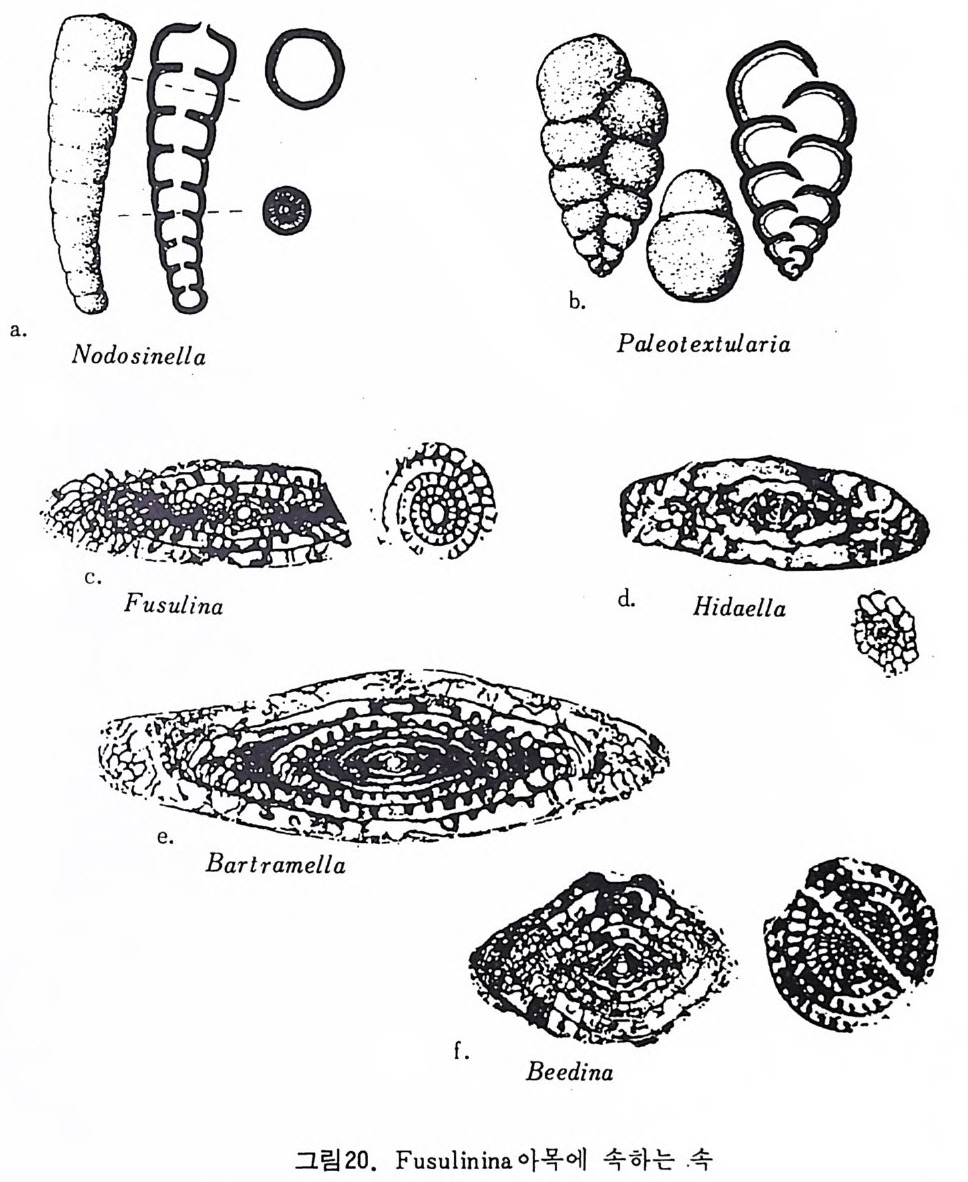
여기에 수록된 분류체계는 Loeblic h and T app an(1964) 의 분류에 기초하 고 있다. 유공충은 각벽의 조성에 의해 5 아목 (sub order) 으로 구분된다. Allog ro mi ina , Textu l arim a , Fusulin i n a , Mi lio l i n a 및 Ro t al ii na 로 이 는 다시 17 개의 상과로 구분되는데 Allo g rom ii na 는 Membranous 혹은 의키던질각을 가 지 며, Tex t ular i a 는 주로 교착질각을, M i li ol i na 는 자기 석 회 질각을, Fusuli- nacea 는 석회 미립질각을 Ro t al ii na 는 유리다공 석회질각을 갖는다. Allo grom ii na 아목 : Membranous 혹은 의키탄질각으로 되었으며, 드물게는 산화철외피, 교착물질을 갖는 것이 있다. L agy nacea 상과 : 단실로 tub ular, round, fl ask 모양을 한다. 고생대에서 현 세에 걸치며 대부분이 저서고착성 (be nth i c sess i le) 이고 일부는 이동성 이다(그림 19-a . b). Te xt ular ii na 아목 : Cemen t에 외부물질이 부착, 죽 교착질각을 갖는다. Ammo disc acea 상과 : 多 宰 이며, non-sep tat e 혹은 p ro t ose pt a t e 이다. Tee- tino 11s t es t s 로서 외층은 대부분이 부착성 물질로 되어 있으며 고생대 에서 현세에 걸쳐 산출된다. 일부는 이동성이나 대체로 저서고착성이 다. Astr o rhiz id a e, Amm odi sc i dae 과가 이에 속한다(그림 19-c). Lit uo lacea 상과 : 多室로 격 벽 이 발달, 연속적 (seria ll y) 혹은 나선상 (s pi rall y)으로 배열한다 . 하나 혹은 여러 개의 구공울 보이며 일부는 치판 을 갖는다. 미립석회질각을 가지며 대부분이 부착성 물질로구성된다. 일부는 의벽공을 가지며, 내부는 비교적 복잡하고 고생대에서 현세에 걸쳐 발달하며 대부분이 저서이동성이며 극히 일부가 저서고착성이다. Rzehakin i d a e, Li tuo lid a e, Textu l ariid a e, Trochammi nid a e, At ax op h rag m- iida e, Orb it ol i n i dae 과 등 이 이 예 속한다(그림 19-d e. f. g. h). Fusulin i n a 아목 : 각벽은 미립 석회질각으로서, 원시적 형태의 경우 미립 상의 방해석으로 되어 있고, 진화한 것은 둘 이상의 분화층으로 되어 있 다. Endoth y r acea 상과 : 多室. 로 구성되었고 격벽이 있으며, 나선상 혹은 연 속적인 배열을 갖는다. 하나 혹은 여러 개의 구공을 보이며 각은 석 회 미립질로 간혹은 의섬유질, 원시적인 double-laye r, 교착성 물질 등 으로 구성된 종류가 있다. 고생대에서 트라이아스기에 걸쳐 산출되고 . 있으며 저서고착성 혹은 이동성이다. Nodosin e llid a e, Colanie ll i da e, Pt ych oclad iida e, Paleote x tu l ariid a e, Semi -
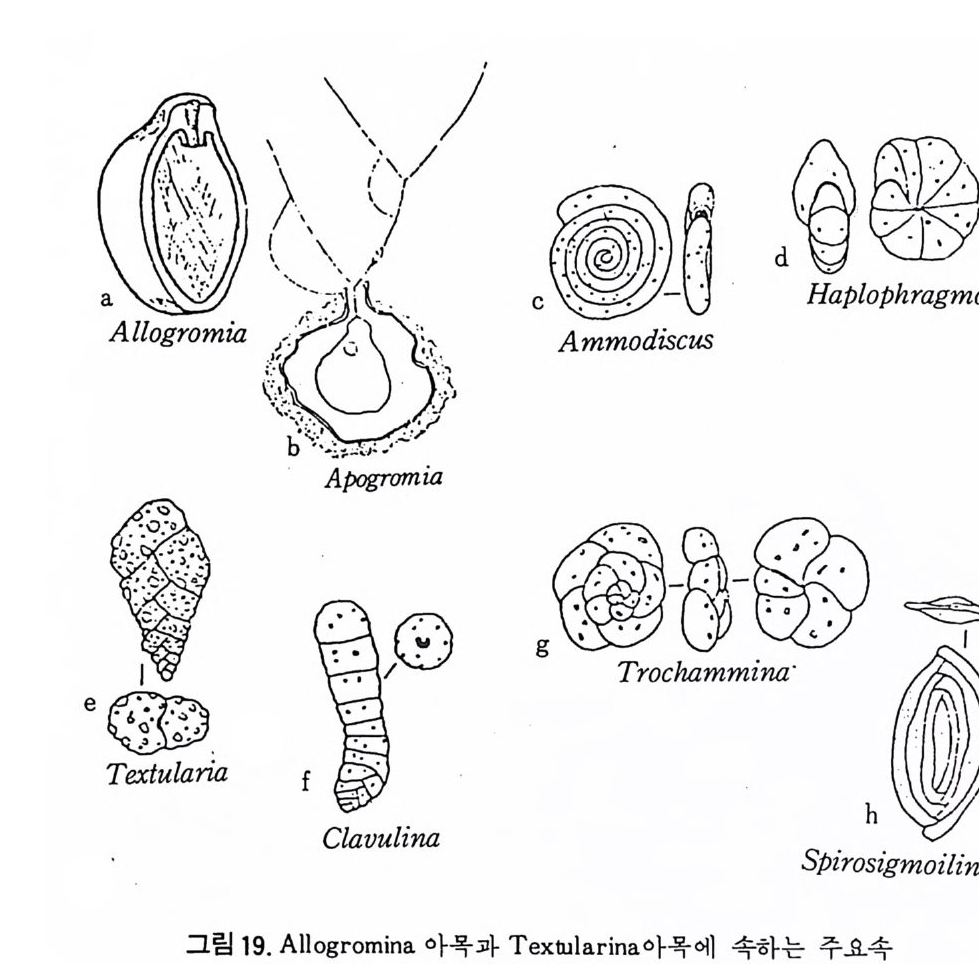 \\'\:[
\\'\:[
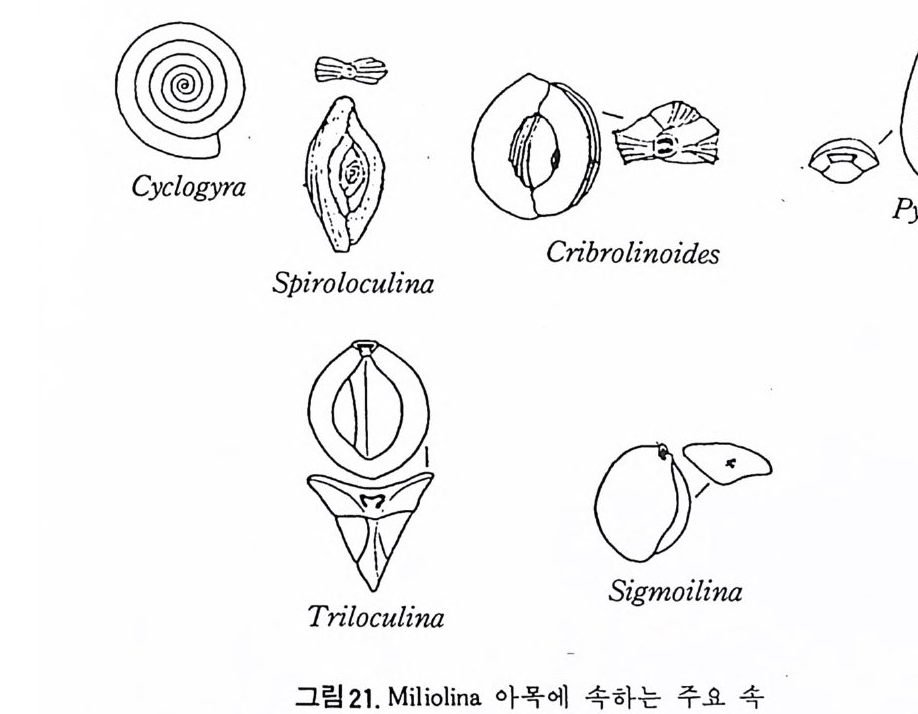
tex tu l ariid iae , Tetr a ta x id a e, Bis e riam mini d a e, Tournaye l lid a e, Endoth y ri - dae, A~chaedi sc id a e 및 Las iodi sc i dae 과 등이 이에 속한다(그림 20-a.b). Fusulin a cea 상과 : 다실로 격벽구분을 보이며, 평연선회상 배열을 갖는 방추형으로 여러 개의 구공을 갖는다. 석탄기에서 페름기에 걸쳐 산 출되며 저 서 이동성 이 다(그링 20-c. d e. f). Mil ioli n a 아목 : 자기 석회질각으로서 초기부분을 제외하고는 무공질로 되 ’ 어 있다. Mil io l acea 상과 : 다실로 대부분이 격벽구분을 보이며 명 屬은 Proto s ep t- ate 형태를 갖는다. 나선 혹은 선회상으로 배열하며 하나의 혹은 다 수의 구공을 갖는다. 일부는 구공치 아나 multip le t oo t hp la t es 를 갖는다. 각은 석회자기질로, b i -la yer 이며 트라이아스紀에서 현세에 걸쳐 산충
 。
。
되 며 저 서 이 동 성 이 다. Fis c herin id a e, Nuberculariid a e, Mi liol id a e, Alveo - l i n i dae 과 등이 이에 속한다(그림 21). Rota lii n a 아목 : 석회질의 다공질 각을 갖는다.
 Cr ibr olin o id e s
Cr ibr olin o id e s
Nodosaria c ea 상과 : 다실로 격벽구분을 보이며, 연속 혹은 나선상의 배 열로, 구공은 하나이며 t erm i nal 상으로 간혹 ape rtu r al chamberle t를 갖는 . 다. 석회 방사상각으로 되어 있고 고생대 말에서 현세에 걸쳐 산충되 고 있 다. Nodosariid a e, Polym orph in i d a e, Glanduli nid a e 과 등 이 이 에 속 한 다(그림 22- a. b). Bulim in a cea 상과 : 다실로 격벽구분을 보이며, 소라나선상 배열을 갖는다. comma-shape d aper t ure 로 때로는 neck 위에 ter mi na l aper t ure 룹 보이며, 전부가 하나의 치판을 갖는다. Calcit ic radi al t es t s 로 구성되었으며 쥬 라기에서 현세에 컬쳐 산출되고, 대부분이 저서이동성(b en t h i c va gile ) 이 나 일부는 부유성 이 다. Turrili n i d a e, Sp h aeroid ini d a e, Boliv i n i t ida e, lslandi el lid a e, Eouv ige rin i d a e, Bulim i ni d a e, Uvig e rin i d a e 科 등 이 이 에 속 한다(그림 22-c. d). Camssmida u- lsi nhaa pcee ad a상pe과r t u: r다e 로실 로치 판격을벽 구갖분는다을. 보이석 며회, 입 복상합각배으영로을 구 성갖 는되 다었 으.C며 o 백악기 (Cre t aceous) 에서 현세에 걸쳐 산충되는 처 서이동성으로 Pleu-
rosto m ellid a e, Caucasin id a e, Cass i dul i n i dae 科 등이 이에 속한다(그림 22 -e. f). Nonio n acea 상과 : 다실로 격벽구분을 보아며, 평연선회상 혹은소라나선 상의 배열을 갖는다. lnte r io m argi na l 혹은 areal a p er t ure 로서 치판은
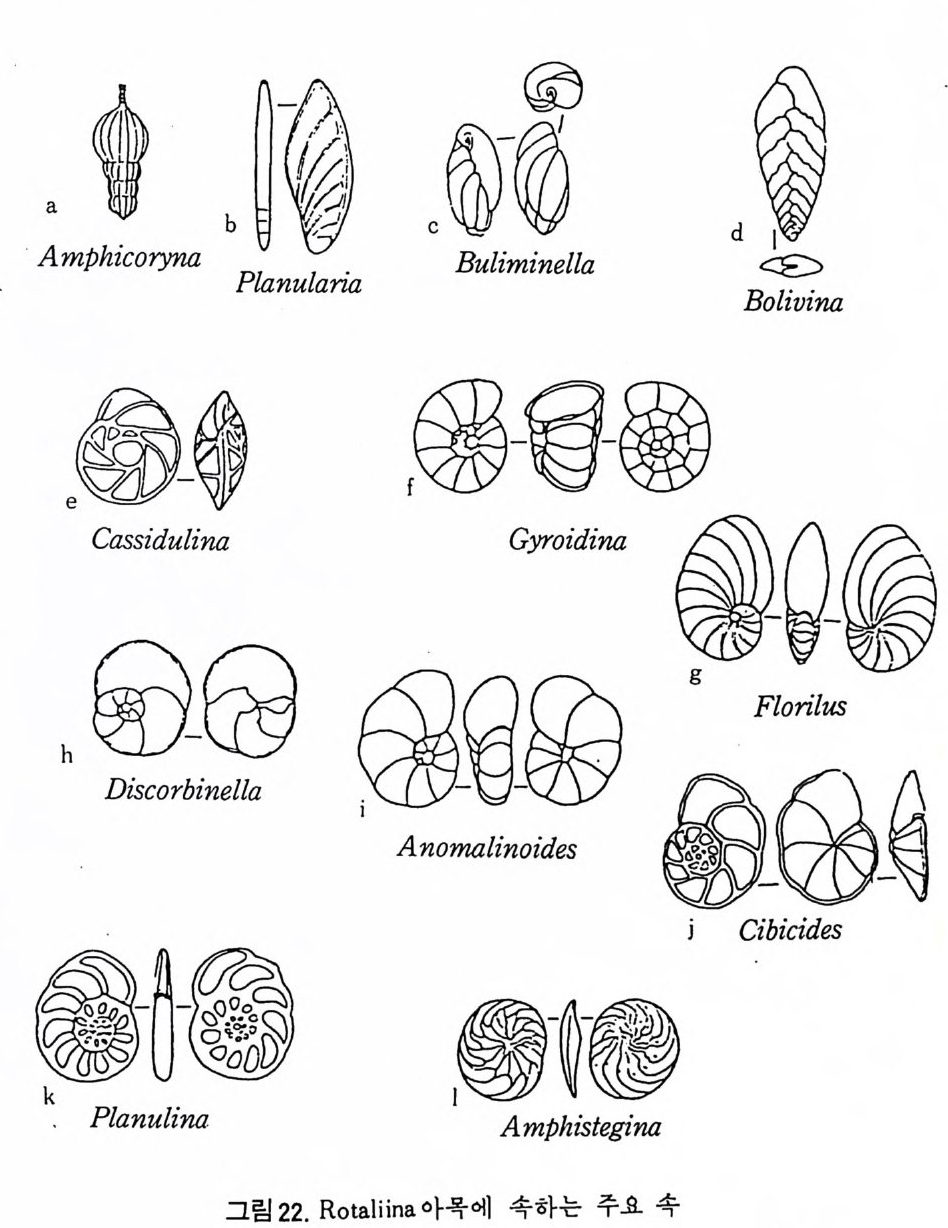 Aam p h i人c o ry na bP l a」n nul凡ari a c uB尤 lim @ @[B
Aam p h i人c o ry na bP l a」n nul凡ari a c uB尤 lim @ @[B
발달되지 않았으며 석회 입상각을 갖는다. 백악기에서 현 세에 걸쳐 산 충되는 저서이동성으로 Nonio n id a e, Alabam i n i da e 科 등이 이에 속한다 (그럼 22- g), Dis c orbacea 상과 : 다실로 격벽구분을 보이며. 소라나선상의 배열을 갖 는다. i n t er i omar gi nala per t ure 로 치판을 갖기도 한다. 석회방사상각으 로 구성되었으며 백악기에서 현세에 걸쳐 산출되 는 처서 이동성으로 Dis c orbid a e, E pi s t omar ii dae 과가 이에 속한다(그링 22). Anomalin a cea 상과 : 다실로 격벽구분을 보이며 , 소라나선상 배 열을 갖는 다. lnte r io m argi na l a per t ure 로 sup ple menta r y ap er t ures 를 동반키도 한 다. 석회 입상각으로 구성되었으며 백악기에서 현세에 걸쳐 산출되는 저서이동성으로 Osangu l ariid a e, Anomal i n i dae 과 등이 이에 속한다(그림 22-i) . Orbit oi d a cea 상과 : 다실로 격벽구분을 보이며, tro chospi ra l annular 혹은 c y cl i ca] 상의 배열을 갖는다. sin g l e aper t ure 로서 극히 일부가. 치판을 갖 는다. 일부 속의 경 우 chamberle t s 의 발달을 보이 며 secon dary chambe- rle t s 이 발달된 것도 있다. 석회 방사상각으로 구성되 었으 며 백악기에 서 현세에 걸쳐 산출되는 저서이동성 및 저서고착성으 로 Ep o nid ida e, Glabrate l lid a e, Amp h is t e g i ni d a e, Cib i c id ida e, Planorbulin i d a e, Cy m balop o r i- dae, Le pi doc y cl i n i dae 과 등이 이에 속한다(그림 22- j, k. I). Globig e rin a cea 상과 : 쥬라기에 출현, 현세에 걸쳐 산출되는 부유성 유공 충으로 다실로 되었으며 격벽구분을 보아며, 나선상 혹은 c y cl i ca] 한 배 열을 갖는다. Inte r io m argi na l a p er t ure 로, access or y 혹은 sup p le me n- tar y aper t ure 를 동반하기도 한다. 석회 방사상각으로 구성되었으며 Hete r ohelic id a e, Planomalin id a e, Schackoin id a e, Rota l ip o rid a e, Globot runcani- dae, Hantk e nin id a e, Globorota l iid a e, Globig e rin id a e 科 등이 이 에 속한다 (그림 23-a b. c. d e). Rota l i ac ea 상과 : 다실로 격 벽구분을 보이며, tro chospi ra l, pla nis p i ra l, cyc - li cal 한 배열을 갖는다. sin g l e 혹은 mult iple aper t ure 로서 too th p la t e 와 sept a) fl a p을 갖는다. 석회 방사상각으로 구성되었으며 백악기에서 현 세 에 걸 쳐 산출, 저 서 이 동성 으로 Rota l iid a e, Calcarin id a e, Elph i d iida e, Nummuliti da e, M iogyp s i n i dae 과 등이 이에 속한다(그링 23- f. g. h). Sp iri ll ina cea 상과 : 다실로, 초실에 연결된 처음 부분은 격벽이 발달되지 않았으며 뒷부분에서 격벽구조를 보인다. 방은 나선형으로 배열되고,
��� )������<�\� l�1��p� ��|�t�DŤ�0���� �8��� x��� ������� ��t�ٳ1�<�\� S pi r i l
l i ni add �e� t��� ��\��(���� 2 3-i). Roberti nacea ���� : ���\�, ����l���D� �t p�� ,�|���� ���X� 0���D� � ���. X�X� l ���<\�� � X���t � �� � ���<�p� @z@� aragonite \� l�1�� ��t�ٳ1<��\�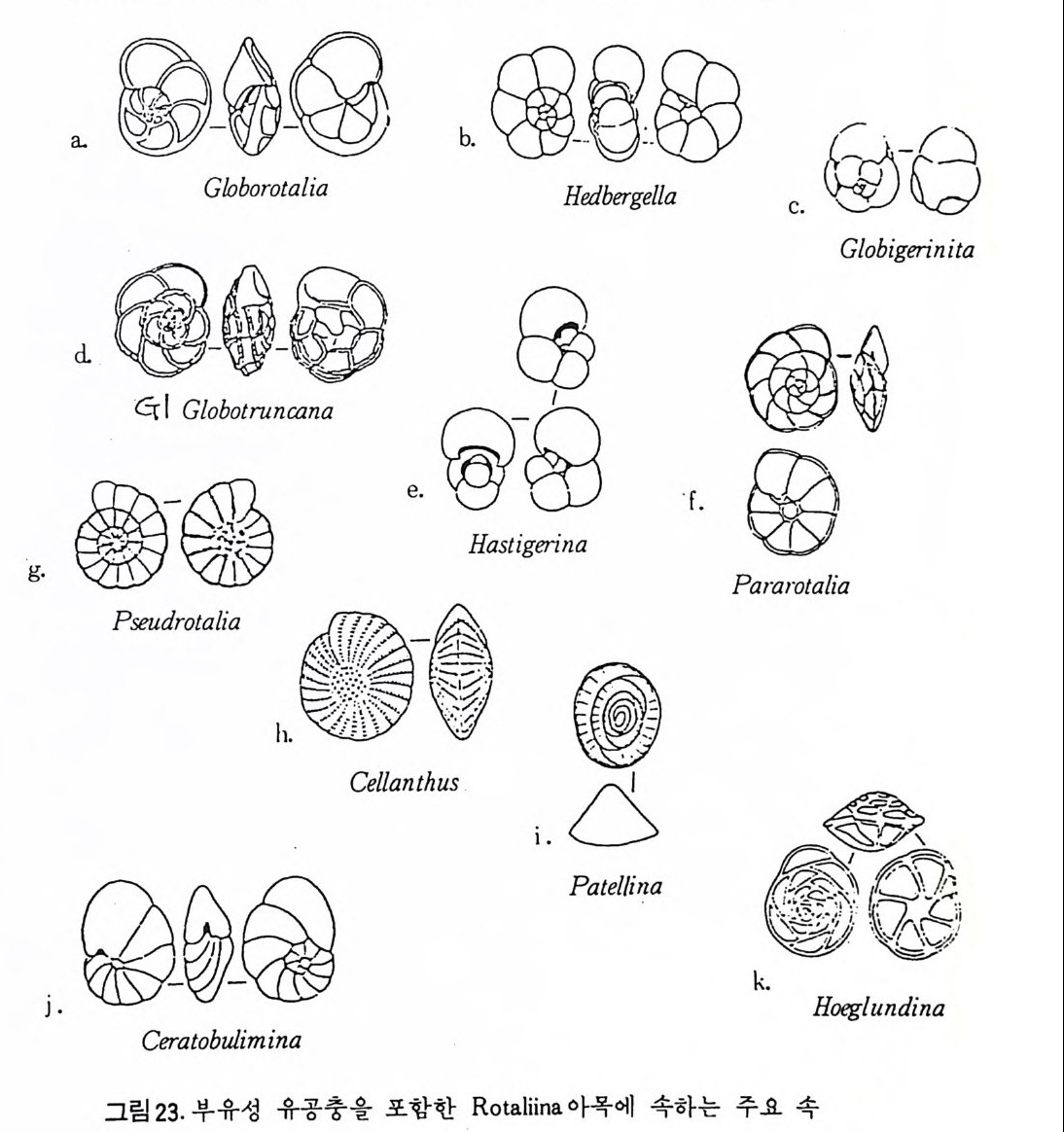 R�o� |�� �t�.ib nC�t eirdae}te oa����b ���l mui i�n�r�Di�8d �a��e ,�Ťrota�li. �}a A@ t.k)�� 23-� �� i.�\��(����T� tlobo�- :-G�a.
R�o� |�� �t�.ib nC�t eirdae}te oa����b ���l mui i�n�r�Di�8d �a��e ,�Ťrota�li. �}a A@ t.k)�� 23-� �� i.�\��(����T� tlobo�- :-G�a.
1 ·4 생태 및 고생태 1 • 4 • 1 생태 유공충의 대부분이 해수에 서식하며 조간대 같은 양은 지역으로부터 대 륙붕을 지나 수심 10,OOOm 를 넘는 해구에 이르기까지, 위도상으로는 적도 에서 국해에 이르는 세계적 분포를 갖는다. 대부분이 저서성이며 일부가 부유성으로서, 현세 유공충의 종이나 개체 의 분포상태는 대륙봉환경에서 가장 잘 연구되고 있는 실정아며, 이 지역 의 저생군집 분포에 대한 지리적 분화가 찰 알려지고 있어 생지리적 연구 에 이 용되 고 있 다. Allo g rom i dae 나 L ag en i dae 과에 속하는 일부 속은 담수성 이며 기수에 서식하는 종류도 있다. 유공충의 군생태학적 연구는 Nor t on(1930) 에 의해 시작되었다고 할 수 있다. 풀로리다 근해에서 서인도제도 근해의 현세퇴적물 중의 유공충의 분 포와 수심, 온도 등을 관련 연구한 결과 유공충은 과(f am i l y)단위로 심도 에 따라 현저한 차이를 보이고 있음을 지적하였다. Natl a nd (1933) 는 이 같은 현 세 유공충군의 생 태 학적 분포를 기 초로 과거 유공충 화석군집의 생식환경 및 이룽 포함한 지층의 퇴적환경 규명을 시도 하였다. Phlege r (1951) 는 생 체군집 ( I iving po pu l ati on ) 과 유해 군집 (dead po pu lati on ) 을 비교 연구하여, 퇴적속도나 사후운반,환경변이 등과 관련시킨 해석을 하였으며, 유공충의 분포가 심도나 온도 등과 같은 생태학적 제한요소 (limi ting fa c t or) 에 의해서만 좌우되는 것이 아니라 좀더 복잡한 요인들과 연관되어 있음을 지적하였다(그림 24 참고). ` 유공충의 생태를 연구하거나, 더우기 고생태를 밝히고 고환경을 복원하 는 데는 많은 난제가 놓여 있다. 유공충 군집에 있어 부유성과 저서성군 사이의 비율, 재형충 (os t racoda) 과 유공충과의 비율, 般의 성분비, 각 科(f am i l y)의 산출바 , 種 다양도 등 은 환경에 따라 변화룰 보이고 있으므로 이들을 이용하여 환경을 유추할 수 있다. 우선 유공충의 분포를 규제하는 환경요인으로 크게 물리적, 화학 적, 지학적, 생물학적인 면을 지적할 수 있다. 물리 적 요인 으로는 수심 ( wate r dep th) , 수온 ( wate r tem p er atu r e) , 정 수 압 (h y dros t a ti c pre ssure), 밀도, 광량 (I ig h t int e n sit y), 물의 운동 및 wa- ter tur bid ity; current sys te m 등을 들 수 있으며 화학적 요인으로는 영분
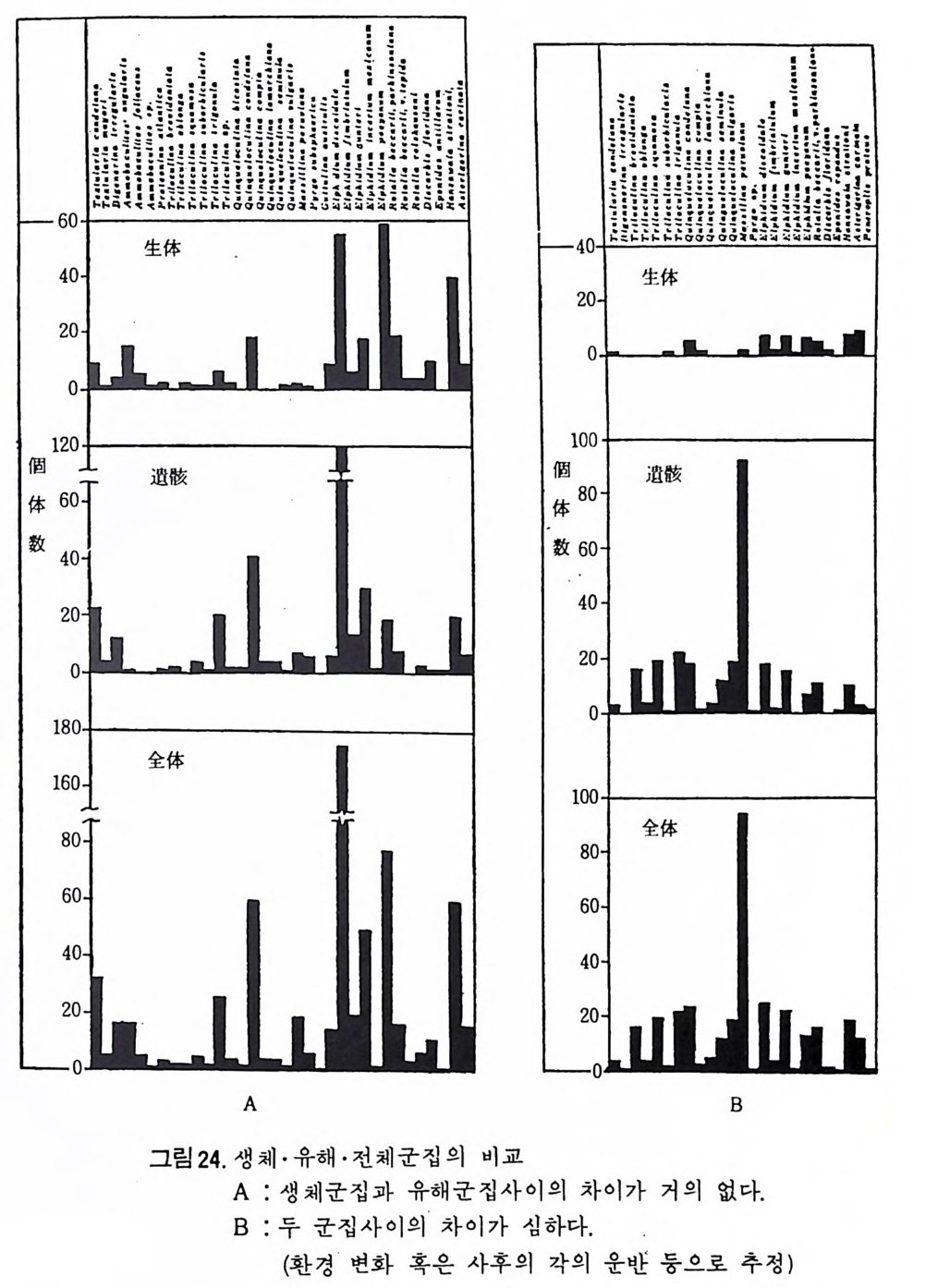 6。 -、-·.p..』••• .• •.- •--·•』•• .`- -• • -· i• .- ..I .. g.-. . • . . . ·『.. •..m. . -•l• v•• . •.• •. --• <•m•••v.•. • · . 生. • I•- 슬.v.•. ••n .• nn·l•••• 4.g体 •. .2• ..••n ·- . •••壘•. n.n. •• -. 4 •• n• 』n n .••••• • • • . ,. ,••.,,.. • . ..ql.m..,소 k..,·• ·. ...,,•, . .. pn••.p .••-..• nIp••4•.•. m•. n.•n•-·. •• -•0 •• •2 ..、.ni.lP• ••.k• • ·. .30 •. n,- .•·•• • 0. •• n•: •-•i.••n••- .• . ·•'• • .••3 0n pp•,•••••.•p .•••• o •. • • np- •-l•••.•• • · :• • 0 ••~.•••n••n••• • •••• •`.<•4•••• ••g. • • .-n• n.9.• •.u • . •n.、7.,mE..••.p •. •: i·:2-•·빽'v1m·.E •4 • - 그m - E역•••• ••--.,h.2 . 급,..mn·`, • . .. .. .,.n.. ..n급i, .mg.n.•:••• •.•.` • .• `.n슬,,..••.n-•N .· .• ..n· •• n 빼rnq ..•. 2 •.n:••-··v.w•-Mmn.•,••• 너•• • • ` • •-nIH•... •. .•,.. • .....l---. . . . . . .·.. ` . . l 4。 I ·-『•l.-•• k. ·• •..-8••-• •p.m• • •. • •2m-•...•n<•. •n』. • • -. •• · nn.••.n •』 .• •• •. •• •n ,n ••• •• ••• I.t.n.·-. . ,•• ••n. • k . k• •••n .-•••n •.n』• `•, ••• . l, .pI...• .·-• • . .or .••.n•.p •••I • -.•.• •n•.o .•.. ,••••, ,,. •,..•.. ,• • .h . ,. p .n•q ••- P • •· • • 0.p. •pn•,•••.2 .••• ••o p .•• •n、..••• .• 0 . .• . `..... . .•.、n,.•.••.•김 p. .•.m,. -mmi,n•. •`촐 •. ••n• •y•, • ..p •• •- , •• •.••y• • •,• ••,• ’m.•••• •‘ m-n••••••• n‘•:•- •• •.•• • nn••• n_•a •• • •••• •• mF r닉 • ••• •••. • - •n• •3•H • • •••• •l·훌 ., v.?.•• •• »••. •••.•i .••-•.•-.•t .,.`
6。 -、-·.p..』••• .• •.- •--·•』•• .`- -• • -· i• .- ..I .. g.-. . • . . . ·『.. •..m. . -•l• v•• . •.• •. --• <•m•••v.•. • · . 生. • I•- 슬.v.•. ••n .• nn·l•••• 4.g体 •. .2• ..••n ·- . •••壘•. n.n. •• -. 4 •• n• 』n n .••••• • • • . ,. ,••.,,.. • . ..ql.m..,소 k..,·• ·. ...,,•, . .. pn••.p .••-..• nIp••4•.•. m•. n.•n•-·. •• -•0 •• •2 ..、.ni.lP• ••.k• • ·. .30 •. n,- .•·•• • 0. •• n•: •-•i.••n••- .• . ·•'• • .••3 0n pp•,•••••.•p .•••• o •. • • np- •-l•••.•• • · :• • 0 ••~.•••n••n••• • •••• •`.<•4•••• ••g. • • .-n• n.9.• •.u • . •n.、7.,mE..••.p •. •: i·:2-•·빽'v1m·.E •4 • - 그m - E역•••• ••--.,h.2 . 급,..mn·`, • . .. .. .,.n.. ..n급i, .mg.n.•:••• •.•.` • .• `.n슬,,..••.n-•N .· .• ..n· •• n 빼rnq ..•. 2 •.n:••-··v.w•-Mmn.•,••• 너•• • • ` • •-nIH•... •. .•,.. • .....l---. . . . . . .·.. ` . . l 4。 I ·-『•l.-•• k. ·• •..-8••-• •p.m• • •. • •2m-•...•n<•. •n』. • • -. •• · nn.••.n •』 .• •• •. •• •n ,n ••• •• ••• I.t.n.·-. . ,•• ••n. • k . k• •••n .-•••n •.n』• `•, ••• . l, .pI...• .·-• • . .or .••.n•.p •••I • -.•.• •n•.o .•.. ,••••, ,,. •,..•.. ,• • .h . ,. p .n•q ••- P • •· • • 0.p. •pn•,•••.2 .••• ••o p .•• •n、..••• .• 0 . .• . `..... . .•.、n,.•.••.•김 p. .•.m,. -mmi,n•. •`촐 •. ••n• •y•, • ..p •• •- , •• •.••y• • •,• ••,• ’m.•••• •‘ m-n••••••• n‘•:•- •• •.•• • nn••• n_•a •• • •••• •• mF r닉 • ••• •••. • - •n• •3•H • • •••• •l·훌 ., v.?.•• •• »••. •••.•i .••-•.•-.•t .,.`
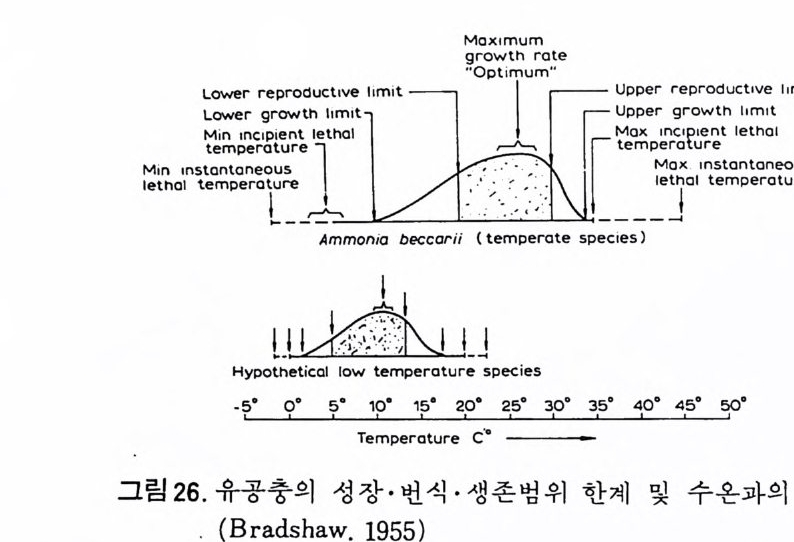
도 (sa l in i ty ), pH , 산소, 질소, 탄산가 스 , 산 화 • 환원電位 有 機탄 소 , 영 양 영류 등을,지학적 요인으로는 해안지형, 해저지 형 , 퇴 적물 의 입도, 해안으 로부터의 거리 등을, 생물학적 요인으로는 공생생 물 , 천 적, 먹이로 이용 되는 식뭉플랑크톤의 양 등을 들 수 있다. A) 심 도 0Nate r dep th) 심도의 변화는 저서성 유공중의 분포에 현저한 영향을 주고 있다 . Sts c - hedr i na (1 952) 는, 북서태평양을 심도에 따라 50m 이상, 50~200m,200~750 m,750~2000m 동 네개로 구분하였으며 그림 25 에서 나타난 것처럼 산 층 유공 충군 역시 다름을 볼 수 있다. Phleg er(1964) 는 멕시코 만 북부 해역을 9 군집대로 구분하였으며 , 각 군 집대의 경계는 구성종의 심도분포의 한계에 의해 결정되는데, 환경조건이 서로 다른 해역에 따라 군집대가 변화하는 것은 당연한 일이다. 이처럼 유 공충의 분포가 심도의 변화에 의해 좌우되고 있으나 절대적인 기준은 될 수 없으며, 여러 다른 환경요인(심도에 따른 빛의 두과정도 • 해류와 밀의 水鬼와의 경계 • 산소용존 • 먹이량 동등)과 복합되어 영향을 주고 있다. B) 온도 (Temp e ratu re) 온도변화는 특히 처서성 유공충의 분포에 커다란 영향을 미찬다. 온도의 변화는 영분도나 산소용존량 등의 이차적인 변화를 야기시키므로 유공충 의 분포와 온도만의 절대적 관계륭 생각할 수는 없으며 항상 다른 요인과 복합시켜 생각해야 한다. 우선 일반적인 경향을 살펴보연, 수온의 연변화는 표층수에서 심해로 갈 수록 적어지며, 중위도 지방에서 가장 크고, 200m 보다 양은 곳에서 가장 큰 계절변이룰 보이고 있다. 유공충은 1°C~50°C 사이의 온도에서 견딜 수는 있으나 성장이 가능한 범위는 l0°C~35°C 이며. 번식은 보통 l8°C~30°C 범위 내에서 이루어지고 있다. 성장의 한계 온도 내에서는 온도가 높을수목 성장률이 높다(그림 26 참조). 수온은 유공충 穀의 크기에 영향을 주는데 좋은 예로 교착질각을 갖는 유공충의 경우 수온이 낮을수록 각이 커지고 있음을 볼 수 있다. Hofk e r (1954) 에 의하면, 殺의 크기는 뭉론 각의 비율도 관계되고 있으며, 방의 높이 는 온도가 높아침에 따라 증가됨을 지적하였다. C) 빛 (Light) 빛이 직접적으로 유공충의 분포를 좌우하지는 않으나 간접적으로 영향
 A B c
A B c
 麟 n:三n i re \\Mg\roo xw,mt heu mr o t e r\::t\\ \:t
麟 n:三n i re \\Mg\roo xw,mt heu mr o t e r\::t\\ \:t
울 주고 있다. 유공충은 해조류나 식물성 플랑크톤을 먹이로 취하므로, 먹 이량은유공충의 개체수 밀도를 결정하는 중요한 요인이다. 빛이 잘 두과 되는 심도 80m 정도의 투광대 (ph ot ic zone) 에 풍부하다. 그 외에도 해조는 저서성 유공충의 생활장소로 제공되기도 하므로, 빛은 간접적으로 유공충 의 분포에 영향을준다. D) 물의 운동 파랑 • 조류 • 해류 등을 포함하는 물의 운동은 심도와 깊은 관련을 갖는 요인으로 유공충에 대해 수력학적 작용을 하며, 간접적으로는 퇴적물이나 먹이량 등을 동해 영향을 미찬다. 좋은 예로서 용승류 지역은 식물성 플 랑크론의 증식을 촉진시키며, 파랑 및 조류는 천해역에, 해류는 심해역에 영향을 크게 미친다. E) 영 도 (Salin i t y) 유공충의 대부분은 35%(}~45% r'의 영분도에서 서식하고 있다. Di sc orb- i no p s i s 屬온 57%( }의 높은 영도에서도 생존하며, 반대로 강이나 河口 (estu a- r y)처럼 15%,1~0. 5%, }의 낮은 영도에서도 극히 일부가 생존하고 있다. 영 도가 낮아짐에 따라 종의 다양도가 낫아지는 경향음 보인다 .
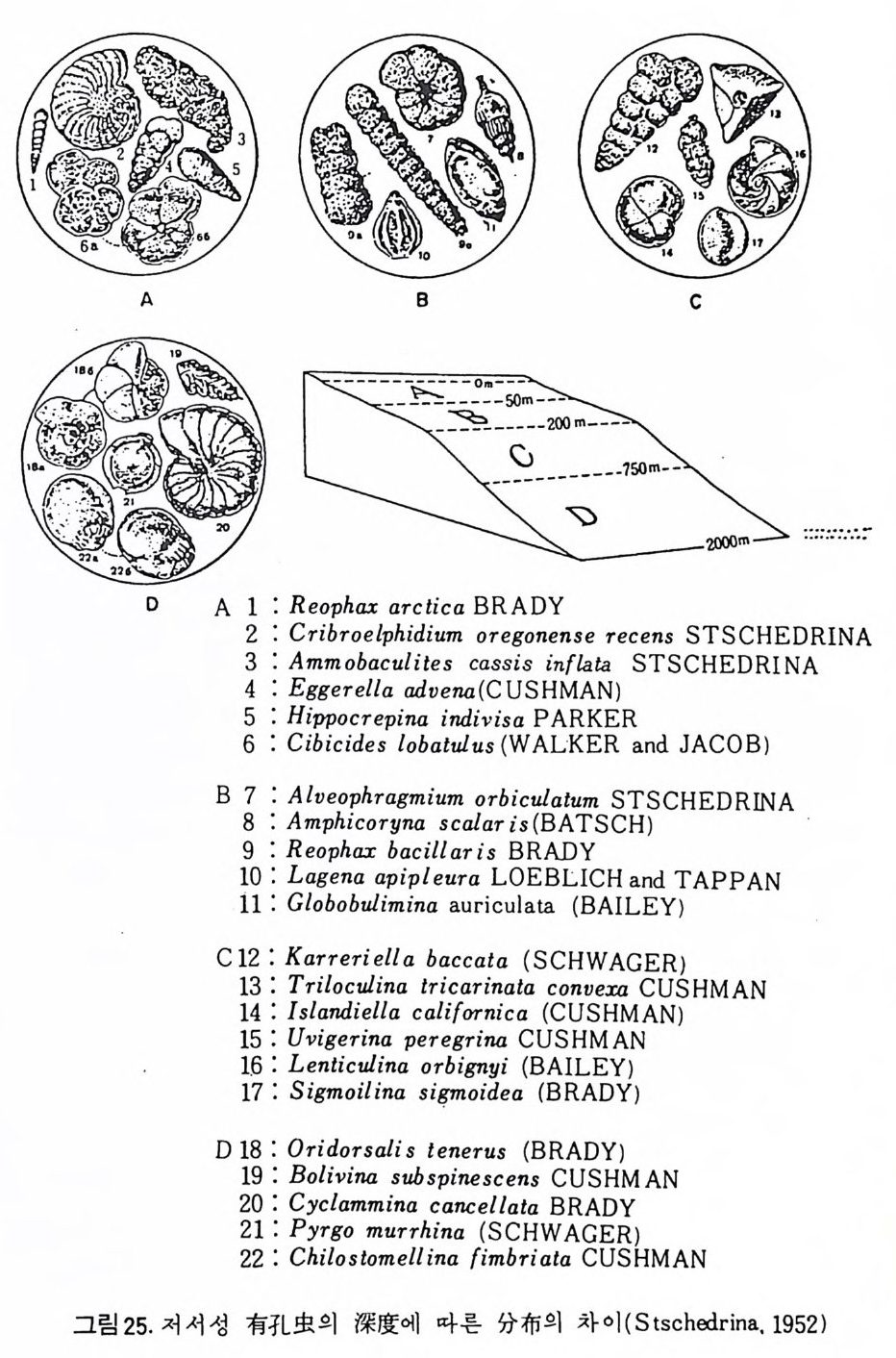
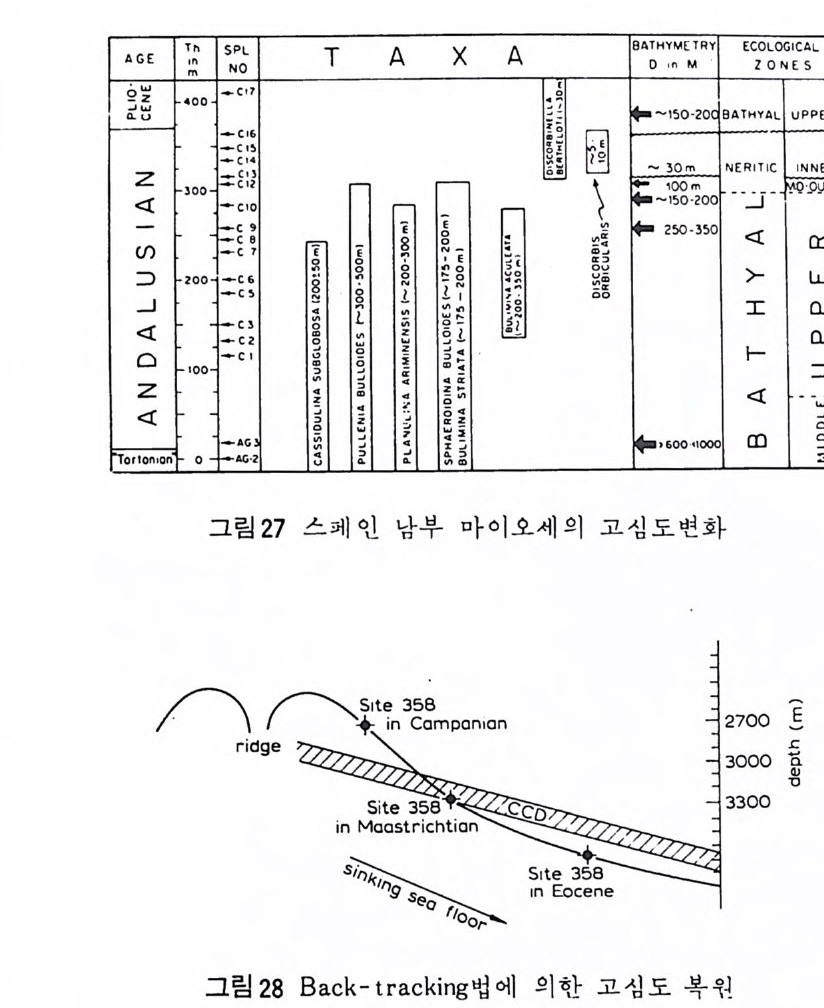
Bradshaw 는 영 도의 변화에 따른 Ammon i a 성 장실험을 동해 7~67 % (, 범 위 에서 성장이 가능하며 13~14% 。 범위에서 번식이 가능하나 20%0 이하에 서는 정상적인 번식이 어렵다는 사실을 지적하였으며, 매우 낮은 영도에 서 는 유공충 각의 크기 가 중어 들며 , 탄산칼슘 (calciu m carbonate ) 분비 작용 이 약화되어 두께도 얇아진다는 사실을 밝혔다. 기수역 같은 변동이 심한 환경에는 인내성이 강한 종만이 생존가능하며 결과적으로 기수역의 유공 충은 분포 범위가 넓다. F) 산소 (Oxy ge n) 용존 산소량은 심도에 따라 변화하고 있으며, 표층수의 경우 광합성식 물에 의해서 산소가 방출되므로 가장 높다. 심층수보다는 중층수의 용존 산소량이 더 낮은데 이는 생물체 산화에 의해 산소가 소비되기 때문이다. 수온이 상승함에 따라 유공충의 대사작용이 활발해지므로 산소 소비량이 많아지나 성장률 및 번식률과 산소량과의 정확한 관계는 밝혀지지 않았다. Lu t ze(1962) 와 Harman(1964) 에 의해 용존 산소량과 유공총 각의 모양, 표면 장식간에 상관관계가 있음이 밝혀졌다. 1 • 4 • 2 고생태 유공충을 이용한 고생태 환경 복원은 해양미고생물학에서 가장 광범위 하고 오랜 전동을 갖는 연구분야이다. 최근의 경향은 주로 고심도복원,고 온도 口갖 기후 복원. 해수의 화학성분 복원 , 고지리의 복원 등 매우 다 양하다. 특히 판구조 및 해양저 확장 이론의 발전에 크게 조력했고 여전 히 새로운 지 체구조론을 보완하기 위하여 유공충이 활용되고 있다. DSDP 사업이 대표적인 경우가 될 것이다. 동일과칭의 법칙에 의거하여 현생종의 생태자료를 과거종의 분포에 적 용함은 최근 여러 학자들에 의해 다소 수정 이용되고 있다. 고환경의 변화 에 따른 생물군의 서식지의 십·도의 적응은수직방향의 이동을고려해야하 기 때문이다. 그림 27 은 Ber gg ren 과 H aq (1976) 이 남부 스페인의 마아오세 (Andalusia n ) 지층의 고심도 변화를 유공충군으로 추정한 것이다. 많은 종 들이 아 당시 새로이 출현하였으므로 이돌의 고생태는 현생 후손들의 생 태와 유사하리라고 추측된다. 심도변화는 주변 육괴의 구조운동을 반영하 며 또한 저탁류의 활동 등을 암시한다. 또다른 고십도 변화의 연구는 back-tr ac ki ng 법으로 불리우는 Mckenz i e 와 Scla t er(1971) 의 실에가 있다. 그림 28 은 DSDP Leg 39 의 세개의 지점으로
서 남대서양의 알젠탄분지에 위치하는 5,OOOm 깊은 해처이다. 이 들은 대 서 양 중앙해 령 (Mi d- At la nti c rid ge ) 에 서 생 성 된 후 냉 각과 함께 침 강하떤 서 오른쪽으로 판 이동을 하는 중 이므로 말기 백악계~초기 제삼기에 걸친 고 십도의 변화 물 시료내의 유공충 군집을 분석함으로써 추적할 수 있다. 또 한 유공충의 유우로서 고 CCD 심도도 대체로 정해진다.
 A-o• G Z 브E 嶋훌 T A x A 8ATDH Y,nM (M TR Y (CZO LOO SGI(C ASL
A-o• G Z 브E 嶋훌 T A x A 8ATDH Y,nM (M TR Y (CZO LOO SGI(C ASL
고온도의 변화는 유공충각 내의 두 가지 안정 동위원소인 0 과 0 의 함 량비로서 추적된다. 오백만년 이전 유공충의 180/1 6()비는 해수의 동위원소 함량을 그대로 반영하고 따라서 이와 직접 관련되는 수온을 지시한다
(Emi lian i & Edwards : 1953 ; Savin & oth ers : 1975). CLIMAP 계획의 일 환으로 플라이스토世의 수온 변화를 연구한 Imbr i e 와 Kip p 등은 요인분석 법 (Facto r Analy tic Meth od) 이 라는 동계 적 인 방법 을 사용하였 다. 심해저에서의 탄산염질 퇴적물의 생성은 이러한 물질의 집적과 용액상 태로의 용해―두 가지 작용의 함수관계에 있다. Ber g er(l971) 는 CCD 보다 훨씬 얕은 곳에 유공충 탄산염이 현저하게 용 해되는 I y socl i ne 이 있음을 인식하고 백악기에는 현재의 CCD 평균깊이인 4, 500m 보다 훨씬 얕은 3, 200m 로 떨어졌다는 사실을 발견하였다 (Ber g er & Wi nt e rer: 1974) . 유공충은 지역적으로 지각운동사를 밝히는 데도 널리 이용되고 있다. 카 리브해분의 경우 플라이오세의 부유성 유공층인 Gwborota l ia mi oc enic a 및 Globorota l ia mul ti camera t a 를 이용하여 350~450 만년 전에 카리브해와 태평 양의 연락이 단절되었다는 사실을 밝혀냈다(그림 29).
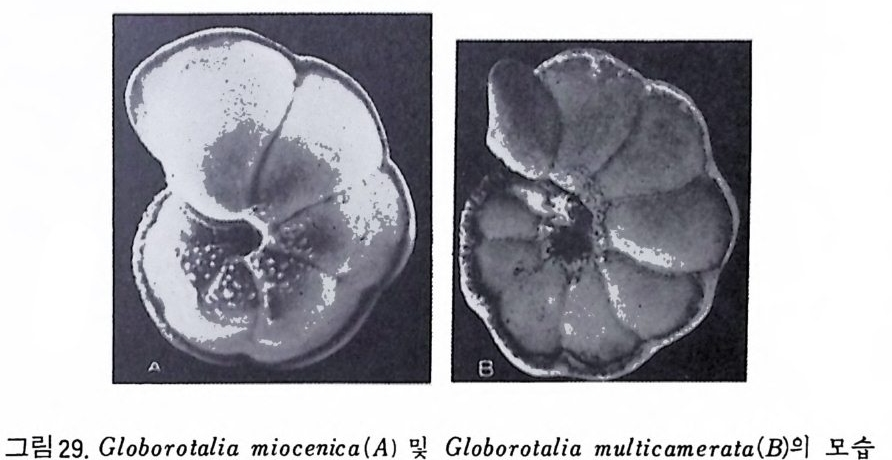 그림 29. Globorota l ia mi oc enic a (A) 및 Globorota l ia mul ti camera t a( B,)의 모습
그림 29. Globorota l ia mi oc enic a (A) 및 Globorota l ia mul ti camera t a( B,)의 모습
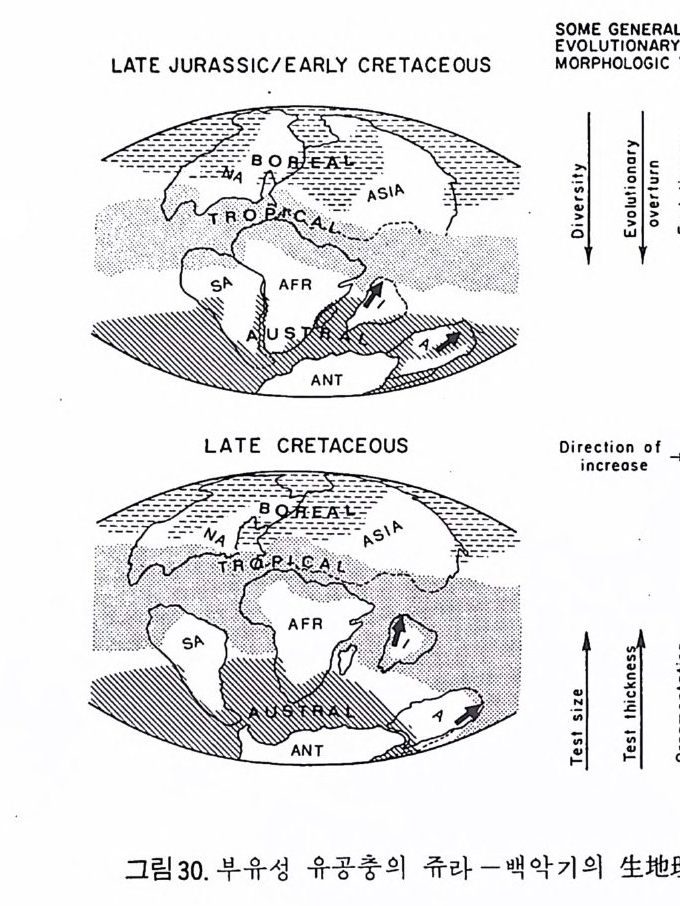
유공충은 대체로 위도에 나란한 분포를 하고 있으므로 이들 군집성분의 변화는 위도에 따른 수온의 변화, 습도 및 풍향계의 변화를 지시하며 나아 가서는 해수 및 대기 순환의 변화를 암시한다. 그림 30 은 쥬라기 및 백악 기의 주요 부유성 유공충의 분포로서 대륙이동에 따른 해류의 변화가 생 물지리에 어떤 영향을 미치는가를 보여주며 또한 종의 분포나 형태학적 변 화와 직접 관련지어짐을 예시한 것이다 (Gordon:1970;D ill e y: 1971).
 SOME GENERALIZED
SOME GENERALIZED
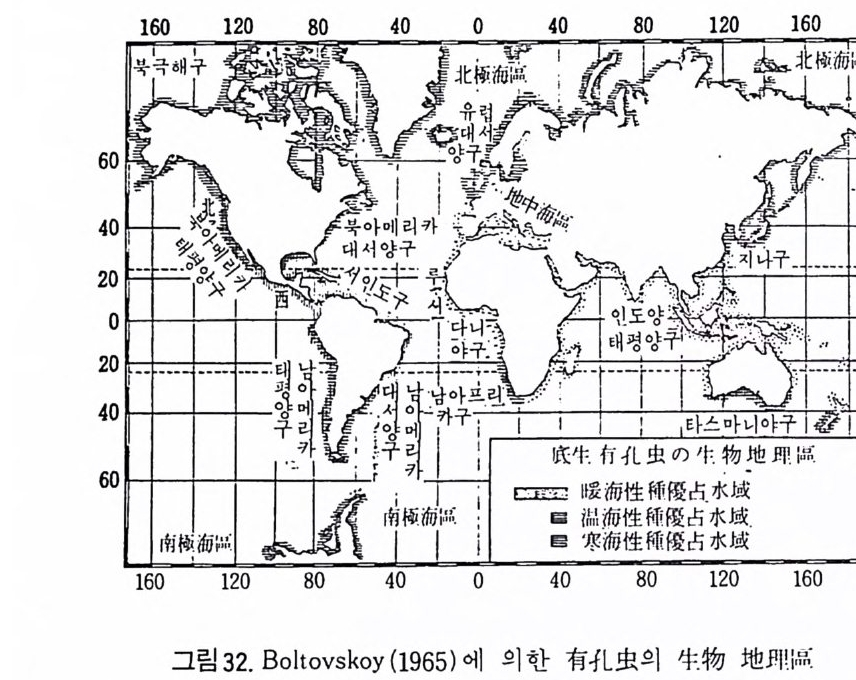
1 • 5 생지리. 생충서 및 진화 1 • 5 • 1 생물지리 현재 해양환경은 저서성 유공충의 지러적 분포에 의해 크게 난해성과 한 해성 · 둘로 구분할 수 있다. Cushman(1928) 은 북국해와 남국해 군집연구를 통해 각각 고유의 종을 갖 는 반면 공통종도 많음을 지적하였으며, 온대 • 아열대 • 열대의 천해역인 경우 유공충군의 분포구분이 명료하여 이 지역을 4 군(지중해 유공충군 ·서 인도 유공충군 • 동아프리카 유공충군 • 인도-태평양 유공충군)으로 구분 하였다 (그림 31 착조) . 저서성 유공충에 대한 조사가 계속됨에 따라 생물
00i06 0 100 1l6O I 80 `_ l 6010L_12 4 L)lOo . l。 20 0.I 20 4< ( t 重0 『 t 1 麟 > 讓 「4 。0。 60 ••• : 기 효' l ',1 가군.,.~f. ` a\ -,- I- - 가 -- t.\ 사 - -• -- -·4:;··f.; t-L .L._L_.-,. 60 } 니 : .. l l I I I I I I Il80 20 l 4麟0 60 &°,-, 100 120 21 4驛0 1휩60 180 160 140 120 3 1圖00 麗80 60 40 42 0圖 0 20 642 \鳳。 :’。 o o 40 20 。 20 40 60 60 20 그림 31 . 저서성 有 1 L 虫의 생물지리구
지 리구의 구분도 세분되는 경 향을 보인다. Bo lt ovsko y (1965) 는 그림 32 과 갇 아 적물지리구를 세분하였으며 난해성 종이 우세한 수역으로서 서인도구, 루시다니아구, 지중해구, 동아프리카구, 인도양-태평양구를, 한해성 종 이 우세한 수역으로 북국해구와 남국해구를, 온대성 종이 우세한 수역으 로서 북아메리카 태평양구, 북아메리카 대서양구, 남아메리카 태평양구, 남아메리카 대서양구, 남아프리카구, 유럽대서양구, 지나구, 타스마니야 구를 들고 있다.
 160 120 80 40 0 40 80 120 160
160 120 80 40 0 40 80 120 160
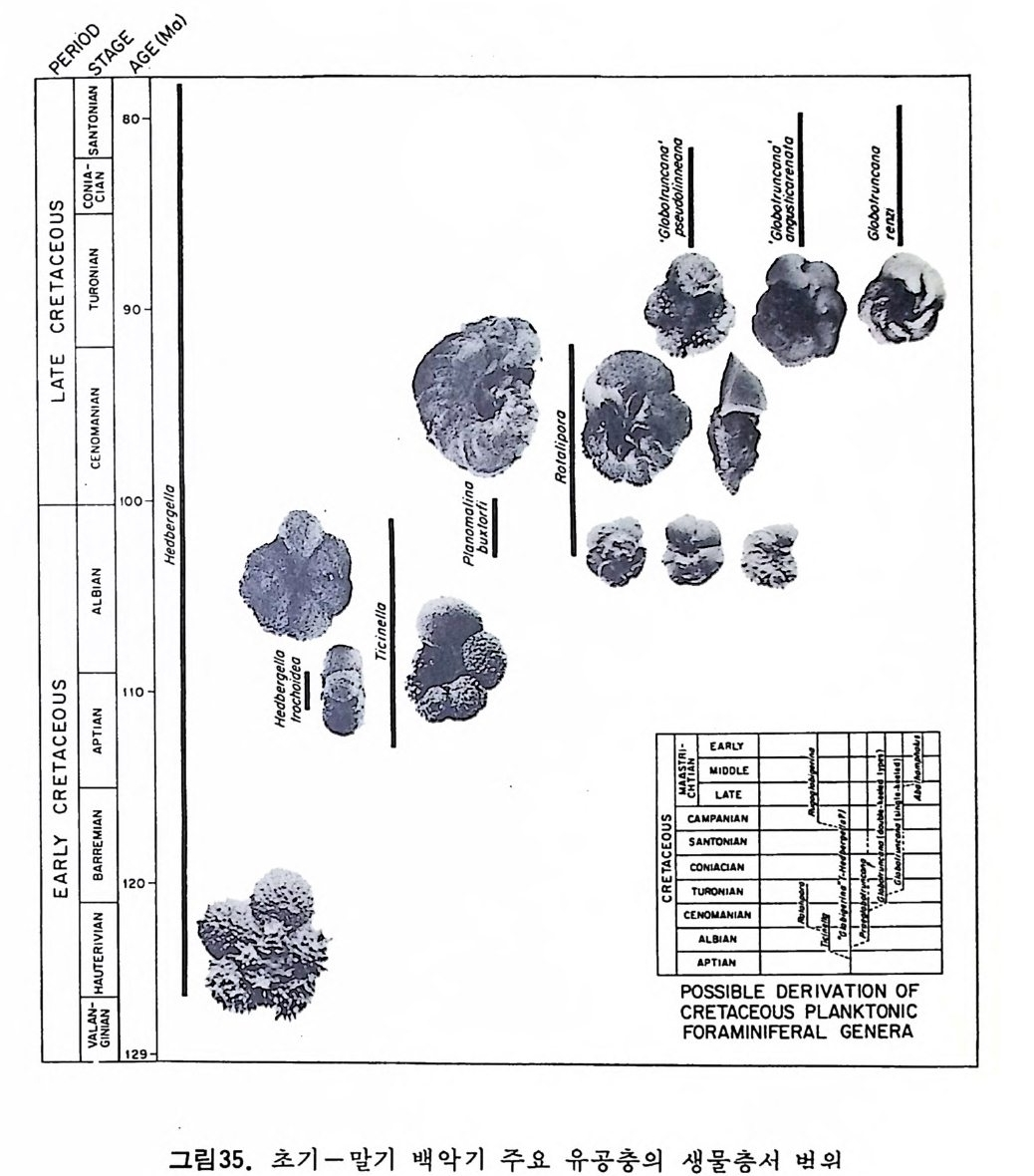
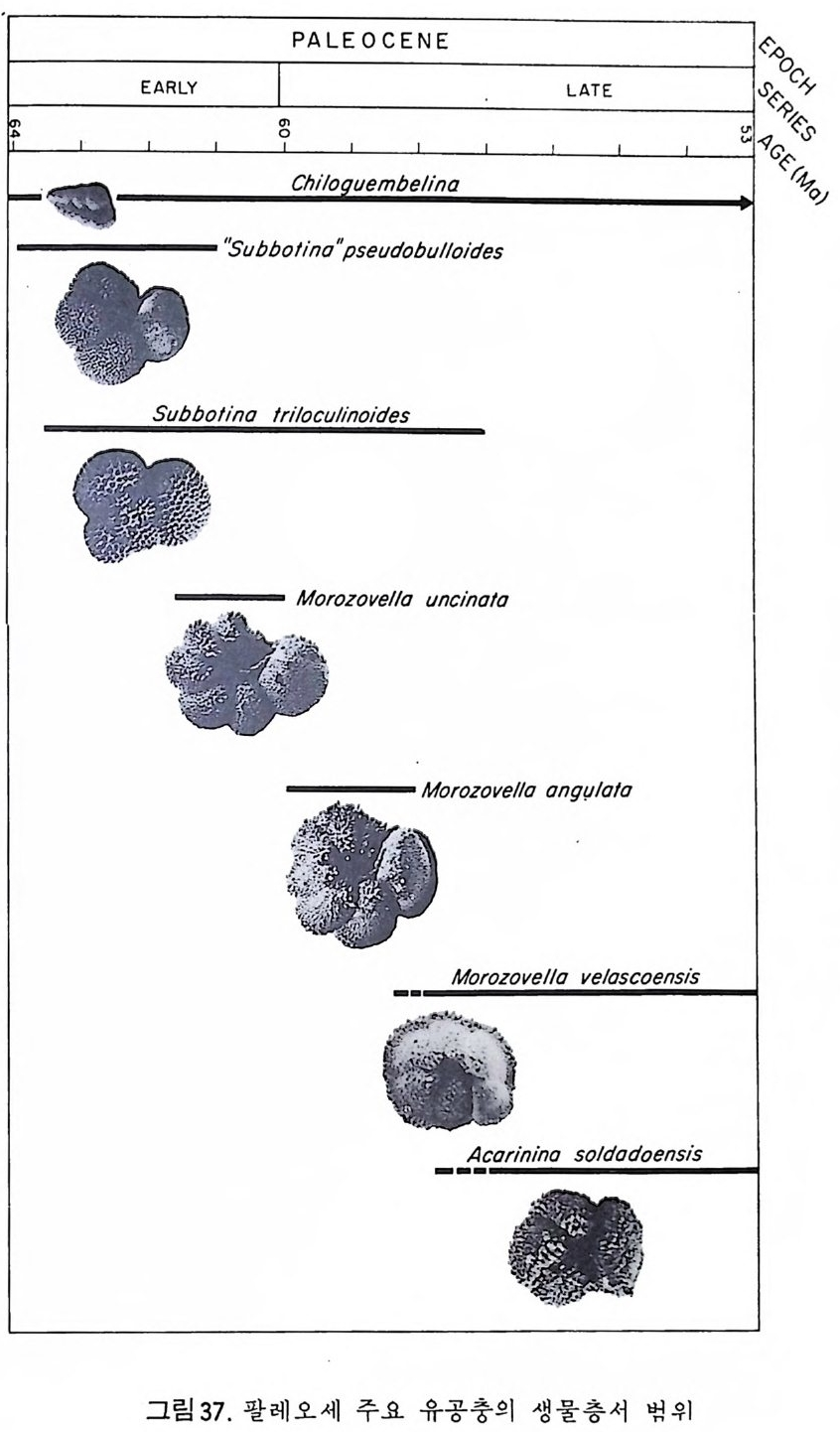
부유성 유공충의 경우, 종의 지리적 분포에 가장 큰 영향을 미치는 환 경요인은 수온이다. 해류에 의해 비대칭성을 띠긴 하나 대체로 적도를 중 심으로 남 • 복위 40° 사이는 난류성 군집이, 이보다 고위도지방은 한류성 군 집 이 분포하고 있다. 남 • 북 한류성 군집의 경우 많은 종이 공통되고 있 으며 수온에 따른 부유성 유공충의 분포를 보면 그림 33 과 같다. 1 • 5 • 2 생 물충서 및 진 화 유공충류는 거의 모든 지질시대의 분대에 이용되고 있으나 그 중 말기 고생대, 후기 백악기와 제삼기에 특히 유용하다.
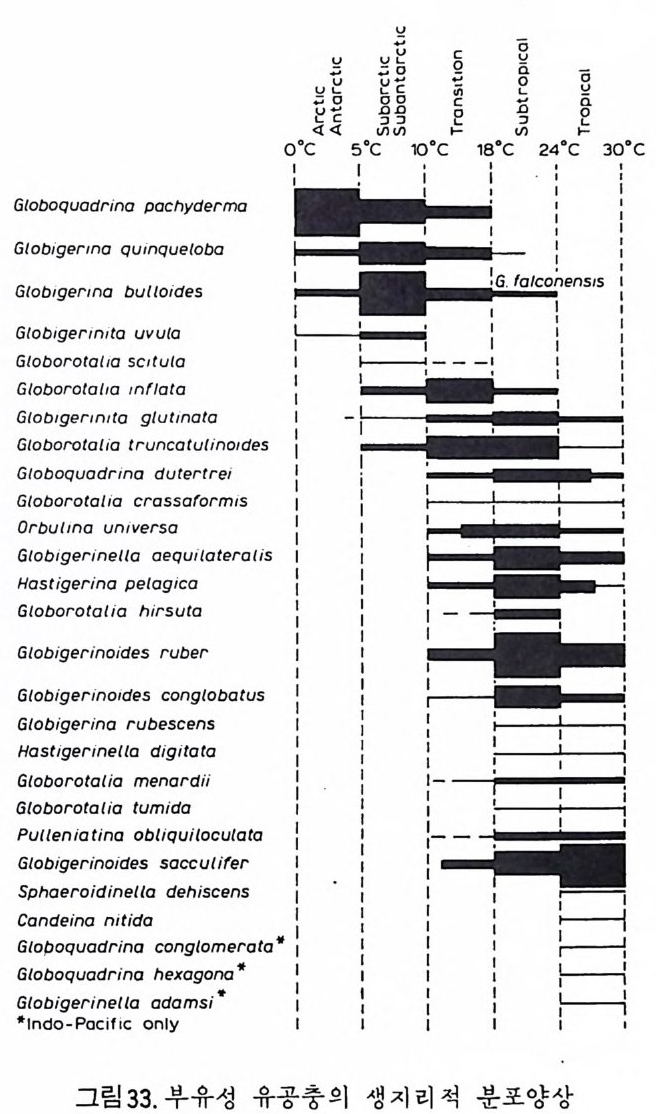 o:' c口 u5n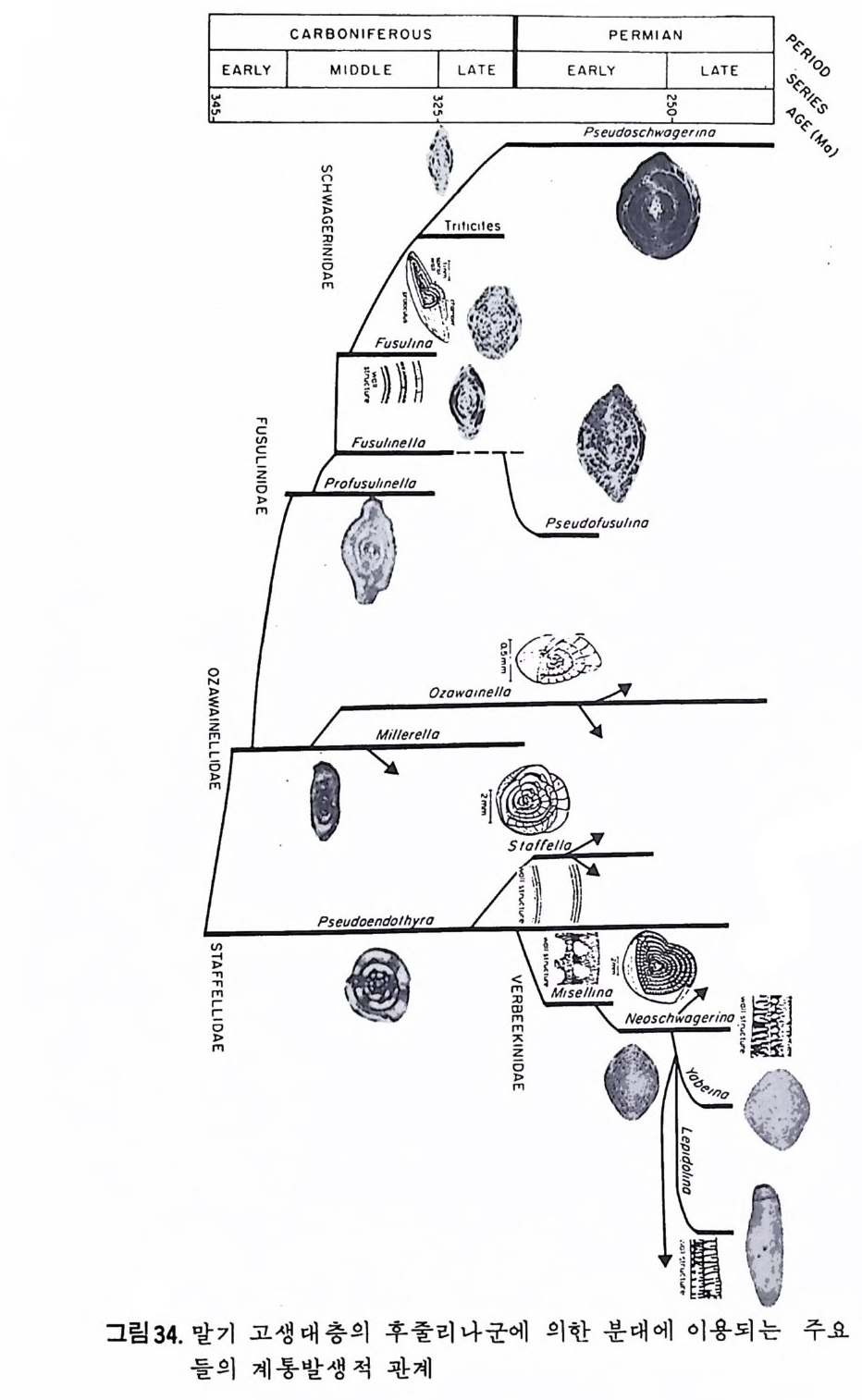 I|~ EARLY CARBIASNMGOCIOEIHNDARWEID F LEER O US 324L8‘· 一LA T E EARLYP E ◄IR fM I0,A~N1 LA。T E %.%?ce令 oq 3o es%
I|~ EARLY CARBIASNMGOCIOEIHNDARWEID F LEER O US 324L8‘· 一LA T E EARLYP E ◄IR fM I0,A~N1 LA。T E %.%?ce令 oq 3o es%
 00 t S3
00 t S3
 LATE CRETACEOUS
LATE CRETACEOUS
 PALEOCENE cA
PALEOCENE cA
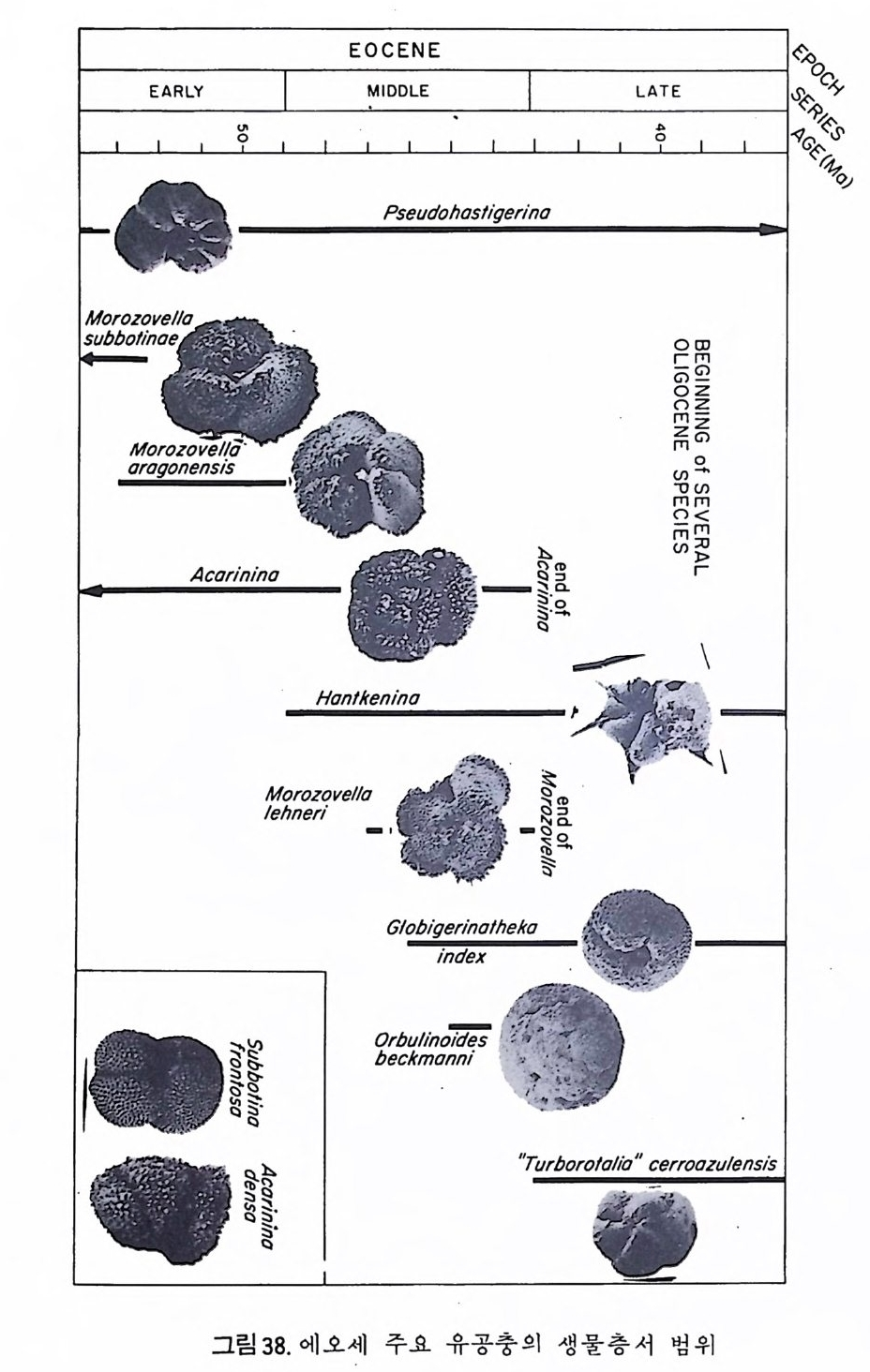 EOCENE 8°%%`
EOCENE 8°%%`
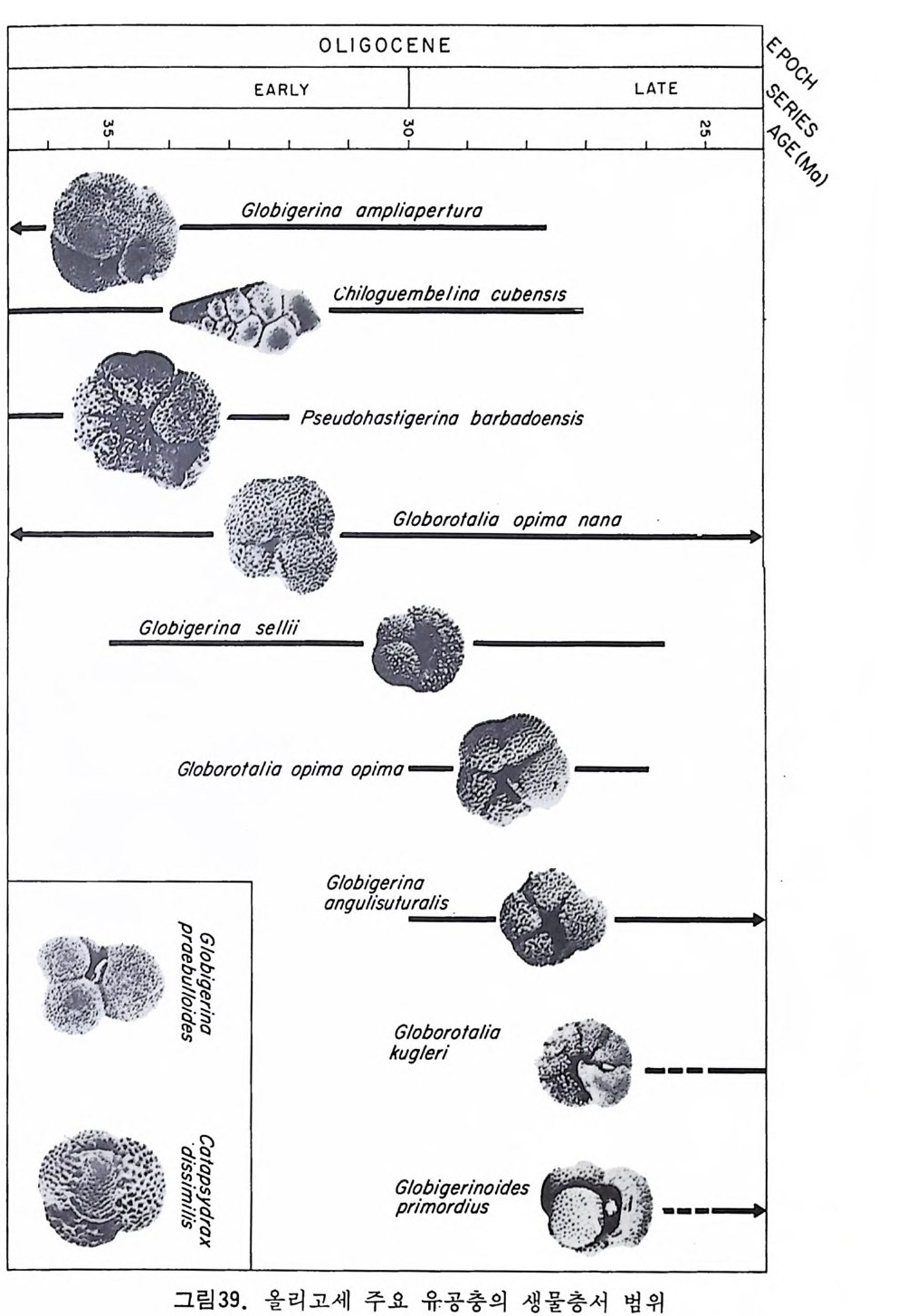 OLIGOCENE 8%v
OLIGOCENE 8%v
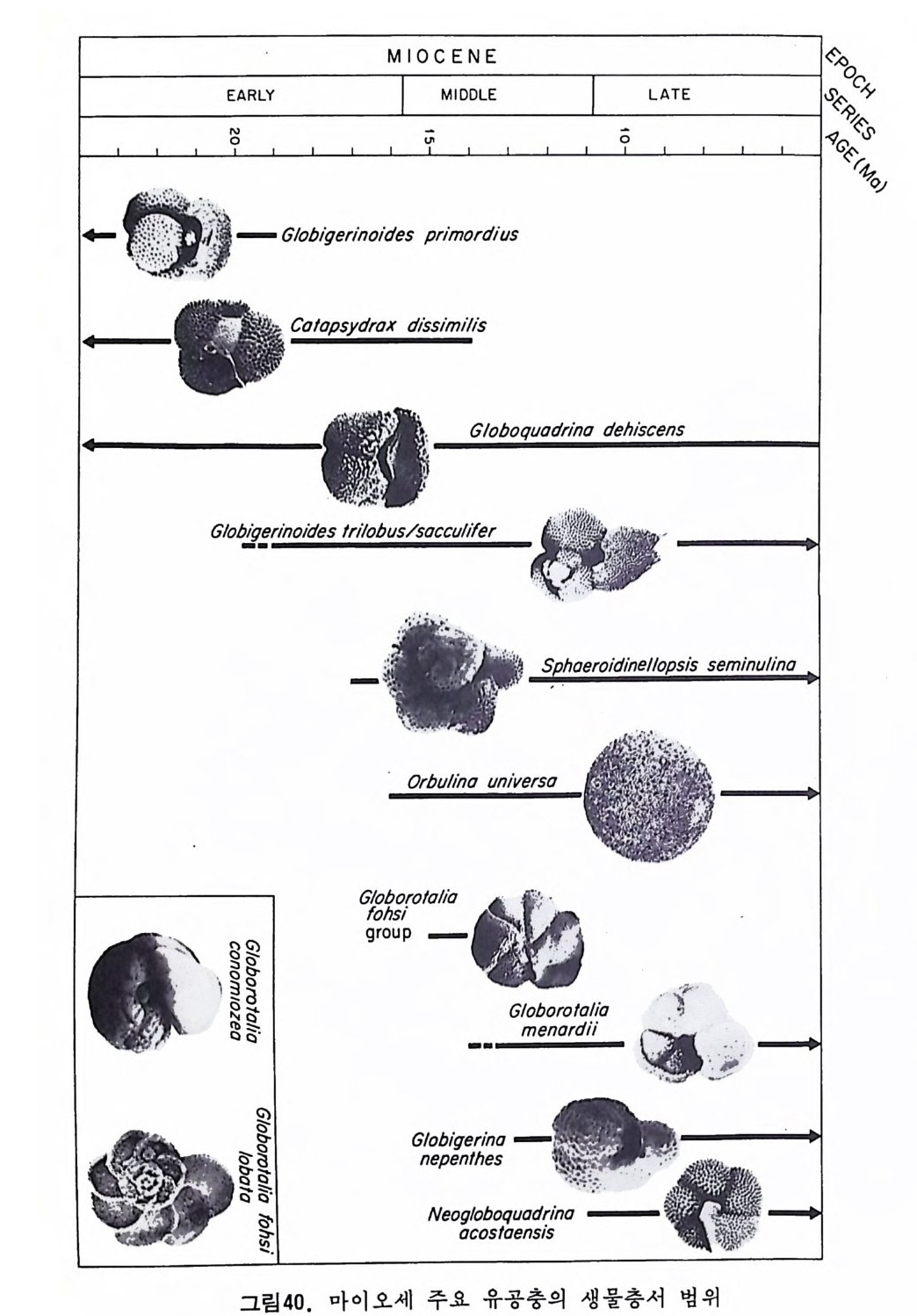 MIOCENE %%%C
MIOCENE %%%C
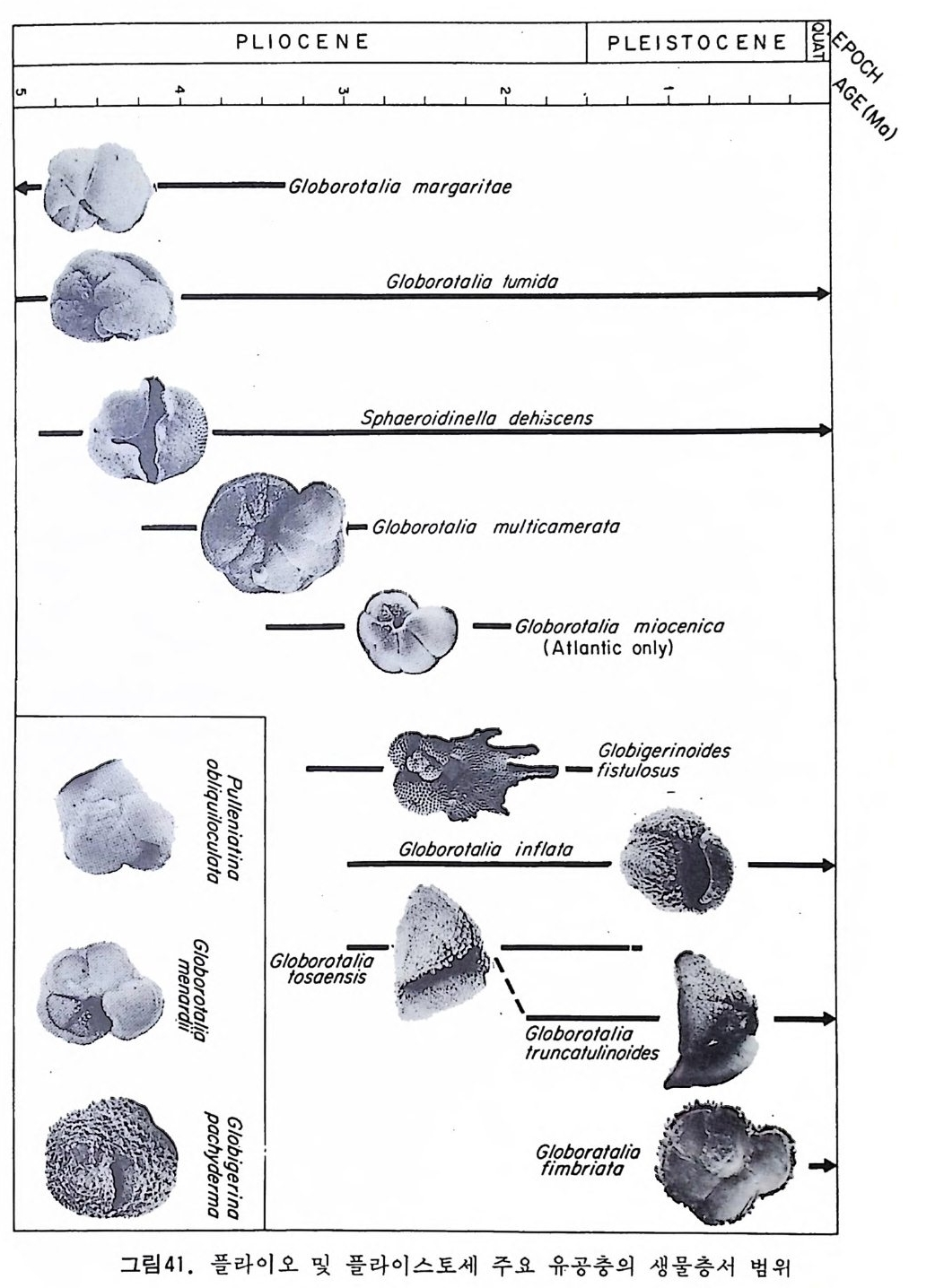 u` 1— 스 PLIOGCloEboN ”r Eol olio morqo rilo ”e PLEISTOC EN E UAF. '%vS. C인 %
u` 1— 스 PLIOGCloEboN ”r Eol olio morqo rilo ”e PLEISTOC EN E UAF. '%vS. C인 %
다 (Ber gg ren,1972). 이러한 분대는 고위도, 중위도 맞 저위도 지역별로 행 해지고 또한 절대 연대치에 의해 보강되고 있다. 최근에는 지자기연대와 도 대비가 성공적으로 시행되어 마이오세까지 거슬러 울라가는 연대표가 작성되었다. 그 밖에도 유공충 화석에서 추출한 아미노산의 변화물 이용 하여 연대를 측칭하는 기술이 개발되어 유공충의 생충서 분대를 보완해 주 고 있 다 (King and Hare, 1972) . 유尹충 진화에 국적인 방산의 시기는 데본기로서 이때 다방성 구조와 새 로운 형태의 석회질벽이 생성되었다. 페름기의 후중리나그룹의 번성기에는 약 5,000 종이 보고되었으며 지역 적인 지각운동과 생태의 차이로 다양화가 이루어졌다. 세계적인 해퇴의 시 기인 트라이아스기를 넘기고 쥬라기에 들어오자 광역적인 해침과 수반된 온난한 기후는 대륙주변해를 이상적인 유공충의 낙원으로 조성하였다. 다양 한 그룹이 각종 서식처에 적응 발전하다가 중생대 말의 대량 절멸의 시기 롤 맞게 된다. 중생대 말의 격변적인 생태계의 변화는 그 원인아 불분명하 나 200-300m 정도의 표층수에 영향을 미쳐서 대부분의 부유성 및 천해의 처서성 유공충에 결정적인 타격을 가했다. 신생대에 들어와 다시 부유성 유공충과 대륙붕 지역의 새로운 처서성 유공충이 재황을 보게 되는데 초 기에는 대형인 Nummul it es 가 특기할 만하고 그 밖에 대부분의 유공충은 신 생대 말의 한냉화 현상에도 불구하고 오늘날과 같은 분포양상을 갖추게 되 었다. 그림 35~41 은 각 지질시대별 주요 유공충그룹의 생충서 분포를 보 여준다. 참고문헌 〈교과서편〉 Cit a, M.B. (1964) : Mi cr op a leont ol og ia, 1-297, La Goria r dia , M ila no. Cushman, J. A. (1950) : Forami ni f er a, the ir . c lassif ica ti on and economi c use, l -605 Harvard Univ e rsit y Press, Cambrid g e , Massachusett s. Galloway, J. J. (19 33) : A manual of Forami ni f er a, 1-483, Prin c ia Press, Bio - omi ng t on , India n a. Glaessner, M. F. (1948) : Prin c ip le s of mi cr op a laeont ol og y, 1-196, Melbourne Univ c rsit y Press, Melbourne. Jon es, D. J. (19 56) : Int roducti on t o mi cr ofo s sils , 1-40 6, Harpe r & B rot hers, New York.
Loeblic h , A. R., Jr . a nd Tapp a n, Helen (19 64) : Treati se on inv e r te b rate Pa- leo nto l o g y, Part C, Proti st a 2 : Sarcodin a chie f l y Thecamoebia n s and Fo- rami ni f er id a , vols. 1 & 2, 1-900, Geol. Soc. Americ a & Univ . Kansas Pr- ess. Matt he s , H. W. (19 56) : Ein f i ihr ung in die Mi kr op a laont ol og ie, 1-348, S.Hi rz - el, Le ip z ig Ne u mann, Made le in e (19 67) : Manuel de mi cr op a leonto l og ie des Forami ni f er es {sy s te m ati qu e-str a ti gr ap h ie ) , 1-297, Gaut hi e r -Vi llar s, Paris . Pokorny , Vladim i r{l9 58) : G rundzUg e der Zoolog isc h en M ikr op a laonto l og ie, Band I, 1-58 2, VEB Deut sc her Verlag der Wi ss enschaft en , Berlin . Pokorny Vla d im i r ( 19 63) : Prin c ip le s of zoolo g ica l mi cr op a laeonto l og y, Vol. I, 1652, Pe rg a mon Pres s, Oxfo rd. Ra uz e r- Che rn o u so va, D. M. and Fursen k o, A. V. (1962) : Fundament a ls of pa - leo n to l og y . General pa rt , P roto z oa, 1-72 8, Israel Prog r am for Scie n t ific Tran s la t i on s, Je rus a le m . Sig a l, Ja cq e s (19 52) : Ordre des Forami ni f er a , In Piv e te a u , Je an : Trait e de pa le o n to l o g ie , tom e 1, 133-3 01, Masso n et Cie , Paris . 〈 논문편 〉 Adams, C.G., 1967. Te r ti ar y Forami ni f er a in the Te th y a n, Americ a n and In- do- Pa cif ic pr ovin c es. Sy s t. A ssoc., Pub!., 7 : 195-21 9. Arhold, Z. M. (19 55) : Li fe his t o r y and cy t ol og y of the for ami ni f er an Allog r o-
mi a lat i co llaris , Univ . Cali f. Pub!. Zoo/., 61, (4), 167 -2 52. As an o, Kiy o shi ( l9 50 -1 952) : Illust r ate d cat a log u e of Ja pa nes e Te r t i ar y sm aller Forami ni f er a, pa rt s 1 가 2 & Sup ple me n t no. 1, Hosokawa Prin t i ng Co., To- kyo . Asano , Kiy o shi ( l956-60 ) : The Forami ni f er a fro m t he aye ars 1873-1876, 1-23 8, Soc. Ec on . Pal. Mi n. , Sp e c. Publ. , no. 9. Be, A. W. H. , 1977. An ecolo g ica l, zoog e og r ap h ic and tax onomi c revie w of Re - cent pla nkto n ic for ami ni f er a . In : A. T. S. Rams ay (E dit o r ) . Oceanic Mi cr o- pa leont ol og y, 1. Academi c Press , London, pp. 1-88 . Berge r, W. and Gardner, J., 1975. On the de te rm i na ti on of Plei s t o c en e ter n- pe ratu res fro m pla nkt on ic fo rami ni f er a. J. Forami ni f era l Res., 5(2) : 102- 114. Berge r, W. and Rot h, P. H., 1975. Oce an ic mic r op a leo n t ol og y; pr og r es s and pr osp e ct. Rev. Geop h y s . Sp a ce Sci. , 13(3) : 561 -6 34. Bergg r en, W. A. (1969) : Rate s of evoluti on in some Ce n oz oic pla nkto n ic For- min i f er a, M icr op a leont ol og y, 15, (3), 351-365. Be rg g r en , W. A. and Phil lips , J. D., 1971 . Infl ue nce of con ti ne n t al drif t on the dis t r ib u t ion of Te r t i ar y be n th o nic for ami ni f er a in the Carib b ean an d M ed it - erranean reg i o n s. Sy m p . Geol. L iby a , Uni v. of Li by a , Trip o li , pp. 263 -2 99. Blow, W. H. (19 69) : Late mid d le Eocen e to Recent pla nkt on ic fo r am i ni f er a l bic s t ra ti gr ap h y, 1st Int e rn. Con(. Plankt on ic Mi cr ofo ssil s , Geneva, 1967, P- roc., 1, 199 -4 22. Bolli, H. M. (1950) : The dir e cti on o 「 co ili n g in the evolu t ion . o f som e Globor- ota l iid a e, Cushman Found. Foram.Res., Cont r. , 1, (3- 4), 82- 89. Bolli, H. M., (1957) :Plankto n ic for ami ni f er a fro m the Olig o ce n e - M i oc ene Cip c - ro and Leng u a for mati on s of Trin i d a d. In : A. R. Loe bl ic h and oth e r s. St u- die s in Forami ni f era . U. S. Natl . Mus. Bull., 215 : 97-125. Bolli, H. M. (1966) : Zonati on of Creta c eous to Plio c en e marin e sedim e n ts based on pla nkt on ic Forami ni f er a, A soc. Venezolana Geol. M i ne r. Pet rol., Bo! In- for m., 9, ( 1 ), 3 -3 2. Bolli, H. M. , Loeblic h , A. R., Jr . a nd Tapp a n, He le n (19 57) : Plankt on ic fo ram- ini f er al fam i lie s Hant ke n in i d a e, 0rbulin i d a e, Globorot al iid a e and Globot rirn can- ida e, U. S. Natl . Mus., Bull., 215, 1-50 . Blow, W. H. (19 69) : Late mid d le Eocene to Recent pla nkto n ic for ami ni f er al bic - str a tig r aph y, J&t Inte r n. Conf pl ankto n ic M icr ofo ssil s, Geneva, 1967, Proc., 1, 199-422. Bolto v skoy , Este b an (19 56) : Di cc io n ario for ami ni f er olo g ico plu rili ng iie, i-1 96, Arge nti na Serv. Hi dr . Publ. Mi sc . no. 1001 . Bolt o vskoy , Es te b a n( l96 5) : Los Forami ni f er os rec ie n te s ;B i ol og ia, me to d os de estu d io , • ap li c a ci6 11 oceanog r ii. fica , 1-510, Edit o ria l Univ e rsit a ria de Buen-os Ai re s, Buenos Ai re s . • Brads ha w, J. S. (19 59) : Ecolog y of livi n g pla nkto n ic Forami ni f er a in the north and eq u ato ria l Pacif ic Ocean, Cushman Found. Foram. Res., Contr . , 10, (2), 25-64. Bronnim ann, Paul and Brown, N. K., Jr . (1956) : Taxonomy of th e Globot r un-canid a e, Eclog a e ge ol. Helv., 48, (2), 503 -5 61 .
Di lle y, F. C., (1973) : Large r for ami ni f er a and seas thr oug h tim e. Sp e c. Pap. P a-
leont o l., 12 : 155 -1 68. Doug la s, R. G., (19 73) : Benth o nic for ami ni f er al bio s tr a ti gr aph y in the Cent ral North Pacif ic, Le g 17, Deep Sea Drill i n g Proje c t. Init ial Rep . Deep Sea Dr- ill . Proj. , 17 : 607-67 2. Eames, F. E., Banner, F. T., Blow, W. H. and Clarke, W. J. (19 62) : Funda- ment al s of M id- Terti ar y str a ti gr aph ic a l correlati on , 1-163, Univ e rsit y Pr- ess, Cambrid g e . Ellis , B. F. and Mes sin a , A. R. (1949 et seq .) : Cata l og ue of Forami ni f er a, vo- Is. 1-45 &: sup p le m ent vols., Americ a n Museum of Natu r al His t o ry, New York. Ellis , B. F. and Me ssin a , A. R. (1965-1967) : Cata l og ue of ind ex large r For- ami ni f er a , vols . 1-3, Ame r ic a n Museum of Natu r al His t o ry, New York. Ellis , B. F., M essin a , A. R., Charmatz , R. and Ronai, L. E. (1968-1969) : Ca- tal og u e of ind ex sm aller Forami ni f er a, vols. 1-3, Americ a n Museum of Na - tu ral Hi st o ry, Ne w York. Emi lia n i. Cesare (19 54) : Dep [h habit a ts of some sp e cie s of pe lag ic Forami ni - fer a as ind ic a te d by oxy ge n iso t o p e rati os , Am. Jo ur. Sci. , 252, (3), 149-158. Eric s on, D. B., Wollin Goesta and Wollin , Jan et (1954) : Coil in g dir e cti on of Globorot al i a tru ncatu l i no id e s in deep - sea cores, Deep -S ea Research, 2, (2), 152 -1 58. llin , Grel l, K. G. (1956) : Proto z o o log ie, 1-284, Sp r in g e r, Berlin . Ha ll am, A., (19 73) : At la s of P~laeobio g eo g ra ph y. Elsev ier . Amste r dam, 531 pp. He dle y , R. H., (19 64) :T he bio l og y of for ami ni f er a. Int. Rev. Ex p. Zool. , 1 : 1- 45. He dl ey , R. H. and Adams , C. G., (1974) :Forami ni ( e ra. Academi c Press, Lond- 011, 276 pp. Hi lt erm ann, He in r ic h (l961) : Bib l io g r aph ie st r ati gr aph is c h wic h ti ge r mik r op a l- aont olo g isc h er Publik a ti on en von etw a 1830 bis 1958, 1-40 3, E. Schweiz e r- bart' s c h e Ve rl ag ., St uttga rt. Hall, R. P. (19 53) : Proto z oolog y, 1-682, Prenti ce -Hall, Eng le wood Clif fs,N .J. Ha m i lt o n , E. L. (19 53) : Up pe r Creta c eous, Tertia r y, and Recent pla nkto n ic Forami ni f er a fro m Mi d- Pacif ic flat - top p e d seamount s, Jo ur. Pal., 27, (2), 204-237. Ho fk e r, Jan , S r. (1950) : Wonder ful anim als of the sea, Forami ni f er a, Ams- Ish t eiw rd aadma, NYaatsu u rfa ulm i si t(, 1 916,4 ) (3:) , Be6n0t h -o n7 9ic. Forami ni fe r a off the Pacif ic coast of Jap a n ref e rred to bio s t rati gr ap h y of the Kazusa Group , Geol. Survey Ja p- an, Rep. , (205) , 1-45 . Jen kin s , D. G., (19 71) : New Zealand Cenozoic pla nkto n ic for ami ni f er a . N. Z. Ge- ol. Surv. Paleont ol . Bull., 42 : 278 pp.Klein p L •II, R. M. (19 38) : Mi oc ene slrati gr ap h ,v of Calif or nia , 1-450, Anwr. As-soc. Pct rol. Geo!., Tulsa, Oklahoma. K 야 11 ig swald, G. H. R. and Wag n er, C. W ., t、
Reis s , Zeev{ l95 7) : The Bil a mellid e a, nov. sup e r fam ., and rem arks 011 Cn•t- aceous glo borota l i ids , Cushman Found. Foram. Res., Cont r., 8, (4). 127- 145. Reis s , Zeev (19 63) : Reclassif ica ti on of pe rf or ate Forami ni f er a, Israel Geo!. Survey, Bull., 35, 1-11 1 . Ross, C. A. (19 67) : Develop m ent of Fusulin i d ( Forami ni f er id a ) fau na! realms . Jo ur. Pa l.eo nt. , 41, (6), 1341-135' 4, 9 tex t -figs . Sait o, T. (1963) : Mi oc ene pla nkto n ic Forami ni f er a fro m Hons hu , Jap a n, Toh-oku Univ . , Sci. Rep ., 2nd ser. (Geo!.) , 35, (2), 123-209. S chu lt z e, M. S . (18 54) : Uber den O rga nis m us de r Polyt lr alami en (Fo rami ni f e- ren ) nebst Bemerkung e n iibe r die Rhiz o p o den in Allge m e in e n, 1-68, Wi llr e- Im Eng e lmann, Leip z ig _ Si m on, W., et al. (1962) : Leit fos s il ie n de r Mi kr op a laont ol og ie; ein Abris s Ire - raus g e g e b en von ein e m Arbe it sk r ei s deu t sc lr er Mi kr op a laonto l og e n, 2 vols., 1-43 2 • Gebruder Bornt raeg e r, Be rl in . Scott , G. H. 1958. Di st ri b u t ion of po p u lati on s of fos sil ior ami ni f er a. New Ze al and Jo ur. Ge o/. Geop h y s ic . , Vol, 1, no. 3, p. 474 -4 84. Takaya nag i, Yokic lr i, Nii t sum a, Nobuaki, and SakaL Toy o sab uro 0968) : Wall mic r osI ruc tu re of Globorot al ia tru ncatu l in o id e s (d' Orbig n ;•), Tohoku Univ ., Sci. Re p. , 2nd ser. (G e o!. ) , 40, (2), 141-170. Th al mann, ·H. E. 0960) : An ind e x to tire ge nera and sp e cie s of the Forami ni - fer a , 1890-1 950, 1-39 3, G. Vand e rb il t Foundat i on , S tan fc r
Towe, K.M., and Cif ell i , Ric h ard(1967) : Wall ult r as tr u ctu re in th e cal ca re- ous Forami ni f er .a : crys ta l log r aph ic as p e cts and a mode l fo r cal ci f i ca ti on , . Jo ur. Pal., 41, (3), 742 -7 62. Uchio , Takay a su (19 61 ) , Ecolog y of livi ng _ benth o nic Forami 11i fer a fro m the Sa n Die g o , Califo r ma area, Cushman Found. Foram. Res., Cont r ., Sp e c . Pub!. Uj5i i,e , 1H-7i r2o .s h i ( 1962) : Int rocluclio n to st a t i st i ca l fo ram i n if 타 al zon a t i o1 1 , Geo/. Soc. Jap a n, Jo ur., 68, (803) , 431-45 0. Von . Koenig s w ald, G. H. R., ~meis . J. 0., Bunin g , W. L. an d Wag n e r, C. W., (1963): Evoluti on ary Tre 뼈 s in For am i ni ( e ra. Else v ie r . Ams te rd a m , 355 pp. Wagi 1e r, C. W., 1964. Manual of Large r Forami ni ( e ra. Ba1aafs c h e Pe ·t r ol e - um Mi j., The Hag u e, 307. pp. Walt on , W. R. (1964) : Recent fo ram i ni f er a l ecolo g y and pa le o ecolo g : v, . In lmb-
rie , Jo hn, and Ne w ell, Norman, edi t o rs, Ap pr oa ch es to pa le o ecolo g y, Jo hn Wi le y and Sons , New York, 151-23 7. Wood , Alan(1948) : The slr uc tu re of 1he wa ll of lhL' res t in th e For am i ni f e- ra; its value in class H ica 1i o1 1, Ge o! , Soc . Lon do n, Jo ur., 5 4, (2) , 229- 2 55. 2 介形 虫 (Ost ra coda) 2 • 1 서언 개 형 충은 Linn aeus (1758) 에 의 해 Monoculus 라 불 리 던 절 족동물 (Eu arth r- o poda)문 중 갑각강 (Crusta cea ) 에 속한다. 개형충의 분류학적 연구는 사실상 0.F.Mull e r (1 776 ) 에 와서 해수에 사 는 개형충을 Cy th ere, 담수에 사는 개형충을 C yp r i s 라 이름한 데 서 부터 시 작되었다고 할 수 있다 . 그 이 후 , 19 세기에 들어서면서부터 황 발한 화석 개형충류의 연구로 현재 약 1, 200 屬 이상의 분류군을 갖 는 大亞 綱이 되 었 다. 개형충은 보통 0.15~2mm 정도의 현미경적 크기를 가지며 2 매의 背甲 에 의해 둘러싸여 있다. 해수에 사는 현생종 중 G ig an t oc yp r i s 속은 25nm 정 도의 크기 를 가지 며 특히 고생 대 의 Lepe rdi tiida 目 중 심 한 경 우는 80mm 이 상에 달하는 종류도있다. 캄브리아기의 해수로부터 시작되어, 석탄기에는 담수에까지 생식범위를넓혔다. 그 이후 끊임없는 진화과정을 거쳐 오늘날에 이르게 되었으며, 현 세에는 국지방에서 적도에 이르기까지의 해수 • 기수 • 담수 …습지 둥의 다 양한 환경에 적응 서식하고 있다. 해수의 경우 해안가로부터 심해저에 이 르기까지 , 심도로 보면 1,5 00 fat homs(2,800m) 의 깊이까지 분포한다. 개형충의 거의 대부분은 저서성이며, 아주 드물게 부유성 종과 그외 다 론 갑각류나 어류 • 양서류 등에 기생 혹은 공생하는 종도 있다. 특히 부 유성 개형충 갇은경우는 각이 아주 얇아 화석으로서의 보존이 어려워 고 생물학적으로는 거의 무시되고 있다. 이처럼 극히 드문 예를 제외하고는 배갑이 화석으로서 잘 보존되는 석희질 각인데다, 다양한 환경에 적응하 고 있으며 , 현미경적 크기로 풍부히 산출되고 있어 다른 갑각류에 비해 고 생물학적 • 층서학적 • 고생태학적 연구에 많이 이용되고 있다 . 또한 다른 갑각류와는 상당히 달리 오랜 자질시대를 거치는 동안 연속적 전화양상을 보이고 있어, 이들은 고생물지리 (p aleob i o g eo g ra ph y)나 지역간 층서대바 등 에 이용된다. 유기체 (so ft bod y)를 포함한 생물체 전체를 모르고서 그 일부인 배 갑을 이해한다는 것은 사실상 무리지만, 화석상으로 穀만이 보존되고 유기체 는 보존되지 않으므로 고생물학의 경우 어쩔 수 없이 생물학적 기준과는 달라질 수밖에 없으며, 주로 각에 의존하여 생물의 기능형태학(f unc ti onal morph o log y), 분류학(t axo_ r1 o my), 생태학 (ecolo gy), 계동 발생학(p hy lo g e~y ) 등을 연구하고 있는 실정이다. 개형충류의 연구 발달 상황을 간단히 살펴보면 다음과 같다. 0.F.M i.i ller(1776) 에 의해 현생종에 대한 체계적인 연구가 시작되었으며, Sars (1866) 에 이 르러 서 야 분류기 초를 세 워 크게 4 그룹 (Myo docop a, Po d- ocopa , Platy co pa , Cladocop a) 으로 구분되 었 다. 개형충의 유기체라든가 형태학적 적응성 (morp h olog ica l adapt a ti on ) • 생태 ·생식 • 계통발생학적인 연구는 G.W.Muller(1894) 에 이르러서 시작되었다 고 할 수 있다. Brady (1868) 와 그외 공동 연구진은 개형충 연구의 새로운 방향을 제시 해 주였으며 현생종과 화석종 사이의 비교연구에 의한 고생태학적 해석을 시도하였다. 그 외에도 Cannon (1 925) 의 기능형태학적 연구, Elo f son(1941) 의 현생종 에 대한 생태학 연구, Har t mann(1968) 의 생물학적 연구 동을 들 수 있다. 반연에 1813 년 화석 개형충에 대한 최초의 언급이 있었고, Jo nes(1849~
1901 ) 에 의 한 J L 생 대 가 1 형충 연구 . 그 외 엿엿 연구가 있었으 나 실질 적으 로는 19 2 0 년 이 후 에 석 유 문 세와 함께 황밥한 연구 성 향을 보 이 고 있 다 . Zalanyi (1925) , Alexander (1933) , Sy lv este r -Bradley (1941) , Trieb el (19 41) 예 의해 각의 형태학 연구의 발전을 보았으며 1950~1960 년대에 걸쳐 분 류학 ( t axonomy ) 이 중요시 되 었 다. 그후 이 분류학을 기초로 많은 응용 고생물학적 연구가 진행되고 있으 며 Puri ( 1965) , Neale (1969) , Oertli (1971) , Swain (1975) 그외 많은 학자들 에 의해 활발한 연구 경향을 보이고 있다. 2 • 2 생활사 절족동물 r' J의 갑각綱에 속하는 개형충은 한 쌍의 각에 둘러싸였으며 외 관상으로 二枚貝조개와 흡사하여 Mussel shrim ps 혹은 Muschelkrebse 라고도 붕린다. 이 각에 둘러싸여 보호되고 있는 유기체는 패각 전 길이의 2/3 가량을 차 지하고 있으며 두부 (ce p halon) 와 흉부(t horax) 둘로 구분된다. 두부에 는 an t ennu i es( 혹은 제 1 ante n nae) , . ante n nae (혹은 제 2 ante n nae) , mand ibl es, maxill ae 등이 쌍을 이루며 붙어 있고 흉부에는 제 1 • 제 2 • 제 3 의 tho racic le g s 가 부착되어 있다(그림 1 참조). 두부는 크게 for ehead , uwer lip, h ypo s t ome 으로 구분해 볼 수 있다. an t ennules 와 an t ennae 가 부착되어 있는 fo rehead 는 두부의 위쪽·앞부분을 차지하고 있으며 눈을 보호하는 역할을 한다. An t ennules 는 수영 과 감각기 관으로서 사용되 며 ante n nae 는 이 동에 사용된 다. Forehead 바로 밀이 u pp er . l ip으로, 죽 복부의 전반부위가 된다. 밑에 입 이 있으며 아무런 부속기관도 부착되어 있지 않다. H ypo s t ome 은 up pe r l ip의 뒷부분으로 그 사이 에 입 과 man di ble 이 있으며 h~s t ome 의 측면부에 m axi llae 가 발달한다. Man di ble 은 음식물을 저작하는 역할을 하며 m axi llae 는 저작과 순환의 두 가지 역할을 맡고 있다• Ante n nules, ante n nae, mand ibl e, m axi llae 등의 부속기 관 ( awend ages ) 은 기 능이나 구조 모양등이 다소 툴리나 근본적으로는 3 부분으로 구성되어 있 다. 죽, 이 들 3 부분은 pro to p odite, endop odite, exo podit e 로서 그림 2 와 같으
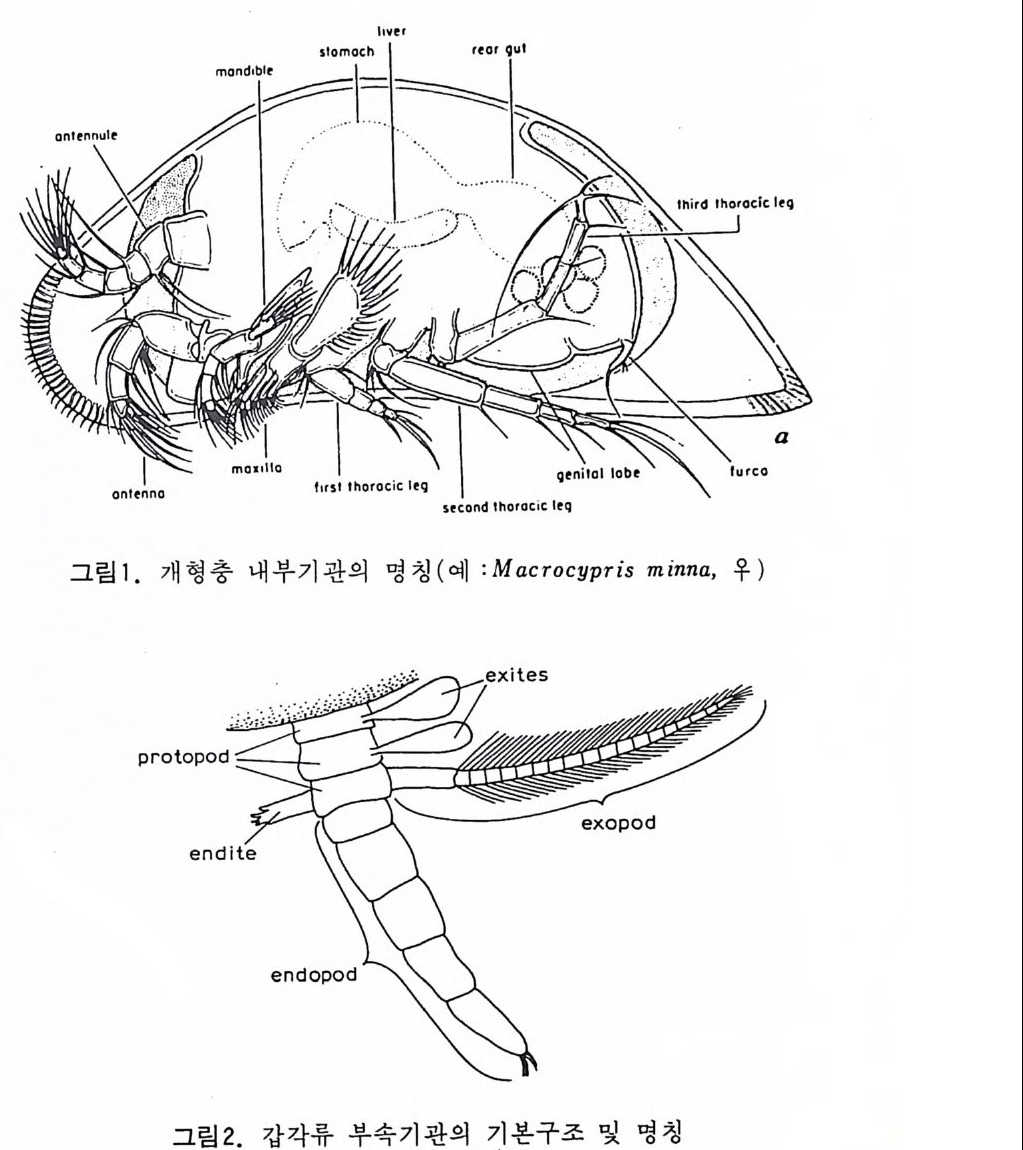 slomoch 'I. , rror gu t
slomoch 'I. , rror gu t
며 이들 3 부분은 모두 몇 개의 p odomeres 로 구성되고 있다 . 흉부는위·장·생식기관등등의중요한내부기관을거의 모두 포함하 고 있으며 3 쌍의 th oracic le g s 가 부착되어 있다. 생식기관은 3 번째 th oracic leg s 뒤에 위치하며 가장 뒤쪽에 f urcae 한쌍
이 자리하고 있다. 대부분의 해수 _. ... 의 정우 재 1 Ihoracic l e g s 는 이동에 사용되며 Cy pr id ida e 같은 담수종의 정우 음식섭취 기관으로 사용된다. 제 2 tho rac ic le g s 는 개형충의 대부분을 차지하는 Podoco p a 의 정우 걷고 · 기고 • 고착하는데 사용되고 있다. 제 3 tho racic l e g s 는 이동하는 데 이용되는 종도 있고 때로는 몸을 씻는 데 사용하는 종류도 있다. Furcae 는 사실상 부속기관 (a pp en dag es) 에는 포함되지 않으나 이동기능을 가지며 혹은 다른 기관에 의해 몸체륭 이동시에 보조기능을 갖는다. 다론 부속기관처럼 많은 pod omeres 에 의해 구성되지 못하고 단순한 한 개의 막 대모양을 하고 있다. 유기 체는 exoskele t on 이 라는 외 벽 으로 싸여 있으며, 이 exoskele t on 의 최 외측막은 키틴질층 (cJ 1 iti 11ous la y er) 으로 내측막은 표피층 (epi dermal lay e r) 으로 구성되어 있다. 유기체륭 둘러싼 이 exoskele1011 은 내측벽(i nner lamella) 과 외측벽 (out er lame lla ). 즉 du p li calures 라는 이중벽을 이루며, 이 du p l i ca t ures 의 최외측벅 말하자면 외부와 접하는 부분은 딱딱한 석회질막을 이루며 얇게 키탄질 막 으로 싸여 있다. (그림 3 참조). 이처럼 키탄질 막으로 피복이 된 석회질막은 개형충의 각인 valve 룽 안 들며 양쪽 valve 를 일 컬 어 cara p ace 라 한다. 이 두 valve 가 접 하는 부위룹 cardi na l mar gi n 과 ext ra cardi na l margi n 둘로 구분한다. Cardi na l mar gi n 은 동쪽의 hin g e mar gi n 을 말하며 ext racardi na l margi n 은 동쪽을 제외한 전 • 후부와 복부의 가장자리를 일컬으나 형태에 관한 단원 에서 설명키로 하고 생략한다. 유기체가 성장함에 따라 옛 석회질 껍징을 벗어 던지고 새로운 각이 표 피세포에 의해 분비되는, 즉 탈피현상 (moull i n g)을 갖는다. 따라서 생장선은 없는 것이 보통이다. 그러나 고생대의 Cr yptop hy llus 속의 경우 탈피한 유생 의 각이 순차적으로 殺頂部에 남아 생장선상의 구조를 형성한다(그림 4 참 조). C yt heracea 의 대부분은 9 개의 幼殺(i ns t ars) 을 갖는다. C yp r idi n i dae 에 속 하는 명명 종은 6 개의 탈피단계를 보이며, Haloc yp r idi dae 의 경우 7 단계,즉 종에 따라 다소 차이를 가지며 성장한다.
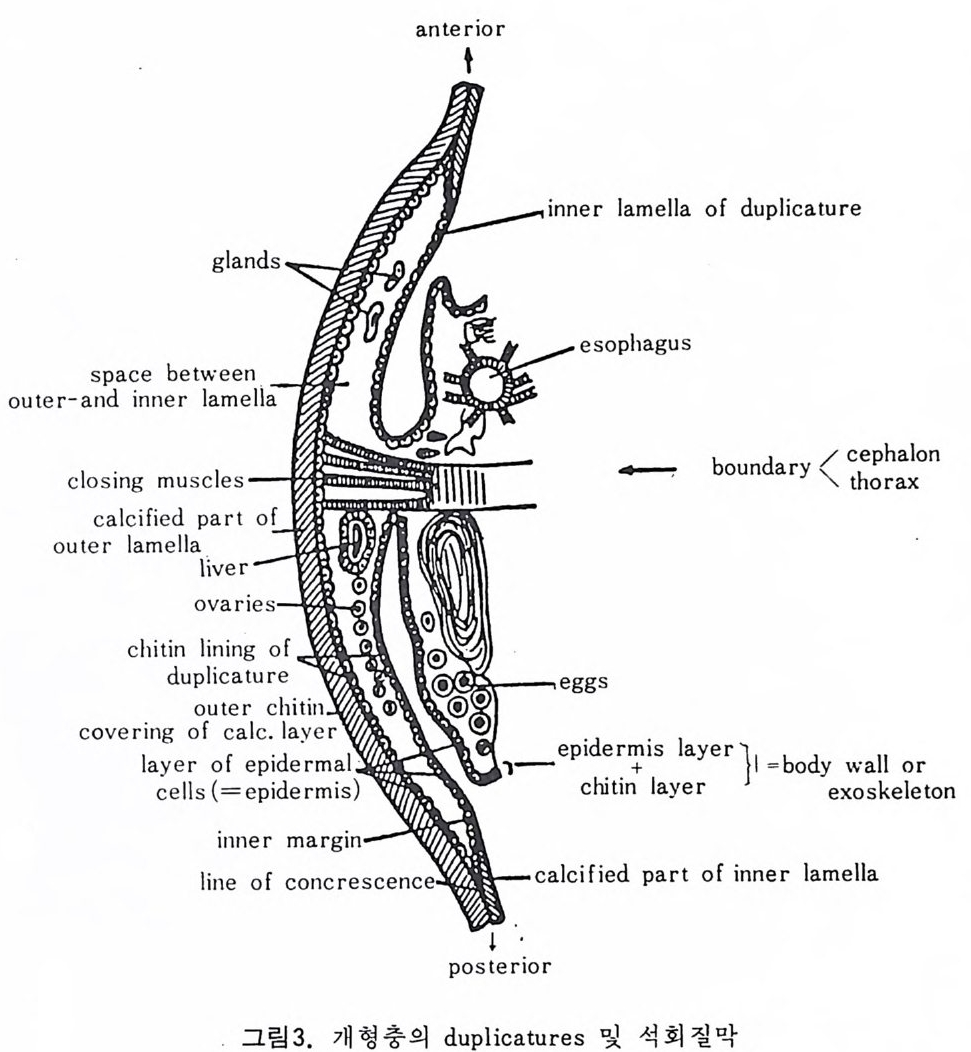 ante rio r
ante rio r
다소 차이룰 갖는 탈피현상에 의해 유생은 성체 (adu lt)로 자라게 되는데 이때 유생과 성체 사이에는 크기, 비율, dup li c a tu r e, hin ge 등등 많은 차이 륭 보인다. 따라서 유각에 대한 연구를 병행하여 개체발생 (on tog en y)을 밝 혀 보고 동시에 계동발생(p hy lo gen y)학적인 견지의 고찰을 요한다. 일반적으로 담수종의 경우 해수종보다 빠른 성장속도를 보인다. 또한 부속기관의 발생 역시 종의 각 탈피단계에 따라 차이룽 갖는다(그 링 5 참조).
 그림 4. 순차적인 幼生殺울 보여주는 Cryp top h y ll us obot o i de s. (각의 단면)
그림 4. 순차적인 幼生殺울 보여주는 Cryp top h y ll us obot o i de s. (각의 단면)
동쪽에 발달된 치 아구조 (hing e m ent) 에 의 해 두 valve 가 맞뭉려 있으며 , 또 한 이 치구조를 따라 좁고 길게 cu ti cle 질 인대 O iga men t)가 발달되어 있다. Valve 는 유기체와 각을 연결하고 있는 근조직 (muscles) 에 의해 열리고 닫히게 되는데, 죽 내전근 (adduc t or muscle) 이 수축시 인대는 이완하며 valve 는 닫히 게 된다. Mand i ble 과 연결된 숱한 근조직은 각의 동쪽 부위 |(dorsal margi n) 혹은 전복부 (an t eroven t ral) 에 연결되고 있다. 穀에 근조직이 부착시 각 표면에 돌기같은 자국이 생기는데 화석의 경 우 유기체는 보존되지 않으며 자국인 근혼 (muscle scars) 만이 남는데, 이들 은 화석을 분류하는데 무엇보다도 중요한 역할을 담당하고 있다. 근혼은 穀의 어 느 부위 에 있느냐에 따라 크게 중앙근혼장 (cent ra l muscle scar fiel d) 와 배 부근혼 장 (dorsal muscle scar field ) 둘로 구분되 는데 자세 한 것은 형태에 관한 부분에서 다루기로 한다. 그 외 개형충의 소화계나 순환· 신경계 등을 살펴보기로 하자. 보통 개형충은 입 • 식도 • 위 • 항문을 갖는다. Podoco pi da 目예 속하는 종 류는 짧은 장을 가지며, 간혹은 간장 O i vers) 에서 소화액을 분비하는 종류 도 있다 . 순환은 유기 체 막이 나 아가미 (gills) , 그 외 각 부속기관에 부착된 진동판 (vibr ato r y pla te s ) 에 의 해 이 루어 진 다. My od ocop ida 目 에 속하는 개 형 충에 서 는 심장이 큰 역할을 한다. 7 사형충의 신경계는 뇌 (cerebrum ), 복부의 신경절(ga n g li a), 환식도 신경 절 (cir c umesop h a ge al g an g li on) 로 구성된다. 신경은 뇌로부터 눈, ante n nules, ante n nae 등의 각 기관으로 전달되며 운동신경은 보통 환식도 신경절 (c i r cumesop h a ge al gan g li on ) 으 로부 터 ante n nae 등에 전 달된 다. 감각기관으로서 큰 역할을 담당하는 눈의 경우를 보면, Podoco pa 에 속하 는 대부분의 종이 만안을 가지 며, M y odoco pa 의 C yp r idi n i dae 에 속하는 현생 종은 정가운데의 눈 양옆에 한 쌍의 측눈을 갖는다. 많은 종은 눈을 갖지 않고 있으며 간혹은 앞쪽의 기관이 비슷한 가능을 갖기도 한다. 개형충의경우언제나암·수의구분이있다.암·수모두한쌍의 대 칭성 생식기관을 가지며 일부는 체내에 일부는 제외에 위치한다. 암 컷 의 경 우 난소 (ovarie s ) , 자궁 (ut er i) , 질 (vagi na e) , 정 자 저 장소 (semi n- al rece pt ales) 를 가지며, 수컷의 경우 고환(t es t es), vasa defe r enti a, 음경
(pen es) 등을 갖는다. 간혹 Zenker's or ga n 을 갖는 종류도 있다. 암·수의 구분을 갖는다 하여 모든 개형충류가 양성생식을 하는 것은 아 니다. 때론 처녀생식 (p ar t heno g enes i s) 에 의해 형성되며, 어떤 담수종의 경 우 지역적 기후에 따라 생식조건을 달리하는 현상을 보이기도 하는데 이 룰 지리적 처녀생식이라 한다 그외 환경요소에 따른 개형충의 생물학적 적응성 및 반응, 영향력 등은 생태에 관한 부분에서 언급하기로 한다. 2· 3 형태 및 분류 2 • 3 • 1 형태 개형충은 수차의 탈피현상을 거쳐 성장하며 마지막 단계에서 같 은 종이 라도 암 • 수에 따른 동종이형 현상을 갖는다(그림 6 참조).
 0.5 mm
0.5 mm
따라서 탈피 로 인한 각 성 장단계 상의 幼穀과 性的 二型 性 현 상 (sexual di- mor ph i sm) 에 의한 형태학적 차이룰 고려해야 한다. 그 외 에 도 crumi na l dim orph is m , ant ral dim orph is m 등을 들 수 있 다. 형태학적 차이에는 외형적 차이는 물론 내형적 차이도 있다는 점을 고 려하면서 우선 가장 기초적인 각의 전·후 위치관계 (or i en t a ti on) 부터 설명 하고자 한다 . 개형충의 외형만을 간단히 보고서 전 • 후 관계를 밝혀보면 그림 7 에서와 같이 고생대 종의 경우 brood po uch 나 medi an lobe 가 있다든가 마름모꼴 외 형의 경사 방향 쪽이 후면에 해당한다. 중생대_신생대 종의 경우 측면에 서 볼 때 높이 가 최대 치를 갖는 쪽이 보통 앞이 되며, 위에서 볼 때는 반
 ORIENTATION OF CARAPACE
ORIENTATION OF CARAPACE
대로 뒤쪽이 최대치를 갖는다. 그외 뾰족한 끝이 있다든가 날개구조 (alae) 가 발달해 있는 쪽이 후면이 된다(그림 7 참조). 그 외에도 안결절 (ey e tur bercle) 이 발달되어 있다든가 표면장식 드0드 .0. Q근_. 보고서도 전·후 관계를 알 수 있으나 좀더 뒤에서 자세히 다루기로 하고, 세부적인 설명에 앞서 간단히 개형충의 측면부 각 부위의 위치에 따른 명 칭과 형태학적 명칭을 살펴보기로 한다. 전 부 (ante rio r ) 와 후부 (pos te rio r ) , 배 부 (dorsal) 와 복부 (vent ra.l ) 로 크게 구 분해 볼 수 있는 7 사형충의 경우, 좀더 작은 부위의 위치를 정확히 표현하 기 위하여 13 으로 세분하였다(그림 8 참조). 그 외에도 경우에 따라 내전 근혼 (adducto r muscle scar) 의 앞부분을 지 적 할 때 pre adducto ria l area, 뒷부 분을 지 적 할 경 우 pos t adduc t or i al 라는 식 으로 pr e-나 po st- 를 넣 어 사용되 고있다. 외형은 보통 卵形, 타원형, 정방형을 가지며, 때론 만장방형, 부등변사 변형, 원형을 갖는다.
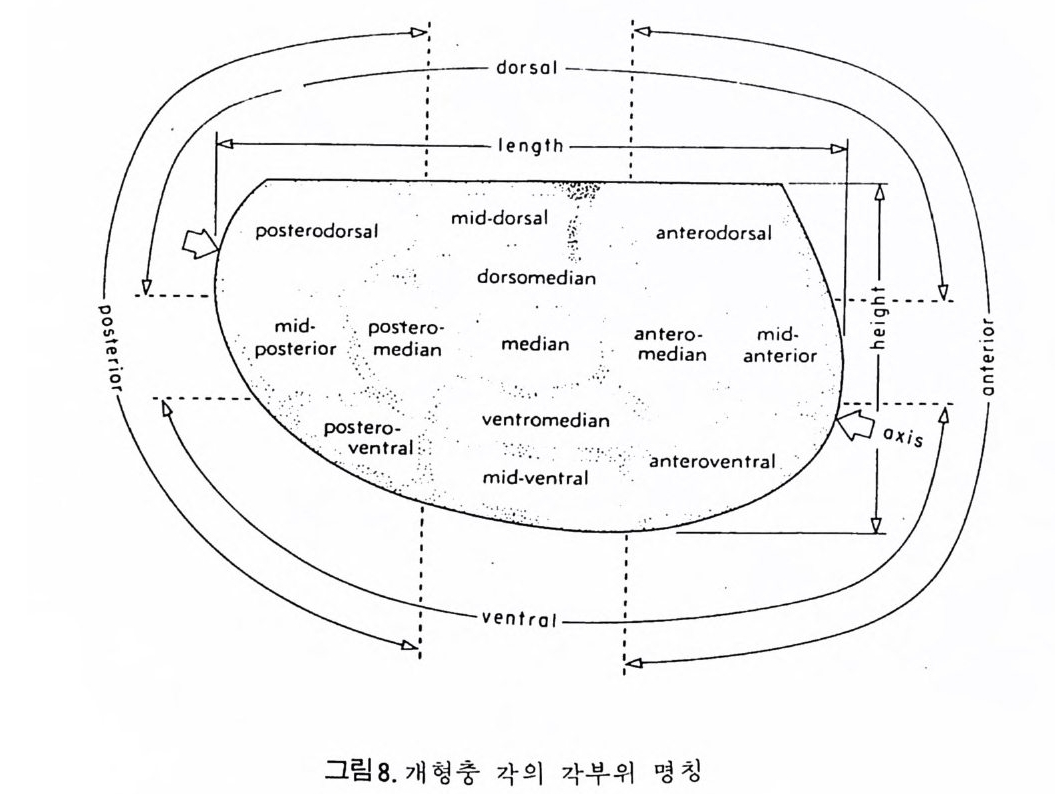 /v/ / dleonrsg atlh — —’‘’’’’’’
/v/ / dleonrsg atlh — —’‘’’’’’’
매연 (dorsal e dg e) 은 볼록하거나 직선이며 복연 (vent ra l mar gi n) 은 볼록, 직선, 오목한 모습을 보인다. 치 아주변부를 dorsum 이 라 하며 , 이 끝부분의 殺의 윤곽선 이 급변하는 부 위 륭 cardi na l corners 라 한다. 각의 양끝과 dorsal border 가 만나는 이 car- dina l corners 는 예각을 이루거나 혹은 둔각을 이루며 이는 분류에 중요한 보조역 할을 맡는다(그림 9 참조). 그 외에도 최고 높이룰 보이는 부분이 각의 전 • 후 • 중앙부 어느 쪽에 위 치 하는가에 따라서 pre p le te , pos tp le te , am p le t e 로 구분한다. Pr e p le t e 는 穀의 전반부에, p os tp le t e 는 穀의 후반부에, am p le t e 는 중앙부 에 위치하는 경우롤 일컫는다 . S\ vi n g이 란 최고 높이룽 갖는 부분이 각의 맨끝에 가까운 경우를 말하며, 따라서 p re p le t e 나 p os tp le t e 殺안이 이룽 갖게 된다. 두 valve 가 닫힐 때 맞물리는 부위를 접촉대 (conta c t mar gi n) 라 하며 크 게 두 가지로 구분되나 좀더 뒤에서 설명키로 한다. 穀 표연은 아우런 장식이 없이 미끈한 종류부터 복잡한 표면장석을 갖 는 종류 등 다양하다. 간단히 표연장식에 대해 알아 보면 다음과 같다(그 링 10 참조). 각의 표면 장식 으로는 gra nules, pu stu l es, str i a e , costa e , pits, spi ne s, reti cu la 등을 둘 수 있다. 표면 전체 혹은 일부가 아주 작고 도톨도톨한 돌기모양을 하며 밀집되 어 있을 때를 g ranules 라 한다. 돌기가 좀더 커지고 현저히 튀어 나왔을 때를 pu stu l ae 혹은 pa pill ae 라 한다. S t r i ae 란 가늘고 긴 중구조로서 En t omos i s 같은 종류는 표면 전체가 평행 s t r i ae 로 이루어 져 있다. r i b 이 좀더 쿨 경우는 coas t ae 라 한다. P it s 란 표떤이 움푹웅푹 패여든 구조로서 벽공관(p ore canals) 처럼 완전 히 구영 (op en in g ) 이 뚫린 것 이 아니 라 단지 패 인 자국에 지 나지 않는다. 고생대에서 현생종에 이르기까지 가장 많은 비율을 차지하는 것은 reti - cula t e 로서 막대 (b ar) 가 서로 교차하며 그 사이는 함몰부 (de p ress i on) 가 보 이는, 즉 격자상이다 . 이때 막대 (bar) 의 모양이 넓고 편평하며 상대적으로 함울부의 비율이 줄어들연서 이는 pun cta te 상태로 옮아간다. 감은 종류라도 탈피현상에 의해 표면장식이 툴리므로 분류에 있어서는 언제나 주의하여야 한다. Lobe 란 殺의 일부가 위로 튀어나온 부분응 망하며 반대로 움푹 들어간
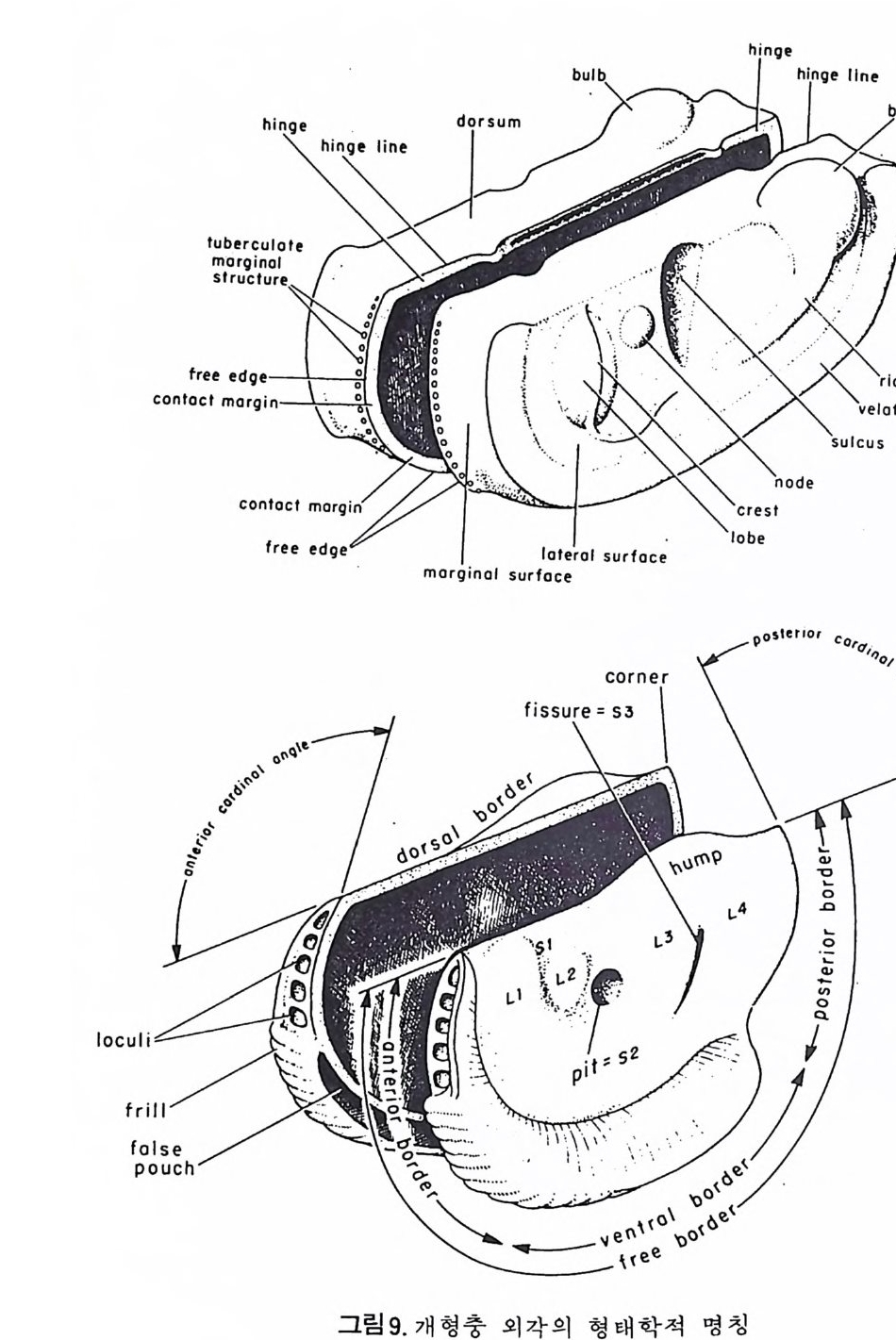 h.’ n -/
h.’ n -/
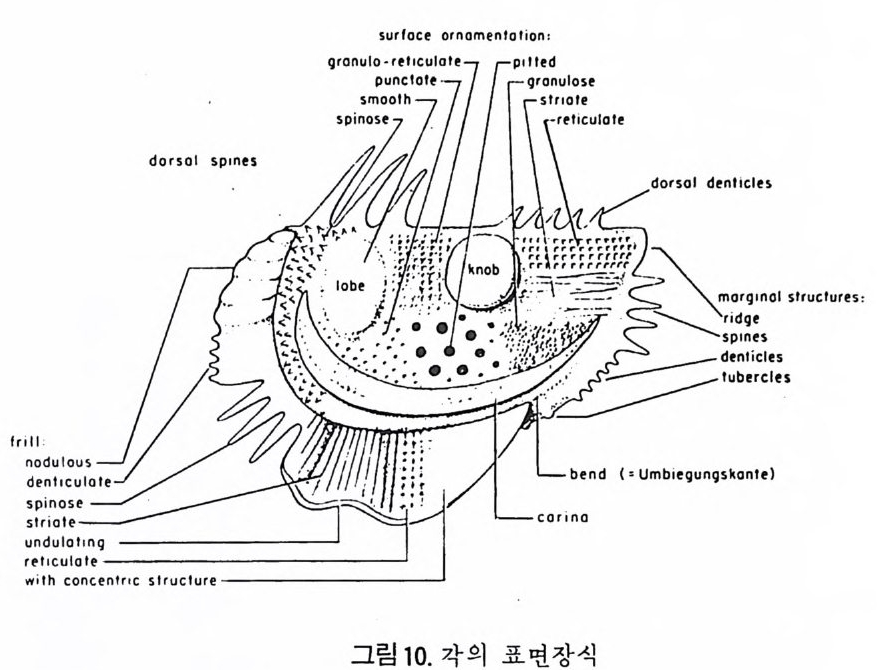 ,urloce ornom en t o to o n,
,urloce ornom en t o to o n,
부위 륭 s u lci라 한다. Sulc i가 길게 발달될 경우는 fur rows 혹은 fiss ures 라 고 일컫는다. 상승부위인 lobe 의 반대편 쪽.즉 각의 안쪽의 경우는 움푹 들어가 있으 며 이는 유기체의 해부학적인 것과 깊은 관련을 갖는다. 따라서 분류에 많 이 이용되어 왔으나 사실상 언제나 내부기관의 영향에 의하는 것만은 아 니므로 많은 혼란을 일으켜 왔다 . 편 리 상 lobes 는 전 부 (ant e rio r ) 쪽부 터 후부 (po ste rio r ) 방향으로 L1, L2, L3, L. 로 구분한다. 4 개의 lobes 를 모두 갖는 것을 qu adrilo b ate valve, 3 개의 ]obes(L1·L2•L3) 룽 갖는 것을 tri lo b ate valve, 2 개의 lobes(L1·L2) 를 갖는 것을 bilo b ate valve 라 한다. Sulc i의 경우도 역시 앞에서부터 S1, S2, S3 로 부르며 unis u lcate , bis u lc- ate , t r i sulca t e 라는 명 칭을 사용한다. 가장 중요한 것은 S2 로서 이는 내전근의 위치를 지시한다(그림 11 참조). 같은 속에 속하는 개형충의 경우라도 lobes 나 sulc i의 수가 다소 툴리므
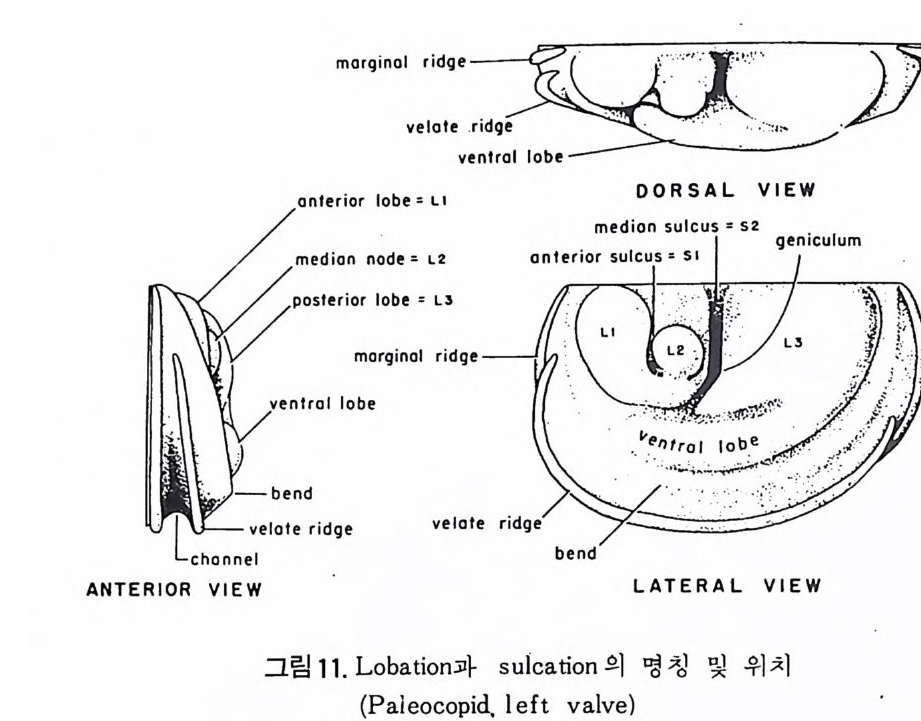 mor91noIve:: ndq . \ ]J
mor91noIve:: ndq . \ ]J
로 이것만으로는 분류의 기준이 될 수 없지만 다른 것에 의한 분류에 있어 보조역할로서 아주 중요하다. 표면장식에 대해서는 이 정도로 간단히 끝내고 각의 복부 (ven t ral area) 및 배 부 (dorsal area) 에 관해 살펴 본 다. 두 valve 가 열릴 때 自由緣(fr ee e dg e) ’ 을 따라 열리게 되는데 이는 치아 가 발달된 배부 (dorsal p ar t)의 양끝에서 전부 (an t er i or pa rt) • 복부 (ven t ral pa rt) • 후부'i> os t er i or par t)에 걸치는 부분을 일컫는다. 이때 자유연의 가 장 밑부분. 죽 복부를 ven t er 라 한다. 그러나 복부의 형태학적 구조는 비단 복부에만 한정되는 것이 아니고전 후부를 부분적 혹은 전부에 걸쳐 연결되어 있으므로, 이 경우를 advente r 라 하며 양 부위를 포함한 복부를 말한다. Valve 가 닫히게 될 때 두 valve 가 맞닿는 부분을 접촉대라 하며 접촉되는 특성에 따라 크게 둘로 구분해 볼 수 있다. 죽 두 valve 중 한쪽이 다른 한 쪽 valve 보다 클 경우는 작은 것을 겹 쳐 덮게 되는데 (overla ppi n g) 만일 비
슷하거나 갇을 때는 겹치는 현상을 볼 수 없다. 접 촉대 는, 자유연을 따라 발달한 selv ag e 라는 r idg e 에 의 해 간단히 c六L 걷口 링으로써 끝나는 경우가 있는가 하면 비교적 복잡한 접촉대를 갖는 것도 있다(그링 12 참조)•
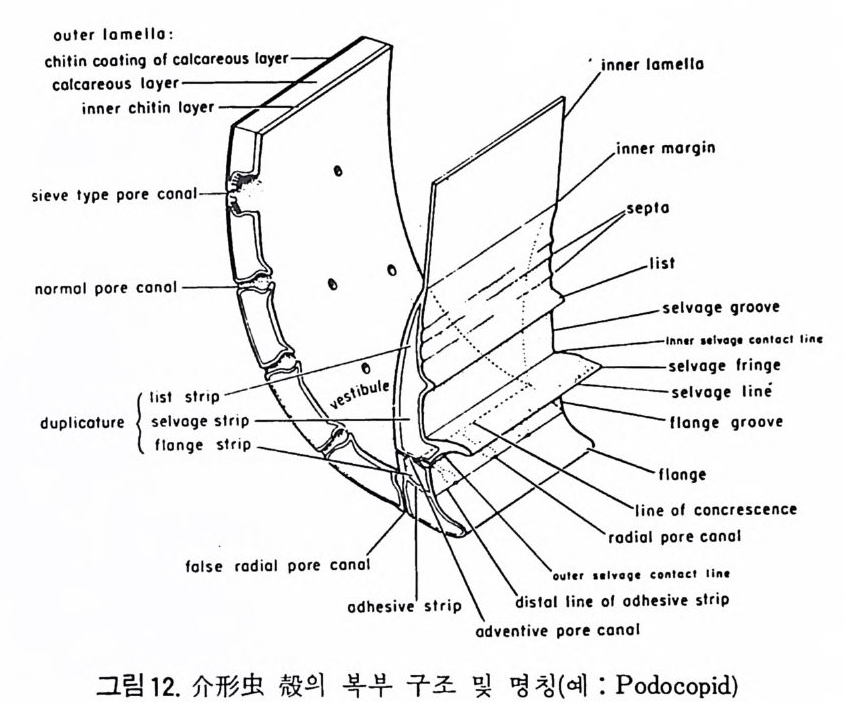 out er lomello·
out er lomello·
각이 닫힐 때 s elv ag e 는 반대편 각의 g roove 와 맞물린다. 일부 개형충의 경우 내측 벽이 석회화에 의해 du pli ca t ure 를 만들게 되는 데 이 경우 접촉대는 좀 복잡한 양상을 갖게 된다. Selva g e 보다 좀더 안쪽에는 소규모의 r idg e 형 태를 가진 li s t가. 바깥측에 는 fl an g e 가 발달한다. 이때 fl an g e 는 내측벽에 발달되었다고 하기보다는 외 측벽에 발달한다고 봉이 타당하다 . Selva g e 와 flan g e 사이에는 움푹한 flan g e g roove 가. sel vag e 와 lis t 사이에 는 selvag e g roove 가 발달하는데 이 런 구조를 갖는 이 du p li ca t ure 는 외측벽 과 봉합선 (line of concrescence) 이 라는 키 탄질 성분에 의 해 연결되고 있다. 또한 두 내 • 외측벽 사이의 맞닿는 부분에 가늘고 긴 몇 개의 구멍이 방 달하는데 이 룽 방사벽 공관 (radi al por e canal) 이 라고 한다.
벽공관의 경우는 외측벽면상에 수직으로 뚫린 수직벽공관 (normal po re canal) 과 두 벽 접촉부위의 방사 벽공관 둘로 구분되며, 이는 분류에 중요 한 구실을 하게 된다. 벽공(p ore) 의 수나 배열상태 그 외에도 벽공 자체내의 구조(예 뭉 들면 격자상 (s i eve typ e) 이냐 아니면 단지 한 개 구멍이냐), 벽공내어! 탈의 유 • 무라든가 등등 숱한 다양성을 갖게 되는데, 최근들어 분류에 있어 뺄 수 없는 중요한 구실을 하고 있다(그링 13 참조). 살아 있는 개형충의 내측벽과 외측벽 사이의 공간을 ves ti bul e 이라 한다. 복부구조는 개형충의 종류에 따라 너무나도 다양하며, 어떤 종류의 정 우 여 러 rid ge, fri ll , flan g e 동을 갖게 된다. 형태의 다양성으로 인해 Jaa nusson( 19 57) 같은 사람은 velate , his r ia l ,ca- rin a l st ru ct u re 등의 명 칭을 사용하며 여 러 형 태룽 표시 하고 있는데 모든 7 사형충에 적용되지 못하기 때문에 많은 문제성과 혼란점을 갖는다. 배부는 指가 발달되고 있는 동쪽부분, 즉 cardi n~ I mar gi n 을 말한다. 배부가 전 • 후부와 연결되는 부분을 cardi na l corner 라 하며 치구조선 01 i n ge line ) 과 측면부 연 장선과의 이 루는 각을 cardi na l an g le 이 라 한다. 예 각을
 ·--·- · ·:~ , .,
·--·- · ·:~ , .,
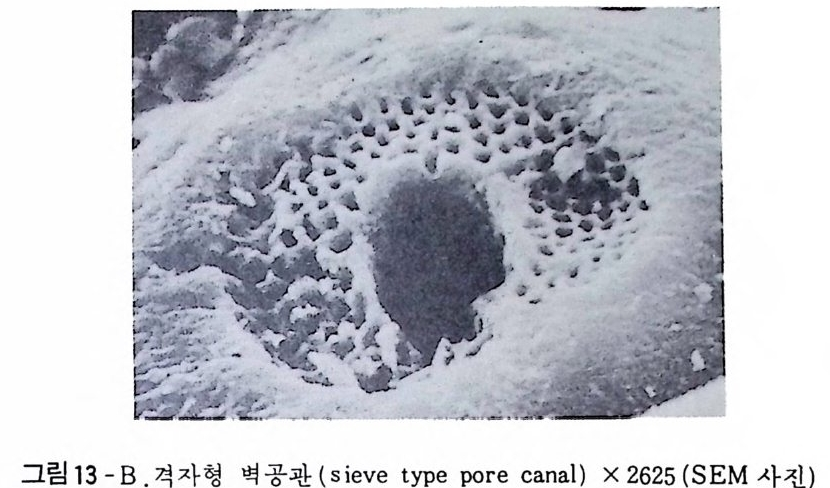 그림 13 -8 겨자형 벽 공관 (sie v e type po re canal) X 2625 (SEM 사전)
그림 13 -8 겨자형 벽 공관 (sie v e type po re canal) X 2625 (SEM 사전)
이 루며 만나거 나 둔각을 이 루는데 Ba ir d ia나 C yt herella 처 럼 각을 측정 키 어 려운 정우도 많다 . 치아 주변부 릅 통칭 hing e mar gi n 이라 부른다. 배부에서 무엇보다 중요 한 것은 여시 치아구조로서 너무도 다양하며 특히 화석으로 보존이 좋은 까닭에 분류학상 가장 중요하다고도 볼 수 있다. 자세한 것은 좀더 뒤에 근혼과 함께 별도로 다루기로 하며 그 외에도 배부는 각의 or i en t a ti on 을 밝히는 데 큰 여할을 한다 . 개형충의 형태학적 고찰에 있어 가장 중요하다고 봉 수 있는 근혼과 치 아구조에 대해 살펴보기로 한다. 우선 근혼이란, 개형충이 살아 있을 당시의 근조칙이 穀에 부착되어 穀 의 개폐 및 부속기관의 운동을 맡아보던 것으로서 죽음과 동시에 유기체 는 없어지고 흔적만 남아 있는 것인데 이 근조직의 흔적을 이용한 형태기 능학적 해석이나 그 밖의 많은 연구를 통하여 많은 생물학적 해석을 할 수 있으며 무엇보다도 분류에 있어 중요한 역할을 하고 있다. 근혼은 valve 내측에서 관찰되며, 穀내의 분포 위치에 따라 중앙 근혼장 및 배부근혼장 둘로 구분된다(그림 14 참조). 각각에 위 치하는 근혼은 기원 (or igi n) 을 달리한다. 우선 중앙근혼장은 내 전 근혼군과 mand ibu lay gro up 및 fro nta l gro up 으로 구성되어 있다. 내전 근혼군은 각의 개폐를 담당하던 내전근의 자국(근혼)으로서 다른
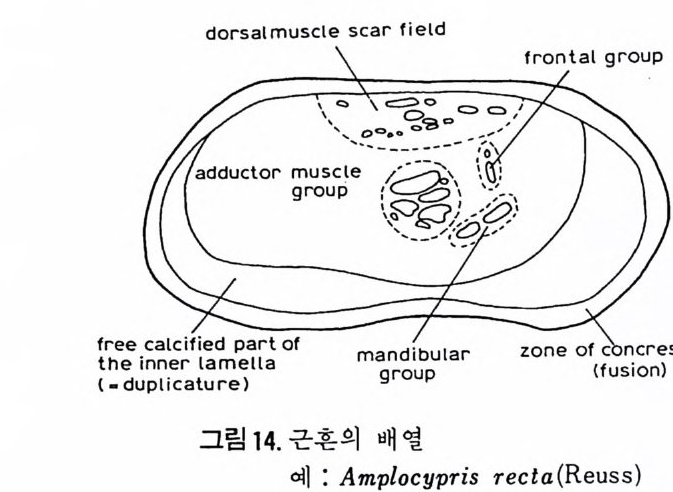 dorsalmuscle scar fie ld
dorsalmuscle scar fie ld
근혼보다 분류에 중요한 역 할을 맡는다. 이 근혼의 전복 부 (an t ero-ven t ral ) 에 mand i bular 이 위치하며 이는 근조직의 자국이라기보다는 키틴질로 된 sup por t rods 의 자국이 다. 이 위에 fron t al g rou p이 위 치 하며 이는 mand ibu lar muscle 과 키 탄 질 endo - skele to n 에 연경된 근조직의 자국이라 할 수 있다. 중앙 근혼장을 이루는 세 g rou p의 근혼과는 별도로 간혹 몇몇 종류 는 fr on t al 과 adduct o r g rou p사이에 독립된 근혼을 갖는데 이를 ful cral scar 라 한다. 배부근혼군은 매우 다양하며, 그 기원에 대한 연구는 아직까지 멸명 종에 국한되어 연구되고 있는 실정이다. 근혼 중 가장 중요한 것은 내전근혼으로서 화석화 과칭에서 가장 보 존이 잘 되고 있으며 이는 과(fa m i l y)를 분류하는 기준으로 이용되고 있다. 때론 屬이나 種의 구분에도 이용된다. 근혼에 의한 분류는 고생대 개형충보다는 중생대 • 신생대의 경우에 더 욱 중요한 역할을 한다. 근혼은 개체발생에 따른 다양성을 보이고 있는 경우도 있음을 고려해 야 한다. 따라서 같은 종이라도 성체와 유체에 따른 양상이 틀린 경우도 있으므로 분류에 있어 신중해야 한다. 다양한 모습을 보여주는 근혼의 모습을 예로 들어본다(그림 15 참조).
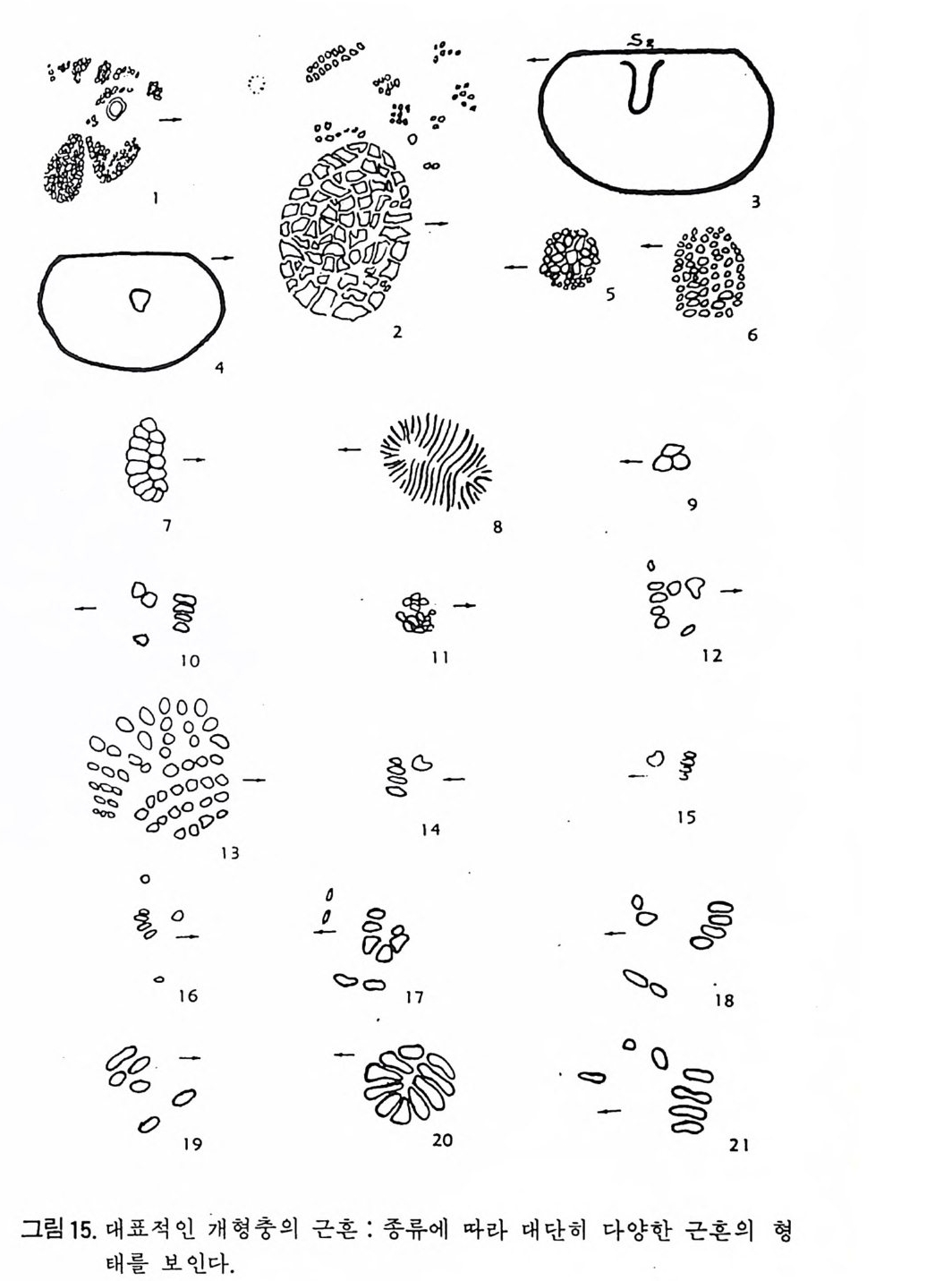 <을i떄 ’ 'r8 짜…I I Q- - .. ... . 00念。 8Oooo 守ao쯔72W 2 oo一• ; •。• ·• •{ -鷄 U令- o懷g綠OO &禎oOO 36
<을i떄 ’ 'r8 짜…I I Q- - .. ... . 00念。 8Oooo 守ao쯔72W 2 oo一• ; •。• ·• •{ -鷄 U令- o懷g綠OO &禎oOO 36
치아구조는 분류에 있어 아주 중요한 요소로서 이미 절멸 되 고 없는 Ar- cheoco pi ds 라든가 그외 t-.l yo docop ins , Cladocoµns 등에는 전 혀 방당되 지 않 고 있다. 이는 가장 원시적인 것으로서 그 이후 진화과정운 거쳐 다양한 모습을 보여주고 있는데 각 치아구조 사이의 연속적인 전화모습 등은 전혀 분 수가 없다. 단지 단편적이고 독립적인 양상으로 한 개체내에서도 탈피 현상 과정에서 치아구조도 따라 변하는 종류가 있으므로 種의 구분에 많 은 혼란을 가져다 준다. 개체발생뿐만 아니라 계통발생 천상을 보이여 다 양한 형태륭 갖는 치아구조는 분류상 속의 구별에 가장 중요한 역할을 담 당하고 있다. Zalany i (1929) 에 의 해 처 음으로 치 아구조에 대 한 형 태 분류 (morph o log ica l classif ica ti on ) 가 이루어 졌으며 그는 den t a t e 와 nonde n ta t e 둘로 구분하였다. Schwey e r (1940) 에 의 해 g le i ch t e il ig e 와 versch i eden t e il ig e 로 구분되 였으며 Bold(1946) 에 와서 Podocop ida g rou p의 치 아구조는 셋으로 (adon t ·tax odont ·he- ter odont) 나뉘었다. 계속 발전과정을 거쳐 더욱 세분화되었으며, 예룹 들 자연 Malkin (1953) 에 의 해 adont ·desmodont ·tax odont • crasid o nt • he te r odont • cryp tod ont • archid o nt 의 일 곱으로. Sy lv es te r .:. Br adley (19 56) 에 의 해 lop h o - dont _· m erodont • ento m odont • lobodont • schiz o dont • amp h id o nt 여 섯으로. H01ve 와 Laurencic h 0958) 에 의 해 백 악기 종 치 아구조를 adont • pr io n odont • lop h odont • merodont • ento m odont • lobodont • amp h id o nt • schiz o dont • go ng ylo - dont 로 구분. 그중 merodont 를 pa leomerodont • holomerodont • anti m erodont • hemi m erodont 로 am p h i don t도 pa ramp h id o nt • hemi am p h id o nt • holamp h id o nt 등 좀 더 세분하였다 . 그 외에도 많은 개형충학자에 의해 숱한 이름으로 구분되 고 있는데 같은 모습의 치아구조에 다른 이름이 붙여진 정우, 또 같은 이 름이라도 학자에 따라 이름붙인 기준이 툴리다든가, 그 외에도 이름의 한 계성, 즉 광의로서의 한계와 협의로서의 한계가 툴리므로 통일성이 없어 실 제상 문제점이 많다. 치아구조의 분류 • 형태 • 특징을 설명하기 이전에 우선 가장 기초적인 치 아의 각부 명칭을 익히기로 한다. - 그림 16 에서 보는 바와 같이 우선 치아구조는 위치에 따라 크게 셋으로 구분 (ante r io r • me dian • pos te rio r ) 되 며 med ian elemen t는 다시 an t erom edi an 과 p os t erome di an 으로 구분된 다. 치 아구조는 bars, gro oves, tee th , socket_s 등으로 구성 되 는데 우선 bars 란 r idg e 라고도 하며 좁고 길게 발달된 돌출부라고 할 수 있다. 이 돌출부
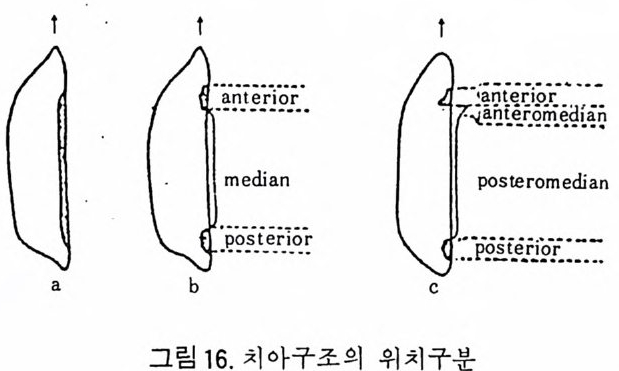 -t -
-t -
에 반하여 좀고 길게 움푹 패인 홍을 g roove 라 한다 . 따라서 r idg e 는 반대편 穀에 발달한 g roove 와 맞물리게 되는데 표면이 매끈한 경우와 둡니처럽 꺼 끌꺼꿀한 경우가 있다. 톱니 같은 것이 발달한 경우를 crenula t e 로 표현하 며 이는 접촉대에 수직 혹은 수평면에 발달한다(그림 17 참조) .
 •'
•'
Tee t h 란 이빨처럼 뾰족나온 것이며 이에 반해 쏙쏙 들어간 것을 socket 라 하는데 양각이 맞물릴 때 t ee t h 는 socke t와 맞물리게 된다. 그 외에도 치아구조와 관련되는 많은 이름이 있으나 설명은 생략하기로 하고 그림으 로 대치하기로 한다(그림 18 참조). 현재 는 h i n g emen t를 크게 adont • merodont • am p h i don t로 구분해 쓰고 있으며 merodon t속에 lop h odont • merodont • desmodont • Pseudadont • p en t odon t가 구별되며 am p h i don t는 다시 ento m odont • .sc hiz o dont • 협 의 의 amp h id o nt • g on gy lodon t로 구분되고 있다.
 solvog o• r mo 『gi n f\lon 'g e selvag e
solvog o• r mo 『gi n f\lon 'g e selvag e
Lop h odont 특징 을 갖는 것 으로서 는 By tho cy the rin a e, . C;y the rurin a e, Para- doxosto m ati na e 등이 있다. Xesto leberid i n a e, Cy the rop ter in a e, Cy the rin a e 등은 me·ro d ont typ e 을 보여 주며 Neocyt he rid e id i n a e 와 Eucy the rin a e 의 경 우 desmodont hin g e ment 를, Krit hin a e 의 경 우 pse udadont type 을, Pecto c y the rin a e 는 pe nto d ont type 을 보 여준다. Arnph id o nt 중 가장 원시 적 인 en t omodon t에는 Pro g onoc yt her i nae 가 속하며 Sch i zoc yt her i nae 는 schiz o dont type을, Trach y leber i d i nae 와 Hemi cy the rin a e 는 amp h id o nt typ e 을, Loxoconch i nae 는 go ng ylo dont type의 h ing emen t를 갖 는다. 이 들 h ing e 의 · 모습을 도표로 보면 다음과 갇다(그림 19, 20 참조).
 /\/\\
/\/\\
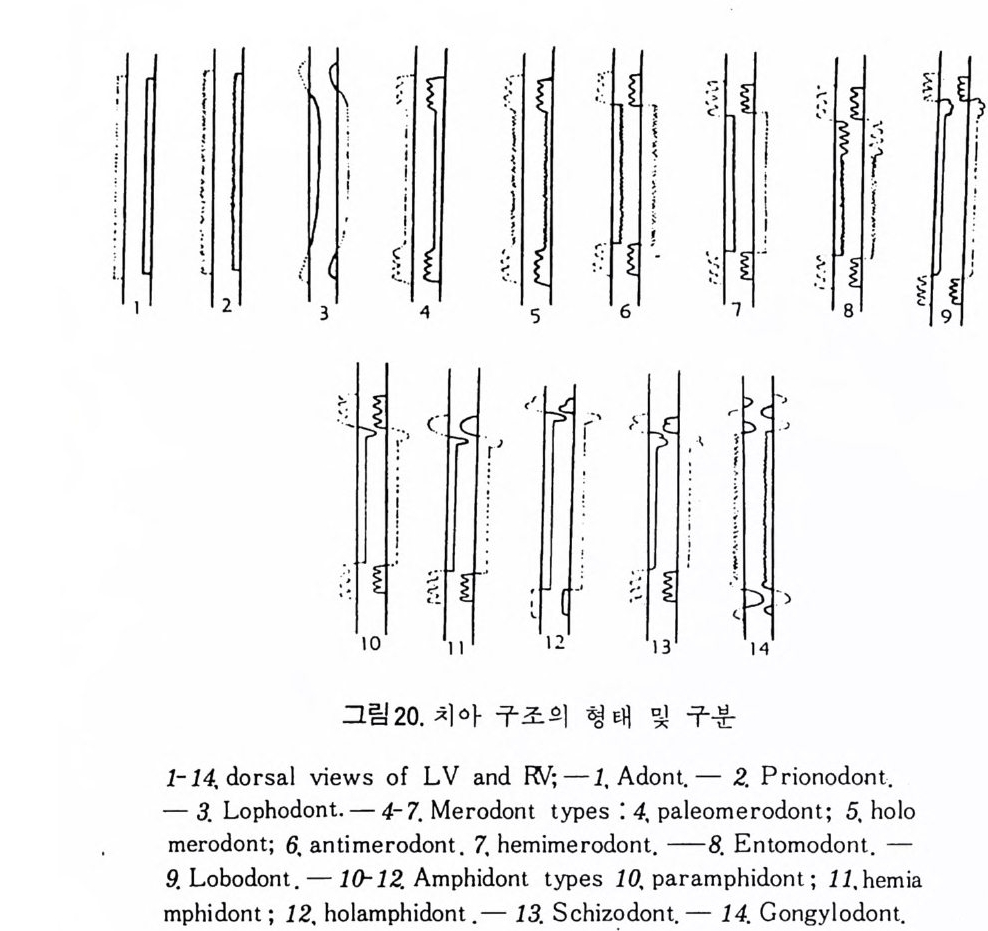 I .It...••'I.,.,l•., , ‘ . .'`., . ` 2 :..`,.\.. 3 r c.L•..'.•., ...,,1J.’ , f... ) . . 1 \ \;广口l4 r.,45 一 ltu '.,,,•.•’1 z'., _ . ;.` .. ..;... . .-J . 6. . tJ g_r ., /F.‘ ,. 홍홍 `· `V'`‘``:tC,` 、. -...'‘.v..,?•,.,.1、(f• . • • • 집—집 ,_i3 ` J 1..`/.v.'` ‘1 l1홍f j홍8 r` i;5f Y_ ,
I .It...••'I.,.,l•., , ‘ . .'`., . ` 2 :..`,.\.. 3 r c.L•..'.•., ...,,1J.’ , f... ) . . 1 \ \;广口l4 r.,45 一 ltu '.,,,•.•’1 z'., _ . ;.` .. ..;... . .-J . 6. . tJ g_r ., /F.‘ ,. 홍홍 `· `V'`‘``:tC,` 、. -...'‘.v..,?•,.,.1、(f• . • • • 집—집 ,_i3 ` J 1..`/.v.'` ‘1 l1홍f j홍8 r` i;5f Y_ ,
2·3·2 분류 가]형충의 분류 는 대부분 부 속 기관과 H 사강의 형 태 학 에 의 존한 다 . 분류는 개형충 개체에 대한 감정 (i den tifi ca ti on) 과는 달리 개체군 을 다루 는 것으로서, 많은 여러 특징들을 평가하여 선조부터 현 생 자 손 에 이 르 기 까지의 연관관계,즉 같은 유사성을 갖는 개체군을 묶어 하나의 계통표 를 만들어내는 작업이라 할 수 있다. 이러한 분류작업에 의해 명 개체군 을 구별해 보 았을 때 , 각 개체군을 구 성하고 있는 여러 개체는 그 개체군이 공통으로 갖는 특성을 갖 고 있어 다 론 개체군과 쉽게 구별하는데 큰 도움울 주는 형질을 언제나 가지고 있다 고는 한정지을 수 없다.
따라서 실제로 분류하는 데 있어서는 각의 모양이나 크기 • 치아구조·근 혼양식 (muscle scar pa tt e rn) •ornamen t a ti o n-번연대·벽공관 • 性的 2 型등의 특징을 동시에 고려해야 한다. 어떤 분류군을 구별하는 데 있어서 매우 중요한 형질이 다른 분류군에 는 전혀 적용될 수 없는 경우가 허다하다. 또한 어떤 시대의 분류군을 구별하는데 절대적인 역할을 하는 형질이 다 론 시대에는 무용지물인 경우가 많다. 한 예로 고생대 종은 주로 sulcati on • lobati on • ornamen t a ti on 에 의존되고 있으며 중생대와 산생대 종의 분류에 있어 매우 중요한 치아구조나 근혼 양 식 은 거의 무시되고 있는 실정이다. 이처럼 개형충을 분류하는 데 있어서 시대와 개체에 따른분류기준에 차 이가 있으므로,죽 절대적 기준이 없는 만큼 학자들에 따른 분류양상도 다 양하다. 간단히 몇몇 학자에 의한 분류양상을 보면 다음과 같다. Sar s (1 866 ) 에 의해 재형충은 크게 4 개의 아목으로 구분되었다. Podocop a, M~,o d oc op a, Cladocop a 및 Pla ty co p a 로서 주로 현생종에 의존하 여 구분하였으며 고생대 화석종은 전혀 포함되지 않고 있다. Ulr i c h ( 1894 ) 는 고생대 재형충에 대해 분류기준을 세웠으며 개형충목 (Orde r Os t ra c oda ) 하에 13 개의 科 (Le p cr ditii dae • Ento m i da e • Cy pr id inid a e… … Darwi 1111 lid a c) 로 구분하였 다. Jo nes (19 01) 에 의 해 실루리 아紀 (S i lur i an) 의 개 형 충이. Bassler 와 Kellett (1934 ) 에 의해 고생대종이 분류되였으며, 그 외에도 Swart z( 1936), Hen- 11 i 11 g smoe11(1953) 에 의한 고생대종의 분류가 있다. 그 외에도 많은 학자에 의해 분류되고 있었으나 현생종과 화석종을 모 두 취급한 분류로서는 V.Pokorn y(1 958) 가 처음이다. 그는 처음으로 치아 구조 • 근혼 (dup li c a tu r c • 二形) 동의 중요성 을 고려 하여 분류기 준을 만든 것 이다. 간단히 Pokorn y에 의한 분류룹 보면 개형충아강 (Subclass Ostr a coda) 은 Lcp e rdi tiida • Bcy r ic h ii da • Podocop ida • My o docop ida 등 4 개 목 (Order) 으 . 로 구분되고 있다. 현재 개 형 충亞綱 (Subclass Os t racoda) 은 Archeocop ida , Lep e rdit ico p - ida , My o docop ida ,Bey r ic h ic o p ida , Podoco pi da 동 5 개 의 목 (order) 으로 분류 되고 있으며 그 외에 분류학적 위치가 확실치 않은 1 개 의 목이 있다. 7 서형 충의 분류는 표 1 에 종합되어 있으며 아목 수준까지의 각 분류군에 대한·
분류학적 특칭은 다음과 같다. 표 1 개형충의 분류 1. ORDER ARCHEOCOPIDA Prin c ip a l fam i lie s : Bradoriid a e, Hip po nic h ariid a e. 2. OHDER LEPERDITICOPIDA Prin c ip a l fam i ly : Lep e rdit iida e. 3. OHDEH MYODOCOPIDA A. Suborder My o docop ina i. Sup e rfa m i ly Cy pr id i n a cea. Prin c ip a l fam i lies : Cy pr id i n i d a e , Cy li n d roleb eri- did a e, Cy pr id i n e llid a e, Sarsie l l ida e, Ruti de rmati da e. 8. Suborder . Halocy pr ifo r mes i. Sup e rfa m i ly Halocy pr id a cea. Fami ly Halocy p r id i d a e. ii. Sup e r fam i ly Thaumato c yp r id a cea. Fami ly Thaumato c yp r id i d a e. iii. Sup e rfa m i ly Ento m oconchacea. Fami ly Ento m oconchid a e . C. Suborder Cladocop ina Prin c ip a l fam i lie s : Polyc op ida e, Ento m ozoid a e. 4. OIWEH BEYRICHICOPIDA A. Suborder Hollin o morph a i. Sup e rfa m il y Hollin a cea. Prin c ip a l fam i lie s : Ct en onote l lid a e, Ct en oloc u lin i d a e, Hollin e llid a e, Hollin i d a e, Tetr a dellid a e, Qu adrij ug a to r id a e, Tvae r e n e ll id a e, Eup r im i tiida e. ii. Sup e rfa m i ly Euryc hil in a cea. Prin c ip a l fam i lie s : Euryc h il in i d a e , Oep iki i da e. iii. Sup e rfa m i ly Prim it iop s acea. Fami ly Prim i tiop s id i d a e. 8. S uborder Bey r ic h io m o rph a i. Sup e rfa m i ly Bey r ic h ia c e a. Fami ly Bey r ic h ii da e. C. Suborder Bi no dic o p ina i. Sup e rfa m i ly Drep a nellacea. Prin c ip a l fam i lie s : Aechmi ni d a e, Bolliid a e, Drep a nell ida e. 5. ORDER PODOCOPIDA A. Suborder Meta c op ina i. Sup e rfa m i ly Healdia c ea. Princ ip a l fam i lie s : Healdii da e, Bair d io c y pr id ida e, Pachy d omellid a e, S igilli id a e ( = S aip a ne ttida e) . ii. Sup e rf am i ly Thlip s uracea. Prin c ip a l fam i lie s : Thlip s urid a e, Bufi ni d a e, Ro- po lonellid a e, Qu asill i t ida e. B. Suborder Podocop ina i. Sup e rfa m i ly Cy the racea, Prin c ip a l fam i lies : Tric o rnin i d a e, Berounellid a e, By tho cy the rid a e, Paradoxosto m ati da e, Gloria n ellid a e, Cy the rurid a e, Lep toc - ythe rid a e, Cy the rid e id a e, Loxoconchid a e, Xesto l eberid i d a e, Krit hi d a e, Prog - onocyt he rid a e, Proto c y the rid a e, Trachy l eberid i d a e, Hemi cy the rid a e, Cy the r -ett ida e.
ii. Sup e rfa m i ly Bair d ia c ea. Prin c ip a l fam i lie s : Bair d ii da e, Beecherellid a e. iii. Sup e r fam i ly Darwi nu lacea. Fami ly Darwi nu lid a e. iv. Sup e rfa m i ly Cy pr id a cea. Prin c ip a l fam i lie s : Cy pr id i d a e Cy pr id o p s id a e, Macrocyp r id i d a e, Ponto c yp r id id a e, Candonid a e, Ilyo cyp r id i d a e, Cy pr id e id a e. C. S uborder Platy c op ina i Sup e r fam i ly Kloed. e nellacea, Prin c ip a l fam i lie s : Kloedenellid a e, Glyp top le u-urid a e. ii. Sup e r fam i ly Cy the rellacea. Prin c ip a l fam i lie s : Cy the rellid a e, Cavellin i d a e. 6. UNCERTAIN ORDER Suborder Ki rk by o cop ina Sup e rfa m i ly Ki rk by ac ea. Prin c ip a l fam i lie s : Ki rk by ida e, Amp h is s it ida e, A rcy z onid a e. (1) Archeocop ida 목(그림 21) 캄브리아기에서 오오도비스기에 걸쳐 산출되는 7 사형충으로서 모든 캄브 리아기 이후 개형충의 선조이리라 생각되어진다. 좌 • 우 양각의 크기는 거 의 같으며 길고 직 선 인 배 연 (dorsal margi n) 과 아주 볼록한 복연 (ventr a l mar gi n) 을 갖고 있다. 키틴질 혹은 약하게 석회화되었거냐 인산영화된 각 울 갖고 있으며 치 아구조는 없다. 몇 몇 속들은 복안 (comp o und ey e s) 이 리 라 생 각되는 t ubercles 를 갖고 있다. 穀의 표면은 매끄러 운 것, 주름진 것, rib s , sp in es 혹은 cres t s 를 갖는 것 둥이 있으며, inn er lamella 는 석회화 되어 있지 않다. (2) Lep e rdit ico p id a 목(그림 22) 오오도비스기 전기에서 데본기에 걸쳐 살던 7 사형충으로서 비교적 두껍게 석회화된 각을 갖고 있다. 좌 • 우 양각의 크기는 다르며 자유연(fr ee edg e ) m울a r따gi 라n) .큰 뚜 렷각한이 배작 각은 (d각or을sa l a겹n쳐g l es싸),고 볼 록있한다 .자 유연길(고f r e직e 선m인ar gi치 n아) 을연( 갖hi고 n g있 e 으며 각의 후반부가 더 불룩하고, 더 높다. Du p li ca t ure 는 발달되어 있지 않 고 안결절 (ey e tub ercles) 이 각의 전반부에 있다. 내 전 근혼군 (adduct o r muscle scar gr oup ) 은 작은 근혼들이 한데 모여 각의 중십 부보다 앞쪽에 넓은 연적을 차지하며 발달하고 있으며. 많은 경우는 근혼의 수효가 200 개 이상에 달하며 직 경 이 각의 높이의 1/3 정도를 차지한다. 전부근혼군(fr on tal muscle scar g rou p)은 내 전근혼군의 앞, 안결절 아래에 있으며 많은 수 의 근혼들로 이루어져 있다. 실루리아기와 데본기 종들의 전부근혼군은 V 자형으로 이루어져 있다.각의 표면은 대체로 매끈하다. 저서성.
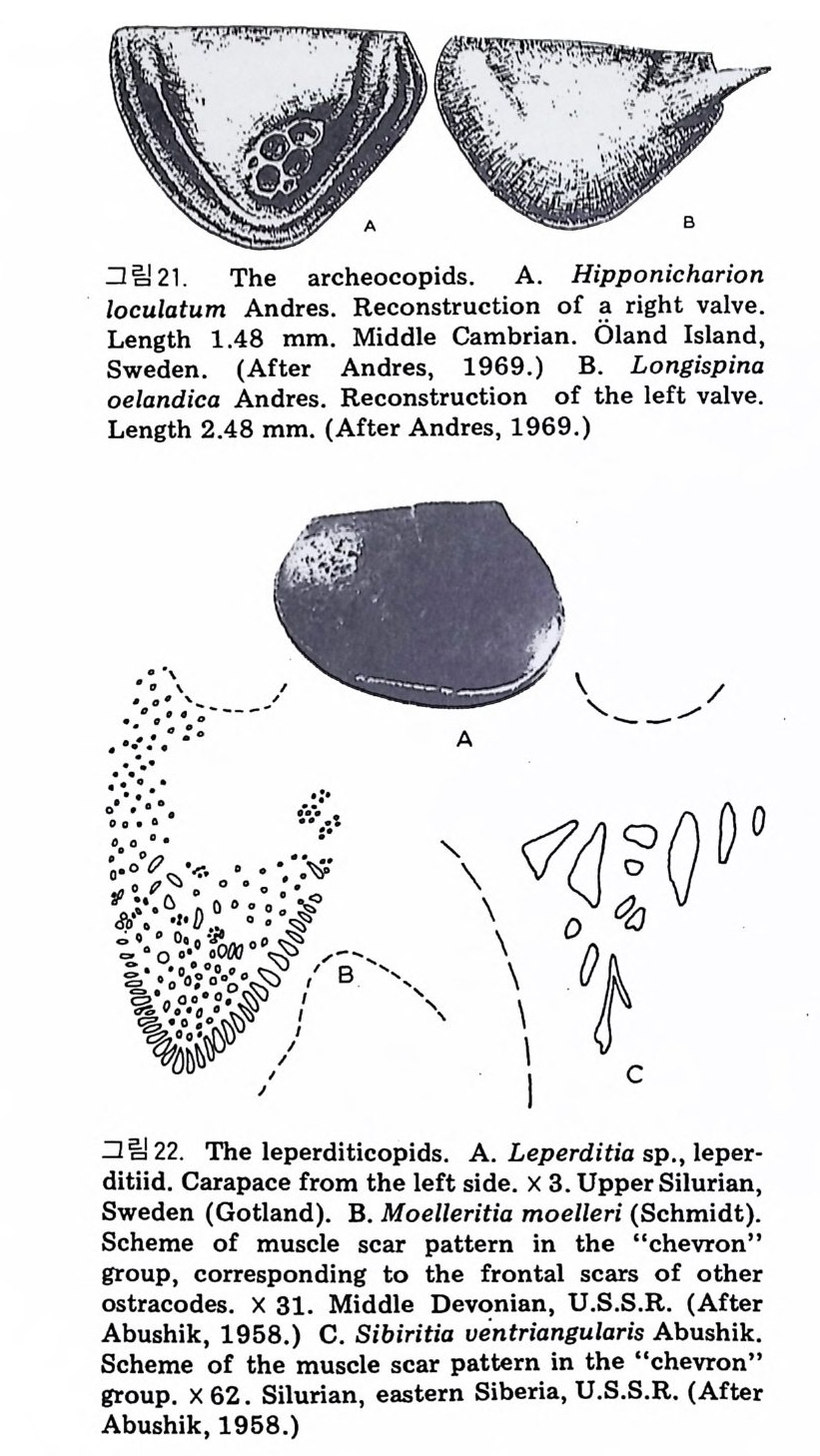 그 림 21. The archeocop ids . A. Hi ppo nic h ario n
그 림 21. The archeocop ids . A. Hi ppo nic h ario n
 틀싸? 4 % f 4- ,3· e
틀싸? 4 % f 4- ,3· e
(3) My o docop ida 목(그림 23) 오오도비스기에서 현세에 걸쳐 살고 있다.형태적인 다양성이 크기 때문 에 이 目 의 일반적인 특칭을 서술한다는 것은 불가능하다 . 대체로 각의 복 연은 볼록하며 in ner lamella 의 주변부는 석회화되어 있다• . 어 떤 종류들은
an t ennae 의 출입과 운동을 용이하게 하는 rostr a l in c i sure 를 갖고 있다. 현 생종에서는 second an t ennae 의 exo p od it e 가 여러개의 마디로 이루어져 있 어 유영생활에 적합하도록 되어 있다.어떤 廊들에서는 강한 성적 이형 현 상을 보인다.一돈 현생종들은 해양성인데, 그들 중 어떤 것들은 원양성이 며 다른 것들은 해저표면이냐 퇴적물내에서 서석한다. 대체로 각의 석회화 정도가 약하여 화석으로 보존되는 경우는 드물다. A. My o docop ina 아목 (그림 23) 각은 키 틴 질 혹은 석회 질로 되어 있으며 보통 rostr a l i nc i sure 와 rostr u m 을 갖고 있다. 십장과 복안을 갖고 있으며실루리아기부터 현세까지에 걷 쳐 살고 있다. B. Halocy pr if or mes 아목 백악기부터 현세에 걸쳐 살고 있으며 키틴질 혹은 약하게 석회화된 각 울 갖고 있다. 각의 배연부는 직선이며 표면은 대체로 매끈하다. 십장은 있 으나 눈은 없다. 내전근혼군은 좁고 간 근혼들이 몇 개의 열로 배열되어 이루어져 있다. 이들은 모두 해양에서 원양생활울 한다. C. Cladocop ina 아목 난형 내지 원형의 각을 갖고 있으며 짧은 배연과 치아연을 갖고 있다. Inner lamella 의 석회화된 부분은 비교적 넓으며, rostr a l i nc i sure 는 없 거나 아주 미약하다. 내전근혼군은 몇 개의 커다란 근흔들로 이루어진 둥근 형태물 보인다.눈과 심장은 없으며 한쌍의 th oracic le g s 만 갖고 있다. 해 양에서 서식한다. (4) Bey r ic h ic o p ida 목 (그림 24, 25, 26) 오오도비스기부터 트라이아스기에 걸쳐 산출되는 7 사형층으로서 대부분 이 고생대에 한정되어 산출되기 때문에 해양마고생물학적인 중요성은 제한 되어 있다. 각의 석회화는 찰 되어 있으며.다소 적선이고 간 cardin a I margi n 과 볼록한 extr a cardi na l mar gi n 을 갖고 있다. 많은 속둘이 현저한 loba ti on 과 sulca ti on 을 보인다. Inner lamellar 는 석회화되어 있지 않다. 몇 과에서만 근혼이 나타난다. Crum ina l d i mor ph i sm 과 antr a l d itn) r p h i sm 은 이 目 에 서 만 나타난다. 어 떤 것들은 sculpt ur al di mo rphi sm 이나 pr op o rti on al di mor p h i sm 을 보이며.다른 것들은 d i mor p h i sm 을 보이지 않는다. 해양성. A. Holl ino morph a 아목 (그립 24) Ant r al d i mor p h i sm 을 보여 주는 것 이 특칭 이 다. 그러 나 몇 몇 종류들은 dim orph is m 을 보여 주지 않는다•
 ,O.5 m m, l O.5 m m I
,O.5 m m, l O.5 m m I
B. Be y r ic h io m orph a 아목 (그림 25) Crumi na l d i mor p h i sm 을 보여 주는 것 이 독칭 이다. H i s ti um 은 없다 . 실루 리아기부터 석탄기 하부까지에 걸쳐서 산출된다.
 I 1mm l
I 1mm l
C. Bi no dic o p ina 아목 (그림 26) Med ian sulcus 의 앞과 뒤 의 배부에 node 를 갖는 속들이 여 기 에 포함된 다.대부분의 속들이 d i mo r p h i sm 울 보이지 않는다 . 오오도비스기부터 페름 기까지에 걸쳐 산출된다.
 I ,
I ,
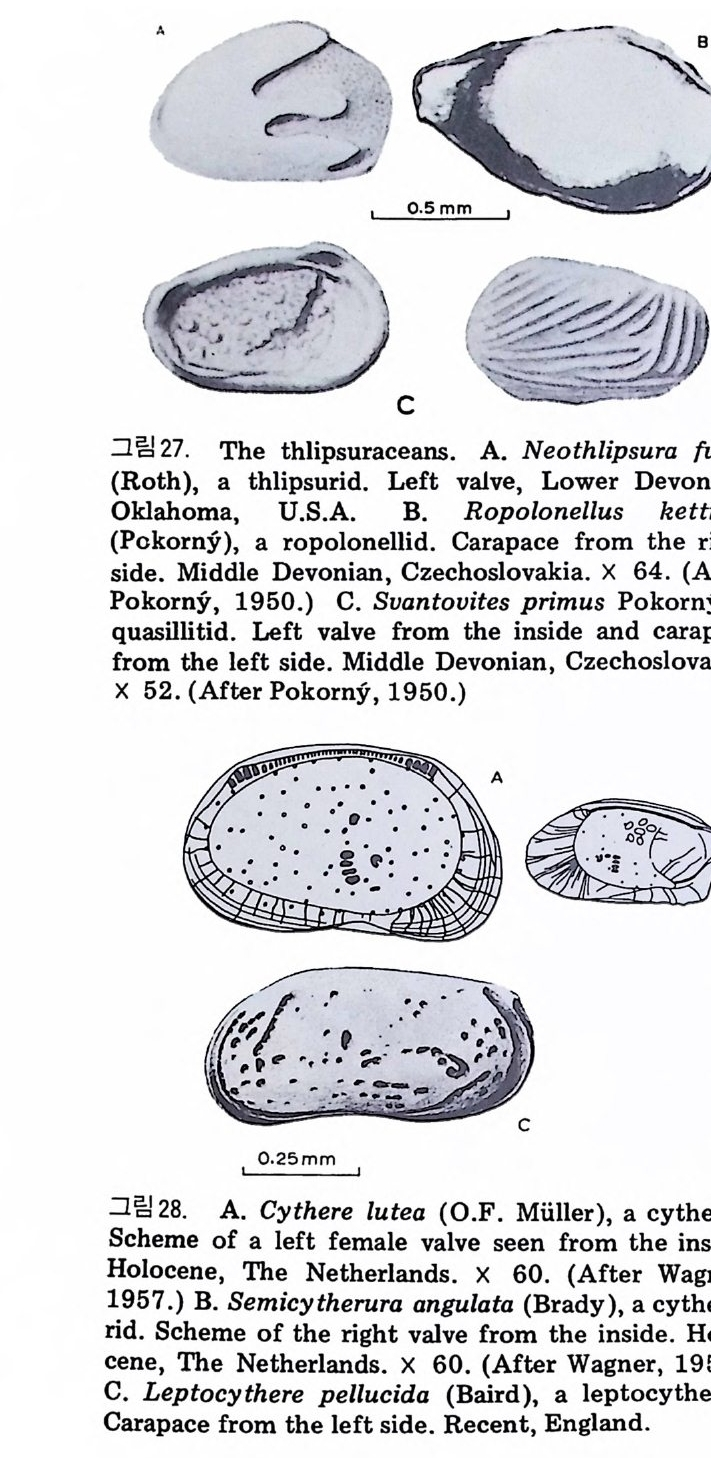 C
C
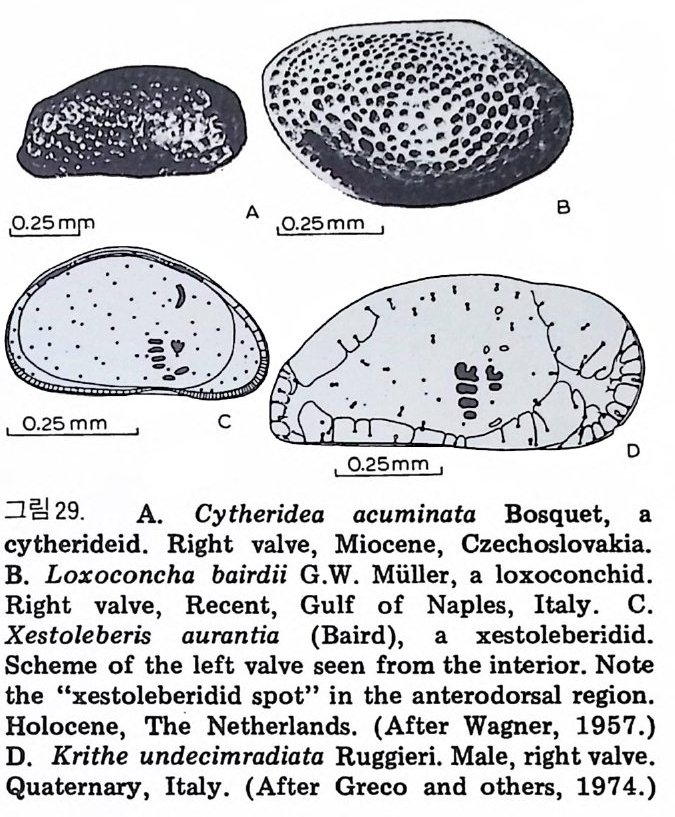 ‘ B
‘ B
(5) Podocop ida 목 (그림 27-32) 오오도미스기부터 현세에 걸쳐 산출되는 개형충으로서 석회화된 각을 갖 고 -: . 1. 으며 rostr al i nc i sure 는 없다. 내전근혼군은 원지 형 태 에 서 는 많은 수 의 근혼들로 이루어져 있지만 전화된 형태에서는 근혼의 수가 상당히 강소 되어 있다.심장은 없다.해양성 내지 당수성. A. Meta cop ina 아목 (그림 27) 많은 수의 근흔들이 뭉쳐서 원형의 내전근혼군을 형성하고 있다. B. Podocop ina 아목 (그림 28-32) 각의 중복연 (mi d- vent ra margi n) 이 안쪽으로 만곡되 어 있다. Dup lica t- ure 는 대체로 넓고, ves ti bule 은 있거나 없다. 내전근혼의 수는 감소되어 있다.해양성 내지 담수성.오오도비스기부터 현세까지에 걸쳐 산출되고 있다. C. Platy c op ina 아목 각은 강하게 석회화되어 있으며 좌 • 우 양각의 크기 는 서 로 다르다. Inner
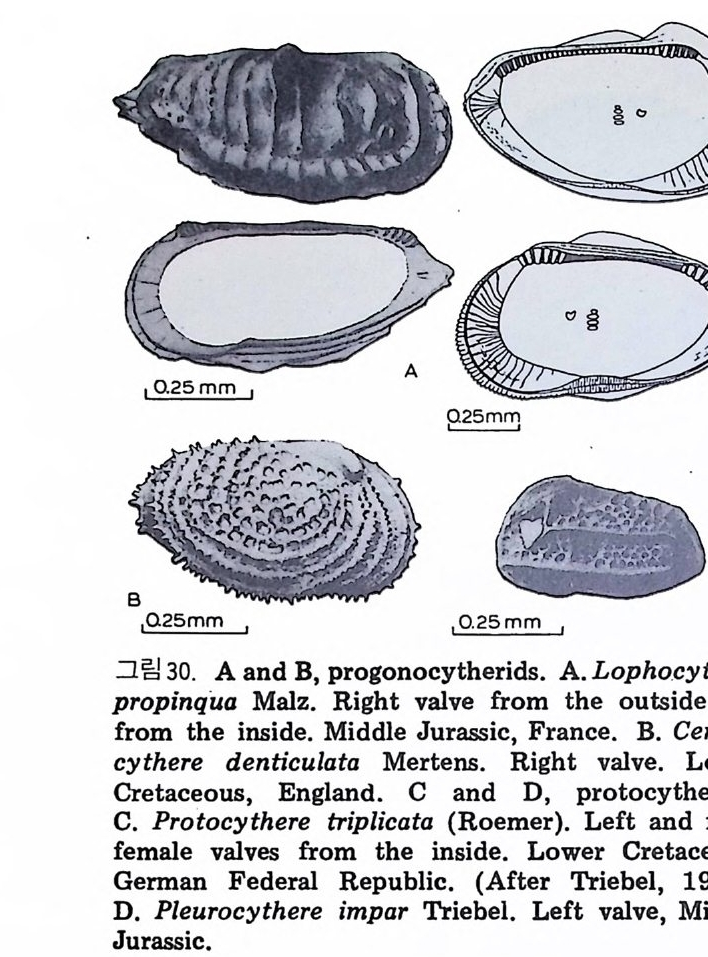 ' 25mm,
' 25mm,
(6) 미정 목 lamellar 는 석회화되어 있지 않거나 주변부만이 석회화되어 있다. Conta c t g roove 가 자유연(fr ee mar gin) 을 따라서 나타나며 치아연(hi n g e mar gin)을 따라 서도 혼히 나타난다. 성적이형 현상은 자성 (f emale) 각의 후부가 불룩하게 팽 창된 형 태로서 나타난다. A. Kir k by o cop ina 아목 (그림 33). K i rkby oco pi na 아목은 Be y r i ch i oco pi da 목에 포함시 켜 왔으나 최근에 어 떤 학자들은 이들을 Podoco pi da 목에 포함시키고 있다. 이들의 특칭은 다음 과 같다: 길고 직선인 배연, 약간 볼록한 복연, adducto r scar pit( Ki rk -bya n pit) 그리고 격자상 구조를 갖는 각의 표면. 오오도비스기 부터 트라 이아스기에 걸쳐 산층된다.
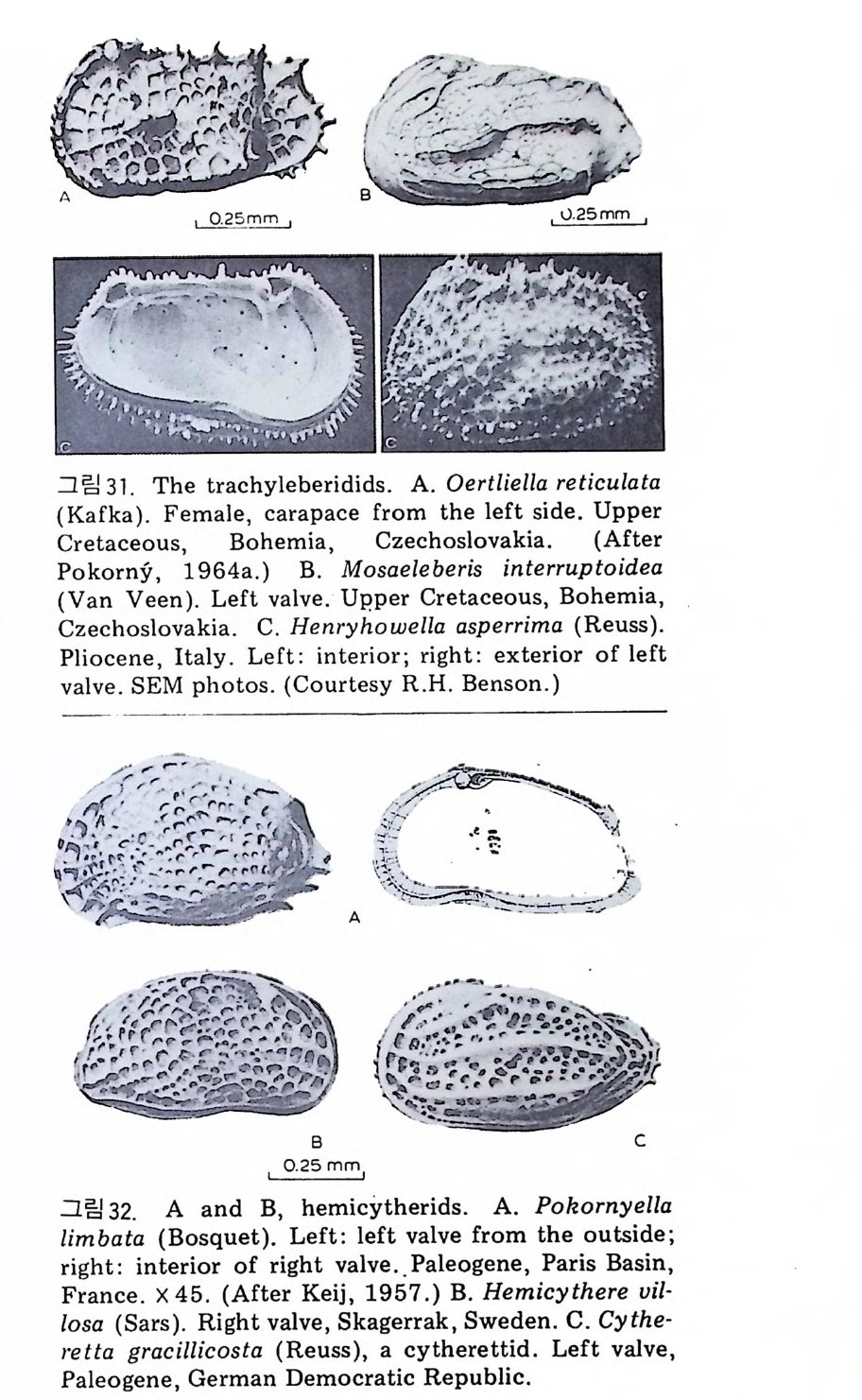 r
r
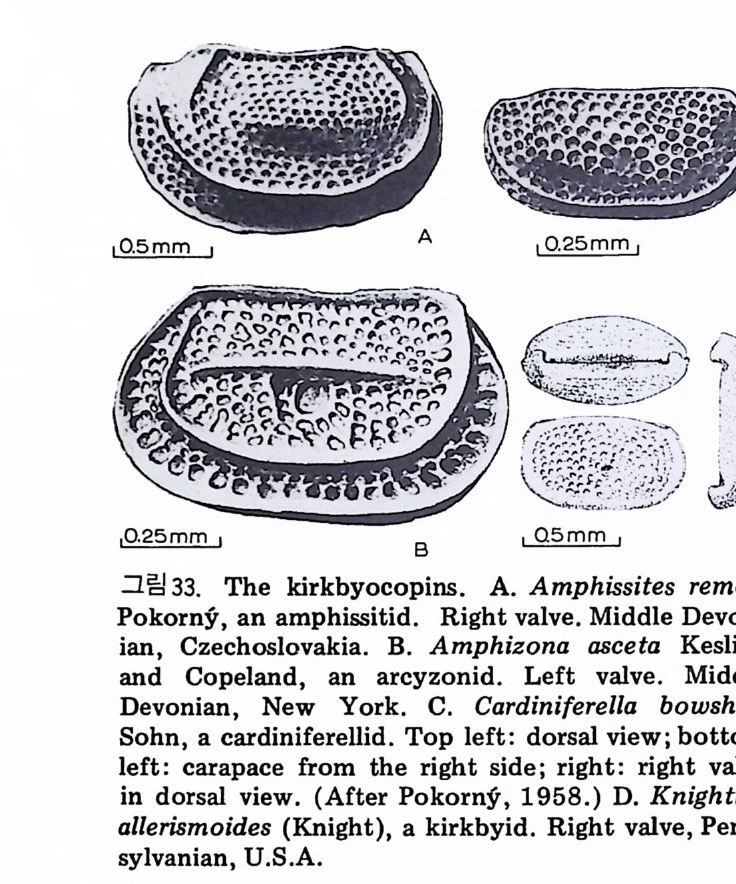 D
D
2·4 생태 및 고생태 개형충은 원래 해수로부터 시작되었으며 현재도 대부분의 종이 해수에 서식하며. 저서성이거나 부유성이다. 석탄기에는 담수에까지 서식범위를 넓혔으며, 오늘날은 적도에서 국지방에 이르기까지의 해수 • 기수 • 담수 • 습지 등의 다양한 후!-정에 적응하고 있다 . 해수의 경우 성해처 (hyp e raby ss al) 에 이 르 기까지. 심도로는 2,800m 정도의 깊이까지 분포한다 . 식물에 부착.동물
에 기생 혹은 공생하는 종류, 지하수내에 서식하는 종류에 이르기까지 다 양한 환경에 적응하고 있는 만큼 각 환경에 대한 이들의 형태적 특성 역시 다양하다. 그러면 우선 고생태를 복원하는데 있어 기본 자료가 되는 개형충과 환 경의 각 요소와의 적응 관계를 좀더 세부적으로 살펴보기로 한다. A) 영 양 (Nut ri ti on ) 개형총의 많은 종이 해서식물이나 작은 동물을 먹이로 취하고 있다. An - nelid s , Turbellaria n s, Nemerte ans 혹은 작은 갑각류 동이 대체로 먹이대상이 되고 있고, 간혹은 부패된 동식물의 일부조각을 먹는 종도 있으며, 퇴 적물을 무선택적으로 섭취하는 종( l im n iv orous) 도 있다. 일부 종은 口部가 찌르거나 빨기 좋도록 변이된 형태를 갖고 있어 식물즘을 흡수한다. 약 30 屬 정도논 공생응 하는 종류로서 다른 갑각류의 . 부속기관 혹은극피동물의 몸체에 붙어 살며, 그 외에도 어류의 아가미 등에 붕어 기생하는 종류도 있 다. 이 처 럼 개 형 충은 nut rit ion al s y s t em 면에 서 볼 때 filt e r -fe e d ing typ e 과 ckp o sit -fl'L'd ing t v p e 을 · 다 갖는 다양성을 보여주고 있다. B) 영 도 (Salin ity) 영도는 개형충의 분포양상에 가장 큰 역할을 하는 환경요소로서, 특히 개형충의 유기체의 생리작용에 결정적 영향을 미친다. 해역 가까이 서식 하는 종의 경 우 대 체 로 영 도에 대 한 적 응다양성 이 높은 (eury ha lin e ) 경 향을 보이는데, 이는 찾은 강수현상에 의한 영분도의 변화에 기인한다. 담수에 서식하는 종은 해수종과 분류학적으로 구별된다. 그러나 멸 종류는 두 수 역에 공존한다. 영분도와 개형충과의 관계에 대한 연구 중 Elofs o n(1941) 과 Wa g ner (1 957) 에 의하연, 영도 10% 。1 가지는 영분도 저하에 따라 감소하다가, 그 이하에서는 급격히 감소한다. 영도 3~10%, 범위내의 경 우 種數는 극소가 되고 반대로 개체수는 많아지는 경향이 있다. 일반적으 로 영도 3% 。 이 상이 되떤 담수종은 소실하는 경 향이 있다. Lago on s, estu - arie s , marshes 등의 기수환경에서는 광영성 種 내지는 기수성 種에 의 한 독특한 분포상응 봉 수 있다. 잉반적으로 유공충이나 다른 해수성 동 물군의 경우 영도가 감소함에 따라 접차 사라져버리는 데 반해, 개형충의 정우는 다론 종군의 모습으로 바뀌어 존재하므로 기수 환경 연구에 큰 여 향유 한다. 기수환경에서는 종수가 극히 작으며 심지어는 단일종만이 우 세하게 방당하는 경우도 봉 수 있다. 한 예로 Ostr a code Sea 라고 봉리 는 Azov Sea 의 정우름 보면 만일종이 lm 2 당 14,000~30,000 마리에 달한다.
C) 온도 (Te mp e ratu re) 개형충의 어떤 종류는 다양한 온도에 적응하는 광온도성 (euryt he rmal) 인 데 반하여. 어떤 것은 온도의 변이에 아주 예민하며 좁은 온도범위 내 에 서 만 서 식 하는 협 온도성 (ste n oth e rmal) 을 갖는다. 분류학적 으로 볼 때 처위도 지방의 천해에 사는 개형충류가 고위도 군보다 비교적 높은 다양 성을 보이며 분포한다. 한류성 _심해종을 ps y c hrop h il ic , 한류성 -천해종 을 cryo p h i li c , 난류성 종을 the rmop h il ic 라 칭 한다. D) 심도(D e pt h) 개형충과 심도만의 관계를 추정하기는 어렵다. 왜냐면 심도의 변화는 다 른 환경요소(퇴적양상 • 온도 • 빛 • 먹이 등)의 변이룽 동반하기 때문이다. 대체로 불안정하고 수심이 얕은 지역에서의 개형충의 다양도나 일도는 좀 더 수심이 깊고 안정된 외해성 환경에서보다 낮은 경향을 보인다. 심도 의 중가에 따른 가중압력 은 종의 수직 적 분포에 방해 자 (ba rrie r ) 역 할을 하 므로 특정된 소심도범위 (s t enoba t h i c) 내의 적응현상을 보여 야 할 것 이 다. 그러나 실제상으로는 많은 개형충류가 바교적 넓은 수심에 걸쳐 분포하는 광심도성 (eur y ba t h i c) 의 경 향을 갖는다. E) 저 질 (Substr ate ) 저질의 특성은 개형충류 분포 및 구성에 큰 역할을 한다. 개형충은 대부 분이 퇴적물표면 (e pifa una) 혹은 퇴적물속에 서식하는 種( i n fa una) 으로서. 서식하는 장소의 퇴적물 입자의 크기나 성분 모양 및 견고성에 따라 분포 를 달리한다. 일반적으로 입자가 작은 퇴적물내에 분포하는 개형충은 껍 질이 매끄럽고. 굵은 퇴적물의 경우는 표면이 거친 경향이 있다. Remane 과 Elo f son 에 의 해 s ilt와 cla y속에 서 식 하는 종을 endop e lose, sand 속에 서 식 하는 종을 enchp sa mmon, cla y와 s ilt표면에 서 석 하는 종을 epi pe lose, sand 표 면에 서식하는 종을 e pip sammon 으로 칭하기로 제안되었다. F) 빛 Oigh t) 빛이 개형충에 미치는 영향에 대해 많은 사람들이 사육실험을 통해 연 구해 왔다. 빛을 강하게 비춰 주면 개형충은 그림자 쪽으로 이동하며. 반 대로 캉캄한 곳에서는 빛 쪽으로 이동하는데, 이같은 추광성은 먹이나 이 성에 대한 반응보다 강한 것으로 보고되고 있다. 그외에도 감광시켰을 때 성장속도가 저하하는 현상, 幼期의 개형충류의 성장은 적색광 밑에서 느리고 청색 밑에서 빠르다는 사실도 밝혀져 있다. 개형충류는 이처럼 영분도, 온도, 심도. 처질, 광 동의 환경조건의 변
 . ¢= 1. 6 』 aua 1ew 야 1 ? l~!ueq 3 aw 6u!sea 』 :>u1
. ¢= 1. 6 』 aua 1ew 야 1 ? l~!ueq 3 aw 6u!sea 』 :>u1
화에 대하여 비교적 우수한 적응력을 갖는 편이나 한편 꽤 민감한 반응을 보인다고 할 수 있다. 그 외에도 p H 에 대한 개형충류의 반응성을 보면. 아 칙까지 연구결과가 거의 없는 형편이긴 하나. Klu g h ( 1927) 에 의하여 C yp r i a 와 C y or i no t us 같은 담수성종이 . pH 4 에서 2 일간. p HlO 에서 7 일간. pH 6 ~pH 8 사이에서는 건강히 생식됨이 밝혀졌다. 개형충을 이용하여 고환경을 연구·복원하는 데는여러 가지 방법이 있 다. 우선 개형충 자체에 대한 연구로 현생종에 대한 생활양식이나, 생물학 적 연구를 기초로 하여 환경요소에 직접 관련시켜 비교하는 연구법을 들 수 있다. 외각구조에 대한 기능 형태학적 해석을 시도항으로써 시간 • 공 간변이에 따른 형태학적 변이룰 비교하기도 한다(그림 34) . 생식의 방법, 性比, 분포양상 등 종의 집단 구조를 보거나 종다양도. 시 공간적인 분포 패턴의 변화 등 군집구조를 연구함으로써 고생태학적 정보 룰 얻을 수 있다. 이 밖에도 개형충과 함께 산출되는 화석군으로부터 간 접 적인 환경 정보를 얻거나 화석의 보존상태 , C/V 比 (carapa c e 수 /valv e 수) A/ J比 (adu lt수/j uven i le 수). 사후운반 등 자료로서 보완하기도 한다 (Sohn, 1962 ; Benson, 1974 , 1975 ; Hartm ann, 1973 등) . 2· 5 생지리 및 생충서 현생 혹은 신생대의 개형충은 전세계에 걸쳐서 해수, · 담수 및 기수로부 터 산출되고 있으며 산출되는 개형충 종군의 지리적인 유사성에 의해 명 개의 생지 리구 (bio g e o g ra ph ic realms) 로 구분된다. Celt ic, Boreal, Lusit an ia n Transatl a nti c, Carib b ean, Calif orn ia , Jap o n ic , Nova Zealand ic 등은 해수성 개 형충의 특성에 의해 구분한 것이며 Holarcti c, Neotr o p ica l, Et hi o p ian ,Aust- ralia n 등은 담수성 종에 의 해 구분된 것 이 다? 그 외 에 도 lnd o-pa cif ic, West Af rica n, Pata go nia n , Peruv ian , Panamania n 등이 있으나 개 형 충군의 특성 에 대해서는 잘 알려쳐 있지 않으며 동양권의 개형충에 대해선 거의 미지의 상태이다. 그림 35 는 태평양지역의 개형충에 의해 구분된 12 개의 牛地理區 륭 보여주고 있다. 개형충은 지리적 분포가 좁기 때문에 부유성 유공충과 같은 범세계적 분 *Ce lti c 은 영국 및 복해지역을, Boreal 은 노르웨이, 그린란드 및 복대서양지역 을 . Lu- s it an i an 은 지중해지역을. Transa t lan ti c 은 북미의 동부해역을, Car i bbe a n 은 Trin i d a d 와 멕시코만 및 풀로리다 납부 룬 일컫는다 .
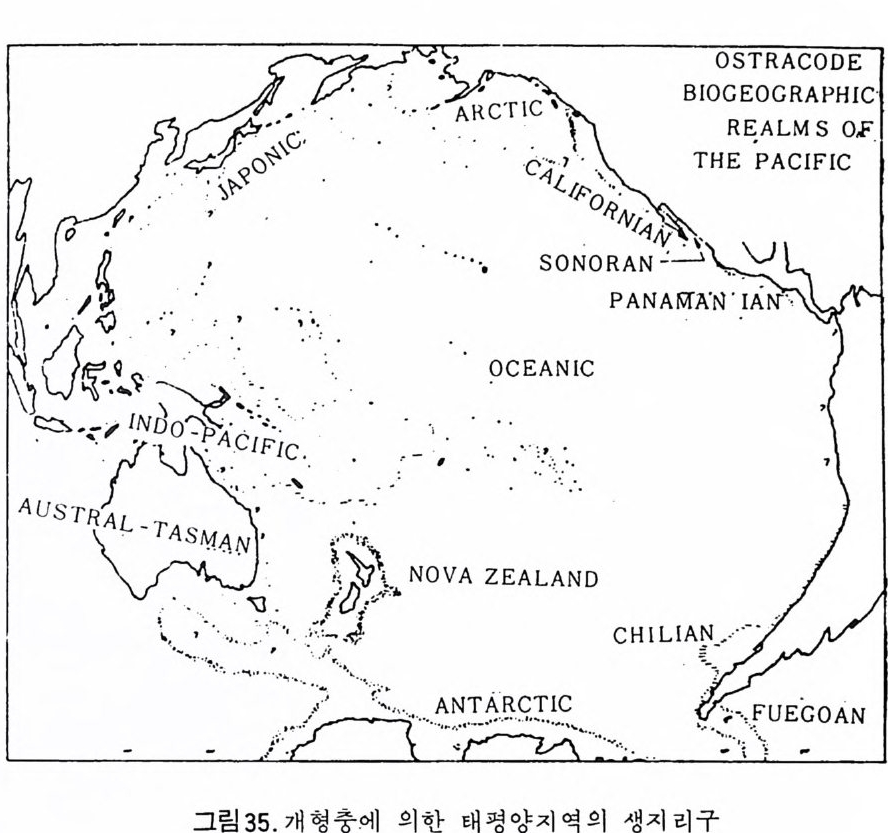
포를 나타내는 것과는 상당히 성질이 다르다. 저서성 개형충은 부유성 종과는 달리 쉽게 지리적 장벽을 넘어 분산되기 어려운 한계성을 가지고 있는 데도 불구하고 고지리를 밝히는 중요한 역 할울 한다. 특히 고심도나 고위도를 알아내는데 중요한 역할을 맡고 있 어 고지 리의 변화상을 추적 하는데 많은 도움을 준다. 그 외에도 pa leocir c u- lati on 및 고기후의 좋은 지시자이기도 하다. 개형충 중 특히 광범위한 지 역적 분포를 갖는 심해성 종에 대한 연구는 생지리에 많은도움을주며 특 히 고지리 복원에 큰 기여를 한다. 범세계적인 심해성 개형충의 연구는 Benson 과 Sy lv este r -Bradley ( 1971) 의 지중해의 제 3 계 연구나 Van den Bold(1974) 의 카리브해역의 연구가·대 표적이다. 이들 연구의 특성은 해저 분지의 발달과 개형충 군집의 적응,이
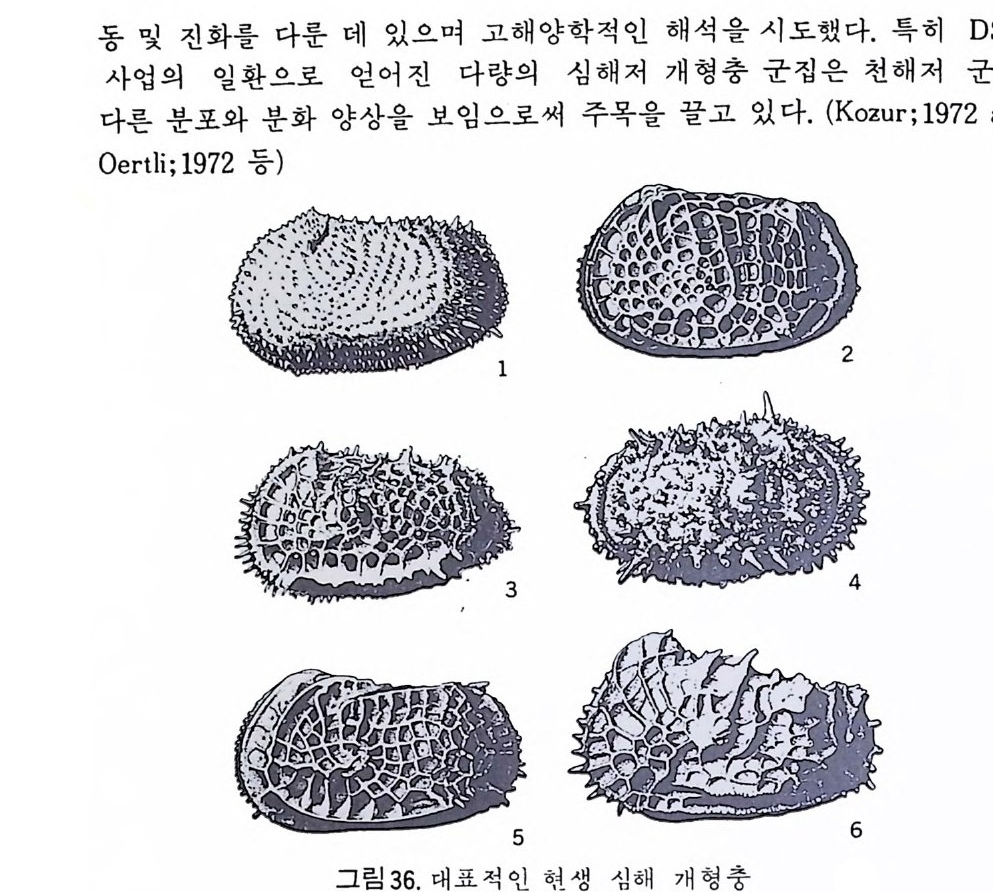 동 및 진화를 다룬 데 있으며 고해양학적인 해석을 시도했다. 특히 DSDP
동 및 진화를 다룬 데 있으며 고해양학적인 해석을 시도했다. 특히 DSDP
개형충 가운데서 부유성으로는 유일한 Haloc yp r idi na 亞 目은 껍질의 석회 화 정도가 약하여 화석으로 보존되지 않는다. 따라서 광역적 대비에는 그 다지 효과적이지 않으나 다량으로 산출되고 있는 화석 종 중에는 눈에 뛸 정도로 현저하게 형질이 시대와 함께 변하는 것이 많으므로 일정 지역내 의 층위를 밝히는 지시자로서 유효하다. 이러한 시준화석은 각 시대의 고지리 및 고기후의 영향을 강하계 받았을 뿐만 아니라 이것이 발견 되는 지역의 암상에도 지배받았다. 지질시대를 걸치는 동안 개형충 화 석의 형태변화를 보연 중생대 및 고생대에 걸쳐 가장 현저하며 신생대에 는 일반칙으로 그다지 두드러지지 못하다 .. 이유는 여러가지로 생각해 볼
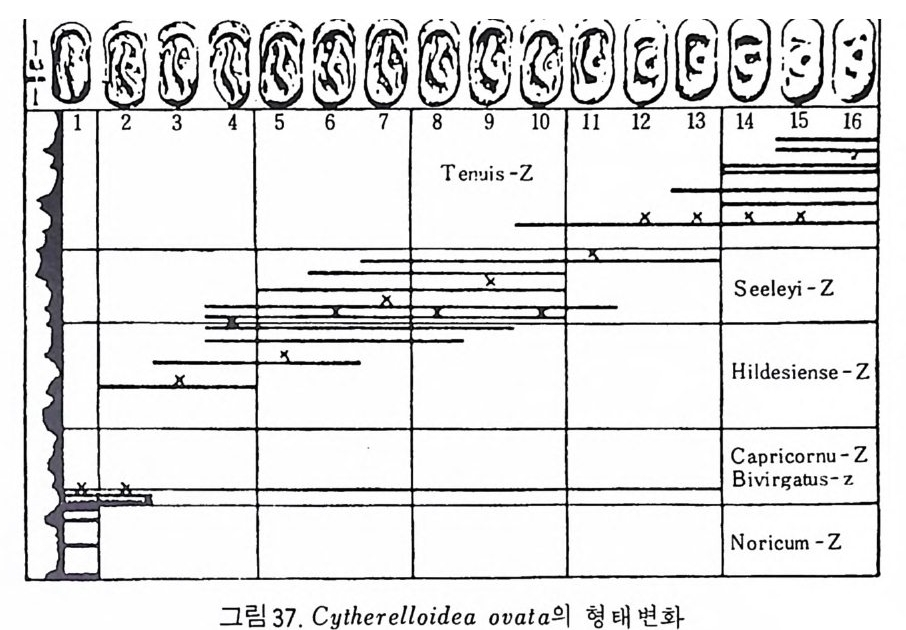 通麟懿還關驛釋誌)劇
通麟懿還關驛釋誌)劇
수 있겠으나 우선 개형충 자체의 형태변화 혹은 진화속도가 신생대에 비 해 중 • 고생대에 빨랐다고 볼 수 있으며 한편은 지질시대 구분 자체에 있 어 시간간격의 차이름 들 수가 있다. 영국의 오도비스기에서 제 3 기에 걷친 각 지질시대의 개형충의 시준화석이 고지리에 대한 설명 및 범위와 함께 완성되었으며, 암상에 지배되기 때문에 생긴 약접을 보충하기 위해 종의 진화계통에 기초하여 분대 (zona ti on) 를 하였다.그러나 분포범위 전체에 결 쳐 종의 출현 및 소멸에 대한 정확한 층위학적 자료의 부족 및 이주의 식 별문제, 생태적 변화 및 진화적 변화의 구별 등 문제점을 많이 가지고 있 어 활발히 연구되고 있지는 않다. 개형충은 비해성층의 분대에 가장 유효한 시준화석 역할을 한다. 영 국의 Purbeck-Wealden 층을 C yp r i dea 속에 의 해 T it hon i an 에 서 Barrem ian 까지 10 개로 분대하였으며 이는 이전에 같은 C yp r i dea 속에 의해 구분된 독일 의 Purbeck-W ea. Iden 층과 대비되어졌다. 북 • 남미 양 대륙의 경우, 양대륙 의 분리문제와 관련 연구되고 있다• 담수성 개형충의 알이 건조시 25 년 도 견단다고 알려질 정도로 건조에 강하며 바람이나 기타 다른 요인에 의 해 널리 분산되어 현생종 중 세계적인 분포를 갖는 것도 있다. 그림 37 은 백악기 Cy the relloid e a ova /a의 형태변화를 보이는 그림으로 시대에 따른 접 진적 변화를 볼 수 있다. 주요 개형충 속들의 지질시대 별 분포는 표 2 와 같다.
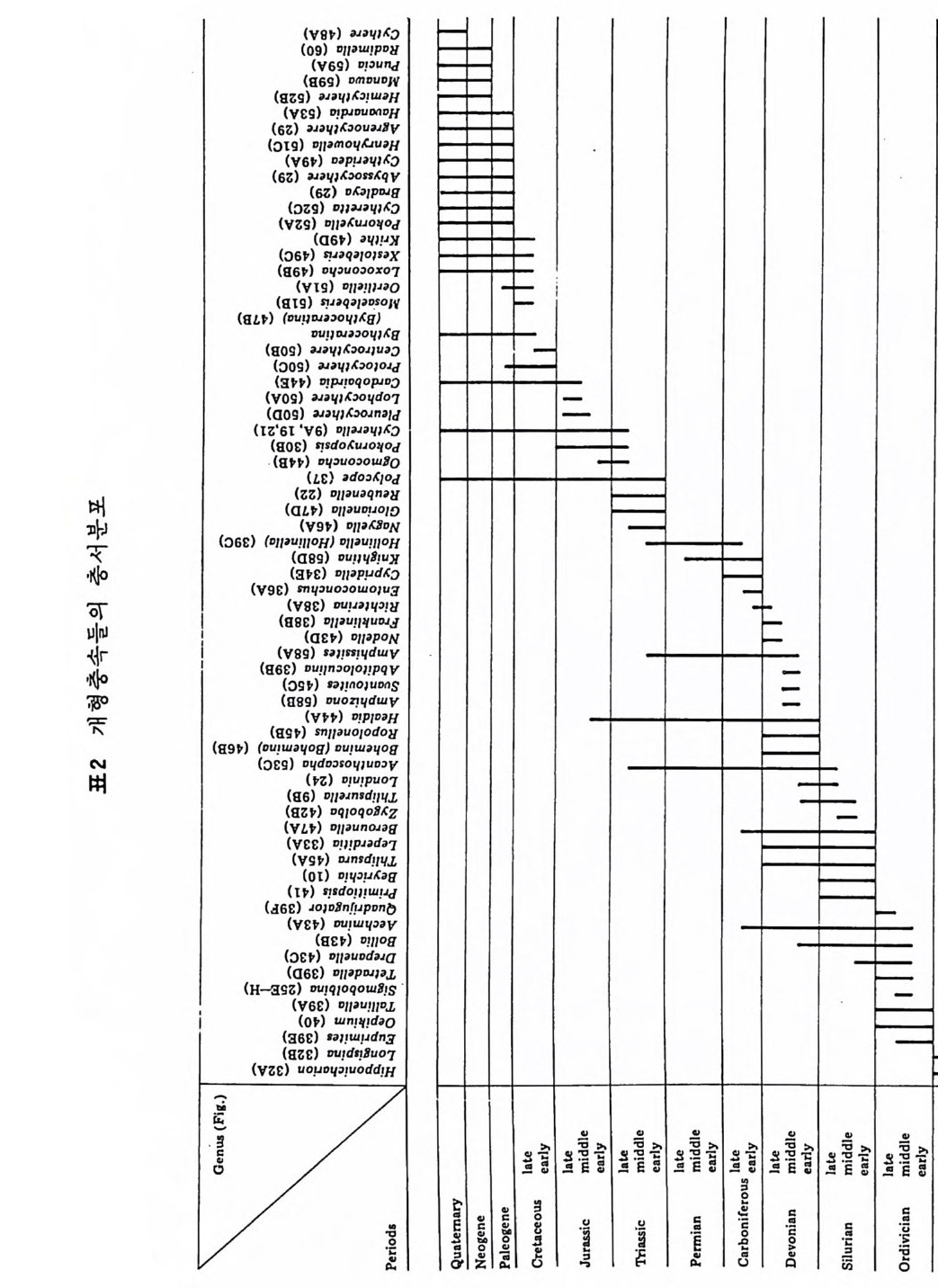 (V8V) ,,aw O
(V8V) ,,aw O
참고문헌 Coreye ll, H.N., 1963. Bi bl iog r aph ic Index and Classif ica ti on of th e Mesozoic Os- tra coda. Univ e rsit y of Dayt on Press, Ohio , 1175 pp. (2 vols.) (Refe r ence wo- rk on Mesozoic Ostr a coda describ ed pr io r to 1962. Includes classif ica ti on outl ine ,di ag n o ses of sup r asp e cif ic tax a, selecte d fau na) list s , bib l io g r aph y. c ata l: >- gue of spe c ie s , ge nera and fam i lies .] G rekoff , N., 1960. Ap e rcu sur Les Os tra codes {os sile s . T echnip Paris , 97 pp. [In tro ducti on to th e stu dy of fos sil ostr a code~.) Hartm a nn, G., 1966, 1967, 1968, an in pr in t . Ostr acod a.ln : Dr. H. G. Bronns Kia - ssen und Ordnung e n des Ti er reic h s. Band 5 : Arth rop o da, 1 . Abt. Crusta cea. 2 Buch, I V. Teil . Akademi sc he Verlag s ge sellschaft Geest urrl Portig . Leip z tg . 568 pp. (3 volumes) . [B asic neonto l og ica l tre ati se on ostr a codes. ) Harttn a nn,G .and. H . S., P u ri, 1974. S uin m ary of neonto log ica l andp a leonto l og ica l clas-
sif ica ti on of Ostr a coda. Mi tt. Hamburg. Zool. Mus. Inst. , 70:7-73.[A na tt em p t at clas si f ica ti on of ostr a code ge nera and hig h er ta xa based both on the ir s- oft pa rts and carap a ce fea tu res. Only th ose fam il ie s havin g Recent rep r ese nta ti ve s are inc luded. ] Howe, H. V., 1971 . Prelim in a ry lis t of new ostr a cod tax a 1961- 19 71 . M'elan ge s, Mus Geosi . , N o. 3 : 28 pp. Howe , H . V., 1972. Correcti on s _a nd add ition s to pr elim i na ry lis t of new ostr a co d tax a 1961-1971 . Melang e s, Mus. G eosci. , No. 5. [Pu blic a ti on comp le menti ng Ostr a cod Taxonomy by the same auth o r. ] Howe. H. V., and Lanreocic h , L., 1958. Intr o ducti on to the S tu dy of Creta c eous Ostr a coda. Louis i a n a St ate Univ e rsit y Press , Bato n Roug e , La., 536 pp. (Des- crip tion and fig u r es of all spe _ci e s of Creta c eous ostr a codes pu blis h ed up to the end of 1956 as well as the releven bib l io g r ap h y. ] Moore. R.C. and Pit ra t, C.W . (Ed ito rs).1961. T reati se on Inverte brate Paleon- log y. Part Q. Arth rop o da, 3, Crusta cea. O str a coda. Geolog ica l Socie t y o( Am- eric a and Univ e rsit y of Kansas Press, Lawrence, Kansas , 4 42 pp. (Imp o rta n t refe r ence book on Ostr a coda. ] Oertli . H.J. , 1963. Mesozoic Ostr a cod Faunas of France. Brill , Leid e n, 57 pp. [A - survey of th e str a ti gr aph ic a l seq u ence of Mesozoic ostr a codes of France. ] Pokorhy , V., 1965. Prin c ip le s of Zoolog ica l Mi cr op a laeonto log y. 2. Perga man Pr- ess, London. 465 pp. (Translati on of German edit ion . ) Van Morkhoven, F.P.C.M., 1962, 1963. Post- P alaeozoic Ostr a coda. Their Morph - olog y. Taxonomy and EcoMmi c Use. Elsevi er , Amste r dam, Volume I , General , 204 pp. Volume II , Generic Descrip tion s. 478 pp. Alexander, C.L .1 933. Shell str u ctu re of the ostr a cod ge n us Cy the rop ter on,and fos sil spe cie s fro m th e Creta ceous of Texas. J.P aleonto l ., 7 : 181-21 4.Andres, D., 1969. Ostr a coden aus de m mi ttl e r en Kambriu m von Oland . Le th a ia , 2 : 165-18(). Bassl e r, R. S. and Kellet , B ., 1934 . Bib l io g r aph ic ind e x of Paleozoic Ostr a coda. Geo!. Soc. Am., Sp e c. Pap. , No. 1 : 50() pp. Be rson, R. H . 1959. Ecolog y of Recent oslraco() d es of the Todos Santo s Bay re gion ,Baja Califo r nia , Mexic o : Univ Kans as, P aleont. cont r. , Arth r op o da, Art. 1, p. 1 -8 0 (Law renc e) Benson, R.H., 1971. A new Ce no zoic de ep - se a ge nus, Aby s so. cyt h ere(Crusta c ea : Ostr a coda : Trachy le b erid ida e) wi th descrip tion s of five new spe cie s . Smi - ths on. Contr i b . Paleobio l ., 7 : 25p p. Benson, R. H, 1972. The Br 떠 Le ya pro blem, wi th descrip tion s of tw o new ps yc hr osph eric ostr a code spe cie s , Ag r enocy th ere and Poseid o nami cu s( Ostr a coda : Crusta c ea) . Smi ths on. Contr i b . Paleobio l ., 12 : 138p p. Benson, R.H., 1974. The role of ornament a ti on in the des ig n and fun c ti o n of the ost r a code. Ceosci. Man,6-47-51 : Benson, R.H., 1975a. The orgi n of tlie ps y c hrosph ere as rec ord e d in cha ng e s of deep -s ea ostr a code assemblag e s. Leth a ia . 8 : 69 一 83. Benson, R.H., 1975b. Morph olog ic sta b il it y in Os tr a coda. Bull. Am. Paleonto l., 65 : 13-46. Benson, R.H. and Sy lv este r -Bradly, P.C., 1971 .D e ep - se a ostr a codes and the tr - ansfo rmati on of ocean to sea in the Teth y s . In : H.J. Oe rtl i ( Edit or } . Pafe o eco l-Ogie d Ostr a codes, Pau 1970. B ull. C entr e Rech. Pau SNPA. 5 (sup pl) 63 一 91 . Blumenste n g e l, H., 1965. Zu Taxio n omi eu nd Bio s tr a ti gr aph ie verkie s elte r Ostr a c- oden aus dem Thurin g e r Oberdevon. Ereib e rge Forschung s h., C 183 : 127p p. Brady, G.S., 1968 A monog r aph of tlie Recent Briti sh ostr a coda. Trans. Linn . Soc. Lond.,26 : 353-495. Brady, G. S., 1880. Rep o rt on the Ostr a coda dredg e d by H. M . S. Challeng e r du-rin g the ye ars 1873-1876. Rep. Sei. Res. Voy a g e H. M . S. Challeng e r. Zool. 1 . qt, 3 : 184p p. Cannon, H.G., 1931 . On tl1e anato m y of a marin e Ostr a cod, Cy pr id i n a ( Do loria ) lev is Skog s berg, Di sc overy Rep. Combr 社g e. 9 : 435 -4 82·. Cannon, H.G., 1933. On the fee di ng mechanis m of certa in marin e ostr a cods, Trans. R. Soc. Edin b ., 5 7 : 739-764. Cannon, H. G., 1940. On the anato m y of Gig a nto c y pr i s m'u l leri Di sc overy Rep. Cambrid ge . 19 : 185 -2 44. El0fs o n, 0., 1941 . Zur Kenntn i s der marin e n Ostr a coden Schwedens mi t besond- erer Berucksig u ng des Skage rraks. Zool. Bid r . U p ps ., 19 : 215-534. Gramm, M.N., 1972. Marin e Trassic ostr a codes fro m South Prim orye . On the evoluti on of the ac:H u cto rs of cavellin i d s . In : Proc. Int Geol. C on gr ., 23rd. Cze· choslov akia 1968, pp. 135 -1 48. Grundel, J., 1962. zur Phy lo g e neti k und Taxio n omi e der Ento m ozoid a e (Ostr a co-
da) unt e r Ausschluss der Boucii na e. Geolog ie, 11 : 1184 -1203. Ha11ai, T.1957. a. St ud ie s on the Ostr a coda fro m Ja pa n, 1. Subf am i ly Le pt oc y 1h, rin a e 11. Subfa m . Univ . T oky o , Jo ur., sec . 2. v. 10, pt3, p,4 31-468. pl. 7-10. Ha11ai. T. 19:: >7 . b. S111rli( 'S 0.' ; L' Q, \' .•• : ,:~ ·: 딴 (rnm .Ja p a n. TI Subfa m i ly Pc cto c y th- c ri11 a c 11 . .,;u b fam . Univ . Toky o . Jou r ., scc. 2. v. 10. p: , 1, p. .l ( :.· • : n . pl. 11. Hanai. T. 1957. c. St ud ie s on the Ostr a coda fro m Ja p a n. ill Subfa m i ly C_. • .. ,ri - 11al . G.W. Muller and Cy 1 herop l eri1 1 ae. 11. subfa m . Univ . Toky o . Jo ur. sec. 2, I'< • .. IJ,pt. l, p. 11-36 . pl. 2-4. Ha na i, T. 1959, St ud ie s on the Ostr a coda (rom Jap a n : 4-fa m i ly Cy th erid c id a e Sars, 1 925. Un iv . Toky o . Jo ir ., sea . 2, v. 11 pt. 3, p. 291-3 08 .. pl. 16-18 , 41ext -figs Hardin g ,J.P .,1955. T he evo lu t ion of ter r es I ria l habi1 s in an os tr a cod .I n:Sy m p - osiu m on Orga nic Evoluti on Bull Nat. H i st Inst. S ci. I ndia , 7 : 104-106. Ha rd in g , J. P., 1 965. C rus ta c ean cu1 i cl e wi th ref e r e nce lo the o.,;I racode carapa - cc. Pubbl. S tn. Zoo!. Nap o li, 33 (.,;u p pl. ) : 9 -31. Ha r di ng , J. P., 1966. My o d o cop a n os lraco ds fro m the gills and nosl rils of fi.,;h e.,;. In : H. Barne. ,; (E dil o r) . Some Cont em p o rary St ud ie ; in Marin e Scie n ce. Allen and Un w i n, Lo n don, pp. 369 -374. Ha rl ma n, G., 1963. Zur Phy lo g e n ie und Sy s te m alik der Os lr acoden. Z. Zoo/. S y s t. Ev ol uti on s(orsch., 1 : I -154. Ha rlm a nn , G., 1973. Zurn ge ge n warlig e n .St an d de r Er for schung de r Ostr a coden inl e rsti tiel l e r Sy s le m e . Ann. Sp e leol.,28 : 417-426. . Ha zel, J .E ., 1970. Allanli c con ti ne n ta l she lf and slop e or the Unit ed St at e s -Ostr - . acod e zoog e og r ap h y in 1he So u· 1h e rm Nova Scolia n and North e rn Vi rg i ni a n rau n al pr ov in c e s. U. S .G e o! . Surv., P rof. Pap. , 529 -E. Ha 1. e l, J.E ., 1971 . Os lr ac od e bio s tr a ti gr ap h y of the Yorklown Formati on (up pe r M- ioc e ne and low e r Plio c e ne ) of Vi rg i ni a and Norlh Carolin a . U. S. Geo!. Surv., Prof. Pap. , 704 : 13p p. Ho rn ib r oo k, N. de 8., 1963. The New Zealand ostr a code ra mi ly Puncii da e. Mi er -op a leont o log y, 9 : 318 -320. Howe , H.V., 1971 . Ec ol og y or Ame r ic a n tor ose Cy th crid c id a e. In : H.J. Ocrtl i (E - dit or ) , Pal'eo 'e c olog ie d' O st racodes, Pau 1970. Bull. Centr e Rech., P au-SNPA. 5 (su p pl. ) : 349 一 359. Hulin g s , N.C. and Puri, H .S .., 1965. TIH) ecolog y or shallow wate r ostr a codesor the wes t coast or Florid a . Publt. Si n. Zoo!. Nap o li, 3 3 (sup pl) : 308-344. Jaa nusson, V., 1957. Mi dd le Ordovic i a n oslraco des or centr a l and south e rn Swe- den. Bull. G eo!. Ins t . Up ps ., 37 : 173 -442. Keij, A.J. , 1957. Eocene and Olig o cene Oslracoda or Belgi um . Mem. Inst. R. Sci. Nat. Belg. , No. 136 : 210p p. Keslin g , R. V., 1969. Cop u lalory adap l ati on s in os lr acodcs.3. Adap ta ti on s in s om e• exti nc l ostr a codes . C ont rib . M11s. Paleonto l . Univ . Mi ch ., 22 : 273 -312. Keslin g , R.V&Rog c rs, K.J. 1957. Si1 .c , Lobalio n , relate sl ruc lu rcs, and ornamc'-
111 a1 io11 in sorr ic UL'~• r ic h iid os t rac o dc. ~ : Ja ur. Paleonl. vol. 31 . No. 5. p. 997-1009 pl, 127 -1 30. (Tulsa) . KL•,;]in g , R.V. anJ Huss e y, R.C., 1953. A ne w fam i ly and ge n us of os lr acodaf ro m I 恥 Ordovic i a n Bi ll 's Creek sha le of Mi ch ig a n. Cont r ib . Mus. Paleonlol .. Univ . Mi ch ., 1 1 : 77 一 95. Kil c ny i,T .I., 1971, Some basic qu es ti on s in 1hc pa laeoec ol og y of ostr a cods In : 1-I. J. Oc r1 li ( Edit or) , Paleoecolog ie d'Ost racodes, Pau 1970. Bull. Cent re Rech. PAU-S NPA. 5(sup pl.) : 31-44. K ing m a.J .T h. 1948. Cont rib u t i on lo the knou ied g e of th e Youn{I ' Caenozoic O st r acoda fro m th e Malay a n Reg ion . Univ . Ut re cht, Holland . Kornic k er, L.S ., 1966. Morph olog y, onto ge ny and int r a spe cif ic varia t i on of Sp ina - cop ia, a new ge nus of my o doco pid ostr a cod (Sarsie l lid a e) . Smi ths on. Conlrib . Zool.,8 : 50p p. Kornic k er, L. S. and Solm, I. G., 1974. Evoluti on of th e Ento m oconchacea. In : Int. Sy m p . Evoluti on Post Paleozoic Ostr a coda, Hamburg 1974(abstr a ct) . Kozur, H., 1972a, Ei ni g e Bemerkunge n zur Sy s te m ati k der Os tr a coden und Besc- hreib u ng 1ie u er Platy c op ida aus der Tria s Ung a rns un der Slowakei. Geol. P- al~ont o l.M itt.ln nsbruck,2 : 1-27. Kozur, H., 19726. Di e Bedeutu n g tr ia s sis h er Ostr a coden fur str a ti gr ap h is c he und pa laookolog isc lie Unte r suchung e n. Mi tt. Ges. Geo!. Bergb austu d.,21 : 623-660. Kozur, H., 1974. Di e Bedeutu n g der Bradoriid a Vorlauf e r der po st- k ambris c her Ostr a coden. Z. Geol. W iss . Berlin , 2 : 823-830. Kozur, H. ai1d Mosti er ,H., 1972. Di e Bcdeutu n g der Mi k rofo s sili e n fur str a ti gr a- ph is c l1e pa laookolog isc he und pa llio ge og r ap h is c he Unt er suchung e n in der Trias Mi tf. Ges Geo!. Bergb ausfu d .,21 : 341-360. Kris l an-Tollmann, E., 1973. Zur Ausbil d ung des Schlie s smuskelf el des bei tr ia d i- schen Cy the re IIid e a (Ost racoda) . Neues Ja hrb. Geol. Paliio n lol. M onats h ., 1973 : 351-373. Krommelbein , K., 1 955. Arte n der Gatt un g e n Condracy pr is und Pachy d omella im Mi t1e l-Devon. Senckenberg. Leth aea,36 : 295-310. Lang e r, W., 1973. Zur Ult ra str u ktu r . Mi kr omorph olog ie und Taph onomi e des Ost _ racoda-Carap a x. Palaeonto g r aph ic a , Abt. A.144 : 1-54 . Mar1i1 1 sso11, A., 1965. The Si lu ro-Devonia n ge nus Nodib e y r ic h ia and fau nally ass- ocia 1 ed Kloedenii ne s. Geol. F aren. Slockh. Forh.,87 : 109-U 언 Mar1i ns s011, A., 1967. The successio n and correlati on of os t racod` 、 fau nas in the Sil u ria n of Got la nd . Geo!. F aren. St oc kh. Forh., 8 9 : 350-386. McKenzie , K.G., 1967a. Saip a nellid a e : A new fam i ly of po docop id Ostr a c oda. Cr- uslaceana,13 : 103-113. McKenzie , K.G., 19676. The dist r i b u 1io 1 1 of Caenozoic marin e Ostr a coda frn m the Gulf of Mexic o lo Auslralasia . In : C.G. Adams and D.V. Ag e r (Edil o rs) . rl sp e els o/ Teth y a n Bi og e og r aph y. Sy s l. Assoc. Publ. L and., 7 : 219 -238.
McKe n zie , K.G., 1969. Dis c u s sio1 1 . l11:J .W .Ne al e (Edit or ) , The Taxonomy , Morph ol-
og y and Ec ol og y of Rece nt Ostr ac oda. Oliv e r and Boy d , Edin b urgh , pp. 48 4 -4 85. Menz ie s ,R . J. , G eor ge , R.Y. and Rowe . G .T., 1973. Aby s sal Env ir on ment and Ecol- ogy of the World Oce an s. J oh n W ile y , New York, N.Y., 488p p. Muller , G .W., 1894. Di e Ostr a coden des Golfe s von Neap e l und der ang r enzend en Mee re sabschnit te. Faun und Flora.d e s Golfe s von Neap e l, 21, Monog r . 8. Frie d lande r, Berlin , 404p p. Ne al e, J.W . (Ed it or ) , 1969. The Taxonomy , Morph olog y and Ecolog y of Recent Ostr a coda. O liv e r and Bo yd , Edin b urgh , 553p p. Oe r tli , H .J. (E dit o r ) . 1971 . Colloq, u iu m on the Paleoe co log y of Ostr ac odes. P au 1970. Bull. C entr a Rech. Pau-SNPA 5 (sup pl, ) : 953p p. Oer tli , H.J. , 1 97 2 . Ju ras sic Os tr a code s of DSDP Le g 11 (si t es 100 and 105) -P rcl i m i na r y acc ou nL ln : C. D. Hollis t e r , J. L Ewi ng and oth e rs, Init ial· R ep o rts of the De ep Se a Drill in g Proje c t.1 1 .Gove rn me n t Prin t i ng Of fice , Washti ng ton . D. C., p p. 645- 657 . Po k o rn y , V., 195 4 . Zaklady zoolog ick e mi kr op a leonto l og ie. Akademi a, Pragu e, 65l pp . Pok o rn y, V., 195 7 . The ph y lo m o r ph o g e n y of the hin g e in Podocop ida (Ostr a coda. PoC k orur ns y ta ,c Ve .a, )1 9a5n8d . Gitrsu nbdez a u rg in e g d0e11 r tzhoe o lotagx iicohn eonm y M. Ai k cr otap aU li inoinv .t o lG oego ile. ,, 1 119.5 D7 t s: c 2h2. pV p.e r!. PoW ko isrn s .y, , B V e. ,r l 1 i9n 5, 49 5. 3Hpi p n.g e an d fre e margi n str u ctu r es of som e S ilu ria n ostr a cods PoA koc tran y U , Vn :i, v1. 9 G64e6 o. l T. ,h 1e9 5p9h y: l3o g2 e1 n- e3t4i c1 . lin e s of Cy th ereis marssoni Bonnema. 1941 (O s tr a co d a. C rus ta c ea) in the Up pe r Creta c eous of Bohemi a. Cze ch oslovakia Acta Uni v. C arol. Ge o l., 1964 : 255-274. Purl, H . S. (E dit or ) . 1965. Os tr a cods as ecologi ca l and pa laeoecolog ica l ind ic a to rs Pubbl. S t n. Zool. N ap o li, (su p p l. ) : 612p p. Purl, H .S., B o n a du c e, G. and Ge r va s io , A. M., 1969. Di st r i b u ti on of Os tr a coda in the Me d it er ran ea n . In : J. W. Neale (E d itor ) , The Taxonomy , Morph olog y and Ecolog y of Rece n t Ostr a coda. O liv e r and Boy d, Ed inb urgh , pp. 356-411. Ra b ie n , A ., 1 954. Zur Ta xio n omi e und Chronolog ie der oberdevonis c hen Ostr a cod- en. Abh. Hess. L ande sa mt . Bodenf or sch., 9 : 268p p. Rug gie r i,G ., 1971 . Os tr a c o da as cold clim ate ind ic a to r s in the Ita l ia n Qu ate r nary. In : H. J. Oe r tl i ( Ed it or ) . Paleoecolog ie d' Ostr a codes, Pau 197 0. Bull. Cent re Rech. Pau-S NPA . 5 (su pp l , ) : 285 -2 93. Sars, G .O., 1922-1928. An Account of the Crusta c ea of Norway, IX. Ostr a co- da. Be r ge n, 277p p. (11 9 pls . ) Schne id e r, G . F., 1960. Fauna ostr a kod niz h netr i a s ovy k hotl o zhenii Prik a spu skoi pniaz dm ney ni n Kosa tzi. a kInh s: taG n e. oTlorg. iK ao minpe lfet ke sg na .z oYnuozshnnoosi t Gyeuo g!a . ESk. sSp . Se d.iRt s. iTi , uArkkamde. n Nias ut ak n iS .ZSa.-S.R., 5 : 287-303. Sclnmi ko v, E. I., 1969. Novoe semeis t v o rakus l~
coccos p here 를 이루고 있으며 죽은 뒤에는 이들이 분리되어 해저 퇴적물내 에 분산되어 퇴적된다 . 이들 타원형 또는 원형 조각들은 크기가 1~15µm 정 도의 coccol it hs 이 다.
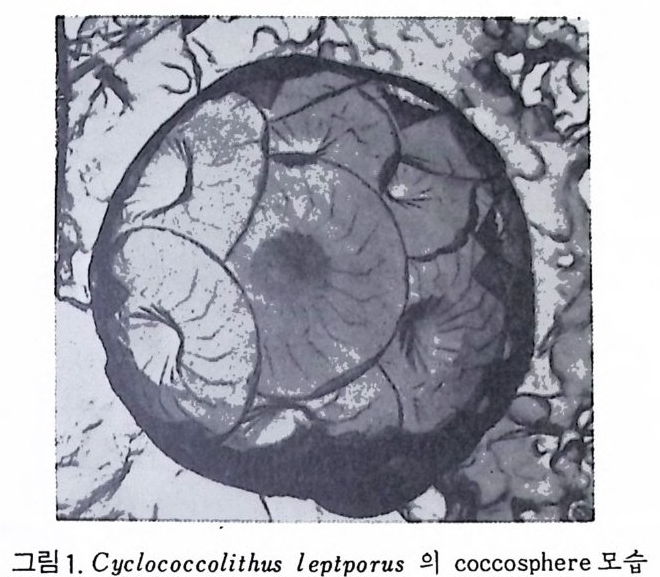 그림 1. Cy cl ococcoli thu s l ept po rus 의 coccosph ere 모습
그림 1. Cy cl ococcoli thu s l ept po rus 의 coccosph ere 모습
같은 해조류의 일종으로 형태가 전혀 다르지만 전동적으로 난노플랑크 돈에 포함시 켜 온 그룹들 중에 서 aste r olit hs 혹은 di scoas t ers 라고 불리는 것들이 있다. As t erol it hs 는 높은 종 다양도와 열대 -아열대 해역에 풍부한 생물충서학적 가치 때문에 많이 연구되고 있다. 1836 년 최초로 coccol it hs 와 &scoas t ers 를 연구한 사람은 독일의 Ehrenberg 로서 R i.ig en 의 백악충에서 발견된 이들을 무기 기원으로 생각했다. Huxle y는 1858 년 대서양의 심해처 연니로부터 이들을 확인하고 역시 무 기물로 보았다. 1861 년에 와서 야 비로소 Wall i ch 와 Sorb y에 의해 이들이 생물이라는 것이 밝혀졌고 1891 년 유명한 H.M.S.Challe ng er 호의 탐사보고 서 에 서 Murra y와 Renard 는 난노플랑크론을 포함한 석 동물성 플랑크론을 주 목하였다. 20 세기에 들어와서 Lohmann 을 위시한 생물학자 및 고생물학자 둘의 연구 결과를 집 약한 nan no--pa leonto l og y “가 발간되 었 다. 1930 년 대 에 노르웨이 학자들의 연구에 의해 생물학적인 윤곽이 드러나게 되었으며 1954 년 Bramle tt e 와 R i edel 은 중생대와 신생대 지층의 범세계적 지충대비
에 이들의 이용 가능성을 제시하였다. 이들은 특히 제 3 기층을 대상으로 하여 열대-아열대-온대 기후 지역 사이의 정교한 생물 충서학적 분대와 대바에 크게 성공하였다. 난노화석은 약 1. 000 배의 광학현미경을 동해 관찰된다. 해저 퇴적물이 나 부유물은 화학약품으로 찰 세척해 내고 원심분리기에 의해 난노호杉4 크 기의 입자들만으로 분리하여 간단한 스미어술라이드 (smear sl i de) 를 만들 어 현미경 관찰이 실시된다. 현미경의 편광 장치는 화석의 광학적 특성을 밝혀 종을 감별하는 데 큰 도움을 준다. 투사 및 주사 현미경 (TEM 및 SEM) 은 광학현미경 하에서 식별되지 않는 초미 형태를 감별하는데 최근 많 이 이용되고 있다. 앞으로 다루게 될 형태와 분류의 장에서는 주로 전자 현미경 아래에서의 특칭들만을 논하게 될 것이다. 또한 생충서학적으로 중요한 화석군에 관한 소개는 광학현미경 사전을 첨부항으로써 실제 응용 에 도움이 되도록 하였다. 3 • 2 생활사 대부분의 현 생 해조류는 ha pt onema 라고 부르는 편모기구룽 가지 고 있으 며 해조류인 Ha pt ophy ceae 강의 특징인 유가질 인편으로 표연이 둘러 싸여 있다. 유기질 인편에 석회질판을 철석시키는 능력으로 해조류는 Hapt o - p hy ceae 의 나머지 그룹과 기타 다론 해조와 구분된다 (그링 2 참조). 식물성 플랑크톤의 한 종류이며 바다의 먹이 사슬의 일원으로서의 중요 성에 비해 이들에 대한 배양을 통한 연구는 아직 미미하다 .• 그 중 Emi lia - nia huxley i(Lo hmann)Hay and Mohler 는 비 교적 배 양하기 가 용이한데 이들은 원양과 근해의 도처에 생식하며 열대, 아열대 몇 고위도 지역에도 넓게 분 포한다. 이러한 접둘이 이 종의 생물학적 정체물 실험실에서 밝힐 수 있 는 이접으로 작용하였다. E. huxle yi의 세포는 그림 3 과 같이 핵, 두 개 의 영록체, mi toc hondria , die- tyos om 또는 Golbi appar atu s 그리 고 ''bod y -X 라고 불리는 공포체 동을 포 함하는 원형질로 구성되어 있으며 원형질은 두 개의 이중막으로 싸여져 있다. 엽록체는 광합성 작용을 하는 엽록소를 가지 며 광도에 따라 그의 모 양이나 위치를 번화시킬 수 있다. M it ochondr i a 는 에너지를 생산하는 산화효소를 함유하고 있으며 diet y o- some 혹은 Golbi appa ra t us 는 아마도 세포벽 물질을 생산하는 데 참여하 는 기능을 가졌으리라고 믿어전다.
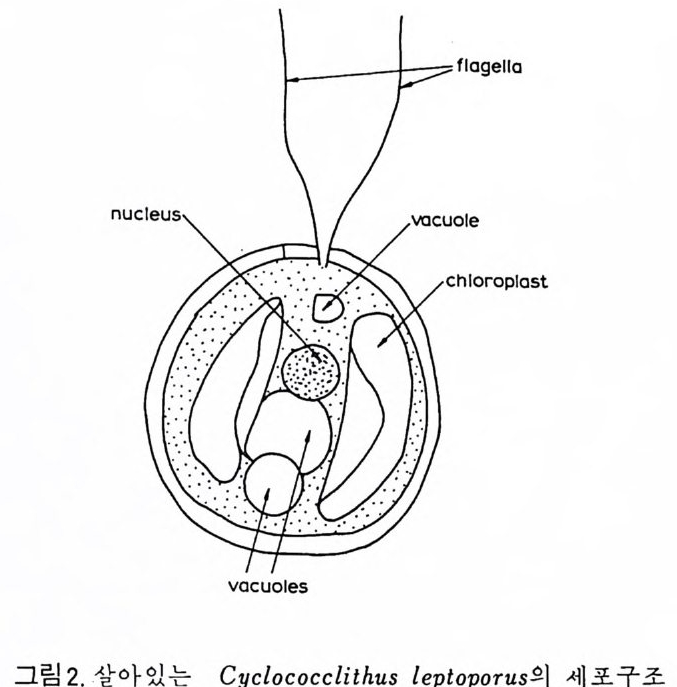 fla g el la
fla g el la
E.hu. rle y i 의 여러 성장단계에 속하는 개체의 단면을 조사해 보면 coccl- it hs 의 생성온 원형 질 안에서 일어난다는 사실을 알 수 있다. 어 떤 종류의 coccol it ho ph ores 는 2 기 의 생활주기를 가진다 . 편모체를 갖는 운동기와 비운동기가 교대로 나타난다. 비운동기에는 세포가 coccoli ths 물 질을 생산하지만 운동기에는 cocco lit hs 가 없거나 있더라도 비운동기에 생 산된 것과는 다론 형태의 것이다. 때문에 가끔 이들이 다론 종으로 감별 된 예도 있다. Coccolith u s pel ag icu s C-Na llic h ) Sc hi ller 를 연구한 Paasche (1968) 에 의 하면 성적 융합이 일어나면, 비운동기에는 핵이 염색체를 완전한 쌍으로 갖는 dip - loid s t a g e 를, 운동기 에 는 전 자의 경 우의 반수에 해 당되 는 쌍을 짓 지 않은 염색체를 갖는 hap lo id s t a g e 를, 죽 두 형태의 생활주기를 갖는 사실을 밝혀 냈다 . . 따라서 cocco lit ho p hores 는 유공충이나 아아도 방산층과 같이 이 형의 생활 주기를 갖는 것 같다.
 § 노
§ 노
해조류는 대부분 광독립 영양체이지만C.p ela gi cus 의 운동기에서 이물질 인 박테리아나 더 작은 해조를 섭취하는 것이 관찰되었다. 또한 일부 종류 에서는 종속 영양체적인 경향이 있는 것으로 믿어진다. 실험실에서 E.huxle yi는 매 19 시간마다 세포 분열이 일어났으며 자연상 태에서는 대서양의 하루에 1. 2 배에서 흑 해의 하루에 4.8 배까지 번화가 십 하다. 분열과 coccol it hs 의 생산은 수온이나 영양 영류의 양과 뚜렷이 관 계가 있음이 알려졌다. Cocco lit hs 를 이루는 CaCO J는 보통 방해석으로 결정되지만 아라거나이 트인 경우도 있다 . E.huxle .는 배양기에서 세 가지의 동질이상의 광물로 구성되나 화석의 경우 다론 성분들은 대단히 불안정하기 때문에 방해석만
이 확인된다. Coccoli ths 의 결 정 작용은 대 체로 두 가지 형 태 가 있는데 hete rococcolit hs 와 holococcol it hs 가 그것 이다. 전자는 방해 석 이 서로 다른 형 태와 크기를 가진 이소절정으로 이루어전 것이며 후자는 대체로 능연체나 육각 프리즘 의 형태로 광물의 원래의 정벽을 유지하는 경우를 말한다. 앞서 설명한 대로 · 대부분의 coccol it hs 는 비운동기에 만들어지는데 운동 기에는 세포내의 에너지가 편모운동에 소모되어 절정작용을 위한 체내의 화학작용아 어렵기 때문인 것으로 보인다. Coc co l it hs 의 기능에 대해서는 여러가지 설이 있다. 그 중 가장 중요한 것은 광선음 모으는 작용과 이오片곤 대조적으로 잉여 광선음 반사 차단시 키 는 작용 이 다 . coccoli t h s 의 렌 즈상 형 태 (convexo-concave) 로 보 아 집 광 의 작용이 유리한 생각 같으며 특히 두광대의 하부에서는 유리한 생태학적 장미로 추측 된다. D i scoa s t e rs 는 풍라이오世에 절멸한 그룹이 기 때문에 그들의 결정작용음 추적하기가 용이하지 않다. 이들의 형태는 coccol it hs 에 미하여 대단히 복 삼하지만 미소구조는 오히려 단순하다. 이들은 방해석을 관상으로 정충 해서 외각의 형태롤 만든다. 3 • 3 형태와 분류 석 회 질 난노풍랑크톤은 c~cco lit h 나 그와 유사한 관음 갖는 다양한 형 태 상의 특칭응 갖는그룹들로구성된다. Di scoas t ers 와갑이 형태적으로 완 전히 다론 그룹들도 coccol it hs 와 같은 크기를 가지 기 때문에 함께 연구되 는 것이 보통이다. 형태의 다양성 때문에 이들에 대한 분류 체계는 아직 붕완전하다. 이들이 동물과 식물의 특칭음 함께 가지는 관계로 분류상의 낭점이 더욱 가중되었으나 현재 고생물학자들은 대체로 이들음 식뭄로 보 고 석뭉명명 규약에 따라 기재를 하고 있다. 식뭉학자들은 해조류룽 황금색 식물 황금색조류강 (Coccol it ho p h y ceae) 의 Helio l it h ae 目에 포함시키고 있다. 평의 상 이들은 다음과 같이 셰 종류로 냐누어전다 (표 1 참조). (a) 해조류와 분명한 관련이 있는 무리 (b) Nanno lit hs 에 속하지만 해조류는 아닌 무리 (c) Incerta e sedis 속둘 각종에 대한 기재를 비롯한 분류학적 자료는 Farin a cci (1969 년 이후)의
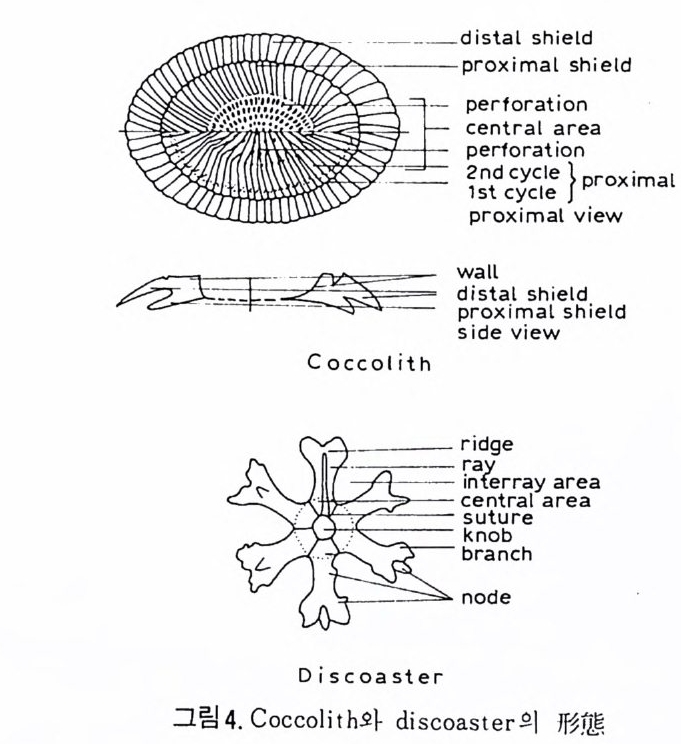 --_ ____ _ d ist a l shie l d
--_ ____ _ d ist a l shie l d
카타로그와 Loeblic h & Tapp a n (19n6 년 이후) 의 The annot a t e d ind ex and b i bl i o g ra p h y'’에 상세히 설명되어 있다. 다음에 소개하는 주요 그룹들은 전자현미경 아래서의 특칭과 사전음 함 께 첨부하였다. 표 4 와 표 1 과 같이 우선 크게 세 그룹으로 냐누고 다음 주요科의 수준에서의 표정을 요약하였다 . A. Coccol it ho p ores 와 이에 유관한 nannolit hs (1) Arkhang e lskie ll i d s-중 생 대 에 속하는 과로서 두 개 혹은 네 개 의 cy cl e 울 가지는 타원형이 특정이다(그림 5). (2)Coccol it h i ds- 중생대와 신생대에 산출되며 다양한 형태를 보이는 이 중의 판구조가 특칭이다 (그림 6,7,8,9). 13) Podorhabd id s-중 생 대에 산출되 고 이 중의 판구조를 가전다 (그림 10). 14) Rhabdosph aerid -타 원 혹은 원형 의 한 두개 의 판으로 구성된 신생 대 산(그림 11).
표 1 석회질 난노플랑크론의 주요 그룹 A. COCCOLITHOPHORES AND RELATED NANNOLITHS 21.. CAorkccanolgi etlh sik d is e l l ids FFaammii llyy ACorkcchoalni gthe al sckeiae el l aPceoaceh eB u(Mk 지es o(zMoiec s aonzdo iCc ) e nozoic ) Fami ly Prin s ia c eae Hay and Mohler (Cenozoic ) Fami ly Heli co sp h aeraceae Black (Cenozoic ) 3. Podorhabdid s Fami ly Podorhabdaceae Noel (Mesozoic ) 87465 ..... SSPRZytoyh e nrgap aotb ch o dd oas iospns psco h pi hlad iha ets aehr rieidi dr dsi s ds s FFFaa•imm~n• iiiii lllll yyyyy SSPERyotih e frnafp a etbc ho l dolas iosptnphs hopa hal ci hae t eearhaaeriaec ro ca en ceaRaaeeceae eie Lna Leh eLe amemBrmmdlmtaem cer(kermM mrma(eaMnsannonen zsn o(o C(izCc( oeC) en iecn o)no z ozozoicoi c)i c ) ) am i ly Zy go dis c aceae Hay and Mohler (Cenozoic ) B. NON- CO CCOLITHOPH0RE NANNOLITHS 1. Braarudosph aerid s Fami ly Braarudosph aeraceae Defi an dre (Mesozoic and Cenozoic ) 2. Cerato lit hi d s Fami ly Cerato l i tha ceae Norris (Cenc,z o ic ) 3. Disc oaste r id s Fami ly Dis c oaste r aceae Vekshin a (Cenozoic ) 4 . Fascic u li thi d s Fami ly Fascic u li tha ceae Hay and Mohler (Cenozoic ) 5 . Heli ol i thi d s Fami ly Heli ol i tha ceae Hay and Mohler (Cenozoic ) 6. Lit ha str i n i d s Fami ly Lit ha str i n a ceae Thie r ste i n (Mesozoic ) 7. Li tho str o mati on id s Fami ly Lit ho str o mati on aceae Haq (Cenozoic ) 8. Sp h enoli thi d s Fami ly Sp h enoli tha ceae Defi an dre (Cenozoic ) 9. Thoracosp h aerid s Fami ly Thoracosp h aeraceae Schil ler (Mesozoic and Cenozoic ) C. GENERA INCERTAE SEDIS 1. Isth molit hu s (Cenozoic ) 4. Nannoconus (Mesozoic ) 2. Mi cr orhabdulus (Mesozoic ) 5. Triq u etr o rhabdulus (Cenozoic ) 3. Mi cu /a (Mesozoic ) (5) Pont o sph aeri d s-이 중관이 접합되는 타원상 구조를 가지며 신생 대충 에서만 산출된다(그림 12). 16)S t ep hanol it h i ds- 다양한 기하학적 구조를 가지는 중생대산(그림 13). 17) Sy ra cosph aerid s-신 생 대에 산출되 여 중앙부의 결정 침들이 특칭 이다 (그림 14l. 18)Z ygodi sci ds- 중생대. 신생대산으로 타원형 연변벅에 im bric a te d 구조 가 특칭 이다(그림 15).
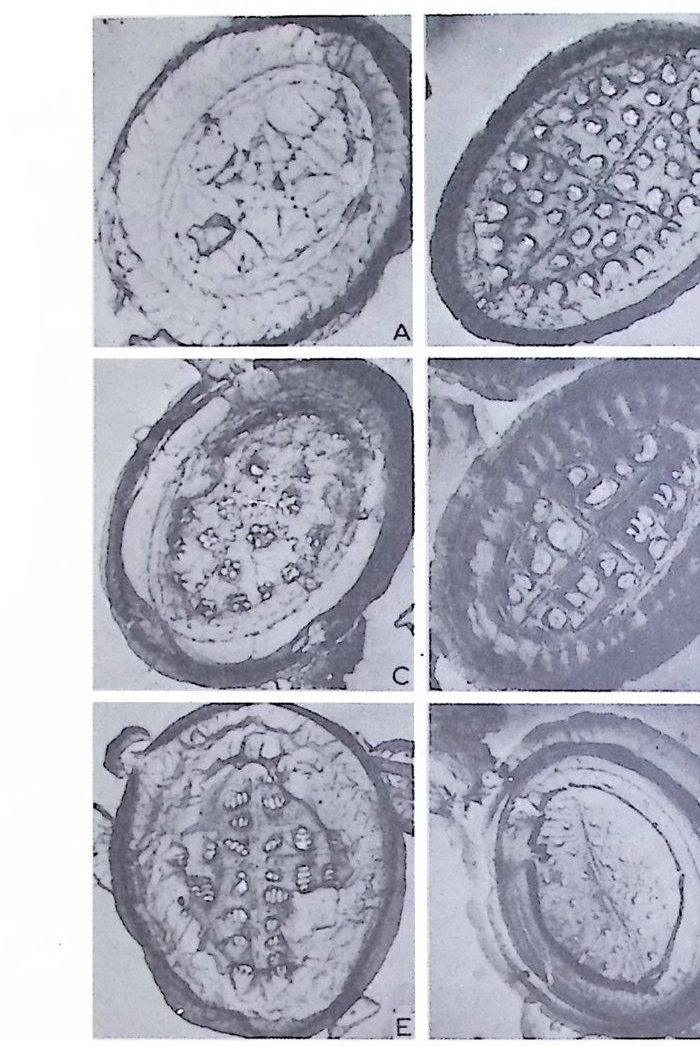 %r` ;\4>-.
%r` ;\4>-.
그림 5. Arkhang e lskie l lid s A. A rkhang e lskie l la cym bif orm i s Vekshin a _. pr oxim al view . X 5 .120. B. C. A. spe cil l ata Bukry_ 8 Di s ta l vie '< X4,320. C. pr oxim al view . X4 ,640. D. Broznsonia pa rca(St r adner). B ukry, dis ta l view . X 5. 600. E. Gartn erag o costa tu m Bukry, dis ta l view . X 3,600. F. Kam- ptn eriu s pe rciv a lli Bukry. pr oxim al view . X 3 ,360. (Af ter Bukry, 1969. )
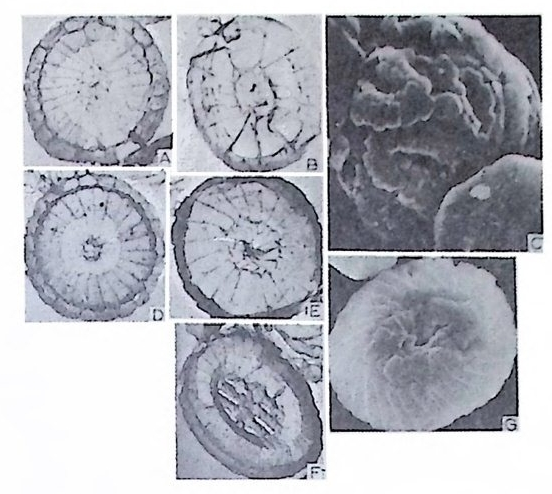
그림 6. 중 생 대 Coc col i t h id s A-C. Bi sc utu m consta n s (Gorka) Black. A. Proxim al view . X Z ,925. B. Di st a l view . X3 ,825. C. C.o c cosph ere. X3 .4 00. D. E Di sc orhabdus rot a to riu s(Bukry) Thie rste in . D. Prox- im al vie w . X3 ,825. E. Di s ta l view . X6 ,075. F. Sollasit es horti cu s (S t r adner and oth ers) Black. dis ta l view . X4 ,05 0. G. Watz nauria barnesae(Black) Perch Ni e lsen. dis ta l view . X 4 ,050. (A. B. D, E. and F aft er Bukry, 1969. )
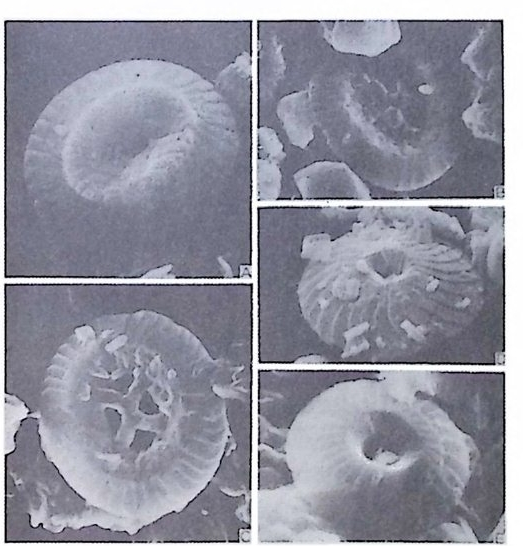
그립 7. 신생대 Coccolith ids A. Coccolith u s pe lag icu s (Walli ch ) Schil ler ,dis t a l view . X3,250. 8. Cruc ipl acolith u s ten uis (S tra dner) Hay and Mohler. dis ta l vie w . X 2,250. C. Chia s molith u s calif orn - icu s (Sulliv a n) Hay ·an d Mohler, dis t a l vie w . X3,000. D. Cy cl ococc-olith u s lep top o rus (Murray and Blackman) Kamp tne r, dis t a l view . X 6 , 250E. Umbi lico sph aera sib o g a e (Weber-Van Bosse) Gaa- rder, dis t a l vie w . X5,000.
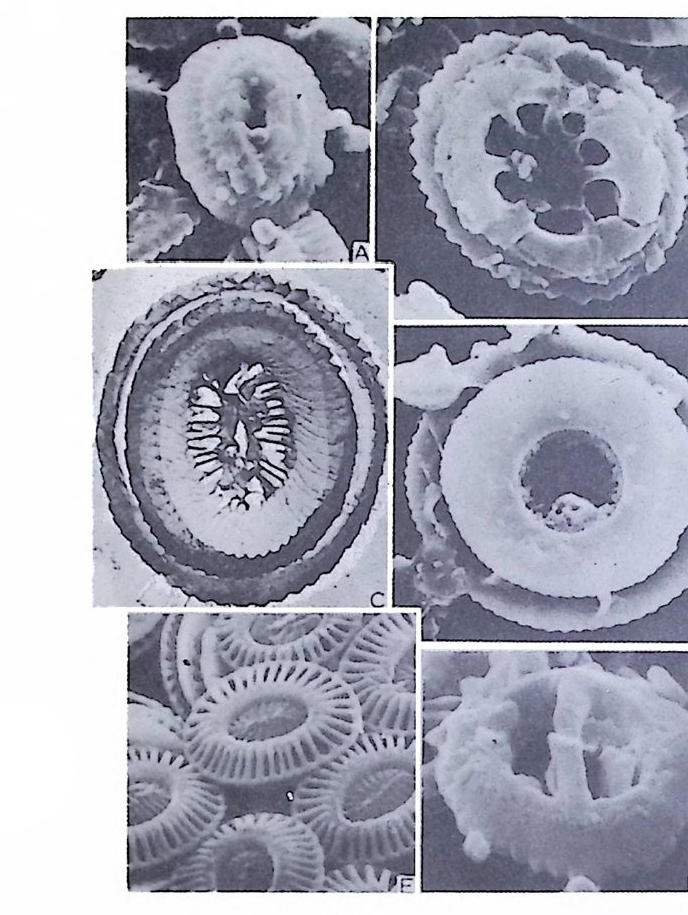
그림 8 . 신생대 Coccolit hi d s A. Prin s iu s marti ni i (P erch-Ni e lsenl Haq. dis ta l view , X3 ,300. B. Toweiu s crati cu lus Hay and Mohle r. dis - tal view . X4 ,400. C. Di c ty o coccit es sp. , p r oxim al vie w. X6 , 87 5. D. Reti cu lof en estr a dic ty o da(Defl an dre) Hay and Mohler. pr oxim - al view . X3 ,300. E: E mi li a n ia huxley i(L ohmann) Hay and Mohler. dist a l view s. X6 , 60 0. F. Geop h r:Jo cap s a oceanic a Kamp tn er. dis ta l vi ew. X 11. 000.
<,.-t`A \ f 、 ,' ,/ ,.. \ r.•i
 그림 9 . 신생 대 Coc co li t h id s Fami ly He li c o sp h ae ra c eae.A . Helic o sp h ae r a
그림 9 . 신생 대 Coc co li t h id s Fami ly He li c o sp h ae ra c eae.A . Helic o sp h ae r a
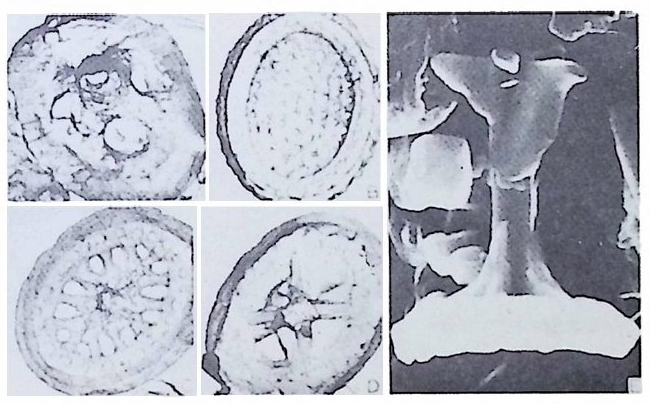 '!':':' -• •. ;\ { (/; '
'!':':' -• •. ;\ { (/; '
그림 10. Pndorhabdid s . A. Podorhabdus gr an u/at1 1 s( R e i1 1 hard11 Bukr~·. d is - ral \'iew Y '.L f iSO . 8. Crib r osp h aera ehrenberg i Arkhang e lsk~-- pr oxi m al vie w . X3 ,650. C. cret ar habdus erenulatu s Bramle tt e and Marti ni . dis ta l \,j e w. X4 ,800. D. E. Predi sc osp h ae ra creta c ea (Arkha 11g c ls k v l Gartn er. D. Di s ta l vie w . X 4 ,800. E. Si de vie w . X4 ,320 . (A -0:aft e r Bukry, 1969. )
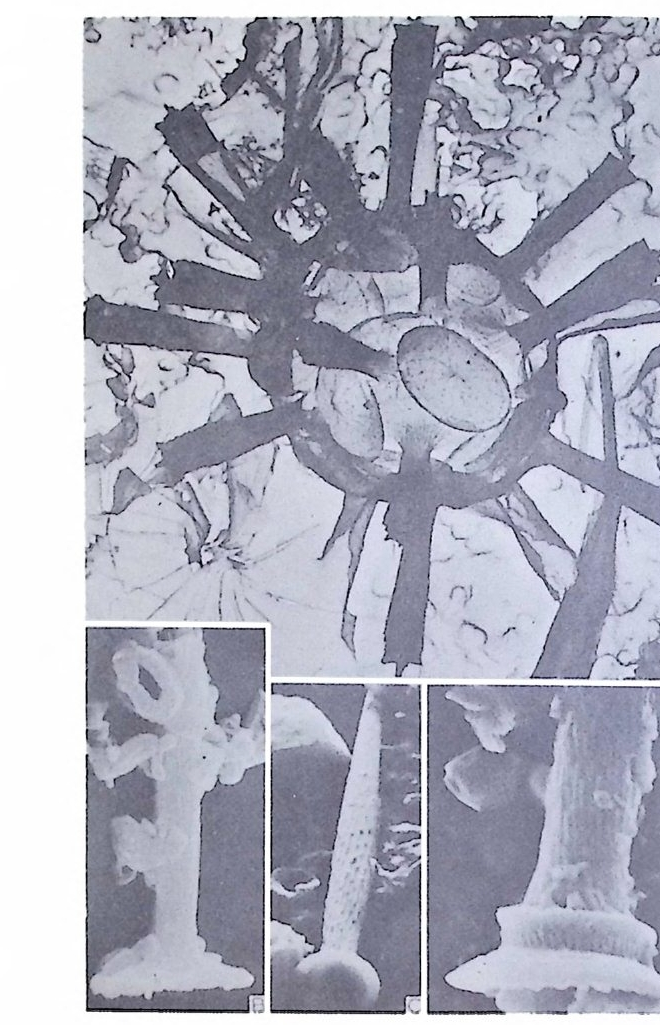
그림 11. Rhabdosph aerid s . A. A coccosp h ere of Rhabdosp h aera clavig e ra Murray and Blackman. X3 .710. (Photo :H. Okada\ . B. Rhabdosp h - aera pr ocera Marti ni , sid e view . X4 ,770. C, D. Blackit es sp ino - sus(Defl an dre) Hay and Towe. C. Ti tle d view of comp le te sp e c- im en. X3.180. D. Si de view of basal pla te and pa rt of the ste m . X4 ,770.
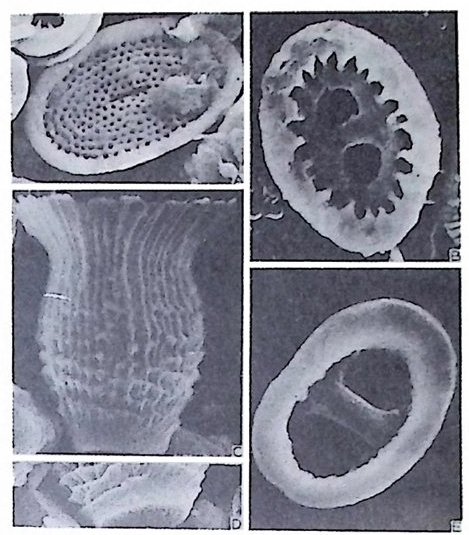
그림 12. Ponto sph aerid s A. Ponto s ph aera jap o nic a (Takaya m a) Haq_ dist a I ,rie w. X3 ,640. B. Transversop o nti s pu lcher(Def la ndre) Hay, M- ohler and w_a de.p r oxim al view . X3 , 12 0. C. D. Scyp h osph aera p- ulcherrim a Defl an dre. C. Si de view . D. Ti lt e d pr oxim al view sh- owi ng pe r for ate d centr a l area. X 2 ,600. E. Lop h odolit hu s nascens Bramlett e and Sulliv a n. dist a l view . X2 ,860.
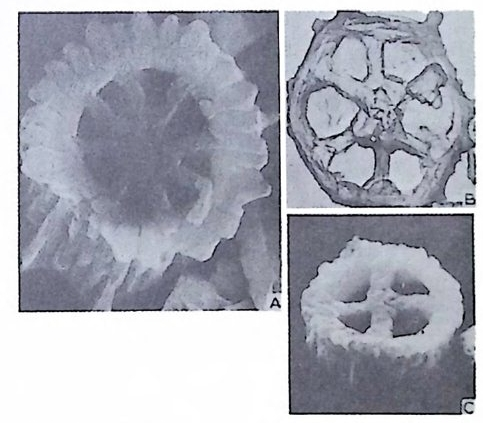
그림 13. St ep h anoli thi d s . A. St ep h anolit hi o n la{{it tei Noel. X8,100. (Ph- oto : H. Thie rste in ) B. Corollit hi o n exig u um S t radner. X 8 ,625. (Af ter Bukry, 1969. ) C. Cy li n d ralit hu s coronatu s Bukry. x4 .875. (After Roth and Thie r ste i n . 1972. )
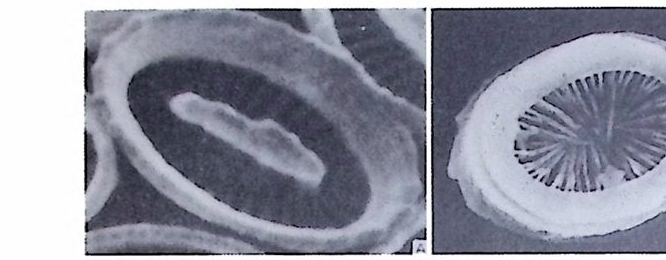
그림 14. Svracosph aerid s . A. Sy ra cosp h aera lami na Le cal -S ch landc r. d1 -sta l view . X6 ,300. B. S. pu lchra Lohmann. pr oxim al \ 어 ew. 니 .50 0 .
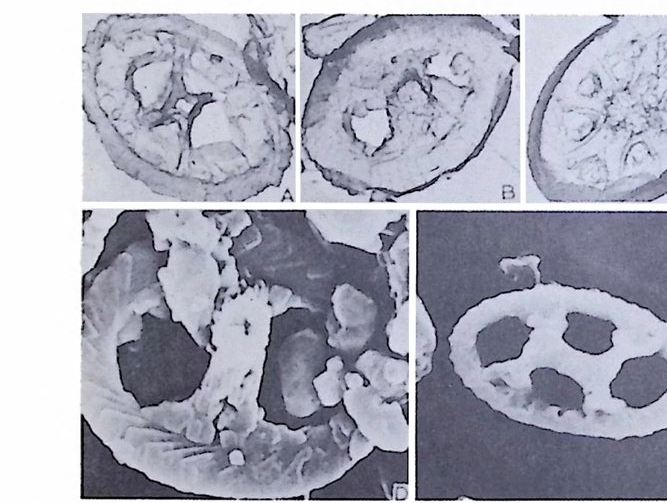 4
4
그림 15. Zy go dis c id s . A-C. Mesozoic Fami ly Ei ffel lit haceae. A. Ei ffell i- thu s tu rris e if feli ( D efl an dre) Rein hardt, dis ta l view . X 3 ,430. B. Chia s to z yg u s in tu rratu s(Rein hardt) Bukry. dis t a l view . X3 .020. C. Va ga lap illa octo radia t a ( Gorka) Bukry, dis ta l view ,X 5 ,200. (A C : aft er Bukry, 1969. )D-E. Fami ly Zy go dis c aceae. D. Zy go dis c Nus e oscigo mc cooild iet hs e sB rdaumbilue s tt (eD aenfl da n Sdurell)i v a Bnl.a ckd.i ds i t sa lt a vl ievw ie.w X. 4X . 2 1 6, 806. 0. E.
B . 非 Coccolit ho p h ore nanrolit hs (1) Braarudos p her i ds- 중생대 및 신생대에 산출되는다섯개의 절정단위 로 구성 되 는 pe n t al i t hs 가 득칭 이다 (그립 16). (2)Di sc oaste rid s-디 스크 모양이나 별모양의 as t erol it hs 로서 현생 종 중 양 적으로. 가장 중요한 그룹이다 (그림 17). (3)C e ra t ol it h i ds- 후기 신생대에만 산출되는 말 발굽모양의 nanno 화석 (그림 18). (4) Fascic u lith id s-반 시 린다형 의 신생 대 산 (그림 19). (5) H e li o l it h i d s - 두개의 인접하는 크기가 다른 판과 방사상구조륭 가진다 ( 그 럼 20). ( 6)L it ha s t r i n i ds- 중생대산으로서 다섯개의 별모양의 결정구조로이루어 침 (그림 21). ( 7 ) L it hos t roma ti on i ds- 신생대에 산출되는 두껍고 삼각형의 각을 가전 다 ( 그림 22). (8) Thoracosph aeri ds -중 생 대 와 신생 대 에 산출되 는 구 형 이 특 칭 이 다 (그 림 23 ). (9) S p henol it h i ds- 둥근 다번형 의 구조를 갖는 신생 대 산(그림 24)
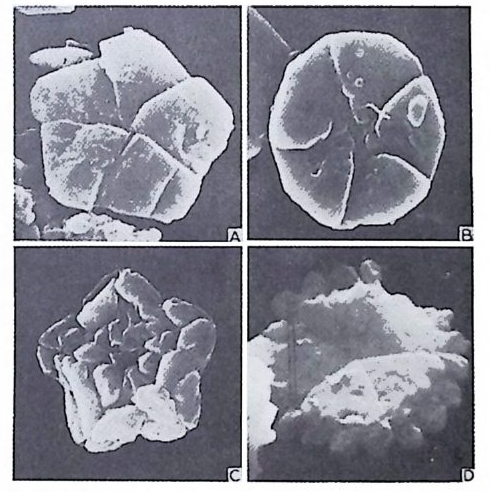
그림 16. Braarudosph aerid s . A . Braarudosph aera big e lowi (Gran and Ba- arud) Def la ndre. x3 ,135. B. B. disc ula Bramlett e and Rie d el. x 2,850. C. Mi c ranlholit hu s obt u sus St ra dner. X3 ,200. (Afte r Thi- erste in .lCJ 'll. )0. Pemma pa p ill atu m Marti ni . X3 ,420.
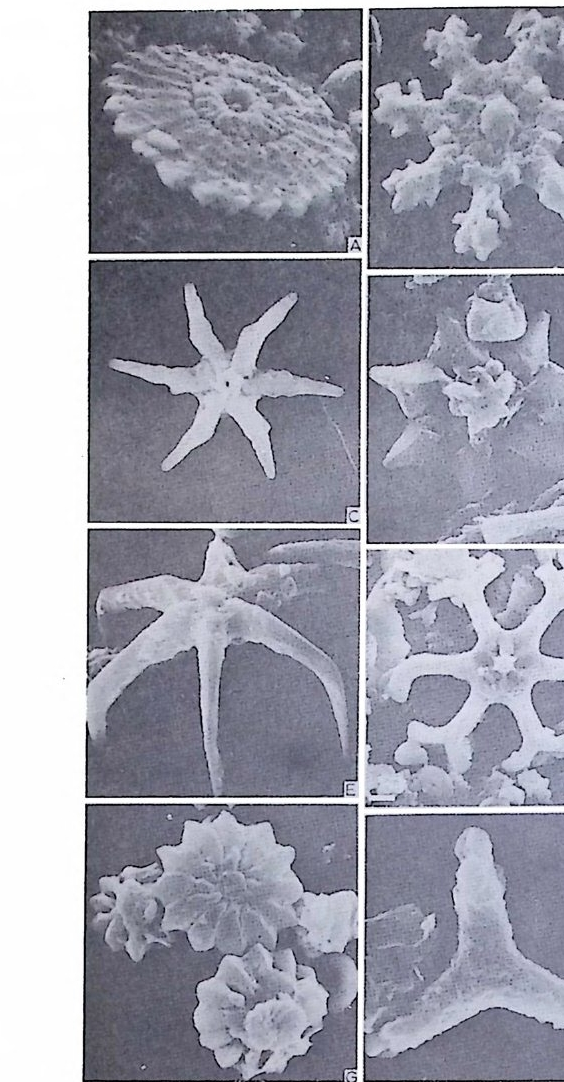
그림 17. Oi sc oaste rid. -;_ A. Di sc oasler mult ira dia /1 1s Bramkt tc and Ri ed el. X3, 000. B. D. mi ru s Def la ndre. ><2.500. C. D. I()doensis Bram- lett e and Ri ed el. X2,700. D. D. saip a nensis Bramlett e and Ri e - del. ><2,500. E. D. brouweri Tan Si n Hok . >< -1 ,000. F: D .-c h allen - ge ri Bramlett e and Ri ed el. X3,250. G. Di sc oasler()id ( 's kuep pe ri (St r adner)Bramlett e and Sulliv a n. >< 1.5()0. H. T rih r achi at u s or/- hosty lu s Shamrai. X2 ,500.
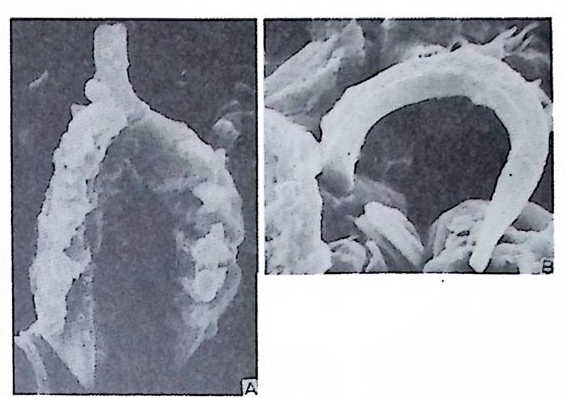
그림 18. Cerato l it hi d s. A.C erato l it hu s aff . acutu s Gartn er and Bukry _ X 2 .475. B. Amauroli thu s deli ca tu s Gartn er and Bukry_ X4 .125.
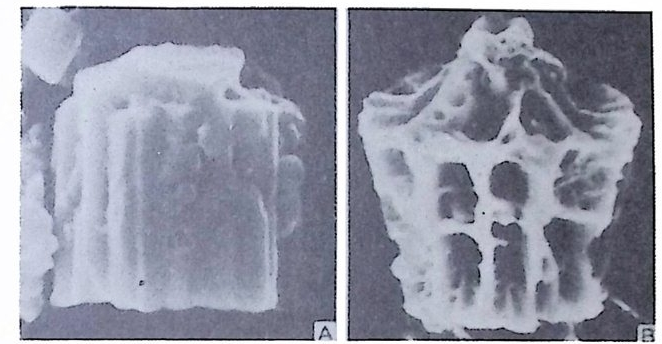
그림 19. Fascic u lit hi d s . A. Fasculi thu s tym p a nif orm i s Hay and Mohler. sid e view . X4 .550. B. F. schaubii Hay and Mohler. o bliq u e s•d e vi ew . X2 .100.
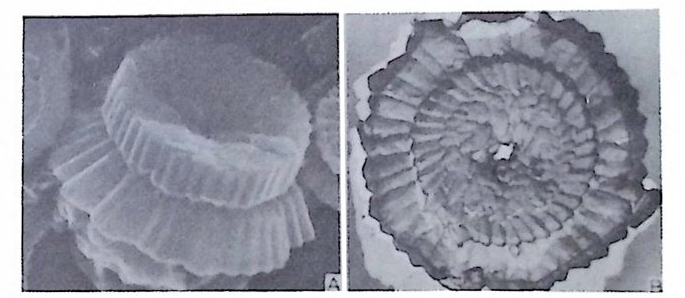
그림 20. He l i ol it hi d s . A. Heli oli thu s sp. , obliq u e sid e view . X 3 ,000. B. H. klei n p e ll i Sulliv an,_t o p vie w. X3 ,000. (B : ph ot o , H. Okada)
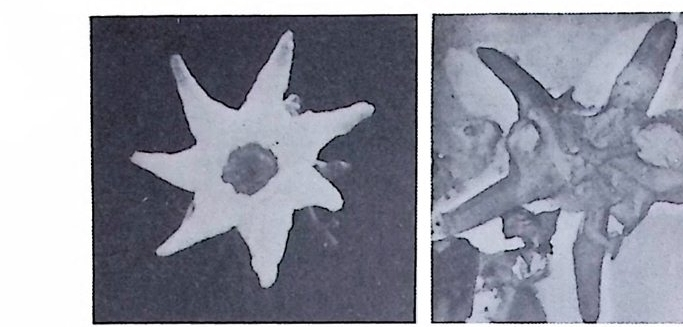
그림 21. Li tha sl rin i d s . A. Litha str in u s gr il li S t radner. X 3 .590. (Af 1 e r Forchheim er. 1972. ) B. Bukrya ste rhay i(B ukry) Prin s . X 4 .450. (A f1e r Bukry, 1969. l
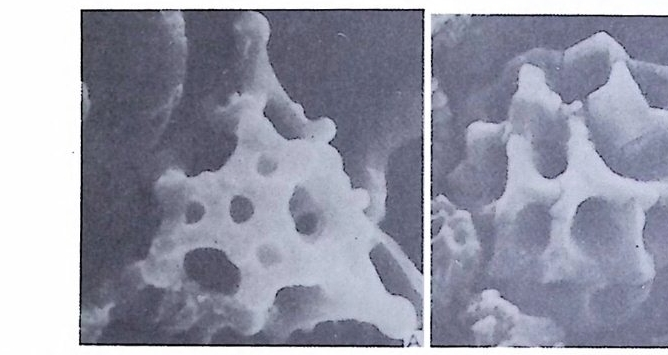
그림 22. Litho str o mati on id s . A. Li lh oslromalio n pe rdurum De fl a ndrc. X 3 , 120. B. Polyc ladolil h us op e rosus Def la ndre. X 5 ,S80.
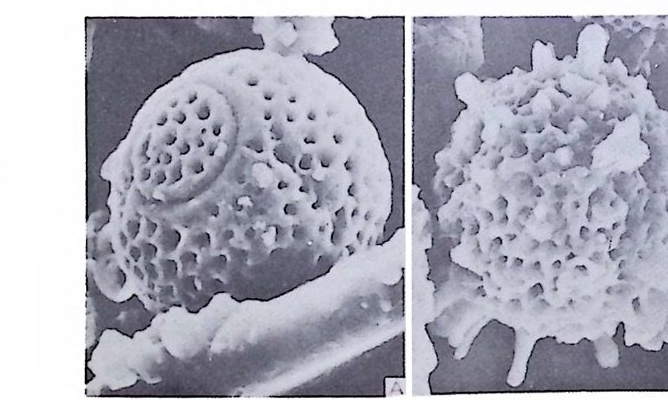
그 림 23 . Thoracosph aera sp .
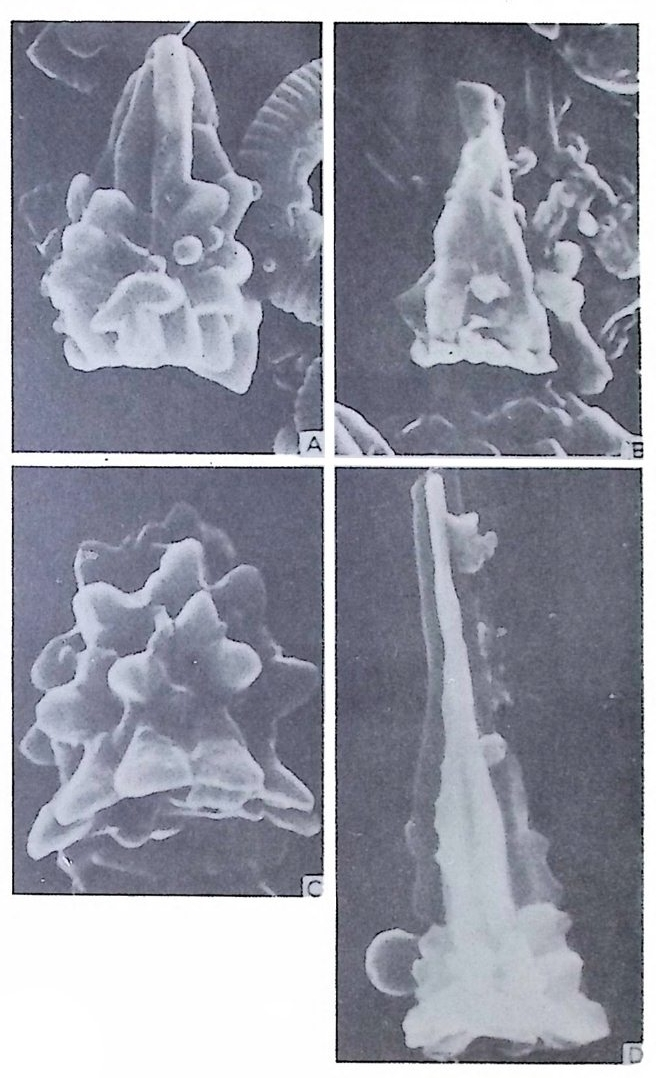
그림 24. Sp h enolit hi d s . A. Sp h enolit hu s r 따i ans Defl an dre. X4 ,500. B. S. dis te n tu s ( M arti ni ) Bramlett e and Wi lc oxon. XS ,625. C . S.mor- i(o rmi s( Bronnim ann and St ra dner) Bramlett e and Wi lc oxon. x 5 ,625. D. S. hete romorph us Defl an dre. X5 ,625.
C. Incerta e sed i s 속들 (D Isth molit hu s ® M ic rorhabdulus @ M ic ula @ Nannoconus @ Trig u et ro rhabdulus 둥 다섯 속이 알려 져 있다(그립 25, 26).
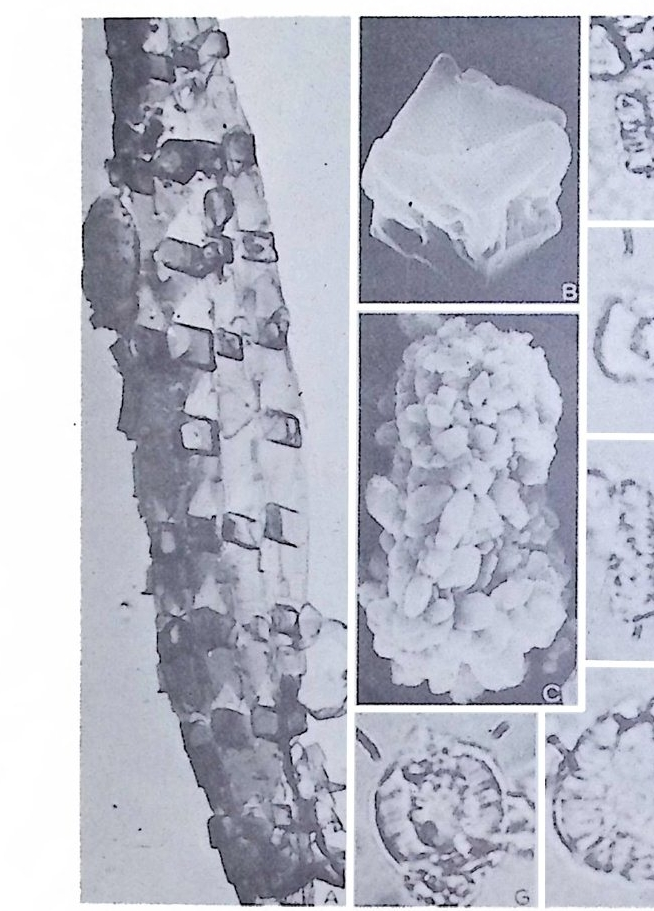
그림 25 . 주요 중생대 inc erta e sedis nannolit hs . A. Mi cr orhabdulus belgi cu s Hay and Towe. X8 ,000. B. Mi cu la sta urop h ora(G arde t) St ra dner.X 4 ,950. (Photo : H. T hie rste in . )C 一 H. Nannoconus sp p_ C. Overgr o wn out e r surfa c e of Nannoconus sp . D. N. mi nu tu s GB.r onNn. i m g alno nb. uElu. sN B. rtri.iu n in t iim i aBnnro. nHi.m N a.n nu.:a s Fsa. iNi . Berloonnng ia m tu asn nB. r u(Dnn— im H a. n Xn. 1, 1 00. aft e r Maniv i t. 197 1. )
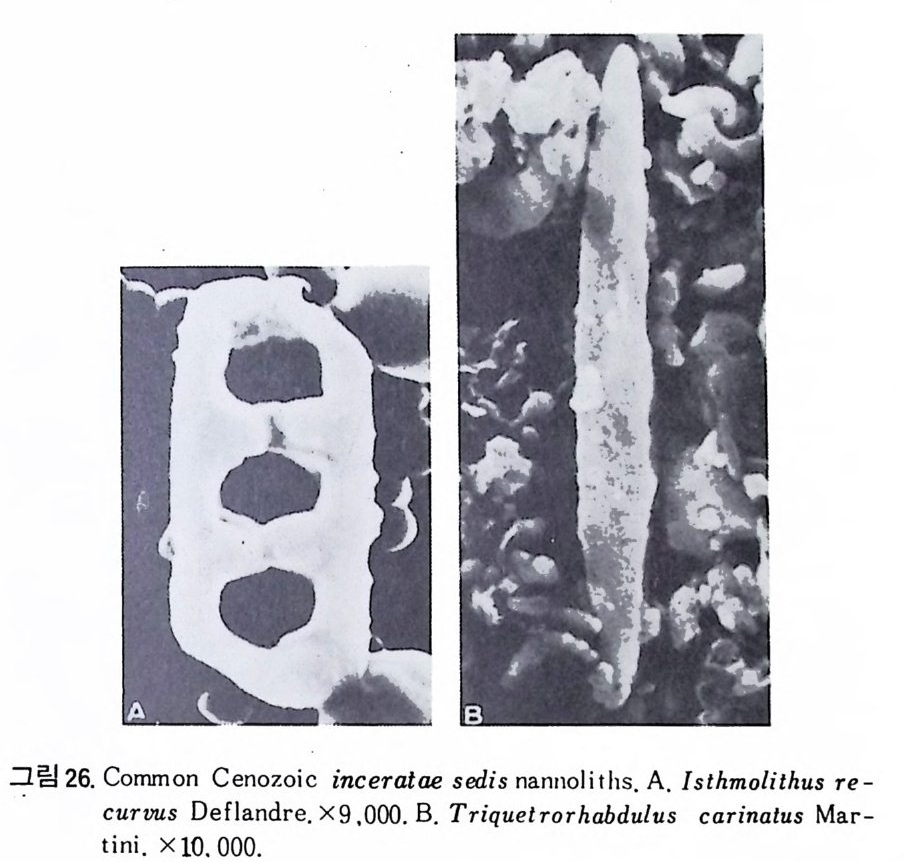 p., .
p., .
3 ·4 생태 해조류는 부유성 해양생물로서 원양으로부터 조간대 및 내륙 석호에 이 르는 생활권을 가진다. 특히 하구나 석호 등 해안 지역에는 해수의 평균 영도보다 크게 높거나 낮은 영분을 갖기 때문에 이들의 영도 범위가 넓 음을 알 수 있다. 앞서 예를 든 Em ilia nia huxle yi의 경우 45% 。 ~16% 。의 범위를 가진다. 이들은 광합성을 하기 때문에 수면에서 100~150m 까지 서식하며 특히
50m 수십에서 분포가 최대에 이른다. 150 종 이 상의 현생 종이 보고되 었으나 범 대 양적 인 분포는 아직 충분히 알려져 있지 않다. 양적으로.는 해류의 용승지역에 많은 영양분이 공급되 어 높은 생산이 이루어지므로 이들이 밀집되어 서식한다. 고생산역은 위 도 45° 의 남북지 역과 적도대를 따라 좁게 분포되고 있으며 풍부한 무기 영류는 coccol it ho p hores 를 위시한 석물성품랑크톤의 번성을 유도하고 이 는 또 동물성플랑크톤의 생산을 높게 하는 원인이 된다. 플랑크톤의 채 집과 해 저퇴 적물의 연구를 통해 서 McIn ty re 와 Be ( 1967 ) 는 대서양의 생지리를 위도와 기후로 특칭지워지는 다섯 개의 생물구로 구 분하였다. 또한 선택된 종들의 풀랑크톤 채집에 의한 위도별 분포를 퇴적물 내의 종의 분포와 비교해 본 질과 12,000 년전 마지막 빙기가 끝 난 이후해수가 급격히 따뜻해졌음을 밝혀냈다. McInt_ vr e & 외 (1970) 는 또한 태평양에서 네개의 생물지 리구륭 확인했으 며 각각은 수온 요소에 특히 예속적이어서 이러한 자료들이 고기후 및 고
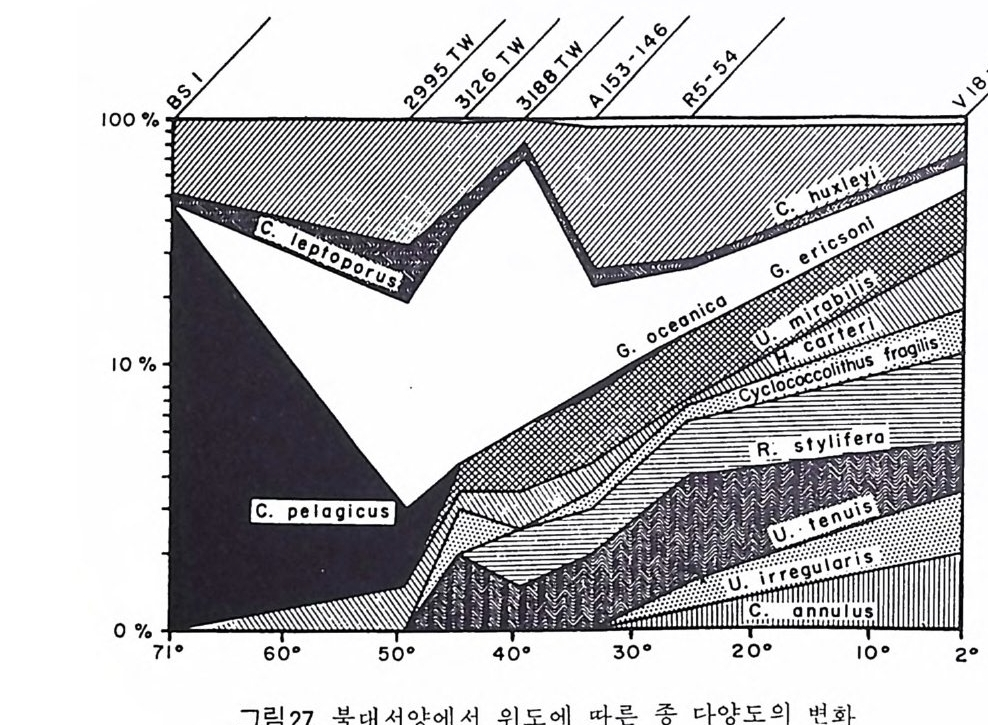 10%
10%
해양학적인 해석에 대단히 가치가 있다는 사실을 알게 되었다. Okada 와 Honjo (1973) 는 복부와 중앙태 평 양을 남북으로 자르는 단면에 따른 분포를 연구하여 표면온도와 표면해저류에 의해 이들의 분포가 결정 된다고 주장했다. 3 • 5 고생지리와 고기후 난노플랑크톤의 고생물지리와 고기후를 복원하려면 먼저 퇴적물 내의 이들의 분포양상을 밝혀야 한다. 이들 화석군이 생물군집 (b i ocoenos i s) 을 대변하는가 하는 문제는 많은 요소들에 의해서 철정되지만 가장 중요한 것은 탄산영 의 용해 현상이다. Coccol it h- 방해석은 개체가 죽은 후 아래로 떨어지연서 CaC03 로불포 화되어 있는 수충을 지날 때 용해된다. 그러나 대부분의 cocco lit hs 는 이 둘을 먹고 사는 동물의 fec al p elle t s 에 섞 여 서 하강하기 때문에 많은 양이 용해 작용을 연할 수 있다. 바닥에 떨어 지 연 fec al p elle t s 는 박테 리 아의 작용으로 곧 분해 되 어 그 속 에 들어 있 던 coccoli ths 는 용해 되 기 시 작한다. 수 심 이 대 체 로 4 km 가 넘 으
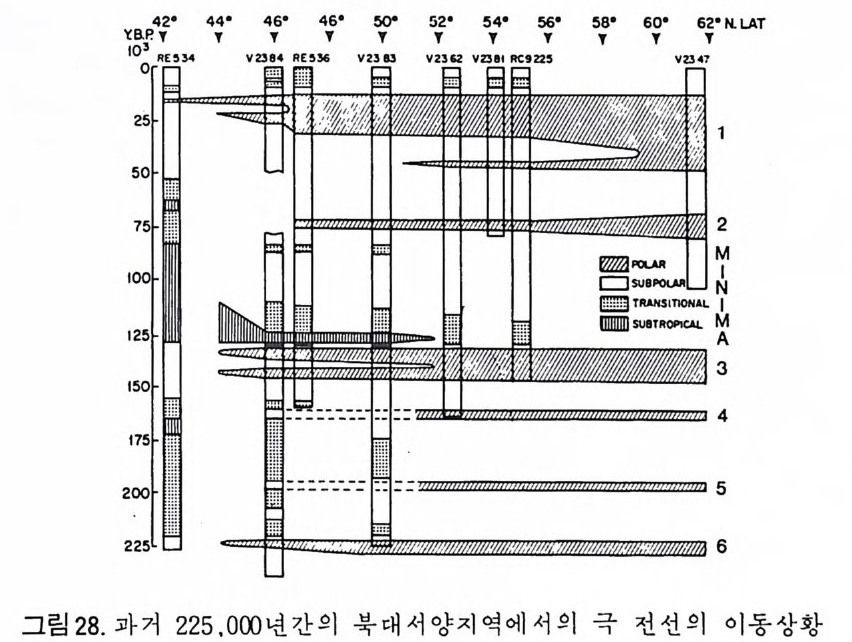 YaP. 4V 2 · 4V4 · 4V6 · 4V6 . gV 5Y2 • 5v4 · 3v· 5v8• 6v0• 6v2· N. 나 T
YaP. 4V 2 · 4V4 · 4V6 · 4V6 . gV 5Y2 • 5v4 · 3v· 5v8• 6v0• 6v2· N. 나 T
 COOL WARM
COOL WARM
떤 용해 작용이 국십해서 종다양도가 급격히 떨어지며 C.C. D. 수십인 5km 이하에서는 탄산영질 퇴적물을 찾아보기가 힘들다 . 속성작용도 역시 화석군집의 성질을 다소 변화시킨다 . 결정연 위에 방 해석 의 재 결정 현상이 일어나 nannol it hs 의 형 태를 변형 시켜 서. 감정에 어 려움을 주기도 한다. 따라서 분석 대상을 비교적 얕은 퇴적물에 국한시키 고, 군집내에 아직도 성세한 형태학적 구조를 갖는 nannol it h 군집을 선택함
으로써 이러한 오류를 피할 수 있다. 현재 해양의 해조류의 생태학적 자료를 과거 지질 시대에 적용시컬 수 있는 시대적 한계는 현재와 군집의 구성이 갇은 흥적세 를 넘지 못한다. 한 예로서 McIn ty re 외 (1972) 는 북대서양에서 채취한 시추코아로부터 nanno - l it hs 와 부유성 유공충의 군집변화를 조사해서 국전선의 지리적 변화를 추 적 하였다 . 그들의 연구 결과는 과거 225,000 년 동안 10· 이상의 위도상의 기 후 변 화 가 북대서양에서 일어났음을 지시한다(그림 28 참조) . 제 4 기 빙하시대 이전의 고생물지리와 고기후 복원은 이들 그룹의 진 화속도 가 빠르므로 현재 군집과의 비교룽 통해서는 불가능하다. Ha q과 Lohmann(1976 ) 은 대서양의 팔레오世와 올리고世의 난노풀랑크톤 군 집을 12 개의 시간대로 나누고 그들을 고기후의 지시자로 상아 위도에 대 한 선 호 도 룽 조사하였다. 고위도, 중위도 및 저위도 군집으로 구분한 난 노 식 물 군 으로 부터 다섯번의 위도의 변화가 탐지되었다(그림 29 참조) . 3 • 6 생물충서 및 진화 석회질 난노 플 랑크돈은 중생대 쥬라기초에 처음으로 출현해서 쥬라기 중 기 까지는 비교적 종다양도가 낮은 상태로 존속했다. 후기 쥬라기 Oxfo r di an 과 백악 기 의 A pti an~Alb i an 에 두 번의 큰 다양화가 일어나서 백악기 말 의 Maa s t r i ch ti an 에는 매우 풍부한 산출을 보인다. 그러 나 곧이어 대규모 의 해양생물의 절멸현상에 의해 신생대 제 3 기 초인 Dan i an 에는 4~5 종이 살아 남았을 뿐이다 . 잠시 동안 비교적 작은 유전자풀을 형성하면서 저속 도로 전화하다가 후기 팔레오世와 초기 에오世에 급격한 종분화가 다시 일 어나 신생 대 최대의 종다양도를 형성하였다. 이 시 기는 처음으로노 di scoas t er 가 출현하여 번성한 현상과 일치한다 . 그 후 종다양도는 급격히 떨어지다가 두번째의 방산현상이 중기 마이오 世에 일어났으나 홍적세까지는 계속 종의 다양도가 다시 감소하게 되었다. 풍부한 개체수와 짧은 생존기간은 난노플랑크돈이 훌륭한 시준화석이 되는 조전이며 지역간의 넓은 지충대미를 가능케 한다. 특히 저위도 지역 에서, 이들은 중생대 및 신생대 지층에 유용하다 .. 현재까지 연구된 생충서 차료를 다음 그림들 (30,31 ,3 2) 에 요약하였다. 난노 플 랑크톤의 전화 연구는 두 가지 관접에서 추구되어 왔다. 즉 하나 는 어떤 그룹의 계동 전화에 나타나는 전화의 경향과 다론 하나는 분류학 적 전화의 속도에 관한 것 이다.
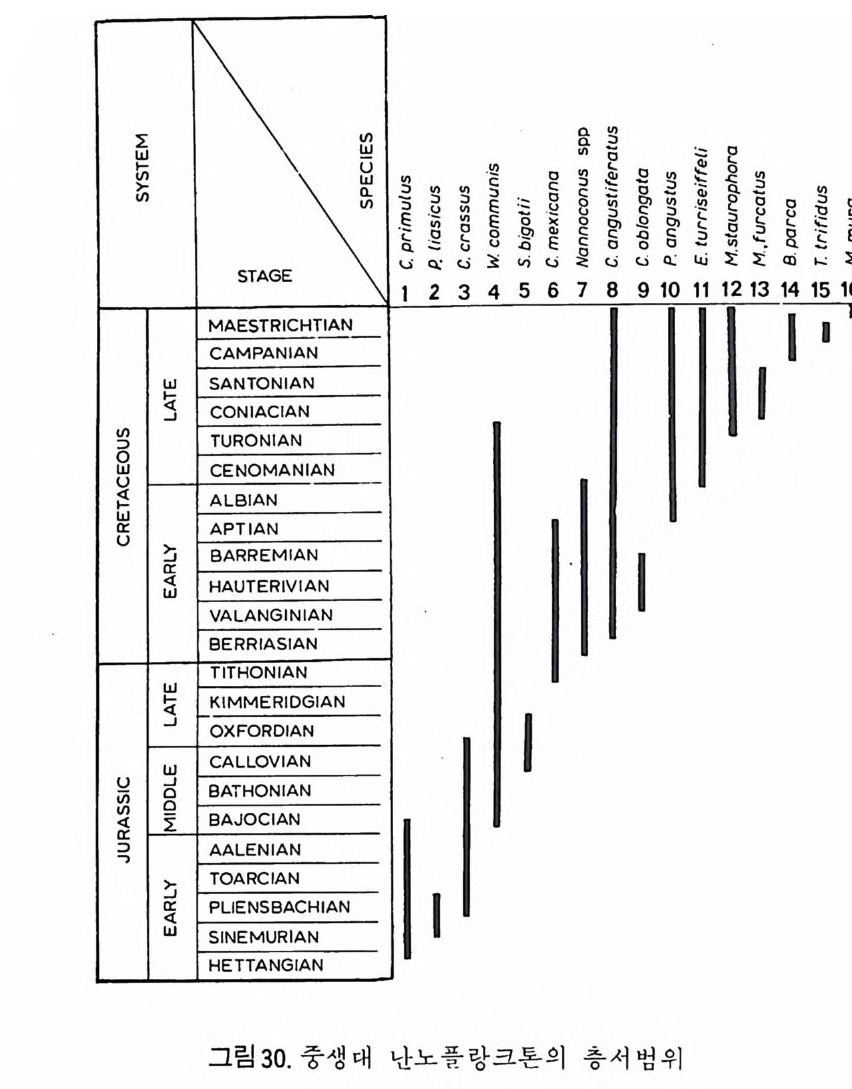 i8[ :巳 3 MSHCACVCAAALOAAEBNULEMNNSAITTSTOPIAANOETAAMNCGGRRNN AIIEIIIAIVCANN ANIHNIINAAA T N NNIA N 짧센Vl t홍I 법`2 3 3나H 조4 5 H=6 g70홍.용 능웅!!o8응I,눕 H흉0I9 1 1r0鬱 1,노1녹 니1I조2 1덩5조I3 Rot l8「'15〈t~訂1 4_
i8[ :巳 3 MSHCACVCAAALOAAEBNULEMNNSAITTSTOPIAANOETAAMNCGGRRNN AIIEIIIAIVCANN ANIHNIINAAA T N NNIA N 짧센Vl t홍I 법`2 3 3나H 조4 5 H=6 g70홍.용 능웅!!o8응I,눕 H흉0I9 1 1r0鬱 1,노1녹 니1I조2 1덩5조I3 Rot l8「'15〈t~訂1 4_
Gartn er(1969a) 는 하부 제 3 기층에서 산출되는 cocco lit h 의 일종인 Ch- i asmol it hus 속 내에 두 개의 계통선이 있음을 밝혔다 . 이들의 종분화와 지 질시대가 그림 33 에 표시되어 있다. 신생 대 의 Hel i cos p haera 屬울 연구한 Haq ( 1973a) 은 이둘의 전화현상에는 세 가지 의 뚜렷한 전화경향이 있음을 알아내 었다. 또한 Prin s ( 1971) 는 di scoas t er 그룹을 연구해 두 개의 계통을 뚜렷이 확인하였다. 학자들은 최근에 들어와서야 계동발생의 경향에 대한 연구를 위해 여러
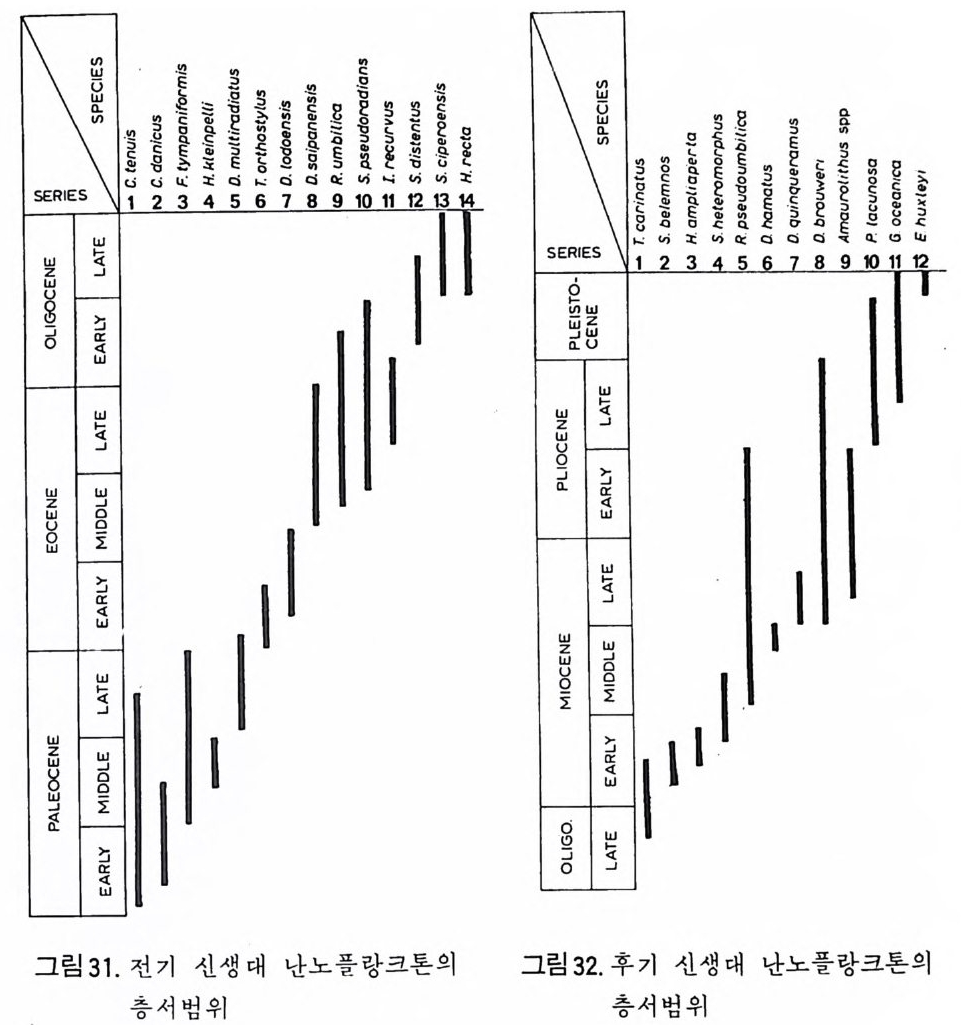 SE3RNIE3SQ owvw0il1 iv> 3J s· 3mu1as·2 1:n7 o 1EfEsuu 0o pkdaHH옹 4l큽E A Uso.nJ15 301s pnPM1p5·o끔1o70 1- 1 s1 0ns0gS|lEs8uJg aOndg .ull069 E pn n5Ealo sJIpnnA1:> '0sStJn 1 - s!꿈1·cgJ1o31 s응S ! P, o -g_JW114 SE3R’NI|E3멉uS o >3i- J4 d sn SOUE0Ja~ad Iao -nJs•O. .dn , E EG8P0), ~.,l qsJ nE1n0fosnE pnb9nE EE sOdaM nmJsgn bsOPcmJmqc sogoulog8C>n9 o mmd3 으0Mld
SE3RNIE3SQ owvw0il1 iv> 3J s· 3mu1as·2 1:n7 o 1EfEsuu 0o pkdaHH옹 4l큽E A Uso.nJ15 301s pnPM1p5·o끔1o70 1- 1 s1 0ns0gS|lEs8uJg aOndg .ull069 E pn n5Ealo sJIpnnA1:> '0sStJn 1 - s!꿈1·cgJ1o31 s응S ! P, o -g_JW114 SE3R’NI|E3멉uS o >3i- J4 d sn SOUE0Ja~ad Iao -nJs•O. .dn , E EG8P0), ~.,l qsJ nE1n0fosnE pnb9nE EE sOdaM nmJsgn bsOPcmJmqc sogoulog8C>n9 o mmd3 으0Mld
그룹의 계통의 경향에 관한 자료를 이용하기 시작했다 . 많은 계통들은 뚜 렷한 선조형이 없거나 형태학적으로 중간형태의 것을 갖지 않은 채 갑자기 출현하는 경우가 많다. 분류학적 반도율은 전화의 속도를 지시한다. 모든 화석 종들이 기재되고 보고된 뒤, 시간에 대한 종의 빈도율을 산출할 수 있 다 . 분류군의 첫번째 발생과 마지막 소멸을 자료로 시간_빈도곡선이 만들어지고 이로부터 발 생률과 소멸룹이 산출된다. 실제로는 분류에 대한 개인적인 견해차와 동
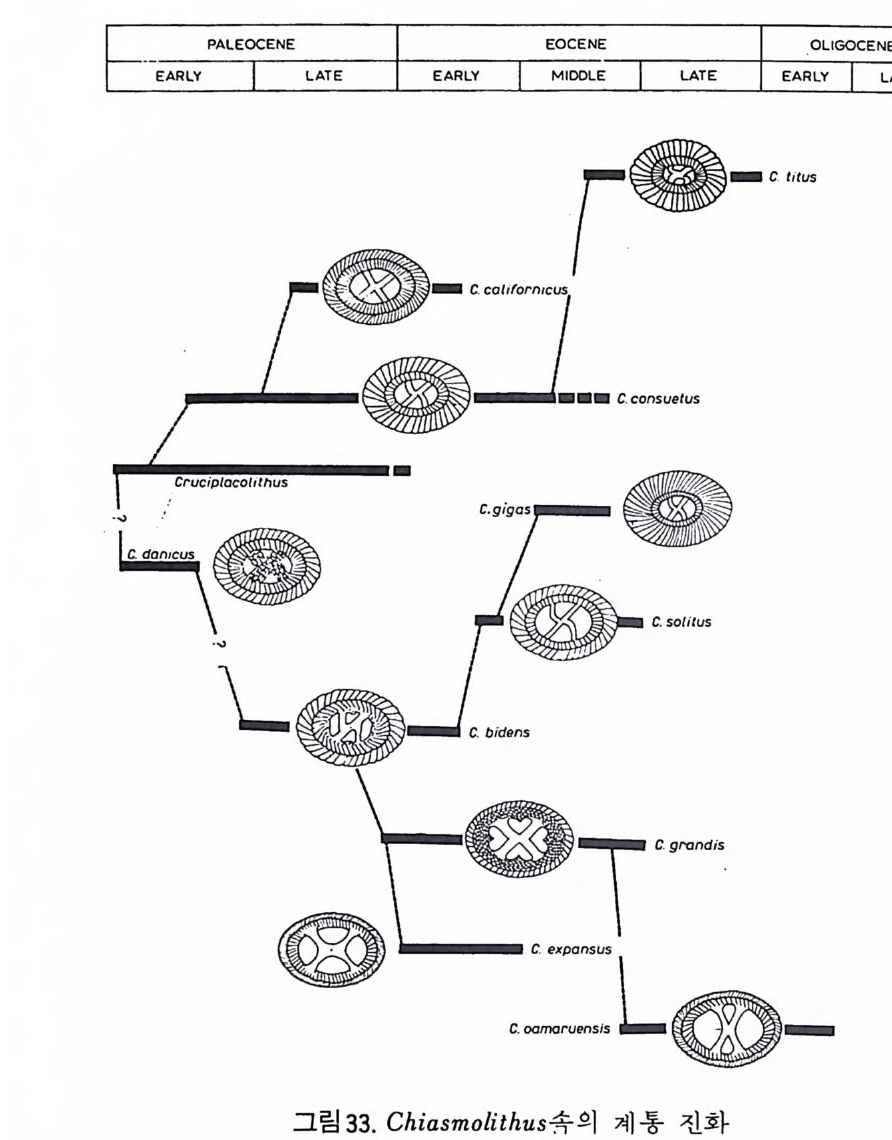 ·-E A RLY PALEOCENE LATE EARLY EMOICDEDNLEE LATE EAR.OL yL IGO| CENLEA TE
·-E A RLY PALEOCENE LATE EARLY EMOICDEDNLEE LATE EAR.OL yL IGO| CENLEA TE
의어 문제 그리고 화석의 보존과 속성작용 등에 의한 혼란으로 진화의 속 도를 연구함에는 장애가 많다. Ha q(l 973b) 은 산생 대 의 cocco lit hs 와 disc oaste rs 의 전화움을 연구하여 이들이 팔레오世에서 초기 에오世에 크게 발전했으며 (그림 34,35 참조) 평 군적 으로 coccol it hs 는 매 30. 000 년에 한 種이 . di scoas t ers 는 매 50. 000 년 에 한 種씩이라는 매우 빠른 종분화를 했음을 밝혀내었다.
 s6 :卜|. _I; I| ’/. 1,N ,E. /I , /E^ 「JI I;'1c\ |' N` : /I\OE L IG\| \O\C \ ELN \E EM IOC1E MN IE L g-4.d’ ,-T 'II'
s6 :卜|. _I; I| ’/. 1,N ,E. /I , /E^ 「JI I;'1c\ |' N` : /I\OE L IG\| \O\C \ ELN \E EM IOC1E MN IE L g-4.d’ ,-T 'II'
보충교재 Blac k, M., 1965. Coccolit h s, Endeavour, 24 : 131-137. [A short, in t ro duct- ory and beaut ifull y ill ustr ate d arti cl e on nannolit hs .] Fa2r8i,n a c1c97i, 0. A.P, ro1c9.7 1I .I RPloaunnkd t ot naib c le Coonn [c. aR lcoamree,o u2s :n 1an3n4o3p- l1a 3nk69t o. n [, SR uommme,a Srye p ot f. 2n3a--
nnop la nkt on ter mi no log y in Eng li sh , French, Ge rm an, Ita l ia n , Rus sia n and Sp a nis h .] Noe l, D.- , 1965. Sur !es coccolit he s du Ju rassiq ,ue Europ e en et d'A(riq ,ue du Nord. C. N. R. S:, Paris , 209 pp. (29 pls .) Noel, D., 1970. Coccolit he s creta ces de la Craie camp a nie n ne du Bassin de Paris . C. N. R. S., Paris , 129 pp. (48 pls .) [T hese tw o pa p e r s pr esent ul- tra str u ctu re of Ju rassic and Creta c eous nannolit hs . Numerous tra nsmi ss i- on and scannin g elect ron mi cr og r ap h s of the M esozoic sp e c ie s are illu s! ra- te d .] Paasche, E., 1968. Bio l og y and ph y s io l og y of coc co lit h op h orid s . Ann . Rev. Mi - c7: o bio l., 22 : 71-86. [A summary of all avail a ble data on the sub je c t till 19 67.] Perch-Ni el s en, K., 1971 . Durchs ic h t Terti ar er Co ccoli th e n . Proc. II Plankt - • onic Con{. Rome, pp. 939-980. [S ummary of Ce n o zo ic na n no fo s sil ta xonomy ;valid and inv alid tax a and morph olog ica l comp a ris o n s th rou g h hand-drawn ill- ustr a ti on s of tax a.] Rein h ardt, P., 1972. Coccolit he n, Kalkig e s Plankt on seit Ja hrmi ll io n en. Die Neue Brehm-Bi ich erei, A. Zeim se n Ve rl ag , W itten b e rg , 99 pp. [A sh ort int ro ducto ry tex t bo ok on nannof o ssil s in German lang u ag e .) Schil le r, J., 1930. Coccolit hi n e a e. In : L. Rabenhorst' s Kryp tog a me n -Flora,1 0 . Akad. Verlag s ge sellschaft , Leip z ig . pp. 89-263. [M orph o lo g y and Phy s io l - olog y of all liv ing coccolit ho p h ores known unti I the 1920' s.] 화석목록 Farin a cci, A., 1969 and late r . Cata l og u e of Calcareous Nanno(ossil s . Ediz i - oni Tecnoscie n za, Rome, vols. 1 to 7. [T his comp ila t i on giv e s t he o rig ina l descrip tion and illu st ra ti on s of all tax a at ge ne r ic and sp e c ie s lev e l wi tho ut comment s about valid i t y of names, etc . Volumes pu blis h e d int e rm i t ten t ly . ] Loeblic h Jr. , A. R. and Tapp a n, H., 1966. Annot a te d ind e x and bib l io g r ap h y of the calcareous nannop la nkto n . I . Phy c olog ia, 5 : 81-21 6. S ubs eq u e n t Nos . of the same : II : 1968, J. Paleonto l., 42 : 584 -5 98 ; III : 1969, J. Paleonto l ., 43 : 568-58 8 ; N :19 70, J. Paleonto l., 44 : 558 -574 ; V : 1970, Phy c olog ia, 9 : 157 -1 74 ; VI :1971, Phy c olog ia, 10 : 315 -339 ; Vll :1973, J. Paleont ol., 47 : 715- 759. [Th ese and fu tu re numbers ind ex all nannop la nkt on tax _a wi th co mment s about the ir non-valid i t y under Bot an ic a l or Zoolog ica l Code s if not de s cri- bed pr op e rly. Also inc ludes up -to -d ate bib l io g r aph y on narmo p la nkt on .] 참고문헌 Black, M., 1963. The fine str u ctu re of the mi ne ral pa rts of Coccolit ho p h or-
ida e. Proc. Linn . Soc. Land., 174 : 41-46 . Berge r, W. H., 1973. Deep - sea carbonate s : evid e nce for a coccolith lys oclin e . Deep - Sea Res., 20 : 917-92 1 . Bramlett e, M. N. and Wi lc oxon, J . A., 1967. Mi dd le Terti ar y calcareous nan- nop la nkto n of th e Cip e ro sect i on , Trin i d a d, W. I. Tulane St ud . Geol., 5 : 93 -131. Bukry, D., 1969. Up pe r Creta ceous coccoli ths from 1:ex as and Europ e . Univ . Kansas Paleonto l . Contr ib . , 51 : 1-79 (40 pis. ) . Bukry, D., 1971 . Cenozoic calcareous nannofo s sils from th e Paci fic Ocean. Trans. San Di eg o Soc. Nat. Hi s t. , 16 : 303-328. Bukry, D., 1973. Low-lati tud e coccoli th bio s tr ati gr aph ic zonati on . In : In_it i al Rep o rts of th e Deep Sea Dril ling Proje c t, XV. U. S. Government Prin t i ng 0 ffice , Washin g ton , D. C., pp. 685 -7 03. Bursa , A. S., 1965. Di sc oaste romonas calcif ero us n. sp. , an Arcti c relic t s e- creti ng Di sc oaste r Tan Sin Hok, 1927. Grana Palyn ol., 6 : 147-165. Ce pe k, P. and Hay, W. W., 1969. Calcareous nannop la nkto n and bio s tr a ti gr - aph ic subd iv isi o n of th e Up pe r Creta ceous. Trans. Gulf Coast Assoc. Ge- ol. Soc., 19 : 323-33 6. Edlawnadr dPsa, leAo.g eRn.,e . 19P7r1o . c.A IIc aPlcla arneko tu osn inc a nCnoonp{ l.a , n kRtoo m n ez, opnpa.t i3 o8n 1o-f 4 1 t9he. New_ Eng - Forch h e im er, S., 1972. Scannin g electr o n mic r oscop e stu di es of Creta ceous coccolith s from th e Kop ing s berg Borehole no. 1, SE Sweden. Sver. Geol. Unders., Ser. C., no. 668 : 141 pp. Gaarder, K. R., 1962. Electr o n mic r oscop e stu die s on holococcolith o p h orid s . Ny tt. Mag. Bot. , 10 : 35-51(12 pis. ) Gartn er, S., 1969a. Phy lo g e neti c lin e ag e s in the Lower Terti ar y coccolith ge - nus Chia smolith u s. Proc. N. Am. Paleonto l. Conv., G : 930-957. Gartn e r, S., 19696. Correlati on of Neog e ne pla nkto n ic for ami ni f er a and calc- areous nanno fo s sil zones. Trans. Gulf Coast A ssoc. Geol. Soc., 19 : 585 - 599. Gartn er, S. and Bukry, D., 1969. Terti ar y Holococcoli ths . J. Paleonto l., 43 : 1213-1 221 . Haq , B. U., 1973a. Evoluti on ary tre nds in th e Cenozoic coccolith o p h ore ge nus, He li c o p o nt o sph aera. Mi c rop a leonto log y, 19 : 32-52 . Haq, B. U., 19736. Transg r es sio n s , clim ati c chang e and th e dive rsit y of cal- careous nannop la nkto n. Mar. Geol., 15 : M25-M 30. Haq, B. U. and Lohmann, G. P., 1976. Early Cenozoic calcareous nannop la n- kto n bio g e og r aph y of th e At la nti c Ocean. Mar. Mi c rop a leonto l., 1 : 119-194. Hay, W. W., 1965. Calcareo us nannofo ssils . In: B. KummeI and D. Raup (Edit - ors) , Handbook of Paleo n t o log ica l Techniq ,ue s. W. H. Freem an, San Fr- ancis c o, Calif. , pp. 3-6.
Hay, W. W. and Mohler, H. P., 1967. Calcareous nannop la nkto n fro m Early Teriia r y rocks at Pont Labau, Franee and Paleocene-Early Eocene corre- lati on s. J. Paleonto l ., 41 : 1505 一 1541 . Hay, W. W., Mohler, H. P., Roth , P. H., Schmi dt , R. R. and Boudreaux, J. E., 1967. Calcareous nannop la nkt on zonati on of the Cenozoic of the Gulf Coast and Carib b ean-Anti llea n area and tra nspo rta ti on correlati on . Trans. Gulf Coast Assoc. Geol. Soc., 17 : 428-48 0. Honjo , S., 1976. Coccolit hs : pr oducti on , tra nsp o rta ti on , and sedim ent a ti on . Mar. Mi cr op a leont ol ., 1 : 65-79 . Isenberg, H. D., Doug la s, S. D., Lavin e , L. S. and Weis s fe l lne r, H., 1967. Laborato ry stu d ie s wi th coccolit ho p h orid calcif ica ti on . Proc. Int. Con/, Tr- op ica l Oceanog r aph y, Mi am i , Fla., pp. 155 -1 77. Maniv i t , H., 1971. Les nannof os sil es calcair e s du Creta ce fra nsais (de I'A p tien au Dania n ) , Essae de bio zonati on app u y e e sur Les str a to ty pe s. The sis , Paris Univ e rsit y, 187 pp. (32 pis. ) . Mant on , I. and Leedale, G. F., 1969. Observati on s on t he rnic r oanato r ny of Coccoli thu s pe lag icu s and Cric o sp h aera cart e rae, wi t h sp e cia l re fe re n c e t o the orig in and natu re of coccolit hs and sc ales, J. Mar. Bi ol . Assoc., U. K. , 49 : l_-1 6. Marti ni . E., 1971 . • St an dard Terti ar y and Qu ate r nary cal ca r eo u s nanno p la nk- t on zonat i on . Proc. II Plankt on ic Con/. Rome, pp. 739 -7 85. McInty r e, A. and Be, A. W. H., 1967. Modern coccolit ho p h o r id a e of the At l- anti c Ocean-I. Placolit hs and cy r to l it hs . Deep - Sea Res., 14 : 561-5 97. McInty r e, A., Be, A. W. H. and R:ich e, M. B., 1970. Modern Pacif ic cocco- litho p h orid a : A pa leonto l og ica l the rmomete r . Trans.N . Y. A cad. Sci. , 32 : 720- 731 . McInty r e, A., Ruddim an, W. F. and Jan t ze n, R., 1972. Sout hw a r d pe n e t ra ti - ons of t he North At lant ic Polar F ront : fau na) and flor al evid e nc e of large - scale surfa c e wate r mass movement s over the last 225, 000 ye a rs. Deep - Sea Res., 19 : 61-77. Okada, H. and Honjo , S., 1973. The dis t r ib u ti on of oceanic coc col it h op h ori- ds in the Pacif ic. Deep - Sea Res., 20 : 355-374. Parke, M. and Adams, I., 1960. The mot ile (Crys ta l lolit hu s hy a li nu s Gaarder and Markali) and non-moti le ph ases in the life his t o ry of Coccolit hu s pe la- gicu s (Wallic h ) Schil le r. J. Mar. Bi ol . Assoc. U. K., 39 : 263-2 74. Prin s , B., 1969. Evoluti on and st ra ti gr aph y of coccolit h in i d s fro m th e Lower and M idd le Li as . Proc. I Plankt on ic Con/., Geneva, 2 : 547-55 8. Prin s , B., 1971 . Sp e culat i on s on relat i on s, evolut ion and st rat i gr aph ic dis t r- ibu t ion of dis c oaste rs. Proc. II Plankt on ic Con/., Rome, pp. 1017-1037. Roth , P.H., 1970. Olig o cene calcareous nannop la nkto n bio s t rati gr ap h y. Eclog a e Geol: Helv., 63 : 799-881 . Rot h, P. H. and Berge r, W, H., 1975. Di st rib u ti on and dis s oluti on of coccol-
ith s in the sout h and cent ra l Pacif ic. In : Di ss oluti on of Deep - Sea Carbon-
ate s . Cushman Found. Foram. Res., 13 : 87 -113. Rot h, P. H. and Thie r ste i n , H., 1972. Calcareous nannop la nkt on : Leg 14 of the Deep Sea Drill i n g Proje c t. In : Init ial Rep o rts of the Deep Sea Drill - ing Proje c t, XIV. U. S. Government Prin t i ng Of fice , Washin g ton , D. C.,pp . 421-48 5. S t radner, H. and Papp , A., 1961 . Terti ar e Di sc oaste rid e n aus Oste rreic h und de ren st rat i gr ap h is c he Bedeut un g . Jah rb. Geol. Bundesanst. (W ien ) ,7: 1-160. Thie r st e i n , H. R., 1971 . Te nt a ti ve Lower Creta c eous calcareous nannop la nkt - ton zonat ion . Eclog u e Geo!. Helv., 64 : 459-4 88. Thie r ste i n , H. R. , 1973. Lowe r Cret a c e ou s calc a reous nannop la nkt on bio s t ra - t igr ap h y . Abh. Geol. Bundesansl. (W ien ) , 29 : 52 pp. Thie r ste i n , H. R., 1974. Calcareous 11a1111op la 11kt o1 1-L eg 26 of the Dee p Sea Drill i n g Proje c t. In: lni lial Rep o rls of the Deep Sea Dril l in g Proje c t, XXVI. U. S. Government Prin t i ng Of fice , Was h in g ton , D.C. ,p p. 6 19-667. Thie r st e in , H. R., 1976. M esozoic calc a reou s nan11op la nkt on bio s t r ati gr aph y of marin e sedim e n t s . Mar. M icr op a leonlol., 1 : 325-36 2. Wata b e, N. an d W ilb ur, K. M. , 1966. Ef fec t s of tem p e rat u r e on gr owt h ca- lcif ica t ion and coccoli t h fo rm in Coccoli t hu s hu .rle y i (Coccolit h in e ae) . Limn - ol. Oceanog r ., 11 : 567-5 75. W ilb ur, K. M. an d Wata b e , N., 1963. Exp e rim ent al stu d ie s on calc if ica ti on of the alga Coccoli t hu s huxley i. Ann. N. Y. Acad. Sci. , 109 : 82-112. W ise , S. W. and Ke lt s , K. R., 1972. Inf er re d dia g e ne t i c his t o ry of a weakly sil ic i f ied deep sea ch alk. Trans. Gulf Coast A ssoc. Geol. Soc., 22 : 1-77 . 4 石灰藩 (C alcareous Al g a 러 4 • 1 서언 오늘날 열대나 아 절대의 대 · · +·봉 에 퇴적되고 있는 석회질 퇴적물로부터 우리는 많은 종류의 저서성 두조와 홍조류가 형성시킨 CaC03 로 이루어진 쇄설물을 발견할 수 있다. 아러한 것들은 과거 지질시대의 지층에서도많 이 발견되는데 이 중에는 最古의 화석군에 속하는 것들도 있다. 석회조는 고생태와 생물충서학적 가치 때문에 미고생물학에서 중요하다. ·조류는 탄 산염질 광물의 생산자로서뿐만 아니라 초 (ree f)의 골격을 이루며 세립질 입 자들을 묶어 놓는 퇴 적 작용에 중요한 몫을 한다. 深는 수생 광합성 식물을 총망라하나 특히 석호]조는 이 중에서 방해석질 골격을 갖는 그룹을 칭한다. 석회조의 연구는 관습상 플랑크본인 해조류를 제외한 저서성 형태에 국 한한다. 1700 년대부터 시작된 석회조 연구는 20 세기 초에 이르러 비로소 현재 우리가 알고 있는 지식의 바탕이 되는 업적들이 발표되기 시작했다. 그러 나 화석 석회조에 관해서는 최근까지도 찰못 이해되어 왔는데 이는 현존 하는 상동 후손이 없기 때문에 더욱 심했다. 지난 20 년 동안 탄산영 퇴적 학의 큰 발전에 힘입어 이들의 지질학적 해석과 분류학적 정체가 밝혀지 면서 미화석으로서의 가치가 높아졌다. Joh nson(1961) 은 화석 석회조의 분류를 정리했으며 계속해 서 카타로그“ 동을 저술했다. 최근 Gi ns burg 외 (1971) 는 석회조의 지 질학적 관접둘을 요 약했다. 석호]조 화석의 연구는 암석의 박편을 제작하여 10~150 배의 배율인 광 학현미경으로 실시한다. 박편을 투과한 광선은 외부의 형태뿐만 아니라 내부의 골격의 구조도 관찰할 수 있게 해준다. 대부분의 석회조류는 수 mm~ 수십 cm 의 크기물 갖지만 이들은 죽은 후 잘게 분리되며 또한 세부구 조률 보기 위해서는 현미경이 필요하기 때문에 미화석으로 취급된다. SEM( 전자현미경)은표면구조와내부구조를 관찰할 때 필요하다. 완전하게 고화된 퇴적물 안에 있는 석회조 화석은 속성작용을 받아 세부구조가 붕 괴 될 수 있기 때문에 SEM 을 통한 연구에 있어서 제한된 사용 범위를 갓 는다. 4 • 2 생활사 조류는 수생이며, 독립영양성이고 비관다발석물이다. 염상체라고 불리 는 몸체는 뿌리, 줄기 및 잎으로 분화되어 있지 않고 모든 세포들이 광합성을 한다. 조류는 크기, 형태, 세포의 구조, 생화학 그리고 생식에 있어 종류에 따라 매우 다양한 특칭을 가진다. 세포의 구성에 있어서 조류는 원시적 인 원핵세포(p rocar yo t es) 와 분화된 진핵세포 (eucary o t es) 의 두 가지가 있다. 청록조는 박테리아나 바이러스와 같이 DNA 가 세포막으로 세포질과 격리되어 있지 않은 원핵세포를 갖는다.반 면에 다른 종류들은 세포막으로 둘러싸안 염색체 핵을 가진 진핵세포를갖는다. 저서성 석회조류는 인위적으로 묶여전 그룹이므로 그들의 생물학적 특 성은 대단히 다양하다. 따라서 그들의 골격상의 형태를 분류에 적용할
때는 각 그룹이 독자적안 영역을 갖는다는 접을 잊어서는 안된다. 현재 생존하는 10, 000 종의 조류 중 약 5~10% 가 어느 정도라도 석회 질화된 것이며 이들은 거의 소수의 홍조와 녹조 과에 속한다. 대부분 의 비 해 성 인 석 회 조는 Charo p h yt a 에 속한다. CaC03 를 침전시키는 현상은 광합성을 하는 동안 CO2 가 표면부에 추 충되는 과정에서 기 인한다는 연구 보고가 있다. 또 다론 현상은 세포간의 물질교대 및 선택적인 탄산영의 흡수와 관련이 있다는 것이다. CaC03 가 침전되는 장소는 조류의 종류에 따라 다르며 이 접이 화석조 류의 보존에 중대한 영향을 미 찬다. 침전되는 장소는 : G 세포내, @)세포 벽, @ ) 세포부 ,L, @ ) 식물체의 표면부로 나눌 수 있다. Corall i naceae( 홍조) 과는 방해석 형태로 침전시키며 마그네슘이 다소 격 자 사 이 에 포함된다. 한편 Dasyc l adaceae 및 Co diac eae (녹조) 과에 속하 는 종류들은 애라거나이트의 형태로 침전된다. Charo p h yt a 의 생식기는 방 해석질이다 . 석회조류의 광물성분은 화석화 과정에서 각각 다른 정도의 속성작용 을 보여 결국 보존의 질에 차이를 초래한다 . 4 • 3 형태와 분류 현재 사용되고 있는 석회조의 분류는 ®세포의 표면이나 세포의 내부 혹은 세포들 사이에 CaC 야를 침전시키는 능력이 있는 종류, @)비석회 질 설모양 그리고 코코이드조 (cocco i d al ga e) 가 만든 유기질 막 위에 미세 한 탄산영질 퇴적물이 기계적으로 집적되어 형성된 박충리 구조를 갖는 형태 등 두 가지로 나눈다. 대부분의 전자와 같은 골격질 석회조는 현존 하는 생물학적 분류 체계에 적절하게 포함되어 분류된다. 현생조류는 색 소, 먹이를 비축하는 방법, 영색체의 초미구조, 그리고 편모의 종류 동 에 의해 구분된다. 많은 조류는 분명한 색의 차이로 일차적인 분류가 실 시된다. 구조나 생석계 둥은 현생조류의 재차분류에 열쇠가 된다. 화석 종의 경우도 현생종의 분류체계를 그대로 따르며 표 1 은 주요 그룹의 분 류표이다. 화석종에 대한 지석은 불완전하고 단편적이간 하지만 계속해서 새로운 자료들이 보충되고 있다. 이들 중에는 현재 살고 있는 후손이 없어서 조 류의 전화에 관한 연구를 어렵게 하는 것도 있다. 비골격질 조류인스트 로마톨라이트 (s t roma t ol it e) 는 여러가지로 해석되어 그들의 분류도 생물 학적 인 것과 비생물학적 인 것 이 있다. 1883 년 최초의 고생물학적 기재가
표 1 화석 석회조의 분류 Ap pr oxim at e number I some common ge nera pr in c ip a l ge nera 1lum CYANOPHYTA (blue-g re en alg a e)l5 I Gi rva nclla Class CYANOPHYCEAE 1yl u m RHODOPHYTA ( red alg a e) Class RHODOPHYCEAE Order CRYPTONEMIALES Fami ly SOLENOPORACEAE 21 pa rachaete le! Fami ly CORALLINACEAE 5 ASo rclehnaoepo ol irt( hI o t h amniu m Subfa m i ly MELOBESIEAE Litho t ha mniu m (crusto se corallin e s) Li tho p h y ll um Litho p o rell(I Subfa m il y CORALLINEAE 5 MCoerlonblleisn ia ( I (arti cu late d corallin e s) Ja ni( I Fami ly UNCERTAIN 5 A rchaeolit ho p h y ll um (ancestr al corallin e s) Cuneip h y c us ph y lu m CHLOROPHYTA (gr een alga e ) Class CHLOROPHYCEAE Order SIPHONALES Fami ly CODIACEAE 15 Orto nella Penic i l l us Halim eda Order DASYCLADALES Fami ly DASYCLADACEAE 30 A cet ab ularia Cy m op o lia Das y cl 따 us Mi zz ia Neomeris ph y lu m CHAROPHYTA Class CHAROPHYCEAE Order CHARALES 15 Chara Trochil isc us
Cryp toz oon pr ol i (erum 에서 비롯되어 그후 이들에 대한 분류는 생물학적인 것보다는 기하학적인 형태, 죽 체내외의 형태에 의해 실시되어야 한다는 의견이 지배적이다. 그러나 s t roma t o lit e 의 분류는 현재 뚜렷한 체계가 채 택되지 않은 상태에 있다. 4 • 3 • 1 골격 질 석 회 조 ( Skeleta l Calcareous Alg a e ) 1 ) 남조식물 (Cy an op h y ta) 선캄브리아대 이후 남조류는 가장 혼한 해성 및 비해성 조류이지만 극 히 일부의 종류만이 생화학적 침전물을 남간다. 개체들은 모두 현미경 관 찰 대상이며 접액질 껍질 속에 자리잡고 있다. 많은 종류들이 미세한 퇴 적물을 집적시켜 스트로마톨라이트와 같은 박충리 구조를 만든다. 대부분 의 화석종들은 형태가 너무 단순하기 때문에 분류하기가 어려우며 가장 혼한 화석은 G i rvanella 속으로 분철되지 않은 10~50µm 의 반경을 가지 며 캄브리아 전기에서 후기 백악기까지 생존하였다. 絲狀의 골격질 남조 류는 다른 물체에 외피를 형성하여 결핵체상이나 기둥모양으로 발전한다.
 ~5:.•#;.\§. •- .,:...: .•::_ 따?소 •. : ·.. ?:.'· :`;;': · ,' , 출,:·•e . :.•. s;.. I• ,%\ •: t¢.. .. . • · .•I、., ., 心•군 ·.\·.:. x•• .요,`, %,..、E\ t; .& \\전. •.칼 •步••.•?. %..\ 수` ., `:. _다• 龜: 론.-··?;.`·r?: • ; ' f_..모'. :. .:앗.:..`. · .호 · · ..효•정,합&.f'·?요 ·;;'k건:t;. ;;:`\ . ;:;• :, .'V•.:.:: ·.·;'. 홍:· · •. ` ·감:나 소:. 홍·`•'’.,홍•나,. 슈'.암., . .. :;.' ...걸;..::· .:;.;.? : .、·..k.~; \ :요·\ ,정 :,.'\: • ; :· `엇:-:정o.:S혼 \F0· . 스, ._`` ·:·· .
~5:.•#;.\§. •- .,:...: .•::_ 따?소 •. : ·.. ?:.'· :`;;': · ,' , 출,:·•e . :.•. s;.. I• ,%\ •: t¢.. .. . • · .•I、., ., 心•군 ·.\·.:. x•• .요,`, %,..、E\ t; .& \\전. •.칼 •步••.•?. %..\ 수` ., `:. _다• 龜: 론.-··?;.`·r?: • ; ' f_..모'. :. .:앗.:..`. · .호 · · ..효•정,합&.f'·?요 ·;;'k건:t;. ;;:`\ . ;:;• :, .'V•.:.:: ·.·;'. 홍:· · •. ` ·감:나 소:. 홍·`•'’.,홍•나,. 슈'.암., . .. :;.' ...걸;..::· .:;.;.? : .、·..k.~; \ :요·\ ,정 :,.'\: • ; :· `엇:-:정o.:S혼 \F0· . 스, ._`` ·:·· .
2 ) 홍조석물 (Rhodoph yt a) 대부분의 홍조류는 독특한 내부 구조를 가지며 세포벽 안쪽 혹은 세포 벽돌 사이에 상당한 양의 석회분이 침전되어 있어서 세포의 구조와 생식기 관 둥이 잘 보존되어 있다. 화석 석회조는 세 가지로 분류되는데 (D Coralli na ceae, ® Solenopo r aceae 그리 고 ® `‘선조형 coral li nes’' 동의 그
룹들이 있다. Cor 색ina ceae 科는 쥬라기에 출현하여 현존하는 대 1 부분의 석회조를 포함 한다. 전동적으로 이들은 성장형에 따라 ®외피를 만드는 형태와 홍 )ar ti culat e d-se gm ent e d 형 태로 구분해 왔다. 현생종의 분류는 형태상의 득칭으로 일곱 개의 아과(亞科)로 나뉘나 이것이 그대로 화석종에도 적용 가능한지는 불분명하다. 따라서 재래적인 분류가 보존이 불량한 화석그룹에서는 편리한 것 같다. 외피형 그룹은 다양한 성장형을 가져 박충리상 皮型, 결핵체상, 대단히 불규칙한 괴상 및 분지형 둥 다양하다. 대부분은 수 mm 에서 수 cm 에 이르며 저질에 단단히 부착되어 있거나 간혹 해저에 자유롭게 성장하기도 한다 (그림 2,4) .한편 체질형 그룹은 5~20cm 의 키로 직립하며 분절되어 있다 (그림 3,5) . 외피형 조류는 초를 형성하는데 시멘트 역할을 하기도 하며 반 대로 이돌은 쇄설물로 붕괴되기도 한다. 섬세한 체질형 조류는 퇴적될 때 모래 크기의 파쇄 울로 세립화되는 것이 보통이다.
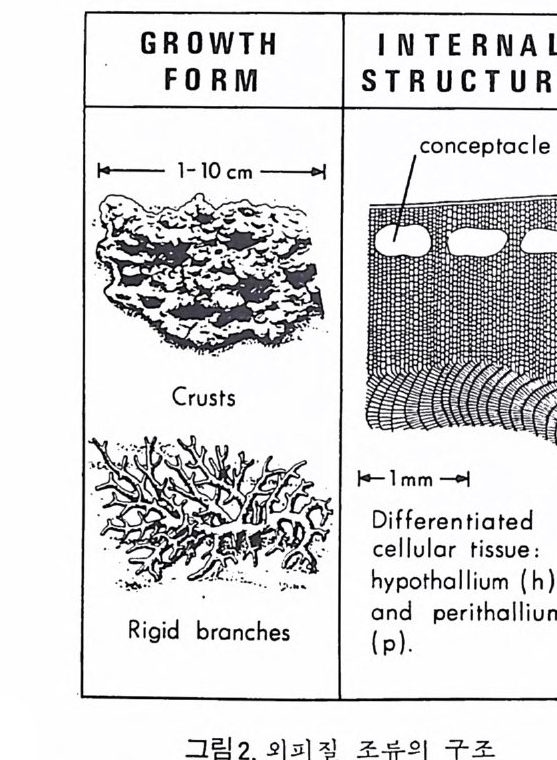 GR OWTH INTERNAL
GR OWTH INTERNAL
 그림 3 . 체정 형 조류 의 구 조
그림 3 . 체정 형 조류 의 구 조
Soleno p orac e aeH 는 후기 캄브리아기에서 팔레오世까지 생존한 석회조류 로서 외피형, 원형의 결핵체를 이루는 수 mm 에서 수 cm 의 크기를 가진다. 세포의 구조는 한 종류의 조직형을 가지며 생석기관을 갖추지 못했다. 屬 수준의 주요 특칭은 수직의 세포로 이루어전 줄내에서 수평층 구조나 분
 00 µ
00 µ
할 상태 둥이 포함된다 (그림 6 참조). A11eestr a l . cor al. l ine s 은 후기 고생대에 출현해 펜실바니안紀에 혼한 종류 로서 현생 Coral lin aceae 그룹과 내부 구조가 유사하기 때문에 〈선조형〉이란 표현이 붙게 되었다. 이들은 보통 외피형과 수직 체철형의 형태를 가지며 석회암의 중요한 구성원이다. 미씨씨피안紀에서 중기 페름紀에 생존했던 Archaeolith o p hy ll um 은 현재 살고 속인 L it ho phy llum 과 유사한 세 포 구 조를 갖고 있다 (그 림 7 참조).
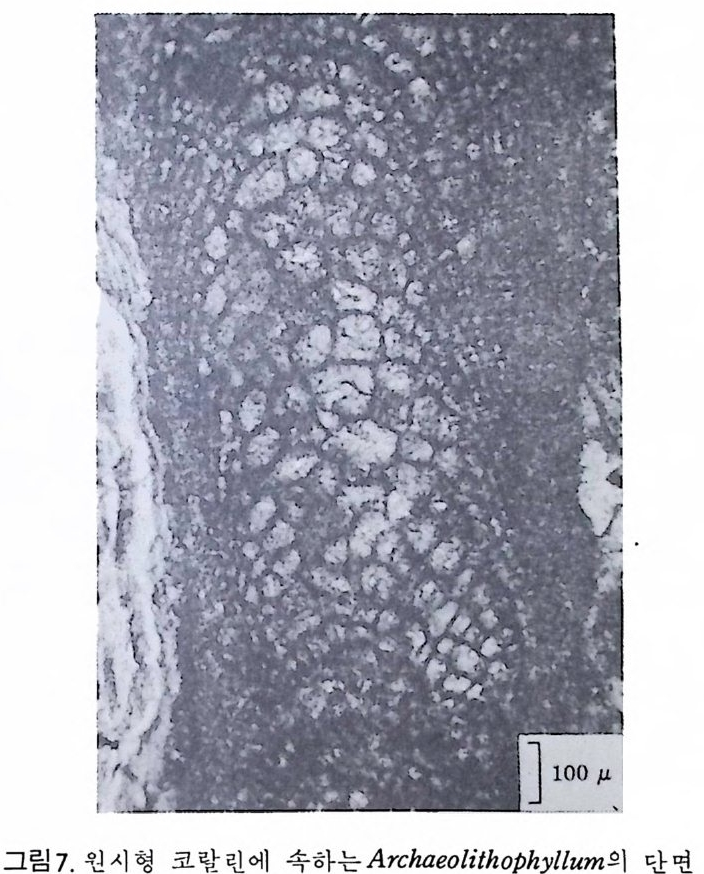 그림 7. 원시형 코탈란에 속하는 Archaeol ith o p hyllum 의 단연
그림 7. 원시형 코탈란에 속하는 Archaeol ith o p hyllum 의 단연
3 ) 녹조석물 (Chloro phyt a) 탄산영 물질을 침전시키는 바다의 녹조류는 C ocli aceae 과 및 Dasyc l ada- ceae 科 둥 두 종류가 있다. 탄산영 광물질의 성분이 다양해서 영상체는 애라거나이트의 침으로 되어 있고 영상체의 주형은 CaC 야의 침전물로 형
성된다. 내부 사상체와 小 1L 의 구조 외에 각각의 분절의 형태를 기준으로 분류를 실시한다. 열대와 아열대에 서식하는 Pen i c i llus 와 같은 녹조류의 미세한 광물질 관은 이 지역 해처 퇴적물의 주요 성분을 이루기도 한다. Cod i aceae 科에 속하는 종류들은 두 가지의 성장형태를 갖는데 외피형 혹은 결핵체형 아니면 직립분철형이 그것이다(그림 8). 결핵체형은 보 동 고생대에 출현하고 중생대 및 신생대에는 수 cm 높이의 직립 분절 및 분지형의 골격에 튜브상의 사상체들이 엮어져 있다(그림 9 참조). Das y cladaceaeH 는 칙립, 분절, 분지형이며 수 cm 높이까지 성장한다. 대부분의 종들은 대형의 中央枝와 이를 둘러싸고 있는 작은 방사상 가지 의 다발로 이루어지며 대부분의 화석종들의 경우 속이 비어 있는 원통 혹 은 공모양의 구조를 가전다(그림 10,11 참조). 생식기관인 s po r amgi a 는 줄기 나 가지 옆에 달려 있으며 석회화되어 보존된다. 이 그룹의 분류는 석묻 전체의 윤곽, 분절의 종류, 가지의 배열과 갯수, 그리고 생식기관의모양 과 위치에 의하여 실시한다.
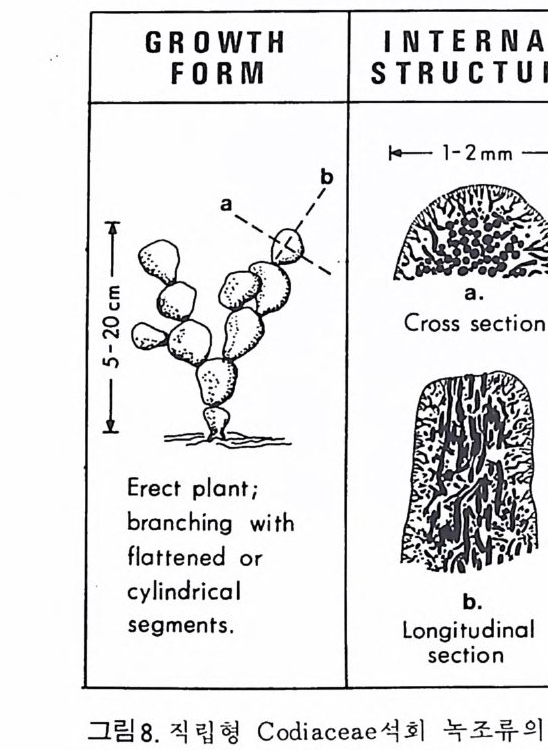 GR O WTH INTERNAL
GR O WTH INTERNAL
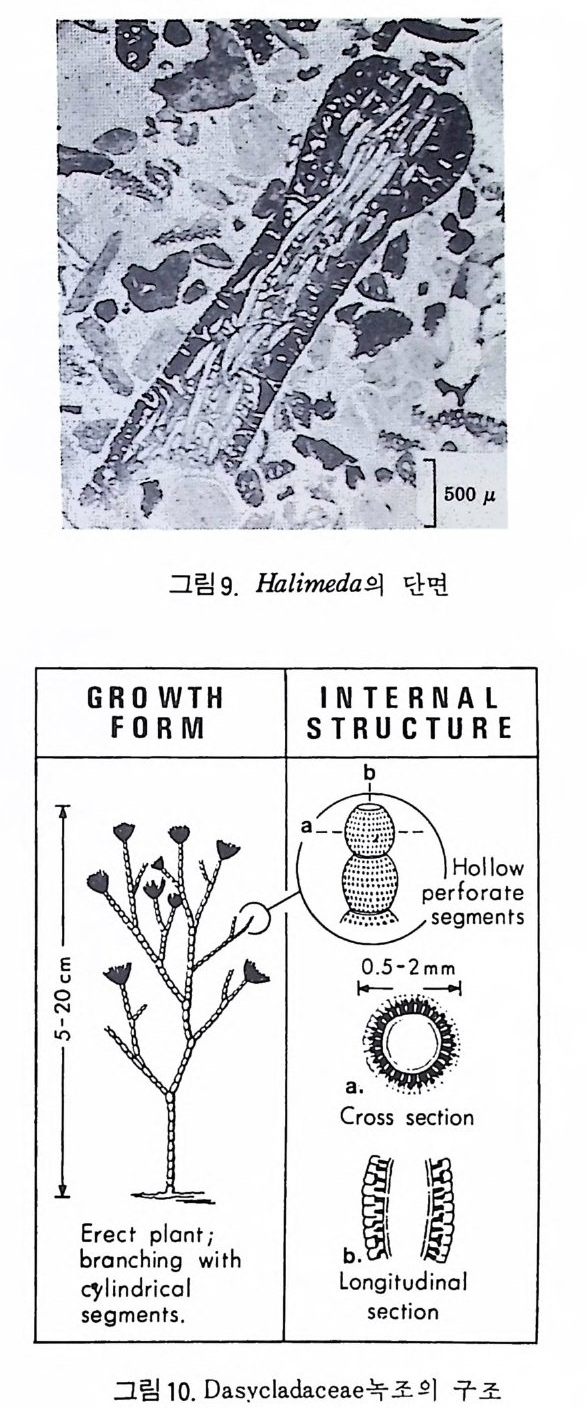 그림 9. Hal im eda 의 단면
그림 9. Hal im eda 의 단면
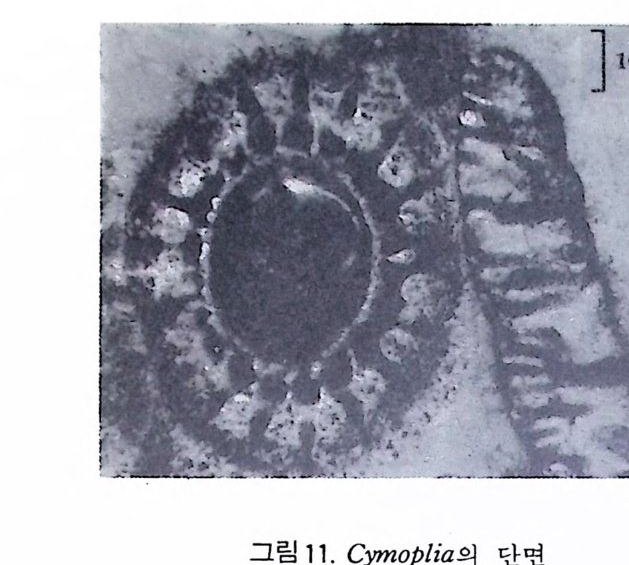 그림 11. Cy m o p lia 의 단면
그림 11. Cy m o p lia 의 단면
4 ) Charoph yt a 카로피타는 생리적이나 생화학적으로 녹조와 유사하지만 그 형태가 전 혀 다르기 때문에 독립 된 문을 이룬다. 크기 는 수 cm 에 서 l m 에 이르며 습 성과 외부의 구조는 고동식물과 비슷하다. 대부분의 종류는 gyr og onite 또 는 oo go n i um 이라고 불리는 생식기관만이 석회질화되어 화석으로 산출된 다. 이들의 크기는 0.5~ 1mm 이며 와상으로 튜브가 감겨 전체적으로는 타
 `-· , · -之姿〉국~- -· ,,,, _ .
`-· , · -之姿〉국~- -· ,,,, _ .
 그림 13. Charo p h yta의 석회화된 생식기관
그림 13. Charo p h yta의 석회화된 생식기관
원체를 이룬다. 현생 charo p h yt es 는 대부분의 해양 저생성 석회조류오R 근 달리 담수와 기수 환경에 서식한다. 그러나 이들은 하천에 의해 운반되어 연해 퇴적물에 유입될 수도 있다•. 화석으로 후기 실루리아기 이후부터 발 견된다(그림 12,13,14 참조). 4 • 3 • 2 St ro mato lite 조류성 스트로마톨라이트 (s t roma t ol it e) 는 수 mm 에서부터 대규모의 생물 퇴적 구조까지의 다양한 크기를 갖는다. 현미경적인 남조류가 미세한 탄 산염 질퇴 적물을 포획해서 뚜렷한 박충리 구조를 만들지만 조류 자체는 석 회질화되지 않는다. 그들의 외형은 대단히 다양해서 판상 박충리, 도움 상, 기둥상 등이 있고 어떤 것은 가지를 갖거나 손가락모양을 하기도 한 다(그림 15 참조).
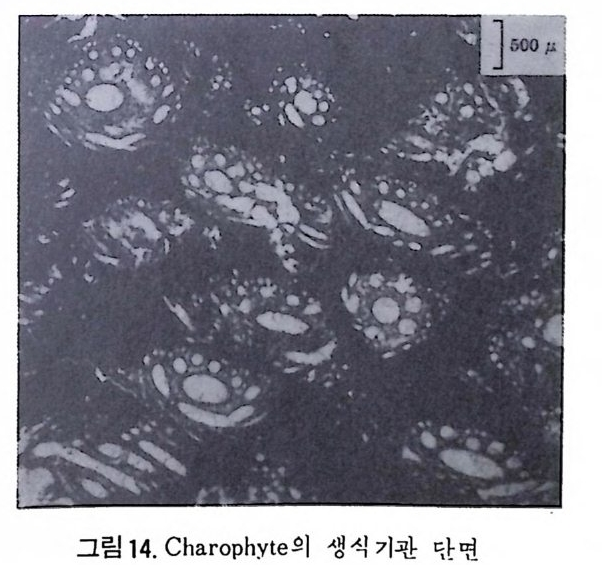 -`•P'• -· . 5`.`,.` . .` ;\}...,.'. . i、·..、`‘b.j ‘.-8ig 0 .-'r·』'V\ `.g s--:.* 1` ._.\、잃 亨- • .oO를 .`.J O ,
-`•P'• -· . 5`.`,.` . .` ;\}...,.'. . i、·..、`‘b.j ‘.-8ig 0 .-'r·』'V\ `.g s--:.* 1` ._.\、잃 亨- • .oO를 .`.J O ,
 '\,~ --
'\,~ --
Oncol it e 란 동심원상으로 형성된 박충리를 말하며 대부분의 스트로마톨-라 이트는 0.5~ 1mm 의 박충리로 이루어 진 수 cm 의 크기를 가진다. 현촌하는 조류 스트로마톨라이트에 관한 연구는 이들의 생물학적 현상과 물리적인 환경 변화와의 관계를 밝히는데 크게 공헌하였다. 기하학 적인 형 태를 기준으로 한 분류가 이들과 같은 비골격질 석회조류에 맞 는방 법인것 으로 보인다. 4 • 4 생태와 고생태 다음과 같은 현생조류의 그룹과 생태 환경과의 관계에 대한 정보가 화석 석회조류와 고환경을 추정하는데 도움을 준다. 녹홍카남〈그조조조로룹류류류피〉 타 ————해해해담 성성성 수 /::또 〈파초바환는 질해경 혹 성기>혹은 수 : 은 성암점 석토접토질질저질저 질질저 질 그림 16 은 신생대 조류의 대륙붕 주변의 환경에서의 분포 양상이다. 카 로피타는 비해수에서 성장해서 석회질 물질은 호수에 대량 퇴적된다.녹조 는 여러 환경에서 s t roma t o lit e 를 형성하지만 대부분은 해양 분지나 영호 의 얕은 연변부에 국한된다. 심해에서 생성되는 스트로마톨라이트와 다수의 담수종이 역시 보고되었다. 골격질 석회 녹조류는 대부분이 천수의 석호에서 성장하여 그지역 되적
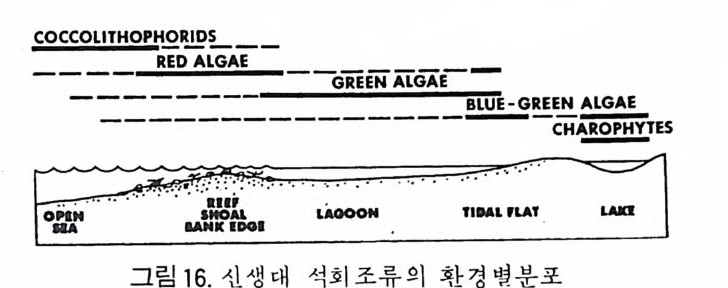 ·CO -C·CO·L IT-, H.O P,-HR,OE- DR IAD· 'LS:-G A :-E'’, •:- ,. GREEN A_L GA-E ' :- -CH-A-ROP-HYTE S
·CO -C·CO·L IT-, H.O P,-HR,OE- DR IAD· 'LS:-G A :-E'’, •:- ,. GREEN A_L GA-E ' :- -CH-A-ROP-HYTE S
물의 중요한 성분이 된다. 외피형 코랄린 홍조류는 해수면에서부터 230m 수심까지 서석하지만 주로 암석질 저질과 應性 환경에 분포한다. 외피질 형과 분김 형 코탈란 그룹은 노출된 고에너지 환경에서도 생존한다. 석회 질 부유성 조류인 해조류 는 성해저의 주요 퇴적물 성분이지만 대륙봉 지 역과 연안 지역에서도 역시 퇴 적된다 . 수십과 위도별 분포는 속이나 종에 따라 대단히 제한돼 있다.그러나 아 직 현생 조류에 대한 생태학적 선호도가 충분히 연구되어 있지 않기 때문 에 화석종을 가지고 고생태 환경을 추적하기가 어렵다. 석회질 해조류의 지리적 분포는 대체로 다음과 같다. 〈 그룹 〉 〈지리적 분포〉 S t roma t ol it e 를 포함하는 남조류 열대-국지방 외포 1 형 코랄린 조류 열대—극지방 분철형 코랄란 조류 열대一한냉一온난 C cxli acean 녹조류 열대 — 아열대 D as y cladacean 녹조류 열대 ―온난대 Ade y (1970) 는 북대서양의 외피형 코랄린 조류의 분포에 관한 연구를통 해 위도에 따른 수온의 번화, 깊이에 따른 수십의 변화가 중요한 환경요 소임을 밝혔다(그림 17 참조). 또한 데본魚란의 석회질 確를 형성하는 석회조류를 연구한 Wray 0972) 는 환경에 따라 독특한 그룹이 산출됨을 보고했다. 그러나 이 종류들이 현 재는 모두 절멸되었기 때문에 현생종들과의 관계는 불분명하다. 그림 18 에서 우리는 특정한 그룹의 석회조류에 의해 퇴적상이 추적될 수 있는 가능성을 볼 수 있다. 따라서 이러한 자료는 적어도 다론 저생성 화
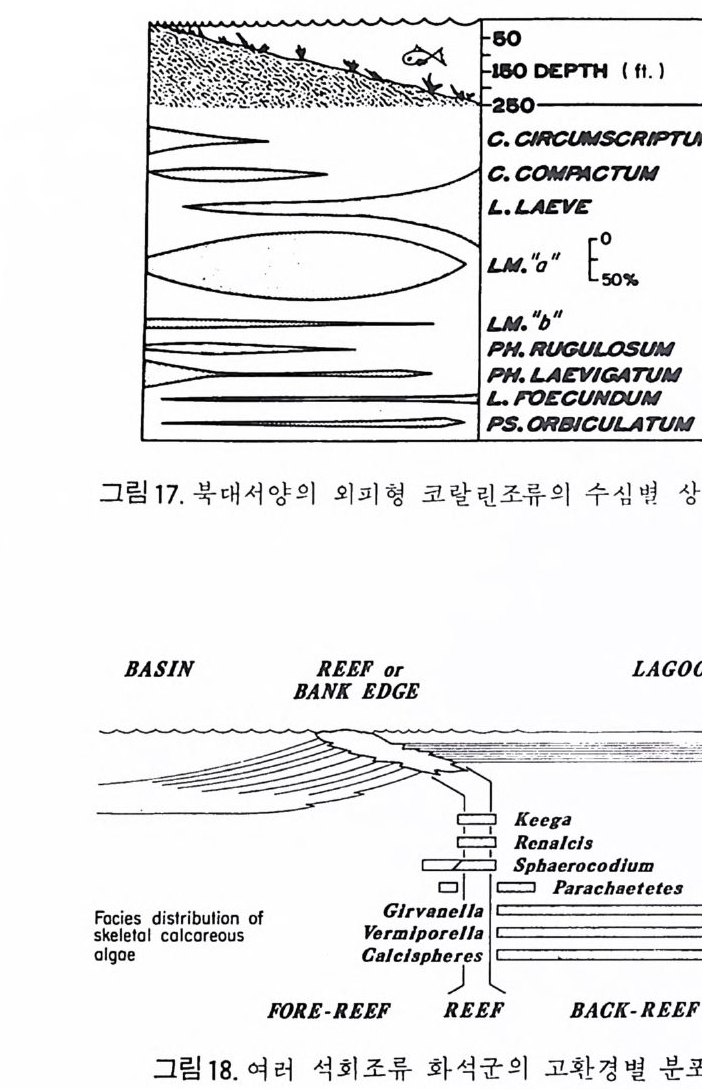 so
so
석군의 고생태학적 환겅 추적을 보완해 주는 역할을 한다. 4 ·5 생물충서 저생 석회조류의 주요 그룹들은 긴 생존기간을 갖기 때문에 생물충서학 적 응용이 상당한 제약을 받는다. 십한 경우 어떤 원시적인 남조류는 수 백만년 동안 거의 번화가 없이 생존한 예도 있다. 또한 난류성 석회조류와 는 대조적으로 저생종은 좁은 서식지에만 분포함으로써 광역적인 충서대
비가 어렵다 . Wray (1 971) 는 현재와 과거 지질시대에 초의 형성에 중요한 역할울한 주요그룹의 충서적 범위를 정리하였다. 그림 19 와 20 은 주요 속 들의 생존 시대를 표시한 것으로 지질시대별 주요 군집을 인식할 수 있다. 이러한 시간에 따른 산출 순서를 보면 C cxli aceae 와 Das y cladaceae 의 두 과는 아마도 초기 캄브리아기나 선캄브리아대에 공통 조상형으로부터 유래 된 것 으로 추측된다. Soleno poraceae 는 중기 고생 대 에 Ancestr a l corallin e 으로부터 전화한 것으로 보인다. 또한 Corall i naceae 의 선조형은 불분명 하지 만 Solenopo r aceae, Ancestr a l corallin e 혹은 다론 원시 형 홍조류로부터 유래된 것으로 보인다.
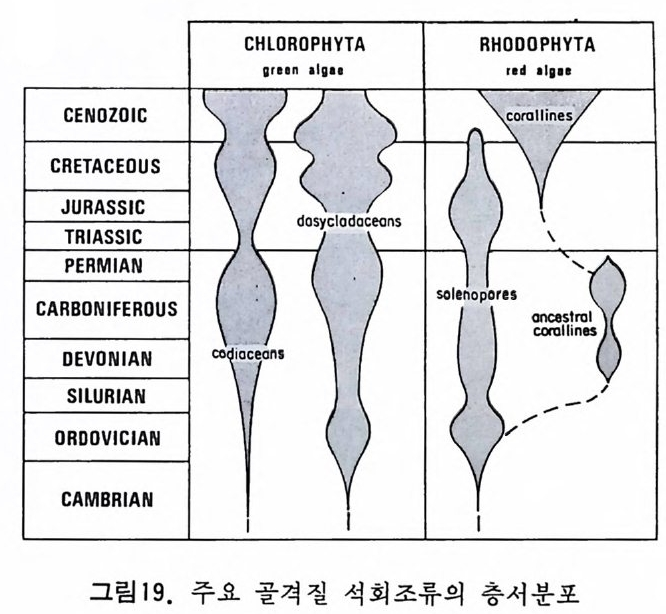 CHLOROPHYTA RHOOOPHYTA
CHLOROPHYTA RHOOOPHYTA
보충교재 Daw_s on , E. Y., 1966. Marin e Bota n y : An Intr o ducti on . Holt , Rin e hart and Wi ns to n, New York, N. Y., 371 pp. [T ext- b ook overv iew of ph ys io l og y, ec- olog y and geo g ra ph ic dis t r i b u ti on of marin e pla nt s; emp h asis on bent hi c alga e . Includes a brie f chap te r on calcareous alga e.] Fritc h , F. E., 1935-1945. The St ru ctu re a 뼈 Rep r oducti on of the Alga e .Ca-
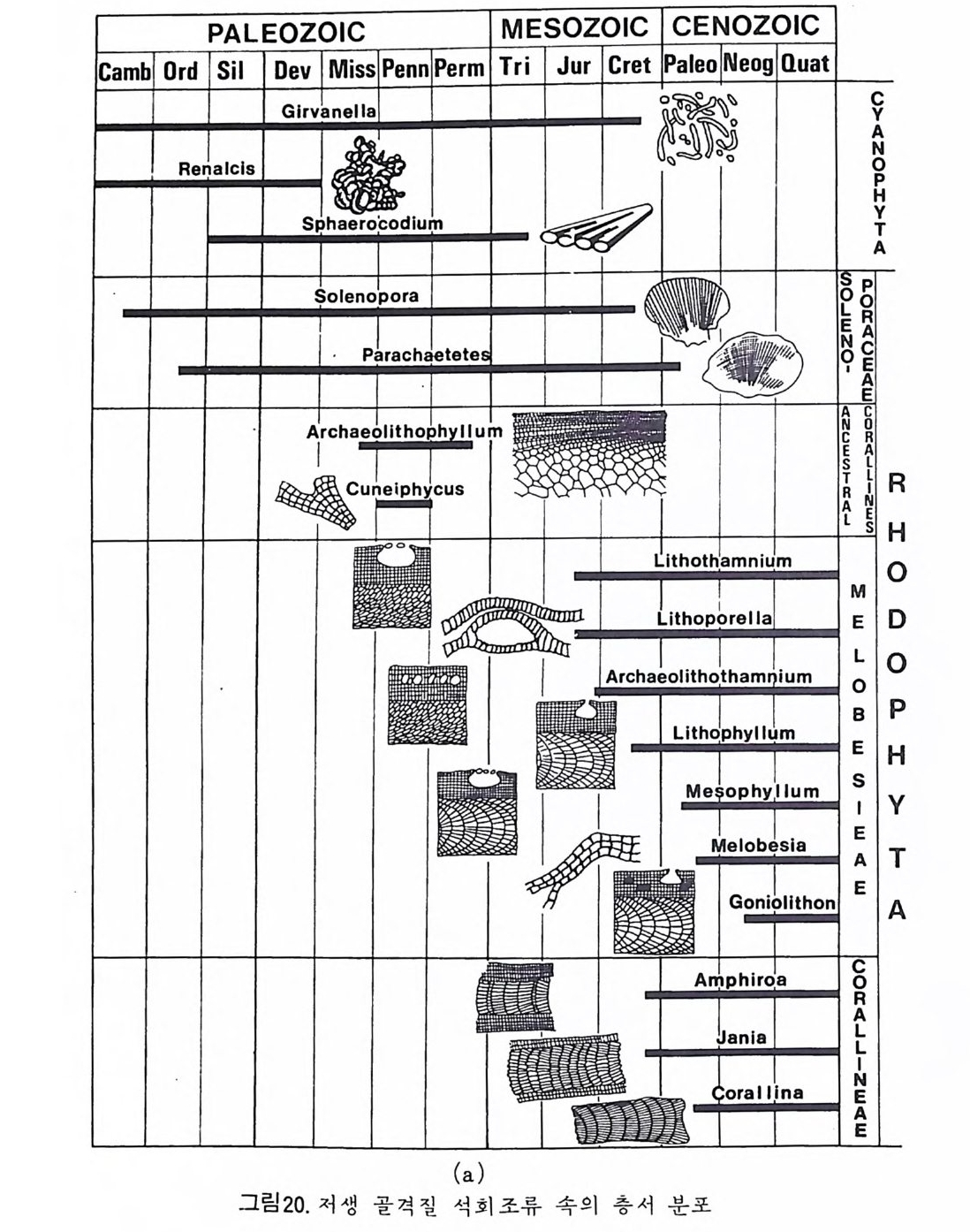 Cambi ORrde riIl Pa S lAci ll Ls |I D0GeZv VII', a M lC i s sH드|! Penn Il P e— 『 rn ~TE 『iS OJuZ r OICI 『C e t 邑l°C렇PE焉N핫O Z~OIC t CpHYY ATNAO
Cambi ORrde riIl Pa S lAci ll Ls |I D0GeZv VII', a M lC i s sH드|! Penn Il P e— 『 rn ~TE 『iS OJuZ r OICI 『C e t 邑l°C렇PE焉N핫O Z~OIC t CpHYY ATNAO
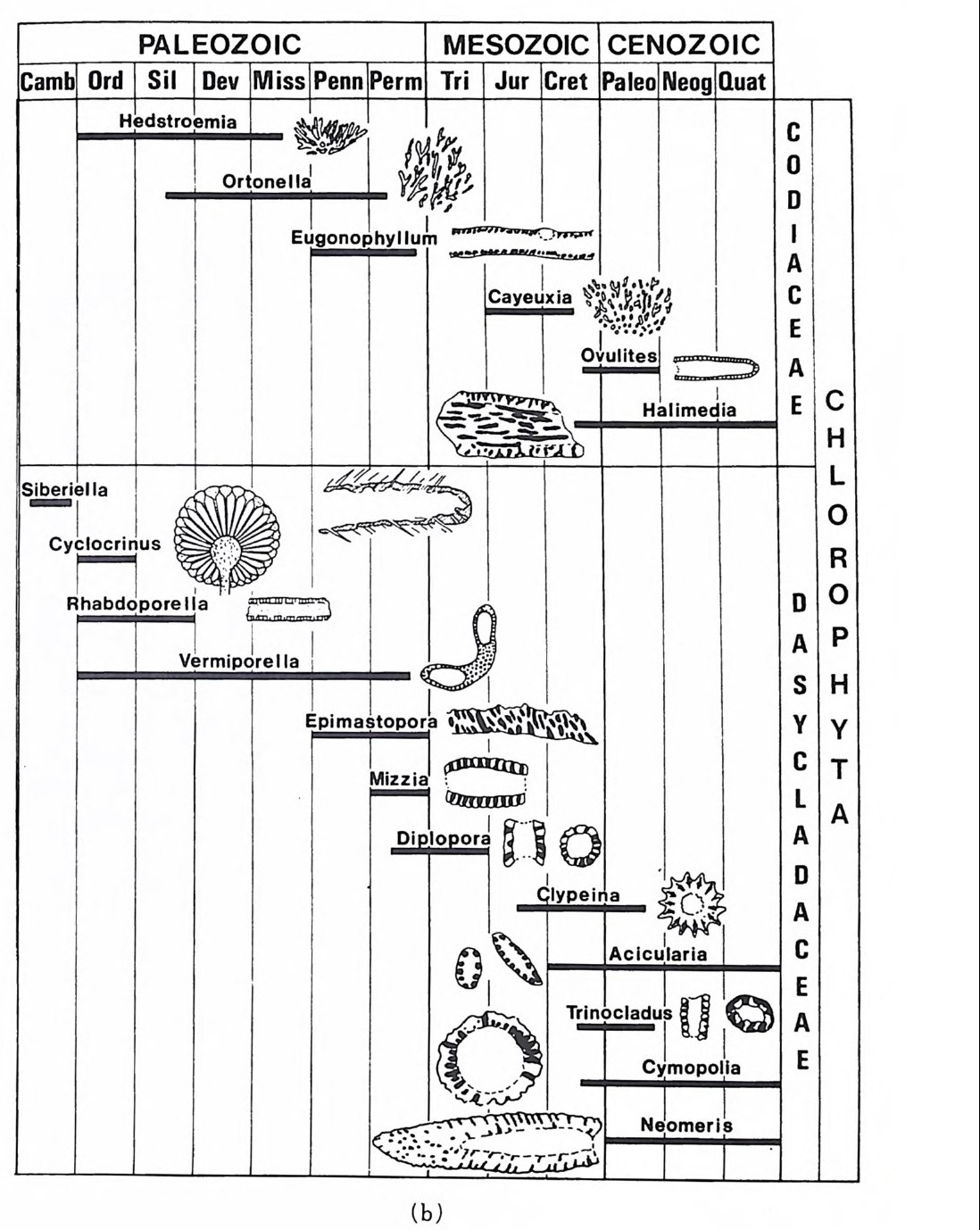 PALEOZOIC MEISOZIOlC IC ENI OZ I IC
PALEOZOIC MEISOZIOlC IC ENI OZ I IC
mbrid g e Univ ersit y Press, London, Vol. I (19 35) , 791 pp. : Vol. 2 (19 45) , 939 pp. [M a;i si v e l \',O volume comp ila ti on of world's lit er atu re on morph olog y and rep r oducti on of alga e. St ill a usefu l and in clusiv e refe rence work on as- pe ct s of th ese subje ct s; obvio u sly defi ci e n t in modern advances in ph y s io l og y and classif ica ti on .) Gi ns burg, R., Rezak, R. and Wray, J. L., 1971 . Geolog y of Calcareous Al- ga e: Not e s (or a Short Course. Univ ersit y of Mi am i , School of Marin e .an d At m osp h eric Scie n ce, M iam i , Fla., 64 pp. [U p -to -date summ ary ( il l ust - rate d out line fo rmat ) of vario u s asp e ct s of calcareous alg a e. Emp h asis on th e ir use in env iro nment al in te rpr eta ti on of carbonate sedim ent s .] Jo hnson, J. H., 1961 . Li m est o ne-Buil d in g Alga e and Alga l Li m esto nes. Colo- rado school of Mi n es, Golden, Colo., 297 pp. [D escrip tion s, ill ust r ati on s,g e - olog ic rang e s and ge og r ap h ic dis t r ib u t ion of im p o rta n t ge nera of fos sil and ge og r aph ic dis t rib u t ion of im p o rta nt ge nera of fo s sil and recent calcareous alga e. Also chap te rs on clas sif ica ti on , chemi ca l com p o s it ion and ecolo g y.] Walte r, M. R. (Edit o r) , 1976. St r omato lit es . Else vie r, Amste rdam, 790 pp. (Comp ila ti on of 43 st u die s on all asp e ct s of st ro mato li te s; met h odolog y_, sy - ste m ati cs , bio l og y, bio s t r ati gr ap h y , and th eir use in pa leoenvir o nment a l and basin analys is . ] Wray, J. L., 1977. Calcareous Alga e. Elsev ier , Ams te rdam , 185 pp. [C om p - rehensiv e illu st r ate d tex t -r efe rence coverin g majo r gr oup s of . marin e and non-marin e calcareous alga e, Des cr ip tion s , classif ica ti on , and st r a ti gr ap h ic and envir o nment al dis t r ib u t ion ; also chap ter s on pa leoecolog y, sedim ent -pr o- ducin g alga e and alga l fac ie s in tim e.] 참고문헌 Adey, W. H., 1966. The dis tr ibu t ion of saxic o lous crusto se corall in e s in th e north weste rn North At Iant ic. J. Phy c ol., 2 : 49-54 . Adey , W. H., 1970. The ef fec t s of lig h t and te mp e ratu re on gr owt h rate s in boreal-subarcti c crusto se corallin es. J. Phy c ol., 6 : 269 -2 76. Adey, W. H. and Jo hansen, H. W., 1972. Morph olog y and ta xonomy of Cora- llin a ceae wi th sp e cia l refe rence to Clath romorph um, Mesop h y l lum and Neo- po lyp o rolit ho n ge n. nov. (Rhodop h y c eae, Cryp to nemi al es). Phy c olog ia, 11:159 _ l80. Adey, W._H . and Macin t y r e, I. G.,· 1973. Crusto se corallin e alga e:a r.e -evalu- Arantoi otnt , inH . thJ. e gane odl oPga icua tl a rsdc.,i e n Fc.e Gs.. KG,e o1l,9 70S.o c.C aAlcmif .i cBa util ol.n, i8n4 p:l a8n8t 3s .- I9n0 4: .H . Sc- hrader (Ed it or ) , Bi o log ica l Calcif ica ti on : Cellular and Molecular A sp e ct s .
Ap ple t on -Cent u ry- Croft s, New York, N. Y., pp. 375-446. Cloud, P. E. and Semi kh ato v , M. A., 1969. Prote r ozoic str o mato l it e zonati on ; Am. J. S ci. , 267 : 1017 -1 061 . Davi es , G. R., 1970. Alga l-lami na te d sedim ent s, w_e ste r n Austr a lia . In : B.W. Log a n, G. R. Davie s , J. F. Read and D. E. Cebulski, Carbonate Sedim ent- ati on a 뼈 Envir o nment s, Shark Bay, Weste rn Austr a lia . Am. Assoc. Pet. Geol. Mem., 13 : 169 -2 05. Garrett , P., 1970. Phanerozoic str o mato l it es : noncomp e ti tive ecolog ic rest r ic - tion by gr azin g and burrowi ng anim als. Scie n ce, 169 : 171-173. Gebelein , C. D., 1969. Di st r ib u t ion , morph olog y, and accreti on rate of Rece-nt sub ti da l alga l st r om ato l it es , Bermuda. J. Sedim ent. Petr o l., 39 : 49-69. Hof m ann, H. J., 1969. At tri b u t es of str o mato l it e s. Can. Geol. Surv. Pap. 6 9 一 39 : 58 pp. Hof m ann, H. J., 1973. S t romato l it es : characte ris t i cs and uti lit y . Earth - Sci. Re v. , 9 : 339 -3 73. Joh ns on , J. H., 1943. Geolog ic im p o rta n ce of calcareous alga e wi th annot a te d bib l io g r ap h y . Colo. School Mi ne s Q. , 38(1) : 102 pp. Joh n s on , J. H., 1957. Bib l io g r ap h y of fos sil alga e: 1942-1955. Colo .. School Mi n e s Q. , 52(2 ) : 92 pp. Jo hns on , J. H., 1967. Bi bl io g r ap h y of fo ssil alga e, alga l lim esto n es, and the ge olo g ica l wo rk of alga e , 1956-1965. Colo. School Mi ne s Q., 62(4): 148 pp. Kumme l, B. and Raup , D., 1965. Ha 뼈 book of Paleont ol og ica l Techniq ,ue s, W. H. Free m an and Co., San Francis c o, Calif. , 852 pp. Le m oin e , M., 1911 . St ru ctu re anato m i qu es des Melobesie e s. Ap pli c a ti on ii la classi f ica ti on . Inst. Oceanog r . Monaco Ann., 2(1 ) : 215 pp. Le w i n, J. C., 1962. Calcif ica ti on . In : R. A. Lewi n (Edit or ) , Phy s io l og y and Bi oc hemi st r y of Alga e. Academi c Press, New York, N. Y., pp. 457-465. Log a n, B. W., 1961 . Cryp to zoon and associa t e d str o mato l it es from the rec- en t, Shark Bay, Wes te r n Aust ra lia . J. Geol., 69 : 517-533. Log a n, B. W., Hof fm an, P. and Gebelein , C. D., 1974. Alga l mats , cryp tal - ga l fab ric s , and str u ctu res. In : B. W. Log a n, J. F. Read, G. M. Hag a n, P. Hof fm an, R. G. Brown, P. J. Woods and C. D. Gebelein , Evoluti on a 뼈 Di a- ge nesis of Qu ate rnary Carbonate Seq, u ences, Shark Bay, Weste m Austr a -lia . Am. Assoc. Pet. Geol. Mem., 22 : 358 pp. Maslov, V. P., •1 956. lskop a emy e isv estk o v ye .v.o dorosli S. S. S. R. (Fossil calcareous alga e of the U. S. S. R.) .Akad. Nauk S. S. S. R., Tr. Inst. Geol. Nauk, 160 : 301 pp. Maslov, V. P. et al., 1963. Vodorosli ( Alga e) . ln : Yu. A. Orlov, Osnovy pa le- ont ol og ii. Moskva Akad. Nauk S. S. S. R., 14 : 19-312. Monty , C.L .V ., 1967. Di st ri b u t ion and str u ctu re of recent str o mato l it ic alg- al mats , easte rn Andros Island , Bahamas. Soc. Geo!. Bel(!. Ann., 90 : 55 -100.
N1•mcjc , F., 1959. Paleobot a nik a . Praha, Nakladalest v i Cesk. Akad. Ved, 1 : 402 pp. Pi a, J., 1920. Di e Sip h oneae verti ci l la t ae vom Karbon bis zu r Krei d e . Zoo!. Bot . Ges. W ien Abh., 11(2). Rid i n g , R. , 1975. Gi rv anella and oth er alga e as dep th ind ic a to rs. Leth aia , 8 : 17:: l-1 79. St oc kman, K. W ., Gi ns b urg, R. N. and Shin n , E. A., 1967. The> pr od u c ti on of lim e mud by alga e in sout h Florid a . J. Sedim ent. Pet rol. , 37 : 633 -6 48. Surdam, R.C.and Wray, J.L ., 1976. Lacust r in e st ro mato li t c s, Eoc e ne Gre- en Ri \'e r Formati on , W yo mi ng . In:M. R. Walte r(E dit o r ) , St ro mat o li tes .El- se\· ie r , • Ams t e r dam, pp. 535-541 . Wray, J. L., 1971 . Alga e in reefs th rou g h tim e. North Am. Paleont o l. Conv., Chic ag o , 1969, Proc ., J : 1358 -1 373. W ray , J. L., 1972. Envir o nme n ta l dis t rib u t ion of calc a r e o us alg a e in Up pe r DL'vonia n ree f comp l ex es. Geo!. Rundsch., 61 : 578 -584. 5 放散虫 (R adio l aria ) 5 • 1 서 언
현생이언의 전기간 동안 방산충은 어느 미화석군보다 도 완벽한 보존과 분포윤 보인다 . 세포막에 의해 분할왼 내부 • 외부의 구조 로 다른 원생동 물들과 분별이 된다. 또한 이 세 포막 의 여러가지 특징에 의해 더 욱 세분 된다 . 그러나 골격부의 성분과 기하학적 형태의 차이가 이러한 세 보 학적 인 상호관계윤 지시하므로 경 질부에 관한 연구만으로도 분류가 사능하나. 지질학자들은 방산층의 한 그룹인 p ol y c y s ti nes 웅 집중 적으 로 연구하있으 며 여기에 s p umellar i ans 와 nassellar i ans 가 포함된다 . 또 다 본 주요 그-;,;-은 t r ipy leans 이다. 이들의 대 표적인 종류가 그림 1 에 소개되어 있다 . 현생 방산충의 골격은 바정질 오 팔린실 리카로 구성되어 있으며 제 3 기 이 후의 화석의 성분이기도 하다 . 제 3 기 이전의 화석에서 가끔 다른 광물 로 치환된 표본들 이 발견되지만 방산층은 항상 규 질 의 성분을 고수했 다 고 믿어진다. 방산충과 밀접한 찬척인 acan t har i ans 는 셀레스타이트 성분을 가지는데 퇴적암에는 보존되지 못하는 풍랑크론이다 . 그들 은 완전히 대창적인 20 개 의 스카인을 가지는 정으로도 방산층과 구별된다. IIcl i oZ()an 도온 대칭적인 방 @o0 @}
@o0 @}
산 구조를 가지는 담수 원생동물로서 방산층과 비슷하지만 화석으로 보존 이 가능한 골격을 갖고 있지 않다. Ehrenber g는 최초로 방산층 을 광범위하게 연구한 사람이며 M ikr cg e olog ie (18 54~ 18.5 6 ) 에 많은 방산층 을 수록했 다. 19 세 기 중영 Muller 와 Hertw ig 둥은 방산 층 의 분류학적 위치와 내부 구 조를 연구하는 기 초 를 닦았다 . Haeckel 은 1887 년 H.M .S. Challen g er 호의 표본 으로 모노그라 프 를 작성했다. 1950 년대의 R i edel 은 십해저 연니로부터 육지의 여러 곳에서 방산층을 수집하여 Haeckel 의 분류체계를 재정리하 는 작업을 주도했으 며 Khabak ov 나 Lipm an 그리고 Pe t rushevska ya 등에 의해 새 로운 분류군 간의 관계나 계몽 이 확립되었다. 방산층 을 암석이나 퇴 적 물 시료로부터 추출하기 위해서 는 적절한 분해 약품(예 :Tc t r asod ium p:v ro p hos p ha t이 을 물 에 녹여 부은 뒤 끊이는 작업부 터 시작된다. H202 가 유기물 질을 산화시키며 거품을 일으켜서 기계 적으로 암석을 분리시키기 위해 첨 가된다. 그후 석회 질 물질을 제거하기 위해 HCI 이나 CH3COOH 를 이용하여 산처리를 시 킨 다. 물 속에서 분해되지 않는세 일과 같은 암석은 전조시킨 후 등유에 참 가서 완전히 침투시키고 다시 등 유를 매고 물에 담그는 방법 을 취한다. 62µm 의 채로 처리한 나머지 농축 물을 slid e g lass 에 키펫팅하여 Can a
동화작용 및 이화작용의 역할을 맡고 있다. 중앙캠술의 세포막은 종에 따라 다양한 색깔을 띠나 대체로 노랑이나갈 색이 혼하다. 세포막의 성분은 키틴질에 가깝고 여러 개의 구멍이 있어서 물을 교환하기도 하며 내형질로부터 뻗어 나온 僞尼의 통로이기도 하다 .
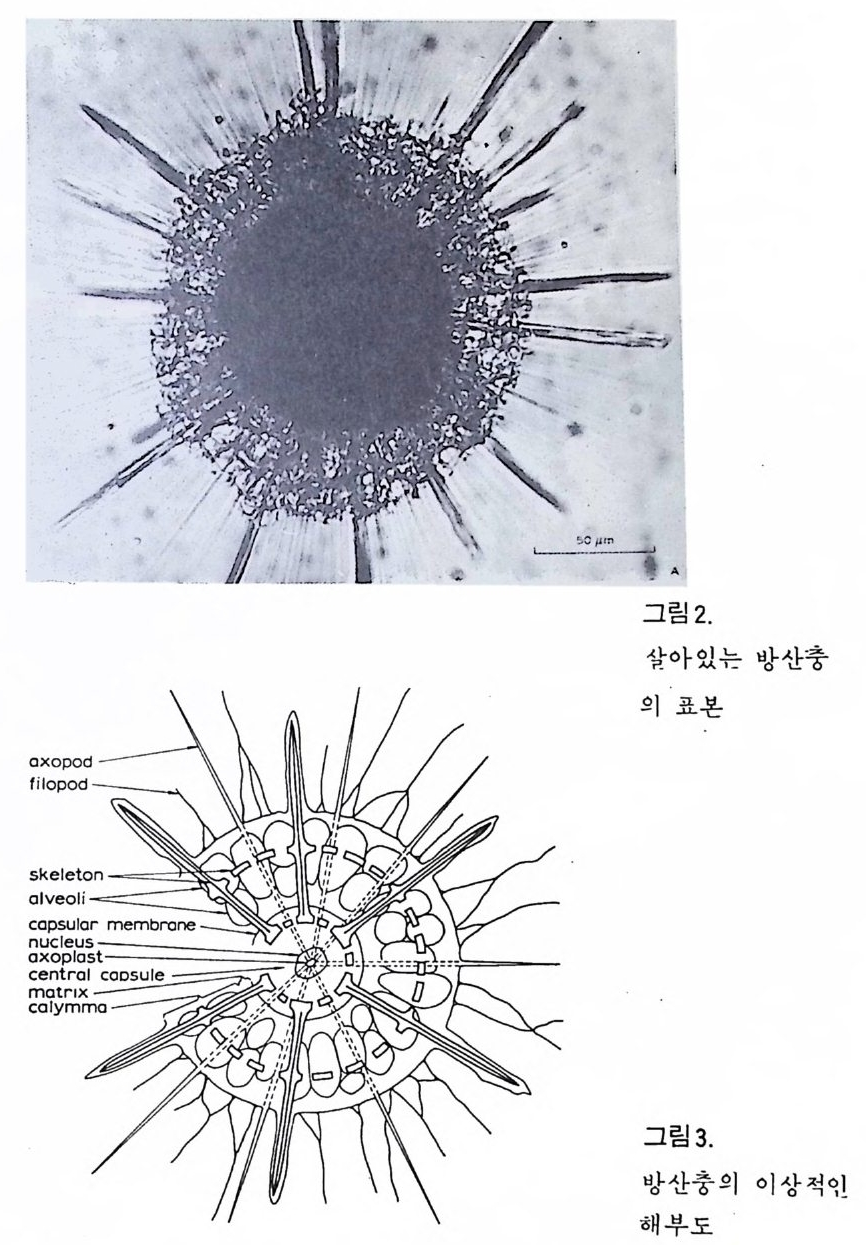 ' SC µm -’
' SC µm -’
외형질은 소위 페포 (alveol i)로 채워져 있으며 , 외부로 동십원상의 거품 질인 cal ym ma 안에 간신히 보이는 타원상의 구조를 이룬다. 이 부분은 수 압을조절하는기능을하는것으로생각된다. 외형질의 좁은부분인 ma t r i x 는 cal.vmma 와 중앙캠슐을 분리하는 역할을 한다. 외 형 질에 서는 가끔 소 화가 되는 과정의 먹이로 보이는 규조의 세포막이나 Zooxan t hellae 가 발견 되기도 한다. 방산층은 보통 두 종류의 위족을 보유하는데 하나는 내형질로부터 나와 서 방사상으로 뻗는 axo podi a 로서 중심의 a.x o p las t에 삽입되어 있다. Axop o- di a 와 axop las t의 형태는 분류에 대단히 중요하다. 얇고 섭세한 fi lo podi a 는 외연부의 세포질의 단순한 연장체이다. 이들은 방사상이며 뿌리부분은: 접 합되어 있는 경우도 있고 먹 아를 잡을 때는 한데 묶인다. 방산층의 생활사를 관찰하기는 매우 어려우나 대체로 단순한 세포분열 이 주를 이루는 것 같다. 유공충과 같이 유성과 무성생식의 과정을 겪으 며 일생은 수일~수개월인 것으로 믿어진다. 대부분 7 의 방산층은 골격을 갖는데 이 경질부는 세포질 내에 특히 거의 가 외형질에 싸여 있다. 따라서 겅절부는 해수와 직접 접촉하고 있지 않 기 때문에 살아 있는 방산층은 결코 용해되지 않는다 . 5 • 3 형태와 분류 대부읍본의 방산층은 경 질부를 보유하고 있으며 이는 연한 세포질로 보호 되어 있기 때문에 해수와 직접 접축함으로써 생길 수 있는 용해작용을 면 한다. 골격 의 기본 구조는 bar 와 s pi ne 으로서 한 po lyc y s ti ne s 그룹의 경우 s pi ne 의 조합으로만 이루어진 것을 s pi cule 이라고 한다. 대부분의 방산층 골격구조는 복잡하며 뚜렷한 세포벽이 인지된다. 세포 u4 의구조는격자상. 스폰지상 그리고 다공질 각 등 세 가지 형태가 있으며 그림 4 와 같다 . 구형은 s p umellar i ans 의 가장 보편적 인 특정 이며 방사상의 s pi ne 이 잘 발 달된다. Nassellar i ans 는 공통적으로 대칭축을 보유한다. 표 1 은 Ri ed el (1971) 이 제안한 분류체계에 가장 가까운 것으로 대부분으] 현생 및 화석 방산충을 포괄한다. S p umellar i a 목에 속하는 여러 과를 구분하는 기준은 전곡의 윤곽 및 벽 의 구조이며 Nassellaria 目은 그림 5 와 같은 기본 골격구조의 유사성에 의 해 분류한다. 科들 간에는 독특한 구조가 발달되어 소공의 크기, 형태 및 배열이 중요한 열쇠가 된다.
 (A) 격자상세포벽
(A) 격자상세포벽
표] 방산 충의 분류 체계 Phy lu m PROTOZOA Class ACT I_N OPOOA Calkin s Subclass RADIOLARIA Mi .ill er Sup e rorder TRIYPLEA Hertw i g Sup e rorder POLYCYSTINA Ehren b e r g . em e n d. Ri ed el Order SPUMELLARIA Ehren berg Fami ly ENTACTINIIDAE Rie d el Fami ly OROSPHAERIOAE Hae ckel Fami ly COLLOSPHAERIDAE Muller Fami ly ACTINOMMIDAE Hae ckel. em e nd. Ri ed el Fami ly PHACODISCIDAE Hae ck e l Fami ly COCCODISCIDAE Hae ck el Fami ly SPONGODISCIDAE Hae ck e l. em e nd . Ri ed el Fami ly IIAGIASTRIDAE Pe ssagn o Fami ly PSEUDOAULOPHACIDAE Ri ed el. emen d. Pessagn o Fami ly PYLONI1DAE Hae ck e l Fami ly THOLONIIDAE Haec k e l Fami ly LITHELIIDAE Haecke l 0rder NASSELLARIA Ehrenberg Fami ly PLAGONIIDAE Haec k el. em e nd . Ri ed el Fami ly ACANTHOOESMIIDAE Hae ckel Fami ly THEOPERIDAE Hae ck el. emend. Ri ed el Fami ly CARPOCANIIDAE Haec k e l. emend. Ri ed el FFaammii llyy PATMEPRHOIPCYONRDYATCHIIDDAAEE RHi aede cekl el. em e n d. Ri ed el Fami ly ARTOSTROBIIDAE Ri ed e l Fami ly CANNOBOTRYIDAE Haeckel. emend. Ri ed el Fami ly ROTAFORMIDAE Pes s agn o Radio laria in certa e sedis Fami ly ALBAILLELLIDAE Defl an dre Fami ly PALAEOSCENIDIIDAE Ri ed el
 v
v
5 • 4 생태 방산충은 모든 해양에서 서식하며 모두 난류성이라고 알려져 있으나 다 론물체에 부착해 있을 가능성이 있다. 그들은 거의가 원양성이며, 아 직 연안성인 것은 알려져 있지 않다. 수심에 따른 개체수의 번화가 그림 6 에 표시되어 있다. 해구와 같은 심해에도 일부 종류가 서석하고 있음이 알려졌다. 방산충은 공생조류인 Zooxan t hel J ae 의 숙주 역할을 하여 이들로부터 광합 성에 의해 생산된 영양분을 공급받고 대신 호흡 작용의 산물인 CO2 로 공 생생물의 대사를 돕는 것으로 보인다.
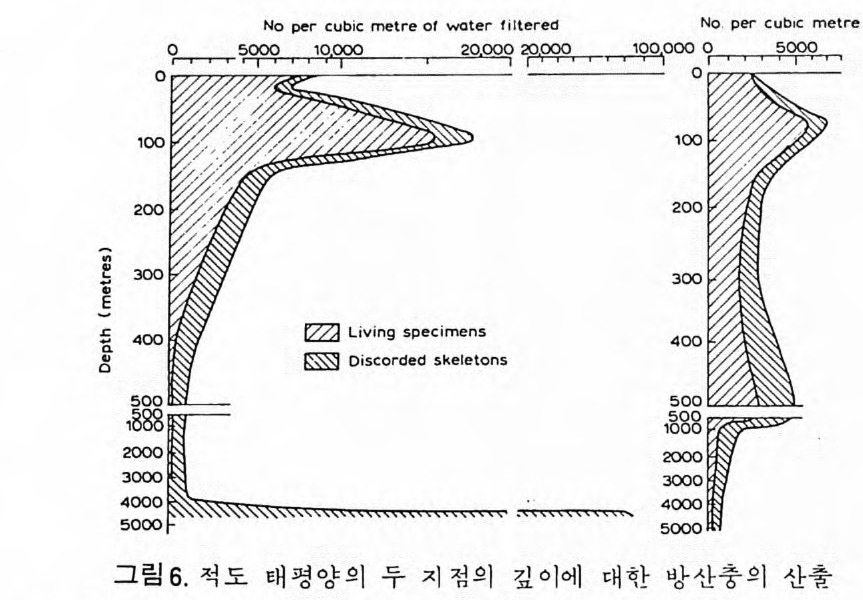 No per cubic met re 01 wat e r fil te red No pe r cubic met re
No per cubic met re 01 wat e r fil te red No pe r cubic met re
현재 알려진 지식으로는 방산층의 섬세한 형태는 부유성 습성에 적응하 기 위한 것으로 보인다. 방사상의 s pi nes 와 무수한 axo pods는 수중에서 의 유동을 위한 것이고 세포질 내의 유적들은 부력울 증가시키는 구실을 하 는 것 같다. Brandt (1895) 는 방산층 의 페 포 체 (alveolar comp le x) 에 는 CO2 로 포화되 어 있는 물이 들어 있고 이를 조철함으로써 부력을 조철한다고 주장했다. 이 러한 생각은 방산층이 수직 이동을 함으로써 환경에 적응한다는 종래의 관 찰을 합리화시키는 것이다. 이 밖에도 alveol i의 조절에 의한 수직 운동에
관한 여러 견해들이 발표되었으나 아직은 불확실한 것들이다. 플랑크론 표본으로부터 생물의 지리적 분포가 연구된 예가 매우 적은반 떤 퇴적물 내의 방산층의 성분과 그 위의 해수의 환경 요소들의 관계가 많 이 연구되어 있다. Petr us hevskaya ( 1967, 1971b~d) 와 Renz(1973) 는 해양에서의 방산층 군집 의 분포는 수괴의 특징과 깊은 관련이 있으며 특히 해류의 순환이 주요한 역항울 한다고 주장했다 . 수직적 인 분포도 역시 표층, 중충 및 십충 해류 의 이동과 밀접한 관계를 맺는다. 특히 양국 지방에는 유사한 군집이 분 포하며 어떤 것들은 적도 지방까지 표충수를 따라 연속되기도 한다 . 퇴 적물에 관한 많은 연구는 표층수의 특성 이 방산층의 분포에 가장 중 요한 영향을 준다는 사실을 증명한다. 5 • 5 고생태 및 고해 양학 방산층은 해양의 중요한 규질 퇴적물의 성원이며 20~30% 의 방산충이 주가 되는 생물질 실리카로 구성된 泥質 퇴적물은 방산층 연니를 만돈다. 따라서 방산층은 해수의 실리카 순환에 중요한 역할을 한다. Jo hnson(1974) 의 실험에 의하면 수시간이나 수주 정도연 방산충은 해수에서 완전히 용 해되며 수심이 1, 000m 이내의 sil i c a -po or 수층에서는 더욱 십하다. 퇴적물 내에서도 용해는 계속되는데 이는 물의 포화도와 깊은 관계가 있다. 퇴적 률이 높은 곳에서는 규질 화석의 용해도가 낮은데 이는 빠른 매몰과 유기 성 규질물의 부분 용해에 의한 간극수의 완충작용이 골격으로부터 해수를 차단하기 때문인 것으로 보인다. 화산작용은 칙 접 간접으로 s i l i ca 의 퇴적을 도와주는데 비 정 질 상태로 직 접 침전되기도 하지만 규질 생물체의 생산을 촉진시킨다. 그러나 근본적 으로 해저의 s ili ca 퇴적은 적도 지역의 광역적인 영양영류의 해양학적 순 환에 힘입은 플랑크론의 생산에 비교할 것이 못 된다. 따라서 이 지역은 방산층 이외에도 석회질 난노플랑크론이나 유공충의 왕성한 생산이 촉진되어 C.C. D. 보다 얕은 해저에서는 오히려 방산층의 번성이 가리워진다. 방산층은 북태평양이나 남극대륙의 주변에서는 상대적으로 다른 생웅보 다 우세하여 규조 다음의 서열을 차지한다(그림 7 참조). 대양의 중심부는 육원 쇄설물의 퇴적륭도 낮고 플랑크론의 생산성도 낮 기 때문에 바닥의 생물 골격은 장시간 해수에 노출되어 완전한 용해 작
 J(6'
J(6'
용이 초래된다. 결과적으로 이 지역은 유기물이 없는 원양성 갈색 접토 (pela g ic brown clay) 가 주요 되 적물을 이룬다. 해양의 시공간적인 상태에 따라 방산충의 함량은 다소 변화가 있으며 대륙봉 지역에서는 매우 적거나 없는데 이는 `육원의 퇴적물의 함량이 많기 때문 0l · 다. 방산층 쳐어트 (Rad i olar i an Cher t)는 쥬라기의 Te t h y s 해와 같은 곳에서 잘 알려져 있다 . R adi olar it e 와 현재 원양성퇴적물의 비교는 지사를 밝히 는 데 중요한 작업이다. 방산층 쳐어트가 원양성 석회암을 피복하고 있다 떤 C.C.D. 의 변화를 추정하여 고십도의 해석이 가능하다. 대양저의 중앙 해령 부근의 비교적 얕은 해저 에서 퇴적된 석회질 퇴적물은 해저확장에 따라 이동되어 해령의 기슭에 이르면 해양지각이 석으면서 수십이 깊어져 서 규질 퇴적물로 相 맏 化를 할 것이다. 과거의 수심변화는 현재의 C.C.D. 깊이의 자료를 근거로 추정한 것인데 실제에 있어서 지질 시대의 C.C.D. 는 당시의 기후나 해수의 성격에 따라 다소 달랐 을 가능성도 있다. 대륙사면에서 일어나는 저탁류에 의한 t urb idit e 는 화석군집을 재배열시 켜 거 꾸로 퇴적물 기원을 추적하게 하기도한다. 지체 구조적으로 활발한 지 역 인 해 만에 퇴 적 되 는 melan g e 층에는 r adi olar it e 가 포함될 수 있으므로 판 의 충돌 경계부를 복원하는 데도 한몫을 맡는다. 다른 미화석군집과 함께 방산층은 수심이나 해수의 성질 둥 고해양학적 인 해석에 큰 기대를 걷 수 있는 그룹이다. 화석군집을 이용한 퇴적환경 의 복원은 화석군과 현생 생물군의 엉격한 생태학적 비교를 통해 지사의 많은 문제들 을 푸는 작업이다 . Bramlett e( 1946) 은 켈리포니아의 마이오세 지층인 Mon t ere y충을 연구하 여 이 지층이 정상적인 대륙붕-보다 얕은 퇴적환경의 산물이라는 사실을 화석군집을 연구함으로써 밝혀냈다. 실·제 이와 유사한 퇴적지가 캘리포니 아 주변의 한 분지에서 발견되어 방산층이 풍부한 보통 깊이의 비교적 연 안성인 분지의 한 예로 알려졌다(표 2 참조). 범세계적인 공간 분포와 높은 종다양도 때문에 방산층은 훌륭한 고기후 의 지시자이다. 현재 북태평양이나 남극해의 석회질 되적물이 적은 지역 이 특히 방산충류 고생태학 연구의 최척지라고 할 수 있다. N ig r i n i(1 970) 는 퇴적물 내의 방산층 군집과 해수의 온도와의 상관관계 로부터 수온의 지수를 산출하였다. 이 지수를 여러 사람들이 각지역의해
표 2 과거시대와 현재의 방산층 퇴적층의 특 칭 Terrig e nous Average rate Associa t e d comp o nent of sedim enta t i on mi cr ofo ss il s dia t o ms Radio l aria n o6ze low 1 m/m.y. for ami ni f er a calcareous nannop la nkto n for ami ni f era Radio l arite s low 1 m/m.y. calcareous nannop la nkto n dia t o ms ? dia t o ms Santa Barbara Basin moderate 1000 m/m.y. for ami ni f era calcareous nannop la nkto n dia t o ms Monte r ey Formati on moderate 1000 m/m.y. for ami ni f era calcareous nannop la nkto n 저시추 코아에서 관찰되는 시대별 방산층 군집의 번화에 적 용하여 해수의 고온도의 변화를 추적하였다. Moore (1973) 는 지 난 24, 000 년 전보다 오늘날 오레곤 해 역 이 약 4°C 의 온도상승이 있음을 밝혔고 Sachs (1973) 는 과거 幻 0, OO0 년 동안 중앙 아국대 태평양의 수온이 현재보다 약 10°C 정도 낮은 상태였었다고 계산해냈다. 규질 미화석이 풍부한 퇴적물의 대상분포는 그 지역 해수의 높은 생물 생산을 반영하며 이것은 과거의 광역적인 고해양학적 환경 을 복원하는 데 좋은 도구가 된다. R i edel 과 Funnell (1964) 은 태평양의 제 3 기 미화석 분 포를 도화하여 당시 해류의 방향과 위치가 오늘날과 거의 같았다는 사실 을밝혀냈다.그러나세부적으로는후기제 3 기에적도해류계의 분포가 좁게 나타났는데 이것은 당시 냉각된 해수의 강한 순환에 영향을 받은 것 이 아닌가 추측된다. 1964 년 이후 해 양처 확장 현상을 위 의 자료에 적 용 시켜서 동태평양 해령의 서쪽 적도의 해처가 계속하여 복서쪽으로 이동함 에 따라 퇴적물 내의 미화석의 군집에 이상적인 분포가 야기된 것을 알게 되었다. 5 ·6 생물충서 방산층은 계통발생적 상호 관계가 아직 불분명하기 때문에 훌륭한 지층
대비의 도구가 되어오지 못했다. 그러나 최근의 연구는 이들이 장기의 지 층 대비에 유용하다는 결론을 얻었다. 세부적인 화석대가 신생대에 정립 되었고 더 오래된 화석군에 대해서도 성공적인 결과가 기대된다. 선캄브리아대나 캄브리아기의 방산층 기록에는 아직 불확실한 문제가 남아 있으나 오르도비스魚 C 이후의 지층에는 잘 보존된 방산층 화석군이 발 견되 었다 . 전형 적 인 고생 대 방산층은 albail lel id s , pa laeoscenid s 그리 고 int e rn al s pi cules 을 갖는 s p떠 ellar i ans 의 일부가 포함된다. 앞으로의 고생대 방산층의 연구는 이미 잘 연구되어 있는 팔석류나 코 노돈 트 화석군의 생물 충서를 기반으로 해서 방산층의 충서 확립에 중점 을 두어야 할 것이다. 트라이아 스 기의 방산층은 거의 알려져 있지 않으나 쥬라기 지층에는잘 보존된 화석이 많이 산 출 된다 . 백악기의 방산층은 이미 이 시대 21 지층 및 시대 구분에 활 용되기 시작했다 . 특히 다른 종류의 미화석이부족한지 충에서 는 방산층이 대단히 중요한 역할을 한다 . 백악기 지층에서는 현재 해저에서 채 취한 시추 코아(예 : D. S. D. P.) 와 육지의 해성층 사이에 대 비가 시도되었다. 제 3 기와 제 4 기충으] 방산층은 다론 시대보다 훨 씬 세밀하게 연구되어 현재 모든 시대별 시준화석군이 잘 정리되어 있다. 신생대의 생층서 체 계는 주로 해저 퇴적물 표품으로부터 얻어졌다. 이 샘플들은 또한 석회질 미화석 을 함유하고 있어서 찰 알려져 있는 부유성 유공충이나 석회질 난 노 플랑크론의 충서 체계와의 상호관계도 잘 연구되 었다. 특히 D. S. D. P. 표품들이 각 해역에서 채취되어 시공간적인 분포 양상이 알려졌고이러한 생물대는 고치자기 편년과 함께 엮어져서 고해양학 연구에 주요 자료가 된다. Ha y s(l965) 는 남국해의 풀라이오-풀라이스토世의 십해저 퇴적물을 분 대하였다. 이것은 뒤에 해저의 고지자기 연구에 이용되고 다시 방사능-고 지자기 편년과 비교되었다. 그 후 각 해양에서 화석군집, 절대연대 맛 고 지자기 등 세 가지의 연구 분야가 협동하여 지사학적인 업적이 날로 쌓여 가고 있다.
보충교재 Camp b ell, A. S., 1954. Subclass Radio l aria . In : R. C. Moore (Edit or ) , Treat- is e on Inverte brate Paleont o log y. Proti st a 3. Geol. Soc. Am., New York, N. Y., and Kansas Univ . Press, Lawrence, Kans a s, pp. D11-D 163. (A convenie n t versio n of Haeckel's sy s te m wi th some late r addit ion s and mi no r revi si o n s. Should be used wi th caut ion because of errors and th e unrelia b i- lity of ag e rang e s comp ile d fro m older lite r alure.) Foreman, H. P. and Ri ed el, W. R., 1972. Cata log u e of Polyc y s ti ne Radio /aria . Serie s 1 (1834-1900), Vol. 1 (Mey e r, 1834-Bury, 1862), Parts 1 and 2. Sp e cia l Publi ca ti on , Americ a n Museum o( Nat u ral Hi s to ry, New York, N.Y. [T his cata l og u e cont a in s orig ina l descrip tion s of and subseq u ent refe r ences to ge nus and sp e cie s -level ta xa covered in chronolog ica l order fro m earlie s t works. Subseq u ent volumes are in pr ep a rati on .) G runau, H. R., 1965. Radio l aria n cherts and associa te d rocks in sp a ce and ti- me. Eclog a e Geol. Helv., 58(1 ) : 157 -208. [S ummariz e sg e og r ap h ic and st r- ati gr ap h ic dis t r ib u t ion of radio l aria n rocks th roug h out th e world. Di sc u sses t heir (deep - wate r) orig in main l y _i n te rms of sedim ent o log ica l-g e ochemi ca l evid e nce and associa t ion wi th oceanic basalts .) . Haeckel, E., _18 87. Rep o rt on th e Radio l aria collecte d by H. M. S. Challeng e r durin g th e ye ars 1873-7 6. Rep . Voy a g e Challeng e r,, Zool. 18 : clxxxv ii+ 1803 pp. (140 pis. , 1 map ) . [T he fu ndament al monog r ap h on radio laria n ta xo- nomy . An ind is p e nsable refe rence work, but should be used wi th awareness _of th e arti fici a l it y of th e classif ica ti on , and th e in adverte nt inc lusio n of reworked older sp e ci m ens in what he to ok to be Qu ate rnary sa~p l e lude s ext ensiv e rev iew of bio l og y and dis t rib u t ion .] Hollande, A. and Enju m et, M., 1960. Cy tol og ie, evolut ion et sy s te mati qu e des Sp h aeroid e s (Radio l air e s). Arch. Mus. Nat. Hi s t. Nat. , Ser. 7. 7:7-134. [A key refe rence on radio l aria n bio l og y. S tr esses cy t olog y of th e sp h eric a l sp u mellaria n s but in cludes observati on s on some nassellaria n s as well. Pro- po ses revi se d classif ica ti on of th e for mer gr oup based on sof t anato my .] Pct r ushevskay a , M. G., 1971a. 야 th e nat u ral sy s t e m of_ po lyc y s ti ne Rad iol a-
ria (Class Sarcodin a ) . In : Proc. Second Plankto ni c Con(., Roma, 1970 Ed- izi o n i Tecnoscie n za, Rome, pp. 981-99 2. [T his summary of a revis e d radi- olaria n classif ica ti on wi ll serve as an in t roducti on , in Eng l is h , to ext e nsiv e work on radio laria n sy s te mati cs , morph olog y, and bio g e og r aph y .] Rie d el, W. R. and Holm, E. A., 1957. Radio l aria . In : J. Hedg pe t h (Edi t o r) , Tr- eati se on Marin e Ecolog y and Paleoecolog y, I . Geol. Soc. Am., Mem. 67 : pp. 1069 -1072. [T his annot a t e d bib l io g r ap h y on radio l aria n ecolog y is a go od int ro ducti on to th e monog r ap h s result ing fro m th e gr eat oceanog r aph ic exp e dit ion s of th e late nin e t e enth -early tw ent iet h cent u rie s .] Ri ed el, W. R. and Sanfi lippo , A., 1977a. ·ce nozoic Radio laria . In: A. T. S. Ra-msey (Edit or } , Oceanic Mi c rop a leonto log y. Academi c Press, New York, N. Y., in pr ess. [T hese tw o pa p e rs by Rie d el and Sanfi lip p o summariz e nume- rous stu die s on radio l aria n sy s te m ati cs and bio s t rati gr aph y, th e second rev- iew i ng radio l aria n knowledg e app li ca ble to pa leonto l og y.) Rie d el, W. R. and Sanfi lip po , A., 19776. St r ati gr aph y and evoluti on of tro p - ica l Cenozoic radio l aria n s. In : W. R. Rie d el and T. Sait o {Edit or s} , Marin e Plankto n a 뼈 Sedim ent s . Proc. Thir d Plankto nic Con{., Kie l . 1974. Mi cr o- pa leont o log y Press, in pr ess. S tre lkov, A. A., Khabakov, A. V. and Lip m an, R. Kh., 1959, Podclass Radi o- laria iii luche vik i . In: Yu. A Orlov(Edit or ), Osnovy Paleonto log ii. Akad. Nauk Sof. SP.a Sle. oRn.t, o Mlogo ys.k oIws,r aveoll . PVr,o gp r pa.m 3 6f9o-r 48S2c.ie n Etin fgic li s Th raTnrsalnastli oant i so,n :J Fer uu 따sa alemmen, 7t a2l8s pp. [T he Eng li sh tr anslati on is a valuable summary of exte nsiv e Russia n ra- dio l aria n inv esti ga ti on s whic h are large ly unknown to weste rn readers. Alt- houg h th e Haecke li an sy s te m is foll owed, th e tre atm ent of all aspe cts of radio l aria n stu dy is qu it e comp r ehensiv e .] 참고문현 Ads he ad, P. C., 1967. Collecti on and laborato ry main t e n ance of pla nkto n ic fo- rami ni f er a. Mi c rop a leonto log y, 13(1 ) : 32-40 (pls . l-2) . Barth e l, K. W., 1970. On the dep o sit ion of th e Solnhof en litho g r aph ic lim est- one (Lower Ti tho nia n , Bavaria , Germany ) . Neues Ja hrb. Geol. Palaonto l., Abh., 135 : 1-18 (2 te xt- fi gs , 4 pis. ) . Berge r, W. H., 1968. Radio l aria n skeleto ns : soluti on at dep ths . Sc ien ce, 159 : 1237 一 1238. Berge r, W. H. and Wi n te rer, E. L., 1974. Plate str ati gr aph y and th e fluc t- uati ng carbonate lin e . In : K. J. Hsii and H. C. Jen ky n s (Ed ito rs) , Pelag ic Sedim ents on Land and U 뼈 er th e Sea. Int. Assoc. Sed im ento l., Sp e c. Pu- bl. No. 1. Black~II, Oxfo r d, pp. 11-48 (26 te xt- fi gs ) . Bosellin i , A. and Wi n te rer, E. L., 1975. Pelag ic lim esto ne and radi ol arit e of the Teth y a n Mes ozoic : a ge neti c model. Geolog y, 3(5) : 279-282(2 tex t -fi- gs ). Bramlett e, M. N. . 1946. The Mont e rey for mati on of Cali for nia and th e orig in of its sili c e ous rocks. U. S. Geol. Surv., Prof. Pap. , 212 : 57 pp. Brandt, K., 1895. Bi ol og isc he und fau nis ti sc he Unte rsuchung e n an Radio l arie n und anderen pe lagi sc hen Thie r en; 1. Unt e rsuchung e n iibe r den hy d rosta ti sc - hen Ap pa rat von Thalassic o llen und colonie b il d enden Radio l arie n . Zool. J야 rb., Sy s t. , 9 : 27-74 .
Cachon, J. and Cachon, M., 1971. The axop o di al sys te m of Radi o laria Nass- ellaria . Orig in, orga nis a ti on and relati on wi th the oth er cell orga nelles. Gen- era! consid e rati on s on the macromolecular orga nis a ti on of the ste reop la sm of acti no p o ds. Arch. Proti st e n kd., 113 : 80-97 (9 tex t - f igs ., 9 pis. ) [in F r- ench wi th Eng li sh summary. ] Cachon, J. and Cachon, M., 1972a. The axop o dia l sys te m of Radio l aria Sp h - aeroid a e. I. Cent ro axop la sti da e. Arch. Proti st e n kd., 114 : 51-64 (7 tex t- figs , 9 pis. ) [in F rench wi th Eng li sh summary. ] Cachon, J. and Cachon, M., 1972b. The axop o dia l sy s te m in Radio l aria Sp h a- eroid a e. II. The Periax op la sti da e. III. The Cryp tax op la sti da e (Anaxop la sti da e). N. The fu sules and the rheop la smi c sy s tem . A rch. Proti st e n kd., 114 : 291- 307 (5 te xt- fi gs , 8 pis. ) [in French wi th Eng li sh summ ary. ] Calvert, S. E., 1974. Dep o sit ion and diag e nesis of sili c a in marin e sedim ent s. In: K. J. Hsu and H. C. Jen ky n s (Edit or s), Pelag ic Sedim ent s On Land and Under the Sea. Int. Assoc. Sedim ent ol ., Sp e c. Publ. No. 1. Blackwell, Oxfo r d, pp. 273 -2 99 (7 tex t- fi gs .) . Casey, R. E., 1971 . Radio l aria n s as ind i ca to rs of pa st and pr esent wate r ma- sses. In : B. M. Funnell and W. R. Rie d el (E d ito rs) , The Mi c rop a l.eo nto log y of Oceans. Cambrid g e Univ e rsit y Press, London, pp. 331-341 . Casey, R. E., Partr id g e , T. M. and Sloan, J. R., 1971 . Rad iol aria n life spa - ns. morta l it y rate s , and seasonality ga in e d fro m Rece~t sedim ent and pla - nkto n samp le s. In : Proc. Second Plankt on i c Con{., Roma, 1971. Ediz i o n i Tecnoscie n za, Rome, pp. 159 -1 65. Chip ping , D. H., 1971 . Paleoenv iro nment al sig n if ica nce of chert in the Franc- isc an Formati on of weste rn Califo r nia . Geol. Soc. Am., Bull., 82(6) : 1707 -1711 (1 tex t- fig.) . Defl an dre, G., 1960. A pr op o s du develop pe ment des recherches sur les ra- dio l air e s fos sil e s. Rev. Mi cr op a leonto l., 2(4) : 212 一 218 (pl. 1) . Dum it r i c 죠, P., 1970. Cryp to cep h alic and cryp to th oracic Nassellaria in some Mesozoic dep o sit s of Romania . Rev. Roum. Geol., Geop h y s ., Geog r ., Ser. Geol., 14(1) : 45-124 (27 pis. ) . Dunham, J. B. and Murph y, M. A., 1975. An occurrence of well-p r eserved _Ra d iol aria from the ·upp e r Ordovic i a n (Caradocia n ) of cent ra l Nevada. J. Pa l.e onto l ., 50(5) : 882-88 7 (2 tex t- fi gs ., 1 pl.) . Foreman, H. P., 1963. Up pe r Devonia n Kadio l aria fro m the Huron member of the Ohio shale. Mi cr op al.eo nto l og y, 9(3) : 267 -304 (pis. 1-9) . Foreman, H. P., 1966. Two Creta c eous radio l aria n ge nera. Mi c rop al.eo nto lo- gy. 12(3) : 355-35 9 (11 tex t- fi gs .) . Foreman, H. P., 1968. Up pe r Maestr i c h ti an Radio l aria of Califo r nia , Pa l.ae o-
nto l . A ssoc., London, Sp e c. Pap. , no. 3 : pp. l-82 (pls . 1-8, tex t- fi g. l) . n Forte y , R. A. and Holdsworth , B. K., 1972. The olde st known well-p r eservedRadio l aria . Bull. Soc. Paleont o l. Ita l., 10( 1 ) : 35-41 (pis. 10, 11, tex t- fig.I ). Frien d , J. K. and Rie d el, W. R. 1967. Cenozoic orosph aerid radi ol aria n s from tro p ica l Pacif ic sed im ent s, Mi c rop a leonto log y, 13 : 217-23 2. Garris o n, R. E., 1974. Radio l aria n cherts , pe lagi c lim esto nes, and igne ous ro- cks in eug e osy n clin a l sedim ent s. In : K. J. Hsu and H. C. Jen ky n s (Edit or s) , Pelag ic Sedim ent s On Land and Under th e Sea. Int. Assoc. Sedim ento l., Sp e c. Pub!. No.·l. Blackwell, Oxfo r d, pp. 367-396 (6 tex t- figs .). Garris o n, R. E. and Fi sc her, A. G., 1969. Deep - wate r lim esto nes and radi o- lari tes of the Alpi ne Ju rassic . In : G. M. Fried man (Edit or ) , Dep o sit ion al En-
vir o nments in Carbonate Rocks, a Sy m p o siu m . Soc. Econ. Paleonto l. Mi n- eral., Sp e c. Pub!., No. 14 : pp. 20 -5 6. Goll, R. M., 1968, 1969. Classif ica ti on and ph y lo g e ny of Cenozoic Tris s ocy c l- ida e (Radio l aria ) in th e Pacif ic and Carib b ean basin s ; Part I, J. Paleonto l., 42(6) : 1409 -1 432 ; Part II , J. Paleonto l., 43(2) : 322-33 9. Goll, R. M. and Bj< f>r klund , K. R., 1971 . Rad iol aria in sur fac e sedim ent s of the North At la nti c Ocean. Mi c rop a leont o log y, 17(4) : 434-45 4 (8 tex t- figs. ) . Goll, R. M. and Bj< f>r klund , K. R., 1974. Rad iol aria in surfa c e sedim ent s of the South At la nti c. Mi c rop a leonto log y, 20(1)38-75 (16 te xt- figs .) . Harpe r, H. E. and Knoll, A. H., 1975. Sil ic a , dia to ms and Cenozoic rad iol ar- ian evolu t ion . Geolog y, 3(4) : 175-1 77 (1 tex t- fig.) . Hay s , J. 0., 1965. Radio l ari_a and late Tertia r y and Qu ate rnary his t o ry of An- ta rcti c seas : Bi ol og y ,of th e Ant a rcti c Sea II . Anta rct. Res., Ser. 5, pp. 125 -184. Hay s , J. 0., 1970. S tr ati gr ap h y and evoluti on ary tre nds of Rad iol aria in Nor- th Pacif ic deep - sea sedim ent s. In : J. D. Hay s (Ed itor ) , Geolog ica l Investi g- ati on s of th e North Pacif ic. Geol. Soc. Am., Mem. 126 : 185-218(pl .1 ). Hay s , J. D. and Op d y k e, N. 0., 1967. Anta rcti c Rad iol aria , mag n eti c rever- sals, and clim ati c chang e . Scie nce, 158 : 1001-1011. Heath , G. R., 1974. Di ss olved sili c a and deep - sea sedi m ent s. In:W. W. Hay (Edit or ) , St u die s in Paleo-Oceanog r aph y. Soc. Econ. Paleonto l, Mi n eral., Sp e c. Pub!., No. 20 : 77 一 93 (11 te xt- fi gs .) . Holdsworth , B. K., 1969. The relati on ship betw een th e ge nus Albail l ella De- flan dre and the cerato ik i s c id Radio l aria . Mi c rop a leonto log y, 15( 2 ) : 230-2 36 (pl. 1, te xt -fig . l) . Holdsworth , B. K., 1971 . The cerato ik s cid natu re of th e radio l aria n Lap ido p - isc um pivi t ea ui De flan dre. Mi c rop a leont o log y, 17(2) : 244 -2 48 (pl. 1, t ext -fi gs 1-2). Jo hns on, D. A. and. Knoll, A. H., 1974. Radio l aria as pa leocli m ati c ind ic a to rs: Pleis t o c ene clim ati c fluc tu ati on s in th e eq u ato ria l Pacif ic Ocean. Qu ate rna-Joh ryn s oRne, s.,D . 4A :. 2a0n6d- 2K 1n6o l(l3, 1Ae x. tH -.,f ig1s .9,7 5 2. pAisb. s) o . lut e ag e s of Qu alernary radio l a-ria n datu m levels in the eq u ato ria l Pacif ic. Qu ate rnary Res., 5 : 99 -110 ( 2 tex t -figs. , 1 pl. ). Jo hnson, T. C., 1974. The dis s olut ion of sil ic e ous mi cr ofo s sil s in sur fac e sed- im ent s of the easte rn tro p ica l Pacif ic. Deep - Sea Res., 21 : 851-864(6 te- xt -f igs ., 2 pls .) . Klin g , S. A., 1971 . Di m orph is m in Radio l aria . In: Proc. Second Plankt on ic Con(., Roma, 1970. Ediz i o n i Tecnoscie n za, Rome, pp. 663 -672 (5 pls .) . Klin g , S. A., 1976. Relati on of radio l aria n di s tr ib u t ion s and subsur fac e hy d r- og r ap h y in the North Pacif ic. Deep - sea Res., 23 : 1043-1058 (8 figs .) . K rug li k o va, S. B., 1969. R adio l ya rii v po verkhmostn o m sloe osadkov severnoi po lovuny Ti kh og o Okeana. (Radio l aria in the sur fac e lay e r of se d im ent s of the north ern half of the Pacif ic Ocean.) In: Ti kh y Okean. M ik ro(lora i mi - kro(auna v osadkakh Ti kh og o Okeana. Nauka, Moscow, pp. 48-72 . Li si t zin , A. P., 1971 . Di st rib u t ion of sil ic e ous mi cr ofo s sils in suspe nsio n and in bot tom sedim ent s. In: B. M. Funnell and W. R. Rie d el (Edit o rs) , The Mi - crop a leont o log y of Oceans. Cambrid g e Univ e rsit y Press, London, pp. 173- 195. Moore, T. C., 1971 . Radio l aria . In: Init ial Re p o rts Deep Sea Dril l in g Proje - ct, Leg 8, pp. 727-77 5. Moore Jr. , T. C., 1972. Mi d- Terti ar y evolut ion of the radio l aria n ge nus Cal- ocy c lett a. M icr op a leont o log y, 18(2) : 144 -152 (pl s. 1, 2) . Moore Jr. , T. C.; 1973. Late Pleis t o c ene-Holocene oceanog r ap h ic chang e s in the north e ast e rn Pacif ic. Qu ate rnary ltes ., 3 : 99-10 9 (6 t ext -figs .) . Ni gr in i , C., 1967. Radio l aria in pe lag ic sedim ent s fro m the India n and At lan - t ic Ocea11s. S crip ps Inst. Oceanog r . Bull., 11 : 125 pp. Nig r in i , C., 1968. Radio l aria from easte rn tro p ica l Pacif ic sedim ent s. Mi c r- opa leont o log y, 14(1) : 51-53 (16 t ext -figs ., 1 pl.) . Nig r in i , C., 1970. Radio l aria n assemblag e s in the North Pacif ic and the ir ap - pli ca ti on to a stu d y of Qiat e rnary sedim ent s in core V20-130. In: J. D Ha- ys ( Edit or ) , Geolog ica l Investi ga ti on s of the North Pacif ic. Geo!. Soc. Am., Mem. 126 : 139 -1 83 (4 pls .) . Nig r in i , C., 1971 . Radio l aria n ·zo nes in the Qu ate r nary of the eq u ato ria l Pa- cif ic Ocean. In: B. M. Funnell and W. R. Ri ed el (Edit o rs), The Mi cr op a le- ont ol og y of Oceans. Cambrid g e Univ e rsi t y Press, London, pp. 443-491. Ni sb et, E. G. and Pric e , I., 1974. Si lic e ous tu rbid i t e s:bedded cherts as re- depo sit e d, ocean rid g e -deriv e d sedim ent s. In: K.J . Hsu and H.C. Jen ky n s (Edit o rs) , Pelag ic Sedim ent s On Land and Under the Sea Int. Assoc. Se-
dim ento l., Sp e c. Pub!. No. 1- Black well, Oxf o rd, pp. 351-36 6. Pessagn o Jr. , E. A., 1970. The Rot a fo rmi da e, • a new fam i ly of Up pe r Cret- • aceous Nassellaria (Radio l aria ) fro m t he G reat Valley seq u ence, Califo r nia Coast Rang e s. Bull. Am. Paleont o l., 58 (257) : 1-32 (9 pis. ) .Pessagn o Jr. , E. A., 1971 . Ju ras sic and Creta c eous Hagi as tr i d a e from the Blake-Bahama Basin (Si te 5A, JO IDES Leg I ) and the Great Valley seq u -
ence, Calif or nia Coast Rang e s. Bull. Am. Paleonto l ., 60 (264) : 1-83 (pis. 1-19). Pessa g n o Jr. , E. A., 1972. Creta ceous Radio l aria . Part I : The Phaseli for - mid a e, a new fam i ly , and oth er Sp o ng o dis c acea fro m the . Up pe r Creta c eous po rti on of the Great Valley seq u ence. Part II : Pseudoaulop h acid a e Rie d ,.J fro m the Creta c eous of Califo r nia and the Blake-Bahama Basin (JO IDES PeLses ga g n1 o) . JBr. u, l lE. . AAm,, . 1P97a6l.e onRtao d li.o, l a6r1ia (n2 7z0o) na: t2i o6n 7 -a3n2d8 s (ptir sa . t2i g2r- a3p1 hy) . of the upp e r Creta c eous po rti on of the Great Valley seq u ence. Mi cr op a leonto l og y, Sp e c. Pap. , No. 1. Petr u s h e v skaya , M. G., 1962. Znachenie rosta ske leta radio l ya rii dlya sis t e m - ati ki . (Th e im p o rta n ce of sk eleta l gr owt h in radio l aria n s for the ir sy s te m at- ics .) Zoo/. Zh., 41( 3 ) : 331-341 . Petr u s h e vsk ay a , M. G., 1967. Radio l ya rii ot ry a dov Sp u mellaria Nassellaria an- . tar kt ich e sko i obla s t i (A nt ar cti c spu mellin e and nassellin e radio l aria n s.) In: E. N. Po vlo vs k ii (E dit o~ ) . Issledovaniy a · Fauni Morei, 3( 12) . ?ool. Inst. , Ak- ad. Na uk . S. S . S. R., Le n in g r ad, 186 pp. (102 tex t- fi gs .) . Pe tr u s h e vsk ay a , M. G., 1971a. Radio l ya rii Nassellaria v pla nkto n e Mi ro vog o Oke an a. (N a ssell aria n Radio l aria in the pla nkto n of the world ocean.) In: B. E. By k hovs k ii (E dit o r} , Issledovaniy a . Fauny Morei, 9( 17) . Zoo!. Inst. ,A k- ad. Nauk . S. S. S. R., Lenin g r ad, 295 pp. (146 tex t -figs .). Petr u s h e vsk ay a , M. G., 1971b. Sp u mellaria n and nasse ll aria n Radio l aria in the pla n k t on and bo ttom sed im ent s of the Cent ra l Pacif ic. In: B. M . Funnell and W. R. Rie d el (Edit o rs) , The Mi cr op a leont ol og y of Oceans., Cambrid ge Uni- versi t y Press, London, pp.3 09-317(6 tex t -figs .). Pet r us h e vskay a , M. G., 1971c. Radio l aria in the pla nkto n and Recent sed im e-nts fro m the India n Oce an and Ant a rcti c. In : B. M. Funnell and W. R. Ri ed el (Edit or s} , The Mi cr op a leont ol og y of Oceans. Cambrid g e 'U-ni v e .r si t y Press, London, pp. 319 -3 29 (6 tex t -fi gs .) . Pet r us h evs kay a , M. G. and Koslova, G. E., 1972. Rad iol aria : Leg 14, Deep Sea Drill i n g Proj ec t. In: D. E. Hay e s, A. C. Pim m and oth ers, Init ial Rep - orts of the Deep S eaDrill in g Proje c t, L eg 14: 495-648(41 pis. ). ·Po korny, V., 1963. Prin c ip l es of Zoolog ica l Mi cr op a leonto l og y. Perga mon Pr- ess, London. 652 pp. Renz, G W., 1976. The dis t r ib u ti on and ecol og y of Radio l aria in the Cent ral Pacif ic pla nkt on and su r fac e sedim e nt s. Scrip ps Inst. Oceanog r ., Bull., 22: 1-26 7 (17 tex t -figs ., 8 pis. ) . Resche tn j a k , V. V., 1955. Verti ka lnoe ras p e delenie radio l ya riy Kurilo -Kamcha- tsk oy Vp a din y . (Verti ca l dis t ri b u t ion of radio l aria n s in the Kuril e -Kamchatk aTrench.) Tr. Zoo!. Inst. , Akad. Nauk S. S .S.R., 21 : 94-101(1 pl.). Ri ed el, W. R., 1957. Radio l aria : a pr elim in a ry st r ati gr ap h y, Rep . Sweri ish De-ep- Sea Ex pe d., 6(3 ) : 59 -96 (pis. 1-4) . Rie d el, W. R., 1971. Sy s te m ati c classif ica ti on of po lyc y s ti ne Radio l aria . In:B. M. Funnell and W. R. Rie d el (Edit o rs) . The Mi c rop a leonto log y of Oceans. Cambrid g e Univ e rsit y Press, London, pp. 649 -6 61 . Rie d el, W. R. and Funnell, 8. M., 1964. Tertia r y sedim enl cores and mi cr ofo - ssils from th e Pacif ic Ocean flo or, Q. J. Geo!. Soc. Lond., 120 : 305 -3 68 (pis. 14-32 ) . Rie d el, W. R. and Sanfi lip p o , A., 1974. Radio l aria fro m the sout h ern India n Ocean, DSDP Leg 26. In: T. A. Davie s , B. P. Luy e nd y k and orh e rs, Init ial Rep o rts of Deep Sea Dril l in g Proje c t, Leg 26 : 771-813 (15 pis. ) . Sachs, H. M., 1973. Late Pleis t o cene his l o ry of rhe Norrh Pacif ic; cvid e nce fro rr :- qu anti tat i ve stu d y of Radio l aria in Core V21-173. Qu ate rn ary Res., 3(1) : 89-93. Souta r, A., 1971 . Mi cr op a leonrolog y of anaerobic sedim enls and rhe Calif or nia Current. In. B. M. Funnell and W. R. Ric d e l ( Edir o rs) , The Mi c rop a leont o lo- gy of Oceans. Cambrid g e Univ e rsit y Pres s, London , pp. 223-23 0 (1 te xl- fig., 5 pis. ) . St a dum, C. J. and Li ng , H. Y., 1969. Trip yle an Radio l aria in deep - sea scdim - ents of the Norweg ian S_e a . Mi c rop a leonto log y, 15(4) : 481-489 (1 pl.) . St ei n m ann, G., 1905. Geolog isc he Beobacht un g e n in den Alpe n. l]. Die Schar- dt' s che Uber falt un g s th eorie und die ge olog isc he Be deut un g dcr Ti e fs e cabsa- lze und der op h io l it isc hen Masseng e ste i n e . Ber. Natu r(orsch. Ges. Freib urg. 16 : 18-67. St ei n m _ann, G., 1925. Gi bt es fos sile Ti ef s e eablage rung e n von erdg e schic h t lic - her. Bedeut u ng ? Geo!. Rundsch., 16 : 435-468. St u rmer, W., 1963. Mi kr ofo s sile n in den Mornscheim er Schic h t e n. Geo!. Bl. N. 0. Bay e rn, 13(1) : 11-13 (1 pl.) . Theye r, F. and Hammond, S. R., 1974a. Paleomag n eti c po larily seq u e nc e and radio l aria n zones, Brunhes to Polarity Ep o ch 20. Earth Planet. Sci. Lett ., 22 : 307-319 (7 tex t -figs .) . They e r, F. and Hammond , S. R., 1974b. Cenozoic mag n eti c tim e scale in de-cp- sea cores:comp le ti on of th e Neog e ne. Geolog y, 2(10) : 487-492,4 tex t- figs .) . Thomp s on, D'Arcy W., 1942. On Growt h and Form. A New Edit ion . Cambr- idg c Univ e rsil '; I.. Press, London, 1116 pp.
6 바다 珪藩 (Marin e Di at om s) 6 • 1 서언 바다 규조는 석회질 미화석이 결핍되어 있는 고위도 지역의 고생물학적 문제를 푸는 데 특히 유용하다. 규조류는 생물충서학과 고생태학의 중요 한 연구 대상이다 .• 그럼에도 불구하고.이 분야는 아직 충분한 연구가 부족한 편이다. 규조류는 해수의 화학성분, 고영도, 고수심, 고온도 및 영양영류에 관한 정보를 주며,퇴적물 내의 규조류는 고해류의 위치와 방향을 아는 데 귀중 한 열쇠의 역할을 한다. 규조는 광합성을 하는 단세포 식물이며 어떤 그룹은 준군체 생활을 하 지안 단독으로 살 수도 있다 . 그들은 부유성이거나 고착성이며 해조류와 함께 바다에서 가장 풍부한 식물성 플랑크론이므로 먹이 사슬에 중요한교 량역할을 담당한다. 규조는 원양. 연안 및 당수 환경에 모두 잘 적응한다. 화석 규조는 19 세기 후반에 처음 연구되어 분류학에 많은 공헌을 해왔 다. 1920 년대 의 중기에 가서야 이들의 충서학적 용도가 모색되 었으며 미 국, 소련. 일본 둥지에서 활발한 연구가 시작되었다. 이런 연구는 1875 년부터 시 작된 The Schmi dt Di at o m A tl as” 와 1927 년부터 나온 Hus t edt 의 논문들을 기초로 한 것 이다. 근년의 심해저 시추코아는 특히 규조의 생 충서학에 큰 활기를 불러일으켰으며 앞으로도 해양 퇴적물의 연구는 규조 의 중요성을 더욱 증대시킬 전망이다. 6 • 2 생활사 규조의 세포는 원형질 물질로 채워져 있고 핵은 한쪽 편에 치우쳐 위치 한다. 핵은 작고 여러 형태를 보이며 더 작은 nucleol i가 핵 `속에 자리잡 고 있다. 핵 의 주변을 유색 체 (chromato p h ores) 가 둘러싸고 있으며 이들 은 종에 따라 숫자, 크기, 모양이 다양하다. 바다에서 규조군은 유색체에 의해 독특한 색을 띤다. 일 년 중 한때 규조류는 지방질 물질을 생산해서 몇 개의 구형체의 형 태로 세포 안에 저장한다. 아마도 이것은 겨울 동안 생존하기 위한 양분 의 저장 방법으로 생각된다. 규조류는 fru stu l e 이 라고 하는 외 각부를 가지 며 이 는 두 개 의 valve 로 구 성되어 그 안의 원형질을 감싸고 있다. Frus t ule 은 오팔란 실리카로 이루
어졌으며 큰 valve 를 ep ithe ca, 작은것 을 hy pot heca 라고 부른다. 두 valve 는 접촉부가 gi rdle 이라고 부르는 얇은 원형대로 접합되어 있고 보통 화석에 서 이들은 분리되며 gi rdle 은 보존되어 자주 관찰된다. 현미경 아래서 규조를 관찰할 때는 valve fa ce 를 위로 술라이드에 고정 시키는데 이것을 valve vi ew 라 하며 대부분의 분류는 이를 통해 이루어전 다. 규조의 대칭성은 세 개의 축을 따라 형성된다 . valve 의 장 축 방향의 apica l axis, 직각인 tr ans apica l axis 그리고 두 개의 valve 중십을 지나는 perv alval axi s 가 그것이다. 보통 화석은 완전한 상태의 f rus t ule 이 없으므 로 앞의 두 개 축의 대칭성이 논의된다. Hendey ( 1964) 의 연구에 의하면 세포벽은 두 가지 의 형 태를 가지는데 실 리카절 단일충으로 되어 있는 형과, 두 개의 충 이 검으로 있고 그 사이를 직각인 실리카 slats 로 엮어 져 있는 locular wall 형 이 있다. 규조는 단순한 세포 분열에 의해 증식된다. 분열하기 전에 ep ithe ca. 9-} h YJX)t heca 가 열려지면서 두 valve 가 유리된다. 원형질이 노 출 된 쪽으로 새 로운 valve 가 생성되어 먼저 있던 valve 와 짝을 이룬다. 이때 새로 생기는 쪽은 hypot heca 가 되 기 때문에 분열을 거듭할수록 각 의 크기 는 감소된다 (그림 1 참조)· 크기의 감소와 함께 각의 형태와 표면 구조에도 번화가 수 반된다. 이리하여 최소의 크기와 생명력을 유지할 때까지 분열이 진행되 거나 어떤 경우에는 적당한 수준에서 일단 멈추어 auxos po re 를 형성한 후
 ·=기 |H Gl |D CI IF E 广 B
·=기 |H Gl |D CI IF E 广 B
다시 원래의 크기로 환원하여 새과정을 반복한다. 생식 주기 의에도규조 는 한 생육기에서 다음 생육기까지 살아 남을 수 있는 능력이 중요하다. 대 부분의 종들은 조건이 나쁠 때 무거운 포자륭 형성해 바닥에 안주하다가 다음 생육기에 활동을 시작한다. 규조류의 영양영류로 중요한 것은 물의 인산질, 질산질 및 규산질 물질 이다. 야외 및 실험실의 관찰에서는 이 세 가지 물질 중 한 가지만 결핍 되어도 규조류는 생석과 성장을 중지한다. 특히 용승류가 있는 해역이나 연해성에는 많은 양분이 모여 큰 군집을 이룬다. 이 밖에도 종에 따라서 는 cobalami n ( 비 타민 B tz ), thiam i n (비 타민 B1 ) , S, Fe, Mn 그리 고 회 원소성 분 둥의 공급이 생식에 중요한 경우도 많다. 6 ·3 형태와 분류 규조의 형태는 대단히 다양하기 때문에 간단한 분류군을 재시하기가 매 우 어렵다. 현재 대부분의 규조학자들은 19 세기의 선배 학자들이 구분한
 그림 2. i(-조류의 외형
그림 2. i(-조류의 외형
두 개 의 큰 그룹인 Cen t rales 와 Pemales 를 분류의 기본으로 삼고 있다. Cen t rales 는 원형, 타원형, 아원형, 삼각형, 사각형 둥 다양한 외형을 가지나 그들의 표면 구조가 거의 중십접을 갖는 배열을 하는 특칭을 가진 E 卜. 반면에 Pennales 는 장축을 갖는 신장된 형 태 에 장축에 직 각 인 두 짧은 축 의 구조의 결합을 갖는다(그림 2 참조). Pennales 는 다시 봉선 (r ap he) 의 유무에 따라 세분되는데 이 밖에도 각의 기하학적 형태 및 표면 구조가 중요하다. Hende y (1964) 는 일곱 개의 〈형태 그룹〉을 제안하고 이들을 다시 두 개 의 기본 그룹으로 묶었다. 이들은 r ap he 나 p seudora phe 의 유무에 따라 대 분된 것 이며 ra p he 를 갖는 것들에는 (D line ar diat o ms @ cuneate diat o ms @ cym b if orm diat o ms © carin o id di a t oms 가 포함되 며 rap he 가 없는 그룹 에는 (D disc oid diat o ms @ gonioi d diat o ms @ solenoid di a t oms 로 세분 하였다 (그림 3~7 참조). 6 ·4 생태와 고생태 해양 환경에서 규조류는다양한적응울보인다. 저생 규조는 연해에 살며 고착성이거나 이동성이며 천해에 사는 포식생물의 중요한 먹이가 된다. 어떤 종류는 접액을 분비하거나 해저 표면에 단단한 매트를 형성해서 되 적물의 침석이나 번호H 문 방지하기도 한다. 부유성 규조류는 원양성이거나 연안성이며 전자는 생활사의 여러 과정 울원양에서보내는반면후자는 생활사내에 플랑크론과저서성인단계 를 거치므로 물이 얕은 대륙붕 지역에 서생한다. Hendey( 1 964) 는 연안성· 플랑크론을 holop la nkto n ic, melop la nkto n ic 그리고 ty chop lank t on i c 한 습성을 갖는 종들로 구분하였다 (그림 8 참조). 원양에는 이러한 다양한 군집들이 함께 발견되는 경우가 있는데 이는 해저류에 의 해 수 km 이상 운반된 것으로 보인다. 규조류는 부유물의 상태로 해수 중에 포함되어 있다가 해저류나 표층류 에 의해 상당한 거리를 이동할 수 있다. 한 예로 남극해의 규조가 적도 지방까지 확산되는 경우도 있다. 또한 지역적으로 다른 용해도 때문에 퇴적물 내의 화석 군집도 교란을 받는다. 예로서 적도 지방에서는 용해도가 높은데 한 가지 종만 발견되는 표본은 다 론 종들이 모두 녹아 버린 것으로 추정할 수도 있다. 이러한 bio c oenosis
 Cocconci ,
Cocconci ,
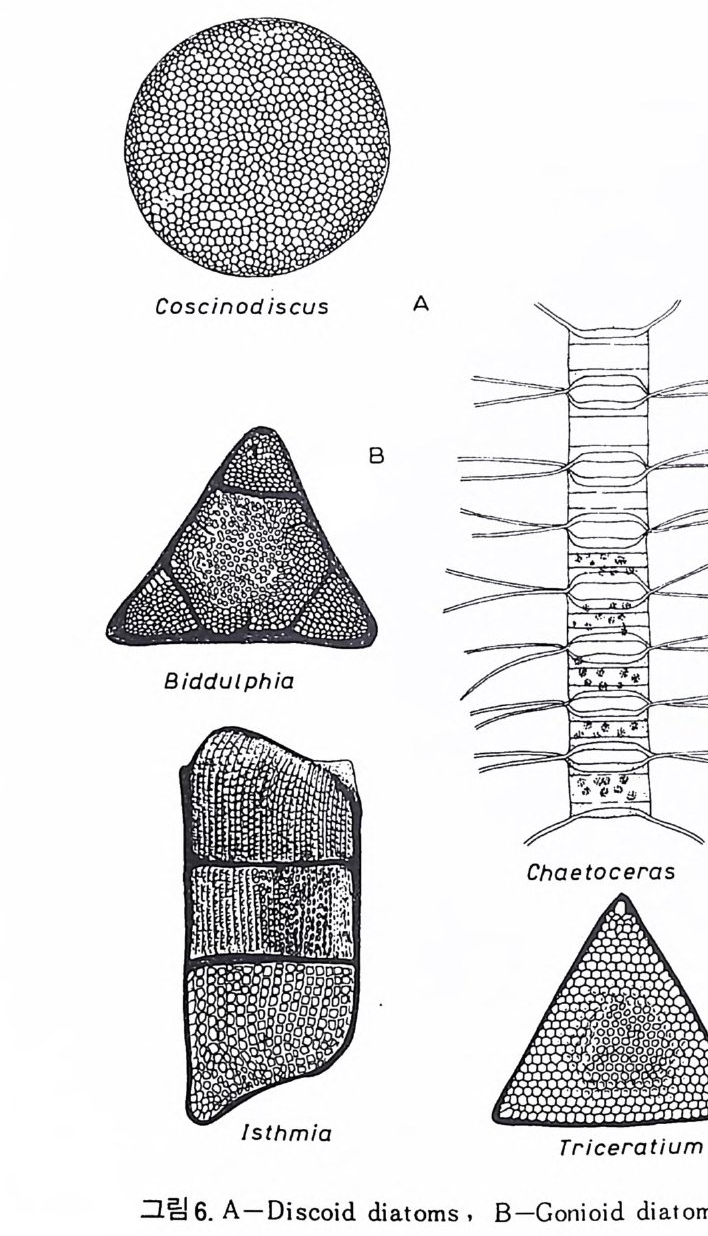 Coscin odi sc us A \
Coscin odi sc us A \
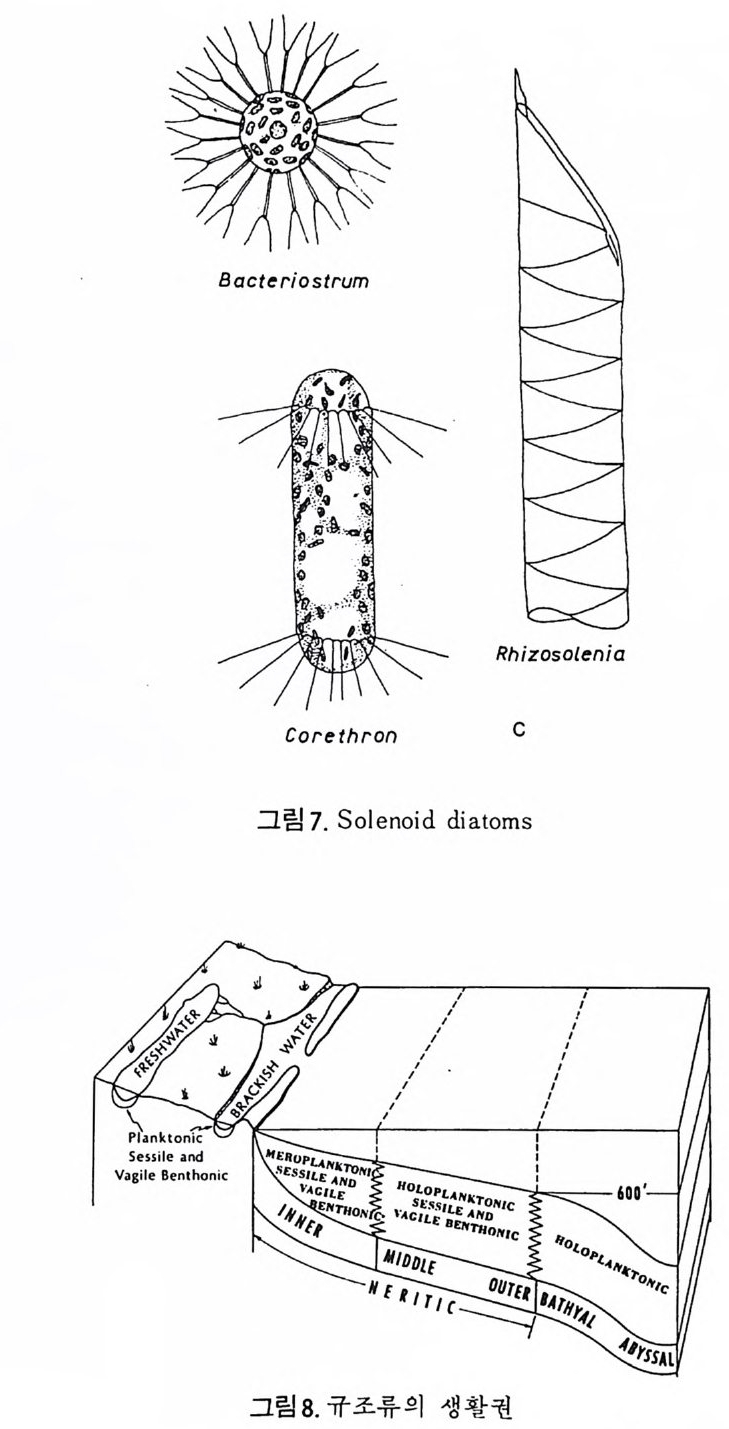 Bacte rio str u m
Bacte rio str u m
와 th anato coenosis 양쪽의 교란은 고생태를 연구하는데 대단히 불리하며 따라서 규조의 고생태학은 〈정량적인〉 고생태학이 되어야 한다. 좋은 결 과를 얻어내기 위해서는 여러 가지의 통계적 처리가 불가피하다. 한 샘플 내의 성분 분석을 정량적으로 실시하기 위해서는 최소한 300 개 체 이상의 규조를 선별해야 하며 종의 다양도가 높으면 그만큼 대상 개체 의 수를 늘인다. 종에 따론 상대적인 성분비는 시간이나 공간에 따라 그 변화가 추적된다. 규조 군집의 번화를 초래하는 여러가지 생태 요소들의 상대적인 중요도를 밝히기 위해서 Td 의 값을 이용한다. Td=~Xl O O 여기서 X wi즌 난류 규조종의 전체수. Xe 는 한류 규조종의 전체수이며 따는 규조 온도로 난류와 한류 군집의 상대적인 양을 표시한다. 규조 군집의 구성원 을 위도, 수괴, 그리고 수온 둥과의 관계로써 밝힐 팔요가 있다. 화석군집을 남북번화에 따라 종수의 그룹으로 구분하기 위 해서는 reccurrent 그룹분석을 한다. aab = ./ NAJNAB B ./ Na1 여기서 NA 는 종 A 의 산출 총량, NB 는 종 B 의 산출 총량, 그리고 J AB 는 종 A 와 종 B 의 총량이다. NB>NA 이고 aab~0.5 이면 종의 묶음이 유사성을 보이는 동일 그룹에 속한다고 간주된다. 화석군 (assoc i a ti on) 을 산정하기 위해서는 군집분석 (cluste r anal y s i s) 을 사용하며 Imbr i e 와 Kip p( 1971) 으| Bio l ogi ca l Resp:> n se Model 이 앞으로 많이 사용될 전망이다. 이는 요인분석의 방법을 이용해서 어떤 환경요소에관련 된 종의 그룹을 알아낸다. 수온이 플랑크돈의 분포에 가장 큰 영 향을 주므로 수온의 종속성이 주로 탐구되지만 이 방법은 영도나 양분공급과의관계를 밝히는 데도 이용된다. 대부분의 연구는 풀라이스토세의 수온 번화를 규명하는 데 집중되어 왔 다. 예를 들면 Kanay a 와 Koi_z u mi (1966) 는 recurrent gro up anal y s i s 와 Td 의 값으로 북태평양의 수온 번화를 추적했다. 한편 적도 태평양 지역을 대상 으로 구해진 Td 의 값을 다론 방법들에 의해 밝혀진 수온 번화 결과들 과 비교되었다(그림 9 참조). 동부적도 태평양과 같이 용승류가 있는 곳은 영양 공급이 잘 되는 환경
을 조성해서 각의 크기가 현저하게 커진다. Kol~ e (l954) 가 관찰한 한 종의 경우크기가 10 배 이상의 번화가 있었 다. 이렇게 물의 순환과 용승류의 발달은 규조의 크기와 밀접한 관련을 갖 는다. 빙하기에 해류의 순환이 신속해지고 이에 따라 활발한 용승작용 이 있었던 때의 규조는 크기가 증대했을 것으로 추정된다. 앞서 말한대로 저탁류와 같은 해저류는 퇴적물을 이동시켜 고생태학적 해석을 교란시키는 경우가 있다. 화석 군집의 보촌 상태를 관찰하고 또 퇴적물의 퇴적구조를 조사하면 이들이 원지성인지 아닌지를 알 수 있다. 반면에 남대서양의 해저 시추 코아속에 보존된 규조 화석군의 경우처럼 기수성 규조류는 해수준의 변동을 비롯해서 수온과물의 화학성분 변화둥 을 추정 가능케 해준다. 화석 군집 의 번화로부터 추정된 해 안선 번화동 생 태 환경 변화는 나아 가서 기 후 의 지시자 역할을 한다. 담수종들이 해양 퇴적물 내에 포함되어 있을 경우 이들은 육원 쇄설물 기원임을 지시한다. 호수와 같은 육수에 살던 담수종이 기후의 변화로 건조되면 바람에 의해 해역으로 운반될 수 있다. 해저 코아에서의규조군 집의 성분 번화 륭 면밀히 검토하면 육지의 기후 번화를 추 측 할 수 있다. 규조의 만우] 되적물량 당 개 체수 및 종의 수는 표면수의 생산량을 지시 한다. 종 다양도는 수백 개체 륭 임의 로 선 별 해 종 수 로 다 양도 룽 대신 해왔는 데 이 방법은 종간의 상대적인 개체수의 성분비가 무시되었으므-로 우리 는 새로운 종 다양도 지수를 개발했다. Shannon-W i ener 의 다양도 지수는 s Hs= -~ P, log e P, £ = I 이다. 이때 S 는자유롭까 선별된 300 개체 중에 포함된 종의 수, P t는 0~ 1. 0 으로 산출된 i번째 종의 상대적 인 양, 그리고 l o g e P; 는 P i 의 로 가리 즘 이다. H 의 값이 높으면 다양한 종의 분포를 지시하며 0 이 되면 단일종을 의 미한다. 적도 태평양에서 빙하기 동안의 높은 다양도는 해수의 활발한 순 환을 지시한다. 이 밖에도 해류에 의한 규조류의 확산을 이용해 서 고해 류를 추적할 수도 있다. 6 • 5 생물충서 및 진화 플랑크론 규조유는 시간-충서 대비에 훌륭하게 이용된다. 생물계의 위

 Di /AT OM ZOINI ES
Di /AT OM ZOINI ES
기타고 불리는 백악기와 고제 3 기의 경계부에는 여러 규조류의 소멸과 출현이 교차되는데 Pennales 그룹이 이때 처음 나타났다. 특히 마이오세와 그 이후의 지층들에 대한 생충서학적 연구는 Schrader(1973) 에 의해 실시 되었다. 대륙봉지역과 십해저는 모두 규조류를 함유하므로 두 지역의 지 충들의 대비가 가능하다. 일반적으로 신제 3 기 해성충에서 형성되는 규조군집은 마이오세 초기의 저위도, 중위도등 지역과 시대에 따라 독특하게 형성되며 많은 표준화석 둘은 시대 구분의 열쇠가 된다.
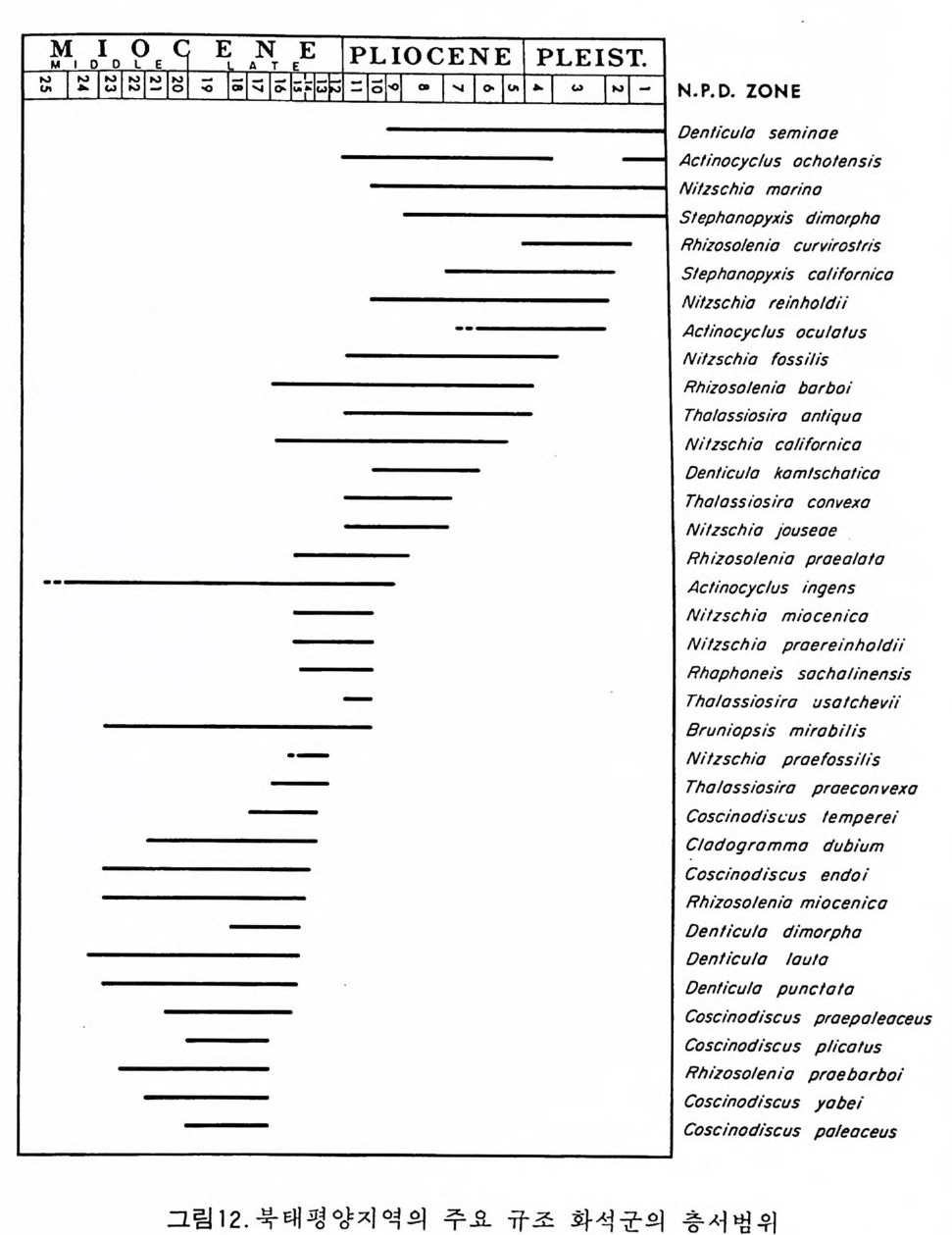 MM I 0 I
MM I 0 I
각을 갖는 규조는 중기 백악기부터 발견된다. 이때 이미 종이 다양한것 으로 보아 중생대 이전에 출현하였지만 각이 없는 원형질 상태로 생활했을 지도 모론다. 백악기의 갑작스러운 각질 규조의 출현은어떤 생화학적인 원인보다는 선택적인 해수에 의한 용해현상에 기인하는 것으로 보인다.백 악기 의 규조는 Cen t rales 이며 Pennales 는 고제 3 기 에 출현하여 에오世에 다 소 번성하였다 . T app an 과 Loeb li ch(1971) 는 Cen t rales 의 확산이 에오世에 안정되었고 제 3 기 동안 마이오세를 매고는 대체로 번화가 없다는 것을 지적했다. 반면 에 Pennales 는 마이오세 이후 지수적 인 증가을 해 서 현재 Cen t rales 를 흰 싼 능가하게 되었다. 전자는 주로 저서성인 반면 후자는 부유성이다 . 현재의 대부분의 담수종은 p enna t e 형이며 이들은 에오世에 해수로부터 기수를 거쳐 담수로 이동한 것으로 보인다. 불행하게도 규조는 시간이 지남에 따라 구조의 복 잡 성이 나타나지 않기 때문에 구조의 번화를 충서학적인 도구로 쓰지는 못한다. 규조의 지리적 격리는 유전자 풀 성분의 번화를 통하여 어떤 형질의 번 화를 초래한다. 부유성 군집인 겅우 많은 종들이 특정의 수괴에 제한적으 로 서식함으로써 어느 정도의 지리적 격리가 야기된다. 그러나 설 명이 어 려운 정은 같은 수괴에서 종의 전화가 일어나는 것이다. 단지 장기간에 걷찬 미미한 환경 요쇼의 번화가 형질 번화의 원인이 아닌가 추 측 하고 있 을 분이다. 후기 마이오世에서 현세까지 적도 태평양에서 6 종의 규조의 진화를 관찰할 수 있다. 그 중 한 종은 현재까지 생존하지만 나머지는 모 두 철멀하였으며 평군 생촌기간은 50 만년 정도이다. 이러한 사실로 새로 운 종은 다소 유전적으로. 불안정하거나 환경의 접전 적 인 번화에 의해 전화 가 일어난 것을 알 수 있다. 보충교재 lle11de y , N.l. , 1964. A11 i11t ro d 1 1c to ry acc ou nt of the sma ll e r alga e of Brit ish coast a l wat e rs. Fi sh . Invest. Ser. 4, Part 5, Bacil l ario p hy c eae( Di at o m s) , pp.l -317(London). [A lt h oug h a11 accou111 of dia t o m s in the north east At la- nti c, thi s book has an exc el len t int ro ducti on to dia t o m tax o n omy , bio l og y, ec-olog y a11d meth o dolog y.] Huste d t, F., 1927-1964. Die kie s el alge n Deut sc h lands , Oste r re i c h s un
enzenden Me er es-Gebie te . In: Dr. Rabenhorst' s Kryp tog a men-Flora van De-
uts chland, Oeste rreic h s und der Schweiz . 7, I , Sect. 1-5, pp. 1-920. (19 27 -1 930) ; II , Sect. 1-5, pp. l-73 6 (1931-1937) ; ill. Sect. l 一 3, pp. 1-556 (19 61-1964) . Le ip海 [A source book in liv i n g marin e and fre sh-wate r dia t o ms. Morph olog y and ecolog y are str essed .) Koiz u mi , I., 1975. Ne og e ne dia to ms from th e Weste r n Margi n of the Pacif ic Ocean, Le g 31, De ep Sea Drill ing Proje c t, Init ial R ep. Deep Sea Drill in g Proje c t, 31 : 779-81 9. [A key pa p e r st r essin g bio s tr a ti gr ap h ic correlati on of Eq u ato ria l and North Pacif ic dia t o m zonati on s.) Pat r ic k , R. and Re im e r, C., 1966. The dia to m s of th e Unit ed St a te s, 1-M o- nog r . Acad. Nat. Sci. Phil a ., 13 : 1-688. [A book on fre sh-wate r dia t o - ms wi t h excell e n t int rod u ct o ry cha p te rs on vario u s aspe cts of dia t o m bio l o- gy and dis t rib u t ion . ) Si m onsen, R. (E dit o r ) , 1972. Fir s t sym p o siu m on re cen t and fos sil marin e di- ato m s . Nava He dwi gia, 39 : 1-294. [S ome fiftee n cont r ib u ti on s on vario u s aspe ct s of fo ssil and liv i n g marin e dia t o ms.] 참고문헌 ArEr_hx pe en diu ist , i o nG (. 1, 91497 5—2 .1 9S 4 8e )d, i m R e en p t . , c5o r:e 1s -8fr9om. th e Eas t Pacif ic. Swed. Deep - Sea BloInw:, P.W B . rHo.r,m i1 m9 6an9n. aLnadt e H .M Hi d. d Rlee nEz o (cEed niet ot ros} ,r ecPernoct . pFla in rks tto nIinc t . b iCo s otnr [a . t iP glra anpkh t y- . onic M ic rofo ssil s , Ge nev a 1967 . Brill , Leid e n, 1 : 199-42 1 . Burck l e, L. H., 1971 . Correl a ti on of Late Cenozoic marin e secti on s in Jap a n and th e Eq u ato ria l Pacif ic. Trans. Proc. Paleont o l. Soc. Ja p. , N. S ., 82 : 117-128. Burckle, L. H., 1972. _La te Cenozoic p] ankto nic dia to m zones from th e easte - rn Eq u ato ria l Paci f i c. In : R. S im onsen; (Edit or ) , Sy m p o siu m on Recent and Fossil Marin e Di a to ms. Nova Hedwi gia, 39 : 217-256. Burckle, L. H., 1975. Di a to m evi de nce bearin g on th e correlati on of th e Rel- izi a n sta g e , Califo r nia . Pac. Geo!., 9 : 33-34 . Burckle, L. H. and Bi sc ay e , P., 1971 . Sedim ent tra nspo rt by Ant a rcti c Bo- ttom Wate r th roug h th e ·ea ste rn Rio Grande Ri se . Annu. Meet. Geo!. Soc. Am., pp. 518 -519 (abst r act) . Donahue , J., 1970. Di a to ms as Qu ate rnary Bi o str ati gr aph ic and Paleoclim at- ic Indic ato rs in Hi gh Lati tud es of th e Pacif ic Ocean Thesis , Columbia Un- ive rsity , New York, pp. 1-23 0. Emi lia n i, C., 1966. Paleote m p e ratu re analys is of th e Carib b ean cores P6304 - 8 and P6304-9 and a ge neraliz e d tem p e ratu re curve for th e pa st 425, 000ye ars. J. Geo!., 74 : 109 -1 26. Eric s on, D. and Wollin , G., 1968. Pleis t o cene clim ate s and chronolog y in de- ep- sea sed im ent s. Scie n ce, 162 : 1227-1234. Hays , J. D., Sait o, T., Op d yk e, N. D. and Burckle, L. H., 1969. Plio c ene-
Pleis to cene sedi m ent s of the Eq u ato ria l Pacif ic: Their pa leomag n eti c, bio s t- rati gr ap h ic and cli!l la ti c record. Geol. Soc. Am. Bull., 80 : 1481-1514. Hendey, N. I., 1964. Int ro ducto ry account of the smaller alga e of Brit ish co- asta l wate rs. Fi sh . Invest. , Ser. 4, Part 5 , Bacil l ario ph y c eae( Di a to ms) ,pp. 1-317{London) . Imb2·ie , J. 떠i d Kip p. N., 1971 . A new mic r op a lconto log ica l me th od for qu anti - tal iv e pale oclim ato log y: app li ca ti on to a late Pleis t o cene Carib b ean core. In: HK.a vKe. nT, uCreoknnia .n, p p(E. di7t 1or— ) ,1 8L1 a. te Cenozoic Glac ial Ag e s. Yale Univ ersit y, New Jo use, A. P., Koroleva, G. S. and Naga eva, G. A., 1963. St ra ti gr ap h ic and pa leog e og r aph ic research in the Ind ian Ocean secto r of th e Ant a rcti c. Oke-anol. Issled., 8 : 137-1 60 (in Russia n ) . Kanaya , T. and Koiz u mi , I., 1966. Int e rpr eta ti on of dia to m th anato coenosis from th e North Pacif ic app li e d to a stu dy of core V20-130 (stu die s of a de-ep- sea core V20-130, Part N). Tohoku Univ . Sci. Rep ., Ser.2 , Geo!., 37 : 89-130. Kil h am, P., 1971 . A hyp o th esis concernin g sil ic a and th e fres h-wate r pla nkt- onic dia t o ms. Li m nol. Oceanog r ., 16 : 10-18. Koiz u mi , I., 1968. Terti ar y dia to m flor a of Og a Penin s ula, Akit a Prefe ctu re. north east Ja p a n. Tohoku Univ . Sci. Rep. , S er, 2 Geol., 40 : 170-24 0. Koiz u mi , I., 1972. Marin e dia to m flo ra of th e Plio c ene Tats u nokuchi Forma- tion in Fukushim a Prefe c tu re. Trans. Proc. Paleo~t o l. Soc. Ja p ., N. S., 86: 340-359. Kolbe, R. W., 1954. Dia t o ms from eq u ato ria l Pacif ic cores. Sued. Deep - Sea Ex p ed iti o n{ 1947 一 1948), Rep ., 6 : 183-196. Kozlova, 0. G., 1964. Di a to ms of th e India n and Pacif ic Secto rs of th e An- ta rcti c . .A cad. Sci. U. S. S. R., Inst. Oceanol. , Israel Prog r am for Scie n ti - fic Translati on s, Je rus alem, pp. 1-191. McCollum, D. W., 1973. Neog e ne ge nus Trin acria as a str ati gr ap h ic marker in sout h ern ocean sedim ent s. Ant a rct. J. U. S., p. 198. Mi tch ener, C. U. and Sokal, R. R., 1957. A qu anti tat i ve app r oach to a pr ob!- em in classif ica ti on . Evoluti on , 11 : 130 -162. Nakaseko, K., Koiz u mi , I., Sug a no, K. and Maiy a , S., 1972. Mi cr obio s tr ati - gr aph y of th e Neog e ne for mati on s in th e Nadaura area, Toy a ma Prefe ct- ure, Ja pa n. Geo!. Soc. Ja p. , Bull., 78 : 253-264. Schrader, H., 1973. Cenozoic dia t o ms from the North east Pacif ic, Leg 18. In : Init ia l Rep o rt of th e Deep S ee Dril l in g Proje c t, Leg 18 : 673 -797.S im onsen, R., 1969. Di at o m s as ind ic a to r s in estu a rin e envir o nment s. Vero{{. Inst. Meeresfo r sch. Bremerhaven, 11 : 287 -2 92. Si m onsen, R., and Kanay a, T., 1961 . Not es on the marin e sp e cie s of the di- ato m ge nus Denti cu la Kut z. Int. Rev. Gesamt en Hy d robio l ., 46 : 498-513. Tapp a n, H. and Loeblic h Jr. , A. R., 1971 . Geolog ic imp li c a ti on s of fos sil p h - ytop la nkt on evoluti on and tim e-spa ce dist r i b u ti on . In-: R . Kosanke and A. T. Cros s (Edit or s) , Sy m p o siu m on Palyn olog y of the Late Cre t ace 야 s and Ea- rly Terti ar y. Geol. Soc. Am. , Sp e c. Pap. 127 =· 247 -3 40. W or nardt, W. W., 1969. Dll l.io m s , pa st, pr esent and futur e. In: P. Bronnim a- n:1 :::.mt I!. H. Renz (E d itor s) , Proc. Fi rs t Int. Con{. Plankt on ic Mi cr of os s- ils , Geneva, 1967. Brill , Leid e n, 2 : 690-714. 7 珪質靴毛蟲 및 에브리디안 (S ili co fl a g ella t es & -Ebrid i a ns) 7 • 1 서언 규질 편모충류와 ebr idi ans 는 석회질 미화석이 없거나 충분치 않은지층 에서 고해양학이냐 고기후의 복원에 중요한 역할을 하는 플랑크톤이다. 백 악기 초 기에 출현하여 현재에 이르기까지 별로 형태적인 변화가 없이 생 존해 왔다. 현재 해양에서 규질 편모충은 전체 식물성 플랑크론중에는 바 교적 작은 성분 그룹을 이룬다. 화석 규질 편모충과 ebr idi ans 는 함께 산출되며 관습적으로 함께 취급 연구되어 왔다. 근본적인 골격 구조의 차이분만 아니라 이 두 그룹에는 다론 중요한 차이가 있다. 전자는 법세계적 분포에 광합성을 하는 독립 영양체인 반면 후자는 보통 온난~한냉 해역에서만 생존하며 또한 발색 단을 갖고 있지 않기 때문에 종속 영양체이다. 다론 대부분의 미화석 의 경우와 같이 독일의 Ehrenber g가 19 세 기 중영 현생과 화석종을 관찰 기재하였다. 1890 년 Bo ga r t에 의해 규질 편모충은 규조나 방산층으로부터 분리되어 S i l i co flag ella t a 目으로 독립하였다. 그 후 광범위한 해저 시료의 연구로 1960 년대까지 분류체계가 잡히고 최근의 연 구는 주로 그들의 고생태 및 생물 충서에의 응용에 집중되고 있다. 늦은 전화 속도에도 불구하고 규질 편모충은 후기 신생대의 고위도 지역의 충 서 대비에 성공적인 역할을 하고 있다.
7 • 2 생활사 규질 편모충은 단세포이며 20~50µm 의 크기를 갖는 해생 편모충이다. 두명한 원형질의 주요 부분은 규질인 골격 안에 위치하고 있지만 위족, g- ranules, 유색체를 포함하는 외층과 단 한 개의 편모는 골격 밖에 위치한 다. 세포는 둥근 핵과 광합성을 위한 황색이나 녹색의 유색체 를 포합한다 (그림 1 참조). 골격은 오팔린 실리카로 구성되어 있으며 種에 따라 형태가 단순한 고 리상부터 복잡한 도움상까지 다양하다. 그러나 기본적인 고리 (basal rin g ) 룰 모두 보유하면서 좀더 복잡한 부분을 한쪽 면에 형성시키므로 대부분 의 종들은 basal r ing에 대해 비대칭형이 된다. 따라서 전 체 적인 외형은반 구상으로 되고 방사상 대칭형을 이 룬다 . 골격의 기본을 이 루는 튜브상의 막대는 속이 비어 있으며 대부분 표면은 평평하지만 종류에 따라서는 고 배율 전자 현미경 아래서 격자상 혹은 r idg e 의 구조가 보인다(그림 2, 3 참 조)
 그림 1. 규질 편모충인 Di.ct y o cha fibul a Ehrenber g의 살아있는 세포
그림 1. 규질 편모충인 Di.ct y o cha fibul a Ehrenber g의 살아있는 세포
 그림 2. 튜브상 막대 구조 를 그림 3. Di ct y o cha fibu la
그림 2. 튜브상 막대 구조 를 그림 3. Di ct y o cha fibu la
유색체의 촌재는 이들이 독립 영양성으로노 광합성에 의해 석량을 제조한 다는 것을 뜻한다. 그러나 위족을 이용해서 먹이를 포획할 가능성도 있으 나 배양 실험에서는 관찰되지 않았다. 녹조로 된 공생석물이 어떤 종에서 발견되었으며 이들이 영양분을 섭취하는 데 도움을 주리라고 사료된다. 규질 편모충은 식물형의 분열을 하는데,모체와 꼭 같은모습으로 새로운 골격 이 생성되는 basal ring쪽으로 원형질 물질이 유출되는 모습이 관찰 되었다. 아직 다른 형태의 생식은 알려져 있지 않지만 이들도 다론 식물
성 편모충류에서와 같이 유성생식의 가능성도 있다. 규질 편모충은 되적물로부터 직접 smear sl i de 를 제작하여 관찰하기도 하며 되적물 내의 모든 다론 성분들율 화학적으로 제거시킨 후 규질 성분 만을 남겨 이들을 농축시켜 관찰한다. 상세한 시료 처리법은 Mandra 의 ( 1973) 를 참조하라. 7 ·3 형태 및 분류 규질 편모충류는 플랑크론 조류의 한 上움으로 황녹조류강의 한 目으로. 취급한다. 여러가지 기재적인 용어들이 그립 5 에 소개되어 있다. L ipp s(1970) 는 이들의 기능형태학적인 분석을 토대로 하여 분류체계를 정리하였다. 그의 주장에 의하면 전정한 규질 편모충은 8 개 의 그룹이 있 다. 다른 것들은 기본구조에 있어서 차이가 있을 뿐만 아니라 오히려 규 조나 방산층과 더 유사성이 있다.
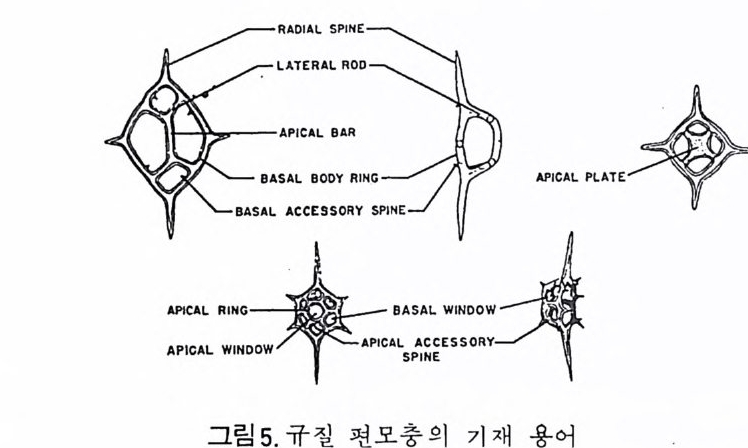 「 ............ -- V • • .,.4
「 ............ -- V • • .,.4
7 ·4 생태 및 고생태 . 대부분의 해양 미부유성 생물과 같이 규질 편모충은 그 군집의 조성과 번성은 해수의 온도나 영도 그리고 영양분의 공급에 밀접한 관계를 가전 다. 이들은 범세계적 인 분포를 보이며 수온이 위도에 따른 군집 형성에 가 장 중요한 요인으로 보인다. 용승류가 형성되는 곳은 영양 공급이 좋을 분만 아니타 실리카 성분이
풍부한 저류층이 공급되므로 이들의 골격을 형성하는 데 유리하다. 해수 의 용승 작용은 대기와 해수의 순환과 밀접한 관련이 있으므로 과거지질 시대의 해양의 생산성이나 기후 번화와 같은 고생태학적인 환경 복원에 이 둘 그룹은 중요한 도구가 된다 . Gemein h art (1934) 는 D i cty ocha 와 Di sl eph a: nu s 腦둘이 각각 온난해수와 한 냉 해 수에 적 응하는 사실을 관찰한 후 Dic tyo cha I Di st ( 'ph a: nu s 의 비 율을 현 세 퇴적층의 고기후 해석에 이용하였다. 이 원리는 해저 시추 코아의 연 구에도 그대로 적용되어 수온이나 해류의 번화 등을 추적할 수 있다. 많은 학자들은 수온 대 屬의 구성비율을 그래프로 표시하여 고생태 자료로 삼 는다 . 최근 Poelchau(1974) 는 생물 군집이 단순히 형성되는 것이 아니라 포식 자에 의한 선택적 포석, 수괴 사이의 생산성의 차이와 번화 그리고 종간 의 고유현상 둥의 요인 등이 군집 의 성분에 중요한 번화를 일으킨다고 주 장했 다. 이러한 복잡성에도 불구하고 규질 편모충은 아직도 고기후와 고생산성 연구에. 특히 정성적인 해석에 있어서 중요한 역할울 한다. 7 • 5 생물충서 및 진화 가장 오래된 규질 편모충은 초 기 백악기의 지층에서 발견되었다. 후기 백악기엔서는 D icty och 社 s, Corb i sem ia s 및 Vallacer t a 가 혼히 발견되며 Vall- acer t a 와 L y ramula 屬 은 후기 백악기 의 시준화석 이다. 다론 모든 종류들은 제 3 기에 모두 출현해서 nav i culo p s i d 는 초기 팔레 오世, d i s t e p han i ds 는 중기 에오世, 그리고 canno pi l i ds 는 울리고世부터 산 출 된다 . Han nait es 屬은 현재까지 에오世에서만 보고되었다. 모든 주요 속둘은 Val/,a c erta, Hannait es 및 Corb i sema 를 제 외 하고는 현세 까지 계 속 번성 하고 있다. 규질 편모충류의 전화 속도는 늦은 편이기 때문에 생물 충서학적 분대 는 다소 어려움이 있다. 계속해서 새로운 종들이 보고되고 있으므로 종의 출현과 소멸을 기준으로 한 전화 속도의 측정이 가능하다. 알려진 연구에 의하면 진화속도는 팔레오世에 다소 빠르고 다음은 중기 마이오세 - 풀라 이오世가 그 전 후기에 비해 비교적 빠르게 나타난다.
생충서학적 분대의 어려움에도 불구하고 다론 미화석군이 없을 때는 규 질 편모충류는 중요하게 쓰인다. 현재에는 마이오-풀라이오세의 상세한 분대를 기대해 볼 수 있다. 7 ·6 에브리디안 규질 편모충과 같은 크기의 범위에 속하는 에브리디안은규질의 골격을 갖는 해양 부유성 편모충이다. 살아 있는 세포는 두 개의 부동의 편모를 가지며 유색체를 갖지 않는다. 골격은 듀브상이 아니며 성분은 방산층과 유사한 그룹임을 앙시한다. Loeb li ch 외 (1968) 는 에브리안이 쌍편모조류와 유사한 핵과 부등의 두개 의 편모를 갖는 점으로 보아 규질 편모충류에 더 가까운 것으로 생각하고 독립된 網으로 묶여야 한다고 주장했다. 7 • 6 • 1 생 활사 및 생 태 학 그림 6 에서 보는대로 세포내의 큰 핵으로나 L 터 두 개의 편모가 뻗어 있 고 골격은 완전히 원형질 안에 위치한다. 이들은 식물과 같은 형태와 유
 그림 6. 에브리디안의 세포 모식도
그림 6. 에브리디안의 세포 모식도
성 생식의 두 가지 방법으로 번식하는 것으로 보인다. 전자의 경우 먼처 새로운 골격이 모세포 안에 형성되고 이어서 다론 세포 구성분이 두 개의 분리 된 개 체로 분열한다. 가끔 새로운 골격이 분리되지 않고 이중 골격을 만드는 경우도 있다. 유성 생석이 있으리라는 추측은 간혹 화석에서 두세 개의 정상적인 개 체의 골격 위에 놓이는 種皮가 발견되기 때문이다. 에브리디안은 종속 영향체이며 주로 규조를 먹고 산다. 대체로 규질 편 모충류보다 혼치 않고 종도 다양치 못하다. 그들은 단지 온난 -한 냉 해수 에서만 비교적 자주 발견된다. 이들의 지리적인 분포에 관해서는아직 알 려진 바 없다. 7 ·6 ·2 주요 속과 생물충서 골격의 형태는 tr ir a d ial 혹은 t e t r axi al 대칭형이다. 어떤 경우골격이 완 전치 못하거나 화석에서는 파편으로 분해되어 나타나므로 분류 체계를 세 우기가 어렵다. 가장 혼한 종류는 t heca t e 형으로 고제 3 기까지 거 슬러 올라간다. 그들은 에오世와 마이오世에 가장 빈번하다 (그림 8 참조). 한편 대부분의 non- t heca t e 형은 t heca t e 형 의 부분적 인 골격 이나 불완전한 형태 들 로서 Cardw f ol i a 屬이 대표적이며 팔레오世-마이오世에 산출된다. 그 러나 이 종류는 현재 쌍편모조류의 규질 외골격인 것으로 밝혀졌다(그림 7 참조) .
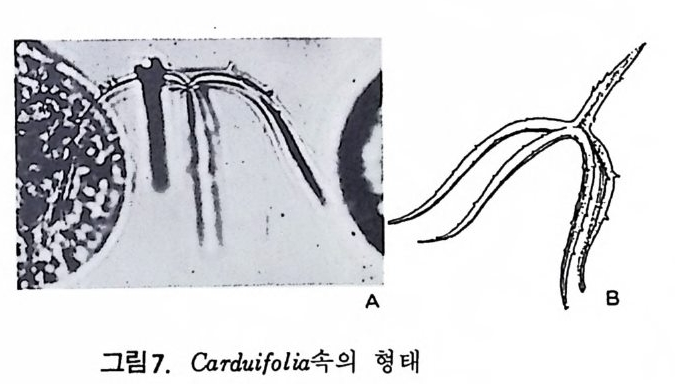 _. -~• ~ `·
_. -~• ~ `·
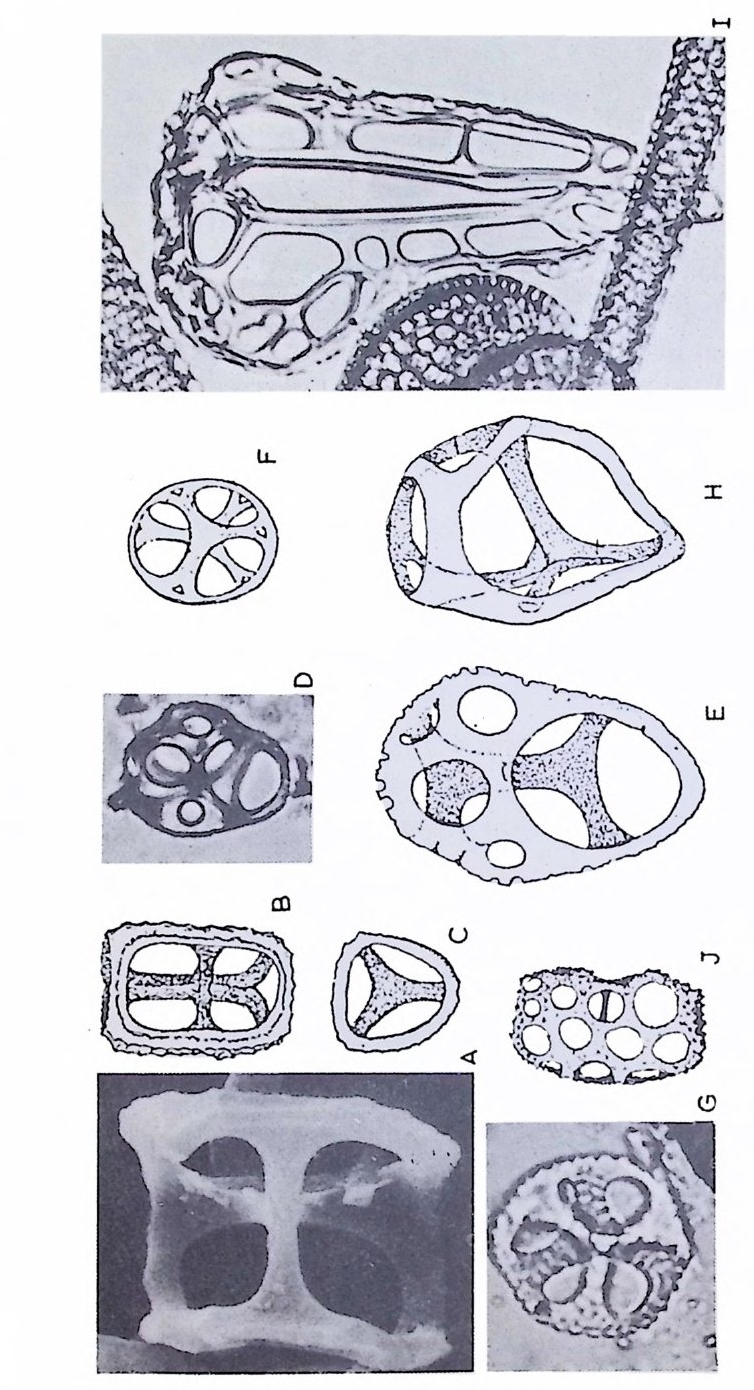
보충교재 Lip ps , J. H., 1970. Ecology and evoluti on of sili co fl ag e llate s. Proc. N. Am. Pa l.eo nto l. Conv., G : 965-99 3. [A go od ge neral int r o ducti on to th e subje c t and a criti ca l revi ew of sili c o fl ag e llate fun cti on al morph olog y and valid i t y of vario u s gen era.] Loeblich III: A. R., Loebli ch , L. A., Tapp a n, H. and Loeblich Jr. , A.R., 1968. Annota te d ind ex of fos sil and Recent sili co fl ag e llate s and ebrid ian s wi th de- scrip tion s and illus tr ati on s of valid l y pr op o sed tax a. Geo/. Soc. Am., Mem. 106 : 1-31 9. [B rie f in tr o ducti on s to sili c o fl ag e llate and ebrid i a n bio l og y, his - to ry of stu d y and ge ologi ca l dis t r ib u t ion is foll owed by descrip tion s of tax a in the orig ina l langu a ge in whic h th ese tax a were firs t describ e d. Orig ina l ii- lustr ati on of spe cie s are also inc luded in pla te s. Includes comp r ehensiv e bi- blio g r aph y unti l 1965.] 참고문헌 Bukry, D., 1974. S t rati gr ap h ic value of sili c o fl ag e llate s in non-I rop ica l reg i- 011s . Ge o! . Soc. Am. Bull., 85 : 1905 一 1906. Bukry, D., 1976. Sil ic o fl ag e llate and coccolith str ati gr aph y, south easte rn Pa- cif ic Oce an , De ep Sea Drill i n g Proje c t. !nit. Rep. D.S.D.P., 34:715-735. BuLkergy , 1D6., aDned e p F oSse tae rD, rJil.l i Hn g ., P19ro73je. c tS. i!l inc io t f.l aRg ee lpla. tDe s. Sa.nDd .Pd.i,a t1 o 6m : s8t1r 5a -ti8 g7r1 a.p h y, Bukry, D. and Fos te r, J. H., 1974. Sil ico fl ag e llate zonati on of Up pe r Creta c -eous to Lower Mi oc ene deep - sea sedim ent s. J. Res. U. S. Geo!. Surv., 2 : 303-310. Cie s ie l ski, P. F., 1975. •Bi o s t r ati gr ap h y and pa leoecolog y of Neog e ne and Oli- go cene sili c o fl ag e llate s fro m cores recovered durin g Anta rcti c Leg 28, De-ep Sea Drill ing Proje c t. !nit. Rep. D. S. D. P., 2 8 : 625-691. Ci es ie l ski, P. F. and Weaver, F. M., 1974. Early Plio c ene tem p e ratu re chan- ge s in th e Ant a rcti c Seas. Geolog y, 2 : 511-515. Defl an dre, G., 1934. Nomenclatu re du squ elet t e des Ebria c ees et descrip tion de qu elqu es for mes nouvelles. Anan . Proti st k d ., 4 : 75-96. Defl an dre, G., 1950. Cont rib u t ion I'etu de des sili c o fl ag e llid e s actu els et fo- ssils . Mi c roscop ie, 2 : 72-108; 191-210. Freng u elli, J., 1940. Consid e racio n es sobre los sili c o fl ag e lados f6 sile s . Rev. Mus. La Plata , Ser. 2, 2 : 37-112. Gemein h ardt, K., 1930. Sil ic o fl ag e llata e . In : L. Rabenhorst, Kryp tog a men-Fl-ora von Deuts chland, Oste rreic h u 뼈 der Schweiz , 10. Akademi sc he Verla- gs ge sellschaft , Leip z ig , pp. 1-87.
Gemein h ardt, . K ., 1934. Die Sil ic o fl ag e llate n des sUdat l ant isc h e n Oze an s. Wi ss . Erge bn. Dt sc h. At la nti sc hen Exp e d. Mete or:• 1925-1927, 12 : 274-312. Hovasse, R., 1932a. Not e pr elim i na ir e sur !es Ebria c ees. Soc. Zoo!. Fr. B u- fl., 57 : 118 -1 31 . Hovasse, R., 1932b. Trois i e m e not e sur !es Ebria c e es. Soc. Zoo!. Fr. Bull., 57 : 457 -476. Hovasse, R., 1934. Ebria c ees, Din o fl ag e lles et Radio l air e s . C. R. Acad. Sci. Paris , 198 : 402 -404. Je nudrer zjoe sw cisl kla i,t i oJn . sP d. eafni dn e Zd abryi l l os ,i li c Go . fl Aag. , e ll1a9t7e 2 .c liLa na gt ee s Pilne ias t o scu eb nae n t.p aar lcetoi ct e d m epe ep r -a ste- a core. Deep - Sea Res., 19 : 327 -329. Li ng , H. Y., 1970. Sil ic o fl ag e llate s from cent ra l North Pacif i c core sed im e n ts . Bull. Am. Paleont o l., 58 : 85-129. Lin g , H. Y., 1972. Up pe r Creta c eous and Cenozoic sili co fl ag e ll ate s and eb rid - ian s. Bull. Am. Pa l.eo nto l., 62 : 135 -229. Mandra, Y. T., 1969. Si lic o fl ag e llate s : A new too l for th e stu d y of Ant ar cti c Tertia r y clim ate s . J. Anta rct. Res., 4 : 172-1 74. Mandra, Y. T. and Mandra, H., 1972. Paleoecolog_ v and ta xo n o m _v of sili c o fl - ag e llate s fro m an Up pe r Mi oc e ne dia t o m i te near San Fe lip e , Baja Califo r nia , Mexic o . 0cc. Pap. Ca l.if. Acad. Sci. , 99 : 1-35. Mandra, Y. T., Brig ge r, A. L. and Mandra, H., 1973. Che m i ca l ext racti on tec - hniq u es to free fos sil sili c o fl ag e llate s fro m marin e sedim e n ta r y rocks. Proc. Calif . A c 교. Sci. , 39 : 273-284. Marshall, S. M., 1934. The Sil ic o fl ag e llata and Ti nt i nn oin e a . Br. Mus. (N at. H i - st. ) , Great Barrie r Reef Exp e d. 1928-1929, Sci. Rep ., 4 : 623 -6 64. Marti ni , E., 1971 . Neog e ne sil ico fl ag e llate s from the eq u ato ria l Pacif ic. /nit. Rep . D. S D. P., 7 : 1696 -1 703. Marti ni , E., 1972. Sil ico fl ag e llate zones in the late Olig o c ene and early Mi oc -ene of Europ e . Senckenberg. Leth aca, 53 : 119-122. Marti ni , E., 1974. Sil ico fl ag e llate -zo nes in the Eocene and early Olig o cene. Senckenberg. Leth aea, 54 : 527 -532. Norris , R. E.,· 1967. Alga l consorti um s in marin e pla nkto n . In:V. Kris h namur- th_ v (Edit o r), Semi n ar Sea, Sa l.t and Plant s. Cent ra l Salt and Marin e Ch-em. Res. Inst. , BhaV11ag a r, India , pp. 178-189. Perch-Ni el sen, K., 1975a. Late Creta c eous to Pleis t o cene sil ic o fl ag e llate s from the sout h ern SW Pacif ic, D. S. D. P. leg 29. !nit. Rep . D. S. D. P., 29 : 677 -721. Perch-Ni el sen, K., 1975b. Late Creta c eous to Pleis t o cen e archaeomonads, eb- rid i a n s, endoskeleta l din o f lag e llate s and oth er sil ic e ous mi cr ofo s si l s f rom the subanta rcti c S. W. Pacif ic, D. S. D. P. Leg 29. !nit. Rep. D. S. D. P., 29 : 873-907.
Pe rc h-Ni el sen, K., 1976. Ne w sil ic o fl ag e llate s and a sili c o f la g e ll ate zonati on in no r th Europ e an Paleocene and Eocene dia t o m i te s. Bull. Ge o! . Soc. De n. , 2 5: 27-40. Po elc h au, H. S., 1974. Holocene Si li c o f la ge llat es of the North Pacif ic, The- ir Di st r ib u ti on 터 Use (or Paleote m p e ratu re Dete r mi na ti on . Thesi s , Uni- ver s it y of Calif or nia at San Die g o , 165 pp. Sc hu lz, P., 1928. Be it ra g e zur Ke nn t ni s fos s ile r und rez ent e r Sil ic o f la g e ll ate n . Bot . Arch., 21 : 225-29 2. Ty n an , E. J., 1957. Si lic o fl ag el late s of the Calve r t for mati o n (M ioc e ne ) of Ma -ryl a n d. Mi c rop a leonto l og y; 3 : 127-136. Van Valk e n burg, S. D. and Norris , R. E., 1970. The gr ow t h and morph o lo g y of the sil ic o fl ag e ll ate Di ct y o cha fibu la Ehren b e rg in cul tu r e. J. Phy c ol., 6 : 48- 54. Worn a rdt , W. W., 1971 . Eo cen e, Mi oc en e and Plio c e n e ma ri n e dia t o m s and sili c o fl ag e ll a te s stu d ie d wit h the scan nin g ele c t ron mi cr oscop e . Proc. II Pl- ank t on ic Con [., Rome , 2 : 127 7 -1300. 8 코노돈트 (C onodont s) 8 • 1 서언 인 산염질 미화석 중 분포가 가장 넓고 생물 충서학적으로 중요한 그룹 은 코 노돈트이다 . 이들은 200µm~6mm 의 크기를 갖는 미지의 생물의 겅 질부이다. 캄 브리아기로부터 트라이아스기까지 산출되며 이 구간의 생충 서 분대는 코노돈트가 가장 중요한 역할을 한다. 시준화석으로서의 가치 는 이들의 형 태가 대단히 다양하며 범세계적 분포를 하는 데서 비롯된다. 1856 년 Pander 에 의해 처음 발견된 후 꾸준한 연구가 지속되 어 오다가 1920 년대와 1930 년대 초에 북미에서 큰 연구 봄이 일어났다 . 이어 분류 체계에 발전이 있었고 1970 년대의 연구는 주로 하부 오르도비스 -하부 캄 브리아기의 시대와 상부 석탄-상부 트라이아스기의 생충서학에 중정을 두고 있다. 상부 백악기와 쥬라기의 코노돈트는 지금 이동되어 온종류로 생각되고 있다. 코노돈트는 인 회 석의 미결정들이 혼재되어 있는 유기질 기질로 구성되 어 있다. 찰 보존된 표본은 백색-호박색에 두명하지만 속성 작용에 의해 대부분은 회색, 흑색 또는 갈색으로 착색되었고 불두명하게 변했다. 인산영질 미화석은 세일을 세 척 함으로써 혹은 석회암울 품은 초산이나
포름산으로 녹여내어 추출해낼 수 있다. 이들은 보통 석회질 미화석보 다 적게 산출되므로 많은 양의 표본이 필요하다. 8 • 2 형태 및 분류 형태상으로 네 가지의 주요 그룹이 구분되며 이들의 외형과 각 부분의 명칭이 그림 1 에 소개되어 있다. CD Si m p le cone ® Bar-ty pe conodont s @ Blade-ty pe concxlont s @ Platf orm conodont s 한 개체에 대한 기재에는 위치에 대한 방향 표시가 대단히 유용하다 . 전 후 및 상하의 방향을 인석하는 기준으로는 den ti cle 이 좋다. 안쪽이나 바깥 쪽의 방향은 pla tf or m type에서 장축의 굴곡에 따라 정해진다 . 기 타 종류에 따라 독특한 표면 구조가 발달하는데 특히 감별에 중요한 것들이 그림 1 에 소개되어 있다 . 내부 구조는 속 이상의 분류분만 아니라 기능 형태의 분석에도 중요하 다. 처음 핵은 하나의 薄 膜으로 중앙에 자리잡고 계속 성장하면서 박막상 으로 겹검이 커진다. 最古 의 형태들의 성장薄膜둘은상부쪽이끊어져 있 으며 (그림 2 참조) Concxlonti form es 亞 目 의 상부 박막둘은 절단되 지 않고 하부 basal p la t e 에는 훨씬 작은 미립자들이 발달되어 있다(그림 3 참조) . 대부분의 코노돈트는 하부가 파여져 있는데 두 가지의 파인 형태는 분 류학에 중요하다. Basa l ca vity는 성장이 계속되는 동안 지속적으로 형성 되는 형태이고 pit는 어느 정도 성장이 지속된 후에는 더 이상 확대되지 않으므로 작은 형 태를 이룬다. 코노돈트의 개체발생사는 薄片 울 제작하여 관찰한다. 초 기 단계는 sim p - le-cone typ e 이며성장이 계속됨에 따라 여러 모양의 den ti cles 이 추가되며 성장 박막이 정해전 방향에 따라 누중된다 . 이러한 부등성장 (an i some t r i c g row t h) 의 결과 성장이 가속되는 방향이 개체 성장기 동안 번화하여 특 수한 형태를 만들어 분류하는 데 기준이 되게 한다. 표면의 미세한 구조 를 관찰하여 분류에 이용하기 위해 SEM (전자 현미 경 ) 이 자주 이용된다. Hin d e (1879) 는 세일의 총리면에서 관찰된 여러 종류의 코노돈트를 관 찰하여 이들이 같은 동물의 부분품이라고 했다. 대체로 정온한 환경에서 퇴적된 코노돈트군은 그들이
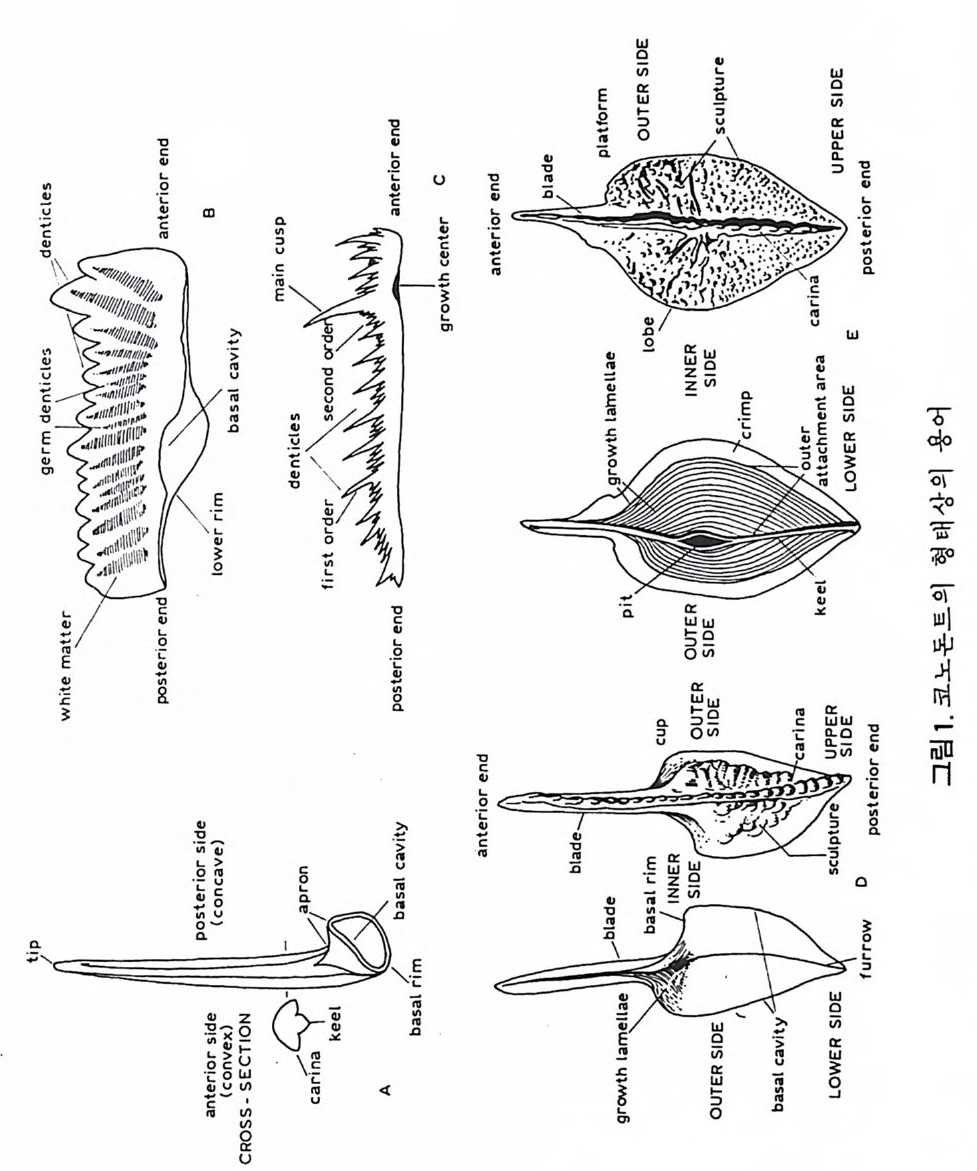 itaro e dnner B yt isabla vac pau cmn,s i tcselden /\ tena irondre c gwroertnc he tj: t r ionderena dlae ttr amo 훑麟멸 ~EURTD ESI 二\\:\u IENNI E.DSI itponders ero 어용 의상
itaro e dnner B yt isabla vac pau cmn,s i tcselden /\ tena irondre c gwroertnc he tj: t r ionderena dlae ttr amo 훑麟멸 ~EURTD ESI 二\\:\u IENNI E.DSI itponders ero 어용 의상
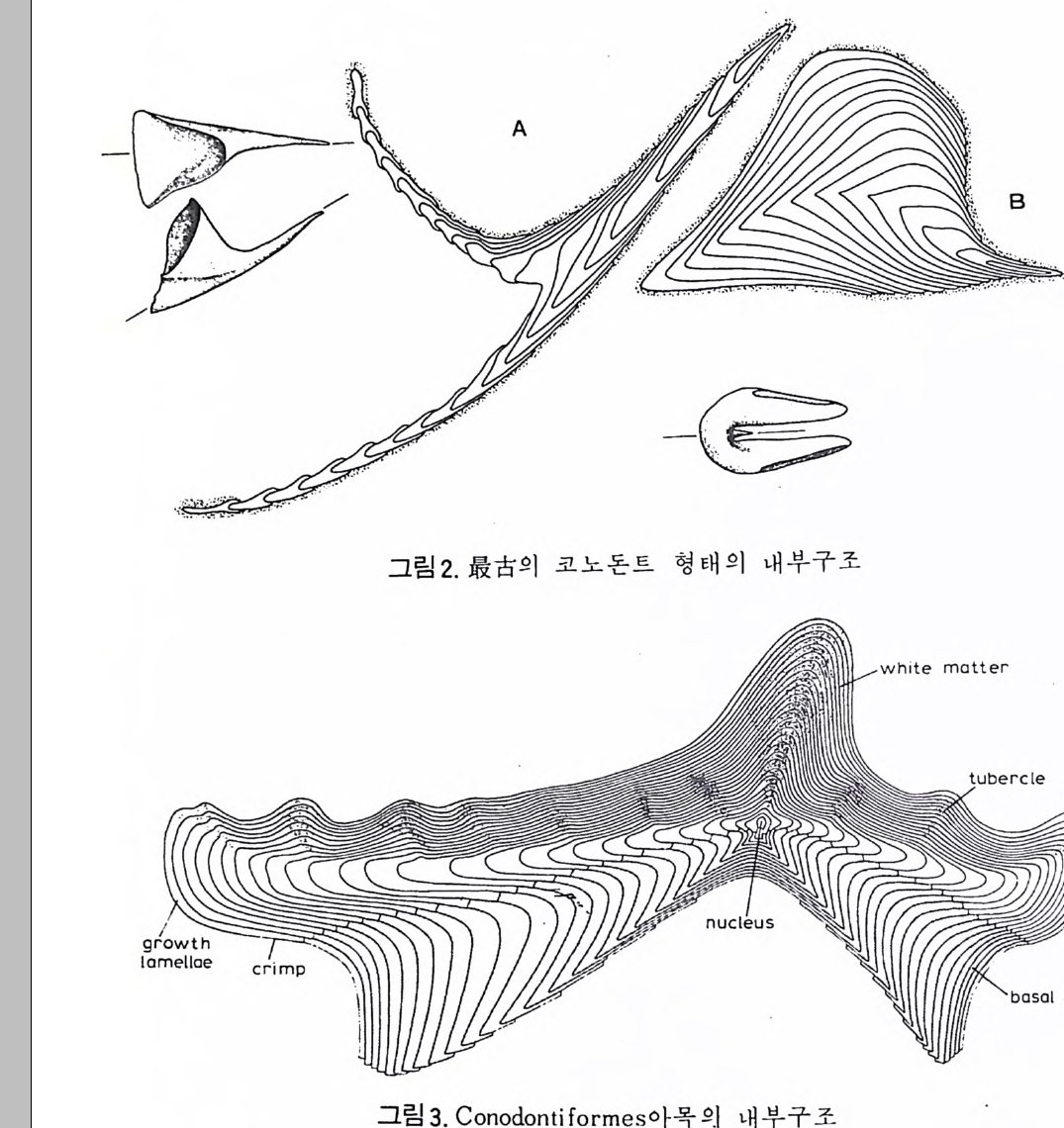 A . • '
A . • '
인 군집은 ®많은 부분이 좌 • 우와 같이 쌍으로 발견되고, @) 한 종류의 몇 쌍들이 다론 한 개 이상 종류의 쌍들과 함께 수반되어 나타나며, ® 여러 군집간의 성분이 다양하며 여러 분류만우]들 사이에 홍 어전 부품들 의 형태가 다양하다(그림 4 참조).
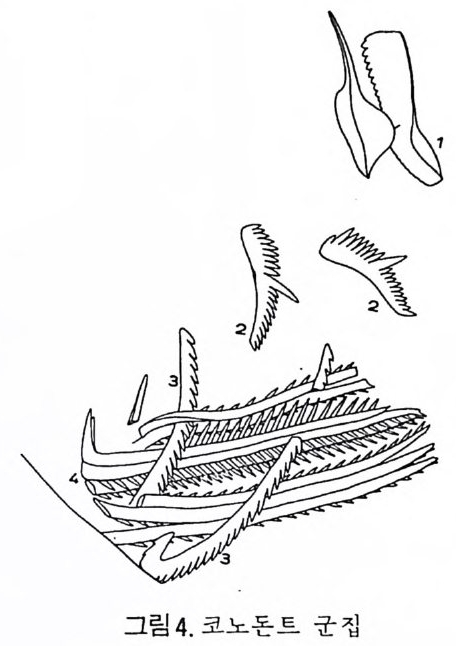 \
\
이러한 군집들은 실제로 회귀하게 발견되지만 완전히 섞인 부품들을가 지고 통계적인 방법을 써서 코노돈트의 각부분을 이론적으로복원시킬 수 도 있다. 한때 코노돈트 (conodon t=희 랍어로 cone t oo t h 의 뜻) 는 그 이름과 같이 치아로 생각된 적도 있었으나 여러가지 증거로서 부정되었다. 각二부분의 형태나 배열로 보아 코노돈트는 지지기관이나 혹은 움켜주 1 거나 걸러내는 작용을 하는 기관일 가능성이 있다. 코노돈트의 조직학적 연구에 의하면 이는 조직을 지탱하는 역할뿐만 아 니라 인산영질 물질을 일시 집적시켰다가 나중에 다론 기관을 만들 때 쓰 거나 혹은 다른 목적을 위해 사용하는 역할을 했던 것으로 보인다. 코노돈트는 광범위하고 다량이 산출됨에도 불구하고 아직 분류학적 위 치를 정하지 못하고 있다. 초기 고생대에 주요 계동 발생학적 발전이 이 루어지고 고도로 분화가 진행되었지만 조직의 특성, 외부의 형태뿐만 아 니라 산출 범위까지도 어떤 화석 및 현생 그룹과도 유사한 접이 없다. 다음은 주요 코노돈트 屬들의 사전음 지질시대의 순서로 배열하였다(그 링 5-11 참조).

그림 5. 캠브리아기 코노돈트 A. Furn- is hin a fur nis h i Muller. X45. B. West e rga ardodin a tr ic u sp ida ta Muller. X75.

그림 6. 오르도비스기 코노돈트 A. Seal- opd us rex, Li n dstr o m. X45. B. Acont iod us sp. X40. C. Parois t - odus pa rallelus Li nd str o m. X40. D. Panderodus sim p le x( Branson and Mehl). X90. E. Cordy lo dus ang u latu s Pander. X60. F. Py go - dus anserin u s Lamont and Li - ndst ro m. X55. G. Amorph og n ath - us sp. X60. H. Belodin a Comp r - essa ( B ranson and Mehl ) . X 60.
그립 7. 산루 리 아 기 코노돈트 A. /crio d in a irr eg u laris Branson and Branson. x 30. (Af ter Rexroad, 1967.) B. Ap s id o g n ath u s tub ercu/atu s Wall ise r. X 66. C. Pte r osp a th o dus amorph og n ath o id e s Wall ise r. X 60. D. Kockelel/ a varia b il is W allis e r. X 23. (Af ter Wallis e r, 1964.) E. Sp a th og n ath o dus ste i n h ornensis Zie g l e r. X 60.
그림 8. 페름기 코노돈트 Gondole/l a rosenhran- tzi . x 70. (Af ter Bender and St o p pe l, 1966.)

그림 9 . 데본기 코노돈 .§. A. Icrio d us sp . X 45. B. Polyg n ath u s asym metr i c u s Bis c hoff and Zi eg l e r. X 30. C. Ancy ro g r.at h us io waensis Young qu is t. x 46. D. Ancy r ode/l a rot un dil o ba (Brya nt) . X 46. E. sPpaa l mth a otdo u lesp ci so shtaa ts u ssi (MBir lalinesro na)n. dx 4M5i .ll ier . x 46. F. Bi -
그림 10. 석탄기의 코노돈트 하부석탄기 : A.Sip }ion ode-
Ila dup li c a ta ( Branson and Mehl). X50. B. P - seudop ol yg n ath us sp. X 65. C. Scaliog n a th us an- choralis B ranson and M ehl. X 60. D. Gnath odus semi gla ber Bis c hoff . X 60. 상부석 탄기 : A. Adeto g n a th us giga nte u s (Gunn- el) . X 55. B. Idio g n ath o dus sp. X 55.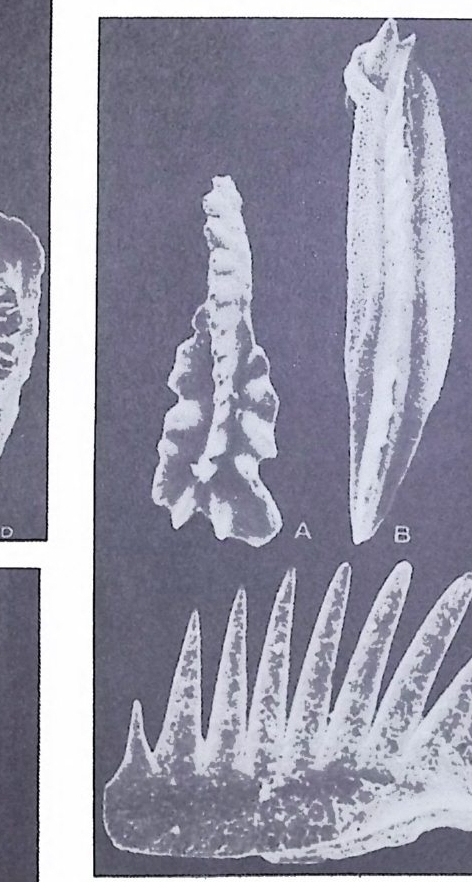
그립 11. 트라이아스기의 코노돈트 A.Ep igo ndole/la ab- nep tis( Huckrie d e) . X 90. B. N 싹 ondole /l a mo- mberg m 퍄 pro vo Kozur. X 90. C. Neosp a th o dus cristo g o // i(H uckrie d e) . X 150.
8 • 3 고생태 코노돈트 동물은 海生이며 유영성이었다고 믿어진다. 많은 屬이나種들 은 원양성이며 따라서 해류를 따라 여러 대륙으로. 확산되어 그 분포가 복 미, 유럽, 호주 둥지까지 이론 것으로 생각된다. 최근의 고생물지리학적 연구에 의하면 이들은 고생태 복원분만 아니라 동물구를 설정하는 데도 유용하다는 사실이 밝혀졌다. 현생 모악동물 (Chae t o gn a t ha) 의 생활방식이 코노돈트의 생태학적 모델로 제안되었다. 모악동물은 조류나 해류를 타고 이동하는 전형적인 부유성 동물로서 수심에 따른 뚜렷한 생활권을 형성한다. 코노돈트도 역시 이러한 수직적인 분포 양상을 가졌으리라 믿지만 실제 로 화석 군집에서 이룰 밝히기는 어렵다. 두 가지의 이론적인 수직 분포 의 모델 이 그림 12 에 제시되었다.
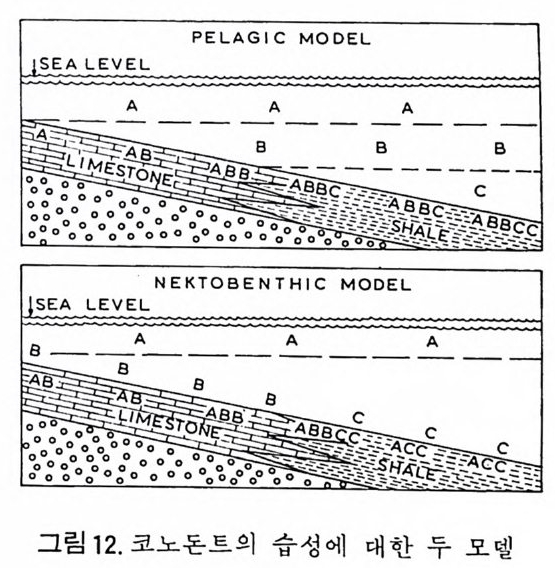 PELAGI C MODEL
PELAGI C MODEL
8 • 4 생물충서 및 진화 코노돈트의 충서학적 가치는 전세계적인 지층 대비가 가능한 접으로 대 번이 된다. 주요 그룹의 충서 범위가 표 8 에 제시되었다. 진화경향은 아직 충분히 알려지지 않고 있다. 초기의 코노돈트는 거의 가 유기질 성분으로 되어 있으며 경우에 따라 영산에 의해 소량의 인산
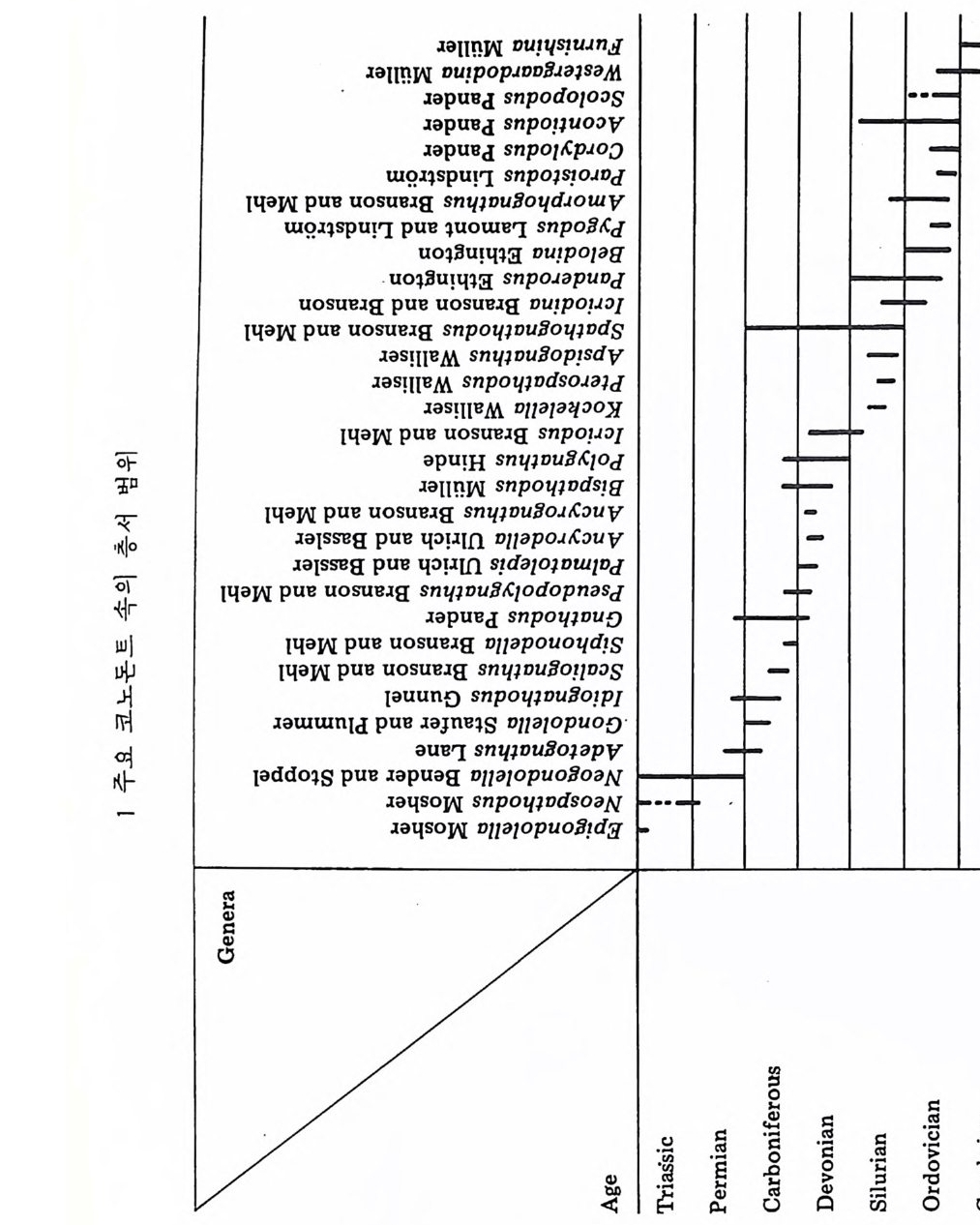 』하1nw 0U! 외 unn g
』하1nw 0U! 외 unn g
영 성분이 용해되어 나가도 그들의 형태는 보존된다. 또한 원시적인 그 룹에서는 코노돈트의 본체와 저부의 기관이 분리되지 않아 대단히 단순한 모양을 가진다. Sin gle cone 상은 가장 원시 적 인 형 태 이며 basal cav ity가 제 한되는 방향으로 번화해 간다. 코노돈트는 특히 계통형태발생 (ph y lo mor~ ph og en esis ) 의 연구에 적합하다. 두족류나 일부 유공충류에서 관찰되는 것 과 감이 개체발생의 초기 단계가 계동발생에 나타나기도 한다. 이러한연 구는 박편을 만들어 시행한다. 보충교재 Hass , W . H., Rhodes , R. H .T., Muller, K. J. and Moore, R. C., 1962. Cono- dont s. In: R. C. Moore (E dit o r) , Treati se on Inverte brate Paleonto l og y, pa - rt W. Geo!. Soc. Am., Ne w York, N. Y., and Kansas Univ . Press, Lawren- ce, Kans as, pp. 3-69 . [A revis e d edit ion of thi s t reati se volume is in pr ep - arat i on .] Li nd s t ram, M., 1964. Conodont s. Elsevi er , Amste r dam, 196 pp. [G enerali nt - roduc ti on .J Lin d st ro m, M. and Zi eg le r, W.(~dit or s), 1972. Sy m p o siu m on Conodont Ta-xonomy . Ge olo g ica et Palaeont ol og ica , Sonderband 1, Marburg, 158 pp. Muller , K. J. and Nog a mi , Y., 1972. Growt h and fun cti on of conodont s. In : Proc. 24th Int. Geol. Cong r. , Mont re al. 1972, Sect. 7, pp. 20-27. [R ev ie- ws his _t o log y.] Pande r, C. H., 1856. Monog r aph ie der (ossil e n Fi sc he des sil u ris c hen Sy s t- ems der russis c h-balti sc hen Gouvernement s. Konig !. Akad. W.s s., St . Pete - rsburg, 91 pp. [T he earlie s t and classic a l work on conodont s. ] Rhodes , F. H.T. (Edit o r} , 1973. Conodont Paleozoolog y. Geol. Soc. Am., Sp e c. Pap. , 141 : 296 pp. Seddon, G. and Sweet, W. C., 1971 . An ecolog ic model fo r conodont s . J. Pa- leont ol ., 45 : 869-88 0 .. [ P aleoecolog ica lm odel of conodont anim al.] Sweet, W. C. and Bergs t ro m, S. M. (Edit o rs), 1971 . Sy m p o siu m on Conodont Bio s tr a ti gr ap h y. Geol. Soc. Am. Mem., 127 : 499 PP. [U p -to -date inf o r mal- ion on bio s t r ati gr ap h y of conodont s by 30 con t r ib u t o rs.) 화석목록 Ash, S. R., 1961 . Bi bl io g r aph y and ind ex of conodont s, 1949-1948. Mi cr op a -leont olog y, 7 : 213 -2 44.
Fay, • R. 0., 1952. Cata l og u e of conodont s. Univ . Kansas Paleonto l . Contr i b . , Verte brata , 3 : 206 pp. Ellis o n Jr. , S. P., 1962. Annota te d bib l io g r ap h y, and ind ex, of ·co nodont s. Univ . Texas, Pub!., Nr. 6210: 128 pp. Ell iso n Jr. , S. P., 1963. Sup ple ment to annota t e d bib l io g r aph y, and ind ex of conodonts . Texas J. Sci. , 15 : 50-67 . Elliso n Jr. , S. P., 1964. Second sup ple ment to annota t e d bib l io g r aph y, and in- dex, of conodonts . Texas J. s·c i. , 16 : 216-2 42. Ell iso n Jr. , S. P, 1967. Thir d sup ple ment to annot a te d bib l io g r ap h y , and ind ex, of conodonts . Texas J. Sci. , 19 : 5 -3 4. Zie g le r, W. (Edit o r) , 1974, 1975. Cata l og u e of Conodonts . Vol. 1, 504 pp. Vol 2, 404 pp. (fur th er volumes fo rth c omi ng ) . [W ell-ill u s tr a te d com p ila t i on of ind ex conodont s. ] 참고문헌 Barne s, C. R. and Nowland , G. S., 1975 . Conodont s: A thi n g of the pa s t and of th e fu tu re. Geosci. Can., 2(2) : 85-89. Bender, H. and St o pp e l, D., 1965. Perm-Conodont en . Geo!. Ja hrb., 82:331- 364. [pl. 14 -16.) B ranson, E. B. and M eh], M . G., 1933 -3 4. Conodont s I udie s , 1-4. Mo. Un- iv. St ud ., 8 : 349 pp. (pl. 1-28 .] Clark, D. L. and Mt ill e r, K. J., 1968. The bas al op e nin g ol conodont s . J. Pa-
leont ol ., 42 : 561-57 0. Fis h er, D. W., 1962. Small conoid a l she l ls of uncerta i n aff in i t ies . In: R. C.Mo- ore (Edit o r) , Treati se on Inverte brate Paleont ol og y, Part W. Ge o! . Soc. A m., New York, N. Y., and Kans as Univ . Press, Lawren ce, Kans a s , pp. 99- 143. G ross, W., 1966. Kle in e Schup pe nkunde. Neues Ja hrb. Geol. Paliio n t ol . A bh., 125 : 29 -4 8. • Hi nd e, G. L., 1879. On conodont s fro m the Chazy and Cin c in n ati gr oup of the Cambro-Sil u ria n , and fro m the Hami lt o n and Genesee-shale div i s i o n s of the Devonia n , in Canada and the Unit e d St a te s . Geo!. Soc. Lond., Q.J., 35(art. 29) : 351-36 9. [pl.1 5 -1 7.] Hy m an, L. H., 1959. The Inverte brate s, 5. McGraw-Hi ll, New York, N. Y.,783 pp. Matt h ews, S. C., 1973. Lapw orth e llid s fro m the Lower Cambria n St re nuella Lim esto n e at Comley, Shrop s hir e . Paleont ol og y, 16 : 139-1 48. [pl.8 -9. ) Muller, K. J., 1964. Ostr a coda (Bradorin a ) mi t ph osph ati sc hen _Ge h ausen aus dem Oberkambriu m von Schweden. Neues Ja hrb. Geol. Paliio n t ol . Abh., 121:1-46. Muller. , K. J. and Clark, D. L., 1967. Early late Devonia n conodont s fro m the Sq u aw Bay Lim esto ne in Mi ch ig a n. J. Paleonto l., 41 : 902-919. [pl. 115-118.] Muller, K. J. and Nog a mi , Y., 197- 1. Uber den Fein b au der Conodonte n . Mem. Fae. Sci., Ky o to Univ . , Ser. Geol. Mi n eral., 38(1) : 1-87 . [pl.1 -22 .] Rexroad, C. B., 1967. St r ati gr aph y and conodont pa leonto l og y of th e Brass£- iel d (Sil u ria n ) in th e Cin c in n ati Arch Area. Geol. Surv. Ind., Bull., 36 : 1- 64. [pl . 1-4] . Schmi dt , H., 1934. Conodont e n-Funde in ursp r tin g l ich em Zusammenhang. Pala- onto !. Z., 16 : 76-85 . Sc hm i d t, H. and MUi ler , K. J., 1964. Weit er e Funde von Conodonte n-Grup p- en aus de m oberen Karbon des Sauerlandes. Palaonto l. z., 38 (3/4) : 105- 135. Sc ott , H. W., 1934. The zoo l o g ica l relati on ship s of the conodonts . J. Paleonto nto l., 8 : 448-45 5. Swe et, W.C. and Bergs tr tlm , S. M ., 1972. Multi el ement tax onomy and Ordov ici a n con odont s. Ge ol og ica et Palaeonto l og ica , Somlerband 1 : 29 -42. Ulric h , E.O. and Bas ~l e r, R.S., 1926. A _cla ssi f ica ti on of th e too t h -lik e fos sil e s, con odon ts , w i th dec rip tion s of Americ a n Devonia n and Mi ssis s ip pian spe cie s . U. S. Natl . Mus. P roc., 6 8 : 1-63 . [pl.l -11. ] Wallis e r , O. H ., 1 964. Conodont e n de s Sil u rs. Abh. Hess. Landesamt . Bodenfo r sch., Wi es baden,41 : 106 pp. [pl.1 -32 .) 9 칫又報毛 藩類 (D i no f la g ella t es) 9 • 1 서언 요즈음 근해에서 적조 현상이 일어나 해양 생태계를 파괴하는 경우를, 종종 보게 된다. 적조는 바로 작은 단세포 식물인 쌍편모 조류의 번성기 에 일어나는 현상으로 그들의 세포에서 독특한 적색 색소가 물에 붉은 빛 울 띠 게 하는 것 이 다. 또한 그들은 pa ralyt ic shellfi sh poiso n” 이 라고 하는 치사적인 독소를 분비하므로 해양 생물에게 대단한 피해를 입힌다. 어떤 종류는 발광성으로 일찍부터 잘 알려져 있다. 현생 쌍편모 조류는 만세포이며 5µm~2mm 의 크기를 갖는 조류의 일 종이다. 그들은 해수나 담수에서 부유성인 경우 먹이 사슬에 중요한 일 익을 담당한다. 소수는 해처의 모래에 살기도 하며 또 어떤 것은 공생이
나 기생하는 경우도 있다. 대부분의 종류들은 유색체를 보유하여 광합성을 하는 독립영양체이나, 어떤 것은 동물과 같이 음식을 소화시키는 종속 영양체인 것도 있다. 쌍편모 조류는 영양 단계와 포낭 단계를 포함하는 비교적 단순한 생활 샤를 갖는다. 포낭체는 포분질 (s po ro po lle ni n) 과 같은 튼튼한 유기질 물질 로 형성되어 있기 때문에 화석화될 수 있다. 화석으로 발견되는 쌍편모 조류는 모두 포분질 포낭이며 이는 육상 식 물에서는 호 H 손과 포자를 둘러싸는 포분질에 해당된다. 화석군집은 페름기에서 현세에 이르는 해성층에서 계속 발견되고 있다. 화석상에서는 항상 acrit ar chs, ta sm anitids , 호남는 및 포자와 함께 산출되는 데 호 H 손, 포자를 매고는 모두 포낭의 외벽이다. 이들 미화석은 퇴적물로 부터 영산이나 불산을 이용해서 광물 성분을 제거한 뒤 골라낼 수 있으며 주로 호 H 손학자(p al yno lo gi s t) 의 연구 대상이다. 이들 화석들은 모두 pa lyn - omo rp hs” 라고 불리며 특히 dino fl age llate s, a~rit ar chs 그리 고 tas ma nitids 를 acid - in s oluble or orga nic- walled mi cr opl a nk ton ” 이라고 칭한다. 표 9 는 dino fl age llate s (Py rr hoph y ta) 의 분류상 의 위 치 를 표시 한 것 으 로 이 중 Di no ph y s ia l es, Gy mnodinial es 그리고 Perid inial es 동 세 目 이 화석 기록에 남아 있다.
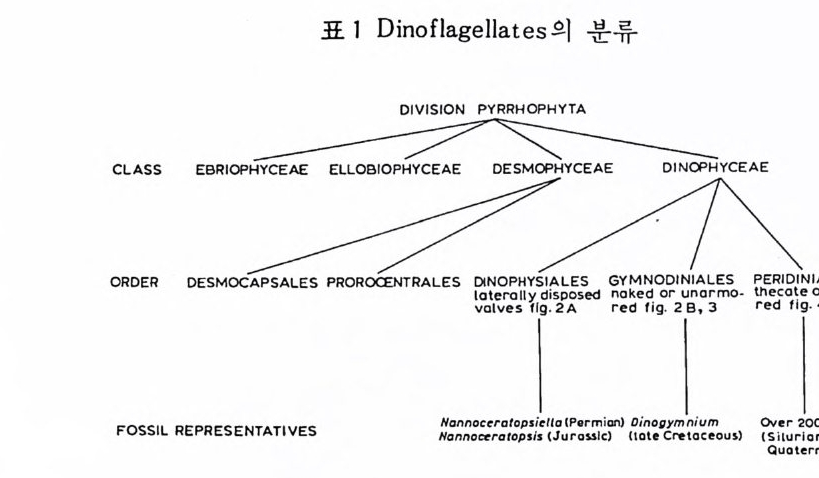 표 1 D i no fl a g ella t es 의 분류
표 1 D i no fl a g ella t es 의 분류
9 ·2 형태 및 분류 9 • 2 • 1 현생종 쌍편모 조류는 단세포 식물로서 원형질체의 바깥 쪽에는 유색체가, 안쪽 에는 핵과 공포가 자리잡고 있다(그림 1 참조).
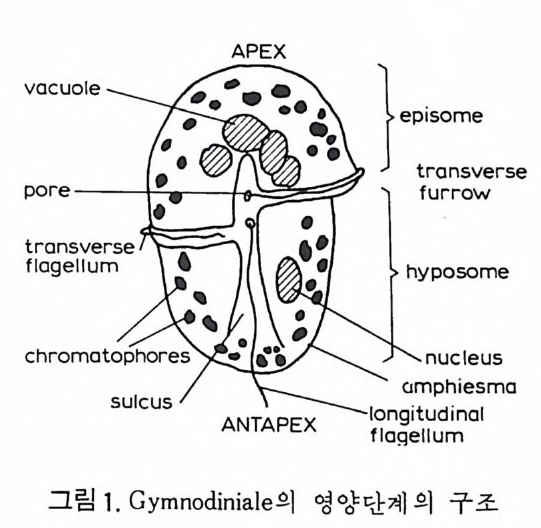 APEX
APEX
유색체는 황색이나 갈색이며 개체에 따라 독특한 색조가보이기도한다. 먹이는 전분 혹은 밝은 색을 띠는 지방질로서 저장된다. 큰 공포인 p usules 는 외부와 섬세한 관으로 연결되어 있어 배설기관의 역할을 하는 것으로 보 인다. 원형질체는 두개의 편모를 가지며 세포벽의 편모공으로부터 뻗어 있 다,편모공이 있는 부분에서, 핵을 제외한 원형질인 세포질은 다소 접성이 적고 자주 위족을 만든다. D i no p hy ceae 는 세포벽의 형태로 구별이 되는데 운동 단계에서 이들은외 각에 am ph i esma 라고 하는 세 포 보호막을 가지 며 또한 길 이 가 다론 두 개 의 편모를 발달시킨다. 어떤 종류에서는 보호막이 일련의 판들로 이루어져 있 어 t heca 라고 불린다. 두 개의 편모는 서로 직각으로 sulcus 나 fu rrow를 따라 발달하는데 장축 편모는 sulcus 룹, 그리 고 교축 편모는 c ingu lum 이 라 고 하는 g roove 를 따라 세포를 한 바퀴 둘러싼다(그림 1 참조).
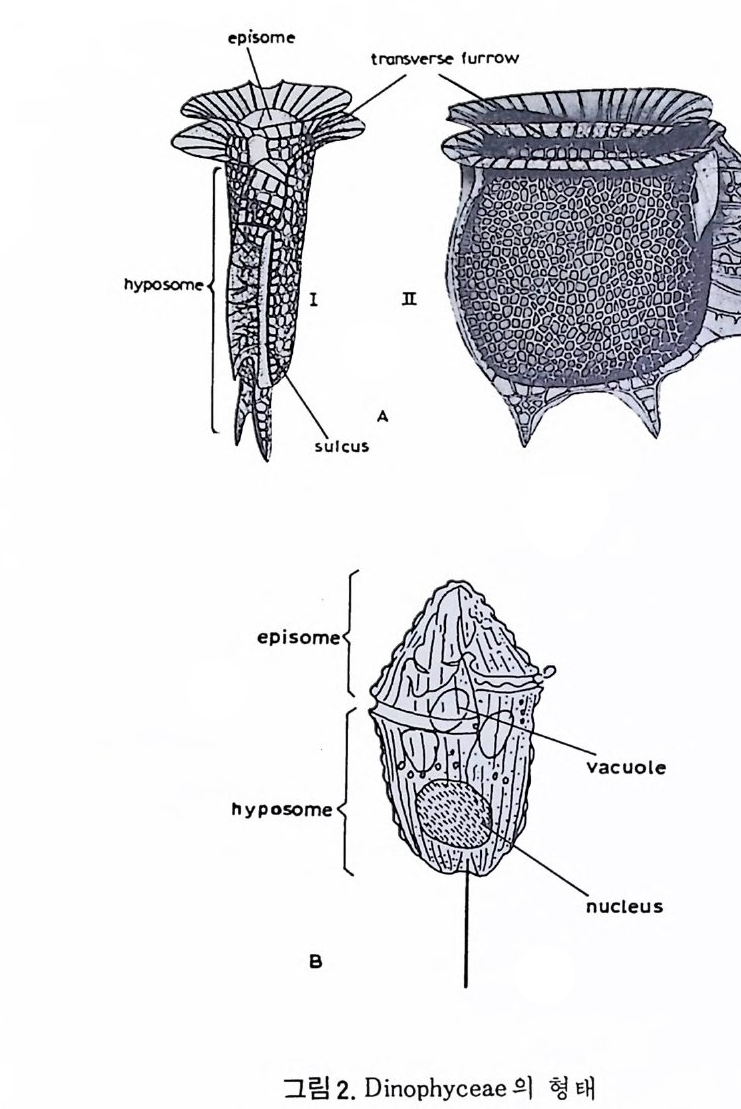 epis o me tra nsverse fu rrow
epis o me tra nsverse fu rrow
C ingul um 은 세포를 전부인 e pi some 과 후부인 h yp osome 으로 구획 짓는데 sulcus 는 대체로 후부에 제한 분포한다. Theca 는 횡적으로 유교판의 연결로 이루어져 있으며 판들의 수나 위치 는 분류에 대 단히 중요한 제 표 (tab ulati on ) 로 표시 된다. Kofo id (1907, 1909) 는 판의 명칭 체계를 만들었으며 현재 이 방법이 널리 쓰이고 있다. 그림 3 에서 보는 바와 같이 t heca 의 열과 번호는 Ko foi d 의 알파벳 체계 에 따
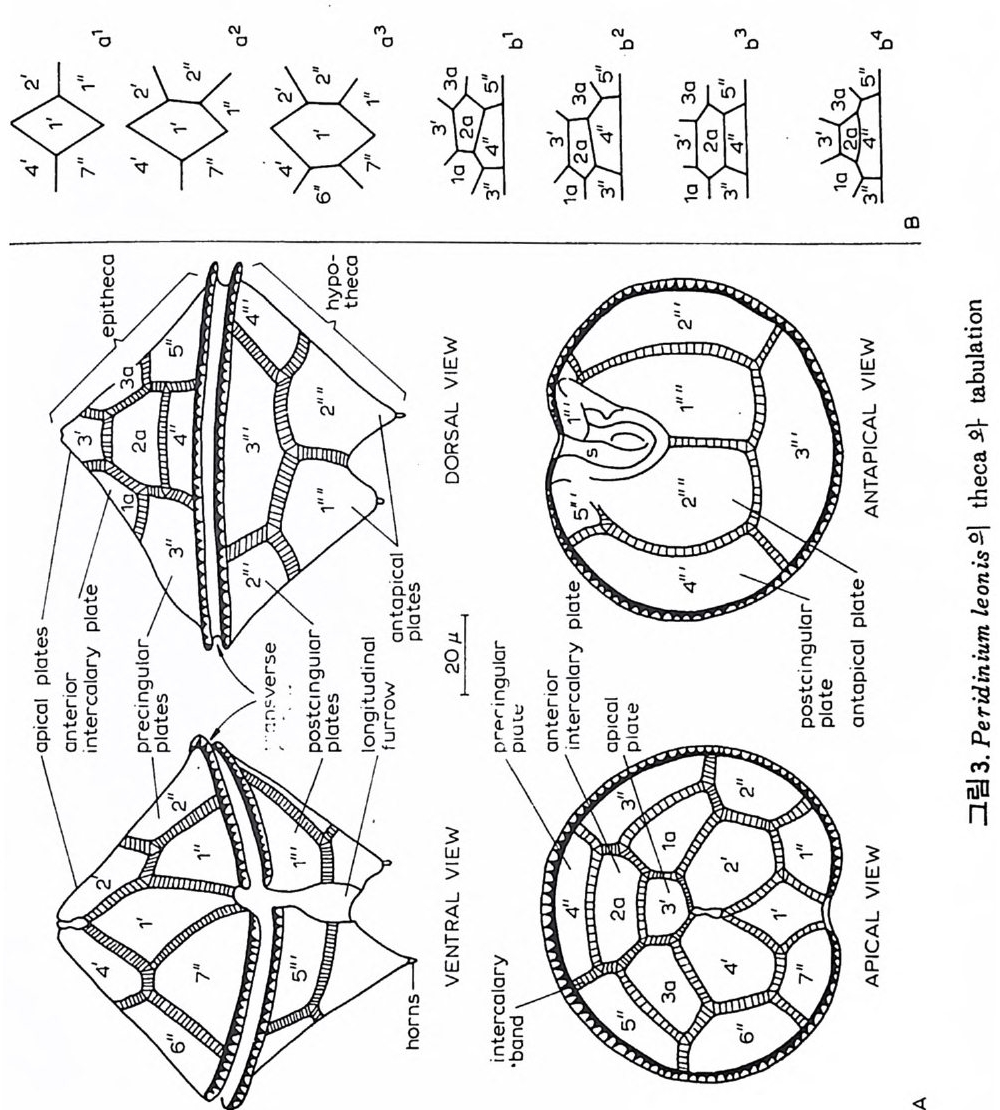 al a230 , bb 3
al a230 , bb 3
라 명명된다. 판의 표면부는 평활한 것부터 미소 구조까지 다양하게 관찰된다. 판들의 경계는 타일 모양으로 시멘트되어 있으며 성장은 판의 경계부인 su t ure 에 서 i n t ercalar y bands 의 형태로 일어난다. 9·2·2 화석종 모든 화석들은 포낭으로 보존 발견된다. 그들은 중생대 및 신생대뿐만 아니라 약 40~ 년이 지난 사일루리아 암석에서도 추출된다. 중생대 후기와 산생대에는 담수 화석종도 발견되지만 대부분은 해수종으로서 접토 , 세일, 실트암 혹은 니앙에서 주로 발견된다. 대부분의 di nocy s t s 는 25~250 µm 의 직 경 을 가지 며 구형, 타원체 형 및 신장된 형 태 둥 다양한 모습을 갖는다. 많은 종류가 쌍편모 조류의 득칭 적 인 구조인 tab ulati on , cin g u lu m 혹은 sulcus 둥을 보유하기 때문에 접두 어 인 p ara 를 붙여 서 pa rata b ulati on , pa racin g u lu m, pa rasulcus 그리 고 pa ra- pla te 등의 용어를 사용할 수 있다. 어 떤 di noc y s t s 는 spi ne s 이나 둘기 (pr o- cesses) 와 같은 구조를 가지며 50µm 이내의 크기를 갖는 horns 와 함께 di noc y s t s 의 방향을 정하는데 유용하다. D i noc y s t s 는 운동 단계 의 형 태 와 아주 유사하든지 혹은 완전히 다를 수 도 있다. 돌기의 유무에 따라서 두 개의 주요 형태인p rox i ma t e 와 chora t e-
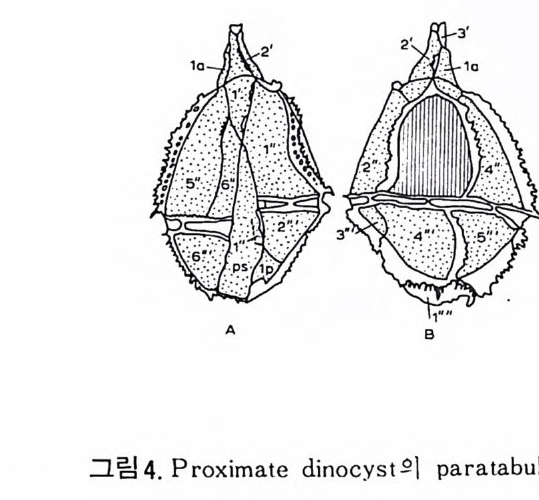 A
A
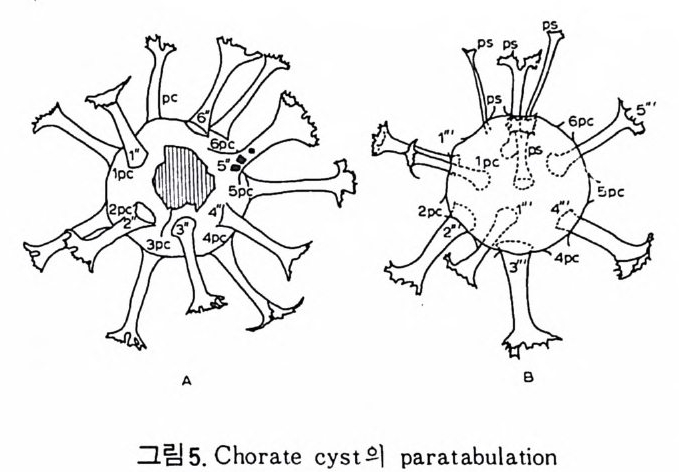 A B
A B
c y s t s 로 구분할 수 있다. Proxi m ate c y s t s 는 돌기가 없고 c y s t의 모습이 거 의 t heca 의 것과 일치한다. 이것은 아마도 t heca 와 접촉하면서 생성된 것으쵸 L 보인다(그림 4 참조). 반면에 chora t ec y s t는 강력한 장식품인 들기를 가진다(그림 5 참조). 세번째 형태는 cavate cy s t로서 내벽과 외벽은 부분적으로 접합되어 있 으며 두 벽 사이에는 공간이 자리잡고 있다(그림 6 참조). 이것은 형태학적 으로 유용한 형태일 뿐 유전적인 유연관계를 암시하는 것으로받아들여져 서는 안된다. D i noc y s t의 외벽은 한 개 혹은 그 이상의 박충으로 한 개의 혹은 겹으
 그림 6. Cavate cys t 의 형 태
그림 6. Cavate cys t 의 형 태
로 포낭을 이루며 이들 외벽 사이는 공간으로 서로 격리되어 있다. 이들 세포벽, 포낭 그리고 공간의 입체적인 형태는 적당한 접두사와 접미사롤 환용해서 표 2 와 같이 표시하며 그림 7 은 이들의 도해이다. 표 2 D i noc y s t s 의 세포벽 용어 Meanin g Combin a ti on Defi ni t ion Prefi x aut o p h ragm sin g le wall auto - sin g le auto cy s t sin g le body auto c oel sin g le cavit y endop h ragm inn er wall endo- mner endocy s t inn er body endocoel inn er cavit y mesop h ragm mi dd le wall meso-mi dd le mesocy s t mi dd le body mesocoel mi dd le cavit y pe rip h ragm oute r wall pe ri- oute r pe ric y s t oute r body pe ric o el oute r cavit y Suff ix auto p h ragm sin g le wall -ph ragm wall emnedsoopp hh rraaggm m imnni ded rl ew awlal ll pe rip h rag m oute r wall auto cy s t sin g le body -cy s t body emnedsooccyy s s t t minin ded rl eb obdoyd y pe ric y s t oute r body auto c oel sin g le cavit y -coel cav ity menedsooccooeell minin ded rl eca cvaivt yit y pe ric o el oute r cavit y
 outoc y s t endocy st
outoc y s t endocy st
di n oc y s t 의 h o rn 은 api ca l, ant ap i ca l 혹은 lat e ral 위치에만 있으므로 di no c y s t 의 방위 를 정 하 는 데 편 리하고 최대수는 다섯 개 이다 . 돌기는 기 둥 상 이나 spi ne 의 형태이며 p ara p la t e 의 경계부에 위치한다. 이들 둘기는 여러가지 모양으로 배열하여 일련의 pr ocess com p lex 를 이룬다. 둘기의 배 열, 형 태 그리고 위치 는 種이나 屬 울 분류하는데 중요한 형태 요소이다 (그림 8 참 조). Ev itt (l~l) 는 많은 di noc y s t s 의 표면에 규칙 적 인 모습의 구멍 이 있음을 발견하고 archeo py le 이라 명명했다 . 이 구멍은 한 개 혹은 그 이상의 pa ra- p la t e 가 빠져나와 생간 것 인데 어떤 경우에는 p ara p la t es 의 경계부인 sut- ure” 가 함몰된 것도 있다. 여러가지 형태의 archeo py le 은 유전적으로 결정 된 종의 특수한 형질이다(그림 9 참조). 표 1 에서와 같이 모든 알려진 화석 di noc y s t s 는 Di no p hy c eae 강에 속한다. 두 개의 속들이 D i no p h y s i ales 와 Gy mnodini a l es 目에 포함되고 200 屬이 넘는 대 다수는 Penid ini a l es 目 에 속한다. 屬과 種에 대한 분류 체계는 독자적인 두 가지가 현 존한다. 한 가지는 화석의 체계로 호 H 손학자들에 의해 제안된 것이고 다른 하나는 현생 영양
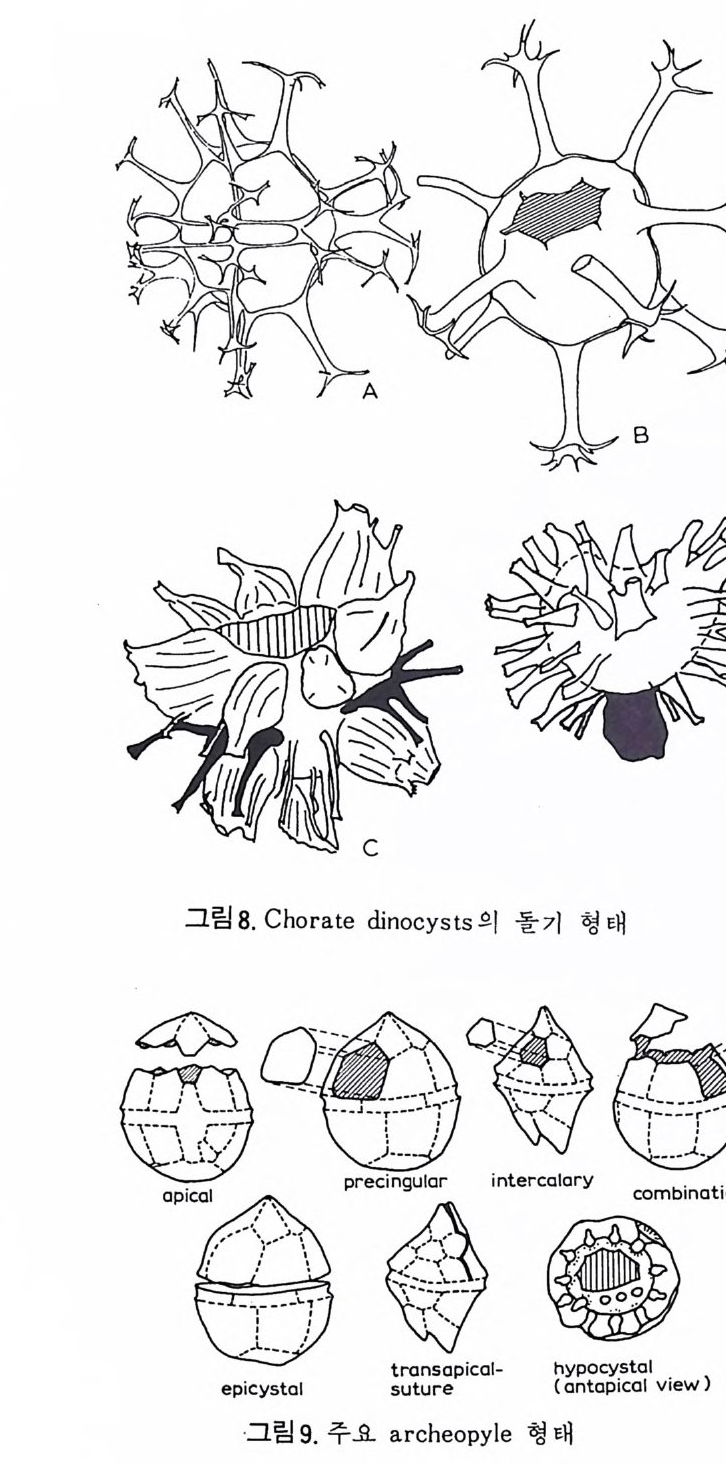 D
D
단계의 표본에 의한 체계로 생물학자 들에 의해 개발되었다. 모든 생황사 의 단계를 포괄하는 하나의 통합된 분류 체계가 완성되기에는 아직 요원 한 감이 있다. 9 • 3 생태 현생종의 생태에 관한 자료는 매우 미약하여 안지 그들의 분포 양상의 윤곽이 알려져 있으며 그것도 대부분이 바다에 사는 유동 단계의 종들에 국한된다. 유동 단계에서 쌍편모 조류는 두광대에서 광합성을 하는 독립 영 양 체 이 다. 수온의 적 응도에 따라 이들은 광온성 (euryt he rmal ) 인 종류와 협 온 성 (s t L'llo t hermal) 인 종류로 구분되는데 후자는 주로 온수에 제한되어 산다. Cera ti um 屬은 해수의 인산영 함량과 해류에 의해 그 분포가 영향을 받 으며 개체수의 번화는 계절을 탄다. 보통 늦은 봄이나 여름에 여러 배로 중석되는데 이 시기는 규조의 초]번성 시기의 직후이며 영양영류의 양이 최대가 되는때와 일치한다. 이때 개체수는 o.,l ' 1 리터의 해수에 10~ 마리 정도에 이르며 보통 해수가 적색이나 황색으로 보인다. 대부분의 Cera ti um 屬의 種둘은 협영성 (ste n ohalin e ) 이나 G y mnod i n i um 과 Per iai n i uml 썩은 담수에서 해수까지 광범 위하게 분포한다. 그러 나 이 두 屬 둘의 종들은 경우에 따라 특정한 환경에 제한된다. 현재 바다의 석물성 풍랑크론 중에서 쌍편모 조류는 규조 다음으로 풍 부한 그룹이다. 이들은 먹이 사슬에 일차적인 중요한 단계에 속하며 유공 충과 같은 원생동물에서 고래와 같은동물에 이르기까지 먹이의 공급원 이 된다. 쌍편모 조류의 생한사에는 세 가지 뚜럿한 상이 있다. 가장 혼하게 관 찰되는 것은 황동 단계 혹은 영양 단계의 단세포이다. 이는 분열에 의해 서 무성 생 식 이 된다, 두번째 상은 유성 생 식 으로서 Cera ti um 의 두 종에 서 관찰된 바, 두 개의 크기가 서로 다른 배우자(gam e t es) 가 결합하여 수명 이 짧은 접 합포낭 (zy gotic cys t ) 을 형 성 한다. D i noc y s t라고 불리 는 휴지 포낭 (res ti n g c y s t)은 세번째 상으로서 영양 단계에서 셀룰로즈질인 세포 벽에서 형성된다. 이러한 휴지 포낭은 화석에서 약 40 종 이상 기재되었다. 포낭울 생산하는 주요 그룹은 Gon y aulax 와 Per i d i n i um 屬으로 이들은 화석 상에도 혼히 발견된다. 피낭 (enc y s t men t)은 보통 최번성기에 일어나며 amp hies ma 안에서 형성된 포낭은 조만간 비활동 포낭울 떠나 바닥으로 가 라앉는다 (그 림 10 참조) .
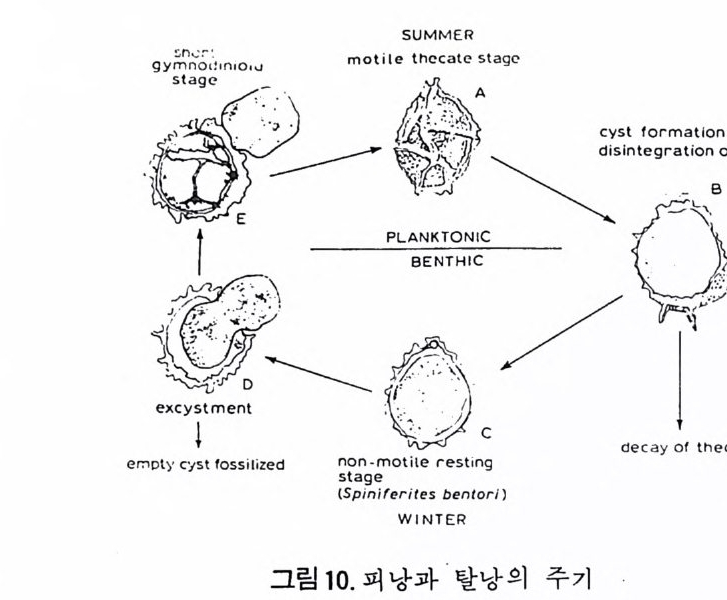 :gy s;o; t;n a~. ;ogc ,e·· .: , n , o,J - --. -----m-o-t i le S Ut:h M cMcaE t Rc st a g e 二 삶'&言,;껏:,a ::ec a
:gy s;o; t;n a~. ;ogc ,e·· .: , n , o,J - --. -----m-o-t i le S Ut:h M cMcaE t Rc st a g e 二 삶'&言,;껏:,a ::ec a
많은 포낭둘은 원형질을 겨울 동안 보호하여 생존케 하며 동면 기간은 다양하다. 포낭의 발아에 적당한 조건은 수온에 의하기 때문에 주로 40m 이하의 수심을 갖는 대륙붕 지역이 적당하다 . 그러나 400m 정도에서 발견되는 종 속영양체인 쌍편모 조류들 중에도 피낭울 할 수 있는 능력이 있는 것들이 있다. 대다수의 포낭들은 연안대 안쪽에 집중되어 사는 종들에 의해 생산 된 것이다. 탈낭 (exc y s t men t)은 archeo py le 을 통해 원형질이 유출됨으로써 이루어전 다. 그림 10 에서와 같이 포낭으로복-터 해방이 된 자유 유영하는 초 기 단계 는 매우 섬세한 세포막과 장축 편모를 보유한다. 이러한 unif lage llate sta - g e 로부터 교축 편모가 생 기 고 즉시 활동을 하는 gymnodini o i d s tag e 로 이 행되며 계속해서 유동 단계가 오는 것으로 추측된다. 9 ·4 생지리 및 고생태 Cera ti um 屬의 분포는 알려 져 있으나 이 속에 속하는 해 양종들은 포낭울 만들지 않는다.반연에 포낭성이고 찰 알려져 있는 약 40 종의· 생태는 ' 별로 연구 된 것이 없다. 현생 di noc y s t s 의 분포에 관한 연구 (W illiam s, 1971) 가 북대 서 양의 현세 되적물을 대상으로. 시행되어 대부분의 종들은 범세계적인분포
를 하지만 상대적인 함량(개체수)은 기후에 의해 제한되어 번화가 있음을 알아내었다(그림 11 참조). 제 4 기 퇴적층의 연구에 의하면 dino cy s t 군집은 고기후와 고환경에 관 한 상당히 정확한 정보몰 준다는 걷론을 얻었다. Dale (1968) 은 영국의 한 시추코아로부터 다섯 개의 군집을 확인하고 제 4 기 빙기와 간빙기의 교차 에 따론 해류의 순환에 관한 역사를 추적했다.
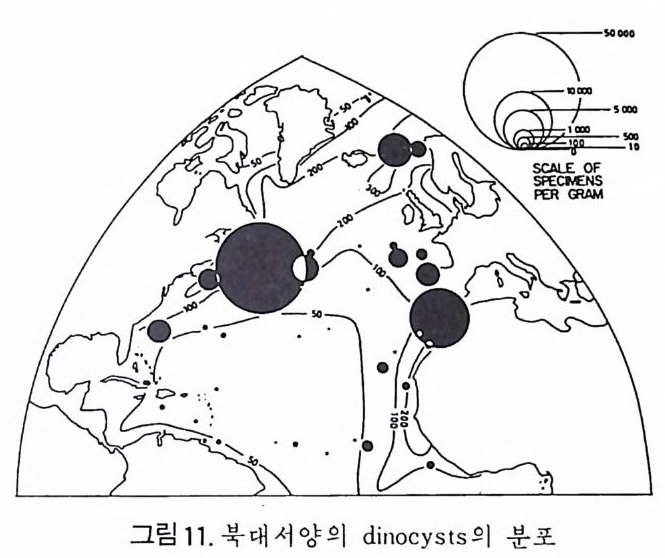 ’<
’<
Downie 의(1 971) 는 남부 영국의 에오世 초기 지층을 연구하여 세 개의 군 집 이 각각 원양환경 (go ny au lacacean). 연 oJ : 환경 (acrit ac h) 그리고 석호 또 는 기수 환경 (p er idi n i acean) 을 지시한다고 결론지 었다. 이러한 군집의 수 직적 번화는 이 지역의 해침과 해되의 역사를 추적할 수 있었다(그림 12 참조). 중생대 d i noc y s t s 의 형태적인 특성으로 원양과 연안 환경의 차이를추적 하며 호 H 손·포자를 포함한 p al yn omor phs 의 상대량의 변화를 해안에서부터 의 거리를 복원하는 고지리적 정보로 이용할 수도 있다. 또한 새로운 종류 와 오래된 것등로 오영된 호格수군집에서 원지성 인 종류와 이동된 종류를구 분할 수 있다면 이들의 상대적인 함량을 지역에 따라 비교함으로써 추정 된 해안선으로부터의 거리를 앙아낼 수도 있다.
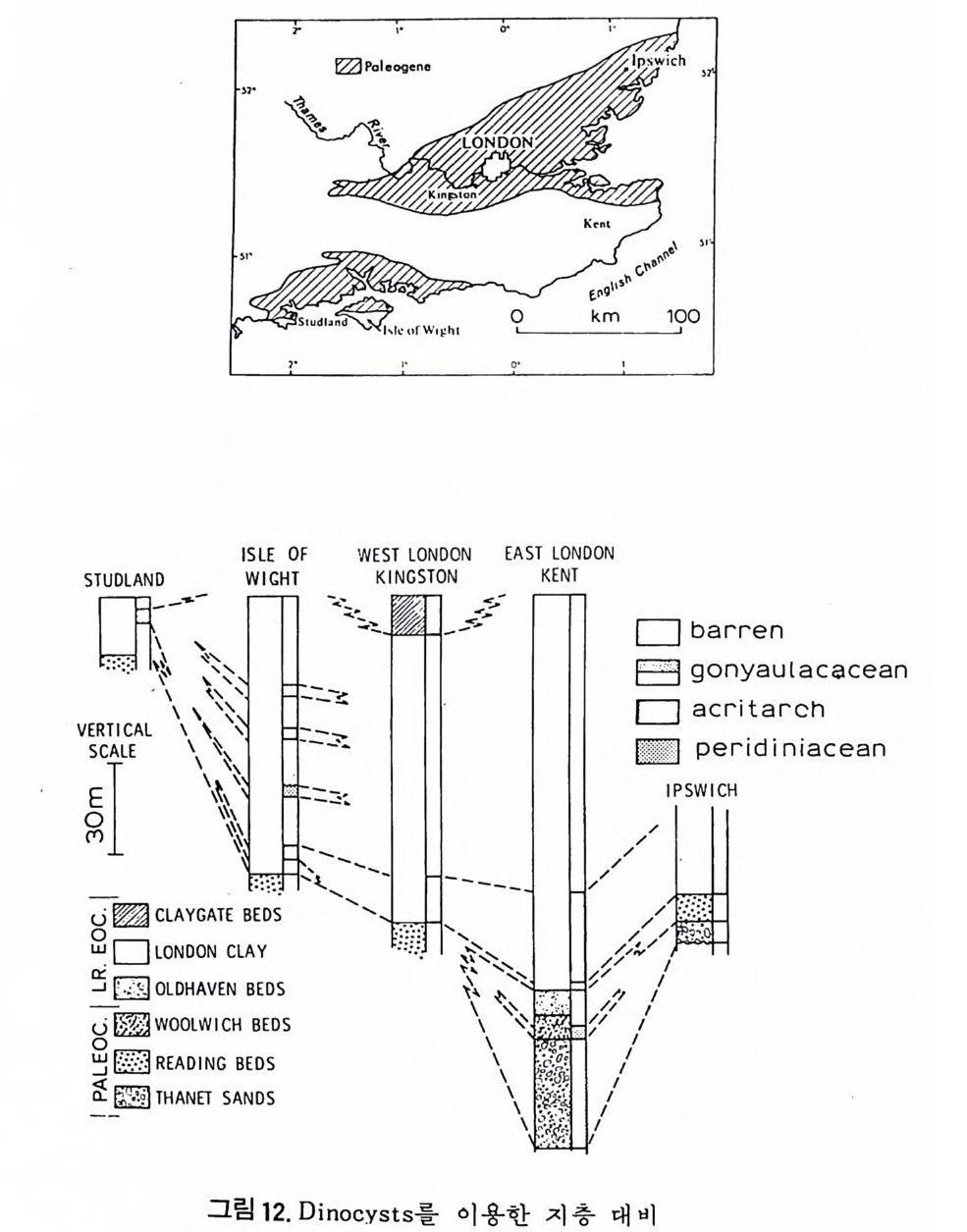 ...\ '’?.
...\ '’?.
반면에 과거의 견해와는 달리 담수성 di noc y s t s 둘이 후기 백악기에도 생 존했었다는 사실들이 밝혀지면서 di noc y s t s 의 존재가 반드시 해성층을 지 시한다고 봉 수 없게 되였다. 9 ·5 생물충서 및 진화 이 철에서는 편의상 화석 dino cy s t s¾ p ara t abula i on 과 archeo py le 의 형성 울 근거로 해서 일곱 개의 계통으로 나누어 설명키로 한다. 가장 중요한 것들은 대부분의 pro xim at e or chorat e cy s t s 를 가지는 Gony a ula cyst a , cava- te cy st s 가 대 부분 인 Perid inioi d 그리 고 Cerati oid 및 C)t :lo ne ph eli um 둥 이 있 다. 이들의 충서 법위는 표 3 과 같다.
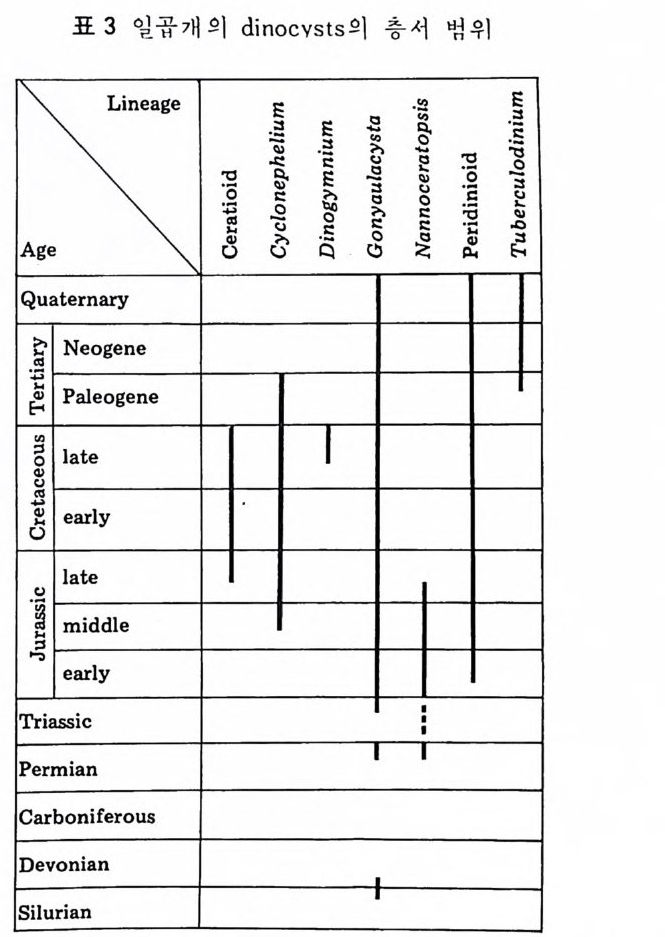 표 3 일곱개 의 d i nocvs t s 의 충서 범위
표 3 일곱개 의 d i nocvs t s 의 충서 범위
Wi ]]iam s (1977) 는 중생 대 와 신생 대 의 시 준화석 그룹으로 30 개 의 concur-rent ran ge zones 와 12 개의 subzones 를 설정 했다. 이 에 따르면 각 zone 괴 subzone 은 중생대와 초기 제 3 기에는 범세계적인 분포를 보이는 화석군으 로, 말기 제 3 기는 국부적인 지역에 분포하는 화석군으로 이루어진다. 제 4 기에는 국소수의 di noc y s t s 가 존재하므로 충서 분대는 이들의 상대 적 함량에 의해 정해진다. 현재는 약 40 여 종이 피낭 단계를 갖는다는 사살이 알려 져 있다. Wall (1971 은 휴지낭을 갖主· 현생종을 네 개의 그룹으로 나누었는데 이들은 Gony au lru< Peridi nium , Ceratiu .n, 그 리 고 P y ro ph acus 그룹 이 다. 배양키 안에서 조류는 네 가지 단계의 군체 크기의 번화가 관찰된다. 즉 초기의 정체기. 두번째의 세포 분열기. 세번째의 안정기 그리고 마지막의 쇠퇴기 둥을 거치는 동안 군집은 한 번의 크기상의 순환을 이룬다(그림 13 참조). Zeuner(1916) 는 모돈 생물은 전화하는 동안 이와 유사한 번화를 겪 는다고 생각했다. 상대적으로 활발치 못한 분화현상 뒤에는 폭발적인 분 화가 뒤따르며 철정기 혹은 안정가물 지나 쇠퇴기로 변화하며 한 윤회는 5 천만년이 소요된다고 했다. Tapp a n & Loeblic h (1970, 1972) 는 ,Z e uner 윤 회관을 지질시대룽 지내는 식물성 플랑크톤에 적용시켜서 특히 · 우성생식 울 하는 경우에 주목하였다. 또한 폭발상은 작은 dino cy s ts , 쇠퇴 상에는 큰 di noc y s t s 로 대표된다고 생각하였다. 초기 정체상은 사일루리아-초기 쥬라 기에 해당되며 폭발상은 초기 밋 중기 쥬라기에 철정을 이루고 쇠퇴기는 후기 쥬라기-초기 백악기에 맞는다고 주장했다. 두번째 순환은독자적인그룹돌에 의해서 초기 백악기에 폭발기를 맞아 절정에 이르기까지 25m~y . 동안 지속되었으며 후기 백악기에 쇠퇴기를 맞 논다. 세번째 폭발형 후기 팔레오世에서 초기 에오世까지 지속되고 쇠퇴 형은 현세까지 계속되고 있다는 것이다(그림 14 참조). 한 순환의 쇠되상은다음 순환의 폭발상으로 연속되며 이러한 폭발적인 상들은 연해의 최대 확장기와 일치한다. 다론 주요환경 요소의 변화도역 시 폭발의 원인으로 작용할 수 있다. 화석 자료에 의하면 새로운 di noc y s t속은 매 1. 4m. y.마다 출현하며 평균 屬의 수명은 42.4m. y.로 계산된다. 현생속들을 포함해서 고생대 이후의 한 屬의 전화 속도는 〈매 460,000 년에 한 속〉으로 산출되며 이러한 급속한 전화는 이들이 이상적인 생물충서 분대의 도구임을 암시한다. Ea t on(1971) 은 남동 영국의 에오世 지층으로부터 몇 가지 di noc y s t s 의 계
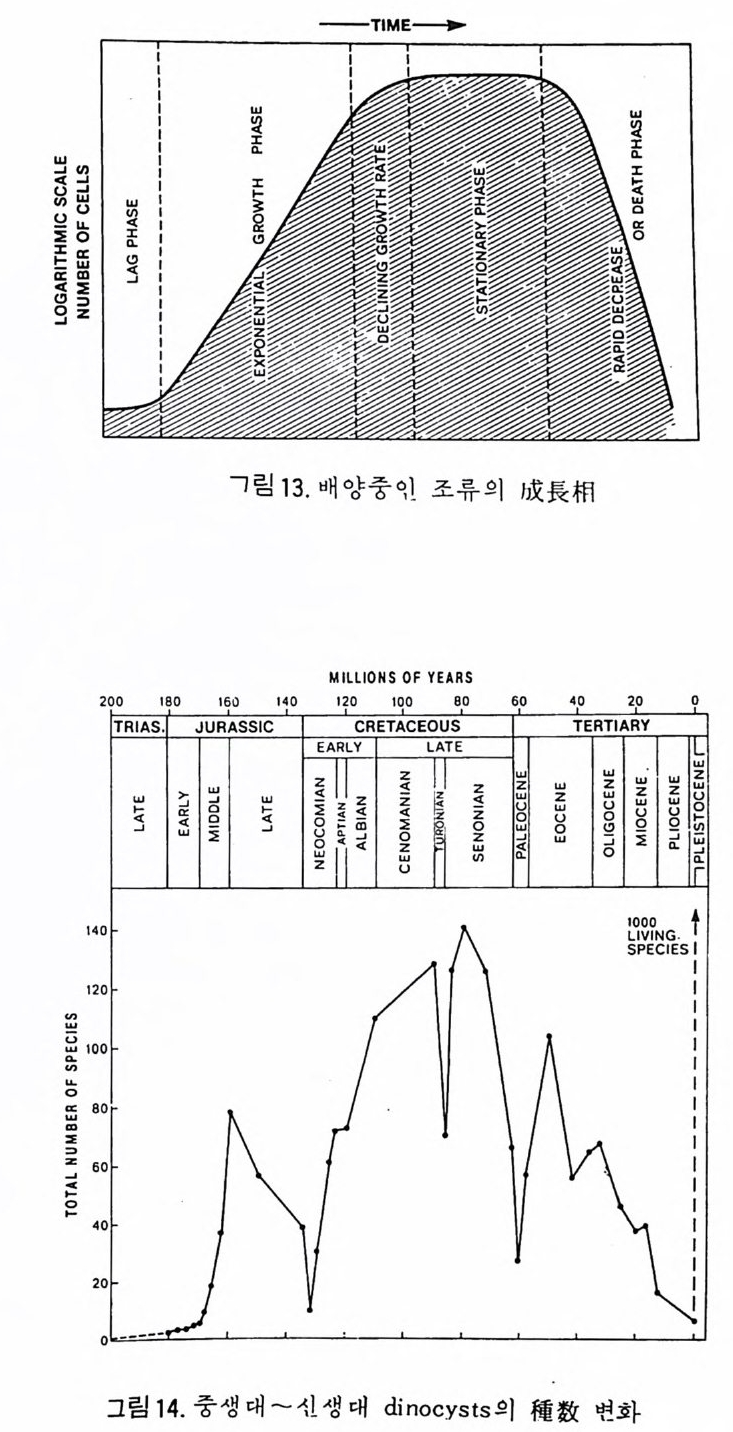 TIM
TIM
동전화를 설정할 수 있었다. 이러한 계통을 따른 형태의 번화는생물 충서 학적 가치를 지닌다. Archeop yle , su t ure 둥의 계통적 변화가 이에 속한다. 보충교재 Downie , C, and Sari e ant, W. A.S., 1966. The morph olog y, te rmi nolog y and clas s i f - ica ti on of fos sil dino fl ag e llate cy s ts . In : R.J. Davey , C. Downie , W. A . S . Sa- crjy e sa tns t. Ba unldl . GB .rL. .M W~si . llN iaa m t. sH, iS st t .u d (Gie es o /o.)n , SMueps pol.z o3i c : HaJn-d 1 C7a. i n[ Mo zoodice m dinc oo n(lca egpe tlsl a toe f dino cy s t morph olog y wi th pa rti cu lar reg a rd to wall lay e rs and th e in fo r mal gr oup s in clud ing chorale, pr qx i m ate and ca vate cy s ts .] Ev itt, W.R., 1967. Di no f lag e llate stu d ie s IL The archeop yle . St an f or d Uni v. Pub/., Geo!. Sci. , 10(3) : 1-83. (The sta ndard refe r ence on arch eop y le s in dino cy s - ts. ] Ev itt, W.R ., 1969. Di no f lag e llate s and oth er orga nis m s in pa ly n olog ica l pr ep a ra- tion s. In : R.H . Tschudy and R.A. Scott (E d itor s), Asp e ct s of Palyn olog y Wi - ley- Inte rscie n ce. New York, N. Y., pp. 439-479. [A b rie f disc ussio n of th e mo-
rph olog y of modem din o fl ag e llate s and the ve g e ta ti ve sta g e is fol lowed by a relati ve ly deta i l e d secti on on the morph olog y· of fos sil dino cy s ts . ) Frit sc h, F.E., 1965. Class VI . Di no p h y c eae (Perid ini e a e) . In : F.E. Frit sc h, St r- uctu r e and Rep r oducti on of th e Alga e, Cambrid g e Univ ersit y Press , London. pp. 664 구 20. [D i sc usses th e classif ica ti on , morph olog y and rep r oducti on of modem dino p h y c eae. Presents a deta il e d. but slig h tl y outd a te d, account ; from the bio l og ica l view p o in t. ] Li s te r , T.R. , 197- .(). The acrit ar chs and chit ino zoa from th e Wenlock and Ludlow _Se rie s of th e Ludlow and Mi ll ic h op e areas, Shrop s hi r e. Palaeonto g r . Soc. (M- onog r .) , 124 : 1 一 100, [T he ter mi n olog y, in clud ing a gl ossary, classi f ica ti on an- bio l og ica l aff ini t ies of th e acrit ar chs is out li n ed. A det a il e d secti on deals wi th excy s tm ent op e nin g s '.'The th eory th at acrit ar chs are th e resti ng cy s - ts of naked dino fl ag e llate s is for cefu ll y pr esent e d. This pu bli ca ti on is re- Loceobmli mche _,nJd re. ,dA .f oRr. aunnd . i Ln soieg bhl ti c ihn tIo I I,r Aec.eRn.,t 1 9thq e6 o. r Iine d se xc otnoc etrhn ei n g ge nthe eras,e suobrgg ae nnies rm a,s .] and secti on s of th e Py r rhop h y ta. St ud . Trop . Oceanog r . Mi am i , No. 3 : 94 pp, [T he in tr o ducto ry chap ter outl ine s th e sup r age neric classi f ica ti on , habit at, morph olog y and life cy c le of th e dino fl ag e llate s . It is aconcis e account whic h in cludes deta i l s ga rnered from many sources.] Loebli ch III, A.R., 1970. The amp h ie s ma or dino f lag e lia t e cell coverin g . N. Am. Paleonto l. Conv., Chic ago , 1969, Proc. G, pp. 869-92 9. [A deta il e d secti on on the comp os it ion and str u ctu re of th e cell wall of th e veg e ta tiv e sta g e andresti ng cys t, and cell div isi o n and a comp r ehensiv e sup r age neric classif ic- ati on . ] Muir , M.D. and Sarje a nt, W.A.S., 1971 . An annota te d bib l io g r aph y of th e Tasm-
anaceae and of relate d liv ing for ms {Alga e : Prasin o p h yc eae) . In : S. Ja rdi ne (Ed itor ) .Les acrit ar ches. Edit ion s C.N.R.S., Paris , pp. 56 一 117. (Rath er heavy for the stu dent but a usefu l and uniq u e work on th e ta smani t ids .] Sari e ant, W.A. S., 1969. Mi c rofo s sil e oth er tha n po llen and spo res in pa lyn olog - ica l pr epa rati on s. In : G. Erdtm an (Ed itor ) . Handbook of Palyn olog y :Mo rph olo- gy, Taxonomy , Ecolog y. Munksg a ard, Cop e nhage n, pp. 165-20 8. [The morph olo- gy, habit at and life cyc le of modern din o fl ag e llate is revie w ed. T he morph olog y coyf s tths , e adndin o tchy e s tvs a riiso u csa tse ug bo d riiv zi es di o n asc.c Tohrdei nsg e ctoti opn r ooxni mt h a et e a ,c crhito arra cthe s aanndd ctaavs amtea -nifi ds is pr esent e d in a very readable sty le .] Sarje a nt, W. A. S, 197(J. Xanth i d i a , Pali no sp h eres and 'Hy s tr i x ' : A revi ew of t- he stu d y of fos sil unic e llular mi c rop l ankto n wi th orga nic cell walls. Mi c ro- scop y, London, 31 : 221 一 256. [A n in te resti ng account of the his t o ry of th e stu dy o_f f o ssil din ocy s ts wit h brie f bio g r aph ic al sketc hes of many of the e- arlie r scie n ti st s who stu di es th ese orga nis m s. Also dis c usses veg e ta ti ve st- age , cys t relati on ship , acrit ar chs and ta smanit ids .1 Sarje a nt, W.A. S., 1974. Fossil a 따 Livin g Di no f la g e llate s . Academi c Press, Lon- don, 182 pp. [M orph olog y,li fe cyc le and ecolog y of liv ing dino fl ag e llate s is dis c ussed in consid erable deta i l fol lowed by th e his to ry of stu dy and morp - holog y of fos sil din o fl ag e llate s. The ph il o sop h y and pr oblems of classif ica ti. .: on of liv ing and fos sil din o fl ag e llate s accomp a ny an Ap pe nd ix , give n over so- lely to one of th e exis ti ng classi f ica ti on s for din ocy s ts . Prepa rati on te ch' ni - qu es are also dis c ussed in an Ap pe nd ix. ] Wall, D. and Dale, B., 1968. Modern din o fl a g e llate cys ts and evoluti on of th e P- erid in ia les. Mi c rop a leonto log y, 14 : 265-3 04. (The veg e ta ti ve sta ge , cys t rel- ati o nship is now known for over 30 spe cie s of modern dino f lag e llate s. The concep t of lin eag e s, as based pr im aril y on archeop yle type , is a new fiel d of stu dy .] 참고문헌 Bargh oorri, E.S. and Ty l er, S.A., 1965. Mi c roorga nis m s from th e Gunfl int ch- ert. Scie n ce, 147 : 563 -577. Barss, M.S. and Wi ll ia m s, G.L ., 1973. Palyn olog y and nannofo s sil pr ocessin g te - chniq u es. Geol. Surv. Can., Pap. , 73-26 : 1-25 (pis. 1-2) . Cookson, LC. and Ei se nack, A., 1960. Mi c rop l ankto n fr om Austr a lia n Creta ceous sed im ents . Mi cr op a leonto log y, 6 : 1-18. -Cramer, F.H., 1970a. Dis t r ib u t ion of selecte d Si lu ria nacrit ar chs : Rev. Esp.Mi c rop a leonto l., Sp e c. No. 1 : 1-202 (pis. 1-23 ) . Cramer, F.H., 1970b. Mi d dle Sil ur ia n conti ne nt al movement esti m ate d from ph - ytop l ankto n -fa c ie s tra nsg r essio n . Earth Planet. Set. Lett ., 10 : 87-93 . Cramer, F.H., 1971. A pa ly n ostr a ti gr aph ic model for At la nti c Pang e a durin g Si lu ria n tim e. Bur.R ech. G eo! .M i n. ,Mem.73 : 229 一 235. Cramer, F.H. and Di ez de Cramer, M.D.C.R., 1972. North Arreric a n Si lu ria n p- alyn of a cie s and th eir spa ti al arrang e ment : Acrit ar chs. Palaeonto g r aph ic a , Abt. B, 138 : 107-180 (pis. 31-36) . Deunff , J., 1971 . Le ge nre Polye dryx iu m Deunff . In : S. Ja rdi ne (Edit or ) . Les A crit ar ches. Ed i tion s C.N. R. S., Pari s,_p p . 17-49 . Downie , C., Ev itt, W,R. and Sarje ant, W.A.S.,1963. Di no f la g e llate s , hy s tr ic h osp - heres, and the classi f ic ati on of th e acri t ar chs. S ta n{ord Univ . Pub!., Geo!. Sci. , 7(3) : 1-16. Downie , C., Hussain , M.A. and Wi ll ia m s, G.L ., 1971 . Di no f la g ll ate cy s t and acr- i tar ch associa ti on s in th e Paleog en e of sout h east Eng l and . Geosci. Man, 3 : Ea2t9o 급n , G5 (.p Ll s.,. 1l 9-712 . )A . morph og e neti c serie s of din o f la g e llate cy s ts from the B ra- cklesham Beds of the Isle of Wi gh t, Hamp s hir e , Eng l and . In : A. Farin acci ( Ed i tor ) , Proc. Second Plankt on ic Con{., Roma, 1970. Ed iz io n i Tec n oscie n za, Rome, 1 : 355-379 (pl s . .1- 4) . Ehrenberg, C.G., 1838. Uber das Massenverha. l tn is der ie tz t leb end en Ki e sel- In- fu sorie n und Uber ein neues lnf u sorie n Cong l omerat al s Pol i rschi e fe r von Ja str a ba in Ung a rn. Abh. Preuss. Akad. Wi ss ., 1 836 : l!J9 -135 (pis. 1-2) . Eis e nack, A., 1963. Hy s t ric h osp h a. r en. Bi ol . Rev., 38 : l!J7 -139 (pl s. 2 -4 ) . Ev itt, W.R., 1961. Observati o ns on the morph olog y of fo ssil din o f la g e llate s. M- i crop a leonto log y, 7 : 385 -4 2(J ( pi s. 1 -9 ) . Ev itt, W. R., 1963. A dis c ussio n and pr op o sals concernin g fos sil din o fl a g e llate s , hy s tr i c h osph eres and acrit ar chs, I_a nd IL Proc. Natl . Acad. Sci. , Wash.,49 : 15 8 -164 : 298 -3 (J2 . Ev itt, W.R., 1967. Di no f lag e llate stu die s . II. The archeop yl e. St an fo r d Univ . Pu- bl., G eo!. S ci. , l!J(3 ) : 1-83 (pi s. 1 -9 ) . Ev eitst. ,P Wri.vRa _.t,E '1 9p7u5 b.l iMc aatni uoan l, Sfot ar n afo rT de aUcnhiivn ge rsCito yn,f C er aelnifc .e on Fossil Di no f la g e llat- Ev itt, W.R.and David son, S. E., 1964. Oi no f la g e llate stu die s. I. Oi no f la g e llate c- ys ts and th ecae. St a nfo r d Univ . Puhl., Geol. Sci. , l(J(l) : 1-12 (pl. 1) . Frit sc h, F. E., 1965. The str u c tur e and Rep r oduct ion of th e Alga e. Cambrid g e Univ ersit y Press, Cambrid g e , 791 pp. Gocht, H. and Netz e l, H., 1974. Reste relektr o nenmi k roskop is che Unt e rsuchung e a am Panzer von Perid in iu m (Oi no f lag e l lata ) . Arch. Proti st .e n kd., 1 16 : 381-4H J. G6rka, H., 1969. Mi c roorga nis mes del' Ordov ici e n de Polog n e. Palaeonto l. Pol., No. 22 : HJ2 pp .
Graham, H.W., 1942. St ud ie s in the morph olog y, tax onomy and ecolog y of th e P- erid ini a l es. Carneg ie Inst. W ash. Pub!.,No. 542 : 129 pp. Gray, J. (coord. ), 1965. Techniq u es in Palyn olog y :Part III. In : B. Kummel and D.M. Ruap (Edit or s) , Handbook of Paleonto log ica l Techniq ,ue s. Freeman and C o. , S an F ~anci s co, C al i f. , pp. 469 -7 (J6 . Jux , U., 1968. Uber den Fein bau der Wandung bei Tasmanit es Newt on . Palaeon- tog r aph ic a , Abt. B., 124 : 112-124. Jux , U. 1969. Ober den Fein bau der Zy s te n wandung von Pachy sp h aera marshal/- iae Parke, 1966. Palaeonto g r aph ic a . Abt. B, 125 : 104-111 . Kofo id, C.A ., 19CJ 7 . The pla te s of Cerati um with a not e on th e unit y of th e g- enus. Zool. A nzeig e r, 32 : 177 -183. Kofo id, C.A., 19(J9 . On Perid i n iu m ste cni Jo rge nsen, wi th a not e on th e nomencl- atu re of th e skeleto n of th e Perid ini d a e. Arch.Proti st e n kd., 16( 1 ) : 25-47. Kofo id, C.A. and Skog s berg, T., 1928. The Di no fl ag e llata : the Di no p h ys oid a e. H- arvard Univ . , M us Comp . Zool., M em., 51 : 766 pp. Le b our, M V., 1925. The Di no f lag e llate s of North ern Seas. Mar. Bi o l. Assoc., U. K., London, 25 (Jpp. 25(J Li st e r, T. R ., 197(Ja . The met h od of op e nin g , orie n ta ti on and morph olog y of th e Trem ado cia n acri t ar ch, Acanth o dia c rodiu m ubui Marti n. Yorks. G eo!. S oc., P - roc., 3 8 (1 ) : 47 -55 (pl . 5). Li s te r , T . R., 197(Jb . A rmnog r aph of the acrit ar chs and chit ino zoa fro m th e W- en loc k and Ludlow Se ri e s of th e Ludlow and Mi ll ic h op e areas, Shrop s hir e . Palaeonto gr . S oc. (M onog r .) , pa rt 1 : 1 一 l(J(J (pis. 1-13). Loe bl ic h III, A.R., 197(J. The amp h ie s ma or din o f la g e llate cell coverin g . N. Am. Pal!?o nto l . Conv., C hic a go , 1969, Proc. G, pp. 869 -929. Loeblic h Jr. , A.R. and Tapp a n, H., 1969. Acrit ar ch excy s tm ent and surfa c e ult r- astu c t u re wi th descrip tion s of some Ordov ici a n tax a. Rev. Es;:,. Mi c rop a leon - to l., 1 一 45 -5 7. Mant e ll , G.A. , 1 845. Not es of a mi c roscop ica l exami n ati on of th e Chalk and F- lin t of sout h east Eng la nd wi th remarks on th e Anim alculi tes of certa in Te rti a ry and modern dep o sit s. Ann. Mag . Nat. Hi s t. , 16 : 73-88. Mart in, F., 1969. Les Acrit ar ches de I'O rdov ici e n et du Si lu rie n Beig e s. Inst. R. Sci. Belg. , Mem., 16(J : 175 pp. Newt o n, E.T., 1875. On Tasmanit e and Austr alia n Whit e Coal Geol. Ma g., S- er. 2, 2(8 ) : 337-34 2 (pl. l(J) . Parke, M., 1966. The ge nus Pachy s ph aera(Prasin o p h y c eae) . In : H. Barnes (Ed ito r). Some Conte m p o rary St ud ie s in Marin e Scie n ce. Allen and Unwi n, London, pp. 555 -5 63. S aj_i e ant, W.A. S., 1961 . Mi c rop la nkto n fro m th e Kellaway s Rock and Oxfo r d C- lay of Yorkshi r e. Palaeonto l og y, 4 : 8(J- 118. Sarje ant, W.A.S., 1967. The str a ti gr ap h ic a l dis tr i b u ti on of fos sil dino f la g e llat-
es. Rev. Palaeobot. Palyn ol., 1 : 323 -3 43. Schop£ , J.W . and Bargh oorn, E.S., 1967. Alg a -lik e fos si l s fro m th e Early Pre- cambria n of Sout h Af rica . Scie n ce, 156 : 51J8 -512. St ap li n , F.L. , 1961. Reef contr o lled dis t r i b u ti on of Devonia n mi cr op la nkto n in Al berta . Palaeonto l og y, 4 : 392 -4 24 (pl s . 48 -5 1 ) . Tapp a n, H.an d Loeblic h Jr. , A.R., 197(J. Geobio l og ic im p li c a ti on s of fos sil ph y t - opl a nkto n evoluti on and tim e-spa ce dis tr i b u t ion . In : R.M. Kosanke and A.T. Cross (Edi t o rs) , Sy m p o siu m on Palyn olog y of the Late Creta c eous and Ea- rly Terti ar y. Geol. Soc. Am., Sp e c. Pap. , 127 : 247 一 34( J. Tapp a n, H. and Loeblic h Jr. , A. R., 1 971 . Sur fac e sculpt ur e of the wal l in Low- er Paleozoic acri t ac hs. Mi cr op a leonto l og y, 17 : 385-41 0. Tapp a n, H. and Loeblic h Jr ., A.R., 1972. Fluctu a ti ng rate s of pr oti st a n evolut- ion , div ersif ica ti on and exti nc ti on . In : B.L . Mamet and G.E.C. Weste r ma~ nn (Edit or s) . Sect. 7, Paleont ol og y, Int. Geol. Cong r ., 24t h, Montr e al, 1972, p p. 205-213. Vozzhennik o va, T.F., 1965. Vvedenie v izu chenie isk op a emy k h pe rid i n e ev yk h vodo rosley. (Intr o ducti on to the stu d y of fos sil pe rid ini a n alga e.) Akad. Nauk S. S.S.R., Si b. 0 님 .,.Ins t. Geol. Geofi z:, Tr., 1 56 pp. Wall, D., 1962. Ev ide nce fro m Recent pla nkto n reg a rdi ng the bio l og ica l aff ini t - ies of Tasmanit es Newt on 1875 and Leio s p h aerid i a Ei se nack 1958. Geol. Ma g., 99 : 353-362 (pl. 17) . Wall, 0., Mi cr op la nkto n, po llen, and spo res fro m the Lower Ju rassic in Brit ai - n. Mi cr op a leonto l og y, 11 : 151-19() (pis. 1-9) . Wall, D., 1971 . Bi ol og ica l pr oblems concernin g fos sil i z a ble din o fl ag e llate s. Geo sci. Man 3 : l-15 (pl s. l-2). Wall, D. and Dale, B., 1968. Early Pleis to c ene dino fl ag e llat_e s fro m the Roy a l Socie t y Borehole at Ludl1am. Norfo lk New Phy tol ., 67 : 315-32 6 (pl. 1 ) . Wall, D. and Dale, B., 197(J. Liv ing hys tr i c h osph aerid din o fl ag e llate sp o res fro m Bermuda and Puerto Ri co . Mi cr op a leonto l og y, 16 : 47-58 (pl.1 ) . Wetz e l, 0., 1933 .. D ie in orga n is c her Subsstea nd zi m eenrhtpa le tte r n o egn r aMp hi iks r c ohfeo ns suiln id e n stdr ea st i b-- alti sc hen Kreid e fe u erste i n s, mi t ein e rn gr aph is c hen Anhang. Palaeonto g raph ic a , 7 7 : 147 -186 : Palaeonto g r aph ic a . 78 : 1-ll(J (p i s. 1 -7 ) . Wi ll ia m s, 0.8. , 1971. The occurrence of din o fl ag e llate s . in marin e sedim ents . In : B.M. Funnel I and W. R. Ri edel (Ed i to rs) , Mi cr op a laeont olog y of the Oce-ans. C ambrid ge Univ ersi t y Press, London, pp. 231-24 3. Wi ll ia m s, G.L ., 1977. Bio s tr a ti gr ap h y and pa laeoecolog y of Mesozoic - Cenozoic dino cys ts . In : A.T. S . Ramsay (Edi t or ) . Oceanic Mi cr op la eonto l ogy _ Academi c P ress, London. Zeuner, F.E., 1946. Dati ng the Past, A11 Intr o duct ion to Geochronolog y. Meth u -en and Co., London, 444 pp. (24 pis. ) .
10 花粉과 胞子 (Pollen & Sp or e) 10 • 1 서 언 해양 미고생물학적 견지에서 보떤 호 H 슨과포자는 유일한 타지성 (alloch 노 th onous) 화삭이다. 해양 화분학은 20 세기 중엽 해양학의 발전과 더불어 크게 성장했으나 충분한 자료의 부족으로 아직은 충서학이나 고생태학적 참재 능력이 개발되지는 못한 상태에 있다 . 대부분의 논문들은 지표 샘풍 이나 시추 코아 중의 제 4 기 화석을 주로 다룬 것이었고 최근에 이르러서 야 국지적으로 해역의 화분과 포자에 관십이 높아졌다. 화 분립 은 종 자석물의 남성 생식기관(mi cro gam e t op h ytes) 으로서 꽃식물 이나 혹은 속씨식물의 꽃 방 그리고 겉씨식물의 작은 홀씨 주머니 (mic rosp - o ran g기 a ) 에서 나 오는 것 이다. 꽃 가루받이와 수정이 끝난 후 씨가 될 밑씨 안에서 씨눈이 형 성된다. 침엽수의 화분은 펜실바니아기에서 그리고 속씨 식 물 의 화분은 초 기 백악기에서 처음으로 발견되었다(그림 1 참조) .
 그림 1. 화분
그림 1. 화분
포자는 조류. 균류. 이끼류 및 양치석물과 같은 하동식물이 생산한다. 사일루리아기의 지층에서 처음으로 확인되었으며 원시적인 관다발식물의 포자는 동형 홀씨이거나 혹은 이형 홍씨일 수도 있다. 남성의 작은 홀씨
 그림 2. 포자
그림 2. 포자
와 여성의 큰 홀씨를 생산하는 이형 휼씨 형대는 후기 데본기에서 이미 발견된다(이 경우 작은 홀씨는 화분과 같다)(그랑 2 참조). 동형 홀씨 식물에서는 포자가 동연을 하고 나서 양성의 생식 기관을 만 들든지 혹은 남성 및 여성 생식 기관을 갖는 두 개의 식물로 발달하기도 한다. 호 H 본과 포 ·자 를 현미 경으로 감별하기 위해서는 oil im mersio n ob j ec ti ve 로 x. 950~ 1, 500 정도의 고배율이어야 한다. 추출 방법은 시료의 기질과 연구 자에 따라 다양하다. 암석과 되적률 둥에 따라 다르지만 대체로 3~5cm3 정도의 양이면 충분하고 육지기원의 泥質岩이 좋다. 육지의 생태 환정 번 화를 규명하고자 할 때는 샘플의 간격이 촘촘한 것이 좋다. 시료 채취는 오영울 방지하기 위해 극히 조십해야 하며 시료의 보관도 신경을써야한다. 자세한 시료 처리 방법은 Gray ( 1965) 의 Handbook of Paleonto l ag ica l Te- chru q, ues 을 참고하면 된다. 10 • 2 형태 및 분류 10·2· l 화분 화분과 포자는 그들의 크기. 모양. 개구 (ap e rtu re). 표면 장식 둥의 형 태 와 외벽의 구조적 득칭들에 의해서 감별된다. 대다수의 호 H 손은 20~80µm 의 크기를 가지며 간혹 lOµm 이하나 200µm
이상인 경우도 있다. 대체적인 크기는 종류에 따라 일정하지만 시료의 처 리 방법에 따라 약간씩 크기가 번화되기도 한다. 예를 들어 큘리세란 쟁 라 틴에 화분을 담그어 술라이드를 만들면 약간 부푸는 반면 같은 시료를 실 리콘류에 넣으면 약 30% 정도 크기가 감소된다. 방사상으로 대칭적인 형태의 여러 가지 변형이 보이며 그 중 회전 타원 체형 이 좀더 보편적 이다. 그림 3 에서와 같이 극과 적도의 크기 관계는 일 반적인 형태 그룹을 정하는데 기준이 된다. 측방으로 대칭인 불축한 모습 의 화 분은 원칙적으로 두 그룹에 속하는데 Monoco ty ledons( 백합)과 겉씨 식물이다. 후자는 기포나 날개가 달린 다공질 화분을 생산한다.
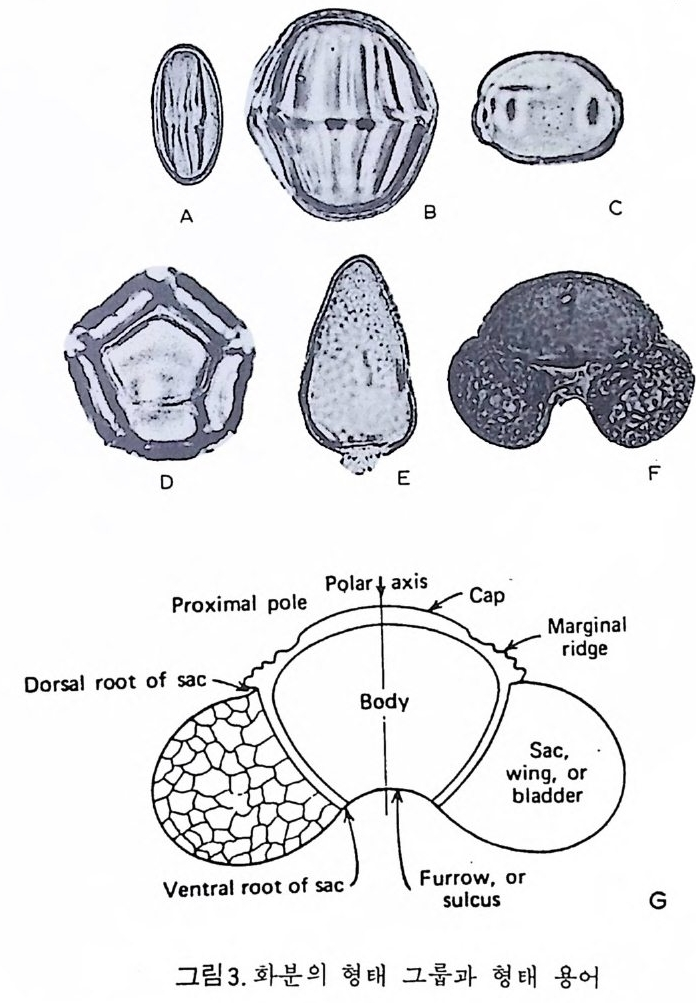 A :
A :
화분 판별에 가장 기본이 도!는 것은 개구의 수와 모양이다. 일반적으로 i so di ame t r i c 한 형태를 p ore 라고 하며 길쭉하고 홈과 같은 형태를 col pi라 한다. 개구가 없는 것 (ina p e rtu rate ). 한 개를 갖는 것 (monopo late or mon- ocolpa te ) , 여 러 개 가 있는 것 (dipor ate , tri p o rate , ste p h anop o rate , or pe rip o- rate ) , 많은 col pi를 갖는 것 (dic o lpa te , tri c o lpa te , s tep h an o c olp a t e, hete r ocol- pa te ,or syn c olp a te ), 혹은 다수의 p ore 와 colpi 를 갖는 것 (dico lpo rate , tr ic o l por ate , ste p h anocolpo rate , pe ric o lpo r ate ) 등 이 있 다.
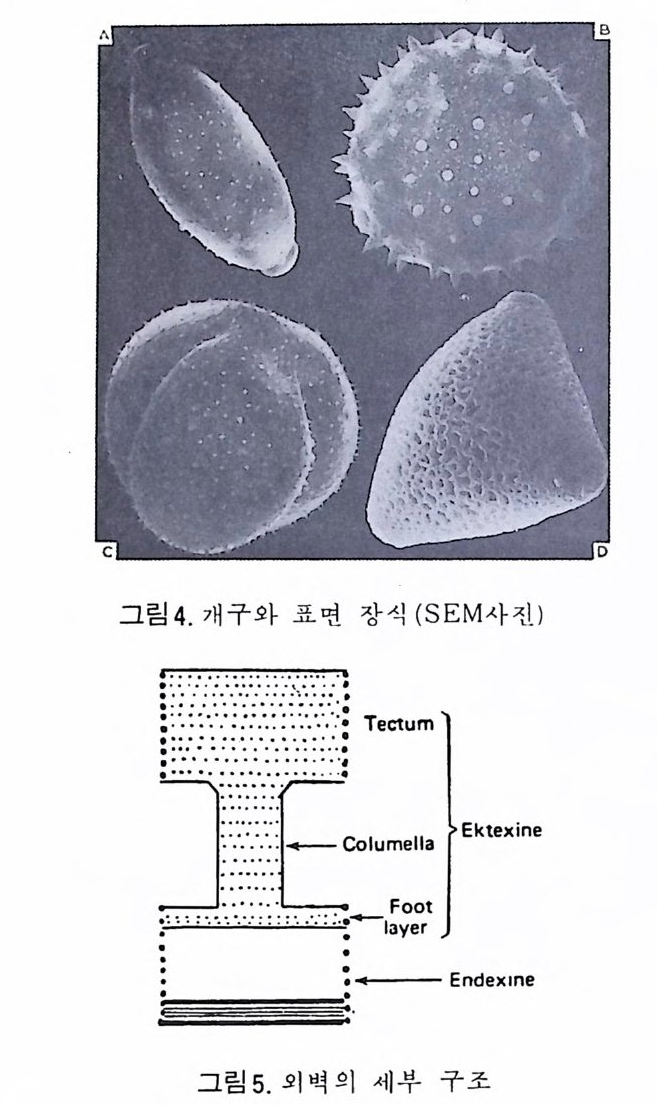 그림;.. 4.. .J.. 개.. ..[구. .와. . 표 면尸….. ...….장 석 二―[ ( SEJrM Ek 사 etEe x전m. m e ) d nx e
그림;.. 4.. .J.. 개.. ..[구. .와. . 표 면尸….. ...….장 석 二―[ ( SEJrM Ek 사 etEe x전m. m e ) d nx e
화분을 감별하는데 가장 큰 진단 가치가 있는 것은 표면 장식과 외벽의 구조일 것이다. 약 12 가지의 장식이 알려져 있는데 이 중에는그림 4 와같 은 것들이 있다. 호냥의 외벽 구조는 그림 5 와 같이 ek t ex i ne 과 endex i ne 으로 구분되며 광 물질과 유기물의 복합체로 구성된다. 내벽은 원형질과 핵을 둘러싸는 셀 물로즈질과 쉽게 파괴되어 화석화되지 못하는 여러 성분으로 되어 있다. 10 • 2 • 2 포자 포자는 구형. 사면체형 또는 신장된 형태를 보이며 성숙되면 한 개씩 혹 은 네 개씩 묶여 산출된다. 신생대의 포자는 호 H t과 비슷한 크기를 갖지 만 더 오래된 포자들은 2mm 에 달하는 것도 있다 . 표면 의 scars 혹은 laesurae 는 한 개 (monolete ) 이 거 나 세 개 (tri le t e ) 혹 은 없 는 (alet e) 경우도 있다. 포자의 벅도 화분과 같이 i n ti ne 과 ex i ne 이 있 으며 p er i ne 이라고 불리는 외벽이 발달하는 경우도있다(그림 6 과 7 참조). 표 면 장식 과 구조는 호 H 본과 같으며 monole te 형보다는 tr ile t e 형 에 더 발달 되어 있다. 고생대나 중생대의 포자에는 crassit ud es, sacci 또는 te et u rn 의 연 장 구 조 등 이 혼 히 보 인다.
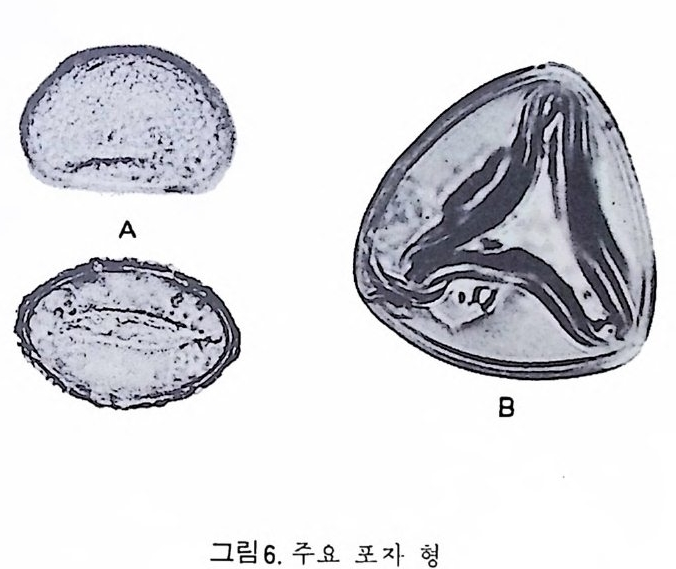 .-.~ ':! ' '' -a.,·,': 도.) ~ 끌 : . s,,. ^” . /
.-.~ ':! ' '' -a.,·,': 도.) ~ 끌 : . s,,. ^” . /
 , . . , ' 흐 5 0
, . . , ' 흐 5 0
10· 3 군집의 형성 해저 퇴적물 속의 화분과 기원지에서의 석물 군집이 생산하 는 화분 사 이의 관계는 매우 복잡하고 다양한 요소들에 의해 파악될 수 있다 . 중요한 요소들 중에는 생태학적인 것과 확산 운반되는 방법 및 속성 작 용 이 포함 된다. 호 H 손과 포자의 기원지에서 식물 군집을 제한하는 환경 요소에 는 기후, 지 형 외에도 토양. 지하수, 화재 및 인간과 같은 국부적 인 요소들이 있다 . 어 느 지역의 식물의 번성은 호目손 군집에 의해 개괄적으로 추정될수 있다. 그 러나 각 식물종의 상대적인 분포량이 그들이 생산하는 화분의 상대량과 그 대로 일치하지는 않는다(그림 8 참조).
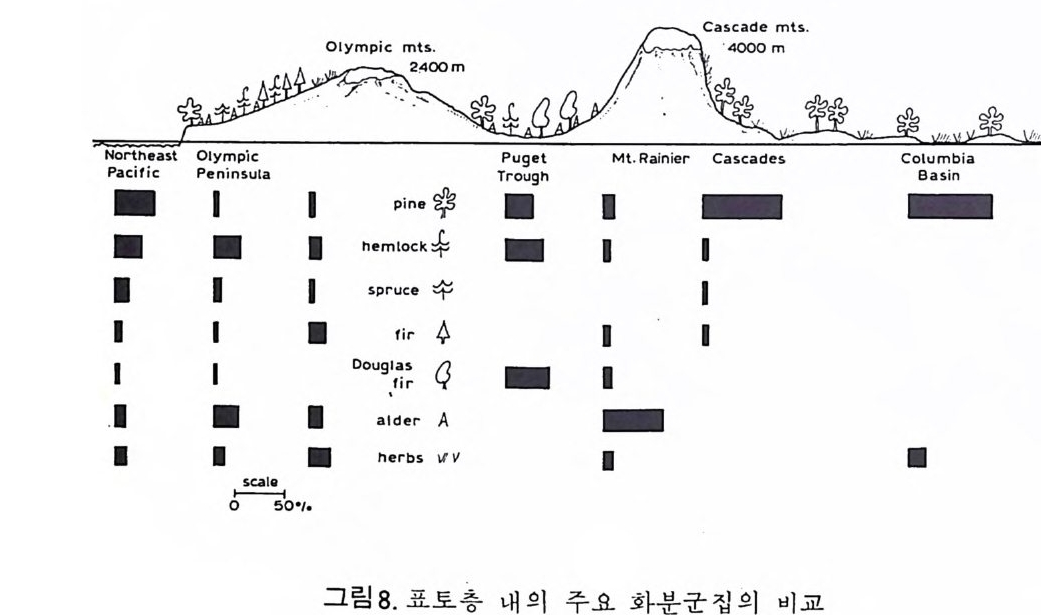 三’-I’II■ ’’I■l■ 〈’’I■■I Dno s uepan gm r lelu 'd aflir ic esor br e rc s- k 三 半니나q4\AI V --:-’’’I -’ \ \-•
三’-I’II■ ’’I■l■ 〈’’I■■I Dno s uepan gm r lelu 'd aflir ic esor br e rc s- k 三 半니나q4\AI V --:-’’’I -’ \ \-•
화산, 퇴적 및 파괴 작용은 화분·포자 군집에 선별적으로 작용해서 이 들을 적당히 차페시킨다. 초기의 운반은 바람에 의해서 (anemo ph i lous), 동 물에 의해서 (zoog a mous) 그리고 물에 의해서 (hy d roph il o us) 이루어지며 이 러한 확산은 10~150km 를 넘지 않는다. 어떤 호 H 손은 바람에 의해서 해변가에서 대륙붕- 끝까지 운반될 수도 있 다. 공기에 의한 확산은 대기의 순환과 호 H t립의 항공 역학적인득칭과 깊 은 관련이 있다. 대륙 연변부의 공기 중의 호 H t 함량은 대단히 낮은 것으 로 보아 바람에 의한 운반은 십해저 퇴적물에 크게 영향을 주지 않는 것 같다. 하천에 의한 운반이 해양 환경으로 호 H 손을 운반하는 데 가장 중요한 것 으로 보인다 . 하천에 의한 운반이 공기에 의한 것보다 9 배 이상이라는 자 료가 있다. 특히 태평양의 경우 하천의 영향이 뚜렷하다(그림 9 참조) .
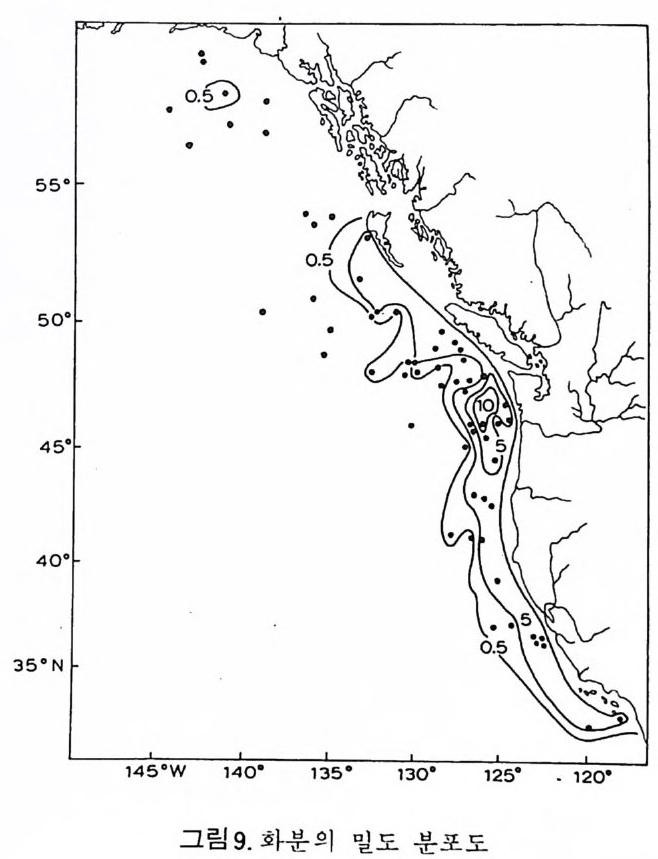 . 0.0 .
. 0.0 .
호봐립이 물에 들어가 젖으면 세립 씽트나 접토와 같은 방식으로 운반 되고 퇴적된다. 자료는 충분치 않지만 호只본과 포자의 수리역학적인 특칭 이 종에 따라 다르다는 것 이 관찰되었다 . 부유성 퇴적물을 조사해 보면 화 분 군집 이 국지적인 환경보다는 수계 분지 전역의 광역적 인 식물상을 대 번한다는 사실도 밝혀졌다. 해양에서는 하천에서와 마찬가지로 호目손은 육원 부유성 물 질과 같은 작 용을 해서 해안에서 멀어질수록 화분의 양은 계속 감소한다. 그러나 복동 태평양에서의 관찰에 의하면 대륙붕에서 적고 대~ 사면이나 대륙대에서 는 많으며 분지에서는 최소의 집중을 보이는 것이 알려졌다. 이러한 현상 은 니질물이 대륙붕 외대를 통과하여 사면에서 해저류에 의해 운반되고 침 전되는 현상과 밀접한 관계가 있는 것으로- 보인다 (Baker, 1973) . 복동 태평양에서의 호 H 손의 집중 분포양상은 부분적으로 표충류의 역할 을 앙시한다. 오리노코 삼각주 지역에서도 대륙 붕 퇴적물 내의 화분 분포는 표충 해류에 의해 영향을 받았다. 그림 10 과 같이 100 개의 시추 코아의 최상부의 화분음 조사해 본 정 과 소 나무가 무성한 육지 부근이나 소나무가 살지 않는 육지 근처의 해역 공히 전체 화분 함량의 최소한 30% 를 소나무 호 H! 이 차지한다. 이러한 현상은 복서로 흐르는 알라스카 해류와 남쪽으로 흐르는 캘리포니아해류에 의한 확산임을 앙시한다 . 화분과 포자의 상이한 파괴작용도 역시 화분이 생성되는 때부터 선택적안 요소로 작용한다 . 호수, 소택지 혹은 실험실과 같은 육지 환경에서 화분 과 포자의 화학적 파괴 작용이 관찰되어 표 1 에 요약되어 있다. 박테리아나 균류의 황동도 또한 파괴작용에 가담한다. 이러한 차별적 파괴 현상은 e xi ne 의 화학. 광물 조성, 포분질의 화학적 구조의 차이, 그 리고 혹은 포분질과 호냐집단 사이의 관계와 깊은 관련이 있다. 산화 작용, bio t u r bati on 그리고 속성작용 (d i a g enes i s) 은 해성충의 유기 탄소의 분포에 영향을 주고 나아가서는 화분과 포자의 함량에 번화를 준 다. 미국 북서부의 대륙 연변부의 해저 퇴적물에서 바교적 높은 화분함량 과 유기 탄소 함량의 최대치는 용존 산소 함량의 최소치와 뚜렷한 상 관 관계가 있다. 흑해에서도 Traverse (1974) 에 의해 이상적으로- 높은 화 분의 함량이 주목되 었는데 이는 최소한 閉海性 인 퇴 적환경과 깊은 관련이 있는 것으로 보인다. 지질시대의 탄산영질 퇴적물에 화분의 결핍 현상은 부분적으로 산화작용 그리고/혹은 퇴적작용과 관계가 있어 보인다. 현재
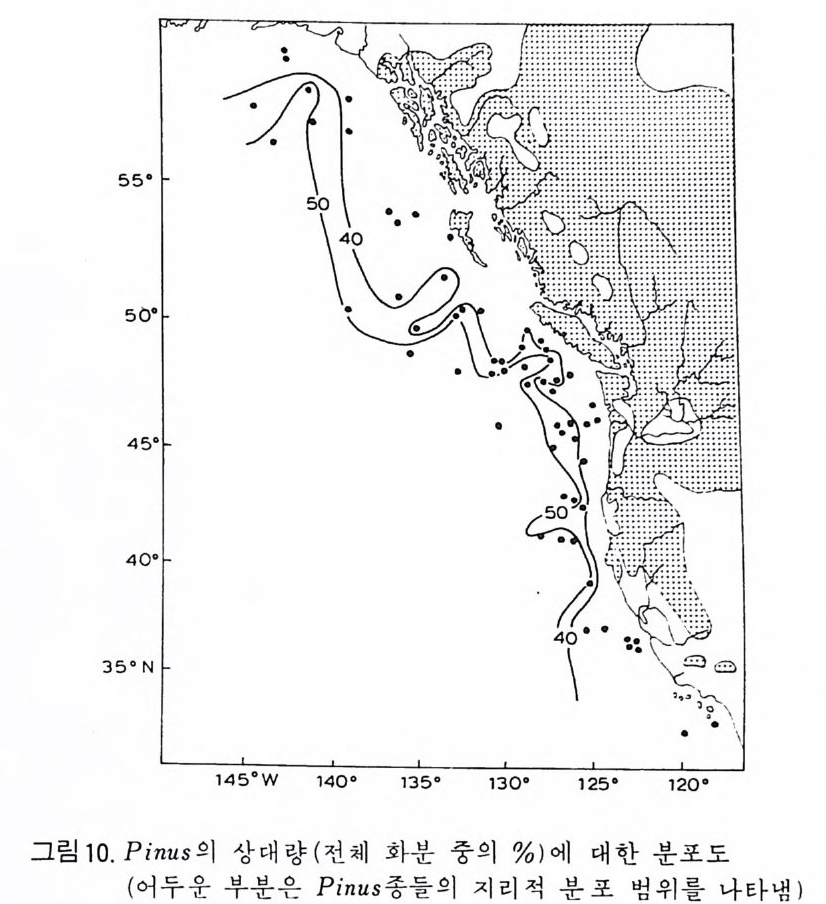 三
三
Bahamas 와 같은 탄산영 물질이 퇴적되는 지역에는 화분이 풍부한데 함량 의 극대치는 세럽질 퇴적물 속에서 나타난다. 속성 작용을 받는 동안 화분립은 황철석. 망간 광물 혹은 기름으로 채 워지는 수도 있다. 시간이 지나면서 포분질은 색상의 전호 H t 겪는데 이러 한 번화는 형광이나 반사 기술로 탐지된다. 화분과 포자는 sa fr a ni n- _ O 에 영색아 되며 영색되는 정도가 지질시대와 대바된다는 사실이 S t anle y (1966b) 에 의해 밝혀졌다. 고생대나 중생대의 화분립은 이 물감을 쉽게 받아들이지 못하여 황색이나 갈색으로. 남는 반 떤 제 4 기 호 H 손은 영색이 되어 현세의 것과 같은 색을 연다. 이러한 색깔 의 번화를 이용하여 퇴적물의 호 H 손 군집 중 오렌 것들과 퇴적 당시의 호 HE 둘을 골라낼 수도 있다 .
 표 1. 주요 화분의 화학적 카괴
표 1. 주요 화분의 화학적 카괴
10·4 생물충서 및 고생태 10·4·l 수평적 분포 호봐과 포자가 모석물을 떠나 복잡한 과정을 동해 종국적으로 해저 퇴적 물에 매몰된 후 이들은 대체로 기원지의 석물상을 반영한다고 보아진다. 다 시 북태평양을 예로 들면 육지의 석물상과 해저 퇴적물의 화분상은 좋은 대미가 가능하다. 그림 11 에서 14 까지는 호 H 손의 상대적인 함량을 각 시료 당 전체 화분의 총량에 대한 4 가지 주요 호目손군의 백분율(%)을 도시한 것 이다. 이들은 각각 Tsuga he t ero p hy lla( 그림 11), P i cea( 그림 12), Sequ oia (그 림 13). 그리고. Q uerc 露(그림 14) 이며 이들의 육지의 번성지가 작은 접들로 표시되어 있다. 일반적으로 호 H 손 군집은 해안과 내륙의 광역적인 식생을 혼합한 것이며 분포 범위는 영구 水系에 따라 다르다. 하천의 부재에 따른 주요 식생울 대변할 화분군의 결핍현상은 여러 곳 에서 확인되었다. 복서 아프리카의 건조한 지역은 영구 하천이 없는 관계 로 遠海 퇴적물에 호 H 손을 공급하지 못한 예가 그 대표적 인 경우이다 (Koreneva, 1971) .
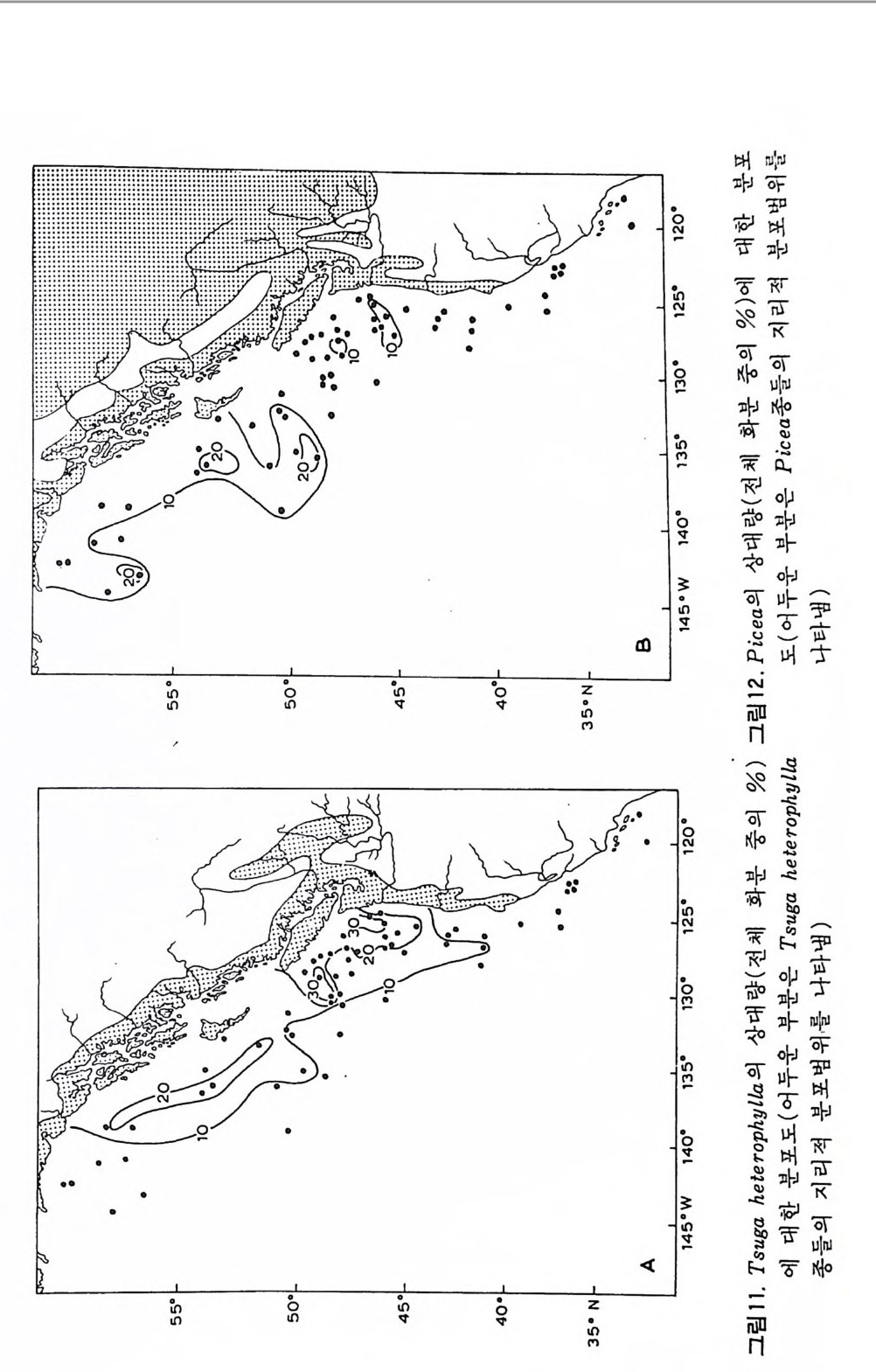 \\\\ ,:·J·: ~t . . .~- . ..•• • ... .. ` · \ B01·2 • 125•013 ·5 311 ·04•5W14 포 분대한상 의ac의 증i분화 e체(량 대% 에) 분위범포를-(P리은분부 운지두의 들적 종어eac i )냄타
\\\\ ,:·J·: ~t . . .~- . ..•• • ... .. ` · \ B01·2 • 125•013 ·5 311 ·04•5W14 포 분대한상 의ac의 증i분화 e체(량 대% 에) 분위범포를-(P리은분부 운지두의 들적 종어eac i )냄타
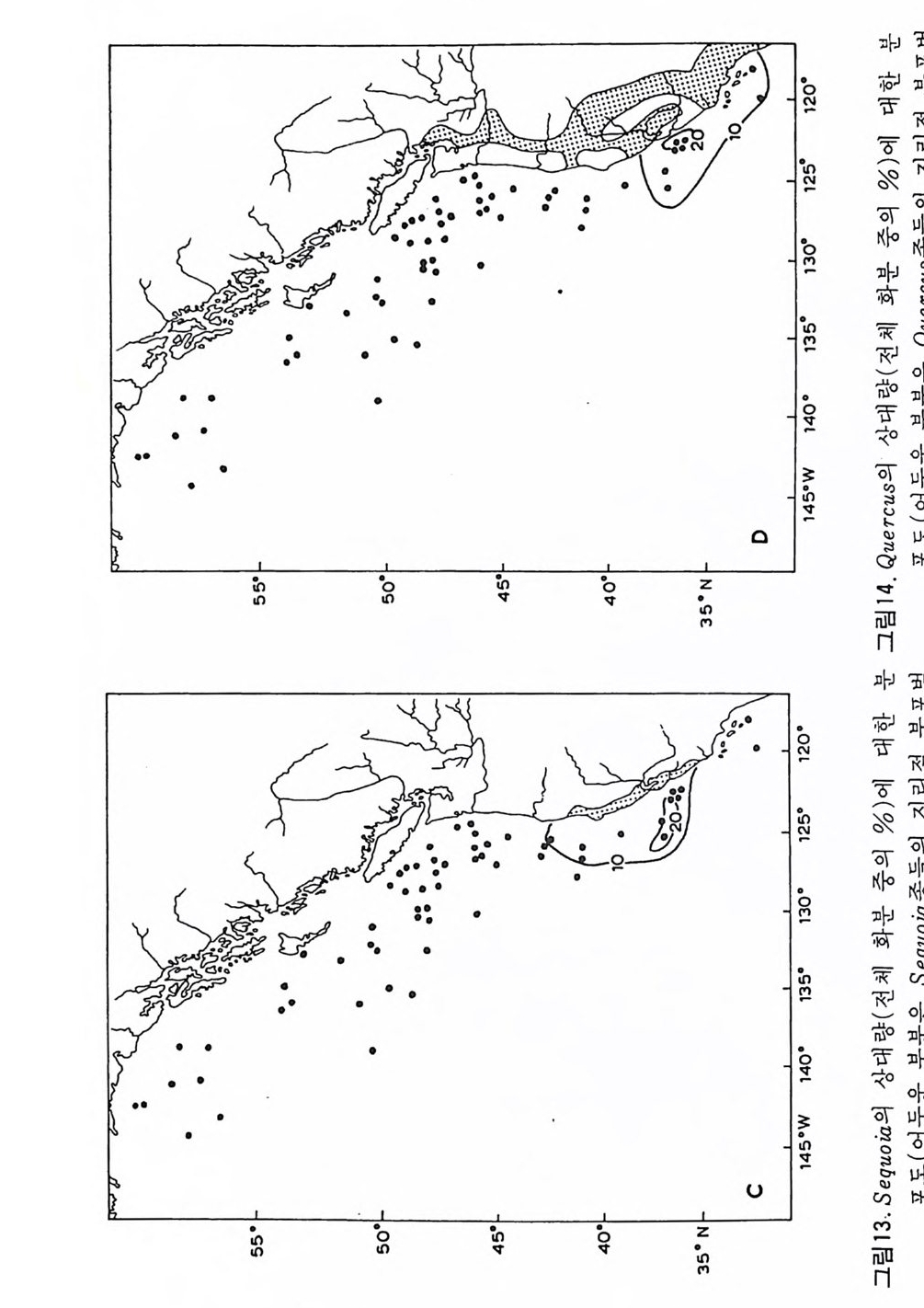
10·4 ·2 수직 적 분포 주요 호넘본과 포자군의 충서 범위는 표 2 에 제시되었다. 소수의 연구 논문이 해양의 제 4 기 이전의 pa l yn omo rp hs 에 관해 발표되 었다. 중생대와 제 3 기의 해저 퇴적물의 대비와 분대가 지역적으로노 화분 •• 포자에 의해 실시되었다. 예륭 돌연 남동 호주의 G ipp sland 분지의 40 개 이 상의 대륙붕 시추 코아륭 분석하여 분대를 실시하였는데 약 150 종의 군집 둘의 수직적 변화를 기초로 한 것이다. 남극해의 시추 코아의 화분을 분석한 결과 후기 울리고世에 초기 빙하 기의 석물상이 존재했음을 알아내었다. 호 H 손과 포자는 생물 충서학적인 도구로 사용될 뿐만 아니라 고기후학적 인 도구이기도 하다. 복서 대서양과 복동 태평양의 해저 코아의 호 H 손 군집 울 분석해 본 결과 적어도 70,000 년 전까지는 이들과 당시의 육상 석물군 집의 대비가 용이하였다. 와성론주의 서부 한 소택지의 화분 군집과 원해 대륙사면의 군집이 그림 15 에 비교되었다. 이들의 호 H 손 군집 분대는 잘 들어맞고 있다. 그림 15(A) 는 서부 Olym pic Thn i nsula 의 후기 빙기 석물상으로 목초 (Gr amin eae) 와 사 초 (Cy pe raceae) 가 주가 되는
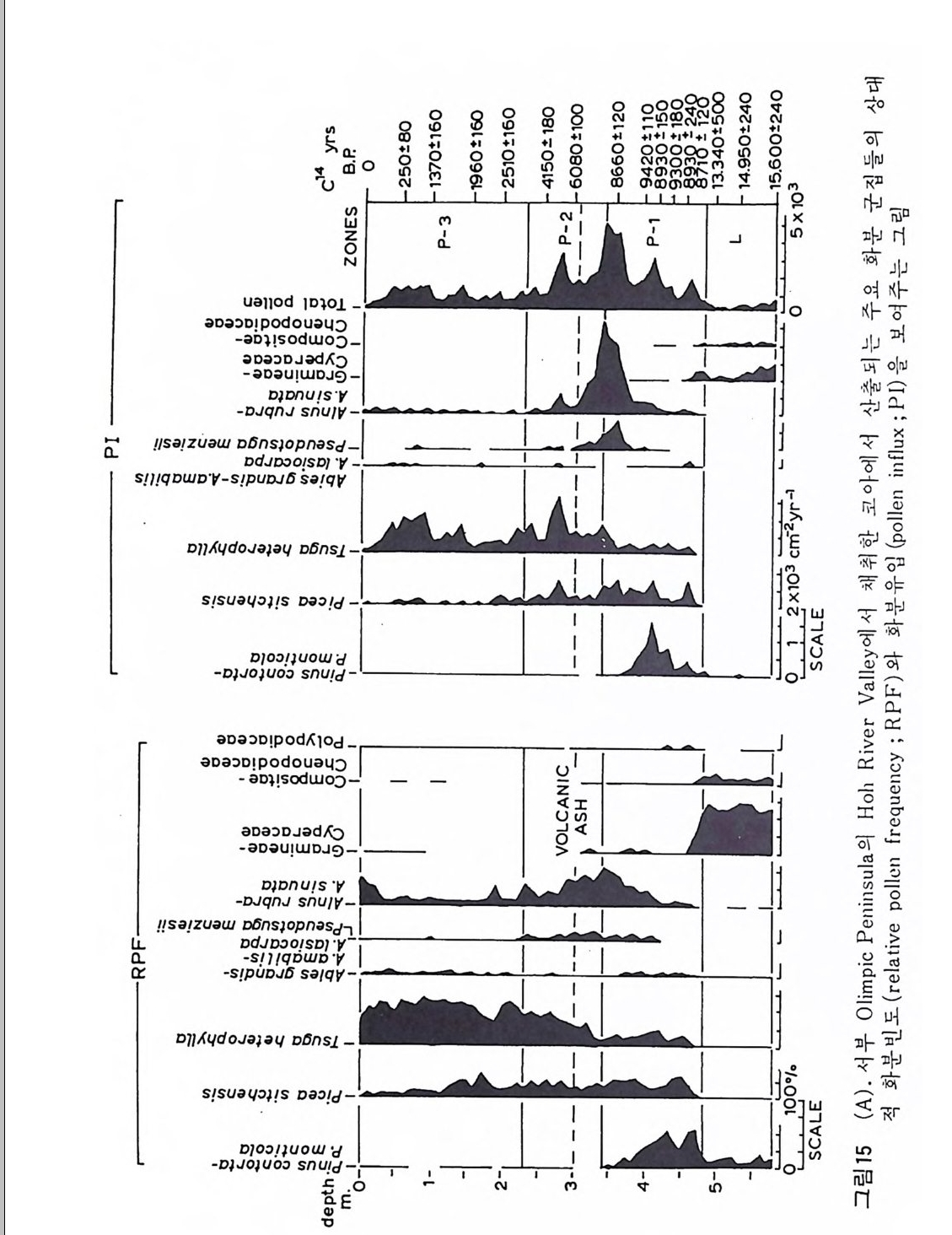 4, · c·s yr 8P .. 。 28 50!017063 !106190016 t 102561 0t 4510으18 0 86000 10t 860016 2t 49021이0 5!39801 0 3090 .18!0 ??;g8접8 1 304050 t.3 1.04 45092t 집대 들상의
4, · c·s yr 8P .. 。 28 50!017063 !106190016 t 102561 0t 4510으18 0 86000 10t 860016 2t 49021이0 5!39801 0 3090 .18!0 ??;g8접8 1 304050 t.3 1.04 45092t 집대 들상의
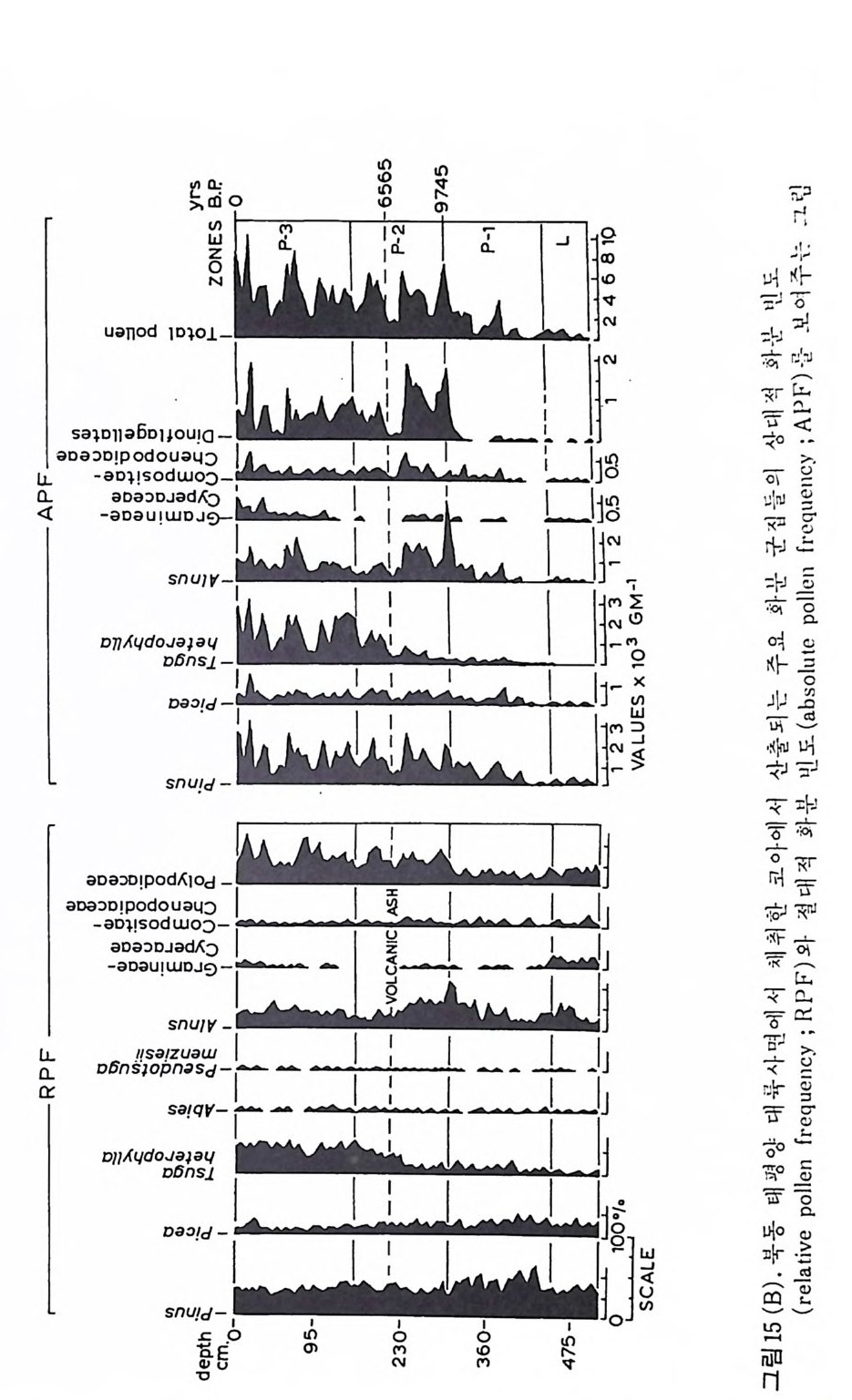

쌍편모 조류와 부유성 유공충/방산충의 비율의 급격한 번화와 일치한다. Heusser 외 (1975) 에 의하면 호1-¾. 방산층 그리고 산소의 동위원소 변화로 추정된 과거 80,000 년 동안의 기후 번동이 서로 일치한다. 육지에서 과거 석물상에 관한 정보가 불충분하거나 없을 때 해저 퇴적물 이 대용품으로 사용된다. Orego n slop e (D.S.D.P. sit e 175) 에서는 과거 920.000 년 동안의 호 H 손 군집이 30,000 년전 이후 것과 다르다는 사실이 밝혀 졌다. 보충교재 Facg r i. K . an d Ive rsen. J. . 1975 . Te xt bo ok of Pollen Analys is . Hafn e r. New York. N. Y. . 295 PP. [N ewedit i on of the comp a ct. i n fo r mati o n-fi lied basic pa lvnolog v tex t. Inc lu de s kevs to po llen ide nt ifica ti on . ] Kap p_R .0 .. 1969. How to know po llen and Sp o res. B rown and Co .. Dubuq u e. Iowa. 249 PP. [In e xp e nsiv e .i llu s t rate d Ke v to majo r po llen and sp o re gr oup s_ l Manto n . A. A. (E dit o r). 1966. Marin e pa lvnolog v _M ar. Geol.,4 (6) : 395-582.(Con- 1ai 1 1s ge n e ra l arti cl e s as well as sp e ci fic stu d ie s fro m vario u s of the wor- Id. l Tsch udv. R. H. and Sc ott . R. A .. 1969. Asp e ct s of Palyn olog y. W ile v. New York. N. Y. . 510 PP. (A n excell ent com p ila ti o n bv fou rte e n aut h orit ies on vario u s a- sp e c1s of pa lvnolog v rang ing fro m the role of po llen and sp o res in the pla - n1 Ki ng d om to the g~ olog ic re cord of pa lvnomorp h s.1 참고문헌 Bake r . F. T .. 1973. Di sl rib u 1i on and comp o sit ion of su s p e nd ed sedim ent in bot t- om wat er s of the Washi ng to n cont in e nt al sh elf and slo p e , J. Sedim ent. Petr - ol. . 43 : 812— 8 21. Cross. A. T .. Thomp s on. G. C. and Zait ze ff . J. 8 ., 19fi 6. Source and dis t ri b u ti on of pa lyn omorp h s in bot tom sedim ents . sout h ern pa rt of Gulf of Calif or nia , M- ar. G eo!., 4 : 467— 5 24. Da\ris .R .B .and Webb 111,T.,1975.The cont em p o rarv dis t ri b u t ion of po llen in easte r n North Ame r ic a : acomp a ris o n wi th veg e ta ti on . Qu at e rnary Res., 5 : 395- 4 34. Dvakows ka. J., 1948. The po llen rain on th e sea on 1he coast of Greenland. B- ull. Acad . Polon. Sci, Lett r ., Ser. 8, 1 : 25 三 13 Erdtm an, G .. 1969 Handbook of Palyn olog y. Hafn er. New York, N. Y .. 486p p,
Florer, L. E., 1972Q u ate r narv pa leoecolog v and str a ti gr aph y of the sea cli f fs. \.\!este r n Olvmp ic Penin s u la, Washin g t on . Qu ate r nary Res.,2 : 202- 2 16. Franklin , J. F. an Dvrness, C. T., 1969 Veg e ta ti on of brego n and Washin g t on . U. S. Dep . Ag r ic . Fores/ Serv. Res. Pap. PNW-80, 216 pp (Po rt l and, 0 re.) . Grav, J. . 1965. Tech n iq u es in pa lyn o)og y. In : B. Kummel and,D., Raup (Edit o rs) . Handbook of Paleont o /og ica l Techniq u es. F reeman and Co., San Fran -
cis c o, Calif ., pp. 471-7 08. Groot, J. J., 1966. Some observati on s of po llen gr ain s in susp e nsio n in the est- uary of th e Delaware Ri ve r. Mar. Geol., 4 : 4()9 -41 6. Habib , D., 1968. Sp o res, po llen and mi cr op la nkto n fro m the Horiz o n Be ta Out c- rop. Scie n ce, 162 ; 148(J - 1481. Hansen, H. P .. 1947. Postg l acia l fo res t suc c essio n , clim ate , and ch ronolog y in the Pacif ic North w e st. Trans. A m. Phil o s. Soc., 37 :.1 3 ( J PP. Hansen, H. P .. 1949. Pollen conte n t of moss po ls te rs in re la ti on to fo res t com - po sit ion . Am.Mi dL Nat. , 4 2(2) : 473-479. Harlett, J. C. and Kulm, L. Q., 1973. Susp e nded sedi m ent tra nsp o rt on th e north e - rn Orego n conti ne nt al shel f. Bull. Geo!. Soc. A m., 84 : 3815-3826. Ha\,jn g a , A. J., 1964. lnvest i ga ti on in to the di ffer enti al cor ros io n su sc ep t ibi l i t y of po llen and spo res. Pollen Sp o res, 6 : 621-63 5. Heusser, C.J. , 1969. Moderr~ po llen spe ctr a fro m the Oly m p ic pe n in s u la, Was h in g ngt on . Bull. Torrey Bot. Club,96 : 4(J7 -417. Heusser, C. J., 1973. Modern po llen spe ctr a from Mt . Ranie r , Washin g ton . North w est Sci. , 4 7 : 1-8. Heusser, C, J. and Florer, L. E., 1973. Correlati on of marin e and con ti ne n ta l Q- uate r nary po llen records fro m the north east Pacif ic and we ste rn Was h in g t- on. Qu ate r nary Res., 3 : 661-67 !J . Heusser, L. E., Shackleto n, N.J. , Moore, T. C. and Balaam, W. L., 1975. Land and marin e records in the Pacif ic North east durin g the last gla cia l in te rv al. G-eol. Soc. Am, Abstr . Prog r .,7 : 1113-1114. Hop k in s , J. S., 195(J. Di ffer enti al floa ta ti on and dep o si t ion of coni fer ous and de- cid u ous tre e po llen. Ecolog y,3 1 : 633 - 641. Kemp , E. M. and Barrett , P.J. , 1975. Ant a rcti c gla cia t i on and early Terti a ry ve- ge ta ti on . Natu re, 258 : 5(J7 -5( J8 . Kemp , E.M . and Harris , W.K., 1975. The vege ta tion of Terti ar y isl ands on th e Nin e ty - East Rid ge . Natu r e, 258 : 3(J3 -3fJ7 . Koreneva, E. V., 1957. Sp o re-po llen analys is of bott om sedim ents of th e Sea of Okhots k . Tr. Inst. Okeanol. Akad. Nauk S.S.S.R., 22 : 221-25 1. Koreneva, E.V., 1964. Di st r i b u ti on and pr eservati on of po llen in sedim ents in the weste r n pa rt of th e Pacif ic Ocean. Tr. G eol. I nst. Akad. Nauk S.S. S. R., l(J9 : 1-88 . Koreneva, E.V., 1971 . Sp o res and po llen in Med iter ranean bott om sedim ents . In: B.M . Funnell and W.R. Ri e del(Edit or s), The Mi c rop a leonto log y of Oceans. Cambrid ge Univ ersit y Press, New York, N.Y., pp. 361-37 1 . Lublin er-M i a nowska, K., 1962. Pollen analys is of th e sur fac e samp l es of bot to- m sed im ents in th e Bay of Gdansk. Acta Soc. Bot. P olon.,31 : 305-312. McAndrews, J.H and Power, D.M., 1973. Palyn olog y of th e Great Lakes, th e su- rfac e sed im ents of Lake Onta rio . Can. J. Earth Sci, HJ : 777-792. Muller, J., 1959. Palyn olog y of recent Orin oco delta and shelf sedim ents . Mi cr - opa leonto l og y, 5 : 1-32 . Peck. R., 1973. Pollen budg e t stu di e s in a sm all Yorkshi r e catc h ment. In : H.J. B . Bi rk s and P.G . West (Ed itor s), Qu ate rnary Plant Ecolog y_ Wi le y, New York, N. Y., pp. 43 -f JJ. Ro ssig n ol-St r ic k , M., 1973. Pollen analys is of some sap r op e l lay e rs from th e deep- sea flo or of the easte rn Medit er ranean. !nit . Rep. Deep . Sea Dril l . Proj XIII : 971-991. Sang s te r, A.G. and Dale, H.M., 1964. Pollen gr ain pr eservati on of under-rep r es- en te d spe cie s in fos siI spe c tr a . Can.J .B ot. , 42 : 437-449. St a nley, E.A, 1966a. T he app l ic ati on of pa ly n olog y to oceanog r aph y wi th refe r - • enc e to th e north weste rm Nt la nti c. Dee,r Sea Res., 13 : 921-939. St a nley, E.A., 19666. The pr ob lem of rew orked po llen and spo res in marin e s- ed i m ents . M ar. G eol., 4 : 397 -4 (J8 . S tan ley, E.A., 1969. Marin e Palyn olog y. Oceanog r . M ar. B i o l. A nnu. Rev., 7 : 277 - 292. S tov er, L.E . and Evans , P.R., 1973 . Up pe r Creta ceous Eocene spo re-po llen zon- ati on , off sh o re Gip ps land Basi n , Austr a lia . Geol. Soc. A ust. Sp e c. Publ. ,4 : 55- 72. St o ver, L.E . and Partr i d g e , A.O., 1973 . Terti a ry and late Creta ceous spo res arrl po llen from the Gip ps land Basin , sout h easte rn Austr a lia . R. Soc. Vic . Proc., 85(2) : 2 37 -2 86. Thomp s on, 0., 1972. Paleoecolog y of th e Pamlic o Formati on , S t. Mary' s County , Maryl a nd. Thesi s , Rutg e rs Univ ersi t y, New B runswi ck , N. J. Trave rse, A., 1974. Palyn olog ic in vesti ga ti on of tw o Black Sea cores. AAPG, Mem., 2 (J : 381 -3 88. Traverse, A. and Gi n sburg, R.N., 1 966. Pal 팟 nolo gy of th e surfa c e sedim ents of the Great Bahama Bank, as r~late d to wate r movement and sedim ent a ti on . Mar. Geol.,4 : 417-459. Wi ll ia m s, G.L. and Brid e aux, W., 1972. Palyn olog ic analys es of cored sed im ents from th e Grand Banks, Newf ou ndland . Am. A ssoc. S tr ati gr . P alyn ol., Abstr . 3 따 Meet. , 136.
한글색인
기 개형충 I2, I4, IO4-50 생활사 106 - 12 연구사 104 - 6 소화신경계 IIl 형 태 l12 - 28 표면장식 115 근혼, 치아구조 I2I 분류 128 - 42 생 태 142 - 6 생충서 146 - 50 계통 진화 33 고 생물지 리 28 고 생태 학 25, 28 고 해양학 28 곤드 와나 29 골적 질 석 회조 193 공생식물 249 국세지 질학 총회 20 군집 분석 238 • 규 3 조 12, 14, 231 - 44 연구사 231 생 황사 231 -3 형 태 • 분류 233 -4 생 태 • 고 생 태 234 - 4° 생충서 • 진화 꾹 0-4 규질 편모충 I2. l4, 작 7 -52 연구사 객 7 생황사 248-5 0 형 태 • 분류 250 생 태 • 고 생 태 250 -l 생 충 서 • 전 화 25 I -2 L 난노 플랑크돈 I3 , 29 , I56 - 85 생황사 158 一 &
연구사 156 -7 분류 • 형 태 I6I - 76 생 태 177 - 8 고생지 리 ·고기후 179 - 81 생충서 • 진화 181 -5 남조식물 I93 C: 단접식 모델 35 대진화 33 도관계 63 T亡 조., .己T 14 □
먹이사슬 16 먹이순환 26 모악 동물 265 목초 3°3 멸종 33 미국 충서학 명명위원회 19 t:t 방사성 동위원소 20-2 1 . 32 방산층 12 , 14 , 29 , 2IO - 23 생활사 212 -4 형태 • 분류 214 -8 생태 218 -9 고생태 • 고해양학 219 - 22 생충서 222-3 방해석 II 병렬진화 38 분대 19 人 사초 3°3 사후 이동 26생물 군집대 21 생물 충서단위 19 생물충서학 19 생존대 19. 20 석 회조 12, 14, 189 - 207 연구사 190 생활사 190 -l 형 태 • 분류 191 - 202 생 태 • 고 생 태 202 -· 4 생충서 204 -7 선태동물 14 시상화석 13 . 26 시준화석 I3 , 231 쌍편모 조류 12, 14, 2&; -86 형 태 • 분류 269 -7 9 생태 279-80 생지 리 • 고생 태 280 -3 생충서 • 진화 283 -6 。 아미노산 생화학적 연구 40 알라스카 해류 298 암권 26 양치식물 291 에브리디안 252-4 생물생태 252-3 생충서 분류 253-4 연안환경 281 영구수계 300 오리노코 삼각주 298 요인 분석 법 85, 238 원생동물 45 위족충류 45 유공충 12, 14, 29, 45-98 연구사 45-7 생활사 47-52
각 52-5 방 56-6o 구공 63-6 분류 68-77 생태 78-83 고생태 83-5 생지 리, 충서 86-8 X 자연도태설 32 적응방산 34 적응수령 38 전 성 가 (acme) 21 점이적 모델 35 접합포낭 279 정향전화 37 제너윤회관 284 종간 전화 33 종다양도 지수 각° 종펴 253 중앙근혼장 I 객 지자기 충서 40 지질대 I9 진핵세포 I90 진화학 32 天 쳐어트 29 충서단연 21, 23 충서대비 2I 포 판 구조론 29 팡게아 29 페어 34 폭발적 진화 34
표 준군집 2I 표준화석 2I 프 테로포드 I3 E 타지 성 화석 29I 탈피 현 상 Io8 투광 대 26 t 해양생 태계 27 해 양생 태 학 25 허족 46 홍조식물 193 화분 • 포자 12, 14, 291 - 307 형 태 • 분 류 292 - 5 생 충 서 • 고 생 태 300 -7 화석 개형충 105 화폐석 46 흑해 298
영문색인
A Acant h aria n s 210 Acrit ar chs 270 Albail le lid s 223 Alabami ni d a e 76 Allog r omi da e 78 Alveoli 63 Alveolin i d a e 72 Ammodis c ace a 71 Amp h is t e g ini d a e 76 Amp h ie s ma 271 Ance!!I ral corallin e s 197 Anomalin a cea 76 Anomalin i d a e 76 Archaedis c id a e 72 Archaeolit ho p h y ll um 197 Arkhang e l skie l l id s 162 Asth e ro lit h s 157 As t rorhiz i d e a 69 At a xop h rag m i ida e 71 B Back-t r ackin g 83 Balsam 212 Bey ri c h ic o p ida 136 Bey r ic h ii da 1틀 29 Be:v r ic h io m orph a 136 Bif or ami na ta 69 Bin o dic o p ina I36 Bi os t rat igr ap h ic unit 19 Bio - z one 19 -2 0 Boliv i n i t ida e 74 Ry th ocvth e r in a e 126 811lim i 11 a('ra 74 Bulim i ni d a e 74 811lim i 11i de a 69
C Calcareous Alga e 189 -207 bio l og y 190-1 morph olog y 191 clas sif ica ti on 192 eco log y 202 pa leo e c olog v 203 bio s t rati gr ap h v 204-7 ske le t a l C. Alga e 193 -2 01 st romato l it e 201 Calca reo us Nonnop la nkt on 156-85 bio l og y 158-61 morph olog v & cla s s if ic a t i on 161 -7 6 eco lo g y 177 -8 pal eo s t ra t igr ap h y 179 pe.le o c lim at e 180 -1 bio s tr a ti gr ap h v, ev o lut ion 181 -5 Calcarin i d a e 76 Carduif ol ia 254 Cass id u lin a cea 74 Cassid u lin i d a e 75 Ce n t rales 234 Cerati oi d 283 Cerati um 199 , 284 Ce ra to b alim i ni d a e 77 Charop h y ta 200 -1 Chia s m olit h us 183 Ci bi c i d i d a e 76 Coccolit h id s 162 Coccolit ho p o res 156 Colanie l lid a e 71 Conodont s 257-6 7 cmloasrpsh i f o icloa gt iy o n > 258-64 pal eo e co log: v 264 bio s tr a ti gr ap h y -26 4
evoluti on 266 Conodont ifor mes 258 Corallin a c eae 191, 197 Corbis e m a 251 Corbis e m i ds 251 Corato l it hi d s · 171 Cordia c e a e 197 , 198 , 205 C r.vo p h il ic 144 Crus h man lab o r a to ry 17 C. S. R. S 25 Cy c loc occoli thu s lep tpo rus 157 Cv elo n e ph e li um 283 Cy m b alo p o r id a e 76 Cy pr id e a 149 Cy p r id ini d a e 108 , 111 Cy t h e r ace a 108 Cy the r e ll o id e a ova ta 150 Cy t h e r in a e 126 Cy t h e ro p te r in a e 126 Cvth e r urin a e 126 D Das _v c la dac ea e 197 -8 , 205 Deate rofo r ami na ta 69 Dia t o ms 12, 26, 231 -44 bio l og y 231 morph olog y, clas s if ica ti on 233 ecolog y 234 -6 pa leoe c olog y 236 -40 bio s t ra ti g r ap h y, evo lut ion 객 0-44 Di ct y o cha 251 Di ct y o chid s 251 Din o fl ag e llata 12. 26, 269-86 morph olog_ v , cla s sif ica t ion 269 - 79
fos sils 274 -278 ecolog y 279 bio s tr a ti gr ap h y , pa leoecolog y 28o-6 Din o p h y si a l es 270 , 277 Di no p h y c eae 271 Di sc oaste rs 161 , 183 Di sc obacea 76 Dis c obid a e 76 Di sc obid e a 70 Dis c oas te rid s 171 D/V Glomar challeng e r 15 E Ebrid i a n s 252 bio l og y, ecolog y 252 bio s tr ati g r aph y 253 Ekte xin 295 Elph id i i da e 74 Emi lia n ia huxley i 16o , 177 Endexin e 295 Endot h y r acea 71 Endoth y r id a e 72 Endoth y r id e a 69 Eourig e rin i d a e 74 Ep ist o mariid a e 76 Ep o nid i d a e 76 Evoluti on 32 Exti nc ti on 33 F Fascic u lit h id s 171 Fi sc he r in i d a e 73 Forami ni f er a 12. 13, 14, 45 -9g bio l og _v 47 - 52 te st 52 - 55 chamber 56 -6o
ap e rtu re 63 -66 classif ica t i on 68 -77 ecolog x 78 -83 pa leoecolog y 83 -85 hio s t ra t igr ap h v 88 - 98 Frustu le 231 Fusulin a cea 72 Fusulin i n a 54. 71, 89, 91 G Gi ga nt oc y pr is 104 Gi nk g o 37 Glabrat e llid a e 76 Glandulin i d a e 74 ' G lobig e rin a cea 76 Globig (' rin a aochy d erma 68 Globig e rin i d a e 76 Globorara_lii d a e 76 Globorot al ia menardii 23 ,, mi oc enic a 85 multi ca merala 85 t runcat ulin o ie s 68 Globot runcanid a e 76 Gondowana 껑 Gony o ulax 284 Gy m nadin i a l s 270 H Haloc~•p r id i d a e 105 Haloc:,pr id i n a 148 Halocvp r if or mes 134 Hannait e s 151 Hant kenin i d a e 76 Hap to nema 158 Helic o sp h aera 183 Helio l it h ae 161
Heli ol it h id s 171 l-lel io zoan 210 Hem: vc :v 1 herin a e • 126 Herodo111s 14 Helerohelic i d a e 76 H. M. S challeng e r 15 Holop la nklonic 234 Hollin o morph a 136 Index fos sils 21 I sla ndie l l id a e 74 Kr it hin a e 126 L Lage nid a e 78 Lag e nid e a 69 Lag_ v nace a 71 LC 'pl?r dit ico p ida 131 L<.'p e rdi1 i id a 104_ 129 Lepi do c:v c lin i d a l? 76 Li 1h asri11 i ds 1711 Li 1h osp h c> r 28 Li lh osI romal ion id s 171 Li 11 1olact> a 71 Li tu olid e a 69 L. 0. C 25 Loxoco11chi1 1a <.' 129 M Melop la nkt o nir 234 Meta cop ina 136 Meta seq u oia 37 Mi lio a cea 72 Mi lio l acea 58 Mi lio l id e a 69 Mi lio l in a 72
M iog yps in i d a e 7b Monocot: vle dons 293 Monoc\1lus 104 Mvodoc o p a 105 Mvdocop ida III , 129, 133 Mus ch e lk rebse 106 Mus sel sh rim p s 106 N Nas scll aria 214 Nas sell aria n s 210 , 217 Nc o c v t he r id e id i n a e 123 ~o d os a ria c e a 74 Nod o sa riid a e 74 No n io n ac ea 75 No n io n id a e 76 Nube rc u la rii da e 71 N'um mulit es 14, 91 Nummulit ida e 76 。 Oi l im m('rsio n obje c t ive 292 Oncolit e 202 Orbit o id a cea 76 Orbit ol in i d a e 71 Osang u lariid a e 76 Os t racoda 12, 14_ 104-5 0 his t o r_v of st u d_v 105 bio l og _v 106 - 12 ornament a t ion 115 -8 vent ral dorsal area 118 classif ica t ion 128-42 ecolog _v 142 -6 bio s t rat igr ap h _v 146-5 0 p Palaeoscenid s 223
Paleot c xt u larii de a 74 Pang a ea 29 Palyn omorph s 270 Pect oc y th erin a e 126 Pennales 234 Perid i n i a l es 270 Perid i n i o i d 283 Perid i n i u m 284 Pi ce a 301 Pi nu s 299 Pi ne st e p pe 3 연 Planomalin i d a e 76 Planorbulin i d a e 76 Plat v cop a 105 Plat yc op ina 137 Pleuros t o mellid a e 75 Podocop a 105 Podorhabid s 162 Podocop ida 129 , 138 -9 Polyc y s t ine s 210 , 214 Pollen, Sp o re 12. 291 -3 07 morp h olog y classif ica t ion 292-4 sp o re 295 bio s rrat igr ap h y 300 pa leoecolog y 301 - 2 Pon to s p h aerid s 162 P rog o nocy t he rin a e 126 Prot o f o ram ina t a ~ Prot o zoa 45 Psy c hrosp h cric 144 Pt e rop o ds 13 Pt yc hocladii dc a 71 Py r op h acus 284 Q Qu ercp s 302
R Radio l aria 12. 13, 210-2 30 lo1i s1 or:-,, of stu dy 210-1 1 •bio l og v 212 morph olog y, clas sif ica t i on 214-8 eco log v 2I8 pa leo e co log y 219-2 2 b ios t ra t igr aph y 222 -3 Rhabdos p h aerid 162 Rhiz o p o da 14 Robert i n acea 55 Robeti na cea 77 Robetu n id a e 77 Rota lia c ea 76 Rota l iin a 73 Rota l ii de a 69, 70 Rota li ida e 76 Rota l ip o rid a e 76 Rzehakin i d a e 71 s S afr an in - O 309 Sarodin a 45 Schackoin i d a e 76 SEM 40 Semi ter tu lariid i a e . 72 Sequ oia 302 Sil ic o fl ag e llata 12, 13 적 7- 52 bio l og y 각 8 - 50 morph olog y, classif ica ti on 250 ecolog y, pa leoecolog y 250 bio s tr a ti gr aph y 251 Solenop o raceae 195 Sp iri li na cea 76 Sp iri ll ini d a e 55, 77 S ph ac roid i n i d a e 74 Sp h enoli thi d s 171
Sp u mallaria 214 Sp u mellaria n s 209 -1 0 , 212 , 223 St r aba 14 St ro mato li te 201 Sup e rpo sit ion 19 T Tasmanit ids 270 Tetr a sodiu m py r op h osph ate 212 Tetr ata xid a e 72 Textu lariid a e 71 Textu lariin a 71 Thecate 253 Thoracos ph ae r id s 171 Tournaye l lid a e 72 Trachy le berid i n a e 126 Trip yle ans 210 Tsug a hete rop h y ll a 301 Turri l ini d a e 74 Ty c hop la nkto nic 234 u Uv ige rin i d a e 74 V Vallacerta 251 X Xesto leberid i n a e 126 z Zenner 윤회 284 Zonati on 19 Zy go dis c id s 163
白光浩 서웅대학교 문리대 지 질학과 0식 동대학원 종업 서독 키일대학교에서 이학박사 학위 동력 자원연구소 선임 연구원 미국 스탠포드대학 객원교수 현재 고려대학교 이과대학 지질학과교수 저서 더구과학』 (I ·II) 논문 「페르시아만의 현생개형류에 대한 연구」외 다수 ,III i'I L 대우학술총서 자연과학 30 해양 □ I 고생물학 1986 년 6 윙 1 일 찍 음 1986 년 6 월 15 일 펴 냄 지은이 白光浩 펴낸이 朴孟浩 펴낸곳 民音社 우편대체계좌번호 010041-31-523282 110 서울 종로구 관철동 44-1 734-4234 • 6110( 편집부) 734-2000, 735-8524( 영업부) *카본은 바꾸어 드립니다 . 값 6,400 원
대 우학술총서 • 자연과학 1 소립자와게이지상로작용 金鎖쥴 뭄/값 3.600 원 2 動力學繼 李炳昊/값 5.400 원 3 질소고정 宋承達 콥/값 2,800 원 4 相轉移와臨界現象 김두절 뭄/값 2,800 원 “計媒作用 陳宗植 뭄/값 2.800 원 6 뫼스바우거分光學 玉恒南 뭄/길 2.800 원 7 국가량원소의영양 昇正子著/값 6.500 원 8 *慕 1 t틀纂와 有農틀熹 1 t合物 尹能民 뭄 / ~l 5. 000 원 9 抗生物質의 全合成 姜錫久 著/값 9.000 원 10 국소적 형태의 A tiy ah-S ing er 지표이론 지동표 著 / 값 2 .800 원 11 Muco po ly saccha 『i des 의 生 1 t릎 및 生物理툰 박준우 뭄 / 값 3. 800 원 12 ASTROPHYSICS ( 天體物理學 ) 洪承!서 著/값 4 . 700 원 13 프로스타글란딘합성 숲한표著 /~l3 . 600 원 14 천연물화학연구법 禹沮植 폼 / 값 5. 300 원 15 脂訪營養 金淑홍 콩/값 6.300 원 16 結晶化유리 金炳歷 콥 / ~l 4. 500 원 17 高分子의化學反廳 趙羲煥 꼴/값 4, CXX)원 18 과학혁명 金永植 홀/값 3,200 원 19 .11 地흩 童基弘 홀/값 4, (XX)원 20 정보이른 한영열 꼴/값 4,500 원 21 原子核反應論 鄭雲赫흄/값 8,500 원 22 禮壇力學 金相哲 흄/값 4.700 원 23 분자궤도이른 이익준흄/값 3. 었)원 24 반응속도 른 정경훈훌/값 4,000 원 25 미분위상수학 李鉉求 흩/값 2,500 원 26 자가공명방법 조성호 블/값 4.700 원 27 1i라스마 를 리학파 빠웅할 죄덕인 흩&t 4, 虹원 28 천문민측파 분석 李時雨 훌RJ. 4, a x)원 29 석탠庫 4 지 변환 기술 金相敦 불 /값 6, 5 00 원 30 海洋畵古生物學 白光浩 불 값 6, 4 00 원 31 편미분방정삭른 김종식 흄 / 값 4, 00)원 32 大統-題諭 소광 섭 불 /값 5. 0 00 원 33 金 屬電 子系의 多體理 論 金德洲 著/값 6, 60C 원 34 복합재료 權肅 1.:: 著/값 5,20