
장현갑 서울대학교 심리학과 및 동대학원 졸업 (문학박사) , 가톨릭의대 생리학교실 및 서울의대 약리학교실 연구원 , 서울대학교 심리학과 교수 및 New York Sta t e Instit u te for Basic Research 객원교수 역 임 . 현재 한국심리학회 생물·생리심리학회 회장, 한국신경생물학회 부회장, 영남대학교 심리학과 교수 . 저서로 『 격리성장과 행동장애』 외 3 권, 역서로 『생리심리학 』 외 3 권 , 논문으로 “Conseq ue nces of ablati ng the
![]()
 생뭄심리학
생뭄심리학
 생물심리학
생물심리학
책 머리에 심리학사에 의하면 생리심리학이란 강좌와 『 생리심리학 원리 』 라는 교과서가 심리학의 창시자인 분트 W. Wund t에 의해 최초로 개설된 강좌였고 교과서라고 한다. 그 후 100 여 년이 지난 오늘날에도 생리심리 학은 심리학의 가장 핵심 기초과목으로 다루어지고 있다. 이것은 생리 또는 생물심리학이 과학으로서의 심리학이 시작되는 태동기부터 오늘 날에 이르기까지 심리학의 기초가 된다는 사실을 강조한 것이라 하겠다. 오늘날 뇌와 행동 사이의 관계를 연구하는 생물심리학은 생리학, 해 부학, 유전학, 발생학, 약리학, 내분비학, 분자생물학, 면역학 등 광 범위한 분야의 생물과학적 발견과 방법에 의존한다. 이처럼 다방면의 생물과학적 방법과 지식을 수용하여 행동과 정산과정을 과학적으로 설 명한다는 것은 대단히 어렵고도 중요한 일이다. 필자는 이 책을 집필 하는 데 있어 뇌의 작용에 관한 최신의 지식을 가능한 평이하게 서술 하려고 하였지만, 이렇게 다양하고 난해한 생물과학의 지식을 체계적 으로 평이하게 다룬다는 것이 천학비재인 저자로서는 너무나 큰 난제 였다. 이 작업을 수행하는 데 4 년 여의 시간이 소요되었지만 여전히 불만스럽다. 이 책은 모두 14 장으로 구성되었는데 이런 내용 구성은 기존의 생물
심리학 교과서의 전형적인 준거를 따른 것이며, 이 책에 소개한 내용 들도 최근에 출판된 이 분야의 대표적인 교과서의 내용과 큰 차이가 없다. 보다 다양하고 입체적인 그림을 인용하여 이해를 도우려 했지만 출판 사정상 여의치 못한 아쉬움이 있다. 이 책을 펴내면서 고마움을 표시해야 할 분이 한두 분이 아니다. 이 책이 나올 때까지 계속 관심과 용기를 주신 고려대학교 김기석 선생님 과 김현태 교수, 서울대학교 이춘길 교수, 충남대학교 손전훈 교수 그 리고 영남대학교 정봉교 교수께 고마움을 표한다. 그 밖에 생물 • 생리 심리학회의 회원 여러분의 관심과 격려에도 고마움을 잊을 수 없다. 더구나 지루한 원고 작성과 교정과정의 피곤한 작업과정에서 헌신적으 로 도와준 박순권 선생과 황성혜 양의 고마움은 두고두고 잊지 못할 것이다• 이 책이 나오게끔 연구비를 지원해 주신 대우재단의 배려와 민음사 편집부의 여러분께도 고마움을 표한다.
1994. 12. 영남대 생리심리학 연구실에서 장현갑
생물심리학
차례 책 머리에 • 5 제 1 장 뇌와 행동 -- 11 1 뇌 ―― 환상의 컴퓨터 • 12 2 심신관계론 : 역사적 변천 • 14 3 심신관계론 : 과학적 증거 • 20 4 생물심리학의 연구방법 • 23 제 2 장 신경원과 시냅스 -- 31 1 신경원의 구조 • 32 2 신경교세포 • 35 3 신경세포막 : 유동성 모자이크막 • 37 4 신경원의 전기신호 • 38 5 시냅스 전도 • 53 제 3 장 신경 계통의 해부학 -- 63 1 말초신경 계통 • 64 2 중추신경 계통 • 72 3 뇌 속의 주요 구조 • 74 제 4 장 뇌의 발달과 노화 -- 83 1 뇌의 발달단계 • 84 2 손상에 대한 신경조직의 반응 • 99 3 노화 • 104제 5 장 시각 -- 133
1 눈의 구조 • 114 2 시각계통의 해부학 • 115 3 망막에서의 시각전환 • 119 4 이중이론 • 122 5 수용야 • 124 6 X, Y 및 .W세포 • 125 7 LGN의 반응 특징 • 127 8 색채지각 • 128 9 뇌피질 신경원 : 세부특칭탐지기인가, 공간빈도분석기인가? • 132 10 양안시 • 136 11 기타 시각영역 • 137 제 6 장 기타 감각 -- 147 1 청각 • 147 2 평형감각 • 157 3 미각 • 158 4 후각 • 160 5 체성감각 • 162 6 통각 • 167 제 7 장 운동 -- 175 1 근육 • 176 2 척수 • 180 3 뇌간 • 184 4 운동피질 • 186 5 기저핵 • 191 6 소뇌 • 193 제 8 장 갈증과 기아 -- 197 1 갈증 • 198 2 갈증의 이중결핍이론 • 2013 기아감 • 206
4 신진대사의 양상 • 207 5 기아감과 포만감의 신호 • 208 6 과식증 • 213 7 의측 시상하부 증후군 • 217 8 체중의 조절과 조절점 • 218 제 9 장 성 행동 -- 221 1 호르몬 • 222 2 호르몬과 발달 • 230 3 호르몬이 성 행동 발달에 미치는 영향 • 240 4 태아기 테스토스테론 투여가 인간행동에 미치는 효과 • 244 제 10 장 수면과 꿈 -- 249 1 수면의 측정 -- EEG • 250 2 밤 수면 • 252 3 수면장애 • 256 4 수면의 뇌 과정 • 258 5 주기성 수면 • 262 제 11 장 정서 행동 -- 267 1 정서란 무엇일까? • 267 2 정서와 자율신경 계통 • 271 3 공격성 • 276 4 쾌중추 • 281 제 12 장 학습과 기억 -- 285 1 학습과 기억의 세포학적 바탕 • 286 2 척추동물의 뇌에서 일어나는 고전적 조건화 • 294 3 장기증강 • 295 4 건망증 • 297제 13 장 뇌반구와 인지기능 -- 307
1 좌우 뇌반구의 해부학적 차이 • 308 2 뇌반구의 특수성 • 309 3 전두엽 • 317 4 반구 우세성과 손잡이 • 320 5 실어증 • 324 6 대뇌 우세성의 발달과 피질의 기능단위 • 329 제 14 장 정신장애 -- 333 1 정신분열증 • 334 2 감정장애 • 347 3 불안장애 • 355 4 아동기 장애 • 360 참고문헌 • 363 찾아보기 • 393제 1 장 서장: 뇌와 행동 마음과 신체 또는 마음과 뇌는 서로 어떤 관련성이 있을까? 이른바 심신관계론 (m i nd-bod y pro blem) 또는 보다 현대적인 의미에서 뇌와 행 동 사이의 관계 (brain - behavio r p roblem) 를 밝혀 보려는 이 의문은 인 간의 많은 의문들 가운데 가장 오래 된 의문이며, 또한 가장 풀기 어 려운 의문의 하나였다. 오늘날 이 물음을 탐구하는 전문 영역의 심리학을 생물심리학 (bio lo g ical psy c h olog y) 또는 생 리 심 리 학 (ph ys i o lo g ical psy c h olog y) 이 라 부르며, 이 심리학은 최근 신경과학 (neurosc i ence) 의 놀라운 전보에 따 라 엄청난 발전을 이루었다. 예컨대 생물심리학자들은 마음과 뇌는 어 떤 관련성이 있으며, 어떤 특정한 마음이나 행동을 일으키는 뇌의 실 체는 어디이며, 감각적 정보들은 뇌의 어떤 부위에서 어떻게 처리되고 통합되는지, 학습과 기억, 수면과 꿈 또는 언어나 인지 같은 고등정신 현상들이 뇌와 어떤 관련이 있는지에 대한 과학적 해답을 추구한다. 나아가 생물심리학자들은 우울증 환자, 정신분열증 환자 또는 난폭한 폭력배와 같은 심한 정신장애자가 뇌 기능의 이상과 어떤 관련이 있으 며, 불면증이나 불안신경증과 같은 신경증 환자라든가 스트레스에 의 한 심신증(p s y chosoma ti c dis o rder) 환자의 예방과 치료를 연구하기도 하며, 뇌 손상 이후에 오는 행동장애의 개선을 연구하기도 한다. 지난 40 여 년 동안 행동을 생물학적으로 이해하려는 생물심리학의
지식은 실로 많이 진전되었으며, 오늘날 생물심리학은 신경과학의 주 요 영역의 하나로 취급되고 있다. 그 예로 미국 신경과학회 (So ci e ty for Neurosc i ence) 는 17,500 명의 회원을 가진 큰 학회인데 그 중 심리 학자는 1990 년의 경우 회원수나 연구업적 면에서 세번째로 큰 그룹을 이루고 있으며, 젊은 학자들만을 고려한다면 가장 큰 그룹으로 부상되 어 있다. 한국의 경우도 1989 년 한국심리학회 산하에 생물 • 생리심리 학회가 창립되었으며, 1992 년에는 한국신경생물학회와 한국신경과학회 가 창립되었는데, 이 두 학회에 생물심리학자들이 다수 참여하고 있다. 이 장에서는 생물심리학의 전통적 관심이었던 심신관계론의 전개과 정과 현대 생물심리학의 위상, 생물심리학의 연구방법 등을 중점적으 로 소개한다. 1 뇌-환상의 컴퓨터 20 세기 후반 컴퓨터의 개발로 인류의 삶은 엄청나게 변화되었다. 그 러나 오늘날 현존하는 최고 성능의 컴퓨터보다 훨씬 앞서가는 환상의 컴퓨터를 한번 생각해 보자. 죽, 수백억 개의 정보들이 저장되고 처리 될 수 있으며, 정보를 한 체계로부터 다른 체계로 재정리할 수도 있 고, 나아가 환경 속에 존재하는 수많은 종류의 상이한 자극에 대해 파 장을 맞출 수 있고, 의부 자극이 없더라도 언어를 자발적으로 생성할 수 있고, 또한 과거에 한 번도 구사하지 않았던 문장을 말할 수 있고, 문법 구조도 이해할 수 있는 고성능의 컴퓨터를 생각해 보자. 바로 이런 환상의 컴퓨터가 인간의 뇌이다. 비록 인간의 뇌는 지금 까지 자연이 만들어 놓은 가장 놀라운 생물학적 체계이지만 부족한 점 도 많다. 이 환상의 컴퓨터도 우주 속에 존재하는 방대한 범위의 에너 지를 모두 다 받아들이지 못한다. 예컨대 가시광선 밖에 있는 광선 에 너지를 볼 수도 없고, 가청범위 밖의 소리도 듣지 못하며, 어떤 독가 스의 냄새도 말지 못하며, 단 10 분 동안만 산소 공급이 중단되어도 영
원히 쓸모 없게 된다. 또한 이 놀라운 정보처리 능력과 분석기술도 감 정이 격앙되거나 침울할 때라든지, 뇌 속의 화학적 조건이 조금만 이 상해져도 엉망이 될 수 있다. 이처럼 우리의 뇌가 항상 안정적인 기능 만 수행하는 것은 아니다. 그렇지만 우리의 뇌는 수십 년 전에 있었던 사건들을 회상해 낼 수도 있고, 한 번도 경험하지 못한 미래의 사건을 예측할 수도 있으며, 미운 감정을 느낄 수도 있고, 사랑의 감정도 느 낄 수 있으며, 환상적인 꿈도 꿀 수 있는 컴퓨터이다. 오늘날 이런 놀라운 마음의 세계를 생성시키는 곳이 뇌라는 것을 의 심하는 사람은 없다. 그러나 과거에는 마음이 자리하는 곳이 뇌가 아 니라 심장(우리말에 마음이 자리하는 장기란 의미에서 심장이란 말이 유래 되었듯 서양에서도 감은 생각)이란 견해(예컨대 아리스토텔레스)가 오랫동 안 지속되어 왔다. 마음이 자리하는 곳이 뇌라는 이론적 근거를 최초 로 제시한 사람은 프랑스의 철학자요 수학자였던 데카르트(1 596-1650) 였다. 데카르트는, 실체는 두 개의 상이한 유형으로 존재한다고 믿었 는데, 그 중 하나는 물질적 실체 (ph ys i ca l ma t er i al) 로서 이것은 뇌를 포함하는 우리의 신체적인 속성이고, 또 하나는 마음을 포함하는 비물 질적 실체 (nonp hy s i c al ma t er i al) 라고 했다. 이러한 두 개의 실체론을 이원론 (dual i sm) 이라 하는데, 데카르트는 이 두 실체가 서로 만나 상 호작용을 하는 장소가 뇌 속의 송과선 (pine al gla nd) 이 라고 주장했다. 오늘날 대부분의 생물심리학자들은 데카르트와 달리 일원론 (mon i sm) 을 믿는다. 다시 말해 우주에는 단지 한 종류의 실체만이 존 재하며, 뇌를 비롯한 우주의 삼라만상은 이 하나의 실체에 의해 움직 이는 것이라는 견해이다. 그러나 사고, 환상, 기억 등과 같은 정신적 사건들에 대한 견해는 학자마다 다르다. 예컨대, 마음이란 단지 뇌의 활동을 설명하기 위해 사용하는 단순한 개념에 불과하다는 견해, 마음 이란 자의식 (se lf -awareness) 이라는 특칭적 면을 드러내는 한 과정에 불과하다는 견해, 마음이란 복잡한 뇌 속에 있는 수백억 개의 신경세 포들이 만들어내는 신경망 조직에 의해 파생되는 속성 (emer g en t p ro perty)에 불과하다는 견해, 그리고 마음이란 뇌의 작용에 수반되는
별 의미 없는 부산물에 불과하다는 견해들이 제시되어 있다. 우리는 앞으로 뇌의 기능에 관한 많은 상이한 면들을 알아보게 될 것이다. 예컨대 개개 정보들이 뇌 속의 신경세포 내에서나 사이에서 어떻게 처리되며, 뇌 속의 화학적 조건이 바뀌어 정신장애가 생겼을 때 정보의 흐름에 어떤 장애가 생기는지, 그리고 감정이 변화될 때 일 어나는 신경계통상의 변화라든지, 배고품이나 목마름이 일어날 때 이 를 어떻게 알아차리는지, 나이에 따라 뇌가 어떻게 발달하며, 노화나 손상에 의해 뇌에 어떤 변화가 일어나는지 등도 알아볼 것이다. 또한 학습과 기억이 일어날 때 뇌에는 어떤 변화가 생기며, 단편적인 운동 요소들이 어떻게 복잡한 운동으로 통합되는가 하는 점에 대해서도 알 아볼 것이다. 2 심신관계론 : 역사적 변천 2.1 고대로부터 근대까지의 변천 뇌와 행동 사이의 관계를 밝히려는 심신관계론은 고대로부터 현대에 이르기까지 철학, 심리학, 생물학 또는 의학 분야의 중요한 관심거리 였다. 비록 뇌와 행동과의 관계를 실험적으로 연구하게 된 것은 200 여 년 정도의 짧은 역사에 불과하지만, 이 문제는 고대 그리스 시대부터 학자들 사이에 많은 관심과 논쟁을 불러일으켰다. 기원전 5 세기경 히포크라테스(기원전 460-380) 는 간질은 〈 뇌장애 〉 에 서 기인되며, 쾌락, 환희, 우울, 익살과 같은 정신현상도 눈물처럼 뇌 에서 솟아나오는 것이라고 했다. 위대한 고대 그리스의 철인 소크라데 스(기원전 469-399), 풀라톤(기원전 428-348) 및 아리스토텔레스(기원전 384-322) 는 우리가 어떻게 객관세계를 인식하며, 사고, 감정, 환상과 같은 심리적 세계가 도대체 어떤 것인가에 관해 깊은 사색을 했었다. 이 시기의 철학자들은 어떤 특정 사건을 그들이 가정하는 한 가지 원
리로부터 추론해 나가는 연역적 사고를 주된 방법으로 사용했다. 그러 나 과학이란 새로운 관찰들을 근거로 하지 않고서는 존재할 수 없다. 따라서 이들 고대 철학자들의 생각이 신경과학에 기여한 정도는 철학 이나 논리학의 발전에 기여한 정도와 비교할 때 극히 미미한 것에 불 과했다. 고대 그리스 문화가 쇠퇴한 후부터 르네상스 시대가 전개되기까지의 오랜 기간인 중세 암흑시대에는 현세보다 내세에 관심을 두는 기독교 신학이 지배적이어서 인간 문제와 같은 현세 문제는 큰 관심을 끌지 못했다. 1543 년 Andreas Vasa li us(1514-1564) 가 죽은 사람의 뇌를 해 부하여 관찰한 뇌의 해부학은 신경과학의 역사로 볼 때 매우 중요한 것이었지만 당시에는 별 관심을 끌지 못했다. 흔히 현대 과학은 17 세기부터 시작되며, 요하네스 케플러 (1571- 1630)I 갈릴레오 갈릴레이 (15 64-1642) 그리고 아이작 뉴턴 (1642-1727) 과 같은 3 대 과학자에 의해 시작되었다고 한다. 그러나 이들은 모두 물리 학자였고 기계론자들이었다. 이들과 비슷한 시기에 등장한 윌리엄 하 비 (1 578-1657) 가 1628 년 〈 혈액 순환의 원리 〉 를 발견했는데 이 발견은 생물학사상 획기적 사건이었다. 더구나 1620 년 프랜시스 베이컨 (1561-1626) 에 의 해 제 시 된 〈 귀 납법 〉은 많은 상이 한 관찰 자료를 바탕 으로 일반화를 시도하는 논리적 방법으로서, 아리스토텔레스에 의한 〈 연역법 〉 의 맹점을 보완할 수 있었기 때문에 과학 연구를 위한 새로운 논리적 방법으로 수용되었다. 2.2 생리학의 태동기 생리학은 심리학보다 훨씬 일찍 발달한 과학이다. 그러나 윌리엄 하 비가 1628 년 〈 혈액 순환의 원리〉라는 획기적인 생리학의 원리를 발표 하였지만, 신경 계통의 기능에 관한 연구는 19 세기가 시작될 때까지도 별로 없었다. 19 세기에 접어들어 영국 사람 Charles Bell (1 774-1842) 과 프랑스 사람 Fran<;ois Mage n die ( 1783-1855) 가 거 의 같은 시 기 에 각기
독립 적 으로 〈 감각 정 보는 척추의 후근 (dorsal root) 을 통해 중추신경 계 로 전달되고, 운동 정보는 척추의 전근 (ven t ral roo t)을 동해 말초근육 으로 전달된다 〉 는 이론바 〈 벨-마장디 법칙 〉 을 발표하였다. 이것은 곧 신경 계통에는 상이한 기능을 하는 영역이 독립적으로 존재한다는 사 실을 시사한 것으로서, 뇌와 척수의 기능을 알아보기 위한 많은 후속 연구를 자국한 계기가 되었다. Bell 과 Ma g end i e 의 발표가 있은 후 몇 년 사이 에 , 신경 계통의 성 질에 관한 많은 주요 발견들이 제시되었는데, 그 중 특히 중요한 발견 은 Herman Von Helmhol t z (1 821-1894) 에 의한 감각생리학적 발견이 다. 그는 시각에 있어서 색채 지각과 관련하여 삼원색 이론(t r i chromatic th eo ry)을 제시하였고, 청각이론으로 장소설(p lace th eo ry)을 제시하였는데, 이것은 오늘날까지도 큰 영향력울 갖는 감각의 주요 이 론으로 간주되고 있다. 2.3 다윈과 진화론의 영향 1859 년 찰스 다윈 (1809 - 1882) 은 인간에 관한 기존의 관점을 뒤흔들어 놓은 『 종의 기원에 관해서 (On the Ori gin of Sp e ci es) 』 란 책울 출판했다. 이 책은 그 이전에 출판된 어떤 책보다도 인간 자신에 대한 관점에 깊 은 충격을 주었다. 진화론 (evolu ti on i sm) 이라 불리는 다윈의 이론은 오 늘날의 신경과학에도 많은 영향을 주고 있다. 이어 1872 년에 출판된 『 인 간과 동물의 정 서 표현 (&pre s sion of the Emoti on s in Man and Ani- mal) 』 이란 책에서 그는 인간과 동물의 정서표현, 특히 안면 표정에 있 어서 어떤 일관된 연속성이 있다는 증거를 제시하였다. 이 책은 오늘 날의 정서 연구에 흔히 인용되는 주요 참고서의 하나가 되었다. 〈모든 種의 동물들 사이에는 어떤 연속성이 있다 〉 고 주장한 다윈의 견해는 생물학자나 심리학자들로 하여금 정서나 사고와 같은 이른바 〈인간적 속성 (human a ttri bu t es) 〉조차도 생물학적 바탕을 갖는다는 견 해를 유도했을 뿐만 아니라, 동물에게도 학습이나 추리와 갇은 고등정
신현상이 존재한다는 생각을 갖도록 하였다. 다윈 이후 과학자들은 인 간적인 것과 동물적인 것을 엄격하게 구분할 수 있는 객관적 기준이 없다는 견해를 갖게 되었는데, 이런 경향은 신경과학이나 심리학에서 도 마찬가지였다. 2.4 심리학의 태동기 심리학이라는 새로운 과학을 태동시킨 최초의 심리학자는 독일의 Wi lhe lm Wund t(1 832-1920) 였다. 비록 Wund t는 자기 스스로 생리십리 학이라고 불렀던 실험심리학을 창설하였지만, 20 세기가 시작되기 이전 까지 엄격한 의미에서의 생리심리학 연구는 Wund t의 실험실을 포함하 여 세계 어느 곳에서도 볼 수 없었다. 그럼에도 불구하고 Wund t는 그 가 공식 적 으로 학문을 시 작한 1853 년부터 사망한 1920 년까지 70 여 년 간 491 개 의 연구주제 에 53, 735 쪽 분량의 방대한 연구물을 쏟아 내 었 고, 십리학 역사상 최초의 심리학 실험실을 독일의 라이프치히 대학에 설립한 공적을 남겼다. Wundt가 사용했던 아이디어들은 오늘날에는 잘 통하지 않는 단순한 역사적 사건에 불과하다. 왜냐하면 그는 의식을 요소 (elemen t)로 분석 하는 것이 심리학의 주된 과제라고 했으며, 의식을 분석하는 수단으로 자신의 연구실에서 훈련받은 사람들에 의한 내관법 (intr os pe c ti on) 이란 방법을 사용했는데, 이 방법은 주관적 경험 분석의 테두리를 크게 벗 어나지 못했기 때문에 현대 과학의 방법론과는 많은 차이가 있었다. Wund t의 가장 뛰어난 제자였던 Edward T it chner (1 867-1927) 가 미 국으로 건너가 코낼 대학에서 내관법을 중심으로 한 분트의 심리학을 펼치려고 하였지만 크게 성공하지 못했다. 왜냐하면 미국 사람들은 진 화론을 바탕으로 하는 보다 역동적이고 적응적인 기능주의 심리학을 더 선호했기 때문이다. 기능주의 심리학은 하버드 대학에 미국 최초의 심리학 연구실을 설립했던 Wi lliam J ames(l842-1910) 에 의해 시작되었 고, 시카고 대학의 An g ell(1869-1949) 과 Joh n Dewe y (1859-1952) 에 의
해 공식적으로 선언된 심리학인데, 이들은 심리학적 지식을 응용분야 특히 교육장면에 적용하는 데 많은 관심을 가졌다. 이러한 관심은 미 국의 심리학 특히 학습심리학의 발전을 가져와 심리학 연구에 쥐나 고 양이와 같은 동물을 사용하여 학습의 기본법칙을 찾는 새로운 연구들 울 자극하게 되었다. 이러한 기능주의자의 연구방법은 현대 십리학 중 학습심리학, 동물심리학, 생물심리학 등의 기초 심리학의 연구에 큰 도움을 주었다. 동물을 사용하여 학습연구에 선구적인 역할을 한 심리학자로 Edward Thomd i ke(1874-1949) 를 빼놓을 수 없다. 그는 컬럼비아 대학 에 서 고양이 를 사용하여 시 행 착오 학습 (tria l -and-error learnin g ) 이 라는 중요한 학습이론을 제시하였는데, 이 이론은 그 후 Sk i nner 에 의한 조 작적 조건화란 중요 학습이론을 탄생시킨 계기가 된다. 또한 비슷한 시기에 러시아의 생리학자 Ivan Pavlov(1849-1936) 는 개를 이용하여 조건반사란 학습이론을 제시하였다. 이 두 사람의 조건화 연구방법은 오늘날까지 인간이나 동물의 학습을 연구하는 데 중요한 방법론적 이 정표가 되었다. 2.5 현대 생물심리학 1950 년대부터 오늘에 이르기까지 40 여 년 동안 과학의 전보는 놀랄 만큼 빠른 속도로 이루어져 왔다. 20 세기 전반까지도 심리학과 생리학 은 상호간의 정보교환이나 공동연구 없이 개별적으로 발전되어 왔다. 제 2 차 세계대전 전까지만 해도 심리학자들이 신경 계통의 기능에 관심 을 가전 경우는 극소수에 불과하였고, 신경생리학자가 행동 연구에 관 심을 가진 경우도 극히 적었다• 그러나 1950 년대 이후 이런 단절의 벽 은 무너지기 시작하였다. 이때부터 미국이나 유럽 선전국의 대학원에 서 생리심리학 또는 심리생물학이란 프로그램이 등장하여 심리학과 생 리학의 공동관심과 공동연구가 시작되었으며, 이와 같은 경향은 지금 까지도 지속되고 있다. 이러한 공동 프로그램은 매우 성공적인 결과를
표 1-1 신경과학 연구와 관련된 업적으로 노벨 생리학상을 수상한 사람들 1904 Ivan Petr o vic h Pavlov : 소화의 생리학과 관련된 연구 1906 Cami llo Golgi & Santi ag o Ram6ny Caja l : 신 경 계 의 구조에 관한 연구 1932 Charles Scott Sherrin g t on & Edg ar Doug la s Adria n : 신 경 원의 기 능에 관 한 연구 1936 Henry Hallett Dale & Ott o Loewi : 시냅스의 화학적 전달물질에 관한 연구 1944 Joh n Erlange r & Herbert Spe n cer Gasser : 단일 신경원에 관한 연구 1949 Eg as Monit z : 정 신수술법 의 개 발 1961 George von Bekesy : 청 각기 제 에 관한 연구 1963 Joh n Carrew Eccles, Alan Lloy d Hodg k in & Andrew Fie ld in g Huxley : 신 경충격과 시냅스에서 일어나는 사건에 관한 연구 1967 Ragn a r Granit , Haldane Keff er Hart line & George Waid : 시 각의 기 본적 처리과정에 관한 발견 1970 Jul iu s Axelrod, Ulf Svante von Euler & Bernard Katz : 신 경 계 내 에 서 신 경과 근육 사이의 화학적 전달물질의 발견 1973 Karl von Frisc h, Konrad Lorenz & Nik o laas Tin b erge n : 동물행 동학 분 야의 발견 1977 Rog er C. L . G uil lem i n, Andrew V. Schally & Rosalyn S. Y alow : 뇌 의 펩 티드 호르몬의 발견과 성분분석 연구 1979 Allan MacLeod Cormack & Godfr ey Newbold Hounsfi el d : CAT (comp ut e r ize d axia l tom og rap h y ) 의 발전에 공헌 1981 David Hunte r Hubel, Torste n Nil s W i es el & Rog er Wolcott Sp erry : Hubel 과 Wi es el : 시각계에서 단일 신경원의 반응에 관한 연구 Spe r ry : 좌우 대뇌반구의 기능 편재화에 관한 연구 1986 Rit a Levi- M onta l cin i & Sta n ley Cohen : 신경 성장요인과 신경계의 발달 에 대한연구
낳게 되어 지금은 미국을 비롯하여 캐나다, 영국, 프랑스, 독일, 일본 그리고 오스트레일리아 등지로 확산되고 있다. 1988 년 신경과학회 (Soc iet y for Neuros ci ence) 에서 출판한 북미 지역 에서 행해지고 있는 〈 신경과학 훈련 프로그램 목록〉을 보면 이런 공동 프로그램 이 실시되고 있는 연구소가 311 개소이고, 339 개의 박사학위
프로그램이 소개되어 있다. 한편 표 1-1 에는 20 세기 이후 신경과학 연 구로 노벨상을 수상한 학자들과 그들의 주요 업적을 소개하였다. 이 표를 통해 우리는 20 세기 동안 신경과학이 어떻게 발전되어 왔는가를 참고할 수 있을 것이며, 여기에 소개한 노벨상 수상자들의 업적은 이 책의 후속되는 관련 부분에서 자세하게 언급될 것이다. 3 심신관계론 : 과학적 증거 지금까지 우리는 뇌와 행동 사이의 관계성을 논의한 여러 견해들이 어떻게 변화되어 왔는가를 살펴보았으며, 오늘날 이 문제는 신경과학 의 한 분야인 생물심리학에서 주로 다루어진다는 것을 알게 되었다. 마음과 뇌 사이의 관계를 구체화시켜 실험적으로 규명한다는 것은 매 우 어려운 일이며 어쩌면 불가능한 일인지도 모른다. 왜냐하면 많은 사람들이 사용하는 마음 (m i nd) 이라는 개념의 구체적인 실체를 밝힐 수 없으므로 이를 실험적으로 증명하는 것은 더욱 어려운 일이기 때문이 다. 따라서 뇌 또는 신경계통이라는 신체적 속성을 통해 마음의 세계 에 간접적으로 접근할 수밖에 없다. 이런 간접적 접근법은 과학적으로 결정적인 증거를 끌어낼 수는 없지만 마음과 뇌의 관계를 알아보는 데 는 많은 도움을 준다. 여기에서는 이 방법을 사용하여 마음과 뇌의 관 계를 밝힌 몇몇 연구들을 살펴보기로 하자. 동물의 뇌 피질에 미약한 전류를 홀려 자극하면, 이 전기자극에 의 해 행동변화가 일어난다는 사실이 1870 년 독일의 과학자인 Fr it sch 와 Hit z ig에 의해 처음 보고되었다. 최근에는 동물의 뇌 속에 미리 여러 개의 미세한 전극을 삽입해 두고 원격 자극조절기를 통해, 움직이고 있는 동물들의 뇌에 전류를 흘려 보낼 수 있는 방법까지 등장하게 되 었다. 이런 방법은 특히 호세 델가도(J ose Del g ado) 에 의해 많이 발전 되었는데 그에 의해 이루어전 대표적인 연구들을 살펴보자. 델가도 (1969) 는 원숭이의 뇌 피질 아래에 있는 어떤 부위에 전극을 심어놓고
이 전극을 통해 5 초 동안 전기자극을 가한 결과, 이 원숭이는 10-14 초 동안에 다음과 같은 행동을 보여주었다고 한다. 〈 자극이 종료된 후 지 금까지 행하고 있던 행동을 갑자기 멈추고, 안면 표정이 바뀌며, 머리 를 오른쪽 방향으로 돌리고, 뒷발은 땅에 짚고, 앞발은 들어올린 후 오른쪽 방향으로 회전하기 시작한다. 이어서 방의 중앙에 마련해 놓은 막대기 위에 앞다리를 울려놓고 그 위로 타고 울라갔다가 내려온 후, 으르링거리면서 위협적인 모습을 보여준다. 그 다음에는 다른 원숭이 롤 공격하고, 이어 다정한 자세로 동료 원숭이에게 접근하면서, 마지 막으로 처음에 보였던 정상행동으로 되돌아온다. 〉 이런 자극을 20,000 번이나 되풀이해도(나중에는 실험자가 지쳐서 더 이상 못할 상태까지) 원 숭이는 앞서와 거의 똑갇은 순서의 행동을 반복하였다. 또 하나의 홍미로운 뇌 자국 실험을 살펴보자. 이번에는 뇌의 어떤 부위를 전기적으로 자국하면 공격성이 억압되는 경우이다 (Del g ado, 1963). 4 마리의 원숭이를 한 우리에서 사육한다. 그 중 알리 (Al i)라고 부르는 수컷은 덩치가 크고 매우 공격적인 원숭이이고, 사라 (Sarah) 라 는 원숭이는 덩치가 큰 암컷이며, 루 (Lou) 는 덩치가 작은 수컷이고, 엘사 (Elsa) 는 덩치가 작은 암컷이다. 평소 알리는 사라와는 친하게 지 내며, 작은 수컷인 루에겐 별 관심이 없고, 작은 암컷인 엘사에겐 공 격적으로 대했다. 실험자는 알리의 뇌 속의 미상핵 (caudate nucleus) 이 란 부위에 전국을 심어놓고, 알리가 공격성을 보일 때 이곳을 자국하 면 공격성이 멈춘다는 사실을 발견했다. 이번에는 우리 속에 지렛대불 장치하여 이것을 누르면 알리의 미상핵이 자국되어 공격이 멈추어지도 록 했다. 이 우리에 사는 원숭이들도 이 지레를 누르면 알리의 공격적 행동이 멈춘다는 사실을 오랜 시행착오 끝에 알게 되었다. 그런데 이 지레를 누르는 원숭이는 항상 공격을 당하던 작은 엘사였다. 알리가 엘사를 공격할 때마다 엘사는 지렛대가 있는 곳으로 뛰어가 지레를 눌 러 알리의 공격을 멈추게 하였으며, 나중에는 엘사 스스로 알리 앞에 서 무서운 표정을 지어 알리의 화를 돋구고선 재빨리 지레를 눌러 알 리의 공격을 멈추게 하는 행동까지도 학습하게 되었다.
이번에는 인간의 뇌 피질을 전기적으로 자국했을 때 나타나는 행동 상의 변화를 알아보자. 동물의 경우와는 달리 인간의 뇌를 전기적으로 자극할 수는 없다. 그러나 뇌 피질의 손상에서 오는 간질의 발병부위 를 찾아내, 이를 제거하기 위하여 뇌 피질을 자국하는 수가 있다. Bic k fo r d, Dodg e 및 Ui hle i n (1 960) 의 보고에 의하면 간질환자가 신문을 보고 있을 때 뇌 피질의 어떤 부위를 자국하면, 자국이 가해지고 있는 동안 신문을 접었다가 편 후, 이를 왼쪽으로 돌리고 가장자리를 만지 는 따위의 행동이 자극이 계속되는 동안 되풀이된다고 한다. Heath (1 963) 는 하루 종일 참만 자려 하는 기면증 (narcole p s y) 환자를 대상으 로 뇌 피질 자극법을 적용하였다. 한 환자의 뇌 속에 전국을 고정시켜 놓고 휴대용 자극 장치를 통해 잠이 오려고 할 때마다 환자 스스로 버 튼을 누르도록 하였다. 이렇게 함으로써 이 환자는 스스로 자극 장치 를 조작하여 잠을 쫓을 수 있게 되었다. 만약 잠이 드는 순간 버튼을 누르지 못하여 잠에 빠지게 되면, 곁에 있는 친구가 환자 대신 버튼을 눌러주어 잠을 깨울 수도 있다. 이런 사실을 근거로 Hea t h(l964) 는 멀 지 않은 장래에 심한 우울증이나 통증을 가전 환자들의 고통을 없애기 위해 뇌 속에 있는 〈쾌 중추〉를 전기적으로 자극하는 방법을 사용하게 될 것이라고 예견했다. 그는 실제로 부천의 질병과 이에 따른 자신의 책임감 때문에 몹시 고통을 받고 있는 한 정신과 환자의 뇌 속 중격핵 (sep tal nucleus) 을 환자 자신도 모르는 사이에 원격 조정으로 전기자극 울 가하면, 자극을 받고 난 15 초 후 갑자기 힘을 내면서 자신의 여자 천구를 유혹하기 위한 계획을 말하기 시작했다는 사례도 보고하였다. 이처럼 뇌에 가한 전기적 자극이 이상한 감각과 운동의 출현은 물론 감정적인 변화까지도 일으킬 수 있음을 볼 때 뇌가 바로 마음을 산출 하는 곳이라고 말할 수 있는 것이다. 그러나 뇌를 자국할 때 나타나는 이런 결과는 단지 뇌 자극이 마음의 변화를 야기할 수 있다는 믿을 만 한 증거를 제시해 주는 것일 뿐 반드시 그렇다는 최종 결론을 제시하 는 것은 아니다. 비록 이런 연구결과들이 심신관계에 관한 결정적 증 거는 아니지만 심신관계에 관한 새로운 이해의 장을 열게 한 것은 틀
림없는 사실이다. 4 생물심리학의 연구방법 신경과학자들이 사용하는 연구방법들은 연구과제, 연구자 자신의 훈 련 배경 또는 관심 영역에 따라 상이하다. 신경 계통아나 뇌의 분비 계통에 이상이 있는 환자들에게 관심을 갖는 신경과학자들은 파킨슨 (Parkin s on) 병이나 알츠하이머 (Alzheim e r) 병과 같은 신경 계통이나 신경전달물질 체계에 이상이 있는 환자를 집중적으로 연구한다. 또 다 른 신경과학자들은 중풍이나 뇌세포의 감염에 의해 생기는 언어장애나 기억장애를 연구함으로써 특정 뇌 부위의 기능을 탐색하는 수도 있다. 이런 경우에 속하는 실제 연구사례들을 후속되는 장들에서 볼 수 있을 것이다. 정상적인 뇌의 기능을 알아보기 위해 뇌 장애 환자를 대상으로 연구 하는 전통은 옛날부터 있었다. 그러나 최근에 들어와 이런 연구기법은 각종 전자장비의 발달에 따라 놀라울 정도로 많이 진보되었는데, 전산 단충조영 술 (comp ut e r ize d axia l tom og rap h y : CT) 이 라는 기 법 은 바로 이런 예에 속하는 것이다. 이런 기법의 계속적인 발전과 심리학적 분 석기술이 더 발달된다면 언어, 기억, 인지와 같은 고등정신과정도 뇌 의 기능으로 설명할 수 있을 것으로 기대된다. 다음에는 현대 생물심리학 분야에 보편적으로 사용되는 연구방법, 죽 뇌 손상 기법, 뇌 기능 탐지법, 뇌 자국법 및 뇌 조직학적 기법들 울소개한다. 4.1 뇌 손상 기법 실험실에서 사육하는 쥐, 고양이 또는 행스터와 같은 동물의 신경 계통 가운데 어떤 특정 부분을 손상시키고, 그 후 나타나는 행동상의
변화를 관찰하는 연구방법이 신경과학의 중요 연구방법으로 일찍부터 사용되어 왔다. 어떤 특정 뇌 부위를 실험적으로 손상(l es i on) 시키거나 절제 (abla ti on) 한 후에도 어떤 기능들이 정상적으로 유지된다면, 이 기 능은 손상되지 않고 남아 있는 뇌의 기능에 의한다고 할 수 있지만, 손상 후에 특정 행동 기능이 상실되거나 영향을 받는다면 이 결과는 손상된 뇌의 기능 때문이라 할 수 있다. 그러나 후자의 경우 특정 행 동 변화가 뇌 손상에 따른 부작용 때문이 아니라는 점을 입증하기 위 해서는 적절한 수술통제 방법이 필요하다. 뇌 손상 기법에는 장단점이 있다. 사람의 경우는 중풍과 같은 우연 적 사건으로 일어나는 뇌 장애나 교통사고에 의한 뇌 장애 환자를 관 찰하는 수밖에 없지만, 뇌 손상법은 다양한 피험 동물을 대상으로 의 도된 특정 뇌 부위에 국한시켜 손상의 위치나 크기를 조정할 수 있는 이점이 있다. 또 한 가지 장점으로는 손상 이후 동물이 어떤 행동은 할 수 있고, 어떤 행동은 할 수 없는가를 다양한 실험을 통해 장기간 에 걸쳐 체계적으로 알아볼 수 있다는 것이다. 그러나 이 손상법에는 단점도 있다. 예컨대 손상 이후 일어날 행동 변화를 알아보기 위해 어떤 방법을 적용하며, 어떤 내용을 알아보아야 할 것인지를 결정하기 위해 많은 노력과 주의가 필요하다. 또한 뇌 손 상 이후의 기능 회복이 매우 역동적인 양상을 보인다는 점이다. 예컨 대 손상된 축색이 회복될 때 새로운 가지를 형성하기 때문에 손상 전 과는 다른 새로운 시냅스 연결이 이루어질 수 있다는 점이다. 따라서 손상 후 전혀 예기치 않았던 기능상의 변화가 일어날 수도 있다. 이런 단점에도 불구하고 뇌 손상법은 뇌의 기능을 이해하는 데 실로 엄청난 정보를 제공해 주고 있다. 뇌를 손상시키는 방법에도 여러 가지가 있다. 가장 혼한 손상방법은 전기적 손상법이다. 이 방법은 손상시킬 뇌 부위와 칙집 접촉되는 전 극의 끝부분은 노출하고, 나머지 전극 부분은 절연한 전극(흔히 가는 철선을 사용)을 뇌 속에 삽입하고 이 전극을 동해 뇌에 전류를 흘려 파 괴하는 방법이다. 뇌 속 깊은 곳에 있는 어떤 부위에 전극을 삽입하기
위해서는 먼저 동물을 마취시키고, 뇌 고정 장치 (ste re ota x ic ins tr u ment) 에 동물의 머리를 고정시킨 후, 두개골에 작은 구멍을 뚫고, 이 구멍 울 통해 전극을 삽입한다. 이때 전극의 삽입 위치와 깊이 등은 뇌 해 부도에 따라 미리 결정되어야 한다. 전극이 정확하게 삽입되면 직류전 류롤 홀려 손상시키게 되는데, 손상의 크기는 전류의 강도와 적용시간 에 따라 달라진다. 흡입법 (as pi ra ti on) 이라는 뇌 손상법도 있다. 이것은 손상시키려는 뇌 부위에 미세한 유리관이나 주사기와 갇은 피펫울 삽입하고, 이 피 펫울 통해 뇌 조직을 부드럽게 빨아들이는 방법이다. 이 방법은 뇌의 표피 부분인 신피질 부위를 손상시킬 때 주로 사용된다. 화학물질을 특정 뇌 부위에 주입하여 뇌 조직을 손상시키는 화학적 손상법 (chemi cal les i on) 도 있다. 이 방법은 어떤 신경세포체만 선택적 으로 손상시키고, 신경통로에는 아무런 영향을 끼치지 않는 특수 화학 물질을 사용하는 기법이다. 앞서의 전기적 손상이나 흡입수술법은 신 경 세포체와 신경통로 양자를 모두 손상시 키 지만 kain i c ac i d 나 ibo - ten ic a ci d 와 같은 특정 화학물질을 뇌 속에 주입하면 세포체만 선택적 으로 파괴될 뿐 축색은 영향을 받지 않는다. 화학적 손상법에서는 작 은 미세관을 전극 대신 삽입하고, 이 관을 통해 소량의 화학물질을 주 입하여 손상시킨다. 제 2 장에서 자세히 언급하겠지만, 뇌 속에는 화학적으로 상이한 수십 여 종의 신경전달물질 (neuro t rans mitt er) 이 있어서 뇌의 다양한 기능을 수행하게 한다. 이 화학물질들은 한 신경세포와 다른 신경세포가 서로 연결되는 시냅스 (s yn a p se) 라는 곳에서 분비된다. 이 물질이 분비되어 두 신경세포 사이에 상호작용이 일어나면 한 신경세포로부터 다른 신 경세포로 정보가 전달된다. 지난 몇 년 동안 몇 가지 신경독소 (neuro t ox i n) 를 사용하여 뇌 속의 신경전달물질 체계를 선택적으로 파 괴하는 기법들이 선보이기 시작했다. 예컨대 저자들(장현감, 윤병수 및 박순권, 1992) 이 6-h y drox y do p am i ne(6-0HDA) 이란 물질을 뇌 속의 편 도핵 부위 에 주입 하여 도파민 (dop am i ne ) 과 노르에 피 네 프린 (norepi ne -
p hr i ne) 과 갇은 신경전달물질을 분비하는 신경세포를 선택적으로 파괴 하고 위궤양 발생에 미치는 영향을 검토한 연구 등은 바로 이런 예에 속하는 연구이다. 이 와 유사한 신 경 독소로서 5-6-dih y d r oxy tryptar nin e (5-6-D HT) 이 란 것도 있는데, 이 물질은 세로토닌 (sero t on i n) 을 분비하는 뇌 속의 신경세포만을 선택적으로 파괴한다. 또 아세틸콜린 (ace ty lcho li ne) 이란 신경전달물질만을 선택적으로 고갈시키는 eth ylc holin e rnusta r dazki· n i um i on(AF64A) 이란 것도 있다. 이와 같이 뇌 속의 신경전달물질 체계를 선택적으로 손상시키는 기법은 특정 신경전달물질의 기능을 결 핍시킨 동물의 행동을 연구하기 위해 유용하게 사용될 수 있다. 예컨 대 동물의 뇌 속 특정 부위에 AF64A 를 주입하여 알츠하이머 병과 유 사한 증후를 발생시키는 실험적 모형도 있다 (Sm ith, 1988). 4.2 뇌 기능의 탐지법 신경세포들 사이의 정보전달에는 전기적 또는 화학적 신호가 사용된 다. 전기적 신호는 현대적인 전자장비를 통해 쉽게 측정될 수 있기 때 문에 신경 계통의 활동을 탐지하는 데 많이 이용된다. 뇌의 전기적 활 동을 탐지할 수 있는 기록전극을 두피에 부착시켜 놓고, 뇌의 전기적 활동을 증폭기로 증폭시 켜 기 록한 것 이 EEG (electr o enceph a log ram ) 이 다. EEG 기록은 기록전극 밀에 있는 수백만 개의 신경세포들이 나타 내는 전기적 활동을 복합적으로 기록한 것인데, 이 기록은 뇌의 전반 적 활동을 알아보는 데 매우 중요한 정보를 제공해 준다. EEG 는 수면 또는 각성과 같은 의식 변화에 따른 뇌의 기능 상태를 알아보기 위해 효율적으로 사용될 수 있을 뿐만 아니라 간질과 같은 뇌 장애를 진단 하는 데도 유용하게 사용될 수 있다. 뇌의 전반적 기능 상태보다는 개개의 산경세포가 특정 자극에 대해 어떻게 반응하는가를 알아보려는 경우에는 기록전극을 하나의 신경세 포 속이나 그 세포의 인점부에 두어야 한다. 이러한 〈단일 단위 (sin g le
un it s) 〉 에 대한 전기적 기록은 특히 감각 기능을 담당하는 뇌 부위에 있는 신경세포의 기능을 알아보는 데 매우 값진 것이었다(1 981 년 노벨 생리학상을 수상한 Hubel 과 W i esel 의 연구는 바로 이 기법을 사용하여 얻은 결과이다). 단일 단위기록은 일시적으로 마취시킨 동물에서 얻을 수도 있고, 뇌 속에 영구적으로 심어놓은 전극을 통해서도 기록될 수 있다. 후자의 경우는 자연스럽게 활동하고 움직이는 동물의 뇌 속에서 일어나는 신 경활동을 기록할 수 있기 때문에 매우 유용한 기법이다. 4.3 뇌 자국법 손상법과 기록법에 이은 세번째 뇌 연구방법은 특정 뇌 부위에 교류 전류를 흘려 자극하거나(직류전류를 사용하면 자국 부위가 손상됨), 신경 세포를 흥분시키는 특정 화학물질을 사용하여 자극하는 방법이다. 전 기적 활동을 기록할 때 사용하는 기록전국과 유사한 전극을 이용하여 이번에는 뇌를 전기적으로 자국할 수도 있다. 또한 어떤 화학물질을 이미 삽입된 캐눌러 (cannula : 뇌 조직 속에 꽂아 넣는 미세관으로 조직 속 에서 생성된 어떤 생화학물질을 빼내거나 의부로부터 특정 약물을 주입할 때 사용하는 기구)를 통해 뇌 속에 주입하여 자국할 수도 있다 . 그러나 이러한 자극에 의한 뇌의 활동이 정상적인 뇌의 활동을 반영하는 것인 지 아니면 비정상적인 흥분에 의해 0 일어난 부작용인지 명백하지 않을 때도 있다. 그러나 세심한 실험절차를 사용하면 이런 난점을 극복할 수 있다. 뇌의 활동을 억압하는 국소마취제나 신경안정제와 같은 약물 도 이 방법을 사용하여 뇌 속에 주입할 수 있다. 4.4 조직학적 기법 신경과학 분야에서 이루어지는 대부분의 실험들은 마지막 절차로 뇌 조직의 검색과정을 거쳐야 한다. 19 세기부터 다양한 종류의 색소나 염
료로 뇌 조직을 염색하여 세포구조를 더 명확하게 관찰할 수 있는 방 법들이 추구되어 왔다. 세포조직을 중점적으로 연구하는 신경과학의 한 분야를 조직학 (h i s t olo gy)이라 부르는데, 그 동안 다양한 조직학적 기법들이 발전되었으며, 현재도 이를 위한 많은 연구들이 진행되고 있 다. 크게 보면, 뇌 조칙의 염색법에는 세포체에 있는 단백질을 염색하 는 세포염색법 (cell s t a i n) 과 축색을 구성하는 지방질을 염색하는 섬유 염 색 법 (fibe r sta in ) 이 있 다. 19 세 기 말 이 탈리 아의 뛰 어난 해부학자 카밀로 골지 (Cami llo G olgi ) 는 골지 염색이라는 특이한 염색법을 개발했는데, 이 방법은 신경원이 질산은 (s il ver n it ra t e) 으로 염색될 수 있다는 성질을 이용한 것으로서 거의 1 세기 동안 신경 염색의 기본 방법으로 채택되어 왔다. 골지와 더불어 당대의 가장 뛰어난 스페인의 해부학자 Ramon Y. Ca j al 은 골 지 염색법을 사용하여 뇌는 개개의 독립된 신경원으로 구성되어 있다 는 사실을 발견하였다. 이 발견은 뇌는 조직의 망 (re ti culum of tissu e) 으로 구성된다는 골지의 견해를 부정하는 아이러니를 낳게 되었다. 이 처럼 개개 신경원이 어느 정도의 공간을 사이에 두고 서로 분리되어 있다는 사실은 뇌의 구조와 기능을 이해하는 데 매우 중요한 것이다. 또 하나의 홍미로운 방법은 〈바로 조금 전에 왕성하게 활동했던 뇌 세포가 어느 것이었나〉를 확인하는 방법이다. 신체를 구성하는 세포들 과 마찬가지로 뇌세포도 활동할 때 에너지를 소모한다. 뇌가 활동할 때 사용하는 에너지원은 포도당(g lucose : 포도당이 혈액 내에 있을 때 이 를 혈당이라고도 한다)이다. 활동을 많이 하는 세포일수록 뇌 속에 순 환하는 혈액으로부터 보다 많은 포도당을 공급받게 된다. 2- 디옥시글 루코스 (2-deox yg lucose : 2-DG) 라는 화합물은 구조상으로 포도당과 매 우 흡사하고 혈액으로부터 세포 속으로 잘 흡수될 수 있지만, 뇌세포 는 이 물질을 대사시킬 수 없다. 만약 2-DG 를 동물에게 주사하면 활 동을 많이 하는 세포일수록 포도당과 함께 2-DG 를 많이 흡수하게 된 다. 그러나 뇌세포의 에너지가 되는 포도당과는 달리 2-DG 는 세포 내 에 들어와도 이용되지 못하고 원래 상태로 머물게 된다. 만약 동물에
게 2-DG 에 방사성 탄소동위 원소 (radio a cti ve carbon ato m ) 를 붙여 주사 한 후, 이 동물을 희생시켜 뇌 조직 절편을 작성하고, 여기에 X 선을 두과시켜 촬영하면 2-DG 분자를 흡수한 뇌세포를 확인할 수 있다. 즉 활동이 많았던 세포일수록 2-DG 분자를 더 많이 흡수하여 방사선 사 진에서는 더 검게 나타난다. 이러한 기법은 Louis S okolo ff(1 977) 에 의해 개발된 것으로 바로 조금 전에 가장 왕성하게 활동했던 뇌세포를 찾아내는 데 효과적인 방법이다. 앞서 언급했던 조직학적 기법들은 모두 죽은 조직을 대상으로 검색 하는 방법이다. 그러나 최근에는 신경과 환자들을 대상으로 지금 막 활동하고 있는 뇌의 기능을 국소적으로 확인하는· 방법까지 등장하였 다. 죽, 앞서 본 2-DG 기법의 한 변형으로서 양전자방사단층촬영 (po sitr o n emi ss io n tom og rap h y : PET) 이 란 기 법 이 Raic h le (l983) 에 의 해 개발되었다. 이 방법은 환자들에게 미량의 방사성 포도당을 주사하 고(유해한 영향을 주지 않을 정도의 소량), 이 방사성 물질로부터 방출되 는 양전자를 이용하여 환자의 뇌 활동을 컵퓨터로 촬영하는 방법으로 오늘날 현대화된 큰 병원에서 채택하는 일반화된 방법이다. 가장 최 근에 개 발된 자기 공명 영 상법 (magn e tic resonance ima gi ng : MRI) 이라는 기법도 활동중인 뇌 조직을 관찰하는 방법이다. 이 기법 은 뇌를 강한 자장에 노출시켜 뇌 속의 수소원자를 일정한 방향으로 회전시킨 후 여기에 전자파를 투과시켜 수소원자가 방출하는 전자파의 에너지를 탐지함으로써 뇌의 활동을 영상화시킨다. 이 기법을 사용하 면 아주 해상도가 높은 선명한 영상을 얻을 수 있다.
제 2 장 신경원과 시냅스 19 세기까지는 그 당시 최고 성능의 현미경을 갖고서도 뇌 조직의 미 세한 면들을 관찰할 수 없었다. 당시 과학자들은 한 신경세포와 다른 신경세포 사이에 매우 가늘고 긴 한 가닥의 섬유가 있어 이 섬유가 두 신경 사이를 직접 연결하고 있기 때문에 각각의 신경세포가 독립적으 로 존재 하는 것 이 아니 라는 생 각(예 컨대 Cam illo Golgi ) 과, 이와는 달 리 각각의 신경세포들은 개개의 세포들 사이에 좁은 간격을 두고 서로 떨어져 독립적으로 존재한다는 주장 (Ramm6n Y. Ca j al) 으로 서로 맞서 있었다. 당시엔 누구도 이 두 입장 가운데 어느 하나의 입장을 지지해 줄 수 있는 결정적 증거를 제시할 수 없어서 이 두 사람은 1906 년 노벨 생리 학상을 공동 수상하게 되었다. 그러나 1950 년대에 들어와 전자현미경 이 보급되면서 뇌는 독립적인 각각의 신경세포(하나의 신경세포를 신경 원 neuron 이라고 함)로 구성된다는 사실이 밝혀짐으로써 Ca j al 의 주장이 지지받게 되어 과거의 논쟁이 종식되었다. 모든 신경원은 전기적 신호를 발생시켜 이룰 전달하는 능력과 다론 신경원의 활동을 바꿔놓을 수 있는 화학적 물질을 분비하는 능력을 갖 고 있다. 이러한 전기적 신호를 신경충격 (nerve imp ulse) 이라고 하는 데, 이 신경충격은 신경원의 세포막을 넘나드는 이온의 운동에 의해 발생 된다. 신경충격은 신경 전달물질 (neurotr an smi tters ) 이 라는 화학적
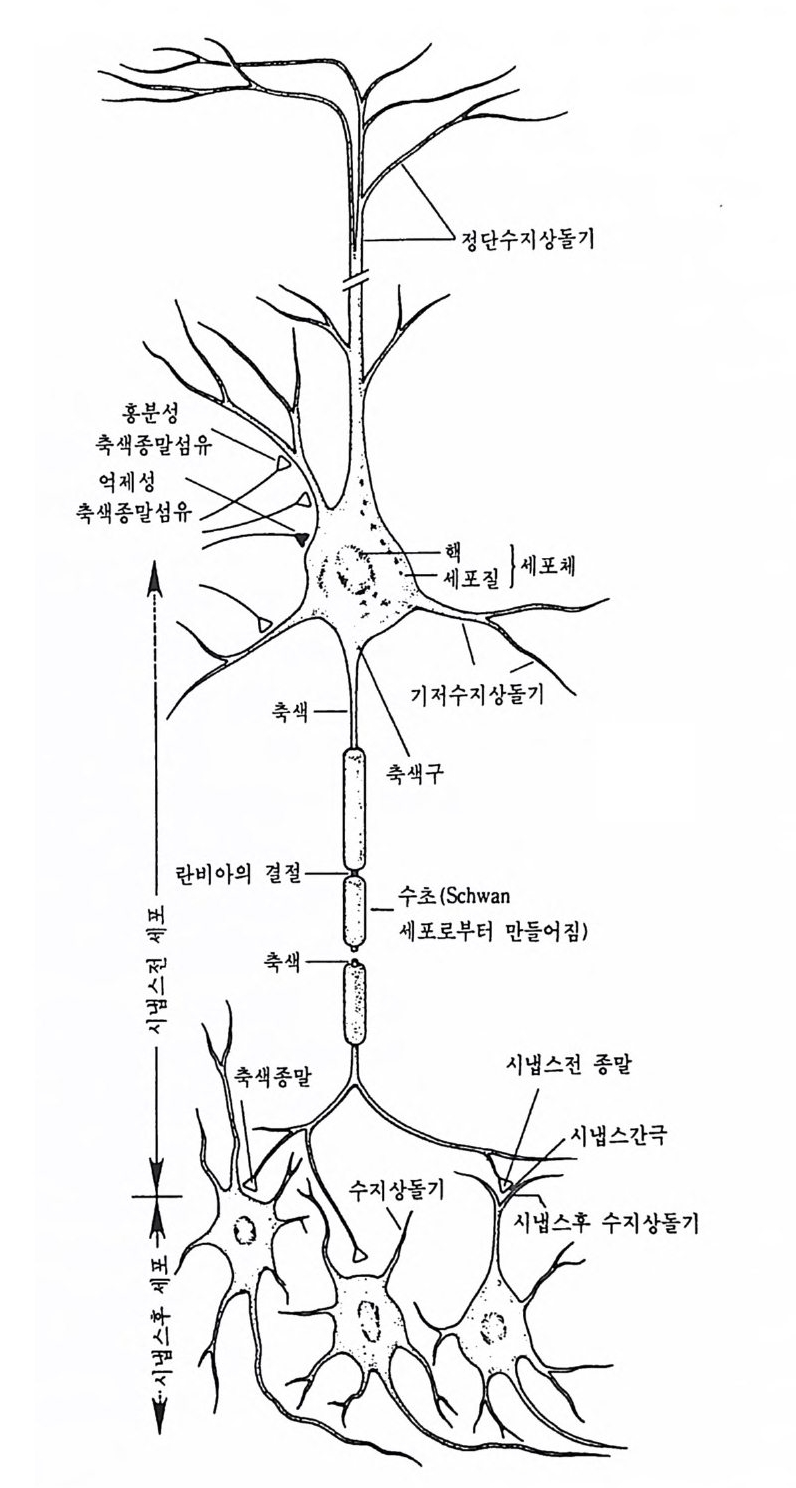
물질에 의해 한 신경원으로부터 인접의 다른 신경원으로 이동한다. 우 리는 이들 신경원들이 어떻게 기능적으로 통합될 수 있는가를 알아보 기 위해 개개 신경원의 특성을 먼저 알아보고 이어 개개 신경원들 사 이의 정보소통 현상을 살펴봄으로써 뇌의 기능을 보다 자세하게 알아 볼 것이다. 1 신경원의 구조 뇌는 여러 가지 상이한 종류의 세포들로 구성되지만, 우리 뇌 속에 서 정보를 수용하여 처리하고 전달하는 기능을 수행하는 세포는 신경 세 포 (nerve cell ) 또는 신 경 원 (neuron) 이 다. 이 장에 서 는 주로 뇌 의 정 보처리 능력, 죽 신경원의 기능에 관해 먼저 살펴보려고 한다. 신경원 은 그 종류에 따라 크기나 모양이 다르지만 공통점도 많다. 포유류의 경우 대부분의 신경원은 세포체의 지름이 10-lOOµm(lµm=10-6m) 정 도로 매우 작은 것이 특징이지만 세포체로부터 멀리 떨어진 곳까지 뻗 어가는 축색은 길이가 lm 이상이나 되는 긴 것도 있다. 인간의 뇌 속에 얼마나 많은 신경원이 존재하는지는 아직 잘 모르지 만 적게는 수백억 개에서 많게는 천억 개 정도까지 추정한다. 신경원 이 정보를 처리, 전달, 저장하는 기능은 다음과 같은 두 가지 특성에 의한다. 즉 첫째는 전기적 신호를 야기하고 전달하는 기능과 둘째는 그림 2-1 전형적인 척수동물의 신경원 개요 이 그림에는 신경원의 주요 구조와 다른 세포와 접촉하고 있는 부위를 나타내 고 있다. 세포체는 핵과 세포질로 구성되며, 수지상돌기 (지저 및 정단)와 축 색돌기를 갖는다. 축색돌기는 길이가 다양한데 lm 이상 되는 긴 것도 있다. 축색돌기가 시작되는 곳을 축색구라 하는데 이곳에서 활동전위가 발생된다. 축색돌기는 주로 수초라는 지질막으로 덮여 있는데 란비아 결절에 의해 군데 군데 찰려 있다. 한 신경원의 축색 분자들이 다른 신경원의 수지상돌기나 세 포체와 접촉하는데, 이 접촉부위를 시넵스라 하며 한 개의 신경원이 약 1,000 개의 다른 신경원들과 시넵스한다.
 흥분
흥분
신경전달물질을 생성하고 분비하는 기능이다. 그립 2-1 에서는 척추동물의 전형적인 신경원의 모습과 다른 세포와 시냅스하고 있는 모양을 예시한 것이다(무척추동물의 경우는 신경원의 수지상돌기가 세포체와 연결되는 것보다 축색과 직접 연결되는 경우가 더 많다). 신경원은 몇 가지 상이한 부분으로 구성된다 . 세포체는 핵 (nucleus) 과 세포가 필요로 하는 물질을 생성하는 기구들로 구성된다. 이 기구들은 핵 주변에 있는 소포체 (endop la smi c re tic ulum) 란 곳에 존 재한다. 소포체는 지질과 단백질로 이루어진 막으로 되어 있다. 소포 체와 관련된 구조로서 리보솜 (r i bosome) 이란 것이 있는데 이것은 단백 질과 리보핵산 (r i bonucle ic aci d : RNA) 의 복합체로 이루어지며 단백질 합성에 매우 중요한 곳이다. 세포체로부터 뻗어나오는 돌기에는 축색 또는 축색돌기 (axon) 와 수 지상돌기 (dendr it e) 가 있다. 수지상돌기와 세포체는 신경원으로 들어오 는 정보를 받아들이는 수용기 역할을 한다. 모든 신경원의 수지상돌기 에는 수지상가시 (dendriti c s pi ne) 라는 작은 돌기가 있는데 이것은 수지 상돌기의 변형이다. 축색은 신경원의 출력부 (ou tp u t)에 해당되며 축색 의 종말부 쪽으로 신경충격을 전달한다. 하나의 신경원은 축색종말 부에서 다른 신경원과 기능적으로 연락되는데, 이 부위를 시냅스 (s yn a p se) 라 한다. 또한 축색은 여러 개의 곁가지를 가지며, 축색종말 부의 경우 일반적으로 수백 개의 곁가지가 있다. 미 소관 (mi cro tu b ule) , 신 경 세 사 (neurofi lam ent) 또는 미 세 사 (mi cro fi la- men t)라는 구조들도 수지상돌기 및 축색돌기와 함께 모든 신경원에서 발견되는 중요한 구조들이다. 이 구조들은 세포의 발달을 유도하고, 세포를 구조적으로 지지하고 세포 내의 산물들을 이동시키는 기능을 한다. 이들 구조들은 크기가 서로 다른데 미소관은 지름이 20-30µm 로 서 그 중 가장 크며, 신경세사는 lOµm 정도이고, 미세사는 lOµm 이 하로 가장 작다. 신경원 내에는 또 다른 주요 구조물로 미토콘드리아 (mit ochon dri a) 가 있는데, 이것은 세포에 에너지를 공급해 주는 데 중 요한 역할울 하며, 그 밖에 골지체 (Gol gi bod y)란 구조는 세포의 대사
활동과 단백질 합성에 중요한 역할을 한다. 축색의 종말부에는 작은 주머니들이 많이 있는데, 이곳에는 신경전달물질이 저장되어 있다. 2 신경교세포 뇌 속에 가장 많이 분포되어 있는 세포는 신경원이 아니라 신경교세 포 (glial cell : glia) 이 다. 교 (룡, glia) 라는 말은 그리 스어 의 아교 (glu e) 라는 말에서 유래된 것으로 개개의 신경원을 붙잡아 매는 성질을 가진 다. 이 신경교세포의 크기는 신경세포의 1/10 정도이나 수적으로는 약 10 배 정도로 수천억 개가 될 것으로 추정된다. 이 세포들은 그 자체로 는 신경충격을 생성시키지 못하지만 신경원들이 고유의 기능을 수행하 는 데 도움을 주며, 뇌 조칙이 손상되었을 때 이를 회복시키는 데도 매우 중요한 기능을 한다. 신경교세포도 형태에 따라서 핍돌기 신경교 세 포 (olig o dendrog lia) , 성 상 신 경 교세 포 (astr o cyt e) , 미 세 신 경교세포 (mi cro g lia) 및 방사 신경교세포 (rad i al gli a) 로 나누어진다. 신경교세포의 중요한 기능 가운데 하나는 신경원의 축색을 둘러싸는 수초 (my el in s heath ) 를 만드는 것 인데 , 이 과정을 수초화라고 한다. 수 초화는 신경충격을 보다 신속하게 전달하는 데 주요 역할을 하며, 축 색들 사이의 전기적 방해를 방지하는 역할도 한다. 말초신경계에서는 슈반 (Schwan) 세포라는 신경교세포가 이 기능을 담당한다. 신경 계통 이 성장하면서 축색이 자라게 되면, 하나의 슈반 세포가 막 자라나는 축색의 한 결절을 감싸게 된다. 이렇게 축색이 계속 자라는 동안 각각 의 슈반 세포가 각 축색의 결절을 감싸게 되므로 하나의 축색은 몇 개 의 결절로 이어지는 듯한 모습을 보이게 된다. 이러한 축색의 결절을 랑비에 결절 (nodes of Ranv i er) 이라고 하는데, 이것은 프랑스의 해부학 자 Ranv i er(1835-1922) 에 의해 처음 명명되었으며 뇌, 척수 또는 말 초신경계에서 두루 관찰된다. 말초에서와는 달리 중추신경 계통에서는 슈반 세포와 유사한 핍돌기 신경교세포가 뇌와 척수에서 수초를 만든다.
인간의 뇌 속에서 수초화가 완성되는 데는 몇 년이라는 긴 시간이 소요된다. 이러한 수초화 과정과 지적 성장 또는 운동기능의 성숙 사 이에는 매우 밀접한 관련이 있다. 수초의 주성분이 지방질이기 때문에 수초화된 신경섬유는 표면이 흰색으로 보인다. 따라서 뇌나 척수에서 수초화된 신경섬유를 백질 (wh it e ma tt er) 이라 부르고, 수초화가 이루어 지 지 않은 세포체나 수지 상돌기 를 회 백 질 (gray matt er ) 이 라고 한다. 성상 신경교세포와 미세 신경교세포는 신경계의 손상이나 감염에 의 해 생성된 찌꺼기를 제거하는 청소 세포들이다. 또한 성상 신경교세포 는 섬유성 성상 신경교세포(fi brous as t roc yt es) 와 원형질성 성상 신경교 세 포 (pro to p l a smi c astr o cyt es ) 로 나누어 지 지 만 이 들 세 포들의 기 능적 차이는 아직까지 밝혀져 있지 않다. 신경원과는 달리 인간 뇌의 신경교세포는 출생 후에도 계속 분열되 어 새로운 신경교세포를 만든다. 뇌 조직의 손상은 청소 세포들로 하 여금 분열하도록 자국하게 하고 분열된 세포는 신경원이 죽거나 제거 된 빈 자리를 채우게 되어 신경교 반혼(gli a scar) 이 생기게 된다. 이러 한 흔적은 신경조직의 손상부위를 확인할 수 있는 유일한 지표가 된 다. 성상 신경교세포는 신경원의 생존과 성장 및 분화에 필요한 물질 을 생성하는데, 이러한 물질을 신경자극 요소 (neuro t ro pi c fa c t ors) 라 부른다. 또한 성상 신경교세포는 신경원을 둘러싸고 있는 체액의 화학 적 요소를 통제하도록 도와주며, 체액의 화학적 영향울 받지 않도록 신경원을 격리시킴으로써 시냅스의 개별성도 보장해 준다. 신경교세포 가 신경원의 생존을 위한 신진대사에 어떤 역할을 할 것이라는 추측은 오래전부터 제시되었는데도 불구하고 아직까지 이러한 가설을 입증하 기 위 한 직 접 적 인 증거 는 미 비 한 상태 이 다 (Ku ffl er, Nic h olls, & Mart in, 1984). 방사 신경교세포는 태아의 발달 초기단계에서 나타나는 특수한 신경 교세포이다. 신경원이 생성된 장소에서 분화될 장소로 이동할 때는 이 신경교세포에 따라 붙어 이동하게 되므로 이 신경교세포는 신경원의 이동에서 안내자 역할을 한다.
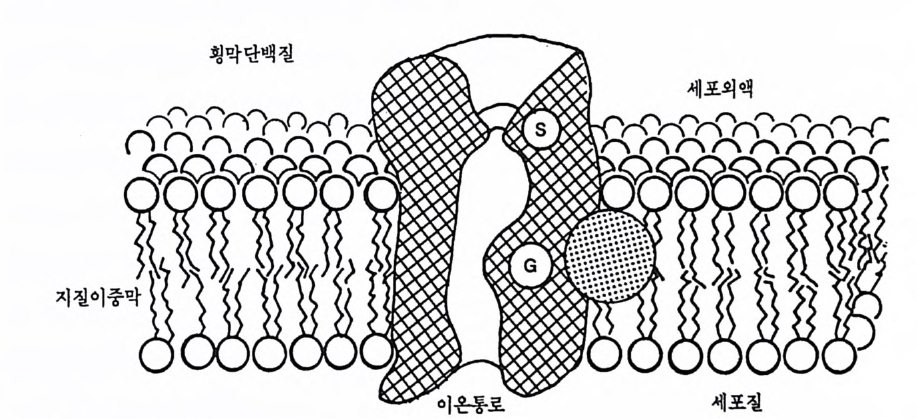
3 신경세포막 : 유동성 모자이크막 모든 정신활동의 바탕이 되는 신경원의 전기적 신호는 어떻게 발생 될까? 이 문제를 알아보기 위해서 신경원 막의 구조와 성질부터 알아 보자. 신경원의 막도 기본적으로는 일반 세포의 막과 유사하지만 몇 가지 점에서 특이하다. 동물 세포의 막은 지방질의 이중충으로 구성되 는데, 이를 지질이중충(lipi d b il a y er) 이라 한다. 이 지질이중충에는 다 양한 종류의 단백질이 곳곳에 부착되어 있으며, 이들 단백질의 배열방 식에 따라 해당충 부위의 생물학적 특성도 달라진다. 이 지질분자들의 결합은 느슨하기 때문에 막은 유동성을 띤다. 지질아중충은 여기에 결 합된 단백질과 함께 유동성 모자이크 모델(fl u i d mosaic m odel) 로 간주 된다. 그립 2-2 에는 신경원 세포막의 유동성 모자이크 모델을 그림으 로 나타내고 있다. 실제의 단백질은 이 그림에서 나타난 것보다 훨씬 더 정교하고 분명한 모습을 가진다.
 지질이횡막단백冒질 〈〈冒:'틀세포의액
지질이횡막단백冒질 〈〈冒:'틀세포의액
그림 2-2 지질이중막 구조 지질용해성 꼬리 부분은 꼬리들끼리 서로 마주보고 있으며, 물분자에 친화성 울 갖는 머리 부분은 세포의액과 세포질 쪽으로 머리를 돌리고 있어 이중막을 이루고 있다. 이 막 사이에 횡막단백질이 박혀 있으며, 그 사이에 이온통로가 있다. 이온통로는 특정이온을 선별, 통과시키는 S 부분과 이 이온통로 전체를 열고 닫는 G 라는 문이 있다. 점을 찍어놓은 원부분이 이온통로의 개폐를 통재 한다.
뇌와 같이 수분이 많은 매체 속에서 지방질 성분은 마치 기름방울을 물에 떨어뜨리면 기름방울이 구형을 이루듯이 스스로 응집된다. 각 지 질 분자의 한쪽 끝인 머리 부분은 물 분자에 대한 친화력을 보이는 반 면 반대쪽의 꼬리 부분은 물 분자에 반발한다. 이러한 물과 지질 분자 사이의 분자적 상호작용 때문에 물 분자가 많이 있는 쪽으로 지질 분 자의 머리 부분이 모여 줄을 서게 된다. 그러므로 지질 분자의 머리 부분은 물과 접촉할 수 있는 신경세포막의 안쪽 면과 바깥쪽 면을 형 성하고, 물 분자에 대한 친화성이 없는 꼬리 부분은 서로 마주보게 되 어 신경세포막의 내면을 형성하여 지질이중충이 구성된다. 이러한 막 의 지질구성 때문에 물 분자나 기타 물에 잘 녹는 수용성 물질들은 막 의 지질 부분을 통과하지 못한다. 세포막을 가로지르는 횡막 단백질 (tra nsmembrane p ro t e i n) 의 주요 기능 가운데 하나는 어떤 물질들을 동 과시키기 위한 이온통로(i on channel) 구실을 한다는 점이다. 막은 신 경원 전체에 걸쳐 획일적으로 구성되어 있지는 않다. 막의 특수 부분, 예컨대 신경전달물질을 방출할 수 있도록 특수화되어 있는 축색종말부 와 다른 신경원으로부터 들어오는 흥분을 받아들이는 수용기 부위인 세포체나 수지상돌기에는 여러 종류의 단백질이 막에 삽입되어 있다. 4 신경원의 전기신호 이제 앞서의 물음, 즉 어떻게 신경원이 행동이나 정신활동의 근본이 되는 신경충격을 야기하는가에 관해 알아보자. 이 물음에 대한 해답을 알기 위해서는 신경세포의 막에서 전기적 신호가 발생되고 전달되는 과정과 이 전기적 신호가 축색종말 부위에서 인접하는 다른 신경세포 로 전달되어 영향력을 미치게 되는 시냅스 전달 과정을 이해해야 한 다. 뇌 속에는 여러 종류의 이온돌이 있지만 신경원의 전기적 신호를 이해하기 위해서는 나트륨 (Na+ : sodiu m ), 칼륨 (K+ : pot a s siu m ), 칼슘 (Ca2+ : calciu m ) 및 염소 (Cl-: chlorid e ) 와 갇은 4 가지 이 온의 특성 을
표 2-1 신경 활동에 관여하는 주요 이온들의 기능 이온 기능 Na+ 신경충격의 발생 (홍분성 시냅스후 전위)에 관여 K+ 신경충격이 발생하는 동안 안정전위의 회복에 관여 : 억제성 시냅스후 전위와도 관련 c1- 억제성 시냅스후 전위와 관련 Ca2+ 시냅스에서 신경전달물질의 방출에 필요 : 홍분성 시냅스후 전위와 관련
아는 것이 중요하다. 표 2-1 에는 이들 4 가지 이온의 기능을 간단하게 나타내었다. 이 4 가지 무기이온은 물과 같은 용액 속에서 분자가 분해 되어 생긴다. 예를 들어 영화나트륨 (NaCl), 죽 소금은 물 속에서 용 해되어 나트륨과 염소라는 원자로 분해되어 전기적 성질을 띤 전하이 온이 된다. 이렇게 되면 염소 이온은 음전기를 띠게 되고 (Cl-), 나트 뮴 이온은 양전기를 띠게 된다 (Na 기. 편의상 양전기를 띠는 이온을 양이온 (ca ti on) 이라 부르는데, 그 이유는 이 이온들이 건전지의 음극 (cath o d) 쪽으로 끌리기 때문이며, 음전기를 띠는 이온을 음이온 (an i on) 이라고 하는데 이것은 양극 쪽으로 끌린다. Na+, K+, Ca2+ 그 리고 Cl- 은 세포막의 내면과 의면에 상이하게 분포되어 있다. 이온은 전하를 띠기 때문에 하전된 전기적 성질에 따라 서로 끌어당기기도 하 고 밀어내기도 한다. 즉, 같은 전기적 신호를 가진 이온들끼리는 서로 밀어내고, 다른 신호를 가진 이온들은 서로 끌어당긴다. 신경원이 전기적 신호를 일으킨다는 사실은 지난 150 여 년 동안 수 많은 실험을 통해 밝혀진 것이다. 신경원의 전기적 활동을 밝힌 연구 결과들은 신경원이 비교적 크고 조작이 용이한 몇몇 무척추동물(특히 오징어의 경우처럼 축색의 지름이 1mm 나 되는 거대신경원 gian t neuron 을 대상으로 연구함)을 연구 대상으로 한 것이었다. 영국 케임브리지 대학 의 Andrew Hod gk i n 과 Alan Huxle y는 오칭 어 의 축색 을 대 상으로 신 경원의 전기생리적 현상을 최초로 연구한 공로로 1963 년 노벨 생리학
상을 수상하였다. 그 후의 많은 연구들에서도 신경원의 전기적 활동을 연구하기 위해서 오칭어의 신경원을 사용했으며, 그 결과 오징어의 신 경원에서 이루어지는 현상이 인간과 같은 포유류의 신경원에서 볼 수 있는 현상과 극히 유사하다는 사실도 입증되었다. 4.1 안정전위 오칭어의 거대축색에 기록전극을 삽입하여, 막을 사이에 두고 일어 나는 전위차를 기록할 수 있다. 신경원이 쉬고 있는 동안 기록된 전위 를 안정 전위 (restin g pot e n ti al) 라 하는데 , 이 때 안정 이 란 말은 신경 원이 신경충격을 발생시키지 않는다는 뜻이지 아무런 활동도 하지 않고 완 전히 쉬고 있다는 뜻은 아니다. 신경원은 안정전위를 발생시키고 이룰 유지하기 위해 끊임없이 신전대사 활동에 필요한 에너지를 소모하고 있다. 오칭어와 같은 연체동물로부터 원숭이와 같은 포유동물에 이르기까 지 대부분 동물의 신경원을 대상으로 한 연구들에 따르면 안정전위에 서는 세포의 내면이 -40mV 에서 —9 0mV 정도의 음전기를 띤다고 한 다. 앞서 언급한 오칭어의 거대축색을 대상으로 한 연구결과들을 살펴 보면 오칭어의 신경원에서 기록되는 안정전위는 _60mV 에서 _70mV 인데 대부분의 교과서에서는 一 70mV 로 기록되어 있다. 안정전위는 축전지의 음극과 양극 사이에 존재하는 전위차와 유사하다. 이 두 전 극 사이를 연결하면 전류가 흐르는 것과 같이 만약 세포막의 안쪽과 바깥쪽을 연결하면 전류가 흐른다. 비록 신경원에서는 세포막의 안팎 이 어떤 전선에 의해 연결되어 있지는 않지만, 이 둘 사이의 전기적 흐름을 이해한다는 것은 매우 중요한 것이다. 신경세포막을 가로질러 흐르는 전류를 이온성 전류(i on ic curren t)라 고 한다. 이 이온성 전류는 전자의 흐름에서 생기는 일반적인 전류와 는 달리 신경 세포막의 이온통로(i on ic channel) 가 열릴 때 이 통로를 통 한 이온의 이동에 의해 발생되는 전류이다. 이런 전류의 흐름을 신경
표 2-2 오징어의 거대 신경원에서 측정한 이온농도 이온 세포 내 농도 세포 외 농도 비율(내/의) 평형전위 Na+ 50mM 440mM 1 : 9 +55mV K+ 400mM 20mM 20 : 1 -75mV c1- 52mM 560mM 1 : 11 -60mV (A-)• 385mM *A 는 아미노산과 갇은 다양한 종류의 유기 음이온
충격 (nerve imp ul se) 또는 활동전위 (acti on p o t en ti al) 라 한다. 막을 사 이에 두고 전위차가 OmV 에 이를 때까지 줄어드는 현상을 탈분국화 (de p olar i za ti on) 라고 하며, 이와는 다르게 전위차가 안정전위보다 더 커지는 현상을 과분극화 (h yp e rp ola ri za ti on) 라고 한다. 따라서 막전위가 -60mV 로부터 -45mV 로 바뀌는 것은 탈분국화이고, -70mV 로부 터 -90mV 로 막전위가 바뀌는 것은 과분국화이다. 탈분극화와 과분 극화는 신경원의 활동 중 가장 보편적인 것이다. 안정전위란 막을 통과한 이온들에 의해 나타나는 화학적, 전기적 힘 의 균형을 의미한다. 표 2-2 에는 오칭어의 거대신경원의 막 내면과 의 면에 존재하는 주요 이온들의 농도가 나타나 있다. 표에서 A.:. 라는 기 호는 아미노산과 같은 다양한 종류의 유기 음이온 (or gani c anion ) 들을 총합적으로 표시한 것으로 어떤 음이온은 아미노산이고, 또 어떤 것은 한 개 이상의 전자를 갖는 단백질이다. 여기서 모든 종류의 음이온 하 나하나를 확인하는 것은 중요한 일이 아니다. 보다 중요한 것은 음전 기를 띤 음이온돌이, 세포의 안쪽에서부터 바깥쪽으로 이동하려고 하 지만 크기가 너무 크기 때문에 세포막이 파열되지 않는 한 이동이 불 가능하다는 것이다. k 를 제의한 나머지 Na+, K\ 및 Cl- 이온들은 모두 크기가 작은 무기 이온둘이다.
4.2 이온통로 안정 상태에서 K+ 은 신경원의 의부보다 신경원 내부에 약 20 배나 더 높은 농도로 분포되어 있다(표 2-2). 이와는 달리 Na+ 는 내부보다 의부에 약 9 배 정도 더 많이 분포되어 있다. 이처럼 Na+ 와 K+ 이온 의 농도상의 불균형으로 인해 신경원의 전기적 신호가 발생된다. 이온 은 지질이중충을 칙접 통과할 수는 없지만, 어떤 조건 아래서는 세포 막에 있는 특수 이온통로를 통해 통과할 수 있다(그림 2-2 참고). 이온통로는 이온의 종류에 따라 통과를 허용할 수도 있고 ' 허용하지 않을 수도 있는 선택성을 갖는다. 축색막의 경우 Na+ 나 K+ 중 어느 하나만을 선택적으로 통과시키는 통로는 있지만, 이 두 이온 모두를 통과시키는 통로는 없다. 그러나 수지상돌기에 있는 이온통로는 선택 적이지 않아서 Na+ 와 K+ 양자를 모두 통과시킨다. 이온통로의 구성 (confi gurat i on ) 은 어느 한 가지 양상으로 고정 되 어 있는 것이 아니다. 죽 이온통로를 구성하는 단백질의 모양이 바뀔 수 도 있기 때문에, 이에 따라 통로의 작용도 바뀔 수가 있다. 더구나 이 런 구조적 변화는 순식간 (0.001 초 이내)에 일어날 수 있다. 대부분의 경우 통로가 열리거나 닫히는 두 가지 방식이 있지만 N 감의 통로는 불활성화(in ac ti va ti on) 라는 제 3 의 방식이 있다. 통로가 열리면 통로가 닫혀 있을 때보다 이온의 통과가 보다 쉽고 보다 빠르게 이루어전다. N 갑 통로의 경우 통로의 폐쇄와 통로의 불활성화 사이에는 약간의 차 이가 있다. 죽 불활성화의 경우는 폐쇄화에 비해 Na+ 에 대한 장벽이 더 커진다. 이온통로의 구성은 막을 사이에 두고 발생하는 전위의 변화에 의해 조정될 수도 있고, 통로에 있는 단백질과 상호작용하는 어떤 화학물질 에 의해서도 조정될 수 있다. 이온통로는 세포막 부위에 따라 밀집의 정도가 다르다. 일반적으로 전압식 통로는 주로 축색 부위에 밀집해 있고, 화학적 통로는 세포체와 수지상돌기에 밀집해 있다. 이온통로는 여러 가지 화학물질에 대한 반응성에서도 차이가 있다.
어떤 약물은 축색에 있는 Na + 의 전압식 통로는 차단하지만, Na + 의 화학적 통로에는 별 다른 영향을 끼치지 않아 N 갑 이온을 통과할 수 있게 해준다. 예를 들어 복어의 내장에 있는 어떤 독물은 축색막에 있 는 Na+ 의 전압식 통로와 결합하여 이 통로를 차단하지만, 이 이온의 화학적 통로에는 별 영향을 미치지 않아 N 갑의 전입을 가로막지 않는다. 이온통로는 세포막 전체에 산포되어 있지만 어떤 곳에는 많이 분포 되어 있고 어떤 곳에는 거의 없다. 수초화된 축색의 이온통로는 주로 랑비에 결절 부분에 분포되어 있고 결절과 결절 사이에는 드물다. 이 온통로는 크기가 너무 작아 현대의 기법으로서는 직접 관찰할 수 없기 때문에 이 통로의 밀도를 알아보기 위해 간접적인 방법을 사용한다. 예컨대 복어에서 발견되는 데트로도독신(t e t rodo t ox i n: TTX) 은 Na+ 통로의 바깥쪽 면을 이루는 단백질 부위에 잘 결합한다는 사실이 발견 되었는데, 오징어의 거대축색 표면에는 µm2 당 평균 100-600 개의 통로 가 있다는 사실이 밝혀졌다 (She p herd, 1983). 한국 사람이나 일본 사람 들은 복어 요리를 좋아하여, 내장이 제거되지 않았거나 불완전하게 제 거된 복어 요리를 먹고 생명을 잃는 경우가 있다. 그 이유는 복어 내 장 속에 있는 TTX 가 N 삼 이온통로를 차단하여 신경통로(그 중에는 호흡조절을 하는 신경통로도 포함)가 마비되기 때문이다. 4.3 전기적 또는 화학적 기울기 안정전위는 막을 통과하는 이온들에 의해 생기는 전기적, 화학적 힘 에 의해 발생한다. 여기서 두 가지 중요한 이온, 즉 Na+ 와 K+o J] 대 해 살펴보기로 하자. 안정 상태에서는 축색에 있는 대부분의 N 갑 통 로가 닫혀 있지만 간혹 N 강 이온이 축색 안으로 들어갈 수 있도록 통 로가 열리는 경우도 있다. 어떻게 해서 이 이온이 축색 안으로 들어갈 수 있을까? 이것은 막을 사이에 두고 존재하는 전기적 또는 화학적 기울기로부터 생기는 두 가지의 물리적 힘이 N 갑 이온으로 하여금 막 의 내면으로 들어가도록 하는 압력으로 작용하기 때문이다.
이 중 첫번째 힘은 전기적인 것으로, 세포막의 안쪽이 음전기를 띠 므로 Na+ 와 갇은 양이온을 세포 내로 끌어들이려는 전기적 기울기 (electr ica l grad ie n t) 가 막을 사이 에 두고 생 기 게 된다. 따라서 만약 N 갑 통로 가운데 어느 하나가 열리게 되면 막 바깥에 있는 N 갑 이온 들은 이러한 전기적 힘에 의해 안쪽으로 끌리게 된다. 두번째 힘은 세 포막의 안쪽과 바깥쪽에 분포된 이 온들의 농도기 울기 (concentr a ti on grad ie n t) 때문에 생긴다. 용액 속에 있는 이온둘은 이동아 자유로워 고농도 쪽에서 저농도 쪽으로 확산되어 갈 수 있으므로 궁극적으로 전 체 용액 속의 이온 농도가 균일하게 유지되는 평형상태에 이를 때까지 이동이 계속된다. K+ 이온의 경우는 N 강 이온의 경우와는 다소 차이가 있다. K+ 이 온의 통로는 신경원이 안정 상태에 있을 때도 열려 있다. 그래서 안정 상태에서 K+ 이온은 Na+ 에 비해 막 두과성이 100 배 정도 더 높다. K+ 이온의 농도는 바깥쪽보다는 안쪽이 더 높기 때문에 농도기울기의 힘은 K+ 이온을 세포막 바깥으로 내보내려는 방향으로 작용하지만, 전기적 기울기의 힘은 이와 반대 방향으로 작용한다• 따라서 이 두 힘 이 서로 갈등을 일으키기 때문에 , 상쇄되고 남은 순수한 힘에 의해 소 량의 K+ 이온만 바깥으로 밀려나게 된다. 4.4 활동전위 안정전위는 신경원이 자극을 받지 않는 한 계속 유지된다. 자연 상 태에서 신경원에 대한 자극은 결국 시냅스 전도로 나타나는데, 이 현 상은 다음에 다루기로 하고 여기서는 신경원에 인위적인 자극을 가했 울 때 일어나는 현상만을 살펴보기로 한다. 그림 2-3 은 하나의 축색 내에서 일어나고 있는 전위의 변화를 시간 경과에 따른 함수로 나타낸 것이다. 하나의 전국은 축색의 표면에 놓 고 다른 하나의 전극은 축색 안에 삽입하여 이 두 전극 사이에 일어나 는 전위 변화를 전압계로 기록한다. 그립 2-3 에서 0 으로 표시한 시간
 +3 0
+3 0
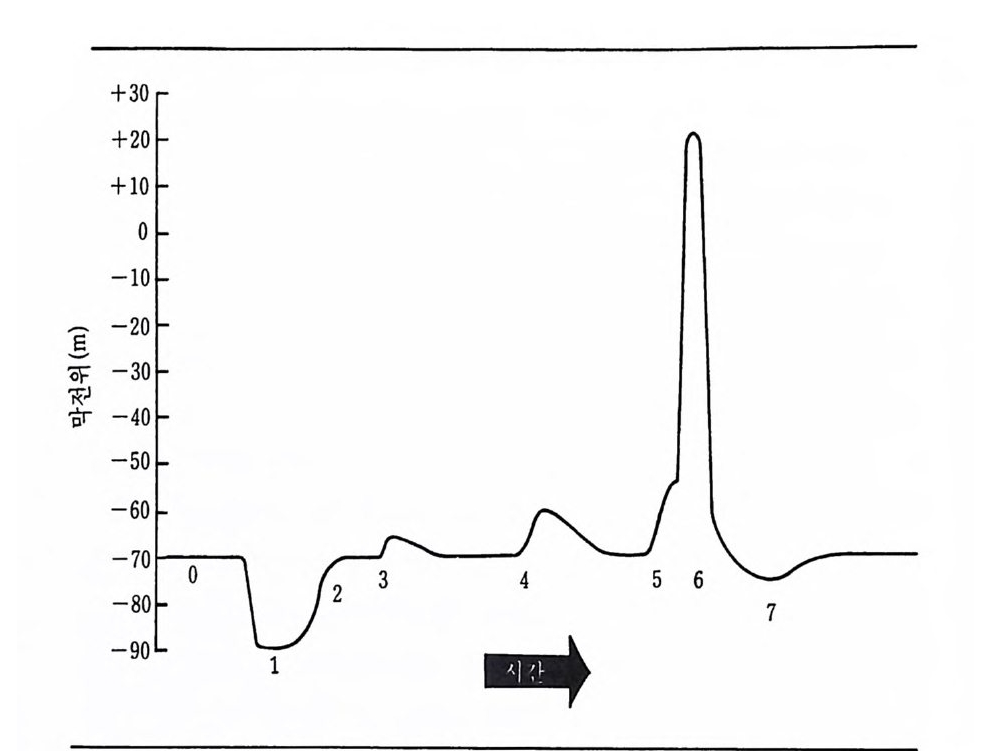
그림 2-3 인위적인 자극을 가했을 때 일어나는 신경원의 막전위 변화 0 은 자국을 가하지 않았을 때, l 은 과분국화 , 3, 4 및 5 는 정도가 다른 탈분 국화, 6 은 활동전위 , 7 은 과분극화임(본문을 참고할 것). 에는 신경원에 아무런 자극도 가하지 않은 상태일 때 나타나는 안정전 위(― 70mV) 이다. 시간 1 에서는 축색 표면의 전극을 통해 미약한 음전 하를 가하면 막 내면의 부적 전하는 더욱 십화되어 음전위가 상승하게 되는데, 이런 변화를 과분국화라고 한다. 시간 2 에서는 신경원에 가한 자극을 멈추게 되면, 즉각적으로 원래의 안정전위 수준으로 되돌아온 다는 사실을 가리킨다. 이번에는 신경원을 탈분국화시킬 수 있는 전류를 흘려보자. 시간 3 에서는 비교적 미약한 전류를 가했을 때 일어난 탈분국화 현상이고, 시간 4 에서는 시간 3 에서보다 약간 더 강한 전류를 가했을 때 일어난 탈분극화 현상이다. 일반적으로 세포들은 수 mV 정도의 미약한 전류 에 의해서도 탈분극이 일어나지만 즉시 원래의 안정전위 수준으로 되
돌아온다. 그러나 시간 5 에서와 같이 다소 강한 전류를 홀리게 되면 이보다 더 강한 전류를 가했을 때(시간 6) 와 마찬가지로 최대한의 전 위발사를 보여주게 된다. 이때 자국은 역치(th reshold) 에 달했다고 하 는데, 역치란 어떤 순간적인 자국이 세포막으로 하여금 신속하면서도 최대한의 전기적 변화를 일으키게 하는 접을 말한다. 비록 세포에 따 라 차이가 있지만 일반적으로 역치는 안정전위로부터 약 15mV 정도 의 탈분극을 일으킬 수 있는 정도의 자극 크기를 말한다. 예컨대 어떤 세포의 안정전위가 ― 70mV 일 때 어떤 자극을 주어 막 내면의 전위 롤 -55mV 정도까지 탈분국화시키면 이 자극은 역치자극이고, 이때 막전위는 신속하면서도 최대한의 전기적 변화를 일으키게 된다. 역치하 자극 (sub th reshold s tim ula ti on) 이란 그림에서 시간 3 이나 시간 4 에서 기록된 것처럼 작은 국소반응을 일으키는 자극이다. 한편, 역치 이상의 자극을 가하면 가해지는 자극의 강도와는 상관없이 시간 6 에서 볼 수 있는 것과 같은 동일한 크기의 반응을 보여준다. 이때 나타나는 반응을 활동전위 (act ion p o t en ti al) 라 하며, 다른 말로는 신경충격 (nerve impu lse) 또는 극파 (s pi ke) 라고도 한다. 일반적으로 수지상돌기에서는 가해지는 자국의 정도에 비례하는 점진성 전위 (grad ed p o t en ti al) 만이 일어나지만, 축색돌기에서는 활동전위만 일어난다. 하나의 주어진 신경세포에서 일어날 수 있는 모든 활동전위는 그 크 기나 모양이 모두 동일하다. 이런 현상을 실무율 법칙 (all-or-none law) 이라 하는데, 좀더 정확하게 말하면 실무율 법칙이란 〈 활동전위의 크기나 모양은 이롤 일으키는 자극의 강도와는 독립적이다〉라는 뜻이 다. 이 법칙에 따르면 한 신경원이 전할 수 있는 메시지는 전보의 메 시지와 유사하다. 죽 전보 송신자가 보다 큰 점 (do t)과 보다 큰 선 (dash) 으로 메시지를 보내고 싶어도 이것이 불가능한 것처럼 한 신경 원이 더 큰 활동전위의 메시지를 보내고 싶어도 보낼 수 없다. 이 양 자의 메시지 송신에서 메시지는 오직 충격 {imp ulse) 과 쉼 (p ause) 의 두 가지 전달방식만을 연속적으로 되풀이하는 것이다.
4.5 활동전위 동안의 분자이동 활동전위가 일어나면 막을 사이에 두고 대치하고 있던 이온둘 사이 에 심한 동요현상이 일어나게 된다. 예컨대 안정상태시 막 바깥에 주 로 존재하던 N 갑 이온은 앞서 언급했던 전기적 기울기와 농도기울기 에 따라 막 안쪽으로 들어오려고 한다. 만약 이러한 N 강 이온돌의 막 내로의 전입이 허용되면 이들은 순식간에 막 내로 진입한다. 안정상태 의 세포막은 N 갑 아온의 진입을 허용하지 않지만 일단 활동전위가 발 생되면 Na+ 의 전입이 순간적으로 허용된다. N 갑의 통로를 형성하는 막 단백질은 전압활성화 (vol t a g e-a cti va t ed) 통로인데, 이것은 다시 말해 막 전위의 차에 따라 Na+ 의 투과성이 달 라진다는 뜻이다. 신경원의 막이 약간만 탈분국되어도 N 갑 통로가 열 리기 시작하며 N 감의 막 내 진입이 쉬워진다. 만약 탈분극이 미약할 때는(역치 이하의 자극인 경우) 증가된 Na+ 이온의 막 내 진입은 K+ 이온의 막 의 배출에 의해 균형울 이루게 된다. 이때 왜 K+ 이온이 막 바깥으로 나갈까 하는 의문이 제기된다. 그 이유는 K+ 이온을 계 속 막 내에 붙잡아둘 수 있게 하는 전기적 기울기가 N 강의 유입으로 약해지기 때문에 K+ 이온이 바깥으로 밀려나게 된다. 그러므로 막의 탈분극화는 Na+ 나 K+ 의 통로를 모두 넓혀준다는 뜻이 된다. 막전위가 역치에 이르게 되면 통로는 더욱 넓어져서, K+ 가 막 밖으 로 유출되는 양보다 더 많은 양의 Na+ 가 막 내로 전입해 들어오게 된 다. 이렇게 막 내로 들어온 Na+ 이온은 세포를 더욱 심하게 탈분국화 시키며, 이에 따라 N 핥 통로는 더욱 넓어지게 된다. 화학적 기울기에 의한 N 갑의 막 내 진입은 막전위가 OmV 를 넘어 +30mV 가 될 때까 지 계속된다(그립 2-3 의 시간 6 에 해당). 막전위가 최고치에 도달해도 N 갑의 농도는 여전히 막 내보다 막 의 에서 높다. 활동전위에 의해 신경원 내에서의 Na+ 의 농도가 높아진다 하더라도 대개의 경우 불과 1% 미만의 증가에 머물고 만다. 따라서 계속되는 농도기울기 때문에 N 갑 이온은 계속 세포 안으로 들어가려
고 하지만 세포 내의 전기적 기울기가 Na+ 의 세포 내로의 전입을 더 이상 허용하지 않는다. 이에 덧붙여 활동전압이 최고치에 이를 무렵에 Na+ 통로는 서서히 닫히기 시작하지만, K+ 통로는 여전히 정상일 때보다 더 넓게 열려 있다. 따라서 Na+ 이온은 더 이상 세포 내 진입이 어려워지지만 K+ 이온은 여전히 세포 내에 더 높은 농도로 존재하기 때문에 막을 떠나 바깥으로 나가게 된다. 막전위의 안정상태로의 복귀는 막 안으로 들어 온 N 갑을 막 바깥으로 추방하고 막 바깥에 있는 K+ 이온을 막 안으 로 잡아들이는 Na+-K+ 펌프 (sod i um- p o t ass i um p um p)에 의해 이루어진 다. 이때 K+ 이온의 막 투과성 증가 때문에 지나치게 많은 양의 양이 온이 세포 바깥으로 유출되어 있고, 또한 Na+-K+ 펌프에 의해 막 내 의 N 감가 막 바깥으로 추방되기 때문에 일시적으로는 안정상태보다 더 심한 분극화, 죽 과분국화 상태가 일어난다(그립 2-3 의 시간 7 의 경 우). 막을 넘나드는 이온의 흐름을 방해하게 되면 신경활동에 심한 변화 가 나타난다. 예 컨대 치과에서 주로 사용하는 노보카인 (novocain e ) 이 나 자일로카인 (x y loca i ne) 과 감은 국소 마취제를 투여하면 이 약물들이 세포막의 N 핥 통로에 부착되어 Na+ 이온의 막 내 진입을 방해한다 (Mi ller, 1985) . 이 런 까닭으로 이 약물이 작용하는 범 위 내 에 있는 모 든 신경원들은 활동전위의 발생이 차단되어 마취효과가 나타나게 된 다. 같은 이유로 이 마취제를 통각정보를 전달하는 감각신경에 주입하 면 동각정보의 전달이 차단되어 통각이 뇌로 전달되지 못하게 된다. 4.6 불응기 막전위가 최고치로부터 안정치로 되돌아오는 동안에 세포들은 새로 운 활동전위를 생성하지 못한다. 왜냐하면 아주 짧은 기간 동안 세포 가 재흥분발사에 저항하기 때문이다. 죽 신경세포는 활동전위가 발생 되고 난 후 약 0.001 초 동안 반응을 하지 않는 불옹기 (refr ac to r y
p er i od) 란 시기가 있다. 불응기는 두 단계가 있는데 첫번째 단계를 절 대불응기 (absolute refr ac to r y p er i od) 라 하며, 두번째 단계를 상대불응 기 (relativ e refr ac to r y pe r i od) 라 한다. 절대불응기 동안에는 아무리 강 한 자극을 가해도 결코 막전위가 발생되지 않는다. 그러나 상대불웅기 동안에는 정상 역치보다 더 강한 자극을 가하면 활동전위가 발생된다. 이 두 불웅기 동안 N 갑 이온의 막투과성은 정상보다 더 낮아지며 K+ 이온의 막두과성은 더 높아진다. 불응기는 한 신경원의 흥분빈도의 최고치를 결정하는 데 중요하다. 만약 어떤 한 신경원의 절대불응기가 0.001 초라고 한다면, 어떤 자극 도 이 신경원에 초당 1, 000 번 이상 활동전위를 야기시킬 수 없다. 만 약 가하는 자극이 더 미약하면 상대불응기 때문에 신경충격 발사반도 는 더욱 낮아질 수 있다. 불응기가 나타나는 시간 간격은 신경원에 따라 차이가 있기 때문에 연구자들은 뇌가 어떻게 행동을 통제하는가를 알아보기 위해 이를 이 용한다. 예컨대 어떤 행동을 지배하는 신경원의 불웅기가 이 행동과 유사한 다른 행동을 지배하는 신경원의 불웅기와 똑같다는 사실이 받 견되었다면, 이 두 행동은 동일한 신경원에 의해 통제받을 것으로 예 상할 수 있을 것이다. 이와는 달리 두 개의 연관된 행동들의 불웅기가 서로 다르다면, 이 두 행동들은 각기 상이한 신경원에 의해 지배되는 것으로 생각할 수 있을 것이다. 4.7 활동전위의 전파 지금까지 우리는 신경원의 어느 한 지점에서 일어나는 활동전위에 대해 언급하였다. 지금부터는 이러한 활동전위가 어떻게 다른 신경세 포로 이동되는지를 알아보기로 하자. 일반적으로 활동전위는 세포체에서 축색이 출발하는 축색구 (axon h ill ock) 라는 곳에서 발생한다. 축색은 불량 도체이기 때문에 축색구에 서 발생한 활동전위가 먼 거리까지 이동되는 방식은 전선을 동해 전기
가 흐르는 방식과는 다르다. 활동전위의 축색 이동은 처음 활동전위가 발생될 때와 마찬가지로 축색의 막을 따라 새로운 활동전위가 연속적 으로 재생되는 방식으로 이루어진다. 활동전위가 일어나면 N 강 이온은 축색의 어느 한 지점에서 막 안으 로 들어가게 되는데, 바로 이 지점의 막 내는 일시적으로 인접 지점의 축색 부위에 비해서 양전하를 띠게 된다. 활동전위가 발생한 지점에서 부터 양전하는 축색을 따라 이동하면서 인접 부위의 막에 탈분극을 일 으킨다. 이런 방식으로 활동전위는 계속 발생되면서 축색을 따라 전달 된다. 하나의 활동전위가 축색을 따라 이동해 가는 것을 활동전위의 전파 (pro p ag a t io n of t he acti on pot e n ti al) 라고 한다. 이 러 한 활동전위 의 전 파 는 곧 하나의 활동전위가 제 2 의 활동전위를 낳고, 제 2 의 활동전위는 제 3 의 활동전위를 낳는 식으로 축색의 종말까지 계속 파급되어 가기 때문에 처음 발생한 활동전위와 마지막 활동전위는 그 강도에서 동일 하다. 활동전위의 전도 속도는 전기 전도의 속도보다 훨씬 느린데, 그 이유는 활동전위가 일어나기 위해서는 N 강 이온의 막 내로의 이동이 이루어져야 하기 때문이다. 지름이 가는 축색에서 일어나는 활동전위 의 전도 속도는 매우 느려 초당 lm 미만에 불과하고, 지름이 굵지만 수초가 없는 축색의 경우는 초당 10m 정도이며, 지름이 굵고 수초가 덮여 있는 축색의 경우는 초당 100m 또는 그 이상인 경우도 있다. 한 때 신경전달 속도가 전기전달 속도와 같을 것이라고 생각한 적도 있는 데 이는 크게 잘못된 생각이었다. 4.7 .1 수초와 도약 전도 척추동물의 경우라도 수초가 없는 축색의 활동전위 전달 속도는 가 장 굵은 섬유일지라도 초당 10m 정도에 불과하다. 이런 속도라면 기 린의 발에서 발생된 신경충격이 뇌까지 도달하는 데는 약 1 초 정도가 소요될 것이며, 비록 덩치가 작은 동물이라 하더라도 초당 10m 정도 의 전달 속도라면 어떤 긴급성이 요구되는 협응적 반응이 일어나기에
는 느린 속도이다. 그러나 축색에 수초가 덮임에 따라 신속한 전달과 협웅적 반웅이 가능하게 되었다. 어떻게 수초화가 활동전위의 전달을 촉진시키는가를 논의하기 앞서 다음과 감은 비유를 고려해 보기로 하자. 가령 어떤 사람이 기계 장치 의 도움 없이 3km 정도 떨어전 곳으로 달려가 서류를 전한다고 하자. 이 경우가 바로 무수초화된 축색을 따라 활동전위가 전달되는 경우이 다. 이 경우에도 메시지는 전달될 수 있지만 그 속도는 매우 느리다. 이번에는 메시지를 야구공 속에 넣어 이 공을 던져서 전하는 경우를 생각해 보자. 한 사람이 3km 나 떨어진 곳까지 한꺼번에 공을 던질 수 는 없지만 여러 사람이 중간 중간 적당한 거리에 배치되어 이 공을 목 적지까지 중개할 수 있다. 이 경우가 바로 수초화된 축색을 따라 활동 전위가 전해지는 경우에 비유될 수 있는데 이렇게 하면 매우 빠른 속 도로 메시지를 전할 수 있다. 척추동물의 축색은 주로 지방질로 된 수초라는 막으로 덮여 있다. 이 수초는 약 1mm 정도의 거리를 두고 수초가 덮여 있지 않은 랑비 에 결절에 의해 끊겨져 있다(그림 2-1 을 참고). 활동전위가 축색구에서 발생되면, 이것은 축색을 따라 첫 수초가 시작되는 곳까지 이르게 되 며 다음엔 이 활동전위가 첫 수초 결절을 뛰어넘어 첫 랑비에 결절까 지 이르게 되고 여기서 다시 다음 수초 결절을 뛰어 넘어 두번째 랑비 에 결절로 넘어오게 되는데, 이런 도약식 전도방식이 축색의 종말까지 계속 이어진다. 첫 랑비에 결절과 두번째 랑비에 결절 사이에서는 활 동전위가 발생될 수 없는데 그 이유는 다음의 두 가지이다. 첫째 두 결절 사이에 있는 수초의 막이 전기적 전달에 대한 저항성을 증가시키 기 때문이며, 둘째는 랑비에 결절에는 Na+ 의 통로가 많이 분포되어 있지만, 결절들 사이의 수초 부분에는 이 통로가 없기 때문이다. 어떤 한 랑비에 결절과 다음 결절 사이의 전도는 전기긴장성 전도 (electr o to n ic conduc ti on) 에 의해 이루어진다. 비록 이 방식이 강도의 손실 없이 메시지를 먼 곳까지 전달할 수 있는 것은 아니지만 전달방 식 중 가장 빠른 방식이다. 신경충격이 첫 결절에서 다음 결철까지 긴
장성 전도로 전달되어도 전하는 여전히 역치 이상이기 때문에 다음 결 절에서도 충분한 강도의 활동전위가 재생될 수 있어서 그 다음 결절까 지 계속 전달될 수 있다. 결절마다 일어나는 활동전위와 결절 사이의 긴장성 전도가 계속 교번하는 것을 도약전도 (sal t a t o ry condu cti on) 라고 한다. 수초가 덮인 축색의 전도속도가 초당 120m 나 되는 경우도 있는 데 이렇게 전도속도가 빠른 것은 도약전도 때문이다. 4.7.2 활동전위 없는 신호전달 활동전위에 관해 지금까지 밝혀진 것은 오칭어의 거대축색과 같은 큰 신경원에서 발견된 사실들이 근간을 이룬다. 일반적으로 긴 축색을 갖는 큰 신경원들은 척수로부터 말초근육까지, 또는 뇌의 한 부위에서 다른 부위까지 등 비교적 먼 거리까지 메시지를 전달할 수 있도록 전 문화되어 있다. 그러나 짧은 축색을 갖는 작은 신경원들은(경우에 따라 서는 아예 축색이 없는 경우도 포함) 아주 짧은 거리 내에서만 메시지 교 환이 이루어질 뿐인데, 이 신경원들은 몇 가지 점에서 큰 신경원들과 차이가 있다. 짧은 축색조차도 갖지 않는 작은 신경원을 국소신경원(l ocal neuron) 이라 하는데, 이 신경원은 바로 이 신경원과 인접한 신경원에게만 정 보를 전달할 뿐 활동전위는 야기하지 못한다. 이 신경원은 크기가 다 양한 막전위의 일종인 점진성 전위 (grad ed p o t en ti al) 를 야기한다. 국소 신경원에 자극을 가하면 가해지는 자극의 정도에 비례하여 탈분극이 일어나는데 이러한 막전위의 변화도 인접 세포로 전달될 수 있다. 즉, 이 점진성 전위는 인접 세포에 전달되면 활동전위와는 판이하게 전위 의 강도가 현저하게 감소된다. 사실 모든 신경원의 수지상돌기에서는 점진성 전위를 보여준다. 그러나 국소신경원의 경우는 이 신경원의 어 디에서도 활동전위가 일어나지 않으며 오직 점진성 전위만이 발생될 뿐이다. 그러나 이러한 점진성 전위도 인접 세포로 정보를 전달하는 데는 충분한 강도의 전위가 된다. 긴 축색을 갖는 큰 신경원은 세포의 한쪽 끝인 수지상돌기에서 정보
롤 받아 이 세포의 다른 쪽 끝인 축색돌기의 종말을 통해 인접 세포로 정보를 보낸다. 그러나 국소신경원에는 이러한 수용부와 전달부 사이 의 뚜렷한 구분이 없고, 세포막의 여러 부위에서 정보를 받아들여 이 를 인접한 다른 부위로 전달한다. 대개의 경우 한 국소신경원의 수지 상돌기로부터 다른 신경원의 수지상돌기로 직접 정보전달이 이루어지 게 된다. 아직 국소신경원에 관한 지식은 단편적인 것에 불과하지만 국소신경원이 정보처리에 매우 중요한 기능을 할 것으로 생각된다. 우 리는 앞으로 국소신경원이 시각정보를 처리하는 예들을 살펴보게 될 것 0l 다. 5 시냅스 전도 신경충격이 축색종말에 이르면 인접하는 다른 세포들과의 정보소통 울 위한 일련의 사건들이 일어난다. 이때 소통이 이루어지는 다른 세 포들이란 신경세포일 수도 있고, 근육세포일 수도 있으며, 내분비세포 일 수도 있다. 한 신경원의 축색종말과 다른 신경원의 수지상돌기, 세 포체 또는 축색과의 접합을 시냅스라고 한다. 시냅스에는 전기적 시냅 스 (ele ct r ic al s yn a p ses) 와 화학적 시냅스 (che mi cal s yn a p ses) 가 있다. 전 기적 시냅스에서는 한 신경원의 신경충격이 특수한 통로를 거쳐 인접 한 다른 신경원으로 직접 전달된다. 전기적 시냅스는 혼치 않으며 화 학적 시냅스가 대부분이다. 시냅스에서 화학적 신호를 전달할지 아니 면 전기적 신호를 전달할지를 결정하는 신경원을 시냅스전 신경원 {pre syn a pt ic n euron) 이라 하며, 정보를 수취하는 신경원을 시냅스후 신 경원{p os t s yn a pti c neuron) 이라 한다. 실제로 거의 모든 신경원은 다른 신경원과의 관계 속에서 불 때 시냅스전 신경원이 될 수도 있고 시냅 스후 신경원이 될 수도 있다.
5.1 화학적 시냅스 화학적 시냅스의 전도방식에 대한 이해는 뇌의 작용방식에 대한 이 해뿐만 아니라 정상 또는 비정상적인 행동을 이해하는 데도 중요하다. 대부분의 향정신성 약물들은 시냅스에서 작용하는 화학적 전달물질들 과 상호작용하며, 정신분열증이나 우울증과 같은 심한 정신병은 화학 적 시냅스의 비정상적인 기능들 때문에 야기된다(이런 문제는 앞으로 자 세하게 논의할 것이다). 그림 2-4 에는 화학적 시냅스의 주요 부분을 나
 스주머니
스주머니
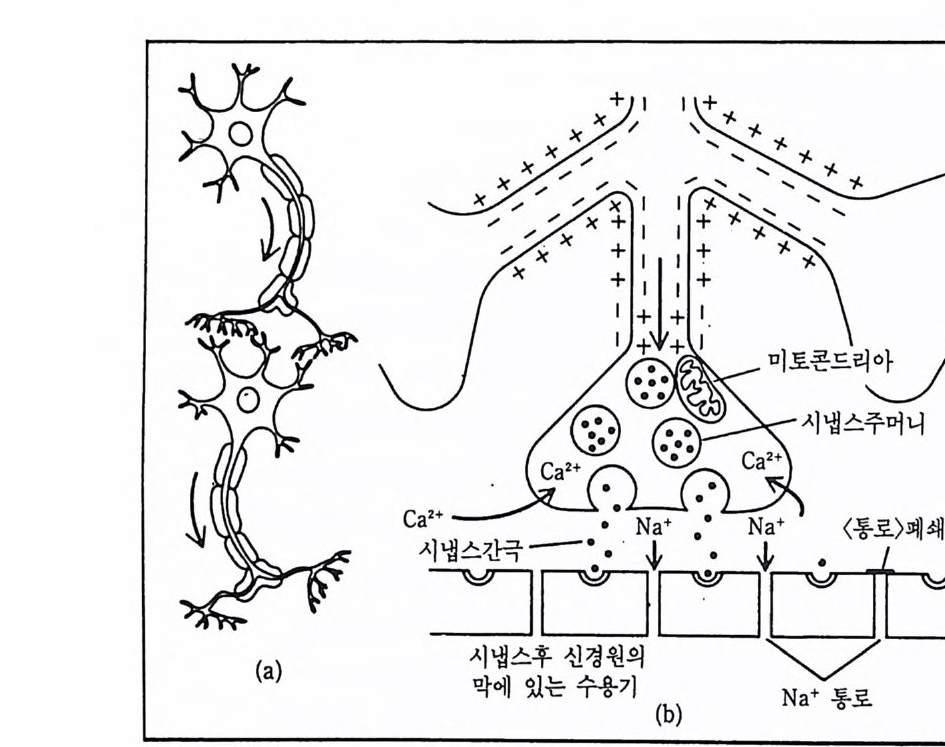
그림 2-4 화학적 시냅스에서의 정보전달 (a) 정보전달은 시냅스전 신경원의 축색종말로부터 시냅스후 신경원의 수지상 돌기나 세포체로 전해진다. (b) Ca2+ 이온이 시냅스전 종말로 들어가 신경전달 물질을 분비시킨다. 죽 시냅스 주머니가 열리고, 그 속에 들어 있던 신경전달 물질이 시냅스 간국으로 홀러나오면 간국을 지나 시냅스후 수용기와 결합하게 된다. 이렇게 되면 시냅스후 신경원의 막에 있는 이은동로가 열리게 되어 Na+ 이온이 이 통로로 들어가게 된다.
타내었다. 시냅스전 종말이 갖는 몇 가지 특징 중 가장 두드러진 것은 이 부분 이 볼록하게 확장되어 있다는 점이다. 축색종말에는 신경전달물질을 함유하고 있는 시냅스 주머니 (syn a p tic ves i cle) 가 있는데 이곳에는 여 러 가지 종류의 신경전달물질 (neuro t ransm itt er) 아 들어 있다. 한편 체 내에는 많은 화학물질들이 있어서 이 물질들이 신경원의 기능에 영향 울 미칠 수 있다. Barchas 등 (1978) 은 〈 인접 신경세포에 정보를 전달 하는 물질 〉 을 신경전달물질이라고 정의했던 재래의 정의를 수정해야 한다고 주장했다. 왜냐하면 물질의 종류에 따라서 신경활동을 증폭시 킬 수도 있고 약화시킬 수도 있기 때문에 신경전달물질이란 개념 대신 에 신경변조물질 (neuromodula t ors) 이란 개념이 더 적합하다고 주장하 였다. 신경전달물질이든 신경변조물질이든 이 물질들은 넓은 의미의 신경조정물질 (neurore gu la t or) 에 속한다. 표 2-3 에는 일반적으로 인정 표 2-3 일반적으로 인정되는 신경전달뭉질의 후보물질 일반적으로 신경전달물질로 인정된 물질 Acety lc holi ne Bio g e n ic ami ne s No repi ne p hr in e (noradrenalin ) Ep ine p hr in e (adrenalin e ) Dop am i ne Seroto n in Hista m i ne 아미노산과 이의 유도체 Gamma-ami no buty ri c aci d ( GABA) Gluta m ate Glyc ine Aspa r t at e Lute in i z in g hormone releasin g hormone (LHRH) 가능성 있는 신경전달물질 Substa n ce P /3- endorp h in Oxy toc in Vasop re ssin Somato s ta t in TRF (thyrot ro p in releasin g fac to r ) Taurin e Met- e nkep ha lin Cholecys t o k in in Leu-enkeph a li n Vasoacti ve int e s tin a l hormone (VIP)
되는 신경전달물질의 목록이 제시되어 있는데 이 물질들은 척추동물의 신경계에서 흔히 발견되는 것들이다. 시냅스전 종말은 시냅스후 산경원의 막에 있는 시냅스후 수용기 (po sts y n a pt ic recep tor ) 라는 특수한 부위 와 서 로 마주 보고 있다. 이 수용기는 신경전달물질이 부착되는 곳으로 단백질의 구조가 변형되어 만들어진 곳이다. 시냅스전 종말과 시냅스후 수용 부위 사이에 시냅스 간극 (s yn a ptic cle ft)이 있는데 이 간극은 20-30run 정도이다. 시냅스는 시냅스전 종말, 시냅스 간극 그리고 시냅스후 수용 부위와 같은 세 가 지 요소로 구성된다. 축색종말과 수지상돌기 사이에 있는 시냅스를 축 색수지 상 시 냅 스 (axodend ritic syn a ps e ) 라 부르는데 , 이 시 냅 스가 가장 많다. 또 다른 유형 의 시 냅스로는 축색축색 시 냅스 (axoaxon ic syn a ps e ) 와 축색 세포체 시 냅스 (axosoma tic syn a ps e ) 그리고 심 지 어는 수지 상수 지상 시냅스 (dendrodendr itic s yn a p se) 도 있다. 5.2 화학적 이온동로 화학적 시냅스에서 일어나는 일련의 사건들은 신경충격이 시냅스전 축색의 종말에 도달하는 순간부터 시작된다. 죽, 이때 시냅스 주머니 속에 있는 신경전달물질이 시냅스 간극으로 분비되며, 분비된 신경전 달물질은 시냅스 간극 내에서 확산되어, 시냅스후 막에 있는 수용기 부위와 결합하게 된다. 다음에는 시냅스후 신경원막의 이온통로가 열 리면서 어떤 이온은 시냅스후 신경원 내부로 들어가고 어떤 이온은 신 경원 바깥으로 홀러나오게 된다. 이때 이온의 유입과 유출은 그 이온 의 전기적 또는 화학적 기울기에 의해 통제된다. 앞에서 본 축색의 경 우와는 달리 이 이온둘의 통로는 전기적으로 작동하는 것이 아니라 화 학적으로만 작동된다. 축색에 있는 전기적 이온통로에 신경전달물질과 갇은 화학적 자극을 주어도 아무런 영향이 없는 것처럼 , 시냅스후 신 경원의 화학적 이온통로를 전기적으로 자극해도 아무런 변화가 일어나 지 않는다.
5.3 효능물질과 길항물질 신경전달물질과 수용기 분자는 열쇠와 자물쇠의 관계처럼 서로 잘 결합된다. 이런 결합은 하나의 특수한 관계일 뿐 결코 고유한 관계는 아니다. 즉, 특수 신경전달물질이 아닌 다른 여러 종류의 화학물질들 도 시냅스후 수용기에 결합될 수 있다. 이런 화학물질들은 정상적인 시냅스 전달기능을 차단시킬 수도 있고, 자연적인 전달물질의 효과를 흉내내는 경우도 있고, 정상적인 시냅스 활동의 작용기간을 연장시킬 수도 있다. 수용기에 결합하여 자연적인 신경전달물질과 극히 유사한 생물학적 기능을 하는 물질들을 그 전달물질에 대한 효능물질 (a g o ni s t) 이라 부른다. 한편 수용기 분자와 결합하여 정상적인 시냅스후 효과를 일으키지 못하게 함으로써 정상적인 시냅스 전도를 차단하는 물질을 길항물질 (an t a g on i s t)이라 한다. 중추신경계에 영향을 미치는 많은 약 물들은 특정 신경전달물질에 대한 효능물질과 길항물질로 구분할 수 있다. 약물이나 기타 다른 화학물질들도 여러 가지 방식으로 시냅스에서 일어나는 사건들에 영향을 미칠 수 있다. 즉 이런 물질들은 신경전달 표 2~4 화학적 전달에 영향을 미치는 약물들 신경전달물질 효능물질 길항물질 No repi ne ph r in e Amp he ta m i ne Reserp ine Tri-c y cl ic a nti de p re ssants Alph a -meth ylt yr os in e Dop am i ne Ap om orp h in e Reserp ine Amp he ta m i ne Chlorp ro mazin e Cocain e Seroto n in Ipro nia z id Reserp ine Acety lc holi ne Phy so stig mine Atr o p ine (musca rinic) Curare (nico ti nic) GABA Muscim o l Bic u cullin e Pic ro to x in
물질의 합성과 방출 그리고 시냅스 간극에 방출된 전달물질의 분해와 흡수 등의 과정을 촉진시키거나 여제시킬 수 있다. 많은 약물들은 시 냅스에서 한 가지 이상의 방식으로 영향을 미친다. 어떤 약물둘은 단 지 한 종류의 신경전달물질에만 영향을 미치는 선택적인 것도 있고, 또는 광범위하게 여러 종류의 상이한 시냅스에 작용하여 화학적 전달 과정 전반에 영향을 미칠 수 있는 것도 있다. 흔히 화학적 시냅스 전 달에 영향을 줄 수 있는 약물들은 시냅스 간극에서 신경전달물질의 양 을 증가시키거나 감소시킨다. 표 2-4 에는 시냅스 전달에 영향을 미치 는 것으로 알려진 몇 가지의 효능물질과 길항물질이 제시되어 있다. 정신약리학(p s y cho p ha tm acolo gy)이란 분야는 주로 약물이 화학적 시냅 스에 미치는 영향을 다루는 신경과학의 한 분야이다. 5.4 전달물질 방출과 Ca2+ 의 중요성 축색종말에는 Na+ 과 K+ 의 통로는 물론 Ca2+ 이온의 통로도 있다. 이 Ca2+ 통로는 전압석 통로로서 신경충격이 축색종말에 도달하면 열 리게 된다. 동로가 개방되면 시냅스 간극에 있는 Ca2+ 가 축색종말 안 쪽으로 신속히 유입되는데, 이것은 Na + 가 전기적, 화학적 기울기에 의해 세포 안으로 둘어가는 양상과 유사하다 . Ca2+ 가 축색종말 속에 들어가면 신경전달물질이 방출된다. 만약 인위적으로 축색종말 주변에 있는 체액둘을 제거해 버리면, 비록 축색에 자극을 가해 신경충격을 일으켜도 축색종말에서 신경전달물질이 방출되지 않는다. 그러나 이 축색을 Ca2+ 가 포함된 용액 속에 담그고 신경충격을 가하면 신경전달 물질이 정상적으로 방출된다. 한편 Ca2+ 울 축색종말 부위에 직접 주사 하면 신경충격이 없음에도 불구하고 축색종말에서 신경전달물질이 방 출된다. 시냅스전 종말에서 이루어지는 신경충격의 기능 가운데 가장 중요한 것은 Ca2+ 의 이온통로를 개방하여 Ca2+ 를 축색종말에 유입시 킴으로써 신경전달물질의 방출을 유도하게 되는 것이다. Ca” 의 축색종말 유입이 어떤 기제를 통해 시냅스 주머니 속에 들어
있는 신경전달물질을 방출시키는지는 아직 미지수이다. 그러나 한 가 설에 의하면 축색종말에서 Ca2+ 가 시냅스 주머니를 간극 쪽으로 이동 시 키 는 수축기 제 (contr a cti le mechanis m ) 를 활성 화시 킬 것 이 라는 것 이 다. 그렇게 되면 이동된 주머니의 막이 종말의 막과 합쳐져서 간극 쪽 으로 개방된 하나의 주머니가 만들어지고 이 주머니에서 간국으로 신 경 전달물질이 방출되 는데 , 이 런 과정 을 세 포의 유출 (exoc yt os i s) 이 라고 한다. 5.5 시냅스후 전위 신경전달물질과 시냅스후 수용기 사이의 상호작용에 의해 발생되는 전류는 이온성 전류이다. 죽 Na+ 는 흥분성 시냅스후 전위 (exc itat o r y pos ts y n a p tic pot e n ti al : EPSP) 발생 에 관여 하는 이 온이 고, K 냈} Cl- 는 억제성 시냅스후 전위 (inh ib i t or y pos ts y n a pt ic p ot e n ti al : IPSP) 발생에 관여하는 이온이다. 이 이온성 전위 발생에 관해서는 나중에 자세하게 논할 것이다. 그러나 여기서 빼놓을 수 없는 것은 이런 시냅스후 전위 나 축색에서 일어나는 신경충격(활동전위) 모두 동일한 이온이 개입되 지만 이 두 전위는 여러 가지 의미에서 서로 다르다는 것이다. 즉, 첫 째, 활동전위 즉 신경충격의 크기는 언제나 일정하지만(실무율의 법 칙), 시냅스후 전위의 크기는 실무울에 따르지 않는 점진성 전위이다. 둘째, 신경충격은 언제나 흥분성으로 작용하지만 시냅스후 전위는 홍 분성도 될 수 있고 억제성도 될 수 있다. 5.6 시냅스 지체 시냅스 전도가 이루어지는 데는 시간이 필요하다. 오징어의 거대축 색을 대상으로 하여 시냅스 현상을 연구한 보고에 의하면 신경충격이 축색종말에 도달하여 신경전달물질을 방출하기까지의 시간간격이 약 0.0002 초 정도인데, 이 시간간격은 주로 Ca2+ 통로가 열리는 데 소요
되는 시간 때문이다. 또한 세포의유출, 유출된 신경전달물질이 시냅스 간극으로 확산되는 과정 그리고 확산된 신경전달물질이 수용기 분자와 결합하는 과정 등에도 그 이상의 시간이 소요된다. 이처럼 시냅스전 신경충격으로부터 시냅스후 전위가 일어날 때까지의 소요시간을 시냅 스 지체 (syn a pt ic dela y)라 하는데 포유동물의 경우 약 0.001 초 정도이다. 5.7 흥분성 시 냅스후 전위 신경전달물질의 주요 기능은 시냅스후 신경원의 막에 있는 이온통로 룰 여는 것이다. 시냅스후 전위의 특성은 어떤 종류의 이온통로가 열 리는가에 따라 달라진다. 시냅스후 전위는 EPSP 가 될 수도 있고 IPSP 가 될 수도 있는데, K+ 의 전도성을 감소시키거나 N 강 통로를 여 는 신경전달물질의 작용은 EPSP 를 발생시킬 수 있다. 막을 사이에 두고 교환되는 이온의 수는 어떤 특정 신경전달물질에 의해 얼마나 많은 이온통로들이 열리는가와 이들 통로의 열린 상태가 얼마나 오래 지속되는가에 따라 달라지며, 이런 과정은 또한 시냅스 간극에 존재하는 가용한 전달물질의 분자 수에도 의존한다. 그리고 시 냅스전 신경원에서 일어나는 탈분극의 수준과 축색종말 내에 유입된 Ca2+ 의 양과 같은 요인들에 의해 가용한 전달물질 분자의 양에 미친 영향들도 고려되어야 한다. 신경전달물질의 작용은 시냅스후 수용기 부위와 시냅스 간극으로부 터 신경전달물질이 제거됨에 따라 멈추게 된다. 신경전달물질의 제거 는 이 물질을 분해하는 효소작용아나 시냅스전 축색종말로 신경전달물 질이 재흡수됨에 따라 이루어진다. 대부분의 시냅스 부위에서는 위의 두 과정이 동시에 일어난다. 바로 이 기제가 시냅스후 전위의 발생기 간을 제한시키고 시냅스 부위에 연결된 수많은 신경회로 사이의 혼란 을 방지해 줄 수 있다. 또한 이 재흡수 기제논 시냅스전 신경원으로 하여금 신경전달물질의 활동을 가능하게 해준다.
5.8 억제성 시냅스후 전위 모든 시냅스가 흥분성인 것은 아니며 상당수는 억제성이다. 억제성 시냅스란 과분국화를 일으키는 시냅스후 전위를 말하는데, 이때 전달 물질의 작용으로 개방되는 수용기 분자의 이온통로는 K+ 이나 c1- 또 는 이 두 이온 모두에 대해 선택적으로 작용하는 통로이다. 이와 같은 과분국화 상태의 시냅스에서 기록한 전압 변동을 억제성 시냅스후 전 위 (IPSP) 라 한다. 왜냐하면 이 전위는 시냅스후 신경원의 막전위를 변 경시켜 신경충격의 발생 역치를 높이기 때문이다. 예컨대, 만약 억제 성 시냅스에서 어떤 전달물질이 K+ 의 동로만을 선택적으로 열면 K+ 이온이 세포 바깥으로 흘러나오게 되는데, 이런 K+ 의 유출은 막 내의 전압을 -75mV 까지 내려가게 하거나 K+ 의 통로가 닫힐 때까지 계속 되어 K+ 이온의 화학적 기울기를 낮춘다. 이런 현상은 과분국화를 초 래하며 결과적으로 역치를 높이게 된다. 시냅스후 전위에 기인되는 막전압과 시냅스후 신경원이 신경충격을 발생시키는 역치 사이의 관련이 중요한데, 여기에는 Cl- 가 관련된다. 표 2-2 에서 보았듯이 오칭어의 거대축색에서 기록되는 Cl- 의 평형전위 는 -60mV 이다. 이 상태에서는 Cl- 의 이온통로가 열려도 이미 이 이 온은 평형상태에 있기 때문에 더 이상의 이온 이동은 나타나지 않는 다. 그러나 c1-통 로와 가까운 곳에 있는 다른 시냅스 부위가 탈분국 울 일으키면 이로 인해 막전압이 -59mV 가 되는 순간 열린 c1-통 로 룰 통해 c1-이 온이 막 내로 유입된다. 이와 같은 Cl- 의 막 내 유입은 막전압이 탈분극 상태에 있는 동안 계속되어 시냅스후 신경원의 흥분 울 방해하고, 전체적인 막전압을 Cl- 의 평형상태 수준으로 유지시킨 다. 그 결과 시냅스후 신경원은 역치보다 수 mV 낮은 과분극상태에 머물게 되고, 이 시냅스는 억제성 시냅스가 된다.
5.9 시냅스 통합 시냅스후 신경원이 흥분성이든 억제성아든 신경충격을 발생시키려면 시냅스후 신경원의 축색구에 있는 충격발생부위 (sp ike init iati ng reg ion ) 에 충분한 정도의 탈분국이 일어나야 한다. 시냅스후 전위는 시냅스 부위와 매우 가까운 지점에서만 막전위를 바꾸어 놓을 수 있다. 신경 충격과는 달리 시냅스후 전위는 스스로 재생될 수 없기 때문에 발생지 점에서 거리가 멀어지면 전위의 크기가 감소되고 효율성도 떨어진다. 비록 특정 시냅스가 충격발생영역에 매우 가까운 곳에 있다 하더라도 이 시냅스 의에 또 다른 시냅스가 동시에 활동을 하지 않으면 충격이 발생되지 못한다. 또한 활동을 하고 있는 시냅스의 위치가 어디인가 하는 것도 대단히 중요하다. 즉 충격발생영역에서 가까운 시냅스일수 록 영향력이 더 커진다. 예컨대 충격발생영역에서 가까운 시냅스는 멀 리 떨어져 있는 시냅스의 영향력을 무시해 버릴 수 있다. 그러므로 충 격발생영역에 가까이 있는 억제성 시냅스는 멀리 떨어져 있는 시냅스, 예컨대 수지상돌기와 같은 곳의 흥분성 시냅스의 영향을 무시할 수 있다. 시냅스후 전위의 지속기간이 수 분에 이르는 경우도 있다. 이처럼 시냅스후 전위가 오랫동안 지속됨으로써 시간적으로 탈분국화와 과분 국화가 서로 합쳐 지는 시 간가합(t em p oral summati on ) 이 이 루어 질 수 있다. 시 간가합 의 에 공간가합 (s pa ti al summati on ) 이 라는 것도 있는데 , 이것은 여러 개의 시냅스후 전위들이 시냅스후 세포 표면의 아주 가까 운 곳에서 일어날 때 그 효과가 가합되는 현상이다. 이런 점진성 전위 에 대한 자세한 증거는 아직 제시되지 않았지만 대뇌 기능에서 매우 중요한 역할을 할 것으로 추측된다.
제 3 장 신경 계통의 해부학 신경해부학자들은 포유류의 신경 계통을 크게 중추신경 계통과 말초 신경 계통으로 나눈다. 말초신경 계통은 다시 두 개의 하위 체계로 나 누어지는데, 하나는 골격근의 감각과 운동을 조정하는 체성신경 계통 이고, 다른 하나는 심장이나 내장과 같은 내장기를 조정하며, 각성이 나 정서상태와 밀접한 관련이 있는 자율신경 계통이다. 우리 인간의 신경계를 구성하고 있는 전체 신경원들 가운데 거의 전부라 할 수 있 는 99% 의 신경원이 중추신경계를 이룬다. 중추신경 계통은 뇌와 척수 로 구성되는데, 인간의 뇌는 자연계에서 발견되는 수많은 조직 가운데 가장 복잡한 조직으로 알려져 있다. 뇌 중에서 최고의 고등정신기능을 담당하는 대뇌피질은 신경 계통 가운데 가장 늦게 발달된 구조물이다. 신경 계통의 조칙 원리를 이해하면 신경 계통의 다양한 기능을 쉽게 이해할 수 있다. 이 장에서는 신경 계통의 주요 구성단위인 말초신경계와 중추신경계 를 중심으로 이들 신경 계통 사이의 상호 연결 및 뇌의 구조와 기능 등에 관한 기본적인 내용을 살펴볼 것이다.
l 말초신경 계통 우리 인 간의 신 경 계 통은 말초신 경 계 통 (pe rip h eral nervous sys t e m : PNS) 과 중추신경 계통 (cen t ral nervous sys t e m : CNS) 으로 구성된다. 중추신경 계통은 뇌 (bra i n) 와 척수 (s pi nal cord) 로 구성되고, 말초신경 계통은 뇌와 척수를 제의한 말초의 체성신경 계통 (soma tic nervous sys t e m ) 과 자율신 경 계 통 (auto n omi c nervous sys t e m : ANS) 으로 구성 된 다. PNS 와 CNS 는 기능적으로 서로 통합되는 방식으로 작용하지만, 우리는 이 두 계통을 구분하여 논의할 것이다. . PNS 는 뇌 신 경 (crania l nerves) , 척 수신경 (spi na l nerves) , 그리 고 자 율신경 계통과 같은 3 가지 종류의 신경 요소로 구성된다. 신경통로(이 롤 구성하는 신경원을 포함)는 신경충격이 전달되는 방향에 따라 구심성 (aff ere nt) , 원심성 (eff ere nt) 그리고 혼합성 (m i xed) 으로 구분된다. 구심 성 신경은 신경충격을 말초에서 중추로 전달하는 신경이며, 이와 반대 로 원십성 신경은 중추로부터 말초 쪽으로 신경충격을 전달하는 신경 이다. 혼합성 신경은 위의 두 가지 방식을 공유하는 신경을 일컫는다. 1.1 뇌신경 척수를 경유하지 않고 뇌로 직접 들어가거나 나오는 12 개의 말초신 경이 있는데 이를 뇌신경이라 부른다. 뇌신경은 뇌의 제일 상부와 연 결되는 신경으로부터 가장 아래 뇌 부위와 연결되는 신경에 이르기까 지 그 순서에 따라 로마숫자의 번호를 붙여 명칭을 표시하는데, 이 순 서는 매우 중요하다. 왜냐하면 뇌신경의 경우 위에서부터 아래 부위에 이르는 개개 신경의 고유 이름보다 고유 번호롤 사용하는 경우가 더 혼하기 때문이다. 표 3-1 에는 12 개의 뇌신경을 뇌 상부에서 하부에 이르는 순서에 따 라 고유 번호, 신경 이름 그리고 이 신경들의 주요 기능을 감각적인 것, 운동적인 것 그리고 감각과 운동의 혼합적인 것으로 대별해 놓았다.
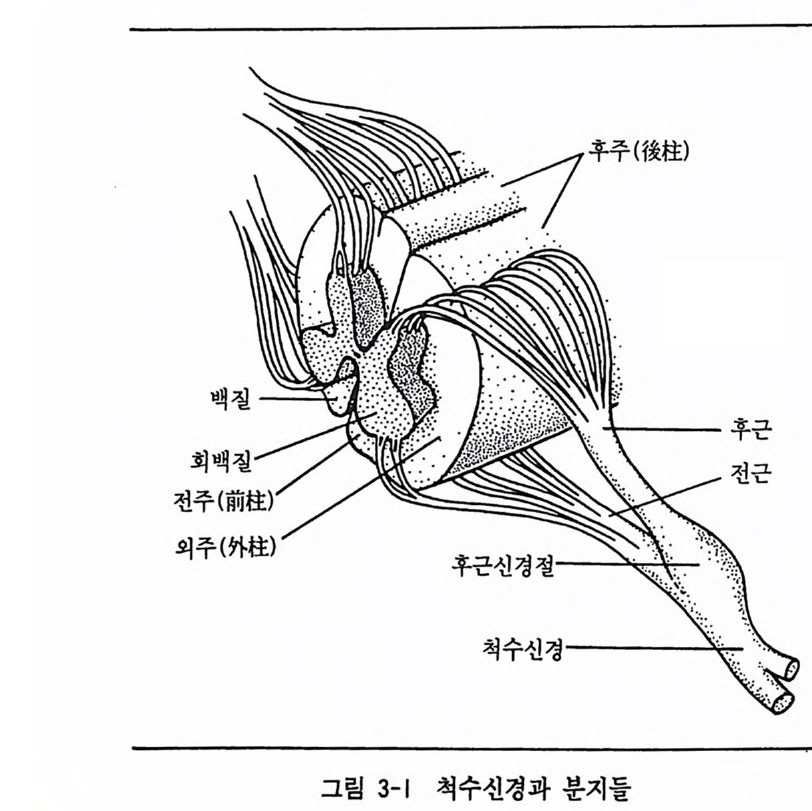
표 3-1 뇌신경의 종류와 기능 요약 뇌신경의 종류 주요기능 I 후구신경 구심성 : 냄새의 지각 II 시각신경 구심성:시각 III 동안신경 원심성 : 안구운동 IV 활차신경 원십성:안구운동 V 삼차신경 혼합성 : 안면감각과 저작운동 VI 의전신경 원심성:안구운동 VII 안면신경 혼합성 : 미각과 안면운동 VIlI 청각신경 구심성 : 청각과 평형감각 IX 설인신경 혼합성 : 마각과 연하운동 X 미주신경 혼합성 : 내부 신체기관의 감각과 심장활동과 갇은 내부 신 체기관의 기능조철 XI 부신경 원심성 :성대와 목운동 XII 설하신경 원심성:혀운동 1.2 척수신경 척수신경은 목의 상부에서부터 등뼈의 꼬리 부분까지 척수의 좌우로 출입하는 31 쌍의 신경을 일컫는다. 이 신경들은 출입하는 척수의 위치 에 따라 크게 5 집단으로 나누어진다. 즉, 목 부위에서부터 꼬리 부위 로 내려가면서, 8 쌍의 경부신경 (cervic a l nerve), 12 쌍의 흉부신경 (tho racic n erve) , 5 쌍 의 요 부 신 경 (lw nbar nerve) , 5 쌍 의 천 골 신 경 (sacral nerve) 그리고 1 쌍의 미골신경 (coccy ge al nerve) 으로 구분된다. 각각의 척수신경은 척수로 들어가는 후근 (dorsal roo t)과 척수에서 나 오는 전근 (ven t ral roo t)으로 구성되는데, 이 두 근은 척수 입구에서 하 나로 합쳐져 하나의 척수신경을 이룬다. 이 신경의 후근은 구심성이고 전근은 원심성이다. 그림 3-1 은 척수신경의 모습을 나타내고 있다. 후 근을 이루는 구심성 신경원의 세포체는 척수신경절 (s pi nal gan g lion ) 속 에 있기 때문에 이 척수신경절을 후근신경절 (dorsal root g a ngli on) 이라
고도 한다. 또한 이들 세포체 주변에는 신경총(p lexus: 신경섭유 및 신경 세포가 그물처럼 얽힌 조직)이라 부르는 신경세포의 망둘이 있는데, 이 것도 말초신경 계통의 일부분이다. 개개의 척수신경은 신체의 특정 부분과 기능적으로 관련되어 있다. 후근은 신체의 특정 영역으로부터 올라오는 감각정보를 뇌로 전달한 다. 이와 반대로 전근은 뇌에서 내려오는 운동정보를 특정 신체 영역 으로 전달하는 기능을 한다. 이와 같이 어떠한 척수신경에 의해 지배 되는 일정 부위의 신체 영역을 체절 (denna t ome) 이라 한다. 서로 인접 하고 있는 체절 사이의 경계는 명확하지 않으며, 때로는 인접 체절들 사이에 서로 중복되는 수도 있다. 그럼에도 불구하고 특정 척수근이 한정된 신체 부분만을 지배한다는 사실은 신경학자들이 척수신경의 장 애 위치를 찾아낼 때 유용한 지표로 사용된다.
 후주(後柱)
후주(後柱)
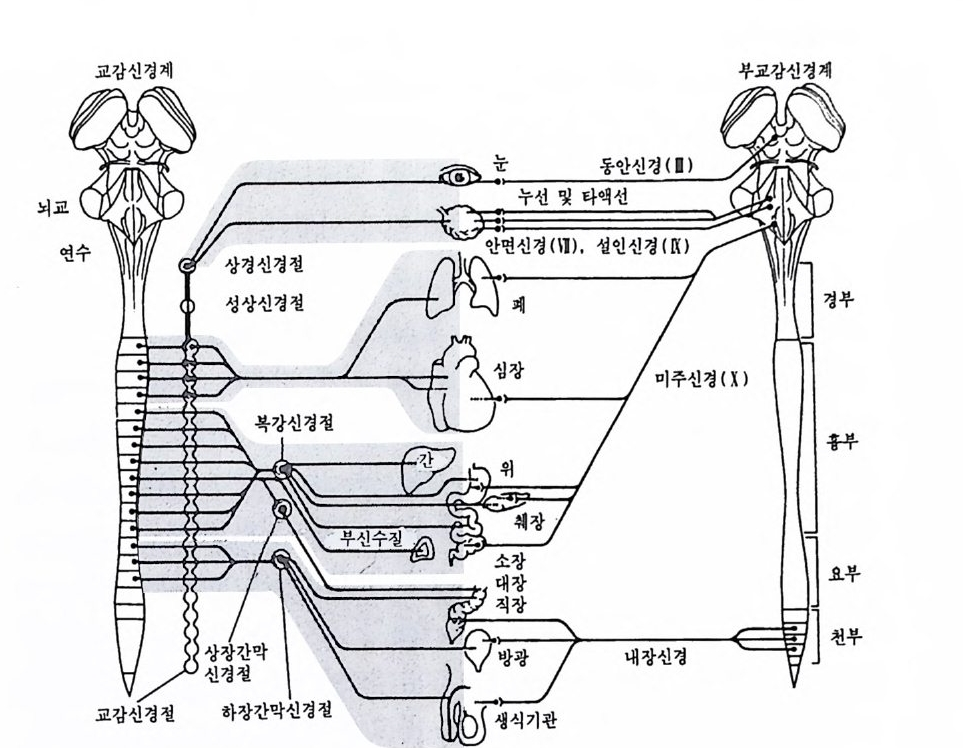 교감신경계 부 교 감신겅계
교감신경계 부 교 감신겅계
그림 3-2 교감신경계와 부교감신경계 일반적으로 자율신경은 절전신경과 철후신경으로 나누어진다. 교감신경계의 절전신경은 척수에서 교감신경절까지이며, 절후신경은 교감신경철에서부터 지 배하는 장기까지이다. 부교감산경은 신경절을 거치지 않고 직접 지배하는 장 기까지 이어진다. 1.3 자율신경 계동 가령 여러분이 대학 입시와 같은 긴박한 상황에 직면해 있다고 하 자. 비록 열심히 공부했지만 자신이 목표로 하는 학과에 지원자가 많 이 몰렸고, 예상되는 커트라인도 평소 자신의 성적보다 더 높을 것으 로 보도되고 있고, 이제 10 분만 지나면 시험이 실시된다고 하자. 이런 상황에 처하게 되면 사람들은 몹시 두려워하고 긴장하여 평소 실력을 충분히 발휘할 수 없게 된다. 손에서는 땀이 나고, 호흡은 거 칠어지고, 심장은 방망이질하며, 뱃속이 울렁거리고 토할 것 같은 느
낌이 들게 된다. 이때 심호흡을 하고, 목을 좌우로 돌리며, 어깨를 올 리고 내리면서 긴장 해소를 위한 노력을 해보지만, 별 효과가 없다. 심장은 계속 뛰고, 손바닥은 땀에 잦고, 위는 계속 역겹다. 불행하게 도 이런 신체 변화는 마음대로 통제하기가 어렵다. 이런 상황은 어려운 시험을 앞두고도 일어나지만, 많은 청중 앞의 무대 위에 서게 될 때도 일어나며, 중요한 운동시합을 앞두고도 일어 난다. 이러한 상황에서 일어나는 복잡한 신체 반응을 불안반응이라고 특칭 지을 수 있으며, 이것은 우리가 어떤 주어진 과제를 성공적으로 수행하려고 애쓰는 상황에서 흔히 경험하는 반응이다. 이러한 반응을 야기하는 신경 계통을 자율신경 계통이라 하며, 이런 반웅들은 자신의 의지적 통제에서 벗어난 반웅이기 때문에 불수의적(i nvolun t a ry)인 것 이 라고 한다. 이 반응은 자율신경 계 통의 교감신경 계통 (sy m p at h e tic nervous s y s t em) 에 의해 지배된다. 그립 3-2 에서는 자율신경계를 이루 는 두 신경기구, 즉 교감신경 계통과 부교감신경 계통(p arasy m p a t he tic nervous s y s t em) 울 예시하였다. 1.3 .1 교감신경 계통 척수를 따라 좌우로 두 개의 큰 신경줄기가 뻗어 내려오는데, 이 신 경줄기가 바로 교감신경계를 구성한다. 이 줄기를 따라 많은 마디둘이 일정한 간격을 두고 전개되는데, 이 마디둘을 교감신경철 (s y m p ath e tic g an gli a) 이라 한다. 이 신경철에는 축색종말과 수지상돌기가 시냅스를 이루는 세포체들이 모여 있다. 척수에서 나와서 교감신경절로 들어가 는 신경원을 절전신경원(p re g an gli on ic neuron) 이라고 하며, 교감신경절 에서 뻗어 나오는 신경원을 철후신경원(p os tg an gli on i c neuron) 이라 부 른다. 교감신경계를 구성하는 모든 절전축색은 흉부에서 나오는 12 개의 척 수신경들과 요부 중 위쪽 3 개에서 나온 척수신경으로 구성된다. 이 신 경들 가운데 어떤 것은 축색의 결가지, 죽 축색측지를 교감신경 줄기 에 있는 다른 신경절로 보내기도 하며, 또 다른 신경들은 축색측지를
신체 내부에 흩어져 있는 다른 교감신경절로 내보내기도 한다. 교감신 경절에서 나온 축색들은 이 신경원이 지배하는 내장기까지 계속 진행 된다. 앞에서 본 것처럼 교감신경 계통의 활동은 스트레스 상황이나 불안 이 촉발되는 상황에서 두드러 지 게 나타난다. 유명 한 생 리 학자 Walte r Cannon(1871-1945) 은 교감신경계의 전반적 기능이 유기체로 하여금 〈 공격이냐 도망이냐 〉 의 둘 중 어느 하나의 반응을 준비하게 하는 것이 라고 했다. 이러한 준비상태는 짧은 시간 동안 보다 많은 에너지를 이 용할 수 있게 해주고, 심장박동을 증가시켜 주며, 혈액을 장기로부터 근육 쪽으로 보내주어 위기상황에 효과적으로 대응할 수 있게 해준다. 교감신경 계통이 보여주는 〈 공격이냐 도망이냐 〉 하는 이 반응은 부 신 (adrenal gla nd) 으로부터 아드레 날린 (adrenalin e ) 이 라는 호르몬을 분 비하게 한다. 부신은 중심부인 수질 (medulla) 부분과 표피 부분인 피 질 (co rt ex) 로 나누어지는데, 수질 부분은 절전교감신경원의 축색에 의 해 직접 지배된다. 수질 내부에 있는 크롬 친화성 세포 (chroma ffin e cell) 에서는 아드레날린이 만들어진다. 죽 교감신경계의 활동에 의해 크롬 친화성 세포가 활동을 개시하면, 이 세포는 아드레날린을 혈액 내로 분비하게 된다. 실제로 부신수질이 분바하는 분비물은 아드레날 린 (에 피 네프린 e pi ne p hr i ne 이 라고도함) 과 노르아드레 날린 (noradrenaline : 노르에 피네프린 nore pi nep hr i ne 이 라고도 함) 두 가지 이 다. 깜짝 놀랐을 때 우리가 경험하는, 이른바 〈아드레날린 쇄도 (adre nali ne rush) 〉라는 현상은 아드레날린이 혈관, 십장 그리고 기타 여러 조직에 미친 영향에 의해 나타나는 것이다. 다시 말해 아드레날린은 심장의 박동을 보다 빠르고 강하게 하며, 근육이나 간 속에 저장해 놓 은 포도당을 끌어내어 에너지화시킨다. 이와 같은 교감신경계의 활성 은 이른바 비특이성 불안 (nons pecifi c aruc i e ty)이라 부르는 막연한 공포 나 긴장 상태에서 주로 일어난다. 일반적인 추정에 의하면 약 5% 정 도의 사람들이 만성 불안이나 지속적인 교감활동의 항전상태를 보여준 다고 하는데, 이런 긴장 또는 스트레스 반웅의 지속은 편두통, 고혈압
과 같은 심장혈관성 질병, 위궤양, 알레르기, 암 등과 같은 각종 신체 질병을 야기시키는 원인이 된다. 이런 질병을 정신신체 질환(p s y cho somati c d i sorders) 이라 하며, 병원을 찾는 전체 환자의 50-70% 가 바로 이런 질병을 보여주는 환자둘이라 한다. 1.3 . 2 부교감신경 계통 부교감신경 계통은 12 개의 뇌신경 가운데 제 III, VII, IX 및 X 의 4 가 지 뇌신경의 축색에서 나온 절전신경원과 두 종류의 천골척수신경으로 구성된다(그림 3-2). 대부분의 내장기는 교감신경계와 부교감신경계 양 자의 지배를 받는다. 부교감신경계는 신체 에너지의 보존과 생성에 관 련된 활동을 한다. 그러므로 부교감신경계의 활동은 심장박동을 느리 게 하고, 심장수축의 강도를 약화시키며, 위와 장의 운동을 촉진시킨다. 비록 부교감신경계의 활동이 교감신경계의 활동과는 상반되지만, 이 두 신경 계통은 어떤 결과를 얻기 위해서는 서로 협동적으로 작용한 다. 이런 경우의 한 예로 성기의 발기와 사정의 경우를 고려해 볼 수 있다. 남성기는 해면처럼 생긴 성기의 조직 속으로 혈액이 쇄도해 들 어감에 따라 발기하게 된다. 이렇게 들어간 혈액은 혈관을 충혈시켜 성기를 크고 단단하게 만든다. 이러한 과정은 부교감신경 계통의 활동 에 의해 이루어진다. 한편 교감신경계의 활동은 발기 과정을 억제시킨다. 남성들이 겨정 거리가 있거나 두려운 일이 있을 때에는 발기가 잘 되지 않고 또한 발 기 상태가 잘 유지되지도 못하는데, 그 이유는 교감신경계의 기능항전 때문이다. 어떤 일에 대한 실패 경험은 그 일에 대한 재시도에서도 똑 같은 실패가 일어나지 않을까 하는 두려움을 만든다. 이러한 불안은 또 다른 불안을 야기하게 되어, 결국 공포를 야기한 바로 그 일은 계 속 실패를 거듭하는 악순환을 낳는다. 그러나 대부분의 남성은 이와 갇은 실패 사례를 잘 극복할 수 있으며, 과거에 경험했던 성적 흥분이 나 쾌감을 회상하여 발기를 회복시킬 수 있다. 발기는 부교감신경계에 의해 이루어지지만 사정은 주로 교감신경계
에 의해 이루어진다. 사정은 정액주머니 (semi na l ves i cle) 와 전립선(p ro · sta t e g land) 에 있는 불수의근의 수축에 의해 일어난다. 그러므로 교감 신경계의 활동은 성 행동 초기에는 이를 방해하지만 성 행동의 종결 과정에서는 매우 중요한 역할을 한다. 여성의 성적 반응에 있어 부교감신경계의 활동은 음핵 (cl it o ri s) 을 확 대시키고 질의 윤활 작용에 도움을 준다. 여성의 오르가즘은 자궁과 질의 근육들이 주기적으로 수축할 때 일어나는 복잡한 반응이다. 1.3 . 3 불수의적 반응의 수의적 지배 일반적으로 자율신경계는 의식적으로 동제되지 않지만 몇몇 예의가 있다. 예컨대 자율신경계는 방광 기능을 통제한다. 부교감신경계의 활 동으로 방광이 수축되면 배뇨가 이루어지는 반면에 교감신경계의 활동 으로 방광 수축이 방해받으면 배뇨가 억제된다. 이처럼 기본적으로는 배뇨 현상이 자율신경계에 의해 지배되지만 경우에 따라서 수의적으로 소변을 참을 수 있다. 소변을 참을 수 있는 것처럼 심장박동이나 혈압 과 같은 자율신경계의 기능도 수의적으로 통제될 수 있다면 건강의 유 지나 질병의 치유에 큰 도움을 줄 수 있을 것이다. 이러한 자율신경 계, 특히 교감신경계의 기능항전과 관련되어 있는 고혈압이나 편두통 과 같은 정신신체 질병의 치료를 위해 동양에서 수천 년 동안 실천해 온 명상 수련이 매우 효과가 있다는 접이 과학적으로 입증되고 있으 며, 이런 명상 실천은 행동의학의 주요 기법 중의 하나로 간주되기도 한다 (Shapi ro , 1985) . 1.3.4 면역체계와 자율신경계 모든 면역체계에는 자율신경이 분포되어 있다. 면역체계는 신체에 침입하는 박데리아나 바이러스로부터 신체를 방어하는 데 중요한 역할 울 한다. 또한 세 포독성 T 세포 (C yt o t ox ic T-cell) 와 같은 면역 체 계 는 체내에서 발생하는 종양이나 암을 공격하기도 한다. 정서와 같은 십리상태와 질병의 발생 사이에 어떤 관련성이 있지 않
을까 하는 의문은 오래전부터 있었고, 최근에 이 문제에 대한 관심이 고조되 어 심 리 신 경 면 역 학(p s y choneuro imm unolo gy) 이 라는 새 로운 학문 분야가 등장하게 되었다. 이 분야의 역사는 짧지만 많은 과학자들이 신경계와 면역체계 사이의 관계를 밝히기 위한 연구에 열중하고 있다. 면역체계를 구성하는 주요 성분들, 죽 면역세포가 만들어지는 골수 (bone marrow) , T세 포가 성 숙되 는 장소인 흉선 (thymu s gla nd) , 그리 고 T 세포를 비롯한 기타 면역성분을 함유하고 있는 비장 (s p leen) 과 림 프절(lymp h node) 등은 교감신경계와 부교감신경계의 신경지배를 받는 다. 한편 이들 자율신경계의 신경세포들이 뇌의 다른 영역에 있는 신 경세포의 영향을 받기 때문에 우울이나 스트레스에 관여하는 뇌의 상 태가 면역체계에 직접적으로 영향을 미칠 수 있다. 이와 같은 신경계 와 면역체계 사이의 직접적인 관련성이 질병에 어떻게 작용할 것인가 하는 의문은 앞으로 밝혀져야 할 중요한 문제이다. 2 중추신경 계통 앞에서 살펴본 것처럼 말초신경계는 신체 전체에 널리 퍼져 있고, 우리 신체의 활동에 매우 중요한 기능을 하지만, 인간이 가지고 있는 전체 신경원의 1% 미만에 불과하다. 나머지 99% 의 신경세포는 뇌와 척수로 이루어진 중추신경계를 구성한다. 척수는 신경조직으로 만들어 전 하나의 길다란 관(t ube) 처럼 생긴 구조인데, 머리쪽 부분으로 연수 (medulla oblon g a t a) 와 연결되며, 꼬리 부분으로는 척수신경의 제일 아 랫부분인 미골신경 (coccyg e al nerve) 까지 연장된다. 척수는 회백질 (gray matt er ) 과 백 질 (whit e m att er ) 로 구성 되 는데 , 회 백 질은 세 포체 , 수지상돌기, 신경교세포 그리고 축색종말이 모인 곳으로, 여기에는 상 이한 신경핵 집단이 많이 있다. 백질은 수초화된 축색에 의해 만들어 전 것으로서 척수를 오르내리는 신경로 (nerve tr ac t)로 구성된다.
2.1 상행신경로 척수의 상행신경로 (ascend i n g nerve t rac t)는 전의측로 (an t erola t era l), 배측로 (dorsal) 그리고 척수소뇌로 (s pi nocerebellar s y s t em) 로 이루어진 다. 척수에 있는 모든 신경로는 좌우대칭의 양측성 (b il a t eral) 이며, 신 경로 개개의 명칭은 이 신경로가 시작되는 곳과 끝나는 곳의 이름을 순서대로 나타낸 것이다. 예컨대 척수 내에 있는 의축척수시상로 (lat e r al spi no th a lami c tra ct) 는 척 수에 서 시 작하여 시 상까지 상행 하는 신 경 로를 말하고, 척 수피 개 로 (spi no te c ta l trac t ) 란 척수에 서 중뇌 에 있 는 피개로 상행하는 신경로를 지칭하는 것이다. 상행하는 대부분의 신경로는 적어도 3 개의 신경원이 하나의 조를 이 룬다. 첫번째 신경원은 1 차 감각신경원인데, 이 신경원은 척수신경의 후근절 (dorsal root g an gli on) 에 세포체가 있으며, 여기서 나온 축색이 척수 내의 어떤 특정 신경핵과 1 차 시냅스 접촉을 하지만, 때로는 축 색 측지를 다른 신경원에 보내어 시냅스를 하기도 한다. 이렇게 척수 내에서 1 차 시냅스 접촉을 한 후 뇌로 향해 상행하는 신경원이 두번째 단계의 신경원이다. 제 2 단계의 신경원은 척수의 반대편으로 교차하여 시상까지 진행되는 경우도 있고, 척수소뇌로를 구성하는 신경원처럼 교차하지 않는 경우도 있다. 제 3 단계의 신경원은 세포체가 시상의 여 러 핵에 있으며 여기서 나온 축색이 대뇌피질까지 연결된다. 그러나 어떤 상행로는 척수신경의 후근에서 시상까지 직접 전입하기도 하는 데, 이것은 상행신경로가 언제나 3 단계의 신경원으로 구성되는 것이 아니라 2 단계로 구성될 수도 있음을 보여주는 것이다. 2.2 하행신경로 뇌의 여러 영역에 있는 신경원으로부터 하행하는 축색이 척수에 있 는 운동신경원과 시냅스하는 경우가 많은데, 이처럼 뇌로부터 척수로 내 려오는 신경로를 하행 신경 로 (descend ing nerve tra ct) 라 한다. 여 기 에
는 두 개의 주요 하행로가 있는데, 좌측 피질척수로와 우측 피질척수 로 (cort ico spi na l trac t) 가 바로 그것 이 다. 피 질 척 수로의 축색 들은 대 부 분이 연수에서 교차한다. 이 피질척수로가 연수에서 교차하기 바로 직 전에 연수추체 (medullary py ram i d) 를 만들기 때문에 피질척수로를 추체 로 (pyram i da l tra ct) 라고 부르기도 한다. 수많은 신경로를 거 천 축색 들 은 척수 내의 어떤 특정 운동신경원으로 수령되는데, 이 특정 운동신 경원을 전각세포 (ven tr al horn cell) 라 하며, 이곳에는 2, 000-10, 000 개의 시냅스 접촉이 이루어진다. 2.3 뇌의 막 뇌는 뇌막 (men i n g es) 이라 부르는 3 개의 막으로 둘러싸여 있다. 제일 바깥쪽 뇌막을 경막 (dura ma t er) 이라 하는데, 이 막은 질긴 것이 특칭 이며 섬유성의 세포충으로 되어 있다. 가장 안쪽에 있는 막은 경막과 달리 얇고 섭세하여 연막(pi a ma t er) 이라 한다. 경막과 연막의 중간에 있는 막을 지 주막 (arachno i d laye r ) 이 라 하는데 , 소주 (tra becul~e) 라 부르는 작은 기둥 모양의 지지조직 섭유가 거미줄처럼 짜여 있 다. 연막과 지주막 사이의 공간을 지주막하공간 (subarachno i d spa c e) 이라고 하는데, 이 공간은 뇌척수액으로 채워져 있다. 뇌막을 관통하 는 혈관들이 뇌의 활동에 필요한 산소, 포도당 그리고 그 밖의 주요 물질을 공급하고, 뇌의 대사과정에서 발생한 노폐물을 처리한다. 3 뇌속의주요구조 인간의 대뇌피질은 고랑과 이랑 같은 많은 주름을 볼 수 있다. 이 고랑을 해부학적으로 열 (sulcus, 복수로 sulc i)이라 하고, 열과 열 사이 의 이랑 부분을 회 (gyrus, 복수로 gyri)라고 한다. 이 열과 회의 생김새 와 위치는 개인마다 다소 차이가 있지만 일반적으로 크게 다르지 않다.
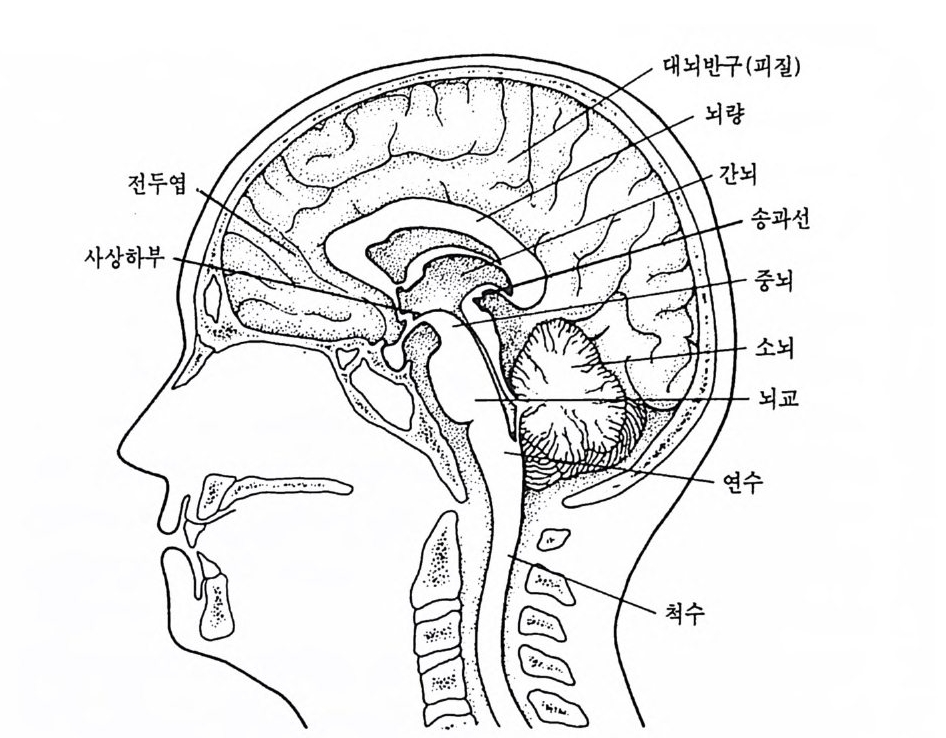 중뇌
중뇌
그림 3-3 인간 뇌의 우반구 내측면 정중선을 따라 두 대뇌반구 사이를 가르면 그림에서 예시된 여러 뇌구조물이 드러난다. 각각의 반구는 전두영, 두정영, 후두영 그리고 측두영과 같이 4 개의 영 (l obe) 으로 나누어진다. 또, 그림 3-3 은 우반구의 내측면 (med i al sur- fa ce) 을 나타낸 것으로, 이를 시상절편 (sa gitt al sec ti on) 이라 한다. 이 단면에서 간뇌 위에 뇌량 (cor p uscallosum) 아라는 백질로 된 거대한 신 경섬유의 다발이 보이는데, 이것은 좌반구와 우반구를 연결하는 축색 다발로 약 200 만 개의 축색이 모여서 만들어진 것이다. 뇌량이 철단된 환자(간찰을 치료하기 위해 간혹 이를 철단하는 경우가 있으며, 뇌량이 잘 린 뇌를 분할뇌라 함)를 대상으로 뇌의 자국정보 처리를 알아본 연구들 에 의해 실로 놀라운 사실들이 밝혀졌는데, 이런 연구성과로 인해 Rog er S perry는 1981 년에 노벨 생리학상을 받았다. 소뇌는 신채운동의 시발, 변경, 지시 및 자세유지 등에 중요한 역할
울 한다. 소뇌와 대뇌 사이에 상구체 (sup er io r co lli cu li)와 하구체 (infe r io r coll icu li ) 그리고 송과선(pi neal g land) 도 있다. 상구체는 시각 계통의 하나로 시각 자국에 대한 안구운동의 방향 정위에 중요한 역할 을 하며, 하구체는 청각 계통의 하나이다. 송과선은 뇌의 중앙에 있는 내분비선의 하나인데, 좌우 한 쌍의 대 칭적인 구조가 아니라 정중선에 있는 단일 구조이다. 이처럼 독특한 송과선의 생김새와 위치가 데카르트에게 매우 충격적인 인상을 주었던 것 같다. 그는 송과선이 마음과 신체가 상호작용을 하는 곳이라고 주 장하였지만, 실제로 송과선은 행스터와 갇은 설치류의 경우에는 생식 주기를 통제하고, 조류의 경우에는 일내활동주기를 통제하는 등 생체 내의 주기적인 기능을 조절하는 일종의 생물학적 시계와 같은 기능을 한다. 시상은 뇌의 중앙부에 있는 것으로 마치 복숭아 씨앗처럼 생긴 신경 구조이다. 이곳은 감각정보의 처리, 운동조철 그리고 기타 전뇌의 기 능을 담당하는 많은 신경원들의 복합체로서, 통각의 지각에도 매우 중 요한 역할을 한다• 시상은 유인원에서 찰 발달되어 있는데 이 구조의 전화는 대뇌피질의 진화와 매우 밀접하게 관련되어 있다. 시 상의 바로 밀에 시 상하부 (h yp o th alamus) 라 부르는 신경구조가 있 는데, 이곳도 신경핵이 밀집된 곳으로 매우 중요한 구조이다. 시상하 부의 전방 경계는 시신경교차 (optic ch i asm) 이고, 후방 경계는 유두체 (ma mmilla ry body) 이 다. 시 상하부에는 체 온 조절이 나 체 내 수분의 조 절, 수면과 각성, 기아와 갈증 그리고 성 행동 등의 조철에 관여하는 신경원과 신경회로를 포함하고 있다. 시상하부 바로 밀에 있는 뇌하수 체 (pituitary g land) 는 내분비선 가운데 가장 중요한 분비선으로 시상하 부와 밀집한 관련이 있다. 시 상과 시 상하부의 전방에는 기저 핵 (basal gan g lia) 이 라 부르는 거 대 한 신경핵 집단이 있는데, 이 부위는 시상이나 기타 뇌 부위에 의해 감싸여 있다. 기저핵에 포함되어 있는 신경핵둘이 모두 같은 기능을 하는 것은 아니지만, 기저핵은 주로 신체 운동의 시발과 조철 기능에
 A
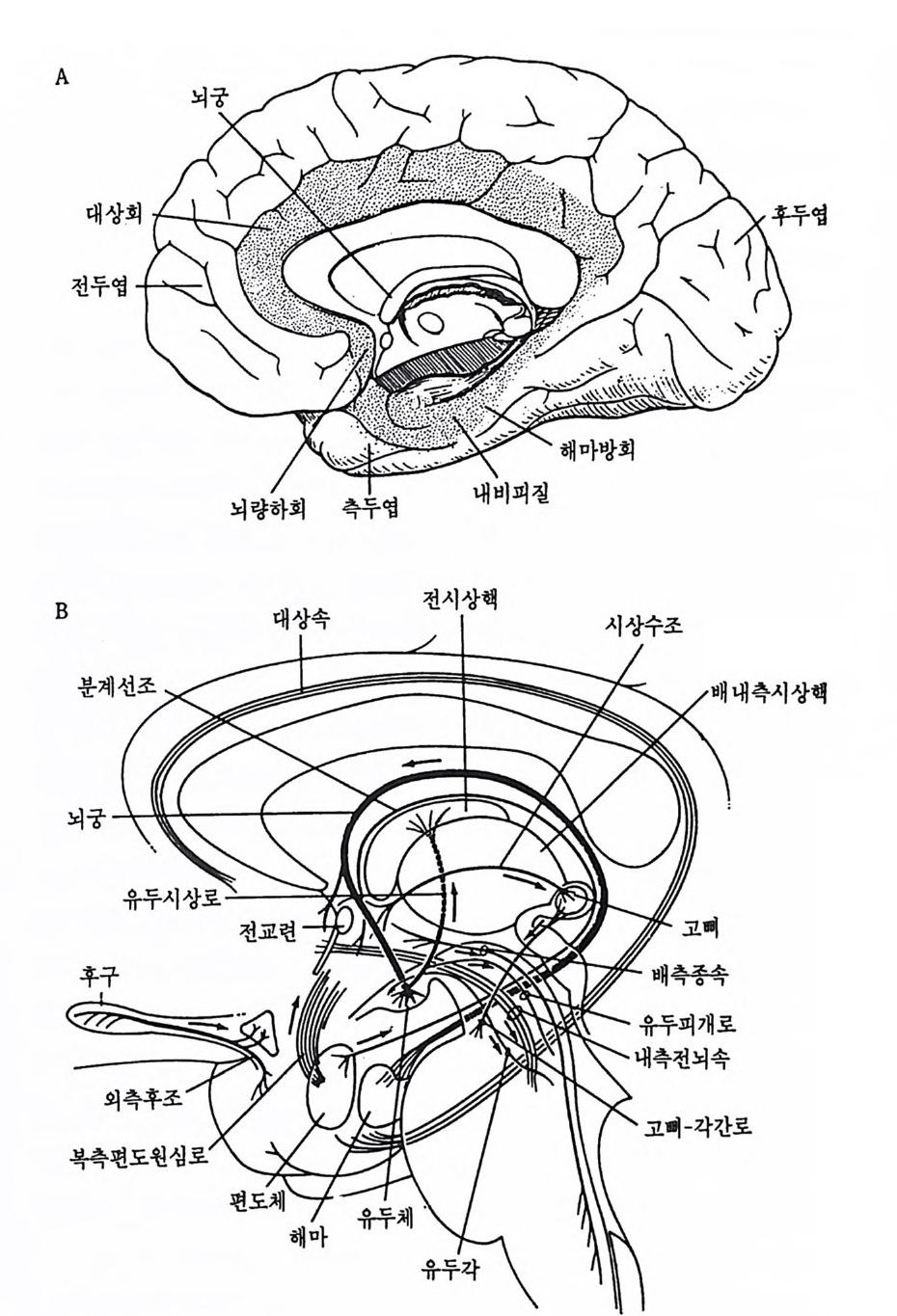
A
그립 3-4 변연계 구조 변연계를 이루는 구조들은 신피질을 비롯한 다른 뇌부위들로 덮여 있다.
관여한다. 시상과 시상하부로부터 미측 방향으로 연수에 이르는 사이 에 뇌간 (bra i n s t em) 이란 곳이 있는데 이곳은 호흡과 감은 생명 유지에 필수적인 기능들을 담당한다. 뇌간의 정중선을 따라 뇌간 망상체 (brain st e m retic u lar fo nna ti on) 란 구조가 있는데 이곳은 각성과 수면, 의식의 변경 그리고 전체 뇌의 통합적 기능과 같은 중요한 심리적 기 능을 통제하는 곳이다.
대뇌피질의 아래쪽에서 뇌간 상부 사이에는 변연계 (limb ic s y s t em) 라 부르는 구조가 있는데 (그림 3-4) , 이 구조에 는 해 마 (hip po camp us ) , 중 격 핵 (sep ta) nuclei) , 편도핵 (amy gd ala) , 대 상회 (cing u la te gyru s) 와 시 상 하부의 유두체 등과 같은 신경핵 집단이 포함된다. 해마는 대단히 복 잡하게 조직화된 구조로서, 마치 바다에 사는 작은 동물인 해마 (sea horse) 를 닮은 듯하여 이러한 명칭이 붙여진 것이다. 해마는 기억의 유지와 저장에 관여할 것이란 견해 때문에 신경과학자들이 많은 관심 을 가지고 연구하는 뇌 구조물이다. 해마는 뇌궁(f om i x) 을 통해 시상 하부의 유두체와 연결되고, 뇌간과도 연결되며 인접 부위의 변연계 구 조인 편도핵과도 연결된다. 편도핵은 측두영의 내면에 있는 신경핵 집 단인데 시상하부 기능과 매우 밀접하게 관련되어 있고, 정서 경험과 표현에 중요한 기능을 담당한다. 3.1 대뇌피질 유인원 뇌의 가장 놀라운 진화적 특칭은 대뇌피질이 고도로 발달되 어 그 크기가 엄청나게 커졌다는 것이다. 특히 인간의 대뇌피질은 다 른 유인원과 비교할 수 없을 정도로 많이 발달되었다. 대뇌피질을 혼 히 신피질 (neoco rt ex) 이라 부르는데 이것은 앞서 본 해마와 같은 구피 질 (old co rt ex) 보다 훨씬 최근에, 죽 늦게 발달된 피질이란 뜻이다. 신피질의 진화 정도와 복잡한 행동의 발달 사이에 어떤 관련성이 있 는가를 알아보기 위해 몇몇 홍미로운 방법들이 제안되었다. 한 예로서 Hein z S t e p han(1972) 이 제안한 〈진화계수(p ro gr ess i on i ndex) 〉라는 개념을 살펴보자. 이 계수는 어떤 주요 뇌 구조물의 진화 정도를 수치로 나타낸 것으로 특정 뇌 영역에 부여하는 수치가 높을수록 이 영역이 다른 뇌 영역에 비해 상대적으로 더 크고 발달되어 있다는 것을 의미 한다. 이 개념에 따른 뇌의 표준수치는 원시적인 식충동물(pri m iti ve i nsec ti vore) 의 뇌를 기준으로 하여 다른 동물의 뇌 진화 정도를 비교하 는 것이다. 이 진화계수에 따르면 인간과 가장 가까운 침팬지의 신피 질은 60 이고, 인간의 신피질은 156 이다. 인간의 신피질이 156 으로 평 가되는 데 비해, 해마는 17, 소뇌는 4, 후구는 1 로 평가된다. 따라서 인간 뇌의 해마와 소뇌는 신피질에 비해 전화 척도상 매우 낮은 뇌 부 위이며, 후구는 가장 원시적인 뇌 부위라는 뜻이 된다. 진화의 압력에 힘입어 신피질은 그 크기가 더욱 커지면서 표면에 고 랑(열)과 이랑(회)이 만들어지게 되었다. 고랑과 이랑으로 주름이 잡 힘으로써 두개골의 용적이 확장되지 않고도 뇌피질의 표면적은 증가될 수 있었다. 인간의 주름잡힌 신피질을 펴게 되면 3,000cm2 정도가 된 다. 뇌피질은 영역에 따라 두께가 1.5- 4.0mm 정도로 조금씩 다르지 만 평균 두께는 2.5mm 정도가 된다. 뇌피질은 부위에 따라 상이한 기능을 통제한다. 20 세기 초 Brod- mann 은 피질의 각 영역을 구성하는 세포들의 구조적인 차이를 바탕으 로 대뇌피질의 여러 영역을 구분하여 표시한 독특한 뇌 지도를 출판하 였다. 그는 뇌피질을 52 개의 상이한 영역으로 구분하고 각기 독특한 구조를 가지고 있는 영역마다 고유한 번호를 부여하였다. 예컨대 시각 의 일차 중추는 17, 이차 중추는 18, 체성감각은 1. 2, 3, 운동 중추는 4 등으로 표시하였다. Brodmann 이 제시한 이 체계는 그 후 많은 연 구들에 의해 타당성이 의심받게 되었지만 뇌해부학의 연구 역사상 매 우 홍미로운 사건으로 간주되어 지금까지 응용되고 있다. 3.2 대뇌피질의 충 대뇌피질은 6 개의 충으로 되어 있는데 표면에서 심부로 들어가면서
다음과 같은 충으로 구분된다. I. 총상충(p lex ifo nn laye r ) : 재일 바깥의 신피질충으로 세포체는 별로 없고 주로 하부층에 세포체가 있는 신경원의 수지상돌기로 구성됨. II. 의과립충 (ex t ernal gran ular lay er ) : 세포체의 모습이 추체를 닮은 작 은 추체신경원으로 이루어전 충. llI. 의추채층 (ex t ernal pyram i dal laye r ) : 중간 또는 큰 추체세포로 이루어 전 충. IV. 내과립충(i n t ernal gran ular laye r ) : 많은 작은 추체세포와 별 모양의 성상신경원으로 이루어진 충. V. 내추체충(i n t ernal pyram i dal laye r ) : 큰 추체세포로 구성된 충. VI. 방추세포충 (s pindle celllaye r ) : 방추체처럼 길다란 모양의 신경세포로 이루어진 충. 추체세포는 성상세포보다 크기가 더 크고, 수지상돌기의 분포도 성 상세포와 다르다. 성상세포의 수지상돌기는 세포체에서 모든 방향으로 골고루 뻗어나가는 경향이 있지만, 추체세포의 큰 정단 수지상돌기 (api ca l dend rit e) 는 세포체의 정접으로부터 뻗어나오는데, 이 돌기에서 나온 분지들은 배측으로 뻗어나가 2 개 이상의 피질충을 넘어가기도 한 다. 한편, 추체신경원의 기저 수지상돌기 (basal dendr it e) 는 세포체의 기저부에서 나와 그 분지를 수평으로 뻗친다. 또 이 세포의 축색은 세 포체의 바닥에서 출발하여 아래쪽에 있는 백질 쪽으로 뻗어나가지만, 축색의 측지들은 옆으로 뻗거나 인접 영역으로 들어가 상행하는 경우 도있다. 신경원의 수지상돌기는 형태에 관계없이 다른 신경원으로부터 둘어 오는 정보를 수용하는 영역이다. 일반적으로 추체신경원의 수지상돌기 는 뇌의 표면을 향해 수칙 방향으로 뻗어나간다. 이 신경원의 정단 수 지상돌기는 제 I 피질층까지 약 2.5mm 정도 뻗어 울라가지만 기저 수지 상돌기는 세포체로부터 200µm 정도밖에 뻗어나가지 못한다. 이에 반
해 성상세포의 수지상돌기는 극히 제한된 좁은 영역에 국한하여 가지 롤 뻗는다. 이처럼 이 두 세포의 수지상돌기나 축색돌기가 뻗어나가는 형태가 서로 다르기 때문에 추체세포는 신피질의 〈 출력세포 (ou tp u t cell) 또는 장거 리 (lon g -d is ta n ce) 세 포 〉 로 간주되 고, 성 상세 포는 어 떤 제한된 영역 안에서 정보를 처리하는 〈 국소회로(l ocal ci rcu it)〉의 기능 울 하는 것으로 간주된다. 3.3 신피질에 대한 입력통로 포유류의 경우 다음과 같은 4 개의 통로를 경유하여 신피질에 정보를 전달한다. 첫번째는 특수감각통로 (s p ec ifi c sensory channel) 이고, 두번 째 는 비 특수 또는 확산동로 (nonspe cific o r diff use channel) 이 며 , 세 번째 는 반대편 반구에서 들어오는 교련통로 (comm i ssural channel) 이고, 네 번째는 동일한 반구에서 들어오는 반구내통로(i n tr ahe mi s p her ic channel) 가 그것들이다. 특수감각통로는 시각이나 청각과 같은 특정 감각신경원의 축색으로 구성되며, 이 축색의 세포체는 시상의 여러 핵에 있다. 이 축색의 대 부분은 피질 제 1V 충에 있는 추체세포에서 시냅스한다. 그러나 그 중 어떤 축색의 종말은 제 III 층의 바닥에서 발견되는 수도 있으며, 어떤 축색의 종말은 성상세포와 시냅스하기도 한다. 비특수 또는 산만통로의 세포체는 뇌간 망상체에 있는 신경원으로부 터 울라오는 정보를 중개한다. 이 세포의 축색은 짧은 측지를 가지며 축색종말은 모든 신피질 영역의 어느 충에서나 발견된다. 이 신경의 구심성 흥분은 대뇌피질 활성화의 정도를 조절하며, 수면으로부터 각 성에 이르는 다양한 의식상태를 매개한다. 뇌간에 있는 청반(l ocus coe - ruleus) 이 이 계통의 주요한 부분이 다. 교련통로는 한쪽 뇌반구에서 다른 쪽 뇌반구로 교차하는 축색다발인 데 전교련 (ante r io r commi su re) , 후교련 (po ste r io r com misu re) 그리 고 뇌 량 (corp u s callosum) 등이 이 에 속한다. 대부분의 경우 두 반구 사이 의
연결은 대칭적이어서 거울에 상이 비치듯이 두 반구의 같은 부위가 서 로 연결되지만 대칭적으로 연결되지 않는 경우도 간혹 있다. 반구내통로는 같은 반구 내에 있는 신경원들 사이의 전달동로이다. 때때로 이 통로는 인접하는 신경원들의 축색측지로 이루어지기도 한 다. 어떤 신경원은 축색측지를 통해 자체의 활동에 간접적으로 영향을 미칠 수도 있다. 이런 영향은 하나의 신경원이 축색측지를 간신경원 (i n te rneuron) 으로 보내 시냅스하고, 간신경원에서 나온 축색이 다시 원래의 그 신경원과 시냅스함으로써 가능하게 된다.
제 4 장 뇌의 발달과 노화 인간의 신체와 뇌를 구성하는 모든 세포들은 하나의 수정란으로부터 만들어진다. 신생아도 성인과 같이 약 1,000 억 개의 뇌세포를 가지고 있는 것으로 추정된다. 뇌가 성장하고 발달하는 과정을 고려해 보면 몇 가지 홍미로운 의문을 제기할 수 있다. 즉, 어떻게 개개의 신경원 들이 서로 연결되어 적절한 기능을 유지할까? 신경연락이 적절하게 이루어지지 못하면 어떤 일이 일어날까? 어떻게 수많은 신경원들이 뇌 속의 자기 위치를 찾아 이동해 갈까? 바로 이런 문제가 이 장에서 다룰 주요 내용이다. 일반적으로 포유류의 뇌 속에 있는 신경원은 출생 후에는 더 늘어나 지 않으며, 후구세포를 제의한 다른 뇌세포는 죽은 후 새로운 세포로 대치되지도 않는다. 그럼에도 불구하고 뇌 손상 이후에도 손상된 세포 나 인접 세포에서 다양한 반응들이 나타나는데, 이러한 재생 반응은 새로운 축색종말이 돋아나거나 광범위한 시냅스 변화 때문이다. 비록 전 생애를 통해 서서히 성장하는 신경원도 있지만, 손상 후 신경회로 가 정상상태로 회복된다는 것은 포유류의 뇌에서는 불가능한 것으로 알려져 있다. 인간의 뇌세포 가운데 상당수는 생애를 통해 죽어가지만, 이 과정은 일반적으로 서서히 그리고 미약하게 일어나기 때문에 급격한 뇌 손상 이 없는 한 우리의 심리과정이나 행동과정에 갑작스런 변화가 일어나
지 않는다. 노인들에게는 심한 뇌 손상이 찰 일어나는데, 알츠하이머 병이라는 뇌 손상은 노인에서 잘 나타나는 대표적인 뇌질병이다. 이 병에 걸린 노인들은 정신적으로나 정서적으로 큰 장애를 보여준다. 1 뇌의 발달단계 태아기 동안 인간의 뇌는 다음과 같은 8 단계를 거치면서 발달된다. 1) 신경판 (neural p la t e) 의 유도(i nduc ti on) 2) 신경원과 신경교가 될 세포의 증식 (pro li fer ati on ) 3) 신경원과 신경교세포가 최종적으로 위치할 장소로 이동 (m igr a ti on) 4) 특정 회로를 형성할 신경원들의 응집 (ag greg a t i on ) 5) 신경 원이 특정 형 태로 발달되는 분화 (d iff eren ti a ti on) 6) 다른 세 포들과 기 능적으로 연 결되는 시 냅스 발생 (syn a p tog e n esis ) 7) 목적지에 도달하지 못한 신경원들이 소멸되는 선택적인 세포 사망 (selecti ve cell death ) 8) 사용 또는 불사용으로 인해 시냅스 연결이 안정되거나 배제되는 기 능적 안정 화 (fun cti on al vali da ti on ) 위와 같은 8 단계의 발달을 거쳐 뇌가 성장하는데, 다음에 각 단계들 울 보다 자세하게 살펴보자. 1.1 신경판의 유도 임신 후 처음 3 주 동안 장차 신경 계통이 될 조직들이 배아의 배측 표면에서 하나의 세포판을 형성하기 시작하는데 이 세포판을 신경판 (neural p la t e) 이라 부른다. 이 신경판이 형성되고 나면 이 판을 구성하 는 세포들과 이 판에서 생성되는 세포들이 장차 뇌세포로 발달된다.
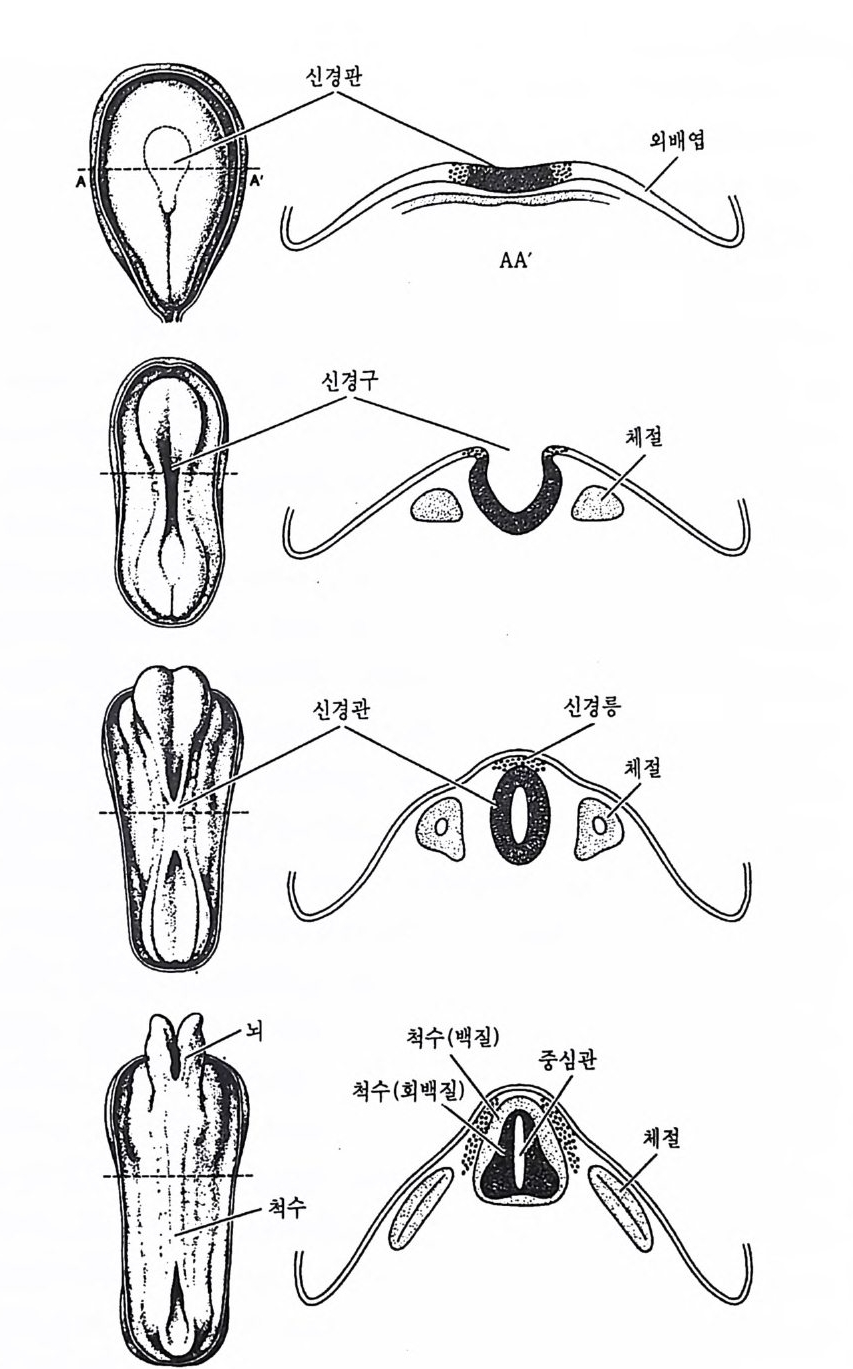 -A
-A
그림 4-1 배아기의 뇌발달 임신 18 일째 신경판이 형성되며, 21 일째 신경구가 보이기 시작하고, 24 일째 신경관이 형성되며 주변에 신경릉이 이루어진다.
신경판이 형성된 칙후 이 판의 좌우에 한 쌍의 두둑이 솟아나게 된다 (그림 4-1). 이 두둑이 접점 자라면 그 사이에 신경구 (neural gr oove) 라 부르는 깊은 홈이 생기며, 두둑이 더욱 자라 서로 맞물리게 되면 그 사이에 신경관 (neural t ube) 이 생기게 된다. 이때 이 신경관은 약 12 만 5 천 개의 세포로 구성되며 이 관이 신경 계통의 바탕이 된다. 이 신경 관의 주변에서 양적인 변화는 계속되지만, 기본형태는 그대로 유지된 다. 즉 척수의 중앙에는 척수관이, 뇌 속에는 몇 개의 뇌실이 나타난다. 어떤 신경세포들은 산경관으로부터 떨어져 나와 신경롱 (neural crest) 울 만들게 되는데, 이 신경릉이 대부분의 말초신경 계통과 부신수질을 이루는 세포들의 발생 장소가 된다. 1.2 세포증식 신경관이 처음 만들어졌을 때는 약 12 만 5 천 개 정도의 세포를 갖지 만, 출생시 인간의 뇌는 적어도 천억 개 이상의 신경세포로 구성된다. 이 숫자는 태아가 자라는 270 일 동안 매분 약 25 만 개의 세포가 평균 적으로 증식된다는 것을 의미한다 (Cowan, 1979). 대부분의 경우 포유 동물의 뇌에 있는 신경원은 출생 직전이나 직후 중석되지 않지만, 쥐 나 고양이의 경우에는 출생 후 어느 일정 기간 동안 뇌세포가 다소 늘 어나기도 한다 (Al tm an, 1967). 그러나 유인원의 뇌에서 새로운 신경원 이 만들어지는 데는 분명한 한계가 있다. 예컨대 Pasko Rakic (1 985) 에 의하면 레서스 원숭이의 경우 소뇌나 해마를 제의한 어떤 뇌 부위 에서도 출생 후에는 신경원이 만들어지지 않는다고 하며, 안간의 경우 에도 레서스 원숭이와 유사한 결과를 보여준다고 한다. 세 포분열 주기 가운데 유사분열 주기 (mi tot i c cy cl e) 는 4 개 의 분명 한 시기를 갖는다. 즉 유사분열기 (mi toti c p hase) 에는 하나의 세포가 두 개의 딸세포 (dau gh t er cell) 로 분열기가 있고, 다음에는 유사분열후간기 (po stm itot ic g a p)라 부르는 휴지기가 있다. 많은 세포들은 이 시점에서 분열을 멈춘다. 다시 분열을 계속하는 세포는 DNA 를 합성하는 S 기 (S
p hase) 로 들어가게 되며, S 기 다음에 유사분열전간기 (pre mi toti c gap ) 가 따르고, 그 후 다시 유사분열이 이루어지는 유사분열 주기가 되풀 이된다. 세포증식 과정은 많은 종의 동물들을 대상으로 연구되어 왔으며, 그 결과로 다음과 같은 일반화가 가능하게 되었다(Ki mble , 1988). 첫째, 망막의 절세포(g an gli on cell) 와 해마와 뇌피질에 있는 추체세 포처럼 덩치가 큰 신경원의 증식은 양극세포나 과립세포와 같은 비교 적 작은 신경원의 증식보다 먼저 일어난다. 일반적으로 큰 신경원은 긴 축색을 가지는 경향이 있으며, 실제로 세포체의 크기와 축색의 길 이 사이에는 상관이 있다 . 이러한 까닭으로 작은 신경원은 짧은 축색 울 가지며, 장래 〈 국소회로(l ocal ci rcu it) 〉 를 이루게 된다. 이와는 달리 보다 큰 신경원은 긴 축색을 가지며 먼 거리를 연결하는 기능을 갖는다. 둘째, 뇌 속에서 진행되는 대부분의 세포증식은 신경관의 내벽을 이 루는 충에서 일어난다. 뇌의 개개 영역은 그 영역 자체의 고유 발달양 식에 따른다. 예컨대 대뇌피질의 경우, 가장 먼저 태어난 신경원들이 대뇌피질의 가장 깊은 내부층을 이루며, 가장 늦게 태어난 신경원이 가장 바깥의 표피충을 이룬다. 그러나 망막의 경우는 이와 달리 먼저 태어난 절세포가 바깥 표피층을 차지하며 그 후에 태어난 간신경원과 수용기가 내면충을 형성한다. 1.3 세포의 이동 기능분화가 일어나기 전의 신경원을 신경아세포 (neuroblas t)라고 한 다. 분열이 끝난 신경아세포는 궁극적으로 자신들이 자리잡아야 할 위 치로 이동하기 시작한다. 정확하게 어떤 요인이 이런 이동 과정에 관 여하는지에 대해서는 아직 찰 모르지만, 아마도 이 시기는 유사분열기 에 세포들이 증식되어 밀려나오는 것과 밀접하게 관련되어 있을 것으 로 추측되며, 이런 세포 방출이 곧 세포의 이동을 야기시킬 것으로 생 각된다. 신경아세포의 이동은 마치 아메바가 이동할 때의 모습과 흡사
하다. 즉 신경아세포의 한쪽 부분이 먼저 이동할 방향으로 뻗어나오 고, 이어 나머지 세포 부분이 끌려가는 모습을 취한다. 그러면 이런 이동성 신경아는 자신이 어디로 가야하는가를 어떻게 알게 될까? 예컨대 대뇌피질의 경우, 모든 신경아는 피질의 여러 충 가운데 운명적으로 결정된 특정 장소로 이동하기 위해 그 세포가 태어 난 곳에서 최소한 수 mm 를 이동해야 한다. 따라서 이 신경아들은 자 신이 가야할 정확한 피질충을 발견해야 하며, 그곳에 가서도 다른 신 경원들과 적절한 시냅스 접촉을 하기 위해 스스로 방향을 잡지 않으면 안 된다. 이동하는 피질 신경아는 특정 형태의 신경교세포, 즉 방사신 경교 (rad i al g l i a) 에 의해 인도된다. 방사신경교세포는 뇌가 형성되는 태아기에만 존재할 뿐 임신 종기가 되면 사라져 버린다. 이 신경교는 신경아가 최종 목적지를 향해 이동해 갈 때 마치 버팀목과 같은 작용 울한다. 방사신경교와 소뇌 구조 사이의 관련성에 관한 주목할 만한 연구사 례들이 발표되었는데, 그 중 한 예로 〈 위버 생쥐 (weaver mouse) 〉 란 사 례가 있다. 위버 생쥐는 신경학적으로 돌연변이가 일어난 생쥐인데, 걸음걸이가 좌우로 왔다갔다 한다는 (weav i n gg a it) 뜻에서 그 이름이 유래된 것이다. 위버 생쥐는 태아기 동안 방사신경교세포가 너무 일찍 사라졌기 때문에 이동하는 피질 신경아세포에 적절한 버팀목 기능을 마련해 줄 수 없었다. 따라서 많은 신경아세포가 소뇌의 특정 위치까 지 이동해 가지 못하며 정상적인 운동 수행에 필요한 시냅스 접촉이 이루어지지 못한다 (Rak ic & Sid m an, 1973). 지금까지 신경학적으로 돌연변이를 일으킨 생쥐 혈통이 자그만치 140 여 종에 달하는 것으로 알려졌는데, 이들 가운데 상당수는 신경아 가 이동할 때 방향을 상실하여 일어난 사례들이다. 그러나 이런 신경 원의 위치가 일탈된 변위성 (ecto p ia) 신경원들 가운데는 선택적 세포 사망 시기(다음 항에서 설명) 동안 이동된 것들도 다소 있다. 정상적인 뇌의 경우에도 이동시기 동안 신경아세포들 가운데 길을 잃어버리는 경우가 3% 나 된다. 이처럼 신경아세포가 적절한 장소로 이동하지 못
해서 생긴 사례가 인간의 뇌에서도 발견되는데, 이런 신경장애 사례에 서는 여러 가지 비정상적 증후가 관찰된다. 예컨대 심한 독서장애를 보이는 실독증 (d y slex i a) 환자의 경우 언어 이해에 중요한 기능을 하는 좌반구의 특정 뇌부위의 세포들이 비정상적으로 배열되어 있음이 관찰 되 었 다 (Geshwi nd & Galaburda, 1985) . 1.4 세포의 응집 이동하는 신경원은 적절한 장소로 이동해야 할 뿐만 아니라 이동하 는 동안 적절한 방향성을 유지해야 하며, 또 이동된 장소에서 다른 세 포들과 기능적으로 적절하게 연결되어야만 한다. 이런 과정을 응집 (ag greg at i on ) 과정 이 라 부른다. 이 과정 에서 각각의 세 포들은 상대 방 세포를 〈 인지 (co gniti on) 〉해야 하는데, 이때 각 세포는 다른 세포의 막 단백질을 인지하는 방법을 취할 것으로 추측된다. 망막, 해마, 소뇌 또는 대뇌피질 세포와 같은 어떤 특정 형태의 신 경원들을 동물의 뇌에서 떼어내 조직배양액에 옮긴 후 이 신경원들을 분리해 두면, 분리된 신경원들은 각기 고유한 형태의 신경원들끼리 재 응집하게 된다 (Ku ffl er 등, 1984). 이 사실은 곧 태아의 뇌에도 이와 유 사한 기제가 있어서 특정 형태의 세포들이 뇌의 특정 층과 신경핵 집 단으로 응집할 것이란 점을 시사해 준다. 이런 응집이 적절하게 이루 어지지 못하면 앞서 본 위버 생쥐와 유사한 신경학적 돌연변이가 생길 것으로 추리 한다. 한 예로 릴러 생 쥐 (reeler mouse) 사례가 보고되 었는 데, 이 생쥐는 걸음걸이가 비틀거리기 (reel) 때문에 그와 같은 이름을 갖게 되었다. 이 생쥐의 피질세포를 조직배양하면 정상적인 생쥐의 피 질세포와는 달리 재응집이 일어나지 않는다고 한다• 1.5 세포의 분화 한 신경원이 그 신경원의 최종 목적지에 도달하게 되면, 이 신경원
은 특정한 형태의 모습, 죽 독특한 형태의 수지상돌기와 특정 수의 축 색분지 그리고 그 신경원 고유의 화학적 전달물질을 갖게 된다. 이 과 정 울 세 포의 분화 (dif fere nti at i on ) 라고 한다. 축색 이 나 수지 상돌기 의 신 경 성장은 신경원 가지의 끝에 있는 성장원추(gr o wt h cones) 의 발달을 통해 이루어진다. 그림 4-2 에는 성장원추가 예시되어 있는데, 성장원 추는 지금부터 약 1 세기 전 스페인의 유명한 신경해부학자 Ca j al 에 의 해 최초로 관찰되었다. 그는 이 성장원추를 발견하고 : 몹시 흥분하여 이 원추를 가리켜 〈 앞으로 뻗어나가면서 부딪치는 장애물을 기계적으 로 제거하는 부드럽고도 탄력성 있는 일종의 돌격무기, 예컨대 성벽을 부수는 기구와 같은 것 〉 이라고 했다 (Ram6n y Caja l, 1917, Kim ble, 1992 에서 인용).
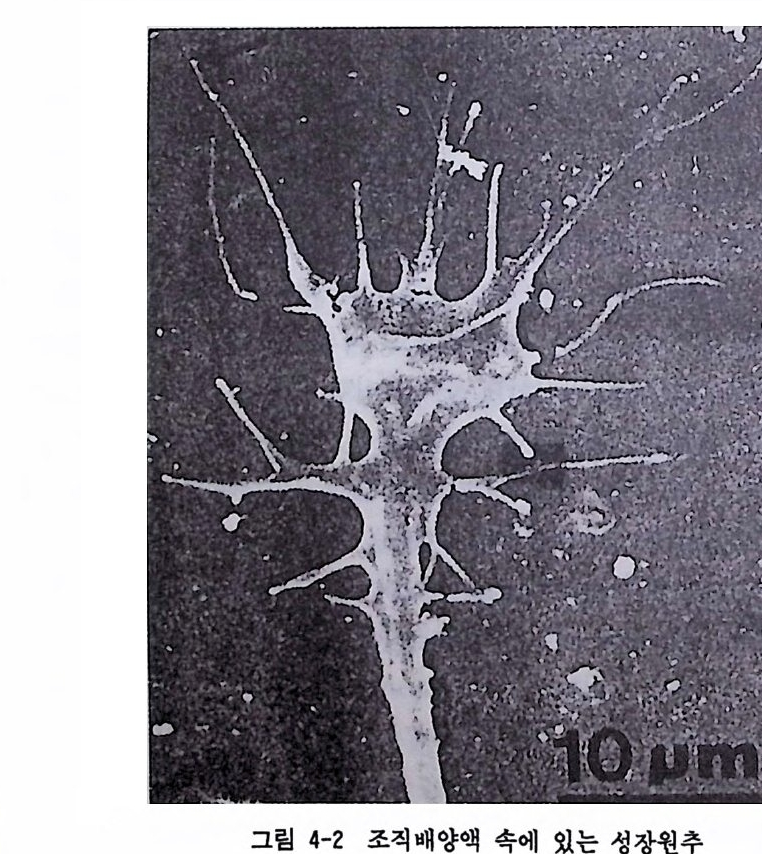 그립 4-2 조직배양액 속에 있는 성장원추
그립 4-2 조직배양액 속에 있는 성장원추
축색과 수지상돌기의 성장원추를 조직배양액 속에 넣고 관찰해 보면 Ca j al 이 언급한 것처럼 매우 힘찬 모습으로 연장과 수축을 거듭하며 앞으로 뻗어나간다. 축색이 배양액 속에서 뻗어나갈 때, 다른 세포와 접촉하는 부위를 떼어내어 관찰했을 때 어떤 물질은 취하고 어떤 물질 은 버린다는 점이 발견되었다. 축색의 성장원추가 적절한 접촉대상을 찾으면 바로 그 접촉 세포와 시냅스를 이루게 된다. 어떻게 신경원들 이 적절한 접촉대상 세포를 인지하게 되는가는 명확하지 않지만 성장 원추와 접 촉하는 세 포의 막에 특정 한 〈 인 지 물질 (recog nition mole- cules) 〉 이 있을 것으로 추측되며, 이 물질이 적절한 시냅스를 형성하는 데 도움을 줄 것으로 예상된다 (Dodd & Jess ell, 1988). 1.6 뇌세포 발달과 환경요인 한 신경원의 성장에 영향을 미치는 마지막 요인은 유전적 요인이다. 그러나 이러한 유전적 요인도 성장하는 세포 주변의 환경요인들에 의 해 조정된다 . 영양실조와 같은 환경조건이 세포의 성장을 변경시킬 수 있는데, 그 이유는 적절한 영양소가 없으면 축색의 수초화가 잘 이루 어지지 않기 때문이다. 인간의 경우 출생 후 여러 해에 걸쳐 수초화가 진행되기 때문에 유아기의 영양실조는 신경 계통의 발달에 영속적인 영향을 미친다. 그러나 이러한 초기 장애가 회복될 수 있다는 증거도 있다. 예컨대 어린 시절에 심한 영양실조를 경험한 한국전쟁의 고아들 이 미국 가정으로 입양되어 적절한 영양 공급을 받은 결과 학교성적과 지능에서 미국의 정상아동 이상으로 회복되었다는 사실이 보고되었다 (Rosenzweig & Leim an, 1982) . 호르몬의 불균형도 뇌 발달에 영 향을 미 친다. 크레 틴 병 (creti ni s m ) 에서 볼 수 있는 신경 계통의 십한 장애는 주로 갑상선의 부적철한 분 비 때문에 생긴다. 이 의에도 많은 물질들이 태아 발달에 십각한 장애 를 일으킨다고 한다. 그 중 대표적인 물질로 알코올을 둘 수 있다. 알 코올은 신경 계통의 이상을 야기하며, 지적 기능이나 행동에도 이상을
야기 한다. 태 아알코올증후군(f e t al alcohol syn d rome : FAS) 은 1973 년 Kenneth J ones 와 Davis Sm it h 에 의 해 처 음 명 명 된 증후인 데 , 이 중후 는 어머니가 아기를 임신하고 있는 동안 알코올을 과다하게 섭취하여 생긴 것으로서, 출생 후 아기는 신체적으로나 정신적으로 심한 장애를 보여준다. 장애의 정도는 어머니가 섭취한 알코올의 양과 섭취 시기에 따라 다양하게 나타난다. 인간의 경우 대표적 인 증상은 I Q가 40-50 에 불과한 정신지체를 비롯하여 과잉활동성 (hyp era cti vit y ) 그리고 경우에 따라서는 우울 증상도 보여주며, 안구가 비정상적으로 작고, 코와 입 술 사이의 인중이 지나치게 긴 안면 표정과 뇌의 크기가 정상에 비해 현저 히 축소된 소뇌 증 (mi cre nceph a ly 또는 mic ro enceph a ly) 을 나타내 기 도 한다. 임신중인 흰쥐에게 알코올을 섭취하게 하면, 이 어미로부터 태어난 새끼둘은 어떤 반사작용(예컨대 등을 바닥에 닿게 눕혀진 동물이 스스로 일어나는 반사)의 성숙과 눈울 뜨는 시기가 느려지며, 이유 직 후에 행한 좌우변별 학습에도 장애를 보인다고 한다. 한편 임 신중인 쥐 에 게 MAM (meth yla zoxy m eth a nol aceta te ) 이 란 물 질을 두여해도 신경 계동의 발달이 억제되어 소뇌증이 나타난다. 장현 갑, 정봉교 그리고 박순권(1 988) 의 연구에 따르면, 출생 전에 MAM 에 노출된 쥐의 뇌 크기는 정상 뇌의 60% 정도로 작아지고, Hebb- Wi lliam s 미로학습의 장애와 과잉활동성을 보였다. 또한 출생 전에 MAM 에 노출된 경험을 가진 동물들은 스트레스에 대한 반응성도 달 라진다. 죽 정상적인 동물들은 약한 스트레스를 받은 후 전기충격에 대한 도피 수행에 결함을 보이지 않지만, 출생 전에 MAM 에 노출되 어 신경계가 손상된 동물들은 스트레스의 강약에 관계없이 전기충격 직후의 도피 수행에 심한 결함을 보였다 (Chan g 등, 1989). 이상과 같이 신경계의 구조적 기형과 기능적(행동적) 장애를 관련시 켜 연구하는 분야를 행동기형학 (beha vi oral t era t olo gy)이라고 하는데, 이 분야는 신경과학 영역에서 발달 장애의 생물학적 원인을 규명하는 최신 연구영역의 하나이다.
1.7 신경전달물질의 선택 분화하는 산경원이 취해야 하는 중요한 〈 결정 〉 중 하나는 어떤 종류 의 신경전달물질을 선택하는가 하는 문제이다. 개개의 신경세포는 유 전적 지시에 의해 여러 가지 또는 모든 신경전달물질을 생산할 수 있 도록 계획되어 있지만, 실제로 각 신경원이 분화된 후에는 한 가지 전 달물질만 분비하는 것이 보통이다. 자율신경계의 신경원을 대상으로 한 실험들에 의하면 분화하는 신경원을 둘러싸고 있는 주변 환경이 이 신경원이 분비하는 분비물질의 종류를 결정하는 데 중요한 역할을 한 다고 한다 (Patt er son, Pott er, & Furshpa n , 1978) . Pa tt erson 과 그의 동료 둘의 연구에 의하면, 배아기에 있는 쥐의 신경릉에 있는 세포를 떼어 내어 비신경세포인 다른 체세포와 섞어 조칙배양하면 이 신경릉의 세 포들이 콜린성 (cho li ner gic) 기능__미주신경을 거쳐 십장박동을 느리 게 하는 기능_을 보인다고 했다. 한편 이 세포를 다른 체세포와 섞 지 않고 자연스럽 게 그냥 두면 노르아드레 날린성 (noradrenalin e rgi c) 기능_정상적으로 심장을 지배하는 흥분성 신경원을 통해 십장의 박동을 증가시키는 기능 _ 울 보인다고 했다. 따라서 이 결과들은 조직배양액 속에 들어 있는 세포의 종류에 따라 원래는 노르아드레날 린성 신경기능이었던 것이 콜린성 신경기능으로 변화될 수 있음을 시 사하는 것이다. 1.8 시냅스 발생 뇌의 활동은 시냅스 회로를 통해서 나타난다. 시냅스 발생 (s yn a pt o g enes i s) 이라 부르는 시냅스 형성시기에는 분화중인 신경원이 다른 신 경원과 서로 상호작용함으로써 적절한 상대 세포는 인지하고 부적절한 상대 세포는 배제한다. 이 시기에 형성되는 모든 시냅스가 영구적으로 생존하는 것은 아니며, 또한 시냅스 발생은 출생 후 뇌가 성장 • 발달 하는 동안에도 계속된다. 임신 기간이 비교적 짧은 동물의 경우는 출
생 후에도 상당한 양의 시냅스 발생이 이루어진다. 예컨대 쥐와 같은 동물은 출생 후 12 일에서 30 일 사이에 시냅스 발생 수가 무려 10 배나 더 증가한다고 한다 (Ja cobson, 1978) . 이처럼 쥐나 고양이의 경우는 출생 후 일정기간 동안 새로운 신경원 의 생성으로 뇌의 회로가 더욱 풍성해지고 복잡하게 된다. Al t man 과 그의 동료들(1 967) 은 출생 후 어느 일정기간 동안 대뇌의 특정 영역에 서 세포분열이 일어나고 있음을 방사성 촬영으로 증명하였다. 이렇게 출생 후 새로 형성되는 신경아세포들은 세포체가 작고 축색이 짧은 이 른바 〈미세신경원 (m ic roneuron) 〉 인데 이 세포들은 해마, 후구 또는 소 뇌를 구성하는 신경회로 속에 통합된다. 이처럼 고양이나 쥐의 해마, 후구 또는 소뇌에서는 출생 후에도 새로운 신경원의 가세나 대치가 상 당 기간 동안 계속된다. Al tm an 은 소뇌에서 새로운 신경원의 가세가 성숙한 동물의 운동기능 발달과 관련 있다고 시사했지만, 이러한 출생 후 신경원의 증가가 무엇을 의미하는지는 아직 잘 알려져 있지 않다. 1.9 화학적 친화력 가설 어떻게 신경원이 정확하게 시냅스 접촉을 이루어갈까? 예컨대, 망 막에 있는 절세포의 축색이 어떻게 뇌 속으로 진행하여 적절한 신경구 조와 시냅스 접촉을 이룰 수 있을까? 포유류와 척추동물의 시각 계통 이 질서정연한 점 대 점 (po in t to p o i n t)의 대응방식으로 조직화되어 있 다는 것은 이미 많은 연구자들에 의해 증명되었다. 비록 이 문제에 대 한 논의는 지금도 계속되고 있지만, Rog er S p e rry (1963) 에 의해 제안 된 화학적 찬화력 가설 (chemoa ffi n ity h yp o th es i s) 이 이 문제를 설명하는 데 가장 설득력 있는 가설이다. Sperry는 시냅스 접촉이 선택적으로 이루어지는 이유는 개개 신경원 의 화학적 기능분화 때문이라고 주장하였다. 이 가설에 따르면, 개개 의 신경원은 그 자체의 독특한 화학적 동질성을 발달시켜 나간다고 한 다. 그에 의하면 망막에서 뻗어나온 개개의 절신경원은 뇌 속에서 이
신경원과 화학적으로 가장 적절한 신경원을 찾아내어 이와 짝짓게 된 다는 것이다. 화학적 친화력 가설을 지지해 주는 근거들은 주로 개구 리의 시개 (op tic t ec t um) 에 있는 신경원을 손상시킨 후 이 신경원이 재 생되는 모습을 관찰한 연구에서 얻어졌다. 개구리와 같은 양서류는 신경조직의 재생력이 놀랄 정도로 뛰어난데 이러한 재생능력은 어린 동물에서는 말할 것도 없고 성숙한 동물에서 조차도 현저하다. 포유류의 경우는 시신경을 절단하면 그 후 영구적으 로 완전한 시맹이 되지만 , 개구리나 도마뱀과 같은 양서류는 비록 시 신경이 절단되어도 재생이 가능하여, 이 동물의 시각피질에 해당되는 시개의 신경원과 새로운 시냅스 접촉이 이루어진다. 개구리의 경우 이 러한 시각 재생이 일어나는 데는 약 한 달 정도가 소요된다. 즉 이들 의 시신경을 절단한 후 정상 위치에 놓아두면 한 달쯤 지나 정상적으 로 시각이 회복된다. S p e rry는 이러한 개구리의 시신경 재생능력을 이용하여 여러 가지 실험을 수행하였다. 예컨대 한 실험에서 그는 시신경을 차단하고 개구 리의 안구를 180 도 회전시켜 망막의 배측 부분이 복측 부분이 되도록 위치를 뒤바꾸었다. 다시 말해 망막의 앞쪽 반이 안구의 뒤쪽 반이 되 도록 위치를 바꿔놓은 셈이다. 이런 수술을 하고 나서 한 달 정도의 회복기간을 주어 시신경과 시개 사이의 연결이 재생될 수 있게 한 후, 시야의 여러 방향과 위치에 파리와 같은 미끼를 제시하여 이에 대한 반응을 알아봄으로써 개구리의 시각이 어떻게 달라졌는가를 관찰하였 다. 이렇게 수술된 개구리는 미끼가 앞쪽으로 다가왔을 때는 반응하지 않고, 안구가 180 도 회전된 뒤쪽 방향에서 다가왔을 때 정확하게 반응 하였다. 이 동물은 계속 180 도 회전된 시야를 보였는데, 이런 반응은 수백 번의 재학습을 시켜도 교정되지 않았다. 이런 현상은 다음과 같은 사실을 시사한다. 죽 재생된 절세포의 축 색이 시개를 향해 뻗어간 후, 절단 전에 시냅스 접촉을 맺었던 시개의 같은 장소에서 시냅스 접촉이 새로 형성되었다는 사실이다. 그러므로 이러한 새로운 위치, 다시 말해 망막의 앞쪽 반에서 들어온 정보가 이
제는 망막의 뒤쪽 반에서 들어온 정보로 해석되어 뒤쪽에서 접근하는 미끼에 대해서만 반응하게 된 것이다. 위와 유사한 연구결과가 안구를 좌우로 서로 맞바꾼 경우에서도 일어났다. 이런 사실들은 곧 재생하는 신경원이 시개에서 처음 접촉했던 시냅스 위치로 되돌아온다는 것을 의미하는 것이다. 1.10 선택적 세포의 죽음 수십억 개의 신경세포가 임신중의 발달과정 동안 죽어간다. 이러한 방대한 신경세포의 죽음은 정상적 발달과정의 한 현상이다. Cowan 등 (1984) 에 의하면 태아기 동안 생성되는 모든 신경원들 가운데 약 50% 가 출생 전에 죽는다고 한다. 출생 전에 이렇게 많은 세포들이 정 상적으로 성장하지 못하고 죽고 만다는 사실은 낭비적인 것처럼 보이 지만 이러한 선택적인 세포의 죽음은 신경회로의 조직화에 도움이 될 수도 있다. 시냅스 접촉을 울바르게 하지 못하는 신경원일수록 손상되기 쉽고 살아남지 못할 가능성도 높다. 대부분의 경우 세포의 생존은 한 신경 원으로부터 다론 신경원으로 연결되는 시냅스 사이의 자국물질에 의한 다. 다시 말해 연결되는 세포들은 살아남고, 연결되지 못한 세포들은 죽게 된다. 그러므로 선택적인 세포의 죽음이란 신경회로의 한 구성부 분이 되지 못하는 세포는 제거되어 버리는 것을 의미한다. 이처럼 신 경아가 이동하는 동안 길을 잃어버리게 되면 이를 제거해 버리는 것이 뇌의 기능을 증진시키는 데 도움을 주는 것이다. 1.11 기능적 안정화 마지막 발달과정은, 사용되는 시냅스의 접촉은 더욱 안정되지만 사 용되지 않는 시냅스는 약화 또는 배제된다는 이른바 기능적 안정화 (funct io n al valid a ti on ) 단계 이 다. 만약 이 런 안정 화가 이 루어 지 지 않으
 三
三
그림 4-3 볼수영과 피질동 (co rti cal barrel) (a) 왼쪽 봉수염 하나하나가 오른쪽 피질통 하나하나에 상응한다. (b) 와 (c) 에서 볼수영 한 줄씩을 출생 직후 제거하면(검은 원) 상응하는 피질통이 사라진다. (d) 만약 모든 볼수염을 제거하면 상응하는 피질통이 모두 다 사라진다. 면 신경원이 사라지게 된다는 대표적인 예가 생쥐의 체성감각피질에서 관찰되었다. 만약 생쥐의 볼수염 (wh i sker) 을 지배하고 있는 감각신경 원을 생애 초기에 파괴하면 이 신경원들과 시냅스를 이루고 있는 통 (barrel) 처럼 생긴 감각피질 부위가 완전히 사라지고 만다. 즉, 이 피 질세포들은 말초감각 수용기와 정상적인 시냅스 접촉을 이루지 못하므 로 기능적 안정화가 이루어지지 못해 죽어버리는 것이다(그림 4-3). 1.12 뇌피질의 풍요화 어린 시철의 환경과 경험의 차이가 뇌의 구조적 변화를 야기한다는 일련의 실험 결과가 Rosenzweig 등을 중십으로 한 캘리포니아 대학교 (버클리) 연구진에 의해 보고되어 주목을 끌었다. 이들은 그립 4-4 에 서 볼 수 있는 것처럼 3-4 마리의 쥐를 표준적 사육조건에서 키운 경우 (sta n dard colony : SC 조건), 동료 없이 단 한 마리의 쥐를 별다른 자
극 없이 키운 궁핍조건(i m p over i shed colony : IC 조건), 그리고 여러 마 리 의 쥐 를 다양한 장난감이 있는 조건에서 키 운 풍요조건 (enric h ed colony : EC 조건)으로 나누어 생후 23 일 이유 후부터 성숙할 때까지 사육하였다. 이들은 이런 사육조건의 차이가 쥐들의 대뇌피질에 구조 적 변화를 야기한다는 사실을 발견하였다. 죽, 다른 뇌 부위는 이 세 집단 사이에 별 차이가 없었지만, EC 조건에서 키운 쥐의 대뇌피질이 다른 두 집단의 피질보다 더 무거웠다. 다시 말해 EC 조건의 쥐는 IC
 (b)
(b)
조건의 쥐보다 피질 무게가 5% 더 무거웠고, SC 조건의 경우보다는 2% 정도가 더 무거웠다 (Rosenzwe ig & Leim an, 1988). 대뇌피질 가운 데 특히 차이가 나는 부위는 후두 부위의 시각피질이었고, 이런 현상 은 특히 EC 조건의 쥐에서 두드러지게 나타났다. 이러한 무게 증가는 주로 피질신경원의 기저 수지상돌기의 증가와 이에 따른 세포체의 비 대에 기인되는데, 평균적으로 EC 쥐는 IC 쥐에 비해 수지상돌기의 길 이가 20% 정도 더 길다. 이처럼 수지상돌기가 20% 길어지면 한 신경 원 당 시 냅 스 수가 2, 000 개 이 상 더 많아진다 (Greenoug h, 1975) . 또한 이러한 성장조건의 차이는 성숙 후의 학습능력에도 큰 차이를 야기한 다 . 예컨대 격리조건에서 성장한 생쥐는 표준조건에서 성장한 생쥐에 비해 미로학습, 조건회피학습 등의 수행 저조를 보여주며, 이러한 학 습 저조가 변연계, 특히 해마-편도 복합체의 신경전달물질의 기능 이 상에서 기인될 가능성이 시사되고 있다(장현감, 1984). 이상에서 본 것처럼 세포의 증식으로부터 시냅스 발생에 이르기까지 일련의 단계를 거쳐 뇌는 조직화되어 간다. 만약 환경자극이 정상적으 로 주어진다면 유전정보에 의해 명령받은 기본적인 신경회로는 안정화 되어 유지, 지속된다. 그러나 수지상돌기의 성장이나 시냅스 수정은 변화하는 환경조건에 적응하여 바뀔 수도 있다. 이러한 과정이 학습과 기억의 생물학적 기초 과정이 되는데 이는 후속되는 장에서 자세하게 논의할 것이다. 2 손상에 대한 신경조직의 반웅 신경세포나 신경교세포가 유사분열의 한 주기가 끝나면 신경교세포 는 이 주기로 되돌아올 수 있지만, 신경세포는 되돌아올 수 없다. 이 렇게 두 세포의 재생력이 다르다는 것을 알게 되면, 뇌가 손상된 후 신경원의 재생이 어렵다는 것을 쉽게 알 수 있다. 현재까지 알려진 지 식으로 볼 때 포유류의 뇌에 있는 성숙한 세포가 죽게 되면 후구에 있
는 후각신경을 제의한 다른 어떤 신경세포도 새로운 세포로 대치될 수 없다. 세포의 죽음은 한평생 계속되지만 특히 나이가 많아질수록 더 심해진다. 나이가 많아침에 따라 세포의 죽음이 가속화되면 정신적 또 는 행동적으로 많은 변화가 일어나게 된다. 2.1 축색 손상에 대한 반응 만약 한 신경원의 세포체가 회복될 수 없을 정도로 심한 손상을 입 으면 수지상돌기와 축색을 포함하여 세포의 모든 구성물이 죽게 된다. 그러나 만약 세포체가 죽지 않으면 손상된 축색은 여러 가지 형태의 재생반응을 보여준다. 만약 축색이 세포체로부터 다소 떨어진 곳에서 절단되어 세포체가 죽지 않았다면, 축색, 세포체, 세포체와 수지상돌 기 사이의 시냅스 접촉부 그리고 심지어는 시냅스후 세포체의 반응성 에도 변화가 일어나게 된다. 다음에는 이런 반응성 변화의 대표적 예 룰 몇 가지 경우로 나누어 고려해 보자. 2.2 전향성 변화 세포체로부터 절단된 축색분절은 곧 소멸되기 시작한다. 죽 세포체 로부터 더 이상의 영양 공급을 받지 못하기 때문에 몇 시간 이내에 죽 고 만다. 만약 축색이 수초로 덮여 있는 경우라면 수초가 먼저 떨어져 나가게 된다. 이렇게 죽게 된 축색이나 수초는 노폐물을 처리하는 특 수한 신경교세포들에 의해 찌꺼기로 처분된다. 양서류와는 달리 포유 류의 뇌에서는 기능적 재생이 이루어지지 않지만 중추신경계에 있는 신경원의 수초가 악화되어도 말초에 있는 원심성 신경원의 수초는 계 속 살아 있기 때문에 축색의 재생을 유도하는 반응이 계속될 수 있다. 만약 손상당한 세포의 시냅스후 세포가 근육세포인 경우 이 근육세 포는 곧 마비된다. 손상 직후에는 근육의 긴장성이나 탄력성이 상실되 는 정도이지만 몇 주 동안 새로운 신경지배가 이루어지지 않기 때문에
이 근육세포는 위축 (a t ro p h y)되고 만다. 만약 팔이나 다리가 골절되어 깁스를 해보았다면 깁스를 풀고 난 후 어느 기간 동안 근육이 위축되 어 있음을 경험하게 되는 것은 바로 이런 이유이다. 이러한 근 위축은 시냅스전 신경원에 의해 분비되는 어떤 자극요인 (tro p ht c fa c t or) 의 감 소에 의한다. 대부분의 경우 신경원과 골격근 사이에는 아세틸콜린 (ACh) 이 매개하는데, 이 ACh 가 바로 신경전달물질인 동시에 자국요 인이다. 그러므로 신경과 근육 접합부에서 ACh 의 효과를 차단하는 물 질을 주입하면 신경이 절단된 것과 근본적으로 동일한 효과가 나타난 다. 그러나 근 자체를 직접 전기적으로 자극하여 근 위축을 방지할 수 도 있는데, 이것은 곧 근육이 신경지배 없이도 직접 수축할 수만 있다 면 근 위축은 일어나지 않는다는 것을 보여주는 것이다. 시각 계통에 있는 신경원들도 위의 경우와 유사한 반응을 보여준다. 만약 시신경을 절단하면, 시신경의 축색이 끝나는 시상의 의측술상체 (lat e r al gen ic u late bod y)에 있는 신경원들에 심한 위축 현상이 일어나 고 잇달아 죽게 된다. 이러한 전향성 (ante r og rad e) 시냅스 변성은 다음 수준의 시각 계통에서도 일어난다. 즉 의측술상체에서 뻗어나온 2 차 축색들이 종결되는 시각피질의 IV 충에 있는 신경원들도 죽게 된다. 이 러한 연속적 시냅스 변성 효과는 기본적으로 모든 감각신경 계통에서 관찰되지만, 뇌의 모든 부분에서 공통적으로 일어나지는 않는다 (Kelly, 1981b) . 2.3 후행성 영향 만약 한 축색이 세포체에서 매우 가까운 곳에서 절단되면 거의 모든 신경원의 구성물들은 궁극적으로 죽게 된다. 그러나 절단된 곳이 세포 체에서 상당히 멀리 떨어진 곳이라면 세포체 쪽의 축색은 살아나고, 짤린 부위 바깥의 축색과 축색측지들은 접합을 이루고 있는 부위까지 차츰 죽어가게 된다. 한편 절단된 축색 부위에서는 봉합이 일어나고 여기서 섬세한 축색의 싹이 새롭게 돋아나게 된다. 이와 동시에 세포
체에서는 다음과 같은 일련의 변화들이 2-3 일 사이에 나타나게 된다. 즉 세포체는 수분을 흡수하여 크기가 3 배나 더 커지고, 세포핵은 세포 의 한쪽 가장자리로 이동하기 시작하며, 세포체 내의 소포체에 복잡한 변화가 일어난다. 이 변화는 RNA 와 새로운 단백질의 합성 등이 주된 것인데, 이것은 새로운 축색이 발아할 준비롤 하고 있다는 것을 의미 한다. 이러한 축색과 세포체에서 일어나는 후행성 변화는 3-4 주 동안 계속된다. 2.4 축색의 발아와 시냅스 개조 위에서 본 것처럼 만약 한 신경원이 축색의 손상에도 불구하고 계속 생존한다면 세포체와 연결된 축색부위에서 새싹이 돋아난다. 말초신경 원에는 손상된 신경원이 거의 정상적인 길이로 재생되거나 목표 세포 와 새로 시냅스 접촉을 이루고 나면 자라던 축색 싹은 더 이상 자라지 않는다. 축색이 발아되는 것은 일종의 신비에 속한다. 이러한 뇌 속의 축색발아는 별다른 효과가 없거나, 유해한 효과가 있거나(만약 축색발 아가 찰못되었을 때) 또는 유익한 효과가 있을 수도(만약 축색의 발아가 효과적으로 시냅스 접촉을 대치시켜 줄 경우) 있다. 비록 이 의문은 해결 되지 않았지만 Ra i sman 과 그의 동료들의 연구에 의하면 시냅스 개조 (syn a pt ic remodeli ng ) 라 부르는 과정 을 통해 축색 발아가 새로운 시 냅스 연결을 형성할 수 있을 것으로 생각된다 (Ra i sman & Fie ld , 1973). Ra i sman 과 F i eld 는 전자현미경을 이용하여 50,000 개 이상의 시냅스 를 연구하였는데, 이들은 해마와 시상하부 사이의 신경 접합이 이루어 지는 쥐의 중격핵을 중심으로 연구했다. 시상하부로부터 울라오는 내 측전뇌 속 (media l for ebrain bundle : MFB) 과 해 마로부터 나오는 축색 인 뇌궁채 (fim br i a- fo m i x) 가 중격핵에서 시냅스를 이룬다. Ra i sman 과 F i eld 는 이 두 신경로가 중격핵에서 서로 상이한 방식으로 시냅스하고 있다는 사실을 알게 되었다. 죽 MFB 를 통해 온 축색종말은 중격신경 원들의 세포체나 수지상돌기 양자에서 시냅스하지만 뇌궁채를 통해 들
어온 축색종말은 오직 중격신경원의 수지상돌기에서만 시냅스한다는 것이다. 이러한 시냅스 방식을 알아내고 나서 이들은 여러 마리의 쥐를 사용 하여 이 두 신경로를 하나씩 순차적으로 잘랐다. 죽 먼저 축색 하나를 절단하고, 절단된 축색의 싹이 나울 수 있든 시간을 주기 위해 몇 주 간을 기다렸다가 다른 하나의 축색을 절단하였다. 그 후 축색종말에서 변성이 일어나도록 2-3 일 간의 시간적 여유를 준 후 동물을 희생시키 고, 두 개의 절단 축색 중 늦게 절단한 축색에서 어떤 변화가 일어났 는가를 중점적으로 알아보았다. 그 결과 먼저 절단된 축색에서는 손상 이후 생긴 축색의 변성물질들이 신경교세포에 의해 이미 깨끗하게 치 워져 있었다. 즉 뇌궁채 손상에 앞서 MFB 를 먼저 절단한 경우는 MFB 손상에 따라 공백이 생겼던 시냅스 부위가 뇌궁채의 새로운 축 색종말로 가득 채워졌다. 절단의 순서를 위의 경우와 반대로 했을 때 도 먼저 절단된 뇌궁채의 시냅스 공백이 새로 돋아난 MFB 의 축색에 의해 채워져 있다는 사실을 발견하였다. 요약하면, 선행된 손상에 의한 축색종말의 공백 부위가 다른 축색의 새싹으로 채워진다는 것인데, 이는 중추에서도 시냅스의 개조가 일어 난다는 것을 뜻하는 매우 중요한 증거가 된다. 그런데 위의 두 신경원 은 각기 상이한 신경전달물질을 이용한다. 즉 뇌궁채는 콜린성 물질을 전달물질로 사용하고, MFB 는 노르아드레날린성 물질을 전달물질로 사용한다. 따라서 이 두 신경원을 수용하는 중격신경원의 수용기막에 서 새로운 신경전달물질을 수용하기 위한 기능 분화가 일어났다는 사 실도 알 수 있다. 이러한 시냅스 개조에 따라 행동상 어떤 변화가 일 어나는지는 아직 명확하지 않지만, 쥐의 경우 새로운 축색 싹이 돋아 날 수 있는 힘은 나이가 들면서 약화된다고 하는데 (McWi liam s & Lyn c h, 1983) , 이 사실은 어 린 동물들이 나이 든 동물들에 비 해 뇌 손 상 이후 기능 회복이 잘 된다는 사실과 어떤 관련성이 있을 것으로 추 측하게 한다.
3 노화 20 세기 후반에 들어오면서 영양의 개선, 의약의 발전, 보건위생의 증진 등에 따라 평균 수명이 연장되고 있다. 우리나라의 경우도 평균 수명이 70 세 이상으로 증가되면서 노령인구가 급속하게 늘어나 노화에 따른 심리 • 생물학적 관심과 연구가 증가되고 있다. 여기서 우리는 노 화에 따른 뇌 변화와 뇌 장애 그리고 이에 따른 심리적 기능의 변화를 살펴보기로 하자. 3.1 정상적 노화 나이가 들면서 신체와 함께 뇌도 노화된다. 노화가 진행됨에 따라 뇌는 신경원을 상실하게 되며, 이에 따라 뇌의 크기도 줄어들게 된다. 뇌 속의 신경원이 상실되는 것은 출생 전부터 시작되는 일이지만 35 세 까지는 그 정도가 미미하다. 그러나 나이가 많아지면 신경원의 상실이 더욱 늘어나게 되는데, 특히 75 세 이후부터 신경원 상실은 더욱 심하 게 된다. Brod y (l955) 에 의하면 90 세 노인의 뇌신경원 수든 유아의 뇌 신경원 수의 절반에도 미치지 못한다고 한다. 이러한 세포 상실은 뇌 무게의 감소를 야기하는데, 실제로 45 세 이후부터 뇌의 무게는 계속 감소된다. 이러한 사실에도 불구하고 신피질에 있는 어떤 신경원들은 평생을 통해 계속 성장해 가는 경우도 있다. Buell 과 Coleman(1979) 은 뇌 장 애가 아닌 다른 원인으로 사망한 평균 51 세의 성인들, 평균 80 세에 죽 은 노인들, 그리고 노인성 치매라는 뇌 장애 때문에 사망한 평균 76 세 의 환자들, 이렇게 세 집단의 뇌를 부검하여 뇌피질 신경원의 수지상 돌기의 변화를 비교했다. 결과에 의하면 뇌 장애 없이 사망한 집단(나 이분포 44-92 세)에서는 정단 수지상돌기가 많이 관찰되었는데, 이들에 서는 이 정단 수지상돌기가 1 년에 약 0,2lµm 씩 성장했다고 한다. 그 러나 이러한 수지상돌기의 성장이 노인성 치매 환자의 뇌에서는 전혀
관찰되지 않았다. 이 연구의 결과는 정상적인 노인은 비록 뇌가 노화 되더라도 신경원이 살아 성장하고 있는 데 반해, 노인성 치매 환자의 경우는 신경원의 상실과 함께 성장이 정지된다는 것을 시사한다. 노화가 진행되면서 상실되거나 손상되는 피질세포는 주로 운동피질 에 있는 Betz 세 포라고 부르는 비교적 큰 추체 세포들이 다. Scheib e ! , Tom iy asu 와 Sche i be!(1977) 은 74 - 102 세 사이에 사망한 7 명의 노인의 뇌를 부검하였더니, 전체 Betz 세포 가운데 약 75% 정도에서 수지상 돌기의 가시들이 상실되었고, 기저 수지상돌기가 위축되어 있었지만, 피질 주변에 있는 다른 신경원의 상실은 30% 정도에 불과했다. Betz 세포는 사지 가운데 주로 다리의 근육운동 특히 신근의 이완운동을 매 개하는 신경원이다. 이처럼 나이가 들어가면서 Betz 세포가 상실되기 때문에 노인들은 흔히 다리의 통증이나 경직, 운동장애 등을 많이 보 이게 된다 . 3.2 노화에 따른 인지적 변화 최근 수 년 동안 노인과 장년 사이의 인지적 기능에 어떤 차이가 있 는가를 알아본 연구들이 많이 보고되었다. 이런 연구들은 주로 학습과 기억울 연구대상으로 삼았다. 앞으로 학습과 기억의 장에서 자세하게 다루겠지만, 기억에는 3 가지 종류가 있다. 즉 전화 다이얼을 돌리는 동안 전화번호를 기억하는 것과 같은 단기기억 (short -ten n memo ry)과 두 종류의 장기 기 억 (lon g -ten n memory ) , 즉 세 상사의 이 치와 지식 등 울 기 억 하는 의 미 기 억 (semanti c memory ) 과 생 애 의 사건들을 기 억 하는 삽화기억 (ep iso dic memo ry)이 있다. 그런데 70 세 이전의 정상적 노인 들에서는 단기기억에는 큰 장애가 없지만, 장기기억에는 장애가 나타 나기 시작한다. 그래서 이들은 며칠 전에 읽은 책의 내용이나 경험했 던 사건의 내용을 잘 기억하지 못한다. 그 후 노화가 진행될수록 장기 기억뿐만 아니라 단기기억까지도 영향을 받게 된다.
3.3 비정상적 노화 대부분의 노인들은 노화가 되더라도 지적 능력에 십한 손상이 일어 나지 않지만 소수의 노인들에서는 지적 손상이 십하게 일어나는 수도 있다. 65 세 이상의 노인 가운데 약 5% 정도가 심한 인지적 장애를 보이는 치매 (dementi a) 환자이다. Coy le , Pric e 그리고 DeLong (19 83) 에 의하면 미국의 경우 65 세 이상의 노안 가운데 5% 는 십한 치매 환 자이고, 그 밖에 10% 의 노인은 경미한 치매 환자라고 한다. 치매로 진단받은 환자 가운데 약 70% 정도가 알츠하이머 병이라는 보다 심한 인지적 장애를 보여준다. 치매 환자들이 보여주는 초기 증후의 특칭은 최근의 사건에 대한 삽화적 기억에는 장애가 심하지만, 어린 시절의 사건 등과 같은 먼 과거의 기억에는 큰 장애를 보이지 않는 점이다. 알츠하이머 병으로 진행되면 글을 읽거나 쓰는 능력의 장애, 의식의 전반적 혼미, 언어장애, 숫자 계산의 장애 등이 나타나기 시작한다. 이런 치매 환자들은 찰 흥분하고, 환각이나 피해망상과 같은 증상도 나타낸다 (Co y le 등, 1983). 3.4 알츠하이머 병에서 잘 나타나는 변화 알츠하이머 병으로 죽는 환자가 늘어나면서 이들의 뇌를 부검하여 뇌의 조직학적 변화를 체계적으로 알아보는 연구들이 많이 수행되었 다. 이 병에 따른 뇌 변화는 다양한데 대체로 다음과 같은 5 가지가 많 이 나타난다. 3.4 .l 세포상실 알츠하이머 환자들이 보여주는 가장 두드러진 변화는 뇌피질에 십한 세포 상실이 일어난다는 점이다. 건강한 노인의 경우에도 젊었을 때와 비교하면 30% 정도의 뇌피질 세포가 상실되지만 알츠하이머 환자들의 경우 세포 상실이 무려 50% 이상 일어난다고 한다 (Samora j sk i, 1976).
이러한 세포 상실은 주로 인지적 기능을 담당하는 전두영이나 측두영 의 피질 부위에서 일어나기 때문에 지적 기능에 장애가 생기는 것은 극히 당연한 일이다. 3.4 .2 수지 상돌기 의 번 성 알츠하이머 환자의 뇌 조직을 검사한 연구들에 의하면 피질신경원의 수지상돌기가 심하게 변성되어 있다고 한다. 예컨대 Scheib e ! 등 (1975) 의 연구에 따르면 환자의 실제 나이와는 관계 없이 치매의 정도가 심 해질수록 수지상돌기의 변성이 더욱 두드러지게 나타난다고 한다. 수 지상돌기의 변성은 특히 신피질의 제 m 충에서 많이 관찰되며, 특히 이 돌기의 기저부나 수평부에서 두드러지게 일어난다(그림 4-5). 이 수지 상돌기들은 피질 내에 있는 신경원들과 시냅스하는 돌기이며, 유아기 에 최종적으로 분화된 돌기들이다. 그러므로 가장 늦게 만들어전 이 돌기들이 가장 먼저 변성되는 셈이다. 이렇게 변성되는 수지상돌기들 은 고도의 추리, 복잡한 생각, 기억 등과 같은 피질 활동과 밀집한 관 련이 있는 것으로 추측된다. 세 포의 신 경 관 (neurotu b ules) 이 나 신경 세 사 (neurofi lam ent) 가 퇴 화되 면 수지상돌기들도 점차 비틀어지고 변성될 가능성이 있다. 알츠하이
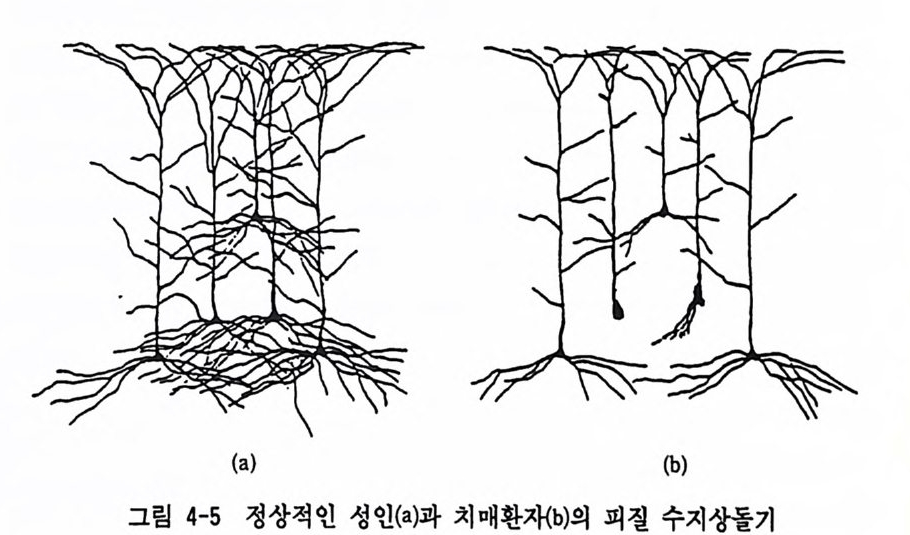 (a) (b)
(a) (b)
머 환자들 가운데 특히 40 대 말이나 50 대 초반에 이 병에 걸린 환자들 은 수지상돌기에 또 다른 형태의 병적 변화가 나타난다. 죽 이들의 뇌 에서는 노인 환자들에서 보편적으로 볼 수 있는 수지상돌기의 퇴화뿐 만 아니라 세포체가 병적으로 부풀어오르고, 기저부나 정단부의 수지 상돌기에서 가시가 상실되며, 특히 어느 한 부위의 수지상돌기가 무질 서하게 크게 성장하는 병적 성장의 모습도 보여준다. 이러한 병적 변 화가 이들의 인지적 기능장애를 일으키는 주원인으로 간주된다. 3.4 .3 신경섬유의 엉킵 신경관이 뒤틀리거나 꼬이는 현상을 신경섬유의 엉킴 (neurofi br ill ar y t an g le) 이라 하는데, 이것이 치매 환자에서 두드러지게 잘 관찰되는 또 하나의 증후이다. 이 엉킴 현상은 알츠하이머 환자의 뇌에서 보편적으 로 관찰되는 것이지만, 대표적인 정신지체 환자인 다운 (Down) 중후군 환자에서도 관찰되며, 이런 엉킴은 이들 신경원 내에서의 정상적인 물 질 흐름을 차단하여 정상적인 기능을 못 하도록 하는 것이다. 정상적 으로 노화된 노인의 뇌에서도 가끔 관찰된다. 3.4 .4 노인성 반점 나이가 매우 많은 노인이나 알츠하이머 환자의 뇌에서 관찰되는 또 다른 특칭으로 노인성 반점 (senil e p la q ue) 이라는 것도 있다. 이 반점 부위를 골지 방식의 세포염색법으로 염색하면 약간 작고 검은 반점으 로 나타난다. 이 반점은 축색종말이 변성된 것인데 면역체계에 의해 생성되는 면역글로블린(i mmuno g lobu li n) 과 유사한 아밀로이드 (am y · loid : 전분과 유사한 구조적 특성을 가진 복합물질이며 당단백질로 생각되 지만 아직 생화학적 조성은 정확하게 밝혀지지 않았다)라는 복잡한 단백질 울 포함하고 있다. 3.4 .5 신경전달물질의 장애 알츠하이머 환자의 뇌 속에 어떤 종류의 신경전달물질 수준이 병적
으로 변화되는가는 아직 명확하지 않다. 그러나 지금까지 발견된 사실 가운데 ACh, 노르아드레날린, 5-HT 그리고 소마토스타틴 (soma t os t a ti n) 과 같은 물질들의 분비 수준이 병적으로 낮아진다고 한다 (W inb lad 등, 1985) . 이 들 신경 전달물질 가운데 특히 ACh 의 분비 수 준이 가장 낮아진다고 한다. ACh 의 수준은 ACh 의 전구물질인 콜린과 아세틸 조효소 A 로부터 ACh 를 합성 하는 효소인 choli ne acety lt ra nsfe r ase (ChAT) 의 분비 수 준을 측정함으로써 알 수 있다. 정상대조집단에 비해 알츠하이머 환자 집단에서는 이 효소의 분비 수준이 60-90% 밖에 되지 않는다 (Co y le, Price , & Delong, 1983). 뇌피질에서 콜린성 신경원의 세포체들은 기저 전뇌 콜린성 체계 (basal for ebrain choli ne rgic s y s t em) 라 부르는 전뇌의 기저부에 있는 매우 좁은 영역에 밀집되어 있다 (Pr ic e 등, 1985). 이 기저 전뇌 콜린성 체계에 있는 신경원들은 대뇌피질과 해마의 여러 영 역으로 축색을 보낸다. 기저 전뇌 콜린성 체계에 포함되는 중요 뇌 구 조에는 내측중격 (media l sep tum ), 브로카 대 (dia g o n al band of Broca : dbB), 그리고 메 이네르트 기저 신경 핵 절 (nucleus basalis o f Mey ne rt : nbM) 등이다. 내측중격과 dbB 는 주로 해마로 두사되고, nbM 은 신피 질의 여러 영역으로 산만하게 두사한다. nbM 에 있는 신경원의 상실이 뇌의 광범위한 영역에서 볼 수 있는 ChAT 의 감소에 매우 중요한 역 할을 하는 것으로 알려져 있다. 따라서 nbM 에 있는 콜린성 신경원의 축색에 변성이 일어나면 뇌피질의 ChAT 수준이 감소된다• nbM 의 신경원 상실은 다른 형태의 치매 환자에서도 관찰된다. 예컨 대 파킨슨 병의 하위범주에 속하는 치매성 환자들에 있어서도 nbM 에 서 선택적인 세포 손실이 나타난다. 또한 다운 증후군에 속하는 노인 환자들이 보여주는 지적 손상도 nbM 및 nbM 과 관련되는 뇌 영역의 콜린성 신경원의 변성과 불가분의 관계에 있다는 증거들도 있다 (P ri ce 등, 1985). 비록 알츠하이머 병에서 다론 뇌 부위의 변성도 문제시될 수 있지만, 특히 기저 전뇌 콜린성 체계에서 나타나는 현저한 세포 손 실과, 이 손실에 따른 뇌피질이나 해마를 포함하는 광범위한 뇌 영역
에서 콜린성 축색의 변성이 알츠하이머 병의 지적 손상을 야기하는 데 매우 중요한 요인으로 생각된다. 3.5 콜린성 가설과 치매의 치료 해마나 뇌피질에서 콜린성 신경지배가 감소됨에 따라 알츠하이머 병 의 여러 증후가 야기된다는 제안을 이른바 콜린성 가설 (cho li ner gic h yp o th es i s) 이라고 한다. 이 가설에 근거하여 뇌에 ACh 의 공급량을 늘 려중으로써 환자의 인지적 기능을 부활시켜 보려는 시도가 많이 행해 졌다. ACh 는 혈액이나 소화기관 속에서 분해되기 때문에 이를 직접 말초로 주사하거나 구강을 통해 복용해서는 효과가 없다. 그러므로 가 장 흔히 사용되는 방법은 ACh 의 전구물질인 콜린을 환자가 먹을 음식 에 첨가하는 것이다. 이와 유사한 방법으로 콜린성 물질인 레시틴 (l e cithi n) 이란 물질을 복용하게 하는 방법도 사용된다. 이처럼 ACh 의 전구물질을 많이 복용하게 함으로써 이 환자들의 뇌 속에 아직 살아 있는 콜린성 신경원이 보다 많은 ACh 를 생산하게 함으로써 시냅스 회 로에 보다 많은 양의 ACh 가 전달물질로 이용될 수 있게 하려는 방법 이 시도되고 있다. 그러나 이런 기대는 크게 성공하지 못했다. 이런 예견을 실험적으로 연구한 논문들을 개관한 Bar tus 등 (1985) 의 연구에 의하면, 이러한 전 구물질의 처치가 한두 사례에서는 지적 기능을 약간 향상시켰으나 대 부분의 연구에서는 알츠하이머 환자의 기억기능을 증진시키는 데 실패 했다고 한다. 이러한 전구물질 투여가 지적 기능의 개선에 실패한 이 유를 지적하는 견해도 제시되었다. 예컨대 nbM 에 생존하고 있는 콜린 성 신경원이 수적으로 너무 적어 잉여 투여한 콜린을 이용하는 데 실 패했을 것이라는 견해, 콜린성 신경원의 콜린 섭취기능이 적철하지 못 해 잉여 콜린이 이 신경원 속으로 잘 들어가지 못했을 것이라는 견해, 또는 비록 많은 ACh 가 합성되었다 하더라도 이의 시냅스 방출과 방출 후 시냅스후 수용기와의 상호작용에 문제가 생겼을 수도 있다는 견해
둘이 제시되었다. 미국의 경우 노인성 치매 환자가 약 100 만 명에 이 르고 이들의 가정치료비로 매년 약 60 억 달러가 지출되는 것으로 추정 되고 있다. 이런 막대한 치료비 지출 때문에 노인성 치매 환자의 치료 는 사회적으로 십각한 문제로 대두되고 있으며, 이에 따라 많은 치료 연구가 진행중이다. 그 동안 알츠하이머 병의 약물치료를 위한 몇 가지 약품이 선보였 다. 그 중 많이 쓰이는 약물은 시냅스에서 ACh 를 무기력하게 만드는 효소인 AChE (acety lc holi ne ste ra se) 의 기 능을 억 제 하는 피소스티 그민 (ph ys o stig mine ) 이 란 약물이 다. 이 약물을 동물에 게 두여 하면 ACh 의 시냅스 작용을 연장시키는 것으로 알려져 있지만, 이 약물을 알츠하이 머 환자에게 투여하여 인지적 기능의 향상이 일어나는지의 여부를 알 아본 실험 연구는 그렇게 많지 않다 (Crook, 1985). 앞서 본 ACh 의 전 구물질을 투여하면 별다른 부작용이 나타나지 않지만, 피소스티그민과 같은 AChE 길항물질을 투여하면 심한 부작용이 나타난다. 따라서 현 재로서는 이 약물을 알츠하이머 병의 치료에 이용하는 데는 많은 제한 이 따른다. 한편 피라세탐(pi race t am) 이라는 약물이 알츠하이머 병의 치료에 웅 용되기도 하는데, 이 약물은 피소스티그민과 같은 항콜린효소 (an ti cho li nes t erase) 가 아니라 시냅스전 축색종말에서 ACh 의 방출을 촉진시 키는 약물이다. 이 약물은 흰쥐와 같은 실험동물을 대상으로 하는 실 험에서 단독으로 사용할 수도 있고, 콜린과 혼합하여 사용할 수도 있 다. Bart us 등(1 982) 은 콜린과 혼합된 피라세람을 주사받은 흰쥐의 기 억능력이 처치를 받지 않은 대조군의 기억능력에 비해 다소 좋아진다 는 사실을 보고하였다. 그러나 아직 이 약물을 알츠하이머 환자에게 직접 사용하여 성공했다는 사례는 보고되지 않고 있다.
제 5 장 시각 광파장 에너지가 눈의 특수한 수용기에 도달하면 전환(t ransdu cti on) 과정을 거쳐 신경세포의 전기적 에너지로 변화된다. 전환이란 감각 계 통에서 일어나는 한 현상으로 어떤 형태의 에너지가 다론 형태의 에너 지로 변화되는 과정을 말한다. 신경충격은 눈의 망막에서 출발하여 뇌 의 여러 영역으로 전달된다. 미세 기록전극을 사용하여 여러 뇌 영역 에서 일어나는 전기적 반응을 기록한 연구를 통해 우리는 뇌가 어떻게 시각자국의 여러 가지 특성들, 다시 말해 모양, 색깔, 위치 등을 지각 하게 되는가를 알 수 있게 되었다. 시각 계통의 전반적 조정기능을 설명하기 위해 두 가지 주요 이론이 제시되었다. 첫째는 계열처리이론 (se ri al pro cessin g th eo ry)인데, 이 이 론에 따르면 얼굴재인과 같은 복잡한 시각정보는 이 자극에 대한 반응 성이 높은 신경원들로 구성된 여러 뇌 영역들이 칙렬로 연결된 회로를 통해 단계적으로 처리된다고 한다. 둘째는 병행처리이론(p arallel pro cessin g th eo ry)이다. 이 이론에서는 계열처리이론과는 달리 얼굴과 같은 복잡한 자극의 지각은 시각자극의 여러 가지 측면을 개별적으로 처리하는 상이한 뇌 영역들이 거의 동시에 활동함으로써 이루어진다고 설명한다. 앞으로 이 두 이론의 구체적인 차이를 살펴볼 것이다.
l 눈의 구조 척추동물의 눈에는 광선에너지를 신경에너지, 죽 신경충격으로 바꾸 어 주는 망막 (re ti na) 이란 매우 엷은 세포충이 있다. 이 망막세포충은 두께가 300-400run 정도밖에 되지 않지만, 여기에는 수백만 개의 신경 원이 매우 복잡한 시냅스를 이루고 있다. 망막에서 발생한 신경충격은 시신경 (op tic nerve) 을 통해 뇌로 전달된다. 뇌는 이러한 신경충격의 흐름을 해석하며, 이 해석의 결과로 지금 무엇을 보았는가를 알 수 있 게 된다. 망막은 안구의 뒷면에 붙어 있다. 망막의 전면에는 젤리 (g e lli)와 같 은 물질인 수정 액 (vit er ous humor) 이 있고, 이 수정 액 과 눈의 제 일 앞 부분 사이에 렌즈(수정체)가 있다. 렌즈에 연결된 몇 개의 작은 근육 들이 렌즈의 형태를 조절하여 망막 위에 광선의 초점이 맺히도록 해준 다. 눈의 전면에는 수양액 (aq ue ous humor) 이라는 액체가 들어 있으며, 눈의 제일 바깥 표면 쪽에는 각막 (cornea) 이란 구조가 있다. 이것은 굴곡면을 갖는 구조로서 두께는 중앙부가 약 0.5nm 정도이고 가장자 리는 약 1.0nm 이다. 인종에 따라 파란눈, 노란눈, 검은눈 등 눈동자 의 색깔이 다른데, 그 이유는 홍체(i r i s) 가 가진 색소의 차이 때문이 다. 홍체는 매우 얇고 정교한 근육조직으로 중심부에 있는 동공{p u pil) 의 크기를 조철한다. 광선이 눈 안에 들어갈 때는 각막, 수양액, 동공, 수정액을 거쳐 마 지 막으로 망막에 이르게 된다. 광선 수용기는 간상체 (rod) 와 원추체 (cone) 인데 이것은 눈의 제일 뒷부분인 망막에 있다. 그러므로 광선이 광선 수용기에 도달하려면 망막에 붙어 있는 많은 다른 종류의 세포들 을 거치지 않으면 안 된다. 이 세포들은 모두 투명하기 때문에 이들 세포를 통과할 때 에너지의 손실이 거의 일어나지 않는다(그립 5-1). 시신경을 이루는 축색이 안구를 출발하는 망막 지점을 맹점 (b li nd s p o t)이라 한다. 이 맹점에는 광선 수용기가 없으므로 이곳에 어떤 시 자극이 도달하여도 시각 경험이 일어나지 않는다. 그럼에도 불구하고
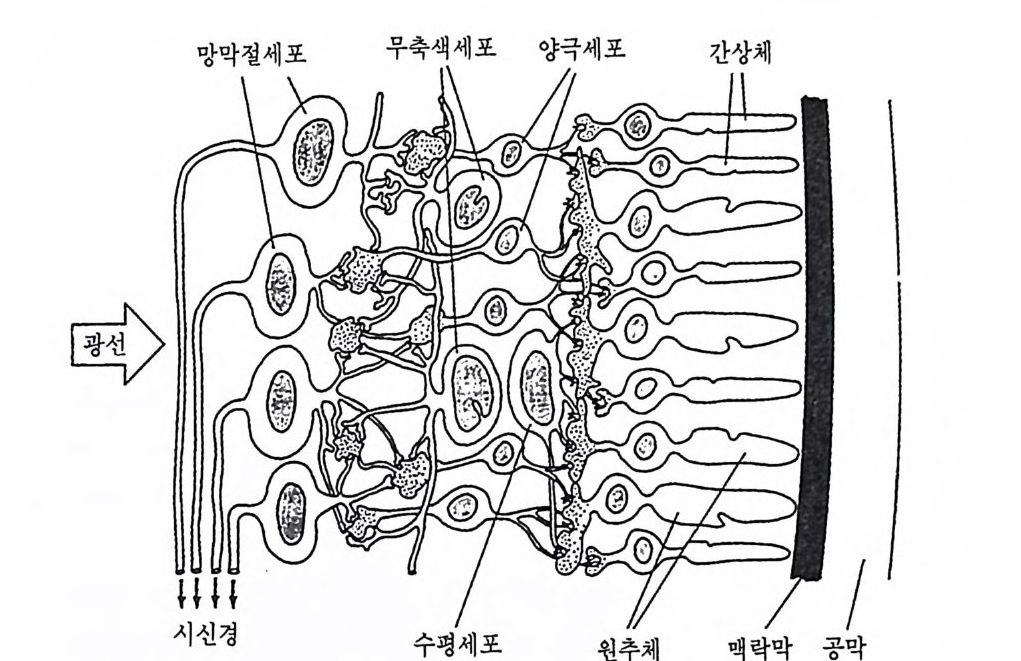 무축색세포 양국세포 간상체
무축색세포 양국세포 간상체
그림 5 키 포유류의 망막구조 광선 수용기인 간상체와 원추체가 수평세포와 양국세포와 일차 연결하고, 이 어 무축색세포와 망막절세포와도 연결되어 시신경을 이룬다. 우리는 일상적으로 맹점을 잘 인식하지 못하는데, 그 이유는 이 맹점 부위를 뇌가 기능적으로 채워주기 때문이다. 2 시각 계통의 해부학 뇌 구조 가운데 많은 부위들이 시자극의 정보처리에 개입한다. 현재 활동중인 신경원들을 탐지해 내는 기법인 2- 디옥시글루코오스 (2-deox y glu cose) 법을 사용하여 시각의 대뇌 구조를 찾아보면, 전통적인 해부 학에서는 시각과 관련이 없는 것으로 알려진 뇌 구조들이 시자국에 반 웅을 보인다. 이러한 뇌 구조에는, 전두영의 여러 부분들과 기저핵의 미 상핵 (caudate ) 과 피 각(p u t amen) 까지도 포함되 지 만, 여 기서는 시상의
의측술상핵 (lat e r al gen ic u late nucleus: LGN) 과 후두영의 시각피질을 중 십으로 언급할 것이다. 2.1 외측술상핵 시신경은 망막절세포의 축색들로 이루어진다. 인간이나 레서스 원숭 이의 경우 약 1 백만 개의 절세포축색이 모여 시신경을 이룬다. 대부분 의 시신경 축색들은 시상의 의측술상핵로 들어가 그곳에서 시냅스하지 만, 약간의 시 신 경 축색 들은 중뇌 의 상구체 (sup er io r coll icu lus) 나 시 상 하부의 시교차상핵 (sup ra chia s matic nuclei : SCN) 으로 들어가는 경우도 있다. 그림 5-2 는 망막과 의측술상핵 사이의 연결을 나타낸 것이다. 코 부 위의 안쪽 망막에서 출발하는 시신경축색은 건너편 뇌의 의측술상핵으 로 투사되는데 , 이 런 두사방식 을 대측 (con t rala t era l) 투사라 한다. 한 편 망막의 바깥쪽 부위에서 출발하는 축색은 같은 쪽의 의측술상핵에 무사되는데, 이런 두사방식을 동측(ip s il a t era l) 투사라 부른다. 이러한 기본 두사방식은 모든 포유류에 공통적인 현상이지만, 대측과 동측의 투사비율은 동물의 종에 따라 다르다. 유인원의 경우는 이 두 방식의 두사비율이 50 : 50 정도이다. 망막절세포에서 출발한 시신경의 축색이 뇌에서 교차되는 지접을 시교차 (optic ch i asm) 라 하는데 이곳은 뇌의 복측에 위치한다. 이러한 해부학적 배열 때문에 왼쪽 시야에 나타난 사물의 상이 절반 은 오른쪽 뇌로 들어가며, 반대로 오른쪽 시야의 반은 왼쪽 뇌로 들어 간다. 이와 같은 원리를 이용하여 두 대뇌반구 사이의 연결구조인 뇌 량을 절단한 환자를 대상으로 좌반구와 우반구의 기능을 독립적으로 연구할 수 있게 되었다. 유인원의 경우 개개의 LGN 은 약 1 백만 개 정도의 신경원으로 구성 되며, 6 개의 분명한 충으로 구성되어 있다(그립 5-3). 제일 안쪽 두 개 의 충 (1 층과 2 층)에 있는 세포들은 바깥쪽에 있는 나머지의 4 개충 (3,
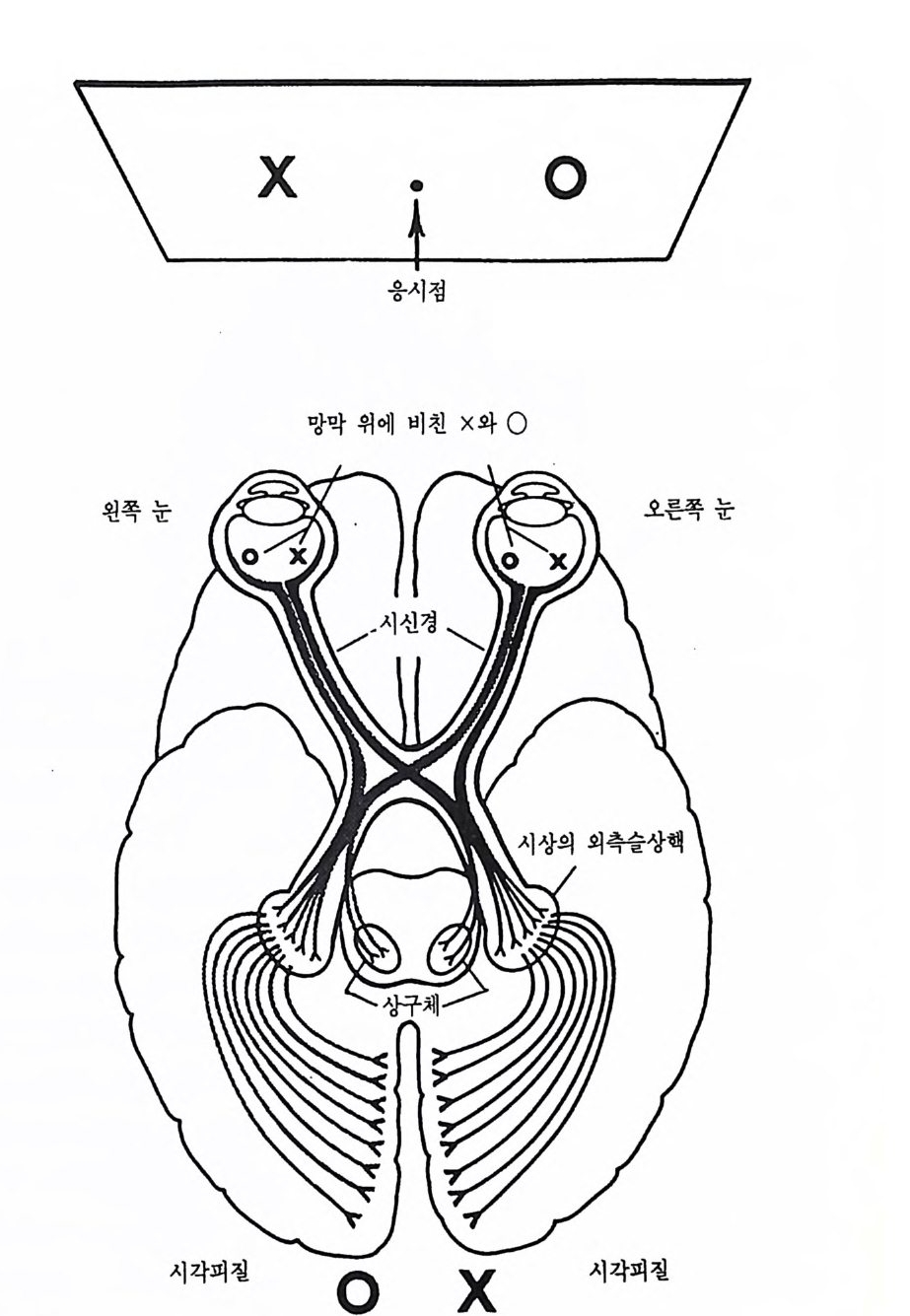 x • 。
x • 。
그림 5-2 인간의 시각동로 망막의 오른쪽 반에서부터 온 입력 (X) 은 우측 시상의 의측술상핵과 우측 피 질로 두사된다. 반대로 망막의 왼쪽 반에서부터 온 입력 (0) 은 왼쪽 시상의 의측술상핵을 지나 왼쪽 시각피질로 무사된다 .
 ‘ .... ,,..\i,, `· `' . . .i ' . 서 ,·.., i .. 'f'. ..• .` 를 ..’ 广 t • \ .. .. .. i\ ^ . . .. . ,,. [\:.'J \. ,,, ]’. '
‘ .... ,,..\i,, `· `' . . .i ' . 서 ,·.., i .. 'f'. ..• .` 를 ..’ 广 t • \ .. .. .. i\ ^ . . .. . ,,. [\:.'J \. ,,, ]’. '
그립 5-3 레서스 원숭이의 외측술상핵 6 개의 세포충으로 구성되어 있다. 4, 5, 6 층)의 세포들보다 크기가 더 크다. LGN 에 있는 세포들 가운데 1, 4 및 6 층에 있는 세포들은 건너편의 눈에서 온 축색들과 시냅스 접 촉을 하고 2, 3 및 5 층에 있는 세포들은 같은 편의 눈에서 온 축색들 과 시냅스 접촉을 한다. 2.2 대뇌피질의 시각영역 대부분의 LGN 에서 나온 축색은 대뇌피질의 시각영역으로 두사된 다. 원숭이의 경우 LGN 에서 출발하는 축색의 99% 가 1 차 시각피질에 서 시냅스를 이룬다. 이 피질영역은 Brodmann 의 분류 체계에 따르면
17 번 영역에 해당되며, 염색하면 줄 모양의 띠가 보이므로 선조피질 (str ia t e co rt ex) 이라고 한다. 한편 LGN 에서 출발하는 극소수의 축색은 선조피질에 인접해 있는 Brodmann 18 번 또는 19 번 부위에서 시냅스 를 이룬다. LGN 에서 시각피질로 진행하는 축색들은 측지를 뻗어 많 은 시각피질의 신경원들과 시냅스를 이루는데, 이러한 시냅스의 대부 분은 대뇌피질의 IV 및 VI 층에서 이루어진다. Brodmann 17 번에 있는 신경원들은 인접의 18 및 19 번의 시각피질이나 피질하 시각구조인 LGN 또는 상구체 의 표면충으로 다시 투사한다. 3 망막에서의 시각전환 시각은 광선이 망막에 있는 광선 수용기를 자국함으로써 시작된다. 광자극에 칙접 반응할 수 있는 수용기는 망막에 있는 간상체와 원추체 뿐이다. 망막에는 이 두 세포 의에도 수백만 개의 다른 세포들이 있지 만, 이 세포들은 간상체와 원추체에 의해 유발된 시신경 활동에 의해 작용하는 세포들일 뿐 광자극에 직접 반응하지 않는다. 광자극이 신경 활동으로 전환되는 과정은 1967 년 노벨상을 수상한 George Wald 와 그의 동료들에 의해 많이 연구되었다. 여기서는 Wald(1968) 의 연구를 중심으로 언급하기로 한다. 광선이 원추체와 간상체를 자국할 때 즉각적으로 일어나는 변화는 이 세포들 속에 함유되어 있는 감광성 물질의 분자적 구조변경이다. 이 감광성 물질들을 광색소(p ho t o pigrn en t)라고 부르는데, 이 물질의 분자구조가 바뀌면 이 물질의 색깔도 바뀐다. 간상체에 있는 감광성 물질을 로돕신 (rhodo p s i n) 이라 부르는데, 이 물질에 대한 연구가 가장 활발하게 이루어졌으며 이와 유사한 물질이 원추체에도 있다. 로돕신은 복잡한 단백질인 옵신 (o psi n) 과 보다 작은 구성물인 레티 닌 (re ti nene) 이라 부르는 두 가지 단백질로 구성되어 있다. 레티닌은 구조적으로 비타민 A 와 유사하다. 그립 5-4 에서 볼 수 있는 것처럼
로돕신의 분자들은 간상체의 의부 분절을 이루는 일련의 원판의 막 속 에 들어 있다. 이 원판들은 간상체로부터 계속 떨어져 나가지만 새로 형성되는 원판에 의해 대치된다. 이와 유사한 대치과정이 원추체에서 도 일어난다. 정상적으로는 이렇게 떨어져 나간 원판은 망막에 있는 정맥혈관을 통해 제거되지만 색소성 망막염 (reti ni t is p igm en t osa) 이란 병에 걸리면 원판이 떨어져 나가는 과정이 병적으로 이루어진다. 이 병은 유전성이며 망막 위축, 망막혈관 약화 그리고 시야 축소 등을 일 으켜 결과적으로 심한 시각장애를 유발시킨다. 지금까지 척추동물의 광수용기를 연구한 실험결과들에 의하면 광선 에 대한 간상체의 전기적 반응은 안정 막전위로부터 과분국화된 수준 으로 바뀐다는 사실이 밝혀졌다(어떤 무척추동물에서는 광선에 대한 광수 용기가 탈분극화를 야기하는 경우도 있다). 죽 어두운 상태에서는 척추동 물의 간상체의 표피분절에 있는 세포막은 Na+ 의 통로를 열어주게 되 어 N 강가 간상체 내부로 유입되고, K+ 이온은 반대로 간상체 내부에
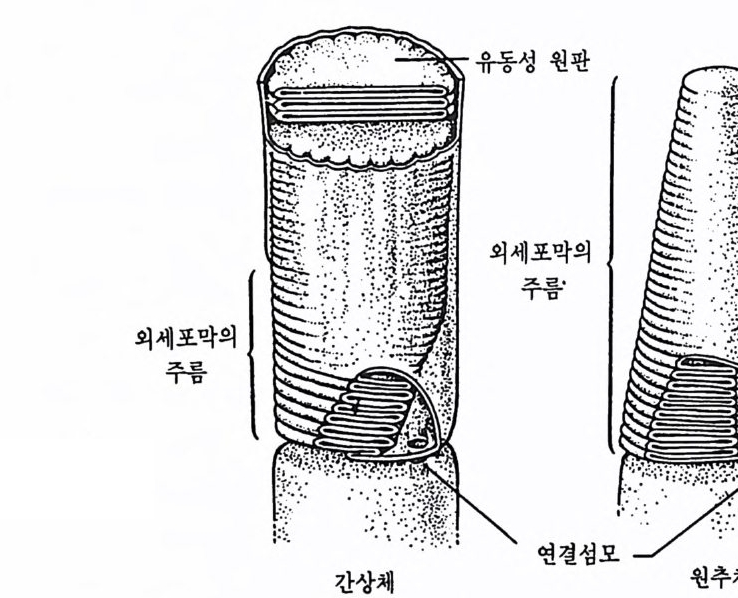 의세포막의
의세포막의
그림 5-4 원추체와 간상체의 모습 의세포막에 둘러싸여 있는 원판 속에 광색소들이 들어 있다.
서 바깥으로 유출되 어 이 른바 암전류 (dark current) 라 부르는 작은 전 류가 발생하게 한다. N 갑 통로는 cGMP(cyc l ic GMP) 란 물질에 의해 계속 열려진다. 이때 광선이 로돕신 분자를 치게 되면 이 분자의 모습 이 변화되어 다른 단백질을 활성화시키게 되고, 그 결과 cGMP 를 분 해시켜 N 강 동로를 닫게 됨으로써 N 갑가 간상체 안으로 들어가는 것 을 막게 된다. K + 의 바깥 유출은 계속되지만 이에 대응하는 N 갑 유 입이 더 이상 이루어지지 않으므로 막전위는 음성전하를 띠게 된다. 따라서 광선에 대한 간상체 막의 반웅은 결과적으로 과분국화를 야기 한다. 예컨대, 바다거북이의 간상체에 빛을 비추면 막전위가 _45rnV 의 안정 전위 로부터 —65r nV 까지 20rnV 이 상 . 과분국화를 야기 하게 된 다고 한다 (Schnapf & Bayl o r, 1987) . 광선에 대한 간상체의 반응이 과분극화된다는 접은 예상치 못한 일 이다. 왜냐하면 앞에서 우리는 과분국화는 곧 반응을 억제하는 것으로 보았기 때문이다. 그러나 간상체에서 일어나는 사건은 이와 다르다. 어두운 상태에서 간상체는 미량의 신경전달물질을 이와 시냅스하고 있 는 다론 신경원을 향해 계속 분비한다. 간상체로부터 이런 신경전달물 질 분비는 신경충격 없이도 일어난다. 실제로 간상체나 원추체는 신경 충격을 일으키지 못하고 점진성 반웅만을 야기할 뿐이다. 간상체에 의 해 분비되는 신경전달물질은 인접 세포의 시냅스 물질을 인접 세포의 시냅스후 수용기의 성질에 따라 억제성 또는 흥분성으로 작용하게 할 수 있다. 만약 신경전달물질의 분비·효과가 다음 세포에 대해 억제성 울 보이게 하려면 광선효과가 이런 안정화된 억제성을 감소시키는 이 른바 탈억제 (d i s i nh i b iti on) 를 일으켜야만 한다. 탈억제는 기능적으로 흥분성과 같다. 만약 간상체가 다음 세포를 흥분시킨다면 광선은 다음 세포에 억재적으로 작용할 것이다.
4 이중이론 이중이론 (du plicity th eo ry)은 간상체와 원추체가 시각 활동에서 서로 상이한 작용을 한다는 것으로, 다른 말로 시각에는 〈 이중 (double) 〉 의 수용기 또는 두 개의 수용기 체계가 있다는 것을 가리키는 것이다. 비 록 간상체에는 로돕신이라는 단 한 가지 유형의 감광색소만 있지만, 원추체의 경우는 그 종류가 세 가지이며, 이들은 각기 상이한 세 가지 의 감광색소를 갖는다. 원추체에 있는 세 가지 상이한 감광색소는 옵 신 부분의 차이에서 기인되며 레티닌 부분은 모든 유인원에서 동일하 다. 원추체에 이처럼 상이한 세 가지 감광색소가 존재한다는 것은 곧 원추체는 간상체와는 달리 상이한 광파장에 대해 반응할 수 있다는 것 울 의미한다. 간상체와 원추체의 또 다른 차이로는 간상체는 어두운 광선 아래서는 효과적으로 작용하지만 밝은 광선 아래서는 작용하지 못한다. 반면 원추체는 낮처럼 밝은 곳에서는 효과적으로 작용하지만 어두운 광선 아래서는 작용을 하지 못한다. 또 원추체 계통은 밝은 광 선 아래에서 사물의 섭세한 면을 볼 수 있는 시력 (vis u al acu ity)이 간 상체보다 훨씬 뛰어나며, 간상체는 이와 달리 광선이 없는 어두운 곳 에서 사물을 찾는 감수성 (sens iti v ity)이 원추체보다 더 높다. 인간의 눈에는 약 1 억 개의 간상체와 500 만 개의 원추체 그리고 약 100 만 개의 절세포가 있다 (She p herd, 1983). 평균적으로 하나의 절세 포로 폭주해 둘어오는 광선 수용기의 수령률은 간상체의 경우 100 : 1, 원추체의 경우 5 : 1 정도가 된다. 망막의 중앙부, 즉 중심와(fo vea) 에 는 간상체가 거의 없지만 원추체는 많이 밀집되어 있으며 수렵률은 1 : 1 정도가 된다. 따라서 이곳에서 시력이 가장 높지만 오직 광선이 충분한 낮 동안에만 그러하다. 중심와에는 약 5 만 개의 원추체가 있으 며, 망막의 의곽 부위로 나갈수록 간상체가 더욱 많아진다. 망막의 바 깥 가장자리의 광수용기는 모두 간상체로 이루어져 있다. 망막의 가장자리에 있는 간상체는 절세포에 대한 매우 높은 수렵률 울 보여준다. 망막의 말초 부위로 나아감에 따라 간상체가 밀집되어
있기 때문에 어두운 곳에서 광선을 받아들이는 감수성은 이곳에서 더 욱 증가하게 된다. 시신경에 하나의 신경정보를 일으키기 위해서는 적 어도 몇 개의 광수용기들이 동시에 작용하지 않으면 안 된다. 몇 개의 광수용기에 빛이 동시에 작용하면 이것이 하나의 절세포로 수령되어 이 절세포가 흥분하게 된다. 따라서 수령비율이 높을수록 어두운 곳에 서도 신경신호가 쉽게 발생된다. 4.1 망막의 기타 세포들 망막에는 간상체, 원추체와 절세포 의에도 · 수평세포 (hor i zon t al cell), 양국세 포 (bip o lar cell) 그리 고 무축색 세 포 (amacrin e cell) 가 있는데 (그림 5-1 참고), 이 세포들은 매우 복잡한 방식으로 연결되어 있다. 쉽게 이 해하기 위해 망막에 있는 세포들을 수평적 기능을 하는 세포군과 수직 적 기능을 하는 세포군으로 나누어볼 수 있다. 수직적 기능 세포는 광 수용기로부터 뇌로 흥분을 전달하는 기능을 담당하는 세포들인데 간상 체와 원추체, 양극세포, 절세포 등이 여기에 포함된다. 수평적 기능을 담당하는 세포는 무축색세포와 수평세포이다. 이 두 세포들은 광수용기충과 절세포충에서 신경흥분을 좌우 수평방향으로 전파하는 역할을 한다. 이러한 수평적 정보통로는 시각계통에 있는 모 든 세포들이 시각수용 분야에 참여할 수 있도록 하는 데 중요한 역할 울 한다. 유인원의 망막에는 약 500 만 개의 수평세포와 약 1 천만 개 내지 1 억 개의 무축색세포가 있다. 수평세포와 무축색세포는 모두 축 색다운 축색을 갖지 않는 것이 특색이다. 유인원의 망막에는 너무나 작은 세포들이 조밀하게 짜여 있어서 최 근에 와서야 이곳에서 일어나는 전기적 활동을 기록할 수 있게 되었 다. 망막에서 일어나는 전기적 활동은 주로 도롱뇽과 두꺼비와 같은 양서류에서 얻어졌다. 망막에서는 무축색세포와 절세포에서만 신경충 격이 발생된다. 망막에 있는 다른 세포들은 제 2 장에서 이미 언급한 것 처럼 시냅스후 전위와 극히 유사한 점진성 전위만 보여준다.
5 수용야 포유류의 망막에 있는 개개 세포로부터 반응을 직접 기록하기는 어 렵지만, 시신경을 형성하는 망막절세포의 축색으로부터 전기적 활동을 기록하는 것은 크게 어렵지 않다. 이러한 축색의 전기적 활동은 1953 년 Ste v en Ku ffl er 에 의해 고양이에서 처음 기록되었다. Ku ffl er 의 연 구는 하나의 시 각세포가 지 배 하는 시 각의 수용야 (recep tive field ) 의 형 태를 이해하는 데 많은 도움을 주었다. 시신경 계통에서 한 신경원의 수야란 그 세포의 발사율이 광선에 의해 영향을 받을 수 있는 망막 상의 영역으로 정의된다. 고양이의 절세포의 전형적인 수용야는 원형이며, 이 원형은 다시 중 심부와 주변부로 나누어진다. Ku ffl er 는 지름이 0.2mm 가 되는 백색 광선을 망막의 여러 부위에 비추면서 시신경로에서 개개 신경원의 반 옹률을 기록하였다. 실험 결과, 반응의 양상이 다른 두 종류의 절세포 가 있음이 발견되었다. 첫째 유형은 흥분 - 중심 (on-cente r ), 억제 - 주변 (off -su rround) 세포이 다. 이 세포의 수용야의 중심부에 광선을 바추면 흥분하지만 주변부에서 광선을 비추면 억제한다. 또한 주변부에서 광 선을 제거하면 흥분이 일어나고, 중심부에서 광선을 제거하면 억제가 일어난다. 두번째 유형 의 세 포는 억 제 -중심 (off -c ente r) , 흥분-주변 (on-s u rround) 유형의 세포이다. 이 유형의 세포 반응은 첫번째 유형의 세포 반응과 정반대이다. 표 5-1 에는 이 두 세포의 특칭을 요약하였다. 개개의 절
표 5-1 홍분一중십 및 억제-중심 세포의 반응 양상 흥분 - 중심 세포 억제-중심 세포 중심부 주변부 중심부 주변부 광선제시 흥분 억제 억제 흥분 광선재거 억제 흥분 흥분 억제
세포는 바로 인접한 절세포들에 비해 각기 조금씩 상이한 수용야를 갖 는다. 하지만 전반적으로 이 세포들의 수용야는 상당 부분 중복된다. 수용야 개념은 시각 계통 전체를 통해 모든 신경원에 공통적으로 적용 될 수있다. 6 X,Y 및 W 세포 포유류의 망막에 있는 절세포는 X,Y 및 W 와 같은 3 종류로 나뉘는 데, 이 세포들의 작용방식은 서로 다르다. 먼저 X 와 Y 세포는 1966 년 Enro t h-Cu g ell 과 Robson 에 의해 발견되었고, W 세포는 S t one 과 Ho ff man 에 의해 1972 년에 확인되었다. 표 5-2 에는 이 3 종류의 절세 포가 보여주는 반응 특성을 예시하였다. 가장 보편적인 세포는 X 세포 로 전체 시신경의 55% 를 차지한다. 그 다음은 W 세포로 40% 를 점하 며 Y 세포는 5% 에 불과하다. 이러한 X,Y,W 세포가 의측술상핵에도 존재한다는 사실이 확인되었다. X 세포는 비교적 제한된 부위에만 수지상돌기를 가지는 중간 크기의 세포로, 전도속도는 비교적 느리다. 이 세포들은 LGN 에서 1 차적으로 시냅스한다. X 세포의 한 가지 특칭은 공간적 가합이 이루어진다는 것
표 5-2 고양이에 있어서 X, y 및 W 신경절세포의 반응 특성 X Y w 세포체의 크기 」 중 (14-22µm) 대 (30-40µm) 소와 중 (8-22µm) 수지상돌기의 분포 제한된 범위 분포 광범위한 분포 다양한 분포 시신경축색에 대한 비 55% 5% 40% 전도속도 느립 (15-23m/sec) 가 장 빠름 (30-40m/ 가 장 느 립 (2-18m/ sec) sec) 흥분발사특칭 지속적 일시적 다양 운동민감성 낮다 높다 다양 무사부위 LGN LGN 과 상구체 상구세
이다. 죽, X 세포는 여러 부분의 수용야로부터 온 흥분을 골고루 수합 한다. 그러므로 이 세포에는 수용야에 제시된 자극들에 의한 흥분과 억제가 서로 균형을 이루기 때문에, 아무런 반응도 일어나지 않는 무 반응점 (null poi n t) 또는 균형 점 (balance poi n t) 도 존재 한다. 이 처 럼 X 세 포는 크기가 중간 정도이고, 신경충격을 느리게 전도하며, 제한된 수 지상돌기를 갖는다는 특칭 때문에 시각자극의 미세한 특칭을 처리하는 데 매우 중요한 것으로 알려져 있다. 그리고 유인원의 LGN 에 있는 X 세포는 색채지각에도 관여하는 것으로 알려져 있다. Y 세포는 크기가 크고, 상당히 넓은 부위로 뻗어가는 수지상돌기를 가지며, 신경충격을 빠른 속도로 전도하는 세포이다. Y 세포의 축색은 LGN 과 상구체로 두사된다. Y 세포는 움직이는 자국에 잘 반응하기 때 문에 운동의 지각아나 자국 강도의 변화를 지각하는 데 매우 중요한 것으로 간주된다. 시신경 중의 40% 가 W 세포인데, 이 세포는 크기가 작은 것과 중간 정도의 것이 혼합되어 있고 전도 속도는 매우 느리며 다양한 크기의 수지상돌기를 갖는다. 이 세포의 축색은 주로 상구체와 극히 제한된 영역의 LGN 으로 투사한다. W 세포는 움직이는 사물을 시각적으로 추 적하는 데 중요한 역할을 담당한다. 어떤 W 세포들은 공간적 가합을 보여주지만, 그렇지 않은 경우도 많다. W 세포는 특성이 매우 다양하 기 때문에 이 세포를 하나의 독립된 범주의 세포로 간주하는 데 의심 울갖는다. 시신경이나 LGN 에 이처럼 다양한 유형의 세포집단이 존재한다는 사실 때문에 시각 계통에는 몇 개의 병행하는 통로가 있어서 형태, 위 치, 운동 및 색채와 같은 시자극의 여러 측면들이 상이한 신경원들의 조합에 의 해 처 리 될 것 이 라는 병 행 처 리 가설 (pa rallel pro cessin g hyp h oth e- s i s) 이 제기되었다.
7 LGN 의 반응 특징 LGN 에 대한 주된 입력은 망막의 절세포이다. 유인원의 경우 망막 절세포에 대한 LGN 의 신경원의 비율은 약 1 : 1 이 된다. 그러나 이 비율은 하나의 절세포가 반드시 하나의 LGN 세포와 시냅스한다는 것 을 의미하는 것은 아니다. 실제로 개개 절세포의 축색은 몇 가닥의 측 지를 뻗어 몇 개의 상이한 LGN 세포와 시냅스하는데, 이러한 형태의 시냅스 배열을 산개 (d i ver g ence) 라고 한다. 이처럼 망막과 같은 감각표면의 상이 LGN 과 감은 뇌의 특정 영역 에 점 대 점의 방식으로 투사되는 것은 다른 감각의 경우에도 마찬가 지이다. Kuff ler 등(1 984) 은, 관련된 정보를 처리하는 데 관여하는 신 경원들은 서로 무리를 짖는 경향이 있는데, 이것이 감각정보의 처리에 있어 하나의 일반화된 원리라고 주장했다. 산개식 시냅스 배열 외에도 개개 LGN 의 신경원은 몇몇의 상이한 절세포로부터 오는 입력정보를 수용하기 도 한다. 이 런 수용형 태 를 수령 (converge n ce) 이 라고 한다. 한 편 LGN 의 신경원들은 시각피질과 같은 다른 뇌 부위에서 들어오는 정보도 받아들이는데, 이러한 피질로부터의 정보수용은 LGN 신경원 들의 반웅 특성을 바꿔놓을 수 있다. LGN 신경원의 수용야도 망막절세포의 수용야와 극히 유사하다. 즉 LGN 신경원도 망막절세포처럼 원형 또는 타원형의 수용야를 가지며, 중심-주변 방식으로 조직되어 있다. 그러나 LGN 신경원의 중심-주변 방식의 조칙 형태는 절세포의 경우보다 더욱 분명하며, 억제 영역이 보다 효과적으로 작용한다. 그러므로 LGN 의 억제 영역을 자국하면 흥분 영역의 자극에 의해 야기되었던 흥분성을 상쇄시킬 수도 있다. 억제 영역의 이런 작용 때문에 산만한 광선은 LGN 신경원에 별 효과 를 야기하지 못한다. 한편 이춘길 등의 연구에서는 고양이의 경우 LGN 의 일부분을 구성 하는 내 측충간핵 (media l int e r lamin a r nucleus) 이 눈의 반사층인 타페 툼 (tap et u m ) 에 의 해 반사되는 망막 부위로부터 입 력을 받아들임 으로써
야간시각에 관여한다는 사실도 발견하였다 (Lee 등, 1984 ; Lee, Lee, & Malpe li, 1992) . 8 색채지각 우리가 수많은 색깔을 볼 수 있다는 점은 실로 경이로운 일이다. 비 록 시각 계통에 포함되어 있는 많은 신경세포들이 색채지각과 관련 있 지만, 그 중에서 LGN 신경원의 색채 지각 기능이 가장 잘 알려졌다. Gouras (19 81) 에 의 하면 인 간은 선홍색 (brig h t red) 으로부터 자청 색 (vio l et blue) 에 이르기까지 200 여 가지의 상이한 색채를 구별할 수 있 다고 한다. 유인원들은 인간과 극히 유사한 색채감각을 갖지만 그 밖 의 다른 포유동물이 진정한 의미의 색채감각을 갖는다고 할 수는 없 다. 그러나 조류, 어류 또는 벌과 갇은 무척추동물들도 광파장을 구분 하는 능력은 보여준다. 스펙트럼에는 ,,선이나 X 선처럼 매우 짧은 파장을 갖는 파동으로부 터 매우 긴 라디오 파동과 갇은 장파장에 이르는 실로 다양한 파장이 있다. 그러나 우리가 볼 수 있는 것은 400-700run 사이에 속하는 극히 제한된 에너지뿐이다(그림 5-5). 가시 스펙트럼상 파장이 짧은 에너지 는 400run 급의 청자색 (blue-v i ole t)이며 파장이 긴 것은 700mn 의 적색 • (red) 이다. 유인원의 눈에는 3 가지의 상이한 광색소를 갖는 3 가지 종류의 원추 체가 있다. 개개의 광색소는 가시 스펙트럼상 각기 상이한 부분의 파 장에 민감한 반응을 보인다. 어떤 원추체는 약 440mn 정도의 광선에 가장 높은 민감성을 보이는데 이 원추체를 청색광 원추체 (blue cone) 라 부르며 정상적인 색채 지각에서 이 영역의 광선을 녹청색으로 본다. 두번째 종류의 원추체는 535mn 의 다소 긴 파장에 민감하게 반응하는 데 , 이 를 녹색광 원추체 (gree n cone) 라 한다. 세 번째 의 원추체는 570 run 의 긴 파장에 민감한 유형인데, 이를 적색광 원추체 (red cone) 라 한
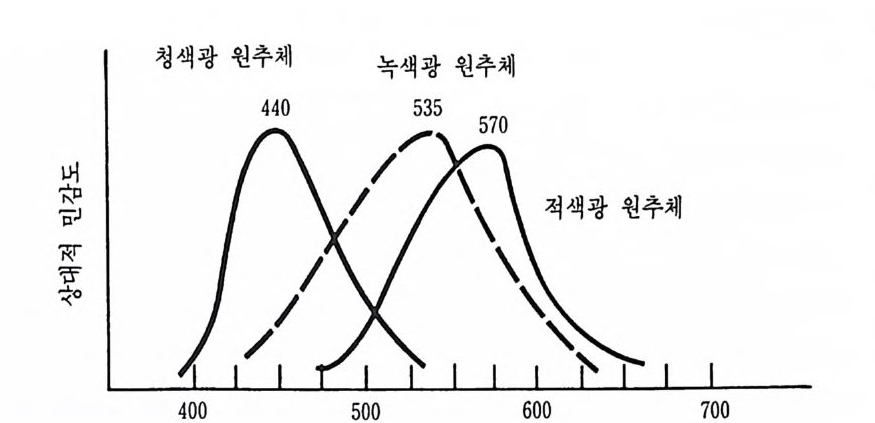 청색광 원추체 녹색광 원추체
청색광 원추체 녹색광 원추체
광파장 (nm) 그림 5-5 유인원의 망막에 있는 3 종류 원추체의 상대적 민감도 다. 이 3 가지 원추체 의 민감곡선은 다소 중복되 어 있다. 예 컨대 500 nm 를 갖는 광선은 이 3 종류의 원추체 모두에 영향을 미친다. 우리의 뇌가 이런 신경충격으로부터 어떻게 색채 경험을 할 수 있게 되는가에 관해서는 LGN 신경원을 논의할 때 다시 언급하게 될 것이다. 8.1 색채의 대립세포란? 스펙트럼상 어떤 제한된 범위 안의 광선(색채광선)을 사용하면 상이 한 광파장을 구분할 수 있는 신경원들이 LGN 에는 포함되어 있다는 사실을 알 수 있다. 이 분야의 연구를 선도한 사람이 Russell De Valo i s 와 그의 동료들이었다 (De Valois & Jac obs, 1968 ; De Valois & De Valois , 1975). 이들은 레서스 원숭이의 LGN 에 있는 개개 신경원들이 상이한 광파장을 갖는 광선에 다르게 반응한다는 사실을 발견하였다. 예컨대 초당 25 번의 자발적 발사를 보이는 LGN 에 있는 한 특정 신경 원은 적색광을 제시하면 초당 50 번의 신경충격을 발사하는 데 반해 녹 색광을 제시하면 초당 1-2 회 또는 전혀 반응하지 않는다. 반응이 흥분 성이든 아니면 억제성이든 반웅의 크기는 동물이 암순응되어 있는 정 도, 광선의 강도 그리고 적용되는 광선의 정확한 파장 등에 따라 달라
진다. 전체적으로 보아 이 분야의 연구들에서는 LGN 에 있는 대부분의 신 경원들은 어떤 범위의 광파장에 대해서는 흥분 반응을 보이지만, 이와 다른 범위의 광파장에 대해서는 반응을 보이지 않는 경향이 있다. 이 러한 신경원들을 색채대립세포 (color opp o nent cell) 라 부르는데, LGN 에 있는 신경원들의 약 75% 가 이에 해당한다. De Valo i s 는 이들 신경 원은 망막의 절세포로부터 입력을 받아들이며 망막에 있는 3 종류의 원 추체 가운데 2 종류의 원추체에 의해 영향을 받는다고 했다. 예컨대 적 색-홍분, 녹색-여제 신경원은 적색광 원추체로부터는 흥분성 입력을 받고, 녹색광 원추체로부터는 억제성 입력을 받지만, 청색광 원추체로 부터는 아무런 입력도 받지 않는다. 그 이후의 연구들에서는 청색광 원추체로부터 흥분을 받는 \ 소수의 LGN 신경원도 있다는 사실을 시사 하고 있는데, 이 세포들은 적색과 녹색 입력에 대해서는 모두 억제적 이다. 또 다른 신경원들에는 녹색-홍분, 적색-억제 세포들이 있다. 청 색광 원추체는 혼치 않지만 중심와에 수용야가 있는 신경원이 여기에 속한다. 이런 현상은 진화적 원인으로 해석할 수 있다. 죽 청색광은 이보다 더 긴 광파장을 갖는 다른 광선보다 분산이 더 잘 되며, 광선 이 분산되면 시력을 약화시키게 된다. 유인원에게는 높은 시력이 생존 의 가능성을 높여주기 때문에, 청색광 원추체가 유인원의 망막 중심와 에 극소수로 존재하는 것으로 추측할 수 있다 (Gouras, 1981). 8.2 LGN 신경원의 이중기능 개개 LGN 신경원은 매우 제한된 수용야를 가지므로 망막상의 어떤 제한된 부분의 자극에 대해서만 반웅을 보인다. 그러므로 이 세포들은 시 야상의 명 백 한 공간적 위 치 (spa t ia l locati on ) 만을 부호화할 수 있으 며, 이러한 반웅 특칭은 자국의 모양 (sha p e) 을 탐지하는 데 이용될 수 도 있다. De Valois 등의 연구에 의해 우리는 LGN 이 색채를 부호화 할 수 있다는 것을 알게 되었다. 그러면 이 LGN 신경원들이 모든 부
호화 과제를 동시에 수행하는가? 이에 대한 해답은 1981 년 시각 계통 신경원들의 반응 특성을 연구한 공로로 노벨상을 수상받은 Torste n W i esel 과 David Hubel 의 연구에 서 찾아볼 수 있다. 이 들은 De Valo i s 가 밝힌 색채의 대립세포가 공간적 위치를 명확하게 부호화하는 동물인 명주원숭이 (s pi dermonke y)의 LGN 신경원에도 있다는 사실을 발견하였다. 이 발견을 접하고 이들은 만약 이 원숭이의 시각 계통에 두 가지의 상이한 기제가 있어서 하나의 기제는 공간적 정보를 담당하 고, 다른 하나는 색채정보를 담당한다면, 이 두 기제의 작용이 동일한 신경통로에서 이루어지는 것인지, 아니면 서로 구분되는 별개의 신경 통로에서 이루어지는 것인지를 규명해 보아야 할 것으로 생각했다 (W ies el & Hubel, 1966). 연구를 거듭한 끝에 이들은 위의 두 가지 작용이 동시에 이루어진다는 사실을 발견하였다. 이들은 16 마리의 원 숭이에서 244 개의 LGN 신경원의 반웅 특징을 검색한 결과 LGN 에는 3 가지의 분명한 세포군이 있다는 증거를 제시하였다. 제 1 유형은 결합세포 (comb i na ti on cell) 로 전체의 77% 정도를 차지한 다. 이 세포들은 중십-주변식 조직과 함께 색채대립 특성도 동시에 보 였다. 예컨대 이 세포의 수용야 중심부는 어느 일정 범위의 광파장에 의해 흥분되었지만 주변부는 상이한 범위의 광파장에 의해 억제되었 다. 그 밖에도 몇 가지 다양한 세포들이 발견되었는데, 적색 흥분_중 심 (red on-cente r ) , 녹색 억 제 -주변 (gree n off- su rround) 이 전체 의 35% 였 고, 적색 억제-중심, 녹색 흥분-주변이 18%, 녹색 흥분-중심, 적색 억제-주변이 16%, 녹색 억제-중심, 적색 흥분-주변이 6%, 그 밖에 청색 흥분-중심, 녹색 억제-주변을 보이는 세포도 있었다. 이러한 결 합세포들은 이 세포들의 중심-주변 조직의 결합에 의해 위치와 모양뿐 만 아니라 대립적인 색채 입력에 따라 색채감도 부호화할 수 있다. 예 컨대 적색 흥분-중심, 녹색 억제-주변 신경원은 스펙트럼상 적색 부분 에 해당하는 광파장을 갖는 단색광이 수용야의 중심부의 한 점에 제시 될 때 최대로 흥분될 것이며, 또한 바로 이 세포는 수용야의 주변 영 역에 들어오는 녹색광에 최대로 억제될 것이다.
W i esel 과 Hubel 에 의해 검토된 세포 가운데 약 16% 는 제 1 유형 세 포의 중심-주변식 조직은 보이지 않았으나 색채대립 반응은 보였는데, 이 세포들을 제 2 유형의 세포 (Typ e II cell) 라고 한다. 이 세포 유형에 도 두 가지 중요 하위 유형이 있는데, 하나는 녹색-홍분, 청색-억제이 고 다른 하나는 녹색-억제, 청색-홍분의 세포이다. 7% 정도의 소수 세포는 오직 중십-주변식 조직만을 보이는데, 이 세포들을 제 3 유형의 세포 (Typ e III cell) 라 한다. 이 세포들은 다양한 광파장에 대한 상이한 민감성을 보이지는 않고 단지 시자극의 모양과 위치만을 부호화한다. 9 뇌피질 신경원:세부특징탐지기인가, 공간빈도분석기안가? 최근에 들어와 대뇌피질의 시각영역에 있는 신경원들의 반응 특징에 관해 많은 관심이 집중되고 있다. 이에 관한 두 가지 견해가 제시되었 는데 , 그 하나는 세 부특칭 탐지 모형 (fea tu r e dete c to r model) 이 고, 다른 하나는 공간빈도모형 (spa t i al freq u e ncy model) 이다. 세부특징모형이 먼 저 제안되었기 때문에· 이 모형부터 살펴보자. 9.1 세부탐지모형 Hubel 과 W i esel 은 대뇌피질의 상이한 시각영역에 있는 신경원들의 반응 특칭을 일련의 실험둘을 통해 광범위하게 연구하였다 (Hubel & Wi es el, 1962 ; 1965b ; 1979) . 이 들의 초기 연구는 주로 고양이 를 대 상으 로 했으며, 그 후에는 레서스 원숭이를 주로 사용했다. 비록 이 두 종 의 동물에서 얻은 결과 사이에는 다소 차이가 있지만, 시각피질 신경 원의 반응 특칭 사이에는 유사성도 많다. Hubel 과 W i esel 은 전극이 삽입된 피질층에 따라 상이한 유형의 세포들이 있음을 발견하였다. LGN 신경원의 축색은 주로 17 번 영역의 IV 및 VI 층으로 두사되며,
제 IV 충의 피질신경원은 LGN 과 극히 유사한 반응 특징, 다시 말해 명 백한 중심-주변 조직 형태를 갖는 원형의 수용야를 보였다. 그러나 제 II, III, V 및 VI 충에서는 이와는 분명히 다른 수용야를 보였다. 9.1 . 1 모서리 탐지기 피질의 IV 충에 있는 신경원들은 특정 각도로 기울어전 선분이나 슬 릿 광선 (s lit ligh t: 좁은 틈을 통해 제시되는 빛)에 잘 반응하는 경향이 있다. 이 세포들을 모서리 탐지기 (edg e de t ec t or) 라 하는데, 그 이유는 각을 이루는 자국, 죽 모서리가 있는 자극에 가장 잘 반응하는 세포들 이기 때문이다. 17 번 영역에 있는 모서리 탐지기는 특정 방향으로 기 울어진 자극에 특히 민감하다. 이 영역에 있는 대부분의 세포들은 10°-20° 정도의 방향성을 갖는 자극에 선택적으로 반응한다. 9.1 .2 단순 및 복합세포 Hubel 과 W i esel 은 미세 기록전극을 사용하여 다양한 시각피질 영역 울 탐색한 결과 세포 반응에서 차이를 보이는 두 개의 하위 영역이 있 음을 알게 되었다. 이들 중 하나는 단순세포 (s imp lecell) 군이고 다론 하나는 복합세 포 (comp le x cell) 군이 다. 단순세 포군은 주로 111 층에 위 치 하는데 자극이 세포의 수용야의 특정 위치에 자리잡거나 어떤 선호적 인 각도와 방향성을 가지고 있을 때 잘 반응하지만 수용야 주변에서 움직이는 자국에는 반응을 억제한다. 이보다 더 많은 신경원들이 복합세포군을 이루는데, 이들은 17, 18, 19 번 영 역 의 제 II , III, V 및 V1 층에 서 주로 발견된다. 복합세포들은 자극의 정확한 위치에 대해서는 민감하게 반응하지 않는다. 그러나 만 약 자극이 정확하게 모서리가 있는 것이라면 수용야의 위치에 관계없 이 이 세포군은 모서리에 대해 잘 반응한다. 복합세포들은 일반적으로 흥분 또는 억제 영역을 별도로 갖지 않는다. 복합세포가 특히 잘 반웅 하는 자극은 수용야를 가로질러 움직이는 모서리 자국이나 슬릿 광선 이다.
적당한 길이의 선분과 같은 특수 자국에 선호성을 보이는 과복합세 포들 (h yp ercom p lex cell) 도 발견되었고, 두 개의 선분이 특정 각도로 만나는 자국에 특히 민감하게 반응하는 세포(구석 탐지기)도 관찰되었 다. 17, 18 및 19 번 영역에 있는 개개의 신경원은 특정한 방향성을 갖 는 각도에 대해서는 반응성을 보이지만, 자극의 기울기가 선호 범위를 벗어나면 반응을 멈춘다. 신경원에 따라 선호하는 방향성이 약간씩 다 르지만 전체적으로 거의 균등한 방향성을 가진다. 시자극의 다양한 세 부특징에 민감하게 반응하기 위해서는 여러 종류의 반응세포가 별도로 존재해야만 이 세부특칭탐지모형이 타당성을 가질 수 있다. 사실 유인 원의 피질에는 약 1 억 개 정도의 시각세포가 있기 때문에 신경원의 양 적인 면에서는 이 모형에 문제점이 없다. Hubel 과 W i esel 은 동물에 게 여 러 가지 약물을 사용하여 마취 시 키 거 나 운동을 못 하게 마비시킨 후에 실험을 실시하였다. 이러한 약물처 치는 실험중에 나타날 수 있는 통증을 경감시키기 위한 것이었지만 제 시되는 자극에 대한 지각적 각성의 정도를 경감시킬 수 있다. 그러므 로 이 연구들은 의식이 없는 상태에서 나타나는 뇌피질 신경원의 반응 울 연구한 것이므로 여기에서 얻어진 결과를 정상적인 조건 아래에서 일어나는 것으로 일반화하기에는 어려운 점이 있다. 그러나 그 후의 보다 발전된 연구들, 예컨대 영구적인 기록전국을 장치한 원숭이를 대 상으로 깨어 있을 때 시각피질에서 나타나는 반응들을 측정한 Wurt z (1969) 의 연구결과도 앞서의 Hubel 과 W i esel 의 연구결과와 유사하다 는 점이 확인되었다. 9.2 공간빈도모형 Hubel 과 W i esel 에 의해 제시된 세부특칭탐지모형이 시각피질신경원 의 반웅 특칭을 설명하는 유일한 모형은 아니다. 최근에 들어와 이 모 형 에 대 한 대 안적 모형 (Camp be ll, 1974 ; De Valois & De Valois , 1 980) 이 제시되었는데, 이 모형을 공간빈도모형 (spa t ia l freq u e ncy model) 이라
 그림 5-6 공간빈도 모형을 설명하는 창살자극 그림
그림 5-6 공간빈도 모형을 설명하는 창살자극 그림
한다. 그림 5-6 의 자극은 평행선들로 구성된 창살 모양의 도형이다. 이 시각 자극은 시야상에 어느 일정한 거리를 두고 몇 개의 흑백선이 교번한다. 다시 말해 이 자극의 특칭은 시야에서 일정 각도를 유지하 는 흑백선분의 교번 정도, 죽 공간빈도 (s p a ti al fr e quenc y)를 가지고 있 다는 점이다. 상이한 공간빈도를 지각하는 데는 실제 빈도뿐만 아니라 흑백 영역 사이의 대비 정도와 기타 요인들 때문에 상당한 개인차를 보인다. 대 부분의 피험자들은 10 의 시각공간 내에서 창살 자극의 공간빈도가 1 에 서 5 사이일 때 가장 민감하게 반응한다. 일반적으로 시각공간을 각도 로 표시하는데 우리가 똑바로 서서 왼쪽 끝에서 오른쪽 끝까지의 전시 야를 180° 로 표시한다. 시각의 예민성, 죽 시력은 한 사람이 간파할 수 있는 공간빈도의 상한계를 측정함으로써 알아볼 수 있다. 일반적으 로 사람의 시력은 10 의 시각공간당 약 48 번의 주기이며 (Camp be ll & Robson, 1968), 레서스 원숭이도 유사한 방법으로 측정해 보면 인간의 경 우와 유사한 결과를 보인다 (De Valois , Morga n & Snodderly, 1974) . 시각피질에서 Hubel 과 Wi es el 방식의 자극으로 검사했울 때 단순 또는 복합세포로 분류될 수 있었던 바로 그 세포들이 창살 자극으로 검사했을 때는 빈도분석기로 분류될 수 있었다. 이런 사실은 고양이 (Maff ei & Fio r enti ni, 1973) 와 원숭이 (De Valois & De Valois , 1980) 의 경
우에도 동일하게 나타났다. 이처럼 신피질의 시각 부위에 있는 신경원 의 반응특칭으로 Hubel 과 W i esel 은 세부특징탐지모형을 제시했고, Cam p bell 과 DeValois 등은 공간빈도모형을 제시했다. 10 양안시 약물투여 상태에서 동물의 시각을 실험해 보면 두 눈이 서로 다론 시야를 갖는다는 사실을 알 수 있다. 마취한 동물에서는 공간 속에 있 는 하나의 점에 두 눈의 초점을 맞추도록 해주는 신경구조나 근육구조 가 정상적으로 작용할 수 없기 때문에 개개의 눈은 시야의 중앙부에 약간의 중복이 있을 뿐, 각각의 독립된 시야를 갖는다. 한쪽 눈에서부 터 온 자극에만 반응하는 신경원을 단안성 (monocular) 신경원이라 하 며, 두 눈에서 온 자국에 대해서 반응하는 신경원을 양안성 (bin o cular) 신경원이라 한다. 두 눈에서 들어온 많은 입력들은 피질에 있는 개개의 산경원에 수령 된다. 고양이의 경우 17 번 영역에 있는 전체 세포의 약 80 .% 가 양안성 신경원이다. 원숭이의 경우 단순세포의 경우는 국소수가 양안성 신경 원이지만, 복합세포는 50% 정도가 양안성 세포이다. 양안성 세포의 경우에도 한쪽 눈이 다론 쪽 눈에 비해 세포의 활동이 더 효과적인 경 우가 있는데 , 이 런 조건을 시 각의 우세 성 (ocular domi na nce) 이 라 한다. 새끼 고양이의 경우 양안시의 정도는 생후 처음 몇 주의 민감기에 경험하는 환경자국에 의해 변화될 수 있다. 죽 만약 새끼 고양이에게 불두명한 콘택트 렌즈를 부착하여 이 시기 동안 다론 사물을 볼 수 없 게 하면 이 고양이의 시각우세성이 병적으로 발달하게 된다. 시각을 폐쇄시킨 눈에서도 기본적인 기능은 이루어지지만, 렌즈를 제거한 후 에 이 눈에 자극을 주면 국소수의 시각피질세포만이 반응을 보인다. 이런 방법으로 한쪽 눈을 폐쇄시켜 입력정보를 차단하면 두 눈의 우세 성이 변화되어 폐쇄되지 않은 쪽 눈의 우세성이 비정상적으로 커진다.
11 기타 시각영역 11. 1 양안시의 결함 : 실험적 사시 사시 (s t rab i smus) 는 시선을 수의적으로 통제할 수 없는 시각장애로 정상과는 달리 두 눈이 병행하여 움직이지 않는다. 사시는 두 개의 상 울 만든다. 사시를 가전 아동들·은 두 눈을 번갈아가며 사용하지만 어 떤 아이들은 한쪽 눈만 사용하기도 한다. 이런 경우 사용하지 않는 눈 의 기능이 유지된다 하더라도 시력은 점점 낮아지게 된다. 이처럼 사 용하지 않는 눈의 시각이 상실되는 것을 약시 (amblyo p i a ex ano p s i a) 라 한다. Hubel 과 W i esel 은 사시를 실험적으로 연구했다. 이들은 생후 4 개월 된 4 마리의 새끼 고양이에게 두 눈의 병행운동을 지배하는 근육 하나 를 절단하여 인위적으로 사시를 만든 후 좌우 번갈아가면서 한쪽 눈으 로만 사물을 볼 수 있도록 습관화시켰다. 이렇게 처치한 후 3 개월이 지나 검사를 했더니 두 눈 모두 정상적인 시력은 보여주었다. 그러나 이 새끼 고양이들(생후 7 개월)의 시각피질에 있는 신경원의 활동을 기 록했더니 79% 의 세포가 단안성이었는데, 이것은 정상적인 경우 20% 가 단안성인 것과 큰 차이가 있었다. 사시의 경우 각각의 눈에서 오는 정보는 정상이지만 두 눈이 서로 협력하여 동시적으로 작용하지 못하 는 것이 잘못이다. 이처럼 두 눈 사이에 정상적인 협력이 이루어지지 않으면 시각피질이 양안작용에서 단안작용으로 바뀌게 된다. 시각피질에 동시 입력이 이루어지지 않으면 양안성 신경원이 줄어든 다는 발견에 이어 Hubel 과 W i esel 은 또 다른 실험을 실시했다. 이 실 험에서 이들은 눈의 근육을 절단하지 않고 콘택트 렌즈를 사용하여 하 루는 왼쪽 눈을 차단하고, 다음날은 오른쪽 눈을 차단하는 실험절차를 3 개월 동안 실시하였다. 3 개월 후 두 눈의 시각은 모두 정상적이었지 만, 80% 의 세포가 단안성을 보여주어 앞서 인위적으로 사시를 일으켰 던 실험결과와 유사하였다. 이 결과는 시각피질에 있는 신경원의 정상
적 양안시는 두 눈이 공간 속에 있는 같은 점에 대해 동시에 초점을 맞추는 기능에 크게 의존된다는 것을 의미한다. 시각피질에 있는 신경원들의 급진적 변화가 언제나 일어나는 것은 아니다. 고양이의 경우는 생애 첫 5 주에서 6 주 사이가 취약성이 큰 시 기인 것 갇으며, 원숭이의 경우는 6 주에서 10 주 사이인 듯하며 (LeVa y , Wi ese l, & Hubel, 1980), 인간의 민감기는 출생 후 수 개월째부터 시작 하여 1 년에서 3 년 사이에 절정기에 이른다고 한다 (Banks, Aslin , & Lets o n, 1975). 양안기능에 대한 표준 실험실 방법을 사용한 연구들에 의하면 사시의 출현이 늦으면 늦을수록 정상화되는 경우가 더 많았다 고 한다. 죽 사시의 출현이 2 살 이전에 시작되면 사시교정수술을 받아 도 양안기능은 계속 비정상성을 보이는 경우가 많다는 것이다. 11. 2 방향선호성 의 발달 자국의 방향선호성을 지배하는 해부학적 연결도 양안시의 경우처럼 유전적으로 결정되고 환경조건에 따라 변화되는가? 이것 역시 양안시 의 경우와 비슷하다는 결과가 제시되었지만 아직은 명확하지 않다. 예 컨대 시각경험이 전혀 없는 새끼 고양이의 시각피질에서도 방향선호성 울 보이는 신경원이 발견되었지만 성숙한 고양이의 신경원에 비해서는 방향선호성이 낮다 (Hubel & Wi es el, 1963 ; Str yk er & Sherk, 1975). 비정상적 시각 경험에 의해 뇌피질에서도 이에 상응하는 변화가 생 긴다는 증거가 있다. 예컨대 만약 새끼 고양이를 한쪽 눈으로는 수직 선만 볼 수 있게 하고, 다른 쪽 눈으로는 수평선만 볼 수 있도록 특수 안경을 씌워 몇 주 동안 사육하면 그 동안 입력을 받아왔던 시각피질 신경원은 그 동안 노출되었던 방향성에만 반응한다(Hi rsch & Sp ine lli ., 1971). Blakemore 와 Coo pe r(1970) 도 이와 유사한 실험을 하였다. 이들은 두 마리의 새끼 고양이 중 한 마리는 하루 몇 시간씩 수평선이 그려진 원통 속에 넣고, 다른 한 마리는 수직선이 그려진 원통 속에 노출시켰
다. 그리고 나머지 시간 동안에는 암실에서 키웠으며, 자신의 신체를 보지 못하도록 특수 목도리를 씌웠다. 5 개월 후에 연구자들은 고양이 의 _ 시각피질 신경원의 활동을 기록하였다. 기록 결과 피질 신경원은 과거에 노출되었던 자국의 방향성과 일치하는 선과 모서리에 대해서만 반응하도록 성장되었음이 밝혀졌다. 이 연구를 비롯하여 그 후 수행된 여러 연구결과에서도 발달 초기의 시각 경험이 시각피질 신경원의 반 응성에 결정적인 영향을 미찬다는 사실이 거듭 발견되었다. 시각피절 속에 있는 신경원들이 명확한 방향성을 갖는 자극에 더 잘 반응한다는 사실에 근거하여 다음과 감은 두 가지 가설, 즉 배양모형 (ins tr u cti on model) 과 유전 설 정 모형 (ge neti c pro g ram mi ng model) 이 제 기 되었다. 배양모형에 따르면, 시각피질에 있는 신경세포들은 어떤 특정 방향성을 갖는 선분이나 각에 반응성을 보일 수 있도록 미리 유전적으 로 설정되어 있는 것이 아니라 생애 초기의 시각적 경험에 의해 배양 된다고 한다. 죽 생애 초기의 민감한 시기에 노출되었던 특정 방향성 울 가진 시각 자극이 다시 나타나면 그 자극에 쉽게 반응한다는 것이 다. 그러나 이 모형은 어떤 신경원은 어떤 방향성을 갖는 자극에만 민 감하게 반응하고, 또 다른 신경원은 또 다른 방향성을 갖는 자극에만 민감하게 반응하게 되는 이유를 설명하지 못하는 약점이 있다. 한편 유전설정모형은 방향성 선호를 전문화시켜 주는 해부학적 연결 이 이미 유전적으로 계획되어 있다고 설명한다. 이 모형은 시각적 경 험을 하지 못한 어린 고양이의 뇌피질 속에도 방향선호성 신경원이 존 재한다는 사실을 잘 설명해 준다. 11. 3 측두엽의 시각영역 유인원의 경우 후두영 의에도 시각에 관여하는 뇌 부위가 많다는 점 이 밝혀졌다. 그 중 하나가 측두영의 복측 부위에 있는 측두하 피질 (infe r io r tem p or al cort ex ) 이 다. 이 부위 가 시 각에 중요한 역 할울 한다 는 것은 Hein r ic h Kluver 와 Paul Buc y (1939) 의 고전적 연구로부터 시
사되었다. 이 연구자들은 메 스칼린 (mescali ne ) 과 갇은 환각제 의 작용 부위 에 관 심을 가졌다. 이 연구자들은 측두영에 종양을 가진 간질 환자들이 간 질발작에 앞서 순간적으로 환각을 경험하기 때문에 측두영이 환각에 관여할 것이라고 생각했다. 그래서 이들은 원숭이의 측두영을 제거하 고, 수술 후 회복과정을 지켜보고 있었다. 그런데 놀라운 것은 이 원 숭이들은 사물을 보지 못하는 시맹이 아닌데도 불구하고 사물을 인지 하거나 확인하지 못한다는 것이었다. 이들은 이런 현상을 정신맹 (ps yc h ic blin d ness) 이 라고 불렀는데 , 이 런 증세 는 측두하피 질 이 제 거 되 면 나타나는 현상이다. 측두하피질에는 사람의 얼굴이나 원숭이의 얼굴과 같은 얼굴자극에 선택적으로 반응하는 〈 얼굴탐지 (fac e dete c to r )> 신경원이 있다는 사실 이 밝혀졌다 (Bruce , Desim o ne, & Gross, 1981 ). 그림 5-7 에서는 이 신경 원이 반응성을 일으키는 자국과 반응성울 일으키지 않는 자극을 예시 하였다. 이 신경원은 다른 어떤 자국들보다 얼굴자극에 더 잘 반응한 다. 이 신경원들의 정확한 역할이 무엇인가에 관해서는 아직 논쟁중이 지만 인간의 경우 측두영이나 후두영이 손상되면 아주 기묘한 시각장 애, 즉 가까운 친척들의 얼굴마저도 알아보지 못하는 장애를 보이는 수가 있다. 어떤 환자들은 사진에 있는 자기 자신의 얼굴이나 거울에 비친 자신의 얼굴마저도 알아보지 못하는데 이런 현상을 얼굴실인증 (pro sop ag n o sia ) 이 라 한다. 원숭이 사회와 같이 복잡한 사회생활이 이루어지는 상황에서는 상대 방의 얼굴표정을 신속히 알아차리는 일이 매우 중요하다. 따라서 어떤 특정 신경원 집단이 얼굴인지 기능을 잘 하도록 진화되어 왔을지도 모 른다. Bruce 등 (1981) 은 일련의 실험을 통하여 유인원의 측두영에는 얼굴재인 신경원이 존재한다는 사실을 증명하였으며, 이 발견 의에도 측두영과 편도체에서 얼굴선택 신경원이 발견된다는 또 다론 보고들도 있다 (Rolls, 1984 ; Perrett 등, 1985).
 殿 翁
殿 翁
그림 5-7 얼굴 자극에 잘 반응하는 측두엽에 있는 한 신경원의 자극에 대한 반응특징. 얼굴 속에 눈을 없애거나, 얼굴 부분들을 산만하게 하거나, 얼굴이 아닌 손과 같은 자국을 제시했을 때는 반웅이 사라지는 것에 주목할 것.
11. 4 기타 시각영역 뇌의 많은 부위가 시각자극에 의해 영향을 받는다. 시각 계통의 어 떤 부분은 다른 행동을 조절하기 위해 시각정보를 사용한다. 예컨대 시상하부의 시교차상핵 (sup ra chia s matic nuclei : SCN) 은 눈으로부터 온 시각정보를 이용하여 밤과 낮의 주기와 같은 환경적 조건에 적합하도 록 행동의 일내주기를 조절한다. 행스터는 망막의 절세포로부터 나온 축색 가운데 약 1% 정도가 SCN 에 두사되는데, 망막에서 SCN 으로 두사되 는 이 신 경 로를 망막시 상하부로 (reti no hyp o th a lamic tra ct) 라고 한 다. 망막시상하부로가 손상되면 시각 식별에는 장애가 없지만 행동을 밤 낮의 주기에 맞추는 기능에 장애가 나타난다. 반면 망막시상하부로나 SCN 은 손상시키지 않고 망막에서 LGN 으로 가는 시산경로나 시각피 질을 손상시키면 행동상 실명 상태가 나타나지만, 환경자극의 변화에 따른 주기적 행동에는 영향이 나타나지 않는다. 시 각 계통의 또 다론 중요 구조는 상구체 (supe ri o r coll icu lus) 이 다. 이 구조는 시개(t ec t wn) 라 부르는 중뇌 영역의 배측표면에 위치한다. 시개의 바로 전방 쪽으로 시각기능을 담당하는 또 다른 구조로 전시개 영 역 (pre te c t al area) 이 란 곳이 있는데 , 이곳에는 빛 에 대 한 동공반사에 관여하는 회로들이 있다. 상구체는 표피부와 십층부의 두 부분으로 나누어진다. 표피부를 구 성하는 세포충은 망막의 Y 세포와 W 세포로부터 울라온 시각정보를 수 용한다. 또한 이곳은 일차시각피질, 일차시각인접피질 그리고 전두안 야(fr on t al eye fi eld) 라고 부르는 전두영 부위에서 온 정보도 수용한다. 상구체의 심층부는 망막에서 오는 정보를 직접 받지 않고, 뇌의 다양 한 영역들, 예컨대 시각피질, 청각피질, 운동피질 또는 피질하구조로 부터 온 정보를 수용한다. W urt z 와 Albano(1980) 는 상구체의 심부층과 표피충 사이에는 기능 상 차이가 있다고 주장한다. 죽 표피충은 시각과 관련 있지만, 심부층
은 자국에 대 한 정 향반웅 (orie n ti ng respo n se) 과 안구운동, 특히 단속성 운동 (saccades) 과 관련 있는 신경원을 갖고 있다. 단속성 운동이란 시 야 속에 관심을 끄는 새로운 목표자극을 향해 섬세하면서도 신속하게 시각 고정점을 계속 변화시키는 안구운동을 지칭한다. 원숭이의 상구체 표피충에서 개개 신경원의 활동을 기록해 보면 수 용야의 크기가 조금씩 다른데, 이 수용야들은 자극의 크기, 모양, 방 향 또는 운동속도 등에 큰 영향을 받지 않는다. 특정 신경원들이 어떤 특정 자국에 대한 반응으로 나타내는 신경충격의 수는 그 자국이 동물 에게 어떤 의미를 부여하느냐에 따라 달라진다. 예를 들어 각성상태의 원숭이에게 단속성 운동을 유발시킬 수 있는 목표자극을 제시하면 이 자국을 선호하는 상구체의 특정 신경원만이 반응을 나타낸다. 그 후 학습시행이 거듭되어 피험동물이 그 자극에 주의집중을 많이 하게 되 면 이 신경원에 대한 반응비율도 상대적으로 증가된다. 또한 고양이 를 대 상으로 한 이 춘길 동 (Lee, Rohrer, & Spa r ks, 1988) 의 연구에 의하면 단속성 운동 전후에 많은 수의 상구체 신경원들이 활성화되며, 이 단속성 운동의 크기와 방향은 활성화된 신경원들의 공 헌이 벡터 평균치에 의해 결정된다고 한다. 그리고 단속성 운동의 속 도는 활성화되는 신경원의 수에 따라 결정된다고 한다 (S p arks, Lee, & Rohrer, 1990) . 상구체는 다양한 양상의 자국들에 대한 정향반응에도 관여한다. 죽 심부충에 있는 대부분의 신경원들은 눈의 정향반웅뿐 아니라 두 가지 이상의 감각양상의 입력정보에도 반응한다. 이 신경원들이 바로 단속 성 운동과 관련하여 반응하는 신경원들인데, 이 세포들 가운데 70% 이상이 두 가지 이상의 상이한 감각양상에 반응하는 다중기능세포들이 다. 아러한 사실들을 기초로 할 때 상구체는 시각자극 자체에 반응한 다기보다는 주의집중을 요구하는 다양한 자극에 대한 정향반응에 관여 하는 기구라고 할 수 있다.
11. 5 뇌 손상과 시각 뇌 손상 환자를 대상으로 한 사례연구가 많이 보고됨에 따라 시각 계통에 대한 이해도 증가되었다. 일반적으로 이런 경우의 뇌 손상은 신경해부학자가 규정하는 해부학적 부위에만 국한되지 않는다. 그런데 도 불구하고 사례연구를 거듭하다 보면 임상적 증후와 뇌 손상 부위 사이에 어떤 일관성이 있다는 사실을 알게 된다. 예컨대 일차시각피질 (17 번 영역)이 손상되면 이 피질영역과 기능적으로 관련되어 있는 시야 의 어떤 부위에 맹점이 생긴다. 손상이 17 번 영역 전체에 걸쳐 일어나 면 완전한 시맹 상태가 되지만, 손상 부위의 크기가 작으면 시야상에 작은 부위에만 맹접이 일어나는 이른바 암점 (sco t oma t a) 이 발생된다. 시각피질의 손상이 곧 시맹 (b li ndness) 을 야기한다고 일반화하는 것 은 잘못이다. 암접 안에서도 약간의 시각기능이 이루어지지만 환자 자 신은 이런 시각적 감각을 의식하지 못한다. Weis k rantz 등 (1974) 은 뇌 종양을 제거하기 위해 시각피질을 거의 완전히 제거한 한 환자의 사례 를 보고했다. 이 환자는 뇌 손상 영역에 상응하는 시야의 부위에 밝은 빛이 제시되어도 사물을 보지 못했다고 한다. 그런데도 실험자가 시맹 영역 안의 어느 곳에 자극이 제시되고 있는가를 추측해 보라고 하면 이 환자는 우연수준 이상으로 그 위치를 잘 맞추며, 더구나 제시한 자 극이 x 인가 아니면 0 인가를 알아맞춰 보라고 했을 때도 아무것도 보 이지 않는다고 하면서도 계속 우연수준 이상으로 잘 맞추었다. 이처럼 의식적인 시각 경험 없이 시자극을 식별하는 이른바 맹시 (b li nds ig h t)는 상구체에 의해 중계되는 현상으로 추측된다.
11.6 계열처리 가설 대 병행처리 가설의 재음미 지금까지 우리는 시각 계통을 논의하면서 광선 에너지가 망막의 광 수용기에 미치는 영향으로부터 측두영에 있는 얼굴탐지 신경원의 기능 까지 살펴보았다. 시각 계통의 어느 한 부분에 대한 입력은 곧 다음
부분에 대한 출력이 된다. 전통적으로 시각 계통은 조잡한 원초적 정 보가 망막에서 출발하여 LGN 으로 투사되고, 이곳에서 어느 정도 처 리된 정보가 17 번 영역으로 들어가며, 이어 18 및 19 번 영역을 거쳐 마지막으로 측두영과 같은 대뇌피질에 이르는 일련의 계열처리체계로 간주되어 왔다. 계열처리가설 (ser i al pro cessin g t heo ry)은 몇 가지 가정을 전제로 한 다. 죽 시각의 1 차 피질 수용영역은 17 번 영역뿐이며, 18 번이나 19 번 과 같은 피질영역은 2 차 처리영역으로 간주하고, 1 차 또는 2 차영역으 로 분류될 수 없는 피질영역도 있는데, 이를 연합피질 (asso ci a ti on co rt ex) 이라 한다. 연합피질은 해부학적으로는 시상으로부터 직접 입 력정보를 받지 않는 피질로서 이 영역들은 유인원 특히 인간에서 양적 으로 크게 발달되어 있다. 쥐와 갇은 설치류는 연합피질이 매우 작고, 육식동물은 좀더 크고, 유인원에서는 매우 크다. 전통적으로 시각에 관한 이론은 주로 계열처리가설이 중심 가설로 받아들여져 많은 실험 적 증거들이 이 가설을 지지해 왔지만, 1980 년대 이후 이 가설에 대한 대안가설로서 병행처리가설이 등장하여 주목을 끌고 있다. 병 행 처 리 가설 (pa rallel pro cessin g the ory ) 의 주장과 이 를 입증하는 근 거들은 주로 Merzen ic h 와 Kaas(1980) 에 의해 제시되었다. 이들은 시 각의 병행처리가설은 〈 지각이란 많은 위상적 감각표상들이 거의 동시 적으로 활동함으로써 나타나는 결과 (Perce pti on is a pro duct of a nearly sim ulta n eous acti va ti on of a number of top o g r al) h i c sensory repr e senta - ti ons) 〉 라는 견해에 그 근거를 둔다. Merzenic h -Kaas 모형은 새로운 신경해부학적 기법을 사용한 실험결과에서 나온 것인데, 이는 과거에 우리가 알고 있었던 것보다 훨씬 다양한 망막표상이 뇌피질에 존재한 다는 사실을 시사한다. 예컨대 부엉이원숭이의 경우 후두피질에 10 개 의 분명한 위상적 망막지도가 표상된다는 사실이 증명되었으며, 레서 스 원숭이의 경우도 여러 개의 시야가 표상된다고 한다 (Van Essen & Maunsell, 1983). 따라서 시각정보를 수용하는 1 차 피질영역이 단 하나 라는 종래의 견해는 이 발견들에서 지지되지 않는다.
시각피질이 아닌 다른 감각피질들도 여러 개의 1 차 피질 수용영역을 갖는다는 사실이 밝혀졌는데, 체성감각의 경우는 7 개 이상, 청각의 경 우는 4 개 이상의 피질영역이 있다고 한다 (Merzen ic h & Kaas, 1980). 미 각과 후각의 경우도 피질수용영역은 있지만 감각수용표면에 대한 전정 한 의미의 공간적 수용야는 없는 것 같다. 이처럼 최근에 알려진 수용 야들은 단순히 서로를 복사하는 수준에서 작용하는 것이 아니다. 시각 피질영역은 해부학적 또는 기능적으로 볼 때 약간씩 중복되기는 하지 만 독특한 방식의 해부학적 연결을 이루고 있고, 고유한 반응속성을 가진 신경원들을 포함하고 있는 것 같다. 이러한 해부학적 발견으로 인해 전통적인 연합피질에 대한 개념이 흔들리게 되었다. 궁극적으로 중심열 뒤에 있는 모든 피질은 여러 종 류의 감각수용야로 구성되기 때문에 과거에 연합피질로 분류되었던 빈 공간이 점차 좁아지게 되었다. 따라서 과거에는 연합피질이 보다 진화 된 종의 뇌피질에서 더욱 넓어진다는 견해가 지금에 와서는 감각정보 처리에 관여하는 피질의 수용영역의 수가 더 늘어난다는 가설로 바뀌 고 있다.
제 6 장 기타 감각 우리 인간은 앞 장에서 보았던 시각 의에도 청각, 촉각, 온도감각, 미각 그리고 후각과 같은 각종 감각을 자극하는 여러 형태의 에너지에 반응할 수 있고, 신체 균형의 변화를 감지하는 평형 감각도 가지고 있 다. 우리가 이러한 특정 형태의 에너지에 반응할 수 있는 능력은 그 형 태의 에너지를 신경활동으로 전환(t ransduc ti on) 하고, 전환된 신경충격 을 적절한 뇌영역으로 전달(t ransm i ss i on) 하며, 전달된 신호로부터 의 미를 축약 (abs t rac ti on) 하는 일련의 기능에 의한다. 뇌로 들어오는 감 각정보 가운데 어떤 것들은 전혀 의식되지 않은 채 자동적인 반사반응 만을 일으키는 경우도 있다. 우리는 이 장에서 시각을 제의한 몇 가지 감각들의 자국 속성, 감각기구의 구조, 전환과정, 뇌에 이르는 주된 통로 그리고 고위 뇌중추에서 이러한 신호로부터 의미를 축약해 내는 과정을 살펴볼 것이다. 1 청각 소리를 들을 수 있는 능력은 귓속에 있는 수용기에 일차적으로 의존 하며, 이 수용기는 청신경 (VID 신경)을 통해 뇌간, 시상 및 신피질에 있
는 여러 청각정보 처리영역으로 신호몰 전달한다. 소리를 전달하는 에 너지의 변화는 공기, 물 또는 뼈와 같은 탄성 매체를 통해 전파되는 기계적인 동요, 즉 진동이다. 1.1 소리의 물질적 • 심리적 성질 자연적으로 생성되는 소리의 파동은 매우 복잡하지만, 단순파동이든 복합파동이든 모든 파동은 기본적으로 빈도(fr e q uenc y), 진폭 (am pli tud e) , 복잡성 (comp le xit y) 이 라는 3 가지 속성 을 갖는다. 소리 의 빈도란 초당 이루어지는 파동의 수로 표시되며, 빈도의 단위는 독일의 물리학 자 Hein r ic h Her t z(1857-1894) 의 이름을 따라 Hz 로 표시한다. 정상적 청각을 가진 젊은 사람은 20-20,OOOHz 의 소리를 둘을 수 있지만, 나 이가 많아질수록 높은 빈도의 소리를 듣기가 어려워진다. 박쥐, 고래 또는 돌고래와 같은 동물은 100, OOOHz 이상의 고주파 소리도 들을 수 있다. 파동의 진폭은 주어전 파동의 높낮이, 죽 파저와 파고 사이의 거리 로 표시된다. 이것은 소리의 강도, 즉 소리의 압력에 해당하는 것으로 데시벨 (de ci bel: dB) 이란 단위로 나타낸다. 일상적으로 자주 들을 수 있는 몇 가지 소리에 대한 강도, 즉 dB 이 표 6-1 에 나타나 있다. 또 하나의 개념으로 파장 (wavelen gt h) 이란 것이 있는데, 이것은 연속적인 두 파동 사이의 거리, 죽 두 파동의 파고 사이 거리를 지칭하는 것이다. 음의 강도와 관련 있는 십 리 적 특성 을 소리 의 세 기 (lou dness) 라고 한 다. 음의 강도 (inte n sity ) 는 음압기 (sound level mete r) 라는 특수 장치 를 사용하여 측정할 수 있지만, 음의 세기는 소리를 듣는 사람의 심리적 특성에 따라 달라전다. 소리의 세기는 sone 이란 단위로 측정되는데, 1 sone 이 란 1, OOOHz 에 40dB 의 소리 크기로 지 각되 는 단위 를 말한다. 음의 강도가 소리 크기의 주요 요인이 되지만 음의 빈도 또한 중요하 다. 소리의 높이, 죽 고저{pit ch) 는 빈도수와 주로 관련 있다. 음색 (ti mbre) 이란 같은 높이와 같은 세기를 갖는 두 음 사이의 차이룰 나
표 6-1 일상 생활에서 쉽게 들을 수 있는 소리의 dB 소리 Dec ibe ls (dB) 조용한방 40 일반적인 가정 50 일상의 대화 60 고속도로에서 50m 떨어진 지점 70 자명종시계 80 분주한거리 85 대형 화물차, 시끄러운 대화 90 지하철 97 오토바이나 소형 착암기 소리 100 잔디 깍는 기계 소리 110 록음악 115 트랩에서 둘리는 제트기 소리 117 발사중인 로켓 180 타내는 것으로서 소리의 질이라고 할 수 있으며, 이 음색은 소리자극 의 복잡성에 따라 달라진다. 1.2 귀의 해부학 인 간의 귀 는 의 이 (exte r nal ear) , 중이 (mi dd le ear) 및 내 이 (inn er ear) 의 세 부분으로 구성된다(그립 6-1). 의이는 귓바퀴와 의청도 (ear canal) 로 구성된다. 읍의 파동이 의청도로 진행되면 먼저 고막 . (tym p a nic membrane) 을 두드리게 된다. 고막은 의청도를 막고 있으 며, 의이와 중이를 갈라놓는 구조이다. 중이의 특칭은 청소골 (oss i cle) 이라는 세 개의 섬세한 뼈 구조로 되어 있는데, 이것은 고막에서 일어 나는 진동을 내이에 있는 난원창 (oval w i ndow) 으로 전달하는 역할을 한다. 고막으로부터 난원창에 이르는 이 세 가지의 청소골은 순서에 따라 추골 (malleus) , 침 골 (inc us) , 등골 (sta p e s ) 이 라 부른다. 청 소골은
전동을 고막으로부터 난원창으로 전달하는 동시에 귓속의 임피던스 정 합 (imped ance matc h in g ) 에 중요한 기 능을 한다. 임피던스(im p edance) 란 에너지의 흐름을 저지하는 것으로서, 이 개 념은 전기적 흐름이나 기계적 진동 모두에 적용될 수 있다. 귓속의 경 우 진동에 대한 저항은 공기로 차 있는 중이보다는 체액으로 차 있는 내이에서 더 크다. 그러므로 내이의 체액은 중이의 공기보다 임피던스 가 더 크다고 할 수 있다. 만약 이러한 내이와 중이 사이의 임피던스 차를 최소화시켜 주지 못하면, 고막을 치는 음파 에너지의 약 97% 는 반사되어 버리고 단 3% 만이 내이로 전달될 수 있다 (Evans , 1982a). 그 러나 임피던스 정합 때문에 고막을 치는 음파 에너지의 약 60% 가 내 이로 전달될 수 있다. 임피던스 정합은 고막의 면적이 난원창의 면적보다 17 배나 더 크기 때문에 가능하다. 즉 넓은 고막 부위에서 보다 좁은 부위인 난원창으 로 압력을 집중시켜 줌으로써 진동의 강도를 높여준다. 청소골의 지레 작용도 부분적으로 임피던스 정합에 기여하는데, 이 작용에 의해 압력 이 기계적으로 약 1. 3 배 증가된다 (Kell y, 1981a). 이러한 두 가지 요인 의 상승작용으로 고막에 가해진 압력은 약 22 배. (17X l. 3) 증가된다. 청 소골에 부착되어 있는 근육이 수축되면 중이의 임피던스 정합특성이 변경되어 난원창에 가해지는 진동의 강도를 낮추게 된다. 매우 강한 소리에 의해 반사적으로 촉발되는 이러한 음압의 감소는 lOOHz 당 30 dB 정도나 되어 매우 중요한 방어기제가 된다. 청소골에 부착된 이들 근육둘은 제 V 및 VII 뇌신경에 의해 지배된다. 제트기 엔진 소리와 갇 이 대단히 높은 소리는 내이의 수용기를 파괴할 수도 있는데, 일단 수 용세포가 상실되면 대치되지 않기 때문에 청각장애가 생기게 된다. 1.3 내이 : 와우각 내이는 소리를 듣는 기관인 와우각 (cochlea) 과 평형감각을 담당하는 난형 낭 (utr icle ) , 구형 낭 (saccule) 및 3 개 의 삼반고리 관 (semi circ ular
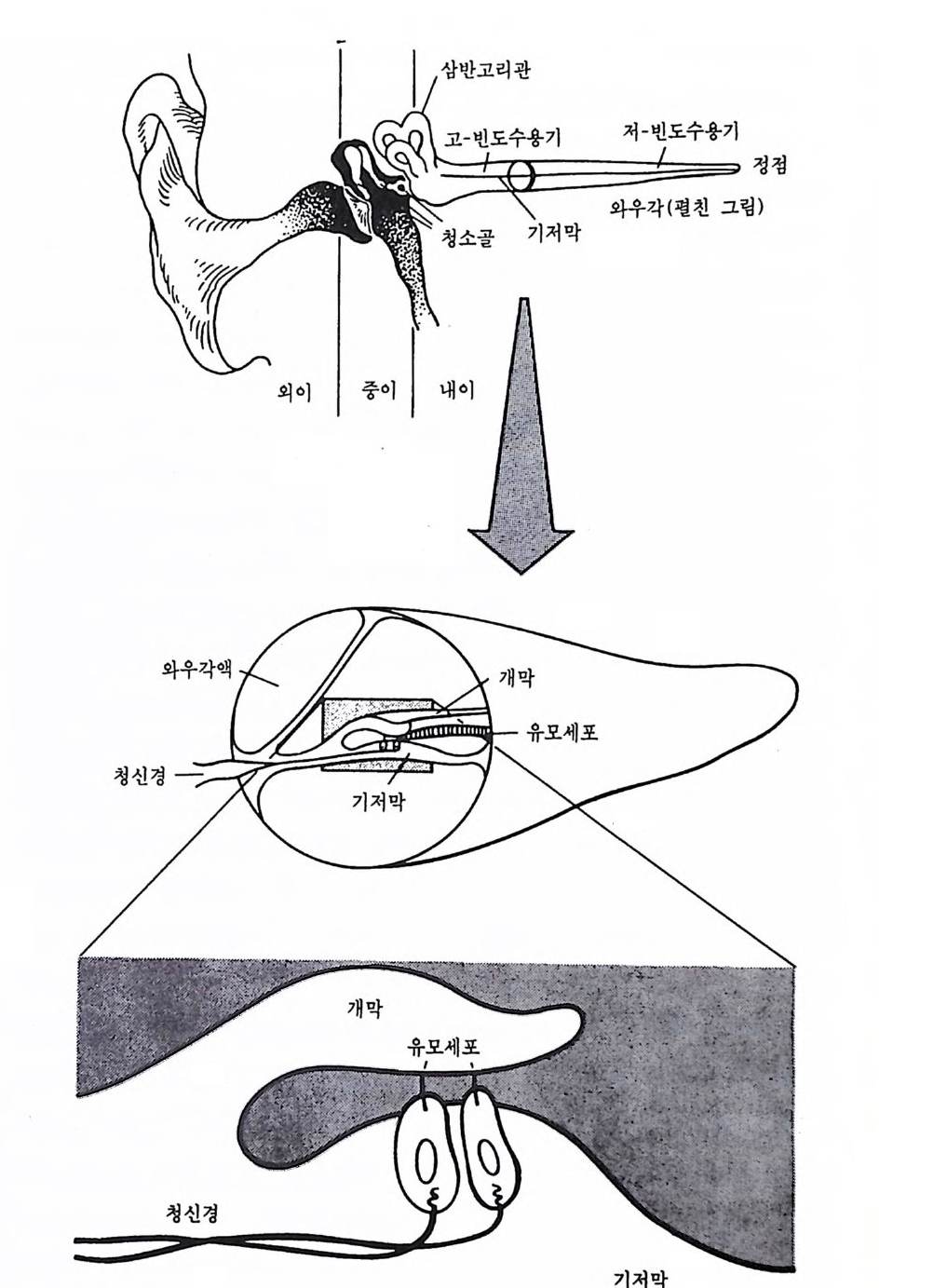 의이
의이
그림 6-1 청각기구 소리의 진동은 의청도, 고막, 청소골, 난원창을 거쳐 와우각으로 전달된다.
canal) 으로 되어 있다. 와우각은 마치 작은 달팽이 껍질처럼 생겼는데 (cochlea 란 말은 달팽이 껍질이란 라틴어에서 유래된 말이다), 인간의 귀 에 있는 와우각은 약 두 바퀴 반 정도 감겨 있으며, 이를 똑바로 펴면 약 3.5cm 정도의 길이가 된다. 이 관의 내면은 몇 개의 막에 의해 전 정 계 (scala vestib u li ) , 고실 계 (scala tympa ni) 및 중간계 (scala media ) 와 같은 3 개의 통로로 나누어진다 (scala 란 계단 또는 통로를 가리키는 라틴 어이다). 고실계의 입구는 난원창과 인접하는 곳으로 매우 엷고 탄력 성 있는 영역으로 청소골의 마지막 구조인 등골로부터 음의 압력이 난 원창을 거쳐 내이로 들어오는 곳이다. 중간 통로인 중간계에는 와우각 내의 체액전동을 신경충격으로 전환 시켜 주는 기능을 담당하는 감각수용기가 있다. 이곳에 있는 감각수용 기를 유모세포 (ha i r cell) 라고 하는데, 이것은 이의 지지세포들과 함께 중간계의 밑바닥, 죽 기저막 (bas il ar membrane) 에 파묻혀 있다. 그림 6-1 에는 청각기 관, 죽 코르티 기 관 (or g an of Cort i) 의 중요 구조를 나타 내고 있다. 기저막은 기저부 (lOOµm) 에 비해 정단부 (500µm) 가 훨씬 더 넓고 기저부가 정단부보다 더 딱딱하다. 코르티 기관의 수용기는 4 열 의 유모세포로 구성되는데, 이들은 기저막 위에 배열되어 있으며 위로 는 개막(t ec t or i al membrane) 이란 막으로 덮여 있다. 1.4 와우각 내에서의 전환 등골이 움직여 난원창을 진동시키면 이 전동은 와우각 내의 체액에 의해 즉각적으로 기저막에 전달된다. 청각에서 기저막의 운동이 어떤 역할을 하느냐에 대한 이해는 1961 년에 노벨 생리학상을 수상한 George von Bekes y (1899-1972) 에 의해 주로 이루어졌다. Bekes y는 인 간의 사체에서 와우각을 적출하여, 등골의 운동이 난원창으로 전달된 후 기저막에서 어떤 순서를 거쳐 음파 이동이 이루어지는가를 알아보 는 특이한 실험을 실시하였다. 청각을 일으키는 신경활동은 기저막의 운동에 의해 유모세포에 가해
지는 힘과 유모세포의 섬모를 덮고 있는 개막에 의해 유모세포에 가해 지는 힘이 서로 결합되어 일어난다. 예컨대 당신이 혼들거리는 현수교 위에 두 발을 밀착시키고 서 있으며(유모세포), 머리털은 움직이지 않 는 천장(개막)에 묶여 있다고 가정해 보자. 천장은 교량과 분리되어 움직이지 않기 때문에 교량이 혼들릴 때마다 머리털은 당겨지고 비틀 리게 될 것이다. 유모세포의 바닥은 혼들리는 기저막에 파묻혀 있고, 유모세포의 정단부에서 솟아나온 섬모(cili a) 는 바로 그 위에 있는 개 막에 의해 덮여 있다. 유모세포의 섬모가 기저막의 불균형적인 운동과 개막에 의해 당겨지고 비틀리어 변형되면 유모세포에서 수용기 전위 (recept or po t en ti al) 가 발생한다. 수용기 전위는 제 2 장에서 이미 보았던 시냅스후 전위와 여러 가지 의미에서 유사하다. 이 수용기 전위가 어 떻게 발생되는지는 아직 명확하게 밝혀져 있지 않지만 유모세포에 가 해지는 힘, 즉 전단력 (shearin g fo rce) 에 의해 이온통로가 열리고 (K+ 와 Ca2+), 그 결과로 이온의 유입이 이루어져 유모세포가 탈분극화될 것 으로 추측된다 (Huds p e th, 1983). 유모세포는 신경원이 아니므로 축색이 없고 신경충격도 야기할 수 없다. 그러나 유모세포에서 생긴 수용기 전위가 유모세포로부터 신경전달물질을 방출시켜 이 세포와 시냅스하 고 있는 와우신경원에 영향을 미칠 수 있다. 인간의 와우각에는 약 15,000 개의 유모세포가 있는데, 그 중 3,000 개는 내유모세포이고, 12,000 개는 의유모세포이다. 이 유모세포는 청 신경 (VIII 신경 ) 의 한 가닥인 와우신경 (cochlear nerve) 을 구성 하는 축색 과 시냅스한다. 와우신경은 약 50,000 개의 신경원으로 구성되는데, 그 중 약 90% 가 내유모세포와 시냅스하며, 약 2-3 천 개의 와우신경원이 의 유모세포와 시냅스한다. 뇌로 흥분을 전달하는 구십성 신경원 의에도, 뇌간에서부터 뻗어나온 1,800 개의 원심성 신경원도 있다. 이 원십성 축색은 와우신경의 구십성 신경원이나 의유모세포와 시냅스한다; 이 원심성 신경원은 유모세포의 활동을 억제하여 높은 소리에 의한 과잉 자극으로부터 유모세 포를 보호한다는 증거 가 있다 (Gul ic k, Ges- cheid e r, & Frisi n a , 1989) .
1.5 장소설 소리의 빈도가 변함에 따라 기저막 내에서의 최대 진폭 위치가 달라 진다는 사실은 소리의 고저지각을 설명하는 두 가지 이론 중 하나인 장소설(p lace th eo ry)의 기초가 된다. 이 이론에 따르면 난원창과 연결 되는 와우각의 기저부는 고주파에 반응하고, 와우각의 정단부에서는 저주파에 반응한다는 것이다. 비록 소리의 빈도가 달라지면 기저막 내 의 반응 위치가 달라지지만, 이동음파가 일으키는 최대 진폭 위치가 너무 넓기 때문에 인간의 민감한 고저 식별 능력을 설명하는 데는 문 제가 있다. 정상적인 청각 기능을 가진 사람들은 주파수의 차이가 3-4 Hz 정도만 되어도 두 음을 식별할 수 있다. 그러나 아동하는 음파는 넓은 부분을 자국하기 때문에 미세한 차이를 가진 두 음에 대해 각기 상이한 반웅 위치를 가정하기는 어려우며, 이러한 현상은 음의 주파 수가 낮을 경우 (800-900Hz) 특히 현저하다. 그러나 고주파의 경우 (4, OOOHz-5, OOOHz) 는 기저막에서 이동하는 음파의 정점이 대단히 민감 하게 구획화되므로 장소설은 상당한 설득력을 갖는다. 최근 Khanna 와 Leonard (1 982) 는 다양한 주파수에 대한 기저막의 정조(t un i n g)는 일찍 이 Bekes y가 시사했던 것보다 훨씬 더 예민하다는 사실을 예시하였 다. 그러므로 장소설은 높은 주파수를 갖는 음의 식별을 설명하는 데 는 타당한 것으로 생각된다. 1.6 제사설 소리의 고저지각에 대한 또 하나의 중요 이론은 와우신경에 있는 상 이한 신경원들이 상이한 빈도의 음에 반응한다는 데 기초를 둔 것이 다. 이 생각은 음의 빈도가 몇 백 Hz 정도가 되는 소리자극에는 합당 하지만, 그 이상의 빈도를 갖는 소리에 대해서는 신경원의 반응에서 불웅기가 있기 때문에 합리적으로 설명하기가 어렵다. 앞 장에서 이미 본 것처럼 신경원의 반응에서 불웅기는 한 신경원이 신경충격을 발생
시킬 수 있는 빈도의 상한계를 규정하게 된다. 예컨대 어떤 신경원의 절대불웅기가 0 . 001 초라고 한다면 이 신경원은 아무리 1 강한 소리가 제 시되어도 반응할 수 있는 최대 반응빈도는 500Hz 를 넘지 못한다. 따 라서 음의 고저지각이 단순하게 음의 빈도에 따른다는 생각은 합당치 못하다. 이런 난점을 해결하기 위한 한 방법이 Ernest Wever(1949) 에 의해 제안되었다. Wever 는 와우신경 내에 기능적으로 밀접한 관련성을 가 진 신경원 집단들이 제사적 (volle y)으로 일제히 흥분을 발사함으로써 소리의 빈도를 부호화할 수 있다고 주장하였는데, 이 견해를 제사설 (volley theo ry ) 이 라고 한다. 이 이론에서는 음의 강도지 각도 개 개 신경 원들의 반응률이 증가함에 따라 부호화될 수 있다고 주장한다. 실험동 물에서는 이러한 과정이 일어난다는 직접적 증거가 제시되어 있지만, 청신경에 있는 신경원은 약 5 , OOOHz 까지는 이런 방식의 제사적 흥분 이 가능해지면 최고위 수준의 뇌에서는 음의 빈도에 대한 부호화의 상 한계가 이보다 훨씬 낮아 대체로 1, OOOHz 정도의 음파까지만 부호화 할 수 있다고 예상한다 (Golds t e i n, 1984). 따라서 앞서 언급한 장소설은 1, OOOHz 이상의 높은 빈도수의 음의 지각에 비교적 잘 적용될 수 있고, 낮은 빈도수의 음의 지각에는 제사 설이 더 잘 적용될 수 있을 것 같다. 다시 말하면, 제사설은 일반적으 로 우위 빈도 (domi na nt freq u e ncy) 가 . 5, OOOHz 이 하인 청 각신호에 대 한 고저지 각을 설명하는 데 유용하며 특히 1, OOOHz 이 하의 빈도수를 갖 는 소리지각을 잘 설명해 준다. 일반적으로 가청범위 내에 있는 음의 고저지각을 설명할 때 1,OOOHz 미만의 저빈도의 음은 제사설로 설명 하고, 5, OOOHz 이 상의 고빈도의 음은 장소설로 설명 하며, 1, 000-5, OOOHz 의 음은 이 두 이론을 혼합시켜 설명한다. 비록 이론상으로는 그럴 듯 해 보이지만 이 이론으로 음의 고저지각에 관한 모든 현상을 다 설명 할 수는 없기 때문에 아칙도 많은 보완이 이루어져야 한다 .
1.7 소리의 중추통로 와우신경핵은 배측 부분과 복측 부분으로 나누어지며, 와우신경핵으 로부터 뇌 의 청 각 부위로 진행 되는 상행투사에 는 대측 (con t rala t eral) 및 동측(ip s il a t eral) 연결의 두 가지 방식이 있는데, 인간의 경우 대측 연결이 기능상 보다 우세하다는 점이 밝혀져 있다(자세한 것은 제 13 장 에서 논의). 청각기구의 상행성 연결에는 소뇌, 하구체, 뇌교와 연수 의 여러 부분, 시상의 내측술상체, 피질두사 영역의 여러 부위들이 포 함된다. 청각피질 영역은 주로 측두영의 Hesch! 회 (Hesch fs gyru s: Brodmann 41 번, 42 번에 해당)에 위치한다 . 시각 계통과 유사하게 청각 계통에도 감각표면 (기저막)에 대한 중다 표상이 신피질 수준에서 나타난다. 이것은 주로 청신경의 축색측지가 고위청각 영역으로 상행하면서 더 많은 신경원으로 갈라지기 때문이 다. 예컨대 원숭이의 청각 계통에는 30,000 여 개의 청신경원이 있고, 와우신경핵에는 약 190, 000 개 정도의 신경원이 있으며, 시상의 내축슬 상체에는 400, 000 여 개의 신경원이 있다. 그리고 피질 수준에서는 최 소한 1 천만 개 이상의 신경원이 있는 것으로 추측된다 (Green, 1976). 1.8 귀머거리 완전한 귀머거리는 희귀하다. 귀머거리의 99% 는 소리를 크게 지르 면 최소한 어떤 가락을 들을 수 있다고 한다. 청각장애에는 신경농과 전도농의 두 종류가 있다. 신경농 (nerve dea fn ess) 은 와우각, 유모세포 또는 청신경의 장애에 기인된다. 이 장애자는 소리의 고저 특히 높은 주파수의 소리를 듣는 데 어려움을 보인다. 이 장애는 청각보조장치의 도움으로 상실된 청각 울 다소 보상받을 수 있다. 신경농은 유전적일 수도 있고, 다음과 갇 이 임신중의 장애에 의해서도 생길 수 있다. 예컨대 어머니가 임신중 에 매독, 풍전, 기타 전영성 질환에 걸렸을 경우나 임신중 여러 가지
독물에 중독되었다든가, 갑상선 분비활동에 장애를 보였을 경우 등이 다. 성인의 경우는 높은 소리에 장기간 노출되면 청각장애가 생길 수 있다. 전도농 (conduc ti ve dea fn ess) 은 중뇌의 ' 청소골이 고장나 와우각에 음 파를 전달하지 못해 생기는 청각장애이다. 이 장애는 중뇌에 어떤 감 염성 질병이나 종양발생으로 인해 생긴다. 전도농은 일시적인 경우도 있으며 계속 나타나면 의과수술에 의해 교정될 수 있다. 전도농 환자 들은 정상적인 와우각과 청신경을 가지고 있기 때문에 중뇌를 우회하 는 소리는 둘을 수 있다. 예컨대, 이 환자들은 두개골의 뼈를 통해 와 우각에 전달된 자신의 목소리를 들을 수 있다. 2 평형감각 출렁거리는 보트 위에 서서 날아가는 새의 모습을 사전 찍는다고 상 상해 보라. 이런 일울 하려고 할 때는 전정계통 (ves ti bular s y s t em) 이란 감각기구가 작용한다. 평형계통은 지구중력에 대해 신체를 직립시켜 주고, 눈의 운동을 도와주는 감각을 일컫는다. 2.1 평형기관 앞서 본 와우각 의에도 내이에 붙어 있는 섬세한 감각기관인 삼반고 리관(th ree semi circ ular canals) 을 비롯하여 이와 인접하고 있는 난형낭 (utr ic l e ) 과 구형 낭 (saccule) 이 라 부르는 감각기 관들이 평 형 기 관을 이루 는 주요 기구들이다. 삼반고리관은 특히 머리의 움직임을 감지하는 데 중요한 기능을 담당한다. 삼반고리관은 난형낭 및 구형낭이 협동하여 끊임없이 머리의 위치에 관한 정보를 신경 계동에 제공해 준다. 개개의 반고리관은 각기 상이한 평면을 이루고 있어서 머리가 어느 한쪽 방향으로 기울게 되면 이 관 속에 있는 감각기구가 영향을 받게
된다. 이들 반고리관 속에는 팽대부 (am p ulla) 라 부르는 불룩한 영역이 있는데, 이 팽대부 안에는 롱 (c ri s t a) 이라고 하는 관의 형태를 갖춘 감 각기구가 있고, 여기에 유모세포가 밀집되어 있다. 이 유모세포의 섬 모들은 삼반고리관의 천장을 향하고 있으며, 팽대정 (cu p ula) 이라 부르 는 젤라틴성 물질로 둘러싸여 있다. 전정 계통에 있는 유모세포의 특이한 접은 머리가 움직이는 방향에 따라 수용기 전압이 탈분국화 또는 과분국화된다는 것이다. 개개의 유 모세포 중 보다 크기가 큰 섭모를 운동섬모 (k i noc ili um) 라 부르며, 보 다 작은 섭모를 부동섭모 (s t ereoc il a) 라 부른다. 만약 머리가 한쪽 방향 으로 움직여서 부동섭모가 운동섬모 쪽으로 정확하게 기울어지면 유모 세포의 수용기 전압은 탈분국 상태에 이른다. 이렇게 되면 신경전달물 질이 방출되고, 이와 관계 있는 전정신경계의 신경원들의 흥분 발사율 이 증가된다. 이와는 달리 만약 머리운동에 따라 부동섬모의 기우는 방향이 운동섬모를 비켜나게 되면, 수용기 전압은 과분국 상태에 이르 고, 그 결과 신경전달물질의 방출 바울이 정상 이하가 되며 전정신경 계 신경원의 발사율도 저하된다. 전정신경에는 전정기관의 유모세포로부터 신호를 수용하는 약 20,000 개의 신경원이 있다. 이 신경원들의 축색은 제 VIU 신경에 포함되며, 뇌 간에 분포된 여러 전정신경원들에서 시냅스를 이룬다. 전정핵 (ves- tibu lar nucle i)에서 나온 신경원들은 하행성으로 척수와 연결되기도 하 고 상행성으로 전뇌와 연결되기도 한다. 3 미각 모든 종의 동물들은 그들의 환경 속에 산재하는 화학적 물질에 반응 한다. 포유류에서 미각수용기는 혀에 집중적으로 분포되어 있다. 미각 수용기는 무리지어 이의 지지세포들과 함께 미뢰(t as t e bud) 속에 위치 하고 있다. 인간의 경우 개개의 미뢰에는 약 50 개 정도의 수용기세포
가 있으며, 이 수용기의 수명은 10-14 일 정도에 불과하다. 죽은 수용 기는 지지세포로부터 분화되어 나온 새로운 수용기세포에 의해 계속 대치된다. 나이 든 사람들은 젊은이에 비해 이러한 수용기 대치가 매 우 느리며 미뢰의 수도 적다. 이런 이유로 나이 든 사람은 미각이 둔 감하다. 타액에 의해 용해된 물질이 미각수용기를 자극한다. 일반적으로 미 각은 후각에 의해서도 많은 영향을 받는다. 비록 미각이 미각수용기의 막에 음식물 분자가 작용함으로써 일어나는 것으로 알려져 있지만, 맛 의 감각작용이 이루어지는 전환과정에 관해서는 아직 잘 모른다. 모든 미각은 흔히 기본 미각이라 부르는 단맛, 짠맛, 신맛 및 쓴맛 이라는 4 가지 감각의 혼합으로 간주된다. 그러나 개개의 기본 미각에 해당하는 4 종류의 기본 수용기가 존재하지 않는다는 것이 확실해졌다. 미각에 관한 연구에서, 신맛은 수소 이온 (H+ )의 출현과 관계 있고, 짠맛은 Na+, K+ 이온에 관련되어 있는 것으로 간주된다. 키니네 (q u i n i ne) 는 쓴맛을 야기 하는 강력 한 자극이며 , 단맛은 다양한 종류의 자당 (sucrose) 과 그 밖의 여러 물질들에 의해 야기된다. 그러나 이러한 물질둘을 사용하여 특정 미뢰를 선택적으로 자국시키고, 미각동로에서 개개 신경원의 반응을 측정해 보아도 특정 자극 물질에만 반응하는 특 정 미각수용기를 발견할 수는 없다. 예컨대 Carl P faff man(1959) 은 미각수용기와 연결되어 있는 고삭신 경 (chorda ty m pan i)에 있는 대부분의 신경원이 실험에 사용된 모든 자 극 (H+ 이온을 포함한 몸은 염산, NaCl, 키니네 그리고 자당)에 대해 반응 한다는 사실을 관찰하였다. 약간의 세포들은 P faff man 이 〈최상자국 (best s tim u li)〉이라 언급한 특정 자국에 보다 특징적으로 반응하였지만 단지 하나의 미각자국에만 반응하는 신경원은 발견되지 않았다. 고삭신경에 있는 개개의 신경원들은 적어도 하나 이상의 미각수용기 와 시냅스하며, 개개의 수용기 또한 하나 이상의 신경원과 시냅스를 이룬다. 고삭신경은 제 VII 뇌신경인 안면신경과 합쳐져 뇌간의 고속신경 핵으로 두사되며, 이곳에서 나온 2 차 신경원이 뇌의 여러 부위로 두사
된다. 예컨대 원숭이의 경우 고속핵에서 나온 일부 축색은 시상의 복 후내 측핵 (ventr a l pos tr e io r media l nucleus) 으로 진 행 되 며 , 이 핵 에 서 나 온 신경원은 전두영의 후측 부위에 있는 도( 島 , i nsula) 라는 영역과 두 정엽의 체성감각피질의 〈 안면(fa ce) 〉 영역으로 축색을 보낸다. 한편 고속의 신경핵에서 나온 다른 축색은 피질하 영역인 의측 시상하부, 편도핵 그리고 뇌로 두사된다. 4 후각 미 각과 마찬가지로 냄 새 감각, 즉 후각 (ol fa c ti on) 도 후각수용기 신경 원의 막에 분포된 수용기와 물질분자 사이의 상호작용에 의해 이루어 진다. 후각수용기는 축색을 갖고 있는 완전한 형태의 신경원이다. 인 간의 경 우 후각수용기 는 후각상피 (olfa c to ry epi the liw n ) 이 며 비 강 내 에 위치한다. 개처럼 후각기능이 고도로 발달된 동물에서는 후각상피 조 직이 상대적으로 크다. 후각상피는 몇 종류의 세포들로 구성되는데 수 용기세포는 물론 지지세포도 포함하고 있고, 이 지지세포는 수용기세 포가 죽게 되면 새로운 수용기세포로 분화된다• 후각 수용기의 평균수 명은 약 35 일에 불과하며 죽은 수용기세포를 대치하기 위한 새로운 세 포가 계속 만들어진다. 포유류의 경우 출생 후 새로운 신경세포가 만 들어지지 않는 것이 일반적 원칙이지만 후각세포만은 예의이다. 후각수용기에는 후각상피를 덮고 있는 점액 (mucus) 쪽으로 뻗어가는 6-8 개의 섬모가 있다. 점액은 지지세포로부터 분비되며, 공기에 포함 된 물질들을 용해시켜 수용기와 직접 접촉할 수 있도록 해준다. 인간 은 대략 1 억 2 천만 개의 후각수용기 세포를 갖는 데 비해 독일산 셰퍼 드는 20 억 개 의 수용기 를 갖는다 (Mozel, 1987) . 후각자극에 의 해 발생 되는 전도과정은 아직 잘 알려져 있지 않지만 점액에 용해된 물질들과 후각수용기의 섬모막 사이의 상호작용에 의해 이온통로의 성질이 변경 된 결과 활동전압이 야기될 것으로 추측된다.
척추동물의 경우 특정한 냄새에 대한 특정 수용기가 별도로 존재하 는 것 갇지 는 않다. Gordon Sheph erd (19 83) 에 의 하면 도롱뇽의 후각 계통에는 비특정의 일반 수용기밖에 없으며, 이 수용기는 10 여 가지의 상이한 냄새에 모두 반응한다고 한다. 그러나 이런 상이한 냄새는 후 구 (ol fa ctry bulb) 의 여러 장소에서 개별 신경활동을 야기한다고 한다. 이것은 곧 냄새의 종류에 따라 신경활동의 공간분포가 달라진다는 후 각의 〈장소설(p la te th eo ry)〉울 가정하게 하는 근거가 된다. 만약 상이 한 냄새가 후구의 상이한 신경원을 흥분시키고 이 신경원들이 개별적 통로를 따라 고위 뇌중추에 이른다면, 고위 중추에서도 상이한 장소에 서 이들 흥분들이 부호화될 것으로 추측할 수 있다. 후각수용기의 축색은 뇌의 바닥에 있는 후구 속으로 진행된다. 인간 의 경우 후구는 전두영 밑에 위치한다. 후구에 있는 수용기 세포의 축 색은 많은 세포가 밀집되어 작은 공처럼 생긴 사구체(g lomerulus) 안으 로 들어가 그곳에서 사구체 세포들과 매우 복잡한 방식의 시냅스 접촉 울 한다. 후구에 있는 세포의 주된 형태는 승모세포 (m it ral cell) 와 소 방세포(t u ft ed cell) 이다. 유인원의 경우 이 두 유형의 세포축색은 후구 의 출력 부를 이 루어 신 경 충격 을 의 측후삭로 (lat e r al olfa c to r y tra ct) 를 거 쳐 전이 상피 질 (pre pr i fo rm co rt ex) 로 보낸다. 전이 상피 질로부터 다시 의 측 시 상하부 (lat e r al hyp o th a lamus) 와 시 상의 배 내측핵 (dorsal media l nucleus) 으로 두사된다. 이 영역에서 나온 흥분은 전두영의 안와전두피 질 (orbit of r on ta l co rt ex) 로 전달된다. 이와 달리 쥐와 같은 동물은 후구 로부터 시 상하부와 편도핵 의 피 질내 핵 , 중격 의 대 각대 (dia g o n al band) 및 전천공질 (an t er i or per fo r ate d subs tn ce) 과도 신경연결을 이룬다. 상이한 냄새를 구분하는 능력은 나이가 들어감에 따라 약화된다. 2 천 여 명의 피험자를 대상으로 한 연구에서 65-80 세 사이의 피험자들 가 운데 적어도 반 이상이 후각의 지각검사에서 심한 손상을 보였으며, 80 세 이상 노인 가운데 75% 이상이 후각 손상을 보였다 (Do ty 등, 1984). 무척추동물이나 척추동물 모두 어떤 특수한 분비선으로부터 신체 바 깥으로 신호물질을 분비한다. 이들 동물들은 이 물질을 공중에 직접
방출하는 수도 있고, 환경 속의 어떤 대상물체에 묻혀두는 수도 있다. 이러한 물질을 페로몬(p heromone) 이라 하는데 일반적으로 이 물질들은 휘발성 물질이어서 바람을 타고 먼 곳까지 운반될 수 있다. 페로몬은 특히 생식 상태나 세력권과 관련되는 신호물질이라는 점이 밝혀졌다. 유안원도 페로몬을 분비하는지는 아직 명확하지 않다. 어떤 연구자 둘은 레서스 원숭이 암컷이 성적으로 유혹적인 페로몬을 분비한다는 증거 를 제 시 하였지 만 (Mi ch ael & Keveme, 1970) , 이 와는 달리 부정 적 결과를 얻은 경우도 있다 (Gold fo o t 등, 1976). 유인원의 성행동은 대단 히 복잡하며 연구대상이 되는 동물에 따라 성행동이 판이하게 나타난 다. 따라서 페로몬이 유인원의 사회적 상호작용에 미치는 영향에 대한 확실한 증거는 아직 없다. 인간의 경우에도 페로몬을 분비한다는 몇몇 보고가 있지만 (Ro g el, 1978), 결정적인 증거는 아직 없다. 개, 소, 쥐 또는 코끼리와 같은 페르몬을 방출하는 동물들은 서골비 강기 관 (vomeronasal orga n ) 이 라 부르는 제 2 의 후각기 관을 갖는다. 이 기관은 입과 코가 맞닿는 곳에 위치하고 있다. 서골비강기관의 수용기 는 앞서 본 후각상피에 있는 수용기세포와 유사하게 후구로 축색을 보 내고, 후구에서 나온 이차신경원 축색은 편도체와 시상하부로 투사된다. 5 체성감각 피부는 적 어도 두 가지 성 분, 죽 외 피 (ep ide rmi s) 와 내 피 (derm i s) 로 구성된다. 또한 피부는 털이 있는 유모 부위 (hau ry)와 털이 없는 무모 부위(g larbrous) 로 나누어진다. 피부감각수용기는 형태가 매우 다양하 지만, 일반적으로 압각에 민감한 기계적 수용기 (mechanorece pt or) 와 온도감각에 민감한 온도수용기 (th ennorece pt or) 로 나눈다. 이 러 한 피 부 감각수용기는 다른 종류의 수용기와 더불어 내장, 근육, 건 그리고 신 체의 관절 부위에 주로 분포되어 있는데, 이를 통틀어 체성감각 계통 (somest he tic sys t e m ) 이 라 부른다.
피부수용기는 피막에 쌓여 있는 것도 있고 그렇지 않은 것도 있다. 피막이 없는 비피막수용기를 일반적으로 유리신경말단(fr eenerve endin g ) 이 라 부르며 , 이 수용기 가 담당하는 피부 부위 에 수지 상돌기 가 분포되어 있다. 피막수용기는 세포의 수용 부위 주변에 특수 형태의 모자와 같은 모습을 갖추고 있어서, 이것이 전환과정을 용이하게 해준 다. 체성감각 계통의 수용기는 감각양상(예컨대 촉각이나 온각의 민감 도)의 속성에 따라 민감도, 신경충격의 전달속도 그리고 어떤 하나의 불변자극에 대한 적응률에서 각기 상이하다. 5.1 유리신경말단 유리신경말단은 피부, 근육 그리고 장기를 비롯한 모든 신체 부위에 서 발견된다. 어떤 것은 촉각에 반응하고 또 다론 것은 온도에, 또 어 떤 것은 유해로운 자극에 반응하며 동시에 위의 두 가지 또는 세 가지 자극 모두에 반응하는 수용기도 있다. 이런 이유로 유리신경말달은 중 다감각수용기 (po lys e nsory recep tor ) 라고 부르기도 한다. 대개 의 경우 유리신경말단은 적응속도가 느리며, 지름이 매우 가늘고, 수초가 엷게 덮인 신경원으로 신경전달속도도 비교적 느리다. 그러나 어떤 것은 수 초가 두껍게 덮여 있고 큰 축색을 갖고 있어 신경충격의 전도속도가 매우 빠른 것도 있는데, 이를 A 델타 (A-de lt a) 섭유라고 하며 30m/ 초 이상의 속도로 신경충격을 전달한다. 이와는 달리 지름이 작고 수초화 가 이루어지지 않은 축색을 C 섬유라고 하는데, 이들은 2m/ 초 이하의 느린 속도로 충격을 전달한다. 이러한 두 종류의 수용기는 손가락이 바늘에 찔렸을 때 경험하는 두 종류의 상이한 통각 경험과 관련 있는 것으로 생각된다. 죽, 이 경우 에 즉각적으로 느끼는 급속 통증(fa s t p a i n) 은 A 델타 섬유에 의해 매개 되는 것이고, 반면 시간이 좀 지난 후 서서히 타는 듯이 아픈 통증은 C 섬유에 의해 매개되는 것이다. 감각수용기에서 감각과 신경활동 사이의 상관은 인간을 대상으로 직
접 연구할 수 있다. 즉, 깨어 있는 피험자의 피부 속에 기록전극을 삽 입한 후 여러 가지 자국을 피부에 가하여 이때 일어나는 감각을 보고 하도록 한다. 유리신경말단은 모든 종류의 피부감각을 전달할 수 있다는 사실이 증명되었다. 예컨대 유리신경말단만 가지고 있는 안구의 각막을 자극 하여 온각, 냉각, 촉각 및 통각과 같은 여러 가지 감각을 야기시킬 수 있다 (Ganon g, 1977). 그러므로 유리신경말단은 가벼운 기계적 변형 (촉 각), 온도 변화 또는 손상에 대해 각기 특이하게 반응할 수 있다. 5.2 피막수용기 여러 종류의 피막수용기 가운데 대표적인 것은 파치니 소체(p a ci n i an co rp uscle) 인데, 이것은 피부의 유모 부위와 무모 부위, 근육 또는 장 기의 결합조직에서 발견된다. 파치니 소체를 둘러싸고 있는 여러 겹의 피막을 조심스럽게 벗겨내면 이 소체의 돌기에서 소체의 활동을 기록 할 수 있다. 만약 이 돌기에 충분할 정도의 압력을 가하면 탈분극이 일어나고, 수초화된 돌기의 첫번째 마디에서 신경충격이 발생된다. 이 충격은 이어 이 소체획 두번째 돌기를 따라 척수로 들어간다. 모든 피 부수용기들은 양극성 세포로서 한쪽 돌기는 수용종말인 피부 쪽으로 뻗어 있고, 다론 한쪽 돌기는 척수로 투사한다. 파치니 소체는 진동감각과 같이 반복되는 자극에 대해 잘 반응한다. Mi ch ael Merzen i ch 와 Thomas Harr i n gton(1969) 은 원숭이와 인간의 유모 부위에 진동자극을 가하여 두 종류의 수용기가 진동감각에 관여 한다는 점을 발견하였다. 죽, 저빈도의 진동감각은 유리신경단이나 모 낭 (ha i r follicle s) 의 활동에 의 해 수용되며 , 고빈도의 진동은 파치 니 소 체에 의해 수용된다. 파치니 소체는 저빈도의 진동에 대해서는 반응을 하지 않지만 초당 200 회 이상의 고빈도 전동에 대해서는 매우 잘 반응 한다. 고빈도의 진동에 잘 반응할 수 있는 것은 주로 이 소체를 둘러 싸는 지지조칙의 피막 특성 때문이다. 죽 이 다층 피막은, 지속되는
압력에 의해 일어나는 서행성 변화는 제거해 버리고, 대신 압력자극의 시작과 종결시 일어나는 급속변화에는 반응을 보이도록 한다. 5.3 운동감각수용기 뼈를 둘러싸고 있는 조직이나 뼈와 관련 있는 조직 그리고 신체의 관절 부위에는 관철의 위치와 관련되는 감각정보를 중추신경계로 두사 시키는 유리신경말단이나 그 밖에 다른 종류의 수용기가 있다. 바로 이 입력정보가 근육의 수축 상태와 건의 긴장 상태에 관한 정보들과 더불어 운동을 통제하고 자세를 조절하는 데 사용된다. 5.4 체성감각통로 피부감각수용기의 돌기들은 합쳐져서 후근을 통해 척수로 들어간다. 그러나 일단 척수 안으로 들어오게 되면 체성감각 계통은 전의측체계 (ante r olate r al sys t e m ) 와 배 주-내 융대 체 계 (dorsal column-media l lemnis c al s y s t em) 로 나누어진다. 이 두 체계에 있는 신경원들은 척수로 들어온 일차 구심성 흥분을 받아들이는 시냅스후 신경원들이다. 전의측체계는 이미 오래전에 절멸해 버린 고생동물들에서 찰 발달되 어 있는 것으로 보아 계통발생적으로 매우 일찍 발달된 것으로 간주된 다. 이 체계는 지름이 매우 가늘고, 엷은 수초와 흥분 전달이 느린 축 색을 갖는 신경원으로 이루어진다. 이 신경원들은 중추로 향해 상행하 는 동안 여러 차례 시냅스를 이룬다. 전의측체계는 주로 열기, 냉기, 통증 등의 감각을 전도한다. 이 체계의 명칭은 수용기의 흥분이 척수 에 진입하자마자 척수신경원과 시냅스를 이루고, 시냅스후 척수신경원 이 척수의 전의측 부위에서 뇌로 축색을 보내기 때문에 붙여진 것이 다. 이 체계의 축색은 척수의 중앙선을 넘어 반대쪽 부위에서 상행한 다. 이 체계 가운데 일부는 뇌간의 망상체에서 여러 차례 시냅스를 이 루며 다른 일부는 시상의 내층핵 (intral ami na r nucle i)에서 시냅스한다.
또한 전의측체계의 일부 축색들은 척수의 한 영역인 교상질 (subs t an ti a g ela ti nosa) 에서 종말을 이루기도 하는데, 이 교상질이 통각전도의 신 경기제가 된다는 증거가 많이 제시되어 있다. 배주-내융대체계를 이루는 축색들은 계통발생학적으로 보아 최근에 만들어진 것으로 간주된다. 이 체계의 축색은 전의측체계의 축색에 비 해 지름이 굵고 두꺼우며, 수초화되어 있고, 신경전도도 더 빠르다. 이 축색은 초당 70m 의 속도로 충격을 전달한다. 이 체계에 속하는 신 경원은 척수의 배측 부위를 통해 뇌로 흥분을 투사하며 연수 부위에서 시냅스한다. 연수에서 시냅스 후 출발하는 2 차 신경원들이 내융대 (media l lemn i scus) 를 이룬다. 내융대 신경원들은 정중선을 건너 시상 의 복후의 측핵 (ventr a l pos te r io r late r al nuclei) 과 의 측신 경 핵 으로 투사된 다. 이 부위들이 체성감각 전도에서 가장 중요한 시상 부위들이다. 이 시상 부위의 신경핵둘은 두정영의 체성감각피질로 두사된다. 이처럼 체성감각 계통의 구심성 축색이 정중선을 넘어 반대편의 반구로 두사 되기 때문에 신체 표면의 표상은 대측이 된다. 5.5 체성감각피질 시각과 청각의 경우에서처럼 체성감각피질의 감각표면도 매우 복잡 한 지도를 보여준다. 체성감각의 일차수용 영역은 두정엽의 앞 부위, 죽 Broadman 1, 2 및 3 번 영역에 해당된다. 3 번 영역은 다시 3a 와 3b 로 나누어지는데, 이들 영역은 여러 종류의 유인원에 있어서 신체표면 울 거의 완벽하게 표상한다. 이 네 가지 영역을 이루는 개개 영역은 신체표면의 감각을 단순하게 표상하는 것뿐만이 아니라 내적 조정, 신 경원의 반웅 특성 그리고 다른 뇌 부위들과의 시냅스 연결 등에서 차 이를 보여준다. 체성감각 계통이 보여주는 조정원리 가운데 하나는 감각수용기를 많 이 가지고 있는 신체표면일수록 대뇌피질에서도 보다 넓은 영역을 차 지한다는 점이다. 즉 입술, 혀, 안면 그리고 손가락 끝과 같은 신체
부위는 많은 감각수용기를 가지기 때문에 이들 신체 부위를 표상하는 뇌피질 면적도 넓다. 반면 몸통과 같이 비교적 둔감한 영역들은 피질 의 표상영역도 작다. 최근 체성감각 계통의 피질투사는 피부에 있는 수용기에서 변화가 일어나면 국적으로 바뀌어질 수 있다는 사실이 보고되었다. Mi ch ael Merzen ic h 와 그의 동료들 (1984) 에 의하면, 원숭이의 앞다리의 발가락 하나를 절단하면 이 절단된 발가락으로부터 오는 정보를 처리하는 데 관여했던 피질영역이 사라진다. 그 대신 매우 작은 영역의 신경원들이 없어전 발가락 쪽의 정보를 처리해 주거나 나머지 발가락의 정보를 처 리하는 피질영역의 활동이 더욱 확장된다고 한다. 이 사실은 뇌피질에 서 시냅스의 변경이 일어날 수 있다는 것과 피질의 신경연락이 감각입 력의 변화에 순응하기 위해 바뀔 수 있다는 점을 시사하는 것이다. 피질신경원들의 반응을 탐색하는 과정에서 피질이 기둥 모양의 주 (columns) 로 구성되어 있다는 하나의 해부학적 원리가 발견되었다. 체 성감각피질의 기둥 조직은 1957 년 Moun t cas t le 에 의해 처음 발견되었 으며, 1959 년 Powell 과 Moun t cas tl e 에 의해 재확인되었다. 이들은 기 록전극을 피질 표면에 수칙으로 삽입하여 신경원들의 반웅 특성을 알 아본 결과 동일 수직선 상에 있는 여러 신경원들이 매우 유사한 기능 적 특성을 보여준다는 사실을 발견하였다. 예컨대 동일한 피질원주를 구성하는 거의 모든 신경원들이 가벼운 촉각, 심한 압력 또는 관절의 회전 등에 대해 유사한 반응을 보여준다. 그러나 전국을 직각이 아닌 다른 각도로 비스듬히 삽입하면 반응성이 상이한 여러 유형의 세포가 발견된다. 6 통각 대부분의 사람은 통증을 느낀다. 그러나 가끔은 통증을 전혀 느끼지 못하는 사람들도 있는데, 이들은 심한 상처를 입어도 아픔을 느끼지
못하므로 정 상적 인 생 명 유지 가 불가능하다. 따라서 정 상적 인 통각은 생명 유지를 위한 경고 기능을 제공한다. 그 밖에도 통각은 주의를 끌 게 하고 놀라게 하고 도움을 줄 사람을 찾게 하고 고통스런 자극을 회 피할 수 있도록 동기화시켜 주기도 한다. 아직 통각에 관한 연구는 부 족하지만 최근 몇 가지 홍미로운 사실들이 발견되었다. 특히 최근 발 달한 심 리 학의 한 분야인 행동의 학 (behav i oral medic ine ) 에 서 통각울 중 요한 분야로 다루고 있다. 6.1 통각의 동로 통각의 신경통로에 관해서는 많은 연구가 이루어졌다. 통각을 전달 하는 말초신경은 A 델타 섬유 또는 C 섬유 중 어느 하나에 속한다. A 델타 섬유를 보통 대섬유(l ar g e fibe r: L fi ber) 라 하며 C 섬유를 소섬유 (small fibe r : S fibe r) 라 한다. L 섬 유는 정교하게 수초화되 어 있고 S 섬 유는 수초화되어 있지 않다. 이 두 종류의 신경섬유는 바늘이 피부 깊 숙이 들어갈 때 느껴지는 두 종류의 통각, 즉 먼저 참깐 동안 따끔하 게 느껴지는 통각과 잇따라 타는 듯한 심한 통각에 관여한다. 즉 처음 의 따끔한 통각은 첫 자극에 의해 유발되는 순간적인 신경충격을 초당 15m 정도의 빠른 속도로 이동시키는 L 섬유에 의하고, 뒤이어 지속되 는 타는 듯한 통각은 계속적인 자극에 의해 이어지는 지속적인 신경충 격을 초당 lm 정도의 느린 속도로 전달하는 C 섬유에 의한다. L 섬유 나 S 섭유는 통각 이의에 촉각이나 온도 감각과 같은 정보도 전달한다. 통각을 담당하는 고등 뇌중추 부위에 관해서는 아직 잘 모르지만, 뇌 간의 망상체와 이와 관련된 내측 시상핵 (media l tha lamic nuclei) 이 중 요한 곳으로 간주된다 (Case y, 1982). 사실 뇌간과 시상의 중앙축 주변 에 있는 신경핵둘이 통각이나 통각의 억압에 관여한다는 실험결과들이 많이 제시되고 있지만 통각에 대뇌피질이 관여한다는 증거는 많지 않 다. 변 연계 의 한 구조인 대 상피 질 (cin g u la te cort ex ) 이 통각 흥분을 수 용하여 통각의 정서적 측면을 매개한다고 하는데, 특히 이 피질에 엔
도르핀의 수용기가 밀집해 있다는 점은 주목할 만하다 (K i mble, 1992). 6.2 내인성 아편 : 동각제어 물질 심한 스트레스 상황에 처할 때 통각이 전적으로 사라지는 무통 상태 가 나타나는데, 이를 스트레스성 무통 (s t ress- i nduced analge s ia : SIA) 이라 칭한다. 이런 무통은 모르핀이나 헤로인과 같은 아편성 물질과 유사한 효과를 갖는 신체 내의 어떤 펩티드성 물질의 방출에 기인되는 것이다. 이런 신체에서 발생하는 내인성 (endog en eous) 모르핀을 엔도 르핀 (endor ph i nes) 이라 한다. 엔도르핀은 뇌뿐만 아니라 기타 신체 내 부에서 여러 가지 기능을 한다. 이 물질의 기능에 대해서는 많은 연구 가 이루어지고 있다. Freder ic kson 과 Gea ry (l982) 에 의하면 1976 년에 서 1981 년 사이 의 5 년 동안 엔도르핀과 관련 있는 논문이 무려 7, 500 편이나 발표되었다고 한다. 우리는 여기서 엔도르핀 연구의 최근 추세 룰 몇 가지 측면에서 살펴볼 것이다. 1970 년대 초반 몇몇 실험실에서 실험동물의 뇌 속에 아편성 물질에 대한 특수 수용기가 존재한다는 사실을 발견하게 되었다 (Pe rt& Sny de r, 1973). 이런 발견으로 아편과 흡사한 몇 가지 자생물질들이 뇌 속에 존재할 것이며, 이 아편 물질에 대한 특수 수용기가 뇌 속에서 전화되어 왔을 것이라고 믿게 되었다. 그 후 1973 년 가울 스코틀랜드 에 있는 애버딘 대학 연구진들이 뇌 속에서 아편물질 하나를 발견하였 고, 그 후 2 년 사이 에 또 다른 물질을 하나 더 발견하였다 (Hu g hes, 1975; Hug he s 등, 1975). 연구자들은 이 물질들을 뇌 속의 물질이란 뜻 으로 엔케 팔린 (enkeph a lin ) 이 라 칭 하였다. 이 두 가지 뇌 속 물질은 모 두 5 개의 아미노산으로 이루어진 펩티드인데, 5 개의 아미노산 가운데 하나가 서로 다르기 때문에 그 중 하나를 류-엔케팔린 (leu-en· ke p ha li n) 이라 부르고, 다른 하나를 메트-엔케팔린 (me t -enke p hal in)이 라 불렀다. 이 두 물질은 최초로 분리된 내인성 아편물질이었지만, 그 후 몇 년 사이에 수많은 연구가 이루어져 위의 두 물질 이의에도 몇
가지 내인성 아편물질이 새로 발견되었다. 이런 물질들은 뇌하수체나 뇌 속에서 발견되었으며, 심지어는 부신이나 내장기에서도 발견되었 다. 우리 몸에서 생성되는 아편성 펩티드는 다음과 같은 3 가지 기본 화 학 계열 중 어느 하나에 속한다. 이런 상이한 엔도르핀은 거대한 전구 물질 분자의 분열에 의해서 생성되는 것으로 생각된다 (Ak il 등, 1984 ; Khachatu r ia n 등, 1985) . 이 세 가지 의 엔도르핀 계 열은 가장 큰 전구 물질 분자로부터 가장 작은 물질 분자로 분열되는 방식에 따라 명칭이 붙여진다. 첫째가 엔도르핀, 둘째, 엔케팔린 그리고 셋째가 디노르핀 (d yn o rp h i n) 이다. 날록손 (naloxone) 은 모르핀의 주된 길항물질로서 뇌 의 엔도르핀 수용기와 결합하여 아편을 대치한다. 따라서 아편중독자 에게 날록손을 주사하면 회복이 가능하다. 6.3 엔도르핀 분비와 동각억제의 증거 엔도르핀이 통각을 억제한다는 증거는 이것을 환자에게 직접 두여하 여 통각이 경감되는지의 여부를 관찰해 보면 알 수 있다. 그러나 이것 은 두 가지 기술적 문제를 가지고 있어서 실험하는 데 어려움이 있다. 엔케팔린의 효력은 효소작용에 의해 급격히 떨어지기 때문에 효과적으 로 투여할 수 없으며, 베타-엔도르핀은 혈액 내에서 오랫동안 작용하 지만 혈-~장벽을 통과할 수 없기 때문에 임상적 적용에 어려움이 많 다. 그래서 최근에는 이들 물질과 유사한 화학물질을 개발하여 실험적 으로 적용한 결과 이들 유사 물질이 통각제어에 효과가 있다는 점이 보고되 었다 (Frederick son & Geary , 1982) . 칙접적인 증거가 부족하지만 많은 간접적인 증거에 의해 엔도르핀이 통증 억제에 중요한 역할을 한다는 견해가 제시되고 있다. 통각 억제 를 알아보기 위해 두 가지 중요한 기법들이 사용되고 있는데 그 중 하 나는 자국생 성 무통 (stim u lati on -pr o duced-analge s ia : SPA) 이 고, 다른 하나는 스트레 스 유발 무동 (s t ress- i nduced analge s ia : SIA) 이 다.
SPA 란 마취하지 않은 동물이나 사람의 특정 뇌 부위에 전기자국을 가하여 통각을 억 제 시 키 는 것을 말한다. 이 방법 은 1969 년 Reyn o lds 에 의해 처음 보고되었고 그 후 많은 연구자들에 의해 널리 적용되었 다 (Ma y er 등, 1971). SPA 를 생성할 수 있는 뇌 부위는 뇌의 중심관 및 뇌실 주변의 중뇌수도주위 회백질(p e ri a q ueduc t al gray ma tt er) 과 이 와 인접한 뇌간의 일부 영역이다. 뇌간이나 간뇌의 여러 신경핵과 해 부학적으로 연결되어 있는 고속핵을 자극하면 통각억제 효과가 나타나 며, 이 효과는 고속핵 자극에 의해 중뇌수도주위 회백질이나 거대 봉 선핵을 활성 화시 킴으로써 나타난다고 한다(손진훈과 오애령, 1992). 이 들 뇌 영역을 자극하여 생성되는 통각제어의 측정방법은 뜨거운 철판 위에 울려놓은 동물이 앞발을 들어울리는 반응이나 꼬리치기 반응을 통해 측정할 수 있다. 무통의 정도는 마취를 하지 않고도 실험동물을 복강수술할 수 있을 만큼 충분하며, 사람의 경우에는 말기 암환자의 심한 통각을 제어하는 데 이용될 수도 있다고 한다 (Hosobuc hi, Adams, & Lin c hit z, 1977 ; Ric h ardson & Aki!, 1977) . 이 런 암환자들은 자 신의 뇌 부위에 장치된 전국을 통해 스스로 고통을 통제할 수 있다고 한다. SPA 효과가 날록손 투여에 의해 경감될 수 있다고 하는 사실은 뇌의 직접적인 전기자극에 의한 통각 경감이 엔도르핀에 의해 매개된 다는 강력한 증거가 된다 (Aki! 등, 1984). 반면 뇌의 전기자극에 의해 매개되는 통각제어 중 어떤 것은 날록손을 두여해도 경감되지 않는다 고 하는데, 이것은 비아편성 신경회로가 SPA 에 관련되어 있음을 시사 하는 것이다. 엔도르핀의 통각 억제에 관한 두번째 계열의 증거는 SIA 실험에서 제시되었다. 예컨대, 찬물 속에 동물을 빠뜨려 강제적으로 수영을 시 키는 따위의 스트레스를 부과하면 통각 자극에 대한 동물의 반응성을 낮출 수 있다. 쥐의 통각 자극에 의한 반응성을 낮추는 SIA 효과는 모르핀 투여 효과와 유사할 정도로 강력하다. SIA 는 날록손의 효과를 역전시킬 수 있고, 뇌의 엔도르핀 수준을 증가시킬 수도 있다. 인간에 있어서 SIA 의 증거는 반복적인 스트레스(예컨대 전기충격이 제시된다는
경고신호가 반복적으로 제시되는 경우)가 전기충격에 대한 반사의 역치를 유의미하게 높인다는 증거 (W iller , Dehen, & Cambie r , 1981) 에서 찾을 수 있는데, 이러한 SIA 효과는 5m g의 날록손을 투여하면 사라진다. 전쟁, 사고, 운동경기 또는 기타의 긴급상황에서는 심한 상처를 입 어도 통각을 별로 느끼지 못한다는 사례들이 많이 보고되어 있다. 이 런 경우 긴급상태가 지난 후에야 비로소 고통을 느끼기 시작한다. 모 든 SIA 가 날록손에 의해 역전되지 않는다는 사실은 SIA 가 아편체계와 비아편체계 두 가지 과정에 의해 이루어진다는 사실을 의미한다. SIA 의 아편성 체계를 설명할 수 있는 한 가지 특칭적인 현상은 학습된 무 기 력 (lea rned helpl e ssness) 현상이 다. 학습된 무기 력 이 란 도피 불가능한 전기충격에 노출된 경험을 가졌던 동물은 그 후 전기충격을 피할 수 있는 상황이 제시되어도 도피학습을 할 수 없는 것을 말한다 (Ma i er 등, 1983). 앞서 기술한 상황 의에도 무통을 야기하는 상황은 많다. 예컨대 무 아경지, 최면상태, 침술, 성적 흥분 또는 지속적인 운동 도중에도 통 각을 느끼지 못한다. 그 밖에 환자 자신이 어떤 의료처치가 효과가 있 다고 강력하게 믿을 때도 통각이 억압되는데, 이런 효과를 위약효과 (pla cebo e ff ec t)라고 한다. 이런 다양한 종류의 통각억압과 엔도르핀의 작용을 알아보기 위한 연구가 활발히 수행되어 왔다. 이 분야의 연구 가 새롭게 진행됨에 따라 이해의 폭이나 정도가 달라지고 있다. 현재 로서는 침술의 효과나 위약의 효과가 엔도르핀 분비에 의해 매개되는 것으로 간주되고 있지만, 최면성 무통은 이와는 다른 것으로 간주된 다. 그 밖에 무아경지나 성적 흥분 고조시의 무동이 과연 엔도르핀에 기인되는지 여부에 대해서는 아직 어떤 결론도 내릴 수 없다. 위약효과에 관한 연구들 가운데 Lev i ne 과 그의 동료들에 의한 연구 (1979) 가 홍미를 끈다. 이 연구에서는 치아를 뽑은 직후 약리학적으로 는 아무런 작용도 하지 않는 위약을 취한 젊은 피험자들 가운데 약 1/3 정도가 통각이 억제되었다고 보고했다고 한다. 그런데 이들에게 위약 울 줄 때 이 약은 최근에 개발된 강력한 전통제라는 지시를 주었으며,
이런 위약효과는 날록손 두여에 의해 역전되어 통각이 다시 나타났다 고 한다. 한편 위약은 취했지만 유의미할 정도로 동각 억제가 되지 않 았다고 보고했던 피험자는 날록손 두여에 의해 통각이 더 높아지지는 않았다. 물론 이런 유형의 연구는 앞으로 더 많이 이루어져야 하겠지 만 현재로 볼 때 위약효과는 엔도르핀 방출에 따른 결과로 볼 수 있 다. 침 (acu p un t ure) 이 통각을 제어하는 데 효과가 있다는 것은 오래전부 터 알려져 온 일이다. 침에 의한 무동은 날록손 투여에 의해 역전될 수 있다고 한다 (Aki! 등, 1984). 그러나 과연 침이 내인성 아편물질을 직접 방출케 하는지 아니면 단순한 위약효과인지에 대한 의문이 제기 되었다. Pomeranz(1987) 에 의하면 침의 시술은 칙접 내인성 아편을 방출시킨다는 증거가 있으며, 동물을 사용하여 침에 의한 무통을 야기 할 수 있다고 하였다. 이 분야의 연구는 통각제어의 방법으로 침을 시 술하는 동양의학 쪽에서 과학적으로 검토해 볼 만한 연구 대상이라 할 것이다. 6.4 달리기의 즐거움과 엔도르핀 엔도르핀 연구에서 얻어진 초기의 성과들 가운대는 성적 오르가슴, 주자들의 기분 그리고 각종 종교행사에서 볼 수 있는 도취감과 같은 각종 즐거움이 엔도르핀 방출에 기인되는 것이 아닌가 하는 것이었다. 이런 연구들의 주된 관심은 조깅이나 마라돈과 같은 힘든 달리기를 하 면서 느끼게 되는 주자들의 즐거움이 엔도르핀과 어떤 관련성이 있지 않을까 하는 것이었다. J anal 과 그의 동료들에 의한 연구 (1984) 는, 심 한 달리기 끝에 얻어지는 기분 변화와 충동 감소가 엔도르핀 분비에 기인된다는 실제 증거를 제시하였다. 이론적 관점에서 엔도르핀이 성적 흥분 고조와 오르가슴에 관여할 것이란 가설은 많은 홍미를 끈다. 모르핀과 같은 어떤 특정 물질의 신 속한 방출이 순간적인 쾌감을 야기시킨다는 것은 그럴 듯하다. 아편중
독 환자들이 성적 무관심을 보이는 경우가 많다는 것은 잘 알려진 일 이며, 이는 아편을 취함으로써 자연적 엔도르핀 분비가 대치되어 버리 기 때문일 것으로 설명할 수 있다. 갑작스런 엔도르핀 분비는 남성의 경우 오르가슴 뒤에 일시적으로 따르는 불응기 현상과 관련 있다고 하 는 증거들도 단편적으로 제시되고 있다. 일반적으로 날록손은 인간의 성행동에 영향을 미치지 않는 것으로 생각되지만, 흰쥐의 경우는 성적 으로 무기력한 성활동을 증가시킨다는 보고도 있다.
제 7 장 운동 앉은 자리에서 일어나 양팔을 머리 위로 쭉 뻗고 이어 옆으로 뻗은 후 이를 모아 앞으로 뻗어보자. 이런 운동이 쉽게 이루어지는 것 갇지 만, 이처럼 간단한 운동도 신피질, 기저핵, 소뇌, 시상 그리고 척수 등에 있는 수십억 개의 신경원이 작용함으로써 나타나는 것이다. 또한 이 동안에는 심장박동과 혈압이 변화되는 것은 물론 수많은 근육도 활 성화된다. 이런 운동을 수행하기 위해 특별한 노력이 필요한 것은 아 니지만 여기에는 복잡한 시간 조절과 시냅스 연결이 요구된다. 파킨슨 병에 걸린 환자들은 단순히 일어서거나 물컵을 집어드는 것과 같은 아 주 간단한 활동조차도 수행하기 어렵다. 앞서 살펴본 것처럼 감각수용기는 환경 속에 존재하는 여러 형태의 에너지를 신경충격으로 변화시키지만, 움직일 때는 위와 반대과정이 일어난다. 죽, 움직이기 위해서는 신경충격의 에너지가 근육과 골격을 움직일 수 있는 기계적인 에너지로 전환되어야 한다. 운동신경원에 의 해 골격근이 흥분하여 수축하면 이에 연결된 뼈를 움직임으로써 운동 이 일어난다. 신경계에 의한 운동통제는 복잡하며, 여기에는 척수의 반사기제를 비롯한 신피질, 기저핵 및 기타 뇌 구조의 신경회로 등이 관여한다. 이들 신경회로의 활동으로 인해 나타나는 마지막 결과가 근 육수축이므로 근육에 관해 먼저 살펴보기로 하자.
1 근육 근육에는 골격근 (skele t al muscle), 평활근 (smoo th muscle) 그리고 심 장근 (card i ac muscle) 의 세 가지 유형이 있다. 골격근은 골격에 부착되 어 있고, 평활근은 내장기관과 혈관벽에서 발견되며, 십장근은 심장에 서만 볼 수 있다. 대부분의 관찰 가능한 행동은 골격근의 운동으로 이 루어지기 때문에 먼저 골격근을 중점적으로 살펴보자. 골격근 조직의 기본 단위는 근섬유 (muscle fi ber) 인데, 이것은 배아기 동안 여러 개의 근육세포들이 합쳐져서 형성된 것이다. 근육조직의 가장 뚜렷한 특칭은 수축할 수 있다는 것이다. 근육의 수축은 근섬유 속에 있는 다양한 단백질의 모양이 변화됨으로써 나타 난다. 근육세포는 미오신 (m y os i n) 과 액틴 복합체 (ac ti n com p lex) 라는 두 가지의 주요 단백질 구조를 가지고 있으며, 아 둘은 서로 붙어 있 다. 액틴 복합체와 미오신은 근육이 흥분할 때 마치 노를 짓는 것처럼 서로 옆으로 스치면서 미끄러진다(그림 7-1).
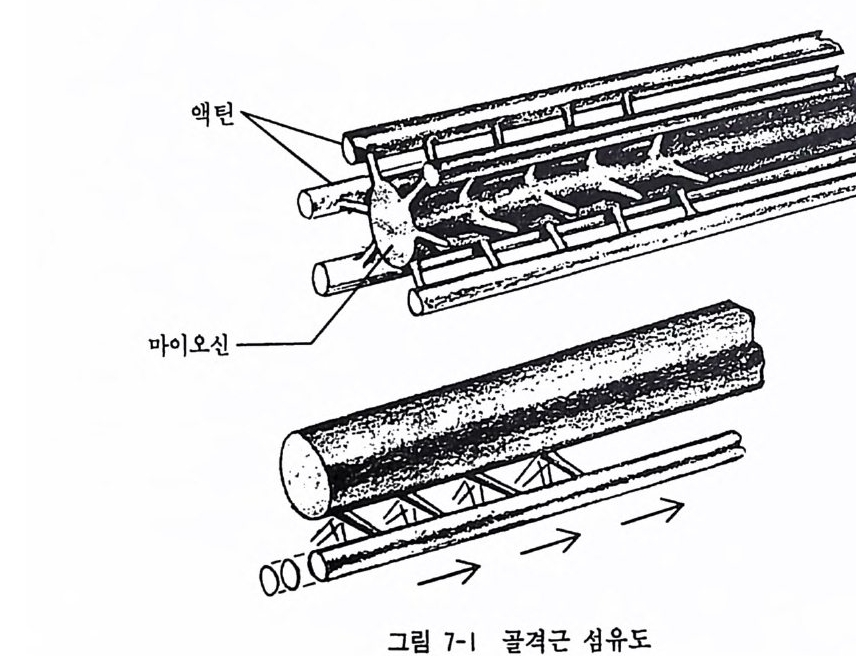 액틴
액틴
골격근은 근육의 수축속도에 따라서 속근(fa s t muscle), 완근 (slow muscle) 및 혼합근 (m i xed muscle) 의 세 가지 형태로 나누어진다. 짧은 시간 동안 강한 수축을 할 때 사용되는 속근은 완근에 비해 혈액과 미 오글로빈 (m yo g lob i n : 완근의 에너지 전달과정에 관여하는 단백질)을 적게 함유하고 있어서 엷은 색을 띠는데, 안구운동을 조절하는 근육이 속근 이다. 완근은 장거리 달리기를 할 때 사용되는 다리근육과 같이 지속 적인 운동에 관여하는 것으로 미오글로빈의 농도가 높고 혈액 공급이 풍부하게 이루어져 붉은빛을 띤다. 완근은 산소를 주 에너지로 사용하 는 이 른바 호기 성 과정 (aerobic pro cess) 에 의존하는 데 반해 , 속근은 산소를 필요로 하지 않는 무기성 과정 (anaerobic p rocess) 에 의존한다. 그러나 무기성 과정은 산소를 빌려서 사용하기 때문에 운동 후에 숨을 가쁘게 몰아쉬는 것과 같은 생리적 반응으로 소비된 산소를 보충해야 한다. 그러나 마라톤 경기를 할 때와 같은 특수한 경우에는 완근도 무 기성 과정의 에너지를 사용할 수 있다. 혼합근은 속근과 완근 모두를 포함하는 근육이다. 1.1 운동단위 눈, 안면, 혀 그리고 목의 근육을 제의한 일반적인 신체근육은 척수 의 전근을 통해 나온 운동신경원에 의해 활성화된다. 전형적으로 하나 의 운동신경원은 하나 이상의 근섭유와 연결된다. 하나의 운동신경원 과 아와 연결된 근섬 유들이 하나의 운동단위 (moto r un it)를 형 성 한다. 하나의 운동신경원에 의해 활성화되는 ' 근섬유의 수에 따라 운동의 정 확성이 결정된다. 예를 들어 눈의 운동을 조절하는 동안근 (oculomo t or muscle) 의 경 우는 개 개 의 운동단위 에 3 - 6 개 정 도의 근섭 유가 포함되는 데, 이런 이유로 신속하게 안구의 초점을 맞추는 운동, 즉 단속성 운 동 (saccades) 이 가능하게 된다. 한편 대되부에 있는 큰 근육의 운동단 위는 2,000 개 정도의 근섭유를 가지는데, 이처럼 한 운동단위에 많은 근섬유가 포함되기 때문에 걷고 달리는 데 유용하게 사용될 수 있다.
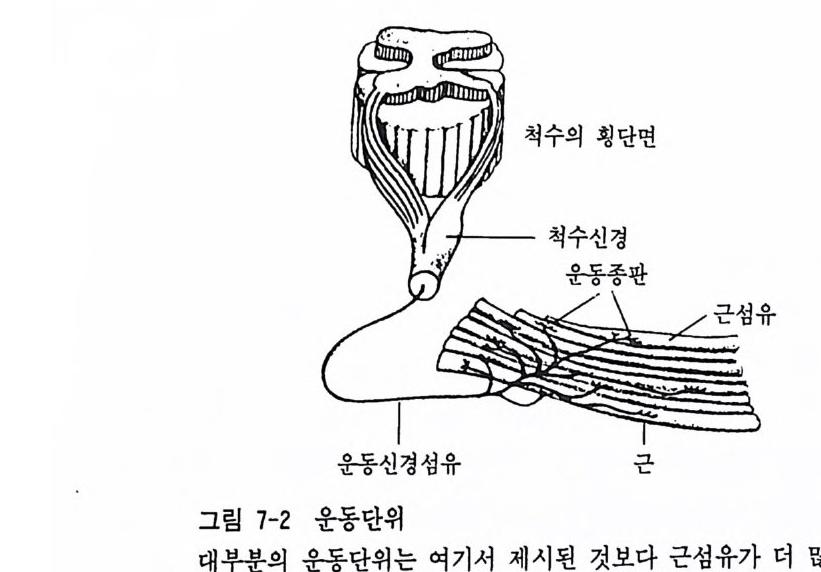 척수의 횡단면
척수의 횡단면
근섬유의 수가 적은 운동단위들은 손가락, 입술 그리고 혀에서 발견되 며, 근섬유의 수가 많은 운동단위들은 팔, 다리 그리고 등에서 발견된 다 (그림 7-2). 척추동물의 근섭유는 오직 수축반응만 보인다. 즉 하나 의 운동신경원이 활성화되면 그 운동단위에 포함되는 모든 근섬유가 수축한다. 척추동물의 근육 이완은 운동신경원의 흥분발사율이 감소되 어서 이루어지지만 무척추동물의 근육은 억제성 신경원(i nh i b it ory neu- ron) 의 지배를 받는다. 1.2 근육의 활성화 근섬유의 수축은 운동신경원에서 분비되는 신경전달물질의 작용으로 나타난다. ACh 는 모든 척추동물의 운동신경원에서 분비되는 전달물질 이다. 운동신경원의 축색종말과 근섬유 사이의 접합부를 신경근 연접 (neuromuscluar jun cti on ) 이 라고 한다(그립 7-3) . 신경 원과 마찬가지 로 근육도 ACh 에 의해 자극될 때 세포막 내의의 전위차를 유발한다. ACh 가 시냅스후 수용기와 결합하면 이온통로가 열리고 Na+ 과 K+ 이
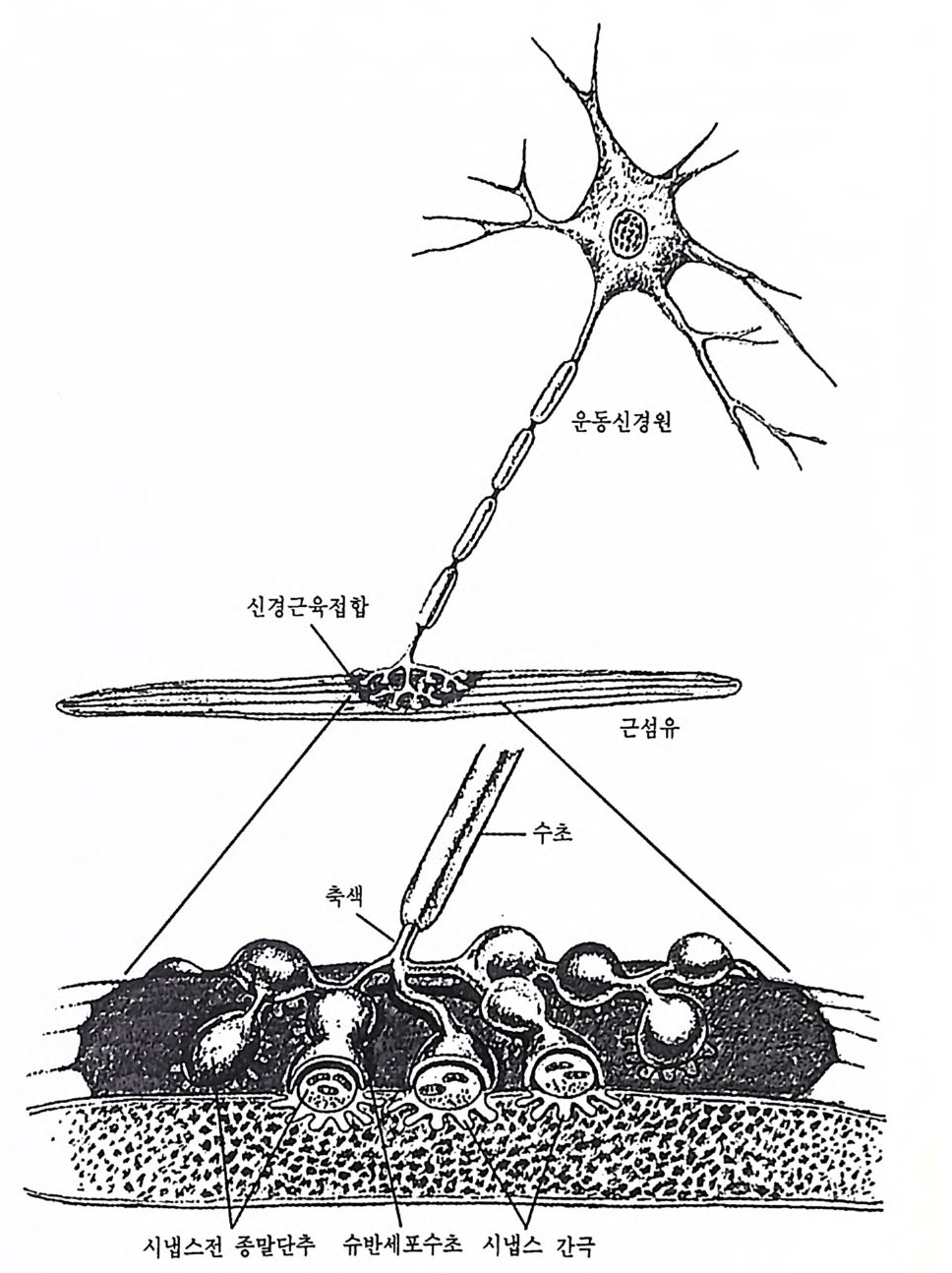 신경원
신경원
그림 7-3 신경근 연접도
근육세포의 세포막을 통과하게 된다. 근육세포의 세포막 내의에서 활 동전위가 기록된다는 것은 세포막이 탈분극화되었음을 의미하는 것으 로, 이것은 K 떡 방출보다 Na+ 의 유입이 더 많기 때문에 나타나는 현상이다. 그러나 실제로 근육세포 내의 근원섬유 (m y o fi ber) 가 수축을 야기 하는 것은 근섬 유의 근소포체 (sarcop la smi c reti cu lum) 에 저 장된 칼 슘 이온의 방출 때문이다. 근형질세망은 모든 근섬유에서 발견되는 그 물 모양의 구조이다. 근육세포 내에서 이용 가능한 칼슘의 양이 증가 되면 액틴 복합체 단백질이 미오신을 스치면서 미끄러지게 된다. 이렇 게 되면 근원섬유의 길이가 짧아져 근섬유가 수축되고, 나아가 근육이 이와 연결된 골격을 움직이게 된다. 코를 굵는 것과 같은 아주 간단한 운동에도 신경계의 여러 영역이 관여한다. 영장류의 경우 운동에 관여하는 뇌의 기본 영역은 척수, 뇌 간, 운동피질 그리고 전운동피질(p remo t or co rt ex) 의 네 영역이다. 이 둘 기본 영역 의에 소뇌와 기저핵 (basal g an gli a) 도 운동의 시작과 조 절에 기여한다. 2 척수 운동신경원의 세포체는 척수의 전각에 있으며, 이를 전각세포 (ventr a l horn cell) 라 부른다. 모든 운동은 바로 이 신경 원에서 일어나 는 활동에 기인된다. 인간의 경우 운동신경원의 수는 약 40 만 개에 불 과하나 이들 운동신경원에 영향을 미치는 신경원의 수는 수십억 개에 이론다. 운동의 통제는 여러 개의 운동신경원으로 구성되는 최종 공통 로(fin al common p a th)를 통해 이루어지게 된다. 2.1 반사 운동통제라는 측면에서 볼 때 척수의 또 다른 특칭은 척수반사
(spi na l re fl ex) 를 담당하는 신경회로가 존재한다는 것이다. 반사 (re fl ex) 란 피부나 관절에 자극이 가해질 때 자동적으로 나타나는 선천 성 운동을 가리킨다. 반사의 한 가지 좋은 예가 신장반사 (s t re t ch refl ex ) 이다. 골격근은 신장될 때 자동적으로 수축하게 되는데, 이를 신장반사라 한다. 이것은 근육조직의 탄력성 때문에 나타나는 것이 아니다. 이런 사실은 근육조직을 활성화시키는 운동신경원의 축색을 절단해 보면 쉽 게 증명되는데, 이 경우에는 근육이 신장되어도 더 이상 수축되지 않 는다. 또한 운동신경원의 축색이 손상되지 않더라도 근육에서 척수로 들어오는 감각신경원 축색이 절단되면 신장반사는 이루어지지 않는다. 이 실험결과는 신장반사를 일으키는 데는 감각 입력과 운동 출력이 모 두 필요하다는 것을 의미하는 것이며, 이것은 모든 반사에 적용된다. 가장 쉽게 관찰할 수 있는 신장반사의 한 예가 술개건 반사 (knee jer k refl ex ) 이 다. 무릎 바로 아래 에 위 치 하는 건 (ten don:) 을 두드리 면 이 건에 연결되어 있는 사두근(q uadr ic e p s muscle) 이 수축되어 무릎이 갑자기 앞으로 움직이게 된다. 이 수축은 건이 자국을 받은 후 약 0 . 05 초쯤 나타나는데 , 이 정도의 반웅 잠재시간은 술개 건 (knee- ten don) 에 가해진 자극에 대한 반응으로 수의적으로 . 다리를 움직 이는 데 최소한 0 . 250-0 . 300 초가 소요되는 것과는 차이가 있다. 슬개건 반 사에는 건을 두드리는 신호를 전달하는 감각신경과 사두근으로 향하는 운동신경이 접합하는 단 하나의 시냅스가 관여하는데, 이를 단일 시냅 스성 반사 (monos yn a ptic refl ex ) 라고 한다. 그러나 대부분의 척수신경 원은 하나 이상의 시냅스를 포함하는데, 이를 다중 시냅스성 반사 (po lys y n a pt ic refl ex ) 라 한다. 2.2 근육 속의 감각기 관 반사는 근육이나 근육과 연결된 건으로부터 유입되는 감각 입력에 의존한다. 근육에는 근육의 길이, 근육의 신장 정도 그리고 근육의 긴
장 정도에 관한 정보를 끊임없이 뇌와 척수로 보내주는 미세한 감각기 관이 있다. 근육의 길이와 신장의 정도를 검색하고 보고하는 감각기관 은 두 가지 형태가 있는데, 그 중 근육의 길이에 관한 정보를 계속 수 취 하는 감각기 관을 근방추기 관 (muscle spi nd le orga n ) 이 라 한다. 2.3 근방추기관 근방추기관은 근섭유 속에 묻혀 있는 미세한 감각기관이다(그림 7-4). 골격근에는 두 가지의 상이한 근섬유가 있는데, 하나는 수축하 여 골격을 움직이게 하는 큰 근섬유이고, 다른 하나는 작고 약한 근섬 유로 근방추기관 내에서 발견되는 것이다. 이들 두 가지 유형의 근섬 유를 구분하기 위 해 큰 근섬 유를 방추의 근 (extr a fu s al muscle) 이 라 부르 고 근방추기관 내에 있는 작은 근섬유를 방추내근(i n t ra fu sal muscle) 이 라 부론다. 또한 방추기관에는 두 가지의 근방추 수용기가 존재한다. 죽 윤나선형 수용기 (annulospi ra l rece pt or) 는 Ia 군 구심성 통로를 통해 정 보를 전 달하고, 꽃가지 형 수용기 (flow er-spr a y recept or ) 는 II 군 구심 로를 통해 척수로 입력정보를 보낸다. Ia 구심로는 주로 근육의 길이
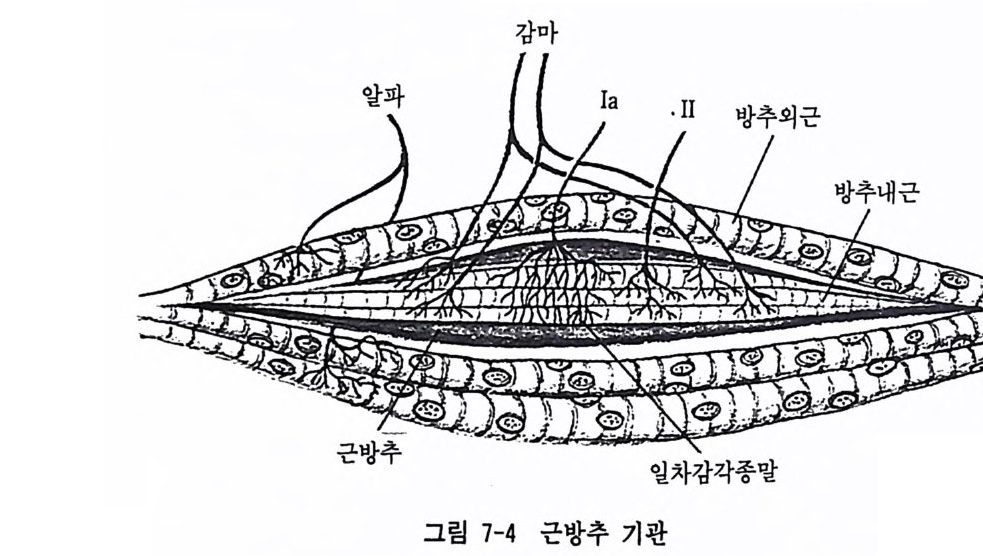 감마
감마
와 관련되고, II 군 구십로는 갑작스런 운동과 관련된다. 신장수용기 (str e tc h recep tor ) 는 크기 가 큰 방추의 근을 활성 화시 키 는 운동신경원과 흥분성 시냅스를 형성한다. 방추내근은 크기가 큰 방추 의근과 평행으로 연결된다. 이것은 방추의근섬유가 신장될 때마다(근 육 이완이 될 때) 크기가 작은 방추내근섬유 또한 신장된다는 것을 의 미하며, 방추의근섬유가 수축하면 방추내근섬유 또한 수축한다. 방추 내근섬유는 너무 약해서 운동을 일으키지는 못하고 근육이 신장되는 정도에 대한 정보를 중추신경계에 전달하는 제한된 역할만 수행한다. 방추의근이 이완되어 길이가 늘어나면 방추내근섬유도 신장되어 방추 기관에서 척수로 향하는 구심로에 신경충격을 발생시킨다. 이런 입력 이 동일한 근육의 방추의근섬유를 지배하는 운동신경원에 흥분을 야기 한다 . 그 결과로 운동신경원의 흥분발사를 더욱 빨리하여 방추의근섭 유를 수축하게 한다. 이와 같은 수축이 일어날 때는 방추의근은 짧아 지며 방추내근섬유에 대한 신장도 약화된다. 방추내근심유의 길이가 짧아져 신장수용기의 구심로에서 나오는 구심성 입력이 감소되기 때문 에 운동신경원의 흥분도 낮아져 근육은 다시 이완된다. 이런 선택적이 고 자동적인 반사체계가 근육의 길이를 일정한 범위로 유지시켜 근육 긴 장 (muscle ton e) 을 유발한다. 2.4 상호억제 정상적인 운동은 굴근과 신근 사이의 협조에 의해 이루어진다. 우리 가 걸을 때 다리를 들어올리고, 사지를 몸통 가까이로 끌어당길 때 사 용하는 근육이 굴근이다. 굴근은 중력의 힘과 상반되는 방향으로 작용 한다. 반대로 신근은 사지를 몸통으로부터 멀리 뻗치게 하므로 중력의 힘과 같은 방향으로 작용하는 경향이 있다. 따라서 걸음을 걷는 것과 같은 조화로운 운동을 수행하기 위해서는 굴근과 신근이 동시에 수축 되어서는 안 된다. 굴근과 신근이 동시에 수축하게 되면 사지가 뻣뻣 하게 굳어 지는 근긴장 (s p as ticity) 상태를 초래 한다. 반사는 억 제 성 간
신경원을 척수의 신경회로에 편입시킴으로써 신근이 활성화될 때 굴근 울 억제시키는데 (반대과정도 마찬가지임), 이 과정을 상호억제 (reci pro cal i nh i b iti on) 라고 한다. 따라서 술개건 신장반사의 경우에 술개전을 가볍 게 두드리면 대되부의 신근이 수축됨은 물론 하지의 신장을 방해하는 굴근이 억제된다. 척수 수준에서도 선천적인 신경회로가 다양한 근육 집단의 수축을 적정수준으로 유지시킨다. 척추동물들은 단순한 〈 반사 로보트 〉 가 아니며, 대부분의 운동은 엄 격한 의미에서 일련의 반사로 분해되지 않는다. 그럼에도 불구하고 이 런 반사를 생성시키는 신경회로는 우리가 행하는 모든 운동에 적용된 다. 반사는 척수의 반사회로를 활성화시키는 고위 뇌 영역을 내려온 신경충격을 통해 전반적인 운동형태로 짜여진다. 정상작으로 반사는 고립된 상태로는 작용하지 않지만, 신장 반사, 철수 반사 (w ith drawal refl ex ) , 그리 고 굵기 반사 (scratc h refl ex ) 와 같은 반사는 척 수 가 절단되어 고위 운동중추에서 내려오는 모든 입력정보가 차단된 동 물에서도 나타난다. 이처럼 척수가 뇌로부터 내려오는 모든 입력정보 와 격 리 되 어 있을 때도 나타나는 반사를 척수반사 (spi na l refl ex ) 라고 한다. 3 뇌간 뇌간에서도 운동을 통제한다. 앞 장에서 본 것처럼 전정계에서 나온 정보는 먼저 뇌간신경핵에서 처리되고, 그 후 자세 조정을 위한 운동 명령이 척수의 운동신경원으로 전달된다. 권두경기중 턱을 타격당해 머리가 뒤로 젖혀져 뇌간이 손상되면 혼수 또는 마비상태가 나타나고 심한 경우 생명을 잃기도 한다. 뇌간의 직접적인 손상이나 척수와 뇌간을 분리시키는 손상은 척수에 여러 가지 반응을 일으킨다. 인간에서 뇌간의 미측 끝부분의 척수를 철단하면 모든 수의적 운동이 상실되고, 절단 부위 이하의 모든 감각
이 없어지며, 수 주 이내에 모든 척수반사가 사라지는데, 이런 상태를 척수 쇼크 (spi na l shock) 라고 한다. 척수 쇼크 이 후 척 수반사는 서 서 히 되 살아나며 , 가끔 과장된 형 태를 보이 기도 하는데 , 이를 반사항전 (hy per- refl ex ) 이 라고 한다. 척수가 심 하게 손상된 환자의 기능을 회복시키는 것은 많은 연구에도 불구하고 아직까지는 불가능한 것으로 알려져 있 다. 그러나 실험동물을 대상으로 하여 조직이식기법을 사용한 연구들 에서 다소 성공적인 경우도 있다 (Gash & Sladek, 1988). 3.1 제뇌경직 상구체와 하구체 사이의 뇌간이 절단되면 제뇌경직 (decerebrate r igi d ity)이라는 현상이 나타난다. 이런 현상은 약 1 세기 전에 동물을 대 상으로 Sherr i n gt on 이 라는 영국의 신경 생 리 학자에 의 해 처음으로 밝 혀졌으며 뇌간이 손상된 사람에서도 관찰된 바 있다. 제뇌경직의 특징 은 항중력근 (an tig av ity muscle, 대부분의 동물에서는 신근)이 비정상적으 로 만성적인 수축을 보인다는 점이다. 그러므로 몸과 사지의 신근이 계속 경직되고 활성화되어 뻣뻣하게 굳어진다. 3.2 중추운동 프로그램 대부분의 운동은 반사적인 것이 야니라 의식적으로 수행되는 수의적 인 것이다. 걷기, 삼키기 그리고 달리기와 같은 수의적인 운동에는 중 추 프로그램이 개입되는 것으로 생각된다. 특정 운동이 중추 프로그램 의 통제를 받느냐 받지 않느냐 하는 것은 그 운동이 독립적으로 수행 될 수 있는 정도에 따른다. 걷기를 예로 들어보자. 뇌간의 미측 끝부 분의 척수가 절단된 고양이를 움직이고 있는 트레드밀 위에 울려놓고 뇌간의 한 작은 부위를 전기자극하면 척수가 손상된 이 고양이가 정상 적으로 걸을 수 있다. 이 경우 자국에 의해 유발된 걸음걸이의 리듬은 분명히 선천적인 중추운동 프로그램에 기인된 것이지 전류의 파동수와
관련된 것은 아니다. 비록 이런 걷기는 트레드밀의 움직임에 의해서가 아니라 뇌간의 자극에 의해 생기는 것이지만 걸음걸이의 형태는 트레 드밀의 속도와 기울기에 따라 변화된다. 걷기에 관여하는 운동 프로그램은 척수에 위치하는 것으로 생각되는 데, 그 이유는 척수가 절단된 동물의 척수를 자국해도 동일한 반응이 유발될 수 있기 때문이다. 더구나 개개의 다리들이 나름대로 걷는 운 동을 보여줄 수 있는 것으로 보아 개개의 다리는 독립적인 프로그램을 가지고 있는 것 같다. 이런 걷기에는 감각 입력이 필요 없는데, 이것 은 후근의 감각축색을 절단하여도 자극에 의해 나타나는 걷기가 사라 지지 않는다는 실험에 의해 입증된다. 위에서 언급한 바와 같이 뇌간 에는 자국을 받을 때 걷기를 위한 척수운동 프로그램을 촉발시킬 수 있는 신경원이 있다. 나아가 뇌간운동 촉발영역뿐만 아니라 척수의 운 동 프로그램까지도 직접 활성화시킬 수 있는 다른 뇌 영역(운동피질) 또한 존재할 것이라고 생각할 수 있다. 여러 가지의 독특한 운동 프로그램도 존재한다는 증거들도 보고되었 는데 , 여 기 에 는 고양이 의 삼키 기 (Doty , 1951 ; Doty & Bosma, 1956) , 설 치류의 안면 문지르는 행동 (Fen t ress, 1983) , 그리고 발기 와 사정 과 같 은 생식행동 (Ha rt, 1978) 을 위한 운동 프로그램 등이 포함된다. 4 운동피질 1870 년에 Gusta v Fr it sch 와 Edourad Hit z ig라는 두 독일인은 개와 원숭이의 전두피질의 특정 영역을 전기자국하면 사지의 운동이 나타난 다는 사실을 발견했다. 그 후 몇몇 과학자들과 신경외과 의사들도 오 랫동안 운동피질의 기능을 연구하였는데, 그 중 유명한 사람이 캐나다 의 Wi lde r Pen fi eld 이다. Pen fi eld 와 그의 동료들은 뇌수술 동안 환자 의 뇌를 자국하여 일어난 운동반응의 지도를 만들었다 (Penfi eld & Rasmussen, 1950). 보통 이 런 수술은 국소마취 아래서 이루어 지 기 때 문
 .... .v..·
.... .v..·
그립 7-5 인간 뇌피질의 운동 영역 뇌의 횡단면으로 본 한쪽 반구의 운동 영역. 개개 신체 영역에 할당되는 피질 의 위치와 상대적 크기를 신채운동뇌도로 나타내었다.
에 환자는 자극의 효과를 알아보려는 의사의 물음에 의식장애 없이 대 답할 수 있다. 이런 자국은 자극을 받는 뇌 영역에 따라 여러 가지 의 식적 감각이나 운동을 야기시킬 수 있으며, 이런 방식의 뇌 전기자국 은 뇌의 건강한 조직과 병든 조직을 식별하는 데도 중요하다. 자극의 결과가 그립 7-5 에 제시되어 있는데 이를 신체운동뇌도라 한다. 운동 피질은 뇌의 중심렬 바로 앞부분에 위치하는대, 이것은 Brodman 의 4 번 영역과 거의 일치하는 곳이다. 그림 7-5 에 제시된 바와 같이 전기자극에 의해 유발되는 운동이 운 동피질상에 마치 〈작은 사람〉 또는 〈운동소인 (mo t or homunculus) 〉의
모습을 닮은 신체부분들로 표상될 수 있다. 신체운동뇌도는 특정 신체 부위의 운동에 관여하는 운동피질 신경원의 수는 그 신체 부위에서 수 행되는 운동의 정확성과 비례한다는 사실을 보여주고 있다. 4.1 조직화 운동피질을 구성하는 신경원의 축색은 뇌의 여러 부분으로 투사된 다. 하나의 중요한 출력 통로가 피 질 척 수로 (cort ico spi na l tra ct) 인 데 , 이 것은 척수로 직접 하행하는 운동피질 축색으로 이루어져 있으며 척수 에서 가지를 뻗어 간신경원, 알파 운동신경원 또는 감마 운동신경원과 시냅스를 형성한다. 또한 피질척수로의 신경원들은 뇌간신경핵의 운동 조철신경원과도 시냅스를 이룬다. 피질척수로는 추체로(py ram i dal t rac t)라고도 부르는데, 그 이유는 피질척수로의 축색이 연수에서 뇌의 반대 편으로 건너 가 연수추체 (medullary pyra mi d) 를 형 성 하기 때 문이 다. 실제로 피질척수로에 있는 백만 개 이상의 축색들 중에서 운동피질의 신경원에서 기인되는 것은 불과 30% 정도에 불과하며 나머지 축색은 전두영 (Brodmann 의 6 번 영 역 ) 과 두정 엽 (Brodmann 의 1, 2, 3 번 영 역 ) 과 같은 인접 피질영역에서 나오는 것이다 (Ghez , 1985). 앞에서 Brodmann 1, 2 및 3 번 영역이 신피질의 1 차 체성감각을 구 성한다는 사실을 알았다. 그리고 이 감각영역이 운동을 통제하는 영역 으로 축색을 보낸다는 것을 보고 의아하게 생각했을 것이다. 그러나 운동에 관여하는 뇌 영역이 근육과 피부수용기로부터 정보를 받아들이 는 것은 당연하며, 실제로 운동피질은 피부, 관절 그리고 건수용기로 부터 입력정보를 받아들인다. 운동피질과 감각피질 사이의 구분은 절 대적인 것이 아니라 상대적인 강조일 뿐이다. 우리가 뇌의 작용에 대 해 공부하면 할수록 뇌의 다양한 성분들이 통합되어 있다는 사실을 알 게 된다. 뇌의 체성감각 및 운동영역 사이의 밀접한 관계가 이의 한 예인데 운동피질신경원의 축색은 피질척수로 의에도 운동과 관련되는 뇌의 여러 부위로 두사된다. 예컨대 운동피질에서 나온 축색은 시상,
소뇌, 미상핵 그리고 뇌간의 망상체 등으로 두사된다. 운동피질로 유입되는 입력정보는 운동통제와 근육, 피부 및 관절에 서 나온 감각정보 사이에 밀접한 관련성이 있음을 보여준다. 운동피질 신경원은 체성감각정보를 중개하는 시상의 복의측핵으로부터 입력을 받아들이고, 체성감각피질의 신경원들도 운동피질로 향해 축색을 보낸 다. 실제로 운동피질의 어떤 영역이 자국되면 이와 연결된 일련의 근 육에서 운동이 야기되는데 이때 자국되는 운동피질의 신경원들은 운동 에 관여하는 근육, 건 그리고 피부에 있는 감각수용기로부터 감각정보 를 수용한다. 시각, 청각 및 체성감각과 관련되는 피질영역과 마찬가 지로 운동피질도 뇌 표면과 수칙을 이루는 기능적 주(柱, column) 를 기 초 단위로 한다. 따라서 어떤 수직 주 내에 있는 신경원들이 전기적으 로 자국되면 이와 관련되어 있는 근육에서 운동이 나타나게 된다. 4.2 운동피질 손상의 영향 인간의 운동피질 손상은 뇌일혈 (s t roke) 또는 기타 의상에 의해 나타 난다. 뇌일혈은 어떤 뇌 부위에 혈액 공급이 차단될 때 나타나는 신경 학적 증후이다 (Kolb & Whis h aw, 1990). 만약 뇌일혈이 뇌의 어느 한쪽 부위에 발생하게 되면 그 부위와 반대쪽 신체에 부분적 또는 전체적 마비가 초기 증상으로 나타난다. 그 후 몇 주가 지나면 마비는 약화되 고 근긴장, 즉 경련이 일어난다. 이 단계에서 반대쪽 팔은 계속적으로 구부러지지만 반대쪽 다리는 계속적으로 신장된다. 회복의 정도는 뇌 일혈의 정도와 그 밖의 조건에 따라 달라진다. 시간이 지나도 뇌일혈 의 영향을 받은 부위의 말초근육(손, 손목, 발 그리고 발목)은 힘이 없 고 어색하며, 조정능력이 잘 이루어지지 않는 등의 후유증이 계속된 다. 손의 운동이 어색한 것은 손가락 하나하나의 개별적 운동이 불가 능하기 때문이다. 그러나 운동피질에 십한 손상이 있어도 영구적인 마 비가 나타나지 않는다는 사실은 운동피질에서 나오는 정상적인 입력정 보가 없어도 뇌의 다른 영역에 의해 어느 정도 운동이 이루어질 수 있
음을 보여주는 것이다. 이런 사실로 미루어볼 때 운동피질의 주된 기 능은 손과 발의 섬세하고 통합된 운동을 야기하고, 정확하고 신속한 운동을 촉진시키는 것이라는 점을 알 수 있다. 4.3 단일운동 신경원의 활동분석 과거 30 년 동안 과학자들은 각성 상태에 있는 원숭이가 어떤 간단한 운동을 수행하고 있는 동안 피질척수로상에 있는 단일 신경원의 활동 울 기록하기 위한 기법을 개발해 왔다. Edward Evar t s 라는 연구자가 이 방법을 처음 소개했는데, Evar t s (1 968) 는 원숭이가 돌쩌귀가 달린 막대기를 좌우로 움칙이고 있는 동안 피질척수로의 신경원에서 일어나 는 활동을 기록하였다. 즉 이 연구에서 한쪽 방향으로의 운동은 손목 을 굽히는 일이고, 다른 쪽 방향으로의 운동은 손목을 펴는 일이다. 막대를 정확한 위치로 움직이는 데에 요구되는 힘이 증가 또는 감소됨 에 따라 신경원의 흥분발사율도 달라진다. Evar t s 는 피질척수로에 있 는 신경원의 흥분발사율은 손목의 운동방향에 대한 함수가 아니라 손 목근육에 의해 사용되는 힘에 대한 함수로 증가되거나 감소된다는 사 실을 발견하였다. 4.4 추론발사 피질척수로의 일부 신경원은 뇌간의 신경원과 시냅스를 이룬 후 소 뇌로 두사된다. 이 통로는 앞으로 곧 일어날 운동에 대한 사전정보를 소뇌로 보내고, 소뇌는 이 정보를 사용하여 예측되는 몸의 움직임에 부합되도록 자세를 (특히 머리와 눈) 조정하게 된다. 이와 같은 〈수축이 일어날 것이란 예고〉는 안정된 시각세계를 유지하기 위해 머리와 눈의 운동을 조절하는 데 특히 유용하다. 이런 형태의 내적 신호를 추론발 사 (corollary dis c harge ) 라고 한다.
4.5 실행증 만약 최근 뇌일혈을 경험한 환자에게 성냥불을 입으로 불어 끄는 시 늄을 해보라고 하면, 비록 환자가 이 지시에 협조적이며 청각에 아무 런 이상이 없음에도 불구하고 잘 수행하지 못한다. 그러나 이번에는 미리 성냥불을 켜서 환자 앞으로 내밀면 환자는 신속히 그것을 불어서 끌 수 있다. 이 행동만 두고 보면 환자가 장난을 하고 있는 것인지 아 니면 이것이 뇌일혈에 의한 비정상적 행동결과인지 궁금하다. 위에서 살펴본 운동장애가 심한 뇌일혈 후에 공통적으로 나타나는 실행증 (a p rax i a) 의 한 예이다. Norman Geschw i nd(1975) 는 실행증이 란 운동강도의 빈약함, 운동의 조정 불능, 감각 상실 또는 행동 요구 에 대한 이해 불능 또는 주의 결핍 등의 원인으로 일어나는 운동장애 가 아니라 학습된 운동장애라고 정의하였다. 실행증은 언어를 이해하 지 못하는 실 어 증 (aph a sia ) , 진행 성 마비 (pa resis) 또는 뇌 손상에 의 해 나타나는 불완전한 마비와는 구분되어야 한다. 실행증은 운동피질의 칙접적인 손상에 의해 생기는 것이 아니라 두정 또는 전두피질의 손상 에 의해 나타나는 것이 보통이다. 5 기저핵 기저핵도 운동을 시작하고 실행하는 데 중요한 역할을 하는 것으로 알려져 있다. 기저핵은 미상핵 (caudate nucleus), 담창구(g lobus pal - lidu s) 그리고 피각(p u t amen) 으로 이루어져 있다. 그 밖에 기저핵의 운 동기능과 밀접하게 관련되어 있는 구조로서 시상하핵 (sub th alam ic nucleus) 과 뇌 간의 혹질 (substa n ti a nig ra) 이 있다. 그림 7-6 에 서 도 볼 수 있는 바와 같이 기저핵은 시상 가까이 위치하며 신체운동의 시작과 조절에 중요하다. 기저핵은 운동피질, 시상 그리고 뇌간 구조들과 일 련의 복잡한 신경고리로 연결되어 있지만 기저핵으로부터 척수의 운동
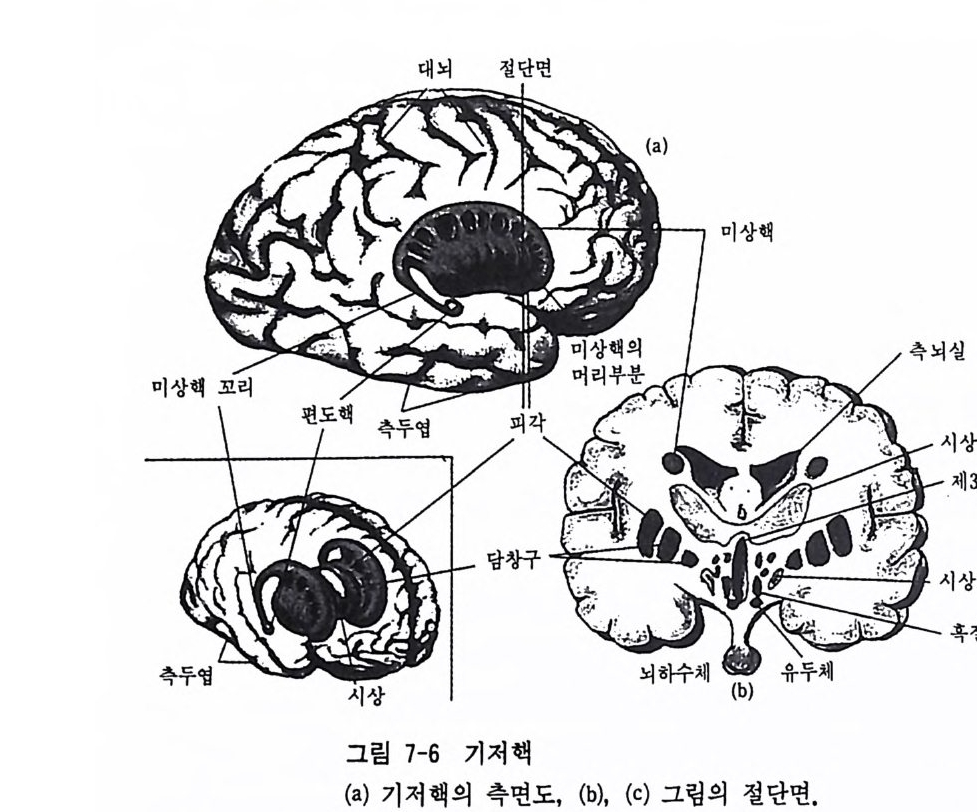 대뇌 절단면
대뇌 절단면
신경으로 직접 두사되는 통로는 없다. 기저핵이 운동에 관여한다는 가장 확실한 증거는 이들 구조에 장애 가 있는 환자들에서 찾아볼 수 있다. 기저핵의 장애와 관련되는 가장 혼한 질병은 파킨슨 병과 헌팅턴 병이다. 파킨슨 병 환자는 사지와 몸 통의 불수의적인 떨림(t remor) 을 보이고 운동의 시작과 수행이 느리며 발을 질질 끄는 걸음걷이를 보인다. 이 병에 걸리면 기억력의 저하와 갇은 인지적 결손도 함께 나타난다. 이 병은 미상핵으로 신경분포를 굴民 흑질의 퇴화 때문에 발생한다. 흑질의 신경세포에서 주로 사홍되 는 신경전달물질은 도파민 (do p am i ne) 이다. 파킨슨 병을 치료하기 위해 L-DOPA 라는 도파민의 전구물질을 사용하면 증세가 상당히 호전되지 만, 치료효과의 지속시간이 짧고, 반복사용시 효과가 떨어지며, 목과 얼굴 등의 근육이 불수의적으로 뒤틀리는 부작용도 나타나기 때문에
궁극적인 치료약물로 사용되기에는 부적당하다. 최근에는 세포이식 (t ransp lan t a ti on) 으로 파킨슨 병을 치료하려는 시도가 이루어지고 있 다. 환자 자신의 부신수질 (adrenal medulla) 세포를 미상핵으로 이식하 면, 이식된 세포들이 소량의 도파민을 분비한다. 그러나 이와 같은 실 험적 수술의 효과가 아직 확실히 입증된 것은 아니다. 한편 헌팅턴 병은 사지와 안면근육을 통제할 수 없을 정도로 빠르게 움직이는 질병이다. 이 병은 진행성 퇴화 질환으로 걷기, 사고, 언어 및 기타 인지 기능에 심한 장애가 나타난다. 기저핵에서 ACh 와 GABA 라는 신경전달물질을 분비하는 세포집단의 퇴화가 이 병의 발 생과 관련되어 있는 것으로 밝혀져 있다. 현재까지는 적절한 치료방법 이 없지만 동물모형을 사용하여 신경이식을 시도한 실험들은 약간의 기능회복을 보고하고 있다 (Isacson 등, 1985). 헌팅턴 병은 유전성이며 30 대 후반이 나 40 대 후반에 주로 발병 되 어 10-15 년 이 내 에 사망한다. 헌팅턴 병은 베네수엘라의 마르카이보 호수지역에서 가장 많이 발견되 었기 때문에, 연구자들은 이 사실을 바탕으로 이 병의 유전적 증거를 추적할 수 있었다 (Gusella 등, 1983). 헌팅턴 병의 유전자는 4 번 염색 체에 위치한다. 유전자 검사로 이 병의 유무몰 파악할 수 있는데, 이 검사는 나이에 상관 없이 언제든지 가능하다. 이 병의 유전자는 우성 이며 이 유전자를 가진 모든 사람은 이 병에 컬리게 되지만 환자의 형 제, 자매라도 이 유전자를 보유하지 않으면 병에 걸리지 않는다. 6 소뇌 소뇌를 구성하는 신경원은 크기가 작아 그 무게가 대뇌 무게의 10% 에 불과하지만 신경원의 수는 대뇌의 1/2 에 해당한다 (Ghez & Fahn, 1985). 소뇌는 3 개의 영으로 구성되어 있고, 이들 영은 다시 10 개의 더 작은 소영 (l obules) 들로 나누어진다. 소뇌의 기능을 간단히 요약하는 것은 쉽지 않지만 일반적으로 운동,
신체균형 그리고 근육긴장을 조절하는 것으로 알려져 있다. 소뇌기능 의 두 가지 특징은 그 작용이 의식에 영향을 주지 않고 자동적으로 수 행된다는 것과 그 작용이 간접적이라는 것이다. 소뇌로 들어가는 감각 정보는 대뇌피질에 의해 수용된 감각정보와는 달리 어떤 의식적 감각 도 야기하지 않는다. 두번째 특칭인 간접적 작용이란 소뇌의 출력정보 가 직접 운동신경원에 도달하지 않는다는 것이다. 소뇌에서 나가는 모 돈 신경정보는 신피질이나 뇌간과 같은 운동체계에서 나온 신호를 조 정 또는 조절하는 기능을 한다. 소뇌로 들어가는 입력정보는 모든 감각을 표상하는 수용기뿐만 아니 라 뇌간과 대뇌 신피질의 신경원에서도 나온다. 구심성 정보를 소뇌로 전달하는 신경원의 축색들이 합쳐져서 소뇌각 (cerebellar p eduncle) 을 형성하고, 이것이 소뇌와 뇌간을 연결한다. 소뇌의 출력정보는 운동에 관여하는 대뇌피질 영역과 전정신경핵과 같은 뇌간의 신경원으로 연결 된다. 소뇌의 세포들은 정확한 기하학적 형태로 3 개의 소뇌피질충에 배열되어 있다. 소뇌를 구성하는 신경원은 성상세포 (s t ella t e cell), 바구 니 세 포 (basket cell) , 퍼 킨 지 세 포 (Purkin j e cell) , 골지 세 포 (Golgi cell) , 그리고 과립세포(gr anule cell) 의 5 가지로 구분된다. 이들 중 퍼킨지 세 포만이 소뇌 밖으로 축색을 내보낸다. 그러므로 소뇌의 출력정보는 인 간의 경우 약 700 만 개에 이르는 퍼킨지 세포에 의해서만 전달되는데 퍼킨지 세포의 출력정보는 모두 억제성이다. 퍼킨지 세포의 수지상돌 기는 10 만 개 이상의 수지상가시를 갖고 있어서 소뇌의 다른 신경원들 과 20 만 개 정 도의 시 냅 스를 형 성 한다 (Shep he rd, 1979) . 소뇌의 손상은 그 위치와 손상 정도에 따라 여러 가지 효과를 보여 주지만 일반적으로 사지운동과 안구운동을 방해하고 균형운동에 장애 를 초래하며 근육의 긴장을 감소시킨다. 운동피질을 손상시킬 때와는 달리 소뇌 손상은 근육수축의 강도에는 큰 영향을 미치지 않는다 (Ghez & Fahn, 1985) . 소뇌의 기능에 관한 생각은 과거 100 여 년 동안 여러 가지 가설로 제안되어 왔고, 이 가설에 대한 수정이나 새로운 가설이 끊임없이 제
시되고 있다. 다양한 가설에서 발견되는 공통점은 소뇌가 의도된 운동 과 실제 운동을 서로 비교하는 기능을 갖고 있다는 것이다. 앞서 제시 된 추론발사라는 개념에 의하면, 지금 막 시작되려는 운동에 관한 정 보는 소뇌로 보내지며, 그 후 소뇌의 회로들은 의도된 운동 유형과 실 제적인 운동근육과 관절에 의해 생성된 되먹임을 서로 비교하게 된다. 이들 사이의 불일치나 수행상의 오류는 뇌간이나 운동피질로 전달되는 소뇌의 명령에 의해 교정될 수 있다. 소뇌에 관한 또 하나의 가설은 소뇌가 탄도운동 (ba lli s tic movement) 에 관여한다는 것이다 (Kornhuber, 1974). 탄도운동은 공간 속의 한 고 정점을 향해 이루어지는 것이다. 이 운동은 되먹임을 줄 수 없을 정도 로 빨라 운동의 궤도에 영향을 줄 수 없다. 안구의 운동이 탄도운동의 한 예인데 안구운동은 소뇌가 손상되거나 황폐화되면 경련이 일어나거 나 느려진다 . 소뇌의 퇴화는 나이가 많은 사람들에게 공통적으로 나타 나며 비타민 B1 결핍과도 관련 있다. 한편 숙련된 운동은 오랜 동안의 학습을 필요로 하는데, 숙련된 기 술을 학습하는 동안에 소뇌회로가 변화된다는 증거들도 제시되어 있다.
제 8 장 갈증과 기아 동물과 마찬가지로 인간도 끊임없이 체액을 보충하지 않으면 안 된 다. 갈증을 느끼는 데는 어떤 신경활동이 관여하는가? 갈증을 설명하 는 주요 이 론으로 이 중고갈이 론 (double depl e ti on the ory ) 이 있다. 이 이 론에서는 갈증이란 세포 의 공간에서의 체액량과 세포 내 공간에서의 체액량을 동시에 고려해야 한다고 강조한다. 뇌와의 상호 정보교환을 통해 이 두 체액구획의 상태가 감지되며, 이 두 구획 가운데 어느 한 쪽에서라도 수분이 결정적 수준 이하로 떨어지면 수분보존반응과 갈증 이 동시에 촉발된다고 한다. 또한 유기체는 끊임없이 에너지를 취하고 이를 저장하지 않으면 안 된다 . 일반적으로 음식 섭취와 체중은 밀접하게 관련된다. 기아감과 포만감을 야기하는 신호자극을 찾기 위해 많은 연구들이 행해졌다. 그 중 한 이론은 포도당이 뇌에서 기아감과 포만감을 야기하는 결정적 물 질이라고 주장하지만 아직 그 증거는 명확하지 않다. 한편 체중을 어 떤 결정점 (setp o in t) 수준에 맞추어 조절한다는 가설도 제시되어 있지 만 모든 인간이 자신의 체중을 완벽하게 조절하지는 못한다. 비만증에 관련된 문제들과 비만증의 원인 등을 알아보기 위해 동물을 사용하여 시상하부를 손상시칸 후 일어나는 비정상적 체중 증가와 섭식 양상의 변화 등을 관찰하는 실험적 모형도 제안되었다.
1 갈증 뜨거운 태양 아래서 몇 시간 동안 테니스를 치고 나면 아마도 한 잔 의 냉수가 그렇게 요긴할 수 없을 것이다. 바로 이런 상황하에서는 물 에 대한 욕구가 너무도 강렬하여 곧바로 마실 수 있는 음료수가 없다 면 거리로 나가 청량음료라도 사마셔야 할 판이다. 만약 찬 음료가 없 다면 미지근한 물이라도 마시지 않을 수 없다. 요컨데 갈증이 증가하 면 갈증을 풀기 위해 어떤 행동을 취하려는 욕구가 생긴다. 난파선에 서 살아남은 생존자들이나 사막에서 길을 잃고 방황했던 생존자들의 경험에 따르면, 갈증이란 단순한 목마름이나 입안건조와 같은 불쾌감 정도에 그치는 것이 아니라 오직 물만을 생각하게 되고, 이 갇증이 채 워지지 않으면 불가피하게도 죽음에 칙면하는 심각한 것이라고 한다. 그러면 갈증을 일으키는 신호는 무엇인가? 건조해진 입이나 혀에서 발생하는 자극이 갈증을 일으키는 것일까? 아니면 체내의 수분이 일 정 수준 이하로 떨어질 때 이를 감지하는 신호가 갇증을 일으키는 것 인가? 이에 관한 의문은 실험실 연구를 자극하게 되었고 ., 이에 관한 몇 가지 설명도 제시되었다. 대부분의 동물은 스스로 체액 수준을 정확하게 조절한다. 정상적 조 건 아래서 체액의 변동 범위는 1% 정도에 불과하다. 많은 동물들의 물마시기는 일내주기성 (circ adia n rh ythm)을 보인다. 예컨대, 행스터는 주로 밤에만 물을 마시는데, 계속 어두운 곳에 두어도 물마시기의 일 내주기성을 보인다. 일내(ci rcad i an) 란 라틴어로 약 하루라는 말(약 : circ a, 하루 : d i eo) 에서 유래되었다. 70k g의 체중을 가진 보통 성인은 하루 약 2.7 리터의 수분을 섭취하는데, 이 가운데 약 절반은 직접 물 울 마셔서 취하고 나머지 반은 음식물을 통해 취한다. 한편 방출하는 수분의 반은 오중을 통해 방출되며, 나머지 반은 대변이나, 땀 그리고 호흡에 의해 방출된다. 어떤 동물은 전혀 물을 마시지 않는데, 이런 동물들은 필요한 수분을 전적으로 음식물을 통해 섭취한다. 바다에 사 는 포유류인 해마, 바다표범 그리고 사막에 사는 많은 동물들이 이에
포함된다. 1.1 일차적 물마시기와 이차적 물마시기 물마시기는 일차적 물마시기와 이차적 물마시기로 구분한다. 일차적 물마시기란 출혈이나 발한과 같은 수분 고갈에 대한 반응과 영분 섭취 에 대한 반응으로써 물을 마시는 것을 말한다. 혈장 속에 함유된 Na+ 와 c1-이 온의 비율은 약 0.9% 이다. 이 이온둘의 농도가 0.9% 이 상 인 용액 울 고장액 (hyp e rt on ic soluti on ) 이 라 하고, 0 . 9% 이 하인 용액 을 저 장액 (hy po to n ic soluti on ) 이 라 한다. 이차적 물마시기란 체액 균형과는 관련 없는 기타 조건들의 결과로 써 생긴다. 예컨대 뇌의 어떤 영역을 전기적으로 자극하여 일어나는 물마시기 반응은 바로 이 경우이다. 건조한 음식을 먹을 때 이 음식을 삼키기 위해 물을 마시는 수가 있고, 음식을 먹을 때 갈증과는 전혀 관계없이 습관적으로 물을 마시기도 한다. 극히 드물기는 하지만 어떤 정신장애자들은 병적으로 물을 많이 마신다. 이러한 여러 가지 형태의 이차적 물마시기를 과학적으로 설명하기란 대단히 어렵다. 대개의 연 구자들은 일차적 물마시기에 주된 관심을 두고 있다. 죽 어떤 조건이 물마시기를 야기하며, 또한 어떤 조건이 이룰 멈추도록 하는가에 주된 관심을 기울이고 있다. 대부분의 사람들은 입 안이 건조해지는 것이 갈증을 유발시키는 조 건이라고 생각한다. 이와 같은 생각은 구강의 건조가 갈증이라는 충동 과 물마시기란 행동을 야기하는 단서가 된다는 견해이다. 갈증에 관한 구강건조이론 (d ry mouth th eo ry)은 일찍 이 Walt er Cannon(1919) 에 의 해 공식 적 으로 제 안되 었는데 , Cannon 은 타액분비 가 감소되면 구강이 건조해지고 이어 갇증이 생겨 물마시기를 시작한 다고 하였다. 이 이론은 상식적인 추론에 기초한 것으로서 실험실의 연구가 시 작되 면서 곧 설득력을 상실하고 말았다. 그러나 Cannon 의 생각처럼 갈증을 야기하는 많은 조건들 가운데는 입 안과 목구멍의 타
액분비 감소가 포함된다는 것도 사실이다. 즉 수분이 결핍되거나 출혈 이 심한 경우 또는 건조하거나 소금기가 많은 음식을 섭취한 후에는 입이나 목에서 타액 분비가 감소된다 . 그러나 입 안이 건조하다고 해 서 반드시 갈증이 일어나는 것은 아니며, 어떤 경우에는 계속 입이나 목구멍을 적셔주어도 갈증이 그치지 않는다. 구강건조이론이 지니는 두 가지 약점을 보다 자세하게 알아보기로 하자. 구강건조이론에 따르면 타액 분비를 감소시키는 조건도 갈증을 유발 시킬 것으로 예견된다. Te it elbaum 과 E p s t e i n(1962) 은 쥐의 의측시상 하부를 손상시키면 타액 분비가 현저히 감소되며, 이와 동시에 마시는 물의 양도 감소된다고 하였다. 또한 Rolls 와 Rolls(1982) 는 선천적으로 타액선이 없는 어떤 한 젊은이의 사례를 소개했는데, 이 사람은 깨어 있는 동안 계속 조금씩 물을 마셨지만 정상인보다 마시는 양이 더 많 지는 않았다고 한다. 이러한 관찰의 결과에서 알 수 있는 것은 구강건 조가 갈증을 야기하는 하나의 요인은 될 수 있지만 마시는 물의 양을 설명하는 데는 적합하지 못하다는 것이다. 구강건조이론에 관한 또 다른 반론은 다양한 동물의 입이나 목구멍 에 물은 주입하지만 섭취한 수분이 체내로는 들어가지 못하도록 한 특 이한 실험방법을 사용한 연구에서 나왔다. 이 실험들에서는 식도나 소 화관에 인공적으로 누관(fi s t ula) 을 장치한다. 식도에 누관이 장치된 개 의 입과 목구멍으로 수분을 주입하면 입과 목구멍은 계속 젖어 있지만 수분이 직접 위에 미치지는 않는다. 왜냐하면 주입한 수분이 한쪽에 열려진 누관을 통해 바깥으로 흘러나오기 때문이다. 이 동물들은 입이 나 목구멍이 젖어 있는 데도 불구하고 엄청난 양의 물을 마신다. 이 동물은 식도 밀에 삽입한 또 다론 누관을 통해 위 속에 물을 주입해 주지 않으면 죽게 된다. 그 후 Bellows(1939) 와 Adol p h(l950) 는 초기 의 이 실험들을 좀더 정교하게 하여 수분의 양을 측정할 수 있도록 했 다. 즉, 끝이 열린 누관을 통해 개에게 수분을 박탈한 후 물마시기를 허용했더니 이 개들은 정상동물과 비슷하게 물을 마시고 멈추었다가는 수 분 이내에 다시 물마시기를 시작하였다. 이런 행동은 물을 개의 위
에 직접 넣어주지 않는 한 계속 지속되는데, 이와 유사한 결과가 원숭 이에서도 발견되었다 (Madd i son 등, 1980). 그러므로 비록 건조해전 목 구멍과 입으로부터 발생하는 감각들이 물을 마시게 하는 데는 영향을 미치지만 보다 높은 중추기제가 물을 마시는 양을 결정하는 것 같다. 2 갈증의 이중결핍이론 우리 신체 내의 체액은 세포 내 구획 (int r a celluar com p a rtin en t)에 있 다가 세 포 의 구획 (extr a celluar comp ar t m ent) 으로 넘 어 갈 수 있고, 경 우에 따라서는 그 반대 현상도 일어날 수 있다(그림 8-1). 세포 의 구 획 에 있는 체 액 은 세 포와 세 포 사이 에 있는 간질액 (int e r stit ial fluid ) 과 혈장 내에 있는 체액으로 나눌 수 있다. 전체 체액 중 67% 가 세포내 액이고 나머지 33% 가 세포의액인데, 세포내액은 체중의 40-50% 를 차 지하며 세포의액은 체중의 20-25% 를 차지한다. 대부분의 세포의액은 세포와 세포 사이의 공간, 즉 간질에 위치하는 간질액이다. 혈장 내의 체액은 매우 정교하게 감지된다. 혈액은 전체 체액의 7% 에 불과하지만 아주 미세한 혈액량의 변화라도 물마시기나 신체의 수 분보존기제에 변화를 야기할 수 있다. 갈증의 이중고갈이론 (double depl e ti on the ory of thi rs t)에서는 세포 내에서나 세포 의 구획에서의 수 분의 변화가 물마시기에 영향을 미치고, 이 영향이 곧 갈증의 욕구를 일으키게 된다고 주장한다 (E p s t e i n, Fit zs im o ns, & Rolls, 1973). 2.1 세포 탈수화 대부분의 체액은 세포 내 구획에 있고, 이곳에서의 체액 상실은 갈 증과 물마시기를 야기하며, 잇따라 수분을 보존하기 위한 생리적 반응 이 일어난다. 죽 항이뇨 호르몬 (an ti d i ure tic honnone: ADH) 이 뇌하수 체 후영에서 분비되며, 이 호르몬은 신장에 작용하여 오중을 농축함으
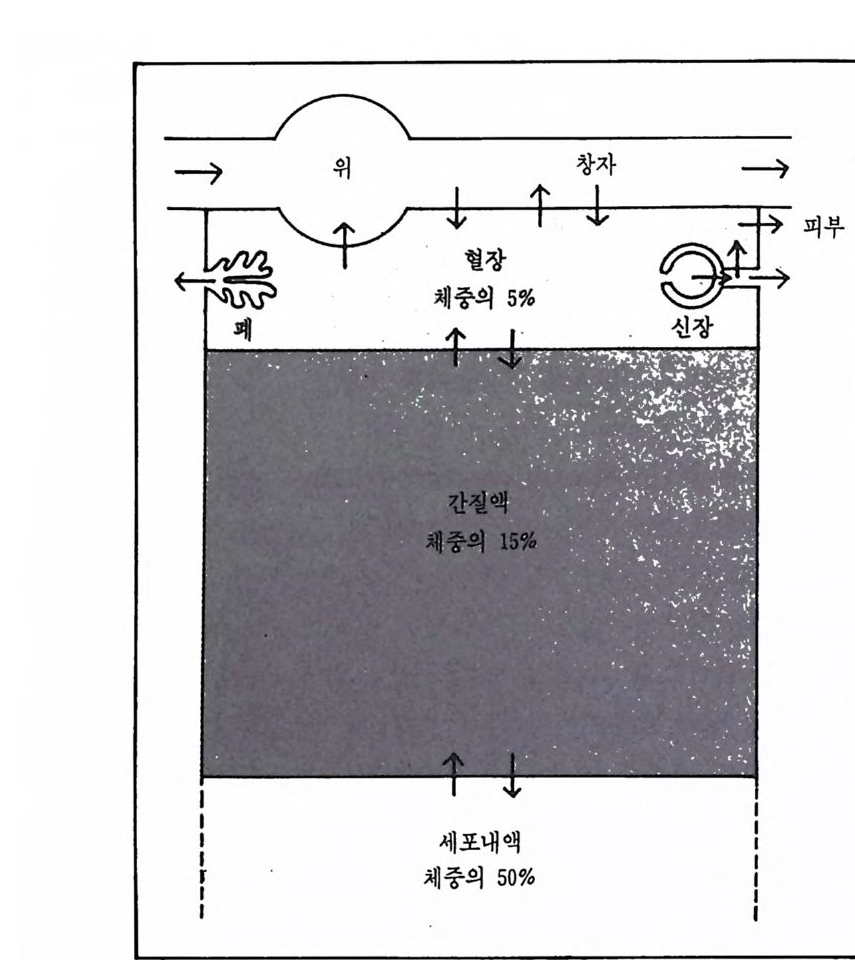 -국' 위창자 -
-국' 위창자 -
그림 8 기 주요 체액 구획 체액은 세포 안에 존재하거나(세포내액), 세포 바깥에 혈장이나 간질액으로 존재한다. 로써 수분의 체의 배출을 억제한다. 이와 같은 수분보전은 수분이 결 핍되는 동안 매우 효율적으로 사용되는 기제이다. 그러므로 갈증과 물 마시기는 세포의 탈수화에 따른 반응으로 일어나게 된다. 세포 탈수화 신호는 뇌 속에 있는 신경원에 영향을 미칠 것으로 추 측되어 왔지만 이에 대한 직접적 증거는 1953 년에 처음 제시되었다. Beng t Andersson(1953) 은 이미 물을 잔뜩 마신 염소의 전측 시상하부 (ante r i or hyp o th alamus) 에 소량의 식영수 고장액을 주입하였더니 엄청
난 양의 물을 마시게 된다는 사실을 발전하게 되었다. 죽, 고장액의 식영수 주입은 삼두현상을 통해 세포 내에서 세포 의로 수분방출을 야 기함으로 세포 탈수화를 일으킨다. 세포막을 자유롭게 통과할 수 없는 용해된 물질 분자가 한쪽 세포막에 농축되어 삼두압 (osmo tic pre ssure) 울 증가시키게 되면, 물 분자는 막을 통해 농도가 낮은 곳에서 농도가 높은 곳으로 이동하기 시작하여 막 내의의 압력이 같아질 때까지 계속 이동하게 되는데, 이러한 수분의 흐름을 삼두현상 (osmos i s) 이라 부른다. 세포 내로부터 수분이 방출되면 세포는 수축된다. 뇌 속에 있는 특 수세 포 —— 이 를 삼투압수용기 (osmorecep tor ) 라 부름 —— 의 용적 감소 현상이 뇌에 고장액을 주입했을 때 발생하는 물마시기 현상을 설명하 는 이론적 근거가 된다. 이와는 반대로 저장액은 수분을 세포 내로 진 입시켜 세포를 팽창시켜 중으로써 갈증을 낮추게 된다. 세포막을 재빨 리 자유롭게 통과할 수 있는 물질들은 삼두압에 영향을 주지 않으므로 갈증에 효과를 미치지 못한다. 요소가 바로 이러한 물질의 하나인데, 이것을 뇌 속에 주입해 주어도 물마시기에 아무런 영향을 미치지 못한 다. 이것은 세포 탈수화가 갈증과 물마시기를 야기한다는 견해들을 가 장 잘 지지해 주는 것이라 하겠다. Andersson 의 실험결과는 흰쥐를 피험동물로 사용한 Peck 과 Novin (1971) 의 연구결과와도 일치한다. 이 연구자들은 흰쥐의 시상하부에 자당과 같은 고장액을 주입하여 갈증과 물마시기를 야기시켰다. 이 실 험에 의하면 세포 탈수화 반응을 일으키는 세포는 오직 전측 시상하부 바로 앞쪽에 있는 의측시삭전(l a t eral pre op tic : LPO) 영역에서만 발견 되었다고 한다. LPO 에 있는 수용기들은 삼두압보다 세포 탈수화에 보 다 민감한 반응을 보이지만, 이 수용기들 역시 삼두수용기라 부른다• 앞서 언급한 것처럼 세포 탈수화는 갈증과 물마시기를 야기할 뿐만 아니라 뇌하수체 후영에서 항이뇨 호르몬을 방출시키기도 한다. ADH 는 신장에 작용하여 수분의 투과성을 증가시켜 혈류 속으로 수분을 재 홉수시킨다. 이런 과정에 의해 오중의 농도는 높아지고 수분이 체내에 머무는 시간도 증가한다. ADH 를 생산하고 방출하는 신경원은 LPO 에
있는 삼두수용기와는 다르다. 세포 탈수화는 포유류의 갈증 욕구를 유 발하는 가장 강력한 요인으로 간주되는데, 포유류의 종에 따라 다소의 차이는 있지만 쥐의 경우 64-69%, 개의 경우 72%, 레서스 원숭이의 경우 85% 정도가 세포 탈수화에 의한 물마시기 행동을 보여준다. 그 러나 사람의 경우는 아직 정확한 증거가 제시된 바 없다 (Rolls & Rolls, 1982) . 2.2 저혈량증 비록 세포 탈수화가 포유류의 물마시기 유발에 중요한 요인으로 간 주되고 있지만, 이 요인 의에도 갈증을 설명하는 다른 요인들이 있다. 출혈의 경우처럼 세포 탈수화가 없어도 세포의 구획으로부터 수분이 빠져나가 생 기는 저 혈량증 (h yp orvolem i a) 과 같은 것도 갈증을 유발시 키는 주요 자극이 될 수 있다. 그 밖에 발한도 세포 내 구획보다는 세 포 의 구획의 수분방출이 심한 경우인데, 이것 또한 갈증을 유발하는 자극이 된다. 저혈량증 갈증은 두 가지 신호의 개입에 의해 일어난다. 첫째 신호 는 신장에 대한 혈액의 공급이 감소될 때 발생된다. 죽 혈액량이 감소 되면 신장에서 레닌 (ren i n) 이란 효소가 분비된다. 레닌이 혈액 속에 들 어 가 순환되 면 중간 생 성 물인 안지오텐시노겐 (ang iot e n sin o g en ) 을 거 쳐 안지오텐신 II 라는 호르몬으로 바뀌게 된다. E p s t e i n 과 그의 동료들은 소량의 안지오텐신 II 를 이미 물을 충분히 마신 쥐의 뇌 속에 투입하 면 그 쥐가 즉각적으로 엄청나게 많은 물을 마신다는 사실을 발견하였 다. 1973 년 S i m p son 과 Rou tt enber g는 안지오덴신 II 의 뇌 수용기는 뇌궁 바로 아래에 위치하며 뇌실로 두사하는 작은 구조인 뇌궁하기관 (subfo rn ic al orga n : SFO) 이 라는 증거 를 제 시 하였다. 이러한 초기의 선도적 발견에 자극을 받아 SFO 가 과연 안지오텐신 II 의 주요 수용기인지 또는 안지오텐신 II 가 정상적인 물마시기에 중 요한 역할을 하는지를 알아보기 위한 많은 실험들이 시도되었다. 많은
연구자들은 SFO 가 안지오덴신 II 의 유일한 수용기라는 점에 대해서는 다소의 논란을 제기하였지만 위의 두 견해를 지지하는 결과를 보여주 었다. 안지오덴신 II 는 또한 부신피질에도 영향을 미쳐 알도스태론이 란 호르몬을 분비시키기도 하는데, 이 호르몬은 신장에 작용하여 Na+ 이온의 체내 유지를 증가시키는데 이렇게 증가된 Na+ 는 수분을 혈류 속으로 되돌려 놓는 역할을 하게 된다. 저 혈량증은 또 다른 종류의 신호도 야기 한다. 압력수용기 (baro- rec pt or) 라 부르는 압력에 민감한 수용기가 있는데, 이 수용기는 심장 벽이나 뇌에 혈액울 공급하는 주요 혈관인 경동맥 (caroti d a rt er i es) 의 혈관벽에서 발견된다. 이 수용기는 이 수용기가 위치하고 있는 혈관이 나 심장벽의 신장 정도에 매우 민감하다. 체액의 상실로 인해 혈액량 이 감소되면 이 압력수용기는 미주신경을 통해 뇌에 신호를 보내게 된 다. 이 신호는 물마시기를 야기하며 이와 동시에 ADH 분비도 촉진시 킨다. 그림 8-2 에는 갈증의 이중고갈이론을 요약해서 보여주고 있다.
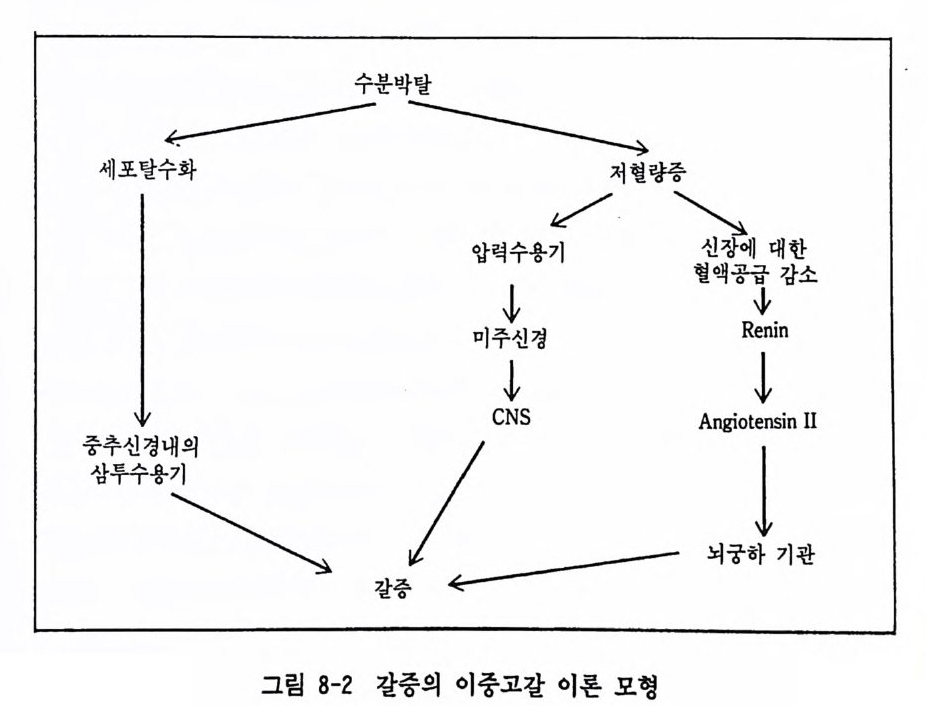 세포탈수~화 --수분박-탈 -~/저혈 呼\
세포탈수~화 --수분박-탈 -~/저혈 呼\
3 기아감 동물은 에너지를 저장하고, 생존에 필요한 물질을 취하기 위해 먹지 않으면 안 된다. 사람들이 왜 특정 음식을 선호하여 취하려 하며, 얼 마만큼의 양을 적절량으로 취하려 하는가에 관해 몇 가지 견해가 제시 되어 있다. 여기서는 주로 기아감, 음식물 섭취량 및 체중 조절에 미 치는 영향에 관해 알아보려 한다. 음식물의 섭취 조절에 관한 많은 연구둘은 주로 어떤 요인이 기아감 울 일으켜 섭식을 시작하게 하며, 또 어떤 요인이 포만감을 느끼게 하 여 섭식을 멈추게 하는가에 관해 집중되어 왔으며, 이 두 문제와 관련 하여 체중조절 문제가 또한 관심을 끌어왔다. 동물들은 무제한적으로 많은 음식물을 취할 수 있는 조건 아래 두어도 제한된 범위 내에서 자 신의 체중을 조절할 수 있지만, 사람의 경우 비만이란 문제가 생긴다. 앞으로 기아와 관련된 주제 속에서 항상성 (homeos t as i s) 이란 개념이 널 리 사용될 것인데, 이 개념은 유기체가 내적 안정성을 유지하려는 경 향성을 의미한다. 이 개념을 체중조절에 적용해 보면, 어떤 기본적인 조절점 (se tp o int)에 근접시켜 체중을 조절하기 위해 다양한 생리적 과 정이 개입된다는 것이다. 음식물을 쉽게 취할 수 없는 상황에서는 신 체 내에 에너지의 저장을 계속하면서도 체중 감소가 서서히 이루어지 도록 하기 위한 다양한 생리적 과정이 작용하게 된다. 비만중 환자들 이 정상 체중을 가전 사람들보다 더 많은 음식을 취하지 않는 데도 불 구하고 늘어난 체중을 계속 유지할 수 있는 것은 바로 이러한 항상성 기제 때문이다 (Bra itm an, Adlin , & Sta n to n , 1985). 많은 조건들이 섭식의 항상성을 방해할 수 있다. 동물들과 마찬가지 로 인간도 맛없는 음식보다는 맛있는 음식을 선호한다. 음식물의 질적 가치와는 관계없이 음식물의 맛이 음식물의 섭취와 체중 유지에 많은 영향을 준다. 특정 음식에 대한 맛의 평가는 피험자의 기아감 정도에 따라 많은 차이를 보여준다. Cabanac(1971) 은 음식의 맛이나 냄새의 평가가 배고픔 정도와 체중에 따라 달라진다는 것을 체계적으로 입증
하였다. 예컨대 설탕물을 마시고 난 후는 배고풀 때보다 단맛을 더 낮 게 평가하지만, 자신의 평소 체중을 10% 정도 낮춘 다이어트중인 사 람을 대상으로 이런 실험을 해보면 설탕물을 마시는 양이나 설탕물에 대한 미각의 평가가 별 영향을 받지 않는다. 4 신전대사의 양상 섭식 통제에 관한 논의를 계속하기 전에 음식물을 먹고 난 직후 소 화관 속에서 영 양소가 흡수되 는 시 기 (absorp tive pha se) 와 이 시 기 가 지난 후 섭식을 하지 않는 무섭식시기 (fas tin g pha se) 동안 일어나는 심 리적, 생리적 반응에 관해 살펴보기로 하자. 4.1 흡수시기 소화가 이루어진 후 혈액 속에 들어간 영양소는 에너지로 쓰이기도 하고 신체조직을 형성하는 기초물질로 이용되기도 하며, 지방질의 형 태로 저장되기도 한다. 신체의 주된 에너지 원천은 포도당으로서 이 포도당은 탄수화물과 단백질로부터 만들어지며, 이것은 에너지로 산화 되거나 저장 형태로 바뀔 수 있다. 과다한 포도당과 아미노산은 근육 이나 지방조직 속에서 당원(g l y co g en) 이란 형태로 저장된다. 또한 당 원은 지방조직 (adip o se tissu e) 속에서는 지방질로 바뀔 수도 있다. 음 식물에 포함된 지방질은 지방조직 속에 저장된다. 당뇨병 환자는 포도 당이나 아미노산을 에너지로 사용할 수 없어서 지방질을 연료로 사용 한다. 포도당이나 아미노산이 신체의 세포 속으로 들어가기 위해서는 인슐린(i nsul in)이란 호르몬이 있어야 한다. 이 호르몬은 혈류 속의 영 양소에 대한 반웅이나 음식물을 먹을 것이라는 기대감에 대한 반응으 로 췌 장 (pa ncreas) 에서 분비 된다.
4.2 무섭식시기 일단 음식을 먹어 영양소가 섭취되고 나면 신체는 음식을 먹지 않는 무섭식시기 동안 저장된 영양소를 사용하여 생존을 유지한다. 무섭식 시기 동안 장 속에 이용할 수 있는 영양소가 없으면 대부분의 신체세 포들은 저장 지방질에서 유래된 유리 지방산(fr ee fatty a ci ds) 과 포도당 울 에너지로 사용하게 된다. 뇌는 오직 포도당만 에너지원으로 사용할 뿐 유리 지방산을 에너지원으로 사용하지는 않는다. 장기간 지속되는 무섭식시기 동안에 뇌는 간의 유리 지방산에서 생성된 케톤 (ke t one) 을 에너지로 사용하기도 하지만 평소에는 거의 전적으로 포도당을 사용한 다. 뇌가 거의 전적으로 포도당을 에너지원으로 사용한다는 사실의 발 견은 섭식이론의 발달에 중요한 역할을 했다. 5 기아감과 포만감의 신호 음식과 관련되는 신호들이 섭식의 개시와 종결에 중요한 역할을 하 는 것으로 간주되어 왔다. 이런 신호들은 음식물의 섭취량을 조절할 때나 장기간에 걸쳐 체중을 조절할 때 중요한 역할을 한다. 앞서 갈증 이론에서처럼 섭식이론에서도 Walte r Cannon 은 위의 수축이 기아와 포만감의 신호가 될 것이란 극히 상식적인 이론을 제안하였다. Cannon 과 Washbum (1 912) 은 피 험 자에 게 측정 기 구가 붙어 있는 바 람 빠진 풍선을 삼키게 한 후 이어 이 풍선에 바람을 넣어 위가 수축 하는 상태를 기록할 수 있도록 하였다. 이 연구자들은 피험자가 주관 적으로 배고픔을 보고할 때마다 위의 움직임이 거의 동시에 나타난다 는 사실을 발견하게 되었다. 그러나 그 후의 여러 연구들에서는 이와 감은 위의 수축이 배고품이나 음식물 섭취량의 조절에 영향을 미치지 않는다는 증거들도 많이 제시되었다. 예컨대 대부분의 위를 제거한 위 암 환자들도 배고픔을 느낀다고 하며 (Wage n ste e n & Carlson, 1931), 위
와 뇌 사이를 연결하는 유일한 신경로인 미주신경 (vagu s nerve) 을 절단 시킨 경우에도 배고픔이 경감되지 않으며 음식물 조절에도 영향을 미 치지 않는다고 한다 (Thom p son , 1980). 이것은 곧 위의 수축운동이 없 어도 배고픔과 섭식행동이 일어날 수 있다는 것을 시사하는 것이다. 5.1 포도당의 신호 역할 포도당은 섭식의 통제에 중요한 것으로 주장되어 왔다. 앞서 언급한 것처럼 장기간의 무섭식기간을 제의하면 뇌세포가 사용하는 유일한 연 료는 포도당이다. 다론 신체 부위의 세포들은 유리 지방산과 케돈체 (keto n e body) 도 연료로 사용한다. 뇌 세 포는 충분한 양의 에 너 지를 저 장하지 못하기 때문에 정상적 기능을 유지하기 위해서는 포도당의 안 정적인 공급을 필요로 한다. 비록 일반 신체가 굶주린 상태에 있다 하 더라도 뇌는 계속 포도당을 공급받아야 한다. 뇌세포와 일반 신체세포 사이의 또 하나의 차이는 대부분의 뇌세포들은 혈액으로부터 포도당을 취할 때 인슐린을 필요로 하지 않는다는 접이다. 뇌의 활동에 포도당이 중요한 에너지원이 된다는 견해에 따라 기아 감의 주요 이 론인 포도당안정 이 론 (glu costa t ic the ory ) 이 Jea n Maye r (1953; 1955) 에 의해 제안되었다. 이 이론에 따르면 기아감과 포만감은 신체가 포도당을 이용하는 비율에 따른다는 것이다. 즉 음식을 먹고 난 직후는 적철한 양의 포도당이 신체의 세포 내에서 충분히 이용될 수 있어서 포도당 이용비율이 높아지고 포만감이 생기게 된다. 이와는 달리 음식을 취한 후 몇 시간 동안 음식을 취하지 않으면 대부분의 포 도당은 이용되어 버렸거나 저장되어 버렸기 때문에 포도당 이용률이 낮아지게 되어 그 결과로 배고품이 나타나게 된다. Ma y er 는 포도당 이용 정도를 탐지하는 유용한 방법으로 세포에 영 양소를 공급하는 동맥 속의 포도당 수준과 세포의 노폐물을 싣고 가는 정맥 속의 포도당 수준을 서로 비교하는 방법을 제안하였다. 이러한 측정방식을 A-V 비율 (a rt e ry -ve in ra ti o) 이라고 칭한다. 만약 정맥에
비해 더 높은 수준의 포도당이 동맥에 있다면(고 A-V 율), 포도당의 이용이 적절하게 이루어지고 있으며 포만감이 생기게 된다. 그러나 만 약 동맥과 정맥 사이의 포도당 수준이 매우 작은 차이밖에 보이지 않 는다면(저 A-V 율), 소량의 포도당만이 세포 내에서 이용되고 있기 때 문에 배고픔이 일어나게 된다. 몇몇 실험에 의하면 이러한 이론은 매 우 적절한 근거를 가진다고 한다 (Thom p son, 1980). 더구나 Ma y er 의 이론은 당뇨병 환자들의 경우에도 잘 들어맞는다. 비록 당뇨병의 발생기제는 복잡하지만, 대부분의 당뇨환자는 적절한 양의 인슐린을 생산하지 못한다. 인슐린은 신체세포가 포도당을 취하 기 위해 필요로 하는 호르몬이기 때문에 인슐린 치료를 받지 않은 당 뇨환자들은 비록 신체세포가 포도당을 필요로 하고 있지만 바정상적으 로 높은 혈당 (blood glu cose) 수준을 보인다. 그러므로 이 환자들의 동 맥 포도당 수준은 높지만 이 높은 포도당을 신체세포가 이용하지 못하 므로 A-V 비율은 매우 낮을 수밖에 없다. Ma y er 의 이론에 따르면 인슐린 처치를 받지 않은 당뇨환자는 동맥 속에 높은 수준의 포도당이 있음에도 불구하고 계속 배고픔을 느끼게 될 것으로 예측할 수 있는 데, 이런 예견은 사실이란 것이 증명되었다. 나아가 Ma y er 는 시상하부에 있는 특정 세포들이 포도당 수준을 감 지하여 기아감을 느끼게 하고 섭식행동을 일으키게 한다고 제안하였 다. 이처럼 기아감과 섭식행동이 시상하부에 의해 야기된다는 이 견해 는 뇌세포가 포도당을 섭취하기 위해 인슐린을 필요로 하지 않으므로 다른 신체 부위의 상황을 반영할 수 없다는 점 때문에 논쟁거리가 되 고 있다. 그러나 Ma y er 는 최근 시상하부에 있는 극소수 세포들이 포 도당을 섭취하기 위해 인슐린을 필요로 한다는 증거를 발견하였다. 이 세포들은 동맥의 포도당 수준에 영향을 받으며, 일반 신체세포와 극히 유사한 방식으로 인슐린의 출현이나 부재에 따라 이 세포들의 반응성 이 변경된다고 한다 (Oomura 등, 1979). 그러나 모든 증거둘이 Ma y er 의 이론을 전적으로 지지하는 것은 아 니다. 죽, 이 이론에 따르면 굶주린 동물에게 포도당을 두여하면 섭식
행동이 줄어들 것으로 예견되지만, 이 예견은 잘 맞지 않는다. 또한 포도당 수용기가 있는 부위로 짐작되는 시상하부에 포도당을 칙접 주 입시켜도 섭식량이 감소되지 않는다 (Nov i n , 1976). 한편 뇌에 대한 포 도당 공급률은 식사와 식사 사이에 별다른 변동을 보이지 않는다고 한 다. 이와 같이 혈액 내에서 일어나는 미미한 포도당 수준의 변화로써 섭식행동을 설명하기는 어려운 것 같다. 5.2 간의 포도당 수용기 포도당 수용기는 시상하부뿐만 아니라 간 (N iiji ma , 1969) 과 십이지장 에서도 발견되었다 (Nov i n, 1979). Mauric o Russek(1971) 은 간에 있는 포도당 수용기는 혈액 내 포도당 수준탐지와 뇌에 포만신호를 보내는 데 매우 중요한 역할을 한다고 강조하였다. 포도당을 주사하는 부위를 변화시켜 보면, 간 또는 십이지장 수용기 중 어느 것이 포도당에 대한 반응으로 섭식행동에 더 효과적인 영향을 미치는지를 알아볼 수 있다. 간은 주로 간-문맥 순환 (he p a tic-p o rt al circ ulati on ) 체 계 를 통해 혈액 을 공급받는다. 이 순환체 계 에 어 떤 물질 울 두입하면 이 물질이 직접 간 속으로 들어가게 된다. 한편 십이지장 에 장치한 미세관을 통해 포도당을 십이지장으로 직접 주입할 수도 있 다. 십이지장과 간의 포도당 수용기는 서로 다른 수준의 포도당에 반 응하며 , 또한 이 두 수용기는 동물의 굶주린 정도에 따라 섭식에 미치 는 영향이 상이하게 나타난다. 예컨대 토끼를 자유롭게 먹을 수 있도 록 해주는 경우 십이지장에 있는 수용기는 간의 수용기에 비해 포도당 주입에 대한 반응으로 섭식을 억제하는 데 더욱 효과적이다. 그러나 비록 동일한 포도당을 십이지장에 주입하더라도 토끼를 24 시간 동안 절식시킨 경우에는 섭식 억제가 찰 되지 않는다 (Nov i n, 1976). 그러므 로 포만감의 신호로서 포도당의 중요성은 동물의 배고품 정도에 크게 의존되는 것 같다.
5.3 콜레시스토키닌 : 포만물질인가? 최근 수년 동안 십이지장에서 분비되는 호르몬인 콜레시스토키닌 (cholecys t o k in i n : CCK) 이 란 호르몬이 사람을 포함하여 몇 몇 종의 동 물에서 기아감을 감소시키는 물질이라고 보고되고 있다. CCK 처치를 받은 흰쥐는 같은 기간 동안 음식물을 박탈당한 통제 군의 흰쥐에 비해 음식물의 섭취량이 줄어든다는 사실이 1970 년대 초 에 처음 보고되었다. 그 후 이러한 섭식 감소가 이 물질의 두여에 의 한 건강상의 장애 때문이 아니라는 증거들이 Jos eph An ti n 과 그의 동 료들 (1975) 에 의해 제시되었다. 이들은 CCK-33 을 복강에 주입하였더 니 주사받은 쥐는 정상통제 동물에 비해 음식 섭취량이 감소되었을 뿐 만 아니라 섭식 후 나타나는 몸치장 행동과 수면 행동과 같은 포만 행 동조차도 보여준다는 사실을 발견하였다. 후속 실험들에서 이들은 흰쥐뿐만 아니라 생쥐, 토끼, 양 및 원숭이 와 같은 다양한 종의 동물에서도 CCK 를 주입하면 섭식량이 감소된다 는 사실을 발견하였다. 그러나 사람의 경우 CCK 주입의 영향은 간단 하지 않다. 즉 CCK 를 투여받은 피험자는 음식을 더 많이 먹는다는 보고도 있고, 덜 먹는다는 보고도 있다. 그러나 소량의 CCK-8 을 혈 관주사를 통해 서서히 주입하면 섭식량이 감소되는데, 이런 결과는 정 상 체중을 가진 피험자 (K i ss il e ff 등, 1981) 나 비만 피험자 (P i -Sun y er 등, 1982) 모두에서 보고되었다. 만약 이 결과가 반복 증명되고 별다른 부작용이 관찰되지 않는다면, CCK-8( 이것은 소장에서 자연적으로 분비 되는 호르몬임)은 비만증을 치료하는 데 매우 효과적인 물질이 될 것으 로 기대된다. CCK 는 뇌에 직접 포만정보를 전달하는 것이 아니라 소장에 있는 포도당 수용기를 작동시키고, 이에 따른 신경충격이 미주신경을 거쳐 뇌에 전달된다. 이런 견해는 쥐의 미주신경을 철단해 버리면 CCK 의 포만효과가 나타나지 않는다는 실험 결과 (Sm ith 등, 1981 ; Lorenz & Goldman, 1982) 에 의해 증명된다.
6 과식증 19 세기 이후 뇌의 바닥 쪽에 있는 복내측 시상하부 (ven t romed i al hyp o th a lamus: VMH) 에 종양이 있거나 손상이 일어나면 체중조절에 장애가 생긴다는 보고가 있었다. 그러나 자연적으로 일어나는 뇌 손상 은 손상의 위치가 해부학적으로 명확한 경계를 보이지 않는 것이 일반 적이다. 1942 년 He t her i n gt on 과 Ranson 은 쥐의 시상하부 복내측 신경 핵을 실험적으로 손상시켰더니 섭식과 체중조절에 심한 변화가 일어남 울 관찰하였다. 이러한 VMH 증후에 관한 명확한 해석은 아칙도 불 가능하지만 이 사실은 여러 차례 거듭 확인되었다. 비록 이 증후의 구체적 결과들은 실험조건에 따라 다소 달라질 수 있지만, 기본적 양상에서는 전반적으로 일치한다. 만약 맛있는 음식물 울 제한 없이 먹을 수 있도록 해주면 VMH 손상 쥐는 엄청나게 많이 먹는다. VMH 가 손상되면 뇌 수술 후 마취에서 정상으로 회복되기 전부터 많이 먹기 시작한다. 이런 동물은 몇 주 동안 과식을 계속하여 수술 전 체중의 두 배까지 이른다. 이러한 과다한 음식물 섭취를 과식 증 (h yperp ha gia) 이라고 부르는데, 이 과식증은 뇌 수술 후 얼마 동안 지속되다가 점차 수술 전 체중보다 약간 증가된 수준에서 섭취량이 줄 어든다. VMH 증후는 시기에 따라 두 가지 단계로 구분된다. 첫째 시 기 를 역동기 (dy na m ic pha se) 라고 하는데 , 이 시 기 동안에는 과식을 하여 체중이 급속도로 늘어난다. 둘째 시 기를 안정기 (sta t ic p ha se) 라 하는데, 이때는 체중 증가나 음식물의 섭취비율이 수술 전 수준보다 약간 높아전 상태에서 안정을 이룬다. 급속 체중 증가는 주로 신체의 지방질에 기인된다. VMH 쥐의 지방은 전 체중의 74% 정도나 되어 정상쥐의 5-10% 에 비하면 실로 엄청난 양이다 (Grossman, 1967). 6.1 VMH 쥐는 배고품을 느끼지 않는가? VMH 쥐는 과식증에 더하여 매우 까다로운 식성도 보여준다.
Powley (19 77) 는 이 쥐 의 까다로운 식 성 을 〈 음식 물의 칼로리 와는 관계 없이 음식물의 감각적 속성에 지나치게 영향받는 섭식행동 특성 〉 이라 고 정의하였다. 고지방이나 맛있는 음식을 먹인 VMH 쥐는 표준적인 조사료를 먹인 VMH 쥐에 비해 더 많이 먹는 경향이 있지만, 먹이가 몹시 쓰거나 짜면 VMH 쥐는 정상쥐보다도 덜 먹는다. 그러므로 VMH 쥐가 정상쥐보다 배고픔을 더 느낀다고 결론짓기는 힘들다. 한 편 VMH 쥐의 까다로운 식성은 음식물의 촉감에서도 나타난다. 죽, 이 쥐들은 일반 사료를 가루로 만들어주면 정상보다 적게 먹는다. 따 라서 이러한 과식증은 단순한 배고품과는 관계가 없는 것 같다. 6.2 VMH 증후와 인간의 비만증 어떤 연구자들은 인간의 비만증과 동물의 VMH 증후 사이에 어떤 관련성이 있지 않을까 생각해 왔다. 인간의 비만증 환자 가운데 시상 하부의 실제적 손상을 가진 사례가 보고된 적도 있다. Hi rsch(1984) 는 어떤 유형의 비만증은 바이러스가 신경계에 침입하여 시상하부에 서서 히 영향을 미치기 때문에 그 여파로 인해 생긴다는 증거를 제시한 적 도 있다. VMH 쥐가 보여주는 까다로운 식성과 비만증 환자들이 음식물의 감각적 속성에 지나치게 영향을 받는다는 것은 서로 유사한 점이다. 이러한 까다로운 식성 때문에 맛있는 음식을 계속 먹을 수 있는 기회 가 주어지면 비만증 환자는 과식하게 되고 결국 체중이 늘어나게 될 것으로 예상된다. Sta n ley Schach t er(l971) 와 그의 동료들은 약간 비 만성을 보이는 피험자들 (15-20% 정도 과체중)을 대상으로 이러한 가설 울 검증하기 위해 몇 가지 실험을 수행하였다. 이들의 실험결과를 요 약하면, 일반적으로 비만피험자는 정상 체중을 가전 통제피험자에 비 해 밀크 셰이크와 같은 맛있는 음식물을 취할 수 있게 해주면 훨씬 많 이 먹는다. 그러나 밀크 셰이크에 쓴맛이 나는 키니네를 섞어주면 바 만피험자는 정상피험자에 비해 반밖에 먹지 않는다.
그러나 최근에 들어와 한때 Schach t er 의 동료였던 Jud it h Rod i n 은 이 주제와 관련된 여러 연구를 개관하고 비만증 집단뿐만 아니라 모든 사람들이 음식물의 의부적 자국조건에 민감하게 반응한다는 결론을 내 렸다 (Rod i n, 1976; 1978). 이런 견해는 Schach t er 의 가설을 부정하는 것이다. 그러나 일반적으로 약간의 비만성을 보이는 사람(1 5-25% 과체 중)은 정상 체중을 가진 피험자에 비해 음식물의 의적 자극에 보다 민 감한 반응성을 보여주지만, 이와는 대조적으로 심한 비만증을 보이는 사람은 (50% 이상의 과체중) 약한 비만증에 비해 음식물의 의적 자국에 대해 오히려 낮은 반응성을 보인다. 6.3 비만증과 인슐린의 관계 만약 어떤 사람이 정상인에 비해 음식물의 의적 자극에 더 높은 반 응성을 보인다면 이 높은 반응성과 증가된 섭식 욕구와는- 어떤 관련성 이 있지 않을까? Rod i n (l 976) 는 음식물의 의 적 자극에 더 높은 반응성 을 보이는 사람 들은 석쇠 위에서 구워지고 있는 스데이크를 보거나 냄새를 맡고 있을 때 정상피험자보다 더 많은 인슐린을 분비한다는 사실을 발견하였다. 또한 자극에 민감한 피험자들은 체중에 관계없이 다양한 종류의 케이 크의 맛을 볼 때도 보다 많은 인슐린을 분비한다는 점이 발견되었다. 비만증 환자가 더 많은 인슐린을 분비할 것이라는 견해를 지지하는 연 구가 J ohnson 과 W il dman(l983) 에 의해 제시되었다. 이들은 4 명의 비 만피험자와 6 명의 정상체중 피험자의 인슐린 분비 수준을 비교하였더 니 비만피험자는 실제 음식을 제시했을 때나 이상적인 음식물을 생각 하고 있을 때나 모두 더 많은 인슐린을 분비하였다. 홍미로운 것은 비 만피험자는 실제 음식제시 때보다 이상적인 음식을 상상할 때 더 높은 인슐린 분비반응을 보였다고 한다. VMH 쥐에서도 음식물과 관련 있는 의적 자극에 대한 반응으로 보 다 많은 양의 인슐린 분비가 일어날까? VMH 쥐는 정상쥐보다 음식
물 자국에 대한 반응으로 보다 많은 양의 인슐린을 분비한다는 사실이 발견되었는데, 이것은 곧 이 쥐의 과식 및 체중 증가가 인슐린 분비 증가와 직접 관련될 수 있음을 시사하는 것이다 (Thom p son, 1980). 인슐린은 오랜 동안 기아감과 포만감을 야기하는 신호역할을 할 것 으로 생각되어 왔지만 인슐린이 분비되는 상황은 매우 복잡하기 때문 에 이의 역할을 명확하게 알지 못했다. Powle y(l 977) 는 일련의 신체 반웅들이 음식물의 소화와 관련 있다고 강조했는데, 이 반응들은 음식 물이 미처 위에 닿기 전부터 시작된다. Powle y는 이 반응을 두부반응 (ceph a lic r espo n se) 이 라고 불렀는데 이 는 눈, 코, 입 등이 위 치 하는 머 리 부위의 수용기 역할을 강조한 것으로서 타액이나 소화액 또는 인슐 린 분비를 증가시키는 소화반응을 선도하는 반응이다. 소화반응에 덧 붙여 맛없는 음식이나 역겨운 물질을 거부하는 반사적인 두부반응도 있다. Powle y는 쥐의 VMH 손상의 일차적 영향은 긍정적이거나 부정 적 두부 소화반응 양자를 증가시켜 음식물의 감각적 요소에 과장된 반 응을 일으킴으로써 입맛이 까다롭게 된다고 하였다. 이와 같은 증가된 두부반응의 결과로 VMH 쥐는 맛있는 음식은 많이 먹게 되고 맛없는 음식은 덜 먹게 된다. 인슐린의 또 다른 기능은 ·포 도당이나 기타 영양소를 신체세포 특히 지방세포 속으로 많이 유입시키는 것이다. 정상 동물에게 인슐린을 두 여하면 섭식이 증가되며, 인슐린 주사를 계속하면 비만 현상이 생기게 된다. 그러므로 만약 VMH 쥐에서 인슐린 분비가 증가된다면 보다 많은 포도당이 지방세포로 진입하게 되므로 맛있는 음식을 제공하면 비만이 나타날 수 있다. 그러나 만약 맛없는 음식물을 제공하면 비만 이 생기지 않는데, 이런 현상은 일반적으로 관찰되는 보편적 결과들이 다. 이런 견해로 보면 VMH 증후의 3 대 증후一―식욕의 까다로움, 과식, 비만一―는 두부소화반응의 증가와 특히 인슐린 분비와 관련될 수 있다.
7 의측 시상하부 증후군 VMH 손상 쥐가 음식을 많이 먹는다는 사실이 보고된 지 10 여 년 이 지 난 후 Anand 와 Brobeck (1 951) 은 의 측 시 상하부를 손상시 킨 쥐 가 VMH 손상 쥐의 섭식 행동과는 정반대의 행동을 보인다는 사실을 보고하였다. 이 들은 의 측 시 상하부 (lat e r al hyp o th a lamus : LH) 가 손상 된 쥐는 음식물을 먹지도 않고 물을 마시지도 않지만, 수술 후 1 주일 동안만 적절하게 돌봐주면 다소간의 회복이 가능하다고 했다. 후속된 연구들에서는 다양한 종류의 행동, 예컨대 이 동물들은 각종 동기부여 자국들 ___ 음식물, 물, 성적 배우자, 위험자극一一에 대한 반응성이 극적으로 사라진다는 점을 강조하게 되었다. 그렇다고 해서 이 동물들 이 의식 없이 혼미 상태에 빠져 있는 것도 아니고, 움직일 수 없는 것 도 아니며 오칙 의적 자극에 대한 반응성만 낮아진다는 것이다. 이 동물의 회복과정에 많은 관심이 집중되었다. 사실 이 동물의 경 우 회복이란 말은 정확하지 않다. 왜냐하면 이 동물들은 체액을 조절 하는 능력이 새롭게 이루어지는 것도 아니고, 음식물에 관한 반응성이 회복되는 것도 아니기 때문이다. Alan E p s t e i n (1 971) 은, 이 동물은 4 단계를 거쳐 정상 행동으로 회복된다고 했다. LH 증후의 첫 단계에서는, 이 동물에게 인슐린을 두여함으로써 포 도당을 결핍시켜 먹이를 먹게 하거나 체중이 많이 감소된 상황에서 맛 있는 음식만을 먹을 수 있도록 해주어도 전적으로 음식먹기를 거부한 다. 이 동물의 물마시기는 건조한 음식을 먹을 때에만 나타난다. 죽 건조한 음식을 삼키기에 용이하도록 음식을 먹고 난 직후에만 소량의 물을 취한다. LH 쥐의 이러한 물마시기 특칭은 제 1 단계로부터 마지막 단계인 4 단계까지 계속된다. 그러나 정상쥐는 타액 분비량이 충분하기 때문에 음식을 먹을 때 물을 마시지 않으며 먹기 전이나 먹기가 끝난 후에만 물을 마신다. 그러나 LH 쥐는 정상 수준의 타액 분비가 이루어지지 않기 때문에 건조한 음식을 먹는 도중에 물을 마시게 된다. 만약 이 동물에게 액체
성 음식을 주면 물마시기가 나타나지 않는다. 실험실용의 표준사료를 주면 음식물을 한 번 씹을 때마다 물 한 모금씩을 마시게 되어 결국 하루 400-600 회 정도나 마시게 된다. 만약 LH 쥐의 타액선을 제거해 침을 분비할 수 없게 하면 섭식중에 더 자주 물을 마시게 된다. 그러 나 LH 쥐는 자기 자신의 내부체액 요구에 따라 물을 마시는 양을 조 절하지 못한다. 죽 물을 결핍시킨 후 물을 마시게 해주어도 많이 마시 지 않는다. LH 쥐는 근본적으로 물을 마시고 싶어하는 욕구가 없어진 것 같으며, 오직 음식먹기의 보조 수단으로서 물을 마시는 것 같다. LH 증후는 뇌간에서 출발하여 LH 를 거쳐 전뇌로 진행하는 축색들 의 손상에 기인되는 것으로 생각한다. LH 증후군은 노르에피네프린과 도파민성 신경원을 선택적으로 파괴하는 신경독소를 주사하여도 나타 난다. 이 신경로는 뇌 간의 혹질 (substa n ti a nig ra) 에서 기 저 핵으로 두사 되는 흑질선조체속 (n igr os tri a t al bundle) 이 주된 신경로이다 (S t r ic ker & Zig mo nd, 1976) . LH 에 있는 신경 원들의 기능에 관해서 는 아직도 잘 모르지만, LH 증후군은 LH 를 지나가는 축색만을 절단하고 그 신경세 포체를 그대로 두어도 나타난다. 한편 Grossman 과 그의 동료들 (1978) 에 의하면 LH 에 키나산을 투여하여 화학적 손상을 일으킨 경우에도 음식먹기와 물마시기를 거부한다고 한다. 이런 화학적 손상은 신경섭 유에는 손상을 주지 않고 세포체만 선택적으로 파괴하는 방법이다. 따 라서 LH 증후군을 야기하는 데 관여하는 정확한 신경 실체가 무엇인 지는 아직 잘 모르는 셈이다. 8 체중의 조절과 조절점 비록 비만이란 문제는 인간뿐만 아니라 모든 포유류에서 나타나는 것이지만, 일반적으로 성인이 된 후는 매우 좁은 범위 내에서 자신의 체중을 계속 유지해 간다. 그러나 이러한 체중조철은 질병에 걸리거나 비만화가 일어나게 되면 잘 지켜지지 않는다. 대부분의 유기체가 의식
적 노력 없이 풍요로운 먹을 거리에 노출되면서도 특정 체중 수준을 잘 유지한다는 것은 주목할 만한 일이다. 체중의 변동은 주로 신체 지 방질의 가감에 따르기 때문에 많은 연구가들은 체내 지방질의 상태를 알려주는 어떤 종류의 신호들이 섭식량과 체중 수준의 장기 조절에 관 여할 것이라고 시사했다. 이러한 가설은 Kennedy (1 950 ; 1966) 에 의해 제시되었는데, 그는 신체 내의 지방 수준은 뇌 속에 있을 것으로 추측 되는 지방항정수용기 (lip os t a t s) 에 의해 감지될 것이라고 하였다. 그는 지방 축적이 어떤 점에 이르면 음식섭취량이 감소되며, 반대로 지방 축적이 감소되면 지방항정수용기가 배고품을 증가시켜 정상체중 수준 으로 회복될 때까지 음식먹기를 지속할 것이라고 시사했다. 체중조절에 관한 조절점이론 (se tp o i n t t heo ry)은 많은 비판을 받아 왔 다. W i r t scha ft er 와 Dav i s (1 977) 는 조절점이론의 몇 가지 약점을 지적 하고 그 당시까지 보고된 자료를 중심으로 하나의 대안적 모형을 제시 했다. 이 모형의 가치는 음식물 맛의 변화에 따른 음식물 섭취량의 변 화를 고려한 것으로서 이것은 대부분의 조절점이론에서 제기된 주요 문제점의 하나였다. 그러면 어떻게 뇌가 신체 내의 지방량을 알아차릴까? 지방은 혈액 울 통해 순환하지 않기 때문에 지방의 수준을 반영해 주는 어떤 종류 의 물질에 의해 탐지될 것이다. 비록 포도당이 뇌의 주 에너지이지만 신체도 미세한 양의 포도당을 에너지로 저장한다. 대부분의 신체 저장 에너지는 지방으로 저장되어 다른 에너지원이 이용될 수 없을 때 이를 사용하게 된다. 지방은 트리글리세라이드(trig l y cer i des) 의 형태로 신체의 여러 부위 에 저장되며, 유리 지방산과 글리세린(g l y cer i n) 으로 혈류에 방출된다. 이 물질들은 에피네프린과 같은 호르몬의 작용에 의해 저장 지방으로 부터 방출되고, 간에서 글리세린은 포도당으로 바뀌게 되며 이 포도당 은 무섭식 동안 뇌의 에너지로 사용된다. 그러므로 최소한 혈류에 방 출된 두 가지 물질, 죽 유리 지방산과 글리세린이 체내의 저장 지방 상태를 반영하는 물질로 간주될 수 있다.
무섭식, 운동, 스트레스 또는 지나친 추위 등에 노출될 때 혈류 속 의 유리 지방산 수준은 높아진다 (Thom p son, 1980). 유리 지방산이 신 호로 사용될 수 있다는 생각은 Oomura 와 그의 동료들(1 979) 에 의해 제안된 것인데, 이들은 유리 지방산 수준에 반응하는 신경원이 의측 또는 내측 시상하부에 존재한다는 사실을 발견하였다. 그러나 지방 수 준을 감지하기 위해 뇌가 어떤 종류의 신호를 이용하는가에 대해서는 아직 잘 모른다. 아직은 어떤 단일 이론으로 장기간에 걸친 체중조절 을 충분히 설명할 수 없다. 이에 대한 설명적 가설로서 Fr i edman 과 Str ick er (1976) 는 중다요인 가설 (multif actor hyp o th e sis) 을 제 안했는데 , 이들에 의하면 뇌 속의 포도당 수준은 다른 신체 부위에서처럼 심하게 변동하지 않기 때문에 포도당이 기아감이나 포만감을 동제하는 단일한 신호라는 견해를 부정하고, 기아감이란 포도당 , 유리 지방산 또는 아 미노산과 갇은 모든 이용 가능한 에너지가 감소하는 데 대한 반응일 것으로 추리하였다.
제 9 장 성 행동 호르몬은 신체 내의 몇몇 내분비조직으로부터 혈액으로 분비되는 생 화학적 물질이다. 호르몬은 두 가지 방식으로 행동에 영향을 미친다. 첫째는 중추신경계를 포함하여 태아의 발달에 영향을 미친다. 특히 태 아의 고환에서 분비되는 호르몬은 태아가 남성으로 발달되도록 하며, 이 호르몬이 분비되지 않으면 비록 태아가 유전적으로는 남성이라 하 더라도 여성으로 발달하게 된다. 이처럼 태아기 동안 비정상적인 호르 몬 분비는 유전적으로 결정된 성의 특칭을 바꿀 수 있다. 태아기 호르 몬은 인간의 경우 성 호르몬 분비와 그 밖에 성과 관련된 의식 발달에 도 미묘한 역할을 담당한다. 호르몬이 행동에 영향을 미치는 두번째 방식은 사춘기나 · 성인기 때 목표 세포와의 상호작용을 통해서 이루어진다. 목표 세포는 어떤 특정 호르몬에 대해서만 반응을 나타낼 수 있도록 하는 특수 수용기를 갖고 있는 세포이다. 대부분 동물들의 성 행동은 호르몬의 영향을 받는다. 인간의 성욕과 성 행동에 미치는 호르몬의 중요성에 관해서는 많은 연 구가 이루어졌다. 그러나 이 문제에 관해 결론을 내리는 것은 쉬운 일 이 아니며, 아직 인간의 성 행동에 미치는 호르몬의 영향은 확실하게 규명되지 않았다.
1 호르몬 고성능의 컴퓨터가 개발되면서 흔히 인간의 뇌를 컴퓨터와 비교하는 경우가 많다. 그러나 최소한 한 가지 중요한 측면에서 이와 감은 바교 는 오류를 범하고 있다. 즉 전적으로 전기적 신호에 의해 작동되는 컵 퓨터의 마이크로칩과는 달리 인간 뇌의 신경원은 혈류를 통해 끊임없 이 공급되는 다양한 화학물질들에 의해 작동된다는 점이다. 이러한 화 학물질들 가운데 가장 중요한 물질 중의 하나가 호르몬이다. 호르몬 (hormones) 이 란 〈 혈액 을 통해 특정 목표 조직 에 이 르러 이 목표 조직
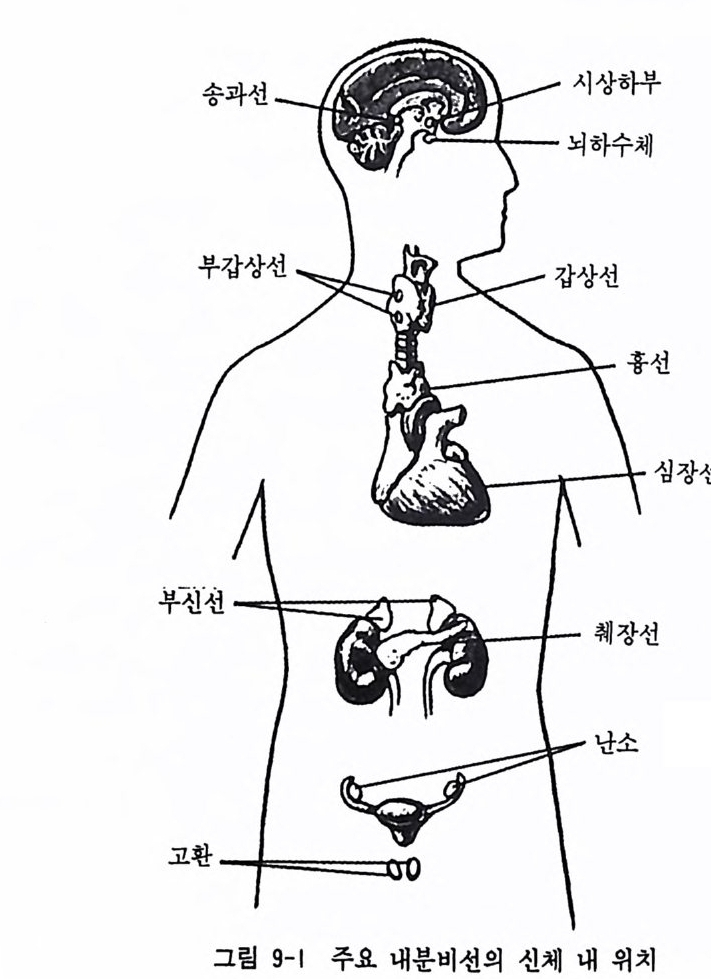 시상하부
시상하부
세포의 활동 변화를 야기하는 화학적 전달자 〉 로 정의된다 (Norr i s , 1980). 호르몬은 신경원을 포함하여 근본적으로 모든 유형의 세포 활 동에 영향을 미찬다. 가장 중요한 호르몬 분비선의 하나는 뇌하수체 전영 (ante r io r pitui t ar y g land) 인데, 이곳은 뇌의 시상하부에 있는 특수 신경원에 의해 분비된 호르몬에 의해 지배받는다. 이처럼 시상하부와 같은 뇌가 뇌하수체 전영을 지배한다는 사실은 호르몬이 행동에 미치 는 영향을 이해하는 데 매우 중요한 의미를 갖는다. 표 9 기 중요 내분비 조직과 그곳에서 분비되는 대표적인 호르몬 뇌하수체 전영 부신피질 ACTH : adrenocort ico tr o p ic honnone cort iso l P-e nd orph i n cortic o ste ro ne GH : grow th hormone aldoste ro ne FLSHH : :I uftoe lil n ici z l e in sg t i hm ou rmlaotin neg hormone a고n환dro g en TSH :p trho ylarocit di n s tim u lati ng h ormone estr o g en 난소 뇌하수 체 후영 estr o g en vasopr e ssin ( anti diu re ti c h ormone : ADH ) pro g es ti ns oxy toc in androg en 갑상선 thyro xin e 췌장 trii o d oth yron in e i nsul i n (P - 세포에서 분비) thyroc alcit on in g luca g on(a - 세포에서 분비) 부감상선 소장 PTH : par ath y r oi d h onnone cholecys t ok in in 부신수질 송과선 epin e p hr in e (adrenalin e ) melato n i n norep ine p hr in e (noradrenalin e ) 심장선 ANF : atri al natr iur eti c f ac to r
신체에 있는 여러 조직에서 호르몬이 분비되는데, 이런 조직들을 총 칭하여 내분비 계통이라고 한다. 그림 9-1 에는 호르몬을 분비하는 조 직 들, 죽 뇌 하수체 {pitui t ar y ) , 갑상선 (thyro id gla nd) , 부신선 (adrenal gla nd) , 췌 장선 {pa ncreas) , 십 장선 (heart ) , 성 선 (go nads) , 송과선 (pine al gla nd) 그리고 시상하부 신경원들이 나타나 있다. 신경 계통의 한 구 조인 시상하부가 내분비 계동의 일부분이 된다는 사실은 이 두 계통이 서로 밀접하게 관련되어 있다는 것을 의미한다. 뇌하수체가 시상하부 에 연결되어 있다는 것은 곧 뇌가 여러 내분비 계통에 영향을 줄 수 있는 최종 공통로 구실을 한다는 것을 의미한다. 표 9-1 에는 중추 내 분비 계통과 여기서 분비되는 대표적인 호르몬의 종류를 열거해 놓았 다. 1.1 목표 조직 신체 내에 있는 대부분의 세포들은 몇몇 특정 호르몬에만 반응한다. 특정 호르몬에 반응하는 세포가 목표 세포가 되기 위해서는 그 호르몬 에 대한 수용기를 가지고 있어야 한다. 이 수용기들은 주로 세포질이 나 세포막에 분포되어 있는 특수 단백질로 구성되는데, 어떤 세포들은 여러 가지 호르몬에 대한 수용기를 가지며, 세포질 수용기와 세포막 수용기를 동시에 가지는 경우도 있다. 1.2 호르몬의 계열 모든 호르몬은 스데로이드, 폴리펩티드 그리고 기타 아미노산이 변성 된 화학적 계열 가운데 어느 하나에 속한다. 스테로이드 호르몬은 특 칭적인 4 고리 구조(fo ur-r i n g s truct ure) 를 갖는데 이 호르몬은 성선과 부신피질에서 분비된다. 스데로이드 호르몬 계열 가운데 특히 전립선 과 남성 성기의 성장과 같은 남성화 기능에 주로 영향을 미치는 호르 몬을 안드로겐이라 하며, 질이나 난소와 같은 여성 성기관의 발달을
자국하여 여성화를 촉진하는 호르몬을 에스트로겐이라 부른다. 에스트 로겐은 자궁 내 태아의 착상을 준비하는 호르몬이고, 프로게스틴은 자 궁벽의 성장을 자극하는 호르몬이다. 부신피질에서 분비되는 스테로이 드 호르몬을 코르티코스데로이드 (cort ico ste ro id ) 라 부르는데 , 모든 스 데로이드는 콜레스테롤에서 합성된다. 여성에서만 유일하게 분비되는 프로게스틴울 제외하고 남성, 여성 모두 앞서 언급된 모든 호르몬들을 규칙적으로 분비한다. 그러나 여성은 안드로겐보다는 에스트로겐을, 남성은 에스트로겐보다는 안드로겐을 더 많이 분비한다. 1.3 스테로이드 수용기 스데로이드는 혈액 내 담체단백질 (carr i er p ro t e i n) 에 부착되어 있지 만 이와 쉽게 분리되어 수용성 지질이 되어 세포막의 지질 부분을 건 너 세포 내로 확산해 들어갈 수 있다. 목표 세포의 세포질 내에 있는 스데로이드 호르몬 수용기가 이 호르몬을 붙잡게 되는데 목표 세포가 아닌 세포에서는 수용기가 발견되지 않는다. 일단 호르몬이 세포질 내의 수용기와 결합하게 되면 이 호르몬-수용 기 복합체는 세포핵 내로 이동하여, 세포핵 내의 DNA 수용체 부위에 결합된다. 여기에서 호르몬-수용기 복합체는 특수한 유전자를 작용시 켜 특정 세포의 생성합성률을 변경시키게 된다. 이러한 과정이 진행되 어 최대의 합성비율에 이르는 데는 몇 시간이 걸린다. 이런 까닭으로 스테로이드 호르몬이 최대 효과를 나타내는 데는 시간이 걸린다• 스테 로이드는 신경원에 직접적인 영향을 미치며 또한 많은 스테로이드 호 르몬이 산경원의 발사율을 변경시킬 수도 있다. 1.4 폴리펩티드 호르몬 대부분의 포유류 호르몬은 두 개 이상의 아미노산으로 구성되는 폴 리펩티드이다. 옥시토신과 같은 작은 폴리펩티드는 아미노산이 선형
고리로 이루어지지만, 성장 호르몬과 같은 큰 풀리펩티드는 대단히 복 잡한 분자구조로 되어 있다. 100 개 이상의 아미노산을 갖는 폴리펩티 드를 단백질이라 하는데, 폴리펩티드는 크기가 크고 지질에 용해되지 않기 때문에 세포 내로 쉽게 들어가지 못하는 대신 세포막에 있는 특 수 수용기와 결합한다. 이러한 결합과정은 1972 년 Su t herland 에 의해 처음 발표되었는데, 그는 폴리펩티드 호르몬이 세포에 작용하는 방식 을 제 2 전령 계통 (second messeng er s y s t em) 이라고 불렀다. 이 가설에 따르면 폴리펩티드는 세포막의 수용기와 결합하여 세포 내의 cAMP (cy cl ic A MP) 수준을 증가시 킨다. 이 러 한 세 포 내 cAMP 수준은
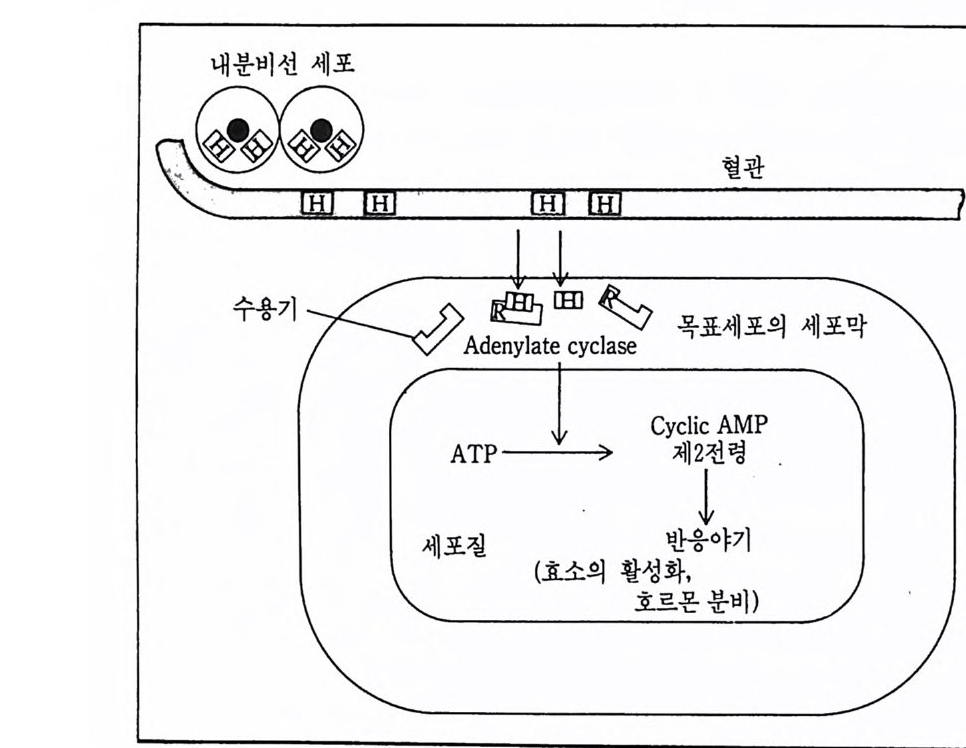 三내분비선 세포
三내분비선 세포
그립 9-2 호르몬 작용에서 제 2 전령의 작용기재 대부분의 폴리펩티드 호르몬이 세포에 영향을 끼치는 모습을 도시화한 것이 다. 호르몬 (H) 이 세포막에 있는 수용기와 결합하면, adeny la te c y clase 의 활 성을 야기하여, 세포질 내의 cyc l ic A MP( 제 2 전령)의 양울 증가시키게 된다 . 증가된 cAMP 는 여러 가지 반응을 야기하게 된다.
adeny la te c y clase 라 부르는 세포막 내에 있는 효소의 작용에 따라 달 라진다(그립 9-2). 그러므로 다양한 세포반응을 야기할 수 있는 cAMP 는 제 2 전령이며 호르몬 자체는 제 1 전령이다. cGMP 와 같은 또 다른 종류의 제 2 전령도 존재하지만 cAMP 와 cGMP 는 모두 하나의 세포 내 에 공존한다. 1.5 기타 호르몬 기타 계열에 속하는 호르몬으로는 티록신(thy rox i n) 과 트리요오드티 로닌(t r ii odo t h y ron i ne) 과 같은 갑상선 호르몬과 송과선에서 분비되는 멜라토닌 (mela t on i n) 그리고 부신수질에서 분비되는 에피네프린, 노르 에피네프린과 같은 것이 있다. 갑상선 호르몬도 스데로이드와 같이 지 질에 용해되어 막을 통해 세포 내로 들어가 세포핵 내의 수용기와 결 합된다. 멜라토닌은 변형된 아미노산으로 구성되어 있으나 이의 수용 기는 잘 알려져 있지 않다. 에피네프린과 노르에피네프린 역시 아미노 산이 변형된 것으로 제 2 전령을 이용하여 목표 세포에 영향을 미친다. 1.6 시상하부 호르몬 인간의 경우, 뇌의 시상하부 바로 아래 가늘게 돌출되어 나온 완두 콩만한 크기 의 조직 이 있는데 , 이 를 뇌 하수체 (pituitary gla nd) 라 한다 (그립 9-3) . 뇌 하수체 란 이름은 르네 상스 시 대 의 해부학자 Andreas Vesa li us 에 의해 처음 명명된 것이다. 그는 뇌하수체는 뇌에서 분비되 는 점액들을 모아 코나 목의 점액으로 배출해 내는 기능을 하는 것으 로 생각했다. 인간의 경우 뇌하수체는 전영 (ante r io r lobe) 과 후영 (po ste r io r lobe) 으로 나누어진다. 그러나 많은 종의 동물들에서는 전영 과 후영 의에 중간염 (int e n nedia te l obe) 이란 것도 있다. 전영과 후영은 서로 다른 배아기 조직에서 생겨나며, 상이한 조절기제에 의해 지배된 다. 특히 후영은 시상하부의 실제적인 연장조직인데 이곳에서 분비되
는 옥시토신과 바소프레신 (vaso p ress i n) 은 시상하부 신경원에 의해 만 들어진다. 이러한 신경분비 (neurosecreto r y ) 신경원의 세포체븐 시상하 부에 있고, 이의 축색이 뇌하수체 후영으로 두사되며, 여기에서 분비 되는 호르몬이 혈액으로 방출된다. 얼마 전까지만 하더라도 뇌 조직과는 별도안 뇌하수체 전영이 어떻
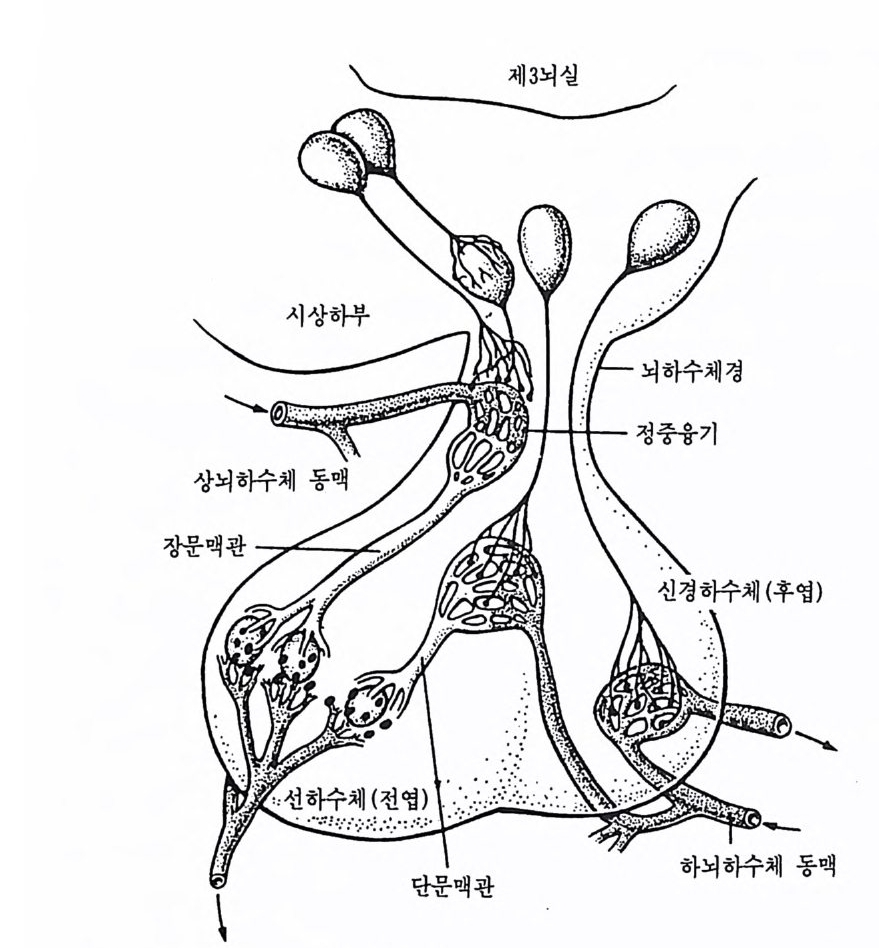 상뇌
상뇌
그림 9-3 시상하부-뇌하수체 연결도 뇌하수체 후영 호르몬은 시상하부 신경핵에 있는 신경분비세포들에 의해 만들 어진다. 이 분비신경원의 축색은 뇌하수체경{pit u it ar y s t alk) 으로 하행하여, 뇌하수체 후영의 정맥 부위까지 이어진다. 뇌하수체 전영은 뇌하수체 문맥계 를 통하여 시상하부 호르몬을 수용한다.
\� 9-2 ����XՀ� 8�t����� t��� ��Q�
XՔ� ̱X�´� �� 8�t��� ����XՀ� 8t���� X1�̱�´ �� 8�t��G�R nH : gonaroopiomdotropin r elaeig snohronme CRH : o crticotropelanocotirnG HGH Prolacint roPin releains gorhmone GHRH : gotr w hhormone lac releasingho mrone GHRIH : growt ohrhoemrnel aee-stininhibiting homnreo(o astsmtoatin) PR H: roplctain reeslnagi hormone TSH � ���p PRIH :pr olatic nrelease-ihinbiigt nhroonme TRH :������ � ��t thyrotoir p neeralisn hgormoneFH LSH, ACTH, p�� �� L��� �����. t� ��@� 0�D�0� ٳH� �� �̥���� ̹䴴�� p���<�\���0� �����. t� p���@� ������<�\��� 0� t�ٳ\� �� �\� 6��� �@�� � 8��Ȳ �@� p���<�\� �p�, ̱X�´� ����� X�t� ������. Geoffrey Harr i s (l 955) �� ̱\���0� ̱X�´� ���<�\� ����� T��Y�� x���� Xt�� ̱X´�� �̱� X �0��� |�� �䲔 ��� ��D� \��\� �ȥLj��. t ��\� �체�Ĭ|� ����XՀ�-̱X�´� 8�� �� Ĭ (hypothalamic-pituitary portal system) |� X�p� , t� �� Ĭ �� ̱ @� ̱X�´� �� ��t�X� <��� �X�D� ���XՌ� XՔ� D��� p��� \ �� 8���X� ݹ<�\� t�踴��� ���\� ��p���t��. 1 968 D� ̱ p���<�\���0� ����XՀ� 8���t��t ��L� �����0� �L��ɔ� Harr i s � 1955 D��� �Hň�X� �$�D� �Ʌ�t� � � �ǔ� ��p�� ���� �. 19 68 D� Roger Gu ill em i n �� Andrew S chall y� �0� ų�� � <�\� ��\���\�@� ��D�지금까지 알려진 바에 의하면 대부분의 시상하부 호르몬들은 작은 폴리펩티드형이다. 표 9-2 에서는 시상하부 호르몬과 이와 관련되는 뇌 하수체 전영 호르몬을 예시해 놓았다. 이 호르몬을 만들고 분비하는 시상하부 신경원은 뇌 속에 있는 다른 신경원으로부터 시냅스 영향을 받으며, 뇌와 뇌하수체 전영 사이의 연결을 이룬다. 그러므로 뇌의 활 동은 전 신체에 걸쳐 호르몬 분비에 직접적으로 영향을 미치는 시상하 부 신경원에 영향을 줄 수가 있다. 이런 사실 때문에 낮시간의 길이 변화가 동물의 배란에 영향을 미칠 수 있고, 어떤 냄새가 호르몬 분비 의 변화를 일으킬 수 있으며, 불안이나 염려가 여성의 월경주기를 바 꿀 수도 있고, 스트레스가 남성의 테스토스테론 수준을 낮출 수 있다 는 사실을 이해할 수 있게 되었다. 2 호르몬과 발달 호르몬은 주로 두 가지 방식으로 행동 발달에 영향을 미칠 수 있다. 특히 안드로겐과 같은 호르몬은 태아 발달시 유기체를 남성 형태로 분 화시키는 데 대단히 중요한 영향을 미친다. 호르몬은 사춘기와 그 이 후의 전 인생에 걸쳐 뇌뿐 아니라 전 신체의 조직에 작용하여 다양한 행동에 영향을 미친다. 생애 초기, 즉 태아기의 호르몬 영향을 발달 영 향 (develop m enta l eff ec t ) 이 라 하며 , 사춘기 와 그 이 후의 영 향을 활성 영 향 (acti va ti on al eff ec t) 이 라 부른다. 태아에 작용하는 호르몬의 발달 영향은 남성화나 여성화를 결정짓는 데 도움을 준다• 성 적 이 형 (sexual dim o rp h is m ) 이 란 남성 과 여 성 이 신 체구조나 행동상에서 많은 차이점을 보인다는 개념이다. 포유류는 호 르몬 분비의 양상, 성 행동 그리고 뇌와 신체의 구조상에서 성적 이형 을보인다.
2.1 호르몬 준비의 주기성 : 여성의 특징 인간을 포함하여 포유류 암컷의 호르몬 분비 특징은 에스트로겐이나 프로게스테론과 같은 성선자극 호르몬과 여러 종류의 너하수체 전영 호르몬을 주기적으로 분비한다는 점이다. 그러나 수컷둘은 이러한 주 기성을 보여주지 않는다. 대부분의 포유류에서 볼 수 있는 호르몬의 주기성 분비는 암컷의 성적 활동의 증가, 즉 발정 (es t rus) 과 관련 있 다. 난소에서 성숙한 난자를 배출하는 배 란 (ovula ti on) 은 발정 과 같은 시기에 일어난다. 이처럼 배란과 성적 활동의 절정시기가 서로 겹쳐짐 으로써 암컷의 임신 가능성은 높아진다. 호르몬 주기성에 따라 자발적으로 배란이 이루어지는 암컷을 가리켜 자발성 배 란자 (s p on t aneous ovulato r ) 라 하는데 , 인 간이 바로 자발성 배 란자이다. 그러나 토끼나 고양이와 같은 종도 주기적인 발정을 보이지 만 이 동물들의 배란은 질벽을 자극하는 데 대한 반웅으로 일어난다. 이러한 종의 암컷을 유발성 배란자(i nduced ovula t or) 라고 부른다. 이 양자의 배란은 뇌하수체 전영으로부터 분비되는 황체 형성 호르몬 (lute in iz in g honnone : LH) 의 갑작스런 분비 에 의 한다. 호르몬 분비 의 성차에 대한 연구는 오래전부터 이루어져 왔는데, 여기서는 1936 년에 Carroll P f e iff er 가 발표한 연구를 중십으로 살펴 볼 것 이 다. 2.2 안구 속의 난소 P feiff er 는 호르몬 분비의 성차에 관심을 갖고 먼저 실험실 쥐의 배 란을 연구했다. 배란은 LH 의 갑작스러운 분비에 의해 일어나기 때문 에 배란을 관찰한다는 것은 비록 간접적이긴 하나 주기성을 알아보기 위한 믿을 만한 지표가 될 수 있다. 암컷의 주기성을 결정하는 요인이 무엇인가를 밝혀내기 위해 P feiff er 는 발달도상의 여러 시기에 걸쳐 수컷이나 암컷 쥐의 호르몬 상태를 조작하였다. 일련의 실험을 통해 P feiff er 는 출생 직후 수컷의
고환을 제거하고, 그곳에 암컷의 난소를 이식시켰더니 이식된 난소에 서 암컷과 유사한 주기성 배란이 이루어진다는 사실을 발견하였다. 죽, 이처럼 수컷에게 이식된 난소는 정상적인 암컷 쥐처럼 4 일 주기의 배란을 보였다. 그런데 이 실험에서는 난소를 이 쥐의 안구에 이식하 여 주었기 때문에 안구의 두명한 체액을 통해 난소의 변화를 명확하게 관찰할 수 있었다. 안구 내에 이식된 난소는 혈관과 연결되었으며, 아 혈관으로부터 영양소와 정상적인 배란 기능에 필요한 뇌하수체 전엽 호르몬을 공급받았다. 수컷 쥐는 출생 칙후 거세되어 정상적으로 남성 호르몬을 분비할 수 없었기 때문에 유전적으로는 수컷임에도 불구하고 암컷과 같은 주기성의 호르몬 분바 양상을 보여주었다. P f e iff er 는, 성숙 후에 거세된 수컷은 무주기성의 남성 호르몬 분비 형태를 그대로 유지한다는 사실도 발견했다. 죽 난소가 안구에 이식된 이 수컷의 건강은 잘 유지되었지만 배란은 보여주지 않았으며, LH 의 갑작스러운 분비도 나타나지 않았다. P f e iff er 는 출생 직후 거세된 쥐 의 관찰을 통해서 이 쥐는 고환 속에 존재하는 남성화 물질이 제거되 었기 때문에 암컷 포유류의 특칭인 〈 본유적 호르몬 주기성(i nheren t hormonal c y cl icity)〉울 보여주는 것이라고 결론지었다. 그러나 성숙 후 고환의 제거는 이러한 호르몬 주기성을 나타내지 않는데, 이것은 이 쥐의 호르몬 변화에 영향을 줄 수 있는 결정적 시기나 민감시기가 이 미 지나가버렸기 때문일 것이라고 추론했다. 현재 우리는 이러한 결정 적 남성화 물질이 데스토스테론이란 것과 결정적 시기가 쥐의 경우는 출생 후 단 며칠 동안이지만 인간이나 고등 포유류의 경우는 이 시기 가 출생 이전이라는 사실을 알게 되었다. P feiff er 는 출생 직후 몇 시간 이내에 수컷의 고환을 암컷 쥐의 목에 이식해 주면 난소는 건강하게 자라지만 주기적인 호르몬 분비와 배란 울 보여주지 못한다는 사실도 발견하였다. 이 쥐의 난소에서 난자는 성장했지만 LH 의 분비가 없기 때문에 배란이 이루어지지 않았다. 이 실험의 결과는 그 후 보다 정교한 실험방법을 적용한 다른 연구자들의 연구에서도 재확인되었다. 이를 통해 우리는 포유류 암컷에서 호르몬
분비의 주기성은 본유적인 것이라는 것을 알 수 있게 되었다. 그러나 이런 기능이 발달 초기의 수컷 고환에서 분비되는 테스토스데론에 의 해 억압될 수 있다는 점도 알게 되었다. 뇌와 뇌하수체 전영 사이의 관련성은 Pfe i f fer 당시까지만 해도 잘 알지 못했다. 그는 뇌하수체가 호르몬 분비를 개시하고 종결하는 지배 선 (mas t er g land) 이라고 추측하였다. 그래서 그는 유아의 고환에서 분 비되는 핵심적인 남성화 물질이 뇌하수체에 직접 작용하는 것이라고 제안하였지만, 오늘날 데스토스테론이 호르몬의 주기성에 미치는 영향 은 이 호르몬이 뇌에 영향을 미침으로써 나타나는 간접적인 것으로 밝 혀졌다. 그러면 이처럼 생애 초기에 데스토스데론이 뇌하수체가 아니라 뇌에 영향을 미찬다는 것을 어떻게 확인할 수 있을까? P feiff er 의 실험에선 이 문제를 해결하지 못했지만 그 이후의 다른 연구자들이 이 문제를 설명할 수 있었다. Harr i s 와 J acobsohn(1952) 은 수컷 쥐의 뇌하수체를 정교한 수술로 때어내어 이를 이미 뇌하수체를 제거해 버린 암컷의 터 어키안 (sella t ur ci ca) 이란 뇌하수체 부위에 이식하였다. 이렇게 옮겨진 수컷의 뇌하수체가 암컷의 시상하부에서 내려온 문맥 체계와 연결됨으 로써 이 암컷 쥐는 정상적인 주기성 배란을 보였다. 이것은 곧 뇌하수 체는 뇌에 의해 지배되며, 뇌가 어느 성의 것인가에 따라 호르몬 분비 의 주기성 또는 무주기성이 결정된다는 것이다. Harr i s 와 Jac obsohn 은 위와 반대의 절차, 다시 말해 암컷의 뇌하수체를 수컷에게 이식하 면 이식된 뇌하수체는 찰 자라며, 뇌하수체 전영의 호르몬 분비 양상 은 수컷의 무주기성을 보여준다는 사실도 관찰하였다. 2.3 암수의 성 행동 유형 포유류의 수컷과 암컷은 서로 상이한 성 행동을 보여준다. 이런 연 구는 주로 실험실의 쥐를 대상으로 연구되었다. 쥐는 암수 간의 성 행 동에 뚜렷한 차이를 보이며, 사람과는 달리 호르몬 분비의 변동과 성
행동 사이에 명확한 관련성을 보여준다. 암컷 쥐의 발정주기 (estr u s c y cle) 는 혈통에 따라 5 일인 경우도 있지 만 전형적으로는 4 일의 발정주기를 보여준다. 4 일 주기와 5 일 주기 사 이의 차이는 5 일 주기의 경우 무발정 (d i es t rus) 일이 하루 더 길다는 점 이다. 이 주기의 처음 2 일 동안에 암컷은 성적으로 무반응적이다. 그 러나 3 일째 초저녁부터 암컷 쥐의 행동에 극적인 변화가 나타나기 시 작한다. 어떤 암컷 쥐는 활동이 증가되기 시작하는데, 이때 일반 활동 성을 측정해 보면 평소에 비해 7 배나 더 많은 활동을 보여준다. 발정 난 이 쥐는 발정 개시 후 24 시간 동안 음식도 적게 먹으며 성숙한 수 컷이 가까이 있으면 성적 관심과 성적 반응을 보이기 시작한다. 만약 수컷이 관심을 보여주면 암컷은 수컷 앞에서 깡충깡충 뛰고 돌진하는 특칭적 행동을 보이고 자신의 성기를 수컷이 탐색하도록 하는 유혹반 응도 보인다. 또한 암컷은 앞으로 돌진했다가 자신의 귓바퀴를 매우 빠르게 움직이는 반응도 보여주는데 이러한 행동들은 수컷을 유혹하기 위한 행동이다. 그 후 수컷이 자신을 따라오면 암컷은 돌진 후 깡충깡 충 뛰었다가 귀를 쫑긋거리는 상동적인 행동들을 되풀이한다. 성 행동 연구의 선구자 중 한 사람인 Frank Beach(1976) 는 이러한 유혹 행 동들을 교태 행동(p roce pti ve behavio r ) 이 라고 부르고, 교태 행 동이란 성적 접촉을 증가시키게 하는 암컷의 성 행동이라고 정의했다. 교태 행동의 세부적 내용은 종에 따라 다르지만 모든 포유류 암컷은 이 행동을 보여준다. 예컨대 고양이 암컷은 발정기의 절정에 이르면 수컷 앞에서 등을 바닥에 대고 몸을 굴리며, 앞발로 땅바닥을 긁고, 그르렁거리는 소리를 지른다. 레서스 원숭이 암컷은 수컷 결에 바짝 붙어 앉아서 수컷의 몸을 쓰다듬어 주며, 자신의 성기를 수컷에게 드 러내 보인다. 유혹 행동에 이 어 나타나는 행동을 수용 행동 (rece pti ve behavio r s) 이 라 하는데, Beach(1976) 는 수용 행동이란 암컷이 수컷으로 하여금 자 신의 등 위에 기어울라와 수컷의 성기를 자신의 질 속에 삽입하여 사 정할 수 있게 허용하는 행동이라고 정의했다. 고양이나 쥐의 주요 수
용 행동을 척추전만(l ordos i s) 반사라 하는데, 이것은 수컷의 성기 삽 입을 용이하게 하기 위해 암컷이 자신의 등을 아래로 낮추고 엉덩이를 위로 둘어울려 활처럼 구부리고 꼬리는 한쪽으로 밀쳐놓는 행동을 말 한다. 이러한 척추전만 반사는 암컷 쥐가 처음 교태 행동을 보이기 시 작한 후 약 6 시간 후부터 나타난다. 죽, 이 주기의 제 4 일째 되는 이론 아침에 일어나는 것이 보편적인데, 바로 이 날이 발정일이다. 발정기 간은 평균 19 시간 정도 지속되는데, 바로 이 기간 동안 배란이 이루어 전다. 발정기에 접어들면 수컷이 자신의 성기를 암컷의 질 속에 삽입하려 는 삽입 행동(i n t rom i ss i on) 이 현저하게 증가된다. 죽 수컷은 몇 번의 재빠른 골반찌르기 (pe lvic th rus t)를 시도한 후 암컷의 등 위에서 내려 온다. 그 후 몇 분 이내에 다시 등 위에 울라가 골반찌르기를 다시 시 도한다. 사정은 등 위로 올라가 여러 차례 골반찌르기를 보여주는 마 지막 시기에 주로 나타나며, 또 사정이 일어날 때는 보다 깊은 골반찌 르기가 나타나는 것이 특칭이다. 교미와 교미 사이의 시간간격은 몇 초에서 몇 분까지 차이가 있치만 앞서와 같은 교미동작이 되풀이된다. 일반적으로 수컷은 몇 시간 사이에 4 번에서 10 번 정도 사정하며 암컷 은 발정기 동안 계속 수컷의 접근을 허용한다. 그러나 이렇게 교미기를 갇이 보낸 한 쌍의 암수를 새로운 주기가 시작되는 첫날, 다시 말해 하루 더 관찰해 보면 전혀 상이한 행동을 보여준다. 수컷은 여전히 그 압컷에 관십을 가지고 암컷에게 접근하지 만, 암컷은 발정기에 보여주던 깡충거림이나 귓바퀴의 움직임 또는 척 수전만 반사를 보여주지 않는다. 그 대신 암컷은 수컷에게 마치 적개 심을 가진 것처럼 이빨을 가는 반응을 보여준다. 수컷이 암컷의 이런 거부태도를 빨리 알아차리지 못하고 계속 추근대면 암컷은 수컷을 향 해 덤벼들며 발로 차기도 하고 물어뜯기도 한다. 이처럼 암수의 성 행 동이 서로 다른 것을 성 적 이 형 (sexual dim o rp h is m ) 이 라고 하는데 , 이 것은 상동적이고 종 특칭적인 것으로서 다론 종의 동물들도 보여준다. 그러나 사람은 많은 동물의 종 가운데 성 행동에 있어서 상동 행동을
가장 적게 보이는 종이다. 2.4 호르몬과 암컷의 성 행동 앞서 본 것처럼 발정주기 동안의 암컷 행동을 현저하게 변화시키는 요인은 무엇일까? 이에 대한 해답은 많은 호르몬둘 가운데 특히 에스 트로겐 때문이라고 할 수 있다. 죽 에스트로겐이 뇌나 신체의 여러 부 위에 영향을 미쳐 이러한 변화를 야기하게 된다. 에스트로겐은 교태 행동이 나타나기 약 24 시간 전, 즉 배란이 일어나기 약 36 시간 전부터 분비 수준이 증가하기 시작한다. 프로게스데론 분비는 에스트로겐 분 비가 최고조에 이른 약 20 시간 후에 최고조에 달하는데, 이때는 최고 의 수용성을 보이기 약 4 시간 전에 해당된다. 발정 행동을 일으키는 가장 강력한 에스트로겐을 에스트라디올 (es t rad i ol) 이라고 한다. 프로게스태론은 암컷이 수컷을 받아들이는 수 용 행동에는 이차적 역할을 하는 것으로 보이지만 교태 행동에는 가장 중요한 역할을 하는 것 갑다. 예컨대 난소를 제거한 쥐는 성적으로 무 반응을 보인다는 사실은 널리 알려진 것이지만, 이 쥐에게 에스트라디 울을 투여하면 수 시간 후 척수전만 행동을 보이기 시작한다. 만약 난 소가 제거된 쥐에게 에스트라디울을 주사하고 몇 시간 뒤에 프로게스 테론을 다시 주사하면 아주 양호한 척수전만 반응이 나타난다. 이 결 과는 에스트로겐 하나만을 주사하여도 척수전만 반응과 같은 단순 수 용 행동은 보여주지만 프로게스데론을 잇따라 추가 투여해 주지 않으 면 교태 행동이 잘 나타나지 않을 것이란 사실을 시사하는 것이다. 에 스트라디올의 주요 작용 부위는 시삭전영역의 주변과 시상하부 그리고 편도핵이며, 프로게스테론의 작용 부위는 중뇌에 위치하는 것으로 밝 혀졌다.
2.5 난소의 주기성 암컷은 행동상의 주기성뿐만 아니라 난소의 주기성도 보여준다. 배 란 (ovula ti on) 은 성적 수용성이 최고조에 이를 즈음 나타난다. 난자의 발달과정과 배란은 네 가지 주요 호르몬, 즉 FSH, LH, 에스트로겐 및 GnRH 에 의한다. 쥐의 경우 이돌 호르몬의 수준은 새로운 발정주 기가 시작되는 제 1 일에 가장 낮다. FSH 수준은 제 2 일 늦게부터 점차 증가하기 시작하여 배란이 이루어지는 몇 시간 전에 최고 수준에 이론 다. FSH 수준이 증가하면 이 호르몬은 난포 (ova ri an follicle ) 와 난자에 작용하여, 난포가 파열될 때까지 난포를 성장시키고 발달시킨다. 배란 이란 난포의 벽이 찢어져 그 속에 있는 난자가 배출되어 나오는 과정 이다. LH 분비는 에스트로겐이 결정적 수준에 이를 때 나타난다. 쥐 의 경우 에스트로겐은 GnRH 의 수준을 직접적으로 증가시켜 뇌하수체 전영에서 분비되는 FSH 와 LH 분비 수준을 높이게 된다. 인간을 포 함한 많은 종의 포유류에서 에스트로겐은 GnRH 에 대한 뇌하수체 전 영의 감수성을 증가시킴으로써 GnRH 의 분비가 많으면 이에 대한 반 응으로 더 많은 양의 LH 와 FSH 가 분비되도록 한다. 2.6 포유류와 뇌하수체 후엽 포유류 암컷이 임신하게 되면 젖을 생산하기 위한 준비로 유선에 변 화가 생기게 된다. 젖의 생산과 분비에 관여하는 호르몬이 옥시토신 (ox yt oc i n) 과 프로락틴(p rolac ti n) 이다. 옥시토신은 8 개의 아미노산으로 이루어져 있고, 뇌하수체 후영에서 분비되는 호르몬이다. 프로락틴은 대단히 큰 폴리펩티드이며 뇌하수체 전영에서 분비되고 유선이 젖을 생성하도록 자극하는 호르몬이다. 일단 젖이 생성되면 젖은 폐포 (alveoli : 젖꼭지 주변에 있는 관) 속으로 들어간다. 젖이 젖꼭지로부터 방출되기 위해서는 폐포 주변의 근육세포들이 수축되어 젖이 신생아의 입 속으로 흘러들어가도록 해야 한다. 이러한 젖의 분비는 신경내분비
반사 (neuroendocr i ne re fl ex) 의 전형적 예이다. 죽 젖꼭지나 그 주변의 신경종말이 자극받으면 신경충격이 발생되고, 이 신경충격은 옥시토신 을 혈류 속에 분비하도록 하는 신경원을 활성화시킨다. 옥시토신이 폐 포 주변의 수축세포에 닿으면 이 세포는 수축하기 시작하여 젖을 분출 하게 된다. 포유반사 (nurs i n g re fl ex) 는 규칙적으로 젖을 주는 자리에 앉는다거나 젖을 달라고 졸라대는 아기를 보는 것과 같은 수유와 관련 된 자국들에 의해 조건화될 수 있다. 2.7 호르몬과 수컷의 성 행동 포유류 암수 사이의 주요 차이는 암컷은 성적 수용성에서 주기성을 보이지만 수컷은 그렇지 않다는 점이다. 그러나 대부분의 동물에서 수 컷이 성적으로 활동성이 높은 시기가 있는데, 이 시기가 곧 교미 시기 이며 수컷은 교미 활동을 위한 준비를 갖추게 된다. 호르몬과 수컷의 성 활동 사이의 관계는 암컷의 경우만큼 뚜렷하지 않다. 일반적으로 수컷은 거세, 죽 고환을 제거하면 교미 활동이 정지 되지만 암컷의 등 위로 울라가는 행동이라든가 암컷의 성기를 찾는 성 과 관련된 행동들은 일정 기간 지속된다. 레서스 원숭이의 경우는 거 세 후 여러 해 동안 계속하여 암컷에게 기어오르기, 삽입, 골반찌르기 와 행동상의 사정까지도 보인다 (Phoen i x, 1976) . 수컷의 성 행동이 호르몬에 의존한다는 명확한 증거가 실험실의 쥐 를 사용한 실험에서 제시되었다. 거세 후 수컷 쥐의 성 행동은 몇 주 동안에 걸쳐 감소된다. 죽, 먼저 사정이 정지되고 이어 삽입 행동이 사라지며 마지막으로 기어오르기 행동이 사라진다. 거세한 수컷의 성 행동은 궁극적으로는 완전히 사라지지만, 거세된 후 성 행동이 사라전 동물에게 안드로겐을 두여하면 일정 회복기가 지나 성 행동이 되살아 난다.
2.8 테스토스테론과 성욕 흔히 데스토스태론(t es t os t erone) 의 수준이 성적 활동의 수준과 관련 있는 것처럼 언급되고 있지만 이런 견해를 지지할 근거는 없다. 거세 후에는 성욕이 줄어들고 테스토스테론을 주사해 주면 다시 회복되지 만, 정상 상태에서 성 행동 자체가 테스토스테론의 수준에 의해 직접 지배된다는 근거는 없다. 대학생을 대상으로 한 연구에서 테스토스데론의 수준과 질문지법으 로 알아본 성적 활동 및 성적 관심 사이에는 상관이 없다는 사실이 보 고되었다 (Mon ti, Brown, & Corriv e au, 1977). 죽 Monti 등의 연구에서 는 테스토스테론의 수준과 수음 사이에는 정적 상관을 보였지만, 다론 형태의 성 행동들과 데스토스테론의 수준 사이에는 아무런 상관도 없 었다. 실험실 쥐의 경우는 성욕과 혈액 내의 테스토스테론 수준 사이 에는 약한 상관관계 를 보여준다 (Damassa 등, 1977). 성기 의 발기 가 불 가능한 남자의 혈류 데스토스태론 수준도 거의 정상적이며, 대부분의 발기부전 환자들에서도 호르몬의 기능 장애는 별로 문제가 되지 않는 다. 예컨대 당뇨병 환자의 발기부전은 순환계 장애 때문이지, 성 호르 몬 장애가 아니다. 미국과 유럽에서는 성 범죄를 저지른 죄인에게 법적으로 거세를 시 행하는 경우가 있는데, 이들을 대상으로 한 David s on, Kwan 그리고 Greenlea f (1982) 의 연구에 의하면 이들은 거세 후 성 행동이나 공격 행동에 관한 중요한 질문에 대해 언급을 하지 않았다고 한다. 의과적 수술에 의해 거세된 사람들은 성적 관련 행동들이 극적으로 약화되고 사라지는 것이 일반적이지만, 몇몇의 경우에는 사정은 불가능하지만 발기나 성 활동은 억제되지 않는다는 보고도 있다. 최근에는 의과적인 거세 방법이 화학적 거세법으로 바뀌고 있다. 시 프로테 론 (cyp ro te r one) 이 나 메드록시프로게 스데 론 (medroxyp ro g es te r one) 과 같은 약물은 고환에서 데스토스데론의 생성을 낮추는 작용을 하는 항안드로겐 (an ti andro g en) 이다. 이 약물들은 전립선이나 남성기와 같은
테스토스데론에 민감한 조직에 작용한다. 어떻게 화학적 거세가 성 활 동을 억압하는지에 대해서는 명확하게 알 수 없지만, 이 약물을 사용 해서 얻을 수 있는 가장 큰 장점은 약물의 효과가 역전될 수 있다는 점이다. 인간의 성 행동은 유인원에 비해 호르몬의 영향을 적게 받는다. 일 반적으로 인간의 경우 남성이나 여성의 성 행동을 체계적으로 호르몬 과 관련지울 수 있는 확실한 증거는 없다. 그러나 성 호르몬의 수준이 매우 낮아지면 성욕이 감소되며, 이런 경우 적절한 호르몬 치료로 성 적 관심을 회복시킬 수 있다. 정상적인 사람을 대상으로 성 행동과 호르몬과의 관계를 연구한 선 행 연구들에서는 연구의 결과들이 서로 일치되지 않는 수가 많았다. 인간의 성욕은 문화, 종교, 성격과 같은 여러 요인들에 영향을 받는 다. 비록 호르몬의 영향을 완전히 배제할 수는 없겠지만 호르몬의 영 향이 모든 개인들에게 동일한 것은 아니다. 물론 보다 적절한 기법과 실험설계를 적용함으로써 호르몬이 성 행동에 미치는 영향을 보다 자 세하게 밝혀낼 수 있을 것이다. 성 호르몬 분비 수준이 극적으로 높아 지는 사춘기가 시작될 때부터 대부분의 청소년들은 성인과 같은 성적 관십과 느낌을 경험하기 시작한다. 비정상적으로 지나치게 일찍 사춘 기가 도래되는 사례도 있는데, 이 경우는 보통 10 세 이전에 사춘기가 시작되는 경우로서, 이 아이들도 성인과 같은 성적 공상이나 행동을 보여준다 (Mone y & Ehrhardt, 1972) . 인간의 성 행동에 관한 과학적 연 구는 이제 막 시작된 분야이며, 앞으로 많은 것이 밝혀질 것으로 기대 된다. 3 호르몬이 성 행동 발달에 미치는 영향 어린 시절 호르몬 투여가 성 행동에 미치는 영향은 여러 연구자들에 의해 보고되었다• 이 분야의 초기 연구들은 주로 Phoen i x 와 그의 동
료들(1 959) 에 의해 이루어졌고, 그들은 주로 기니피그를 실험동물로 사용하였다. 이둘은 임신중인 기니프그에게 남성 호르몬인 테스토스테 론을 주사하면 암컷 새끼들이 성숙 후에 암컷의 성 행동을 보이지 않 고 수컷의 성 행동만 보인다고 했으며, 그 후 쥐를 사용한 Harr i s 와 Lev i ne (1 962) 의 연구에서도 이와 비슷한 결과를 얻었다. 여 성 성 칭 결 여 증 (defe m i ni z a ti on ) 과 남성 화 (masculi ni z a ti on ) 란 개 념 사이 에 는 다소 차이 가 있다. Beach 와 그의 동료들 (1972) 에 의 하면 여 성성징 결여중이란 정상적으로 여성에서 나타날 수 있는 모든 발달 특 징들이 부분적 또는 거의 전적으로 억제되며 남성의 발달 특칭조차도 없거나 아니면 아주 미약하게 나타나는 증후를 가리킨다. 한편 여성의 남성화란 정상적으로 잘 발달한 남성에서만 나타나고 정상적인 여성에 서는 나타나지 않거나 미약하게 나타나는 해부학적, 생리학적 또는 행 동적 특칭이나 특질들이 여성에서 잘 나타나는 것을 의미한다. Whalen 과 Edwards(1967) 는 출생시 어린 수컷 쥐를 거세하면 성숙 후 다른 정상적 수컷이 접근할 때 척수전만 반응을 보인다고 했다. 이 러한 여성화는 생애 초기 며칠 이내에 데스토스테론을 주사해 주면 나 타나지 않는다. 한편 생애 초기에 거세한 수컷 쥐는 성숙시에 발정난 암컷 쥐를 배우자로 짝지어 주어도 수컷의 성 행동을 보이지 않는다. 만약 충분한 양의 테스토스태론을 결정적 발달시기 동안 투여하면, 성 행동은 남성화의 양상으로 발달되는 경향이 있다. 쥐의 경우 이러 한 결정적 시기는 출생 전 하루 또는 이틀 정도로부터 출생 후 6-7 일 까지이다. 출생 후 일주일 이후에 쥐에게 데스토스데론을 두여하는 경 우, 대량으로 투여하면 비록 출생 후 10 일경에도 효과가 나타나지만 소량 두여시에는 별 효과가 없다고 한다 (P faff 등, 1974). 그러나 유인 원의 경우에는 결정적 시기가 거의 전적으로 출생 전의 태아기에만 국 한된다.
3.1 쥐를 대상으로 한 연구 동물을 겁사하는 동안 작용하는 조건에 따라 관찰되는 행동의 종류 도 달라진다. 예컨대 피험동물에게 테스토스테론이나 에스트로겐 가운 데 어느 한 가지의 호르몬을 선택하여 처치한 후, 암컷에게 수컷울 제 시하고 암컷의 행동을 검사하는 경우와 수컷에게 암컷을 제시한 후 수 컷의 행동을 검사하는 경우가 있을 수 있다. 그 밖에 하루 중 어느 시 간에 검사하는가, 사전 검사의 경험 유무, 실험장치의 특성 등과 같은 많은 조건들이 또한 영향을 미칠 수 있다. P faff와 Z igm ond (1 971) 는 출생 칙후 거세한 수컷 쥐를 테스토스데론이나 에스트로겐을 투여한 후 성숙 후에 성 행동 등을 검사했는데, 이 동물들은 성숙한 후에 거 세한 수컷 쥐에 비해 남성 행동이 많이 줄어든다는 사실을 발견하게 되었다. 그러므로 어린 쥐의 거세는 남성성칭 결여를 일으킨다. 출생 직후 테스토스테론을 주사한 암컷 쥐를 같은 시기에 불활성 물 질인 기름을 주사한 암컷 쥐와 서로 바교한 연구도 있다. 데스토스데 론을 주사한 암컷 쥐는 성적으로 왕성한 수컷 쥐와 함께 있게 한 경우 기름을 주사한 암컷 쥐에 비해 여성적 행동을 잘 보이지 않는다. 그러 나 이 두 집단의 암컷 쥐 사이에 남성적 행동의 출현에는 어떤 일관성 있는 차이가 나타나지 않았다. 그러므로 출생 칙후 암컷에게 남성 호 르몬인 테스토스테론을 투여하면 남성적 행동이 나타나는 것이 아니라 여성적 행동만 감소되는 이른바 여성성칭 결여화가 특칭적으로 야기된다. 3.2 유인원 연구 유인원을 대상으로 출생 전 호르몬을 조작한 실험도 많은데, 이 방 면의 연구는 주로 Robert Go y와 Charles Phoen i x 에 의 해 주도되 어 왔 다 (Go y, 1970 ; Phoenix , Jen sen, & Chambers, 1983). 레서스 원숭이는 임 신기간이 약 168 일인데, Go y와 Phoen i x 는 임신 후 40 일에서 134 일 사이에 있는 어미 원숭이에게 테스토스테론을 투여하여 암컷 새끼를
남성화시켰다. 임신중의 어미에게 데스토스태론을 투여하여 태어난 암 컷은 의형적으로는 남성화된 성기를 가지고 있었지만, 내부생식기는 암컷의 특칭을 보였고, 사춘기의 도래도 6 개월 이상 늦어졌지만 월경 은 나타났다. 이 암컷 동물들이 성숙하면서 보여주는 사회 행동은 통제군의 수컷 과 암컷의 중간 형태를 보여주었다. Rosenblum (1 961) 에 의하면 젊은 수컷 레서스 원숭이와 암컷 원숭이 사이에는 사회행동상에 차이가 있 다고 한다. 죽 사춘기 이전의 수컷 원숭이는 같은 시기의 암컷에 비해 더 자주 맞붙어 싸우고, 거친 장난을 치며, 술래잡기 놀이를 하고 위 협적인 자세를 보인다고 한다. Go y에 의하면 남성화된 암컷은 전형적 인 수컷과 암컷 행동의 중간 형태의 놀이모습을 보인다고 한다. 이러한 행동 차이를 동물의 성 호르몬 분비 수준의 차이 때문인 것 으로 보기는 어렵다. 왜냐하면 이 실험의 피험동물은 모두 사춘기 이 전이었으므로 혈액 내 데스토스테론과 에스트로겐의 수준이 아주 낮기 때문이다. 더구나 출생시 정상적인 수컷울 거세하면 사춘기 이전의 3-4 년 동안 전형적이고 완벽한 수컷의 놀이행동이 나타나지 않는다. 따라서 출생 전의 데스토스테론 두여는 남성형의 놀이형태의 발달을 자극하는 방향으로 신경 계통을 변경시킬 것으로 예견된다. 태아기 호르몬 투여에 의해 남성 성기를 가진 암컷 새끼에 대해 어 미 원숭이는 그 새끼를 수컷으로 인지하여 수컷으로 대접하고 수컷의 행동을 강요했을 수도 있다. M it chell (1 968) 은, 레서스 원숭이의 어미 는 암컷 새끼와 수컷 새끼를 특히 생후 3 개월 동안 다르게 다룬다는 사실을 발견하였다. 죽 어미는 암컷 새끼를 수컷에 비해 더 억제적으 로 행동하도록 하고, 보호롤 더 많이 해주며, 처벌도 덜 가한다고 했 다. Go y도 M it chell 의 방식 대로 연구하여 새 끼 의 행동 여 하에 따라 어미가 새끼를 다루는 방식이 달라진다는 접을 발견하였다. 남성화된 암컷 원숭이를 대상으로 성숙 후에 성 행동을 관찰한 연구 도 있다 (Phoen i x 등, 1983). 이들 연구에 의하면 이 동물들의 행동 가 운데 남성행동 유형, 특히 공격성에서는 차이가 있었지만 성 행동의
남성화 경향성은 명백한 차이를 보이지 않았다고 한다. 4 태아기 데스토스테론 투여가 인간행동에 미치는 효과 만약 발생 도중의 태아가 Y 염색체를 가지면 성선은 고환으로 발전 하며 임신 56 일경에 이 미약한 고환조직은 소량의 테스토스데론을 분 비하기 시작한다. 이 테스토스테론은 의성기를 남성화시키며, 고환에 서 는 뮐 러 관 억 제 물질 (Mi llle rian inh ib it ing substa n ce : MIS) 이 분비 되 어 내부 생식기관을 남성형으로 분화시킨다. 데스토스테론은 본유적인 호르몬 분비의 주기성을 억압함으로써 뇌 를 남성화시키는 것으로 추측된다. 인간의 경우에도 태아기 동안 데스 토스테론에 노출되면 애매한 행동 양상이 나타나는데, 다음 두 가지 증후군은 인간에서 특칭적으로 나타나는 예들이다. 4.1 CAH 증후군 태아기 동안 유전적으로 여성 (XX 염색체)인 태아가 비정상적으로 높 은 안드로겐에 노출되는 경우가 있는데, 이런 경우를 선천성 부신비대 증 (cong en it al adrenal hyp e rt ro p y : CAH) 이 라 부른다. 이 중후는 태 아기 에 부신의 기능에 장애가 일어나는 것으로서 부산에서 코르티솔 (cort iso l) 보다 안드로겐이 더 많이 분비 되는 경 우이 다. 이 러 한 안드로 겐의 과다 분비는 의성기와 내부관 조칙의 남성화를 초래하게 된다. 그러나 비록 이러한 증후를 가진 아이가 태어난다 하더라도 의학적인 처치가 가능하기 때문에 더 이상의 남성화가 초래되는 것을 막을 수는 있다. 죽 성형의과적 수술에 의해 남성형 성기를 여성형 성기로 바꿀 수 있으며, 수술 후 정상적인 여자로 키워질 수 있다. 이러한 치료법 이 발견되기 전에는 이런 아동은 남성으로 인지되고 양육되어 왔다. Mone y와 Ehrhard t (1972) 는 수 년 동안에 걸쳐 이러한 CAH 환자들의
심리적 또는 사회적 발달을 관찰하였다. 특히 이 연구자들은 CAH 환 자들이 자신의 성정체감(자기가 남성인가 여성인가), 성관련 행동들 그 리고 성적 관심(동성애와 이성애) 등의 발달에 미치는 영향에 관심을 가져왔다. CAH 증후와 관련된 세 가지 일반적인 특칭을 요약하면 다 음과같다. 첫째, 대개의 경우 CAH 환자들의 성정체감은 자신을 여성으로 본 다는 점이다. 둘째, 아동기의 성관련 행동들은 다소 남성화의 경향을 보인다. 죽 CAH 소녀들은 전형적인 소년들과 같이 게임을 즐긴다. 셋째, CAH 환자들의 성적 관심은 비록 정상적인 여성집단에 비해 동성애를 보이는 경향이 높지만, 아직 확실한 결론을 내리기는 어렵다. 4.2 안드로겐 둔감증후 또 하나의 선천적 장애로서 유전적 남성 (XY 성영색체)의 체세포가 안 드로겐에 대한 반응성이 결여되는 경우이다. 이러한 안드로겐 둔감증 후 (androg en ins ensit ivi ty syn d rome) 를 가진 사람은 음경 도 음낭도 출현 되 지 않는 대 신 다소 확대 된 음핵 (clito r is) 과 마치 자루처 럼 움푹패 인 질과 같은 비정상적인 성기를 갖는다. 고환은 있지만 복강 내에 파묻 혀 있어서 바깥으로 돌출되지 않는다. 이런 성기를 갖고 태어나는 사 람들은 흔히 소녀로 인지되어 양육된다. 이런 안드로겐 둔감증후는 10 대 이전에는 잘 발견되지 않으며, 현재로서는 민감성을 증가시킬 수 있는 방법이 없으며, 사춘기에 여성과 같은 유방과 체형을 갖추는 데 충분할 정도의 에스트로겐이 고환에서 분비되기도 한다. 안드로겐 둔감증후를 가진 사람은 신체 구조의 여성화 정도가 너무 나 완벽하게 이루어지기 때문에 이들 가운데는 패션모델로 성공한 사 람도 있다. Money, Ehrhardt 및 Mas ic a(1968) 는 이런 증후를 나타내 는 14 명의 환자를 연구한 후 〈이들의 심리적 특성은 양자를 잘 육아할 수 있을 정도로 강력한 모성애를 갖춘 여성〉이라고 결론지었다. 이들
의 성적 관심도 정상적인 여성과 같은 방향으로 발달된다. 4.3 생식행동 외의 성차 몇몇 연구들에 의하면 동물세계에서 수컷과 암컷은 생식행동 의에도 많은 차이를 보인다고 한다. 대부분의 경우 이러한 차이는 유기체가 성장하는 결정적 시기 동안 데스토스테론의 출현 여부에 의해 결정되 는 수가 많다. 이 분야에 관한 연구들은 주로 쥐, 생쥐 또는 행스터와 같은 설치류를 대상으로 연구하였다. 연구된 행동에는 이동 행동, 종내 동물 사이의 공격성, 페로몬 분 비, 학습과제 수행능력, 뇌 손상 후의 반응능력 등이 포함되어 있다. 대부분의 연구 결과 설치류 암컷에게 출생 직후에 데스토스데론을 투 여하면 수컷의 행동과 극히 유사하게 발달되며, 이와는 반대로 수컷울 출생 직후 거세하면 암컷의 행동과 유사하게 된다고 한다. 4.4 암수의 뇌 구조 차이 1970 년대 초반 이래 동물 암수의 뇌 구조상에 차이가 있다는 연구들 이 많이 보고되었다. 이런 차이들 가운데 특히 발달시기 동안 호르몬 의 수준 차이가 가장 중요한 것으로 지적되고 있다. 이러한 차이를 처 음 증명한 연구자는 Ra i sman 과 Fie ld (1971 ; 1973b) 인데, 이들은 시삭 전영역에 있는 신경원의 수지상돌기의 전개 양상에 차이를 보인다는 점을 시사하였다. 죽 암수 간에 시상하부 복내측의 주변조직 신경원에 서는 시냅스 양상에 별다른 차이가 없지만, 시삭전영역에서는 특이하 게 차이를 보인다는 것이다. 그 후 Greenoug h 등 (1977) 과 Ay ou b, Greenough 그리고 J uraska(1983) 도 각기 행스터와 원숭이에서 위의 발견과 유사한 결과를 관찰하였다• Gorsk i와 그의 동료들 (1978) 은, 쥐의 경우 내측전시삭핵 (media l pre op tic nucleus : MPON) 의 크기에서 암수 간에 큰 차이를 발견했는
데, 수컷은 암컷에 비해 몇 배나 더 크다고 한다. 더구나 이들은 이러 한 차이가 발달기간 동안의 호르몬 조건과 관련 있다는 사실을 발견했 는데, 죽 수컷을 출생시 거세하면 정상 수컷의 MPON 보다 훨씬 적게 되는 반면, 출생 후 4 일째 암컷에게 1m g의 데스토스테론을 주사하면 정상 암컷보다 훨씬 더 큰 MPON 을 갖게 된다는 것이다. 이렇게 MPON 의 크기가 더 커진다는 것은 신경원의 수가 증가되고 양이 더 커진 것을 의미한다. MPON 이 생식행동이나 성적 이형행동과 관련 있을 것으로 추측되지만, 아직 이에 대한 명백한 증거는 없다. 한편 위에 든 사례 의에도 신경계통의 구조 차이가 성과 관련 있다는 보고 둘이 있다 (Arnold & Gorski, 1984). 인간의 뇌도 남녀 간에 차이가 있다는 보고가 Swaab 와 Fli ers (1985) 에 의해 보고되었는데, 이들은 10 세에서 93 세 사이에 사망한 사람들 가운데 13 명의 남자 뇌와 18 명의 여자 뇌를 대상으로 연구하였다. 이 둘은 이른바 〈 성적 이형 신경핵 (sexually dim o rp h ic nucleus) 〉이라 부르 는 시삭전영역을 탐색했더니 남자가 여자에 비해 약 2 . 5 배 더 크고 약 2 배 정도 세포수가 더 많다는 점을 보고하였다. 또한 이 신경핵의 크 기는 남녀 모두 나이가 증가되면 감소된다는 점도 밝혀졌다. 비록 이 신경핵의 기능이 무엇인지를 명확하게 알 수 없지만 이 부위가 GnRH 분비를 조정하는 뇌 영역인 것으로 추측되고 있다.
제 10 장 수면과 꿈 1920 년대 Hans Ber g er 가 인간의 뇌에서 일어나는 전기활동을 측정 하는 방법을 개발했는데, 이것이 수면과 꿈을 과학적으로 연구할 수 있는 계기를 마련하였다. 오늘날 우리는 전기적 활동 형태로 볼 때 수 면과 꿈이 각성과는 전혀 다르다는 것을 알고 있다. 그러나 꿈과 관련 되는 전기적 활동들은 각성의 전기적 활동 형태와 여러 가지 의미에서 유사하다. 이런 이유로 꿈꾸는 기간 동안의 수면을 역설적 수면이라 부른다. 꿈은 또한 급속 안구운동이란 특징도 보여준다. 급속 안구운 동 (ra pi d eye movement: REM) 수면이란 바로 꿈꾸는 시기의 수면을 지칭하는 것이다. 수면의 단계에 따라 관여하는 뇌 부위의 영역이 상 이하고 신경전달물질도 서로 다르다. 수면과 각성은 규칙적인 주기성운보이는데, 이 주기는 약 24 시간의 생물학적 리듬(일내주기)을 갖는 신경기제의 한 산물이다. 이러한 일내 주기성을 담당하는 신경 부위는 시상하부의 시삭 바로 위에 있는 시교 차상핵 (sup ra chia s matic nucle i)이라는 사실이 밝혀지고 있다. 실험동물 에서 이 신경핵을 제거하면 여러 가지 형태의 생물학적 주기가 깨진 다. 시간대가 바뀜에 따라 일어나는 생물학적 리돔 손상을 제트 랙 (jet lag) 이 라 한다. 일반적으로 우리는 수면이 무엇인가를 잘 알고 있는 것 같지만, 보 다 정교하고 공식적인 개념으로 이를 정의하여 사용해야 한다. 우리는
수면을 운동이 저하되고 자국에 대한 반응성이 감소되며, 뇌의 전기적 활동에 심한 변화가 나타나고, 근육이 최대한 이완되는 상태로 간주한 다. 이와 갇은 정의는 수면을 마취상태, 혼수상태, 최면상태와 같은 표면상으로 유사한 상태들과 구분짓게 하는 데 큰 도움을 준다. l 수면의 측정 一― EEG 사람이나 동물이 실제로 참자고 있다는 것을 어떻게 확인할 수 있을 까? 참자는 사람은 눈을 감고, 규칙적인 주기로 호흡하며 근육은 이 완되어 있다. 그러나 가장 정확하게 수면을 측정하는 방법은 뇌파를 측정하는 방법이다. 뇌파는 뇌의 자발적 전기활동의 변화로서 뇌피질 의 표면충에 있는 수많은 신경원들의 시냅스후 전위가 끊임없이 흘러 나옴에 따라 야기되는 것이다. 이 전기적 활동은 뇌전도 (elec t roence pha log ram : EEG) 란 방법에 의해 측정된다. 1.1 EEG 의 발견 뇌파는 독일의 정신과 의사 Hans Ber g er 에 의해 처음 기록되었다. 그는 두피 위에 장치한 전극으로부터 인간 뇌의 전기적 활동을 측정할 수 있는 가장 기초적인 기법을 개발하였다. 그는 처음에 제 1 차 세계대 전에 참여하여 국소적인 뇌 손상을 입은 환자들의 뇌 활동을 기록하였 다. 이 환자들을 대상으로 Berg er 는 두개골이 손상되어 함몰된 부위 바로 위 두피에 전극을 부착하여 전기적 활동을 기록하였는데, 이렇게 한 이유는 두개골이 없는 곳에 전극을 두면 뇌파가 보다 쉽게 측정될 수 있을 것으로 믿었기 때문이다. 그 후 그는 이 기법을 개량하여 정 상적인 사람의 두개골 위에서도 EEG 를 기록할 수 있게 되었다. 그는 1924 년부터 1929 년까지 계 속된 자신의 연구결과를 1929 년에 발표하였 는데그의 논문이 처음 발표되었을 때 많은 과학자들은 그 결과를 의
심하였다. 그러나 그 후 몇몇 과학자들이 그의 연구결과를 재확인하게 되었으며 EEG 는 뇌 기능을 분석하기 위한 유용한 수단이 된다고 믿게 되었다. 모든 포유동물과 같이 인간의 경우도 EEG 로 기록된 전기 활동의 빈 도는 초당 1Hz 미만에서부터 35-50Hz 에 이르고, 뇌파의 전폭은 5-lOOµV 사이인데, .8H z 이상의 빈도를 갖는 경우는 전폭이 20µV 를 초과하지 않는 것이 일반적이다. 깨어 있는 사람의 EEG 기록은 두 가 지 기본 양상을 보여준다. 죽, 첫째 양상은 8-13Hz 의 규칙적인 동기 성 (syn c hroniz e d) 형 태 를 보여주는 뇌 파이 다. 이 뇌 파를 알파 리듬 (alph a rhyt hm: a 파)이라 하는데, 이의 명칭은 a 란 문자가 그리스어의 알파벳 첫 문자이고 또 이 리듬을 Berg er 가 최초로 발견한 뇌파라는 뜻에서 a 파라고 부론다. 둘째 양상은 대단히 높은 주파수 (15-30Hz) 의 저전압활동 뇌파인데, 이를 베타 리듬 (be t arh ythm: p파)이라 한다. a 파는 깨어 있지만 눈을 감고 있는 이완 상태에서 찰 나타나는 뇌파이 며, /3파는 눈을 뜨고 각성하고 있을 때 잘 나타나는 뇌파이다. 1.2 수면의 단계 EEG 기록을 기초로 할 때, 수면은 일반적으로 5 단계로 나눌 수 있 다. 그 중 처음 단계부터 4 번째 단계까지는 주파수가 7Hz 미만인 서 파 수면 (slow-wave sleep : SWS) 으로 · 분류되며, 5 번째 단계의 수면은 앞서 단계의 그것과 판이하게 다른 양상을 보여준다. 즉, 5 번째 단계 수면의 EEG 는 각성 상태의 EEG 와 유사하다. 이 단계에서는 급속 안 구운동도 나타나기 때문에 이 단계의 수면을 REM 수면이라고도 부른 다. 또한 이 단계의 수면울 역설적 수면(p aradox i cal slee p)이라고도 한 다. 그 이유는 이 단계의 뇌파가 저전압, 고빈도 뇌파로서 보다 느리 거나 높은 전압이 위주가 되는 수면중의 뇌파와는 역설적이기 때문이 다(그립 10-1). 제 1 단계로부터 4 단계까지의 서파 수면은 각 단계로 진행되면서 뇌파
 각성 ``-`-~- 각성
각성 ``-`-~- 각성
하고도 복잡한 내용의 꿈을 꾸었다고 보고하지만, 깨우지 않고 그냥 둔 사람들은 아침에 일어나 지난 밤에 꾼 꿈의 내용을 잘 기억하지 못 한다. 실제로 꿈을 꾸었지만 꿈을 꾸지 않았다고 믿는 것은 바로 이런 이유 때문이다. 2.1 나이에 따른 수면 변화 사람은 나이에 따라 수면의 양이 달라진다. 신생아는 하루 종일 거 의 수면으로 일관하며 수면의 내용은 주로 REM 수면이다. 10 세쯤 되 면 수면의 비율이 성인의 비율과 거의 같아진다. REM 수면의 시간과 비율은 가장 깊은 수면단계인 제 4 단계 수면과 같이 나이가 많아지면서 점차 줄어들며 60 세 이상이 되면 아예 사라지는 경우도 있다 (Kell y, 1981). 현재로서는 왜 이처럼 나이가 많아지면서 수면의 양상이 달라 지는가를 설명할 수 있는 명확한 근거가 없다. REM 수면이 꿈과 관련 있기 때문에 교과서보다는 오히려 통속서에 서 이롤 더 많이 언급한다. REM 수면이 아닌 서파 수면 단계에서도 꿈을 꾸는지에 관해서는 현재 논쟁중에 있다. Demen t (1974) 에 의하면 서파 수면 단계에서 깨워진 피험자 가운데 약 14% 가 꿈을 꾸었다고 하는 데 반해 REM 수면 때 깨워진 피험자는 약 80% 가 꿈을 꾸었다 고 보고했다고 한다. 그러므로 REM 수면이 비 REM 수면보다 꿈과 보다 밀접하게 관련되어 있지만 서파 수면 동안에도 꿈을 꿀 수 있는 가능성을 배제할 수 없다. 이러한 상이한 결과들은 꿈에 대한 정의상의 문제에서 기인될 수도 있다. REM 수면의 꿈은 시각적 상 (v i sual im a g e) 과 꿈꾸는 자신이 포 함되는 화제로 이루어지기 때문에 이 꿈을 심상적 꿈(im a g e ry dream) 이라 한다. 반면 비 REM 은 대낮에 나타나는 사색이나 공상처럼 보다 문제 중심적이며 십상 내용이 적기 때문에 이를 정적 꿈 (s t a tic dream) 이라 한다 (Foulkes, 1966). 그러므로 이러한 상이한 꿈의 내용이 동일 한 꿈의 정의 속에 동시에 포함되어 있다. 이러한 정의상의 문제와 더
불어 피험자에게 꿈의 여부를 알아보는 질문법의 차이 때문에 상이한 결과가 나올 수도 있다는 주장이 있다. REM 수면은 남성의 경우 음경받기와 관련 있고, 여성의 경우는 질 의 분비물 분비와 음핵의 확대와 관련 있다고 한다 (K i es t er, 1980). REM 수면 동안 음경발기가 일어난다는 것은 발기부전증(imp o t ence) 이 심리적 원인에 기인하는 것인지 아니면 생리적 원인에 기인하는 것 인지를 가려내는 데 중요한 방법이 될 수 있다. 예컨대 만약 수면 동 안 받기가 되는 환자라면 각성 도중 이 환자의 발기부전은 순전히 심 리적인 원인에서 일어나는 것으로 볼 수 있을 것이다. REM 수면에서 는 근긴장도가 갑자기 저하되는데 이러한 〈수면성 마비〉는 뇌간에 있 는 청반(l ocus coeruleus) 이란 신경핵의 기능에 의한 것인데, 이 신경핵 은 골격근을 지배하는 척수신경원에 억제성 영향을 미친다. 2.2 REM 결핍과 회복 REM 수면의 기능은 무엇일까? 이에 대한 해답은 아직도 잘 모르 지만 이러한 의문은 많은 연구를 자극하게 되었다. 사람이나 동물을 대상으로 REM 수면이 나타날 때마다 강제로 깨워 REM 을 박탈시키 는 실험을 행할 수 있다. 이러한 강제 각성은 일반적인 수면도 동시에 박탈하기 때문에 통제집단 피험자들에게도 서파 수면을 하는 동안 REM 수면을 박탈한 횟수나 시간만큼 서파 수면을 박탈시켜야 한다. 비록 초기의 연구들에서는 REM 박탈효과가 매우 국적인 효과가 있다 고 보고되었지만, 그 후의 연구들에서는 그 효과가 그렇게 극적인 것 이 아니라고 주장한다. 그러나 한 가지 명백한 결과는 REM 을 박탈당 한 사람은 REM 을 박탈당한 다음날 밤에 보다 많은 REM 을 취함으로 써 박탈된 REM 을 보상받으려는 것 갇다는 것이다. 이러한 REM 회 복은 동물이나 사람 모두에서 나타난다. REM 박탈로부터 초래되는 뚜렷한 장기적 효과는 없다.
2.3 우울증 환자의 수면과 꿈 REM 박탈이 우울증 환자의 치료에 성공적인 방법이 될 수 있다는 보고가 있어 주목을 끈다 (Vo g el 등, 1980). REM 박탈이 장기적으로 유해한 결과를 가져오지 않기 때문에 REM 을 박탈함으로씨 우울증을 치료하는 것은 유용한 방법이 될 수 있을 것으로 기대된다. 정상인과 우울증 환자 사이의 주요 차이는 REM 수면의 총시간에서 차이가 나 는 것이 아니라 REM의 분포상에 차이가 있고, 우울증의 개선 정도와 REM 수면의 분포 사이에 상관관계가 있다는 것이다. 연구자들은 우 울증 환자의 경우 수면주기의 발전자가 손상되거나 약화되어 있기 때 문에 REM 박탈에 의해 이 발전자를 자극하게 되면 우울증이 개선될 것이라고 주장한다. 우울증 치료에 효과가 있는 모든 약물은 근본적으 로 REM 수면 비율을 감소시키지만 전반적인 수면시간은 증가시킨다 는 사실이 발견되었다 (Scherschl ic h t 등, 1982). 그림 10-2 에서는 정상 피험자와 우울증 환자 사이의 수면 양상의 차이를 예시하고 있다
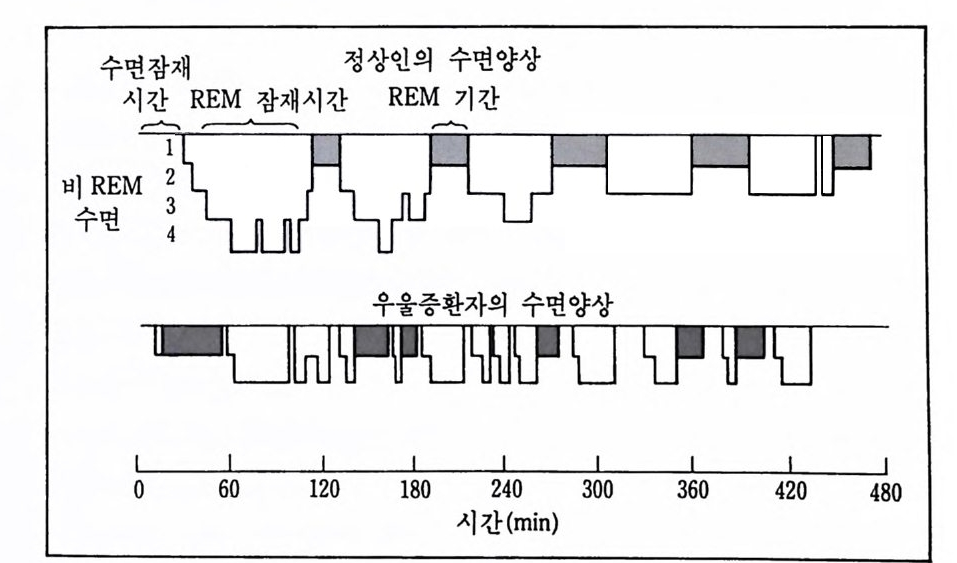 l:I \수R면시간잠:재 R\EM\ 잠 〕재시간\정 상」인RE의M \수기면간양」 상 나 ~lP
l:I \수R면시간잠:재 R\EM\ 잠 〕재시간\정 상」인RE의M \수기면간양」 상 나 ~lP
그림 10-2 정상인(위)과 우울중 환자(아래)의 수면양상 우울증 환자의 REM 수면(검은 부분)이 매우 빨리 시작된다는 접에 주목할 것.
(Gi llin & Borbely, 1985) . REM 수면이 서파 수면보다는 꿈과 더 높은 상관관계가 있다는 발 견은 꿈에 관한 연구를 자극하게 되었지만, 꿈의 기능에 관해서는 아 직도 잘 모르는 것이 대부분이다. 이런 연구들로부터 몇 가지 새로운 사실들이 보고되었다. 예컨대 긴 꿈이라 해서 실제 시간이 더 긴 것은 아니며, 수면중 배뇨는 꿈과는 무관하며 오히려 서파 수면 기간 동안 더 자주 나타나며, 수면중에 일어나 정신 없이 걷는 현상도 3, 4 단계 의 수면에서 더 잘 나타난다는 것이다. 어린이나 어른들이 가끔 꾸는 악몽도 서파 수면 동안에 나타나지 만 가위 눌림 (n ig h t mare) 은 REM 수 면 동안에 나타나는 것이 보통이다(Kim ble, 1988). 3 수면장애 수면장애에는 불면증(i nsomn i a), 수면성 무호흡증 (slee p apn e a), 수 면발작 (narcole p s y) 등이 있다. 이 장애들을 보다 자세하게 살펴보자. 3.1 불면증 대부분의 사람들은 며칠 또는 몇 주일 정도 참둘기가 매우 힘들었던 경험을 해보았을 것인데, 이것이 바로 불면증이다. 우울과 불안이 불 면증을 유발하는 원인으로 간주되기 때문에 불안이나 우울을 치료하는 것이 곧 불면증 치료의 기초가 된다고 본다. 그러나 어떤 수면제도 불 면증 치료에 근본적인 효과가 없으며 수면제 복용은 건강한 사람을 불 면증 환자로 만들 수도 있다. 특히 노인이나 불안신경증 환자들이 단 순한 수면부족감을 채우기 위해서나 충분한 휴식을 취하지 못해 불안 하다고 느끼 기 때문에 벤조디 아제 핀 (benzodia z epi ne s) 과 같은 항불안제 를 수면제로 사용하는 경우가 있는데, 이 경우 어려운 문제가 발생한 다. 수면을 취하기 위해 약물을 취한다는 것은 곧 잠을 청하기 위해
습관적으로 약물을 취하게 되는 악순환을 거듭하게 되며, 드디어는 불 면증을 악화시키고 금단증상까지도 나타나기 때문에 수면제를 취하지 않는 것이 좋다. 불면증을 호소하는 환자들을 대상으로 실험실에서 행한 연구둘을 살 펴보면, 환자들은 그들 자신이 생각하는 것보다 실제로 잠자는 시간이 더 길다는 점이 발견된다. 더구나 불면증 환자들은 실제로 잠에 빠진 시간조차 깨어 있는 상태에서 꿈을 꾸고 있었다거나 잠자려고 노력하 고 있었던 중이라고 생각한다. 수면의 욕구는 사람에 따라 많은 차이 가 나는데 하루 1-2 시간 정도만 자고도 건강을 유지할 수 있는 사람도 있다고 한다. 수면과 관련 있는 신경전달물질은 세로토닌 (sero t on i n) 인 데, 이 물질 합성의 선구물질인 트립토판을 투여하면 보다 쉽게 수면 에 들어간다고 한다 (Har tm ann, 1980). 예컨대 자기 전에 따뜻한 우유 롤 마시라고 하는 경험적 충고는 과학적 근거가 있다. 왜냐하면 우유 는 세로토닌의 선구물질인 트립토판의 원료이기 때문이다. 3.2 수면성 무호흡증과 수면박탈 수면성 무호흡증이란 수면 중 호흡이 참시 동안 중단되는 수면장애 이다. 수면성 무호흡증은 수면자 본인이나 옆에서 이를 지켜보는 사람 에게는 걱정거리가 되겠지만 생명에 심각한 위험은 없다. 이것은 후두 나 기도의 이상에서 기인되는 경우가 많으며 의과수술에 의해 교정받 울 수 있다. 수면성 무호흡증은 비만체격인 사람들에서 특히 많이 발 생된다. 어떤 사람들은 낮 동안 갑자기 수면이 엄습해 와 몇 분 동안 깊은 수면에 빠지게 된다. 이런 경우를 수면발작이라 하는데 아직까지 치료 방법이 알려져 있지 않다. 이런 발작이 운전 도중 일어난다면 치명적 인 결과가 생길 수 있다. 때로는 이런 발작이 길 위에 서 있는 동안에 도 일어나 길 위에 쓰러지는 경우도 있다. 이 수면장애는 유전성이 매 우 강한 것으로 추측된다.
4 수면의 뇌 과정 수면과 각성이 일어날 때 뇌에는 어떤 변화가 생길까? 몇 년 전까지 만 하더라도 대부분의 연구자들은 수면이란 피곤할 때나 뇌가 정상적 인 작용을 하지 못할 때 일어나는 수동적 과정이라고 여겼다. 그러나 오늘날의 신경과학자들은 이러한 견해에 동의하지 않는다. 앞서 본 서 파 수면이나 REM 수면이 뇌의 능동적 활동과정의 산물이라는 증거가 많아 제시되었기 때문에 오늘날은 수면의 여러 상이한 단계가 뇌의 정 상적 활동 상태롤 반영하는 것으로 간주한다. Ber g er 의 EEG 기 법을 채 택하여 연구자들은 참자고 있는 동물의 뇌 에서 직접 EEG 를 기록할 수 있게 되었으며, 여러 상이한 종의 동물들 에서 보여주는 EEG 양상에 어떤 공통점이 있다는 사실도 확인하였다. 이러한 연구들 가운데 벨기에의 신경학자 Fredric Bremer 의 연구 (1936) 는 고전적인 연구로 간주된다. 그는 고양이를 대상으로 뇌를 두 개의 상이한 수준에서 절단한 후 전뇌에서 일어나는 피질의 전기적 활 동을 연구하였다. 그는 뇌간의 상우체 (sup er io r co lli cu li)와 하구체 (inf e r i or coll icu li ) 사이를 절단한 〈 전뇌 격리 (cerveau iso le)> 고양이와 척수와 뇌간 사이 부위를 절단한 〈 뇌격리 (enceph a le iso le)> 고양이룰 실험동물로 사용하였다(그립 10-3). 전뇌격리 동물의 경우는 정상적인 각성 활동이나 안구 활동의 지표가 되는 어떤 뇌파 활동도 피질에서 기록되지 않았고 눈동자는 수축된 상태로 유지되었으며, 서파 수면만 계속 보여주었다. 그러나 모든 뇌간 구조들과 전뇌가 연결되어 있는 뇌격리의 경우에 서는 각성과 수면의 명백한 주기를 보여주었을 뿐만 아니라, 각성 주 기 동안 눈동자가 확대되었고 움직이는 물체를 따라 안구운동도 보여 주었다. 따라서 Bremer 는 수면이란 뇌에 유입되는 감각흥분의 감소에 따라 나타나는 일종의 수동적 과정이라고 결론지었다. 그는 전뇌격리 의 경우는 제 5 뇌신경으로부터 제 8 뇌신경에 이르는 뇌신경의 감각유입 이 차단되어 뇌의 감각유입이 저하되지만, 뇌격리의 경우는 전뇌에 유
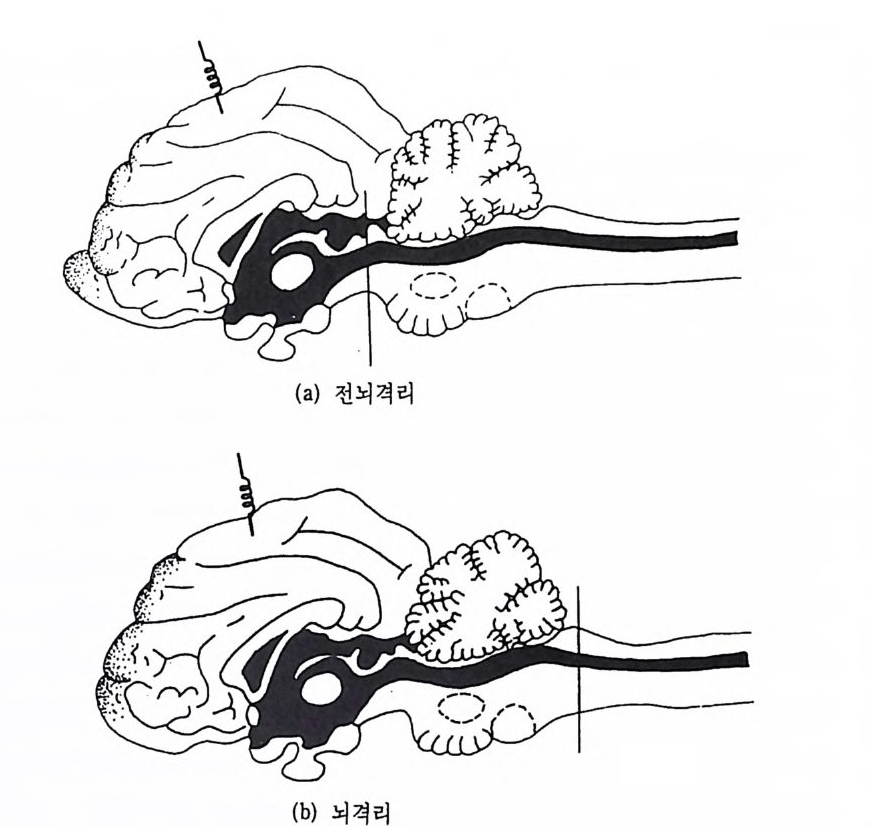 (a) 전뇌격리
(a) 전뇌격리
그림 10-3 전뇌격리와 뇌격리의 뇌절단 수준 입되는 12 개의 뇌신경 모두가 손상받지 않았기 때문에 감각유입이 찰 유지될 수 있어서 이런 차이를 보인다고 했다. 우리는 일반적으로 의 부 감각자극의 유입이 저하되는 조용한 장소나 어두운 밤에 참이 잘 오기 때문에 뇌에 대한 감각유입의 감소가 수면을 야기하는 것으로 생 각하기 쉽다. 그러나 어떤 사람은 밝은 불빛이나 소음 속에서도 잠을 잘 자기 때문에 Bremer 의 이 이론에는 의십이 간다. 수면이 감각자극 유입의 감소에 따른 수동적 과정이라는 이 견해에 대한 실험적 도전이 1949 년 Moruzz i와 Ma g oun 에 의해 제시되었다.
4.1 뇌간 망상체와 전뇌의 각성 뇌간은 단지 몇 개의 신경이 대뇌로 전입하는 장소만은 아니다. 이 곳은 많은 상이한 신경핵과 뇌와 척수 사이를 오르내리는 신경원의 축 색둘이 서로 연결되어 있는 곳이다. 뇌간의 중심부를 따라 새끼손가락 정도 크기의 원통형의 작은 신경핵 집단이 있는데, 이 부위에 있는 신 경핵의 축색과 수지상돌기들은 마치 그물〔綱〕 모양을 하고 있는데 이 부위 를 뇌 간망상체 (brain st e m retic u lar form ati on : BSRF) 라고 부른다. BSRF 에는 약 90 개의 신경핵 집단이 있는 것으로 확인되었지만 이 신 경핵들은 모두 유사한 기능을 하는 것으로 간주된다. 1949 년 Moruzz i와 Ma g oun 은 BSRF 가 다른 뇌 부위를 각성시키는 기능을 갖는다는 사실을 처음 발견하였다. 이들은 BSRF 를 전기적으 로 자국하면 참자는 고양이를 깨울 수 있고, 수면제 두여하의 동물의 서파 수면을 각성파, 죽 저전압의 속파로 바뀌게 할 수 있다는 것을 발견하였다. 그러나 같은 전기자극을 이 부위의 바로 인접 부위에 가 했을 때는 이러한 변화가 일어나지 않는다는 사실도 관찰했다. Moruzz i와 Ma g oun 의 연구가 발표된 직후 Donald L i ndsle y와 그의 동료들 (1950) 은 BSRF 를 손상시킨 동물은 행동상으로나 뇌파상으로 계 속 수면을 보여준다는 사실을 보고했다. 그러나 BSRF 를 남겨두고 이 부위의 바로 인접부위를 손상시키면 이런 수면이나 수면-각성주기에 별다른 영향을 미치지 않는다는 사실도 발견되었다. 이런 실험결과는 수면이 감각자극 유입의 감소에 따른다는 견해를 부정하는 결정적 단 서가 된다. 왜냐하면 BSRF 의 인접 부위의 손상은 와우각, 전정 계 통, 체성감각 계통 등으로부터 유입되는 구십성 감각을 근본적으로 차 단하는 것이기 때문이다. 이런 결과를 보고 Ma g oun 과 그의 동료들은 다른 감각의 유입보다 특히 BSRF 로부터 직접 피질에 이르는 감각입 력이 각성 상태를 유지하는 데 결정적인 것이라는 사실을 알게 되었 다. 후속된 연구들에 의하면 시상의 시상수질판속핵 (intra lami na r nucle i)도 전뇌의 각성에 관여하는 뇌 구조물인 것으로 밝혀졌다.
4.2 뇌의 수면 야기 영역 뇌간에 있는 중추들을 전기적으로 자극해도 수면이 일어난다. 이 사 실은 수면은 뇌간에 있는 신경원들에 의해 조정되는 능동적인 뇌 과정 이란 새로운 견해를 지지하는 것이다. 다시 말해 수면은 각성 상태와 다른 뇌 부위의 활동에 의해 생성되는 자연스런 상태라는 것이다. 그 후 계속된 연구들에서 바로 이 견해를 지지하는 수많은 증거들이 발표 되었다. 참자고 있는 동물의 뇌에서 단일 신경원의 활동을 기록해 보 떤 수면중이라도 전반적인 신경활동 수준이 감소되는 것이 아니라 어 떤 신경원은 4 단계의 수면중에 활동성이 더욱 증가된다는 것이다. 이 런 현상은 수면이 일찍이 Bremer 가 시사한 것처럼 특이성 감각 계통 의 활동수준 감소나 Moruzz i와 Bremer 가 시 사한 것처 럼 뇌 간망상체 의 활동 감소에서 일어나는 것이 아니라 이와는 다른 유형의 뇌 활동 에 의해 일어날 것이라는 사실을 시사해 준다. 4.3 J ouve t의 수면이론 수면의 뇌기제에 관한 주요 이론이 프랑스의 과학자 Mi ch el Jou vet 에 의해 제시되었는데, 이 이론에서는 뇌간에 있는 봉선체계 (ra phe s y s t em) 와 청반(l ocus coeruleus) 과 같은 두 개의 신경구조를 특별히 강 조한다(J ouve t, 1967 ; 1974). 봉선체계는 뇌간의 정중선을 따라 양쪽으 로 배열된 일련의 신경원덩어리로 구성된다. J ouve t는 고양이의 봉선 체계를 선택적으로 손상시키면 며칠 동안이나 계속 참을 자지 않는 불 면증이 나타난다는 사실을 발견하였다. 이 불면증은 점차 약화되지만 정상 고양이와 같은 하루 14-15 시간 수면까지는 회복되지 못한다. J ouve t는 또한 고양이 에 게 PCPA (pa rachlorop he ny la lanin e ) 를 주사하 면 주사 후 40 시간 동안 불면증을 보이고, 200 시간이 지나도 수면이 정상상태로 되돌아오지 않았다고 했다(J ouve t, 1974). PCPA 는 봉선체 계의 신경전달물질인 세로토닌의 합성을 억제하는 물질이다. 이러한
사실에 기초하여 J ouve t는 봉선체계는 각성을 유지하는 신경원을 억압 함으로써 서파 수면을 유도하는 기능을 담당한다고 제안했다. Jou vet 의 이론은 수면 과정을 능동적 과정으로 보는 것이며, 각성은 BSRF 주변에 있는 신경원에 의해 매개되는 것으로 본다. J ouve t는 또한 REM 수면도 설명하였는데, 그에 의하면 뇌간의 청 반이 봉선체계를 억압하여 서파 수면을 멈추게 하고 REM 수면을 유 발하게 된다고 주장하였다. 청반에 있는 신경원의 전달물질은 노르에 피네프린이다. J ouvet는, 청반의 손상은 REM 수면의 출현을 억제하 며, 노르에피네프린을 화학적으로 억제하여도 REM 수면이 나타나지 않는다는 점을 발견하였다. 그러나 다른 연구자들은 이와 다른 결과를 보고하고 있기 때문에 아직 청반과 노르에피네프린이 REM 수면 유발 에 미 치는 영 향은 명 확하지 않다 (Ramm, 1979) . 대부분의 연구자들은 수면이란 뇌간과 시상하부에 있는 신경원에 의 해 일어나는 능동적 과정의 결과라는 데 동의한다. 이 분야의 대표적 최근 이론은 Allan Hobson 과 Robert McCarle y (1977) 에 의해 제안된 이론인데, 이 이론에서는 J ouve t의 이론 가운데 일부를 취하고 여기에 다가 뇌교의 거대세포신경핵 (nucleus gian to c ellularis) 에 있는 콜린성 신경원의 개입을 강조한다. 즉 고양이의 경우 안구운동의 주기적 발사 와 함께 REM 을 보일 때는 거대세포신경핵의 세포들이 특히 활동적인 모습을 보인다는 것이다. 이롤 뇌교-슬상체-시각피질(p ons- g en ic ula t e occ ipital : PGO) 발사라고 하는데, 이러한 거대한 전기활동이 REM 수 면 동안 뇌교, 의측술상체 및 시각피질에서 기록된다. 이 PGO 발사 는 급속 안구운동을 야기하는 것으로 간주한다. 5 주기성 수면 인간의 수면은 어떤 주기적 모습으로 수면과 각성이 되풀이된다 . 인 간은 어두워지면 참자고 밝아지면 깨어 활동하는 경향이 있지만, 어두
워진다고 해서 반드시 잠에 빠지거나 밝아진다고 해서 깨어나는 것은 아니다. 최근에 들어와 인간뿐만 아니라 모든 척수동물들의 수면-각성 주기 는 이 주기 를 작동시 키 는 생 체 시 계 (bio l og ica l clock) 에 의 한다는 증거들이 제시되기 시작했다. 이러한 주기는 일출이나 일몰과 같은 환 경적 사건에 의해 작동되는 것이 아니라 유기체 내의 과정, 죽 내인성 과정에 의해 작동된다. 그러나 이런 내인성 주기는 일출과 같은 환경 적 사건과 동시에 일어나는 것이 일반적이다. 내인성 주기를 유도하는 환경적 사건을 시간 부여자 (ze itg eber) 라 부른다. 어떤 특정 주기가 내인성에 의한 것인지 아니면 단순한 환경사건에 의한 것인지를 알아내는 한 가지 방법은 동물을 환경사건의 단서가 전 혀 없는 특수환경 속(예컨대 시간을 알 수 없도록 조명의 변화가 없는 방 에 동물을 두는 것)에 두고 이런 주기가 일어나는지를 관찰하는 것이 다. 인간의 수면-각성주기가 환경조건과는 무관하게 내인성 주기에 의 한다는 사실이 밝혀지고 있는데, 그 근거는 광선, 온도 및 그 밖의 환 경자극과 격리된 특수 장소 예컨대 켄터키 주의 맘모스 동굴 속이나 프랑스알프스의 지하동굴 또는 세계 도처의 여러 실험실 연구에서 나 왔다. 이러한 연구들에서는 여러 가지 홍미로운 발견들이 제시되었다. 특 히 그 중에는 인간의 수면-각성주기가 24 시간이 아니라 25 시간 주기를 보인다는 점이다. 괘종시계, 태양광선 또는 그 밖의 환경적 단서가 주 어지지 않으면 사람은 보통 때보다 한 시간 늦게 참들고 한 시간 늦게 깨어난다는 것이다. 이러한 25 시간 주기는 나이나 성과 무관하다. 출 생 전 또는 출생 직후 장님이 된 사람도 위에서 본 격리된 사람들이 보여주는 25 시간 주기를 보인다고 한다. 대부분의 사람들이 주말이나 방학이 되면 평소보다 늦게 자고 좀 늦게 일어나는 이유는 바로 이런 25 시간 내인성 주기에 따르기 때문일 것이다. 수면_각성주기만이 유일한 일내주기성은 아니다. 체온, 오중 생성, 성장 호르몬, 코르티솔과 테스토스데론 같은 호르몬도 일내주기성을 보여준다. 일내주기성은 약물의 치료효과에도 중요한 영향을 미친다.
예컨대 난소암울 치료하기 위해 화학요법을 사용하는 경우 환자들의 일내주기 가운데 어느 때 약물을 투여하느냐에 따라 약물의 효과가 판 이하게 달라진다고 한다. 5.1 제트 랙 비행기를 타고 밤낮이 바뀌는 먼 의국으로 여행가면 여러 가지 불쾌 감을 경험한다. 이때 나타나는 피로감, 우울감 또는 기타 일반적 불쾌 감은 시간 변화에 관련되는 신체주기의 혼란 때문에 일어나는 현상으 로 이를 제트 랙(j e t la g)이라 한다. 이것은 신체의 일내주기 시간을 조정하는 시간 부여자가 서쪽에서 동쪽으로 날아갈 때는 빨라지고 동 쪽에서 서쪽으로 날아갈 때는 늦어지기 때문에, 일반적으로 서에서 동 으로 날아갈 때와 같이 주기성이 빨라지는 경우(서울에서 뉴욕으로 가는 경우)가 동에서 서쪽으로 날아가는 경우(뉴욕에서 서울로 오는 경우)보 다 적응이 더 어렵다. 서울과 뉴욕 간은 11 시간 차이인데 이 주기가 완전히 회복되는 데는 약 10 여 일이 소요된다. 이러한 제트 랙의 회복 울 촉진시킬 수 있는 방법에 관한 연구가 시도되었는데, 그 중 하나는 매우 밝은 광선에 노출하는 방법이다. 예컨대, 미국에서 한국으로 왔 울 경우 도착하자마자 태양이 작열하는 해수욕장에서 이틀 정도의 해 수욕만 하면 제트 랙이 극복될 수 있다고 한다. 5.2 시상하부에 있는 생물학적 시계 일내주기가 유기체 자신의 내인성 기제에 의해 작동되는 것이란 사 실이 알려지기 시작하면서 이 생물학적 시계가 뇌의 어디에 위치하는 지를 밝히 려는 연구들이 등장하게 되 었다. Curt R ic h t er 가 눈먼 쥐를 사용하여 이 분야의 연구를 처음 시도하였다 (R ic h t er, 1965). 눈먼 쥐도 정상 쥐와 유사하게 자유롭게 두었을 때 이동활동의 규칙적 주기를 보 여준다. R ic h t er 는 이러한 이동운동의 주기를 파괴시키기 위해 성선,
췌장선, 부신, 뇌하수체 , 갑상선 또는 송과선 등을 제거하는 방법을 시도해 보았지만 쥐의 일내주기는 근본적으로 바뀌지 않았다. 그래서 그는 최종적으로 뇌의 여러 부위를 체계적으로 손상시키는 실험을 시 도하였는데, 그 중 특히 시상하부 복내측핵 주변을 손상했을 때 활동 주기가 무너지는 동시에 물마시기와 먹이먹기의 일내주기성이 사라진 다는 사실을 발견하였다 (R ic h t er, 1967). 몇 년 후 이 주기와 직접적으로 관여되어 있는 시상하부의 신경핵이 정확하게 확인되었는데, 그 핵이 시삭 . 바로 위에 있는 시교차상핵 (sup ra chia s matic nuclei : SCN) 이 었다. Moore 와 E ic hler(1972) 는 SCN 의 손상이 코르티코스데론 (co rtic os t erone : 쥐의 부신피질 스테로이드 호 르몬 중 가장 중요한 호르몬임)의 주기성을 손상시킨다는 사실과 행스터 의 물마시기와 이동 행동의 주기성을 손상시킨다는 사실을 발견했다. 이 연구들이 보고된 후 SCN 에 여러 형태의 일내주기성을 조정하는 생체시계 또는 신경전동자가 있다는 증거둘이 계속 보고되었다 (Moore, 1982). 비록 SCN 이 뇌의 일내주기 계통의 주요 요소이지만 시상하부 바깥 부위에도 또 다른 주요 부위가 있다는 증거둘이 제시되었다. 예 컨대, 종에 따라서는 SCN 의 손상이 체온조절의 일내주기를 손상시키 지만, 레서스 원숭이의 경우는 SCN 이 손상되어도 체온조절의 주기는 물론 코르티솔의 분비주기에도 아무런 영향이 나타나지 않는다고 한다 (Takahashi & Zatz , 1982) . 어떻게 광선이 내인성 주기에 영향을 미치는가 하는 문제는 최근에 와서 상당히 많이 밝혀지고 있다. 망막의 절세포에서 나온 축색들 가 운데 일부는 의측슬상핵으로 진행되지 않고 SCN 으로 진행되는 것도 있다. 이 신경로는 매우 미세한 신경로인데, 이를 망막시상하부로 (reti no hy po th a lamic tra ct) 라 부르며 포유동물의 뇌 에 일반적 으로 관찰 되는 신경로이다. SCN 에서 나온 축색은 일내주기성을 작동시키는 신 경산호를 내보내는 시상하부의 여러 신경핵 영역으로 전행된다. 또한 여 기서 척수로 하행 하여 교감신경 계동의 상경 부절 (sup er i or cerv ica l g an gli on) 로 진행한 후 종국에는 송과선으로 되돌아오는 순환성 선경로
도 있다. 이 순환성 통로는 송과선에서 멜라토닌 분비의 일내주기성을 조절한다. 포유류의 행동에서 멜라토닌의 역할은 아칙 잘 모르지만, 특히 겨울참울 자는 행스터와 갇은 동물의 생식기제를 조절하는 데 매 우 중요한 역할을 하는 것으로 간주되며, 다른 포유류에서도 유사한 기능을 할 것으로 예견된다 (Rusak & Zucker, 1979). 멜라토닌의 분비 수준은 어두운 곳에서 증가되며, 밝은 빛에 노출되 면 낮아진다. 어떤 연구자들은 멜라토닌 분비의 이상이 계절성 감정장 애 (seasonal aff ec ti ve dis o rder : SAD) 와 관련 있을 것으로 생각한다. SAD 란 겨울철이나 이른 봄, 단 며칠 동안 심한 우울감을 경험하는 일종의 우울증이다. 이 장애는 밝은 햇빛을 쬐는 것이 좋은 치료법인 데, 이 병의 치료효과는 밝은 광선이 멜라토닌 분비 수준을 억제하기 때문에 나타난다고 한다 (Le wy 동, 1982) .
제 11 장 정서 행동 19 세기 Wi lliam J ames 는 우리가 경험하는 정서는 자율신경 계통의 신경원들에 의해 지배되는 심장이나 위와 같은 장기로부터 출발한 신 경흥분이 뇌에 전달된 결과라고 생각했다. 이 견해는 정서와 관련된 전통적 견해에 심각한 충격을 가했으며, 그 결과 정서에 대한 과학적 연구가 자극되고, 또한 자율신경 계통이 정서에 중요한 역할을 한다는 사실이 인지되기 시작하였다. 정서에 관한 또 다른 연구 추세는 1950 년대 Jam es Olds 와 Pete r M il ner 가 어떤 특정 뇌 부위를 자국한 쥐는 이 자극을 계속 취하기 위해 열심히 반응한다는 사실을 발견하면서 시 작되었다. 이러한 쾌중추의 발견은 정서 경험과 관련된 뇌 기능에 관 한 새로운 지식의 범위를 넓혀주었으며, 어떤 뇌 영역의 손상에 따른 정서통제 장애에 관한 이해도 가능하게 되었다. 1 정서란 무엇일까? 정서를 보는 관점은 학자에 따라 다르다. 그러나 한 가지 공통된 생각은 정서란 주관적으로 느끼는 경험을 말하고, 흔히 감정으로도 표 현된다. 이 감정은 우리가 흔히 공포, 분노, 불안 또는 즐거움과 같은 용어로 표현하는 것들이다. 우리는 얼마나 많은 종류의 정서가 존재하
는지 알 수 없으며, 이른바 〈기본정서상태 (basic e moti on al s t a t es) 〉 에 관한 견해조차도 학자에 따라 4-10 개 정도로 다르게 언급되고 있다 (Kim b le, 1988) . 정서는 주관적 경험이라는 견해 이의에 생리적 각성, 신체와 안면의 표현반응 그리고 정서관련 행동 등의 세 가지 요소를 포함한다. 다음 항에서 우리는 정서의 생리적 각성과 이 각성과 주관적 경험과의 관련 성을 자세하게 논의할 것이다. 정서 반응은 대부분이 정서 상태와 관 련되고, 자율신경계에 의해 매개되는 것으로 심장박동의 변화(심장고 동), 말초순환계의 변화(말초혈관의 수축에 의해 야기되는 손바닥의 땀 분 비), 땀샘의 활성화(신경성 발한), 침 분비의 억제(구강 건조) 및 소화 기의 변화(위의 운동) 등이 있다. 그 의 에도 부신에 서 분비 되는 아드레 날린 (adrenalin e ) 의 방출과 같은 다양한 호르몬의 변화가 특정 정서상태에서 나타나는 생리적 각성의 원인이 되기도 한다. 이러한 종류의 반응들은 공포나 분노뿐만 아니라 행복이나 환희를 느낄 때도 나타난다. 자율신경계의 교감신경 반응들 이 정서적인 각성과 관련되어 강조되어 왔지만, 부교감신경의 반응도 중요하다. 예컨대 심장박동은 각성시 주로 증가되지만 공포나 전율을 느끼는 상황에서는 오히려 감소된다. 그리고 어떤 극단적인 공포상황 에서는 심장마비가 일어날 수 있고, 불안과 스트레스 아래서는 위산 분비가 증가되며 이것이 위궤양을 일으킬 수도 있다. 심장박동의 감소 나 위산 분비의 증가는 교감신경의 반응이 아니라 부교감신경의 반응 이다. 자율신경 계통의 반응에 관심을 갖는 과학자들을 심리생리학자(p s y chop hy s i o l og ist) 라고 부른다. 십 리 생 리 학자들은 인간의 정서 적 삶을 알 아보기 위한 수단으로 다양한 신체반응둘을 측정한다. 가장 보편적으 로 측정되는 반웅들에는 심장박동, 혈압, 혈류량, 피부나 근육의 전기 적 활동 등이 있다. 그 밖에 가끔 측정되는 정서반웅들에는 호흡률의 변화, 손이나 안면 피부의 온도 변화, 동공의 크기나 소화기의 운동 등도 있다. 혈액을 표본 추출하여 혈장에 함유된 아드레날린, 코르티
솔, 데스토스데론 및 유리 지방산 등의 수준을 측정하여 보다 정교한 반웅을 알아보는 방법도 있다. 이러한 모든 반응둘은 개인이 의식적으 로 통제할 수 없다는 공통점을 가진다. 그러나 심장박동이나 혈압과 갇은 자율신경에 의해 통제되는 반웅들도 조작적 조건화 절차를 사용 한 바이오피드백 (bio f e e dback) 훈련을 동해 피험자 자신이 의식적으로 조철할 수 있다. 그러나 대부분의 사람들이 일반적 정서 상황에서 보 여주는 신체반웅들은 불수의적인 것이 특칭이다. 1.1 초기의 정서학설 신체의 각성과 우리의 주관적 경험 사이에는 어떤 관련성이 있을 까? 정서-유발자국이 신체 각성을 일으키고, 이런 신체 각성이 정서 로 경험되어질까? 아니면 의식적 경험과 신체 각성 양자가 동시에 이 루어지는 것일까? 이와 갇은 물음에 관한 공식적 해답이 1884 년 Wi lliam J ames (l 842-1910) 에 의해 처음 제안되었다. 그는 정서경험은 신체의 각성상태가 되먹임된 결과라는 견해를 제안했다. J ames 는 이 런 생리적 각성은 갑작스런 위험이나 정서를 야기하는 자극에 대해 반 사적으로 나타나는 것이라고 주장했다. 그는 〈우리는 울기 때문에 슬 픔을 느끼고, 떨기 때문에 두려움을 느낀다〉라고 주장했다. 1885 년 덴마크의 생리학자인 Carl Lan g e(1834-1900) 는 J ames 와는 독립적으로 극히 유사한 이론을 발표하였다. 그는 정서를 내장기, 말 초신경, 신체근육에서 되먹임된 결과라고 보았는데, 오늘날 우리는 이 두 견해를 동합하여 정서의 Jam es-Lange 이론이라 부른다. Lan g~근 뇌의 어디엔가 있을 것으로 보이는 혈관운동중추에서 매개되는 혈관의 변화가 정서의 바탕이 된다고 생각했다. J ames 와 Lan g e 의 정서이론은 1880 년대부터 50 년 동안 가장 영향력 있는 정서이론이었지만, 1927 년 Walt er Cannon(1871-1945) 에 의해 본 격적인 비판을 받기 시작했다. Cannon 의 첫 비판은 〈말초장기와 중추 신경의 연결을 차단해도 정서 반응에는 아무런 변화가 일어나지 않았
다〉는 사실에서 출발한다. 그는 J ames 의 견해를 직접 검증하기 위해 고양이의 교감신경절을 제거했다. 이런 수술을 받은 동물들도 수술 후 몇 개월 동안 건강하게 잘 지냈으며 정상적으로 움직이고 행동했다. J ames 와 Lan g e 의 이론에 의하면 이 런 처 치를 받은 동물들은 그들의 뇌가 내부장기로부터 고립되어 있어서 정서 경험을 할 수 없어야 한 다. 그러나 Cannon 은 이 동물들도 정서 반응을 잘 보여주었다고 했 다. 즉 이 고양이는 짖어대는 개를 만나면 으르링거리거나 귓바퀴를 쫑끗 세우며 이빨을 드러내고, 앞발을 들어울려 상대를 칠 준비를 하 는 따위의 다양한 분노반응을 보였다고 한다. 그러나 이 동물들은 교 감신경절에 의해 매개되는 모발의 융기 반응만은 보여주지 않았다. 이 실험의 결과는 J ames-Lan g e 의 정서이론에 대한 의문을 제기하는 계 기가 되었다. 한편 Cannon 은 J ames-Lan g e 의 정서이론에 대한 또 다른 반론을 제시했다. 그는 〈동일한 내장기 상태의 변화는 전혀 상이한 정서상태 나 또는 정서상태가 아닐 때에도 일어난다 〉 라고 반박하였다. 즉 상이 한 정서란 상이한 신체반응과 감각자국에 의한다는 J ames-Lan g e 의 이론을 지지할 수 없다는 것이다. l.2 정서의 인지-각성이론 Sta n ley Schac t er 와 Jer ome S i n g er(1962) 는 정서상태 하에서 나타나 는 신체반응의 역할에 대한 본격적인 연구를 수행했고, 이들의 연구는 그 후 심리학에서 유명한 연구의 하나로 인정되었다. 이 연구자들에 의하면 사람들은 그들에게 일어난 사건에 관한 어떤 종류의 〈인지〉와 그들이 경험한 정서를 호칭하기 위해 어떤 생리적 각성이 필요하다고 하였다. 이들은 정서경험이란 사건에 대한 인지와 생리적 각성으로부 터 되먹임된 사건이 상호결합되어 나타난다고 주장하였다. Schach t er 와 S i ng er 는 시 각의 예 민성을 증가시 키 기 위 해 새로운 비 타민의 효과를 알아본다는 이유를 대고 피험자들에게 아드레날린과 불
활성 물질인 위약(p lacebo) 을 주사하였다. 그 후 피험자들에게 시력검 사를 위해 대기실에서 기다리게 하는 동안 바보스럽고 장난끼 있는 행 동을 하는 즐거운 장면으로 꾸며진 분위기와 이와는 반대로 모욕적인 행동을 함으로써 몹시 화가 나게 하는 장면으로 꾸며진 분위기에 나누 어 대기시켰다. 실험 결과 아드레날린의 약물효과에 대해 올바론 정보 를 제공받은 피험자들의 정서상태는 대기실 분위기의 영향을 거의 받 지 않았으나, 거짓정보를 받았거나 아무런 정보도 받지 못한 피험자들 의 정서경험은 대기실의 분위기에 따라 많은 변화가 있었다. 이 실험 결과를 해석하기는 어렵지만 정서의 인지-각성이론을 지지할 만한 충 분한 근거를 제공하였다. Marshall 과 Z i rnbardo (1 979) 는, Schach t er 와 S i n g er 의 이론은 정서상태에서 신체의 역할에 대한 많은 연구를 자국 하는 데 큰 공헌을 했지만 이 이론은 극히 제한된 의미에서 타당한 것 으로 간주된다고 하였다. 2 정서와 자율신경 계통 행동의 강도는 자율신경 계통에 지배받는다. 교감신경 계통은 신체 로 하여금 보다 강력하고 왕성하게 활동하도록 준바시키며, 부교감신 경 계통은 소화 활동을 증가시키고, 이완과 관련된 신체 활동을 증가 시킨다. 부교감신경 활동을 자극하는 요인은 교감 활동을 촉발시키는 자극의 제거이다. 다시 말해 교감 활동의 자극을 갑자기 제거하면 부 교감계통의 과잉활동성이 반동적으로 뒤따른다. 스트레스 이후 휴식기 간 동안에 위궤양이 찰 발생하는 것은 바로 이런 이유이다. 다음에는 교감신경항전에 따른 신체 변화를 알아보는 거짓말 탐지 , 위궤양 발생 기제, 기타 정신신체질환의 발생 원인을 살펴보자.
2.1 거짓말 탐지기 생리적 각성과 정서경험 사이의 이론적 논쟁은 아직 해결되지 못한 상태지만, 생리적 각성을 측정하는 기법들이 발전되어 거짓말을 탐지 할 수 있게 되었다. 소위 거짓말 탐지기 (lie dete c to r ) 또는 다원기록법 (po lyg rap h ) 이 란 검 사는 범 죄 피 의자를 조사한다든가 유능한 고용인을 선발하기 위한 면접시험에 이용되고 있다. T i ner(l983) 에 의하면 미국 의 경우 매년 50 만 명 이상이 취직, 승진 또는 계속적인 고용계약을 위해 이 검사를 받는다고 하는데, 이 검사의 논리적 가정, 검사과정, 그리고 타당도 등을 알아보자. 다원기록법 검사는 두 가지 중요한 가정 아래서 출발한다. 즉, (1) 정 서상태에서 관찰되는 생리적 각성은 불수의적인 것이다. (2) 죄의식을 가진 자만이 범죄에 관련된 면접을 할 때 생리적 반응을 보일 것이다. 비록 이 두 가정에 대해 확고한 신뢰를 갖는다는 것은 어려운 일이지 만 현실적으로 이 겁사는 널리 사용되고 있다. 다원기록검사법에는 많 은 변형이 있지만 주로 통제질문법 (contr ol que sti on t echn iq ue) 이 많이 사용된다. 통제질문법에서는 검사하기 전에 다양한 사전 면접이 이루어지는데 , 이 사전 면접은 검사자와 피험자 사이에 협동십을 형성시키기 위한 질 문들이 포함된다. 검사시기에는 세 가지 유형의 질문이 주로 제시되는 데, 첫째는 부적절한 질문(예 : 당신의 이름을 어떻게 쓰느냐?), 둘째는 통제질문으로 현재 문제시되는 범죄사건과 직접 연관은 없지만 과거 범죄 사실의 유무를 묻는 것(예 : 당신은 전에 폭행사건으로 피소된 적은 없는가?)이며, 셋째는 관련된 질문으로 현재 문제시되는 사건과 직접 관련되는 질문(예 : 당신은 4 월 7 일 밤 이 사람을 폭행했지 ?)이다. 이 세 가지 질문 중 두번째 단계인 통제질문을 어떻게 구성하는가 하는 문제 가 매우 중요하다. 죽, 이 통제질문은 피험자에게 그 사건과의 관련성 여부를 묻는 것으로 때때로 피험자로 하여금 거짓말을 하게 할 수도 있다. 여러 차례에 걸쳐 질문을 하되 예상할 수 없도록 무선적으로 문
제를 제시하면서 질문이 제시되는 동안 신체반웅들을 측정한다. 만약 피험자가 통제질문보다도 관련질문에 대해 계속 생리적 각성이 더 높 으면 그가 거짓말을 하고 있는 것으로 가정할 수 있다. 반면 관련 질 문보다 통제질문에 생리적 반응이 더 높게 나오면 그는 거짓말을 하고 있지 않다고 가정된다. 만약 특정한 생리적 반응양식이 나타나지 않으 면 이 결과들은 무관한 것으로 간주된다. 2.2 자율신경계와 위궤양 부교감신경계의 과잉 활동에 의해 지나치게 증가된 소화액 분비는 위나 십이지장벽에 있는 세포들에 손상을 주어 이 벽둘이 헐게 되는 데, 이를 궤양 (ulcer) 이라 한다. 실직과 같은 큰 정서적 경험은 위궤양 울 일으킨다고 알려져 있다. 그러나 인간에서 위궤양 발생을 실험적으 로 알아보기 어렵기 때문에 이 문제는 주로 동물 실험을 통해 연구되 어 왔다. 스트레스와 위궤양 발생에 관한 실험적인 연구는 1950 년대 후반 Brady 등에 의한 원숭이의 실험에서 비롯되었다 (Brad y, Port er, Con- rad & Mason, 1958). 이들은 한 쌍의 원숭이를 각기 개별적으로 의자 에 앉혀놓고, 한쪽 다리에 전극을 부착하여 이 전극을 통해 다리에 전 기충격을 가할 수 있도록 했다. 두 마리 원숭이 가운데 한 마리는 이 른바 집 행 원숭이 (executi ve monkey) 라 하는데 , 이 원숭이 가 매 20 초 마다 자기 앞에 있는 지레를 누르면 자기 자신은 물론 동료 원숭이도 충격을 피할 수 있지만, 20 초를 지나 지레를 누르면 두 마리 모두 전 기충격을 받게 된다. 제 2 의 원숭이는 전기충격을 능동적으로 통제하지 못하는 수동 원숭이(p ass i vemonke y)이다. 실험은 하루에 두 번씩, 한 번에 6 시간씩 일주일 동안 계속했다. 실험 첫 회기의 처음 1-2 시간 이 내에 거의 모든 집행 원숭이는 지레를 눌러 전기충격을 피하는 반응을 학습했고, 그 후 실험이 끝날 때까지 계속 성공적으로 전기충격을 회 피하였다.
실험결과에 의하면 집행 원숭이만 위궤양을 보였을 뿐 수동 원숭이 는 위궤양을 보이지 않았다. 왜 이런 결과가 나왔을까? 집행 원숭이 가 더 많이 긴장했기 때문에 위궤양이 더 많았을까? 이 실험의 설계 가 불완전했기 때문에 이 결과를 신뢰하기 어렵다. 왜냐하면 원숭이를 집행과 수동으로 할당할 때도 무선 선정한 것이 아니라 전기충격에 대 한 회피반응의 학습능력에 바탕을 두고 할당했기 때문에 회피반응을 재빨리 학습한 민감한 동물이 위궤양에 쉽게 걸렸다는 가능성을 배제 할반수 없었다. 이런 문제점을 배제하기 위해 Fol t z 와 Mi llet t (1 964) 는 수동 원숭이는 그대로 두고, 집행 원숭이가 성공적인 회피반응을 완전히 학습했을 때, 새로운 집행 원숭이로 바꾸어 수동 원숭이를 계 속 흥분시켰다. 이 실험에서는 집행 원숭이보다 수동 원숭이가 위궤양 을 더 많이 보여주었다. 따라서 위궤양은 단순한 업무집행 때문이 아 니라 보다 심한 정서적 각성경험 때문에 발생한다. 앞서 Brad y의 실 험에서 집행 원숭이는 최소한 6 시간 동안 쉬지 않고 정서적 각성을 경 험하기 때문에 궤양이 생겼고, Fol t z 와 M ill e tt의 실험에선 수동 원숭 이가 더 많은 전기충격을 받고 흥분했기 때문에 궤양에 더 많이 걸린 것이다. 원숭이의 실험과 유사한 궤양실험이 흰쥐를 대상으로도 실시되었다. 원숭이와는 달리 집행 쥐는 계속하여 성공적으로 전기충격을 피하지 못한다. 쥐의 경우 수동 쥐는 전기충격을 통제할 수도 없고, 전기충격 의 시작을 예견할 수도 없어서 집행 쥐보다 위궤양에 더 많이 걸린다 (Weis s, 1968 ; 1971 a, b). 이런 사실로 미루어보아 분명히 위궤양 발생 은 전기충격 그 자체의 수용에서 오는 것이 아니라, 수동적 쥐의 경우 처럼 적절한 대응반응을 하지 못한 데서 비롯될 가능성이 높다. 위궤양은 스트레스를 직접 받는 시기보다는 스트레스기가 지난 후의 휴식시기에 잘 나타난다. 스트레스를 직접 받는 동안은 교감신경계가 활성화되는 시기이며, 그 이후의 휴식시기에는 부교감신경계가 과잉 활동하는 시기이다. 이처럼 부교감신경계가 과잉 활동을 하기 때문에 소화액이 정상보다 많이 분비되어 위나 장의 벽을 이루는 세포들을 손
상시켜 궤양이 일어나게 된다. 위궤양 발생에는 자율신경계를 조정하는 대뇌변연계, 특히 편도체의 역할이 중요한 것으로 간주된다. 즉, 이 부위를 손상시키면 구금에 의 한 위궤양 발생이 감소하고 (Henke, 1980), 갈등-스트레스 조건 아래서 의 위궤양 발생도 억제된다(이종택과 김기석, 1990). 한편 위궤양 발생 에는 도파민과 같은 신경전달물질이 깊이 관여하는 것 갇다. 흰쥐를 대상으로 한 일련의 연구에서 저자들은 할로페리돌 (Halo peri dol) 과 갇 은 도파민 길항물질을 편도 중심핵에 주입하면 위궤양 발생이 촉진되 지만, 도파민이나 도파민 효능물질인 아포모르핀 (a p omo rp h in e) 을 처치 하면 위궤양 발생이 억제된다는 사실을 발견하였다(장현감, 강성군, 배 중철, 1990 ; 장현감과 윤병수, 1991 ; 장현감, 윤병수, 박순권, 1992). 2.3 자율신경계와 기타 정신신체질환 대부분의 사람들은 나름대로의 안정된 자율반응성을 유지한다. 예컨 대 교감신경의 반응성이 매우 높은 사람은 보통 사람보다 자극에 민감 하고 재빠르게 반응한다. 즉 이들은 정서적 표현을 많이 하고, 활동성 이 높고, 대인관계가 복잡하고 혼란스러우며, 침착하지 못한 경향이 있다. 다시 말하면 교감신경계의 반응성은 흔히 말하는 성격과 많이 관련되어 있다. 앞서 언급한 위궤양 의에도 자율신경계 활동과 연관된 많은 질병들 이 있다. 스트레스에 대해 높은 교감신경 반응을 보이는 사람은 심장 병 에 도 잘 걸 릴 가능성 이 높다 (Kran t z & Manack, 1984) . 특히 통제 불 가능한 스트레스는 종양의 성장을 촉진시킨다고 하는데 (Sklar& Anis m an, 1981) , 그 이유는 스트레스가 아드레 날린의 분비를 증가시켜 결과적으로 면역체계의 약화를 초래하기 때문일 것으로 생각된다 (Axelrod & Reis in e , 1984) . 예 컨대 의과대 학생들이 시 험 기 간 동안 스트 레스를 경험할 때 인터페론(i n t e rf erone) 이나 자연살해세포 (na t ural kil ler cell: NK) 와 같은 면역체계의 두 활성인자들의 혈액 내 수준이
감소된다는 사실이 보고되었다 (Glaser 등, 1986). 교감신경계는 흉선이 나 임파절(lymp h nodes) 과 같은 면역계에 직접 영향을 미치기도 한다. 스트레스를 받으면 부산피질에서 코르티코스테로이드 (co rti cos t ero i d) 라 는 호르몬이 분비되고, 이것이 면역기능을 억제시킨다는 실험결과 (Balow & Rosenth a l, 1973) 나 면 역 체 계 와 내 분비 체 계 사이 에 양방적 연 락망이 구성되어 있다는 견해 (Weig en t & Blalock, 1987) 등도 위의 추 측을 지지해 준다. 우리가 일상생활에서 경험하는 다양한 스트레스가 면역기능에 부정 적 영향을 미쳐 질병을 유발시키는 것 같다. 배우자와의 사별이 질병 에 대한 이환율 및 사망률의 증가와 관련되고 (Parkes & Brown, 1972), 시험 스트레스를 많이 받는 의과대학생들이 시험 직전이나 도중에 저 하된 임파구 증식반응을 보인다고 한다 (Dor i an, Key st o n e, Garfin k le & Brown, 1982). 또한 부모와 친밀감이 낮다고 지각하는 사람들에서 악 성종양이 더 빈번히 발생되고, 지나치게 정서 표현을 억제하는 사람들 의 경우에도 암에 대한 면역학적 방어기능이 저하된다. 3 공격성 공격 성 (agg res sio n ) 은 인류의 진화 역 사에서 매 우 중요한 역 할을 담 당한 것이 사실이지만, 만약 현대사회에서 공격성이 없었다면 더 좋은 사회가 될 수 있었을 것으로 생각하는 사람도 많다. 여러 가지 형태로 나타나는 개인 대 개인 사이의 공격성은 이에 관여하는 이들에게 정서 적으로나 신체적으로 큰 상처를 안겨준다. 여기서는 공격성의 몇 가지 형태, 뇌 손상과 공격성 그리고 호르몬과 공격성 등에 관해 살펴보기 로 하자 .
3.1 공격성의 형태 공격성은 여러 가지 상이한 상황에서 일어나며 공격성의 형태도 매 우 다양하다. 대부분의 생물심리학자들은 상이한 형태의 공격적 행동 은 상이한 산경 실체와 호르몬에 의해 지배받는다는 데 동의한다. 따 라서 공격성을 분류하기 위해 여러 가지 상이한 분류체계가 제안되었 다 (Blanchard & Blanchard, 1984). 흰쥐에서의 뇌 손상과 호르몬 투여 가 공격 행동에 미치는 영향을 설명하기 위해 Albe rt와 그의 동료들에 의해 제안된 세 가지 분류도식을 먼저 살펴보기로 하자 (Albe rt & Chew, 1980 ; Albert & Walsh, 198.4 ) . 1) 방어적 공격성 (defe n siv e agg res sio n ) : 방어적 공격성은 지각된 위협 에 대해 직접적 반웅으로 일어나는 공격성을 의미한다. 쥐의 경우 방어적 공격성은 실험자가 쥐의 몸에 유해한 자국을 가하거나 이를 붙잡을 때 나 타나는 반응을 측정하여 점수화한다. 일반적으로, 사람의 손을 타지 못했 거나 또는 손은 탓지만 방어적 공격성을 증가시키는 뇌 수술을 받은 쥐는 실험자의 접촉 자극에 대해 도약반웅, 소리 지르는 반응 또는 무는 반응 등을 찰 나타낸다. 이 쥐들은 흔히 등을 바닥에 붙이고 방어적 자세를 취 하며, 발로 차고 할퀴고 물어뜯는 반응을 보인다. 이러한 방어자세는 다 른 쥐의 공격에 대항할 때도 찰 나타나는 행동이다. 2) 약탈적 공격 성 (pre dato r y agg res sio n ) : 약탈적 공격성은 다른 종에 속하는 동물을 잡아먹기 위해 몰래 접근하여 죽이는 행동을 의미한다. 야 생 둘쥐는 개구리와 같은 작은 동물을 잡아먹는 데 이때 쥐는 개구리의 목덜미를 물어 죽인다. 3) 사회 적 공격 성 (soc ial agg res sio n ) : 사회적 공격성 이 란 사회 적 위 계 를 이루거나 이를 바꾸려 하거나 또는 지속하려는 목적으로 상대방 동물 에게 별 까닭 없이 나타내 보이는 공격성을 의미한다. 이 행동은 쥐의 경 우 암수가 공존하는 무리 속에처 수컷들 사이에서 흔히 나타나는 행동이 다. 수컷 쥐의 사회적 공격성은 그들의 세력권 속에 낯선 수컷울 투입하 여 나타나는 세력권 방어행동이나 음식물을 쟁탈하기 위해 서로 싸우는
행동을 동해 측정한다. 앞서 든 세 가지 유형의 공격성과 관련 있는 뇌 구조는 다음과 같 다. 죽, ® 의 측 중격 (lat e r al sept um ) , @ 봉선 핵 (raph e nuclei) , @ 내 측 시 상하부 (media l hyp o th a lamus) , © 의 측 시 상하부 (lat e r al hyp o th - alamus), @ 편도체 (amy gd ala) 등이다 (P i ne l, 1990). 3.2 뇌 손상과 공격성 위에 예를 든 뇌구조를 양측성으로 손상시키면 방어적 공격, 약탈적 공격 행동에 영향을 미친다. 예컨대 의측 중격, 내측 시상하부 또는 봉 선 핵 의 손 상 은 방 어 적 공 격 성 (Brady & Nauta , 1955 ; Malsbury , Kow & Pfa ff, 1977) 과 약탈적 공격 성 (Albert 등, 1986 ; Marqu e s, Mais bury & Daood, 1979 ; Penot 등, 1978) 울 증가시 키 지 만 , 의 축 시 상하부 와 편도체의 손상은 위의 두 공격성을 약화시킨다 (Woods, 1956 : Ship le y & Kolb, 1977). 이처럼 방어적 공격성과 약탈적 공격성을 매개 하는 뇌 구조는 각기 다른 것 같다. 그러나 뇌 손상이 사회적 공격성에 미치는 효과는 방어적 공격성이 나 약탈적 공격성에 미치는 효과와는 상이하다. 예컨대 의측 중격, 내 측 시상하부 및 봉선핵의 손상은 수컷의 사회적 공격성을 약화시킨다 (Albert , Dyso n & Walsh, 1987 ; Blanchard, Blanchard & Takahashi, 1977) . 방어적 또는 약탈적 공격성과는 달리 많은 종에서 사회적 공격성은 전적으로 수컷에서만 나타나고, 데스토스태론과 같은 남성 호르몬에 크게 의존한다. 예컨대 수컷을 거세하면 한 무리 내에서 보여주던 사 회적 공격성, 즉 사회적 위계상의 우세성이 상실된다 (Albe rt 등, 1986 ; DeBold & Mic z ek, 1984). 그러나 거세된 수컷에게 데스토스데론 울 투여해 주면 상실된 우세성을 되찾게 된다. 위에서 Albe rt가 언급한 세 가지 공격성 이의에도 어미가 새끼를 보 호하기 위해 공격을 가하는 모성적 공격성 (ma t ernal a ggr ess i on) 도 있다
(Svare, 1977). 비록 모성적 공격성은 방어적 형태를 많이 취하지만 사 회 적 공격 성 과 흡사하고 (Albert 등, 1987) , 중격 을 손상하면 다른 형 태 의 방어적 공격성은 증가되지만 모성적 공격성은 증가되지 않는다 (Flannelly 등, 1986) . 3.3 테스토스테론과 공격성 많은 종류의 동물에 있어 사회적 공격성은 암컷보다 수컷에서 더 많 이 나타나는데, 이것은 테스토스데론이 활성화되기 때문이다. 수컷에 서 출생 직후 짧은 기간 나타나는 테스토스테론의 분비는 수컷에 걸맞 는 신경 계동을 발달시키고, 나아가 사춘기 이후 높은 수준의 데스토 스데론을 활성화시키고, 공격행동을 높이게 된다. 이런 문제는 출생 직후 거세한 생쥐 수컷과 암컷에게 테스토스데론을 투여한 후 사춘기 가 지난 후 다시 테스토스테론을 주사해 주면 공격성이 유의미하게 증 가된다는 사실에서 입증되었다. 인간의 경우는 테스토스데론과 공격성 사이에 명확한 관련성이 없 다. 이 의문은 몇 가지 방식으로 연구되어 왔다. 예컨대 피험자들의 공격성 수준을 먼저 측정한 후 혈액을 체취하여 혈장 속의 데스토스데 론을 측정하여 공격성과 데스토스테론 측정치 사이의 상관을 알아본 연구들이 있다. 즉 정상적인 사람의 혈액 내 데스토스테론 수준은 100ml 의 혈장 속에 300-1200mg 정도로 변산이 매우 크며, 또 하루 중의 측 정시간이나 피험자의 건강 정도 또는 스트레스의 정도에 따라 결과가 달라질 수 있다. 그러나 데스토스테론의 수준을 신뢰성 있게 측정하는 것이 불가능하지는 않다. 이 테스토스데론을 정밀하게 측정하기 위해 서는 실험설계가 타당해야 하고 통제집단 선정에 주의해야 하며, 실험 조건의 통제가 찰 이루어져야 한다. 최근에는 이런 조건을 만족시키는 연구들도 많이 보고되고 있다. 공격성을 측정하는 것은 테스토스테론 수준을 측정하는 것 이상으로 어려운 점이 많다. 두 가지 기본적 방법이 알려져 있다. 죽, 첫째 방
법은 Buss-Durkee (1 957) 의 공격성 검사와 같은 자기보고식 평가법이 다. 이 검사는 정상집단의 피험자에게는 사용될 수 있지만, 심한 범죄 를 저지른 죄수들의 공격적 경향성을 측정하는 데는 어려움이 많다. 공격성 평가의 두번째 방법은 동료에 의한 평가, 간수에 의한 평가, 구속기록 등에 의한 평가이다. 또한 공격행동의 개념을 정의하기도 어 렵다. 예컨대 말로 하는 욕설이 음식먹기를 거부하는 행동보다 더 공 격적이라고 말할 수 있으며, 무장강도가 강간범보다 더 공격적이라고 할 수 있을까? 이와 갇은 의문은 해결하기도 어렵고 연구 성과도 미 미하다. 이런 많은 난점에도 불구하고 혈장 내의 테스토스테론 수준과 공격 성 수준 사이에 어떤 상관이 있는가를 알아보는 연구들이 수행되어 왔 다. 일반적으로 정상적인 성인 남자를 대상으로 한 연구에서는 이 두 변인 간의 상관은 낮고, 유의미하지 않지만 정적인 상관을 보인다고 했 다 (Monti , Brown & Corriv e au, 1977 ; Rose, 1978 ; Rubin , 1 982) . 한편 수 감자를 대상으로 한 연구에서 수감자의 데스토스테론 수준과 Buss-Durkee 검사로 측정된 공격성 간에는 상관이 없다고 한다. 그러나 특 수폭력범(예컨대 강간범)으로 기소된 범죄자는 다른 경미한 폭력범에 비해 데스토스데론의 수준이 유의미 하게 높다고 한다 (Rada , Laws & Kellner, 1976) . 그러 나 이 런 유형 의 연구는 아직 많지 않기 때문에 데 스토스태론이 공격성에 미치는 영향은 확실치 않다. 그럼에도 불구하 고 미국이나 유럽의 여러 나라에서 폭력범에게 데스토스데론의 수준을 낮추는 약물을 실제로 두여하고 있다는 사실은 주목할 일이다. 3.4 세로토닌과 공격성 1970 년대에 접어들면서 신경전달물질인 세로토닌 (sero t on i n) 의 분비 감소가 공격행동의 증가를 야기한다는 실험결과들이 제시되어 주목을 끌었다. 이 분야의 연구논 이탈리아의 Valze lli에 의한 일련의 연구가 두드러진다. Valze lli는 어떤 특정 혈통의 생쥐 수컷을 이유 후 4 주
동안 사회적으로 고립하여 사육하면 다른 수컷 생쥐에 대해 심한 공격 성을 나타낸다고 하였다 (Valze lli, 1973) . 어 떤 혈통의 생 쥐 가 격 리 에 의해 공격성이 증가된 것은 뇌 속의 세로토닌 전환이 감소된 데 기인 한다는 것이다. 다시 말해 비록 뇌 속에 있는 세로토닌의 총량은 정상 적이지만 세로토닌의 시냅스 방출에 실패했거나 세로토닌의 활성 장소 에서 합성에 장애가 생기기 때문이라는 것이다. 세로토닌 방출을 억제하는 약물을 사용해도 공격성은 증가한다는 실 험 결과가 제 시 되 었다. 예 컨대 PCPA (pa rachloroph e ny la lanin e ) 라는 세 로토닌 합성을 억제하는 약물을 투여하면 암수 가리지 않고 모두 공격 적이 된다고 한다 (Valze lli, Bernasconi & Garatt ini, 1981). 일단 공격성 이 일어나기 시작하면 세로토닌 수준이 정상으로 돌아올 때까지 공격 성은 지속된다고 한다. 한편 세로토닌의 전구물질인 트립토판이 적은 음식을 먹이면 공격성이 증가하지만 트립토판이나 이와 유사한 화학물 질을 직접 주사하면 공격성은 오히려 줄어든다는 보고도 있다 (Broderic k & Brid g e r , 1984) . 인간 사회에서도 동물의 경우처럼 음식물이 공격성 발생에 영향을 미친다. Mawson 과 J acobs(1978) 에 의하면 옥수수: 주식으로 하는 문화권에서 살인율이 높다고 한다. 이 옥수수는 세로토닌의 전구물질 인 트립토판을 거의 갖고 있지 않기 때문에 옥수수를 많이 섭취하면 세로토닌의 합성이 낮아질 것으로 예측할 수 있다. 하지만 주식으로 옥수수를 섭취하는 것이 곧 공격성을 증가시킨다는 결론을 내리기에는 부적절하다. 4 쾌중추 생물심리학 연구에 있어 하나의 새로운 장을 열게 된 계기가 1950 년 대 캐나다의 맥길 대학에서 일어났다. 1954 년 Jam es Olds(1922-1976) 와 Pete r M il ner 는 「쥐의 뇌 중격과 기타 뇌 부위의 전기적 자극에
의 해 야기 된 정 적 강화 (Positi ve Rein fo rc ement Produced by Electr ic a l Stim u lati on of Sep ta! Area and Oth e r Reg ion of the Rat Brain ) 」 라 는 논 문을 발표하였다 (Olds & Mi lne r, 1954). 이 논문은 그 후 20 년 동안 뇌 의 쾌중추를 확인하려는 연구들을 자국하게 되었다. 대부분의 중요한 연구가 그러하듯 이 연구에서도 행운이 따랐다. 당시 이 대학의 박사 추수과정 연구원 이 던 Olds 와 M i lner 는 충동 (driv e ) 과 동기 (moti va ti on ) 라는 개념과 뇌간 망상체 기능과의 관련성에 관심을 가졌다. 그리하여 이들은 뇌의 각성계통이라고 추정되는 이곳을 전기적으로 자극하여 학 습에 미치는 영향을 연구하고 있었다. Olds 는 사회십리학 전공으로 박사학위를 받았기 때문에 쥐의 뇌에 대한 해부학적인 지식이 부족하였다. 그래서 쥐의 뇌에 전극을 심을 때 뇌간의 해부학적 위치를 잘못 계산하여 전극을 뇌간에 정확하게 삽 입하지 못했으며 게다가 삽입한 전극이 뒤틀리는 바람에 당초 목표로 삼았던 부위에서 몇 mm 떨어진 중격 부위에 삽입하는 실수를 범했 다. 이렇게 중격에 전극을 삽입한 채 자극을 가하게 되었는데 이처럼 중격이 자국된 쥐는 마치 이 자극을 적극적으로 수용받기를 원히는- 것 처럼 행동했다. 이 실험 상황은 동물이 마취에서 깨어난 자유로운 상태였으므로 심 어놓은 전극에 전선이 매달려 있는 채로 자유롭게 움직일 수 있었다. 이런 상황에서 Olds 는 피험동물이 자극을 받은 장소로 되돌아오는 경 향이 있다는 점을 발견하게 되었다. 그래서 그는 뇌에 가한 전기적 자 극의 강화특성을 알아보기 위한 몇 가지 검사를 개발할 수 있었으며, 그 결과 이런 강화 효과를 검증하기 위한 표준방법으로 조작행동 측정 상자를 이용할 수 있게 되었다. 이런 실험절차를 사용하여 Olds 와 그의 동료들은 이 현상에 관해 몇 가지 의문을 알아보려 하였다. 죽, 첫째, 인간도 동물처럼 이런 자극 울 추구하게 될까? 만약 그러하다면 이 자극을 받았을 때 어떤 느낌 이 들까? 둘째, 이 자극은 언제나 쾌감만을 야기하게 될까? 뇌의 어 느 곳을 자국하면 불쾌감이 야기될까? 셋째, 이런 전기자극의 강화
속성은 음식, 물 또는 성적 자극과 같은 자연적인 자국을 받았을 때 나타나는 강화 효과와 유사할까 ? 이런 의문을 알아보는 연구들 가운데 인간의 뇌를 직접 전기자극한 연구보고가 등장하여 홍미를 끌었다. 과연 인간의 뇌를 전기적으로 자 국하는 것이 가능하며, 이와 동시에 환자와 대화할 수 있을까? 사실 이런 자극방식은 간질이나 정신분열증과 같은 질병의 전단이나 치료를 위해 실제 실시되고 있는 방법으로서 특별히 어렵거나 위험하지 않다. 뇌는 그 자체가 전기적 자극을 받아도 통각을 받아들이는 수용기가 없 기 때문에 이미 심어놓은 영구 전극을 통해 자극을 가해도 고통을 느 끼지 않는다. 그러나 이런 자극을 받은 피험자의 언어반응을 해석한다 는 것은 매우 어려운 일이다. 비록 인간의 뇌에 쾌영역이 존재한다는 몇 가지의 단편적 증거가 있 지만 아직 이 증거를 해석하는 데는 많은 문제점이 있다. 인간의 행동 과 인간의 뇌 모두가 매우 복잡하기 때문에 비록 직접적인 증거가 될 수 있는 것처럼 보이는 것조차도 실험적 상황, 환자의 성장 배경이나 성격 특칭과 같은 맥락에 따라 조심스럽게 평가하지 않으면 안 된다. Olds 와 M il ner 의 연구보고가 있었던 바로 그 해 Del gado 를 중심으 로 하는 다른 집단의 연구자들은 고양이를 대상으로 뇌간의 시개영역 (tec ta l area of brain s te m ) , 시 상 및 해 마에 전기 적 자국을 가하면 공포 와 같은 반응이 일어난다는 사살을 관찰하였다 (Del g ado, Robert s & Mi ller, 1954). 그 이후의 연구들에 의하면 포유류의 뇌에는 전기적 자 극에 대해 긍정적, 부정적 또는 중성적으로 반응하는 세 가지 뇌 조직 이 있다는 사실이 알려지게 되었다. 그러나 궁극적으로 대부분의 뇌피질과 소뇌는 전기자극에 대해 중성 적인 반응을 하는 특칭이 있다. 죽 이곳에 전극이 삽입된 동물은 쾌자 극을 받기 위해 지레를 누르지도 않으며 자극을 피하기 위해 지레를 누르지도 않는다. 대부분 뇌의 정서야기 중추는 변연계, 시상하부 그 리고 뇌간이다. 이곳은 뇌피질에 비해 계동발생적으로 오래된 뇌 부위 로서, 이 부위를 자극하면 보상효과를 보여주는 것이 일반적이지만
5% 정도는 처벌효과도 보여준다. 죽, 동물은 이 부위의 자극에 대해 회피성 반응을 보여준다. 특히 이런 처벌효과는 전극이 중뇌의 복측 영역이나 통각과 관련 있는 신경충격을 신체 부위로부터 중추로 전달 하는 신경 섬 유다발인 내 측융대 (media l lemnis c us) 에 위 치 할 때 잘 관찰 된다. 한편 정 적 인 보상효과가 일어 나는 곳은 내측전뇌속 (media l for ebrain bundle) 인데, 이 구조는 뇌의 복측면을 따라 전뇌로 향하는 거대한 축 색다발로 이루어져 있다. 정적인 보상효과가 일어나는 뇌 부위를 전기 적으로 자극하면 자연상태에서 일어나는 보상효과와 유사한 효과가 나 타날까? 이 의문을 풀기 위해 실험실 쥐를 대상으로 많은 연구가 이 루어졌다. 어떤 뇌 영역이 정적 보상효과와 분명하게 관련된다는 증거 는 제시되어 있지만 보상효과를 나타냈다는 증거를 제시해 주지 못하 는 뇌 영역도 있다. 예컨대 의측 시상하부의 자기자극은 기아감을 증 가시 키 지 만 섭 식 행 동은 오히 려 감소시 킨다 (Groves & Schlesin g e r , 1982) . 자기자극을 받은 쥐는 실제 음식을 취하기보다 전기자극을 더 선호하 는 것처럼 행동한다. 왜냐하면 자기자극과 섭식 중 어느 하나를 선택 할 수 있는 상황 하에서, 자기자국에 의해 음식이 계속 제공되어도 쥐들은 자신의 체중이 정상 이하로 떨어질 때까지 음식은 취하지 않 고 자기자국만 받으려 하기 때문이다 (Rou tt enber g & Lin d y, 1965). 또 한 자기자국과 갈증 해결 또는 배우자 접근 간의 선덱을 허용해 준 상태에서도 위와 유사한 결과가 관찰된다.
제 12 장 학습과 기억 학습과 기억의 신경 바탕을 연구하는 분야는 신경과학의 여러 연구 분야 가운데 가장 중요한 연구분야의 하나로 간주된다. 1950 년대부터 지금까지 이 분야에 대한 많은 연구가 이루어져 왔다. 특히 최근 몇 년 사이에 이루어전 이 분야의 연구를 살펴보면, 종래의 실험실 쥐 중 심의 연구로부터 토끼, 고양이, 원숭이와 같은 고등 포유류나 가재나 군소 (A p l y s i a) 와 같은 무척추 하등동물 그리고 인간을 대상으로 한 연 구에 이르기까지 다양한 유기체를 대상으로 연구가 진행중이다. 사람을 대상으로 한 연구는 주로 기억상실과 관련 있는 건망증 환자 를 주된 연구 대상으로 삼아왔다. 이들을 대상으로 한 연구로부터 인 간 뇌의 조정기능이라든지 기억의 저장과 인출을 담당하는 뇌 부위를 알아내는 데 큰 도움을 받았다. 한편 군소나 가재, 쥐 또는 토끼를 대상으로 한 연구로부터 학습이 나 기억에 관여하는 신경원의 구체적 실체를 밝히게 되었고, 나아가 학습과 관련된 시냅스 구조와 기능변화 등도 알아볼 수 있게 되었다.
l 학습과 기억의 세포학적 바탕 학습과 기억의 생물학적 연구에 관심을 가진 과학자들은 인간의 건 망증뿐만 아니라 동물의 학습과 기억에도 많은 관심을 기울였다. 최근 에 들어와서는 바다에 서식하며 매우 느리게 움직이는 무척추동물인 군소와 같은 단순한 유기체를 대상으로 하여 학습의 기초를 연구하고 있다. 이 군소는 소수의 신경원으로 구성되어 있으며, 이들의 신경원 은 척추동물의 신경원에 비해 훨씬 더 크기 때문에 보다 쉽게 다룰 수 있는 이점이 있다. 또한 이런 단순 신경계는 학습이 일어나고 있는 동 안 신경 계통에서 어떤 변화가 일어나고 있는지 알아보는 데 매우 중 요한 단서를 제공해 줄 수 있다. 일반적으로 이런 연구들로부터 세 가 지의 기본 학습현상, 즉 습관화 (hab it ua ti on), 민감화 (sens iti za ti on) 그 리고 고전적 조건화 (class ic al cond iti on i n g)가 성공적으로 연구되었다. 1.1 습관화 흔히 습관화 (hab it ua ti on) 를 〈특별한 주의를 하기 전에는 구두 속에 발이 꽉 차 있다는 것을 알지 못하는 것과 유사한 개념〉이라는 비유를 든다• 그러나 이런 말을 듣는 순간 우리는 구두가 발을 조금 조이고 있다는 느낌을 가져 보지만 이에 대한 주의는 곧 없어지고 만다. 습관 화에 관한 또 다른 예로는 시계가 째깍째깍 하는 소리나 에어컨이 작 동되는 소리를 듣는 경우이다. 습관화는 많은 종의 동물들에서도 관찰 될 수 있으며, 모든 학습에서 공통적이면서 원시적 형태이기도 하다. 습관화는 의미 없는 자극에는 반응하지 않도록 하는 학습이며, 매시간 정각에 울리는 괘종 소리와 감은 반복적인 자국에는 습관화가 되지 않 는 특성도 있다. 습관화는 단순한 피로현상이 아니다. 습관화는 반응의 피로를 야기 하는 데 필요한 자국의 반복률보다 훨씬 낮은 자극의 반복률 아래서도 관찰될 수 있는데, 자극이 조금만 바뀌어도 자국에 대한 반응이 쉽게
되살아난다. 이런 현상을 탈습관화 (d i shab it ua ti on) 라고 한다. 탈습관화 롤 야기하는 하나의 효과적인 방법은 자극을 빠뜨리는 것이다. 만약 규칙적으로 배열된 일련의 자극들에 대해 습관화가 일어났을 때 규칙 적 간격에 있어야 할 자극이 없어지면 이 탈락된 자국에 대해 탈습관 화 반응이 일어나게 된다. 1.2 민감화 또 다른 형태의 원시적 비연합학습을 민감화 (sens iti za ti on) 라 한다. 동물을 잠재적인 위협자국이나 유해한 자극에 노출시킨 후 일정한 시 간이 지나 새로운 자극을 제시하면 이 자극에 대한 반응의 강도가 증 가하게 되는 것을 볼 수 있는데 이것을 민감화라 한다. 공포영화의 제 작자들은 민감화의 위력을 잘 이용한다. 예컨대 어떤 배우가 이 영화 속에서 살인행위를 하게 된다는 것을 관객에게 미리 알려중으로써 영 화 속에서 이 배우가 행하는 일거수 일투족에 신경을 쓰게 한다. 1.3 고전적 조건화 파블로프 (Ivan Pavlov, 1849-1936) 는 러시아의 생리학자로서 처음에는 소화 계통의 기능에 관심을 가졌으며, 소화에 관한 연구로 1904 년 노 벨 생리학상을 수상받았다. 그는 피험동물인 개가 음식물이 입에 들어 가기도 전에 침을 홀린다는 사실을 관찰하였다. 그런데 이런 타액분비 현상은 반드시 실험실시 후 며칠 경과된 후에야 나타나는 것으로 보아 생리적 반응이 아닌 어떤 정신적 반웅, 즉 학습된 것이라고 믿었다. 그는 그 후 조건화를 탐구하는 데 나머지 생애를 다 바쳐 뇌의 신비를 탐색했다. 오늘날 비록 조건화의 뇌 기능에 관한 그의 이론은 고전적 홍미거리가 되고 말았지만, 그가 설정한 기본적인 실험기법은 지금도 세계 도처의 실험실에서 여전히 적용되고 있다. 고전적 조건화의 경우 학습은 음식을 보고 침을 홀리는 것과 같은
무조건반응 (uncond iti oned respo n se : UCR) 이 라고 부르는 본유적 반응과 종소리와 갇은 새롭게 학습된 자극, 죽 조건자국 (cond iti oned stim u lus : CS) 사이의 연합에 의해 나타난다. UCR 은 적절한 자극, 즉 무조건자 극 (uncondit ion ed stim u lus : UCS) 의 제 시 에 의 해 일어 난다. 전 형 적 인 파불로프 실험에서 UCS 는 입 속에 제시하는 고깃가루인데 이것이 UCR, 죽 타액반사를 야기한다. 이 고것가루를 종소리나 광선과 같은 중성적 자극과 밀접하게 연결시켜 몇 번의 시행을 실시하고 나면, 종 소리나 광선 자국만 제시하여도 조건반웅 (cond iti oned respo nse: CR), 죽 타액이 분비된다. 1.4 군소의 습관화, 민감화 및 조건화 컬럼비아 대학의 Eric Kandel 이란 신경과학자는 거대한 바다 달팽이 의 한 종류인 군소를 대상으로 파블로프의 조건화 기법을 적용하여 여 러 해 동안 학습실험을 실시했다• 비록 군소가 보여줄 수 있는 행동목록이 극히 제한되어 있지만 이 동물은 음식물을 찾아서 먹을 수 있고, 자신을 위협하는 침입자에게 혼란을 주기 위해 자신의 몸 속에서 보랏빛 잉크물을 내뿜어 자신을 보호할 줄도 알고, 생식행동도 보여주고 알도 낳으며 활동기와 휴식기 가 구분되는 행동주기도 보여준다. Kandel 은 군소의 아가미 수축반사 (gill wit hd rawal refl ex ) 의 습관화, 민감화와 조건화를 연구하여 학습의 신경기제를 밝혔다 (Kandel & Schwa rtz, 1982 ; Carew, Hawkin s & Kandel, 1983). 그립 12-1 에는 군소 의 모습을 나타내었는데, 이 동물은 아주 정교한 아가미(gill)를 통해 바닷물로부터 산소를 취할 수 있다.’ 이 아가미는 의두막 (man tl e shelf) 이란 막조직에 의해 보호되며, 이 의두막의 종말에는 육질로 된 홉수 관 (sip hon) 이 있다. 이 동물이 앞으로 나갈 때는 아가미, 의두막 그리 고 흡수관이 몸통으로부터 빠져나오게 된다. 흡수관이나 의두막에 가 벼운 촉각적인 자극을 가하면 흡수관, 의두막 그리고 아가미까지 모두
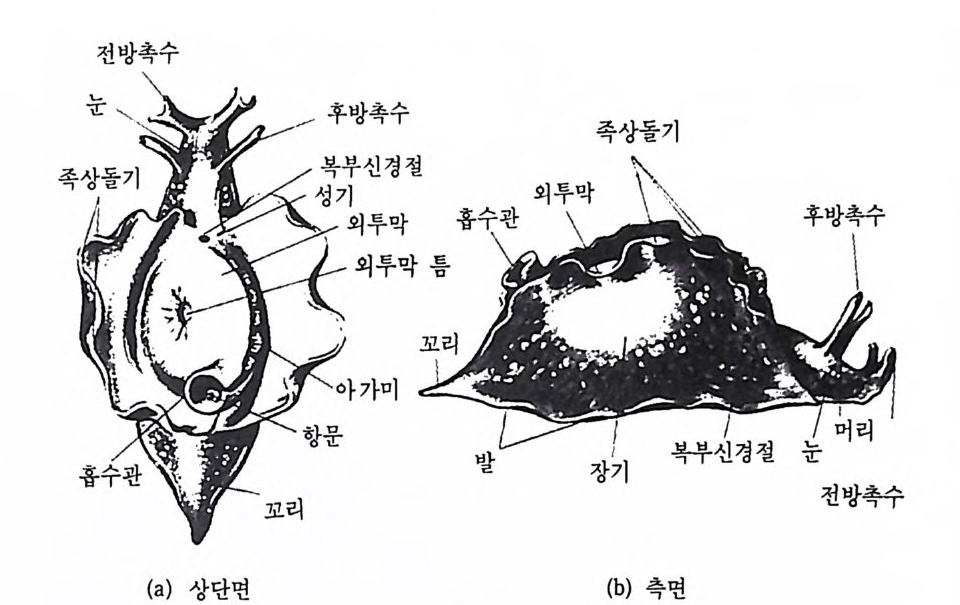 전방촉수
전방촉수
그립 12-1 군소의 모습 성숙한 군소는 길이가 30cm 나 된다. 움츠려들게 된다. 어떻게 이들 구조들이 자극에 의해 움츠려드는가를 밝힌다는 것은, 곧 이러한 반사행동에 관여하는 신경회로체계를 알아 내는 것으로서 그 동안 많은 연구가 수행되었다. 군소의 아가미 수축반사는 9 개의 신경절 가운데 복부신경절 (abdo rni nal g an gli on) 에 의해 주로 지배된다. 이 동물은 하나의 신경철에 약 2, 000 개 정도의 신경원이 있는데 이 모든 신경원 하나하나는 개별적으 로 구분될 수 있다. 이렇게 하나씩 구분된 신경원에 고유번호를 부여 할 수 있어서 어떤 것은 L28 신경원, 또 어떤 것은 M7 신경원 등으 로 구분하여 명명한다. 한 마리의 군소가 갖는 신경원의 총수는 약 20,000 개 정도에 불과하고, 이 동물의 신경회로도 비교적 잘 밝혀져 있고 신경원도 굵어 하나하나 확인될 수 있으므로, 이 동물의 아가미 반사의 습관화, 민감화 그리고 조건화에 관여하는 세포기제에 관해서 많은 정보를 얻을 수 있게 되었다. Kandel 과 그의 동료들은 수년 동 안에 걸친 실험 끝에 이 동물의 아가미 반사 학습에 관여하는 일차장
소가 이 반사를 조절하는 운동신경원들과 시냅스하는 감각신경원의 축 색 종말부에 있다는 사실을 밝혔다. 1.5 아가미 수축반사의 습관화 흡수관이나 의두막이 물을 뿜을 때면 아가미, 의투막 그리고 흡수관 모두 수축하게 된다. 이런 물의 분사에 대한 습관화는 10-15 시행 이내 에 이루어지며, 습관화 후의 수축반사의 크기는 습관화 이전에 비해 약 1/3 정도로 줄어든다. 이런 습관화는 1 시간 이상 지속되는데 Kandel 은 이것을 단기습관화 (sho rt-t enn hab it ua ti on) 라고 불렀다. 그러나 만 약 10 번의 자국제시 중 4 번 정도에서 습관화가 반복하여 일어나는 경 우에는 습관화가 몇 주일 동안 지속될 수도 있는데, 이를 장기습관화 (lon g -ten n hab itu a ti on) 라고 부른다(그림 12-2). 그 후의 실험에서 Kandel 은 이런 수축반사에 개재하는 운동 및 감 각 신경원에 미세전극을 삽입하여 습관화 과정을 알아볼 수 있었다. 수축반사에 관여하는 신경회로가 비교적 간단하기 때문에 이 반사에 관여하는 기본 신경원을 확인할 수 있고, 또한 실험적으로 조작할 수 도 있다. 흡수관에 반복하여 물을 분사시키면 아가미 수축을 야기하게 하는 운동신경원에서 신경충격의 발생 수가 감소된다. 이런 영향은 운 동신경원에서 흥분성 시냅스후 전위 (EPSP) 의 크기가 감소되기 때문인 데, 결과적으로 EPSP 가 줄어들어 운동신경원에 어떤 신경충격을 생 성시키지 못하게 되므로 반사가 일어나지 않게 된다. 한편 이때 감각 신경원을 중점적으로 관찰해 보면 습관화가 이루어질 때 Ca2+ 이온이 감각신경원의 축색종말로 유입이 줄어든다는 사실을 알 수 있다. 이런 현상은 감각신경원이 소량의 신경전달물질을 방출하게 되는 결과를 낳 게 된다. 이처럼 신경전달물질의 소량 방출은 EPSP 의 감소를 초래하 게 되고 운동신경원의 신경충격을 감소시키는 결과를 낳게 된다. 어떻게 반복적인 자극이 감각신경원의 축색종말에서 Ca2+ 이온의 유 입을 감소시키는지는 아직 잘 모른다. 그러나 습관화가 이루어전 동물
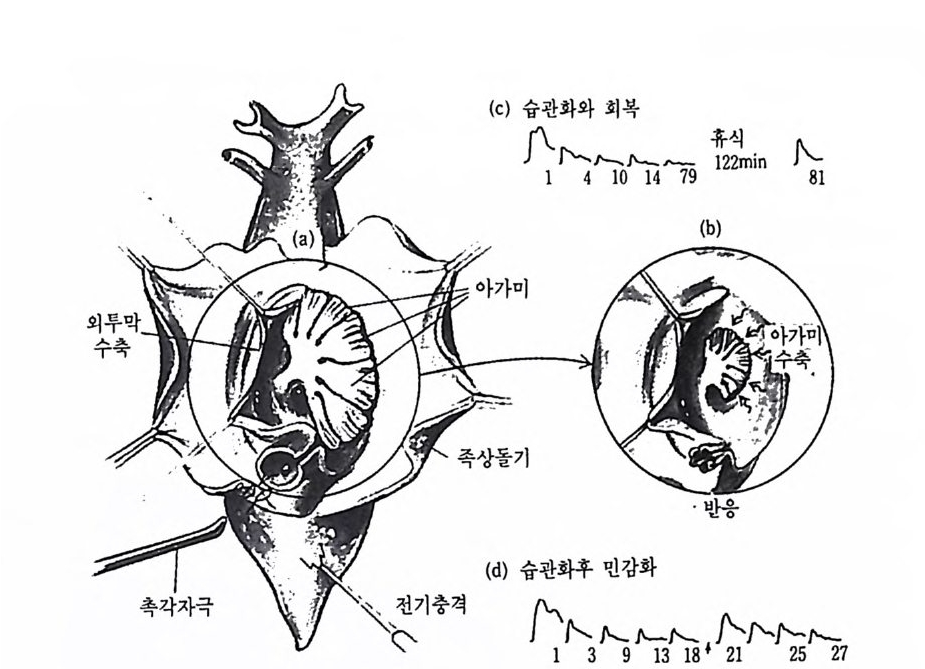 (c) 습관화와 회 복
(c) 습관화와 회 복
그림 12-2 군소의 아가미 수축반사를 연구하기 위한 실험조작 촉각 자국을 홉수관에 적용하면 반사가 일어난다. (a) 수축이 ·일어나기 전 열 려있는 아가미의 크기. (b) 부분적 수축이 일어난 후의 아가미. (c) 몇 번의 습 관화 시행 후 아가미 수축의 정도. (d) 단 한번 꼬리에 민감화를 야기하는 충 격을 준 후 몇 번의 시행 동안 보여주는 아가미의 크기. 의 감각신경원 축색종말에서 나타나는 형태학적 변화는 전자현미경을 통해 알아볼 수 있다. 이 연구에서는 습관화된 동물의 경우는 소위 활 성 영 역 (acti ve zones) 이 라 부르는 축색종말의 특수 영 역 (시냅스 주머니 가 위치하고 있고, 신경전달물질이 방출되는 영역)이 감각신경원의 축색종 말부에서 줄어든다는 사실을 발견하게 되었다 (Ba il e y & Chen, 1983). 이 사실로 미루어보아 반사의 습관화를 기억하는 장소는 아마도 운동 신경원과 시냅스를 이루는 감각신경원의 시냅스전 축색종말부가 아닐 까 생각된다.
1.6 민감화 아가미 수축반사의 민감화는 꼬리 쪽에 강한 전기충격을 가함으로써 야기할 수 있다. 일반적으로 꼬리에 전기충격을 가하면 거의 즉각적으 로 자극에 대한 수축반사가 확대되어 나타난다. Kandel 과 그의 동료 둘은 감각신경원이나 운동신경원에 미세전극을 삽입하여 이 반사의 습 관화와 동시에 민감화 과정도 연구했다. 민감화의 기간은 적용하는 전 기충격의 강도나 빈도에 따라 달라지지만 대체로 몇 시간에서 몇 주까 지 지속된다. 습관화의 경우처럼 민감화를 야기시키는 신경구조도 감 각신경원의 축색종말부라는 점이 밝혀졌다. 그러나 민감화는 여러 가 지 방식으로 습관화와 반대되는 현상을 보여준다. 죽 민감화는 운동신 경원의 EPSP 크기 증가와 관련 있으며, EPSP 크기 증가는 전달물질
 口머리 〈나꼬리
口머리 〈나꼬리
그림 |2-3 군소의 아가미 수축반사에 관여하는 신경회로도 SN24 = 감각신경원 24 ; Fl = 촉진성 간신경원 : El5 = 흥분성 간신경원 : MN6 = 운동신경원 6,
의 방출 증가에 의해 야기되며, 전달물질 방출 증가는 Ca2+ 의 유입 증 가에 의해 일어나며, 민감화에 따라 시냅스에서의 활성영역의 크기와 수도 늘어나게 된다. 습관화와 민감화 사이에 개재되는 신경회로상에는 중요한 차이가 있 다. 즉 민감화 과정 에 는 〈촉진성 간신경 원 (fac ili tat o r y int e m euron) 〉이 개재된다. 이 신경원은 꼬리자극에 의해 야기되는 충격을 감각신경원 에 전달하는데, 이 촉진성 간신경원의 종말은 다시 감각신경원의 축색 종말과 시냅스를 이룬다(그림 12-3). 1.7 아가미 수축반사의 고전적 조건화 군소의 아가미 수축반사는 고전적 조건반사 형태도 보여준다 (Carew 등, 1983). 이 연구들에서 CS 는 나일론 솔을 가지고 흡수판을 잠깐 동 안 자극하는 것이고, UCS 는 아가미의 수축반사를 일으키도록 꼬리에 전기충격을 가하는 것이다. 조건화가 이루어지기 전에는, 나일론 솔 자국은 단지 아가미에 미약한 수축반사만 보여주지만 꼬리에 대한 전 기충격 자극은 민감화를 유발하여 아가미의 수축반사를 포함하여 그 밖의 여러 후속 반웅들도 증가시킨다. 전형적인 조건화 절차에 따라 실험해 보면, 몇 주 동안 지속되는 조 건화는 최소한 15 시행 이내에 이루어진다는 사실을 알 수 있으며, 조 건화에 개재되는 세포기제는 민감화 기제와 유사하며, CS 와 UCS 사 이의 시간관계가 매우 중요하다는 점이 밝혀졌다. 예컨대 CS 를 UCS 에 앞서 제시한 집단(조건화 집단)과 CS 는 제시하지 않고 UCS 만 가한 집단(민감통제 집단) 그리고 CS 에 앞서 UCS 를 먼저 가한 집단(허위조 건화 집단)으로 나누어보면, CS 가 UCS 에 앞서는 조건화 집단에서만 유의미하게 조건화가 찰 일어났다. 가장 조건화가 잘 일어나는 경우는 CS 가 UCS 에 0.5- 1. 0 초 앞서는 경우이다. 이 결과를 정확하게 해석하기 위해서는 더 많은 연구가 필요하지만, 이 연구의 결과는 군소의 아가미 수축반사의 습관화, 민감화 그리고
고전적 조건화에 개재하는 세포기제를 탐색하는 데 매우 유용한 것이 다. 군소의 행동 특성과 이 동물의 신경회로가 비교적 간단하다는 장 점을 이용하여 연구자들은 이처럼 단순한 학습에 개재하는 세포 변화 에 대한 많은 지식을 얻을 수 있으며, 이런 현상을 인간의 학습에도 적용해 보려는 시도가 점차 본격화되고 있다. 2 척추동물의 뇌에서 일어나는 고전적 조건화 파블로프의 고전적 조건화가 척추동물의 뇌 속에서 일어난다는 사실 이 밝혀졌다. 이런 종류의 연구로서는 Ric h ard Thom p son 과 그의 동 료들에 의해 수행되어 온 연구가 대표적인 예가 될 수 있다. 이 연구 자들은 토끼의 순막 반웅의 고전적 조건화에 개재하는 신경회로가 뇌 속의 어떤 특정 영역에 존재한다는 사실을 밝혀냈다. 즉 이들의 주장 에 의하면, 이런 영역이 소뇌 , 해마 그리고 뇌간에 있다고 한다 (Thom p son, Berge r & Madden, 1983 ; Thomp so n 등, 1984 ; McCormi ck & Thomp so n, 1984). 이런 연구에서 관찰된 조건반응은 토끼나 고양이와 같은 동물이 가지고 있는 제 3 의 눈꺼풀인 순막 (n i c tit a ti n g membrane) 의 수축반응이다. 이들 연구에서 적용되는 UCS 는 눈의 각막에 공기를 분 사하는 것이며 CS 는 소리를 제시하는 것이다. 몇 몇 실 험 들 에 의 하면 소뇌 의 치 상-중 간 핵 (denta t e - in t e r po s itu s nucleus) 의 손상은 소리 에 대한 CR 을 소멸시 킨다고 한다. 이 핵 의 손 상은 학습된 반응은 방해하지만 UCS 에 의해 생성되는 무조건 반응인 눈깜빡임 반응은 방해하지 않는다. 김현태, 최준식 및 김기석 (1991) 은 소뇌의 중간핵을 전기적으로 자국하면 순막조건화의 습득은 지연되고 소거는 촉진된다고 하며, 김현태, 류재욱 및 김기석 (1991) 은 소뇌피질 의 단소영 (sim ple lobule) 이 순막조건반응의 출현시간을 조절하는 데 관 여할 것이라는 견해를 제시하였다. 죽, 이 결과는 소뇌(또는 소뇌 일부 분)가 순막반응과 같은 단순 운동학습에 관여한다는 것을 의미하며,
뇌간 주변에 있는 신경원들이 이런 단순 학습에 관여할 것이라는 주장 도 있다. 모든 종류의 학습에 소뇌가 관련된다고 결론지울 수는 없지만, 적어 도 소뇌가 척추동물의 안구반사와 같은 일차적 학습에 관여하는 주요 구조임에는 틀림없다. 또 다른 종류의 학습에 관여하는 신경원 집단들 이 뇌의 다른 부위에 존재할 가능성도 충분히 예견할 수 있다. 이처럼 학습의 종류에 따라 관련되는 신경계의 하위 실체를 찾아내는 것이 생 물심리학의 주요 연구과제이다. 한편 파블로프의 기본 조건화 기법에는 몇 가지 상이한 변형이 있 다. 예컨대 CS 와 UCS 를 시간적으로 겹쳐 제시할 수도 있고(이를 지연 조건화 dela y ed cond iti on i n g라 한다), 이 두 자극 사이에 공벽을 두고 제 시할 수도 있다(이 경우 흔적조건화t race cond iti on i n g라 한다). CS 와 UCS 사이에 약간의 시간 지체를 두면 동물은 CS 의 출현과 UCS 의 출 현 간의 사이 를 기 억 해 야 한다. Thom p son 과 그의 동료들 (1984) 은 눈 깜빡임 반사의 조건화에는 기억이 요구되는 흔적조건화를 알아보았더 니 해마가 결정적인 신경회로의 역할을 한다는 사실을 발견하게 되었 다. 즉 이들에 의하면 해마가 손상되면 지연 조건반응의 학습에는 장 애가 나타나지 않지만 흔적조건화 학습에는 현저한 손상이 나타난다고 한다. 따라서 학습의 종류에 따라 관여하는 뇌 부위도 달라지는 것 같다. 3 장기증강 앞서 언급한 바와 같이 Eric Kandel 과 Jam es Schwart z ( 1982) 는 군 소의 학습에 관여하는 결정적 신경 부위가 시냅스전 부위라고 했다. 특히 이들은 아가미 수축반사를 일으키는 운동신경원을 지배하는 감각 신경원의 축색종말에서 이런 결정적 변화가 일어난다고 믿었다. 그러나 Gary L yn ch 와 Mic h el Baud ry (1984) 는 이러한 학습이나 기 억에 관여하는 결정적 위치가 시냅스전 부위가 아니라 시냅스후 부위
일 것 이 라고 주장하였 다. 이 들은 장기 증강 (lon g -ten n pot e n ti at io n : LTP) 이라는 전위현상에 근거하여 이 가설을 제안했다. LTP 란 유발 전압을 일으키는 신경원의 구십성 통로에 강력한 전기충격을 짧은 시 간 동안 가하면, 그 이후는 표준강도의 전기적 자국에 의해서도 유발 전압의 크기가 더욱 증강하게 되는 현상을 지칭한다. 이때 유발전압이 란 어떤 구심성 통로를 활성화시킬 때 이와 관련 있는 한 무리의 신경 원들이 일으키는 복합반웅이다. 이런 종류의 유발전압은 해마로 진입 하는 주요 구십로인 천공로(pe r fo ran t p a th wa y)를 자극한 후 해마의 치 상회 (denta t e gyru s) 에서 기록할 수 있다. 천공로는 내비피질 (en t orh i nal co rt ex) 을 구성하는 신경원들의 축색들로 구성되며, 이 신경원들은 치 상회의 과립세포에서 시냅스한다. LTP 현상은 수 개월에 걸쳐 지속 되며 학습의 기초가 되는 장기적인 변화와 관련되어 있다. Gary L yn ch 와 Mi ch el Baud ry (l984) 는 LTP 가 치상희 과립세포의 시냅스후 종말에서 칼페인 (Cal p a i n) 이라는 물질을 활성화시키는 데 관 여한다는 증거를 제시하였다. 이들에 의하면, 칼페인은 시냅스후막의 내면에서 포드린(fo dr i n) 이라 부르는 구조 단백질을 파손시키는 것으로 추측되는데, 이 포드린이 파손되면 이들 시냅스에는 신경전달물질을 수용하는 시냅스후 수용기가 더욱 많이 출현될 것이라고 한다. 한편 이 과정은 비교적 영속적으로 작용하기 때문에 일단 이 회로가 활성화 된 후에는 보다 많은 수용 부위가 증가함에 따라 시냅스후 신경원의 반응능력이 증가될 것이라고 한다. 이 가설은, 학습이란 시냅스후 신경원의 생화학적 및 구조적 변화를 포함하는 것 이라고 주장함으로써 Kandel 과 Schwar t z 의 가설과는 다 소 차이가 있다. 물론 시냅스전 또는 시냅스후 양자의 변화가 여러 종 류의 학습 상황에 따라 뇌 조직 내에서 일어날 수 있다. 이 두 가설은 학습에 개재하는 결정적인 신경회로가 시냅스라는 데는 동의한다. 학 습이 현존하는 시냅스 구조의 변화에 기인할 것이라는 견해는 역사적 으로 Ram6n Y Ca j al 에까지 소급되며, 1949 년 Donald Hebb 에 의해 서술된 『행동의 조직 (The Orga n iz a tio n of Behav io r) 』이란 책에서는 학
습이란 〈 새로운 신경세포둘의 집합 (cell assembl y) 〉 이라는 견해가 제시 되었다. 비록 Hebb 에 의해 제안된 생각들을 실험적으로 검증하기는 어렵지만 그가 제안한 일반적 개념과 논의는 학습과 뇌에 관한 현대 생물심리학의 견해를 발전시키는 데 매우 중요한 기여를 했다. 4 건망증 학습과 기억의 생물학적 이해를 위해, 뇌 손상이나 뇌 수술로 인해 학습이나 기억능력에 심한 장애를 보이는 건망증 환자들을 대상으로 한 연구를 살펴보기로 하자. 이런 연구들 가운데 가장 대표적인 예가 신경과학에서 널리 인용되는 H.M 이란 환자의 사례이다. H.M 은 27 세 경 심한 간질발작 때문에 해마 부위에 있는 뇌종양을 의과적 수술에 의해 제거당했다. 그 후 그는 심한 건망증을 보였는데 다음은 Mi lne r (19 70) 에 의 해 보고된 H.M 의 사례 이 다. 나는 내가 아무리 줄거운 일을 경험했거나 또는 슬픈 일을 경험했다 하 더라도 나에게 있어서 하루란 그 자체로 고독하게 끝나고 만다. …… 지금 芸 내가 말한 것이나 내가 행동한 것에 어떤 ’ 잘못이 있지 않을까 걱정 한다. 바로 이 순간 내가 멀쩡한 사람인 것으로 보이겠지만, 조금 전에 무엇이 일어났는지조차 모른다 . 이것이 바로 나를 괴롭히는 것이다. 그것 은 마치 내가 꿈 속을 걷고 있는 것과 같은 기분이다. 나는 아무것도 기 억할 수 없다. H.M 사례처럼 심한 기억상실을 보이는 건망증 환자를 대상으로 한 연구로부터 우리는 기억의 병리현상과 정상적인 기억의 기제를 밝히는 데 도움을 받을 수 있다. 비록 기억상실증 환자를 대상으로 한 연구가 19 세기부터 시작되었지만 기억상실증 환자의 뇌를 부검한 사례는 혼치 않다. 최근에 이르러 건망증 환자를 대상으로 한 행동 연구와 이들 뇌
의 부검연구를 상관시킨 연구가 많이 보고되고 있다. 4.1 건망증이란 무엇인가? 건망증 환자들은 몇 가지 증후를 보이는데, 그 중 가장 혼한 증후가 기억상실이다. 언뜻 보기에 이 환자들은 별 장애가 없는 사람처럼 보 인다. 왜냐하면 이들은 일상적인 대화를 자유자재로 잘 수행하기 때문 이다. 그러나 몇 분 후 이 환자를 다시 만나게 되면 이들이 조금 전 자기와 만났던 사실을 전혀 기억하지 못하고 처음 만났을 때 나누었던 이야기를 되풀이한다는 사실을 목격하고 이 사람들의 기억에 어떤 문 제가 있음을 칙감하게 된다. 건망증 환자들에게 나타나는 기억상실은 과거 사실일 수도 있고, 지금 막 진행되고 있는 사실일 수도 있다. 대 부분의 건망증 환자는 이 두 범주의 기억상실을 동시에 보여주는 것이 일반적이다. 일반적으로 후행성 건망증 (re t ro gr ade amnes i a) 이라고 부르는 기억상 실증은 과거에 일어났던 사건에 대한 기억상실을 의미하며, 진행성 건 망증 (an t ero gr ade amnes i a) 은 현재 일어나고 있는 사건에 대한 기억상 실을 의미한다. 십한 건망증 환자들이란 특별히 고안된 기억력 검사에 서 정상인에 비해 2 표준편차 범위 이하의 성적을 보여주는 환자들을 지칭한다 (Warr ington, 1984 ; Weis k rantz , 1985). 그러나 불행히도 건망증 환자를 분류하기 위한 일반적 원칙이 아직도 공인되지 못하고 있기 때 문에 어떤 사람을 건망증 환자에 포함시켜야 할 것인가 하는 문제는 계속 논쟁거리로 남아 있다. 건망증 환자로 분류되기 위해서는 정신지체를 보여서는 안 되며, 적 절한 지능을 유지해야 하고(기억검사 성적은 예의), 과제수행을 위한 학 습능력도 갖추어야 한다 (Park i n, 1984). 그러므로 지능검사에서 요인별 I.Q 점수는 정상 또는 정상 이상이 될 수도 있다. 이처럼 건망증으로 분류되는 환자들은 다양한 모습을 보여줄 수 있지만, 한 가지 공통적 인 증후는 지능점수에서는 십한 장애를 보이지 않지만 기억에서는 심
한 장애를 보인다는 점이다. 이처럼 건망증 환자가 정상적인 지능을 소유하면서도 기억능력에 심한 장애를 보인다는 것은 매우 역설적이다. 4.2 건망증의 원인 건망증은 여러 가지 원인에 의해 발생할 수 있다. 죽 중풍, 감염성 질환, 만성 알코올 중독증의 후유증인 비타민의 결핍, 사고 또는 뇌 수술의 후유증과 같은 각종 뇌 장애가 건망증의 원인이 된다. 대부분 의 건망증 환자는 다음 두 집단 중 어느 하나에 속한다. 즉, 첫째 집 단은 단순포진성 바이러스 (he rp es sim p le x vir u s) 감염으로 인한 뇌영 (ence p ha liti s) 의 후유증으로 나타나는 뇌염후 건망증이다. 두번째는 만 성 알코울 중독에 의 한 코르사코프 정 신병 (Korsakof fs p syc hosis ) 에 의 한 건망증이다. Korsako ff는 19 세기 러시아의 뛰어난 신경과학자로 만성 알코올 중독인 환자들에게 심한 기억상실이 일어난다는 사실을 최초로 보고했는데, 그 후 이런 환자들의 증상을 코르사코프 정신병이 라고 부르게 되었다. 코르사코프 환자들이 보여주는 뇌 손상은 비타민 B1 의 결핍에 의한 것으로 보인다. 알코올은 소장에서 비타민 B1 의 홉 수를 저지하기 때문에 만성 알코올 중독환자는 비타민 B1 의 결핍이 일 어나기 쉽다. 또한 교통사고나 뇌 수술 후에 건망증이 생기는 경우도 있는데, 앞서 본 H.M 의 사례나 앞으로 볼 N . A 사례는 기억상실과 관 련되는 뇌병리 현상을 과학적으로 언급할 때 자주 등장하는 예이다. 앞서 언급한 H.M 이란 환자는 27 살 때 심한 간질 때문에 뇌수술을 받았는데 그 후 건망증이 생겼다. 비록 수술에 의해 그의 생명은 구할 수 있었지만 심한 진행성 건망증에 이어 심한 후행성 건망증도 나타났 고, 성욕 감퇴와 미각 상실과 갇은 다양한 증후들도 보였다. 1953 년 수술 후 H.M 을 대상으로 한 많은 연구 관찰이나 실험논문이 나오게 되었다. H.M 은 수술에 의해 양측 측두영의 중앙부와 해마의 전축 부 위의 2/3 그리고 이들 구조 주변의 백질과 신피질이 제거되었다. 수술 후 30 년 동안 H.M 은 계속 건망증 증후를 보여주었지만 그의 지능 수
준은 평균 이상이었다. H.M 이 자기가 살고 있는 장소가 어디인지, 누 가 그를 돌보아주는지, 점심은 무엇을 먹었는지도 모르면서 이런 정상 적인 지능을 유지하고 있다는 사실은 신기하다 (Cork i n, 1984). 한편 N.A 라는 사례는 전반적인 기억상실이 아니라 주로 언어적 기 억상실만 보여주는 사례이다. 그는 펜싱 칼이 코 부위를 뚫고 들어가 뇌가 손상되고 이로 인해 건망증이 생겼다 (Teuber, Mi lne r & Vaug hn , 1968). 그는 이 사고에 의해 시상과 뇌의 중앙 부위에 손상을 입었다. 그는 안구운동의 장애도 보여주었는데, 사고에 의해 시개전영역 (pre te c ta l area) 이 손상되었기 때문이다. H.M 과 N.A 는 지금까지 생존 하고 있는 실존인물이기 때문에 아직 이들의 뇌 손상 부위를 정확하게 알아낼 수는 없지만, 최근 살아 있는 환자를 대상으로 뇌 손상 부위를 밝혀낼 수 있는 뇌진단기법이 등장함에 따라 보다 정확한 뇌 손상 부 위가 밝혀질 것으로 전망된다. 최근 몇 년 사이에 건망중에 관한 몇 가지 상이한 이론적 주장들이 제기되었는데, 그 중 두 가지 견해가 특히 주목을 끈다. 첫째는 전망 증의 형태에 따라 관여하는 뇌의 해부학적 회로가 다르므로 건망증 증 후도 다르지 않을까 하는 주장이고, 두번째는 건망증에서 보여주는 장 애를 특징화시킬 수 있는 최선의 방법은 무엇일까 하는 문제이다. 다 시 말해 건망증 환자의 기억 가운데 어떤 것은 상실되며 어떤 것은 남 아 있을까 하는 문제이다. 이 문제에 대해 보다 자세하게 살펴보자. 4.3 특정 기억과 특정 뇌 부위 특정 기억에 관여하는 뇌의 해부학적 위치는 과연 서로 다를까? 앞 서 본 H.M 과 N.A 의 경우처럼 기억상실의 종류가 다르다는 것은 곧 이들의 뇌 손상 위치가 서로 다르기 때문일 수 있다. 이 입장에서는 상이한 유형의 건망증이나 기억에는 서로 다른 뇌 부위가 개입될 가능 성이 제시되어 있다.
이런 해부학적 견해를 반영하는 것으로서 건망증에는 두 가지 종류 가 있다고 주장하는 이론바 〈 이형 전망증(tw o amnesia ) > 가 설이 있다. 이 견해에 따르면 이형 건망증이란 뇌 장애의 부위에 따라 간뇌성 건망 증 (d i ence pha li c amnes i a) 과 내측 측두영성 건망증 (med i al tem p or al lobe amnes i a) 으로 구분할 수 있다는 것이다. 이 가설에 따르면 간뇌성 건 망증은 앞서 본 코르사코프 정신병에서 볼 수 있었던 기억상실이나 N.A 의 사례가여기에 해당된다. 즉 이 환자들은시상하부의 유두체 (mam mil lar y bod i es) 가 주로 손상되고, 그 밖에 시상의 배내측핵 (dorsome- dia l nucleus) 과 정중핵 (mi dl in e nucleus) 의 손상 때문에 기억상실이 일어 나는 것으로 간주한다 (S q u i re, 1982). 저자의 동물실험에 의하면 흰쥐의 경우 유두체의 양측성 손상은 미로 학습의 역전에 장애를 초래하고(장 현감 등, 1988), 시상의 배내측핵 손상도 심한 기억상실을 야기한다는 (장현감 등, 1989) 점 이 밝혀 졌다. 한편 내측 측두영성 건망증에는 앞서 본 H.M 이나 뇌영 후유증 환 자들에서 나타나는 기억상실이 포함될 수 있다. 비록 Weis k rantz (1985) 가 유두체는 뇌궁을 경유하여 해마와 신경 연락을 이루고 있기 때문에 수술에 의한 해마의 손상이 유두체에 심한 신경 손상을 일으킨 다고 지적했지만, 사실상 H.M 은 전형적인 내측 측두영성 건망증 손상 에 의한 건망증 환자이다. 그러나 내측 측두영성 건망증이 유두체의 손상에 기인하는지 여부는 아직도 결론이 나 있지 않다. 왜냐하면 몇몇 보고된 건망증 환자들의 뇌 손상의 위치를 정확하게 가려낼 수 없었다는 점과 뇌 손상의 부위 가 여러 뇌 부위에 중복되어 있다는 점, 그리고 개와 같은 고등 포유 류의 경우 유두체만 제거한 경우에는 왕복회피 과제나 단순한 기억력 검사과제와 같은 비교적 쉬운 과제에서는 장애가 나타나지 않는다는 사실들이 (Kim, Chang & Chu, 1967) 해결되지 않은 숙제로 남아 있기 때 문이다. 따라서 건망증에는 두 종류의 독립된 신경회로가 존재한다고 결론짓는 것은 아직 시기상조인 듯하다. 한편 행동적 근거나 해부학적 근거에서 두 종류 이상의 건망증이 존
재할 것이라는 견해도 제시되었다. 이 견해에 따르면 이 두 집단의 건 망증 환자들 사이에는 인지적으로나 행동적으로 차이가 있다는 것이 다. 예컨대 간뇌성 건망증 환자들은 기억의 부분적 요소들을 모아 어 떤 직관적 기억울 만들어내기가 어렵다는 것이다. 그래서 이 환자들은 ` 허위 기억에 의해 기억의 공백 부분을 대치하는, 이른바 날조화 (confa b ulati on ) 를 보여준다. 이 런 날조화는 특히 코르사코프 정 신병 환자의 초기 특칭이기도 하다. 그러나 내측 측두영성 건망증 환자들은 날조화룰 보이는 경우가 극 히 드물며, 이 환자들은 H.M 의 사례에서 보여준 것과 같은 기억장애 를 보여주는 것이 일반적이다. 개개의 건망증 환자들은 비록 코르사코 프 정신병과 같이 어떤 단일의 진단 범주에 속하더라도 기억장애에 있 어서 개인차가 심하다는 사실을 주목해야 한다. 즉 건망증 유형에 관 한 일반화의 기준이 복잡하기 때문에 연구자들에 따라 주어진 환자를 두고 어떤 대상에 포함시킬 것인지, 아니면 배제시킬 것인지에 관한 기준이 서로 달라질 수 있다. 간뇌성 건망증 환자들은 전두영 손상과 관련 있는 증후를 보이는 경우가 혼하다. 이러한 전두엽 증후 가운데 는 환자가 어떤 한 가지 행동으로부터 다른 행동으로 잘 전환하지 못 하고 이전 행동을 계속 유지하려는 옹고집적 행동(p ersevera ti ve behav i or) 이 포함된다(전두영 손상과 관련 있는 증후들은 제 13 장에서 보다 자세히 논의될 것이다). 뇌영 후유증 환자들에서보다 코르사코프 환자 들에게서 이런 증후가 더 잘 나타난다. 그러나 상이한 유형의 건망증 환자들이 보여주는 이러한 차이점들이 건망증의 정도 차이에 따른 것 으로 간주될 수도 있지만 아칙도 건망증에는 이 두 가지 유형만이 존 재한다는 보장도 없다. 4.4 건망중 환자의 손상 특징은 무엇일까? 건망증 환자의 장애 특칭은 무엇일까? 뇌는 학습하고 기억하는 데 최소한 정보를 부호화 (encod i n g)하고, 저장하며 (sto r age ) , 인출 (re tri-
eval) 하는 세 가지 과정 을 수행 한다. 어 떤 연구자는 코르사코프 건망 증의 결정적 장애가 정보의 부호화 장애라는 견해를 제시하였지만 (Butt er s & Cermak, 1980) , 다른 연구자들은 정 보의 부호화 장애 라기 보 다는 정보를 장기 기억으로 응고화하는 데 어려움을 보여준다는 견해 (Mi lne r, 1966) 를 제시하였다. 한편 또 다른 연구자들은 기억저장으로 부터 정보를 인출해 내는 데 어려움이 있다는 견해를 제시하는 경우도 있다 (Warr i n gt on & Weis k rantz , 1970 ; Wi no cur & Weis k rantz , 1976). 이 처럼 건망증 환자는 기억의 세 가지 과정에 모두 장애를 보여주지만 연구자에 따라 이 세 가지 과정 중 특히 어느 과정의 장애를 강조하는 가에 따라 견해차가 생긴다. 이 세 가지 과정을 보다 자세하게 언급해 보자. 4.5 응고화 장애인가? 코르사코프 건망증과 같이 정보를 부호화하는 데 어려움을 보이는 건망증도 있지만 부호화에 어려움을 보이지 않는 건망증 환자도 많기 때문에 부호화 가설은 설득력이 약하다. 따라서 이 입장은 약화되는 대신에 최근 논쟁은 주로 응고화 가설과 인출가설로 좁혀지고 있다. 기억이 응고화되는 데는 어떤 사건이 일어난 후 일정한 시간이 요구 된다는 가설이 제안되었다. 이 과정은 어떤 한 사건에 관한 기억울 강 화하는 시기로서, 그 사건 이후 다른 사건의 발생으로 인한 기억 방해 를 최소화시켜 주는 것으로 추측된다. 전기경련충격치료 (elec tr oconvul siv e shock the rapy : ECT) 를 받은 환자나 두뇌를 심 하게 강타당한 환 자는 자신의 기억을 응고화하지 못해 뇌 타격 바로 직전에 있었던 사 건을 기억하지 못한다. 경련충격을 가한 동물실험에서도 이러한 처치 를 가한 후 역행성 건망증이 발생된다는 사실이 증명되었고, 학습 후 충격이 가해지는 시간간격에 따라 역행성 건망증의 정도가 달라진다는 사실이 기억응고화 과정의 강력한 증거로 제시되고 있다. 충격적인 뇌 손상 이후 나타나는 역행성 건망증에 관한 옹고화의 설
명은 많은 지지를 받았다 (Russell & Nath a n, 1946 ; McGaug h & Herz , 1972). 이 가설에 따르면 뇌 충격은 진행중인 기억응고를 방해하기 때 문에 최근 경험한 요소들을 응고화하지 못해 역행성 건망증이 일어나 게 된다고 한다. 그러나 응고화 이론이 지지받는 데 있어 가장 큰 걷 림돌은 뇌 충격이나 ECT 이후 상실된 기억이 시간이 지나면서 다소 회복된다는 점이다. 이런 문제점에도 불구하고 H.M 의 건망증 사례를 응고화의 장애로 설명하려는 학자들이 있다 (M il ner, 1966). 이런 생각 은 이 환자가 어떤 대상에 대한 기억을 장기기억으로 저장하지는 못하 지만, 노력을 하면 단 몇 분 정도는 기억할 수 있다는 관찰에서 나온 것이다. 이러한 발견은 H.M 의 건망증을 응고화의 실패로 설명할 수 있는 가 능성을 제시하지만, H.M 에서 나타나는 전반적인 기억장애나 다른 유 형의 건망증을 설명하기는 어렵다. 예컨대 최근에 H.M 은 수술 11 년 전까지 거슬러 올라가는 역행성 건망증까지도 보였다고 한다 (Cork i n, 1984). 더구나 H.M 은 현재 진행되고 있는 사건에 대해 학습하고 기억 할 수 있다는 점도 새롭게 밝혀졌다. 이런 발견들은 현재 제기되어 있 는 응고화 가설로는 건망증의 전반적인 특칭을 설명하기 어려울 것이 라는 점을 시사하는 것이다. 4.6 인출장애인가? 건망증을 설명하는 또 다른 설명개념으로 인출장애가설 (re t r i eval defi cit hyp o th e sis ) 이 있다. 이 가설은 주로 Warr i n gt on 과 Weis k rantz (1970; 1975) 에 의해 제기된 가설로서, 만약 어떤 사람이 어떤 특정 사 실을 현재 기억해 내지 못한다고 해서 그 정보를 영원히 상실해 버린 것이 아니라 그 순간 기억울 인출해 내지 못하는 것을 의미할 수도 있 다는 것이다. 만약 그 후에 그 기억울 인출해 낼 수 있다면 이런 순간 적인 실패는 인출장애로 해석해야 한다. 건망증을 인출장애로 해석하는 입장은 몇 가지 견해에서 지지된다.
건망증 환자가 정상적 기억으로부터 정보를 인출할 수 없다는 개념은 건망증 환자 중 과반수 정도가 거의 완벽할 정도나, 아니면 상당히 의 미 있을 정도로 기억이 회복된다는 사실에 의해 지지된다 (V ict or, Adams & Coll ins , 1971). ECT 를 받은 많은 건망증 환자들 가운데 상당 수에서 유의미할 정도의 회복을 보여준다든가 역행성 건망증이나 진행 성 건망증이 모두 개선된다는 사실은 인출가설의 입장을 더욱 강화시 켜 주는 것이라 하겠다. 또 다론 입장에서 인출가설을 지지할 수 있는 것은, 약간의 암시나 부분적 단서를 제공한다면 건망증 환자에게 여러 종류의 기억과제를 수행하는 데 도움을 줄 수 있다는 것이다. 이런 연구에서는 먼저 피험 자들에게 일련의 미완성 그림을 제시한다. 이런 자극을 계속적으로 제 시하여 조금씩 원도형에 근접하게 되면 종국적으로 단어나 그립을 인 지할 수 있게 된다. 이렇게 계속되는 제시상황에서 정상 피험자는 부 분적 단서만을 계속 제시한 경우보다 훨씬 일찍 자국을 인지하였다. 그 이유는 이러한 부분적 단서나 조언과 같은 것이 기억 속에 저장되 고 있어서 전체적인 도형을 인지하는 데 도움을 줄 수 있기 때문이다. 홍미로운 사실은, 전반적으로 정상인만큼 양호하다고 할 수는 없지만 건망증 환자들도 자극 제시 다음날 극소수의 부분적인 단서를 갖는 자 국들을 정확하게 인지할 수 있다는 점이다. 그런데 이돌은 제시된 자 극을 의식적으로 기억하려고 하지 않으며 실험자를 기억하려고도 하지 않았다. 이와 같은 각성 없는 학습(l earn i n g wit ho ut awareness) 에 몇 가지 유형의 상이한 학습과제에 따라 건망증 환자들이 어떻게 대응하 는가를 앞으로 보게 될 것이다. 4.7 기억 없는 학습 건망증을 설명하기 위한 여러 시도들에서 보여주는 한 가지 어려운 문제는 건망증 환자들이 어떤 특정 학습과제에 대해서는 명백한 기억 을 보여주지 못하면서도 새로운 학습을 할 수 있다는 접이다. 죽, 이
처럼 명백한 각성을 보이지 않고서도 학습할 수 있다는 것은 이론적으 로 해석하기 어려운 수수께끼이다. 앞에서 본 H.M 과 갇은 유명한 건 망증 환자도 언어적으로나 비언어적으로 제시된 기억문제들에 관해 이 전에 그 과제를 보았다는 분명한 의식적 기억 없이도 학습하고 기억할 수 있었다. 예컨대 H.M 에게 별 모양의 도형을 거울을 보고 추적해 나 가면서 그립을 그리게 하면 오류수가 점차 줄어드는 학습을 보여준다. 그러나 정상인과 달리 H.M 은 전날 자신이 수행했던 과제를 전혀 기억 하지 못해 매번 학습할 때마다 과제학습에 관한 새로운 지시를 받아야 만 했다 (M il ner, 1966).
제 13 장 뇌반구와 인지기능 인간의 뇌가 진화된 다른 포유류의 뇌와 다른 특칭은 좌우 뇌반구 사이의 기능 차이이다. 좌우 뇌반구 사이의 기능적 비대칭성(fun c tion al as y mme try)은 인간이 유인원에 비해 훨씬 심하다. 이러한 반구 간의 특수성은 여러 가지 실험상황에서도 관찰되지만 가장 두드러진 예는 간질병으로 인해 좌우 양반구를 연결하는 뇌량을 절제당한 환자 롤 대상으로 한 연구에서 나왔다. 이 환자들을 체계적으로 관찰한 연 구와 또 한쪽 반구만 손상된 환자들을 대상으로 한 관찰에서부터 좌반 구는 우반구에 비해 언어생성에 더 중요한 역할을 하지만 우반구는 시 각, 청각 또는 체성감각 등의 형태재인에 더 중요한 역할을 한다는 사 실이 밝혀졌다. 비록 이 두 반구가 기능상 차이를 보이지만, 서로 협 동하여 활동한다는 사실을 간과해서는 안 된다. 100 여 년 전부터 대뇌 전두영은 미래를 계획한다거나 정상적인 정서 반응을 유지하는 데 중요한 역할을 할 것으로 추측했지만, 전두영 부 위의 해부학적 복잡성 때문에 이런 견해를 결론짓는 데는 어려움이 많 았다. 이 장에서는 좌우 반구의 기능 차이와 언어이해와 같은 고등 인 지기능에 관여하는 뇌반구의 특징을 여러 각도에서 살펴볼 것이다.
1 좌우 뇌반구의 해부학적 차이 인간의 대뇌를 육안으로 관찰해 보면 두 개의 뇌반구가 뇌량 (co rp us calloswn) 이라 부르는 거대한 신경축색다발에 의해 서로 연결되어 있 다는 사실을 알 수 있다. 뇌의 정수리로부터 아래로 내려가면서 두 반 구 사이를 가볍게 벌리면 그 속에 뇌량이 가로놓여 있다. 이렇게 드러 난 뇌량을 예리한 수술칼로 자르면 두 반구는 대칭적으로 갈라지게 된 다. 물론 뇌량 의에도 두 반구 사이를 연결하는 구조로서 전교련 (ante r i or commi ssu re) , 후교련 (po ste r io r commi ssu re) 그리 고 해 마교련 (hip po camp al commi ssu re) 이 란 구조도 있다. 두 반구의 아래 쪽에 위 치 하는 시상, 시상하부 그리고 뇌간까지를 양동분하면 뇌는 전체적으로 보아 좌우가 아주 유사한 두 개의 뇌로 나누어진다. 오랫동안 신경해부학자들은 뇌의 두 반구가 기능이 서로 닮은 거울 상 (m i rror i ma g e) 의 역할을 할 것으로 생각했다. 그러나 최근의 연구들 에 의하면 인간의 좌우 뇌반구의 구조는 동일하지 않고 약간 다르다는 사실이 밝혀졌다. 이런 좌우 반구의 비대칭성은 특히 측두평면(p lanum t em p orale) 이라 부르는 측두영 부분에서 두드러지게 나타난다. 이런 비대칭성은 1968 년 Norman Geschw i nd 와 Walt er Lev it sk y에 의해 발 견되었다. 측두평면은 청각정보처리와 언어이해에 관여하는 측두영의 한 부위이다. 그러므로 이 부위의 해부학적 차이는 좌우 반구의 기능 차와 관련 있을 것으로 생각된다. 심한 난독증 (d y slex i a) 을 보여주는 환자의 뇌를 부검해 보면 측두평면의 신경원이 심한 비정상적 구조를 보인다고한다. Geschw i nd 와 Lev it sk y는 100 명의 성인 뇌를 검사하였는데 그 중에 서 65 명은 왼쪽 측두평면이 오른쪽 측두평면에 비해 더 컸으며, 반대 로 11 명은 오른쪽 측두평면이 왼쪽 측두평면보다 더 컸고, 나머지 24 명은 차이가 없었다고 보고하였다. 그 후의 여러 연구에서도 이런 경 향이 재 확인 되 었다. Sandra Wi tels on (19 83) 은 성 인 뇌 를 대 상으로 연 구한 7 가지의 선행 연구들을 개관했는데, 전체 420 사례 가운데 좌측
측두평면이 우측 측두평면에 비해 더 큰 경우가 71% 였고, 우측 측두 평면이 더 큰 경우는 15%, 양쪽이 동일한 경우는 14% 였다고 한다. 비록 뇌의 비대칭성이 인간의 뇌에서 가장 현저하게 나타나지만 인간 만이 그런 것은 아니다. Le Ma y와 Geschw i nd(1975) 는 몸집이 큰 유 인원(예컨대 고릴라, 오랑우탄, 침팬지) 28 마리의 뇌를 대상으로 연구해 본 결과 그 중 17 사례 (61%) 에서 좌측 측두평면이 우측 측두평면에 비 해 더 크다는 사실을 발견하였다. 그러나 덩치가 적은 기번(gi bbon) 과 같은 원숭이를 대상으로 한 연구에서는 7% 만이 좌우 뇌의 비대칭성을 보였다고 한다. 심지어 조류에서도 좌우 뇌 구조, 특히 노랫소리를 생 성하는 신경핵의 경우 좌우 뇌반구의 크기에서 차이를 보여준다고 한 다 (Nott eb ohm, 1977) . 측두평면의 크기에 있어 좌우 뇌반구의 차이는 인간의 뇌에서 관찰 되는 비대칭성이 가장 두드러진다. 현재로서는 좌우 측두평면의 크기 를 평가할 수 있는 방법이 부검밖에 없지만, 최근 여러 가지 뇌해부학 적 차이 를 알아볼 수 있는 뇌 주사기 법 (brain scannin g tec hn iqu e) 이 소 개되고 있기 때문에 이 방법에 의해 좌우 뇌 동맥의 구조차, 전두영과 후두영 의 크기 또는 측뇌 실 후각(p os t er i or horn) 의 길이 등의 차이를 알아볼 수 있게 되 었다 (W it elson, 1983 ; Kolb & Whis h aw, 1985) . 2 뇌반구의 특수성 앞에서 본 것처럼 인간의 경우 뇌의 좌반구와 우반구는 구조상 차이 가 있고, 적어도 반 이상의 경우 왼쪽 측두평면이 오른쪽 측두평면보 다 더 크다. 이것이 사실이라면 이 두 반구 사이의 기능적 차이도 존 재하지 않을까? 이런 의문에 대한 대답이 지난 1 세기 동안 제시되어 왔다. 아직까지는 이 질문에 대한 일반적인 결론은 좌반구와 우반구 사이의 기능적 차이가 존재하지만 그 차이가 구체적으로 무엇이며, 정 상적인 행동을 이해하는 데 있어서 이 두 반구의 차이가 어느 정도의
의미를 가지는지에 관해서는 계속 논쟁중이다. 최근에 와서 정상인과 뇌 손상 환자를 대상으로 반구 특수성의 정도와 특성을 이해하기 위한 연구가 계속되어 왔는데 여기에서는 의과적 수술에 의해 두 뇌반구가 분할된 환자들에서 나온 증거들을 중심으로 살펴볼 것이다. 2.1 반구 절단 : 분할뇌 환자 좌우 뇌반구가 기능적으로 서로 다르다는 극적인 증거는 심한 간질 발작을 치유하기 위해 뇌 수술을 받은 몇몇 환자들의 연구로부터 나왔 다. 이들 환자들은 수술에 의해 이 두 반구 사이의 주된 연결통로인 뇌량이 절단되었는데, 그 이유는 한쪽 반구에서 일어나는 발작이 반대 쪽 반구로 퍼져나가는 것을 방지하기 위해서였다. 이러한 방식으로 간 질환자를 치료한 최초의 시도는 1930 년대 말 미국의 신경의과 의사 Wi lliam Van Wa g enen 에 의해 이루어졌다. 그 당시에는 이런 수술을 받은 환자들을 대상으로 몇 가지 심리검사를 실시해 보았지만 뚜렷한 영 향을 관찰하지 못하였 다 (Van Wage n en & Herren, 1940 ; Akelait is, 1944). Van Wa g enen 이 행한 수술에서는 대개의 경우 뇌량을 완전히 철단하지 않았기 때문에 부분적인 발작감소 효과밖에 없었다. 1950 년대에 이르러 Rog er S p er ry와 Ronald M y ers 는 두 뇌반구 사 이를 연결하는 뇌량뿐만 아니라 전후교련과 시교차까지 분할시킨 고양 이를 대상으로 일련의 실험을 실시하였다. 이런 처치를 받은 동물을 훈련시키면 두 개의 뇌반구에서는 각기 독립적으로 시각변별 훈련이 가능하다고 한다. S perry와 M y ers 는 어느 한쪽 반구는 다른 한쪽 반 구와 독립적으로 동물의 반응을 통제할 수 있다는 사실을 발견하였다. 이러한 심한 수술을 받은 고양이들은 병에도 잘 걸리지 않는다는 사실 이 판명되었으므로 과거 간질환자를 대상으로 행해졌던 Van Wage n en 의 연구를 다시 시도할 수 있다는 용기를 갖게 되었다. 이런 새로운 시도는 Jos eph Bo g en 과 Phil lip Vo g el 에 의해 실시되었는데, 이들은 뇌량의 완전한 절단은 말할 것도 없고 기타 좌우 연결구조인 전교련,
후교련, 해마교련을, 경우에 따라서는 시상 부위까지도 절단하였다 (Bog en & Vog el , 1962) . 이 환자둘은 Van Wa g enen 의 환자들에 비해 간질발작의 빈도나 강 도가 보다 성공적으로 감소되었지만 이런 심한 뇌 수술은 위험하기 때 문에 실제로 행해진 사례는 그렇게 많지 않다. 1983 년까지 캘리포니아 에서만 모두 15 사례의 수술이 행해졌고, 다른 지역에서도 28 사례의 수 술이 행해졌다 (Gazzan ig a, 1983). 이 중 극소수의 사례에서만 심리검사 의 실시가 가능하였는데, 대부분의 사례에서는 I.Q가 매우 낮게 나왔 으며 광범위한 뇌 장애도 보여주었다. 그러므로 인간을 대상으로 한 좌우 반구 분할증후에 관한 연구결과는 겨우 10 명 미만의 환자를 대상 으로 몇 년 동안에 걸쳐 반복 연구한 사례들에 불과하다. 비록 이 환 자들을 대상으로 한 연구결과로부터 전반적인 일반화를 도출한다는 것 은 불가능하지만, 이 연구들은 인간을 대상으로 개개 반구의 기억능력 과 특징을 알아보는 데는 매우 유용한 자료를 제공했다 .• 이 연구들에 서 Rog er S p e rry는 캘리포니 아 주의 환자들을 주로 다루었고, 다론 지역의 환자들은 Mi ch ael Gazzan ig a 가 다루었다. 이러한 분할뇌 환자 의 연구성과가 공인되어 Rog er S perry는 1981 년 노벨상을 수상하게 되었다. 2.2 반구 사이의 정보교환의 단절 분할증후에 관한 대부분의 연구들은 인간의 시각체계의 해부학적 특 징을 이용한 연구로부터 나왔는데, 제 5 장에서 살펴본 것처럼 개개의 신경원은 시교차란 곳에서 둘로 나누어져 시신경축색의 반 정도는 교 차되어 반대편 부위의 뇌반구로 두사된다. 그 결과 코의 바로 왼쪽(좌 측 시야) 어떤 지점에 참깐 동안 비춰진 자극은 우반구로 두사되며, 코의 오른쪽(우측 시야)에 비춰진 자극은 좌반구에 두사된다. 어떤 자 극이 어떤 쪽의 시야이든 0.2 초 이상 한쪽에 제시되면 피험자의 시선 은 옮겨지기 때문에 두 반구에서 처리될 수 있게 된다. 순간노출기란
장치는 시각자극의 위치와 제시 기간을 조절하는 장치로서, 분할뇌를 연구하는 연구자들뿐만 아니라 일반적인 지각을 연구하는 연구자들에 의해서도 유익하게 사용되는 실험장치이다. 이 분야의 초기 연구에서 매우 놀라운 사실들이 발견되었다. Sp er ry 와 그의 동료들은 단어를 오른쪽 시야에 비춰주면 환자들은 그 단어를 올바르게 읽는 데 어려움이 없지만 똑같은 단어를 왼쪽 시야에 제공하 면(오른쪽 반구로 두사) 그 단어를 읽을 수도 없었고, 스크린에 비춰진 단어와 일치하는 사물의 사진을 언어로서 확인할 수도 없었다. 〈 S p oon 〉이라는 단어를 왼쪽 시야에 제시했을 때 나타나는 전형적인 반응은 〈그것은 바로 반짝이는 섬광이로군요〉, 또는 〈나는 그때 아무 것도 보지 못했어요〉 등이었다. 그러나 이것을 오른쪽 시야에 제시하 면 〈s p oon 〉이 라고 소리 높여 의 칠 뿐 아니 라 〈 s p oon 〉의 사진을 정 확 하게 확인해 낼 수도 있었다. 이처럼 우반구의 기능은 멍청하여 사물을 읽지도 못하고 사진으로도 확인하지 못한다고 결론짓기 쉽다. 그러나 이런 결론은 매우 성급하고 잘못된 것이다. 만약 이들에게 시야 내에 제시하는 단어나 그립의 이 름을 밝히거나 읽으라고 주문하는 대신 시야 밖에 제시된 일련의 사물 둘 가운데 방금 본 단어나 그림과 일치하는 사물을 왼손으로 끄집어 내라고 하면, 환자들은 거의 100% 정도 정확하게 사물을 끄집어낸다. 이 환자들은 계속된 지시에 대해 정확하게 사물을 선택하면서도 매번 그 자신은 아무것도 보지 못했다고 말한다. 우반구는 촉각에 의해 어 떤 사물을 인식할 수는 있지만 적절한 단어를 사용하여 그 사물의 이 름을 알아내지는 못한다. 그 후 여러 연구자들에 의해 이런 상황 아래 서 환자들의 두 대뇌반구는 서로 독립적으로 정보를 처리하고 기억한 다는 사실이 확인되었다. S p e rry는 이런 상황을 아래와 같이 기술하였 다 (Sp erry, 1968) . 정상인들이 보여주는 의식의 흐름은 하나로 통일된 것인 데 비해 이 환 자들의 의식적 각성의 흐름은 두 개로 분리되어 있는 것 같다. 죽, 두 뇌
반구는 각기 독립된 의식을 가지고 있으며, 두 뇌반구 사이의 접촉이 단 절되어 정신적 경험을 공유하지 못한다. 수술 후 두 뇌반구는 각기 분리 된 기억체계를 갖게 되어 한쪽 반구에서 이루어지는 회상과정에 다른 쪽 반구가 참여하지 못한다. 2.3 우반구의 기능 특수성 우반구는 좌반구에 비해 지적 능력에서 일반적으로 열등하지만 어떤 과제의 수행에서는 더 우세할 수도 있지 않을까? 이런 질문에 대한 해답은 분할뇌 환자, 한쪽 반구만 손상된 환자 또는 특수 심리 검사법 을 적용한 정상인을 대상으로 한 연구로부터 나왔다. 이런 증거들에 의하면 어떤 특정 과제 수행의 경우 우반구가 좌반구보다 더 우세한 경우도 있다고 한다. 정상 뇌를 두고 좌반구 대 우반구의 능력을 비교 한다는 것은 찰못된 것일 수도 있다. 왜냐하면 뇌의 경우 두 반구 사 이에 뇌량이라는 연결구조가 있어서 두 반구를 기능적으로 연결시켜 주기 때문에 두 반구의 기능을 독립적으로 연구하기가 어렵기 때문이다. 분할뇌 환자를 대상으로 연구한 Rog er Sperry는 우반구가 좌반구에 비해 특히 시각자극의 형태를 재인하고, 형태를 구성하는 능력에서 보 다 우세한 기능을 보인다고 주장하였다. Gazzan ig a 와 S perry (1967) 는 분할뇌 환자들에게 왼손을 사용하여 삼차원적 투시화법으로 입방체를 그리게 하거나 이차원적 기하형태를 짝지워 토막을 쌓게 하면 아주 재 치 있게 찰한다는 점을 발견하였다. 한 예로서 수술 전 오른손잡이였 던 환자는 수술 후 비교적 단순한 기하학적 도형에 맞추어 나무토막을 쌓는 과제를 주었더니 오른손으로는 힘만 둘이고 성공하지 못했지만 왼손으로는 마치 다른 사람에 의해 지배받는 것처럼 나무토막을 향해 손을 뻗고 이를 주워 모아 쌓기 시작하였다. 이 환자들을 대상으로 원 근법에 의해 집이나 입방체를 그리게 하였을 때도 위와 유사한 증거들 이 나왔다. 이 모든 그림들에서 우반구의 지배를 받는 왼손으로 그려 진 그림이 좌반구의 지배를 받는 오른손으로 그려진 그립보다 우수하
였다. 정상적인 피험자를 대상으로 한 보다 최근의 연구에 의하면 시각형 태의 재인에서 두 반구 사이의 차이는 매우 미묘하다고 한다. Jus ti ne Serge n t ( 19 82) 는 인간의 얼굴을 자극으로 사용하고, 정 상적 인 젊은 사 람을 피험자로 삼아 다음과 같은 실험을 실시했다 . 먼저 순간노출기를 사용하여 익숙하지 않은 얼굴을 0.04, 0.120, 0 . 200 초씩 제시했다. 그 리고 나서 피험자들에게 방금 본 얼굴이 남자 얼굴인가 여자 얼굴인가 알아맞혀 보라고 지시하였다. 대답은 두 개의 단추 가운데 어느 하나 를 선택해서 누르면 되고 반응시간이 측정되었다 . 어느 쪽의 시야에 얼굴을 제시하든 전반적으로 오류의 비율은 6% 정도로 비슷했다. Ser g en t에 의하면 아주 짧은 노출의 경우 얼굴을 좌측 시 야에 제시 하 여 오른쪽 반구로 진행하게 했을 때 반응이 보다 빠르다는 사실을 발 견했다. 그러나 노출기간을 0 . 2 초 정도로 비교적 길게 했을 때는 우세 성이 좌반구 쪽으로 넘어갔다. 즉 오른쪽 시야에 비춰진 얼굴이 왼쪽 시야에 비춰진 얼굴보다 더 빨리 식별되었다. Ser g en t는 시각처리의 초기 단계 에서 우반구가 저 해상도 (low resoluti on ) 자국에 대 한 정 보처 리를 잘 하지만, 보다 많은 정보를 이용할 수 있는 고해상도 자극에서 는 좌반구가 우반구에 비해 얼굴재인이 다소 빠르다고 했다. 어떤 학자들은 이처럼 두 반구가 상이하게 조정되는 것은 자국 유형 때문이 아니라 두 반구의 정보처리 양석이 다르기 때문이라고 . 주장한 다. 죽 좌반구는 분석-계열적으로 정보를 처리하기 때문에 언어와 수 학적 추리에 적합하고, 우반구는 공간-종합적으로 정보를 처리하기 때 문에 형태동각과 전체적인 사고에 더 적합하다고 하였다 (Le vy Ag res ti & Spe rry, 1968 ; Spe r r y, 1968) . 이 모형 에 따르면 좌반구는 시 간 적으로 배열된 불연속적인 항목을 분석하는 특성을 보이는 데 반해, 우반구는 시간적 측면보다 공간적 측면이 보다 우세한 자극을 전체적 인 지 각표상으로 종합하는 데 적 합한 것으로 생 각한다 (W it elson, 1983) . 이러한 특이성이 두 반구의 근본적이며 복잡한 특성을 어느 정도 기술 해 줄 수 있을지는 좀더 지켜보아야 할 것이다.
뇌반구 기능에 관한 많은 정보가 단측성 뇌 손상을 입은 환자의 연 구로부터 나왔다. 캐나다 몬트리얼 신경학 연구소의 Brenda M il ner 는 이런 환자를 대상으로 일련의 연구를 수행하였다. 이 환자들은 간질발 작을 일으키는 뇌의 병리조직을 제거하는 의과수술을 받은 환자들이 다. 따라서 이 환자들은 〈 분할뇌 〉 환자는 아니며 병리적 뇌 조직이 뇌 의 어느 한쪽에 국한되어 있는 환자이다. 수술의 원칙은 비정상적인 뇌 조직은 완전히 제거하지만 정상적인 조직은 최대한 남겨두는 것이 었으며, 비정상의 병리조직이 주로 측두영에 있는 환자들이다. Mi lne r 와 Pe t r i des(1984) 는 얼굴재인을 포함하는 시자극 형태의 재인장애는 좌측 측두영을 손상시킨 경우보다 우측 측두영을 손상시킨 후 보다 자 주 나타난다는 사실을 발견하였다. 또 다른 실험 에서 M i lner 는 우측 측두엽 손상환자는 앨범 사전 중에 서 자기가 보았던 사람을 재인하는 데 많은 어려움을 보인다는 사실을 발견하였다. 좌측 측두영 손상환자나 그 밖에 다론 부위에 신경장애를 가진 환자는 얼굴재인에 어려움을 보여주지 않는다. 영국의 Eli za beth Warr i n gt on 과 그의 동료들은 우측 두정 영 이 손상된 환자도 우측 측두 영이 손상된 환자와 유사한 장애를 보여준다고 했다. Warr i n gt on 은 이런 환자들의 장애를 〈 우측 두정영 지각 부전증후군 (r ig h t par ie t a l imp er cep tion s yn drome) 〉 이라고 불렀는데, 이 환자들은 비록 익숙한 대 상이라도 제시하는 방식이 조금만 이상하면 이를 알아차리는 데 심한 장애를 보인다. Warr i n gt on(1982) 은 이런 장애를 지각 범주화의 장 애, 다시 말해 두 개 이상의 관련된 자극을 하나의 동일한 지각적 범 주로 묶어내는 능력의 장애를 뜻한다고 했다. 따라서 정상적인 상태에 서 우반구는 시각기억의 저장에 필요한 지각적 범주화의 단계를 수행 하도록 하는 데 특별한 역할을 하는 것으로 볼 수 있다. 여러 연구자들은 우반구가 다양한 재인 과제나 시각자국을 정신적으 로 조작하는 과제 수행에서 우세성을 보여준다는 사실을 발견하였다. 이런 과제들 중에는 지도 읽기, 작은 현(弦)을 보고 원의 크기를 판단 하는 과제, 숨은 도형찾기와 같은 과제 등이 있다 {S perry, 1982;
Beaumont, 1983 ; Mi lne r, 1982). 이런 과제수행에서 요구되는 것은 신속 하고 정확한 시자극 형태의 지각기능인데 이런 기능은 분명 우반구가 우세한 것으로 보인다. 시각 형태의 재인에 관한 우반구의 특이성 의에도 우반구는 재인과 관련된 몇 가지 기능에서 좌반구보다 더 우세한 기능을 한다. 그 중 하나가 촉각적 느낌에 의한 대상재인이고 다른 하나는 음의 형태에 관 한 기억이다. Sandra W it elson(1983) 은 촉각재인 검사를 개발했는데, 이 검사에서는 피험자에게 한 번에 하나씩 별다른 의미가 없는 모양의 삼차원 물체를 제시하고 여러 개의 이차원 그림카드들 가운데 이 삼차 원 물체와 같은 것을 선택하도록 하는 것이다. 정상 피험자의 경우 왼 손에 물체를 놓았을 경우(왼손에 있는 촉각수용기는 주로 우반구로 두사 됨) 오른손에 물체를 놓았을 때보다 더 정확하게 구분해 내었다. 이 실험에서 밝혀진 한 가지 홍미로운 사실은 이러한 반구 특수성이 소년의 경우는 6-10 세부터 나타나기 시작하지만 소녀들은 13 세까지 반 구의 우세성이 나타나지 않는다는 점이다. 이처럼 소녀들이 소년에 비 해 뇌의 가소성이 오래 지속되기 때문에 소녀는 소년에 비해 독서에 어려움을 보이는 난독증 (d y slex i a) 이나 언어 사용에 심한 장애를 보이 면서 동시에 사회적 철수가 특칭인 자폐증 (au ti sm) 을 보이는 경우가 보다 적은 것이 아닌가 생각할 수 있다. W it elson 은 소년의 경우 반구 특수성이 더 느리게 전행되기 때문에 소녀는 소년에 비해 좌반구 기능 에 나쁜 영향을 미치는 여러 가지 조건들에 더 잘 견뎌낸다고 주장하 였다. 또 다른 증거들에 의하면 우반구는 멜로디의 재인에도 특수성을 보 여준다고 한다. Brenda M il ner(1967) 는 우측 측두영을 제거한 환자가 좌측 측두영 제거 환자에 비해 멜로디에 대한 기억울 측정하는 음악능 력 검사수행에서 보다 심한 장애를 보여준다는 사실을 발견하였다. 우 측 측두영 제거 환자는 정상 피험자나 좌측 측두영 손상 환자에 비해 동일한 음표를 다른 악기로 연주한 것을 변별하는 음색 변별 (timb re dis c rim ina ti on ) 과제 에 서도 보다 심한 장애를 보여준다.
3 전두영 전두영은 19 세기 중엽 사고에 의해 전두영이 심하게 손상된 Phin e as Ga g e 의 사례가 보고된 후 많은 신경과학자들의 홍미를 끌게 되었다. Ga g e 는 철도 부설 노동자의 작업반장으로 나이는 25 세였다. 노동자들 은 철도 부설에 방해가 되는 바위를 제거하기 위해 폭약을 사용하였는 데 이것이 찰못되어 Ga g e 가 상처를 입었다. Ga g e 를 치료했던 의사의 보고서에 의하면 쇠막대의 파편이 Ga g e 의 눈 바로 아래로 들어가 머 리의 정수리 부분으로 뚫고 나갔는데 이로 인해 그의 전두영 전부가 손상되었다. 그는 이런 사고에도 불구하고 기적적으로 살아났으며 실 제로 참깐 동안만 의식을 상실했을 뿐 사고 후 몇 분 이내에 그의 동 료들과 대화도 나눌 수 있었다고 한다 (Harlow, 1868). 사고 전에 Gage 는 매우 진지하고, 열심히 일했으며 가정에도 충실한 사람으로 평판을 받았지만, 사고 후에는 Harlow 가 기술한 것처럼 성격이 매우 다르게 바뀌었다. 그는 변덕꾸러기로 불손하며, 천박하고 농울 잘하며(사고 전에는 없었 던 습관), 동료들에게 공경받지 못하며, 몇 가지 욕망들이 서로 갈등을 일으킬 때는 참을성이 없고, 충고를 받아들이지 않으며, 어떤 때는 대단 히 고집이 세고, 또 다른 때는 변덕꾸러기로 우유부단하고, 끊임없이 많 은 미래의 계획을 세우지만 곧 포기해 버리고 보다 그럴 듯한 다른 계획 울 고안하기도 했다. 그의 마음이 너무나 급격하게 변했기 때문에 그의 친구나 친지들은 이제는 그가 옛날의 Ga g e 가 아니라고 했다 (Harlow, 1868) . Ga g e 는 그 후 12 년을 더 살았는데 그 동안 서커스단을 따라다니면 서 관중들에게 사고가 일어나게 된 경위를 자세하게 이야기하는 것을 생업으로 삼았다. 전두영이 손상된 모든 사람들에서 Ga g e 와 똑같은 성격 변화가 일
어나는 것은 아니지만, 사회적 관습에 대한 의식의 결여, 계획의 치졸 성, 변덕성과 같은 것이 공통적으로 나타나는 특칭이다. Ga g e 의 사건 이후 전두영이 손상된 환자를 대상으로 한 행동 연구들이 많이 보고되 었다. 전두영은 크고 구조적으로 복잡하기 때문에 전두영이 행동에 미 치는 영향을 자세하게 이해하는 데는 어려움이 많다. 3.1 전두엽의 해부학 전두엽에는 일차운동영역 (Brodmann 4 번 영역)과 이 영역 바로 앞에 전운동영역 (Brodmann 6 번과 8 번 영역의 일부)이 있다. 이 영역 의에도 전두안야(fr on t al e y e- fi eld) 라 부르는 영역도 있는데, 이곳은 안구운동 울 통제하는 매우 중요한 곳이다. 그 밖의 나머지 전두영 영역은 일반 적으로 다음과 같은 세 부분으로, 죽 배의측영역 (dorsolate r al area), 내측영역 (med i al area) 그리고 안와영역 (orb it al area) 으로 나누어진다. 이 영역들은 뇌의 다론 영역들, 죽 전두영 의의 다른 뇌피질, 기저핵, 시상하부 그리고 편도핵 등과 상호 연결되어 있다 (M il ner & Petr id e s, 1984), 아직까지 다양한 전두영 손상 증후들과 이의 해부학적인 하위 구조들 사이에 어떤 명확한 관련성이 있는지는 찰 알려져 있지 않다. 3.2 전두엽 손상 환자들의 종류 전두영의 기능을 찰 모르는 이유가 전두영 손상을 입은 환자수가 적 어 연구하기가 어려운 것이라고 생각할 수도 있지만 사실은 그렇지 않 다. 총탄을 맞은 환자, 종양이 생긴 환자, 중풍 환자들 가운데 전두영 이 손상된 경우가 많다. 그 의에도 1936 년부터 1958 년 사이에 정신치 료를 목적으로 전두영 제거수술을 받은 환자가 자그마치 3 만 5 천 명에 달했는데 (Valenste in , 1980) 이러한 정신수술(p s y chosur g e ry)은 한때 정 신분열중, 우울증 그리고 그 밖의 여러 정신장애를 개선시켜 줄 것으 로 기대되었다.
비록 오늘날도 간혹 정신수술이 시행되는 수가 있지만 이 방법은 거 의 사용되지 않으며 많은 통제를 받고 있다. 이 기법은 1935 년 국제신 경학회에 참석하여 전두영 수술을 받은 한 마리의 침팬지가 수술 후 온순해졌다는 보고를 둘은 포르투갈의 Mon i z 라는 신경의과 의사에 의 해 도입되었다. 미국에도 1950 년 Freeman 과 Wa tt s 에 의해 이 방법이 도입되어 유행되었는데, 이 방법은 어떤 성공적인 치료법이 없어 거의 방치 상태에 있는 절망적인 정신과 환자들을 치료할 수 있다는 기대 때문에 당시 정신과 의사들 사이에서 꽤 유행하게 된 수술법이었다. 그 후 몇 가지 유사한 수술법도 개발되었지만, 공통적인 방법은 전 두영과 시상하부 사이의 산경로를 절단하는 것이며, 보다 최근에는 편 도체와 같은 변연계 영역으로까지 수술이 확대되고 있다. 사실 수많은 수술이 행해졌지만 수술 후의 상태를 제대로 기록한 보고서는 거의 없 다. 그 이유는 수술을 실시했던 신경의과 의사들이 그 후 계속해서 환 자들을 지켜볼 수가 없었고, 환자의 회복 정도를 주관적으로 판단했기 때문에 객관적인 보고가 불가능했다. 3.3 전두염 증후 앞에서 언급한 성격 변화 의에도 많은 다른 변화들이 전두영 손상 증 후 로 보 고 되 었 다 (Kolb & Whis h aw, 1985 ; Mi lne r & Petr ide s, 1984 ; Stu s s & Benson, 1984). 예컨대 여러 가지 종류의 실행증 (a p rax i a) 과 일 반 활동 수준의 변화와 같은 각종 운동장애가 신피질의 운동령과 전운 동령의 손상과 관련되어 보고되었다. 또한 시자극의 탐지장애와 시자 극에 대한 주의집중 장애 같은 증후도 전두영 손상과 관련 있는 증후 이다. 그러나 전두영 손상환자를 대상으로 표준화된 지능검사를 실시해 보 면 지능점수가 두드러지게 감소되지는 않는다 (Kolb & Whis h aw, 1985). 이 환자들은 주의가 쉽게 바뀌거나 검사지상에 있는 어떤 작은 점에 지나치게 주의집중하기 때문에 이 환자들을 대상으로 심리검사를 실시
할 때는 세십한 주의를 기울어야 한다. 한편 이 환자들은 또 다른 인 지검사들에서도 비정상성을 보여주는데, 이들은 여러 종류의 검사에서 정상적인 사람처럼 문제해결을 위해 전략을 바꾸지 못하고 한 가지 행 동전략만을 고집 하는 이론바 옹고집 행 동(p ersevera ti ve behavio r ) 을 보 여준다. 예컨대 이 환자들에게 한 벌의 카드를 주고, 카드의 색깔, 형 태 또는 기하학적 자국의 수에 따라 분류하게 한 후 분류방식을 임의 적으로 바꾸라고 지시하면, 정상 피험자는 쉽게 분류 전략을 바꾸어 적용할 수 있지만 전두영 손상환자는 변화가 어렵다. 어떤 때는 피험 자 자신이 분류 규칙이 바뀌었다는 것을 자기도 안다고 중얼거리면서 도 옹고집적인 행동을 계속 보여준다. 그 밖의 여러 가지 다양한 종류의 검사에서도 전두영 손상환자는 한 가지 종류의 반응을 버리고 다론 반응으로 바꾸는 데 심한 어려움을 보여주는 일종의 옹고집적 사고와 행동을 보여주는 경향이 있다. 또한 이들 환자는 때때로 기억장애도 보여주는데, 이 기억장애가 모든 경우 에 다 나타나는 것은 아니다. 그 밖에 어떤 환자들은 그들의 정신생활 에 많은 어려움을 보여주기도 한다. 예컨대 소수의 전두영 환자들은 캅그라스 증후군 (Ca pgr as s yn drome) 이란 것을 보여주기도 하는데, 이 런 희귀한 증후를 보이는 환자들은 자신의 배우자나 자식들을 마치 자 신의 친척인 것처럼 말하고 행동한다 (S t uss & Benson, 1984). 4 반구 우세성과 손잡이 실어증이 좌반구의 손상과 관련되어 있다는 증거가 처음으로 등장한 것은 1861 년 프랑스의 Paul Broca 란 의사에 의해서였다. 비록 Broca 보다 25 년 전 또 다른 프랑스 의사였던 Da~ 됴 언어장애와 좌반구 손 상의 관련성에 대해 주목했지만 그의 주장을 뒷받침할 수 있는 부검이 실시되지 않았다. Broca 의 발표 이후 그의 발견을 지지해 주는 수많 은 증거들이 제시되었으며 언어이해에 있어서도 언어생성만큼 우세성
의 정도가 높지는 않지만 좌반구가 우반구보다 우세하다는 발견이 제 시되었다. 대부분의 경우 언어생성 중추는 어느 손을 주로 사용하느냐 의 여부와 관계없이 주로 좌반구이다. 그러나 왼손잡이는 오른손잡이 에 비해 우반구나 또는 좌우 양반구가 언어생성 능력에서 다소 양호한 경향이 있다. 실어증 환자로부터 얻은 증거들에 의하면 오론손잡이의 경우 95 - 99% 의 실어증 환자가 좌반구의 손상과 관련 있다고 하는 데 반해 우반구 손상에 의한 실어증은 오른손잡이의 1-5% 정도에 불과하다. 왼손잡이 또는 어떤 손잡이인지가 명확하게 구분되지 않는 사람의 말하기와 언어이해의 편재화 정도는 오른손잡이보다는 미약하다. 예컨 대 Le Ma y와 Geschw i nd (1 978) 는 오른손잡이가 아닌 실어증 환자 가 운데 약 60% 에서 좌반구가 손상되었고, 나머지 40% 는 우반구가 손상 되었다고 하였다. 오른손잡이가 아닌 경우는 오른손잡이에 비해 언어 중추가 우반구에 있는 경우가 더 많다는 증거도 있다. 실어증에서 얻을 수 있는 중거 의에도 손잡이성과 언어의 반구 우세 성 사이의 관계에 관한 자료들이 Wada 검사로부터 나왔다. 이 검사 는 신경의과 의사인 Juh n Wada(l949) 에 의해 고안되었는데, 이 방법 에 의하면 한 번에 한쪽 반구씩을 마취하는 방법을 통해 언어생성에 있어서 반구 우세성을 뇌 수술 이전에 미리 알아볼 수 있다. 이 방법 울 통해 언어생성의 반구 우세성을 미리 알게 되면 가능한 이 부위를 남겨 두고, 수술할 수 있는 계획을 수립할 수 있다. Wada 검사를 실 시하기 위해 뇌의 반구에 혈액을 공급하는 주요 동맥인 좌우 경동맥 속에 작은 관을 삽입한다. 그리고 나서 환자를 눕히고 sodiw n amy tal (강력한 중추신경진정제)을 주사한다. 주사받은 쪽의 대뇌반구는 금방 마취되지만 주사받지 않은 쪽의 반구는 깨어 있게 된다. 이러한 처치 를 하는 동안 환자에게 큰 소리로 숫자를 세도록 한다. 만약 약물이 언어생성에 관여하는 신경회로가 있는 반구 속으로 들어가게 되면 환 자는 약물의 작용이 지속되는 약 5 분 동안 숫자를 세는 것을 정지하고 말도 하지 못한다. 그러나 약물이 언어생성과 무관한 반구로 들어가면 환자는 참깐 동안 머뭇거리지만 곧이어 숫자세기를 계속한다. 이 검사
는 언어생성의 반구 편재성을 알아보는 데 아주 민감하지만 (Wada & Ras- mussen, 1960), 언어의 이해를 알아보는 데는 좋은 검사가 아니다 (Searleman, 1977) . Theodore Rasmussen 과 Brenda M il ner (1 975) 는 262 명의 환자 (140 명 은 오른손잡이, 122 명은 오른손잡이가 아닌)를 대상으로 Wada 검사를 실 시하였다. 이 중 어느 누구도 과거에 좌반구 장애로 고통받지 않은 신 경과 환자들이었다. Rasmussen 과 M il ner 는 오른손잡이의 96% 가 언 어생성에 있어 좌반구 우세성을 보여준 반면 나머지 4% 만이 우반구 우세성을 보여주었다고 했다. 오른손잡이가 아닌 환자들 중에 70% 만 이 언어생성에서 좌반구 우세성을 보여주었고, 15% 는 우반구 우세성 울, 나머지 15% 는 양반구 모두 언어생성에 관여한다고 했다. Ras· mussen 과 M il ner 의 실험결과는 손잡이의 유형과 관계없이 좌반구가 언어생성의 기제가 될 가능성을 시사하지만 오른손잡이가 아닌 환자에 서는 그 비율이 낮아진다는 사실도 시사한다. 앞에서 언급한 것처럼 유인원의 뇌에서도 구조적 비대칭성이 발견된 다. 그러면 유인원의 경우에도 언어생성과 관련 있는 뇌 기제에서의 반구간 비대칭성이 존재할까? 이 문제에 대한 결정적 증거는 없지만 비대칭성이 존재할 가능성은 높은 것으로 시사된다. 일반적으로 동물에서 단측성의 뇌 손상은 전반적인 감각기능이나 학 습 또는 기억과제 수행능력에 십한 영향을 미치지 않는다는 점이 밝혀 졌다. 대체로 이러한 단축성 손상은 뇌 손상과 관련 있는 신체영역의 감각변별력이나 운동과제의 수행력에는 다소 영향을 미친다. Ham ilt on(1977) 은 일련의 연구를 통해 어떤 한 종의 원숭이에서 좌반 구와 우반구의 기능 사이에 어떤 의미 있는 차이도 없다는 견해를 지 지하는 실험결과를 발표했다. 이 실험들은 분할뇌를 가진 레서스 원숭 이를 대상으로 한 것인데, 이 원숭이는 시교차도 절단되어 개개의 반 구가 완전히 독립적으로 검사된 것이다. 얼굴 재인검사를 포함하는 일 련의 검사에서(원숭이는 이 과제를 찰한다), 좌우 반구 사이의 차이가 나타나지 않았는데, 인간의 경우 우반구가 좌반구에 비해 이 과제의
수행에 더 우세했던 것과 비교하면 이 결과는 주목을 끈다. 이 연구에서 홍미를 끄는 한 가지 측면은 비록 시각 재인검사에서 좌우 반구 사이의 차이를 발견할 수 없지만 선호성에서는 좌우 반구가 차이룰 보인다는 점이다. 즉, 천연색 사진(특히 다른 원숭이의 얼굴 사 전)을 보여주고 이를 선택하게 하면 좌반구는 우반구에 비해 사진에 대한 명백한 선호성을 보여주는데 이런 선호성은 어떤 종류의 의적 강 화와는 무관한 것이다. 동물의 소리지르기에 있어서도 반구 우세성이 있는가를 알아본 연구 둘이 있다. 그 중 두 가지 연구는 일본산 짧은 꼬리 원숭이 (Ja pa n ese Macaqu e monkey) 의 종특수성 소리 (spe c i es -spe c ifi c c alls) 를 집 중적 으로 연구한 것이다. 대부분의 유인원과 같이 이 원숭이도 몇 가지 종류의 특이한 소리를 질러댄다. 연구자들은 이러한 소리들을 녹음한 후 이 소리를 다섯 마리 원숭이들에게 오른쪽 귀나 왼쪽 귀로 둘을 수 있게 했더니 오른쪽 귀에 이 소리를 둘려준 경우에는 소리를 명백하게 구분 할 수 있었다고 한다 (Pe t ersen 등 , 1978). 이러한 결과는 사람들에서 발견되는 〈우측 귀 우세성(rig h t ear advanta g e ) 〉 과 극히 유사하다 (Ge ff en & Qu in n , 1984) . 인간 피 험 자들은 단어를 왼쪽 귀에 들려주었을 때보다는 오른쪽 귀에 들려주었을 때 더 정확하게 구분한다. 이것은 언어이해에 관한 좌반구 우세성과 동측성 보다 대측성 청각통로의 우세성이 서로 결합되기 때문이다. 위의 결과와 유사한 증거가 Ric k y e He ffn er(1984) 의 연구에서 나왔 다. 이들은 11 마리의 일본산 원숭이에게 15 가지의 상이한 소리를 변별 하도록 훈련시켰다. 그리고 나서 좌측 또는 우측 상측두희 (su peri or tem p or al gy ros) 를 손상시켰다. 왼쪽 측두엽이 손상된 다섯 마리의 원 숭이는 수술 후 상이한 소리를 변별하지 못했지만 오른쪽 측두영이 손 상된 다섯 마리의 원숭이는 수술 후 어떤 장애도 보여주지 않았다. 인 간의 경우 이에 해당하는 영역이 언어이해와 관련되는 피질이다. 비록 이 연구들이 몇몇 소수의 동물을 대상으로 한 예비적 발견에 불과하지 만 이 발견은 인간뿐만 아니라 유인원에서도 같은 종내의 동료들에게
의미를 전달하는 소리를 이해하는 데 반구간의 편재성이 있음을 시사 하는 것이다. 5 실어증 실어증 (a p has i a) 이란· 뇌 손상으로부터 야기되는 언어장애를 총칭하 는 말이다. 따라서 실어증이란 미약한 말더듬이로부터 굴을 쓰거나 말 하는 데 심한 장애를 보이는 경우까지 실로 다양하다. 실어증의 가장 혼한 원인은 중풍이나 의상에 의한 뇌 손상이다(중풍이란 뇌출혈에 의해 뇌세포가 죽거나 손상되는 경우이다). 뇌 손상으로부터의 회복은 여러 가 지 요인에 따라 달라지는데 아직 이 요인들에 대해서는 잘 모른다. 극히 최근까지 실어증 환자들이 보여주는 뇌 손상 영역에 관한 정보 는 주로 부검 결과에 의존했다. 그러나 뇌와 감이 살아 있는 연한 조 직을 검사하는 새로운 기법이 1973 년 도입되었는데, 이 기법을 컴퓨터 단층촬영 또는 CT 촬영 (comp ut e r ize d tra nsaxia l tom og rap h y : CT) 이 라 부른다. CT 촬영은 방사선 조사장치를 머리를 따라 180 도 회전시키면 서 뇌의 여러 각도에서 얻어전 X 선 사전을 컴퓨터와 결합시칸 것으로 서 미세한 뇌 조직면의 분석이 가능하다. 이 방법에 의해 종양이나 병 적으로 혈관이 확대된 부위를 쉽게 찾을 수 있으며 (Brus t, 1981), CT 촬영은 방사선-불두과 옥도 (rad i o-o p aque i od i ne) 를 정맥에 주사하여 건 강한 조직과 손상된 조직 사이의 대비를 늘려중으로써 그 효율을 높일 수 있다. 최근에 와서 개발된 기법으로 양전자방사단충촬영 (po sitr o n emi ss io n tom og rap h y : PET) 이 란 것 이 있는데 , 이 방법 을 사용하면 살아 있는 뇌의 혈류 활동을 알 수 있다. CT 촬영이나 PET 촬영은 모두 살아 있는 뇌에서 뇌 장애의 위치와 크기를 알아보는 데 유용한 방법이다. 아직 보다 자세하게 밝혀져야 할 것들이 많이 있지만 이 기법은 여러 종류의 실어증 증후와 관련 있는 뇌 부위에 관한 초기 견해를 지지해
줄 뿐만 아니라 〈 상이한 부위의 손상이 상이한 종류의 실어증을 발생 시킨다 〉 는 Damas i o 와 Geschw i nd(1984) 의 개념을 지지해 준다. 가장 최근에 발달된 기법인 핵자기공명촬영 (nuclear magn e tic reso- nance : NMR) 기법은 뇌의 다양한 밀도에 아주 민감하게 반응하게 하 는 기법으로 CT 와 PET 기법을 대치하고 있다. 이 기법은 수소 원자 가 마치 섬 세 한 방적 자석 {sp inn in g magn e ts ) 처 럼 작용한다는 점 을 이 용 한 것이다. 라디오파를 사용하여 뇌 속의 수소 원자를 동시에 회전시 키면 자장이 형성되는데 , 이 자장에 의해 발생되는 미세한 전압을 측 정한다. NMR 촬영은 다발성 경화증 (mul tip le scleros i s) 과 같은 뇌 장 애를 확인하는 데 특히 유용하게 사용된다. 5.1 표현성 실어증과 수용성 실어증 실어증을 분류하는 데는 여러 가지 방식이 있다. 그 중 가장 널리 알려진 구분은 Broca 실어증 (Brocas a p has i a) 이라 부르는 표현성 실어 증 (exp re ssiv e aph asia ) 과 Wemi ck e 실 어 증 (Wernic k es aph asia ) 이 라 부 르는 수용성 실어증 (rece pti ve a p has i a) 이다. 표현성 실어증 환자들은 말을 잘 구사하지 못한다. 죽, 이들은 뇌 손상 이전에 사용하던 단어 둘 가운데 국소수만 사용할 수 있으며 소수의 단어로 분절적인 말만 어렵게 구사한다. 이들은 문법적으로 완벽한 문장을 만들지 못하며, 대명사나 동사의 어미 변화 그리고 접속사를 빼먹어 버리기 때문에 전 보문식 의 문장밖에 구사하지 못한다 . 표현성 실어증 환자는 남들이 하는 말을 듣거나 자신이 스스로 읽은 것을 이해할 수는 있기 때문에 흔히 자기 스스로 난점을 갖고 있다는 것을 잘 알고 있다. 실어증 환자들은 그들의 성대나 기타 다른 언어 생성 근육둘이 마바되거나 약화되어 있지 않다. 이러한 극단적인 예가 말할 때는 사용하지 못하는 단어를 노래로 부를 때는 잘 사용할 줄 아 는 표현성 실어증 환자들에서 관찰된다. 이것은 아마도 노래에 관여하 는 신경기제와 일상적인 말하기에 관여하는 신경기제가 서로 다르며,
이 신경기제들이 서로 다른 반구에 위치하기 때문일 것이다. 1874 년 Broca 가 좌측 전두영에 언어중추가 있다는 증거를 제시하고 난 몇 년 후 Carl Wem i cke 는 좌측 측두영 의 손상에 의 해 나타나는 전혀 다른 종류의 언어장애를 보고했다. 이 실어증을 수용성 실어증 또는 Wemi ck e 실어증이라 부르며, 이 실어증을 야기하는 손상 부위 인 측두영 부위를 Wemi ck e 영역이라 한다. 수용성 실어증에서는 어 떤 특정한 말의 중단이나 단절이 없으며 환자들은 거의 정상인과 같은 속도의 〈말의 흐름 〉 을 보여준다. 그러나 수용성 실어증 환자는 자신의 말을 포함하여 타인의 말을 이해하는 데 어려움을 보여준다. 이들은 복잡하거나 긴 대화를 이해하지 못하고, 받아쓰기를 하지 못하며, 알 맹이 있는 문장을 쓰거나 말하지 못한다. 따라서 이런 환자와 대화한 다는 것은 매우 어려운 일이다. 수용성 실어증 환자의 또 다론 특징은 무의미한 단어를 문장 속에 잘 삽입한다는 것이다. 비록 말의 리듬이 나 흐름에서는 표현성 실어증 환자보다 더 좋아 보이지만 전반적인 대 화 능력은 불량하다. 실어증 환자의 사례를 통해 뇌의 조정기능에 관 해 어떤 것을 알 수 있을까? 상이한 뇌 부위의 손상이 상이한 유형의 실어증을 야기한다는 사실은 곧 언어를 이해하고 말을 구사하는 데 있 어서 뇌의 조정기능을 이해하는· 데 중요하다. 5.2 실어증의 단절이론 1965 년 Geschw i nd 는, 1874 년 Wern ic ke 에 의해 처음 제안된 견해를 대폭 수정, 보완하여 실어증의 단절이론을 제시했다. 이 이론에서는 뇌피질의 여러 영역들이 말과 언어의 여러 측면들울 통제하며, 실어증 은 이들 영역들 사이의 연결이 단절될 때 생긴다고 주장한다. 예컨대 이 이론에서는, 어떤 특정 발음에 개재하는 하위구조와 이 발음을 표 현하게 하는 신경계획 사이에 차이가 있다고 한다. 언어이해의 신경회 로는 좌반구의 22 번과 Wernic k e 영역으로 불리는 40 번 영역이며, 이 영역에는 인접한 청각피질 영역 (41, 42 번)과 후두영의 시각피질 (17,
18, 19 번)로부터 정보가 수령된다. 이 부위의 신경원들에서 나온 축색 둘은 각회 (angu la r gyrus) 라는 피 질 부위 를 통해 Wernic k e 영 역으로 두사된다. 이 이론에 따르면 각회를 건너가는 축색도 있다고 하는데, 이 축색은 두정영에 있는 신경원에서 출발한 것으로서 글을 쓰는 방법 울 전달한다고 한다. 단절이론에 따르면 단순한 하나의 발성을 실제의 말로 전환시키는 데 필요한 신경회로가 Broca 영역에 존재한다고 제안한다. Wernic k e 영역과 Broca 영역은 궁상핵 (arcuate fa s ci culus) 에 의해 서로 연결되어 있으며, Broca 영역에서 나온 신경원들은 인접의 운동피질로 두사되 고, 이 운동피질은 성대와 그 밖의 말하기에 관련되는 발성기관을 작 동시킨다. 한편 노래부르기에 관여하는 신경 프로그램을 포함하고 있 는 운동피질은 다른 부위의 뇌피질과 서로 연결되어 있다. 단절이론에 따르면 중풍이나 뇌 손상은 말하기와 언어에 관여하는 다양한 부위들의 상호 연결을 단절시켜 여러 종류의 실어증을 야기한 다고 한다. 사고로 생기는 뇌 손상은 손상영역이 정확하게 어느 하나 의 영역에만 국한되지 않기 때문에 언어능력에서도 표현성 장애와 수 용성 장애가 동시에 나타나는 혼합성 실어증인 경우가 많다. 이 이론 에 따르면 표현성 실어증 환자는 Wernic k e 영역이 거의 손상되지 않 으므로 언어의 기본구조는 형성할 수 있다고 본다. 다시 말해 이들 환 자들은 자신이 말하려는 의미를 잘 알고 있지만 이런 기본구조를 정상 적인 구어로 전환시키지 못한다. 5.3 실어증의 회복과 치료 실어증 환자들 가운데는 어느 정도 회복이 되는 경우도 있다. 회복 의 과정에 관해서는 잘 모르기 때문에 아직 단일 치료법도 없고, 치료 의 성패에 관해 만족할 만한 설명도 할 수 없으며, 회복에 관여하는 뇌의 변화에 대해서도 잘 이해하지 못하고 있다. 실어증의 치료에 관 한 가장 체계적인 접근은 Andrew Ke rt esz(1979) 가 고안한 방법이다.
그는 실어증 척도(또는 실어증 계수 a p has i a quo ti en t : A Q)를 개발했는 데, 이 척도는 여러 단계의 회복기간중에 있는 환자를 서로 비교하여 점수화할 수 있다. 이 방법을 통해 어떤 성인 실어증 집단 가운데는 일년 이내에 약 21% 의 환자가 회복된다는 사실을 발견하였다. 비록 다른 많은 환자들에서도 다양한 측면의 말과 언어의 이해력이 회복되 었지만 AQ 검사에서는 계속 정상에 비해 낮은 점수를 보여준다. 회복은 손상 후 몇 개월 동안 가장 잘 이루어지며 일년 후부터는 매 우 느린 회복만을 보여준다. 어린이는 성인에 비해 완벽할 정도의 희 복을 보여주는데, 예컨대 32 명의 어린이 실어증 환자를 대상으로 한 연구 (Ala j ouan i ne & Lhennit te, 1965) 에서 1/3 정도의 어린이가 손상 후 6 개월 이내에 거의 완벽할 정도의 자생적 회복을 보여주었으며 1 년 이 내에는 75% 가 회복되었다고 한다. 실어증의 종류가 다양한 것만큼 실어증의 치료방법도 다양하다 (Kert esz , 1979). 대부분의 재 활치료가둘은 그들의 치료방법 을 개 개 환 자의 경우에 맞추어 적용하고 있다. 그럼에도 불구하고 실어증의 치료 에 관한 몇 가지의 공통적인 견해가 있다. 그 중 두 가지 공통적인 방 법이 주로 적용되는데, 그 하나는 자극기법이고 다른 하나는 프로그램 식 교육치료법이다. 자국치료법은 언어와 언어 관련자국을 환자에게 계속적으로 제공하는 방법이다. 좌반구의 중풍으로 우반신이 마비되고 십한 실어증에 걸린 Patr i c ia Neal 이라는 영화배우에게 그의 남편인 Roald Dahl 이 사용한 자국치료 법의 예를 살펴보자. Dahl 은 Neal 에게 잔소리도 하고 농담도 하면서 어떻게든 Neal 이 말할 수 있도록 자극하였다. 그는 자신과 Neal 의 친 구들이 지쳤을 때는 간호원을 고용하여 계속 Neal 로 하여금 말할 수 있도록 자극하였다. 그렇게 하여 4 년 뒤 Neal 은 다시 인기배우가 될 수 있을 만큼 많이 회복되었다. 프로그램식 교수법은 환자에게 단순히 언어자극을 홍수처럼 가하는 것보다 체계적으로 단어와 언어기술을 익히도록 학습시키는 방법이다. 그러므로 환자들은 단순한 문장부터 새로 배워 보다 복잡한 대화까지
익히게 된다. 또 다른 기법들은 주로 실어증 환자들이 가지고 있는 잔 존 언어능력을 최대한 살려 노래를 부를 수 있도록 하는 방법이다. 이 러한 가락음조 치료 (melod ic int o n ati on t hera py)는 환자가 말하고 싶은 내용을 오페라식 대화법으로 노래할 수 있도록 도와주는 방법이다 (Sp ar ks, Helm & Albert , 1974) . 6 대뇌 우세성의 발달과 피질의 기능단위 말하기나 언어에서의 뇌반구 우세성이 출생 후 몇 년까지는 지속될 것이란 몇 가지 견해가 제안되었는데 어떤 견해는 뇌반구의 우세성이 나이와 더불어 발달된다고 하였고, 다른 견해는 뇌반구 우세성은 출생 때부터 나타나지만 열세한 반구도 계속 가소성을 유지하고 있어서 우 세한 반구가 손상을 입을 경우 그 기능이 이행되며 이 가소성은 나이 와 더불어 점점 약화될 것이라고 주장했다. 이런 견해롤 지지하는 많은 증거둘이 생애 초기 뇌 수술을 받았거나 뇌 손상을 입은 어린이를 대상으로 한 연구로부터 나왔다. 이 연구들 에 의하면 뇌 손상이 좌반구에 일어났거나 우반구에 일어났거나 상관 없이 언어발달의 지체에 미치는 영향은 크지 않았다는 것이다. 죽 아 동기까지는 언어의 뇌반구 우세성은 나타나지 않거나 나타난다 하더라 도 대 단히 미 약하다는 것 이 다 (Basser, 1962) . 또 다른 뇌 손상 보고에 의하면 뇌 손상이 출생 후 이른 시기에 일어날수록 정상적인 말과 언 어기술의 발달 또는 회복이 잘 된다고 한다. 이런 사례에 바탕을 두면 생애 초기의 유아는 좌반구나 우반구가 언어능력에서 거의 동등한 능 력을 보여주지만, 어떤 이유인지는 찰 알 수 없지만, 그 후 점진적으 로 좌반구가 우반구에 비해 언어능력에서 보다 우세한 능력을 보여주 는 것 같다. 이 런 두 뇌 반구의 출생 시 동등성 가설 (equ al-at- b ir th hyp o th e sis) 은 최근에 들어와 도전을 받고 있다. 죽 좌반구가 출생시부터 우반구에
비해 보다 우세하다는 증거들이 제시되고 있다. 아동들이 성인에 비해 좌반구의 손상으로부터 보다 잘 회복된다는 사실을 고려한다면 이 동 등성 가설은 아동의 ~} 성인의 뇌보다 가소성이 더 많아서 아동의 우반구는 성인의 우반구에 비해 좌반구가 손상을 입었을 때 우반구 쪽 으로 기능 전이가 더 빨라질 것으로 간주할 수 있다. 이런 가소성은 나이와 함께 점차 감소된다. 아직 확실한 것은 아니 지 만 출생시 우세 성 가설 (domi na nce-at- b ir th hyp o th es i s) 을 지지하는 증거둘이 많이 제시되고 있다. 첫째, 많은 연 구자들이 아동기 뇌 장애에 관한 초기 보고들을 재검토하였더니 처음 주장했던 것처럼 우세성의 접전적 발달이 지지되지 않는다는 점을 발 견하게 되었다(Ki nsboume , 1975 ; Spr i n g e r & Deuts c h, 1981). 더구나 최 근 한쪽 뇌반구에만 손상이 있는 65 명의 아동을 조사한 결과 왼쪽 반 구에 손상이 있는 34 사례 가운데 73% 인 25 사례에서 실어증 증후를 보 여준 데 반해 우반구가 손상된 경우에는 31 사례 중 13% 인 4 사례에서 만 이런 증후를 보여주었을 뿐이다. 이런 통계치는 성인의 경우에서도 비슷하게 나타나 말과 언어의 뇌 우세성은 어른에서와 같은 비율이 어 린이에서도 나타난다는 것을 의미한다. 둘째, 출생시 우세성 가설을 지지하는 또 다른 증거가 정상 아동들 울 대상으로 한 연구로부터 나왔다. Youn g (1982) 은 단어를 알아차리 는 데 있어 오른쪽 귀의 우세성을 보이는 경우가 아동기 초기부터 (대 략 3 세부터) 나타난다고 하였다. 더구나 나이가 들어가면서 이런 오른 쪽 귀의 우세성이 바뀐다는 증거는 발견되지 않는다. 이와 유사한 증 거로서 시각적으로 제시한 단어에 대한 좌반구 우세성을 발견한 연구 도있다. 끝으로 비록 결정적인 것은 아니지만 출생시 우세성 가설을 지지하 는 해부학적 증거도 있다. Sandra W it elson 과 Wazir Pa lli e(1973) 는 생후 3 개월 이전에 죽은 어린이의 뇌를 부검한 결과 79% 의 사례에서 좌측 측두가 더 크다는 사실을 발견하였다. 다른 연구자들도 이와 유 사한 결과를 발견하였지만 이처럼 두드러진 결과는 아니었다. Juh n
Wada 와 그의 동료들(1 975) 은 임신 7 개월부터 생후 8 개월까지 여러 기 간에 걸쳐 사망한 아이돌의 뇌를 검사하였더니 100 사례 중 56% 의 사 례에서 좌반구의 측두평면이 더 컸고, 12% 에서 우반구의 측두평면이 더 컸으며 나머지 32% 는 좌우가 동등하다는 사실을 발견하였다. 임신 10 주에서 44 주 사이의 태아 뇌를 연구한 결과 54% 가 왼쪽 반 구의 측두평면이 더 컸으며, 18% 는 우반구의 측두평면이 더 컸고, 나 머지 28% 는 서로 유사했다고 한다 (Ch i, Dooli ng & Gil les, 1977). 물론 측두평면의 해부학적 차이가 언어의 대뇌반구 우세성과 관련성이 있을 것으로 추측은 되지만 확실하게 관련되어 있다는 증거는 없다. 더구나 이런 해부학적 측정은 쉽지 않으며 연구자에 따라 사용되는 측정방법 은 상이 하기 때 문에 (Wi tels on, 1983) 비록 해부학적 자료와 출생 우세 가 설이 양립될 수 있지만 이것이 문제를 해결하는 것은 아니다• Vernon Moun t cas tl e(1979) 은 포유류의 뇌피질에는 기능적으로 서로 관련되어 있는 세포들이 모여 하나의 기둥〔柱〕울 이루는 기본 기능단 위가 있다고 주장했다. 그는 이 견해를 뒷받침하기 위해 광범위한 피 질영역에 걸쳐서 피질주 (co rtic al column) 라 부르는 기본구조가 존재한 다는 것을 예시하였다. 그는 모든 피질의 기능에 관여하는 신경기제는 이 피질주들 사이의 상호작용과 관련 있을 것으로 추측했다. Mountc a stle .g -, 신피질이 부위에 따라 기능 차이를 보이는데 그 이유 는 각 피질마다 의부와 연결하는 방식이 서로 다르기 때문이라고 하였 다. 즉 신피질의 기능은 어느 곳에서나 지금까지 추측했던 것보다 훨 씬 더 균일하며 포유류 특히 유인원에서 신피질이 크게 확대된 것은 질적으로 상이한 형태의 내적 조정을 가능하게 하는 새로운 형태의 신 경원들이 출현되었다기보다는 동일한 기능의 신경단위들이 수없이 많 이 복제되었기 때문이라는 것이다. 자세히 관찰해 보면 신피질의 표면에서부터 백질에 이르기까지 지름 이 30µm 정도가 되는 한 개의 피질주 내에 약 110 개의 신경원이 있다 고 한다. Moun t cas tl e 은 이를 소주(小柱, m i n ic olumn) 라 불렀다. 그는 이런 작은 기둥이 인간의 뇌피질에 약 4 억 개가 있으므로 인간의 뇌에
는 약 440 억 개의 신경원이 있을 것으로 추정하였다. 물론 피질의 부 위에 따라 이런 소주의 수가 다를 수도 있다. 하나의 기둥 내에 들어 있는 모든 신경원들은 특정 방향성을 가진 선분에 반응한다든가, 유사 한 빈도수의 소리에 반응하는 것처럼 거의 유사한 기능적 특성을 갖는 다. 이처럼 기능적으로 유사한 기둥들이 시각피질, 청각피질 또는 체 성피질에 존재한다. 운동피질도 이런 기능적 기둥으로 조직화되어 있 어서 어떤 수직기둥 속에 있는 신경원을 자극하면 거의 유사한 산체 부위의 운동이 일어난다. 공간적으로 약간 떨어져 있는 기둥들 사이에 도 의측 연결에 의해 서로 떨어져 있는 체계들이 어떤 특정 기능을 수 행할 수 있도록 서로 연결되어 있다. 개개의 기둥둘은 인접해 있는 몇 개의 상이한 기둥들과 모여 하나의 기능단위를 이룬다. 그러므로 뇌 손상에 의해 기등들 사이의 다양한 연결이 차단되면 신경학적 증후가 나타나게 된다. 아직 다양한 실어증의 증후와 이런 기능적 기둥둘 사 이의 관계를 연결시키는 실제적 견해가 제시되지 못하고 있지만 이런 생각은 가능성이 높은 견해라 할 것이다.
제 14 장 정신장애 정신장애는 인간 행동을 이해하는 데 있어 가장 중요하고도 어려운 문제이다. 오랫동안 〈정신병 (men t al ill ness) 〉이라고 간주되어 왔던 정 신분열증이나 우울증과 같은 정신장애가 뇌의 생물학적 장애에 기인된 다는 증거들이 최근에 많이 제시되었다. 물론 모든 정신장애의 근원이 뇌의 장애인 것은 아니지만 최소한 어떤 조건 아래서는 뇌의 비정상성 이 곧 정신의 비정상성을 야기한다는 증거가 점차 축척되고 있다. 이 처럼 정신장애에 대한 생물학적인 접근이 시도되면서 이를 치료하기 위한 약물 사용도 증가되고 있다. 정신장애를 치료하기 위해 사용되는 대부분의 약물은 뇌의 시냅스 전달에 영향을 미치며, 정신장애와 뇌의 특정 신경전달체계 사이의 관련성 규명이 이 분야의 주요 연구주제가 되고 있다. 전세계적으로 헤아릴 수 없을 정도로 많은 사람들이 정신장애에 시 달리고 있다. 미국의 경우 정신분열증 환자가 200 만 명이 넘고, 수백 만 명의 사람들이 우울, 불안 또는 기타 정신적 문제를 가지고 있다. 전문가돌의 추정에 따르면 적어도 인구 10 명당 1 사람 이상이 일생 동 안 정신장애에 대한 전문적인 치료를 필요로 하는 상태라고 한다. 이 들 환자 중 대부분은 정신건강 전문가로부터 주기적인 도움을 받아 생 활할 수 있지만, 일부의 환자들은 입원치료가 필요할 정도로 장애가 심각하다.
정신장애의 진단에 사용되는 중요한 지침은 미국정신의학회 (Arner i can Psyc h ia tr y Asso ci a ti on) 가 발간한 『 진단 및 통계편람 (DSM : Diag. nostic a nd Sta tistical Mannual) 』이다. 이 지침은 24 가지의 다양한 조건들에 대한 진단기준을 담고 있으며, 현재 사용되는 것은 세번째 수정판인 『 DSM III-R (Diag n o stic and Sta t is ti c a l Manual, third edit ion , revis ed , 1987) 』 이 다. 이 장에서 우리는 정신분열증, 감정장애, 불안 및 공포증, 주의력 결 핍장애 및 자폐증과 갇은 대표적인 정신장애의 생물학적 바탕을 알아 볼것이다. 1 정신분열중 가장 심각한 정신장애 중의 하나가 정신분열증 (sch i zo p hren i a) 이다. 정신분열증이란 용어는 스위스의 정신과 의사인 Eug en Bleuler 가 1900 년경에 처음 사용했는데, 그는 이 장애의 기본 양상이 사고와 정서가 현실로부터 분리되는 것이라고 생각했다. 여기에서 분리 또는 분열이 라고 하는 것은 괴기하거나 반사회적인 것으로 보이는 개인의 행동을 유발시키는 것을 말한다. 정신분열중 환자는 정상인들로부터 고립된다 (정신분열증이란 용어가 두 가지 이상의 상이한 성격을 나타내는 사람을 가 리키는 것은 아니다. 이런 사람들은 DSM-m-R 에서는 히스테리성 신경증 의 하위 유목의 하나인 중다-성격장애에 해당한다). 정신분열증으로 진단 받은 남녀의 수는 거의 같으나, 남자의 경우에는 10 대 후반이나 20 대 초반에 전단을 받는 경우가 많고 여자의 경우에는 20 대 후반이나 30 대 초반인 것이 보통이다. 정신분열증에 걸릴 위험도는 전체 인구의 약 1% 정도이다. 1.1 정 신분열중의 중후 정신분열중의 주요 증후는 사고장애, 단조로운 감정, 망상 및 환각
이다. DSM-Ill-R 에는 정신분열중의 또 다론 증후들이 제시되어 있 으나 이 증후들도 위의 네 가지 증후와 관련되어 있는 것으로 간주된 다. 정신분열증으로 진단을 내리기 위해서는 이러한 증후들이 최소한 6 개월 이상 계속 나타나야 한다. 1.1.1 사고장애 정 신분열증의 가장 중요한 증후는 사고장애 (dis o rdered think in g) 이 다. 정신분열증 환자의 사고장애는 이들이 구사하는 언어에서 잘 나타 난다. 이들의 언어는 화제가 급격히 바뀌는 것이 특칭인데, 경우에 따 라서는 한 문장 내에서도 화제의 변화가 나타난다. 이런 산만한 연합 (loo se asso ci a ti on) 은 이들의 언어구사나 쓰기 를 이 해 하기 어 렵 게 하거 나 불가능하게 만들며, 이들을 정상인과는 다른 사람으로 취급받게 만 든다. 1.1 .2 단조로운 또는 부적절한 감정 정신분열증의 두번째 중요한 증후는 감정을 거의 표현하지 못하거나 상황에 맞추어 적절하게 표현하지 못하는 정서장애이다. 예컨대 정신 분열증 환자들은 사랑하는 가족의 죽음에 대해서도 냉담한 반응을 보 이거나 아무런 이유 없이 분노를 표시한다. 1.1 .3 망상 망상 (delus i on) 이란 불가능한 사실이나 도저히 믿을 수 없는 사실을 믿는 것을 말한다. 정신분열증 환자들이 나타내는 망상의 특칭은 자신 들의 생각이 의부의 다른 힘(예를 들어 FBI 나 의계인)에 의해 통제받고 있다는 생각을 하는 것이다. 또 다른 공통적인 망상은 환자 자신이 어 떤 유명한 사람의 쌍둥이 형제라거나 이미 오래전에 죽은 역사적인 인 물과 현재 접촉하고 있다는 생각울 하는 것이다. 망상형 정신분열증 (pa ranoid schiz o p hr enia ) 환자들이 보여 주는 망상은 환자들이 FBI 요 원에게 쫓기고 있다거나, 범죄조직으로부터 협박을 받고 있다는 상황
울 너무나도 자세하게 묘사하기 때문에 처음에는 그것이 그럴 듯하게 들리지만 그들의 생각이 불가능하다는 것이 곧 드러난다. 그러나 이런 망상은 망상을 가진 당사자들이 굳게 믿는 신념이다. 1.1 .4 환각 정신분열증의 또 다른 증후는 환각 (hallu ci na ti on) 의 출현이다. 가장 일반적인 것은 청각적 환각, 죽 환청인데, 때때로 이것은 환자 자신의 머리에서 나오는 소리로 간주한다. 이런 환청은 한 번만 들릴 수도 있 고 여러 차례 들릴 수도 있다. 전형적으로 환청을 일으키는 소리는 환 자가 과거 수행한 일이나, 장래에 해야 할 일에 관한 것이며, 내용상 으로는 비판적인 것이어서 환자 자신의 사고를 압도하는 경우도 있다. 불행하게도 이런 소리는 환자 자신에게는 거역할 수 없는 명령으로 해 석되기도 한다. 경우에 따라 환자는 환청의 소리에 대해 스스로 대답 함으로써 다른 사람들에게는 환자가 마치 독백을 하는 것처럼 보이기 도 한다. 정신분열증 환자들이 후각 및 시각적 환각을 경험하는 경우 도 있지만 청각적 환각만큼 일반적인 것은 아니다. 경우에 따라 이런 증후가 완화되는 시기도 있고 어느 정도 완전한 회복을 보이는 환자들 도 있다. 정신분열증 환자의 망상과 환각의 한 예를 다음 사례에서 살 펴보자. 대학을 졸업한 23 살의 J는 심한 정신병적 증후들을 보였기 때문에 부모 와 함께 병원을 찾았다. 그녀는 6 개월 전에 자신의 최초 직장이었던 무 출판사의 교정직을 그만두고 그 후로는 사람들과 전혀 대화를 하지 않으 려 하고 다른 사람들을 만나려 하지도 않고 자기 혼자서 움츠리고만 있었 다. J가 과거에는 대인관계에 지장을 초래할 만큼 심한 부끄러움이나 불 안감을 보이지 않았기 때문에 이런 행동의 변화는 주위 사람들을 몹시 놀 라게 하였다. 직장을 그만둘 즈음 그녀는 자신에게 무언가 이상한 일이 일어나고 있다는 것을 알게 되었지만 그것이 구체적으로 어떤 것인지는 알지 못하였다. 시간이 지남에 따라 그녀는 접차적으로 자기 자신의 생각
속으로만 몰입하게 되었으며, 존재의 의미나 종교적인 문제에 깊은 관심 울 가지게 되었다. 이런 생각에 매달리게 되자 그녀는 의모에 대해서는 전혀 신경을 쓰지 않게 되어 결국엔 화장도 하지 않고 옷마저 바꿔 입지 않아 몹시 추하게 되었다. 병원에 오기 몇 주일 전부터 그녀는 세상을 종 말로부터 구하는 것이 자신의 임무라는 확신을 갖기 시작하였다. J에 따 르면 다른 사람들은 이러한 절박한 위기감을 느끼지 못하기 때문에 이에 대처하는 데 필요한 조치를 할 수 없다고 하였다. 남들이 모르는 이러한 사실을 그녀 자신만이 알고 있는 것은 초자연적인 힘에 의해 신의 계시가 오직 그녀에게만 전해졌기 때문이라고 하였다. 즉, 어느 이른 새벽에 창 가에 서서 동녕 하늘을 바라보고 있울 때 동쪽 지평선 근방에서 유난히도 반짝이는 행성 하나가 그녀의 눈에 들어왔다. 이 행성을 조용히 바라보고 있을 때, 태양이 지평선 위로 솟아오르면서 찬란한 오렌지색의 광선이 그 행성을 향해 뻗어나갔다. 바로 그 순간 행성이 갑자기 사라지면서 뾰족탑 의 시계가 6 시를 치기 시작하였다. 그녀는 이것이 바로 신이 자기를 선택 한다는 계시라고 믿었다. J는 또한 이런 사명을 방해하려는 적들이 사방 에 있고 그들이 그녀의 마음을 읽을 수 있다고 하였다. 이런 적들의 천리 안적 정탐 행위를 막기 위하여 그녀는 일부러 시시한 행동에 몰두해 왔다 고 하였다. 그녀는 종종 적들이 자신에 대하여 욕하는 것을 둘을 수 있었 고 그들이 그녀의 비밀 사명을 저지하려 한다는 사실까지도 엿들었다. 그 녀는 온종일 이 거대한 전쟁을 수행함에 있어 그녀가 수행해야 할 역할에 대하여 초자연적인 힘으로부터 교육받고 있다고 하였다. 병원에 입원하기 바로 전날 금성으로부터 온 위성사진이 T.V. 뉴스에 보도되고 있었는데 이때 파리 한 마리가 T.V . 화면 위에 날아와 앉았다. 그 순간 그녀는 이 제 더 이상 기다릴 시간이 없다는 것을 알아차리고 자신의 사명을 곧장 행 동으로 옮기 려 하였다 (L i cke y와 Gordon, 1983) . 1.2 정신분열증의 유전적 요인과 기타 요인들 정신분열증의 원인에 관해서는 찰 모르지만 정신분열증을 일으키는 유전적 요인에 관한 증거는 많다. 예를 들어 전체적으로 볼 때 정신분
열증에 걸릴 위험도는 전체 인구의 1% 미만이지만 정신분열증 환자로 진단받은 형제, 자매를 가진 사람이 일생 동안 정신분열중으로 진단될 가능성은 정상인에 비해 6-10 배나 더 많으며, 일란성 쌍생아인 경우 그 가능성은 50 배 정도로 증가된다. 위에서 제시한 증거둘은 정신분열증의 유전적 요인을 강하게 시사하 고 있으나 아직 유전학적 연구가 완성된 것은 아니다. 일란성 쌍생아 의 정신분열증 일치율이 50% 밖에 되지 않기 때문에 그 밖의 다른 요 인들이 작용하는 것이 틀림없다. 그러나 이들 요인이 무엇인지는 아직 밝혀지지 않았다. 예컨대 스트레스를 일으키는 사건이 정신분열증을 유발하는 중요한 요인으로 작용할 것으로 간주되지만 누구도 이에 대 한 확실한 증거를 제시하지 못하고 있다. 또 다른 가설은 정신분열증의 증후를 일으키는 바이러스에 대한 뇌 의 취약성 가설이다. 그러나 이 가설을 지지하는 증거는 부족하다. 예 를 들어 바이러스 가설 (v i ral h yp o th es i s) 을 지지하는 증거는 9 명의 정 신분열증 환자 가운데 1 명 정도는 뇌척수액에 거대세포바이러스 (cy tom eg al ovir u s: CMV) 에 대한 항체를 가지고 있다는 사실이다. 이 에 반해 통제집단의 경우는 비록 혈장에서는 CMV 항체가 발견되지만 뇌척수액에서는 어떤 항체도 발견되지 않는다고 한다. 이 결과는 정신 분열증 환자와 통제집단 간에는 바이러스가 일반 체순환계로부터 보호 막으로 둘러싸여 있는 혈-뇌 장벽으로 침투해 들어가는 능력에 차이가 있다는 것을 시사한다 (Torre y, Yolken & Wi nf r ey , 1982). 정신분열증을 일으키는 유전적 소질로서 바이러스의 침입으로부터 뇌를 보호하는 혈 뇌 장벽의 능력 부족이 그 원인이 될 가능성은 있지만 정신분열증의 원인은 아직 더 연구되어야 할 문제로 남아 있다. 1.3 정 신분열증 환자의 뇌 손상 만약 바이러스가 정신분열증을 일으킨다면 뇌에는 어떤 가시적인 손 상 흔적이 남을 것으로 생각된다. 비록 바이러스와 정신분열중이 서로
관련되지 않았다 하더라도 정신분열중의 증후를 설명해 줄 수 있는 뇌 의 기능장애 또는 뇌 손상은 가능한 생각이다. 뇌의 기능장애가 정신 분열증에 중요한 요인이 될 수 있을 것이라는 논쟁은 거의 1 세기 동안 이나 계속되어 왔다. 컴퓨터 단층촬영법 (CT), 양전자방사단충촬영법 (PET) 또는 자기공명영상 (MRI) 과 같은 신경학적 기법이나 부검결과 에 의하면 정신분열증 환자의 약 1/3 에 해당하는 사례에서 뇌의 비정 상성이 관찰된다고 한다. Larr y Se i dman (1 983) 은 수백 명의 정신분열 증 환자를 대상으로 한 25 개의 개별 연구를 개관하고서, 그 중 20- 35% 에 해당하는 환자들에서 뇌의 구조적인 비정상성이 나타났다고 하 였다. 이 결과는 정신분열증 환자들이 정상인이나 다른 정신의학적 전 단을 받은 환자들보다 뇌 손상의 증후를 더 많이 보여준다는 증거이다. 뇌의 비정상성은 주로 뇌실의 확장 (enlar g ed cerebral ven t r i cles) 과 뇌 의 위축 (cerebral a t ro p h y)으로 특징화된다. 뇌실은 뇌 속의 큰 공간이 며 뇌척수액으로 가득 채워져 있다. 이 두 가지의 비정상성은 거의 동 시에 나타나는 것이 일반적이어서 양자가 서로 관련되어 있는 것으로 생각된다. 뇌실의 확장은 무엇을 의미하는가? 만약 뇌실을 채우고 있는 뇌척 수액으로부터 발생되는 압력이 정상이라면 뇌실의 확장은 뇌실 주변의 뇌 조직 용적을 정상보다 축소시킬 것으로 생각된다. 아것은 뇌의 비 정상적 발달(처음부터 이 뇌 영역이 정상적인 크기로 성장하지 못함)을 의 미하거나 뇌의 위축(정상적인 크기였으나 어떤 원인에 의해 축소되었다)의 결과일 것으로 간주된다. 뇌실의 확장과 뇌의 위축이 동시에 나타난다 는 것은 이상한 일이 아니다. 뇌의 위축이 바이러스 감영에 의한 것일 수도 있지만 그 밖의 조건들도 뇌의 위축을 일으킬 수 있다. 그러나 이러한 뇌의 비정상성이 정신분열증 환자의 1/3 정도에서만 나타나고, 나머지 2/3 에서는 나타나지 않는다는 사실 또한 주목해야 한다. 최근 일란성 쌍생아를 대상으로 한 연구에서, 둘 중 한 명은 정신분 열증으로 전단되었으나 다른 한 명은 정상인 15 쌍의 쌍생아에서(나이 25-44 세, 남 8 쌍 여 7 쌍) 뇌의 비정상성을 검토하였다 (Sudda th 등,
1990). 이들 연구자들은 MRI 를 사용하여, 정신분열증을 보여준 쌍생 아의 뇌실은 확장되어 있으나 정상적인 쌍생아는 그렇지 않다는 사실 울 발견하였다. 즉, 정신분열증 쌍생아는 14/15 쌍에서 정상에 비해 왼 쪽의 측뇌실이 더 컸고, 13/15 쌍에서 오른쪽 측뇌실이 더 컸다. 한편 해마의 전측영역은 14/15 쌍에서 정신분열증 쌍생아의 왼쪽이 더 작았 고, 13/15 쌍에서 오른쪽이 더 작았다. 이런 사실들은 정신분열증이 뇌 의 구조적인 비정상성과관련되어 있음을강력하게 시사하는증거들이다. 1.4 정신분열증의 하위집단 모든 정신분열증 환자들의 뇌에서 비정상성이 발견되지 않는다는 사 실은 정신분열증에 독특한 하위집단이 있을 가능성을 제기한다. 예컨 대 뇌실 확장과 뇌 위축은 정신분열증의 증후 가운데 가장 심각한 음 성 증후 (ne g a ti ve s ympt om) 를 보여주는 환자들에 서 만 주로 나타난다. 음성 증후란 단조로운 감정, 사회관계의 철수와 무감동이 주된 증후인 데, 이들 증후는 정상적인 인간 특성의 강도와 빈도가 심하게 감소되 어 있는 증후를 의 미 하는 것 이 다. 한편 양성 증후(p os iti ve sym pt~m ) 에 는 망상, 환각 및 사고장애와 갇은 비정상성이 포함된다. 표 14-1 은 정신분열증의 양성 및 음성 증후를 보여주고 있다. 뇌의 비정상성과 함께 십한 음성 증후를 보여주는 환자들은 뇌의 비 정상성이 없는 환자들에 비해 약물치료에 대해 찰 반응하지 않는다.
표 |4-1 정신분열중의 양성 및 음성 증후 환각 양성 중후 사고장애 망상 단조로운 감정 음성 중후 사회적 철수 개인적 위생 관리나 몸단장의 장애
또한 이들은 보다 만성적인 정신분열증 환자인 경우가 많으며 개선되 는 경우도 드물다. 이처럼 뇌의 비정상성과 음성 증후를 가진 환자들 이 정신분열증의 독특한 하위 집단을 형성한다. 뇌 장애가 없고 주로 망상, 환각 그리고 사고장애와 같은 양성 증후를 보여주는 하위 집단 도 있다. 이 집단의 경우에는 약물치료의 효과도 잘 나타나고 최소한 부분적인 회복이나 증후가 완화될 가능성도 크다. 1.5 정신분열증의 ·약물치료 1950 년대 이전에는 정신분열증을 치료할 수 있는 성공적인 방법이 없었다. 어쩌다가 병이 개선되거나 심지어는 치료되는 것처럼 보이는 환자들도 있었지만 많은 환자들에게 적용될 수 있는 구체적인 치료방 법은 없었다. 세계 도처에서 수많은 정신분열증 환자들이 최소한의 치 료라도 받기 위해 정신병원에 수용되었지만, 이들은 그곳에서 죽음을 기다릴 수밖에 다른 도리가 없었다. 미국의 경우 1950 년경에 정신병원 에 수용된 정신분열증 환자의 수가 무려 60 만 명에 달했으며, 그 수는 급속히 증가되고 있었다. 1950 년대 중반 프랑스에 서 클로르프로마전 (chlorp ro mazin e ) 이 라는 약 물이 정신분열증 환자를 치료하기 위해 최초로 사용되었다. 1960 년대 초반부터 이 약물은 전세계적으로 사용되었는데, 1975 년경 미국의 정 신병원에 수용된 환자의 수가 25 만 명 정도로 급격하게 감소되었다 (Bassuk & Gerson, 1978). 이 약물은 정 신분열증의 80% 를 차지 하는 양 성 증후를 감소시키는 데 특이한 효과를 보였지만, 이 경우에도 몇 가 지 제한점이 있다. 즉, 이들 항정신병 약물 (an tip s y cho tic dru g)은 정신 분열증을 근본적으로 완치시키는 것이 아니라 가장 성가신 몇 가지의 증후를 일시적으로 감소시킬 뿐이며, 이 약물을 계속 사용하지 않으면 증후가 다시 재발하는 경향이 많다. 대부분의 환자들은 약물이 증후를 완화시키기 때문에 병원에서 조기되원하게 된다. 되원한 환자들은 별 다른 문제 없이 살아갈 수 있기를 기대하지만 이 기대가 충족되는 경
우는 드물다. 약물치료는 환자와 그의 가족을 위한 심리상담, 직업상담 그리고 기 타 지지를 포함하는 포괄적인 치료 프로그램과 함께 적용하면 더욱 효 과적이다. 또한 약물치료에 매우 심각한 부작용이 따를 수도 있다. 그 러나 심리 상담과 함께 클로르프로마진을 비롯한 기타 항정신병 약물 을 적절하게 사용하면 보다 효과적인 치료가 된다. 정신분열증 치료에 사용되는 주요 약물 중 하나가 클로르프로마전이 라는 약물인데, 이 약물은 th oraz i ne 이라는 상품명으로 전세계적으로 팔리고 있다. 클로르프로마진은 페노티아진(p heno thi az i ne) 이라는 약물 족에 속한다. 그 밖에 페노티아진의 상품명에는 Vespr i n , Serenti l, Mellaril, 및 Tin d al 등이 있다. 두번째로 중요한 항정신병 약물족은 티오잔텐(thi oxan th enes) 인데, 여기에는 Tarac t an 이라는 상품명의 약 물이 있다. 세번째는 부티로페논 (bu tyr o p henones) 인데 여기 에는 Haldol 아란 상품명의 약물이 대표적이다. 1.6 항정신병 약물의 부작용 다른 약물들과 마찬가지로 t horaz i ne 이나 Haldol 과 같은 항정신병 약물도 부작용이 있다. 이런 약물을 두여받은 환자 가운데 약 1/3 에서 만발성 운동장애 (tar div e dys k in e sia ) 라는 부작용이 나타난다. 이 장애 는 주로 나이가 많은 사람들에게서 관찰되며 입과 혀에서 비정상적인 불수의 운동이 나타나는 것이 특칭이다. 이들 운동은 혀와 입 안에서 안면경련증(tic)과 같은 경련성 운동이 나타나는 것으로부터 시작된다. 증세가 심해지면서 영향이 파급되어 전 신체의 경련 또는 비틀림 운동 이 나타난다. 이런 운동들은 각성시기 동안에만 계속되며 수면 동안에 는 중지된다. 만발성 운동장애는 사람을 무능력하게 만둘 정도로 심각 할 수도 있다. 약물의 두여량이 적으면 부작용도 감소되는 경향이 있 지만 용량이 낮으면 정신분열증의 증후가 완화되지 않을 수 있으므로 투여량을 결정하는 것은 쉽지 않다. 만발성 운동장애를 호전시키는 몇
가지 약물이 있지만 그 효과가 항상 나타나는 것은 아니다. 아주 드물기는 하지 만 신경 마비 성 악성 증후군 (neurolep tic malig na nt syn d rome : NMS) 이 란 심각한 부작용이 나타나기도 한다. 이 증후군의 특칭은 심한 고열과 순환성 와해 (circ ulato r y colla p se) 로서 간혹 죽음을 초래하기도 하는데, 1980 년까지 모두 60 명의 사례가 보고되었으며 그 중 12 명이 사망했다고 한다 (Gar fi eld, 1983). 이러한 약물의 부작용 때 문에 정신분열증의 증후에 보다 효과적이면서도 부작용이 적은 약물을 개발하기 위한 많은 연구가 진행되고 있다. 1.7 정신분열증의 도파민 가설 많은 증거에 의하면 항정신병 약물이 정신분열증의 양성 증후를 감 소시키는 데 효과적이라고 하는데, 이런 효과는 약물들이 도파민을 신 경전달물질로 사용하는 시냅스에서 신호전달을 차단하기 때문이다. 도 파민 수용기는 D1, D 려 두 가지 종류가 있다. 환자를 대상으로 한 임상 연구와 동물실험 연구의 결과들에 의하면 t horaz i ne 과 Haldol 은 D2 수용기를 차단시켜 치료효과를 나타낸다고 한다. 여러 연구들에 의 하면 이런 약물이 정신분열증의 양성 증후를 감소시키는 효과와 이들 약물이 D2 수용기에서 신경전달물질을 차단하는 능력 사이에는 매우 높은 상관 (r=.90 이상)이 있다고 한다. 따라서 연구자들은 이들 약물 의 항정신성 효과는 이 약물들이 뇌에서 D2 수용기를 차단하는 정도에 달려 있다는 결론을 내렸다. 이런 약물은 D1 수용기도 차단하지만 약 물의 치료효과와 D1 수용기 차단능력 사이의 상관은 유의미하지 않다 고한다. D2 수용기의 차단효과와 임상적 효과 사이에 높은 상관이 있다는 사 실을 바탕으로 정신분열증의 도파민 가설이 제안되었다. 이 가설에 따 르면, 뇌의 도파민 회로(특히 D 2 수용기를 포함하는 회로)의 비정상적인 활동 때문에 정신분열증 증후가 나타난다고 한다. 도파민 신경원의 축 색은 뇌의 여러 영역에서 발견되지만 이의 세포체는 흑질 (sub st an ti a
nig ra) 과 중뇌 피 개 (mi db rain t eg m e ntu m ) 에 밀 집 되 어 있고, 이 밖에 시 상하부의 궁상핵 (arcuate n ucleus) 에서도 도파민 신경 원이 발견된다. 흑 질에서 나온 축색은 전뇌로 두사되며 기저핵에 있는 신경원과 시냅스 를 형성하는데, 이 회로의 장애가 앞서 언급한 파킨슨 병과 관련 있 다. 기저핵의 도파민 수용기가 차단되어 이 영역이 정상적으로 작용하 지 못하면 만발성 운동장애가 야기된다. 특히 이 장애는 도파민 수용 기가 만성적으로 차단됨에 따라 서서히 나타나게 되는 것으로서 도파 민에 대한 신경결핍성 초과민중 (denerva ti on su p ersens iti v ity)에 기인되 는 것으로 생각된다. 신경결핍성 초과민증이란 신경분포가 차단된 근육에서 처음 발견된 것으로, 신경전달물질의 정상적인 공여가 이루어지지 않은 목표 기관 은 시간이 지남에 따라 그 전달물질에 대해 보다 민감해지는 것을 말 한다. 근육에서의 신경결핍성 초과민증은 시냅스후 수용기 분자의 수 가 증가되어 야기되는 것인데 신경원에서도 근육의 경우와 극히 유사 한 기제가 있을 것으로 추측된다. 클로르프로마진의 장기 투여로 인해 도파민 전달이 장애를 받게 되면 기저핵에 있는 신경원의 민감성이 증 가될 것으로 추측된다. 중뇌의 도파민 신경원은 기저핵뿐만 아니라 전두피질, 대상피질 (cing u la te c ort ex ) , 측두영 의 내 비 피 질 (ento r hin a l cort ex ) 그리 고 해 마 의 여러 영역에도 신경을 분포한다. 항정산병 약물은 이런 뇌 영역에 서도 정상적인 시냅스 상호작용을 방해하는 것으로 생각되지만, 도파 민 가설은 아직 제안 단계에 있을 뿐 실험적으로 직접 검증되지 못하 고있다. 1.7 .1 도파민 가설의 제한점 도파민 가설은 항정신병 약물의 치료효과와 D2 수용기 차단효과 사 이에 높은 정적 상관이 있다는 데 근거한다. 이로 인해 정신분열증 또 는 최소한 정신분열증의 정적 증후가 뇌의 도파민 회로의 과잉활동성 에 의해 유발될 것이라는 견해가 제시되었다.
그러나 정신분열증의 도파민 과잉활동성 가설은 많은 도전을 받고 있다. 예컨대 항정신병 약물이 도파민을 차단함으로써 증후를 감소시 키는 것은 마치 아스피린이 열을 일으키는 감염에는 아무런 영향을 미 치지 못하면서 오직 열만 내리게 하는 작용방식과 유사하다는 것과 같 다 (Carl t on & Manowi tz, 1984). 도파민의 과잉활동성이 정신분열증의 증후를 일으킨다는 이 가설은 도파민 활동성을 증진시키는 약물을 두 여하면 정신분열증 환자의 증후를 더욱 악화시키고, 심지어는 정상인 에게도 정신분열증의 증후를 일으킬 수 있을 것이라는 예측을 가능케 한다. 도파민 시냅스에서 도파민 수준을 증가시키는 것으로 알려져 있 는 약물인 암페타민 (am p he t am i ne) 은 시냅스전 신경원에 의한 도파민 재흡수를 차단함으로써 시냅스 간국에서 도파민 분비 수준을 증가시킨 다. 만약 재흡수 과정이 정상적으로 이루어지면 시냅스 간극에 있는 신경전달물질이 시냅스전 신경원의 축색종말로 완전히 흡수된다. 암페 타민을 장기간 사용하는 환자는 망상형 정신분열증의 증후와 유사한 증후가 나타나기도 하지만, 정신분열증의 다른 유형에서 볼 수 있는 증후와 유사한 특징은 나타나지 않는다. 일반적으로 항정신병 약물은 암페타민에 의해 유발된 정신분열증을 치료하는 데 효과적이라고 한다• 도파민 가설의 또 다른 난점은, 항정신병 약물을 다량 두여하면 도 파민 수용기는 몇 시간 내에 차단되지만 정신분열증의 증후가 감소되 는 데는 보통 2-6 주 정도가 소요된다는 것이다 (Schwa rtz & Africa , 1988). 이와 같이 치료효과가 시간적으로 지연되어 나타나는 현상을 치료지체시간(th era pe u tic lag tim e) 이라고 한다. 만약 항정신병 약물이 도파민 수용기를 차단시키는 작용으로 치료효과가 나타난다면, 약물두 여 즉시 또는 단시간 내에 증후가 완화되어야 할 것이다. 그러나 이런 치료지체 현상을 도파민 가설로서는 설명하기 어렵다. 치료지체에 대 한 여러 가지 설명이 제시되었지만 아직 확실한 것은 없다. 치료지체를 설명하는 하나의 가설로서 탈분국봉쇄 가설 (de po la riz a tion blockade hyp o th es i s) 이 있다. 항정신병 약물의 약리작용은 복합적 이어서 시냅스후 D2 수용기를 차단하는 것 의에도 이 약물은 자기수용
기 (au t orece pt or) 도 차단한다. 이 경 우 자기 수용기 란 시 냅스전 신경 원 에 위치하는 도파민 수용기를 가리킨다. 이 자기수용기의 기능은 아직 완전히 밝혀지지 않았지만 시냅스전 신경원에 의해 이루어지는 도파민 의 합성과 분비가 부적 되먹임을 통해 조절되는 것으로 추측된다 (Coope r, Bloom & Roth , 1986) . 항정 신 병 약물에 의 해 자기 수용기 가 차 단되면 처음에는 도파민 분비 신경원의 흥분발사율이 증가되지만 며칠 이내에 발사율이 현저히 낮아지는데, 이것을 탈분극봉쇄 (dep ol ariz a ti on blockade) 라고 한다. 이런 탈분국봉쇄가 일어나는 데는 며칠 또는 몇 주가 걸리기 때문에 항정신성 약물의 치료효과를 설명해 줄 수 있는 것은 시냅스후 수용기의 차단이 아니라 탈분극봉쇄일 가능성을 배제할 수 없다 (Creese, 1983). 그러나 자기수용기 차단이나 탈분극봉쇄는 아 직 충분히 이해되지 못하고 있으며, 정신분열증에 대한 도파민 회로의 관련성에 관한 것은 아직 더 많은 연구가 요구되고 있다. 1.8 정신수술 1936 년부터 1958 년까지 약 20 년 사이에 대략 35,000 명의 환자(이 중 대부분이 정신분열증 환자)들이 정신장애를 치료받기 위해 정신수술 (ps y chosur g e ry)이라 부르는 전두영 절제수술을 받았는데 (Valenste in , 1980), 이것이 정신치료사상 가장 심각한 오류를 범한 사건으로 기록 되었다. 이런 뇌 수술은 지금도 실시되고 있지만 예전처럼 수술 빈도 는 많지 않으며 엄격하게 통제되고 있다. 최초의 전두영 수술은 포르 루갈의 신경의과 의사인 Eg as Mon i z 에 의해 개발되었다. Mon i z 는 전 두영이 손상된 원숭이와 침팬지가 정서적으로 〈진정 (calmi ng ) > 효과를 보여준다는 동물실험 결과를 보고, 정신과 환자의 전두영을 손상시키 는 뇌 수술법을 생각하게 되었다. 여러 번의 시도 후에 Mon i z 는 두개 골을 천공하고, 이 구멍에 특수한 칼을 삽입하여 전두영과 다른 뇌 영 역을 연결시키는 백질을 손상시키는 수술법을 개발하였다. 그 후 미국 의 Walte r Freeman 과 Jam es Wa tt s(1950) 는 Mon i z 의 방법을 개량한
전두백질 절단술(fr on t al leuko t om y)을 고안하였는데, 이 기법이 널리 사용되었다. 이런 뇌 수술이 여러 차례 시도되었음에도 불구하고 수술 후의 경과 룰 알려주는 데 유용한 자료는 거의 없다. 이 수술을 시도한 신경의과 의사들은 환자를 계속 추적하여 관찰하지 않았고, 비록 관찰을 한 경 우에도 객관저인 관찰이 거의 이루어지지 않았는데, 그 이유는 수술의 결과가 좋지 못했기 때문인 것으로 생각된다. 또한 많은 경우에 있어 환자에 대한 수술 전 분석이 미흡하여 수술 결과에 대한 결론을 내리 기가 어려웠다. 비록 전두백질 절단 환자를 객관적으로 연구한 경우에 도 그 결과를 일반화시키지 못했으며, 수술의 전체적인 결과도 명백하 지 못했다 (Benson 등, 1981). 오늘날에는 정신수술이 거의 시행되지 않고 있으며, 수술 대상부위도 전두영이 아니라 편도체나 대상피질이 되는 경우가 많다. 이 수술이 만성적인 통증을 완화시키는 데 효과적 인 경우도 있지만, 정신분열증 증후의 완화에 효과가 있다는 신뢰성 있는 정보는 거의 없기 때문에 정신수술에 대한 강력한 반대 의견이 제시되고 있다. 2 감정장애 감정 장애 (mood diso rder) 는 정 신분열증보다 더 많이 나타나는 정 신병 이다. 감정장애는 다음 두 가지 범주 가운데 어느 하나의 증후를 보여 준다. 하나는 주 우울장애 (majo r depr e ssiv e dis o rder, DSM-m) 에 서 는 단국성 장애 (monop ol ar dis o rder) 라 하고, 다른 하나는 양극성 장애 (bip o lar d i sorder) 이다. 주 우울증후군의 특칭은 일상적인 활동에서 기 쁨이 상실되는 반면 술품이 지나치게 많아지는 것이 특칭이고, 양극성 장애는 조중 (man in a) 과 울증 (de p ress i on) 이 번갈아 나타나는 것이 특칭 이다.
2.1 주 우울중후군 주 우울증후군으로 진단되기 위해서는 적어도 2 주 동안 다음 9 가지 의 증후군 중 최소한 5 가지의 증상이 나타나야 한다. 사랑하는 사람의 사망이나 갑작스런 재앙(화재나 자동차 사고 등) 뒤에 나타나는 증상들 은 지속기간이 상당히 길지 않으면 우울증이라고 진단하지 않는다. DSM- JJI -R 에 의하면 우울증후군의 9 가지 주요 증상은 다음과 갇다. 1. 거의 하루 종일 기분이 침체된다. 2. 정상적으로 줄거움을 주는 활동(성,운동,놀이) 등에 대한 홍미나 줄 거움이 현저히 감소된다. 3. 다이어트를 하지 않아도 체중의 증감 (5% 이상)이 심하다. 4. 불면증 또는 과수면 5. 정신성 운동(p s y chomo t or) 의 흥분이나 침체 6. 피로 또는 활력의 감소 7. 무가치하다는 느낌 또는 지나칠 정도로 부적절한 죄의식 8. 사고 능력이나 주의집중 능력의 저하 또는 결단력의 부족 9. 반복적으로 죽음이나 자살을 생각한다. 2.1.l 주 우울증후군의 약물치료 정신분열증과 달리 주 우울증후군은 약물을 사용하지 않고도 치료가 되는 경우가 있다. 여러 유형의 정신치료가 도움이 될 수도 있고 REM 수면을 박탈하는 것도 우울증을 완화시킬 수 있다. 대개의 경우 약물을 사용하면 회복과정이 촉진되지만, 어떤 경우에는 약물요법이 환자에게 도움이 되지 못하는 수도 있다. 우울 치료에 사용되는 약물 은 여 러 종류가 있는데 가장 중요한 두 가지 의 항우울제는 tri c y c l ic s 과 MAO is ( monoa rnine oxid a se inh ib it or s) 가 있는데 , 전자에 는 이 미 프 라민(imip r ami ne) 이 대표적이고, 후자에는 이소카르복사지드(i socarbox az i d) 와 페노티아전(p heno thi az i ne) 이 대표적인 약물이다.
미국의 국립 정신건강 연구원 (NIMH) 에서 250 명의 환자를 대상으로 16 주간 실시한 한 연구에 의하면, 환자와 주기적으로 만나 병의 경과 롤 살펴보고 환자의 질문에 대답해 주는 임상관리와 함께 이미프라민 을 두여하면 우울증 치료에 효과적이라고 한다. 실제로 이 연구에서는 약물-임상관리 환자집단, 개인심리 치료집단 그리고 인지행동 치료집 단의 3 가지 치료집단과 통제집단으로 위약-임상관리 집단이 연구되었 는데, 위의 3 가지 치료집단이 위약-임상관리 통제집단보다 더 높은 회 복률을 보여주었지만 우울증이 십한 환자의 경우에는 약물-임상관리 집단의 환자들이 다른 두 집단의 환자들보다 더 빠른 치료효과를 보여 주었다 (Elk i n 등, 1989). t r icy cl ic s 과 MAOI 는 뇌 속 중요 회로의 시냅스에서 모노아민성 신 경전달물질의 가용성을 증가시킴으로써 우울증을 완화시키는 것으로 추측된다. 모노아민성 신경전달물질에는 노르에피네프린, 에피네프린, 도파민 그리고 세로토닌이 포함된다. 이들 전달물질 가운데 어느 것이 정서장애와 가장 밀접하게 관련되는지는 아직 분명치 않다. tric y cl ics 와 MAOI 는 모두가 신경전달물질의 가용성을 증가시키지만 작용기제 는 서로 다르다. t r icy cl ic s 는 시냅스전 신경원에 의한 모노아민의 재 흡수를 차단하여 시냅스 간극에서 전달물질의 작용기간을 증가시키는 작용을 하는 것으로 생각된다. 한편, MAOI 는 정상적으로 모노아민을 약화시키는 작용을 하는 모노아민 산화효소 (monoa mine oxid a se : MAO) 라는 효소의 작용을 억제시킴으로써 모노아민의 시냅스 효율성 을 높인다. 모든 항우울제가 tric y cl ic s 또는 모노아민 산화효소 억제 제 계열에 속하는 것은 아니며, 모노아민성 신경전달물질의 가용성에 영향을 미침으로써 우울증 치료효과를 나타내는 것도 아니다. 예를 들 어 이프린돌(ip r i ndole) 이라는 속명을 가진 약물은 모노아민의 재흡수 나 효소의 분해작용은 억제하지 않지만 효과적인 항우울제로 간주된 다. 또한 코카인과 같은 약물은 재흡수를 차단하지만 항우울제로는 효 과가없다.
2.1 .2 항우울제의 부작용 대부분의 다른 약물과 마찬가지로 항우울제도 부작용을 나타낸다. 변비, 오줌의 잔류 그리고 시각이 희미해지는 것과 같은 증후들은 건 강한 사람들에게는 비교적 경미한 문제로 약물을 계속해서 사용함에 따라 사라지는 경향이 있으나, 뜻하지 않게 체중이 증가되는 부작용도 있다. 이보다 심각한 부작용으로는 심장기능 및 혈압의 비정상성이 나 타나 생명을 잃는 경우도 있지만 그 가능성은 매우 희박하다. 그러므 로 환자에게 항우울제를 처방할 때는 과거의 심장질환 여부를 반드시 알아보아야 한다. 특히 나이가 많은 환자들은 우울증과 심장질환을 모 두 가지고 있을 가능성이 높기 때문에 항우울제 처방시에 더 세십한 주의가 필요하다. 정신분열증이나 양국성 우울증을 가진 환자에게 항 우울제를 투여하면 심한 격앙 상태나 조증을 일으킬 수 있기 때문에 보다 정확한 정신과적 진단이 중요하다. 항우울제 처방시에는 의도적 또는 비의도적인 과량 복용의 가능성도 고려해야 한다. 대부분 항우울제의 치사량은 일일 복용량의 10-15 배 정도이므로 환자에게 한 번에 1 주일치 이상의 약물을 처방해 주지 말 아야 한다 (L ic key & Gordon, 1991). 2.1. 3 전기경련 치료 십한 우울증을 치료하기 위해 많이 사용되는 방법의 하나로 전기경 련 치료법 (electr o convulsiv e the rapy : ECT) 이 있다. ECT 의 사용에 대 해서는 많은 논란이 있지만 우울증이 매우 심한 경우 최후 수단으로 사용된다. 따라서 ECT 는 다른 치료법으로 효과가 나타나지 않는 환 자나 자살의 위험이 높은 환자에게만 사용하는 것이 일반적이다. ECT 는 짧은 시간 동안 머리에 전류를 홀려보내는 방법인데, 이때 홀 리는 전류는 환자의 뇌에서 대간질발작과 유사한 전기적 활동을 일으 킬 정도로 강해야 한다. ECT 를 사용하면 근육경련도 나타나는데 이 것은 근육이완기법을 적용하면 없앨 수 있다. 치료 전에 작용기간이 짧은 바비두레이트 (barb it ura t e) 로 환자를 진정시켜 ECT 후에 일어날
결과를 예상할 수 없게 해야 한다. ECT 는 매주 3 회씩 3 주 동안 실시 하는 것이 가장 일반적이다. ECT 는 심한 우울증 환자 중 약 70% 정 도의 환자들에서 치료효과가 나타나며, 정신병적 사고가 동반되는 우 울중에 대해서는 유일한 치료방법이 되지만 다른 정신장애에는 유용하 지 않다. ECT 가 비교적 안전한 치료방법이지만 치료 후 0.1% 이하의 환자가 사망하는 약간의 위험성도 있다. ECT 의 부작용으로는 두통과 치료 직전에 일어난 사건에 대한 기억상실인데, 이 기억상실은 경우에 따라 회복되기도 하고 회복되지 않을 수도 있다. 한 연구결과에 의하면 치 료 전 3 년 동안의 기억이 완전히 사라지는 역행성 기억상실증도 나타 날 수 있다고 한다 (S q u i re, Slate r & Chace, 1976). 이런 기억장애는 전 류가 뇌의 어느 한쪽 면에만 가해지면 감소될 수도 있다. 그러나 이런 단측성 ECT 를 사용하면 우울증 완화효과가 다소 떨어지며, 더 많은 치료횟수를 필요로 한다 (Goldman, 1988) . 정신장애를 치료하는 데 있어서 어떤 치료법의 효용성은 전혀 치료 를 받지 않았을 때의 위험 정도와 비교되어 평가되어야 한다. ECT 를 옹호하는 사람들은 자살의 위험이 ECT 가 갖는 위험보다 더 크다고 주장한다. 우울증 환자의 자살률은 정상인의 약 20 배에 달하는 것으로 추정된다. 2.1. 4 우울증의 모노아민 가설 t r icy cl i cs 과 MAOI 가 시냅스에서 모노아민성 신경전달물질의 가용 성을 증가시킴으로써 우울증의 치료효과를 나타낸다는 사실에 근거하 여 우울증의 모노아민 가설 (monoami ne hyp o th e sis of depr e ssio n ) 이 제 기되었다. 이 가설이 갖는 기본 논리에 의하면, 우울증이란 뇌에서 한 가지 이상의 모노아민성 신경전달물질의 수준이 낮아지는 것과 관련 있고, 반대로 조증은 모노아민의 수준이 지나치게 높아지는 것과 관련 있는 것으로 간주한다. 어떤 연구자들은 노르에피네프린이 가장 중요 한 모노아민성 물질일 것이라고 하였고, 다른 연구자들은 세로토닌이
가장 중요한 것이라고 제안하였다. 또 다른 연구자들은, 우울증은 두 가지 유형이 있는데, 하나는 세로토닌 부족에 기인하고, 다른 하나는 노르에피네프린의 부족에 기인되는 것이라고 주장하였다. 도파민도 모 노아민에 속하지만 많은 연구자들은 이 물질이 우울증에 관여한다는 데는 의문을 가지고 있다. 우울증의 모노아민 가설을 지지하는 발견 가운데 하나의 근거는 레 세르핀 (rese rpi ne) 과 관련되어 나왔다. 1950 년대에 레세르핀은 고혈압 치료에 사용되었는데, 이 약물로 치료를 받은 고혈압 환자 중 10-15% 의 환자들이 심한 우울증을 보인다는 사실이 밝혀졌다. 한편 이프로니 아지드(ip ron i az i d) 라는 약물이 결핵치료를 위해 사용되었는데, 이 약 물을 두여받은 일부 환자들이 노래하고 춤춘다는 사실이 발견됨에 따 라 이 약물이 기분을 극적으로 상승시키는 효과를 가지고 있음을 알게 되었다. 그 후 연구들에 의하면 레세르핀은 시냅스에서 모노아민의 가 용성을 감소시키는 반면 이프로니아지드는 모노아민의 가용성을 증가 시킨다는 사실이 드러나 (Bunne y & Davis , 1965 ; Schil dk raut, 1965) 이들 약물의 효과가 바로 모노아민 가설과 관련 있을 것으로 추측되었다. 2.1. 5 모노아민 가설의 제한점 모노아민 가설에는 비판의 여지가 많다. 예를 들어 다른 항정신병 약물과 마찬가지로 항우울제도 투여한 직후 시냅스에서 모노아민의 가 용성은 즉각적으로 증가되지만 우울증의 치료효과가 나타나는 데는 2- 3 주 정도가 걸린다는 점이다. 또한 새로 개발된 효과적인 항우울제가 모노아민 시냅스에 별다론 영향을 미치지 못하는 경우도 있고, 반면 모노아민의 가용성을 증가시켜 주지만 항우울제로는 효과가 없는 약물 (예, 코카인)도 있다. 한편 어떤 모노아민 가설도 양극성 장애에 대한 리튬영 (lithiwn salt) 의 뛰 어난 치료효과를 쉽 게 설명 해 주지 못한다. 어떤 가설이 유용한 것이 되기 위해서는 검증될 수 있어야 하지만 대 부분의 모노아민 가설은 쉽 게 검증되 지 않는다. Larr y S i ever 와 Kenneth Dav i s(1985) 는 모노아민 가설을 검토하여 보다 자세한 가설
울 제안했는데, 그 내용은 우울증이 모노아민 체계의 저활동성 때문에 생기는 것이 아니라 이 체계의 조절장애에서 비롯된다고 지적했다. 이 가설은 직접 검증될 수 있는 과학적 가설로서 앞으로 많은 연구가 뒤 따를 것으로 기대된다. 2.2 양극성 장애 정 서 장애 의 또 다른 유형 은 양극성 정 서 장애 (bip o lar affe ctive dis o rder) 이다. 이 장애를 가전 환자들은, 우울장애는 물론 조증 에 피 소드도 보여준다. DSM-//1-R 에 의하면 이 장애의 전단에는 다음과 같은 세 가지 기준이 고려된다. 1. 일정 기간 동안 비정상적이거나 지속적으로 고양되고, 과대망상적이 고 쉽게 흥분되는 기분을 보인다. 2. 이 기간 동안 다음 중 최소한 세 가지 증후가 출현한다. a) 지나치게 과장된 자존심과 당당함 b) 수면 욕구의 감소 c) 평상시보다 수다스러움 d) 관념이나 사고의 비약 e) 지나친 주의 산만 f) 목표 지향적인 활동(학교나 직장에서의 성적 활동이나 사회적 활동) 의 증가 또는 정신성 운동의 증가 g) 고통스러운 결과가 쉽게 예상되는 쾌락적인 활동에 몰두, 예컨대 무분별한 상품 구매, 성행위 또는 어리석은 사업두자 등에 몰두한다. 3. 일상의 과업수행이나 대인관계를 방해할 정도로 십한 정서장애의 출현 양국성 장애는 주 우울층후군보다는 출현 빈도가 낮다. 미국의 경우 남자의 10%, 여자의 20% 정도가 일생 중 가끔 최소한 한 가지의 주 우울증후군 에피소드를 경험하지만 양극성 장애를 경험하는 환자는 전 체 인구의 1% 정도에 불과하며, 이환율에서 성차는 거의 보여주지 않
는다 . 2.2.l 리튬 치료 양극성 장애를 치료하는 가장 일반적인 방법은 리듐염을 두여하는 것이다. 1949 년에 호주의 정신과 의사인 Joh n Cade 가 양극성 장애 치 료에 이 약물을 최초로 사용하였다. Cade 는 흥분을 잘 하는 기니피그 에게 이 약물을 주사하여 예측하지 못했던 전정효과를 관찰하였다. 그 는 즉각적으로 이 약물을 조증 상태에 있는 10 명의 양극성 환자와 6 명 의 정신분열증 환자 그리고 3 명의 우울증 환자에게 투여한 결과 조중 환자에서만 증상이 개선됨을 발견하였다(Li cke y & Gordon, 1991 ). 그 후 이 약물은 유럽을 중심으로 사용되다가 오늘날에는 전세계적으로 양극성 장애의 치료약물로 사용되고 있다. 리듐이 모든 환자들에게 효과적인 것은 아니다. 약 20% 정도의 환 자들에서는 증상이 거의 개선되지 않지만 아직 다른 대안적 치료방법 은 없다. 왜 리튬의 치료효과가 환자의 80% 에서만 나타나는지 그 이 유는 아칙 잘 모른다. 조직배양 실험을 한 결과 리듐은 세포에 복합적 인 영향을 미치는 것으로 알려져 있는데, 이노시톨 3 인산(i nos it ol t r ip hosp ha t e) 이라는 제 2 전령체계와 상호작용하는 것 갇다. 세포에 미 치는 이런 효과가 어떻게 양극성 환자의 조중 기분을 낮추는지는 분명 치 않다. 한 가지 가설은 이토시톨 3 인산이 신경전달물질에 대한 대뇌 신경원의 반응성에 중요한 작용을 하기 때문에 리튬이 신경전달물질에 대한 대뇌 신경원의 반응성을 조절하여 뇌의 활동성을 정상화시킬 것 이다. 리듐과 같은 약물이 세포에 미치는 기본적안 효과에 대해 더 많 이 알수록 양극성 장애와 같은 뇌의 기능 부조의 근원을 이해할 수 있 게 될 것이다.
3 불안장애 흔히 현대를 불안의 시대라고 한다. 사람이 살아가는 데는 늘 걱정 거리가 있기 때문에 불안을 생활의 일부분으로 생각할 수도 있다. 그 러나 임상심리학자나 정신과 의사들은 불안이 심각한 문제가 되는 다 양한 심리적 문제를 다루고 있다. 정신분열증이나 우울증 환자들도 불 안을 호소하는 경 우가 있으나, 불안장애 (anxie ty d i sorder) 는 독특한 형 태의 심리장애로 간주된다. DSM - 111-R 에서는 불안장애를 공황장애 (pa nic dis o rder) , 공포증 (ph obia ) , 강박장애 (obsessiv e comp ul siv e dis - order) 및 일반화된 공황장애 (ge neraliz ed pan ic dis o rder) 등으로 분류하 고 있다. 1984 년에 시행된 임상장애의 발병에 관한 한 조사에 의하면 조사대상자 중 8% 이상이 불안장애를 가지고 있는 것으로 나타나 가 장 높은 발병률을 보여주었다 (M y ers 등, 1984) . 3.1 공황장애와 치료 공황발작(p an ic a tt ack) 은 불안 효과가 극단적이고 집중적인 형태를 띠는 것이 특징이다. 공황발작의 증후는 거친 호홉, 현기증, 심장박동 의 증가, 신체 의 떨림 (trem bli ng ) , 발한, 자신이 미 쳐 날뛴다는 강한 공포, 통제감의 상실 등과 같이 다양하게 나타난다. 공황발작은 다가 울 어떤 사건에 대한 예상에 의해 야기될 수도 있고 어떤 특별한 경고 없이도 나타날 수 있다. 발작의 지속시간은 몇 분 정도에 불과한 것이 보통이지만 매우 파괴적이고 불쾌하다. 환자는 자신이 공황을 경험하 고 있다는 것을 인식하지 못하고 단지 강한 불쾌감을 느낄 뿐이라고 생각한다 . 이런 증후들은 교감신경계의 활성화에 의해 파생된 것이다. 공황발작을 경험한 많은 사람들은 공황발작이 닥쳐오면 피하기가 어렵 거나 당황하게 만든 상황에 의해 생긴 만성적인 공포로 시달립을 받는 다. 이 런 조건 가운데 하나가 광장공포증 (a g ora p hob i a) 이다. 광장공포 증은 흔히 집을 떠나거나 혼자 여행하는 것에 대한 공포가 심한 것이
특칭이다. 십한 경우 이 공포증 환자들은 오직 자기 집에만 갇혀 있는 신세가 된다. 공황발작의 특칭은 빠른 심장박동, 발한, 구역질, 한기, 떨림 그리고 잘 조절되지 않는 어떤 것을 행하는 것에 대한 공포 등으 로 발작 초기의 심한 에피소드로는 질식, 실신, 가슴앓이 그리고 무기 력 과 같은 증후들이 나타난다 (Greis t & Jef fe r son, 1988) . 불안을 치료하기 위한 여러 가지 방법들이 시도되어 왔으나 그 중 행동적 치료방법이 가장 유용한 것으로 입증되었다. 이 치료에서 환자 는 보조 치료자의 역할을 하는 친구나 가족이 있는 친숙한 상황에서부 터 공황을 일으키는 상황으로 점진적으로 노출시켜 간다. 약물치료도 많이 사용되는데 노르아드레날린의 재흡수를 차단하는 항우울제가 환 자들에게 도움이 된다. 불안장애 치료에 특히 효과적인 벤조디아제핀 (benzod i aze pi nes) 계열 의 약물들이 최근에 많이 개발되었다. 이 계열에 속하는 대표적인 약 물로는 alpr a zolam (Xanax) , chlordia z ep o xid e (Lib r iw n ) , dia z epa m (Vali wn ) 등이 있다. 벤조디아제핀 계열의 약물결합 부위는 동물과 인 간의 뇌 조직 모두에서 발견된다. 최근 증거에 의하면, 결합부위가 GABA 수용기 복합체 (GABA recept or comp le x) 라는 신경 원의 막구조 와 관련되어 있다고 한다. 앞 장에서 언급한 것처럼 GABA 는 뇌에 분포된 억제성 신경전달물질이며, 뇌의 모든 신경전달물질 가운데 가 장 광범위하게 분포되어 있다. GABA 수용기 복합체는 최소한 4 개의 상이한 영역으로 구성되어 있는데, 첫째 영역은 GABA 의 실제적인 수용기로서 단백질 이온동로와 관련되어 있는 영역이다. GABA 가 이 수용기와 결합하여 개방되는 이온통로는 염소 이온에 대해 특이성을 갖는다. 앞 장에서 본 것처럼 염소 통로가 개방되면 시냅스후 세포의 활동이 억제된다. GABA 수용기의 두번째 영역은 염소 이온통로이 며, GABA 수용기의 세번째 및 네번째 영역은 불안치료에 사용되는 벤조디아제핀이나 바르비투르산염 (barb it ura t e) 에 선택적인 결합부위를 갖는 곳이다. 따라서 이 두 가지 유형의 약물은 GABA 수용기 복합 체의 여러 부분들과 상호작용한다.
벤조디아제핀은 보다 안전하고 부작용이 적기 때문에 바르비무르산 영보다 불안장애 치료에 더 자주 사용되고 있다. 벤조디아제핀은 뇌 전체에 퍼져 있는 시냅스들에서 GABA의 억제효과를 보다 촉진시키 는 작용을 한다. 죽 이들 약물은 GABA 수용기 결합부위와 직접 상 호작용하는 것이 아니라 GABA 수용기와 결합하기 위한 능력을 증가 시킨다. 만약 GABA 라는 전달물질이 없으면 벤조디아제핀도 효과적 으로 작용할 수 없다. GABA 의 광범위한 억제효과가 전체 뇌에 미치 기 때문에 벤조디아제핀의 불안장애 치료효과가 나타나는 것이다. 벤 조디아제핀은 공황장애와 일반화된 불안에 효과적이지만, 강박장애에 는 또 다른 계열의 약물이 효과적인 것으로 밝혀졌다. 뇌 속에는 불안을 일으키는 자생물질도 있다. 이러한 물질이 최근에 와서 분리 되 었으며 , 이 물질을 디 아제 팜 결합억제 제 (dia z epa m bin d in g inh ib i t or : DBI) 라고 한다. 이 물질은 벤조디아제핀의 결합부위에서 벤 조디아제핀과 서로 경합함으로써 벤조디아제핀의 효과를 억제하는 작 용을 한다. DBI 를 실험동물에 주사하면 벤조디아제핀과 갇은 항불안 제의 불안감소 효과를 약화시킨다. 한편 DBI 를 불안 펩티드 (anx i ety p e pti de) 라고 하는데, 이 물질은 시상하부, 해마 그리고 편도체의 신경 원에 고농도로 분포되어 있기 때문에 이들 뇌 영역이 불안과 관련되는 대뇌 부위일 것으로 추측한다. 3.2 강박장애 강박장애 (obsessiv e comp ul siv e dis o rder : OCD) 는 또 다른 유형 의 불 안장애로 불안장애 중 치료가 가장 어려운 것으로 간주되어 왔다. 그 러나 최근에 와서 과거 항불안제로 사용되던 약물들이 OCD 환자를 치료하는 데에도 효과적이라는 사실이 밝혀지고 있다. 강박관념 (obsessio n ) 과 강박행 동 (com p uls i on) 은 비 록 OCD 의 상이 한 측면이 지 만 일반적으로 함께 나타난다. 일반적으로 강박적인 행동은 강박관념에 의해 유발되는 불안을 감소시키려는 일종의 의식(儀式)화된 행동이다.
그러나 환자에 따라서는 강박관념은 있지만 강박적인 행동이 나타나지 않는 경우도 있다. 또한 어떤 환자들은 강박행동을 야기시켰던 강박관 념이 약화되거나 사라진 후에도 강박행동을 계속 보이기도 한다. 강박관념은 반복적인 사고, 심상 또는 충동을 야기하여 개인을 혼란 스럽게 함으로써 불안을 유발한다. 정신분열증 환자도 강박적 사고를 경험하는데 이들은 자신의 사고를 동제하는 강박관념이 의부의 힘 때 문에 생긴 것이라고 여긴다 . 그러나 OCD 환자들은 그들의 강박관념 이 내적으로 생긴 것이라고 인식한다. 강박관념은 혼히 폭력(타인을 해 치거나 죽인다), 오염(악수 또는 문의 손잡이를 만져서 세균에 감영된다), 또는 사회규범의 위반(부끄럽거나 웃음거리가 되는 어떤 것을 말하거나 행 한다)에 대한 사고가 반복된다. 강박적인 의식은 강박관념에 의한 불안울 방어하기 위해 환자가 하 루에도 수십 번 또는 수백 번씩 수행하는 반복적인 행동을 말한다. 그 러므로 하루에 손을 수백 번씩 씻는 것은 타인과 악수를 하거나 문의 손잡이룰 만침으로써 자신의 손이 오염되었을 것이라는 강박관념에서 비롯되는 전형적인 강박행동이다. 비극적인 사건이 일어날지도 모른다 는 강박관념 때문에 가스 난로가 꺼져 있는가 또는 문과 창문이 참겨 져 있는가를 계속 확인하는 강박행동이 나타나기도 한다. 강박관념-강 박의식이 환자의 생활을 지나치게 압도하여 자신의 직업이나 사회생활 이 정상적으로 유지될 수 없을 정도로 심한 경우도 있고, 비록 많은 어려움은 있지만 사회생활에서 자신의 기능을 기본적으로 수행할 정도 로 강박행동의 반복횟수가 최소로 유지되는 경우도 있다. OCD 환자들은 강박적 행동을 통해 즐거움을 느끼지도 못하며, 불 안감소도 피상적이거나 일시적인 것에 불과하기 때문에 강박행동을 반 복한다. 따라서 OCD 는 흔히 언급되는 강박적 도박이나 강박적 음주 와는 다른 장애인 것으로 간주되는데, 그 이유는 이런 행위들은 비록 일시적으로나마 쾌감이나 스릴울 가져다 주기 때문이다. OCD 환자는 인구의 약 2% 에 해당될 정도로 발생빈도가 적지 않다 (Rap a p o rt, 1989).
3.2.1 OCD 의 치 료 OCD 환자의 치료에 도움이 되는 방법은 행동치료이다. 예컨대 오 염에 두려움을 가진 환자의 경우 이 환자가 전화를 받거나 문의 손잡 이를 만질 때 장갑이나 다른 보호장치를 사용하거나 물건을 만전 후 최소한 3 시간 동안은 손을 씻지 못하도록 하는 따위의 행동치료가 큰 도움이 된다. 일단 의식적인 행동이 낮은 수준으로 안정되면 클로미프 라민 (clom ip ram i ne) 을 복용시키는데, 이 약물은 매우 효과가 있다 (Greis t & Jef fe rs on, 1988) . ana fr an il이 라는 상품명 을 가진 이 약물은 다른 tric ycl ic 항우울제와 밀접한 관련성을 가지며 간혹 항우울제로도 사용되지만 우울치료 효과는 다소 뒤떨어진다. 항 -OCD 약물들은 강박관념과 강박행동 모두를 치료할 수 있으며 행동치료보다 더 나은 치료효과를 나타내는 것으로 간주한다. 행동치 료는 강박관념이 아니라 문제시되는 강박행동만을 치료대상으로 하기 때문에 새로운 강박행동이 발달될 가능성은 여전히 남아 있다. 따라서 행동치료와 약물치료를 병행하여 사용하는 것이 OCD 환자에게 더 많 은 도움을 줄 수 있다. 클로미프라민은 OCD 의 한 유형인 강박적으로 자기 의 머 리 털을 잡아당기 는 발모광(t r i cho till oman i a) 치료에도 유용하 다. 이 발모광 환자들은 자신의 머리털을 한 번에 한 가닥씩 잡아당겨 뽑아버려 대머리가 된 환자가 많다. 이 경우 여러 가지 항우울제 중 클로미 프라민만이 치료효과를 나타낸다고 한다 (Ra p a p o rt, 1989) . 양전자 방출사진술을 사용하여 OCD 환자의 뇌를 검사해 보면 전두 엽 그리고 전두영과 기저핵 사이의 신경통로에서 높은 수준의 포도당 대사가 관찰된다. 그러나 이들 통로가 항 -OCD 약물의 영향을 받는지 여 부는 아직 잘 모른다 (Ra p a p o rt, 1989) .
4 아동기 장애 아동기에 나타나는 행동장애도 있다. 그 중 가장 혼한 장애는 주의 력결핍 과잉활동장애 (att en ti on -defic it hyp e racti vit y dis o rder : ADHD) 로 아동의 약 3% 정도에서 나타난다. DSM-III-R 에서는 이 장애에 포함 되는 14 가지의 증후를 열거하고 있으며, 6 개월 동안 이 중 8 가지 이상 의 증상이 나타나면 ADHD 로 전단된다. 안절부철못하고, 가만히 앉 아 있기 어려우며, 쉽게 주의가 산만해지고, 자기의 차례를 기다리지 못하며, 학교생활이나 놀이에 주의를 집중하지 못하고, 타인을 방해하 거나 간섭하며, 타인의 말을 잘 경청하지 못하고, 물건을 잘 잃어버리 고, 주위를 잘 살피지 않고 도로로 뛰어드는 것과 같은 신체적으로 위 험한 행동을 하는 것이 이들 증후에 해당된다. ADHD 는 보통 7 세 이 전에 나타나며, 발병률에서는 소녀보다 소년이 6-9 배나 높다. 많은 경 우 청년기가 되면 증상이 호전되지만 성인이 되어서도 여전히 충동을 억제하기 어려운 환자도 적지 않다. 아동기의 ADHD 증후의 출현과 성인기의 병적인 도박 사이에 높은 상관이 있다는 견해도 제시된 바 있다 (Carl t on 등, 1987). ADHD 의 전단에는 논란이 많다. 교사에게는 비정상적인 것으로 보 이는 아동의 행동이 부모에게는 정상적인 것으로 보일 수 있으며, 어 떤 증후들은 가정환경의 변화나 수면 부족과 같은 원인에 의해 나타날 수도 있다. 그러나 ADHD 가 아동에서 나타나는 매우 현실적이고 공 통적인 문제라는 데는 의문의 여지가 없다. 이 장애의 치료를 위해서 는 가정교육과 행동치료를 병행하면서 환자에게 흥분제를 투여한다. 과잉활동을 보이는 아동에게 흥분제를 사용한 역사는 1930 년대부터 이다. 그 당시 Charles Bradle y를 비롯한 연구자들은 과잉활동성을 보 이는 어린이들에게 벤제드린 (benzed ri ne) 이라는 흥분성 약물을 투여하 여 진정효과를 관찰하였다. 오늘날 ADHD 치료에 가장 널리 사용되 는 약물은 meth y l p h e nid a te ( Ri tali n ) , dextr o amp h eta m i ne (Dexderin e} , magn e siu m pem oli ne (Cy le rt ) 등 세 종류이 다. 이 런 흥분제 가 아동의
과잉활동성 치료에 효과가 있다는 것은 이해하기 어려운 역설적인 현 상이다. 아동의 과잉활동성 치료에 흥분제가 사용된 것은 반 세기가 넘었지만 아직도 흥분제의 작용 양상에 대해서는 잘 알지 못한다. 아 동은 혼히 약물에 대해 성인과는 다른 방식으로 반응한다. 이 약물들 은 노르에피네프린과 도파민이 전달물질로 사용되는 시냅스에 영향을 주는데, 이들 흥분제의 주작용은 시냅스에서 이 전달물질의 재흡수를 차단함으로써 전달물질의 양을 증가시키거나 전달물질의 방출을 증가 시키는 것으로 생각된다. 바로 이 기제가 이들 흥분제의 ADHD 의 치 료효과에도 해당되는지는 현재로서는 알 수 없다. 초기 아동기 에서 나타나는 또 다른 장애로 유아자폐증(i n fa n ti le au- ti sm) 이 있는데, DSM-III-R 에서는 이 장애를 폐장애 (auti sti c diso rder) 로 분류하고 있다. 자폐장애는 2,500 명의 이동· 중 1 명 정도로 비교적 드물게 나타나며 환자의 대부분이 남자아이이다. 이 장애는 매우 심각 하고도 괴로운 장애로서, 비정상적인 언어 발달, 정상적인 사회관계 형성의 실패, 홍미의 대상이나 활동범위의 협소성 등이 특칭적인 증후 로 나타난다. 자폐아동들은 다른 사람이 감정을 가진 존재라는 사실을 알지 못하여 물리적 대상으로 여기며, 동료 아이들이 많이 모인 방에 서도 혼자 논다. 또한 이 아이들은 벽에 머리를 들이받거나 의식화된 운동을 하는 따위의 이상한 몸짓도 보여준다. 언어장애가 공통적으로 나타나는데, 자페아동들은 같은 단어를 반복적으로 이야기하고 대명사 의 사용에서 혼동을 보인다(〈나 〉 라고 해야 할 때 〈 너 〉 라고 한다). 주위에 있는 물건이 옮겨지거나 일상적인 일이 중단되면 심한 긴장을 나타내 기도 한다. 자폐아동들이 반드시 정신지체를 보이는 것은 아니며, 음 악이나 산수 계산과 같은 특정 분야에서는 놀랄 만한 능력을 가지고 있는 경우도 있지만, 일반적으로 전체적인 산술능력은 상당히 결여되 어 있다. 1943 년 Leo Kanner 가 이 장애를 최초로 자세하게 기술하였 다. 자폐증의 칭후는 보통 생후 1 년 이내에 나타난다. 이 아이들은 미소 가 결핍되어 있고 다른 사람이 껴안으면 경직되고 포옹에 대해 별 관
심을 보이지 않거나 거절한다. 신체 접촉에 관심을 보이지 않거나 불 쾌한 반응을 보이기 때문에 부모는 걱정을 하게 되고, 또 이런 반응 때문에 다른 사람들이 자폐아동과 접촉하거나 포옹하는 것을 꺼려한 다. 자폐아동이 성장함에 따라 그들의 비정상적인 언어발달과 사회관 계 결핍은 더욱 심해진다. 자폐증에 대한 진단은 아동이 3 세경에 이르 면 결정된다. 한때 자폐증은 냉담한 부모의 육아태도에서 비롯되는 것 으로 생각되었으나 이 견해는 그 후 큰 지지를 받지 못했다. 이 견해 에 반대하는 주장 중 하나는 자페아동들의 형제들은 보통 정상이라는 것이다. 대부분의 연구자들은 뇌의 어떤 비정상성이 자폐증을 유발시킬 것이 라고 추측하지만 이 가설은 거의 검증되지 않고 있다. 자폐아동들이 심한 언어장애를 보인다는 사실은 대뇌 좌반구의 발달에 문제가 있을 것(F이ein라 등는, 점19을84 ;시 K사ins하 bo지um만e &이를 Hi sc뒷 ~받ck침, 1해98 3주). 는어 떤구 체자적폐인 아 동증들거은는 E없E다G 기록으로 볼 때 클릭 소리에 대해 비정상적인 전기적 반응을 보여주는 데, 이것은 뇌간의 청각중추에서 비정상적인 정보처리가 이루어지고 있다는 것을 의미한다. 이와 같은 비정상성은 자폐 장애의 초기 진단 에 유용하게 사용될 수 있다고 한다 (Fe i n, Skoff & Mi rs ky, 1981). 행동 수정 프로그램이 자폐증 치료에 약간의 효과를 나타내기는 하지만 현 재로서는 만족할 만한 치료방법이 없는 듯하다.
참고문헌 김현태, 류재욱, 김기석 (19 91). 「순막조건화에서의 소뇌 피질의 기능」. 《 한국심 리학회지 : 생물 및 생리 》 , 3, 134- 14 9. 김현택, 최준식, 김기석 (1991). 「순막조건화에서의 적핵과 소뇌 증간핵의 기능」. 《 한국심리학회지 : 생물 및 생리 》, 3, 65-82. 손전훈, 오애 령 (19 92) . 「고속핵 자극에 의해 유발되는 통각억제 에 관련되는 상행 체계」. 《 한국심리학회지 : 생물 및 생리 》 , 4, 14-21. 이종택, 김기석 (1990). 「편도체 중심핵 손상이 갈등 스트레스에 의해 유발되는 위궤양에 미치는 영향」. 《 한국심리학회지 : 생물 및 생리》, 2, 100- 10 8. 장현감(1 984). 『 격리성장과 행동장애 』 . 영남대학교 출판부. 장현갑, 강성군, 배중철(1 990). 「 Halo p e ri dol 및 L-do p a 가 생쥐의 스트레스성 위 궤양 발생에 미치는 영향」. 《 한국심리학회지 : 생물 및 생리 》 , 2, 60-68. 장현감, 박순권, 강성군 (1991). 「 S t ress 성 위궤양 발생 기제에 관한 실험적 연구 : 격 리 성 장동물의 위 궤 양발생 에 미치는 Do p am i ne 성 물질의 영 향」. 《한국심 리학회지 : 생물 및 생리 》 , 3, 11-21 . 장현갑, 윤병수(1 991). 「편도중심핵에 대한 Do p am i ne 성 물질의 두여가 스트레스 성 위궤양에 미치는 영향」 . 《 한국심리학회지 : 생물 및 생리》, 3, 32-40. 장현감, 윤병수, 박순권(1 992). 「편도체 중심핵의 do p am i ne 성 체계가 스트레스 장현갑,위 궤임양호에찬 ,미 치장는경 영희향 」(1.9 89《) 한. 국「십배리내학측 회시지상 :핵 생 손물상 및이 생학리》습,에 4 , 미8치3-는96 . 영향」. 《 한국십리학회지 : 생물 및 생리 》 , 1, 18-25. 장현감, 정봉교, 박순권 (1988) . 「출생 전 MAM 두여가 흰쥐의 중추신경계발달 및 행동에 미치는 영향」 . 『 瑞艦宋大炫敎授 華甲記念論文集 』 , 13-26. Adolph , E.F . (19 50) . Thir s t and its inh ib i t ion in the sto m ach. Americ an Jou rnal of Phys iolo gy , 161 , 374-386. Akelait is, A,J . (19 44) . A stu d y of agn o sis, pra xis , and lang ua ge foll ow ing secti on of the corp u s callosum and ante r io r commi ss ure. Jou rnal of Neurosurg er y , 1, 94-102.
Aki! , H ., Wats o n, S,J ., Y oung, E., L ew is, M .E., K hachatu r ia n , H. & Walker, J. M. (19 84) . Endoge n ous opi oi d s : Biol og y and fun cti on . A nnual Revie w of Neurosci en ce, 7, 223-255. Alajo u anin e , T. & Lhermi tte, F . (19 65) . Acq ui re d aph a sia in chil d ren. Brain , 88, 653-662. Albert , D.J. & Chew, G.L. (19 80) . The sept a! for ebrain and the inh ib i to r y modula- tion of atta c k and defe n se in the rat : A revie w . Behavio r al and Neural Bio lo g y, 30, 357-388. Albert , D,J . Dy so n, E.M . & Walsh, M. L. (19 87) . Comp etitive behavi or : Inta c t male rats but not hyp erd efe n siv e males wit h media l hyp o th a lami c lesio n s sh~re wate r with fem ales. P hys iolo gy and Behavio r , 41, 549-553. Albert , D ,J ., & Walsh, M.L . (1984) . Neural sys t e m s and the inh ib i to r y modulati on of ago n i sti c behavi or : A comp ar is o n of m ammalian spe cies . Neuroscie n ce and Bio b ehavio r al Revie ws , 8 , 5-24. Albert , D,J ., Walsh, M.L, Sie m ens, Y. & Lo 떠 e, H. (1986) . Sp on ta n eous mouse kill ing rats : Gentl ing and foo d depr i v a ti on result i n kill ing behavi or almost ide nti ca l of tha t of rats with media l hyp o th a lamic lesio n s. P hys iolo gy and Behavio r , 36, 1197-1199. Altm a n, J. (19 67) . Postn a ta l grow th and differ enti at io n of t he mammali an brain , with im p li ca ti on s for a morp h olog ica l theo ry of memory. In G.C . Qu arto n , T. Melnechuk, & F.O. Schmi tt(e ds.), The neurosci en ces : A stu dy prog ra m (pp. 723-743) , New York : Rockefe ll er Un ive rsity Press. Anand, B.K . & Brobeck, J .R . (1951) . Hy po th a lami c contr ol of f oo d int a k e in rats and cats . Yale Jou nuzl of Bio lo g y and Medic ine , 24, 123-140. Andersson; B. ( 19 53) . The effe c t of inj e c ti on s of hyp erton ic NaCl soluti on s in dif fere nt par ts of the hyp o th a lamus of go ats . Acta Phys iolo g ica Scan- din a vic a, 28, 188- 20 1 . Anti n, J., Gib b s, J., Holt, J., Youn g, R.C ., You ng, R.C ., & Smi th, G.P. (19 75) . Cholecys t o k in in elici ts the comp le te behavi or al sequ e nce of sati ety in Arnoldr, aAts. .P . f&ou nGuozrls kofi, RC o.Am.p ( 1ar9 a84ti)v .e aGnodn aPdhayls isotleo rg o icida l i nPds uy cch tio olon g oy f, 8 s9tr,u 7c 8 t4u -r7a9l 0s.e x differ ences in t he centr al nervous sys t e m . A nnual Revie w of Neuroscie n ce, 7, 413- 44 2. Aserins k y, E. & Kleitm a n, N. (19 53) . Regu la rly o ccurrin g period s of e ye moti lity, and concomi tan t ph enomena durin g sleep. Scien ce, 118, 273-274.
Axelrod, J. & Reis in e , T.D. (19 84) , Str e ss hormones : Their int e r act ion and regu la ti on . Sdence, 224, 452-459. Ay ou b, D.M., Greenoug h, W.T, & Jur aska, J.M . (1983) . Sex dif fer ences in den- driti c s tru ctu r e in the pre op tic area of t he juv enil e m acaqu e monkey brain . Sdence, 219, 197-198. Bail ey , C.H. & Chen, M. (19 83) . Morph o log ica l bases of long -term habit ua ti on and sensiti za ti on in Ap ly s i a . Scie n ce, 220, 91-93. Balow, J.E . & Rosenth a l, A.S. ( 19 73) . Glucocort ico id s upp re ssio n of m acroph a ge mig rat io n inh ib it ing fac to r , jou rnal of Expe rim e nta l Medid n e, 137, 1031-1041. Banks, M.S., Aslin, R.N ., & Lets o n, R.D . (19 75) . Sensiti ve per io d for the develop- ment of human bin o cular vis io n . Scien ce, 190, 675-677. Barchas, J.D . , A kil , H., E ll iott , G.R ., Holman, R.B , & Wats o n, S.J. (19 78) ..” 底 havio r al neuro-chemi str y : Neuroregu la to r s and behavi or al sta t e s . Scienc e, 200, 964-973. Bart us , R.T ., Dean, R.L., Ponte c orvo, M.J. , & Flick er, C. (19 85) . The cholin e rgi c hyp o th e sis : A his to r ic a l overvie w , current per spe etive , and fut u re dire c- tion s. In D.S. Olto n , E. Gamau, and S. Corkin ( eds.) , Memory dys fun ctio n s : An i n t eg ra t加 of an_im al and human ·research from prec lin i c a l and clini c a l pers p e cti ves (pp. 332-358) , New York : New York Academy of Sci en ces. Basser, L.S . (19 62) . Hemi ple g ia of early onset and fac ulty of s pe e ch wit h spe cial refe r ence to the eff ec ts of hemi sp h e recto m y . Brain , 85, 427-460. Bassuk, E.L., & Gerson, S. (19 78) . Dein s ti ut io n ali za ti on and menta l health ser- vic es . Sci en ti fic A meri can , 238, 46-53. Beach, F.A . (19 76) . Sexual att rac t ivity, pro cep tivit y , and recep tivity in fem ale mammals. Horm,o n es and Behavio r , 7, 105-138. Beach, F.A ., Kuehn, T.E., Spr a gu e , R.H ., & Anis k o, J.J. (1972) . Coit al behavi or in dog s- XI : Eff ec ts of androge n i c s tim ulati on durin g develop m ent on mascu- line mati ng respo nses in fem ales. Hormones and Behavio r , 3, 143-168. Beaumont, J.G . (19 83) . Intr o ductio n to neurop syc hology . New York : Gu ilfo rd . Bellows, R.T . (19 39) . Tim e fac to r s in wate r drink in g in dog s. Ameri can jou rnal of Phys iolo gy , 125, 87-97. Benson, D.F., Stu s s, D.T., Naeser, M.A. , Weir , W.S., Kapl a n, E.F., & Levi ne , H. (19 81 ). The long -ter m ·eff e c ts of pre fr on ta l leukoto m y . Archiv e s of Neurology , 338, 165-169.
Berge r , H. ( 19 29) . 區 r das Elektr e nkeph a log ram m des menschen. Archiv / Ur Psych ia trie und Nervenkrankheite n , 81, 527-570. Bic k fo r d, R.G ., Dodg e, H.W. Jr., & Uihle in , A. (19 60) , Electr o g rap h i c and be- havi or al eff ec ts relate d to dep th stim ulati on in human pat ie n ts . In E.R. Ramey & D.S. 6Doherty (eds.) , Electn 'c al stim u lati on in the unanesth e ti zed brain ( pp.2 48-261) , New York : P. B. Roeber. Blakemore, C. & Coope r, G.F. ( 19 70) . Develop m ent of the brain depe n ds on the visu al env iro nment. Natu re , 228, 477-478. Blanchard, D.C. & Blanchard, R.J. (1984) . Af fec t and agg res sio n : An anim al model app li ed to human behavio r . In D.C. Blanchard & R.J. Blanchard (eds.) , Advances in the stud y of aggr es sion (pp. 1-62) , Orlando, FL : Aca- demi c P ress. Blanchard, R.J., Blanchard, D.C., & Takahasi, L.K . (19 77) . Refl ex iv e figh tin g in the albin o rat : Ag gres siv e or defe n siv e behavi or . Ag gre s sive Behavio r , 3, 145-155. Bog en , J.E . & Vog el , P.J. (1962) . Cerebral commi ss uroto m y : A case rep or t . Bulletin of the Los Ang el es Neurolog ica l Soc iety, 27, 169. Bradley, C. (19 37) . The behavio r of chil dr en receiv ing benzedrin e . Ameri can Jou rnal of Psy c hia try, 94, 577-585. Brady, J.V . & Nauta , W.J. H . (19 55) . Subcort ical mechanis m s in emoti on al beha- vior : The durati on of affe c ti ve chang es foll owi ng sep ta! and habenular lesio n s in the albin o rat. Jou rnal of C omp ara ti ve and Phys iolo g ical Psy ch o- log y, 48, 412-420. Brady, J.V ., Pott er , R.W . , C onrad, D.G., & Mason, J.W . (19 58) . Avoid a nce beha- vior and the develop m ent of g as tr od uodenal ulcers. Jou rnal of the Expe ri- menta l Analysi s of Behavio r , I, 69-72. Braio n an, L.E. , Adli n, E.V. & Sta n to n , J.L. Jr. (1985) . Obesity and caloric i nt a k e: The nati on al health and nutr ition al examin a tio n survey of 1971-1975. Jou rnal of Chronic Disea ses, 38, 727-732. Bremer, F. (19 36) . Nouvelles recherches sur le mecanis m e du sommeil. Comp tes Rendus des Seances de la So ci郞 de Bio lo g ie et d e Ses Fil ia l e s, 122, 460-464. Broca, P. (18 61) . Remarqu e s sur le sieg e de la fac ulte du lang ag e art icu le. Bulletin of Soci ety of Anato m y , Paris ( Ser.2) , 6, 330-357. Broderi ck , P.A. & Bri dger , W. H. (1984) . A comp ar ativ e stu d y of t he eff ec t of L- tryptop h a n and its acety la te d deri va ti ve N-acety l- L-try ptop h a n on rat
muric ida l behavio r . Bio lo g ica l Psy c hia try, 19, 89-94. Brody, H. (19 55) . Orga n iz a ti on of the cerebral cort ex Ill : A stu d y of a gi ng in the human cerebral cort ex . jou rnal of Comp ar ativ e Neurology , 102, 511-556. Bruce, C., Desim one, R., & Gross, C.G. (1981) , Vis u al pro p erties of neurons in a pol ys e nsory area in supe r io r tem p o ral sulcus of the macaqu e . Jou rnal of neurop hy s iolo log y, 46, 369-384. Brust, J.C .M . (1981) , Str o ke : Diag n o stic , anato m i cal and ph ys i o lo g ica l consid e ra- tion . In E.R. Kandel and J.H . Schwart z ( eds.) , Pri nc i ple s of neural scie nc e (pp. 667-679) , New York : Elsevie r . Buell, S.J. & Coleman, P.D. 0979) . Dendriti c grow th in the age d human brain a nd failur e of g row th in senil e dementi a. S ci en ce, 206, 854-856. Bunney, W.E., Jr . & Davis , J.M . (19 65) . Norepi ne ph rin e in dep re ssiv e reacti on s. Archiv e s of General Psy c hia try, 13, 483-494. Buss, A.H . & Durkee, A. (1957) . An inv ento r y for assessin g dif fer ent kin d s of hostil ity. Jou rnal of Consultin g Psyc hology , 21, 343-349. Butt er s, N. & Cermak, L.S. ( 19 80) . Alcoholic Korsakoffs synd rome : An inf o rm a- lion pro ces sing app ro ach to amnes ia. New York : Academi c Press. Bekesy, G. von. ( 19 60) . Expe rim e nts in heari ng . New York : McGraw-Hil l. Cabanac, M. 0971) . Phys i o lo g ical role of ple asure. Sci en ce, 173, 1103-1107. Camp be ll, F.W . (19 74) . The tra nsmi ssi o n of spa t ia l infor mati on throu g h the vis u al sys t e m . I n F.O. Schmi tt & F.G. Worden (eds.) , The Neurosci en ces Thir d Stu d y Prog ra m (pp. 95-103) , Cambrid g e , MA : MIT Press. Camp be ll, F.W. & Robson, J.G . (19 68) . Ap plica ti on of Fouri er analys i s to the vis ib il i ty of grat in g s . j ou rnal of Phys iolo g y, 197, 551- 56 6. Cannon, W.B. (1919) . The ph ys i o lo g ica l basis o f thirs t. Proces sing of the Ro ya l Soc iety of London. Seri es B : Bio lo g ica l Scie n ces, 90, 283-301. Cannon, W.B. & Washburn, A.L. (19 12) . An exp la nati on of hung er . Ameri can ]oural of Phys iolo g y, 29, 441-454. Carew, T.J ., Hawkin s , R.D ., & Kandel, E.R. (19 83) . Differe nti al classic a l cond i- tion in g of a defe n siv e wit hd rawal refl ex in A p lys i a Califo rn ic a. Sci en ce, 219, 397-400. Carlto n , P.L. & Manowit z, P. (19 84) . Dop am i ne and schi zo p h reni a : An analys is of the the ory . Neurosci en ce and Bio b ehavio r al Revie ws , 8, 137-151 . Carlton , P.L., Manowit z, P., McBrid e , H., N ora, R., Swart zb urg, M., & Goldste i n , L. (19 87) . At ten ti on defi cit dis o rder and pat h o log ica l gam blin g. Jou rnal of
Clin i c a l Psy c hia try, 48, 487-488. Casey, K.L. (19 82) . Neural mechanis m s in p a in and analge s ia : An overv iew . In A.L. Beckman (ed.) , The neural bas is of behavio r (pp. 273-283) , Jam aic a, NY : Spe c tru m. Chang, H.K. , Chung, B.K ., Park, S.K ., & Lee, M.H. (19 89) . Alte r ed pa tt er n of shock-moti va te d escape in rats wit h pre nata l ly ind uced Mic re nceph aly. Soci ety for Neuroscie n ce (Abstr ac ts ) , 15, 686. Chi, J.G ., Dooli ng , E.C ., & Gil les , F.H. (19 77) . Left -r ig h t asym me tr ies of the tem p or al spe e ch areas of the hwnan fetus . Archiv e s of Neurology , 34, 346-348. Coop er, J,R., Bloom, F.E., & Roth , R.H . (19 86) . The bio c hemi ca l bas is of neurop ha rmacology (5th ed.) . New York : Oxfo rd Univ e rsity Press. Corkin , S. ( 19 84) . Lastin g consequ e nces of bila t e r al media l tem p o ral lobecto m y : Cli nic a l course and exp erim enta l find in g s in H.M .. Semi na rs in Neurology , 4, 249-259. Cowan, W.M. (19 79) . The develop m ent of the brain . Sci en ti fic Amen·can, 241 , 112-133. Cowan, W.M. , F awcett , J.W ., O'Leary , D.D. M ., & Sta n f ield , B.B . (19 84) . Regr es - siv e events in neurog en esis . S 야 ?nee, 225, 1258-1270. Coy le , J.T ., Price , D.L. , & DeLong, M.R . (1983) . Alzheim er's dis e ase : A dis o rder of cort ical cholin e rgi c inn ervati on . Scien ce, 219, 1184-1190. Creese, I. (19 83) . Classic a l and aty pical antip syc hoti c drug s : new ins ig h ts . Trends in Neurosci en ces, 6, 479-481. Crook, T. (19 85) . Clin ica l drug trials in Alzheim er's dise ase. In D.S. Olto n , E. . Gamzu, & S. Corkin ( eds.) , Memory dysf u n cti on s : An int e g r a ti on of anim al and human research fro m pre clin i c a l and clin i c a l per sp e cti ve s (pp. 428-436) , New York : New York Academy of Sci en ces. Crosson, B. (19 84) . Role of the domi na nt tha lamus in langu a ge : A revi ew . Psy c holog ical Bulletin , 96, 491-517. C6t6 , L. (19 81) . Ag ing of t he brain and dementi a. In E.R. Kandel & J.H . Schwar- tz (eds.) , Pri nc i ple s of neural scie n ce (pp. 547-556) , New York : Elsevi er . Damasio , A.R. & Geschw ind , N. (1984) . The neural basis of lang ua ge . A nnual Revie w of Neurosci en ce, 7, 127-147. Damassa, D.A ., Smit h, E.R., Tennent, B., & Dav ids on, J.M . (1977) . The relati on - ship betw e en circ ulatin g tes to s te r one levels and sexual behavi or . Hor-
manes and Behavio r , 8, 275-286. David s on, J.M ., Kwan, M., & Greenleaf, W.J. (1 982). Honnonal repl a cement and sexualit y in men. Clin i c s in Endocri no logy and meta b olism , 11, 599-623. Dax, M. (18 36) . L 函 ons de la moit ieg au che de l'enc 帥 hale coin c i de nt avec l'oubli des sign es de la pen see. Montp ellier , France : montp ellier pre sse. DeBold, J.F . & Mic z ek, K.A. (19 84) . Ag gres sio n per sist s afte r ovarie c to m y in fem ale rats . Hormones and Behavio r , 18, 177-190. Delga d o, J.M . R( 1969) . Phys ic a l contr o l of the min d . New York : Harp er and Row. Delga d o, J.M . R . (1963) . Cerebral hete r ostim ulati on in a monkey colony. Scien ce, 141 , 161-163. Delga d o, J.M . R. , Robert s, W. W., & Mi ller, N.E . ( 1954) . Learnin g moti va te d by electr ic a l stim ulati on of the brain . Ameri can jou rnal of Phys iolo g y, 179, 587-593. Dement, W.C. (19 54) . Some must watc h whil e some must sleep . New York: W.H. Freeman. Dement, W.C. & Kleitm a n, N.(1955). Incid e nce of eye moti lity duri ng sleep in relati on to varyi ng EEG pat t er n . Federatio n Proceedin g, 14, 216. De Valois , R.L. & Jac obs, G.H. 0968) . Prim a te color vis io n . Sci en ce, 162, 533-540. De Valois , R., M orga n , H., & Snodderly, D.( 19 74) . Psyc h op hy s i c a l stu dies of monkey vis io n III : Spa t ia l lumi na nce contr a st s ensiti vit y tes ts of m acaqu e and human observers. Vis ion Research, 14, 75-81. De Valois , R.L. & De Valois , K.K. (19 80) . Sp at ia l visio n . Annual Revie w of Psycl io lo g y, 31, 309-341 . De Valois , R.L. & De Valosis, K.K. (19 75) . Neural cod ing of color. In E.C. Cart er ett e and M.P. Friec lman (eds.) , Handbook of perc ep tion : Seei ng (Vol. 5, pp. 117-166) , Orlando, FL : Academi c Press. Dodd, J. & Jes sell, T.M. (19 88) . Axon guida nce and the pat t er n ing of neuronal pro je c ti on s in vert eb rate s . Scie n ce, 242, 692-699. Doria n , B.J. , Keys t o n , E., Garf ield , P.E., & Brown, G.M. (19 82) . Aberrati on s in lym ph ocy te subp op u l ati on s and fun cti on s duri ng psy c h olog ica l str es s. Clini c a l and Expe rim e nta l Immunology , 50, 132-138. Doty , R.W . (19 51) . Inf lue nce of s tim ulus pat t er ns on refl ex deg lu ti tion . Ameri can jou rnal of Phys iolo gy , 166, 142-158.
Doty , R.W . & Bosma, J.F . ( 1956) . An electr o my og r ap h i c analys i s of refl ex deg lu ti tion . jou rnal of Neurop hy s iolo g y, 19, 44-60. Doty , R.L., Shaman, P., A p ple baum, S.L. , Gib e rson, R., Sik o rski. , & Rosenberg, L. (19 84) . Smell ide nti fica ti on abil ity : Chang es wit h age . Scie n c e, 226, 1441-1443. Elkin , I., Shea, T ., Watk i n s , J .T . , Im ber, S.D ., Sots k y, S.M . , Coll ins , J .F ., G lass, D. R., Pil ko n is, P.A. , Leber, W. R., Dochert y, J.P ., F ie s te r , S.J ., & Parloff , M .B. (1989) . Natio n al Insti tut e of Menta l Health Treatm e nt of Dep re ssio n Collaborati ve Research Prog ram : General eff ec ti ve ness of tre atm ents . Archiv e s of General Psy c hia try, 46, 971-982. Enroth -C uge l l, C. & Robson, J.G . (1966) . The contr a st sensiti vi t y of reti na l gan gl i on cells of the cat. jou rnal of Phys iolo g y, 187, 517-552. Eps t e i n , A.N . (19 71) . The late r al hyp o th a lami c syn d rome : Its im p lic a tio n s for the phy s i o lo g ica l psy c o log y of hung er and thirs t. In E.S t e l lar and J.M . Spr a gu e (eds.) , Progr e ss in phy s iolo g ical psyc hology (Vol. 4, pp. 263- 31 7) , Orlando, FL : Acadmi c Press. Eps t e i n , A.N ., K is si le ff , H.R. , & Ste l lar, E. (1973) . The neurops yc hology of thi rs t . Washi ngton D.C. : V.H. Wi nst o n . Evans, E .F. (1982) . Basic p hy s i c s & psy c h op h ys i cs of s ound. In H.B. Barlow and J.D . Mollen (eds.) , The sen s es (pp. 239-250) , Cambrid g e , Eng la nd : Cam- brid ge Un ive rsity Press. Evarts , E.W. (1968) . Relati on of p yram i da l trac t acti vity to forc e exert ed duri ng volunta r y movement. Jou rnal of Neurop hy s iolo gy , 31, 14-27. Fein , D., Humes, M ., K apl a n, E., Lucci, D., & Wate r house, L. (1984) . The que sti on of left hemi sp h e re dys f un c tio n in infan ti le auti sm . Psy c holog ica l Bulleti n, 95, 258-281. Fein , D., S koff , B., & Mir sk y, A.F . ( 19 81) . Cli nic al correlate s of brain ste m dys f u nc tio n in auti sti c chil dr en. Jou rnal of Autis m and Develop m enta l D 函 rders, 11, 303-315. Fentr es s, J .C . (1983) . Eth o log ica l models of h ie r archy and pa tt er nin g of spe cies -spe c ifi c behav ior . In E. Sati no ff & P. Teit elb aum (eds.) , Handbook of behavio r al neurobio lo g y : Moti va io n (pp. 185-234) , New York : Plenum. Flannelly, K .J., Kemble, E .D., Blanchard, D .C., & Blanchard, R.J. ( 19 86) . Eff ec ts of sep tal- fo r ebrain lesio n s on mate r nal agg res sio n and mate r nal care. Behavio r al and Neural Bio lo g y, 45, 17-30.
Foltz , E.L. & Mi llett , F.E . ( 1 964) . Exp er im enta l psy c h osomati c dis e ase sta t e s in monkeys . I. Pep tic ulcer-executi ve monkeys . J ou rnal of surgic a l Reaserch, 4, 445-453. Foulkes, W .D. (19 66) . The psyc hology of sleep . New York : Scrib n efs . Frederi ck son, RC.A . & Geary, L.E. (19 82) . Endog en ous opi oi d p eptide s : Rewi ew of ph ys i o lo g ica l, pha macholog ical and clin ic a l aspe c ts . P rog re ss in Neuro- bio lo g y, 19, 19-69. Freeman, W. & Watt s, J.W . (1950) . Human psyc hosurge r y (2nd ed.) . Spr ing f ield , IL : Charles C. Thomas. Fried man, M. I. & Str ic k er, E.M. (1976) . The phy s i o lo g ica l psy c h olog y of h ung er : A phy s i o lo g ica l per spe c tiv e . Psy c holog ica l Revie w , 83, 409-431 . Ganong, W.F. (197 7) . The nervous syst em . Los Alto s , CA : Lang e Medic a l Publi ca - tion s. Garfie ld , E. (19 83) . What do we know about the grou p of m enta l diso rders called schiz o p hr enia ? Part 2 : Diag n o sis and tre atm e nt. Current Conte nts, 17, 5-16. Gash, D.M. & Sladek, J.R. Jr. (1988) . Trans pla nta tio n int o the mammalian CNS. Amste r dam : Elsevie r . Gazzanig a, M.S. (19 83a) . Repl y t o Levy and to Zaid e l. Ameri can Psy c holog ist, 38, 547~549. Gazzanig a, M.S. (19 83b) . Rig h t hemi sp h e re langu a ge foll owi ng brain bis e cti on . Ameri can Psy c hologi st, 38, 525-537. Gazzanig a, M.S: & Spe rry, R.W . (1967) . Langu a ge ~fter secti on of the cerebral hemi sp h e res. Brain , 9 0, 131-148. Geff en , G. & Qu in n , K. (1984) . Hemi sp h eric spe c i al iz a ti on s and ear advanta g e s in pro cessin g spe e ch. P sy c holog ical Bulletin , 96, 273-291 . Geschw ind , N. (19 65a) . Di~o nnecxio n syn d romes in anim als and man : Part 1. Brain , 88, 237-294. Geschwin d , N. (1965b) . Dis c onnecxio n syn d romes in anim als and man : Part 2. Brain , 88, 585-644. Geschwi nd , N. (19 75) . The apr a xia s : Neural mechanis m s of dis o rders of l earned movement. Ameri can Sci en tis t, 63, 188-195. Geschw ind , N. & Levit sk y, W. (1968) . Human brain : Left -righ t asym me tr ies in tem p o ral spe e ch reg ion . Sci en ce, 161 , 186-187. Ghez, C. (19 85) . Volunta r y movement. In E.R. Kandel & J.H . Schwart z (eds.) ,
Pri nc ip le s of neural scie n ce(2nd ed., pp. 487-501), New York : Elsevie r . Ghez, C. & Fahn, S. ( 19 85) . The cerebellum. I n E.R . Kandel & J.H . Schwart z (eds.) , Pri nc ip le s of neural scie n ce (2nd ed., pp. 502-522) , New York : Elsevi er . Gil lin, J.C . & Borbely, A.A . ( 1985) . Sleep : A neurobio lo g ica l win d ow on aff ec ti ve dis o rders. Trends in Ne ur o s ci en ces, 8, 537-542. Glaser, R., Ric e , J., Spe ich er, C.E ., Sto u t, J .C ., & Kie c olt-G laser, J.K. (19 86) . Str es s dep re sses int e r f ero n pro ducti on by leukocyt es concomi tan t with a decrease in natu ral kill er cell acti vity. Behavio r al Neurosci en ce, 100, 675-678. Goldfo o t, D .A ., Krevert z, M. A. , Goy , R.W ., & Freeman, S.K . (1976) . Lack of effe c t of vagi na l lavage s and alip h atic aci ds on eja c ulato r y respo n ses in rhesus monkeys : Behavi ro al and chemi ca l analys e s. Honnones and Behav- ior, 7, 1 -28. Goldman, H.H . ( 19 88) . Intr od uc tion to psy c h ia t r ic tre atm ent. In H.H. Goldman (ed.) , Revie w of gen eral psych ia try (2nd ed., pp. 503-505) , Norwalk, CT : Ap ple to n & Lang e. Goldste in , E .B . (1984) , Sensati on and perc ep tion (2d ed.) . Belmont, C A : Wadswor- th. Gorski, R.A ., G ordon, J.H ., Shry ne , J.E ., & South am , A.M . (19 78) . Ev ide nce for a morp h olog ica l sex differe nce wit hin the media l pre op tic area of the rat brain . Brain Research, 148, 333-346. Gouras, P. (1981 ) . Vi su al sys t e m . IV: color visio n . In E.R . Kandel & J.H . Schwar- tz (eds.) , Pri nc i ple s of neural scie n ce (pp. 249-257}, New York : Elsevie r. Goy, D.M . (19 70) . Expe r im e nta l contr o l of psy c h osexualit y. Phil os op hic a l Transactio n s of the Roya l Socie ty of London. Seri es B : Bio lo g ical Scie n ces, 259, 149-162. Green, D.M. (1976) . An int r od uctio n to heari ng. Hil lsd ale, NJ : Lawrence Erl- baum. Greenough , W.T., C art er , C.S., S te e rman, C., & De Voog d, T.J. (1 9 77) . Sex differe nces in d end ritic p at t er ns in h amste r pre op tic area. B rain Research, 126, 63-72. Greis t, J,H . & Jef fe rs on, J.W . (19 88) . Arucie ty dis o rders. I n H.H . Goldman (ed.) , Rev 神 。if gen eral psych ia try (2nd ed., pp.3 49-364) , Norwalk, CT: Ap - ple to n & Lang e.
Gros.c .m an, S.P. (19 67) . A tex tb o ok of phy s iolg ic a l psych olog y. New York : Wi ley . Grossman, S.P ., Dacey, D., H alaris, A .E ., Collie r , T., & Routt en berg, A. (1978) . Ap ha gi a and adip s ia afte r pre fe re nti al destr u cti on of nerve cell bodie s in hyp o th a lamus. Scie n ce, 202, 537- 53 9. Groves, P.M. & Schlesin g e r , K. (1982) , Bio lo g ical psych ology . Dubuq ue , IA : W.C. Brown. Guli ck , W.L. , Gescheid e r, G.A, & . Frisi n a , R.D . ( 19 89) . Heari ng , phy s iolo g ica l acoustic s, neural codin g , and psych oacoustic s. New York : Oxfo r d Univ e rsity Press. Gusella, ].F., W exler, N.S ., C onneally, P .M., Nay lo r, S.L ., A nderson, M.A., Tanzi, R.E ., Watk i n s , P.C., Ott ina , K., Wallace, M.R. , Sakagu c hi, A.Y ., Young, A. B., Shoulson, I., Bonil la. E., & Mart in, J.B . (1983) . A pol ym o rp hic DNA marker gen eti ca lly link ed to Hunti ng t on 's dise ase. N atu r e, 306, 234-238. Hami lto n , C.R. (1977) . An assessment of h emi sp h e ric spe cial iz a ti on in monkeys . In S. Dim o nd and D. Bli zz ~rd (eds.) , Evoluti on and late r aliza tio n of the brain (pp. 222-232) , New York : New York Academy of Sci en ces. Harlow, J.M . (1868) . Recovery from the pas sage of a n iro n bar throu g h the head. Publica ti on s of the massa.ch usetts Medic a l Soc iety (Bosto n ) , 2, 327-346. Harris, G.W. (19 55) . The neural contr o l of the pituit a ry gla nd. Baltim ore : Wi l- Iiam s and Wi lk in s . Harris, G.W . & Jac obsohn, D. (19 52) . Functi on al graf ts of the ante r i or pituitar y gla nd. Proceedin g s of the Roya l Soc iety of London. Sen·es B : Bio lo g ica l Sci en ces, 139, 263-276. Harris, G.W .& Levin e , S. (19 62) . Sexual dif fere nti at i on of the brain and its expe r im enta l contr o l. Jou rnal of Phys iolo g y, 163, 42-44. Hart, B.L. (19 78) . Hormones, spi na l refl ex es, and sexual behavi ou r. In J.B. Hutc h in s on (ed.) , Dete r mi na nts of sexual behavio u r, Ch ich este r , U.K . : Wi le y. Harmann, E. (19 80) . Eff ec ts of p sy c h otr op i c drugs on sleep : The cate c holam ine s and sleep. I n M.A . Lipton , A. DiM ascio & K.F. Kil lm an (eds.) , PsycJ w ph ar- macology : A gen eratio n of prog re ss (pp. 711-728) , New York : Raven Press. Heath , R.G . (19 63) . Electr ica l self -s tim ulati on of the brain in man. A men·can Jou rnal of Psy c hia try, 120, 571-57 7. Heath , R.G . (19 64) . Pleasure respo n se of human subje c ts to dire ct stim ulati on of
the brain : Phys i o lo g ic and psy c h odyn a mi c consid e rati on s. I n R.G . Heath (ed.) , Role of plea sure in behavio r (pp. 219 - 24 3) , New York : Harpe r . Hebb, D.0. (19 49) . The orga n iz a ft'o n of behavio r . New York : Wi ley . Heffn e r, H .E . & Heff ne r, R.S. (1984) . Temp o ral lobe lesio n s and per cep tion of spe cies ~spe c if ic vocali za ti on s by macaqu e s. Sci en c e, 2 26, 75-76. Henke, P.G. (19 80) . The amy gd ala and restr a in t ulcers in rats . Jou rnal of Comp ara tiv e and Phys iolo g ical Psy ch ology , 94, 313-323. Heth e rin g t on , A.W . & Ranson, S.W. (1942) . The spo n ta n eous acti vity and foo d int a k e of r ats wit h hyp o th a lami c lesio n s. A meri ca n Jou rnal of Phys iolo g y, 136, 609-617. Hirsc h, ]. (1984) . Hy po th a lami c contr ol of app e tit e. Ho~ita l Pracft'c e , 19, 131-138. Hirsc h, H.V.B. & Spi ne lli, D.N. (19 71) . Modif ica ti on of the dis tr ib u ti on of field ori en ta t io n in cats by selecti ve vis u al expo s ure durin g develop m ent. Expe rim e nta l Brain Research, 12, 509-527. Hobson, J.A. & McCarley, R.W . (1977) . The brain as a dream sta t e gen erato r : An acti va ti on -syn the sis h yp o th e sis o f the dream pro cess. A meri ca n fou r- nal of Psyc hia try, 134, 1335-1348. Hosobuchi, Y., A dams, J.E , & Linc hit z, R . (1977) . Pain r eli ef b y electr ic a l stim ula- tion of the centr al gray matt er in humans and its reversal by naloxone. Sci en ce, 197, 183-186. Hubel, D .H . & Wi es el, T.N. (19 62) . Recep tive field s, bin o cular int e r acti on , and func tio n al archit ec tu re in the cat' s vis u al cort ex . Jou rnal of Phys z olog y, 160, 106-154. Hubel, D.H. & Wi es el, T.N. (19 63) . Recep tive field s of cells in str iat e corte x of very you ng, visu ally ine xp er ien ced kit ten s. J ou rnal of Neurop hy s iolo gy , 26, 944-1002. Hubel, D.H . & Wi es el, T.N . (19 65) . Recep tive field s and fun cti on al archit ec tu r e in two non-str iat e visu al areas(l 8 and 19) of the cat. Jou rnal of Neuro- phy s iolo g y, 28, 229-289. Hubel, D.H . & Wi es el, T.N. (19 79) . Brain mechanis m s of visio n . Sci en ti fic Ameri can , 241 , 150-162. Hudspe th, A.J. ( 19 83) . The hair c ells of the inn er ear. S ci en ti fic A meri can , 248, 42-52. Hu gh es, J. (1975) . Isolati on of an endog en ous comp ou nd from the brain wit h
pha macologi ca l pro p er tie s sim il ar to morph i n e . Brain Research, 88, 293-308. Hugh e s, J ., S mi th, T., Koste r lit z, H.W ., Foth e rgi ll, L.A., Morga n , B.A ., & Morris, H.R . (19 75) . Identi fica ti on of t wo relate d pen ta p e ptide s from the brain w it h pot e n t opi at e ago n is t a cti vi ty . Natu r e, 258, 577-579. Isacson, 0., Brundin , P., Gag e, F.H., & BjU rklund, A. (1985) . Neural graf ti ng in a rat model of Hunti ng t on 's dis e ase : Prog res siv e chang es aft er neostr iat a l ibo te n ate lesio n s and stri a t a l issu e graf ti ng . Neuroscie nc e, 16, 799-817. Jac obson, M. (19 78) . Develop me nta l neurobio lo g y (2nd ed.) , New York : Holt, Rin e hart and Wi ns to n . Jam es, W. (18 84) . What is an emoti on ? Mi nd , 9, 188-205. Jam es, W. (18 90) . Pri nc i ple s of psyc hology . New York : Holt. Jam es, W. (1894) . The phy s i c a l basis of emoti on . Psy c ho/o ,gica l Revie w , 1, 516-529. Jan a!, M.N., Colt, E.W . D., Clark, W.C., & G!usman, M. (1984) . Pain sensiti vity, mood and pla sma endocrin e levels in man foll owi ng long -dista n ce running : Eff ec ts of naloxone. Pain , 19, 13-25. Joh nson, W.G. & Wi ldm an, H.E. (19 83) . Inf lue nce of exte r nal and covert foo d stim uli on ins uli n secretio n in obese and nonnal pers ons. Behavio r al Neurosci en ce, 97, 1025-1028. Jou vet, M . (19 67) . The sta t e s of sleep. Sci en ti fic A meri can , 216, 62-72. Jou vet, M. (1974) . !'Monoami ne rgi c regu la ti on of the sleep- w akin g cyc l e in the cat. In F.O. Schmi tt and F.G . worden (eds.) , The neurosci en ces : Thir d stu d y pro g ra m (pp. 499-508) , Cambri dge, MA : MIT Press. Kandel, E.R. & Schwartz, J.H . (19 82) . Molecular bio lo g y of learn ing : Modula- tion of tra nsmi tter release. Sci en ce, 218, 433-443. Kanner, L. (19 43) . Auti sti c dis tu rb ances of aff ec ti ve conta c t. Nervous Chil d , 2, 217-250. Kelly, D.D. (19 81) . Diso rders of s leep and conscio u sness. In E.R . Kandel and J.H . Schwart z ( Eds.), Pri nc i ple s of neural scie n ce (pp. 486-499) , New-York : Elsevie r . Kelly, J.P . (19 81) . Audit or y sys t e m . In E.R . Kandel and J.H . Schwart z(e ds.), Pri nc i ple s of neural scie n ce (pp. 258-268). New York: Elsevi er. Kennedinyg, s G .oCf . (1th9e 5 0R) o.y a Tl hSeo hcy iept yo thof a l aLmoni cd ocno.n tSr eorli e osf f Bo o: d Binito al o kg e i cianl rSatcsi e.n cPerso,c 1e3e7d,-
535-548. Kennedy, G.C. (19 66) . Food int a k e energy balance and grow t h. B ri tish Medic a l Bullleti n, 22, 216-220. Kerte s z, A. (19 79) . Recovery and trea tm e nt. In K:M . Heil lm an and E. Valenste i n (ed.) , Clin i c a l neurops ych olog y, New York : Oxfo r d Univ e rsity Press. Khachatu rian , H., L ewi s, M. E., S chafe r, M.K .H ., & Wats o n, S.]. ( 1985) . Anato m y of the CNS opi oi d sys t e m s. Trends in Neurosci en ces, 8, 111- 11 9. Khanna, S.M. & Leonard, D.G .B . (19 82) . Basila r membrane tun in g in the cat cochlea. S cien ce, 215, 305- 30 6. Kie s te r , E. Jr. (1 9 80) . Image s of the nig h t. Sci en ce, 80, 1, 36-43. Kim , C., C hang, H.K ., & Chu, J.W . (1967) . Conseq ue nces of a blati ng mammi llar y bodie s in dog s. j ou rnal of Comp ar ati ve and Phys iolo g ical Psy c hology , 63, 469-476. Kim b le, D.P. (19 88) . Bio lo g ica l Psy c holog y. New York : HBJ . Kim ble, D.P. ( 19 92) . Bio lo g ical Psy c holog y. New York : HBJ . Kin s bourne, M. (19 75) . The onto g e n y of c erebral domi na nce. I n D. Aaronson and R.W . Rieb er (eds.) , Develop m enta l psych oling u is tics and communic a ti on diso rders (pp. 244-250). New York : New York Academy of Sci en ces. Kin s boume, M. & Hisc ock, M. (19 83) . The normal and devia n t develop m ent of func tio n al late r aliza ti on of t he brain . I n M.M. Hait h & ].]. Camp os (eds.) , P.H. Mussen (Series ed.) , Handbook of chil d psyc hology : Vol.][. I nfa n cy and develop m enta l psych obio lo gy (4th ed., pp. 157-280) , New York : Wi ley . Kis s ile ff , H.R. , Pi- S unye r , F.X ., Thornto n , J., & Smi th, G.P. (1981) . C~ te r mi na l octa p e ptide of c holecys t o k in i n decreases foo d int a k e in man. The Ameri - can Jou rnal of Clin i c a l Nutr ition , 34, 154-~60. Kluver, H. & Bucy, P.C. (1939) . Prelim in a ry analys i s o f f un cti on s of t he tem p o ral lobe in monkeys . Archiv e s of Neurology and Psy ch ia try, 42, 979-1000. Kolb, B. & Whish aw, I.Q. (19 85) , Fundamenta ls of human neurops ych ology (2nd ed.) . New York : W.H. Freeman. Kolb, B., & Whis h aw, I.Q. (19 90) . Fundamenta ls of human neurops yc holoy (3rd ed.) . New York : Freeman. Kornhuber, H.H. (19 74) . Cerebral cort ex , cerebellum, and basal gan g lia : An int r od ucti on to the ir moto r func tio n s. In F.O. Schmi tt & F.G. Worden (eds.) , The Neurosci en ces Thir d Stu d y Prog ra m (pp. 267-280) , Cambrid g e , MA. : MIT Press.
Krantz , D.S. & Manuck, S.B. (1984) . Acute psy c h oph y s i o lo g ic reacti vi ty and risk of c ardio v ascular dis e ase : A revie w and me th olo 엉 cal criti qu e. Psy c lwlog i- cal Bulleti n, 96, 435-464. Kuff ler, S.W . (19 53) . Disc harge pat t er ns and fun cti on al orga n iz a ti on of mam- malian reti na . jou rnal of Neuroph y s iolo g y, 16, 37-68. Kuff ler, S.W., Nic h olls, J.G ., & Mart in, A.R. (19 84) . From neuron to brain ( Znd ed.) . Sunderland, MA : Sin a uer. Kuff ler, S.W ., Nic h olls, J,G ., & Matr in , A.R . (19 84) . From neuron to brain . Sunder- land, MA : Sin a uer. Lang e, C.G. (18 85) . Om Sin d sbevaege l ser. Kj < Ji benhavn. Lee, M.H., Haddad, R., & Rabe, A. (1980) . Develop m enta l Imp ai rm ents in the pro g en y of rats consumi ng eth a nol durin g p_re gn a ncy. Neurobehavio r al Toxi co log y, 2, 189-198. Lee, C., Malpe ll , J.G ., Schwark, H.D. , & Wey an d, T.G . (19 84) . Cat med ial inter - lami na r nucleus : reti no to p y , relati on to tapetum and impli ca ti on s for scoto p i c vis io n . jou rnal of Neurop hy s iolo g y, 52, 848-869. Lee, C., Rohrer, W.H., & Sp ar ks, D.L. (19 88) . Pop ul ati on cod ing of saccadic eye movements by neurons in the supe r io r coll icu lus. Natu re , 332, 357-360. Lee, D., Lee, C., & Malpe ll , J. (19 92) . Acuit y-s ensiti vity trad e-off s in the cat late r al gen ic u late comp le x : Role of the med ial int e r lami na r nucleus in scoto p i c vis io n . jou rnal of Neuroph y s iolo y, 68. Le May, M. & Geschwin d , N. (19 75) . Hemi sp h eric differe nces in the brain s of grea t ape s . Brain , Behavio r and Evoluti on , 11, 48-52. Le May, M. & Geschwi nd , N. (19 78) . Asym me tr ie s of the human cerebral hem i- sph eres. In A. Caramazza and E.B .. Zuri f(e ds.), Lang uage acqu i s ition and langu age breakdown (pp. 311-328) , Baltim ore : Joh ns Hop k in s Un ive rsity Press. Le Vay, S., Wi es el, T.N. , & Hubel, D.H. (19 80) . The develop m ent of ocular domi na nce columns in normal and visu ally depr i v e d monkeys . ]. Comp ar a- live Neurology , 191 , 1-51. Levin e , J.D ., Gordon, N.C., & Fie ld s, H.L . (19 79) . Naloxone dose depe n dentl y pro duces analge s ia and hyp er alge s ia in pos t- o pe r ati ve pai n . Natu re , 278, 740-741. Levy -A g res ti, J. & Spe r ry, R.W . (1968) . Differ enti al per cept ua l capa c i ties in majo r and min o r hemi sp h e res. Proceedin g of the Natio n al Academy of
Sci en ces of the Unit ed Sta tes of Ameri ca , 6 1, 1151. Lew y, A.]., Kern, H.E ., Rosenth a l, N.E ., & Wehr, T.A . (19 82) . Brig h t artif icia l ligh t t rea tm e nt of a manic -d epr e ssiv e pat ie n t wit h a seasonal mood cyc l e. Amen·can jou rnal of Psy c hia try, 139 , 1 496-1498. Lic k ey, M.E . & Gordon, B. (19 83) . Drugs for Menta l Illnes s : A Revoluti on in Psy ch ia try. San Franci sc o : W.H. Freeman. Lic k ey, M.E. & Gordon, B. (19 91) . Medic ine and menta l illn ess. New York : W.H. Freeman. Lind sley, D.B. , Schrein e r, L.H . , Knowles, M.S., & Mag ou n, H.W . 0950) . Be- havi or al and EEG chang es foll owi ng chronic brain s te m lesio n s in the cat. Electr oe ncep h alog ra ph y and Cli nic a l Neuroph y s iolo gy , 2, 4 83-498. Lorenz, D.N. & Goldman, S.A. (19 82) . Vaga l media t i on of the cholecys t o k in i n sati et y eff ec t in rats . Phys iolo gy and Behavio r , 29, 599-604. Lyn c h, G. & Baudry , M. (1984) . The bio c hemi st ry of memory : A new and spe cific hyp o th e sis . Sci en ce, 224, 1057-1063. Maddis o n, S., Wood, R.J., Rolls, E.T., Rolls, B.J., & Gib b s, J. (1 980) . Drin k in g in the rhesus monkey : per ip h eral fac to r s. jou rnal of Comp ar ativ e and Phys iolo g ical Psyc lwlogy , 94, 365-374. Maff ei, L. & Fio r enti ni, A . (19 73) . The vis u al cort ex as a spa t ia l freq u e ncy analys e r. Vis ion Research, 13, 1255- 12 67. Maie r, S.F ., Shennan, J.E ., L ewi s, J.W . , T ennan, G.W ., & Lieb eskin d , J.C . (1983) . The opi oi d / nonop ioi d natu r e of stre ss-i n d uced analge s ia and learned helpl e ssness. jou rnal of Expe rim e nta l Psy c hology : Anim al Behavio r Processes, 9 , 80-90. Malsbury , C.W . , K aw, L.M., & Pfa ff, D.W . (19 77) . Eff ec t s o f m edia l hyp o th a lami c lesio n s on the lordosis respo n se and oth e r behavio r s in fem ale gol den hamste r s. Phys w logy and Behavio r , 1 9, 223-237. Marqu e s, D.M., M alsbury , C.W . , & Daood, J. (1 9 79) . Hy po th a lami c knif e cuts dis so ci at e mate r nal behavi or s, sexual recep tivit y , and estr ou s cyc l ic ity in fem ale hamste r s. Phys iolo gy and Behavio r , 23, 347-355. Marshall, G. & Zim bardo, P.G. (19 79) . Aff ec ti ve consequ e nces of ina dequ a te l y exp la in e d phy s i o lo g ica l arousal. Jou ~/ ofP ersonality and Soci al Psy ch o- logy , 37, 970-988. Mawson, A.R. & Jac obs, K.W . (19 78) . Corn, tryptop h a n, and homi cide . J ou rnal of Ortlw molecular Psych ia try, 7, 227-230.
May er , J. (1 9 53) . Glucosta t ic mechanis m s of regu la ti on of foo d inta k e. New Engl a nd Jou rnal of Medic ine, 249, 13-16. May er , J . 0 955) . Regu la ti on of e nergy , int a k e and the body weig h t : The glu cos- tat ic the ory and the lipo sta t ic hyp o th e sis . Annals of the New York Academy of Sci en ces, 63, 15-43. Maye r , D.]., Wolf e, T.L. , A ki! , H., Carder, B., & Lieb eskin d , J.C . (19 71) . Analge - sia from electr ic al stim ulati on in the brain s te m of the rat. Scie nc e, 174, 1351-1354. McCormic k , D.A . & Thomp so n, R.F. (19 84) . Cerebellum : Essenti al inv olvement in t he classic a lly condit ion ed eye l id respo n se. Scie n ce, 223, 296-209. McGaug h, J.L . & Hert z, M.J. (1 9 72) . Memory consolida tio n . San Francis c o : Albio n . McWi lliam s, J.R. & Lyn c h, G. (19 83) . Rate of s yn a pt ic repl a cement in d enervate cl rat hip po camp us declin e s pre c ipitou sly from the juv enil e period to adult- hood. S ci en ce, 221 , 572-574. Merzeni ch , M. M. & Harrin g t on , T. (19 69) . The sense of f lut t er -VJ 'br ati on evoked by stim ulati on of t he hair y skin o f p ri m ate s : Comp ar i so n of h uman sensory capa c it y wit h the respo n ses of m echanorecept ive aff ere nts inn ervatin g the hair y skin of monkeys . Expe rim e nta l Brain Research, 9, 2 36-260. Merzenic h , M .M. & Kaas, J.H . (19 80) . Pri nc i ple s of o rga n i za ti on of s ensory perc e- ptua l sys te m s in mammals. In J.M . Spr a gu e and A.N . Ep st e i n ( ecls.) , Prog re ss in psych obio lo gy and p hy sio lo .언 .cal psych olog y (Vol. 9, pp. 1-42) , Orlando, FL : Academ ic Press. Merzenic h , M.M., Nelson, R.J., S tr yk er, M .P., Cyn a der, M. S., S chopp m ann, A., & Zook, J.M . (1984) . Somato s ensory' cort ica l map change s foll ow ing digit amp u ta t io n in adult monkeys . Jou rnal of Comp ar ativ e Neurology , 224, 591- 60 5. Mic h ael, R.P . & Keveme, E.B. (19 70) . Pri m ate sex ph eromones of vagi na l ori - gin. Natu re , 225, 84-85. Mi lne r, B . ( 19 66) . Amnesia foll owi ng ope r ati on on the tem p ora l lobes. I n C.W. M. Whit ty and 0.L . Zang will ( eds.) , Amnes ia (pp. 109-1 3 3) , London : Butt ers wort hs. Mi lne r, B. (19 67) . Brain mechanis m s sug ge ste d by stu dies of tem p ora l lobes. In F.L . Darley (ed.) , Brain mechanis ms underlyin g spe ech and langu age (pp. 122-145) , New York : Grune & Str at t on .
Mi ln er, B. (19 70) . Memory and media l tem p o ral regi on s of the brain . In K.H . Pri br am and D.E. Broadbent ( eds.) , B io lo gy of memory (pp. 29-50) , Orlando, FL : Academi c Press. Mi lne r, B( 19 82) . Some cog nitive effe c ts of fron ta l -lobe lesio n s in man. Phil os op hic a l Transactio n s of the Roya l Soc iety of London. Seri es B : Bio lo g ica l Sci en ces, 298, 211- 22 6. Mil ler , N.E.(1 9 85) . The value of behavio r al research on anim als. Ameri ca n Psyc holog ist, 40, 423-440. Mi ln er, B. & Petr ide s, M. (19 84) . Behavi ou ral eff ec ts of fron ta l -lobe lesio n s in man. T rends in Neuroscie n ces, 7, 403-407. Mi tch ell, G.D. (19 68) . At tac hment dif fer ences in male and fem ale infan t mon- keys . C hil d Develop m ent, 3 9, 611-620. Money, J. & Ehrhardt, A .A. (1972) . Man and woman, boy and girl. B altim ore : Joh ns Hop k in s Press. Money, ]., Ehrhardt, A.A., & Masic a , D.N. (1968) . Feta l fem i ni z a ti on ind uced by androg en ins ensit ivity in the tes tic u lar fem in i z i n g syn d rome : Eff ec t on ma rriag e and mate m alism . The Joh ns Hopk i n s Medic a l Jou rnal, 123, 105-114. Monti , P.M., Brown, W. A., & Corriv e au, D.P. (19 77) . Testo s te r one and comp o- nents of agg res siv e and sexual behavio r in man. Ameri can Jou rnal of Psyc hia try, 134, 692-694. Moore, R.Y . (19 82) . The sup ra ch ias mati c nucleus and the orga n iz a ti on of a circ ardia n sys t e m . Trends in neuroscie n ces, 5, 404-407. Moruzzi, G. & Mago u n, H.W. (1949) . Brain ste m retic u lar form ati on and acti va - tion of the EEG. Electr o encep h alog ra ph y and Clin i c a l Neurop hy s iolo g y, 1, 455-473. Mountc a stle , V.B . (1957) . Modalit y and top o g r ap h ic pro p erties of sin g l e neurons of c at's somatic sensory cort ex . Jou rnal of Neurop hy s io/o gy, 20, 408-434. Mountc a stle , V.B. (19 79) . An orga n iz in g pri n c i ple for cerebral fun cti on : The unit module and the dis tr ib u te d sys t e m . In F.O. Schmi tt and F.G. Worden (eds.) , The neurosdences fou rt h stu d y prog ra m (pp. 21-42) , Cambrid g e , MA: MIT Press. Mozell, M.M. (19 87) . Ol fac ti on . In G. Adelman (ed.), Ency c lop ed ia of neurosden- ce (Vol. 1, pp. 874~877) , Bosto n : Bir k h user. My er s; J.K., Weis s man, M., Tis c hler, G.L ., Holzer, C.E., Leaf, P.J ., Orvaschel, H.,
Anth o ny, J.C ., Boy d, J.H ., B urke, J.D ., Kramer, M ., & Sto l tz m an, R (1984) . Six - month pre valence of psy c h ia t r ic dis o rders in thre e communit ies . Archiv e s of General Psy c hia try, 41, 959-967. Nii jima , A. (19 69) . Af fer ent imp u lse dis c harge s from glu corecept or s in the live r of the guine a pig. Annals of the New York Academy of Scienc es, 157, 690-700. Norr is, D.0. ( 1980) . Vert eb rate endocri no logy . Phil ad elph i a : Lea and feb ig e r. Nott eb ohm, F. (19 77) . Asym m etr ie s in neural contr o l of vocali za ti on in the canary . In S. H amad, R .W . Doty , L. Goldste i n , J. Jay n e s, & G. Krauth a mer (eds.) , Late r liza tio n in the nervous syst em (pp. 23-44) , Orlando, FL : Aca- demi c Press. Novin , D. (19 76) . Vi sc eral mechanis m s in the contr ol of f oo d inta k e. I n D. Nov in, W. Wy rwick a, & G, A. Bray (eds.) , Hun ger , bas ic mechanis ms and clini c al imp lica ti on s (pp. 357-367) , New York : Raven Press. Novin , D. ( 19 79) . Some expe c te d and unexp ec te d eff ec ts of glu cose on foo d int a k e. In G.A . Bray (eds.) , Recent advences in obe sity research I (pp. 27-32) , Westp o rt , C T : Food and Nutr ition Press. Olds, J. & Mi lne r, P. (19 54) . Positi ve rein for cement pro duced by electr ica l stim ulati on of t he sept al area and oth e r reg ion s of t he rat b rai n. Jou rnal of comp ar ati ve and Phys iolo g ical Psy ch ology , 47, 419-428. Oomura, Y., Ohta , M., Ishi ba sh i, S., K ita, H., Okajim a , T., & Ono, T. ( 19 79) . Activ ity of chemo-sensiti ve neurons relate d to the neurop hy s iol og ica l mechanis m s of fee di ng . In G.A . Bray (ed.) , Recent advances in obe sity research Il (pp. 17-26) , Westp o rt, CT : Food and Nutr ition Press. Parkes, C.M. & Brown, R.J. (19 72) . Health aft er bereavement : A contr ol led stu d y of you ng Bosto n wido ws and wid o ws. Psych osomati c Medic ine , 34, 449-461. Parkin , A .]. (19 84) . Amnesic syn d rome : A lesio n -spe cific diso rder? Cort ex, 20, 479-508. Patt er son, P.H., Pott er , D.D., & Furshp an , E.J . (19 78) . The chem ica l differ enti a- tion of nerve cells. S cien ti fic A meri can , 239, 38- 47 . Peck, J.W . & Nov in, D. ( 19 71) . Ev ide nce tha t osmorecept or s me diat in g drink in g in rabbits are in the late r al pre op tic area. Jou rnal of comp ar ati ve and Phys w logi cal Psych ology , 74, 134-147. Penf ield , W. & Rasmussen, T. (19 50) . The cerebral cort ex of man. N ew York :
Macmi llan . Penot, C., V ergn e s, M., Mack, G., & Kemp f, E. (1978) . lnte r spe cific agg res sio n and reacti vity in the rat : Comp ar ed effe c ts o f e lectr o lyt ic r aph lesio n s and int r av entr ic u lar 5,7 DHT admi nis tr a ti on . B io lo g y of Behavio r , 3, 71-85. Perrett, D.I ., Smi th, P.A.J ., Pott er , D.D., Mi stl i n, A.J., H ead, A.S. , M i lne r, AD., & Jee ves, M.A. (19 85) . Vi su al cells in the tem p o ral corte x sensiti ve to fac e view and gaz e dire cti on . P roceedin gs of the Roya l soc iety of London. Seri es B : Bio lo g ical Scie n ces, 223, 293-317. Pert , C.B . & Sny de r, S.H. (19 73) . Op iat e recept or : Demonsrati on in nervous tissu e. Scien ce, 179, 1011-1014. Pete r sen, M.R. , Beecher, M.D ., Zoloth , S.R. , Moody, D.B. , & Ste b bin s , W. C. (19 78) . Neural late r aliz a ti on of s pe cies -s p e c i fic vocali za ti on s by Jap a n ese maca- que s. S ci en ce, 202, 324-326. Pfa ff, D., D ia k ow, C., Z ig mo nd, R.E ., & Kow, L.M. (19 74) . Neural and hormonal dete r mi na nts of fem ale matin g behavi or in rats . In F.O. Schmi tt and F.G. Worden(eds.) , The neuroscie n ces thi r d stu dy pro g ra m (pp. 621-646) , Cam- bridg e, MA : MIT Press. Pfa ff, D.W . & Zig mo nd, R .E . (1971) . Neonata l androge n eff ec ts on sexual and non- sexual behavi or of adult rats tes te d under vario u s hormone regi m es. Neuroendocri no logy , 7, 129-145. Pfa ffrna nn, C. (1959) . The aff ere nt code for sensory qua lit y. Ameri ca n Psych olo- gist, 14, 226-232. Pfe i f fer, C.A . (19 36) . Sexual differ ences in the hyp o ph y s e s and the ir d ete n nuna- tion by the gon ads. A meric an Jou rnal of Anato m y , 58, 195- 22 5. Phoeni x, C.H. (19 76) . Sexual behavi or of c astr at e d male rhesus monkeys tre ate d with 19-hyd r oxy tes to s te ro n. Phys iolo gy and behavio r , 16, 305-310. Phoeni x, C.W., Goy , R .W ., Gerall, A.A ., & Young, W.C. (19 59) . Orga n iz i n g acti on of p re nata ll y adm ini s te r ed tes to s te r one pro p ion ate on the tiss ue med iat i ng mati ng behavi or in the fem ale guine a pig. Endocri no logy , 65, 369- 38 2. Phoeni x, C.H. , J en sen, J.N ., & Chambers, K .C. (1983) . Female sexual behavi or disp l a y ed by androg en iz e d fem ale rhesus macaqu e s. Hormones and Beha- vio r, 17, 146-151. Pi- S unye r , X., Kiss ile ff , H.R ., Thornto n , J., & Smi th, G.P. (1982) . C- te n nin a l octa peptide of c holecys t o k in in decreases foo d int a k e in o bese men. Phys iol- ogy and Behavio r , 29, 627-630.
Pomerantz , B.(1987). Acup un ctu r e neurop hy s i o lo g y. In G. Adelman(ed.), Ency cl op ed ia of Neurosci en c e: V ol. 1 (pp. 6-7 ) , Bosto n : Bir k h user. Pomeranz, B.H . 0 989) . Transcuta n eous electr ic a l nerve stim ulati on (TENS) . I n G. Adelman (ed) . Neuroscie n ce yea r (pp. 161-164) , Bosto n : Bir k hauser. Powell, T.P.S . & Mountc a astle , V.B . (19 59) . Some aspe c ts of the fun cti on al orga n iz a ti on of the cort ex of the pos tc e ntr a l gyros of the monkey : A correlatio n of find in g s obta i n e d in a sin g l e unit analys i s wit h cyt oa r- chi tec tu r e. Bulletin of the Joh ns Hop kin s Hosp ital, 105, 133-162. Powley, T.L. ( 1977) . The ventr o media l hyp o th a lamic syn d rome, sati ety , and a ceph a lic pha se hyp o th e sis . Psyc holog ica l Revie w, 84, 89-126. Pric e , D.L. , Cork, L.C., Str u ble, R.G ., Whit eh ouse, P.J ., Kit t, C.A ., & Walker, L.C. (1985) . The fun cti on al orga n iz a ti on of the basal fore brain cholin e rgi c sys t e m in p ri m a te s and the role of t his s ys t e m in Alzheim efs dise ase. In D. S. Olto n , E. Gamzu, and S. Corkin ( eds.) , Memory dysf u n ctio n : An int e gra - lion of anim a l and clin ic a l pers p e cti ves (pp. 287-295) , New York : New York Academy of Scie n ces. Rada, R.T ., L aws, D.R ., & Kellner, R. (19 76) . Plasma tes to s te r one levels in the rapi st. Psy c hosomati c M edic ine, 38, 257-268. Raic h le, M.E . ( 19 83) . Positr o n emi ss io n tom og rap h y . Annual Revie w ofN euro- scie n ce, 6, 249-267. Rais m an, G. & Fie ld , P.M . (1971 ) . Sexual dim orp hism in the pre op tic a rea of t he rat. Sci en ce, 173, 731-733. Rais m an, G. & Fie ld , P.M. (1973a) . A qua nti tat iv e inv estig a tio n of the develop- • ment of collate r al rein n ervati on aft er par t ial deaff er enta t io n of the sept al nuclei. Brain Research, 50, 241-264. Rais m an, G. & Fie ld , P.M (1973b) . Sexual dim o rp hism in the neurop il of the pre op tic area of the rat and its depe n dence on neonata l androge n . Brain Research, 54, 1-2 9 . Rakic , P. ( 1985) . Lim it s of n eurog en esis i n prim ate s . Scien ce, 227, 1054-1056. Rakic , P. & Sid m an, R.L. (19 73) . Sequ e nce of d evelop m enta l abnormali ties lead- ing to gran ule cell defi cit in cerebellar cort ex of weaver muta nt mice . Jou rnal of Comp ar ativ e Neurology , 152, 103-132. Ramm, P. (19 79) . The locus coeruleus, cate c holam ine s, & REM sleep : A cri tica l revi ew . B ehavio r al and Neural Bio lo g y, 25, 415-448. Rap op o r t, J.L . (1989) . The bio lo g y of obsessio n s and comp u lsio n s. Scientific
Ameri can , 260, 82-89. Rasmussen, T. & Mi lne r, B. ( 1975) . Clin i c a l and surgi ca l stu d ie s of the cerebral spe e ch areas in man. In K.J. Zulch, 0. Creutz feld t, & G.C. Galbraith ( eds.), Cerebral localiza tio n (pp. 238-257) , Berlin : Spr i ng e r -Verlag. Reyn o lds, D.V. (1969) . Surge r y in the rat duri ng electr ic a l analge s ia ind uced by foc al brain stim ualtio n . Sci en ce, 164, 444-445. Rich ardson, D .E. & Akil , H. (19 77) . Pain r educti on by electr ica l brain s tim u lati on in man : Part 2. Chronic self- a d mi nis tr a t io n in the per iv e ntr ic u lar gray matt er . Jou rnal of Neurosurge r y , 47, 184-194. Rich te r , C .P. (1965) . Bio lo g ica l clocks in medic ine and psych ia try. Spr i n g f ield , IL : Charles C. Thomas. Rich te r , C.P . (19 67) . Sleep and acti vity : Their relati on to the 24-hour clock. Research Publica t io n -Assoc iation for Research in nervous and menta l Disea se, 45, 8-27. Rodin , J. (1976) . The relati on ship betw een exte r nal respo n siv e ness and the developm ent and main t e n ance of o besity . In D. Novi n, W. Wy rwick a, and G.A . Bray (eds.) , Hun ger , bas ic mechanis ms and clini c a l imp lic a ti on s (pp. 409-419) , New York : Raven Press. Rod in, J. (1978) . Has the dis ti n c ti on betw een int e r nal versus exte r nal contr o l of fee din g outl ive d its usefu ln ess ? In G. Bray (ed.) , Recent advances in obesity research Il (pp. 75-85) , Westp o rt, CT : Food and Nutr it i on Press. Rog el , M .J . 0978) . A criti ca l evaluti on of t he pos sib il i ty of h igh er pri m ate rep ro - ducti ve and sexual ph eromones. Psy c lwlog ical Bulleti n, 85, 810-830. Rolls, B.J . & Rolls, E.T . ( 1982) . Thir st . Cambrid ge , E ng la nd : Cambrid g e Univ e r- sity Press. Rolls, E.T . (19 84) . Neurons in the cort ex of the tem p o ral lobe and in the amy g- dala of the monkey with respo n ses selecti ve for fac es. Human Neu ro - bio lo g y, 3, 209-222. Rose, R.M . (19 78) , Neuroendocri ne correlate s of s exual and agg res siv e behavi or in humans. In M.A . Lip ton , A. DiM ascio , and K.F. Kil lam (Eds.) . Psy cl w- pha macology : A gen eratio n of prog re ss (pp. 541-552) , New York : Raven pre ss. Rosenblum, L.A . ( 1961) . The develop m ent of socia l behavi or in the rhesus monkey. Unp ub li sh ed docto r al dis se rt ation , Un ive rsity of Wi sc onsin , Madis o n.
Rosenzweig , M.R. & Leim an, A.L. (19 82) . Phys iolo g ical psyc hology . Lexin g t on , MA : D.C. Heath . Routt en berg, A. & Lind y, J. (19 65) . Eff ec ts of t he avail ab il ity of r ewardin g sept a! and hyp o th a lami c s tim ulati on on bar-pr e ssin g for foo d under condit ion s of dep ri va ti on . Jou rnal of Comp ar ati ve and Phys iolo g ica l Psy c hology , 60, 158-161. Rubin , R .T . ( 19 82) . Testo s te r one and agg res sio n in men. In J.V . Beawnont and G.D . Burrows(eds.) , Handbook of psych ia try and endocri no logy (pp. 353-366) , New York : Elsevi er Bio m edic al Press. Rusak, B. & Zucker, l. (19 79) . Neural regu lat i on of circ adia n rhyt he ms. Phys iolo g ical Revie w s, 59, 449-526. Russek, M. (19 71) . Hepa t ic recep tor s and the neuroph y s i o l og ica l mechanis m s contr o ll ing fee din g behavi or . In S. Ehrenp re is ( ed.) , Neuroscie nc es Research (Vol. 4, pp. 213-282) , Orlando, FL : Academi c Press. Russel, W.R. & Nath a n, P.W. (19 46) . Trawnatic amnesia . B rain , 69, 280-300. Samorajs k i, T . (19 76) . How the hwnan brain respo n ds to agi ng . jou rnal of Ameri can Geri atrics Soc iety, 24, 4-11. Schachte r , S. ( 19 71) . Some extr a ordin a ry fac ts about obese hwnans and rats . Ameri can Psy c holog ist, 2 6, 129-144. Schachte r , S.& Sin g e r , J.E . (19 62) . Cog nitive , socia l, and ph ys i o lo g ica l dete r m i- nants of emoti on al sta t e . Psy c holog ical Revie w , 69, 379-399. Scheib e l, M.E., Lind say, R.D ., Tomi yas i.l, U., & Scheib e l, A.B. (19 75) . Prog res siv e dendriti c chang es in agi ng hwnan cort ex . Expe rim a nta l Neurology , 47, 392-403. Scheib e l, M.E ., Tomi yas u, U., & Scheib e l, A.B. (19 77) . The agi ng hwnan Betz cell. E xpe rim e nta l Neurology , 56, 598-609. Scherschli ch t, R ., Pole, P., Schneeberge r , J., Ste i n e r, M., & Haefe ly , W. (19 82) . Selectiv e supp re ssio n of rapi d eye movement sleep (R EMS) in cats by typica l and aty pical anti de pr e ssants . In E. Costa and G. Racag ni (eds.), Typ ica l and aty pica l anti de p r essants : Molecular mechanis ms (pp. 359-364) , New York : Raven Press. Schil dk raut, ].]. (1 9 65) . The cate c holami ne hyp o th e sis o f aff ec ti ve diso rders : A revie w of supp o rt ing evi de nce. Ameri can Jou rnal of Psych ia try, 122, 509-522. Schnap f, J .L. & Bayl o r, D.A . (19 87) . How pho to r ecep tor cells respo n d to light.
Scien ti fic A meri ca n, 256, 40-47. Schwartz , S.R . & Afr ica , B. ( 19 88) . Schi zo p hr enic d iso rders. In H.H. Goldman (ed.) , Revie w of gen eral Psy c hia try (2nd ed., pp. 299-315) , Norwalk, CT : Ap ple to n & Lang e. Searleman, A. ( 19 77) . A revi ew of righ t hemi sp h e re linguisti c capa b il ities . Psyc hologi cal bulleti n, 84, 503-528. Seid m an, L.J. (19 83) . Schiz o p hr enia and brain dys f u n cti on : An int e g r at i on of recent neurod iag n o stic find in gs. Psy ch olog ica l Bulletin , 94, 195- 23 8. Serge n t, J . (1982) . Theoretic a l and meth o dolog ica l consequ e nces of v aria t io n s in expo su re durati on in v isu al late r alit y stu d ie s . Percep tion and Psy ch op hy s ics, 31, 451-461. Sheph e rd, G.M. (19 79) . The synaptic orga n iz a tio n of the brain ( 2d ed.) , New York : Oxfo rd Univ e rsity Press. Shapi ro , D .H. (19 85) . Medit at io n and behavio r al medic ine : Ap plica ti on of a self- regu lt ai on str at e g y to the clin ic a l manage m ent of s tr es s. I n S.R. Burchf ield (ed.) , Str ess : Psy c holog ical and Phys ilol og ica l Inte r actio n , Was hi n 안 on Hem isp h ere Pub. Co. Sheph e rd, G.M.S (l98 3) . Neurobio lo gy . New York : Oxp or d Univ e rsity pre ss. Shi ple y, J.E . & Kolb, B. (19 77 ) . Neural correlate s of s pe cies -ty pica l behavio r in the Syr ian gol den hamste r . Jou rnal of Comp ar ati ve and Phys iolo g ic a l Psyc howg y, 91, 1056-1073. Sie v er, L.J. & Dav is, K.L. (19 85) . Overvie w : Toward a dys r egu la ti on hyp o th e sis of depr e ssio n . Ameri can jou rnal of Psy ch ia try, 142, 1017-1031 . Sklar, L.S . & Anism an, H. (19 81 ) . Str es s and cancer. P sy c holog ica l Bulleti n, 89, 369-406. Sm ith, G. ( 19 88) . Anim almodels of Alzheim er's dis e ase : Expe r im enta l choli ne r- gic d enervati on . Brain Research Revie ws , 13, 103-ll8. Smi th, G.P ., Jer ome, C., Cushi n, B.J ., Eter no, R., & Sim ansky, K.J. ( 19 81) . Abdomi - nal vago t o m y blocks the sati ety effe c t of cholecys t o k in i n in the rat. Scien ce, 213, 1036-1037. Sokoloff, L. (19 77 ) . Relati on betw e en phy s i o lo g ica l fun cti on and energy meta b o- !ism in the centr al nervous sys t e m . J ou rnal of Neurochemi stry, 27, 13-26. Spa r ks, D.L ., L ee, C., & Rohrer, W . H. (19 90) . Pop u lati on codin g of t he dire cti on , amp litud e and velocit y of s accadi c eye movements by neurons in the supe - rior coll icu lus. Cold Spring Harbor Sym po sia on Quan tit ive Bio lo g y, 55,
805-8ll. Spa r ks, R., Helm, N., & Albert , M. (19 74) . Ap h asia rehabil itation resultin g from melodic int o n ati on the rapy . Cort ex, 10, 303-316. Spe r ry, R.W . (1963) . Chemoaff ini ty in the orderly grow th of n erve fibe r pat t e1 :1 5 and connecti on s. Proceedin g s of the Natio n al Academy of Scienc es of the Unit ed Sta tes of Americ a, 50, 703~710. Spe rry, R.W ( 19 68) . Hemi sp h eric deconnecti on and unity in conscio u s aware• ness. Americ an Psy c holog ist, 23, 723-733. Sp ring e r , S.P., & Deuts ch , G. (19 81) . Left brain , righ t brain , New York : W.H. Freeman. Squ i re , L.R. (19 82) , The neurop sych ology of h uman memory . Annual Revi ew of Neuroscie n ce, 5, 2 41-273. Squ ire , L.R., Slate r , P.C., & Chace, P.M . (19 76) , Retr og r ad e amnesia foll ow ing electr o- convulsiv e the rapy . Natu r e, 260, 775-777. Ste p h a n, H. (19 72) . Evoluti on of pri m ate brain s : A comp a rati ve anato m i ca l inv estig a ti on . In T. Tutt le (ed.), Thefu n cti on al and evolutio n ary bio lo g y of prima te s (pp. 155-174) , Chic a go : Aldi ne -At he rt on . Sto n e, J. & Hoff ma nn, K-P. (19 72) . Very slow-conducti ng ganglion cells in the cats reti na : A majo r new func tio n al type ? Brain Reseacrh, 43, 610-616. Str ick er, E.M. & Zig mo nd, M. J. (19 76a) . j'Br ain cate c holamin e s and the late r al hyp o th a lami c syn d rome. In D. Novin , W. Wy rwick a, & G.A . Bray (e ds.), Hunge r : Ba sic mechanis ms and clini c al imp lic a tio n s (pp. 19-32) , New York : Gaven Press. Str ic k er, E.M . & Zig mo nd, M. J. (1976b) . Recovery func tio n foll ow ing damage to centr al cate c holami ne -conta i n i n g neurons : A neurochemic al model for the late r al hyp o th a lami c syn d rome. In J.M . Sp ra gu e and A.N . Ep stein (e ds.) , Progr e ss in psych obio lo gy and phy s iolo g ical psych ology (Vol. 6, pp. 121-188) , Orlando, FL : Academi c Press. Str yk er, M. P. & Sherk, H. (1975) . Mod ifica ti on of c ortic a l ori en ta tion selecti vity in t he cat by restr ict e d visu al exp erien ce : A re-exami na ti on . Scienc e, 190, 904-906. Stu ss , D.T. & Benson, D.F. (19 84) . Neurops yc holog ica l stu idies of the fron ta l lobes. Psy c holog ical Bulletin , 95, 3-28. Svare, B. (19 77) . Psyc h olog ica l dete r min an ts of mate r nal agg res siv e behavi or . In E.C. Sim mel, M. E. Hahn, & J.K. Walte r s(eds.), Aggresive Behavi or :
Genetic and neural app r oaches(pp . 129-146), Hil lsd ale,NJ : Lawrence Erl- baum Associa t e s . Suddath , R.L., Ch risti s o n, G.W ., Torrey, E.F., Casanova, M.F., & Wein b erge r , D. R. (19 90) . Anato m i ca l abnormalit ies in the brain s of monozyg o tic tw in s dis c ordant for schiz o p hr enia . New En gla nd jou rnal of Medic ine , 322, 789- 79 4. Suth e rland, E.W. (19 72) . Stu d ie s on the mechanis m of h ormone acti on . Scien ce, 177, 401-408. Swaab, D.F. & Flier s , E. (19 85) , A sexually dim o rp hic nucleus in the human brain . Scien ce, 228, 1112-1115. Takahashi , J.S. & Zatz , M. (1982) . Regu la ti on of c irc adia n rhyt hmicit y . Sci en ce, 217, 1104-1111 . Teite lb aum, P. & Ep st e in , A.N . (1962) . The late r al hyp o th a lami c syn d rome : Recovery of fee din g and drink in g afte r late r al hyp o th a lami c lesio n s. Psych olog ica l Revie w, 69, 74-90. Teuber, H.L. , Mi lne r, B., & Vaug hn , H.G . (19 68) . Persist e n t ante r og rad e amnesia aft er sta b wound of the basal brain . N europs ych o/ogi a, 6, 267-282. Thomp so n, C.l. (19 80) . Contr o ls of eati ng . New York : SP Medic a l and Sci en ti fic Books. Thomp so n, R.F ., Berge r , T.W ., & Madden, J. (1 9 83) . Celluar pro cesses of learn- ing and memory in the mammali an CNS. Annual Revie w of Neurosdence, 6, 447-491 . Thomp so n, R.F. , Clark, G.A ., Doneg an , N.H., Lavond, D.G. , Madden, J., Mamounas, L.A ., Mauk, M.D., & McCormi ck , D.A . (1984) , Neuronal sub- str at e s of basic associa t iv e learn ing . In L.R. Squ ire and Butt er s (eds.), Neurops yc ho/ogy of memory (pp. 424-442) , New York : Guil fo r d . Tin e r, M.L. (19 83. May 9) . Polyg rap h tes ts : Two vie w s [Lett er s to the edit or ] . Washin gton Post, p. AIO. Torrey, E.F., Yolken, R.H ., & Wi nf r ey , C.J. (19 82) . Cy tom eg al ovir u s anti bo dy in cerebrospi na l flui d o f s chiz o p hr enic pat ie n ts dete c te d by enzym e im munoas- say. Scienc e, 216, 892-894. Valenste in, E.S. (ed., 1 980) . The psyc hosurge r y debate . New York : W.H. Freeman. Valzelli , L. (19 73) . The 'iso lati on syn d rome' in mic e . Psy ch o ph armaco/ogi a, 31, 305-320. Valzelli , L., Bernasconi , S. & Garatt ini, S. (1981) . p-C hlorop h eny la lanin e -in -
duced muric ida l agg res sio n in male and fem ale laborato r y rats . Neurop sy- chobio lo gy , 1, 315- 32 0. Van Essen, D.C. & Maunsell, J.H .R. (1983) . Hier archic a l orga n iz a ti on and fun c- tion al stre a ms in the vis u al cort ex . Trends in Neuroscie n ce, 6, 370-375. Van Wag en en, W.P. & Herren, R.Y . (1940) . Surgi ca l div isio n of commi ss ural pat h w ays in t he corp u s callosum. Archiv e s of Neurology and Psy c hia try, 44, 740-759. Vic tor , M., Adams, R.D ., & Coll ins , G.H . (19 71) . The Wernic k e-Korsakoff Syn - drome. Oxfo r d : Blackwell. Vog el , G.W., Vog el , F., M cAbee, R.S ., & Thurmond, A.J. (19 80) . Imp ro vement of depr e ssio n by REM sleep dep ri v a ti on . Archiv e s of General Psyc hia try, 31, 247~253. Wada, J.A ., Clarke, R., & Hamm, A. (19 75) . Cerebral hemi sp h e re asym me tr y in humans. Archiv e s of Neurology , 32, 239-246. Wada, ]. & Rasmussen, T. (1960) . lntr ac aroti d inj e c ti on of sod ium amy tal for late r aliz a ti on of cerebral spe e ch domi na nce. Jou rnal of Neurosurge ry, 17, 266-282. Wage n ste e n, O.H. & Carlson, A.]. (19 31 ) . Hunge r sensati on s of a pa tie n t aft er tot a l gas tr ec to m y . P roceedin g of the Soc iety for Expe rim e nta l Bio lo g y and Medic ine , 28, 545-557. Wald, G. (1968) . Molecular basis of vis u al exc itat io n . Sci en ce, 162, 230-239. Warring t on , E.K. (1982) . Neurops y c h olog ica l stu d i es of obje c t recog nition . Phil os op hic a l tra nsactio n s of the Roya l Soc iety of London. Series B: Bio lo g i- cal Scie n ces, 298, 15-33. Warrin g t on , E.K (19 84) . Recog n it ion memory test. Wi nd sor, Eng la nd : NFER- Nelson. Warrin g t on , E.K . & Weis k rantz , L. (1970) . The amnesic syn d rome : Consoli da - tion or retr iev al ? Natu re , 228, 628-630. Weig en t, D.A . & Blalock, J.E . (1987) . Inte r acti on s betw e en the neuroendocri ne and im mune sys t e m : Common hormones and recep tor s. Immunolog ica l Revie w , 100, 79-108. Weis k rantz , L. (19 85) . On iss ues and the ori es of the human amnesti c s yn d rome. In N.M. Wein b erge r , J.L. McGaug h, & G. Lyn c h(eds.), Memory systems of the brain : Anim al and human cog n it ive pro cesses (pp. 380-415) , New York : Gu ilfo r d .
Weis k rantz , L. & Warrin g t on , E.K . (19 75) . The pro blem of t he amnesic syn d rome in man and anim als. In R.L. Issacson and K.H . Prib r am (eds.) , The hip po camp us : Vol. 2. Neurop hy s iolo gy and behavio r (pp. 411-428) , New York: Plenum. Weis k rantz , L., Warrin gton , E.K ., Sanders, M.D., & Marshall,J. (19 74) . Vis u al capa c i ty in the hemi an op ic field foll owi ng a resric ted occ ipital ablati on . Brain , 97, 709-728. Weis s, J.M . (19 68) . Eff ec ts of c op ing respo n ses on str e ss.J ou rnal of Comp ar ati ve and Phys iolo g ical Psych ology , 65, 251-260. Weis s, J.M . (19 71a) . Eff ec ts of c op ing behavi or in d if fer ent warnin g sig na l condi- tion s on stre s s pat h o log y in rats . Jou rnal of Comp ar ativ e and Phys iolo g ica l Psych ology , 77, 1-13. Weis s, J.M . (197lb) . Eff ec ts of pun is h in g the cop ing respo n se (confl ict ) on str e ss pa th o log y in r ats . Jou rnal of Comp a rativ e and Phys iolo g ical Psy c holog y, 77, 14-21. Whalen, R.E . & Edwards, D.A . (19 67) . Hormonal dete r mi na nts of the develop - ment o f m asculi ne and fem in i n e behavio r in m ale and fem ale rats . Anato m i - cal Record, 157, 173- 18 0. Wi es el, T.N. , & Hubel, D.H. (19 66) . Sp at i al and chromati c int e r acti on s in the late r al gen ic u late body of the rhesus monkey. jou rnal of Neurop hy s iolo g y, 29, 1115-1156. Wi ller, J.C ., Dehen, H., & Cambie r , J. (1981) . Str e ss-in d uced analge s ia in humans : Endog en ous opi oi d s and naloxone-reversib l e dep re ssio n of pa in refl ex es. Scie n ce, 212, 689-691 . Wi nb lad, B., Hardy, ]., Backman, L., & Nil sso n, L-G. (19 85) . Memory fun cti on and brain bio c hemi str y in normal agi ng and senil e d ementi a. I n D.S. Olto n , E. Gamzu, and S. Corkin (eds.) , Memory dysf u n cti on s : An int e g r a tio n of anim al and human research from pre clin i c a l and clin i c a l per s pe cti ve s (pp. 255-268) , New York : New York Academy of Sci en ces. Wi no cur, G. & Weis k rantz , L. (1976) . An inv estig a tio n of pa ir e d-associa t e learn ingin amnesic pat ie n ts . N europs ych olog ia, 14, 97-110. Wi rsh a fter, D. & Davi s, J.D . (19 77) . Set poi n ts .set t ling poi n t s , and the contr ol of body weig ht. Phys iolo gy and Behavio r , 19, 75- 78 . Wi tels on, S.F. ( 19 83) . Bump s on the brain : Rig h t- l ef t anato m i c asym metr y as a key to func tio n al late r aliz a ti on . In S,J . Sega l ow itz (ed.) , Langu age fun c-
lions and brain orga n iz a ti on (pp. 117-144) , Orlando, FL : Academi c Press. Wi tels on, S.F. & Palli e, W. (19 73) . Left hemi sp h e re spe c ia li z a ti on for lang ua ge in the newborn : Neuroanato m i ca l evid e nce of asym m ert y. Brain , 96, 641-646. Woods, J.W . (19 56) . Tami ng of the wil d Norway rat by rhin e nceph a li c l esio n s. Sci en ce, I78, 869. Wurt z, R.H . (1969) . Vi su al recep tive field s of str ia t e cort ex neurons in awake monkeys . jou rnal of Neurop hy s iolo g y, 32, 727-742. Wu rtz, R.H . & Albano, J.E . (19 80) . Vi su al-moto r func tio n of t he prim ate s up erior coll icu lus. Annnual Revie w of neurop hy s iolo g y, 32, 727-742. Young, A.W . 0982) . Asym me try of c erebral hemi sp h eric f un cti on dur ing develop- ment. In J.W .T. Dick erson and H. McCurk (eds.), Brain and behavio r al develop me nt (pp. 168-202) , Surrey, Eng la nd : Surrey Un ive rsity Press.
기 가락음조치료 (melod ic int o n ati on the rapy ) 329 가시 스펙 트럼 (visib le spe c tru m) 128 각성 없는 학습 (lea rnin g witho ut awareness) 305 간뇌 (die n ceph a lon) 75 간뇌성 건망증 (d i ence pha li c amnesia ) 301, 302 간-문맥 순환 (he p a ti c- p o rt al circ ulati on ) 2u 간상체 (rod) u4, u9, 120, 122 간신경 원 (int e m euron) 82 간질 (ep ilep sy ) 14 간질 액 (int e r stit ial fluid ) 201 갈증의 이중고갈이론 (double dep le ti on the ory of thirs t) 197, 201 감각신경 원 (sensory neuron) 290, 293 Gamma-a m i no buty ric aci d( GABA) 55, 57, 193, 356, 357 감정장애 (aff ec ti ve diso rder) 334, 347 강박관념 (obsessio n ) 357, 358 강박장애 (obsessiv e comp ul siv e dis o rder : OCD) 355, 357, 358 강박행동 (com p uls i on) 357, 358 개막(t ec t or i al membrane) 152 거대세포 바이러스 (c yt ome g alov i rus: CMV) 338 거대신경원(gi an t neuron) 39 건망증 (amnes i a) 285, 297 경동맥 (caroti d arte r i es ) 205 경부신경 (cerv ica l nerve) 65 계열처리이론 (ser i al pro cessin g the ory ) u3, 145 고막(tymp an ic membrane) 149 고실계 (scala tympa ni ) 152 고장액 (hy perton ic soluti on ) 199 고전적 조건화 (class ic al cond ition i ng ) 286, 287 골격근 (skele t al muscle) 176 골수 (bone marrow) 72 골지세포 (Gol gi cell) 194 골지 영 색 (Golgi sta i n ) 28
골지 체 (Golgi body) 34 공간가합 (s pa ti al summati on ) 62 공간빈도 (s p a ti al freq u e ncy) 135 공간빈도모형 (spa t ia l freq u e ncy model) 132, 134, 136 공간적 위치 (spa t ia l locati on ) 130 공격 성 (ag gres sio n ) 276 공포증(p hob i a) 355 공황장애 (panic dis o rder) 355 과립세포(gr anule cell) 87, 194 과분국화 (h yperp olar i za ti on) 41, 45, 48, 121 과식증 (h yperp ha gi a) 213 과잉활동성 (hyp er acti vity) 92 광색소(p ho t o pigm en t) 119 광장공포증 (a g ora p hob i a) 355 광파장(lig h t waveleng th) 131 교감신경 계통 (s ymp a th e tic nervous sys t e m ) 67, 68, 268, 271 교상질 (substa n ti a gel ati no sa) 166 교태 행 동 (pro cep tive behavi or ) 234 구강건조이론 (dr y mouth theo ry ) 199 구피질 (old cort ex ) 78 구형낭 (saccule) 150, 157 국소신경 원 (loc al neuron) 52 국소회로(l ocal circ uit ) 81, 87 궁상핵 (arcuate fas cic u lus) 327, 344 국파 (s pi ke) 46 근긴장 (s p as ticity) 183 근방추기관 (muscle spi nd le orga n ) 182 근소포체 (sarcop la smi c retic u lum) 180 근원섬유 (m yro fi ber) 180 근육긴장 (muscle ton e) 183 글루타메이트(g lu t ama t e) 55 글리세린(g l y ce ri n) 219 긁기 반사 (scra t ch refl ex ) 184 급속 안구운동 (ra pi d eye movement : REM) 249, 251, 262, 348
기 계 적 수용기 (mechanoreceoto r ) 162 기능성 비대칭성 (fun cti on al asym me tr y ) 307 기 능적 안정 화 (fun cti on al . valid a ti on ) 96 기면증 (narcole p sy) 22 기억 없는 학습(l earn i n g wit ho ut memory ) 305 기 저 수지 상돌기 (basal dendrite ) 33, 80 기저 전뇌 콜린성 체계 (basal fore brain cholin e rgi c sys t e m ) 109 기저핵 (basal gan g lia ) 76, 180, 191, 318, 344, 359 길항물질 (anta g o n is t) 57, 58 꽃가지 형 수용기 (flow er-spr a y recept or ) 182 L 난독증 (d y slex i a) 308, 316 난원창 (oval win d ow) 149, 150, 152, 157 난형낭 (u t r i cle) 157 날록손 (naloxone) 170 날조화 (co nf abula ti on) 302 남성 화 (masculin iza ti on ) 241 내 과립 충 (int e r nal gran ular laye r ) So 내 비 피 질 (ento r hin a l corte x ) .29 6, 344 내 융대 (media l lemnis c us) 166, 284 내 이 (inn er ear) 149 내인성 아편 (endo g eneous opi oi d s ) 169 내 인 성 주기 (endoge n eous cyc l e) 263 내추체충(i n t ernal pyram i da l lay er ) So 내 측 시 상하부 (media l hyp o th a lamus) •2 78 내 측 시 상핵 (medi al tha lami c nuclei) 168 내측영역 (media l area) 318 내축전뇌속 (med i al for ebrain bundle:: M FB) 102, 284 내축전시삭핵 (media l pre op tic nucleus : MPON) 246, 247 내축중격 (med ial sept um) 109 내측 측두영성 건망증 (me di al tem p or al lobe amn 函 a) 301, 302 내측층간핵 (med ial int e r lamin a r nucleus) 127
노르아드레 날린 (noradrenalin e : nore pi ne p hr i ne 을 보라) 노르아드레날린성 (noradrenali ne rgi c) 기능 93 노르에 피 네 프린 (norepi ne p hr in e : noradrenali ne ) 25, 55, 57, 69, 109, 227, 262, 349, 351, 356 노인성 반점 (sen ile pla q ue ) 108 녹색광원추체 (gree n cone) 128, 130 농도 기 울기 (concentr at i on grad ie n t) 44 뇌 간 (bra i n ste m ) 78, 184 , 283 뇌 간망상체 (brain s te m retic u lar form ati on : BSRF) 78, 260 뇌 격 리 (encepa l e iso l ) 258 뇌고정장치 (ste r eota x ic ins tr u ment) 25 뇌교(p ons) 67, 75 뇌교술상체 -시 각피 질 (po ns-g e n ic u late - occi pital : PGO) 발사 262 뇌궁(forni x) 78 뇌궁체 (fim bri a- fo rnix) 102 뇌궁하기관 (sub foni cal orga n : SFO) 204 뇌 량 (co rpus callosum) 75, 81, 308, 310 뇌 막 (men i n g es) 74 뇌신경 (crani al nerves) 64 뇌 일혈 (str ok e) 189 뇌전도 (elec tr oence p halo gr am : EEG) 26, 250 뇌 주사기 법 (brain scann ing tec hniq u e) 309 뇌 하수체 (pituitary gla nd) 76, 227 뇌하수체 전영 (ante r i or pituitary gla nd) 223, 227-231 뇌 하수체 후영 (po ste r io r pituitary gla nd) 227-229, 237 C 다발성 경화증 (mul tip le sclerosis) 325 다원기록법 (poly g rap h ) 272 다중 시냅스성 반사(p ol y s yn a pti c refl ex ) 181 단국성 장애 (monop ol ar dis o rder) 347 단기 기 억 (short -te n n memory ) 105 단기습관화 (sho rt-te nn habit ua tio n ) 290
단소영 (sim p le lobule) 294 단속성 운동 (saccades) 143 단순세포 (s i m p le cell) 133 단안성 (monocular) 신경원 136 단일 시냅스성 반사 (monos yn a ptic refl ex ) 181 담창구(g lobus pal li du s) 191 담체단백질 (ca rri er pro te i n ) 225 당원 (gly c o g en ) 207 W 세포 125, 126 대각대 (dia g o n al band) 161 대 상피 질 (cing u la te cort ex ) 168, 344 , 347 대 상회 (cing ula te gyrus) 78 대측 (con t rala t eral) 두사 116 도(島 , ins ula) 160 도약전도 (sal t a t o ry conducti on ) 50, 52 도파민 (dop am i ne ) 25, 55, 57, 192, 275, 343 34 5, 349, 352 동공(p u pil) 114 동등성 가설 (eq ua l-at- bi r th hyp o th e sis ) 329 동안근 (oculomoto r muscle) 177 동안신경 (oculomoto r nerve) 65 동측(ip s p la t eral) 두사 116 두부반웅 (ce p ha li c respo n se) 216 두정 엽 (pa rie t a l lobe) 75 등골 (s t a pe s) 149 디아제팜 결합억제제 (diaz epa m bin d i ng inhibi t or : OBI) 357 딸세포 (dau gh t er cell) 86 근 랑비에 결절 (nodes of Ranv ier ) 32, 33, 35, 50 레 닌 (reni n) 204 레세르핀 (rese rpi ne) 57, 352 레티닌 (re ti nene) u9, 122 로돕신 (rhodo psi n) u9, 122
류-엔케 팔린 (leu -enkeph a li n) 55, 169 릉 (C ri s ta) I58 리보솜 (r i bosome) 34 리보핵산(ri bonucle ic aci d : RNA) 34 리븀(lithi wn) 치료 354 릴러 생쥐 (reeler mouse) 89 립 프철 (lymph node) 72 口 만발성 운동장애 (tar di ve dys k in es ia ) 342, 3 선 말초신경 계통(perip heral nervous sys te m : PNS) 35, 63, 64 망막 (re tin a) 114 망막시상하부로 (re tin oh yp o th alam ic trac t) 142, 265 망막표상 I45 망상 (delu si on) 335 망상형 정신분열증(p arano i d sch izo p hr eni a) · 335 맹시 (blin d sig h t) 144 맹점 (b!ind spo t) 114, 144 메스칼린 (mescalin e ) 140 메이네르트 기저 신경핵철 (nucleus basalis of Meyn e rt : nbM) 109 메트엔케팔린 (me t -enke p ha li n) 55, 169 멜라토닌 (melato n i n) 227, 266 면역 글로블린 (immu nog lo buli n) 108 면역체계 (immu ne sys tem ) 71,· 2 76 모노아민 가설 (monoam i ne hyp o th es is of depr e ssio n ) 351, 352 모노아민 산화효소 (monoam i ne oxid a se : MAO) 349 모노아민 산화효소 억 제 제 (monoa mine oxi da se inhibi to r s : MAO is) 348, 349 모서리탐지기 (edge dete c to r ) 133 무축색세포 (amac rin e cell) 123 미골신경 (coccyg e al nerve) 65, 72 미뢰 (tast e bud) 158 미상핵 (caudate nucleus) 21, u5, 191 미 세사 (m ic ro fi lamen t) 34
미세 신경교세포 (m ic ro gli a) 35 미세신경원 (m ic roneuron) 94 미소관 (m i cro t ubule) 34 미오글로빈 (my og l o bin ) 177 미오신 (m y os i n) 176 미주신경 (vagu s nerve) 65, 67, 209, 212 미토콘드리아 (m it ochond ri a) 34, 54 민감화 (sens iti za ti on) 286, 287, 292, 293 n 바구니 세포 (baske t cell) 194 바비듀레이트 (barb it ura t e) 350, 356 바소프레 신 (vasopr e ssin ) 55, 228 바이러스 가설(vi ral hyp othe sis ) 338 바이오피이드백 (bio f e e dback) 269 반구내통로(i n tr ahem i s p her ic channel) 81 반구편재 성 (hemi sp h e re localiz a ti on ) 322 발모광(t r i cho till oma ni a) 환자 359 발정주기 (estr us cyc l e) 234 방사산불두과 옥도 (rad i o-o p a q ue iod in e ) 324 방사성 탄소동위 원소 (rad ioa cti ve carbon ato m ) 29 방사 신경교세포 (rad i al glia) 35, 36, 88 방추내근(i n tr a fu sal muscle) 182 방추세 포충 (spi nd le cell laye r ) 80 방추와근 (ex tr a fu sal muscle) 182 배내측핵 (dorsomed ial nucleus) 161 배의측영역 (dorsolate r al area) 318 배주대융대체계 (dorsal column-medi al lem nisc al sys t e ht) 165 백 질 (wh ite matt er ) 36, 66, 72 Wern ick e 실어증 베타센도르핀 (be t a endorp hin) 55, 170 벤조디 아제 핀 (benzod iaz epi ne ) 356, 357 벨마장디 (Bell-Mage n die ) 법 칙 16
변연계 (lim bic sys t e m ) 78, 275, 283 병 행 처 리 이 론 (pa rallel pro cessin g the ory ) u 3, 126, 145 복내측 시상하부 (ven tr omed i al hyp o th a lamus : VMH) 213 복부신경 절 (abdomi na l habitu a tio n ) 289 복합세포 (comp le x cell) 133 복후내측핵 (ventr al pos te r i or media l nucleus) x59 복후의측핵 (ventr al pos te r i or late r al nucleus) 166 본유적 호르몬 주기성 (inh erent hormonal cyc l ic ity) 232 봉선체 계 (raph sys t e m ) 261 봉선핵 (raph nuclei) 278 부교감신경계동(p aras ymp a th e tic nervous sys t e m ) 67, 68, 70, 268, 271 부동섭모 (s t ereo ci la) x58 부신 (adrenal gla nd) 69 부신경 (accessory nerve) 65 부신수질 (adrenal medulla) 67, 193, 277 부신피질 (adrenal cort ex ) 223,225 부호화 가설 (encodin g hyp o th e sis) 303 분국화 (po lari za ti on ) 48 분자생물학 (molecular bio lo g y) 5 분할뇌 (sp lit brain ) 312, 322 불면증(i nsomn i a) xx, 256 불안신경증 (anx i e ty neurosis) x x 불안펩티드 (anx i e ty peptide ) 357 불옹기 (refr ac to r y period ) 48 불활성화(i nac ti va ti on) 42 브로카 대 (diag o n al band of Broca: dbB) 109 Broca 실어증 325 Broca 영역 327 bic uc ullin e 57 비특수 또는 확산동로 (nons pecifi c or diffuse channel) 81 비특이성 불안 (nonspe cific arucie ty ) 69
人 사구체 (glo merulus) 函 사두근(q uadr ic e p s muscle) 181 사시 (str a bis m us) 137 산개 (div e rge n ce) 127 삼반고리관(thr ee semi circ ular canals) 150, 157 삼원색 이론(tric hroma tic the ory ) 16 삼차신경 (trig e mi na l nerve) 65 삼투압 (osmo tic pre ssure) 203 상경 신 경 절 (sup er io r cervic a l gan g lion ) • 67 상구체 (sup erior coll icu li ) 76, 116, 142 상장간막신 경 절 (sup er io r mesente r i c gan g lion ) 67 상측두희 (sup er io r tem p o ral gyrus) 323 상대 불응기 (relati ve refr ac to r y per io d ) 48 상행신경로 (ascend i n g nerve trac t) 73 색소성 망막영 (reti ni t is pigme nto s a) 120 색채대립세포 (color op po nent cell) 130 서골비강기관 (vomeronasal orga n ) 162 서 파수면 (slow wave sleep ) 252 선천성 부신 비대증 (con g en it al adrenal hyp e rtr o p y : CAH) 244 설인신경 (glo ssop h aryn ge al nerve) 65 설 하신 경 (hyp o g lo ssal) 65 섬유성 성상 신경교세포(fi brous astr oc yt e) 36 성상세포 (s t ella t e cell) Bo, 194 성상 신경교세포 (as t roc yt e) 35 성상신경철 (s t ella t e gan g lion ) 67 성 장원추(gr o wth cones) 90 성적 이형 (sexual dim orp h is m ) 230, 235 세로토닌 (sero t on i n : 5-HT) 26, 55, 57, 109, 257, 280, 349, 351, 352 세 부특칭 담지 모형 (fea tu r e dete c to r model) x3 2, x3 4, x3 6 세포내 구획 (int r ac elluar comp a rtm e nt) 201 세 포독성 T 세 포 (cy tot o x ic T-cell) 71 세포의 구획 (extr ac elluar comp ar t me nt) 201
세포의 분화 (d iff eren ti a ti on) 90 세 포의유출 (exocyt os is ) 59 소뇌 (cerebellwn) 75 소뇌 각 (cerebellar pedu ncle) 194 소뇌증 (m ic rence p hal y or mic ro enceph a ly) 92 소방세포(tuft ed cell) 161 소주 (m ini colwnn) 331 소주 (trab eculae) 74 소포체 (endop la smi c reti cu lwn) 34 속근(fa s t muscle) 177 송과선 (pine al gla nd) I 3, 75 니 ,6 수면발작 (narcole p s y) 257 수면성 무호흡증 (slee p apn e a) 257 수양액 (aq ue ous humor) u4 수용기 전위 (recept or pot e n ti al ) 153 수용성 실어증 (rece pti ve aph asia ) 325, 326 수용야 (rece pti ve field ) 124 수용행 동 (recep tive behavio r ) 234 수정 액 (viter ous hwnor) u4 수지 상가시 (dendriti c spi ne ) 34 수지 상돌기 (dendrite ) 34 수지상수지상 시냅스 (dendrodendr itic syn a ps e ) 56 수질 (medulla) 69 수초 (m y e li n sheath ) 32, 35, 50 수축기 제 (contr ac ti le mechanis m ) 59 수평세포 (hor i zon ta l cell) 123 순막 (nic ititating membrane) 294 순환성 와해 (circ ulato r y collaps e ) 슈반 (Schwan) 세포 35 스테로이드 호르몬 (s t ero i d honnone) 223, 224 , 225 스트레스 유발무동 (s tr ess- i nduced analge s ia : SIA) 169, 170 -1 72, 술개건 반사 (kneej erk refl ex ) 181 습관화 (hab itu a ti on) 286, 293 승모세포(mitr al cell) 161
시 각의 우세 성 (ocular domi na nce) 136 시 각피 질 (visu al cort ex ) 332 시간가합(t em p oral summati on ) 62 시개 (op tic tec tu m ) 95, 142 시개전영역 (pre te c ta l area) 300 시교차 (o ptic chia sm ) 116, 310, 311, 322 시교차상핵 (sup ra chi as matic nucleus : SCN) 116, 142, 249, 265 시냅스 (s yn a pse) 25, 32, 34 시냅스 간극 (s yn a ptic cleft ) 33 . 54, 56 시냅스 개조 (s yn a ptic remodeli ng ) 102 시 냅 스 발생 (syn a pt og e n esis ) 93 시냅스전 신경원(p res yn a ptic neuron) 53, 345 시냅스전종말(p res yn a pti c term i na l) 33, 54, 56, 58 시냅스주머니 (syn a pt ic vesic le ) 54, 58 시 냅 스 지 체 (syn a pt ic delay) 59 시냅스후 수용기 (po sts yna pt ic recept or ) 56, 59 시냅스후 신경원(po s t s yn a ptic neuron) 53 시냅스후 전위 (po sts yna pt ic pot e n ti al) 59 시 맹 (bli nd ness) 144 시상절편 (sag ittal secti on ) 75 시상하부 (h yp o th alamus) 75, 76, 283, 318, 319 시 상하부뇌 하수체 문맥 체 계 (hy po th a lami c-pituitary por ta l sys tem ) 229 시상하핵 (subth a lamic nucleus) 191 시신경 (op tic nerve) 114 시신경교차 (o ptic chia sm ) 76 cAMP(cyc l ic adenosin e monop ho sph a te ) 226 cGMP(cyc l ic gua nosin e monop ho sph a te ) 121, 227 시행착오학습(tri al-and-error learnin g ) 18 신경 결핍 성 초과민증 (denervati on supe r sensiti vity) 344 신경관 (neural tub e) 86, 107 신경교 반혼(gli a scar) 36 신경교세포(gli al cell : glia) 35 신경구 (neural groo ve) 86 신경근 연접 (neuromuscluar jun cti on ) 178
신경내분비반사 (neuroendocr i ne refl ex ) 238 신경농 (nerve deafn e ss) 156 신경독소 (neuro t ox i n) 25 신경롱 (neural crest) 86 신경 변조물질 (neuromodulato r s) 55 신경분비 (neurosecreto r y ) 228 신경섬유의 엉킴 (neurofi br i lla ry tangle ) 108 신경세사 (neuro fi lamen t) 34, 107 신경아세포 (neuroblas t) 87 신경원 (neuron) 32 신경자국요소 (neuro t ro pi c fac to r s) 36 신경 전달물질 (neurotr an smi tter s) 25, 31, 34, 55, 58, 290 신경 조정 물질 (neuroregu la to r ) 55 신경증 (neuros i s) 11 신경총(p lexus) 66 신경충격 (nerve im p ul se) 31, 41, 46, 53 니 ;8 신경 판 (neural pla te ) 84 신장반사 (s t e rt ch refl ex ) 181, 184 신장수용기 (str et c h recept or ) 183 신피 질 (neocorte x ) 78 실독증 (dys l exia ) 89 실무율 법 칙 (all-or-none law) 46, 59 실어중 (a p has i a) 191, 320, 324, 3 기 실어증 척도 (a p has i a quo ti en t : AQ ) 328 실행증 (a p rax i a) 191, 319 심 리 신경 면 역 학(psy choneuro im munolog y) 72 심 신관계론 (m i nd-bod y pro blem) u 십 신증 (ps yc h osomati c diso rder) 11 십장근 (card i ac muscle) 176 。 아가미 수축반사(gi ll withd rawal refl ex ) 288 아드레 날린 (adrenalin e : e pi ne phri ne 을 보라)
아드레날린 쇄도 (adrena li ne rush) 69 아밀로이드 (am y lo i d) 108 아세 틸콜린 (acety lc holi ne : ACh) 26, 55, 57, 101, 109, 193 AChE (acety lc holin e ste r ase) 1 u 아포모르핀 (ap om orp h in ) 57, 275 안드로겐 (androg en ) 223, 224, 225 안드로겐 둔감증후 (androg en ins ensiti vit y syn d rome) 245 안면경 련증(tic) 342 안면신경 (facial nerve) 65 안와영역 (orbit al area) 318 안와전두피질 (orb it o fr on t al cort ex ) 161 안정 전위 (restin g pot e n ti al) 40 안지오텐시노겐 (ang iot e n sin o g en ) 204 안지오덴신 II (ang iot e n sin II) 204, 205 알도스테론 (aldoste r one) 205 알츠하이 머 병 (Alzheim er's dis e ase) 23, 84, 106 Alph a-meth ylt y ros in e 57 암전류 (dark current) 121 암점 (scoto m ata ) 144 암페 타민 (amp he ta m i ne ) 57, 345 압력 수용기 (barorecept or ) 205 액틴복합체 (acti n comp le x) 176 양극성 우울증 (b ip olar depr e ssio n ) 350 양극성 장애 (bip o lar diso rder) 347, 353, 354 양국세포 (b ip olar cell) 87, 123 양성 증후(p os iti ve sym ptom ) 340 양안성 (bin o cular) 신경원 136 양전자방사단층촬영 (po sit ron emi ssi o n tom og rap h y : PET) 29, 324 , 339, 359 억제성 시냅스후 전위 (inh ib i t ory pos ts yna pt ic pot e n ti al : IPSP) 59, 61 억 제 성 신경 원 (inh ib i t or y neuron) 178 얼굴실인증(p roso p a gn os i a) 140 얼굴람지 (fac e dete c to r ) 신경 원 140 S 기 (S pha se) 86 에스트라디올 (es tr a di ol) 236
에스트로겐 (es tr o g en) 223,225,236 A 델타 (A-del t a) 섬유 163 A-V 비율 (a rt ery -ve i n rati o) 209 에피네프린 (ep ine ph r in e : adrenali ne ) 55, 69, 227, 268, 349 X 세포 I25, m6 엔도르핀 (endorp hine s) 169 L-Dop a 1 언 여성성칭 결여증 (de f em ini za t i on) 241 역설적 수면(p aradox ic al sleep) 249, 251 역 치 (thres hold) 46 역 치 하 자극 (subth res hold sti m ulati on ) 46 역행성 건망증 (re t ro gr ade amnesia ) 303 연막(pi a mate r ) 74 연수 (medulla oblong at a ) 67, 72, 75 연수추체 (medullary pyram i d) 74, 188 연합피 질 (associa t i on cort ex ) 145, 146 열 (sulcus) 74 오르가즘 (or g asm) 71 5-6-cl ihyd roxy tryptamine (5-6 - DHT) 26 5-HT(sero t on i n 을 보라) 옥시토신 (oxy toc in ) 55, 223, 228 온도 수용기 (the nnorecep tor ) 162 옵신 (o psi n) 119 옹고집행동(pe rsevera ti ve behavi or ) 302, 320 . Wada 겁사 321, 322 와우각 (cochlea) 150, 260 와우신경 (cochlear nerve) 153 Y 세포 125, 126 완근 (slow muscle) 177 의과립층 (ex t ernal gran ular laye r ) 80 의 이 (exte r nal ear) 149 의청도 (ear canal) 149 의추채층 (ex te rnal pyram i da l laye r ) 80 의측술상핵 (lat e r al gen icu late nucleus : LGN) 101, 118, 127
의측시삭전(l a t eral pre op tic : LPO) 영역 203 의측 시상하부(l a t eral hyp o th a lamus : LH) 161, 217, 278 의 측 중격 (lat e r al sept um ) 278 의 측척 수시 상로 (lat e r al spi no th a lami c tra ct ) 73 의측후삭로(l a t eral olfa c to r y tra ct) 161 의두막 (man tl e shelf ) 288 의피 (ep ide rmi s) 162 요부신경 (lum bar·n e rve) 65 우세 성 가설 (domi na nce-at- b ir th hyp o th e sis ) 330 우울증 (de p ress i on) 11, 54, 333, 347, 352 우측귀 우세 성 (rig h t ear advanta g e ) 323 우측두정 영 지 각 부전증후군 (rig h t par ie t a l imp erc ep tion syn d rome) 315 운동단위 (moto r unit ) 177 운동령 (mort or area) 319 운동신경 원 (morto r neuron) 290 원추체 (cone) 114, 119, 120, 122 원형질성 성상 신경교세포(p ro t o p lasm ic astr oc y tes ) 36 위궤양(g as tri c ulcer) 271, 273, 274 위 버 생 쥐 (weaver mouse) 88 위약효과(p lacebo eff ec t) 172 유기 음이온 (or g an ic ani on ) 41 유동성 모자이크 모델 (flui d mosaic model) 37 유두체 (mammi llar y body ) 76, 301 유리지방산(fr ee fatty aci ds ) 208, 209, 219 유모세포 (ha i r cell) 152, 153 유발성 배 란자 (ind uced ovulato r ) . 231 유사분열기 (mi toti c pha se) 86 유아자폐증(inf an til e auti sm ) 361 유전설정모형 (ge neti c pro g ram ming model) 139 6-hy dr oxy do p amine (6-0HDA) 25 윤나선형 수용기 (annulospi ra l recept or ) 182 음색 (timb re) 148 음색변별(ti mbre dis c rim in a ti on ) 과제 3I6 음성 증후 (neg at iv e sym ptom ) 340
음압기 (sound level mete r ) 148 음핵 (cli tor i s) 71 응고화 가설 (consolid a ti on hyp o th e sis ) 303 의미기억 (semantic memory) 105 2- 디옥시글루코스 (2-deox yg lucose : 2-DG) 28, 115 이미프라민 (im ip ra mi ne ) 348 이 온성 전류 (ion i c current) 40, 59 이온통로(i on channel) 38, 40, 42 이원론 (dua li sm) 13 이중이론 (du pli c ity the ory ) 122 이프로니아지드(ip ro ni az i d) 352 이형 건망증(tw o amnesia ) 가설 301 인 슐린 (ins uli n) 207, 215 인 자각성 이론 (cog nition -arousal the ory ) 270 인지물질 (recog nition molecules) 91 인출장애 (retr iva l diso rder) 가설 3°4 인터페론(i n t e rf erone) 275 일내주기성 (circ adi an rhyt hm) 142, 198, 249, 263 일반화된 공황장애 (ge nerali ze d pan ic dis o rder) 355 일원론 (mo ni sm) 13 입피던스(i m pe dance) 150 x: 자국생성 무동 (s ti mula ti on- p roduced analge s ia : SPA) 170 자기공명영상법 (magn e tic resonance ima gi ng : MRI) 29, 339 자기 수용기 (auto r ecept or ) 345 자발성 배란자 (s p on tan eous ovulato r ) 231 자연살해세포 (na tur al kill er cell : NK) 275 자율신경계통 (au t onom ic nervous sys t e m : ANS) 64, 67 자일로카인 (xy lo cain e ) 48 자폐증 (au ti sm) 316, 334 , 362 장기 기억 (lon g -ter m memory ) 105 장기 습관화(l o ng-t erm habitu a tio n ) 290
장기 증강 (lon g -ter m pot e n ti at io n : LTP) 296 장소설 (pla ce theo ry ) 16, 154, 161 저장액 (hyp o to n ic soluti on ) 199 저혈당중 (h yp ovolem i a) 204 적색광 원추체 (red cone) 128, 130 전각세포 (ven tr al horn cell) 74, 180 전교련 (an t er i or commi ss ure) 81, 308, 310 전근 (ven t ral root) 16, 65, 66 전기경련 충격치료 (elec tr oconvuls i ve shock the rapy : ECT) 303, 350 전 기 긴장성 전도 (electr o to n i c conducti on ) 51 전기적 기울기 (electr ica l grad ie n t) 43, 44 已 ;8 전기적 시냅스 (elec tric al syn a ps e s) 53 전뇌 격 리 (cerveau iso l ) 258 전 단력 (shearing forc e) 15 3 전도농 (conduc ti ve deafn e ss) 157 전두안야(fr on t al eye - fi e ld ) 318 전두영 (fron ta l lobe) 75, 317, 318, 319, 346, 359 전산 단충조영술 (com pu t e ri zed axia l tom og rap h y : CT) 23, 3 걱, 339 전압활성 화 (vol t a g e-ac ti va t ed) 47 전의측체계 (ante r olate r al sys t e m ) 165 전운동피질(p remo t or cort ex ) 180 전이상피질(perprifo rm cort ex ) 161 전정계 (scala vestib u li ) 152, 158, 260 전정계통 (ves ti bular sys t e m ) 157 전정핵 (vestib u lar nuclei) 158 전천공질 (ante r io r perfor ate d substa n ce) 161 전측 시 상하부 (ante r i or hyp o th a lamus) 202 철대불응기 (absolute refr ac to r y per iod ) 48 절세포(g an gli on cell) 87, 122-124 절전신경 원 (pre ga n g lion i c neuron) 68, 70 절후신경 원 (po stg a ng lion i c neuron) 68 점진성 전위 (grad ed poten ti al) 46, 5:z, 59 정 단 수지 상돌기 (api ca l dend rite) •3 3, 8o 정 신 맹 (ps yc h i c blin d ness) 140
정신분열증 (sc hi zo p hren i a) 11, 54, 333, 334, 350 정 신수술 (psyc hosurge r y) 346 정신신체 질환(p s y chosoma tic dis o rders) 70, 271 정 신약리 학 (psych op ha nnacology ) 58 정중핵 (mi dl in e nucleus) 301 정향반응 (or i en ti n g respo n se) 143 제 2 전령 계동 (second messeng e r sys t e m ) 226 제 뇌 경 직 (decerebrate rig idit y ) 185 제사설 (volley the ory ) 155 Jam es-Lang e 이론 269 제트 랙 (jet rag) 249, 264 조절접이론 (se tp o i n t theo ry ) 219 조증 (ma nia) 347, 3 54 주의력결핍 과잉활동장애 (att en ti on -defic it hyp er acti vity dis o rder : ADHD) 360 중간계 (scala me dia) 152 중격 핵 (sept al nucleus) 22, 78 중뇌 (mesenceph a lon) 67, 75 중뇌 피 개 (midb rain teg m e ntu m) 3 선 중다감각수용기 (po lys e nsory recep tor ) 163 중다성격장애 (multip le per sonalit y dis o rder) 334 중다요인가설 (multif ac to r hyp o th e sis) 220 중십와(fo vea) 122 중이 (midd le ear) 149 중추신경 계통 (cen tr al nervous sys t e m : CNS) 64 지방항정수용기 (lipos ta ts) 219 지질막(lipi d membrane) 32 지질이중층(lipi d bila y e r ) 37, 42 진행성 건망증 (an te rogr ade amnesia ) 298, 305 진행성 마비 (pa resis) 191 진화계수(pr o gre ss i on ind ex) 78 전화론 (evolu ti o ni sm) 16 질산은 (s il ver nitrat e ) 28
-K 척 수 (spi na l cord) 64, 75 척수반사 (s pi nal refl ex ) 181, 184 척수소뇌로 (s pi nocerebellar sys t em ) 73 척수쇼크 (s pi nal shock) 185 척 수신경 (sp ina l nerves) 64, 66 척수신경절 (spi nal gan g lion ) 65, 66 척수피개로 (s pi no t ec ta l tra ct) 73 척추전만(l ordos i s) :23 5 천공신경 (sacral nerve) 65 천공로 (perfor ant pat h w ay) :29 6 철수반사 (w ith drawal refl ex ) 184 청반(l ocus coeruleus) 81, :26 1 청색광 원추체 (blue cone) 1:28 , 130 청소골 (oss i cle) 149 체성감각 계통 (somes the tic sys t e m ) . 16:2 , :26 0 체성신경 계통 (soma tic nervous sys t e m ) 63, 64 체 절 (dermato m e) 66 촉진성 간신경 원 (fac ili tat o r y inte m euron) :29 3 총상충 (ple xifo rm laye r ) 80 최종 공통로(fi nal common pat h ) 180 추골 (malleus) 149 추론발사 (corolla ry dis c harge ) 190 추체로(p ram i dal tra ct) 74 추체세포(pyr am i dal cell) 80 축색 구 (axon hillo ck) 49 축색돌기 (axon) 3:2, 34 축색세포체 시냅스 (axosoma tic syn a ps e) 56 축색수지상 시넵스 (axodendr itic syn a ps e) 56 축색종말 (axon ter mi na l) 33, 34, 38, 53, 56, 58, :29 1, 345 축색축색 시냅스 (axoaxon ic syn a ps e) 56 충격 발생부위 (spi ke init i ati n g regi on ) 6:2 췌 장(p ancreas) :20 7
측두평 면 (pla num tem p or ale) 308 측두하피질 (infer io r tem p or al cort ex ) 139, 140 치 매 (dementi a) 106 침골(i ncus) 149 r. 칼페 인 (calpa i n) 296 캅그라스 증후군 (Capgr as syn d rome) 320 케돈 (ke t one) 208, 209 코르사코프 정 신병 (Korsakoff s p sy c h osis ) 298 코르티 기관 (or g an of cort i) 152 코르티졸 (co rti sol) 244 코리티코스테로이드 (co rti cos t ero i d) 225, 276 코카인 (cocain e ) 57, 349 • 콜레시르토키닌 (cholec y s t ok i n i n: CCK) 55, 212 choli ne acety lt ra n sfe r ase(ChAT) 109 콜린성 가설 (choli ne rgi c hyp o th e sis ) x1 0 크레 틴 병 (cretin ism ) 91 크롬친화성 세포 (chroma ffi ne cell) 69 클쿠 근 立 쿠마전 (chlo rp romaz i ne) 57, 341, 342 클로미프라민 (clomi pra mi ne ) 359 키니네 (qu in in e ) 159 E 타페 둠(tapetum) 127 탄도운동 (ba lli s tic movement) 195 탈분국봉쇄 가설 (depo l ari za ti on blockade hyp o th e sis ) 345 탈분국화 (de po la ri za ti on) 41, 45, 47, 158 탈습관화 (dish abitu a tio n ) 287 탈억제 (disin h ibit i on ) 121 태 아알코올증후군(f e ta l alcohol syn d rome : FAS) 92 테스토스테론(t es t os t erone) 239, 240-2 선, 278, 279
테트로도독신(t e tr odo t ox i n : TTX) 43 트리 요오드티 로닌 (triiod oth yron in e ) 227 트립토판(trypt op han) 257 특수감각통로 (s pecifi c sensory channel) 81 티 록신 (thyrox in ) 227 티오잔텐 (thiox anth e nes) 342 工 par achloroph e ny la lani ne (PCPA) 261 파치니소체 (Pa ci nian corp u scle) 164 파킨슨 병 (Parkin s on's dis e ase) 23, 192 팽대부 (am p ulla) 158 팽 대 정 (cup ul a) 158 퍼킨지 세포 (Purk i n j e cell) 194 페노티아전(p heno thi az i ne) 342, 348 페로몬(p heromone) 162 편도체 (amy gd ala) 78, 140, 278, 318, 347 평 형 전위 (equ i li br iu m pot e n ti al) 41 평활근 (smoo th muscle) 176 포도당안정 이 론 (glu costa t ic the ory ) 209 포유반사 (nurs i n g refl ex ) 238 표현성 실어증 (ex p ress i ve aph a sia ) 325 프로락틴 (pro lacti n) 237 피각(p u t amen) u5, 191 피 라세 람(pi race t am) 111 피소스티그민(p h y sos tigmi ne) 57, u1 피 질 (corte x ) 69 피 질주 (cort ical column) 331 피질척수로 (co rtic os pi nal trac t) 74, 188 핍 돌기 신경 교세포 (olig o dendrog lia) 35
중
하구체 (infer i or coll icu li) 76 하장간막신경 절 (infer io r mesente r ic ganglion ) 67 하행신경로 (descend i n g nerve trac t) 73 학습된 무기 력 (lea rned helpl e ssness) 172 할로페 리 돌 (halope r ido l) 275 항상성 (homeosta si s) 206 항안드로겐 (anti an drog en ) 239 항우울제 (anti de p re ssants ) 348- 3 50, 356 항이뇨 호르몬 (an ti d i ure ti c hormone : ADH) 201 항정신병 약물 (an tipsy cho ti c drug) 341 항중력근 (an tigavity muscle) 185 항콜린효소 (anti ch oli ne ste r ase) 111 해마 (h ipp ocam p us) 78, 344 해 마교련 (hippocam p al commi ssu re) 308, 311 핵자기 공명촬영 (nuclear magn e tic resonance : NMR) 325 행동기형학 (beha vi oral ter ato l og y) 92 행동의학(be ha vi oral medi cine ) 168 헌팅턴 병 (Hunti ng t on 's dis e ase) 192 혈뇌 장벽 (blood-brain barrier ) 338 호기성과정 (aerobic pro cess) 177 혼합근 (m i xed muscle) 177 홍채 (iris) 114 화학적 기 울기 (chemic al grad i en t) 43, 58 화학적 손상법 (chemi ca l lesio n ) 25 화학적 시냅스 (che mi cal syn a ps es) 53, 54 화학적 친화력가설 (chemoa ffi n ity hyp othe sis ) 94 환각 (hallu ci na ti on) 336 활동전위 (acti on poten ti al) 32, 14-47, 59 활동전위의 전파(p ro p a g a ti on of the acti on poten ti al) 50 활차신경 (troc hlear nerve) 65 황체 형성호르몬·방출호르몬(l u teini z ing hormone releasin g hormone : LHRH) 55 회(gyrus) 74회 백 질 (gray matt er ) 36, 66, 72 횡 단단백 질 (tra nsmembrane pro te in ) 37, 38 효능물질 (ag on is t) 57, 58 후각상피 (olfa c to r y epi thel iu m ) 160 후교련 (po ste r io r commi ss ure) 81, 308, 311 후구신경 (olfa c to r y nerve) 65 후근 (dorsal root) 16, 65, 66 후근철 (dorsal root gan g lion ) 65, 66, 73 후두엽 (occ ipital lobe) 75, 140 후행성 건망증 (re t ro gr ade amnesia ) 298 흉부신경 (tho raci c nerve) 65 흉선 (thymu s gla nd) 72 흑질 (substa n ti a nigra) 191, 218, 343 흥분성 시냅스후 전위 (exc itor y pos ts yna pt ic poten ti al : EPSP) 59, 6o, 290, 292 히스타민 (his t am i ne ) 55 히 스테 리 성 신 경 증 (hy st e r ic al neurosis) 334
張鉉甲 서울대학교 심리학과 및 동대학원 졸업(문학박사), 가톨릭의대 생리학교실 및 서울의대 약리학교실 연구원, 서 울대 학교 심 리 학과 교수 및 New York Sta te Insti tut e for Basic Research 객원교수 역임. 현재 한국심리학회 생물· 생리십리학회 회장, 한국신경생물학회 부회장, 영남대학교 심리학과 교수. 저서 격리성장과 행동장애' 의 3 권, 역서 ‘생리심리학’ 의 3 권, 논문으로 Consequ ences of ablati ng the mammi llar y bod ies in dog s” 의 50 여 편이 있음. 생물심리학 대우학술총서 • 인문사회과학 82 1 판 1 쇄 찍음 1995 년 3 월 1 일 1 판 1 쇄 펴냄 1995 년 3 월 5 일 지은이 장현감 펴낸이 朴孟浩 편집 당연증 전산조판 동국전산 펴낸곳 (주)민음사 출판등록 1991 년 12 월 20 일 제 16-490 호 135-120 서울시 강남구 신사동 506 강남출판문화센터 5 층 대표전화 515-2000 팩시밀리 515-2007 값 12,500 원 ©장현갑, 1995 철학 • 십리학 /KOC 185 Prin t e d in Seoul, Korea ISBN 89-374-3082-7 94180 ISBN 89-374-3000-2 (세트) * 인지를 불이지 않기로 지은이와 합의하였습니다.
대우학술총서(인문사회과학) 1 한국어의 계통 검방한 44 스페인 문학사 김현창 2 문학사회학 김현 45 영어사 김석산 3 商周史 윤내현 46 어원론 김방한 4 인간의 지능 황정규 47 조선의 서학사 강재언 5 中國古代文學史 김학주 48 한국음악학 아강숙 6 日本의 萬葉集 김사엽 49 中國言語學 문선규 7 現代意味論 이악환 50 스포츠 심리학 8 베트남史 유인선 류정무·이강헌 9 印度哲學史 길희성 51 原始儒敎 김승혜 10 韓國의 風水思想 최창조 52 전쟁론 김홍철 11 사회과학과 수학 이승 훈 外 53 삼국시대의 漠字音 12 重商主義 김광수 유창균 13 方言學 이익섭 54 베르고송의 철학 김형효 14 構造主義 소두영 55 아날학파 김응종 15 외교제도사 김홍철 56 자유주의의 원리와 역사 16 아동심리학 최경숙 노명식 17 언어심리학 조명한 57 국민투표 구병삭 外 18 法社會學 양건 58 한국서지학 천혜봉 19 海洋法 박준호·유병화 59 조선통신사 이원식 20 韓國의 庭園 정동오 60 明末淸初思想 배영동 21 현대도시론 강대기 61 한국의場市 이재하홍순완 22 이슬람 사상사 검정위 62 고려시대의 后妃 정용숙 23 동북아시아의 岩刻畵 황용훈 63 포스트모더니즘의 이론 24 자연법사상 박은정 김욱동 25 洪大容 評傳 김태준 64 韓國의 服飾美 김영자 26 역사주의 아민호 65 정신신경증 이현수
