
박선주 연세대학교사학과 졸업 동 대학 석사, 박사과정 미국 캘리포니아 주립 버클리 대학교 인류학과 박사과정(체질인류학 전공) 현재 충북대학교 고고미술사학과 교수 역서 『 인류의 기원과 전화 』 (1(})2) 논문 「인류기원에 관한 연구 —오스트랄로피데쿠스에 대한 연구를 중심으로 」 「유라시아대륙의 중기흥적세 호미니드화석」 「현생인류기원에 관한 연구」 「한국민족의 체질연구」 등 20 여 편
체질인류학
 체질인류학
체질인류학
두희 그리고 사라, 노플에게 
책 머리에 오늘날의 세상은 옛날 사람들이 생각했던 세상처럼 단순하지가 않 다. 그러나 불과 2, 3 백 년 전까지만 해도 사람들은 자기들이 살고 있 는 세계를 두루 볼 수 있었으며 자기들이 본 현상들을 거의 다 이해 할 수 있다고 믿었다. 아주 단순한 사고를 했던 시대였지만, 이런 사 고의 세계가 계속될 수는 없었다. 사람들은 점차 지구상의 동식물들 이 어디서 시작했으며, 어떻게 과거로부터 현재까지 진행되어왔는지 를 묻기 시작하였다. 특히 지구상에서 가장 복잡한 존재인 인간 자신 에 대해 묻기 시작하였으니, 우리는 언제, 어디서 왔는가? 우리들의 일상생활에서 보이는 행위들은 어떻게 해서 일어나는가? 등등이 그것 이었다. 이런 의문에 대한 실마리는 인류학이란 학문을 통해서 얻을 수 있 다고 생각된다. 인류학 An t hro p olo gy이란 뜻은 고대 희랍어인 anth -rop u s(=man : 사람)와 log ia( =stu d y of : 학)라는 두 개의 단어로 이 루어졌는데, l@l 기를 전후하여 유럽 이의의 세계를 여행한 유럽 사 람들이 유럽 이의의 다른 지역에서 본 사람들의 의모라든지, 그들의 행위 및 그들이 일상생활에서 사용하는 기술들이 자신들과 다름을 알 게 되고 이를 서술하면서 인류학이란 용어가 처음으로 사용되었다. 인류학의 한 분야인 체질인류학 Ph y s i cal An t hrop olo gy은 과거에서 부터 현재에 이르기까지 인류를 포함한 영장류에 대한 생물학상 특칭 을 연구하는 것으로서, 진화 Evolu ti on 를 바탕으로 인류를 연구의 주 대상으로 삼아 인류가 어떻게 변화해 왔는가를 분자단계에서 종의 단
계까지 연구하는 학문이다. 따라서 이 학문은 독자로 종합된 방법론 multid isc ip li n a ry me t hod 울 가지고 있다. 체질인류학에 관한 많은 자료와 연구업적은 이미 l811 기의 유능한 해부학자였던 에드워드 타이슨 같은 학자에 의해 이루어졌는데, 그는 이때에 이미 사람의 몸과 유인원의 몸을 해부하여 직접 비교함으로써 이 분야의 선구자로 알려져 있다 . 그러나 현대 체질인류학의 직접 출 발은 찰스 다윈의 『종의 기원 (The Orig in of Sp ec ie s by Means of Na- tura l Selecti on , 1859) 』으로부터 시작된다. 진화에 관한 다윈의 업적은 그동안 강조되었던 여러 원리와 견해들을 괄목할 만하게 변화시켰 다. 다윈 이전이나 이후에도 오랫동안 체질인류학자들의 연구는 인류 를 〈인종적인 특칭〉에 따라 분류하는 데 모아졌다. 그러나 과거에 인류에 관한 체질 연구는 철저히 유럽 우월주의 입 장에서 진행되었으며, 이에 편승하여 유럽의 식민주의는 인종의 형태 에 따른 편견을 부추기며 이를 해의로 전파시키는 데 크게 기여하였 다. 유럽 사람들은 다른 종족을 연구할 때는 〈과학〉이란 말로 치장을 하였으며, 이들을 유럽 사람들에 비해 열등하다고 단정하였다. 지난 반세기 동안에 발전한 유전학과 고동물학 및 동물행태학 등의 발달과 사회가치에 대한 변화 등에 힘입어 체질인류학자들의 인류의 다양성 에 관한 이해가 급격히 증가하였다. 그 결과 오늘날 체질인류학자들 은 현재 살아 있거나 또는 지금은 절멸된 인류를 연구함으로써 인류 가 환경에 어떻게 적응을 하며 전화해 왔고 또 전화해 갈 것인가를 연구한다. 이 책은 그 학문의 중요성에 비해 학계에 극히 생소한 학문인 체질 인류학을 대학원생과 학부생들에게 이해시키고자 하는 목적에서 쓰여 졌다. 내용은 지금까지 진행된 체질인류학의 연구 성과를 바탕으로 인류의 진화 Human Evolu ti on 를 작은 전화 M i croevolu ti on 와 큰 진 화 Macroevolu ti on 로 나누어 다룬다. 영 장류학 Prim ato l og y 또는 영
장류 진화 Prim ate evolu ti on 와 고인류학 Paleoanth r op o log y, 그리 고 인류의 적 응성 Human ada pt ab ility과 사회 생 물학 So ci ob i olo gy도 체 질인류학에서 다루는 중요 연구분야이며 지질학, 전화생물학, 고생물 학 그리고 비교해부학의 연구 성과가 뒷받침된다. 이 책은 위에서 언급한 영역들을 다시 세분하여 모두 IO 장으로 구 성되었다. 내용을 효과적으로 전달하기 위해 많은 그립과 사진을 넣 었으며, 낱말들은 가능한 한 우리말 사용을 원칙으로 하였다. 이렇게 인간연구의 종합학문인 체질인류학을 의국 대학의 경우에는 위의 내용을 포괄하는 〈체질인류학개론 In t roduc ti on to Phy si c a l An- t hro p olo gy〉이란 과목으로 전 학년에게 교양과목으로 개설하여 학생 둘로 하여금 인간이 무엇이며, 인류가 지구상의 생명체에서 차지하는 위치를 살펴보게 하고 각 분야에 관한 과목을- 대학원까지 설강하고 있다. 현대 과학문명의 발달은 기존의 도덕관이나 윤리관으로는 감당 해 낼 수 없는 새로운 가치가 요구되는 그런 결과를 우리에게 가져다 주고 있다. 실제로 많은 사람들은 생명과학의 연구 성과를 단지 상업 적으로만 인식하고 인간이 과학을 통제할 수 있다고 생각하는데, 체 질인류학은 이런 오만함에서 비롯되는 종교, 도덕, 윤리상 문제(예 : 인류의 기원 문제, 낙태, 정신박약아, 대리모, 유전적 질병 소유자들의 사회수용, 인간의 이기심과 공격성) 등을 깊이 생각케 함으로써 인간 의 존엄성을 다시 한 번 되씹어볼 수 있게 도와준다. 필자가 이 분야의 공부를 하던 미국에는 대학의 수가 2500 여 개가 넘는데, 이 모든 대학에 〈체질인류학개론〉 과목이 개설되어 있다. 이 에 비해 우리나라는 4년 제 대학의 수가 거의 200 여 개에 육박함에도 소위 인류학과란 이름이 붙어 있는 과들은(인류학과나 고고인류학과 등등) 서너 개밖에 되지 않으며, 그나마도 모두가 문화인류학과를 줄 여서 쓰고 있다고 하겠다. 〈체질인류학개론〉 과목이 전공과목으로 개 설되어 있는 학교는 겨우 서너 개 대학뿐이며, 전 학년을 상대로 개
설된 대학은 거의 없는 실정과 십지어 관련학과 대학원에서도 일부의 내용만을 거론하고 있는 현실을 어떻게 설명해야 할지. 앞에서 언급했듯이 인류는 이 지구상에서 가장 복잡한 존재로, 정 말로 인류 자신의 행위를 이해하고자 한다면 인류학의 여러 분야 연 구가 이에 대한 답을 줄 수 있다고 하겠다. 인류는 주어진 환경에 적 응하며 살아가는 능력이 있음을 알 수 있으나 과학문명의 발달은 과 거의 인류가 경험하지 못했던 많은 문제들을 우리에게 제기하고 있 다. 이렇게 복잡한 문제를 안고 있는 인류를 바로 이해하기 위해서는 인류의 연구를 바탕으로 하는 학문, 특히 생물학적 입장에서 연구한 체질인류학의 내용들이 중 • 고등학교뿐만 아니라 모든 대학에서 학생 들에게 소개되고, 대학원에서 전문 연구가 이루어져야 한다. 체질인류 학 감은 자연과학과 인문과학을 연결해서 다루는 과목들이 대학에서 개설되어 학생들을 교육할 때 오늘날 우리의 대학이 참 인간보다는 물질만능을 추구하는 비인간적인 지식인과 기능인만을 양성해 내고 있다는 비난을 면할 수 있다. 이 책이 그런 기대에 작으나마 보뎀이 되기를바라는바이다. 이 책은 여러분들의 도움으로 쓰여졌다. 체질인류학이 무엇인지도 모르는 시절에 이 분야롤 공부하도록 길을 인도해 주신 은사 손보기 교수님, 그리고 체질인류학이란 학문을 일깨워주신 미국 주립 캘리포 니아 버클리 대학교 인류학과의 은사인 F. C. Howell 교수와 T. W 血 e 교수는 내가 잊지 못할 분들이다. 갇이 공부하던 버클리 대학 교 인류연구소의 Ri ch ard Gar y를 비롯한 여러 친구들 또한 나에게 이 책을 쓸 용기를 주었다. 감사의 마음을 전한다. 또한 이 책을 끝내기까지 도와준 여러분들, 특히 원고 타이평부터 교정에 이르기까지 애를 쓴 이은경(충북대 고고미술사학과 대학원생) 님, 그 많은 그립과 사전작업을 해준 조재경, 김윤정(충북대학교 고고 미술사학과 대학원생)님, 뒤에서 격려를 해주신 학과 교수님들, 그리
고 뒤에서 사랑과 희생으로 감싸준 아내에게 감사의 마음을 전 한다. 원고를 읽고 많은 조언과 가르침을 주신 손보기 교수님(단국대)과 최정필 교수님(세종대), 책을 낼 수 있도록 도와주신 서울대학교 인 류학과 전경수 교수님과 대우재단의 여러분께 감사를 드린다. 끝으로 이 책에서는 가능한 한 모든 낱말을 우리말로 쓰고자 노력 하였다. 그 결과 이 책을 읽는 도중 생소하게 느껴지는 낱말들을 자 주 대하게 될 것이다. 더욱 좋은 낱말들을 찾을 수 있게 되기를 바라 며 내용에서 있는 모든 찰못은 저자에게 있음을 말씀드립니다. 청주 개신벌 연구실에서
체질인류학
차례 책머리에 • 5 제 1 장 체질인류학의 전망 17 제 2 장 진화란 무엇인가 33 제 3 장 생명의 바탕 : 유전과 진화 59 제 4 장 영장류 119 제 5 장 뼈대와 이빨 183 제 6 장 인류조상의 발자취 245 제 7 장 인류의 진화 1 : 오스트랄로피테쿠스 265 제 8 장 인류의 진화 1I : 호모Homo의 출현 327 제 9 장 인류의 진화 III: 슬기사람 Home sapiens-393제 10 장 인류의 적응과 사회생물학 433부록 이 책에서 인용된 중요 외래 낱말들 • 465 찾아보기 • 483자세한 차례
책머리에 ·5 제 1 장 체질인류학의 전망 17 인류학의 정의 19 인류학의 여러 영역들 22 체질인류학 23문화인류학 24 고고학 또는 고고인류학 26 체질인류학의 발달의 역사 배경 28 제 2 장 진화란 무엇인가 33 진화론안내 35 전화에 관한 연구배경 36 진화론이전의 시기 36 다윈의 시대 : 찰스 다윈 44 다윈 이후의 시기 : 현대 진화론의 성립 49 창조론과 진화론 51 제 3 장 생명의 바탕 : 유전과 전화 59 인류유전학 : 개인단위에서 일어나는 변화 61 세포와 분자단위 61유전자의 구조와 특성 63
유전자 재결합 71 유전자란 무엇인가 77 집단유전학 : 집단의 유전과 전화의 요인 79 집단유전 79 진화의 요인 81 자연선택의 예 84 자연선택의 또 다른 예 : 어떻게 이해하나 85 큰진화 95 분류학 : 린네의 분류방법 97 큰 진화에 쓰이는 용어와 개념들 101 진화의 유형 104 진화에 관한 지질학의 증거들 108 시간의 측정 : 상대연대측정과 절대연대측정 110 전화계통 그림의 복원 113 비교해부학 114 분자생물학 115 고동물학 117 제 4 장 영장류 119 영장류란 무엇인가 121 영장류의 해부학상 생리학상 특징의 기원 124 영장류의 행위 128 영장류의 몸집과 식성 및 걷기 방법 130 영장류 무리의 사회구조 132 영장류의 분류 136 프로시미안 영장류들 137 엔스로포이드 영장류 151 영장류의 차이점 168 유인원에 관한 일반 이해 176제 5 장 뼈대와 이빨 183
뼈와 이빨 185 뼈의 얼개, 기능 그리고 성질 185 이빨의 얼개와 기능 189 사람의 뼈대 193 기본되는 해부학상 용어 194 뼈대의 이름 198 사람뼈 재기 203 머리뼈 재기 207 몸체뼈 재기 215 남녀 가늠하기 221 나이 찾기 227 키 재기 229 이빨 재기 231 다양한 이빨 형태 234 어금니 도드리 형태 234 인종 나누기 239 제 6 장 인류조상의 발자취 245 이론시기의 유인원들 247 라마피데쿠스:인류의 직접조상인가? 254 분자생물학의 연구 260 침팬지와 인간의 차이 263 제 7 장 인류의 진화 I : 오스트랄로피테쿠스— 265 인류의 기원 : 플라이오/풀라이스토세 호미니드 267 가장 이른시기 인류의 조상:오스트랄로피데쿠스아파렌시스의 발견? 270
하다 : 〈 루시 〉와 가장 이른 호미니드 화석의 발견 273 라에톨리 : 가장 이른시기의 발자국 277 남아프리카 호미니드의 발견 284 타웅 어린아이 화석(오스트랄로피테쿠스 아프리카누스 : 레이몬드 다트의 발견 286 스테르크폰타인 , 크롬드라이,스와르트크란스와 마카판스가트 : 오스트랄로피테쿠스 아프리카누스와오스트랄로피테쿠스 로보스투스 288 오스트랄로피 테쿠스는 사냥꾼인가? 290 동아프리카 호미니드의 발견 295 올드바이 골작 : 오스트랄로피테쿠스 보이세이 296 투르카나 호수 서편 : 오스트랄로피테쿠스 에티오피쿠스 304 인류 기원에 관한 문제들 307 인류의 특징 :두 발 걷기의 시작 312 두 발 걷기에 관한 다윈의 시나리오 314 나머지 여러 가설들 315 호미노이드의 생리상 • 해부학상 특징 317 러브조이의 Attractio n Provisioning 모델 322 로드만 맥켄리의 모델 323 제 8 장 인류의 진화 II : 호모 Home의 출현 327 손쓴사람 Homo habilis의 출현 329 손쓴사람 화석의 발견 329 이른시기 호미니드의 문화와 행위 338 곧선사람 (Homo erectus:立猿人)의 출현 -348 곧선사람의 발견 351 곧선사람의 해부학상 특징 358 곧선사람 화석의 분류의 진화 363곧선사람의 기원지와 이동 366
곧선사람의 문화와 행위 371 인류의 두뇌 : 그 얼개와 기능 373 네안데르탈사람 Homo neanderthalensis의 출현 - 377 최초의 네안데르탈사람 379 네안데르탈사람의 기원과 인류진화상 위치 384 네안데르탈사람의 문화와 행위 387 제 9 장 인류의 진화 III: 슬기사람 Home sapiens一 393 최초의 슬기사람 : 크로마뇽 사람의 발견 395 슬기사람 기원과 발달에 관한 최근의 연구들?- 399 미토콘드리아 핵산 : 유전학자들의 연구 405 고고학상 • 고인류학상의 새로운 증거들 409 후기 구석기문화 421 연모의 특징과 발달 421 후기 구석기사람의 행위 423 유럽의 후기 구석기문화들 424 제 10 장 인류의 적응과 사회생물학 433 인류 : 어떻게 환경에 적응하나? 435 추위에 의한 갈등 437 더위에 의한 갈등 439 고산지대에서 오는 갈등 445 남녀의 성에 따른 차이 449 사회생물학의 소개 453 생물의 집단에서 관찰되는 행위들 454 사회생물학 연구의 문제와 전망 460 체질인류학의 오늘과 내일 463 부록 : 이 책에서 인용된 중요 외래 낱말들 465 찾아보기 483 제 1 장 체질인류학의 전망
제 1 장 체질인류학의 전망
제 1 장 체질인류학의 전망 인류학의 정의 오늘날의 세계는 옛날 사람들이 생각했던 세계처럼 단순하지 않다. 2, 3 백년 전까지 유럽 사람들은 그들이 살고 있는 세계를 쉽게 둘러 볼 수 있었으며, 자기들이 본 세계는 거의 다 이해할 수 있다고 믿었 다. 그들은 당시 주위에 있던 동물과 식물들이 과거에도 있었고, 미래 에도 변하지 않을 것으로 생각하였다. 유럽 사람은 가장 키가 크고 세상의 모든 법칙을 만들었으며, 여자들은 단지 뒤에서 남자들을· 돕 고 아이를 키우며 언제나 더운 음식을 만드는 그런 존재일 뿐이라고 믿었다. 아주 단순한 사고를 했던 세계였으나, 이런 사고가 계속될 수 는 없었다. 17 세기에 들어와 사람들은 지구상의 모든 것들이 왜 존재 하는지, 어떻게 과거로부터 현재까지 진행되어왔는지를 묻기 시작하 였다. 과학자들이 이런 유럽 사람들의 경직된 사고를 변화시키는 데 는 1 세기 이상의 시간이 걸렸다. 19 세 기 에 영국의 지질학자였던 찰스 라이 엘 Charles L y ell 은 지구의 모습은 천천히 지속해서 변화한다고 주장하였다. 찰스 다윈 Charles Dar wi n 과 알프레드 러셀 월리스Alf red Russel Wallace 는 생명체에 관한 진화이론을 세웠는데, 이 이론은 생명체에서 보이는 다양성과
자연환경과 시간에 따른 생명체의 변화에 대해 설명하고 있다. 생명 체에 대한 여러 연구 중 인간 자신에 대한 연구만큼 엄청난 변화가 있는 분야는 없다. 유럽 사회에 있어 l81] 기는 〈 발견의 시대 〉 였다. 많은 탐험가들이 유럽 이의의 세계를 탐험하고 돌아와서, 그때까지 유럽 사회가 경험 하지 못했던 다른 사회와 사람들에 관한 소식을 소개하였다. 그러나 유럽 사람들은 아직도 그들의 생활이 가장 뛰어난 것이며, 그들의 사 고방식과 사회조직이 가장 우수하다고 생각하였고, 유럽 사람들만이 조물주와 닮은 모습으로 창조되었다고 믿었다. 인간이 다른 동물들같 이 하등동물에서부터 전화하였다는 생각은 19 세기 사람들에게는 저주 받을 말로 받아들여졌다.
 巳;'\ .1I ;g! .: ::정;·8::· 〈·으}, i; ` ) ,_, \,• • •,— : :\.-. - , , \.i ,1hi
巳;'\ .1I ;g! .: ::정;·8::· 〈·으}, i; ` ) ,_, \,• • •,— : :\.-. - , , \.i ,1hi
여행자들로부터 이상한 나라에 살고 있는 사람들과 그들의 생활방 식을 전해 들은 유럽 사람들은 이를 단지 재미있는 이야기로만 받아 들였다. 아프리카와 아메리카 대륙에서 데려온 원주민들을 본 유럽 사람들은 비록 의모나 사회조직 및 종교관 등이 자기들과 비슷하지만 이들을 불완전한 존재로 인식하였다. 아메리칸 인디언에 관한 프랑스 자연사학자 조지 드 뷔퐁 Geor g es de Bu ff on 의 묘사는 당시 유럽 사 람들이 이들을 어떻게 생각했는지롤 보여준다 ; 〈 신대륙에서 온 야만 인들은 우리들과 키가 비슷하지만 이는 예의의 일이며, 신대륙에 살 아 있는 모든 생명체는 점점 작아지고 있다는 사실을 알아야 한다. 이 야만인들은 저능하며, 체모와 수염이 별로 없고, 여성에 대한 애정 도 없다. 이들은 달리기가 습관화되어 유럽인들에 비해서 몸이 유연 하고 탄력이 있지만, 반면에 몸이 약하고 감각이 별로 발달되어 있지 못하다• 〉 (사전 1. 1) 이러한 뷔풍의 설명은 아메리칸 인디언에 대한 부정확한 자료 때문이라고 할 수 있다. 그럼에도 불구하고 유럽 학자 들 중 일부는 유럽 이의 지역에 살고 있는 사람들의 생활에 대해 정 확한 지식을 갖고 있었다 . 18 - 19 세기에 들어오면서 유럽 이의의 세 계에 대한 지식이 크게 증가하였다. 오늘날의 인류학은 이 시기에 모 아진 연구자료를 바탕으로 출발하였다. 인류학 An t hro p olo gy이란 뜻은 고대 희랍어인 anth rop u s(=man : 사람)와 log ia( =stu dy of : 학)라는 두 개의 단어로 이루어졌는데 , 이 단어는 l@1] 기에 처음으로 사용되었다. 유럽 이의의 다른 세계를 여행했던 유럽 사람들은 이 세계에서 볼 수 있는 행위와 의모, 또는 그들이 사용하는 기술 등이 자신들과는 다르다는 것을 알게 되었다. 고대 희랍과 로마 사람들은 그들과 인접하고 있는 지역의 사람들의 습관과 생활태도 등에 대해 많은 기록을 남겨놓았다 . 후세의 유럽 사 람들처럼 그들도 이웃 문화와 그들의 의모를 자신들의 입장에서 늘 비교하였다.
로마의 역사학자인 타키두스 (Ta cit us, 55- 1 20 A. D. )는 최초로 인 류학 연구를 한 사람 중의 하나로 꼽히는데, 그도 당시의 도이치 지 방의 사람들을 뷔풍이 아메리칸 원주민을 묘사했던 것처럼, 〈 모두가 지저분하고 무기력하며, 추장들은 얼굴에 보기 흉한 것들을 뒤집어쓰 고 있다 . 〉라고 설명하고 있다. 초기에 희랍 • 로마 사람들은 다른 문화 들과의 차이를 〈 불완전한 표시 〉 로 생각하였다. 따라서 당시 유럽 사 람들이 가지고 있던 유럽 중심주의 내지 우월주의 Et hn ocentr i s m or Cultu ral Se lf -Cen t eredness 를 극복하는 데는 오랜 시간이 걸려야 했다. 인류학의 여러 영역들 우리는 오늘날 서로 다른 환경에서 살고 있는 인류들이 생존을 위 해서 여러 가지 수단과 방법을 사용하고 있음을 알고 있다. 인류학자 들은 인류문화와 생물의 특성 및 환경 사이에서 일어나는 상호간의 작용이 무엇인지롤 연구한다. 그들은 어떤 문화와 인종이 열등하고, 또 어떤 문화와 인종은 우월하다는 식의 내용에는 관심이 없다. 인류 가 살아 남는다는 것은 상대개념으로, 인류학자들은 이것으로 인류집 단이 어떻게 서로 다른 환경에서 생물학상 • 문화상으로 적응할 수 있 는지를측정한다. 인류학은 아주 넓은 범위를 연구대상으로 하고 있다. 이 같은 인류 학은 크게 세 영역으로 나누어지며, 비록 각 영역에서 연구하는 입장 이 조금씩은 다르지만 추구하는 내용은 서로 갇다. 이 영역들은 체질 인류학, 문화인류학 및 고고학으로 나누어전다. 각 영역의 연구는 한 집단이 환경에 적웅해 가면서 나타나는 특성이 다론 집단에서는 왜 나타나지 않는가에 집중된다. 인간은 지구상에서 가장 복잡한 생명체 라고 할 수 있다. 따라서 인간이 정말로 자신을 이해하기를 바란다면
인간행위를 이해해야 하며, 그런 점에서 인류학의 여러 영역들은 이 에 대한 답을 줄 수 있을 것이다. 체질인류학 체질인류학 Ph y s i cal An t hro p olo gy은 과거부터 현재에 이르기까지 인류를 포함한 영장류에 대한 생물학적인 특칭을 연구한다. 연구는 전화 Evolu ti on 롤 바탕으로 인간 자신이 주된 연구대상이며, 인류가 어떻게 변화해 왔는가를 분자단계에서 종단계까지 연구하는 학문으로 독자적으로 종합적인 방법론을 가지고 있다. 체질인류학은 그 연구대 상이 매우 광범위하기 때문에 크게 인류의 전화, 영장류학 또는 영장 류 진화, 고인류학, 인류의 적응성과 사회생물학의 분야로 나누어 연 구한다. 체질인류학의 한 분야인 인류의 전화에서는 인류가 짧은 시간 내에 서 이루어진 작은 전화 M i croevolu ti on 와 오랜 기간에 걸쳐 전행된 큰 진화 Macroevolu ti on 를 연구한다. 영장류들의 분자상의 구조를 비 교 • 연구하여 이들의 관계를 밝히며, 특히 인류를 세포와 분자 단위 에서 연구하는 인류유전학과 집단과 종의 단위에서 연구하여 유전 특 칭을 밝히는 집단유전학, 그리고 인종문제를 다루는 인류의 다양성 연구가 이 분야에 속하며, 이룰 위해서 지질학과 진화생물학의 도움 울받는다. 체질인류학의 또 다른 분야인 영장류학 또는 영장류 진화는 인간이 아니지만 우리와 가장 가까운 다른 영장류의 전화와 행위를 연구하 며, 또한 오늘날 현존하는 다양한 인간들과 인류의 생물학적인 환경 적응방법 등을 연구한다. 모든 인간은 왜 연모를 만들고 말을 할 수 있는지에 대한 답의 일부는 인간과 가장 가까운 영장류인 원숭이와 유인원 특히 침팬지에 대한 생물학상 특성과 행위를 살펴봉으로써 얻
어전다. 고인류학은 생물학상 방법으로 인류의 역사를 다루는 분야로서 호 미니드 화석들을 통해서 이들이 어떤 모양을 가지고 있었는지를 알고 자 한다 . 호미니드 화석들의 연구를 통해서 인류진화를 연구하는 고 인류학자들은 이들의 생물학상 특칭과 행위와의 관계를 밝히고자 하 며 과거 인류의 행위를 복원하는 데 있어 고고학자들에게 도움을 주 고 비교해부학과 사람뼈대학 등의 도움을 받는다. 인류의 적응성 연구는, 오늘날 지구상에 살고 있는 사람들이 여러 면에서 서로 다른 모습, 다른 생활형태를 보이고 있는데, 그런 차이가 나타나는 이유와 또한 전화과정중 집단 사이에 나타나는 유전적 차이 와 환경에 따른 유전적 적응 이의의 방법 등을 연구대상으로 한다. 한편 최근에 체질인류학의 한 분야로 사회생물학을 들 수 있는데, 이 분야는 인간의 행위를 동물들의 행위와 비교하여 인류행위의 원인(예 : 인간의 공격성 등)을 규명하고자 한다. 문화인류학 초기의 문화인류학자 Cul t ural An t hro p olo gi s t s 둘은 대부분 〈원시사 회사람〉둘의 생활을 주요 연구대상으로 하였다. 19 세기 말까지 학자 들은 태평양, 아메리카, 오스트레일리아, 아프리카 및 아시아 등 여러 지역에 존재했던 비유럽 문화들이 유럽의 식민주의와 기술의 영향 아 래 소멸되어가는 것을 깨달았다. 이에 인류학자들은 이들의 문화와 언어 및 사회관계 등의 유산을 보존하기 위해서 구제발굴을 시작하였 다. 그 결과, 비록 많은 문화들이 사라져가기는 했지만 이들의 노력에 의해서 기록으로 남게 되었다. 사라져간 문화들을 연구하는 과정에서 우리는 한 문화가 붕괴됨에 따라 일어나는 질병이나 대학살, 그리고 여기에 수반되어 인류가 느끼는 무기력 등은 문화뿐만 아니라 그 주
변문화와 인간까지도 지구상에서 말살하는 결과를 가져왔다는 사실을 알게 되었다. 최근에 문화인류학자들은 현대 산업사회를 연구하기 위해 원시사회 연구에 사용되었던 현장방법을 이용한다. 이 가운데 중요한 방법은 실제로 현장에서 관찰하는 것으로, 어떤 학자는 주민들과 함께 그들 의 언어를 사용해 가며 생활함으로써 그들의 생활과 사고롤 이해하려 고 한다• 또한 문화인류학자들은 한 사회 안에서 어떤 형태의 행위가 가장 적절한가를 찾고자 한다. 어린이들은 사회화되어가는 과정에서 사회의 규범 등을 배운다. 어 른이 되어가면서 아이돌은 무의식중에 사회 내에서 다른 행위들도 배 우게 된다. 개인은 그 집단의 전통을 따르게 되며, 어느 신을 경배해 야 하는지를 알게 되고, 어떻게 배우는지를 선택하고, 언제 수확울 하 며 또 어떻게 사냥을 해야 하는지 등을 배운다. 이런 규범들을 배움 으로써 이들은 과거 그들이 살아온 환경에 찰 적응하였으며 다음 세 대에게 자신들의 문화를 전달하게 된다. 인류는 배움을 통해서 문화를 소유하게 되는데 이런 배움의 행위는 환경에 따라 변화한다. 이렇게 환경에 따라 변하는· 행위가 궁극적으 로 문화를 변화시킨다는 점에서 행위변화의 유형은 인류학자들에게 중요한 연구분야이다. 문화의 변화는 인간의 생물학상 자리에 커다란 영향을 준다. 예를 들어 농업경제에 기반을 둔 사회에서 갑작스럽게 경제적인 이유로 팔 아야 할 농산물인 목화를 먹는 경우가 있는데, 이는 건강에 해를 끼 친다. 이민 또한 이민자에게 새로운 질병과 음식, 그리고 새로운 짝을 선택할 수 있는 기회를 제공한다. 이와 반대로 어떤 집단의 생물학상 특칭이 문화에 영향을 미치기도 한다• 한 예로 굶주리는 집단의 사람 둘은 영양공급이 좋은 사람들처럼 행동할 수 없다. 또한 생물학상 변 화와 문화의 변화는 독립해서 일어나기도 한다. 인류집단의 문화에
변화가 생기지 않을 경우, 변하지 않는 문화는 환경에 어떤 영향을 미치고, 또한 인류집단과 여기에 속한 개인이 환경에 어떻게 대처하 는가를 분석함으로써 문화인류학자는· 우리들로 하여금 인간 자신을 찰 이해할 수 있도록 도와준다. 고고학 또는 고고인류학 인류학의 또 다론 분야인 고고학 Archaeolo gy은 크게 선사고고학과 역사고고학으로 나누어지는데, 특히 우리가 관심을 갖는 분야는 선사 고고학이다. 선사고고학은 고고학자들의 연구를 과거까지 연장시켜 옛 사람들이 남겨놓은 문화활동을 연구한다. 지금은 소멸된 사회들이 당시의 환경과는 어떤 관계에 있었는지, 또 어떻게 한 문화가 오랫동 안 한 곳에서 지속될 수 있었는지를 연구한다. 그런데 불행히도 이 분야의 연구대상은 당시 문화기술을 보여주는 돌연모와 뼈연모, 토기, 부서전 동물뼈 및 불에 그울린 바위와 같은 완전하지 않은 자료이기 때문에 고고학자들은 이 불완전한 자료들을 통해 과거의 문화를 재현할 수 있게끔 정교한 분석방법 등을 개발해 왔다. 그러나 이런 과학 분석방법들만을 가지고는 당시의 사회조직이나 신앙, 또는 결혼관행 갇은 것들을 정확히 알 수 없다. 그 결과 때때로 고고학자들은 직관에 의존하기도 하며 또한 그들이 남긴 무덤의 흔적 과 불의 사용 및 다른 행위를 나타내주는 증거둘을 종합해서 결론을 내린다. 남겨진 유적 • 유물을 바탕으로 소멸된 문화를 복원하는 데 있어 고 고학자들은 문화인류학자와 체질인류학자들의 도움을 받는다. 오늘날 살고 있는 일부 집단들은 농사에 의해서가 아니라 채소나 과일, 곤충
이나 새알 등을 채집하거나 또는 사냥으로 식량을 얻는다. 약 10000 년 전 농업이 시작되기 전까지 모든 인류사회는 채집과 수렵경제에 의존했었다. 그래서 고고학자들이 조사하는 많은 유적들에서는 수렵/ 채집 인들이 남긴 유물들이 발견된다. 따라서 오늘날 살고 있는 수 렵/채집인들의 생활을 동해서 고고학자들은 선사문화를 이해하고 복 원할수 있다. 선사인류들이 남겨놓은 유물들을 연구하는 것 이의에 당시 사회의 복원은 현대인들의 생활을 이해하는 데 도움을 준다. 인류학자들은 문화발달의 내면을 살펴봄으로써 오늘날 사람들이 독특한 특성을 지 니게 된 이유를 알 수 있다. 또한 고고학 자료들을 통해서 현재 인류 사회에서 보여지는 이민형태나, 인구밀도, 질병 및 식량 획득형태 등 울 이해할 수 있다.
인류학의 여러 분야들* 체질인류학 문화인류학 고고학 인류의 진화 사회문화인류학 선사고고학 큰진화와작은진화 인류유전학 집단유전학 인류의 다양성(인종) 고인류학 (고고학) 역사고고학 비교해부학 (언어학) 사람뼈대학 영장류학 인류의 적응성과 사회생물학 *위는 인류학의 일반적인 분류로 학자들에 따라 고고학을 문화인류학의 범주로 분류하기도 하며 기타 분야를 다르게 분류한다.
체질인류학 발달의 역사 배경과 전망 체질인류학에 관한 많은 지식들은 le1) 기에 얻어졌는데, 이는 에드 워드 타이슨 (Edward Ty so n, 1650- 1 708 A.D. )이라는 유능한 해부학자 가 사람과 유인원을 직접 해부하여 서로 비교 • 연구함으로써 이루어졌 으나 현대 체질인류학의 전정한 출발은 찰스 다윈의 『종의 기원』으로 부터 시작된다. 다윈 이전이나 이후 오랫동안, 체잘인류학자들은 사람 들을 〈인종의 특칭〉에 따라 분류하였다. 체질인류학자들은 어째서 유럽 인은 대개 키가 큰가, 대다수의 아시아인은 왜 마리가 큰가, 사하라 이남지역의 사람들의 피부색은 어떠한가 등을 조사하였다. 화석증거 들도 이와 같은 방법으로 분류되었다. 중국에서 발견된 화석을 자바 에서 발견된 화석과 비교할 때, 만일 시기는 같아도 모양이 서로 다 르면 두 지역에서 나온 화석들은 서로 다른 종으로 구분되었다. 행위 또한 편협하게 해석되었다. 사하라 이남의 주민들은 게으르고, 유태인 은 탐욕스러우며, 아시아인들은 이해할 수 없는 존재로 생각되었다. 그러나 북유럽 사람들에 대한 연구는 별로 없었다. 이때 어떤 한 집 단 내에서 나타나는 다양성은 무시한 채, 집단과 집단 사이에서 보이 는 차이들은 모두 중요시하였다. 유럽의 식민주의는 이런 편협된 견 해롤 부추기면서 이를 해의로 전파하는 데 크게 공헌하였다. 유럽 이 의 지역에 사는 사람들을 연구할 때는 〈과학〉이란 말로 치장하였으 며, 이들을 유럽 사람들보다는 열등하다고 단정하였다. 따라서 유럽 이의의 사람들을 변하지 않는 존재로 단정하는 것은 쉬운 일이었다. 지난 수십 년 사이에 유전학과 고동물학, 동물형태학의 발달과 사 회가치의 변화에 힘입어 과거에 살았고 또 현재에 살고 있는 인류가 다양하다는 사실을 크게 깨닫기 시작하였다. 오늘날 우리는 인류가 매우 다양하다는 사실을 알고 있으며, 학자들은 더 이상 인종 전형성 에 대한 연구는 하지 않는다.
오늘날 체질인류학지들은 실제로 야의에서 사람들을 대상으로 연구 한다. 지금 살아 있거나 아니면 철멸된 사람들 간에, 한 집단 내에서 보이는 변이와 집단 사이에서 나타나는 변이는 모두 홍미있는 연구대 상이다. 인류가 어떻게 진화해 왔으며, 오늘날의 인류는 어떻게 변하 고 있는지는 이들 변이룰 연구함으로써 답을 얻을 수가 있다. 전화는 환경에 적응하는 방향으로 전행된다. 다시 말해서, 사냥을 해서 음식을 쉽게 얻을 수 있으며, 기후나 맹수, 질병 또는 인류가 생 존할 수 있는 능력에 영향을 미치는 모든 종류의 요인으로부터 벗어 났을 때, 진화는 한 방향으로 진행되어간다. 진화라는 관점에서 볼 때, 인류에게서 나타나는 다양성이란 바로 늘 변화하는 환경에 적응 해서 살아 남을 수 있는 능력을 의미한다• 만일 인류에게 다양성이 없다면 인류는 그들이 살고 있는 환경의 변화에 적응할 수 없다. 인류가 환경의 변화에 적응하는 방법은 동물들의 방법과 비슷하다. 인류는 진화를 통해서 생물학상으로 적응하며, 행위를 변화시켜 문화 상으로 적응할 수 있다. 이 두 가지 적응방법은 서로가 밀집한 관계 가 있으며, 인간의 생물학상 적응은 바로 문화상 적응력에 크게 영향 을 미친다. 400 만 년 전에 살았던 인류의 가장 먼 조상은 불을 조절 하거나, 죽은 사람을 묻거나, 또는 서로 의사를 전달할 수 있는 생물 학상 능력이 없었다. 반대로 기술이 고도로 발달하고 도시환경 또한 급속히 발달해 가는 오늘날에 살고 있는 · 인류는 적응이라는 면에서 볼 때 생물학상으로는 별로 발달된 형태가 아니다. 이런 점에서 인류 의 생물학상 특칭과 문화와의 관계는 매우 복잡하며, 체질인류학은 인류가 가지고 있는 지금의 문제들과 연관을 가지고 있다고 하겠다. 최근까지 서양사회는 인간이 자연을 지배할 수 있고 또 하여야 한 다고 믿었다. 인간은 신의 바로 밀에 있으며, 세상과 세상 안에 있는 모든 일들은 신에 의해서 개척되었다고 생각하였다. 오늘날 우리는 서양 사람들 이의의 인류가 오래 전부터 존재했었다는 사실을 안다.
또한 인간은 자연의 일부이자 불가분의 관계에 있는 존재라는 것을 인식하며, 인간은 환경의 영향 아래에서 살아 남는 방법을 배워야 한 다는 사실울 안다. 주어진 환경에 문화상으로 적응한다는 것은 인간 의 생물 특징과 관계가 있다는 사실을 우리는 알고 있다. 자원은 무한하지 않으며, 인간의 몸 또한 완전하게 환경에 적응할 수 없다. 만일 인간에게 참고 견디는 능력이 있다면, 인간의 생물학상 속성과 행위는 서로 상호보충하여 완전하게 되어야 한다. 어떤 학자 들은 인간이 조상으로부터 물려받은 생물학상 속성에서 볼 때, 인간 은 작은 집단으로 나누어 살아가는 삶이 가장 이상형이라고 한다. 만 일 수평방 미터 내에서 많은 수의 사람이 함께 살면 인간은 스스로를 제어하는 능력을 잃게 되고 그 결과 도시에서는 공포와 불안, 그리고 폭력이 증가하는데 이는 바로 생물학상으로 적응할 수 없는 생활여건 때문이라고주장한다. 인류학자들이 알고자 하는 중요한 의문 중의 하나는 전쟁에 관한 인간의 성향에 관한 것으로, 인간의 생물학상 속성과 행위 사이에는 어떤 관계가 있는가에 있다. 전쟁이란 인간이 살아오면서 문화에 의 해서 습득되는 행위일까? 아니면 인간은 본래 동물의 공격적 성향을 가지고 전화해 왔는가? 인간이 만들어내는 가공할 만한 무기는 단지 커다란 이빨과 간 발톱을대신한 문화상 만들어전 대용품인가? 우리는 미래에 올지도 모를 대학살을 미리 예방하기 위해서 체질인류학자들 의 연구대상인 인간에게서 보이는 공격성향의 원인에 대해서 더 많은 연구를 해야만 한다. 인간의 공격성은 과연 유전으로 전달되어 전화 하는가? 아니면 인간이 사회화되어가는 과정에서 배워지는 행위일까? 인간의 두뇌는 엄청난 양의 변화를 수용한다. 따라서 체질인류학자는 이런 변화가 일어날 수 있는 가능성의 한계가 어디까지인가를· 찾아야 한다. 이런 질문들은 실제 문제이며 동시에 학문상의 관심사가 된다.
호미니드 Hom i n i ds 와 호미노이드 Ho mi no i ds : 호미니드는 인 류과 Ho min i dae 의 구성원들을 뜻한다. 인류과는 영장류목에 속 하는 분류 단위로 현재 살아 있는 술기사람과 그들의 절멸한 조상을 모두 포함한다. 호미노이드는 영장류목의 분류 단위인 초인류과 Ho mi no i dea 에 속하는 구성원을 뜻하며 유인원과 절 멸된 선유인원들 및 인류를 모두 포함한다.
 제 2 장 전화란 무엇인가?
제 2 장 전화란 무엇인가?
제 2 장 전화란 무엇인가 진화론안내 전화라는 개념은 생물체를 연구하는 모든 과학의 중심 주제로, 이 개념만큼 인간과 그들이 몸담고 있는 세계롤 바탕짓는 생각도 없다. 살아 있는 모든 생명체에 작용하는 전화는 아직도 완전히 증명되지 않은 하나의 가설이라고 주장하는 학자들도 있다. 그러나 우리는 전 화가 가설t heor y이 아닌 사실fa c t이라는 것을 증명할 수 있는 많은 증거둘을 가지고 있다. 오늘날 살아 있거나, 죽었거나, 또는 사라졌거 나 모든 동물과 식물은 서로 관련이 있으며, 이들은 어떤 최초의 생 명체로부터 시작되었다. 모든 동물과 식물이 서로 관련이 있다는 사실은 불과 100 여 년 전 찰스 다윈과 알프레드 러셀 월리스와 연구에 의해 알려지기 시작했 다. 다윈과 월리스 이전의 자연학자와 철학자들~ 동물과 식물들의 모습은 세월이 지남에 따라 부분적으로 변화된다고 믿었다. 그러나 그들은 동물과 식물에게 변화를 가져오는 요인이 무엇인지를 알지 못 했다. 많은 위대한 생각이 그랬던 것처럼 자연선택에 의해 전화가 일 어난다는 다윈과 월리스의 생각은 그 내용이 복잡하지 않고 간단하 다. 그러나 다윈과 월리스는 자연선택이 진화의 요인이라는 사실은
알았지만 개체변이가 일어나는 원인과 부모의 특칭이 자식에게서 나 타나는 이유는 알지 못했다. 지난 반세기 동안 유전학지들은- 다윈과 월리스가 관심을 가졌던 생 명체를 〈 종〉과 〈 개인 〉 그리고 〈 세포 〉 와 〈 분자 〉 구조까지 연구하여 개 인의 유전적 특칭이 다음 세대에 어떻게 전달되는지를 밝히고 있다. 이돌을 연구하는 전화생물학은 인류전화에 관한 기본틀을 제시하여 준다. 진화에 관한 연구 배경 체질인류학은 인류전화의 원인과 과정, 그리고 전화의 산물인 호미 니드를 주된 연구대상으로 한다. 이를 이해하기 위해서는 인류전화에 관한 연구 배경을 먼저 살펴보아야 한다 . 인류전화에 관한 여러 주장 은 진화론 이전의 시기, 다윈의 시대 그리고 다윈의 시대 이후로 크 게 나누어볼 수 있다. 여기서 전화론 이전의 시기는 다시 계몽주의 이전의 시기(이 시기는 다시 〈주대오-크리스천 전통의 시기 Ju deo-Chris t i an Trad ition > 및 〈중세 • 르네상스 시기〉로 나누어질 수 있다) 와 18-19 세기로 나누어볼 수 있다. 진화론 이전의 시기 계몽주의 이전의 시기 계몽주의 이전의 시기 중 주대오-크리스천 전통의 시기를 지배하 던 관념은 우주와 지구상의 모든 생명체는 오직 하느님 한 분에 의해 창조되었다는 성경의 창세기장에 바탕을 두고 있다. 죽 성경의 창세 기장은 생명의 기원과 존재에 대해서 언급하고 있으며, 또 하느님의
 2. 1 아르마의 대주교 제임스 어셔
2. 1 아르마의 대주교 제임스 어셔
형상을 닮은 인류가 어떻게 시작되었는지를 설명하고 있다. 지구상의 모든 생명체들은 오늘날의 모습대로 태초에 만들어졌으며 〈대홍수〉만 이 이들을 변화시켰다고 주장한다. 성경의 이러한 내용은 유럽 중세 에 이르기까지 절대적인 전실이었다. 17 세기에 들어와서, 비록 성경에 근거하지만, 지구의 생성과 나이에 대해 과학적으로 생각하고자 하는 사람들이 나타난다. 아르마의 대주교인 제임스 어셔(J ames Ussher, 1581-1656 A.D. )는 성경의 구약에 근거하여 지구가 기원전 4004년 에 생겨났다고 주장하였다(사전 2. 1). 그의 계산은 영국 캠브리지 소 재 성 캐더린 대학 학장인 존 라이트푸트J ohn L ig h tfo o t에 의해 더 욱 정밀하게 계산되었다. 그는 기원전 4004 년 10 월 23 일 오전 9 시에 지구가 형성되었다고 발표하였다. 이때까지 유럽인들의 세계관은 유 럽 중십이었다.
한편 르네상스 시기에 오면서 이 시기를 전후해서 유럽 사람들은 세계 여러 곳을 탐험하여 많은 새로운 지식을 얻게 되었다. 콜럼버스 의 아메리카 대륙 발견은 당시 유럽 사회의 세계관에 커다란 충격을 불러일으켰다. 유럽인들은 아메리칸 인디언들을 불완전하게 유럽 사 람을 닮은 존재로 생각하였다. 이들을 통해서 유럽과는· 다른 형태의 사회조직과 종교, 인류사회가 존재한다는 사실을 알게 되었다. 무어인의 유럽 침공, 인쇄술과 총포의 발견은 유럽 사회에 커다란 변화를 가져왔으며, 사상의 발달과 코페르니쿠스 등의 연구에 의해 천체에 대한 이해가 확대되었다. 사람이 더 이상 우주의 중심이 아니 려, 유럽이 더 이상 평평한 모양을 하고 있는 지구의 중심이 아니라 는 중요한 〈인식의 변화〉가 일어나기 시작한 것이다. 18-19 세기 이후 이 시기에 이르러 유럽 사회에서는 자연의 법칙에 대한 관심이 싹 트고 있었으나 아직도 자연의 모든 현상은 조물주의 의도에 의한 것 이라고 믿었으며, 린네는 이 시기 자연계 연구의 대표적 인물이다. 한 편 유럽인들의 과학적 사고에 절대적인 변화를 가져온 대표적인 학자 는 린네 이의에 뷔풍과 에라스무스 다윈, 라마르크, 퀴비에, 라이엘 그리고 맬서스 등을 꼽을 수 있다. 린네 린네 (Carolus Lin n aeus, 1707-1778 A.D. )는 생물계를 생긴 모습에 따라 〈종〉의 단위로 분류하였으며, 동물과 식물의 분류를 통해서 조 물주의 의도가 무엇이고, 피조물들이 언제 만들어졌는지롤 알고자 하 였다. 그는 동물과 식물들은 처음부터 지금과 같은 모습으로 만들어 졌으며 그 이후 변하지 않았다고 생각하였는데, 이런 그의 생각은 1758 년에 발표된 『자연체계 Sy st e m a Natu ra e = Sy st em of N atu re 』(사
전 2. 2) 에 잘 나타나 있다. 이는 아직도 린네와 당시 학자들이 인식 의 한계성을 넘어서지 못했음을 보여주는 것으로써, 이런 사고는 1700 년대 칸트와 버클리, 루소 등의 사상가들에 의한 사고의 혁명과 미국 및 프랑스의 혁명으로 야기된 사회변혁에 의해 변화되었다.
 CAROLI LINN.i EI ••2 ·뻬’
CAROLI LINN.i EI ••2 ·뻬’
 2. 3 프랑스의 박물학자
2. 3 프랑스의 박물학자
 2. 3 프랑스의 비교 해부학자
2. 3 프랑스의 비교 해부학자
뷔퐁 뷔퐁 (Gor g es de Bu ffon , 1707-1788 A.D. )은 프랑스의 유명한 동식 물학자이자 왕의 정원 책임자로, 그는 살아 있는 생명체와 환경은 절 대적인 관계를 가지고 있다고 생각했다. 뷔풍은 각기 다른 종들이 한 공통조상으로부터 발달해 올 수 있다고 믿었으며, 이들은 다른 지역 에 이주를 통해 둘 또는 그 이상의 무리로 나누어진다고 생각했다 . 이때 각 무리는 그 지역의 기후조건에 영향을 받으며, 적응이라는 과 정을 통해 모습이 천천히 바뀐다고 믿었다. 이런 뷔풍의 생각은 다윈 과 매우 비슷하긴 하나 그는 한 종이 다른 종으로 변한다는 진화설 Trans fo r rni sm 을 믿은 것은 아니었다. 에라스무스 다윈 에라스무스 다윈 (Erasmus Darwi n, 1731- 1 802 A.D. )은 찰스 다윈 의 할아버지로 전화라는 개념을 처음으로 생명체의 변화에 도입하였 다. 그는 진화를 자연선택과 성의 선택, 그리고 획득형질의 유전으로 표현하였으며, 인류의 발달 또한 이 개념으로 설명하였다 . 그는 전화 라는 개념은 설정하였으나 그 과정을 설명하지는 못했다. 뷔풍과 에라스무스 다윈의 이러한 생각은 포괄적인 체계를 갖지 못 했는데, 이들의 생각은 라마르크에 와서 비로소 정리되었다 . 라 □ ~르크 라마르크 (Baron Jea n Bap tist e de Lamarck, 1744-1829 A.D.) 는 프랑스 파리에 있는 자연사박물관에서 연구하던 무척추동물학자였다 (사진 2. 3). 유럽 이의의 대륙과 다른 박물관들에서 보내온 자연사 관계자료들을 연구하던 중 그는 5 6,\1]가 되던 1819 년에 생명에 대한 견해가 완전히 바뀌었다. 라마르크는 생명체는 〈사슬〉로 연결되어 있으며, 생명체는 이따금
생겨나서 시간이 지남에 따라 변하며 점차 복잡한 구조로 변해 가다 마침내는 사람과 같은 존재로 변화되었다고 생각하였다. 그리고 그는 화석들을 연구한 끝에 생명체들의 변화를 인정하였다. 라마르크는 변 화의 원인을 생명체들이 〈필요를 인식하게 되었기 때문〉이라고 생각 하였으며, 생명체들은 이런 필요를 살아 있는 동안에 채용하고 이렇 게 얻어진 획득형질은 다음 세대에 전해짐으로써 자식들이 환경에 더 잘 적응할 수 있게 해준다고 생각하였다. 한 예로, 라마르크 자신은 직접 이 실례를 인용하지는 않았지만, 기 린의 기다란 목은 환경의 변화에 따라 음식을 구하기 위해서였다는 다윈의 가설은 바로 라마르크의 생각을 받아들인 결과였다. 물론 이 런 특칭은 다음 세대에 전해질 수는 없다. 그러나 라마르크의 업적은 그 당시 환경이 동물과 식물에 미치는 영향의 중요성을 인식하였고, 환경에 적응하는 생물만이 살아 남을 수 있다는 점을 인식하였다는 데에 있다. 다만 라마르크는 환경이 동물과 식물에게 영향을 끼친다 고는 했으나 어떻게 영향을 끼치는지 그 메커니즘을 이해하지는 못했 다. 퀴l:::l l 에 퀴비에 (Baron George s Cuvie r , 1769-1832 A.D. )는 프랑스의 고동 물학자이자 비교해부학자로, 당시 지구상에 살던 동물들을 비교하여 비교해부학 연구의 장을 열었다(사진 2. 4). 당시 파리를 건설하기 위 해 파리 분지의 석회암을 채취하는 과정에서 암석화된 많은 동물뼈와 이빨들이 발견되었는데, 화석 중 일부는 살아 있는 동물과는 다른, 절 멸된 동물의 뼈로 밝혀졌다. 이때에 생물계에서는 〈철멸〉이란 개념이 성립되어 있었으나, 절멸이 일어난 과정과 이유는 정확히 알지 못했다. 독실하게 조물주에 의한 창조를 믿었던 퀴비에는 종교상의 믿음과 화석의 발견으로 인한 갈등을 해결하는 방법으로 소위 〈카타스트로피
즘 Car t as t rophi sm 〉 가설을 제안하였다. 이 가설에 따르면 과거에 지 구상에는 대절멸이 있었으며, 이어서 새로이 창조된 동 • 식물은 다음 절멸 때까지 변하지 않고 생명이 지속된다고 한다. 최근에는 6500 만 년 전에 갑자기 지구상에서 사라져버린 공룡의 절멸을 이 가설로 설 명하고자 하는 시도가 다시 진행되고 있다. 비록 퀴비에는 창조설을 믿었지만, 동물의 세계를 네 종류(등뼈동 물, 연체동물, 복합동물, 식충류)로 구분하여 이들의 기능적 특칭은 환 경의 적응 때문이라고 생각하였다. 그의 이런 생각은 라마르크의 생 명체는 사슬로 연결되어 있다는 이론과 함께 다윈의 진화설에 영향을 주었다고할수 있다.
 2. 5 『 지질학 원론』의 저서를 남긴 라이엘
2. 5 『 지질학 원론』의 저서를 남긴 라이엘
라 01 엘 라이엘 (Charles Ly e ll, 1797-1875 A.D. )은 영국 스코틀랜드 출신으 로 일찍이 지구의 형성에 많은 관심을 가지고 있었으며(사전 2. 5), 유럽 전역과 미국의 일부지역을 여행하면서 암석층위를 연구하였다.
라이엘은 지구의 모습은 자연적인 요소들, 즉 바람과 풍화작용, 비, 기후의 변화와 계곡을 깊게 파 내려가는 강의 흐름 등에 의해서 천천 히 지속적으로 변해 오늘날과 같은 모습으로 변했다고 생각하였다. 균일설 U nifo rma t er i a ni sm 이라고 불리는 리이엘의 이 가설은 184] 기 초 제임스 후돈(J ames Hut ton , 1726-1797 A.D. : 사전 2. 6) 이 처음 내세 웠으며 이를 라이엘이 가설로 확립하였다. 이 가설에서 주장하는 〈천 천히 그리고 지속적인 변화〉는 오랜 시간을 필요로 하는데, 이런 주 장은 동 • 식물처럼 지구도 변하지 않는다고 믿어왔던 19 세기 초 유럽 사람들의 사고를 무너뜨렸으며, 유럽 사람들은 지구의 나이가 오래 되었다는 주장에 충격을 받았다. 라이엘은 1930 년에 지은 유명한 『지 질학 원론 Pr i nc ip les of Geolo gy』에서 지구의 나이에 대해서는 언급하 지 않았지만 지구의 변화는 아마도 수백만 년이라는 시간에 걸쳐 진 행됐으리라고 추측하였다. 맬서스 맬서스 (Thomas Robert Malth u s, 1766-1834 A.D. )가 저술한 『인 구론 An Essay on the Pr inc ip le of Pop u lati on , 1798 』은 후에 찰스 다 윈과 알프레드 월리스가 독자적인 자연선택이론을 세우는 데 크게 영 향을 끼쳤다. 맬서스는 글에서 인구성장은 자연적인 요인에 의해 제 한을 받지 않고 그 수가 기하급수적으로 증가하여 매세대마다 수가 곱에 달하나, 식량생산은 산술급수적으로 증가한다고 언급하였다. 그 는 생명체들이 그 수를 증가시키려는 충동을 〈생존경쟁〉이라는 방법 으로 나타내는데, 인간은 이런 충동을 인위적인 방법으로 제한한다고 지적하였다. 다윈은 이런 맬서스의 생각을 받아들여 자연선택이 바로 전화의 요 인이라고 인식하였다. 생존경쟁이란 점에서 볼 때 적응도가 높은 개 체는 적웅도가 낮은 개체보다 살아 남을 확률이 높은데, 다윈은 종이
아닌 개체가 서로 작용한다는 점을 인식하고 있었다. 다윈 이전의 학 자들은 종은 변하지 않는 기본단위이며, 종 안에서 개체는 별로 중요 하지 않다고 생각했다. 따라서 이들은 개체에서 일어나는 변화를 관 찰하지 못했으나, 다윈은 개체의 변이를 통해 선택이 어떻게 작용하 는지를 설명할 수 있었다. 다윈은 이런 개체의 특이성을 바탕으로 전 화의 메커니즘을 자연선택으로 설정하였다. 이때 자연선택은 실제로 개체의 적응능력을 불문하고 개체에 작용하는 것이지만, 전화라는 개 념은 집단에 적용되는 것이다.
 2. 6 균일설의 주장에 접근했던 지
2. 6 균일설의 주장에 접근했던 지
 2. 7 비글호 항해에서 돌아온 31 세
2. 7 비글호 항해에서 돌아온 31 세
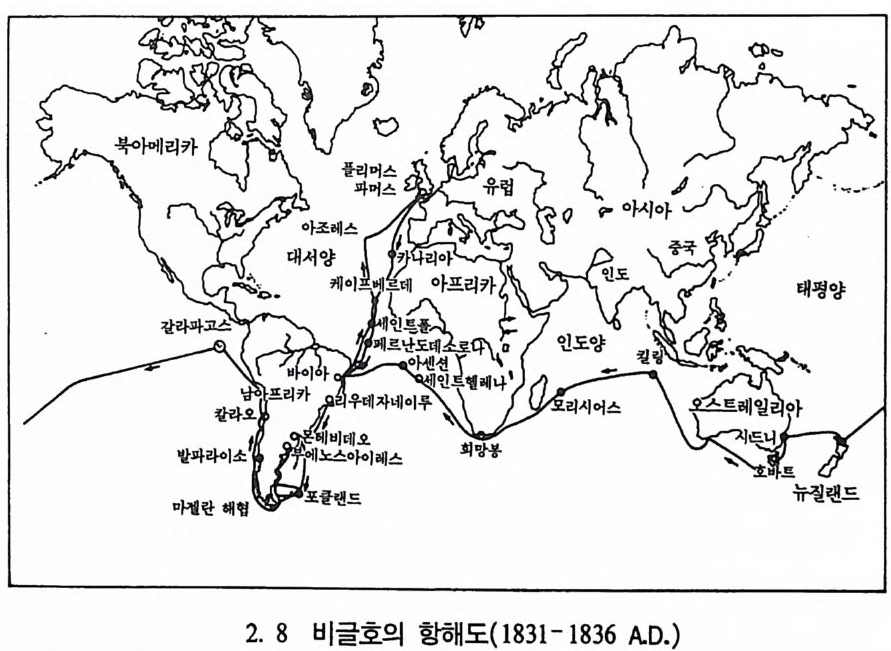
다윈의 시대 : 찰스 다윈 (Charles Darwin , 1809-1882 A.D.) 찰스 다윈은 영국의 부유한 가정에서 태어나 l 伊 1] 때부터 이미 자 연계와 그 존재에 관심을 가지고 있었다. 1825 년에 에딘버러에서 의
학을 전공하고자 했으나 부모의 뜻에 따라 목사가 되기 위해 전공을 바꾸고 목사수업을 위해 캠브리지 대학으로 가게 된다(사진 2. 7). 대학 시절 다윈은 성적이 뛰어난 편은 아니었으나 자연과학에 관한 호기심이 생겨났으며, 그는 여기서 식물학자(꽃가루 전공)인 헨슬로 Henslow 를 만나게 된다. 당시 대학의 연구 분위기는 요즘과는 달라 서 자연에 대한 사람들의 마음은 닫혀 있었고, 자연에 대한 질문과 증명, 사고조차도 법에 저촉되는 일이었다. 단지 소수의 학자들만이 학교 밖에서 자연과학을 강의하고 토론하였다. 헨슬로도 그 중의 한 사람으로, 젊은 다윈으로 하여금 자연과학자의 자격으로 구축함 〈비 글호 〉 에 타도록 추천하였다. 이때 다윈의 나이는 23 세였다. 〈비글호〉 의 함장인 피츠로이 P it zro y는 복음선교인으로, 그는 이 항해를 통해 복음을 선교하고자 하였다. 항해는 5 년 동안 계속되었다(그립 2. 8). 다윈은 이 조사 중에 많은
 r - L
r - L
일들을 관찰하였으며, 다양한 책들을 읽었다. 다윈은 그가 읽은 책들 중 라이엘의 『지질학 원론』에서 많은 영향을 받는데, 이 책은 후돈의 〈균일설 〉 의 입장을 견지하고 있었다. 다윈은 항해중에 기착하는 지역 에서 발견되는 화석과 그 지역의 지리적 변화를- 관찰하였으며, 섬에 살고 있는 동물들을 분류하였다 . 1837 년, 그의 나이 27 세 때 영국으로 귀환한 다윈은 저술을 시작하 였는데, 주로 〈비글호〉 여행에 관한 내용과 산호초에 관한 것들이었 다. 식물과 동물들의 교배와 생태에 관해 연구를 진행시키던 다윈은 1839 년까지 그의 유명한 전화론의 윤곽을 마련하였다. 그 후 1842 년 에 35 쪽 정도의 이론의 초고를 마치고 2 년 후인 1844 년에 그 내용을 24 碑으로 확대하였다. 그러나 당시 사회의 지적상 분위기는 다윈으 로 하여금 발표를 망설이게 하였다. 1858 년에 월리스(사전 2. 9) 가 다윈에게 논문 한 편을 보내는데, 이 논문은 월리스가 맬서스의 인구론을 읽고 말레이 반도에 대해 독자적 으로 연구한 끝에 다윈과 갇은 결론인 〈자연선택 Natu ral Selecti on > 이론을 추출한 내용이었다. 이 논문을 읽은 다윈은 자신이 월리스의 연구를 도용하였다는 소문을 두려워해 자신의 연구를 발표하길 망설 였다. 그러나 다윈의 독자 연구를 이해한 천구들의 주선으로 1858 년 에 다윈과 월리스는 〈린네학회 잡지〉에 합동으로 연구내용을 발표하 였다. 1859 년 다윈의 『종의 기 원 The Orig in of Sp e cie s by Means of Natu - ral Selec ti on 』이 출판되었는데, 출판 첫날로 책은 매전되었다. 그러나 실제로 다윈은 그의 이론을 발표하기를 꺼려했으니, 이는 첫째 다윈 자신이 이론의 발표에 의해 야기될 결과에 두려움을 가지고 있었으 며, 둘째는 자신의 신앙십 때문에, 그리고 끝으로 자신이 관찰할 수 없는 것에 대한 더 많은 증거들을 원했기 때문이었다 . 『 종의 기원』에서 다윈은 인류의 전화에 대해서 아주 짧게 언급하였
 2. 9 박물학자이자 탐험가인 월리스
2. 9 박물학자이자 탐험가인 월리스
 -&T4* ,r • .w• 山 ‘”따 .. ..· ‘m -,,`.,- ..-- ,._ . . , .
-&T4* ,r • .w• 山 ‘”따 .. ..· ‘m -,,`.,- ..-- ,._ . . , .
 2. 11 인간과 유인원과의 관계를 철
2. 11 인간과 유인원과의 관계를 철
 2. 12 다윈을 지지한 토 □~ 헨
2. 12 다윈을 지지한 토 □~ 헨
다 ;
류는 다른 모든 생물과 연관이 있다〉는 주장을 당시의 사람들은 받아 들일 준비가 되어 있지 않았던 것이다. 특히 당시의 만화는 다윈울 신랄하게 풍자하였다(사전 2. 10). 예 : 윌버포스 주교와 토마스 헨리 헉슬리와의 논쟁 윌버포스 : 〈원숭이가 조상이라면 할머니 아니면 할아버지 어느 편이 유인원과 관계가 있는지? 〉 (사전 2. 11) 헉슬리 : 〈나는 우리 조상이 유인원이었다는 데는 조금도 부끄럽 지 않다• 그러나 진실을 감추는 데 인간의 재능을 쓰 는 사람을 조상으로 가졌다는 점이 부끄럽다.〉(사전 2. 12) 다윈의 연구는 아래와 갇이 간단히 설명될 수 있다. 첫째, 생명체는 그들 집단의 규모를 일정하게 유지하기 위한 수보 다 더 많은 자식을 생산한다. 따라서 어린 개체의 사망률이 높다. 둘째, 각각의 개체 안에는 많은 변이 Var i a ti on 가 있는데, 이때 특별 한 특칭을 가전 개체가 환경에 더 잘 적응하며 적응에 실패한 개체는 도태된다. 다윈의 이런 두번째 생각은 맬서스의 〈인구론〉에서 빌려왔 다. 셋째, 자식들은 부모를 매우 닮았는데, 이는 부모의 인자가 자식에 게 전달되어 유지되었기 때문이며, 각 세대는 차츰 생기는 변화에 의 해 적응의 정도를 높인다. 다윈은 이런 변이의 과정과 환경에 더 잘 적응하는 개체의 선태울 자연선택이라고 부르고, 전화의 직접 추전요인으로 파악하였다. 자연 선택이란 개념은 사람들로 하여금 사물에 대한 인식의 혁명을 가져오 게 했다.
다윈의 업적은 전화를 발견하지는 못했지만 전화에 관한 증거와 이 론을 제공하였다는 데 있다. 다시 말해 종들은 뿔뿔이 창조된 것이 아니며, 자연선택이 생물 사이에서 변화의 직접원인이라는 점을 밝힌 데에 그의 업적이 있다. 이때 다윈이 사용한 증거로는 끊임없이 나타 나는 화석들과 화석들의 남은 부분, 살아 있는 동물들의 몸의 구조에 관한 해부학상 비교자료 및 동물 스스로의 번식능력 등을 들고 있다. 이런 것들을 근거로 다윈은 진화론을 주창하였는데, 그는 진화를 〈자 손에서 나타나는 변화 a chang e ·as a descent with mo difi ca ti on 〉로 정의하였다. 이어 1871 년에 간행된 『인간의 후예』 등의 저서는 인류 의 의식에 혁명을 가져왔다. 다윈은 인간이 더 이상 생물세계의 주역 이 아님을증명하였다. 지난 100 여 년간 〈 모든 생명체는 초자연의 힘에 의해 만들어전 것 이 아니라는 이론 〉 인 다윈의 전화론은 인류의 모든 면에 강력한 영향 을 미쳤다. 많은 사람들은 이런 점에서 일찍이 자연선택이 생물 진화 요인의 답이었다고 생각할지 모르나 실제로는 많은 논란이 1930 년대 까지 이어졌다. 다윈은 그의 진화론을 구상하는 과정에서 개체변이가 일어나는 이유를 알지 못해 생물이 살아 있을 때 얻어전 특징들은 다 음 세대에 전해전다는 라마르크의 연구결과에 의존하였다. 그러나 왜 변이가 일어나는지는 멘델 때에 와서야 그 답이 나오게 되었다. 다윈 이후의 시기 : 현대 진화론의 성립 19 세기 이후 많은 학자들은 다윈의 자연선택이 진화상 변화의 주요 요인이라는 가설을 지지했지만, 생물체의 자체 내에서 일어나는 〈필 요 Need 〉가 변화의 직접요인이라는 라마르크의 주장을 포함하는 다른 이론들이 여전히 존재하였다. 특히 멘델의 유전학 연구를 이어받은 새로운 학파가 출현하여, 2 어]기 초반 휴고 드 브리 Hug o de Vr i es 가
이끄는 유전학자들이 새로운 이론의 주역을 담당하였다. 이둘은 진화 의 변화를 가져오는 주요 요인은 유전물질의 변화라고 주장하였다. 이들의 주장은 후에
다윈과 현대 진화론의 적응도 비교 F itt ness Comp a ris o n 다윈의 적응요인 현대 진화론의 적응요인 디론 어린이 사망률 다른교배충동과성패율 Di ffere nt morta l ity in juv en iles Di ffer. mati ng driv e and 다론어른사망률 mati ng success Differ. morta lity in adult sta g e 다른번식률 다른 생존능력 Di ffer. fer ti lity Differ. via b il ity 다른생식력 디론교배충동과성패율 Differ. fec undit y Differ. mati ng driv e and mati ng success
특히 1920 년대에 급격히 발달한 유전학의 연구성과와 다윈의 이론 이 결합되어 소위 〈현대 전화론〉을 정의하였다. 현대 전화론에서는 전화를 〈한 집단의 유전자 구성빈도가 세대가 지남에 따라 변하는 것
a change in allele from one ge nerati on to ano t her 〉 이라고 정의하고 있다. 현대 진화론의 주된 내용은 〈 서로 다른 재생산 d iff eren t rep ro - duc ti on 〉 이라는 개념을 바탕으로 〈 진화점진론 Ph y le ti c Gradualis m > 과 개체 내에서의 변이를 인정하였다. 현대 진화론에서는 돌연변이, 유전자표류, 유전자이동 , 그리고 자연선택을 진화의 요인으로 들며, 이 중 자연선택을 전화를 추전시키는 요인으로 인정하였다 . 1972 년 미국 고동물학자인 스티븐 제이 굴드 S t e phen Ja y Gould 와 니일 엘드리 지 Ni le Eldr i d g e 는 생물체의 전화를 급격한 변화인 소위 계단식 평 형설 Punctu ate d E quili br i um 로 설명하였다. 다윈의 진화점 전론에 따르면 디른 시기에 살던 동물들 사이에 중간형이 화석기록으 로 발견되지 않는 것은 〈 지질상의 기록이 불완전하기 때문t he im - pe rfe c ti on of the geo log ica l records 〉이라고 설명된다. 이에 반해 굴드와 엘드리지는 불완전한 기록 그 자체가 전화상 변 화의 방법을 나타내주는 증거라고 주장한다. 죽 생명체는 한동안 형 태가 변하지 않고 있다가 어느 시점에서 갑자기 변화하기 때문에 중 간형이 나타나지 않는다고 주장한다 . 이들의 가설은 1950 년대에 마이 어에 의해 제기된 〈 타지역 분화 Allo p a tr i c S p e ci a ti on 〉와 같은 가설 로, 새로운 종은 〈 유전혁명 〉 이 일어날 수 있는 고립된 작은 집단 내 에서 일어나며 이때 생겨난 새로운 종은 기존의 커다란 집단의 전체 유전특칭과 섞이지 않는다고 주장한다. 창조론과진화론 다윈이 『종의 기원』에서 인간과 다른 동물은 관련이 있다는 암시 룰 제시함으로써 전화론과 창조론은 생명체 중 특히 인간문제에 관해 심각한 논쟁을 계속해 오고 있다. 여기서는 창조론과 진화론을 비교 하기 전에 먼저 인류사회에 나타나는 신화 m yt h 가 무엇을 의미하는
지를 이해하는 것이 진화론과 창조론을 이해하는 데 도움이 된다. 신화란 〈 밖으로 표출된 역사 사건들이 항상 관습상 이야기 형태로 나타나는 것을 의미하는데, 이때 역사 사건이란 당시 사람들의 세계 관이나 그들이 실제로 행하고 믿고 또 그들이 경험한 자연현상을 표 현한 것〉으로 정의된다. 지구상에 지금까지 많은 사회들이 문화를 가지고 있는데, 이들 문 화는 대개가 나름대로의 탄생신화를 가지고 있음을 알 수 있다. 여기 서는 몇 개의 신화를 살펴보기로 한다. 예 1) 아메리칸 인디언인 체로키족의 신화에 따르면 지구는 본래 물로 덮여 있었다. 어느 날 물 속에 사는 물방개 한 마리가 물 밖으 로 진흙을 밀어 올렸는데, 이 진흙이 최초의 육지가 되었다. 최초로 오누이가 육지에 나타났으며, 어느 날 오빠가 물고기를 한 마리 잡아 서 〈아이를 가져라〉 하면서 누이동생을 때렸다 . 이후 누이동생은 7 일 마다 아이룰 낳았으며 이들이 체로키족이 되었다. 예 2) 중앙아프리카의 야오족의 신화에 따르면 어느 날 카멜론이 낚시를 가서 함정을 만들었는데, 다음날 망에 작은 남녀 한 쌍이 잡 혔다. 카멜론은 이들을 가지고 신 앞에 가서 어떻게 해야 할지를 물 었다. 그러자 신은 카멜론에게 〈그들을 땅에 내려놓고 키워라〉 하고 답하였다. 그들이 자라 결합하여 사람들이 접점 늘어나고 점차 불을 사용하는 법을 배웠다. 세월이 지나 이들은 불을 간직하고 짐승을 좇 아 숲으로 들어가 야오족이 되었다. 예 3) 에티오피아의 신화에 따르면 신이 사람들을 진흙으로 만들었 는데 단단하게 하기 위해서 불에 구웠다. 맨 처음 불 속에서 끄집어 낸 사람은 잘 구워지지 않아 북쪽으로 던졌더니 흰 피부를 가전 유럽
사람의 조상이 되었고, 두번째 사람은 너무 까맣게 구워져서 남쪽으 로 던져 아프리카 흑인의 조상이 되었다. 마지막에 구운 사람은 알맞 은 색깔을 띠게 되었는데 이들이 에티오피아사람의 조상이 되었다. 예 4) 우리에게 친숙한 성경의 창세기장에 따르면 최초의 인류인 아담은 하느님의 형상을 따라 만들어지며, 이브는 아담의 갈비뼈로 만들어졌다 . 뱀의 유혹으로 에덴의 동산울 떠나게 된 후 이들의 후예 가 퍼져 나갔으나 노아의 대홍수 때 방주에 탄 생명만이 살아 남아 다시 지구에 퍼져 나갔다. 창조론자들이란 하느님이 우주와 만물을 창조했다고 믿는 사람들을 말하는데, 이런 점에서 본다면 대다수의 많은 사람들이 창조론자라고 할 수 있다. 그러나 지난 1 세기 동안 전화론에 관한 논쟁이 진행되면 서 창조론자들에게는 새로운 이름이 붙여졌으니, 이들은 모든 종류의 살아 있는 생명은 조물주에 의해 직접 만들어졌으며 어떤 생명도 변 화하지 않았다고 믿기 때문에 반전화론자 An ti -Evolu ti o ni s t s 라고 불 려질 수 있다 . 오늘날 창조론자들은 미국을 중심으로 활동하고 있다. 미국 내에서 창조론은 정통파 그리스도교인들에 의해 주도된다. 정통 그리스도교 주의 Ch ist i an Fundamen tali sm 는 미국 사회운동의 하나로 제 1 차 세 계대전 후 급격히 변해 가는 사회에 대항해서 발생하였다. 이들의 주 의 • 주장은 다음과 같은 5 개의 믿음 강령을 바탕으로 하고 있다. 1. 성 경 내용에 오류는 없음 The Inerrancy of the Bib l e 2. 예수의 신성 성 The Di vinity of Jes us 3. 처 녀 잉 태 The Vir g i n Bir t h 4. 그리스도가 십자가에 못박힘으로 인류의 죄가 속죄받음 The ato n ement of mank ind 's sin s by the crucif ixion of
Chris t 5. 예수의 부활과 최후의 재림 Hi s resurrecti on and eventu al se-cond comi ng 오늘날 미국에서 정통 그리스도교 이념의 리더로 1980 년대 미국 공 화당 전당대회의 강령을 작성했던 제리 파웰 목사 Rev. Jer ry Falwell 를 꼽을 수 있다. 이들은 1960 년대 미국을 휩쓸던 사회 • 사상사의 대 혼란과 나치즘, 허무주의, 공산주의 등의 모든 사상과 모든 반기독교 식 믿음과 행동의 근원은 바로 전화론을 가르쳤기 때문이라고 주장했 다. 창조론 모델에 따르면 모든 생물체들은 아래와 같은 특별한 과정을 통해 나타났다고 주장한다. 제 1 단계에서 창조는 초월된 존재(하느님)에 의해 이루어졌으며, 지 구는 처음부터 완전하게 창조되었고, 이때 지구의 대기권은 안전한 막으로 덮여 있었다. 인간은 특별히 창조되었으며 결코 부패되지 않 았다. 제 2 단계는 지구 위에 몰락과 죽음, 부패가 생겨나고 안전막이 점차 되조하기 시작한다. 성경에서 보이는 뱀에 의한 유혹이 사건의 발단 이다. 제 3 단계는 지구에 대홍수가 도래하였으며 지구는 거의 40 일간 물로 덮여 있었다. 이 기간 중에 지구상의 모든 지질상의 충위들이 만들 어졌으며, 그때까지 지구상에 존재했던 생물 중에 생존자들은 한 배 (노아의 방주)에 있었다. 제 4 단계는 홍수가 물러가고 지구에 존재하던 에너지에 의해 빙하 가 만들어졌으며, 지구를 둘러싸고 있던 안전막이 붕괴되어 생명체의 수명이 감소되었다. 여기서 창조론자들의 전정한 의도에 대한 의문이 제기된다. 창조론
자들의 의도는 공립학교에서 과학을 가르치는 데에 있어서의 변혁, 죽 과학과 창조론을 같은 비중으로 가르치길 희망한다. 그러나 실제 로 그들의 목적은 스코프 시대의 사고와 그 이후에 일어났던 움칙임 을 현재로 끌어오려는 의도이며, 더 이상 성경에 나타나는 신앙상의 측면에서 창조를 이야기하지 않고 대신 〈과학성 창조론 Sc i en tifi c Crea ti on i sm 〉 을 주장한다. 창조론자들은 그들의 견해가 성서식이 아 니라 다윈의 이론처럼 과학이론이라고 주장한다. 창조론자들은· 공립 학교 교과과정에 창조론의 견해롤 도입하고자 하며, 그들은 자기들의 이론이 과학적이고 그들이 오늘날의 갈릴레이라고 표현한다. 이렇게 미국에서 시작된 〈과학성 창조론〉이 최근에 국내에 소개되 었고, 이를 받아들이는 기독교 신자와 학자들이 창조학회를 조직하여 이룰 중심으로 활동하고 있다. 이들은 미국의 창조학회와 창조론자들 의 주장과 활동을 받아둘이고 있는데, 더러는 그들이 주장하는 과학 성 창조론의 내용을 충분히 이해하지 못하고 있는 경우도 있다. 한 예로 창조론을 지지하는 소수의 학자들은 중 • 고등학교 생물교과서를 창조론의 입장에서 서술하고 이룰 공립학교의 교과서로 채택하여 가 르치길 요구하고 있다. 그러나 문제는 창조론이 과학이냐 아니냐 하는 데에 있다. 위에서 언급한 여러 창조 모델 중에 창조론자들의 주장은 명백히 성경의 창 세기를 바탕으로 하고 있다는 점이다. 그러면 힌두교나 불교 또는 체 로키의 모델은 어떻게 이해되어야 하는가 하는 문제가 남게 된다. 이 들을 비교종교학 교실에서 가르찬다면 별 문제가 없으나, 창조론자들 은 자기네의 모델만을 공립학교에서 의무로 가르치길 바라며 이룰 법 으로 제정하기를 바라는 데에 문제가 있다• 그러나 우리나라 헌법에 는 신앙의 자유를 보장하고 종교와 국가를 분리하고 있는데 특정한 종교를 강제로 가르치라는 것은 문제가 있다. 미국 창조학회 회장인 모리스는 성경과 진화론은 조화를 이루는 것
이 불가능하며, 단지 그들의 모델 중 〈 과학이론 〉 만이 교과서에 포함 되어야 한다고 말한다. 그러나 문제는 창조론의 모델 모두가 성경에 근거해서 만들어졌다는 데 있다. 따라서 〈 과학성 창조론 〉 이란 성경에 기초를 둔 정통 그리스도교인들의 견해일 뿐이라고 할 수 있다. 이 갇은 점에도 불구하고 문제의 심각성은 공립학교에서 창조론을 지지 하는 사람들이 급속히 발전하는 지식이나 복잡한 사회를 이해할 만한 준비가 되어 있지 않다는 데에 있다. 창조론에 반해 과학이란 무엇인가? 미국 하버드 대학에서 과학시간 의 역사를 1859 년부터 1950 년까지 조사해 보았더니 놀랍게도 교과과 정에 과학, 수학 또는 그 중에 한 과목만이 매년 매학기에 포함되고 있을 뿐이다. 오늘날에도 단지 10 퍼센트의 학생들만이 과학과목을 수 강하고 있을 뿐이다. 우리나라 대학에서도 같은 실정이다. 학생들은 졸업 때까지 겨우 한 과목의 자연과학개론을 수강할 뿐이다 . 이같이 과학이 오늘날 우리에게 미치는 영향과 공헌은 과거에 우리가 경험하 지 못한 엄청난 양으로 발전하였으나 우리 대학에서는 오히려 과학과 목을 줄여오고 있으니, 우리 사회에 미치는 과학의 효과는 극대화되 어가는 반면 학생들은 이런 효과를 소화하고 이해할 훈련이 되어 있 지 않다. 과학은 어떤 생각, 죽 가설을 설정하고 이를 구체화하는 것이다. 그 리고 이 가설을 비판하는 시험과 실험이 있어야 한다. 과학은 〈절대 진리〉를 다루지 않으며, 비종교이며 어떤 현상을 초월적인 설명으로 대신하지 않으며 관찰을 통해 설명하고자 한다. 이에 반해 창조론은 절대전리를 바탕으로, 질문은 있으나 이에 답하는 대답은 없으니 어 떤 증거나 자료도 그들의 마음을 변하게 할 수 없다. 과학자들은 그들이 찾고자 하는 답울 얻기 위해 질문을 가질 수 있 으며, 때로는 잘못된 해답을 얻게 되는 수도 있다. 과학은 어떤 일들 이 잘못 증명될 수도 있다는 점을 전제로 하고 있다. 그러나 창조론
은 잘못을 증명할 수 없다는 점에서 과학이라고 규정할 수 없다. 미 국에서는 스코프 사건 이후 소련에서 스푸트니크 인공위성을 쏘아 울 리자 이에 자국을 받아 과학교육이 강조되기 전까지 창조론을 〈 사실 〉 로서 가르쳐왔다. 한국에서도 창조론을 과학시간에 가르쳐야 한다는 주장이 창조학회를 중심으로 제시되고 있으며, 중 • 고등학교 생물교 과서에서 다루어져야 한다는 주장파 함께 일부 학자들은 소송을 제기 하고 있는데, 그 내용은 앞에서 주장한 바와 같이 미국 창조론자들의 의견을 따르고 있는 실정이다. 한편 과학성 창조론을 입법화하는 것에 대한 강력한 반대 움직임이 노벨 수상자들을 중심으로 전개되고 있는데 , 이들은 〈 신앙의 생각을 과학시간에 가르친다는 것은 과학교육에 큰 손실을 가져오는 것이며, 이는 과학과 종교 사이에 잘못된 갈등을 불러일으키는 결과를 가져온 다. 나아가 우리 젊은 세대에게 과학적 사고를 갖게 하는 데 문제가 되며, 점차 과학사회에서 일어나는 문제들에 대처하는 우리들의 능력 을 저하시킨다〉라고 하여 과학시간에 창조론을 가르쳤을 때의 문제 점을 지적하고 있다. 끝으로 1981 년에 교황 폴 쩌 1 의 말을 상기해 보자. 그는 〈우주의 창조와 우주론은 사람들과 종교 사이에서 커다란 홍미를 일으켜왔다. 성경 그 자체는 우리에게 어떻게 우주가 시작되었고, 이루어졌는가를 말해 주는 것으로 이는 우리에게 우주 발생에 관한 답을 제시해 주는 것이 아니라 신과 인간과의 그리고 우주와 인간과의 정확한 관계를 제시하는 것이다. 성경은 단지 세상은 하느님에 의해 만들어졌음을 알리는 것이며, 이런 전실(?)을 가르치기 위해 성경이 쓰여전 때에 통용되던 우주론이라는 용어롤 쓴 것이다〉라고 말한 내용을 음미할 필요가 있다.
 제 3 장 생명의 바탕 : 유전과 전화
제 3 장 생명의 바탕 : 유전과 전화
제 3 장 생명의 바탕 : 유전과 전화 인류유전학 : 개인단위에서 일어나는 변화 체질인류학에서는 전화의 과정과 산물을 공부하는데, 이때 변화는 개체에서 일어나나 진화는 개인이 아닌 집단에 적용된다고 앞에서 설 명하였다. 현대 진화론에서는 전화를 〈한 집단 내에 유전자 구성빈도 가 세대가 지남에 따라 변하는 것〉으로 정의한다. 물론 전화는 개체 의 해부학상, 생리학상 변화나 행위의 변화에서도 볼 수 있으나, 실제 는 유전자의 변화에 의해 일어난다. 여기서는 먼저 인간을 인류유-전 학에서 다루는 〈분자단위〉와 〈개인〉 단계로 나누어 살펴본다. 세포와분자단위 유전학에서 개인은 〈세포〉와 세포 내 〈분자〉의 단위로 나누어진다. 다윈 이론의 주요 내용은 변이와 유전, 그리고 적응으로 요약될 수 있다. 이 중 사람을 구성하는 가장 작은 단위인 분자와 세포단위에서 변이와 유전이 어떻게 일어나는지 살펴보자. 다윈은 자연선택이 한 집단 내에서 일어나는 변이의 원인이라고 생 각하였다. 만일 어떤 집단 내에서 배우자를 선택할 때 작은 발을 가
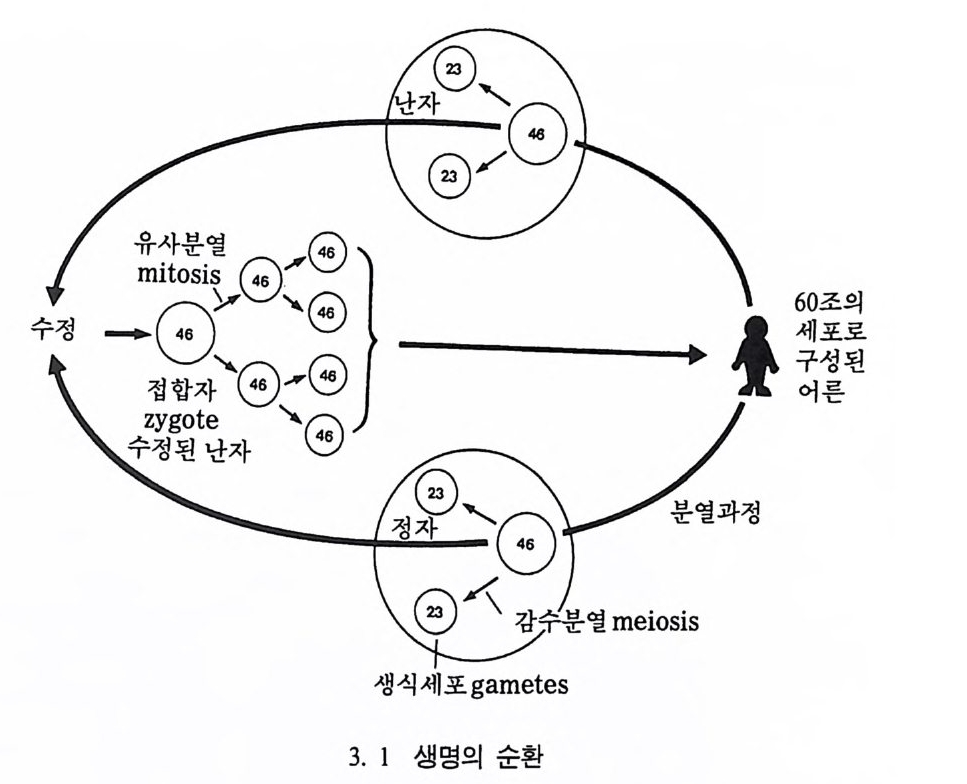
전 사람은 아이를 낳을 수 없다고 한다면, 여러 세대를 지난 후에는 큰 발을 가진 사람들이 많아질 것이다. 시간이 지난 후 작은 발을 가 전 사람들이 다시 만들어질 수 있을까, 그리고 계속해서 더욱더 큰 발을 가진 사람들을 만들어낼 수 있을까? 이렇게 한 집단 내에서 보 이는 변이의 요인이 무엇인가가 바로 다윈이 해결하지 못한 문제였 다. 다윈은 집단 가운데에 변이가 있다는 것을 알았으며, 이러한 변이 들 중 어떤 개체는 다른 개체보다 환경에 더 잘 적응한다는 것을 알 았다. 다시 말해 환경에의 적응도가 높은 개체가 더 찰 번식한다는 뜻으로, 이때 적응도란 한 개체가 더 잘 재생산 할 수 있는 능력을 말한다. 그러나 다윈은 환경에 더 잘 적응하는 원숭이가 어떻게 더 많은 새끼를 나울 수 있는가를 설명할 수는 있었으나, 어떤 원숭이가 어떤 방법으로 더 잘 적응하는지는 알지 못했다. 다윈 당시에는 유전 이라는 것에 대해서 오늘날 우리가 알고 있는 수준에까지 이르지 못 했으므로, 그때 사람들은 . 자식의 해부학상 • 생물학상 특성은 부모의 특성을 섞어놓은 것 Blen di n g Inher it ance 으로 생각하였다. 『종의 기원』이 발간되기 3 년 전인 1856 년에 멘델은 완두콩 관찰을 통하여 유전은 섞임이 아니라 특성이 일정한 규칙을 가지고 나타난다 는 사실을 알았다. 이는 다윈이 결코 알지 못했던 획기적인 발견이었 으나, 학계는 1900 년대 초까지 멘델의 유전에 대한 연구의 중요성을 알지 못하였다. 부모의 유전자들이 어떻게 자식들에게로 전해지는가? 사람은 어른 의 경우 60 조의 세포를 가지고 있는데, 이것은 다시 46 개의 염색체 (c 뇨 omosome ; cbromo=color, soma=bod y)를 가지고 있는 세포와 23 개의 염색체를 가지고 있는 세포로 나누어전다. 23 개의 염색체를 가지고 있는 세포를 생식세포라 하며 정자와 난자가 이에 속한다• 이 정자와 난자가 결합하여 사람이 만들어진다. 수정된 난자인 집합자는
 \::、자 4:60: 조;의
\::、자 4:60: 조;의
어론의 세포처럼 46 개의 염색체를 갖고 있다. 이 접합자는 수없이 많 은 분열을 하여 마침내 하나의 독립된 사람을 만든다(그림 3. 1). 유전자의 구조와 특성 인간의 유전물질은 어떤 구조를 지니고 있을까? 1cm 를 l (H탐손하면 1mm 가 된다. 이 1mm 를 l0 昞꿉!하면 1 마이크로로, 이 길이는 염색 체의 평균길이와 같다. 첫번째 염색체는 오래 전에 현미경을 통해서 착색된 세포 내에서 관찰되었다. 수정된 난자는 약 5660 만분의 1 그램 의 무게를 가지고 있다. 이 수정된 난자가 계속해서 유사분열을 거듭 하여 새로운 생명체를 만들어낸다• 마지막으로 만들어전 사람은 처음 에 수정된 난자보다 약 400 억 배로 커진다. 이때에도 생식세포를 제
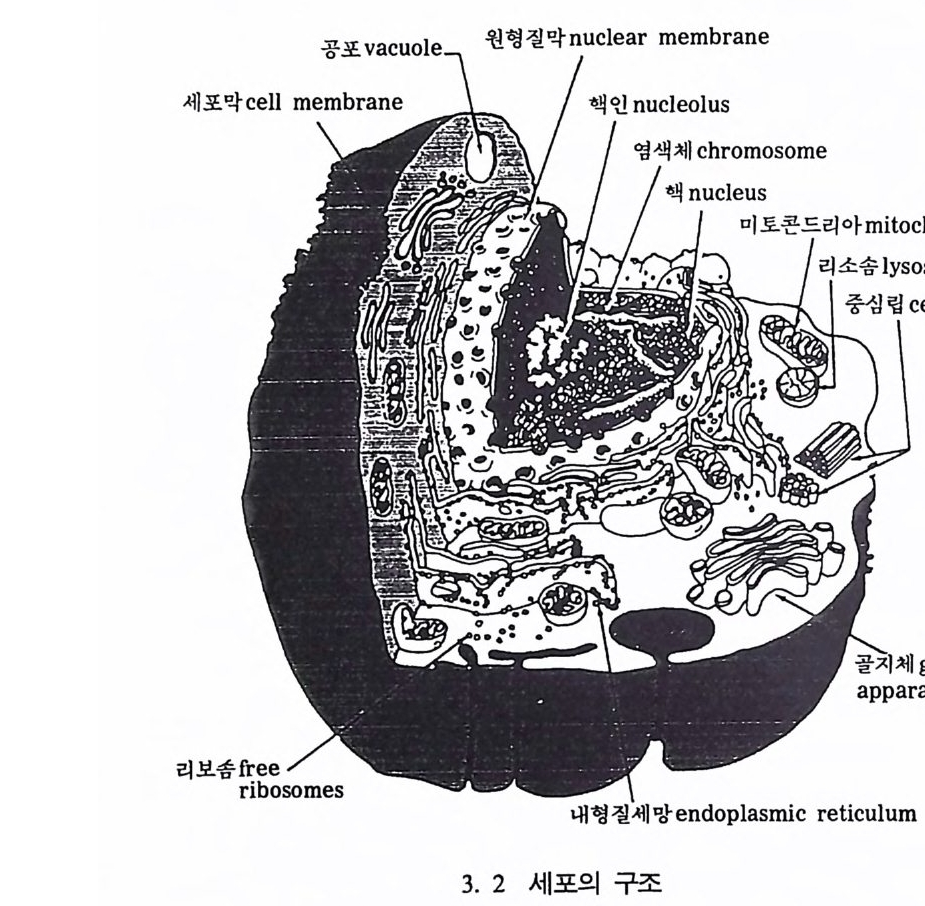
의하고 모든 세포는 여전히 46 개의 염색체를 가지고 있다• 새로운 생 명체를 만드는 정보는 바로 이 염색체에서 발견된다 . 이 정보들은 염 색체 내에 있는 유전물질인 유전인자에 저장되어 있다. 바로 멘델이 말한 유전인자 Gene ti c Par ti cle 이다. 이 유전인자가 생명체의 기능을 조절하며 다음 세대로 자신둘을 전달하는 기능을 가지고 있다(그림 3. 2).
 원형질막 nuclear membrane
원형질막 nuclear membrane
오늘날 우리는 유전자 연구를 통해서 어떻게 이들이 다음 세대로 자신들을 전하는지를 알 수 있으며 그 과정 또한 잘 알려져 있다. 이
와 더불어 유전인자들의 어떤 기능이 생명체의 발육을 조절하는가에 대해서, 그리고 기타 디론· 활동들에 대해서도 많은 연구가 이루어지 고 있으나 그 과정들이 완전히 이해되고 있지는 않디 •. 염색체는 핵산 (DNA : Deoxy ri b o nucleic acid ; 이후 핵산은 DNA 로 표시함)과 단백질로 이루어져 있다. 과거에 유전인자를 포함하고 있는 것이 DNA 인지 아니면 단백질인지에 대한 논쟁이 있었으나, 1952 년에 DNA 가 유전자를 가지고 있음이 알려졌다. DNA 는 지구상 에 있는 수많은 과분자 중의 하나이다.
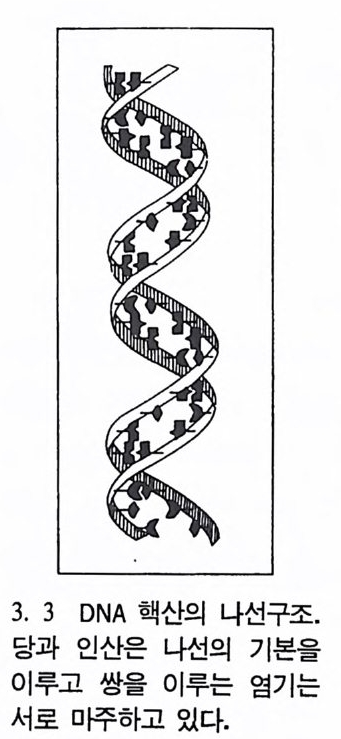 3. 3 DNA 핵산의 나선구조.
3. 3 DNA 핵산의 나선구조.
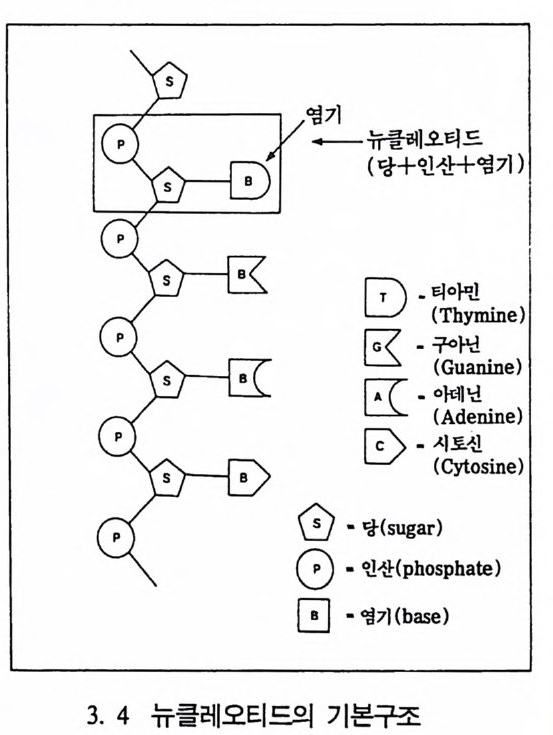 염기
염기
1952년 에 윗슨과 크릭 Wats o n & Cr i ck 이 DNA 의 구조를 그려냈 는데, 이는 생물학에서 아주 중요한 발견이었다. DNA 는 당과 인산염 그리고 영기의 세 개 물질로 구성되어 있는데, 영기는 네 개의 화학
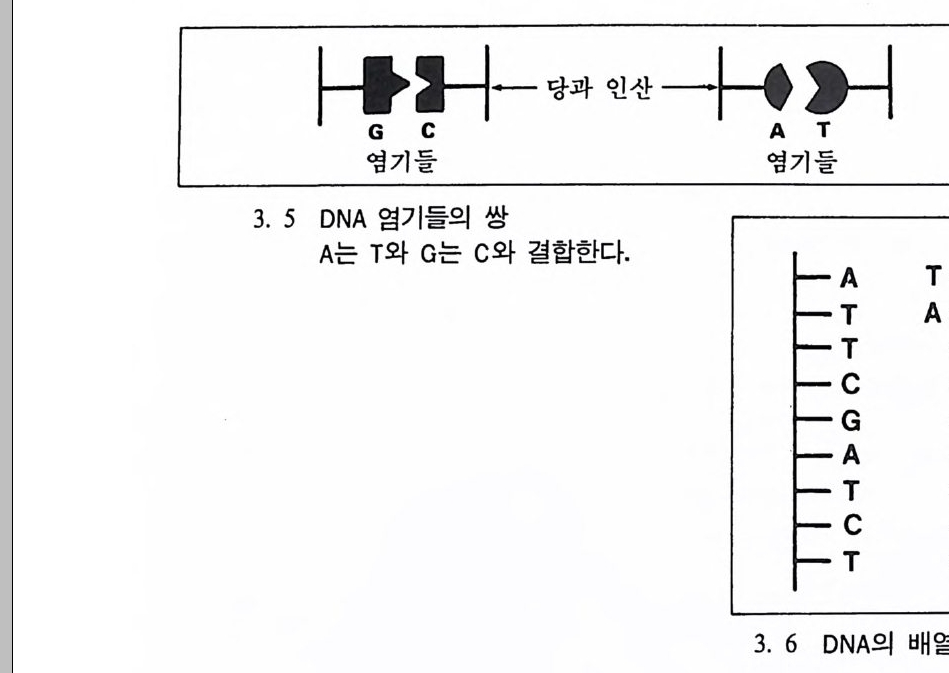 f}-당과 인산一卜' '」
f}-당과 인산一卜' '」
물질로 되어 있다(그립 3. 3, 3. 4, 3. 5, 3. 6). DNA 의 기본구조는 DNA 가 가전 두 가지 중요한 기능을 설명해 준다 : 첫째는 DNA 가 세포에서 어떻게 복제될 수 있는지를 알려주 며, 두번째는 DNA 가 어떻게 세포의 기능을 조절하는지를 설명할 수 있게 되었다. 윗슨과 크릭은 이 연구로 노벨상을 수상하였으나, 당시 사람들은 그들에게는 이해될 수 없는 이런 기초과학의 효율성에 의문 을가졌다. DNA 는 〈생명의 날t he th read of lif e 〉이라고 불린다. 만일 같은 사람의 혀와 췌장에서 세포를 각각 하나씩 떼어내어 DNA 를 비교해 보면 이들이 동일한 성질을 가지고 있음을 알 수 있다. 서로 다른 두 사람의 혀에서 추출한 DNA 는 성질이 약간 다르다. 사람과 침팬지에 서 나온 DNA 는 약 98.4% 가 서로 같은 성질을 보이며, 오직 1. 6% 만 이 서로 다를 뿐이다. 그러나 사람과 도마뱀의 차이는 아주 크다. 다
시 말해서 서로 아주 다른 생명체는 DNA 구조에서 역시 상당한 차 이를 보인다. 사람은 약 30 억 개의 염기쌍이 모여서 된 DNA 들로 이루어져 있 는데, 만일 이들의 길이를 똑바로 잰다면 약 2.7m 가 된다. 각 세포마 다 저장된 정보가 단지 영기에 따라 인쇄된다면 이는 아마도 약 200 개의 맨해튼 전화번호부를 채우거나, 아니면 3 년치 일간신문에 기록 된 양과 갇다고 한다. 이런 뭉치가 바로 사람의 기본구조로, 사람과 같은 유기체는 이 구조로부터 단백질 합성과정을 통해서 만들어진다.
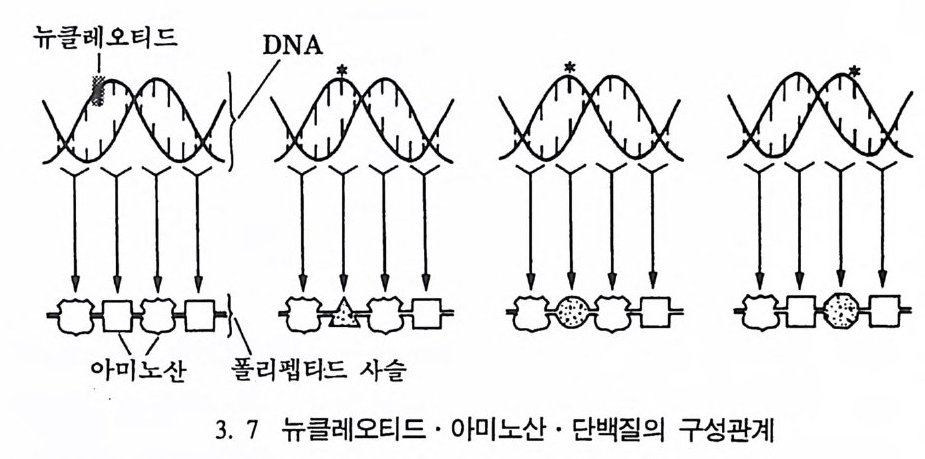 아미노산 폴리펩타드 사슬
아미노산 폴리펩타드 사슬
세포는 단백질을 만드는 공장이며, 단백질은 구조단백질과 효소단 백질로 구분된다. 구조단백질은 유기체의 구조를 결정하는 기능을 갖 고 있으며, 효소단백질은 체내에서 일어나는 화학작용들의 양울 조철 하는 기능을 갖고 있다. DNA 와 같이 단백질은 뉴클레오티드가 모여 서 이루어진 과분자 물질로서, 3 개의 뉴클레오티드가 모여서 하나의 아미노산을 만든다. 인간을 구성하는 아미노산은 모두 20 개가 있다 (그립 3. 7). 단백질은 큰 구조물질이다. 한 예로 헤모글로빈은 600 개의 아미노
표 3. I 아미노산의 종류와 약자 O 卜 □ I 노산 약자 o~ □ I 노산 약자 Alan ine Ala Leuc ine Leu Arg inine Arg Ly s in e Ly s Aspa rag ine Asn Meth io n ine Met Asp a rti c acid Asp Pheny la n ine Phe Cy s te n ine Cy s Prali ne Pro Gluta mic acid Glu Serin e Ser Gluta mine Gln Threon ine Thr Glyc in e Gly Tryp top h an Trp Hi st i dine • Hi s Ty ros in e Tyr Isoleucin e Lle Va line Val * H i s tidi n 타 어린이들에게 필수적인 아미노산이다.
산들로 이루어져 있다. 사람 몸은 50000 개의 단백질들로 구성되어 있 는데, 아미노산들의 배열 순서가 단백질의 특성(기능/모양)을 결정한 다. 단백질 합성은 핵 nucleus 에서부터 시작된다. 전달 RNA(mRNA : masseng er Ri bo nucleic Ac id : 이후 mRNA 로 부름)는 단백질을 만들 기 위해서 세포의 핵에 있는 DNA 로부터 정보를 읽는다. mRNA 는 이 정보를 가전 채 핵 밖으로 이동하여 세포에 있는 리보솜으로 간 다. 이 과정중에 리보솜에 있는 전환 RNA(tR NA : tra nsfe r RNA : 이후 t RNA 로 부름)는 아미노산에 붙어 있는데, mRNA 로부터 정보 롤 받아 이 아미노산을 리보솜으로 전환시킨다. 이때의 리보솜은 생 성되는 단백질의 일부이며, 여기서 염기를 나타내는 낱말인 코드는 모든 생물체에서 똑같이 쓰인다(그립 3. 8). DNA 의 기본은 쌍으로 이루어져 있고 마치 영어 단어처럼 3 개의
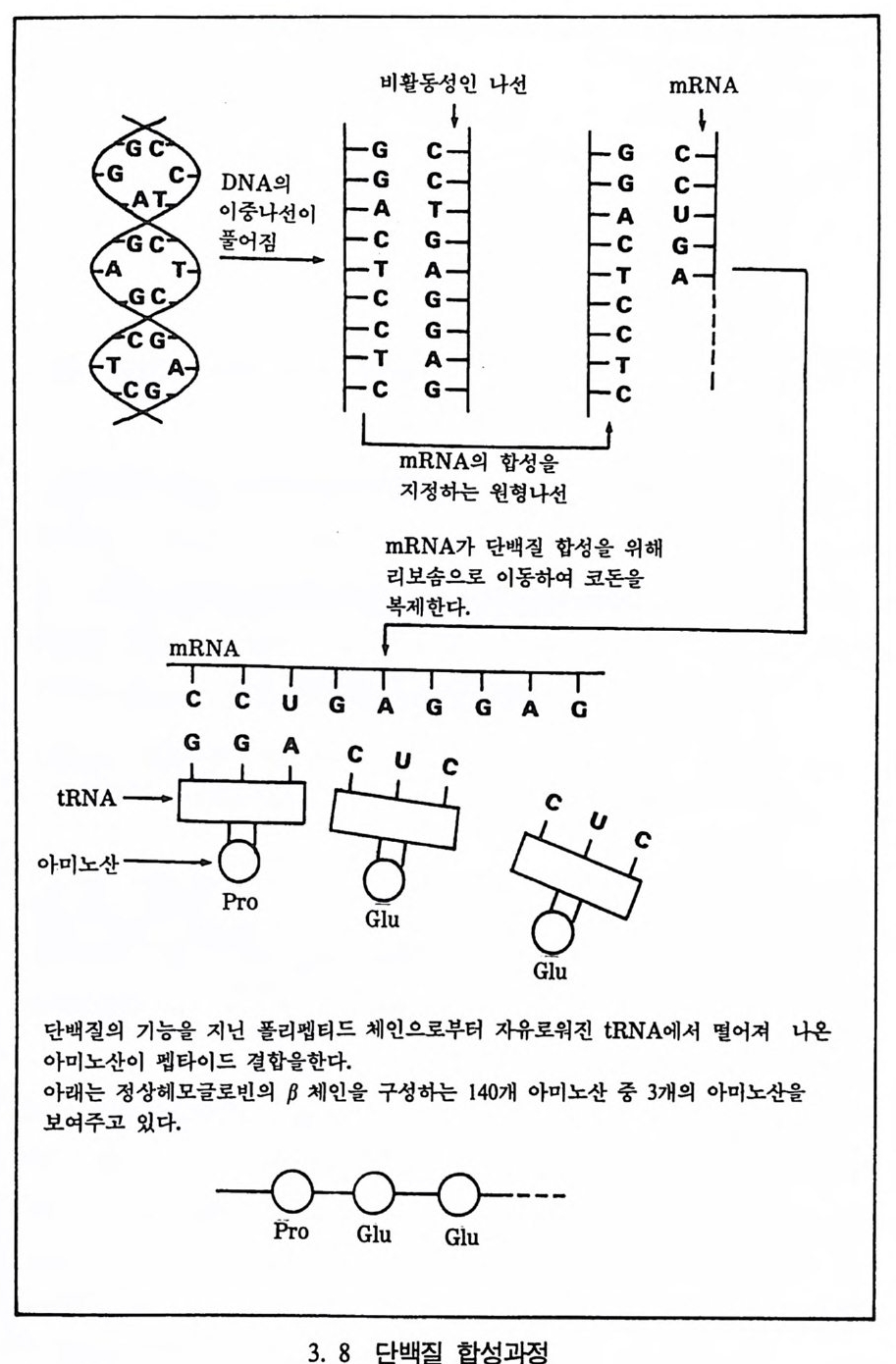 비활동성인 나선 mRNA
비활동성인 나선 mRNA
기본쌍이 모여 하나의 낱말을 만든다. 이 낱말을 코돈이라 부르는데 (예 : UGU, CGC 등등) 각 낱말들은 하나의 특별한 아미노산울 나타 낸다. 이런 체계를 〈트리플리트 코드 : tri p le t · code 〉라고 한다.
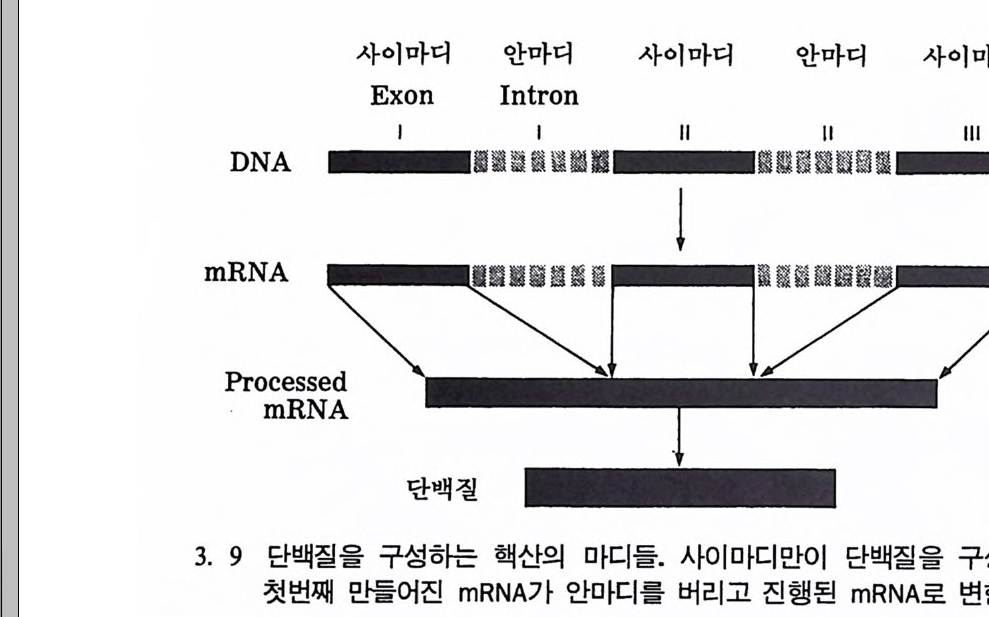 사이마디 안마디 사이마디 안마디 사이마디
사이마디 안마디 사이마디 안마디 사이마디
DNA 를 정확히 이해하는 것은 매우 중요하다. 왜냐하면 이룰 통해 서 어떤 윤리에 관한 일에 결정을 내려야 할 필요가 있기 때문이다. 처음에 DNA에 관한 많은 기초과학 연구는 바이러스와 박테리아를 대상으로 이루어졌다. 박테리아처럼 핵이 없는 세포들을 프로카리오 트라고 한다. 사람의 소화기관인 장에 살고 있는 박테리아인 E.col i를 바탕으로 많은 연구가 이루어져 있다. 이 박테리아는 500 만 개의 DNA 기본쌍을 가지고 있다. 초기의 유전코드에 관한 연구는 대개 이런 박테리아 같은 프로카리오트의 구조에 집중되었으며, 1977 년에 와서야 처음으로 핵을 가전 생명체인 유카리오트에 대한 연구가 시작 되었다. 유카리오트의 경우, DNA 분자 중 어떤 기본쌍은 단백질을
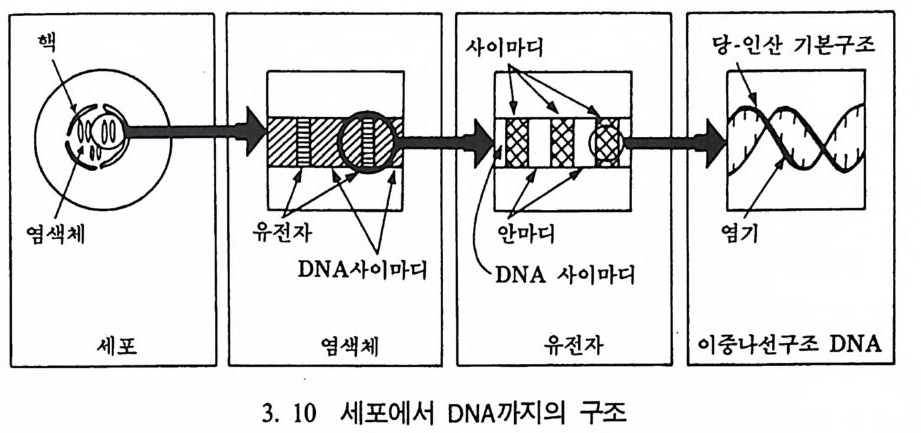 핵사이마디 당 · 인산 기본구조
핵사이마디 당 · 인산 기본구조
생산하지 않는디는 사실이 밝혀졌다. 이렇게 단백질을 생산하지 않는 DNA 의 마디를 안마디라고 하며, 안마디와 안마디 사이에 있는 마디 룰 사이마디라고 한다(그립 3. 9). 바로 이 DNA 의 사이마디들이 단 백질을 만든다. 따라서 유카리오트인 경우 유전자는 염색체의 일부분 이라고 할 수 있다(그립 3. 10). 유전자 재결합 모든 생명체에 있어서 위에 언급한 사실들은 모두가 같다. 그러나 서로 다른 〈종〉둘이 가지고 있는 정보는 서로 각기 다르다. 유전학의
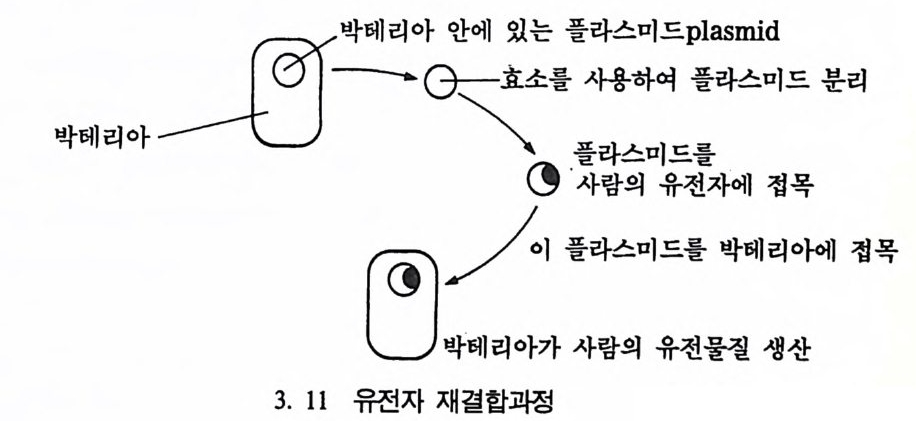 二二雲: :A라 ; ;1 여드p: a라s : nd] 드 분리
二二雲: :A라 ; ;1 여드p: a라s : nd] 드 분리
표 3. 2 1953 년부터 1993 년까지 발전된 주요 유전학의 연구 1953 윗슨과 크릭이 핵산의 구조를 확정하다. 1961 최초로 유전자 코드가 풀리다 . 1973 유전자 재합성을 위한 기술이 특허를 받으며, 바이오테크놀 러지 산업의 단계가 시작되다. 1975 미국 캘리포니아 아시로마에서 열린 국제회의에서 유전학 자와 생물학자들이 모여 유전자 재합성 실험에 관한 지침 을마련하다. 1976 최초의 유전공학 관계 회사인 〈지네테크〉가 샌프란시스코 남쪽 실리콘 밸리예 설립되다. 최초로 암울 유발하는 유전 자를확인하다. 1977 유전자 재합성 방법을 이용하여 박테리아로부터 인간의 호 르몬 생산에 성공하다. 같은 해에 DNA 의 유전 코드를 유 지하는 영기의 배열을 빨리 정할 수 있는 방법이 개발되다. 1978 인슐린을 만들어내는 유전자의 합성이 이루어지다. 1983 중합체 고리 반응법 the po lym e rase cha in reduc ti on 이 개 발되어 특별한 유전자를 빠른 시간 안에 대량생산할 수 있 게 되다• 1985 영국에서 처음으로 인간의 지문에서 유전상 특성을 찾는 방법이 범죄수사에 응용되다. 1987 유전공학의 기술로 재배된 토마토가 처음으로 야의에서 재 배되다. 1988 유전학 방법으로 생산된 동물에 대한 특허가 인정되다. 1990 모든 인류의 유전자 지도와 배열을 찾기 위해 게놈 연구를 위한 국립연구소 The Nati on al Cente r for Human Ge- nome Research 가 미국에 설립되다. 1993. 3 월 루 게 릭 질병 Lou Gehrig ' di seas 궁을 유발하는 유전 자가발견되다.
* 루 게 릭 질병은 미국의 뉴욕 양키즈 구단의 1 루수였다 사망한 루 게 릭 의 이름에서 따온 것으로 이 병은 운동신경계통에 마비가 일어나 3- 4 년 후면 전신이 굳어지면서 죽는 질환이다. 유명한 물리학자인 스티븐 호킹도 이 병에 걸린 환자로 살아 남은 경우이다. 발달은 다른 〈종〉들에서 DNA 를 추출하여 서로 결합시킬 수 있는데 이를 〈유전자 재결합〉이라고 한다. 1972 년에 처음으로 동물들의 DNA 롤 분리하여 디른 동물의 DNA 를 서로 결합할 수 있는 기술이 세상 에 알려지게 되었다. 이 기술의 발달로 사람의 유전자를- 추출하여 박 데리아에게로 옮길 수 있게 되었다(그림 3. 11). 이런 기술의 발달은 물리학에서 원자를 분리시킨 것보다 더 큰 영 향을 인류의 생활과 사고에 미칠 것으로 생각된다. 죽 이 기술을 가 지고 인간을 구성하고 있는 여러 정보들, 예를 들어 인간의 몸 구조 와 크기, 머리칼과 눈의 색깔둘이 왜 다르게 나타나는지를 설명할 수 있을 것이다• 뿐만 아니라 인간의 질병을 유발하는 원인과 지능 및 행위 등을 설명해 줄 수 있는 인자를 한 쌍씩 조사할 수 있으며, 이 들을 다론 동물들의 DNA 와 비교할 수 있다. 1953 년 윗슨과 크릭이 핵산의 구조를 확정한 이후 40 년이 지난 오 늘날 〈생명의 날〉에 관한 수수께끼가 일부 풀리기 시작하고 있다. 그 러나 아직 해결해야 할 많은 문제들이 남아 있다. 죽 유전자 무리들 이 서로 어울려지는 방법이나 식물세포와 동물세포가 서로 다른 이유 등이 그것이다. 부모의 얼굴을 닮은 자식의 얼굴은 어떻게 만들어지 며 사람의 경우 나이와 죽음을 어떻게 조철하는가 등등에 대한 답은 아마 곧 얻을 수 있으리라 생각된다. 유전자 재결합의 기술은 의학분야와 농업분야에서 아주 유용하게 쓰인다. 인슐린이 필요한 사람들을 위해 박테리아를 이용해서 인슐린 을 대량생산할 수 있으며, 인터페론 또한 대량생산되고 있다•
인류 역사상 유전자를 이용하는 방법은 전혀 새로운 발견이 아니 다. 이미 수천 년 전부터 가축과 식용식물들의 수확을 늘리기 위해서 이 방법이 이용되어 왔다. 인류는 알이 큰 옥수수를 보관함으로써 우 수한 종자를 보전하여 생산량을 증가시켜왔는데, 이러한 〈인위선택〉 이 한 방법이다.
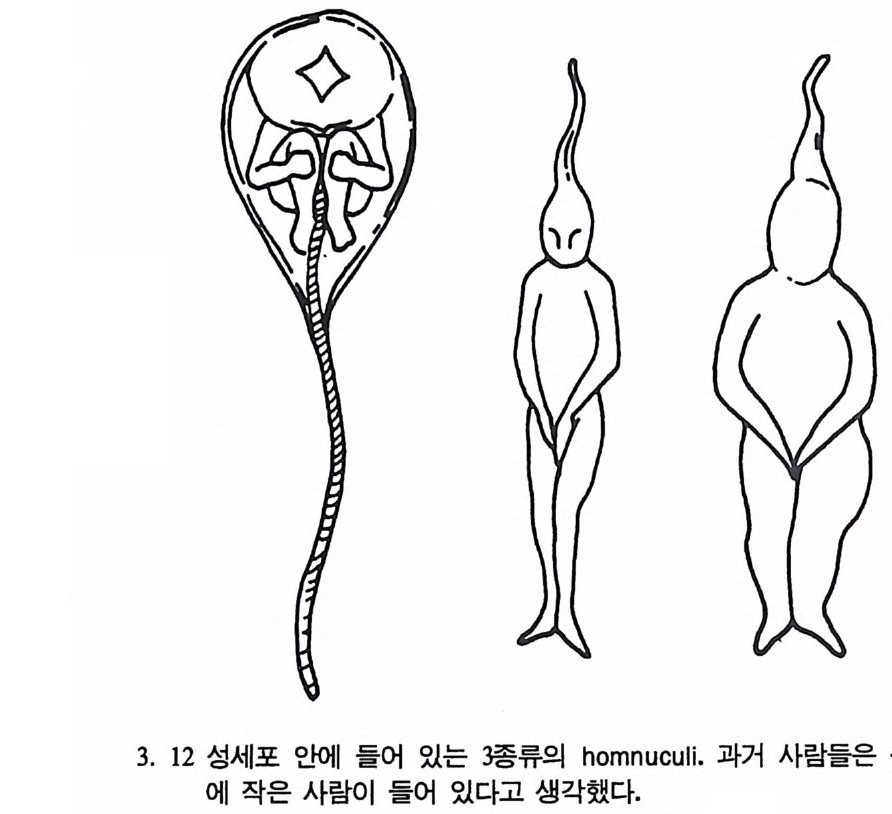 3. 12 성세포 안에 들어 있는 珪동류의 homnuculi. 과거 사람들은 성세포 안
3. 12 성세포 안에 들어 있는 珪동류의 homnuculi. 과거 사람들은 성세포 안
최근에 많은 해충들이 살충제에 대한 면역을 가지고 있음이 밝혀졌 다. 이것도 일종의 전화 과정이다. 해충제가 사용될 때마다 살아 남아 이 해충제에 대한 면역성을 다음 세대에 전하는 해충이 있다. 따라서 농부는 더욱 강하고, 더욱 비싼 해충제를 사용해야 하는데 DNA 합
성기술은 이런 문제를 해결할 수 있다. 한 예로 옥수수 뿌리에 살고 있는 어떤 박테리아는 옥수수 질병에 대항할 수 있는 유전자를 이미 가지고 있다. 이 박테리아를 DNA 합성으로 양산한다면 장차 해충제 가 필요없는 때가 울 것이다. 그러나 유전적으로 변화된 생명체가 환 경에 적응할 수 있는지에 대해서는 많은 논란이 있다. 1981 년에 미국 의 대법원이 인간이 새로운 생명을 만드는 일을 특허로 인정한다는 판결을 내린 이후 2000 년대까지 DNA 합성에 따른 시장가치는 150 - 1000 억 달러가 될 것으로 전망된다. 이의에도 DNA 연구로 최근까지 많은 발견들이 이루어졌다. 그리 스인들은 〈 어린아이의 성을 결정하는 것은 성교시 남자의 흥분 정도 에 달려 있다. 〉 고 믿어, 남자가 흥분할수록 잉태될 아이가 남자일 가 능성이 높다고 믿었다(그림 3. 12). 최근의 연구는 태아가 남자인지를 결정하는 Y 염색체의 위치를 알아냈다. 따라서 앞으로 · 인간은 아이의 성을 원하는 대로 조절할 수가 있을 것이다. 또 다른 연구로는 근육 영양실조를 일으키는 유전자와 이를 가지고 있는 단백질의 발견을 들 수 있다. 이것의 발견으로 이 병에 대한 치료가 곧 가능해질 것이다. 법의학에서는 DNA 정자 식별법을 이용하면 200 억 : 분의 l 의 오차로 강간범을 가려낼 수 있다. 한편으로 DNA 합성에 관한 연구는 과학상, 윤리상 그리고 도덕상 의 논란을 불러일으키고 있다. 과연 사람을 구성하고 있는 모든 유전 자가 밝혀질 수 있느냐 하는 점에서이다. 오늘날 우리는 단지 사람의 유전자 전체 중 0.2% 만 알고 있다. DNA 합성기술이 처음 개발되었 던 1970 년대에 사람들은 기술상의 실수로 어떤 재앙이 생기지 않을까 우려했었다. 그러나 그동안 행해전 수많은 실험 결과 어떤 해로운 일 도 아직까지는 일어나지 않고 있다. 환경 내에서 유전적으로 변화된 생명체에 대해서는 논란이 있는데, 어떤 생태학자는 변화된 생명체는 〈어떤 문제를 야기할 것이다〉라고
하는 반면 어떤 학자들은 〈 문제가 없다 〉 라고 주장하고 있다. 그러나 이 기술이 인체에 해로운 독약물을 생산할 가능성을 배제할 수 없다. 한 예로 1942 년 제 2 차 세계대전 중 독일은 생화학무기들을 생산하기 시작하였다 . 1969 년에 닉슨 대통령은 미국 내에서 모든 공격적인 생 화학무기의 생산을 금지시켰다. 그 후 1972 년에 미국과 구소련은 생 화학무기 생산금지조약협정에 조인하였으며, 많은 나라가 이 조약에 가입하였다. 최근의 소식들에 따르면 미 국방성은 유전적으로 질병을 유발하는 이들 무기 연구를 위해 많은 연구비를 과학자들에게 제공하 고 있다고 한다 . 미 국방성 관리들은 이 생화학무기들이 단지 방어용 이라고 하나, 어쨌든 과학자들은 선택의 기로에 서게 되었다. DNA 를 이용한 기초과학이 인류에게 미칠 영향에 대해 살펴보는 것은 이 연구가 필연적으로 가져오는 결과인 윤리적인 결정(예 : 생 명, 낙태 등)을 내려야 하는 경우가 있다는 점에서 매우 중요하다. 한 예로 헌팅돈병은 성인에게 일어나는 병으로, 신경계통에 이상이 생겨 급격히 회복할 수 없는 상태로 병이 진행되어 마침내는 뇌에 구멍을 만든다. 1983 년에 이 병을 일으키는 유전자를 발견한 이래, 어떤 사람 이 이 질병에 걸릴 수 있나를 알게 되었다. 1987 년에는 우울증의 원 인이 되는 유전자를 발견하였다. 과다 콜레스테롤 H yp ercholes t erole mi a 은 유전적인 병으로, 이는 핏속에 콜레스테롤 양을 증가시켜 심장 마비의 원인이 되기도 한다. 만일 위에 열거한 병들의 유전자를 가전 사람들이 있다고 가정하 자. 그들이 칙업을 갖기 위해 시험을 보았을 때, 만일 회사측이 이들 에 관한 유전적 자료들을 갖고 있다면 회사측은 이둘을 채용할 것인 가 하는 문제가 대두된다. 바로 DNA 를 연구하면서 대두되는 질문은 이런 종류의 정보를 어떻게 처리해야 하는가에 있다.
유전자란 무엇인가? 유전자란 단백질을 만들기 위해서 코드로 이루어전, 정보를 가진 DNA 의 연결체이다. 이 정보는 DNA 에 있는 염기가 나열된 순서에 따라 다르다. 한 쌍으로 이루어져 있는 염색체 내에 유전자가 있는 자리를 좌위 (복수는 로사이 loci, 단수인 경우 로커스 locus) 라 한다. 어 떤 한 유전자의 자리에 다른 정보가 들어가 변한 유전자를 대립유전 자라고 부른다. 만일 두 개의 같은 염색체가 한자리에 쌍을 이루고 있으면(예 : AA 또는 aa) 이때 염색체는 같은 질 Homo zyg ous 을 갖 고 있다고 하며, 만일 그 자리에 서로 다른 염색체가 쌍을 이루고 있 으면(예 : Aa) 다른 질 He t eroz yg ous 을 갖는다고 한다. 인간은 유전자 자리인 한 좌위에 2 개의 대립유전자를 가질 수 있 다. 같은 집단에서도 어떤 개체는 한 좌위에 많은 수의 디론· 성질의 유전인자가 있는 경우가 있다. 그런 예로 초파리를 둘 수 있는대, 초 파리는 눈의 색깔을 결정하는 좌위가 하나 있다. 어떤 초파리는 0~ 자리에 단지 2 개의 대립유전자가 있다. 그러나 초파리 집단 내에서는 모두 14 개의 서로 다론 유전인자가 나타난다. 이 14 개의 유전인자 중 하나가 항상 나타날 때 이를 우성인자라고 하며, 인자들은 표현형과 유전형의 형태로 나타난다. 표현형은 개개인들의 의관으로 보이는 신 체상의 특칭들을 말하며, 유전형은 개인들이 가지고 있는 전체의 유 전형질을나타낸다. 우성인자가 없는 단순한 예로는 MN 혈액체계를 둘 수 있는데, 이 는 껴의 다른 유전자가 하나의 로커스에 의해 조절되는 경우를 말한 다. 이때에 가능한 유전형은 MM, MN, NN의 세 가지가 있다. MM 과 NN 형을 같은형인자라 하며, MN 형을 다른형인자라고 부른다. 이 경우 표현형은 M, N, MN 의 세 가지로 나타난다. 이번에는 우성인자가 있는 경우를 살펴보자. L 과 l 이라는 대립유전
자가 있다고 할 때, 개개인들은 LL, LI 또는 ll 로 나타난다. 이때 같 은형 열성인자 (11) 를 가전 사람은 정상적인 모양의 발을 갖는 반면, 우성인자 (LL, LI) 를 가진 사람은 게발을 갖는다. 사람의 신체적인 특징들인 머리칼색, 피부색, 코의 크기, 보조개 등 은 여러 개의 대립형질을 가전 유전자들이 모여서 그 특칭을 나타낸 다. 만일 우리가 10 개의 다른 좌위들(로사이)을 가지고 있다고 가정 해 보자. 각각의 좌위(로커스)가 2 개의 대립유전자를- 가지고 있다고 할 때, 59049 개의 서로 다른 모양의 표현형이 나타날 수 있다. 이는 다시 말해서 멘델의 유전법칙은 한 집단 내에서 많은 변이의 가능성 을 나타내며, 이것이 바로 다윈이 말한 변이가 생기는 이유를 설명해 준다. MN 혈액형집단을 살펴봄으로써 위에서 언급한 내용을 이해해 보 자. MN 혈액체계의 경우 표현형으로는 M, MN, N 이, 유전형으로는 MM, MN, NN 이 나타난다• 부모가 아이룰 갖는다면 태어날 아이가 지닐 수 있는 MN 혈액형들은 아래와 같다 ; 부모가 모두 MM 형일 때 : 자식은 100% 가 MM 형 부모가 MM과 NN 형일 때 : 자식은 100% 가 MN 형 부모가 MM 과 MN형일 때 : 자식은 50% 가 MM 형이고 50% 가 MN 형 부모가 모두 NN 형일 때 : 자식은 100% 가 NN 형 만일 MM 과 MN 혈액형을 가전 부모가 아이를 4 명 갖고 있다면, 아이들이 가지고 있는 유전형은 어떤 형이라고 말할 수 있을까? 위에 서 살펴본 것처럼 우리는 단지 어떤 혈액형이 얼마만큼 나타날 수 있 는지 그 가능성만을 말할 수 있다.
집단유전학 : 집단의 유전과 진화의 요인 집단유전 지금까지 개인에 있어 유전이 어떻게 일어나는가를 이해하기 위해 세포와 염색체들의 구조와 기능을 살펴보았다. 그러나 전화를 이해하 기 위해서는 집단의 유전을 살펴보아야 한다. 진화라는 개념은 집단 에 적용되는 것이지 개인에게 적용되는 것이 아니기 때문이다. 진화 란 바로 집단이 환경의 다양성과 변화에 대해 적응하는 것을 뜻한다. 다시 말해 자연선택은 개인에게서 일어나나, 진화는 개인이 아닌 집 단에서 나타나는 유전상태의 빈도가 변화될 때를 의미한다. 따라서 한 집단의 유전상태는 그 집단 내에서 일어나는 유전자의 빈도를 조 사함으로써 알 수 있다. 한 집단의 유전상태에 관한 빈도를 알기 위 해서 MN 혈액형집단울 예로 들어보자. 가정 I) 무작위로 충북대학병원에 있는 500 명의 환자들의 기록을 조사해 본 결과 MN 표현형을 가전 환지들은 다음과 같았다 ; M 형이 180 명, MN 형이 240 명, N 형이 80 명으로 나타났다. 우리가 이 자료에서 알 수 있는 사실은 다음과 같다 .• 1. 이 자료의 유전자가 가지고 있는 메시지의 수 : 1000 2. M대립형질의 수 : 180 X 2 + 240 = 600, N 상대형질의 수 : 400 3. M대립형질의 빈도 : 600/ 1000 = 0.6 유전학자들은 한 집단에서 보이는 MN 혈액형 같은 유전상태의
빈도를 표시하기 위해서 p, q라는 기호를 사용하는데, 이때 p가 M 대립형질을 나타내면, q는 N 대립형질을 나타낸다. 따라서 위에서 본 집단의 MN 혈액형의 빈도는 p = 0.6, q = 0 .4로 표시된다. 가정 2) 아이를 갖고자 하는 사람 500 명이 어느 한 곳에서 만 나 짝을 지었다면 이때 서로 다른 대립형질이 정자와 난자에서 나 타나는데, 그 결과 자식들에서 나타나는- 빈도는 아래와 같이 계산 된다.
M(0.6) N(-0.4 ) M(0.6) 0.3 6 0.24 지식들의 빈도 N(0.4 ) 0.24 0.1 6
따라서 만일 짝을 이룬 사람 중 100 명이 임신을 했을 경우 자식에 서 나타날 수 있는 빈도는 MM 형이 36 명, MN형이 48 명, NN 형이 16 명으로 계산되며, 만일 500 명이 임신을 했다고 가정한다면 자식에 서 나타나는 MM형은 180 명이고, MN형이 240 명, 그리고 NN 형이 80 명으로 나타난다. 이 결과는 위에서와 똑같은 빈도로 나타난다. 1908 년에 영국의 수학자와 독일의 물리학자가 한 집단 내에서 유전 인자의 빈도가 나타나는 관계를 관찰하여 〈하디-와인버그 평형 Har- dy - Wein b erg E q u ili br i um 〉이라는 등식을 만들었다. 하디-와인버그 는 자신들의 동식을 p2 +2p q+ q2 = 1 로 나타내었는데, 여기서 p2 = AA 를 의미하고 2 pq는, Aa 를 그리고 q 2 는 aa 를 나타낸다. 그러나 실제로 이 등식은 평형이 아니라 정체를 나타내며, 다음과 같은 조건 내에서만 등식의 성립이 가능하다 ; 첫째, 한 집단 내에서 다른 유전형이 생기지 않아야 한다. 둘째, 그 집단은 커야 한다.
셋째, 집단과 집단 사이에 인구의 이동이 없어야 한다. 넷째, 한 집단 내에서 모든 유전형과 표현형은 갇은 빈도로 나타나 야한다. 위에서 언급된 4 개의 조건들의 성립은 한 집단 내에서 〈 전화 〉 를 인정하지 않을 때에 가능하지만 실제로는 불가능하다. 그러나 이 조 건들은 바로 집단 내에서 〈전화〉가 왜, 어떻게 일어나는지를 설명해 준다. 진화의 요인 현대 진화론에서는 돌연변이 Mu t a ti on 와 유전자 표류 Gene ti c Drif t, 유전자 이동 Gene Flow, 자연선택 Natu ral Selec ti on 을 전화의 요인으 로 규정하고 있는데, 바로 이 4 개의 전화요인들이 하디 - 와인버그 등 식을 성립시키는 4 개의 가정에 변회를- 일으키는 것이다. 등식의 가정 에 변화를 주는 전화요인들은 아래와 같다. 첫번째 요인은 돌연변이이다. 돌연변이는 유전 메시지 자신의 변화 로, 염색체에서나 분자구조에서 일어난다. 다시 말해 돌연변이는 DNA 의 기본구조인 뉴클레오티드를 이루는 영기가 삭제, 첨가 또는 교체 될 때를 말한다; 예) 본래의 구조 : CAT TAG GAT TAA G 가 삭제 : CAT TAG ATT TAA G 가 첨가 : CAT TAG GGA TTA 교 체 : CAT TAG GTT TAA 위의 예들은 단일영기가 어떻게 변하는가롤 보여준다. 이 세 종류 의 돌연변이는 3 쌍의 뉴클레오티드에 의해 만들어지는 아미노산에 변 화를 일으키며, 이어서 변화된 아미노산은 단백질을 변화시킨다. 그러
나 이런 종류의 변화는 자주 일어나지 않으며, 이런 변화가 다음 세 대에 나타나기 위해서는 접합자를 형성하는 일단의 세포에서 변화가 일어나야 한다. 만일 이런 돌연변이가 우리의 엄지손가락에 있는 세 포에서 생긴다면 이 변화는 다음 세대에 영향을 미치지 않는다. 실제 로 돌연변이가 일어날 확률은 적으며, 이렇게 변화를 일으키는 유전 자를 돌연변이형 유전자라고 하는데, 인간의 경우 약 100 만분의 1 의 확률로 접합자에게서 일어난다. 돌연변이는 우성과 열성에서 모두 나 타는데 그 예는 다음과 같다; 예) 돌연변이가 우성인 경우 돌연변이가 열성인 경우 영양실조 색소결핍 Chondody st r o p h y Albin ism @广 ~□ AA @回回?@ 0AA □As. 연:〈 □k 자연돌 자연돌연변이 0 □ 0 ■ AA Aa Aa aa 영양실조 발생 색소 결핍 발생 돌연변이는 실제로 상당히 천천히 일어나기 때문에 대립형질 빈도 수에 영향을 미칠 만큼 큰 원인이 되지 않는다. 유전학자들은 한때 돌연변이 중 많은 수가 오랫동안 주위환경에 잘 적응해 오던, 아주 복 잡한 구조를 가전 생명체에게 돌연한 변화를 가져오기 때문에 생명체 자신들에게 해롭다고 생각했었다. 그러나 실제로 많은 돌연변이는 중 성이며, 극히 소수의 변이들만이 생물 자신에게 유익하다는 사실이 밝혀졌다. 이렇게 모든 생명체에 변이를 일으키는 요인이 돌연변이라
는 사실을 다윈이 살던 시대에는 알지 못하였다. 진화의 두번째 요인은 유전자 표류로, 이는 작은 집단에서 보이 는 〈 모든 유전자 〉 가 어떤 기회에 변화되는 경우를 말한다. 한 예로 두 개의 크고 작은 규모의 집단이 있는데 이때 큰 집단은 50000 명의 주민으로 이루어져 있으며, 작은 집단은 50 명의 주민으로 이루어져 있다고 가정해 보자. 작은 집단의 MN 혈액빈도를 조사해 본 결과 49 명이 MM 혈액형으로 판명되고, 나머지 1 명은 MN 혈액형 울 가지고 있으며, 큰 집단의 경우 49000 명이 MM 혈액형을, 그리고 1000 명이 MN 혈액형을 가지고 있다고 했을 때, 이 두 집단의 경우 N 대립형질의 빈도수는 0.01 이다. 만일 첫번째 집단에 속하는 MN 형의 사람이 사고로 자식을 낳지 못했을 때 다음 세대에 나타나는 N 대립형질의 빈도는 0 이다. 그러나 커다란 집단 내에서 한 명의 MN 혈액형을 지닌 사람에게 사고가 났 울 때, 이는 집단 혈액형의 구성에 큰 영향을 주지 않는다. 이렇게 작 은 집단 내에서 우연히 유전자 구성의 비율에 변화가 생길 때 이를 유전자표류라고한다. 유전자 표류의 다른 형태로 분가유전 Founder E ff ec t을 들 수 있는 데, 분가유전은 작은 집단이 다른 장소에서 새로운 집단을 이루었을 때 일어난다. 이 작은 집단은 단순히 주어전 기회 때문에 커다란 어 미집단과는 아주 다른 유전인자의 빈도를 갖게 된다. 예로 50000 명의 주민으로 구성된 집단에서 50 명이 새 지역으로 이주했다고 가정했을 때, 만일 어떤 기회에 이들이 모두 MM 혈액형을 갖게 된다면 이 분 가집단은 N 대립형질이 전혀 없는 새로운 집단을 형성한다. 전화의 세번째 요인으로 유전자 이동이 있다. 여기서 유전자 이동 은 두 집단 사이에의 이동을 말한다. 예로 A 집단에는 집단 구성원이 100% MM 혈액형을 가지고 있고, E 십단에서는 MN 혈액형이 100% 로 나타난다고 가정했을 때, 만일 A 집단 구성원의 일부가 B 집단으로 이
주했다고 한다면 이때 B 집단에 N 의 빈도수는 떨어져 변화를· 가져온 다. 끝으로 생명의 전화요인으로 자연선택을 꼽는데, 자연선택이 바로 진화의 추전요인이다. 자연선택은 어떤 개체가 환경에 더 잘 적응했 울 때 더 많은 자식을 가질 수 있음을 뜻한다. 다시 말해서 유전자 빈도수의 변화를 의미한다. 자연선택의 예 지금까지 생명체 진화를 일으키는 4 개 요인들을 살펴보았다. 우리 는 전화에 관한 고전적인 예로 페이퍼 나방울 둘 수 있다. 영국에서 살고 있는 이 나방은 수백 년간 흰색에 검은 반점의 날개를 가지고 있었다. 이는 Lig h t Bark 나무에서 살기에 알맞은 보호색으로대 이 지 역에서 사는 새들은 이들을 발견하기가 쉽지 않았다. 이 나방 중 돌 연변이에 의해 검은 날개를 가진 나방이 소수 생겨났다. 그런데 이둘 은 새들에게 쉽게 발견되어 잡아먹히기 때문에 다음 세대로 그들의 유전자를 전할 기회가 매우 적었다. 그러나 영국의 산업혁명 시기중 공장의 연료로 사용하는 석탄에서 나온 매연으로 인해 Lig h t Bark 나무의 색깔이 검게 변하였다. 이런 산업에 따른 검은색화i ndus tr i al mela ni sm 에 의한 자연선택의 결과로 인해 흰색에 검은 반점을 지닌 나방이 새들의 눈에 잘 띠게 되었고, 점차적으로 이들의 수가 감소되 었다 . 환경의 변화가 나방의 적응도에 변화를 가져온 것이다. 자연선택에 대응하는 뜻으로 인위선택이 있는데, 이는 인간이 어떤 생명체를 죽이기 위해서 사용하는 화학품에 대해 내성이 생길 때 일 어난다. 인위선택의 좋은 예로 바퀴벌레를 둘 수 있다. 바퀴벌레는 환 경에 적웅도가 아주 높은 생명체로 암수 한 쌍은 일 년에 약 100 만 마리의 새끼를 낳는다. 미국의 저임금 가정 중 97.5% 가 한 가정당 최
소한 160 마리 이상, 평균 33600 마리의 바퀴벌레를 가지고 있다고 보 고되고 있다. 쓰레기통 옆, 부엌, 식당 데이불, 그리고 가전제품 등에 살고 있는 이들을 잡기 위해 많은 약품들이 개발되고 있으나 이 약품 들에 대한 내성이 생긴 새로운 바퀴벌레가 생겨나는데, 이를 자연선 택에 비해 인위선택이라고 한다. 이렇게 인간이 만든 약품에 내성이 생겨 적응도가 높아졌을 때 이를 인위선택이 일어났다고 한다. 자연선택의 또 다른 예 : 어떻게 이해하나? 전화의 추전요인인 자연선택에 대해 좀더 살펴보기로 하자. 유전자 자리에 두 개의 대립형질이 있다고 가정했을 때, 이 대립형질 중의 하나는 〈우성〉형질을 띠고 있고 다른 하나는 〈열성〉형질을 지니고 있다. 이때 대립형질을 A, 터료 나타낸다면, 가능한 유전형은 AA 와 Aa, 그리고 aa7} 된다. 만일 A 형질이 개인에게 이점이 있다고 한다면, 1000 개의 A 표현형 울 가진 사람과 999 개의 a 표현형을 가전 사람과의 재생산을 비교해 볼 때 A 형질의 빈도가 0.00001 에서 0.99 까지 이르는 데는 2340M] 대 가 걸린다고 한다. 인간의 경우 이렇게 빈도가 변하는 데 걸리는 세 대수로 볼 때 변화의 속도는 매우 느리다고 할 수 있다. 한편 우리가 우리의 조상을 이야기할 때는 대개가 l 伊]대 전후를 말한다. 인간에 있어서 2340Ml 대는 약 50 만 년에 해당된다. 물론 2340 0Ai]대는 진화 라는 개념에서 볼 때 오랜 기간이 아닐 수 있으나(예 : 바퀴벌레), 인 간에게 있어서는 긴 시간이라 할 수 있다. 자연선택은 생물의 전화에 지대한 영향을 미친다. 자연선택이 인간 의 전화에 미치는 예로는 우유 소화와 평형다형현상 Balanced Poly- mor phi sm 의 실례인 주머니섬유증 C y s ti c F i bros i s 과 낫꼴(경상)적혈 구 빈혈증 S i ckle-Cell Ane mi a 이 대표적으로 인용된다.
우유소화 우유는 4% 의 지방과 3% 의 단백질, 5% 의 탄수화물, l% 의 미네랄, 그리고 87% 의 물로 구성되어 있다. 우유에 함유되어 있는 탄수화물 인 락토제는 사람 몸 속에 있는 락타제라는 효소에 의해 분해된다. 사람에게는 락타제를 생산하는 유전자가 있는데, 아이가 세 살 정도 가 되어 젖을 멜 때에는 정상으로 락타제 생산이 중단된다. 이것은 아이 몸에 있는 조절유전자가 구조유전자를 조절해서 생겨나는 것으 로 더 이상 이 효소는 생산되지 않는다.
표 3. 3 집단에 따른 우유의 락토제를 소화시킬 수 없는 빈도수 미국백인 2-19 핀란드사람 18 스위스사람 12 스웨덴사람 4 미국흑인 70-77 아프리카이보족 99 아프리카반투족 90 플라니족 22 타이 사람 99 미국동양계 95-1 00 오스트레일리아원주민 85 * Leher and Libb er, 1976 에서 인용.
만일 어론의 몸에 있는 조절유전자가 구조유전자로 하여금 더 이상 락타제를 만들지 못하게 하면 어떻게 어른들이 우유를 마실 수 있을 까? 세상의 많은 어른들은 우유를 소화시킬 수 없으니 만일 우유를
마셨을 때는 위복통과 설사와 위에 가스가 차는 증세 등으로 고생하 게 된다. 그러나 실제로 우유는 사람에게 중요한 음식으로 많은 사람 둘이 마시고 있다. 다만 어느 집단은 우유를 마시는 데 많은 문제를 지니고 있다(표 3. 3). 지구상에 살고 있는 인간집단에서 보이는 이런 현상을 어떻게 설명 할 수 있을까? 약 1 만 5 천 년 전 인간이 처음으로 우유를 생산하는 동물들을 사육하기 시작하였는데, 그 이전에 살던 사람들은 우유를 마실 기회가 없었기 때문에 몸 안에서 락타제를 만들어낼 필요가 없 었다 . 그러나 우유를 생산해 내는 동물을 가축화하면서 점차 사람들 에게는 어른이 된 후에도 락타제를 생산해야 하는 필요성이 생기고, 따라서 조절유전자에 돌연변이가 생겼는데, 특히 이런 변이는 목축을 하는 집단에게는 절대적으로 필요하였다 . 따라서 목축을 하며 살아가 는 집단의 어른들도 우유를 마실 수 있게 되었다. 어떤 집단이 락타제에 적응한다는 사실은 생물학뿐만 아니라 정치 • 문화상으로도 매우 중요하다. 예로 〈우유는 몸에 좋다〉는 구호는 중 국 사람에게는 좋은 구호가 아니다. 왜냐하면 80% 에 달하는 중국 사 람은 락토제를 분해시킬 수 없기 때문이며, 이런 구호는 유럽 중심적 인 사고의 발상이라고 할 수 있다. 실제로 어른이 우유를 마실 수 있 다는 것 자체가 비정상적인 행위이기 때문이다. 만일 우리가 에티오피아의 굶어 죽어가는 빈민에게 구호물자로 락 토제가 들어 있는 우유를 보낸다면, 그들은 설사와 복통으로 고생을 할 것이며, 아마도 그들은 우리가 도와주려는 것이 아니라 독약을 넣 어 보낸 것으로 생각할 것이다. 따라서 락토제에 면역이 있는지의 여 부는 정치상 그리고 생물학상으로 중요한 의미가 있다. 평형다형현상의 예 : 주머니섬유증Cy s ti c Fi bros i s 과 낫꼴(겸상)적혈 구 빈혈증 Sic k le-C ell Anemi a
표현형 단계에서 하나의 좌위에 두 개의 대립형질이 있을 때 자연 선택이 어떻게 작용하는지를 살펴보자. 아래의 예는 한 좌위에 두 개 의 대립형질이 있는 경우로, 서로 우성이 없는 두 개 인자가 공우성 Codomi nan t을 띠고 있는 경우이다. 아래 보기 중 1 번의 상황은 실제로 일어나지 않는다. 한 집단 내에 서 두 개의 대립형질을 유지하기 위해서는 5 번처럼 집단 내에서 이형 접합체 He t eroz yg o t e 를 선호해야만 가능하다. 이렇게 한 집단 내에서 이형의 형질을 유지하는 경우를 평형다형현상이라고 한다. 평형다형 현상은 한 집단 내에서 두 개 또는 그 이상의 유전인자가 실제로 나 타나는 것으로 그 빈도가 1-2% 보다 더 나타나는 현상을 말한다 . 1 개의 유전자 자리에 2 개의 대립형질인 D 와 I 가 있을 때, -공우성 1. 11 I □ □ □ 변화가없음 2. 11 ■ □ - □ ID □□ ■ 一 4. ■ I - -DD D 나 . 의 손실 양쪽 형질빈도의 초기 평형빈도에 따라 결정 - -선택인자 위의 경우, 3 개의 유전형과 2~3 개의 표현형이 있다. 이중 3 개의 표현형은 공우성이며, 2 개는 우성을 나타낸다.
사람의 경우 많은 유전자 자리는 여러 형태의 대립형질 인자가 있 는데(p ol ym or phi c : 多型), 이는 세계 도처에 평형다형현상이 있음을 뜻하며, 여기서는 대표적인 실례로 주머니섬유증과 낫꼴(경상)적혈구 빈혈증을 살펴본다. 주머니섬유증 이 병은 유전성 질환으로, 사람에게 있어 폐에 점액이 지나치게 분 비되어 호흡곤란과 세균감염으로 결국에는 호흡을 할 수 없게 된다. 이 병은 실제로 적응이란 면에서는 실패를 의미하며, 이 병에 걸린 사람은 3~-ll 정도밖에 살지 못하게 된다. 이 질병을 유전으로 가전 사람은 같은 형의 열성인자 CC 를 가지고 있는데, 동양인과 흑인에게는 별로 나타나지 않으나 백인에게는 자주 나타나 백인 어린아이의 경우 태어날 때 2500 명에 한 명꼴로 이 질병 을 보인다. 이는 빈도수가 2% 이상으로 아주 높다는 사실을 알 수 있으며, 선택이 일어났다면 c 인자는 집단 내에서 줄어들어야만 한다. 그러나 실제로 c 인자는 높게 나타나고 있다. 이를 어떻게 설명할 수 있울까? 1987 년에 발표된 한 연구는 Cc 이형집합체가 폐결핵을 방지할 수 있는 단백질을 생산하는 데 도움을 준다고 보고하고 있다. 폐결핵은 박데리아 질환으로 15 세기 이래로 유럽 인구수의 변화에 막대한 영향 을 주어왔는데, 위의 연구에 따르면 Cc 이형집합는 CC 동형집합체보 다 폐결핵에 저항력이 크며, 따라서 주머니섬유증세가 백인 집단에서 높게 나타나는 이유를 설명해 준다. 낫꼴(겸상)적혈구 빈렬증 피는 실제로 액체 속에 있는 작은 입자들로, 혈액은 피가 흐르는 것을 멈추게 하는 혈소판과 백혈구, 적혈구로 이루어져 있다. 이때 백
혈구 하나에 적혈구가 500 개의 비율로 있으며 lm 떡 혈액에는 약 500 만 개의 적혈구가 있다. 매 드로마다 약 200 만 개의 적혈구가 간장 과 비장에서 파괴되며, 그 결과 필요한 양의 적혈구는 골수로부터 공 급받는다. 적혈구는 접시 모양으로 가운데가 움푹 파여 있다. 적혈구 안쪽에 는 약 2, 3 백만 개의 헤모글로빈이 있는데, 헤모글로빈은 적혈구의
 정상적인 사람의 헤모글로빈의 낫꼴(겸상)적혈구 빈혈중에 걸린
정상적인 사람의 헤모글로빈의 낫꼴(겸상)적혈구 빈혈중에 걸린
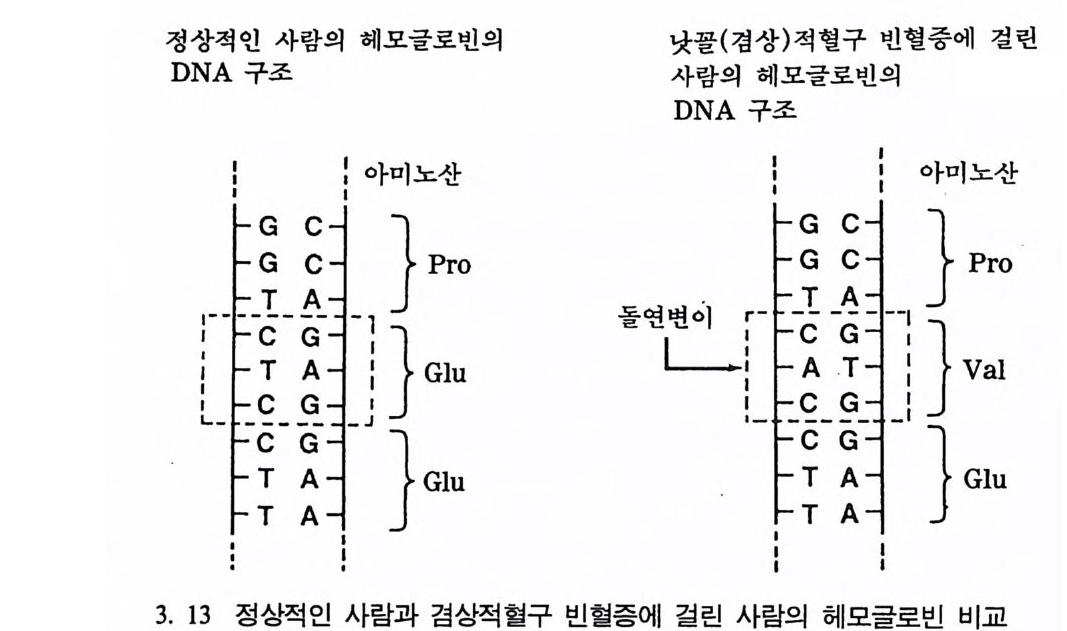
90-95% 를 형성하는 단백질로 아주 복잡한 구조의 분자물이다. 헤모글로빈은 다른 단백질과 같이 단백질 합성에 의해 만들어진다. 헤모글로빈을 가지고 있는 DNA 의 부분은 4 번 염색체 자리에 있는데 약 438 개의 기본영기로 이루어져 있다. 낫꼴(경상)적혈구 빈혈증을 일으키는 헤모글로빈은 정상의 헤모글로빈을 만들기 위해 글루타민산 이 들어가야 할 자리에 염기 자리의 발린 Va li ne 이 대신 들어감으로 써 생겨난 단백질인데, 그 영향은 적혈구에서 나타난다(그립 3. 13).
적혈구는 형체가 없어 물처럼 자유롭게 순환기관을 돌아 다닐 수 있다. 이때 적혈구에 있는 헤모글로빈은 산소를 몸의 각 기관과 조직 둘 그리고 뇌에 공급하는 역할을 하며, 또한 몸의 조직에서 생긴 탄 산가스를 폐로 운반한다. 헤모글로빈은 하나는 정상 그리고 또 다론 하나는 비정상인 27H 의 대립형질을 가지고 있는데, 같은 형의 비정상인 대립형질을 가전 사 람의 경우, 적혈구가 산소를 잃을 때 다수의 적혈구가 반달 모양으로 변하는데 바로 이런 모양의 적혈구가 질병의 원인이 된다. 이렇게 모 양이 바뀐 적혈구들은 핏속에서 잘 움직이지 못하고 서로 엉켜 핏줄 을 막히게 한다. 따라서 살아 있는 몸의 조직들은 산소를 공급받지 못하며 많은 적혈구들이 파괴된다. 이렇게 해서 바로 빈혈증이라고 부르는 증상이 나타나며, 이런 질환을 가전 사람은 몸이 무기력해지 며 점차 쇠약해지게 된다. 두 대립형질은 A 와 S 라고 표시하는데, A 는 정상이며 S 는 비정상 인 대립형질이다. 이때 가능한 유전형은 AA, AS 그리고 SS 가 되며, 대문자는 바로 이들이 공우성임을 표시한다. 이때 AS 를 가진 사람은 정상이나 산소가 부족하며, SS 를 가전 사람은 비장 등의 이상으로 고 생을 하다가 서구 이의의 세계에서는 대개가 결혼 이전에 사망한다. 만일 그들이 결혼하여 자식을 낳기 전에 사망을 하면 대립형질의 빈 도는 급격히 떨어져 마침내 빈도는 0 이 되어야 한다. 그러나 아직도 서아프리카에 있는 부족들은 S 인자를 가전 사람의 빈도수가 0.2 나 되 는 점에서 볼 때, S 형질 인자를 가진 사람이 어떤 점에서는 정상인자 롤 가전 사람보다 유리한 접이 있다고 여겨졌으며 이런 사실은 오래 전부터 알려져왔다. 이렇게 S 형질 인자가 높은 빈도를 나타내는 것은 아주 높은 빈도의 돌연변이가 원인일지도 모른다고 학자들은 한동안 믿어왔었으나, 돌연변이에 의해서라고 하기에는 빈도수가 너무 높았다. 1954 년 영국 사람인 엘리슨이 S 형질의 분포지역을 조사한 결과, 이
 /
/
 `?
`?
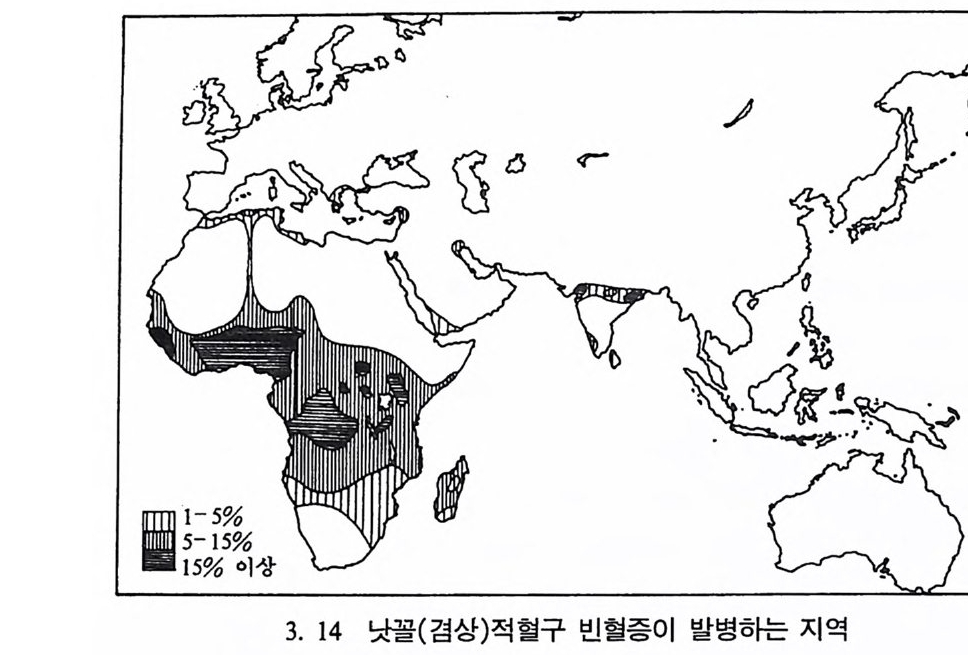
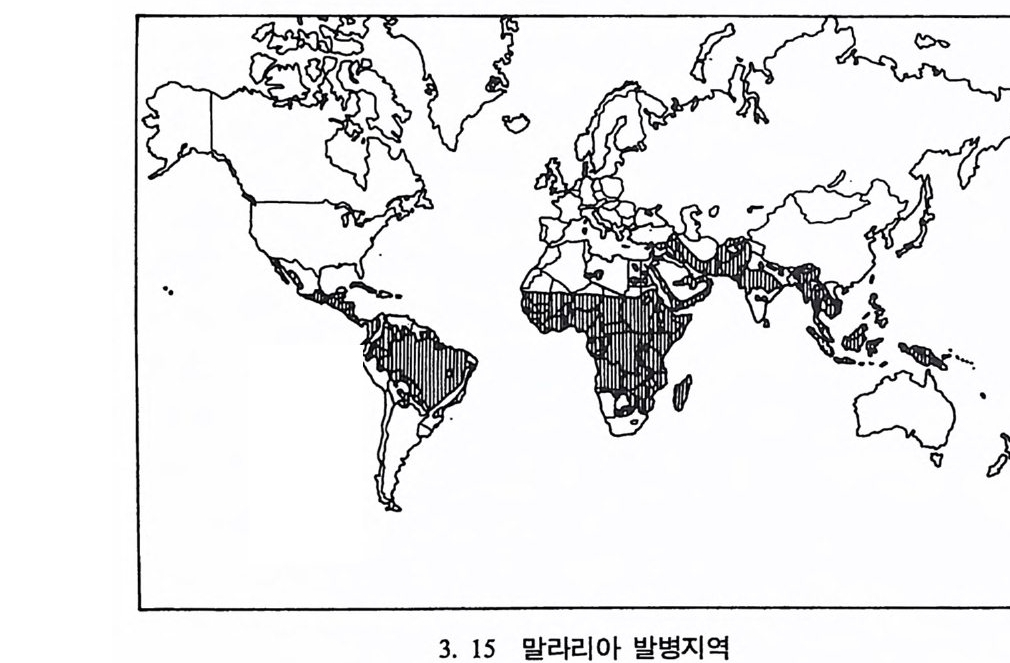
형질은 중앙아프리카지역에 주로 분포되어 있는데 이런 분포는 바로 말라리아가 퍼져 있는 지역과 일치된다는 사실을 알아냈다.(그립 3. 14, 3. 15) 오늘날 지구상에서는 약 20 억 명의 사람이 말라리아에 노 출되어 있으며, 일년에 3 억 정도의 사람이 말라리아에 감염되어 그 중 2, 3 백만 명의 사람이 매년 죽어간다. 말라리아의 감염은 4 개의 풀라스모다움 기생충 중 하나를 가지고 있는 감비 모기 A. g amb i ae 가 사람을 물었을 때에 발병한다. 스포로자 이트 Sp oroz it e 라고 불리는 이 기생충이 모기의 주둥이로부터 핏속으 로 들어가서 약 20¾ ol 지나면 간에 도달하게 되는대, 간은 사람 몸 의 면역체계의 보호를 받지 않는다. 여기서 이 기생충은 분열을 시작 하여 메로조이트 Merozo it e 라는 형태로 변한다. 마침내 세포가 파열되 고 이 메로조이트가 붉은 피톨 속으로 침입하는데, 이때까지 걸리는 시간은 약 2{µ, 순 정도이다. 이들은 핏속에서 계속 그 숫자가 늘어나며 간헐적으로 붉은 피톨을 파괴시키고 계속해서 다론 붉은 피톨 속으로 침투하는데, 말라리아에 걸렸을 때 생기는 오한과 고열은 바로 이 붉 은 피톨들이 파괴될 때 생기는 것으로 여겨진다. 오한과 고열은 말라 리아에 걸린 사람이 빈혈과 뇌에 손상을 일으키거나 아니면 신장이 파괴되어 죽을 때까지 계속되며, 개인의 면역체계나 약을 투여하여 치료가 되어야만 오한과 고열이 멈춘다. 학자들은 낫꼴(검상)적혈구 빈혈증과 말라리아 사이에는 어떤 연관 이 있으리라고 생각하여 15 명의 정상인자를 가진 사람과 15 명의 비정 상인자를 가전 30 명의 지원자들에게 말라리아충을 주사하는 실험을 하였다. 그 결과 정상인자를 가진 사람의 경우 15 명의 지원자 중 14 명이 말라리아에 감염되었고, 비정상인자를 가진 사람은 15 명 중 단 지 2 명만이 감염되었다는 결과를 얻었다. 다시 말해 S 형질이 말라리 아에 대해 저항성을 가지고 있음을- 뜻하는 것이다. 리빙스턴은 인류의 진화에 문화가 미치는 효과를 살펴보았다. 그는
어떤 지역에서는 S 형질의 빈도가 0.2 에 달하며 어떤 지역에서는 전혀 나타나지 않는다는 사실에 주목하여 환경이 이 형질의 발생과 관련이 있다고 생각하게 되었으며, 말라리아의 75% 는 단일 모기에 의해 발 병한다는 사실에 관심을 집중하였다. 곤충학자들에 따르면 말라리아 를 옮기는 모기는 그늘전 곳의 흐르는 물에서는 살 수 없다고 한다. 그러나 햇빛이 비치는 곳에서 자러는· 풀과 함께 고여 있는 물에서는 잘산다고한다. 리빙스턴은 언어학 자료들과 고고학 자료둘을 살펴본 끝에 2000 내 지 3000 년 전에 소위 〈스위던 농업 Sw idd en Ag ri c u ltu re 또는 화전농 업 Slash and Burn Ar g r i cul t ure 〉이 낫꼴예상)적혈구 빈혈증이 높게 나타나는 지역에 널리 퍼져 있다는 것을 알게 되었다. 이런 농업방식 은 열대 삼림지대의 생태계에 결정적인 변화를 가져왔다. 이런 지역 은 처음 몇 년 동안은 농사를 짓기에 아주 좋은 장소였으나 곧 흙 속 에 있는 유기질이 다 없어져 척박한 땅으로 변한다. 물은 이렇게 굳 어전 토양의 표면에 고이고, 나무가 없어 그늘이 생겨나지 않는 땅 위로 햇빛이 비추어 모기가 살기에 알맞는 환경이 되었다. 이런 일들과 더불어 농업은 채집이나 사냥 때보다 더 많은 인력을 필요로 하며 인구의 증가를 수반한다. 농업경제 속에서 늘어난 인구 는 정착생활을 하게 되며, 따라서 마을을 형성하게 되었다. 마울은 풀 로 이엉울 덮은 구조물로 이루어졌는데, 이렇게 풀로 덮은 지봉은 바 로 모기들이 쉬기에 더없이 좋은 장소가 되었다. 그 결과 모기의 서 식처가 된 이들 지역은 일반적으로 말라리아가 들끓게 되며, AS 이형 형질인자를 가전 사람이 이런 지역에서는 더 살기가 좋다고 가늠된 다. 최근에 농업이 소개된 지역에서는 S 형질이 매우 낮은 빈도로 나 타나며, 정착마울이 아닌 이동하는 주거형태를 지닌 피그미족의 거주 구조는 모기가 살기에 적당한 환경이 아니기에 S 형질이 전혀 나타나 지 않는다는 사실은 낫꼴(경상)적혈구 빈혈증이 일종의 환경에 따른
질병임을 알 수 있게 해준다. 비록 SS 의 동형형질을 가전 사람들이 죽더라도 집단 전체로 볼 때 는 빈도수에 별로 영향을 주지 않는다. 오늘날 미국 흑인들의 9% 정 도가 S 형질을 보유하고 있으며, 이들 중 0.25% 가 SS 형질을 가지고 있다고 보고되고 있다• 흑인들이 미대륙에서 살아온 지 200 년이 지난 오늘날 S 형질의 빈도수가 위의 통계수치처럼 떨어졌는데, 이는 바로 S 형질에 알맞는 환경상의 요인이 없기 때문이다. 결과적으로 농경문 화는 인류의 생물학상 전화에 영향을 끼쳤다고 할 수 있으니, 바로 생태계와 문화가 서로 밀접하게 맞물려 있기 때문이다. 큰진화 Macroevoluti on 다윈은 진화를 〈자손에서 나타나는 변화〉라고 정의하였다. 현대 전 화론에서는 이런 변화는 〈서로 다른 재생산〉의 정도에 따라 일어나며 유전인자의 빈도수의 변화로 나타난다고 정의한다. 따라서 한 집단의 유전인자 빈도수의 변화를 연구함으로써 그 집단에서 전화가 어떻게 진행되는가를 알 수 있다. 앞에서 사람의 유전인자의 빈도수가 어떻 게 변하는가를 예를 들어 살펴보았는데, 이들 모두에 자연선택이 작 용함을 알수 있다• 자연선택이란 개념과 용어는 다윈에 의해 시작되었으며, 다윈 당시 에는 자연선택을 〈적자생존〉이라고 생각하였다. 이때 적응도란 말은 진화론적 생물학자들에 의해서 〈다윈의 성공 Dar wini an success 〉이라 는 뜻으로 이해되었는데, 이는 환경에 가장 찰 적응하는 자가 다음 세대에까지 자기의 유전인자를 전달하는 것을 의미한다. 적응도란 이 어지는 재생산과 같은 뜻이다. 적응도는 한 개체가 자기의 유전인자 를 자식에게 전달하는 데 성공할 수 있는 능력을 뜻하며 이는 계산될
수 있다. 지속된 재생산은 한 개체가 짝을 만나는 정도를 뜻하는 것 이 아니라 제 2 세대가 살아 남은 정도를 뜻한다. 제 1 세대의 한 개체가 자기의 유전인자를· 제 2 세대에게 성공적으로 전하기 위해서는 몇 가지 선행되어야 할 조건들이 있다 ; 제 1 세대 개 체가 수태할 수 있는 나이에 도달해야 하며, 많은 수의 좋은 배우자 롤 선택할 수 있어야 한다. 자식은 최대한으로 낳아야 하며, 새끼들이 어른이 될 때까지 많은 양의 에너지를 투자해야 한다. 모든 생명체들은 이 조건들을 어느 정도까지 충족시킬 수 있다. 동 물들은 그들의 진화상 적응도를 극대화하기 위해 여러 가지 방법들을 가지고 있다. 예로 개구리들은 많은 알을 낳는다. 그러나 알이 깨어난 후에는 돌보지 않는다. 따라서 얼룩뱀이 연못에서 올챙이를 잡아먹어 도 어미개구리가 자식을 보호하기 위해서 뱀울 공격하지는 않는다. 원숭이는 반대로 새끼 생산을 극대화하는 방법 대신에 새끼를 키우는 대 에너지를 사용한다. 서로 다른 두 가지 방법을 비교해 볼 때 각각 의 방법은 나름대로의 이점을 가지고 있다. 앞에서 말한 연구들을 불과 수천 년 정도에 걸쳐 일어나는 유전인 자의 빈도수 변화를 관찰하기 때문에 작은 전화 M i croevolu ti on 연구 라고 하며, 짧은 기간의 변화를 통해서 전화를 DNA 수준에서부터 유전인자, 개인, 집단의 단계까지를 연구대상으로 한다. 이에 반해 큰진화는 종 • 속과 같은 단위를 전화의 연구대상으로 다룬다. 변화는 신체의 여러 부분 중 특히 두뇌나 다리 모양, 골반 및 발의 모양 등에서 나타나는데, 이갇이 신체구조의 변화를 일으키는 유전상의 원인은 아직 알지 못하나 신체구조를 만들어내는 코드는 여 러 유전인자와 대립형질이 얽혀 이루어전다는 정도만 알려지고 있다.
분류학 : 린네의 분류방법 여기서는 큰전화의 입장에서 인류의 전화를 살펴보기로 하자. 지 구상에 살고 있는 생물 중 지금까지 약 170 만 종이 알려져 있다. 이 가운데 화분류가 25000 종이고, 등뼈동물이 47000 종, 그리고 75000~ 종 이 곤충류이다. 실제로는 더 많은 생물들이 살고 있을 것으로 여겨지 는데, 특히 열대 삼림지대에는 약 3 천만 종에 달하는 곤충류들이 살 고 있으리라고 여겨진다. 이렇게 많은 생물의 다양성을 연구하는 분 야롤 계통학 S y st ema ti cs 이라고 한다. 분류학 Taxonom y은 계통학의 한 방법으로 인류가 어떻게 전화해 왔는가를 살펴보는 데 절대로 필 요하다. 계통학자들을 포함해서 사람들은 지구상에 살고 있는 생물의 다양 성을 표시하기 위해서 그들 나름대로의 방법들을 가지고 있다 . 그 예 로 아프리카의 칼라하리 지역에는 많은 동물들이 살고 있다. 만일 우 리가 탄 비행기가 이 지역에서 추락했다고 가정해 보자. 배가 고픈 우리에게 기니아 닭은 먹음직스런 음식일 것이다. 그러나 이것을 잡 기는 그리 쉽지 않으며 이 지역에서 살고 있는 부시먼들도 쉽게 잡지 못한다. 이들은 기니아 닭을 말 대신 손짓으로 표현한다. 1700 년대에 다른 문화권에 사는 사람들이 생물을 표시할 수 있는 체계적인 방법에 대해 연구하기 시작하였다. 린네가 처음으로 생물계 롤 분류하여 『자연체계 』 에 소개하였는데, 1730 년 이 책이 처음 출판 되었울 때 린네의 나이 28 세였다. 이 책에서 그는 〈이 세상에는 신이 창조한 만큼의 많은 생물들이 있다 There are as many spe cie s as the Creati on pro duced 〉고 하였다. 그러나 1762 년 판에서 린네는 생 물의 속 Genus 은 변하지 않으나 종 S peci es 은 변할 수 있다고 말하였 고, 1766 년 마지막 판에서는 그간의 생각을 완전히 바꾸어서 〈아마도 새로운 종들이 있을 수 있다〉고 인정하게 되었다. 처음에 린네는 생
물들은 불변성이라고 확고하게 믿었으나, 점차로 그는 하느님이 진화 롤 하는 작은 수의 생물들을 만드셨다고 생각하였다. 린네의 생물에 대한 이런 인식의 변화는 그 자신이 연구한 생물들이 서로 관련이 있 다는 사실을 알고 난 후부터이다. 현대 분류학의 체계는 생물계를 크게 동물계와 식물계로 나눈다. 이들 중에서 같은 이름을 가지고 있는 무리를 덱손t axon 이라고 하는 데, 이는 하나의 종을 나타내며, 이 무리들이 모여서 덱사t axa 를 이 룬다. 이때 서로 연관이 있는 동식물들을 구별하는 기준은 이들이 서 로 나누어 가전 해부학상의 특칭의 유사한 정도에 따라 분류된다. 여기서 현대 분류학체계의 한 예를 살펴보자. 경마장에 가면 말과 기수가 함께 달리는 것을 볼 수 있는데, 이들을 구분하기 위해서 속 과 종의 이름을 사용한다. 이때 속의 이름은 대문자로 나타내고 종의 이름은 소문자롤 쓴다. 속과 종의 이름은 이탤릭체로 표기하거나 밑 줄을 긋는다. 분류체계에 따른 말과 사람의 분류는 표 3. ~ 갇다. 표에서 보면 말과 사람은 둘 다 동물로 같은 계, 문, 강에 속하나 목의 단계에서부터 갈라전다. 이 단계부터 말과 사람은 서로 다른 t axa 이다.
표 3. 4 린네의 분류체계에 따른 말과 사람의 분류 말 사람 계(界 : Ki ng do m) 동물계 Anim a lia 문(門 : Phy lu m) 등골동물문 Chordata 강(綱 : Class) 젖먹 이동물강 Mamma lia 목( 目 : Order) 기제류목 Per i ssodac ty la 영 장류목 Pr i ma t es 과(科 : Fa mily ) 말과 E qui dae 인류과 Hom inida e 속(屬 : Genus) 말속 E q uus 인류속 Homo 종(種 : Sp ecies ) 말 Eq uu s caballus 사람 Homo sap ien s
이렇게 생물의 이름을 정하는 방법은 170 만 종이나 되는 모든 동식 물들을 분류하기 위해서 정해졌다. 이때 각각의 동식물들은 두 개의 이름을 갖고 있는데 이를 〈린네 또는 이중 이름 명명법 L i nnean or Bion om ial Nomencla t ure 〉이라고 한다. 이렇게 생물의 이름을 정할 때는 국제협약에 따라야 한다. 규약에는 두 개의 중요한 규칙이 있는 데, 첫째가 〈우선의 규칙 Law of Pr i or ity〉으로 이는 처음 지어전 이 롬에 우선권이 있으며 한번 정해진 이름은 변경될 수 없음을 뜻한다. 둘째는 〈대표 형태의 규칙 Law of T yp e 〉으로 이는 새로운 종의 이름 울 만들려 할 때는 반드시 대표적 인 표본Typ e S peci emen 이 하나 있 어야 한다는 뜻이다. 이렇게 생물의 이름을 정해 주는 분야를 분류학 이라고한다. 동물의 종을 구분할 때는 크게 두 가지 방법에 따라 이루어전다. 첫째가 형태상의 특칭을 기준으로 하는 방법이다. 이 방법은 희랍의 아리스토텔레스로부터 린네에 이르기까지 사용되던 방법으로 종들을 의모에 따라 구별하였으니, 만일 의모가 아주 다르면 두 개의 서로 다른 종둘로 구분하였다. 오늘날 살아 있는 동물의 경우에는 〈생물학상 종의 개념 Bio l og ica l Sp e ci es Conce pt〉을 적용하여 구분한다. 이때 종의 개념은 동물 암 • 수가 서로 교배할 수 있는지에 달려 있다. 이 개념에 따르면 종이란 서로 짝을 찾아서 새끼를 낳을 수 있는 무리로서 이들은 다른 무리들 과는 짝을 짓지 않는다. 실제로 두 무리의 동물들이 같은 종에 속하 는지의 여부는 쉽게 알 수 있다. 한 예로 두 마리의 새가 서로 비슷 하게 닮았으나 그들이 짝을 찾는 소리가 다르면 이들은 서로 교미를 하지 않는다. 서로가 다론 종이기 때문이다. 동물들에서 이런 차이는 쉽게 알수가 있다. 위에서 말한 방법들은 살아 있는 동물들인 경우에는 문제가 없으나 화석인 경우에는 문제를 발생시킨다. 만일 건 송곳니를 가전 고양이
과에 속하는 머리뼈를 발견했을 경우 이 동물화석이 오늘날에 살고 있는 사자와 같은 종에 속하는 지는 명확히 알 수가 없다. 이렇게 화 석인 경우에는 생물학적인 개념인 짝을 지어 새끼를 낳을 수 있는 능 력을 증거할 직접적인 방법이 없기 때문이다 . 따라서 고동물학에서는 〈옛날종 Paleos p e ci es 〉 개념을 사용하고 있다. 동물의 종을 구분할 때 부딪치는 또 다른 문제는 동물들은 시간이 지남에 따라 진화를 통해서 모습이 변하는 데 반해 린네의 명명법은 변치 않는다는 점이다. 즉 동물은 전화중에 중간형이 나타나는 데 비 해 린네의 명명법은 이들 중간형의 이름을 나타낼 수가 없다. 이 책에서는 영장류 중의 하나인 인류에 대한 연구를 주제로 하고 있다. 따라서 영장류에 속하는 인류는 어디로부터 시작되었는가를 · 살 펴보자(표 3. 2 참조). 인류는 등골동물문에 속하며 아문 단계에서 등 뼈동물로 구분되어진다. 이 등뼈동물 중에서 사람은 젖먹이동물강으 로 분류되며 아강에서 다시 태반을 지닌 젖먹이동물로 분류되고, 이 태반 젖먹이동물 중에서 인류는 영장류목에 속한다. 오늘날 지구상에는 약 180 종의 영장류가 남아 있으며, 인류는 그 중의 한 종이다. 린네는 영장류를 다론 동물의 무리로부터 분리시켜 놓았는데, 이는 인류가 이 무리에 속해 있기 때문이다. 영장류 Pr i ma t e 란 말은 라틴어의
은 길을 따라 진화해 왔다. 전화론의 입장에서 살아 있는 영장류는 전화에 대한 〈 모델 〉 을 제공하기 때문에 우리는 이들을 연구함으로써 인류 조상의 행위가 어떠했는가를 추론할 수 있다. 영장류가 전화해 온 과정을 살펴볼 때 우리가 한 가지 염두에 두어 야 할 점은 오늘날 살고 있는 유인원은 결코 인류가 될 수가 없다는 것이며, 이는 인류와 오늘날 살고 있는 유인원은 공통조상에서 갈라 져 나온 진화의 마지막 산물이기 때문이다. 위에서 큰 전화의 결과에 따라 태어난 동물들에 대해서 살펴보았 다 . 여기서는 태어난 동물 중 살아 남은 후손들에 대해서 살펴보기로 하자. 살아 남은 후손을 측정하기 위해서는 그 동물의 총수효와 지역 상 분포, 그리고 그들이 살았던 기간 등을 가늠해 보아야 한다. 위의 조건들을 인류에게 적용했을 때 위의 두 조건, 죽 인류의 수효와 분 포는 측정이 가능하나, 하나의 생물로서 인류가 기원 이래 살아온 기 간이 다론 동물들에 비해 오래지 않기 때문에 아직은 그 존속기간에 대해서는 알수가 없다. 큰진화에 쓰이는 용어와 개념들 본원화 Pr imiti ve 와 특수화 S p ec iali zed 된 특칭들 : 동물의 큰 전화를 이해하기 위해서 우리는 몇몇 용어와 개념을 정의해야 한다. 먼저 본 원화와 특수화라는 용어를 살펴보기로 하자. 전화생물학에서 본원 화란 뜻은 다른 〈종〉둘 사이에서 서로 비교를 위해 쓰이는 용어로, 죽 상대비교에 사용된다. 그 예로 주머니쥐로 알려진 오포섭 Oppo - ssum : Di de lph is v i rg i n i ana 을 들 수 있다. 이 동물은 오랜 세월 동안 존속했음에도 불구하고 모습이 많이 변하지 않았다. 그 중의 하나로 주머니쥐는 사람처럼 한쪽 발에 다섯 개의 발가락을 가지고 있는데, 이런 모습을 본원의 특칭이라고 한다. 그러나 여기에 비해 오늘날 살
고 있는 말은 단지 하나의 발가락을 가지고 있다. 그래서 말은 주머 니쥐와 비교해 볼 때 특수화된 발을 가지고 있다고 한다. 이렇게 신체의 특수화된 몸의 기관이 특별한 활동이나 생활조건에 적합하도록 변화된 것을 특수화라 하며, 이때 동물은 본래의 적응력 을 상실하게 된다. 예로 중국의 내륙지방에서 살고 있는 판다곰은 본 래가 식육류인데 몸의 여러 기관들이 대나무를 먹고 살 수 있도록 변 하여 손목뼈가 길게 발달하였고 큰 머리뼈와 이빨을 가지게 되었다. 오스트레일리아에서 사는 코알라는 유카리나무를 먹고 살 수 있도록 몸의 구조가 발달하였다. 그러나 오늘날 판다곰과 코알라는 대나무숲 과 유카리나무숲이 줄어듦에 따라 멸종의 위기에 처해 있다. 같은 과 에 속하는 동물들에 비해 적응력을 상실했기 때문이다. 절멸 Ex ti nc ti on : 절멸이란 말은 생태와 밀접한 관련이 있다. 〈종〉 의 입장에서 절멸이란 단지 진화에 실패함을 뜻할 뿐이다. 어떤 동물 에서 유전인자가 남아 있다는 것은 그 동물 중 마지막 한 마리가 살 아 있는 동안에 한정된다• 아래 그림 3. 16 의 왼쪽 그립에서 각각의 점은 다른 동물의 개체를
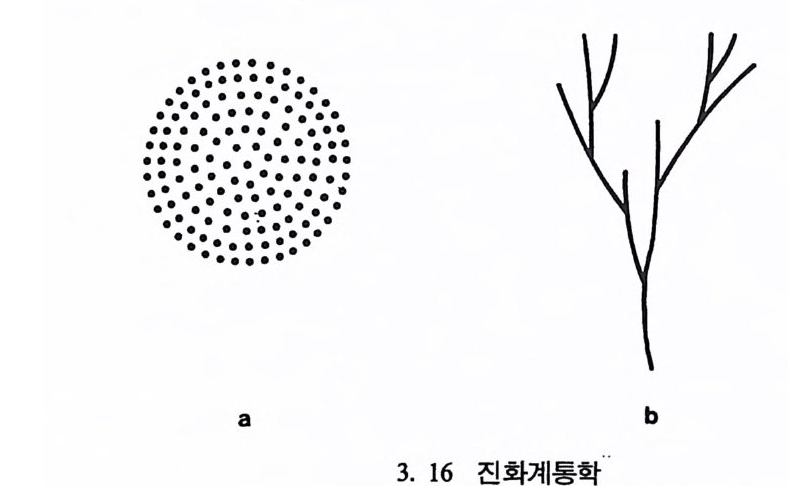 •••.•._•••.••·•_.••• ••• ·••. .; • •.••.•.•• ·• • .•••- •• . .•• ••• •.. • .• ••~••• •. •,•; •-•·••••..•-, • .••••••.·. ..-••.• • .••••.•.:••• - •• .•-•·•.• •••• . •. ••• .••-•_ • ••• .
•••.•._•••.••·•_.••• ••• ·••. .; • •.••.•.•• ·• • .•••- •• . .•• ••• •.. • .• ••~••• •. •,•; •-•·••••..•-, • .••••••.·. ..-••.• • .••••.•.:••• - •• .•-•·•.• •••• . •. ••• .••-•_ • ••• .
나타내는데, 이때 각 동물 사이의 관계는 알 수가 없다. 오른쪽 그립 은 서로 다른 개체들간의 전화상 관계를 보여주는데, 각 동물의 조상 둘은 지금은 절멸되었지만 오래 전에 서로 관련이 있었음을 알 수 있 다. 하나하나의 가지들을 리네지li nea g e 라고 부르는데, 이는 하나의 동물이 시간에 따라 변하는 것을 나타낸다. 이렇게 시간을 따라서 이 루어지는 생물의 전화상 선후관계를 연구하는 분야를 전화계통학 Ph y lo g en y이라고 한다. 종의 분화 S p ec i a ti on ·: 분화란 하나의 〈종〉에서 새로운 〈종〉이 갈 라져 나가는 현상을 의미한다. 이런 종의 분화는 다윈이 전에 이미 생각했고 오늘날에도 활발히 연구가 진행되고 있는 분야로, 분화는 아래와 갇이 3 개의 경우가 있다. 표 3. 5 분화의 방법 시간 첫째 경우, 종 A 종 A : 분화가 일어나지 않음 둘째 경우, 종 A 종 B : 서행전화 ana g enes i s 셋째 경우, 종 A 二 종 A : 파행전화 clado g enes i s 종 B 위의 표에서 분화는 둘째 경우인 서행진화(하나의 종이 접전적으로 다른 종으로 변하는 현상)와 셋째 경우인 파행진화(하나의 종이 시간 이 지남에 따라 둘 이상의 종으로 갈라지는 현상)에 의해 일어난다. 이때 파행진화는 같은 지역에서 일어나는 〈같은 지역 분화 S ym pa tr i c S peci a ti on 〉와 다른 지역에서 일어나는 〈다른 지역 분화 All op a tr i c Sp e c i a ti on 〉로 다시 나누어진다. 〈같은 지역 분화〉는 같은 종에 속하는 동물 중 분화된 두 개 이상
의 무리들이 같은 지역에서 사는 것을 의미한다. 이때 한 동물 무리 는 다른 동물 무리들과는 염색체의 변화나 다른 유전상 변화에 의해 서 분리되고, 이 두 무리의 동물들은 서로 짝을 찾을 수가 없으며, 그 결과분화가 일어난다. 〈다른 지역 분화〉는 하나의 동물 무리가 다른 동물 무리로부터 떨 어져 나갈 때 일어난다. 예로 같은 종에 속하는 세 개의 다론 동물 무리가 있는데 이들 모두가 서로들 사이에서 짝울 구할 수 있다고 (이는 교미에 의해 서로 유전물질을 교환함을 뜻함) 가정해 보자. 만일 하나의 동물 무리가 다론 두 동물 무리로부터 ·격리된다면 이 무리는 시간이 흐름에 따라 변할 것이다. 세월이 지나서 이 무리가 다시 본 래의 두 무리들과 결합할 기회가 주어진다면 이들은 서로들 사이에서 짝을 구할 수 있거나 아니면 짝을 구할 수 없을 수도 있다. 이때 짝 울 구할 수 없는 경우를 분화가 일어났다고 한다. 동물이 짝을 구해 새끼를 낳는 행위는 여러 방법으로 영향을 받는 다. 죽 한 몸 안에 양성을 가지고 있는 경우는 동물이 신체상의 변화 로 인해 육체적인 결합을 막는데, 이 경우를 〈짝짓기 전 방법〉이라고 한다. 〈짝전 뒤 방법〉은 배우자나 접합자가 죽거나 자식을 낳았어도 적응성이 떨어지거나, 아니면 자라면서 발육이 불량한 경우를 말한다. 따라서 분화 이후에 한 지역에서 다같이 살게 되어도 많은 일들이 일 어난다. 진화의 유형 다윈의 진화점진론 (Darw i n i an 또는 Phy le ti c Graduali sm Model) (그 림 3. 17a) 동물의 분화는 생명체가 이 지구상에 생겨난 이후 수없이 일어났으 며, 학자들은 이들 분화가 얼마나 빨리 전행되었는가에 관십을 가져
왔다. 대다수의 사람들이 생물은 한번 창조된 이후에는 변하지 않는 다는 생각이 지배하던 시대에 다윈은 이들과는 반대로 생물은 차츰 변화해 왔다고 믿었다. 다윈은 이러한 고정된 관념을 고치고자 하였 으며, 새로운 종이 생겨난다는 사실을 증명하려고 노력하였다. 다윈의 생각은 당시 시대의 산물이었다. 다윈은 독서를 통해 새로운 지식과 만났으며, 여러 사람들과 의견을 나누었고, 새로운 생물학과 사회이론 의 변화를 인식하고 있었다. 다윈은 자기 이론의 핵심을 그 당시에 널리 퍼져 있던 새로운 생각에서 빌어왔으며, 사람들이 이해할 수 있. 었던 용어를 써서 자기의 이론을 기술하였다.
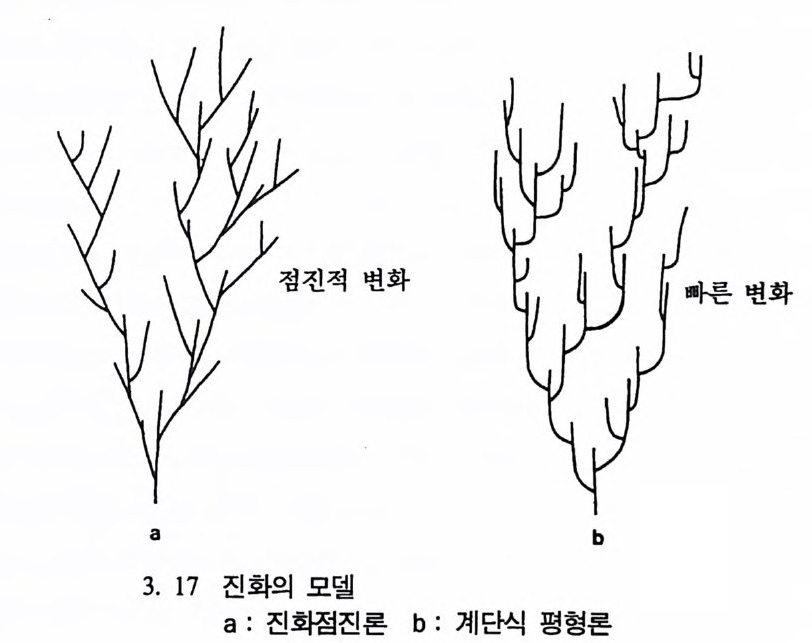 a b
a b
다윈 당시에 〈인류의 문화와 사회는 전보한다.〉는 개념은 널리 인 식되고 있었다. 그는 과거는 차츰 전화해 가는 동물들의 긴 사슬로 이루어져 있다고 생각하였다. 그의 이런 이론을 다윈의 전화접전론이
라고 한다. 다윈은 진화접전론자였다. 그가 지질학상의 기록들을 살펴 보았을 때, 특히 동물화석의 경우, 그는 전화의 중간형을 찾을 수가 없었다. 다윈은 자신의 저서인 『종의 기원』 제 IO 장에서 이렇게 화석 의 중간형이 없는 것은 〈지질학의 불완전한 기록〉 때문이라고 설명하 고 있다. 이런 다윈의 주장은 창조론자들이 다윈의 진화론을 공격할 때 내세우는 근거로 인용되고 있다. 그러나 우리가 염두에 두어야 할 것은 실제로 화석에서 보여지는 불완전한 기록은 진화과정중 어떤 부 분이 없는 것이 아니라 단지 화석의 기록이 부분적으로 없어전 것을 의미한다. 오늘날 우리가 사용하는 현대 전화론은 다윈이 주창한 진화론과 1970 년대 이후에 발달한 유전학 및 생물학의 여러 분야에서 연구된 이론의 결합으로 이루어졌는데, 생물의 변화는 차츰 일어난다는 다윈 의 진화이론에 바탕을 두고 있다. 계단식 평형론 Punc tu a t ed Eq ul i br iu m Model (그립 3. 17b) 19701d 대에 오면서 스티븐 제이 굴드와 니일 엘드리지가 전화론의 한 유형으로서 전화점전론에 대체되는 계단식 평형론을 제시하였다. 이들의 이론에 따르면 전화의 방법과 속도는 다윈의 이론과는 다르게 우발현상으로 빨리 일어나며, 또한 지질학상으로 볼 때 새로운 동물 이 생겨나고, 이후 얼마 동안 변화가 없는 것은 바로 전화의 결과이 지 지질학상 기록의 불완전성 때문이 아니라고 주장한다. 전화에 관한 대다수의 이론들에 따르면 전화는 수천 년에 걸쳐 작 은 무리들에서 일어난다고 한다. 그러나 수천 년이란 시간은 어떤 동 물들의 경우에는 극히 짧은 시간이라고 할 수 있다• 굴드와 엘드리지 는 작은 무리들은 어느 정도 안정되어 변화에 대해 거부반응을 보이 며, 따라서 평형상태에 이르게 된다고 한다. 이들에 따르면 화석이 발 견되지 않는다는 사실은 화석기록들이 없어전 것이 아니며 바로 기록
이 없는 그 자체가 실제 자료로서 다루어져야 한다고 주장한다. 이둘 의 이론은 다윈의 이론과는 정반대의 입장을 나타내고 있다. 전화접전론에 따르면 변화는 시간이 경과함에 따라 천천히 진행되 며, 때로는 진행과정중에 가지를 쳐서 새로운 종이 생겨난다고 한다. 이에 반해 계단식 평형론에 따르면 하나의 종은 상당 기간 동안 변하 지 않는다고 한다. 그러나 같은 종이지만 중십에서 떨어져 변두리에 살던 한 작은 무리가 어느 시점에서 갑자기 아주 빠르게 분리되어 새 로운 종으로 변하는데, 이때 새로 태어난 종은 먼저 살던 종을 대신 하며 상당 기간 동안 변하지 않고 평형을 유지한다고 한다. 굴드와 엘드리지의 연구는 많은 학자들로 하여금 화석들을 다시 살 펴보게 하는 계기가 되었다. 그러나 이들의 연구에 대한 많은 비판이 있으니, 미국 캘리포니아 버클리 대학교의 데이비드 웨이크 Da vi d Wake 교수는 〈 1930 년대와 1940 년대는 유전학과 자연사가 함께 어울 려 전화과정에 대한 새로운 이론들이 소개되었으나 오늘날의 연구는 전화계통도에 별로 영향을 주지 못하고 있다 . 〉고 비평하고 있으며, 미 국에서 발행되는 〈과학 Sc i ence〉 잡지에 무고된 편지 중에 〈화석들만 을 가지고는 전화가 차츰 일어났는지 아니면 갑자기 일어났는지 알 수가 없기 때문에 이에 대한 답은 전화유전학자들에게서 찾아야 한 다.〉고 주장하는 내용이 소개되고 있다. 어떤 학자는 이 계단식 평형 론을 〈시시한 이론〉이라고 혹평하고 있고, 또 다른 학자는 이 모델은 마르크시즘의 산물이며 마르크시스트들의 구성에 따라 진행되었다고 비판을 하고 있다. 이런 비판에 대해 굴드 교수는 〈나는 단지 화석들 에서 나타나는 골을 설명하기 위해서 새로운 이론을 만들었을 뿐이 다〉라고 답하고 있다. 진화점진론과 계단식 평형론에 관한 지난 20 여 년간 논쟁의 결과를 살펴보면, 양쪽의 유형들이 모두 진화과정에서 나타나는 변화를 설명 할 수 있음을 보여주고 있으나 어느 쪽이 더 타당한지를 말하기에는
아직 연구가 더 이루어져야 한다. 진화에 관한 지질학의 중거들 진화생물학자들이 1 만 년 내지 2 만 년이란 시간은 진화상의 변화가 급격하게 일어날 수 있기에 충분하다고 생각하는 데 반해, 인구유전 학자들은 단지 조금씩 변화가 일어날 수 있는 정도의 시간일 뿐이라 고말한다. 우리는 과거에 일어났던 변화의 과정들을 찰 알 수가 없으며, 또한 지금의 자료들을 가지고는 과거에 일어났던 변화들을 자세히 설명할 수도 없다. 과거에 일어났던 일들을 최초로 연구했던 사람으로 스테 노 W. S t eno 를 꼽을 수 있다. 그는 1600 년대 중반에 살았던 인물로 1668 년에 목사 자격을 얻기 전에 이미 해부학을 공부하여 상어에 관 한 전문가가 되어 있었다. 말타 지역에서 그는 암석에 박혀 있는 상 어 이빨들을 발견하였는데, 그는 이 이빨들이 정말로 상어 이빨이며 암석이 되기 전에 그 속에 박혀 있었다고 주장하였다. 스데노는 바위들은 밀에서부터 위로 올라가면서 쌓이는데 이때 가 장 밑바닥에 있는 바위가 제일 오래 된 것이라고 하고, 이런 법칙 울 여적의 법칙 Prin c ip le of Su p er p os iti on 〉이라고 불렀다. 암석과 모 래, 펄둘은 산이나 들에서 냇물이나 강물에 씻겨 강 하류의 삼각주로 옮겨져 쌓였다가 시간이 흐름에 따라 바위로 변하였다고 그는 생각하 였다. 스테노는 되적과 층위 그리고 되적과 충위가 어떤 관계가 있는 지를 연구한 결과 퇴적에서 보이는 충들은 바로 대홍수의 결과로 생 겨났다고 생각하였는데, 이런 주장을 넵튜니즘이라고 한다. 17 奭대 후반에 와서 이런 스데노의 주장은 후돈에 의해 도전을 받는다. 후돈은 지구는 매우 활동적이며 지질상 변화는 되풀이된다고 생각하여 균일설을 세웠는데, 후에 라이엘이 그의 저서인 『지질학 원
리』에서 이 이론을 소개하였다. 후돈의 균일설에 따르면 과거에 지구 상에서 일어났던 자연의 변화과정은 오늘날 자연이 변하는 과정과 갇 은 형태로 변하였기 때문에 오늘날의 증거들을 통해서 과거에 일어났 던 변화의 과정들을 이해할 수 있다고 한다. 이 이론은 후에 다윈울 위시한 많은 학자들에게 영향을 주었다. 1800 년대 초기에, 경제적 이유에서, 천연자원을 찾기 위한 지질담사 가 시작되었다. 당시 자연학자들은 조물주는 후에 사람들이 천연자원 울 필요로 할 것을 미리 알고 있었기에 이 자원들을 바위 속에 넣어 두었다고 생각하였다. 한 예로 1860 년대 미국의 지질학자인 아가지즈 A g ass i z 는 〈빙하는 조물주가 인간을 위해 온대지역의 땅을 비옥하게 하기 위해서 땅을 경작한 방법 God's Great Plou g h 〉이라고 생각하였 다. 이때에 지질학자들은 암석을 분류하고자 하여, 화산암이 제일 처 음으로 형성된 바위이며, 수성암은 그 다음에 형성되었고, 마지막으로 수성암의 일부가 변해서 된 암석을 변성암이라고 분류하였다. 제 3 기 에 만들어진 암석들은 수성암이며, 빙하의 퇴적작용에 의해 만들어전 암석들은 제 4 기에 만들어졌다. 1900 년대 초에 영국의 운하건설자인 스미스 W illi am S mit h 는 운하 건설중에 발견되는 화석들을 수집하던 중, 같은 종류(목을 뜻함)의 동물들의 화석은 항상 같은 충에서 발견된다는 사실에 주목하였다. 스미스는 이렇게 같은 종류의 동물들이 같은 충에서 발견되는 사실을 〈화석 연속의 법칙 Prin c iple of Fauanl Success i on 〉이라고 정의하였 는데, 이 법칙은 오늘날 충위에 관한 연구분야인 충위학의 기본을 이 루고있다.
시간의 측정 : 상대연대측정과 절대연대측정 상대연대측정 Relati ve Dati ng Meth o ds 앞에서 설명한 〈퇴적의 법칙〉과 〈화석 연속의 법칙〉을 이용하여 화석들의 연대를 알 수 있다. 죽 어떤 화석들과 이 화석들이 발견된 바위와의 관계를 비교하면 화석둘의 연대를 가늠할 수 있는데, 이때 바위들만을 비교하는 방법을 〈암석충위학 L it hos t ra tig ra p h y〉이라고 하 며, 바위에 포함된 화석들을 비교하는 연구방법을 〈화석충위학 B i os t ra tigr a p h y〉이라고 한다. 예로 영일만과 남해에 있는 한 지점의 충위를 비교해 볼 때 양쪽 지역의 많은 충이 갇은 성질을 보여준다. 이는 바 로 양쪽의 지층들이 같은 시기에 퇴적되었으며, 여러 차례에 걸쳐 양 쪽 지역의 모습이 같았음을 알 수 있다. 양쪽 지역의 지층에서 발견 되는 동물화석들의 관계를 살펴봄으로써 지층들의 연대를 알 수 있으 니, 만일 서로 다른 세 지역에서 화석을 포함한 지층들이 다음과 같 이 나타난다면, 삼영충 물고기 2 사람 암몬조개 물고기 1 말 해파리 삼엽충 물고기 2 위에서 보이는 서로 다론 지층들은 화석상의 증거에 따라서 지층들의 순서를 다음과 같이 정할 수 있다. 단 이 화석들은 단지 서로의 선후 관계만을 알려준다.
사람 말 물고기 2 물고기 1 삼엽충 암몬조개 해파리 절대연대측정 Absolute Dati ng Meth o ds 원자들이 자연의 힘으로 붕괴되는 과정을 방사성 붕괴라 한다. 바 로 이런 성질을 이용하여 어떤 지역의 암석이나 뼈둘의 나이를 추정 할 수 있는데 이를 절대연대측정법이라고 한다. 이 방법으로는 방사 선탄소 Rad i ocarbon 측정 , 우라늄계 열 Uran ium serie s 측정 , 가열발광 Thermolum ine scence 측정 , 전자회 전반응 Electr o n Sp in Resonance 측정, 아미노산Ami no Ac id Racem iza ti on 측정, 분열자국 F i sson Track 측정, 칼륨 • 아르곤 Po t assum-Ar g on 과 고지자기 Paleoma- g ne ti s 교측정 방법 등이 있다. 철대연대측정법 중 오랜 연대를 재는 데 일반적으로 쓰이는 방법으 로 칼륨-아르곤 K/Ar 방법이 있다. 화산재 등에 포함된 칼륨 가스 40- €:- 온도나 기압에 관계없이 붕괴가 일정하게 진행된다. 죽 포타슘 40 의 50% 가 붕괴되어 아르곤 가스(아르곤 40) 로 변하는 데 걸리는
시간은 13 억 년이며 이 기간을 반감기 hal f -l ife라고 한다. 화산이 폭 발했을 때 분출된 물질들은 액체로 되어 있으며, 모든 가스는 끓어 증발된다. 따라서 순수한 부석암석 Pum i ce 은 칼륨 4 어춘 포함한 석영 울 가지고 있으나 아르곤 4 (H즌 없는데, 이는 화산이 폭발한 후에 아 직 붕괴가 시작되지 않았기 때문이다. 화산폭발로 화산재는 아주 먼 거리까지 날아가게 된다. 이들 화산 재는 바위충에 가라앉는데 고생물학자들은 화산재가 쌓인 충들의 칼 륨 • 아르곤 가스의 비율을 이용해서 화석들의 연대를 측정할 수 있 다. 화산재충 밀에 있는 화석들은 이 총보다 연대가 오래 되었으며, 위에서 발견되는 화석들의 연대는 지충보다 젊다는 것을 알 수 있다. 이런 절대연대측정 방법을 통해서 지질학자들은 지구의 역사에 관한 지식을 얻게 되며, 그 결과 지구의 나이는 46 억 년이란 결론에 도달 하였다. 진화계동그림의 복원 동물의 분류는 동물들이 서로 닮은 정도를 따져서 이루어진다. 동 물의 닮은 정도를 보여주는 계통분화그림 Clado g ram 은 동물들 사이 에 서로 닮은 해부학상 특칭들이 개체들만의 특칭인지, 아니면 공통 조상으로부터 물려받은 것인지에 따라서 결정된다(그림 3. 18). 우리 는 현재 살아 있는 동물들의 특칭을 비교함으로써 동물들의 진화 역 사에 대한 자료를 얻을 수 있다. 위의 계통분화그림은 시간의 앞 • 뒤 관계를 나타내지는 않고 단지 서로의 닮은 정도만을 나타내는 것으로 침팬지와 거미의 조상은 비비 와 침팬지의 조상보다 오래 전에 살았음을 보여주고 있으나, 그때가 언제였는지는 말해 주지 않는다. 여기에 바로 시간의 개념을 도입했
울 때 이를 진화계통족보 Ph y lo g ene ti c Tree 라고 부른다(그림 3. I9). 다시 말해 전화계동그림은 동물이 전화해 오는 과정중에 조상과 후 손간의 관계를 알려준다. 이렇게 동물의 조상-후손간의 관계를 보여 주는 전화계통족보를 만드는 방법과 특성, 방법의 한계 등에 대해 살
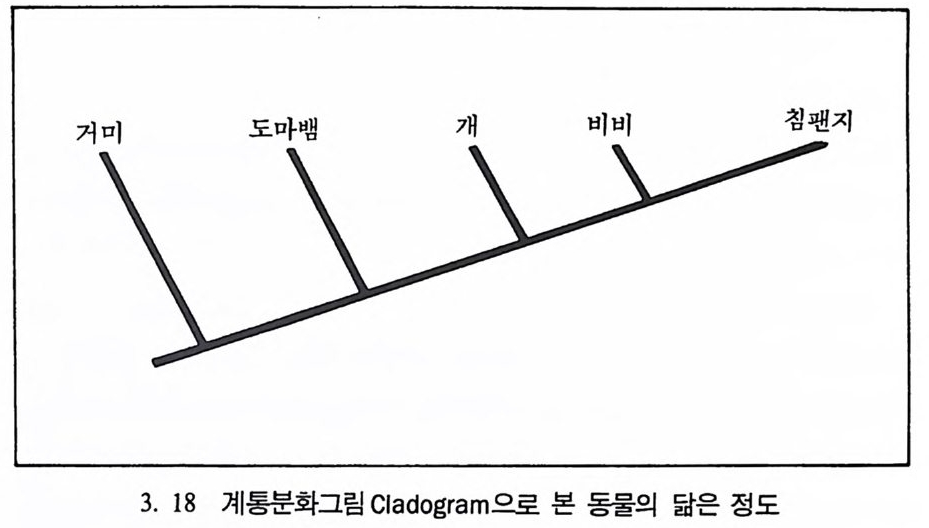 개 비비 침팬지
개 비비 침팬지
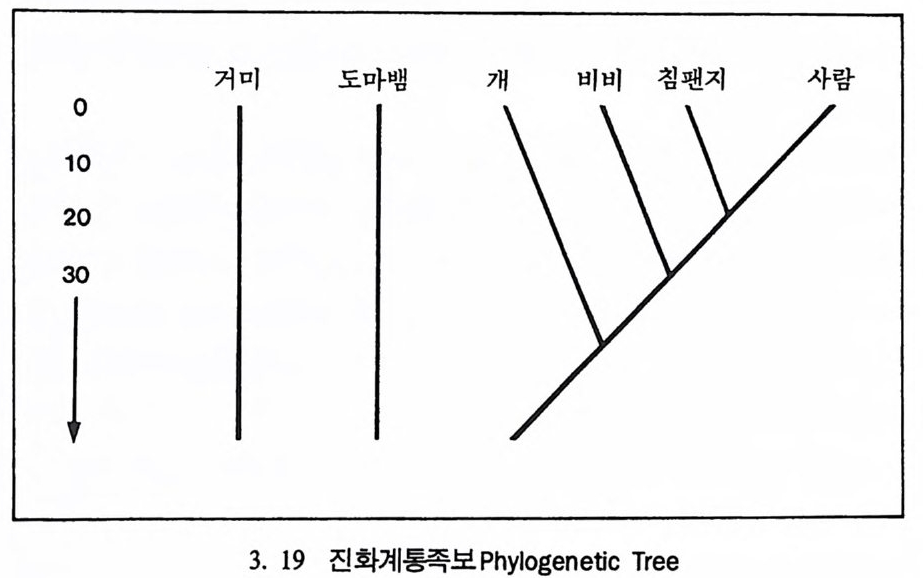 0102030거 미 도마뱀 7ij 비비 침팬지 사람
0102030거 미 도마뱀 7ij 비비 침팬지 사람
펴본다. 일반적으로 전화계동족보는 살아 있는 동물의 해부학상 특칭 을 비교하는 비교해부학, 분자생물학, 그리고 뼈화석을 비교연구하는 고동물학의 비교방법에 의해서 복원된다. 비교해부학 과거부터 일반적으로 사용된 방법으로 근육조직 대 근육조직, 신경 조칙 대 신경조직의 비교 등, 살아 있는 동물의 해부학상 특칭을 연 구대상으로 하는 비교해부학은 조상-후손간의 관계를 정하는 데 매 우중요하다. 비교해부학 연구방법의 한계는 오직 살아 있는 동물에서만 이 방법 의 완전 적용이 가능하다는 점이다. 따라서 서로 닮은 두 동물이 공 통조상으로부터 언제 갈라졌는지는 알 수 없으며, 단지 추측이 가능 할 뿐이다. 서로 닮은 두 동물들을 보았을 때 어느 한 동물도 다른 한 동물의 조상이 아니라는 접을 염두에 두어야 한다. 이는 이들은 전화발전 단계의 마지막 산물이기 때문이다. 따라서 두 동물의 조상 은 이들을 서로 섞어놓은 모양과 같다고 생각하는 것은 단지 상상일 뿐이다. 이 방법의 또 다론 한계는 신체상 서로 닮은 특칭들이 다론 종들 사이에서도 전화되어 나타난다는 사실이다. 그 예로 상어와 돌핀 그 리고 물고기 도마뱀 ich th y o saur : fi sh- li zard 의 지느러미를 들 수 있 다. 이들은 몸의 위와 옆에 지느러미를 가지고 있는데, 바로 옆모습이 서로 닮았다• 그러나 이렇게 닮은 모양은 서로가 관계없이 각각 독자 로전화된 것이다. 이렇게 동물들이 닮은 모양을 갖는 것은 전화상의 관계이거나, 아 니면 각기 독자적인 발달에 의함을 알 수 있다. 어떤 특칭이 같은 조 상으로부터 나와 모양이 닮았을 때를 상동관계 Homolo gy라 하며, 독
자 전화의 결과에 의했을 때를 유사관계 Analo gy라고 한다. 따라서 위에서 예로 본 상어, 돌핀, 물고기 도마뱀의 지느러미는 유사관계임 을 알 수 있다. 이렇게 동물 사이의 상동 • 유사관계는 이들의 겉모습 과 내부구조를 살펴봄으로써 알 수 있다. 분자생물학 이 방법은 동물결 을 구성하고 있는 분자들의 닮은 정도를 찾아보는 것이다. 다양하고 복잡한 구조를 가지고 있는 분자를 이용하여 전화 계통도를 구성하는 데는 아래와 같은 . 방법들이 있다. 면역력 동물들의 면역체계를 이용하는 방법이다. 예로 사람의 단백 질을 토끼에 주입하면 토끼의 면역체계는 사람의 단백질에 대해 특별 한 항체를 만든다. 이 항체들이 사람의 피와 섞이면 완전한 반작용이 일어난다. 만일 말의 피와 섞이면 반작용은 거의 일어나지 않으나, 침 팬지의 피와 섞이면 거의 완전한 반작용 반응이 일어난다. 이렇게 면 역의 방법을 통해 동물들의 멀고 가까운 관계를 추정할 수 있다. 단백질의 배열 사람과 고릴라에서 헤모글로빈 같은 단백질들을 추 출할 수 있으며, 이렇게 추출된 단백질을 구성하는 아미노산의 배열 이 각각 어떻게 이루어져 있는가를 알 수 있다. 예로 사람과 고릴라 의 경우는 단지 몇 개의 아미노산의 배열이 서로 다를 뿐이다. 이 방 법은 DNA 가 동물둘 사이에 어떻게 다론가를 알게 해주는 간접방법 중의 하나이다. 사람과 개의 단백질을 비교하면 그 차이는 더욱 커지 는것을알수있다. 핵산 DNA 교배 이 방법은 서로 다른 두 동물의 DNA 를 녹여 DNA
를 이루고 있는 가닥 st ra i n 둘을 분리한 후, 그 혼합물을 섞어 식힌 다음 두 동물의 DNA 가 어느 정도 서로 혼합되는가를 계산하는 방법 이다. 이 방법으로 조사한 결과, 침팬지와 사람은 1.6 % 정도의 유전 상의 거리가 있는 것으로 밝혀졌는데, 이는 바로 사람의 유전자 중에 서 1000 쌍의 염기를 무작위로 추출했을 때 침팬지는 단지 16 쌍의 영 기가 사람의 것과 다르며, 나머지 984 쌍의 염기는 서로 같음을 뜻한 다. 사람과 개의 경우는 약 25% 가 서로 다르다. 이 방법은 동물 사이의 모든 유전자를 비교할 수 있기 때문에 매우 믿을 수 있는 방법이라고 할 수 있다. 왜냐하면 동물 개개의 전화의 역사는 그들의 유전자 안에 암호로 저장되어 있기 때문에 DNA 가 변 화하는 정도를 계산할 수 있다면 동물들이 분화되어 나간 시간이 계 산될 수 있다. 분자생물락의 문제점 위에서 언급한 방법들은 동물의 진화과정을 이해하는 데 중요한 자료를 제공해 주기는 하나 다음과 같은 문제를 지니고 있다. 죽 동물의 형태상의 변화는 항상 구조상의 변화와 일치 하지 않는다는 점이다 . 게놈에서 일어나는 변화는 DNA 를 구성하고 있는 유전 마디인 안마디에서처럼 동물의 의모에는 나타나지 않는다. 그러나 게놈의 유전상 변화는 상당부분이 일정한 비율로 일어난다. 어떻게 이런 변화가 구조상의 변화 없이 생겨날 수 있는지 의문이다. 돌연변이의 상당수는 중립 neu t ral 의 성질을 갖고 있기 때문에 유 전자의 구조에 아무런 영향을 주지 않으나 단백질들은 서로가 다른 비율로 전화한다. 따라서 단백질에서 찾아지는 분자상의 변화가 전화 과정 관계를 구성하는 데 이용되고 있다. 오늘날 당면하고 있는 문제는 화석과 지질상 자료에서 일어나는 논 란이 분자시계를 계산하는 데에서도 똑갇이 일어난다는 것이다. 죽 분자를 이용하는 방법은 아래와 같은 문제를 가지고 있다.
첫째, 분자상의 연구방법은 오늘날 절멸된 동물에서는 적용될 수 없고 단지 살아 있는 동물에서만 적용된다. 둘째, 이 방법으로는 분화의 순서만을 알 수 있을 뿐, 화석기록에서 필요한 분화가 언제 일어났는지롤 정확히 알 수 없다는 점이다. 한 예로 분자시계는 침팬지와 사람이 수백만 년 전까지 공통조상을 가졌다는 사실은 명백하게 보여주고는 있으나 이들이 어디에서 살았 으며, 어떻게 생겼으며, 무엇을 먹고 그리고 어떤 형태의 사회행위를 영위했는지는 말해 주지 않는다. 그러나 분자시계는 화석자료들과 함 께 사용될 때 좋은 결과를 가져온다. 고동물학 선사시대의 생활을 다루는 학문 중의 한 분야인 고동물학에서는 남 겨진 화석자료들을 이용해서 전화계통 그립을 구성한다. 그러나 실제 로 화석을 남긴 동물들은 극히 일부이기에 연구는 현재 살고 있는 동 물자료에 근거를 두고 있다. 어떤 동물은 일부 아니면 전부가 화석으 로 남는 경우가 있는데 이들 화석자료는 과거의 한 시점으로 우리를 데려가는 시간계산기의 구실을 한다. 화석은 대개가 처음에는 완전한 모양을 갖추고 있지만 시간이 흐름 에 따라 화석의 일부가 없어지면서 점접 본래의 모습을 알기가 어려 워진다. 고동물학은 이런 화석의 본래 모습이 어떠했는가를 밝히는 학문이라고 할 수 있다. 화석은 제한된 정보만을 보여주기 때문에 과 거를 다 밝혀주기보다는 단지 과거에 일어났던 일부분에 대한 정보를 주로 알게 한다는 접을 염두에 두어야 한다. 동물이 죽은 후 화석이 되어 사람이 발견할 때까지는 다음과 같은 단계를거친다
l) 동물의 죽음一 -2) 묻힘一 ➔ 3) 화석화 진행_ -4) 지질상의 변화 一이 화석의 노출_ ➔ 6) 확인 및 수집 -7) 연구·분석 위의 단계 중에서 제 1) 단계에서 제 5) 단계까지의 과정은 동물화석 학 Ta p honom y에서 다루며 제 6), 7) 단계는 고동물학에서 다룬다. 그러나 위의 각 단계에서 쓸모있는 정보들이 없어진다는 점을 알아 야 한다. 예로 얼룩말이 초원에서 죽었을 때를 가정해 보자. 처음에는 주워먹기침승 Scaven g er 들이 와서 죽은 얼룩말의 시체에서 살을 발라 먹는다. 하이에나 같은 짐승은 뼈까지 먹는다. 따라서 얼룩말은 단지 몇 조각의 뼈만 남게 되어 화석으로는 충분하지 못하다. 이 뼈조각들은 화산활동이나 물, 흙, 바람에 의해 땅 속에 파묻힌 다. 동아프리카의 여러 강가에는 죽어서 물에 떠내려온 동물들이 물 속에 퇴적된 상태로 발견된다. 시체는 물이 잔잔히 흐르는 지점까지 떠내려오는데, 이미 이 지역은 전에 떠내려온 동물들의 뼈들로 덮여 있다. 또한 만일 강이 넘쳐 흐르면 강둑에 있던 뼈들도 함께 묻혀서 화석이 된다. 어떤 시체는 식육류나 뜯어먹기짐승에 의해 동굴 안으 로 옮겨져 묻히기도 한다. 이렇게 땅 속에 묻혀 화석이 된 뼈들은 지 각의 변동에 따른 단충작용이나 땅의 ' 표면이 닳아지면서 표면으로 드 러난다. 토목공사나 경지정리 등도 원인이 되어 뼈화석을 땅 위로 노 출시킨다. 이렇게 땅 위로 노출된 뼈화석들은 고동물학지들에 의해 발견되어 연구가 이루어전다. 화석이 되는 것은 뼈뿐만 아니라 나무, 곤충 그리고 알에서 막 깨 어 나오는 공룡의 새끼에 이르기까지 모든 생물체가 화석이 된다. 화 석의 나이는 완전히 알 수가 없는데, 이는 화석이 생기게 하는 요인 이 환경이며 계속 일어나는 지질상의 과정들이 화석이 되는 마지막 조건을 결정하기 때문이다.
 제 4 장 영장류
제 4 장 영장류
제 4 장 영장류 영장류란 무엇인가? 동물의 경우 서로 몸집의 크기가 다른 것은 바로 크기의 다양성으 로 설명되고 있는데, 이런 다양성은 동물의 성장기간이나 삶터, 또는 성이나 특별한 체질상의 차이에 의해서 생겨난다. 동물의 〈종〉을 결 정할 때 한 표본을 기준으로 해서 이루어지는 〈형태에 따른 종의 정 의 Typo log ica l Sp e ci es Conce pt〉는 더 이상 의미가 없기 때문에, 문제 는 인류학자들이 화석에서 이런 다양성을 어떻게 찾아내느냐에 있다. 이 장에서는 영장류의 화석들을 연구할 때 〈종〉의 범위를 어떻게 규 정하는가를 살펴보기로 한다. 영장류는 16 목의 다른 젖먹이동물과 어떻게 구분되는가? 영장류는 몸의 크기라든가, 모습 그리고 행위 등에서 많은 다양성을 보여주고 있기 때문에 영장류라고 규정하는 데 있어 신체의 어느 특칭이 가장 적절한 기준이 되는지는 명확하지 않다. 그럼에도 불구하고 일반적으 로 영장류로 규정할 수 있는 몇 개의 해부학상 • 생리학상의 특칭을 찾을수 있다. 오늘날 살아 있는 영장류 내에서 프로시미안 Prosimi an 영장류와 앤스로포이드 An t hro p o i d 영장류는 서로를 구분하는 특칭이 잘 나타
나지만, 먼 옛날로 돌아가면 갇수록 오늘날 영장류에서 보이는 특칭 들은 잘 나타나지 않는다. 젖먹이동물은 중생대가 끝나고 신생대가 시작되는 약 6500 만 년 전부터 지구상에서 널리 퍼져 살기 시작하였 으며, 영장류의 가장 이른 조상도 이 시기에 발견되었다. 이후 오랜 시간에 걸쳐 영장류들은 환경에 적응하며 자신들의 해부학상 • 생리학 상의 특칭을 간직하였는데, 이들이 지니고 있는 신체상 특징은 다음 과같다. I. 영장류는 일반화된 뼈대를 가지고 있다. 죽 이들은 많은 젖먹이 동물에서는 보이지 않는 빗장뼈와 5 개의 발가락을 가지고 있다. 2. 영장류는 물체를 움켜쥘 수 있는 손과 받을 가지고 있는데, 이때 엄지손가락과 엄지발가락은 밖으로 향하여 있으며, 대개의 영장류 는 손가락과 발가락 끝에 편평한 손톱과 발톱 n ai l 을 가지고 있다 (그림 4. I).
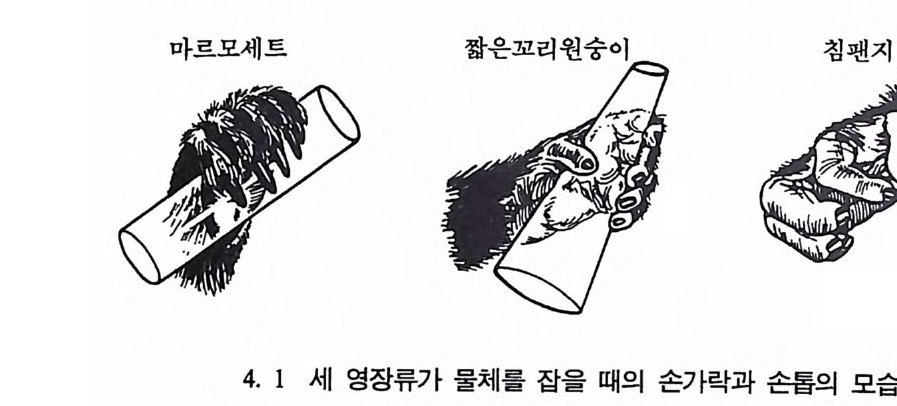 마르모세트 짧은꼬리원숭이
마르모세트 짧은꼬리원숭이
3. 영장류는 발달된 시각과 퇴화된 후각을 가지고 있다. 따라서 눈 은 상대적으로 커지고 뼈로 된 눈굴레로 보호되고 있다(그립 4. 2). 주둥기는 얼굴 쪽으로 줄어들었고, 눈은 정면을 향해 있어 그 결과미간이 좁아졌다.
 눈굼둘레뼈
눈굼둘레뼈
4. 영장류는 다른 젖먹이동물에 비해 커다란 뇌를 가지고 있는데, 뇌에서 시각을 담당한 부분은 잘 발달되어 있으나 냄새를 담당하 는 부분은 잘 발달되어 있지 않다. 5. 영장류는 일반화된 이빨울 가지고 있으며(앞니, 송곳니, 옆니, 어 금니 동), 영장류의 종류에 따라 꼬리가 있고 없으며, 앞팔과 뒤 팔 사이의 길이 비율도 종류에 따라 차이가 난다. 6. 영장류는 일정한 형태의 출산행위를 가지고 있다. 그들은- 적은 수의 새끼를 생산하며, 이들을 키우는 데 많은 시간을 보낸다. 새 끼들은 오랫동안 부모에게 의지하여 배우며 자란다. 한 예로 여우 원숭이 Lemur 는 126 일 만에 출산을 하고, 2 년이 지나면 어른이 되며, 11 세 정도가 되면 죽는다• 또 다른 예로 원숭이들은 임신부 터 출산하는 데 걸리는 기간이 168 일 정도이며, 18 달간의 유아기 를 지나 데까지를 어린아이로 보내며, 2 伊] 정도가 되면 죽는다.
사람은 임신부터 출산하는 기간이 약 266 일 정도이며 , Q1 ] 까지가 유아기, l 心1 ] 까지가 어린아이기이며, 어른이 된 후 약 60 - 70 세까 지 산다. 이렇게 보면 영장류들은 다른 젖먹이동물에 비해 〈 삶의 기간 〉 이 접차 길어지는 것을 알 수 있다 . 영장류의 해부학상 • 생리학상 특징의 기원 〈나무 위 생활 가설 The Arboreal Theor y〉과 〈사냥을 위한 시각발달 가설 The Vid u al Predati on Theory > 위에서 영장류로 정의되는 몇 개의 해부학상 • 생리상의 득칭을 살 펴보았다. 왜 이런 특칭들이 영장류의 전화과정중에 생겨났는가 하는 질문이 대두된다. 이에 대한 답은 1920 년대의 신경해부학자인 스미스 G. E. Smit h 가 처음으로 시도하였다. 그에 따르면 영장류의 조상들 은 나무쥐 Shrew 처럼 생겼으며 나무에서 살았는데, 이렇게 나무 위에 서의 생활이 위에서 언급한 특칭들의 원인이 되었다고 한다 . 그의 〈나무 위 생활 가설〉에 의하면 〈냄새를 맡는 능력〉은 나무 위의 생 활에 별로 도움이 되지 못하였기 때문에 영장류의 후각기관은 퇴화하 였고, 대신 시각이 발달하였는데, 특히 입체시각이 발달하였다고 한 다. 이런 입체적인 시각의 발달로 거리를 정확히 측정할 수 있는 능 력이 생겼고, 눈의 위치도 머리 앞쪽으로 이동하게 되었다고 주장하 였다. 손은 주둥이가 하던 역할을 대신함으로써 정교해지고, 길고 뾰족한 손톱은 점차 없어지고 손톱 앞의 촉감이 발달하였다. 손과 발은 나무 를 움켜잡기에 편리하도록 발달하였다. 따라서 나무쥐 같은 동물이 여우원숭이가 되었을 것이며, 이들이 후에 원숭이가 되었을 것이라고 설명하고 있다. 이런 그의 가설은 한 가지 사실만을 제의하고는 매우
설득력있는가설이다• 한편 영 장류학자인 카트밀 Matt Car t m ill은 스미스의 〈 나무 위 생 활 가설 〉 에 의문을 제기하였다. 그는 다람쥐가 머리 양쪽에 눈이 위 치하고 있으며, 잘 발달된 후각과 가늘고 긴 발톱을 가지고 있음에도 불구하고 나무 위에서 잘 살아가고 있는 사실에 주목하여, 스미스 의 〈 나무 위 생활 가설 〉 을 영장류 진화를 소재로 한 소설이라고 비 판하였다. 카트밀은 스미스의 가설에 맞지 않는 다른 동물을 예로 들 고 있으니, 어떤 오스트레일리아산 마수피얼 Marsu pi al 은 7000 만 년 간 나무 위에서 살아왔는데도 불구하고 뾰족한 손톱과 잘 발달된 후 각을 지니고 있으며, 눈은 머리 양쪽에 위치하고 있어 나무 위에서 사는 생활에도 불구하고 신체상의 변화가 없이 살아왔음을 보여주고 있다며 스미스의 가설에 반론을 제기하였다. 카트밀은 스미스의 가설 일부를 차용해서 자신의 새로운 가설인 〈 사냥을 위한 시각발달 가설〉을 내세웠는데, 그의 가설을 요약하면 아래와 같다 ; 〈 영장류의 조상은 땅 위에 살던 식충류로 나뭇잎 사이로 냄새를 맡 으며 곤충인 음식을 찾으러 다녔다. 밀림지대 땅바닥에서는 먹이에 대한 경쟁이 치열하나, 바로 땅바닥 위층인 카노피에서는 먹이들이 풍부하게 홑어져 살고 있었다. 영장류의 조상은 위층에 있는 풍부한 음식을 얻기 위해서 땅바닥에서 나무 위로 자리를 옮겼다. 그러나 카 노피를 돌아다니며 곤충을 주식으로 하던 식충류들은 나무 위에서 살 면서 딱딱한 열매를 먹는 다람쥐와는 다론 문제를 갖게 되었다. 다람 쥐는 먹을 때 소리를 많이 내지만 먹이는 도망을 가지 않고 제자리에 있었으나, 곤충들은 소리에 매우 민감하게 반웅을 하여 재빨리 도망 울 쳐버렸다 . 따라서 영장류의 조상은 그들의 곤충먹이를 더 쉽게 찾 고자 신체의 어떤 부분을 특칭지워서 발달시켜야만 했다. 그 결과 시 력은 먹이인 곤충까지의 거리를 정확히 측정할 수 있도록 발달했으
며, 발은 나무를 움켜잡을 수 있도록 발달함으로써 두 손으로는 곤충 을 잡을 수가 있었다. 시각은 두 눈을 사용함으로써 더욱 발달되었고, 날카롭던 손돕은 편편하게 변해 나무를 잡는 데 편리해졌다. 〉 카트밀에게 있어 영장류란 사냥을 위해서 시력이 변한 동물을 조상 으로 하는 동물집단으로 간주되고 있다고 할 수 있다.
 t
t
오늘날 지구상에서 살고 있는 영장류는 1804 臼千 정도가 되는데, 대 부분은 북위 25 도와 남위 30 도 사이의 열대지방에 한정되어 살고 있 다. 죽 이들 중 약 80% 정도가 남아메리카와 마다가스카르, 아프리카 및 유라시아의 열대 삼림지대에 걸쳐 살고 있으며, 일부는 사바나와 준사막지대에서 살고 있으나, 오스트레일리아에서는 보이지 않는다 (그립 4. 3, 4. 4). 살아 있는 영장류와 사람은 서로의 관계가 가까우면 가까울수록 서
로가 더욱 닮아 보인다. 동물학자들은 이들의 환경에 대한 적응도를 알기 위해서 반드시 관찰방법을 사용한다. 이들은 동물들이 환경에 서로 다르게 적응하는 것과 신체상의 특칭 사이에 어떤 관계가 있는 지를 알고자 하는데, 이를 연구하는 분야를 환경생물학 Or gani smal B i olo gy이라고 한다.
 드유인원
드유인원
오늘날 많은 생물학자들이 세포나 세포보다 조금 큰 단위에서 연구 롤 진행시키고 있다. 그러나 보다 중요한 것은 동물은 그 전체로서 연구되어져야 한다는 것이다. 그러나 실제로 동물을 전체로서 연구한 다는 것은 생물학에서는 가장 어려운 과제 중 하나이다. 이런 점에서 동물을 환경과 동물과의 관계라는 전체적인 입장에서 연구하는 〈생태 학 Ecolo gy〉은 중요한 연구분야로 인식되고 있다. 생물 중 특히 동물과 그들이 살고 있는 환경과의 관계를 연구하는 (동물)생태학에서 다루는 가장 중요한 대상은 동물과 그들이 섭취하
는 음식과의 관계이다. 이를 위해서 생태학에서 사용되는 몇 개의 기 본용어를 살펴볼 필요가 있다. 한 동물이 사는 장소를 〈본고장 Hab i t a t〉아라고 하며, 〈삶터 (Ni ch e : 생태조건을 가진 터전의 뜻)〉는 이들 이 매일매일 지내는 장소를 말한다. 〈분포구역 Ran g e 〉은 한 종의 동 물이 얼마나 퍼져 있는가를 나타낼 때 쓰인다. 모든 동물들은 그들의 〈삶터〉에 적응한다. 전화란 기회성을 지닌 것으로 환경은 동물에게 전화할 수 있는 기회를 제공하고, 동물은 이 환경에 따라 변한다. 〈원〉영장류 Pro t o-Pr i ma t es 가 진화할 수 있었 던 원인도 바로 환경에 있었으며, 이들은 환경의 이점을 살려서 진화 하였다. 생명체의 전화는 어떤 정해전 목표에 도달하기 위한 것이 아 니라, 기회에 따라 일어나는 것이다. 영장류의 행위 영장류의 식성과 걷기방법, 사회행위는 우리가 가장 관심을 가지고 있는 영장류의 행위에 관한 연구분야인데, 행위란 동물툐 을 특칭지워 주는 것 중의 하나이며, 행위는 · 동물이 살고 있는 모든 환경(사회적 환경과 자연적 환경)과의 상호작용에 의해 이루어전다고 정의될 수 있다. 영장류는 크게 작은 몸집의 프로시미안과 보다 큰 몸집을 가전 앤 스로포이드로 나누어지며, 생활유형에 따라 야행성과 주행성으로 구 분된다. 대다수의 프로시미안은 야행성인 데 반해 많은 수의 앤스로 포이드는 주행성이다. 프로시미안 같은 야행성 영장류는 살아가는 데 주행성 영장류보다 많은 이접이 있다. 먼저 밤에는 식육류로부터 도 망치기가 용이할 뿐만 아니라, 특히 식육류가 주행성인 경우에 위험 성은 거의 없다. 또 다른 이점으로 대낮의 열로부터 보호된다는 점을
들 수 있으며, 또한 곤충 같은 먹이는 밤에 손쉽게 얻어진다는 점이 다. 그러나 이런 이점들이 있는 반면 야행성 프로시미안은 밤에 물체 를 보기가 쉽지 않디는- 단점을 지니고 있다.
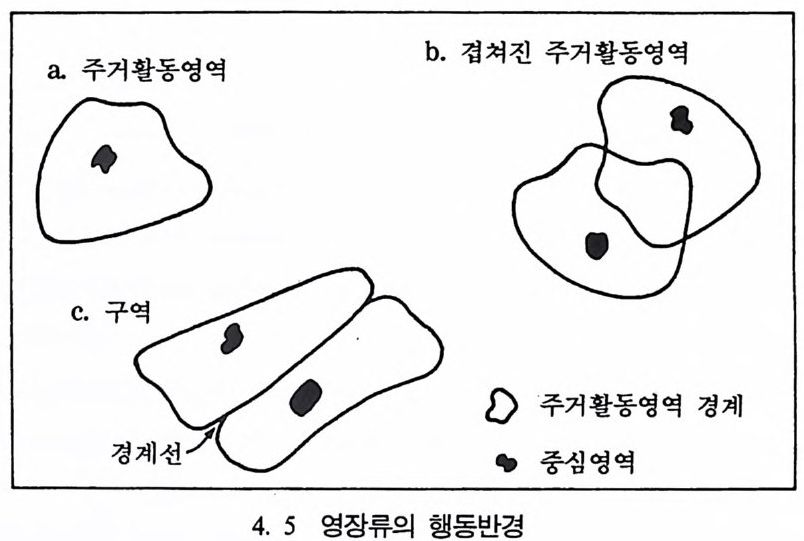 >a 주거활동영 역 b. 겹쳐진 주거활동영역
>a 주거활동영 역 b. 겹쳐진 주거활동영역
한편 영장류를 연구할 때는 이들의 활동과 자연환경과의 관계라는 측면에서 살펴보아야 한다. 이를 위해서는 영장류가 어떻게 땅을 이 용하는지에 연구 초점을 맞추어야 한다(그림 4. 5). 동물이 아침에 일 어나서 잠들 때까지 돌아다니는 구역을 〈한낮의 활동영역〉이라고 하 며, 어른이 되어서 돌아다닐 수 있는 지역의 범위를 〈주거활동영역〉 이라고 한다. 지역이란 말은 특별한 의미로 주거활동영역의 일부를 의미하는데, 실제로 자기가 사용하는 구역을 가리킨다. 중심영역이란 개념 또한 주거활동영역의 일부로 항상 살고 있는 영역의 범위를 말 한다. 영장류의 행위를 보다 정확히 이해하기 위해서는 바로 위에서 언급한 〈땅의 이용〉이라는 측면에서 접근하는 것이 이들의 행위를 보 다 정확히 이해하는 데 도움을 준다. 실제로 대부분의 영장류는 숲속 의 바닥이나 윗부분이 아니면 . 나무 중간부분에서 살고 있다.
영장류의 몸집과 식성 및 걷기방법 영장류의 식성과 걷기방법은 동물의 몸집 크기와 밀접한 관계가 있 으며, 나아가 몸집의 크기는 영장류의 사회구성, 그리고 거주지 등을 연구하는 데 가장 중요한 기준이 된다. 영장류는 종류에 따라 몸집의 크기가 다른데, 그 중 사람의 몸집이 가장 큰 편에 속한다. 영장류의 몸집은 100그 램 정도의 몸무게를 가전 쥐안경원숭이로부터 180 킬로그 램이 넘는 고릴라에 이르기까지 다양하다. 이렇게 다양한 동물 몸집 의 크기가 신체 다른 부분의 성장이나 또는 행위와 어떻게 관계가 있 는가를 연구하는 학문을 〈대비성장측정학 Allome t r y〉이라고 한다.
 ` I `` ``` ` ` ` 晶./. / .. .. ..
` I `` ``` ` ` ` 晶./. / .. .. ..
영장류의 경우 몸의 크기와 음식물은 밀접하게 관련되어 있는데, 일반적으로 영장류가 먹는 음식물은 식충류와 과일류 그리고 잎사귀 로 나누어진다. 만일 어떤 영장류가 식충과 과일 그리고 잎사귀를 각 각 33% 씩 먹는다면 그 영장류의 식성은 정중앙에 있다. 어떤 원숭이 는 잎사귀만을 먹으며, 프로시미안인 어떤 늘보원숭이는 식충류만을 섭취하며, 어떤 원숭이와 침팬지 등의 유인원은 주로 과일을 먹는다.
어떤 원숭이는 과일과 잎사귀를 먹고, 어떤 원숭이는 식충과 과일을 먹는다. 그러나 식충과 잎사귀를 함께 먹는 영장류는 거의 없다(그립 4. 6). 식충류를 먹는 영장류와 식충류와 과일을 함께 먹는 영장류는 작은 몸집을 가지고 있으며, 잎사귀를 먹는 영장류와 잎사귀와 과일을 같 이 먹는 영장류는 몸집이 크다. 예로 몸무게가 1k g인 영장류는 매일 150 마리 정도의 귀뚜라미를 먹어야 하는데, 따라서 몸무게가 10kg 정 도 나가는 식충류 영장류는 매일 1500 마리 정도의 귀뚜라미를 잡아야 만 한다. 만일 한 지역에서 음식물인 곤충의 숫자가 제한된다면, 이에 따라서 식충류 영장류의 몸집 크기도 제한된다. 다론 예로 잎사귀롤 먹이로 하는 영장류의 경우, 먹이룰 찾기는 쉬 우나 소화시키기는 어렵다. 죽 잎사귀들은 씹기가 힘들고 영양분은 셀룰로오스에 함께 섞여 있으며, 가끔 독성을 가지고 있기도 하다. 이 독성은 잎사귀들이 자신을 먹는 동물둘 2 멀리하게 하는 효과있는 방 어수단인데, 이런 성질을 가진 잎사귀를 소화시키기 위해서는 많은 시간과 큰 소화기관이 필요하다. 따라서 잎사귀를 주식으로 하는 영 장류는 커다란 몸집을 지니고 있다. 과일은 영장류가 가장 좋아하는 음식물이며 또한 많은 칼로리를 지 니고 있으나, 단백질의 양이 많지 않으며, 이들은 계철을 탄다는 단점 을 가지고 있다. 따라서 과일을 주식으로 하는 영장류는 작은 소화기 관을 가지고 있으며, 결과적으로 잎사귀를 먹는 영장류보다는 작은 몸집을가지고 있다. 영장류의 몸집은 걷는 방법과 밀접한 관계를 가지고 있다. 어떤 영 장류는 나무 위 에서 네 발을 이용하여 Arboreal Qua drupe d s 이동을 하며, 쥐여우원숭이 같은 프로시미안 영장류는 땅 위를 깡총깡총 뛴 다 Lea pe r. 한편 팔을 사용하여 움직이는 방법 Sus pe nsor y이 있는데,
커다란 몸집을 가전 영장류는 대개가 이 방법으로 이동을 한다. 이런 걷기방법을 가전 영장류 중 두 팔을 사용하여 빨리 움직이는 긴팔원 숭이 같은 영장류를 나무가지타기 Brach i a t or 라고 한다. 영장류 중 가 장 큰 몸집을 가전 고릴라는 땅바닥에서 네 발로 돌아다니고 Terre s tri a l Qua drupe d s, 침팬지는 나무 위에서는 두 팔을 사용하며 땅 위 롤 돌아다닐 때는 네 발을 사용하는데, 이때 침팬지는 고릴라처럼 바 깥 손마디로 땅을 짚고 걷는다: Knuckle : Walki ng . : 끝으로 영장류 중 에서 사람만이 유일하게 땅 위에서 두 발로 걷어 다닌다 B ip edal Locomoti on . 이렇게 영 장류의 각기 다른 〈걷기 방법 〉은 사람과 사람 이의의 영장류를 구분하는 특칭의 하나로 이용된다. 영장류 무리의 사회구조 영장류 사회란 함께 살아가는 영장류들의 무리를 뜻하는데, 이들이 보여주는 사회행위는 영장류가 그들의 사회에 적응하는 중요한 방법 이다. 영장류는 계급사회에서 살고 있기 때문에 한 지도자나 개인, 또 는 집단무리에 의해 통솔된다(사전 4. 7). 무리에는 친구와 적, 그리
떼를 지어 살 때의 이점과 불리한 점 이점 불리한점 배우자선택 용이 배우자선택을 위한 경쟁 사냥의 경우 ; 사냥 대상물의 발견용이 맹수로부터 방어 용이 맹수에게 쉽게 노출 음식 획득 집단에서의 지식 전달 가능 경쟁 음식물의 보호 음식을 얻기 위한 여행
고 일반적인 관계를 나누는 부류 등이 섞여 있으며, 또 무리 중에는 인기가 있는 놈과 없는 놈, 그리고 평범한 놈 등이 있다. 영장류의 어린 새끼들은 그들의 사회에서 살아갈 수 있도록 사회행 위와 더불어 소리와 행동(움직임) 등을 포함한 복잡한 의사소통 방법 울 배워야 한다. 바로 이런 점들이 인간사회에서 일어나는 행위와 같 기 때문에, 인류학자들로 하여금 영장류를 연구하게 만든다. 그러나 영장류 연구를 통해 인류의 행위에 관해 너무 많은 것을 얻으려고 하 는 데는 문제가 있음을 알아야 한다. 가장 작은 몸집의 프로시미안 영장류를 제의하고 대개의 영장류는 무리를 지어서 사는데, 이렇게 무리를 지어 사는 데는 좋은 점과 나 쁜 점이 있다.
 --,.
--,.
영장류의 사회행위의 유형은 아프리카에서 살고 있는 영장류의 무 리 구조연구를 통해 비교적 찰 알려져 있다. 영장류 무리의 구조에 대한 연구를 위해서 〈공간분포〉와 〈무리구성〉의 두 가지의 요인을
살펴볼 필요가 있다. 〈공간분포〉는 이 무리들이 어디에서 살며 어떻 게 분포되어 있는가를 뜻하며, 〈무리구성〉은 무리에 속해 있는 개개 의 숫자나 중류 들을 말한다 . 우리가 아프리카에 가서 영장류~ 사회행위를 관찰한다면, 먼저 혼자서 살아가는 So lit ar y 영장류를 볼 수 있는데, 이들은 서로가 소 리로 신호를 하거나 또는 냄새와 특별한 체액을 땅 위에 홀리는 방법 으로 신호를 전달하기도 한다. 다른 사회형태로 암수짝 Consor t Pa i r 을 들 수 있는데, 이는 수컷 한 마리와 암놈 한 마리가 한동안 함께 사는 형태를 말한다. 이들은 어미 /아비와 자식들로 구성된 가족단위로 이들의 • 결합은(암놈과 수 놈) 오래 지속되는데, 이런 유형의 사회결합은 영장류에는 매우 희귀 한 형태이다. 떼 Con g er y는 결합력이 강하지 않은 무리를 뜻한다. 예로 한 떼의 원숭이들이 과일을 따먹기 위해서 한 나무에 몰려들었다가 과일을 다 따먹으면 다시 흩어지는데, 이런 유형의 사회결합을 말한다. 이의에 암수 섞인 무리 Troo p s 는 많은 암놈과 수놈 그리고 그들의 자식들이 낮에 활동할 때 함께 무리를 이루는 것을 말하며, 숫왕암무 리 Harem 는 수놈 하나에 많은 암놈과 새끼들로 구성된 형태이며, 의 쪽무리 Band 는 오직 한쪽의 성을 가전 놈들로 구성된 무리를 말한다. 위에서 언급한 영장류의 사회구조유형은 비록 모든 종류의 사회유 형을 포함하고 있지는 않으나 영장류가 공간상으로 어떻게 분포되어 있는가를 살펴보는 데는 매우 편리하다. 한때 영장류의 행위는 동물원 등에서 관찰되었는데, 이렇게 동물원 에서 관찰된 행위는 정확하지 못하다는 사실을 알게 된 지는 얼마 되 지 않는다. 그 동안 동물원에서 이루어전 연구는 영장류의 사회행위 에 대하여 잘못 설명되었으니, 우리에 가두어놓은 결과 이들은 비정 상 행위를 나타내었다는 사실이 밝혀졌다. 따라서 학자들은 영장류를
연구하기 위해서 야의로 나가기 시작하였다. 그러나 야의 연구는 먼 저 방법보다 훨씬 진전된 방법이긴 하지만 아직도 잘못된 결론이 나 타나고 있음을 알게 되었다• 오늘날 영장류의 행위를 연구하는 학자 들은 영장류가 가끔 그들이 처해 있는 생태상의 조건에 따라 다른 행 위를 보여준다는 사실을 알게 되었다(표 4-1).
표 4. I 공간상 분포로 본 영장류의 사회유형 침팬지 : 암놈어미와 새끼로 구성되며, 수놈아비는 암놈과 새끼로 부터 떨어져서 의쪽무리에서 산다. 고릴라 : 수놈아비 한 마리와 여러 암놈 및 새끼로 구성되나, 다 론 수놈들은 무리 밖에서 혼자 산다. 새끼수놈은 자라 성인이 되면 집단을 이탈하여 다른 집단의 암놈과 짝을 짓는다.
긴팔원숭이 : 암놈어미와 수놈아비 및 새끼의 가족단위로 이루어 져 있다. 질라다 비비 : 숫왕암무리와 수놈 무리가 함께 뒤섞인 형태의 구 조를 이루고 있다. 부쉬베비 : 어미 하나와 새끼들로 구성된 무리로 이루어져 있으 며, 암놈둘이 사는 구역은 혼자 살고 있는 수놈들의 구역과겹친다.초기 영장류 연구에 따르면 어떤 종의 경우는 배우는 과정 중에 행 위가 특이하게 나타나거나, 아니면 태어날 때부터 그 특칭을 이미 지 니고 있다고 생각하였다. 물론 어떤 종의 행위들은 태어날 때부터 지 니고 있는 것으로 알려져 있다. 예를 들어 흰개미는 태어날 때부터 지 니고 있는 행위의 결과로 멋진 터널과 식민지를 건설한다. 그러나 영 장류의 많은 행위들은 그들이 배워서 얻어지는 것이지 태어날 때부터 지니고 나오는 것이 아니기 때문에 그들의 사회행위는 신축성이 있다.
영장류의 분류 영장류의 분류는 일반적으로 〈아목 단계 Suborder 〉에서 앤스로포이 드와 프로시미안(또는 프로시미)으로 구분하는 방법과 안경원숭이 Tars i ds 와 늘보원숭이 Lor i s i ds 가 어디 에 속하느냐에 따라 하프로라 이니 (Ha p lorh i nes : 앤스로포이드에 안경원숭이를 포함)와 스트렙시라 이니 (S t re p s i r hi nes : 프로시미안에 안경원숭이룰 제의)로 구분하는 방 법이 있다(표 4. 2). 이때 앤스로포이드와 프로시미안의 구분은 일반 적인 특칭인 몸의 크기, 음식, 행위 등을 기준으로 한다. 이에 반해 가지치기방법은 해부학상의 특칭을 기준으로 하는데, 특히 코의 모습 을 기준으로 한다. 위의 두 방법 중 어느 쪽이 더 좋은 방법이라고 말할 수 없는데 그것은 진화가 우리를 위해 일어나는 것은 아니기 때 문 0] 다.
표 4. 2 영장류의 분류 목 아목 과 종 S p ec i es Order Suborder Fa mily 의수 앤스로포이드 인류과 Hom inida e l Anth rop o id s 성 성 원숭이 과 Pong ida e 4 긴팔원숭이과 Hy lo bati da e 9 영 긴꼬리 원숭이 과 Cercop ithe c ida e 82 칼리트리시드과 C allit r ici dae 21 장 세비드과 Cebid a e 30 프로시미안 안경원숭이과 Tars i dae 3 류 Prosim ian 늘보원숭이 과 Lor i s i dae 3 또는 다우벤토니과 Daubento n id a e l 프로시미 짧은꼬리 여우원숭이과 lndrid a e 4 Prosim ii 여우원숭이과 Lemur i dae 17
프로시미안은 야행성 동물로서 사람이 접근하기 어려운 지역에서 살며, 또한 인류와는 닮은 데가 별로 없기 때문에 인류학자들은· 이들 에게 많은 관심을 보이지 않는다. 그러나 오늘날 살고 있는 프로시미 안을 잘 관찰해 보면 상당히 매력있는 동물임을 알 수 있는데, 이는 영장류 역사상 처음 3000 만 년 동안 살았던 영장류는 오직 프로시미 안뿐이기 때문이다. 프로시미안 영장류들 오늘날 프로시미안 영장류 중 여우원숭이와 디우-벤토니과, 짧은꼬 리여우원숭이는 아프리카의 마다가스카르 섬에서 살고 있다. 마다가 스카르 섬은 지구상에서 네번째 큰 섬으로, 길이는 약 1600km7l- 넘 는다. 아프리카 대륙 동쪽에 위치하고 있으며 생태계는 열대 삼림지 대로부터 사바나에 이르기까지 다양한데, 프로시미안 영장류는 삼림 지대에 살고 있다. 이 섬은 약 5000 만 년 전 팔레오세에 아프리카 대 륙으로부터 떨어져 나왔는데, 그 이후 이 섬에 고립된 프로시미안 영 장류가 이 지역에서 번창하였으며, 그 결과 마다가스카르 섬은 오늘 날 프로시미안 영장류 연구의 중심지가 되고 있다. 한편 늘보원숭이 는 아프리카와 동남아시아에서 살고 있으며, 안경원숭이는 동남아시 아에서 살고 있다. 나무뒤주 ITree Shrew 영장류는 아니지만 영장류와 닮은 뒤쥐 Sorex 속에 속하는 나무뒤쥐에 관해 살펴보기로 한다. 이 나무뒤취는 실제 로는 쥐가 아니며 또한 나무에서 살지 않는다. 구대륙의 열대지방에 모두 18 종이 분포되어 살고 있고, 평균 크기는 다람쥐만하며 식충류 이다. 나무뒤쥐는 매우 빨리 성장하여 생후 4 개월 후에는 성숙한 개 체가 된다. 둥근 손톱 대신에 날카로운 손톱, 그리고 긴 코롤 가지고
있으며, 머리 양쪽에 눈이 자리하고 있다. 일반적으로 나무뒤쥐는 영 장류에서 보이는 해부학상 특칭이 별로 없어서 영장류로 분류되지는 않고 있지만, 이들은 영장류의 조상들과 닮았으리라고 여겨진다(그림 4. 8).
 두더쥐
두더쥐
여우원숭이 Lemur 첫번째 프로시미안 영장류로는 여우원숭이를 꼽 을 수 있는데, 이 종류에는 여우원숭이과와 짧은꼬리여우원숭이과, 다 우벤토니과 등의 3 과가 있으며, 이들 모두가 마다가스카르 섬에 살고 있다(사전. 4. 9). 여우원숭이과에 속하는 영장류는 대개가 나무 위에서 살고, 그 중 쥐여우원숭이 Mouse Lemur 가 가장 작으며, 이들은 주로 곤충이나 과 일을 먹고 산다. 이때 쥐여우원숭이는 난쟁이여우원숭이처럼 영양분 울 꼬리에 저장했다가 가뭄이 드는 계철이나 동면 때에 이를 사용한 다. 이들은 야행성 영장류이다. 여우원숭이속에는 많은 종들이 있는데, 이들 중 몸집이 큰 종류는 대개가 주행성이다. 알락여우원숭이는 잡식이며 대부분 땅 위에서 살 고, 붉은여우원숭이는 나뭇잎을 먹고 산다. 이들은 새끼를 한 마리만 낳으며, 영장류의 특칭인 입체적인 시각을 가지고 있으나 아직도 긴
주둥이와 축축하게 젖은 코를 지니고 있다. 여우원숭이과에 속하는 영장류는 앞니와 송곳니가 마치 머리빗처럼 생겼는데, 이런 이빨은 털을 손질하거나 음식을 먹는 데 이용되며, 또 한 나무줄기에서 스며 나오는 고무액 같은 것을 잡아당기는 데 쓰인 다(그립 4. IO).
 4. 9 쥐여우원숭이
4. 9 쥐여우원숭이
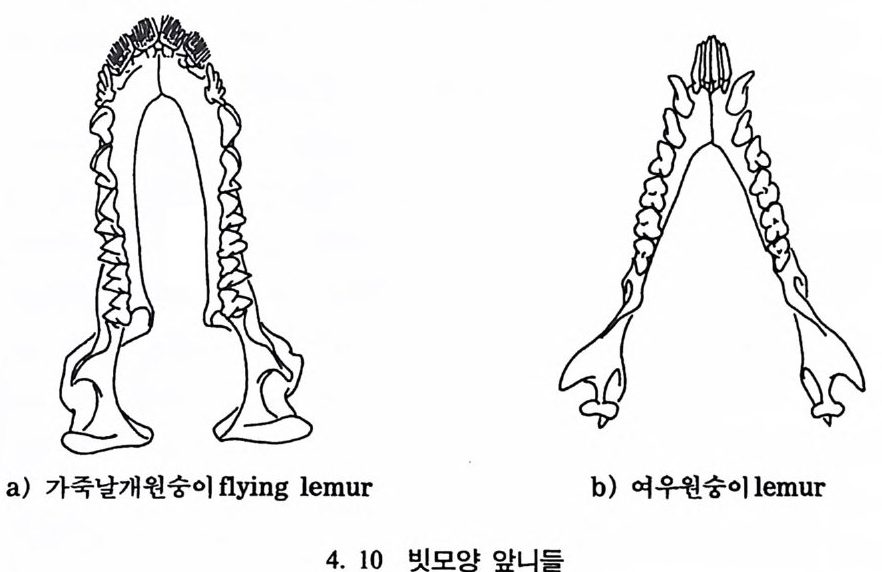 a) 가죽날개원숭이 flying lemur b) 여우원숭이 lemur
a) 가죽날개원숭이 flying lemur b) 여우원숭이 lemur
여우원숭이 중에 가장 큰 무리는 짧은꼬리여우원숭이과에 속하는 종류로 모두 3 개의 속으로 구성되어 있다(그립 4. 11). 그 중 아바하 이 Ava hi가 가장 작은 몸집을 지니고 있고 야행성이며, 이의의 속둘 은 주행성이다.
 a) 인드리원숭이 b) 아이-아이 원숭이 c) 여우원숭이
a) 인드리원숭이 b) 아이-아이 원숭이 c) 여우원숭이
프로시미안 중에서는 인드리 Indr ii가 가장 큰 몸집을 가지고 있는 데, 주로 나뭇잎을 먹고 살며 일부일처제를 유지하고 있다. 인드리 는 〈바로 서서 뛰어오르는 걷기〉 방법을 지니고 있기 때문에 멀리 점 프할 수 있으며 이에 필요한 긴 뒷받을 가지고 있다. 다우벤토니과에 속하는 아이-아이 A y e - a y e 는 여우원숭이 중에서 특히 특수화된 해부학상 특칭을 지닌 프로시미안이다 . 몸무게는 약 2- 3kg 정도이며, 설치류를 닮은 앞니를 가지고 있고 나무 그루터기에 서 산다. 또한 아이-아이는 매우 긴 손가락과 발가락 그리고 뾰족한 긴 손톱과 발톱을 가지고 있는데, 그 중 하나가 더욱 길다. 이 긴 손 가락으로 나무에 구멍을 뚫어 먹이감둘이 움직이는 소리를 듣는다. 그리고는 나무를 갑아낸 다음 긴 손가락을 창끝처럼 사용하여 먹이감 을 찔러 나무 밖으로 끄집어낸다. 이런 특칭 때문에 아이-아이는 딱 따구리 영장류라고도 불린다.
늘보원숭이 Lorie s 늘보원숭이과에 속하는 프로시미안 원숭이는 마 다가스카르 섬 이의에 아프리카와 동남아시아에서 살고 있다. 이 프 로시미안 영장류는 머리빗 모양의 이빨을 가지고 있는 등 여러 면에 서 여우원숭이와 닮은 해부학상의 특칭을 지니고 있다. 모두가 야행 성이며, 땅 위를 매우 천천히 기어서 움직인다(사전 4. 12 ; 그립 4. 13).
 4. 12 수마트라 늘보원숭이
4. 12 수마트라 늘보원숭이
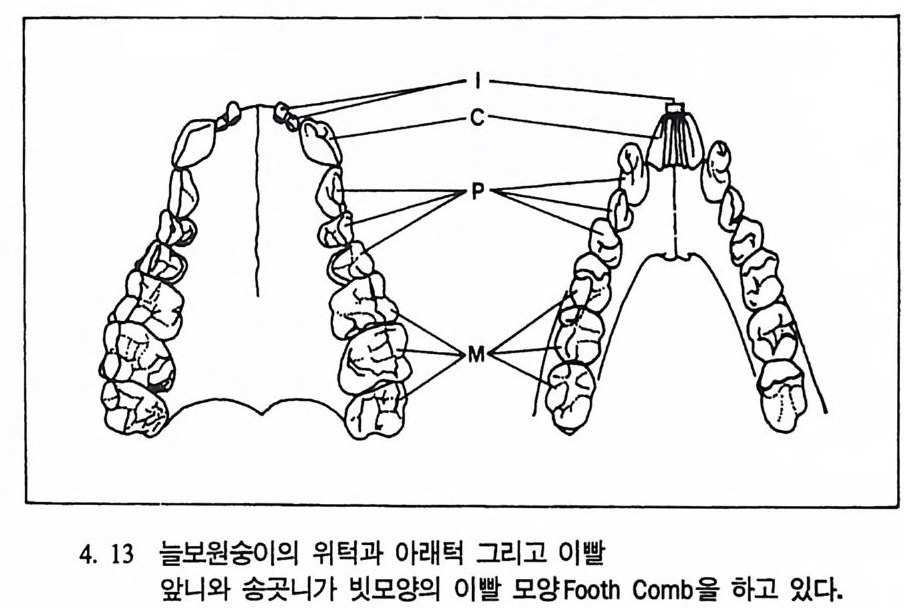 IC
IC
아프리카 대륙에 살고 있는 포토 Po tt o 와 앙완티보 An g wan ti bo 는 천천히 움직이는 접에서는 서로 닮았으나, 포토는 길쭉한 몸매를 지 니고 있는 데 비해 앙완티보는 통통한 몸매를 가지고 있다. 모두 야 행성으로 과일과 곤충 그리고 껌울 먹는다. 몸무게는 몇 그램에서 몇 킬로그램에 이르기까지 다양하다. 부시베비 Bush bab y는 행동이 민첩 하고 재빠른 점을 제의하고는 다른 늘보원숭이와 비슷하다.
 4. 14 안경원숭이
4. 14 안경원숭이
안경원숭이과 프로시미안은 안경원숭이 1 속에 3 종의 안경원숭이로 구성되어 있다(사전 4. 14). 다른 프로시미안들은 윗입술에 붙어 있는 축축하고 윤기있는 코를 가지고 있어 스트렙시라이니로 분류된다. 그 러나 안경원숭이는 앤스로포이드와 같이 마른 코를 가지고 있어 이를 하프로라이니로 분류하고 있다. 안경원숭이의 발가락은 매우 길어(그림 4. 15) 한 나뭇가지에서 다
론 나뭇가지로 뛰어 옮겨가는 데 매우 유용하다. 몸무게는 약 lOOg 정도이며 매일 몸무게의 10% 정도의 음식을 먹는다. 야행성으로 식 충류이다. 눈알이 매우 커서 각각의 눈알 무게는 두뇌 전체보다 더 · 무게가나간다 .
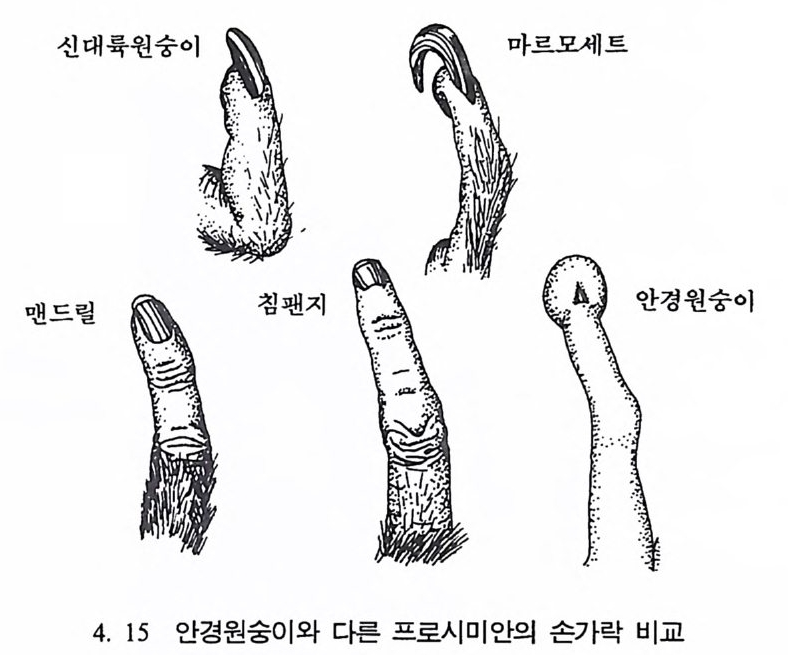 트
트
최초의 영장류 : 프로시미안 영장류의 기원과 진화 영장류의 진화과정 중 전반부의 몇천만 년 동안에는 프로시미안 영 장류만이 살고 있었다. 이들 중 오늘날까지 살아 있는 프로시미안 영 장류의 수는 극히 일부에 속하며, 이들의 과거를 돌아보기 위해서는 이빨, 턱, 뼈 등의 화석을 살펴보아야 한다. 최초의 영장류는 약 6500 만 년 전 공룡들이 살았던 시기에 함께 살던 젖먹이동물로부터 진화되었는데, 지질학상의 연대로는 중생대 말 백악기 말기와 팔레오제의 경계시기에 처음 나타난다(표 4. 3). 그
당시 북아메리카 대륙의 기후는 아열대기후였으며, 오리주둥이공룡
표 4.3 지구의 지질시대 구분 시간 세 (Ep o ch) 대 (Era) 현재 홀로세 (Holocene) 1 만 년 풀라이스토세 (Pleis t o cene) 180 만 년 전 풀라이오세 (Pli oc ene) 신생대 CENOZOIC 500 만 년 전 마이오세 (Mi oc ene) 1500 만 년 전 울리고세 (Ol igo cene) 3500 만 년 전 이오세 (Eocene) 5500 만 년 전 팔레오세 (Paleocene) 6500 만 년 전 중생대 백악기 (Creta c eous) MESOZOIC * 지질학에서 세(世 : E p och) 를 나타내는 용어는 끝에 -cene 이 붙는데, 이는 대 (代 : Era) 의 한 시기를 나타낸다.
Duck- B il led d i.과 트리케라톱스 Tr i cera t o p s, 그리고 키가 15m 나 되 는 티라노사우르스 렉스Ty rannosaurus rex 같은 거대한 공룡들이 지상 에서 살고 있었다. 숲속에서 잘 보이지는 않았지만 최초의 포유류인 마수피얼과 멀티튜베르큐레이트 Mul tit ubercula t es 그리고 식충류 등 이 살고 있었다.
 ?
?
여러 해 동안 학자들은 백악기 /팔레오세 경계를 지구에서 살았던 생명체들의 역사에 중대한 위기가 왔던 시기로 생각해 왔다. 당시 바 다 속에 쌓인 퇴적물을 연구해 본 결과, 많은 플랑크돈과 다른 생물들 이 이 경계시기에 멸종되었음을 알았다. 살아 남아 있던 공룡들도 이 시기에 절멸하였으며, 이 짧은 시기 이후에 다양한 종류의 젖먹이동 물들이 지구상에 나타났다. 오늘날 중생대와 신생대의 경계에서 일어
난 생명체의 대절멸과 젖먹이동물의 출현을 가져온 지질상의 변화가 왜 일어났는지에 관해 많은 가설이 있으나, 아래의 가설들이 많은 지 지롤 받고 있다(사진 4. 16). 아스테로이드 As t ero i d 혜성에 의헌 절멸설 학자들은- 백악기/팔레오세 경계시기에 지구상에서 일어난 생명체의 절멸과 전화는 왜 일어났는 지롤 규명하고자 노력해 오고 있다. 1979 년 미국 캘리포니아 주립 버 클리 대학교의 물리학자인 루이스 알바레즈와 그의 아들이자 같은 대 학교의 지질학자인 월터 알바레즈는 지구상에서는 별로 존재하지 않 는 원소인 이리디움i r idi um 이 백악기 /팔레오세 경계시기의 충에서만 상당량 검출된다는 것을 알았다(사전 4. 17). 이를 바탕으로 알바레즈 부자는 이 시기에 지구상의 생명체를 붕괴시킨 한 커다란 혜성의 존 재를가정하였다. 백악기 /팔레오세 경계시기에 이 혜성이 지구와 충돌하였는데, 이 충돌로 인한 충격과 위력은 약 50 메가톤의 수소폭탄의 폭발과 맞먹는
 `
`
효과를 가져왔으며, 폭발의 결과로 생긴 먼지가 대기를 뒤덮어 태양 열을 차단하자 공룡 같은 냉혈동물들은 절멸하게 되었다고 주장한다. 만일 6500 만 년 전에 대절멸이 없었더라면 공룡 같은 파충류들은 아마 지금까지 살았을지도 모른다는 점에서 대절멸은 생물의 전화과 정에서 매우 중대한 사건이다. 커다란 파충류의 절멸은 결과적으로 인류의 조상을 포함한 다른 생물들이 살아 남을 수 있는 기회를 제공 하는 계기가되었다. 절멸의 주기성 이론 : 니메시스 Nemes i s 함성 한편 다른 - 학자들은· 지 구상에서 생명체 멸종의 주기성에 대해서 연구를 진행하고 있다. 지 구의 역사에서 볼 때 많은 생물체가 멸종되어왔는데, 이런 현상은 약 2600 만 년을 주기로 반복되어 나타난다. 이를 연구하는 학자들에 따 르면 이런멸종의 주기성은 아마도 태양이나 태양계의 조직, 또는 우 주 내에서 진행되는 운동 작용과 관련이 있다고 한다. 즉 우주에서는 태양계에 영향을 미치는 니메시스라는 항성이 있는데, 이 항성은 주
 4. 18 니메시스 항성 이론
4. 18 니메시스 항성 이론
기적으로 우주의 물체를 태양계 밖에서부터 안쪽으로 잡아당긴다. 이 때 잡아당겨져 태양계로 들어온 물체가 지구와 충돌하고 그 결과 지 구상에 생명체의 절멸이 일어나는데, 이런 현상은 일정한 주기를 가 지고 반복해서 진행되어왔다고 주장한다(사진 4. 18). 6500 만 년 전 지구에는 나무가 무성한 아열대기후에 트리케라톱스 와 프데로닥티라 P t erodac ty lar 같은 공룡들이 살고 있었다. 당시 공 룡둘의 발치에서 살던 최초의 젖먹이동물들은 작은 몸집에 뾰족한 코 를 지니고 있었는데, 이들은 마치 오늘날 살고 있는 식충류와 닮은 모습을 하고 있었다. 이 당시의 지구는 기후와 지형이 오늘날과는 달 랐는데, 대륙은 서로 함께 붙어 있었으며 육지와 바다는 고정되어 있 지 않고 움직이고 있었다. 최초의 영장류 화석은 북아메리카 대륙에서 백악기/팔레오세 경계 바로 밀층에서 발견되었다. 발견된 화석은 대개 이빨과 턱 들뿐이며 초기 영장류는 단지 이들의 어금니를 가지고 구분되었다. 최초의 영 장류의 어금니는 그다지 날카롭지 않고 식충류의 어금니 정도로 뾰족 한 모습을 지니고 있다. 최초의 영장류는 북아메리카 대륙의 몬타나 주에 있는 퓨가토리 언덕에서 발견되었는데, 이 화석은 발견된 지명 을 따서 퓨가토리우스g enus Pug ato ri us 속으로 명명되었다. 당시 이 지 역은 온대성 기후를 띠고 있었다. 퓨가토리우스 화석은 뭉뚝한 어금니를 제의하고는 영장류의 특칭을 별로 지니고 있지 않아 일부 학자들은 퓨가토리우스를 영장류로 분류 하지 않으나, 대다수는 최초의 영장류로 인정하고 있다. 영장류의 다양성은 퓨가토리우스와 이를 닮은 영장류로부터 시작되 었는데, 이들을 〈옛 영장류〉 또는 〈플레시아다피형 영장류 (Arch ai c 또는 Plesia d ap iform pr im ate s ) 〉라 한다. 이들 화석은 영 장류의 특칭 울 지니고 있지 않고 너무 특수화된 특칭을 갖고 있기 때문에 카트밀 같은 학자는 이들을 영장류로 구분하지 않고 있으나, 대부분의 학자
둘은 이들을 영장류의 먼 조상으로 받아들이고 있다. 영장류 조상의 뼈둘은 유럽과 북아메리카 대륙에서 발견되는데, 당 시의 이 대륙들은 서로 붙어 있었다. 이 지역에서 얻어지는 뼈들의 40% 정도가 플레시아다피형 영장류의 뼈로 당시에는 모두 6 과가 있 었으나 모두 절멸했다. 속으로 구분된 무리로 플레시아다피스 Ples i adap i s 가 있는데, 이들은 설치류와 같은 특칭을 지니고 있다. 이들은 매우 큰 앞니와 이 앞니 바로 뒤에 간극이라는 이빨 사이의 틈새가 있다. 이들은 커다란 주둥이, 뾰족한 손톱과 발톱을 가지고 있으며 엄 지발가락은 다론 발가락들과 나란하다. 몸무게가 6- 7 kg 정도에 달하 는 종류도 있었다. 팔레오세 초기에 남아메리카 대륙과 인도는 섬이었으며 유라시아와 북아메리카 대륙은 서로 연결되어 있었다. 이 당시의 기후곡선을 보 면 공룡이 살았던 당시의 매우 높았던 기후는 팔레오세에 오면서 기 온이 떨어전 사실을 알 수 있다. 이 시기에 다람쥐나 꾸근꾸뜨 크기 의 초식류와 다람쥐 정도의 식육류 같은 플레시아다피형의 젖먹이동 물들이 급격히 퍼져 나갔다. 플레시아다피형의 영장류들은 모두 커다란 앞니와 이빨 틈새를 가 졌는데, 어떤 종류는 풀을 찰라 먹을 수 있도록 커다란 칼날 같은 이 빨울 가진 턱을 가지고 있으며, 또 다른 종류의 화석은 과일박쥐와 닮은 이빨을 가지고 있다. 이들의 턱은 설치류를 닮았으며 눈굼 안쪽 에 뼈로 된 벽과 눈굼 둘레의 뼈가 발달하지 않아 음식을 씹울 때마 다 근 육 오 동에 의해 눈이 움직였다(그립 4. 19). 〈현대의 여명〉이란 뜻을 지닌 이오세에 오면 떨어졌던 기온이 점차 상승하기 시작하는데, 이 시기의 지구상에는 최초의 짝굽동물(우제류 Arti od acty la r : pig- li ke mammals) 과 홀굽동물(기제류 Peris s od-acty la r : ·ho rse-li ke mammals) 이 출현하였다. 오늘날 살고 있는 동 물의 조상들로서 분류할 수 있는 동물들이 처음으로 나타나기 시작한
시기였다. 이오세 초기에 유럽과 북아메리카 대륙은 여전히 붙어 있었으며, 여기서 살고 있었던 영장류들은 서로 비슷하게 닮았다. 한편 이오세 에 폴레시아다피스형의 동물들은 그 수가 점점 줄어들기 시작하며, 열대지역은 북극 한계선까지 확장되었다. 이오세 중에 유럽과 북아메 리카 대륙은 분리되었으며 이 시기에 현대 영장류의 조상이 처음 지 구상에 나타난다.
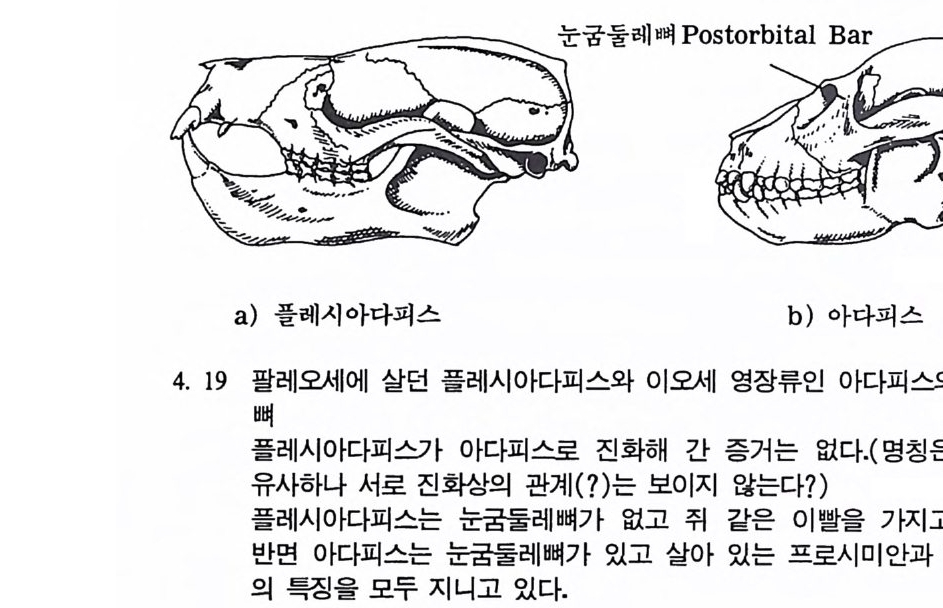 a) 플레시아다피스 b) 아다피스
a) 플레시아다피스 b) 아다피스
이오세 중에 나타난 현대 영장류의 조상은 아다피대 Ada pi dae 와 오 모마이대 Omom yi dae 의 두 무리로 나누어지는데, 이들은 특별한 동물 로부터 생겨난 것이 아니라 플레시아다피형의 동물로부터 생겨났다 . 아다피대와 오모마이대 화석은 작은 앞니를 가지고 있으며, 이빨 틈 새가 줄어들었고, 짧아전 꼬리와 주둥이를 지니고 있다. 엄지손가락과 엄지발가락은 밖을 향해 있으며, 눈 주위에 둥근뼈가 형성되어 있다.
이들 화석에서 보이는 특칭둘이 바로 카트밀이 그의 〈 사냥을 위한 시 각발달 가설 〉 에서 거론한 영장류들이 지니고 있는 특칭이다. 이오세 중에 유럽에 널리 퍼져 살았던 영장류 중 가장 큰 몸집을 가 전 아다피대는 오늘날의 여우원숭이의 조상으로 알려져 있다. 1821 년 퀴비에가 처음으로 이 종류의 화석을 발견하여 아다푸스 Adap us 라고 이름 지었다. 한편 신대륙에는 오모마이드에 속하는 영장류가 널리 퍼져 나갔는데, 그 중 어떤 종류는 아주 큰 몸집을 가졌으며 과일과 나뭇잎을 먹이로 하였다. 이오세 중에 풀레시아다피형의 젖먹이동물은 모두 멸종되었고 이 자리를 아다푸스와 오모마이드가 차지하였다. 오늘날의 여우원숭이는 바로 아다푸스로부터 진화하였고, 안경원숭이는 오모마이드의 한 종 류인 데토니우스 Tu t on i us 로부터 전화하였다. 한편 이 시기에 최초의 앤스로포이드인 폰돈지아 Pondo ngi a 가 아시아의 버마에서 발견되었다. 이 시기에 앤스로포이드가 나타날 수 있었던 것은 당시 지구상에서 번성했던 프로시미안들의 대개가 야행성이었기 때문이었다. 지구의 기온은 이오세 말에 오면 급강하하였으며, 이에 따라 열대 지역이 적도 주위로 줄어들게 되었다. 식생대의 변화에 따라 프로시 미안들의 다양성이 줄어들게 되었으나 아주 멸종은 하지 않고 그 일 부가 오늘날까지 살아 있다. 특히 지구 북반부에서 프로시미안들은 기후의 변화와 설치류의 번성으로 인해 더 이상 북반부 지역에서 살 아남을수가 없었다. 앤스로포이드 영장류 앤스로포이드 영장류에 속하는 원숭이는 지구상에 현재 133 종이 살 고 있다. 이들은 세비드과, 칼리스리시드과 그리고 짧은꼬리원숭이과 원숭이들로 구분된다. 〈고등영장류〉라고도 불리는 앤스로포이드 원숭
이는 프로시미안과는 다론 신체상의 특징을 나타낸다. 앤스로포이드 원숭이는 커다란 골을 가지고 있는데, 골 중에 냄새 를 맡는 부위가 줄어들었으며, 이들은 물체룰 색깔을 지닌 채로 볼 수 있고, 물체를 입체구조로 볼 수 있다• 주둥이는 짧게 줄어들어 냄 새를 맡는 능력이 퇴화했음을 알 수 있다. 앤스로포이드는 프로시미안과는 다르게 주행성 동물로, 이들은 프 로시미안보다 더 넓게 퍼져 살았다. 앤스로포이드는 프로시미안보다 더 발달된 손과 시각을 지니고 있고, 골 사이의 협력기능은 보다 더 발달했으며, 프로시미안보다 더 건 성장기간을 거치게 된다. 뼈대로 볼 때 앤소로포이드는 눈굼둘레뼈가 완전히 붙어 있어, 그 결과 프로시미안과는 다르게 눈알과 턱을 움직이는 힘살둘이 서로 분 리되어 있다. 앤스로포이드의 아래턱뼈는 좌우가 함께 붙어 있는데, 이렇게 아래턱의 좌우 두 부분이 붙는 면을 아래턱 이음새 Mand ibu -lar S ymp h y s i s 라고 부른다. 프로시미안의 경우 아래턱뼈들은 서로 붙 지 않으며 좌우로 나뉘어져 있다. 팔뼈와 다리뼈들은 프로시미안에 비해 거의 비슷한 크기를 하고 있다. 앤스로포이드의 분류와 해부학상 특징 앤스로포이드 영장류는 해부학상의 특칭에 따라 밀아목 In fr aorder 단계에서 납작코원숭이 Platyr r hini와 좁은코원숭이 Ca t arr hini로 구 분된다(표 4. 4). 이러한 구분은 해부학상 특칭에 의한 것이며, 전화 과정에서 서로 다른 지역에서 다른 방향으로 발전해 왔다. 따라서 이 들은 지역에 따라 납작코원숭이는 산대륙 원숭이 New World Mon- ke y s 로, 좁은코원숭이는 구대륙 원숭이 Old World Monke y s 로 나누어 전다. 여기서 신대륙이란 중앙아메리카와 남아메리카를 의미하고, 구 대륙이란 아프리카와 유라시아 대륙을 칭한다. 신대륙 원숭이 인 납작코원숭이는
대륙 원숭이와는 구분되는 납작한 모양의 코를 갖고 있다. 좁은코원 숭이는
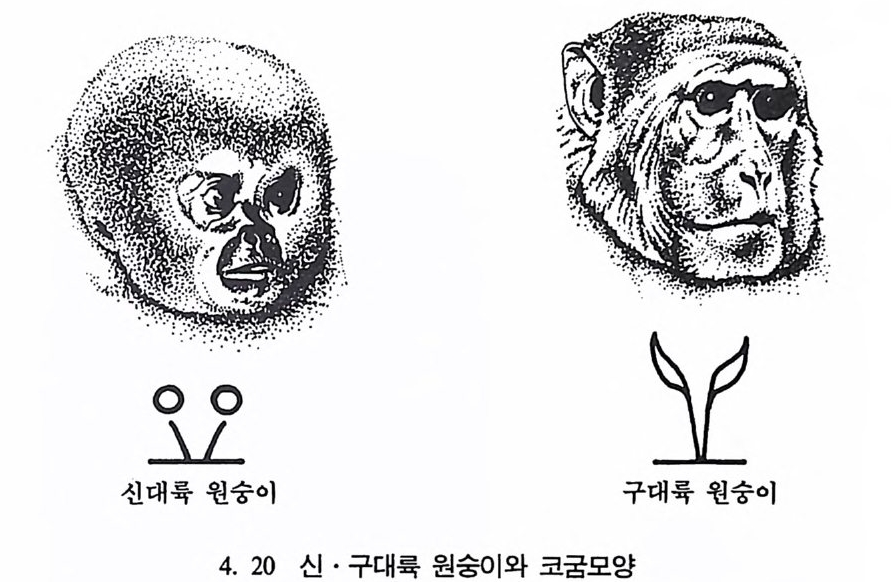 這龜.,` ..'.;.”.., .: 초, . ;編·C’ ., . .','.0 ., .'- : :'·=; :' ,,··' 磁 . ,.· .._' , 조. .학 . '•,h :`
這龜.,` ..'.;.”.., .: 초, . ;編·C’ ., . .','.0 ., .'- : :'·=; :' ,,··' 磁 . ,.· .._' , 조. .학 . '•,h :`
표 4. 4 앤스로포이드 영장류의 분류 아목 앤스로포이드아목 An t hro p o i dae Suborder 밑아목 납작코원숭 01 좁은코원숭이 Inf rao rder Platy rrh i ni Carta r rhi ni | 초과 세보이드초과 짧은꼬리원숭이초과 호미노이드초과 Sup e rfa mily Ceboid e a Cercop ithe coid e ~\ Hom ino id e a 과 세비드과 칼리스리시드과 짧은꼬리원숭이과 Fa mily Cebid a e Call ithr ic i d a e Cerc9p ithe cid a e 아과 콜로바인원숭이 짧은꼬리원숭이 Subfa mily Colobin e Cercop ithe cin e
구대륙 원숭이는 이빨의 구성이 앞니 2, 송곳니 l, 옆니 2, 어금니 3(2: l : 2: 3) 으로 되어 있는데, 이런 구성은 사람의 이빨 구성과 같 다. 이에 반해 신대륙 원숭이는 앞니 2, 송곳니 l, 옆니 3, 어금니 3 (2 : l : 3 : 3) 을 가지고 있다. 구대륙 원숭이와 신대륙 원숭이를 구분하는 또 다른 특징으로 귀에 있는 고막의 위치를 들 수 있다. 구대륙 원숭이는 고막이 머리뼈 안 쪽,죽귀 안에 있는데 반해 신대륙원숭이는머리뼈 표면,죽귀 표 면에 있다(그림 4. 21 ).
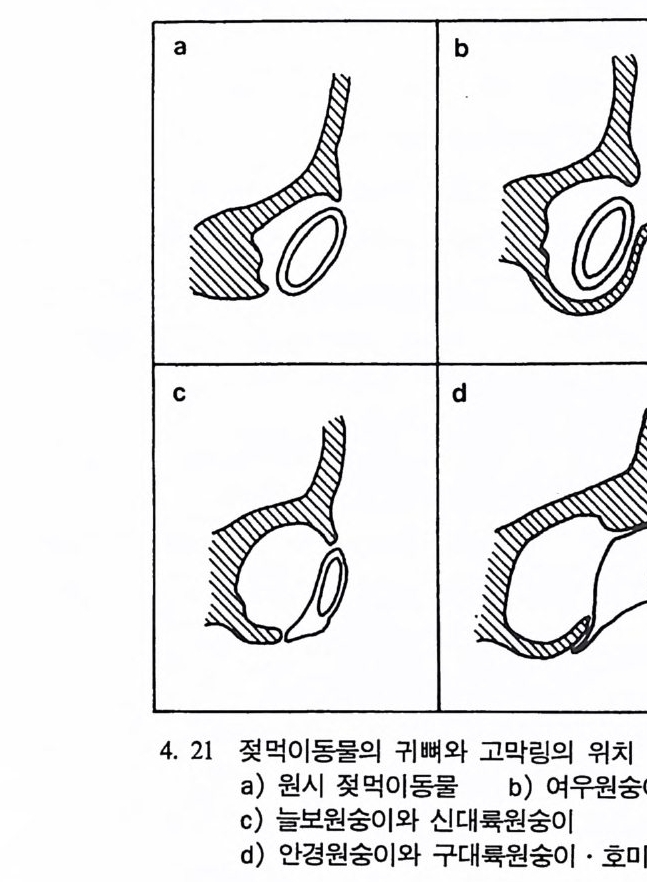 a b
a b
신대륙 원숭이(납작코원숭 01) 신대륙 원숭이인 납작코원숭이는 1 초과(세보이드초과), 2 과(세비드 과와 칼리스리시드과)로 구성되어 있다. 이들은 모두 나무 위에서 살 며, 깡충깡충 뛰거나 팔을 이용해서 이동하거나 네 발로 나무 위를 돌아다니는 걷기 방법을 지니고 있다•
 r.. .Jr .·- <·f. r -'. ,. ... .’ . . 수: - .;
r.. .Jr .·- <·f. r -'. ,. ... .’ . . 수: - .;
신대륙 원숭이는 구대륙 원숭이에서 더 이상 볼 수 없는 앤스로포 이드 전화상의 일반 특칭을 지니고 있으며, 구대륙 원숭이의 조상이 되는 화석 원숭이들과 가장 가까운, 살아 있는 화석이라는 점에서 중 요하다. 나아가 이들은 팔레오세와 이오세 중에 시작된 프로시미안 영 장류가 어떻게 환경에 적응하여 번성했는가를 살피는 데 중요한 실마 리를제공한다. 세비드과 원숭이에는 11 속, 30 종의 원숭이가 있다. 이들은 중앙아메 리카의 멕시코에서부터 남아메리카 대륙의 열대와 아열대 상록수의
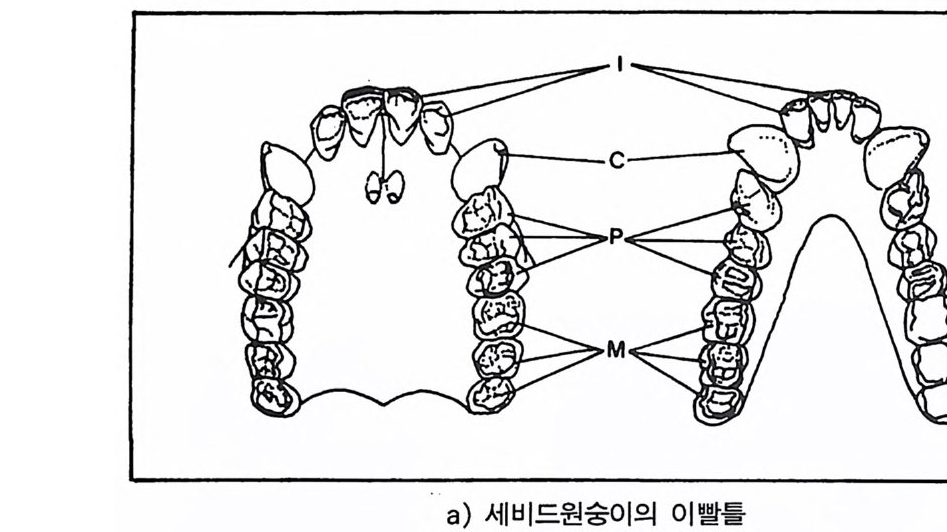 a) 세비드원승이의 이빨틀
a) 세비드원승이의 이빨틀
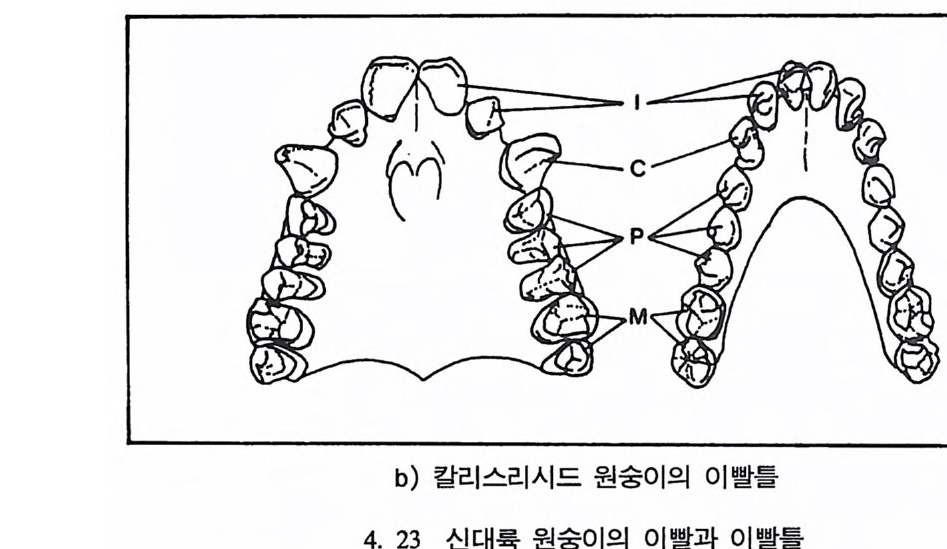 b) 칼리스리시드 원숭이의 이빨틀
b) 칼리스리시드 원숭이의 이빨틀
삼림지대에서 살고 있다. 세비드과 원숭이들은 대개가 나뭇키지를 감 아쥘 수 있는 꼬리를 가지고 있다(사전 4. 22a ; 사전 4. 22b). 몸집은 큰 원숭이의 경우 9kg 정도까지 나가며 세 개의 어금니와 둥근 손톱을 가지고 있다. 이들은 나무 위에서 살며 대개가 낮에 활
동한다. 그러나 다람쥐원숭이 Sq u ir r el monkey , 거미원숭이 Sp ide r monkey, 짖는원숭이 Howler monkey 그리고 부영원숭이 Owl monkey 등은 밤에 주로 활동을 한다. 커다란 눈과 기타 신체상의 특칭둘은 프로시미안 영장류에서 찾을 수 있는 특칭을 지니고 있다(그림 4. 23 a). 칼리스리시드과 원숭이는 좀더 특수화된 모습을 지니고 있다(그림 4. 23b). 이들은 5 속 21 종으로 이루어져 있으며, 중앙아메리카 남부지 역과 남아메리카 대륙의 북부지역에 퍼져 살고 있다. 열대 우림지대 와 회랑을 이루는 삼림 및 사바나 경계지대에 걸쳐 산다. 몸무게는 450 그램이 채 되지 않으며 뾰족한 손톱을 갖고 있다. 과일과 곤충 그리고 껌울 먹는다. 신대륙 원숭이 중 많은 원숭이들은 두 명의 새끼를 갖고 있는데, 주로 수놈이 새끼를 돌본다•
 4. 24 아프리카 파타스 원승이
4. 24 아프리카 파타스 원승이
구대륙 원숭이(좀은코원숭이) 구대륙 원숭이는 짧은꼬리원숭이초과에 짧은꼬리원숭이과가 있으
며, 이는 다시 콜로바인 원숭이와 짧은꼬리원숭이과로 나누어전다. 모 두 82 종의 구대륙 원숭이가 알려져 있다. 구대륙 원숭이는 신대륙 원숭이에 비해 매우 번성한 무리로, 이돌 의 콧구멍은 사이가 넓으며, 반달 모양의 콧구멍을 가지고 있다. 이빨 은 앞니 2, 송곳니 I, 옆니 2 그리고 어금니 3(2: I : 2: 3) 이 있다. 고막은 귓구멍 속 끝에 있으며, 어금니는 바일로포돈트 B i lo p hodon t 도드리를 지니고 있다(사전 4. 24 ; 그림 4. 25).
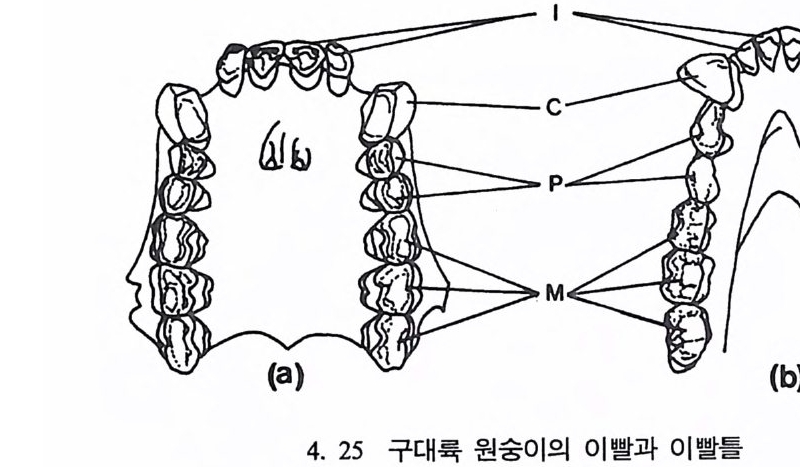 c
c
구대륙 원숭이는 한 쌍의 붉은색을 띤 굳은 엉덩이판이 발달해 있 다. 구대륙 원숭이는 신대륙 원숭이와 달리 땅 위에서 산다. 콜로바인 원숭이는 잎사귀를 주식으로 하기 때문에 해부학상으로는 짧은꼬리원숭이와 다른 특칭을 지니고 있다. 이들은 잎사귀를 갈아서 먹기 때문에 이런 운동을 위해 큰 턱과 찰 발달된 턱근육을 지니고 있다. 어금니는 높고 날카로운 날을 가지고 있는데, 이런 형태의 이빨 날은 잎사귀를 자르는 데 알맞다. 볼에는 주머니가 없으며 뒷다리는 길지 않다. 콜로바인 원숭이들의 내장은 소낭이라는 많은 주머니나 방들로 나
누어져 있는데, 여기에 셀룰로오스녜卜 분해시키는 박태리아가 살고 있 다. 이들은 주로 아시아의 삼림지대에서 서식하며 아프리카에는 1 속 이 살고 있다.
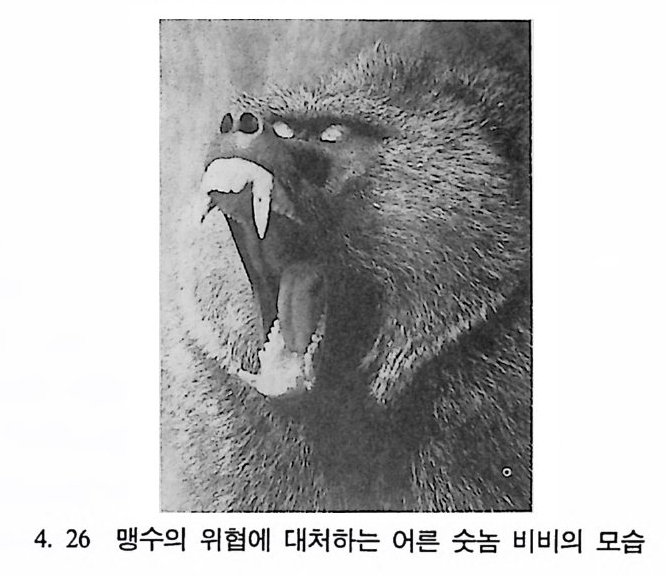 1.
1.
짧은꼬리원숭이는 8 속 45 종으로 구성되어 있다. 이들의 턱은 크지 않고 에나멜 부분이 낮은 이빨을 가지고 있으며 소화기관은 복잡하지 않고 단순하다. 볼에 주머니 같은 부분이 있어 음식을 저장할 수 있 다. 이들은 주로 과일을 주식으로 하며 몸무게는 l.5-4.5k g의 작은 종류로부터 31-36k g에 달하는 큰 종류까지 다양하다. 이외 에 건코원숭이 Proboscis monke y와 랑그르 원숭이 Lang ur mon-key 등의 구대륙 원숭이는 아시아 대륙에서 살고 있다. 비비 원숭이 Baboon 는 많은 종류가 있는데 이들은 영장류 중에서 가장 사회형인 동물로 30-150 마리가 넓은 지역에서 함께 생활한다. 수놈은 아버지로서 중요한 사회적 구실을 하며 새끼들은 오랫동안 보 살핌울 받는다(사전 4. 26 ; 그림 4. 27).
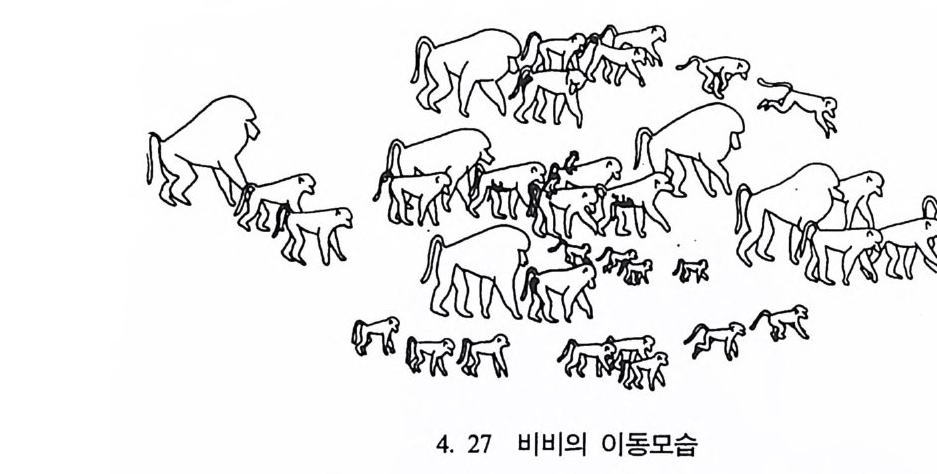 접
접
앤스로포이드의 발견과 지구의 변화 앤스로포이드의 기원과 발달과정을 설명해 줄 수 있는 필요한 화석 의 발견이 별로 없기 때문에 이들이 언제, 어느 집단으로부터 시작되 었는지는 정확하게 알 수 없다. 그러나 일반적으로 앤스로포이드는 이오세 말에 처음으로 나타났다고 추정되며, 지금까지 우리가 알고 있는 최초의 앤스로포이드의 모습은 단지 턱뼈와 이빨둘의 조각을 이 용해서 복원된 것이다. 이오세 말이 되면서 지구의 기온은 점차 떨어져서 지구의 식생대에 변화를 가져왔다. 열대 삼림지대는 적도 부근으로 축소되었고, 이에 따라 영장류들의 삶도 적도 부근으로 줄어들게 되었으며, 그 결과 영 장류의 화석은 적도 부근에서만 발견되고 있다. 이오세 시기의 남아 메리카 대륙과 오스트레일리아는 섬이었으며 유럽과 북아메리카 대륙 은 이미 분리되어 있었다. 듀크 대학의 고인류학자인 사이몬스는 이집트의 기자 남쪽에 있는 피라미드 근처의 엘 파융 저지대에서 울리고세 당시의 유적을 발굴하 였다. 사이몬스와 일행은 화석을 채집하기 위해 사막을 뒤졌고, 그리 고 다음해에 다시 돌아왔을 때 바람에 의해 노출된 유적을 찾게 되었
다. 이 유적과 여기서 발견된 영장류의 화석들을 통해 영장류 연구에 새로운 지평이 열리게 되었다. 울리고세 중에 엘 파융 지역은 열대 우기 삼림지대로, 근처에 늪과 같은 강과 호수가 있었으며 많은 종류의 영장류가 이 일대에서 살고 있었다. 어떤 종류는 안경원숭이 같고 어떤 것들은 앤스로포이드의 화석이었다. 이들 화석둘은 눈굼에 벽둘이 있으며 아래턱은 맞붙어 있었다. 여기서 발견된 화석 중의 하나인 아피디움속g enus Ap idi u m 의 영장류는 주행성으로 과일을 먹고 깡충깡충 뛰어다니는 종류였다. 이집토피데쿠스Agypt op it hecus 라고 불리는 또 다른 화석 영장류는 약 6- 7 kg 정도의 몸무게를 가지고 있는데, 오늘날 남아메리카 대륙 에서 살고 있는 짖는원숭이와 비슷한 걷기 형태를 지니고 있다. 그러 나 이 화석 영장류는 구대륙 원숭이처럼 단지 27 粹의 옆니를 지니고 있다. 한편 파융에서 발견된 다른 영장류는 남아메리카 대륙에서 살고 있 는 원숭이들의 이빨구조처럼 3 개의 옆니를 갖고 있다. 남아메리카 대륙에서 발견되는 화석들은 우리에게 매우 홍미룰 불 러일으킨다. 올리고세에 남아메리카 대륙은 아프리카 대륙 및 북아메 리카 대륙과는 분리되어 섬으로 되어 있었다. 당시에 여기에는 아주 커다란 몸집을 가전 나무늘보 G i an t Slo t h 와 칼날주머니고양이 Saber too th e d Masup ial Cat 등이 살고 있었다. 남아메리카 대륙은 빙하기인 풀라이스토세 말에 와서야 비로소 북 아메리카 대륙과 연결되었다. 남아메리카 대륙에서 발견된 팔레오세 화석들은 아주 좋은 상태로 찾아지지만 영장류의 화석은 하나도 발견 되지 않는다. 최초의 영장류인 데토니우스 화석은 2500-3000 만 년 전의 지층에서만 발견된다. 남아메리카 대륙에서 발견되는 영장류 화석의 주인공들은 어디서 왔는가 하는 문제는 학자들 사이에 많은 논쟁을 불러일으켰다. 남아
메리카 대륙에서 발견되는 영장류의 기원은 오늘날 크게 두 가지 가 설로 대표된다.
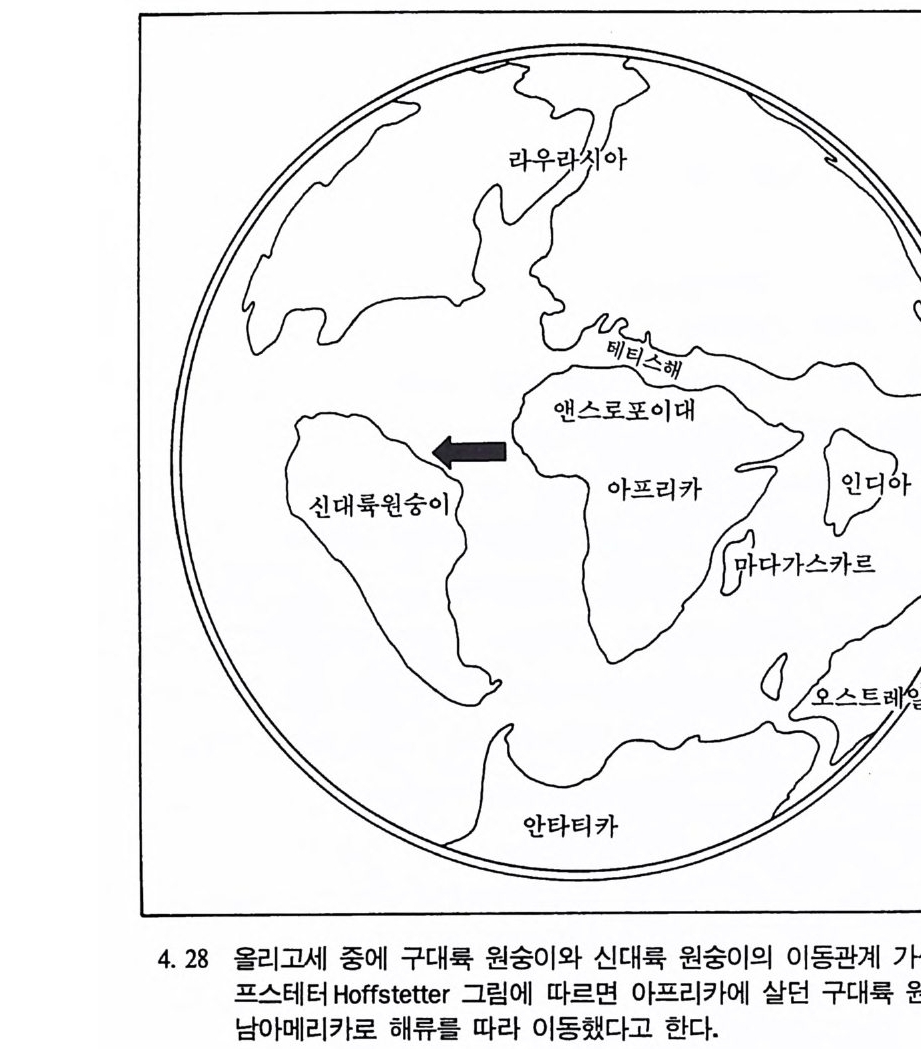 t프리카 / :>t
t프리카 / :>t
첫째 가설은 북아메리카 대륙 기원설로, 이 설에 따르면 영장류는 북아메리카 대륙에서 시작하여 남아메리카 대륙를 연결하는 회랑을
따라서 이동하거나, 대륙 사이에 점점이 이어져 있는 섭둘을 거쳐서 St ep ping St on es 이동하였다는 설이고, 두번째 가설은 아프리카 대륙 기원설로, 울리고세 중에 아프리카 대 륙에서 살던 영장류가 그들이 살던 삼림에서 떨어져 나와 바다에 떨 어전 나무 등을 타고서 해류에 의해 남아메리카 대륙에 도달하여 퍼 져 나갔으리라는 설이다(그림 4. 28). 호미노이드 영장류 호미노이드 영장류는 유인원으로 알려진 긴팔원숭이과와 성성원숭 이과 그리고 인류과인 호미니드과 Ho mi n i dae 로 구성되는데(표 4. 5), 여기서는 인류과를 제의한 유인원들에 관해 살펴보기로 한다. 유인원인 긴팔원숭이과와 성성원숭이과는 각각 작은 유인원과 큰 유인원으로도 불린다. 유인원은 모두 4속 13 종이 있는데, 그 중 9 종이 작은 유인원으로 긴팔원숭이와 큰긴팔원숭이가 여기에 속한다. 유인원 중 4 종이 큰 유인원으로, 보통 크기의 몸집을 한 침팬지인 팬 트로글로다이트 Pan t rog lody t es 와 피그미 침팬지인 팬 팬니스쿠스 Pan pa nis c us, 오랑우탄인 팬 피그미우스 Pan pygmaeu s, 그리고 고릴 라인 고릴라 고릴라 Gor i lla g or i lla 가 여기에 속한다 유인원은 생화학상 및 해부학상으로 사람과 가장 가깝게 닮았으며, 실제로 이들이 가장 두드러지게 영장류의 특칭을 가지고 있다. 비록 영장류의 진화과정 중에 오늘날 살고 있는 유인원이 가장 발달한 형태 인 것처럼 보이나, 주의할 것은 오늘날 살아 있는 형태들은 전화상의 산물이라는 사실을 알아야 한다. 오늘날 살고 있거나 또는 과거에 살았던 유인원을 막론하고 이들에 게서 가장 두드러지는 해부학상의 특칭으로 걷기 방법을 들 수 있는 데, 팔이나 몸을 혼들어 이동하는 방법 (Brac hi a ti on 또는 Sw ing ing ) 이 바로 대표적인 걷기 방법이다.
작은 유인원인 긴팔원숭이과 : 긴팔원숭이와 큰긴팔원숭이 긴팔원숭이속에는 9 종의 작은 유인원들이 있는데 이들은 인도 동 부지역에서 방글라데시,버마, 인도차이나,말레이시아,수마트라,서 부 자바, 보르네오를 거쳐 남중국에 이르는 지역에 분포하고 있다.
표 4. 5 호미노이드 영장류의 분류 아목 Suborder 앤스로포이드아목 Anth rop o id e a 초과 Sup e rfa milly 호미노이드초과 Homi no id e a 과 Fa milly 긴팔원숭이 과 Hy lo bati da e , 성 성 원숭이 과 Pong ida e, 호미니드과 Hom inida e 속 Genera 긴팔원숭이 Hy lo bate s , 성성원숭이 Pong o (l ), 인류 Homo( l) 종 Sp ec ie s 심 판란지 Sy mph alang es , 침팬지 Pan(3) 고릴라 Gorill a ( 1 )
긴팔원숭이와 큰긴팔원숭이는 가장 대표되는 작은 유인원으로 열대 우기 상록활엽수 삼림과 준열대 침엽수 삼림에서 산다. 이들의 얼굴 모양은 매우 다양하며, 두 팔로 몸을- 흔들어 이동한다. 긴팔원숭이와 큰긴팔원숭이는 lm 정도의 키에 매우 가늘고 작은 몸매를 지니고 있 으나, 몸에 비해서 긴 팔과 긴 손목 그리고 작은 엄지손가락을 갖고 있다. 이들에게서 성에 따른 크기의 차이는 볼 수 없으며 모든 유인원 들처럼 꼬리가없다. 긴팔원숭이는 두 팔만을 사용하여 몸을 이동하는 영장류로 대표되
며 땅 위에서는 거의 볼 수 없다. 이들은 대개가 과일을 주식으로 하 지만 때때로 작은 새들과 곤충들 을 잡아먹는다. 가족은 수놈아비 한 마리와 암놈어미 한 마리, 그리고 새끼들로 구 성된다. 새끼는 때] 정도가 될 때까지 함께 지내다가, ~l 정도가 되 면 아비에 의해 살던 지역에서 내쫓긴다. 기본은 자기들의 관할지역 을 철저히 지키는데, 약 40-300 에이커 정도의 지역을 다른 기본의 가족으로부터 지킨다. 서로 이웃하는 관할구역 사이에 사람이 사는 지역은 없다. 지역을 방어하는 방법으로는 대개 소리를 지르나 때로 는 몸싸움을 하는 경우도 있다. 암놈과 수놈이 번갈아가면서 소리를 질러서 대항울 하는데 이를 듀엣 Due t s 이라 부른다. 성에 따른 차이 가 없는 점은 아마도 이렇게 가족을 지키기 위해서 암놈이 수놈과 같 이 행동하는 데서부터 기인한다고 여겨진다. 큰긴팔원숭이는 긴팔원숭이보다 약간 큰 몸집에 검은색 털을 가지 고 있으며, 혀에는 공기주머니가 있는데 이는 특별한 소리를 내는 데 사용된다. 시아망은 절대로 네 발로 걷지 않으며 땅 위에서 두 손바 닥을 벌리고는 두 발로 걷는다. 큰 유인원인 성성원숭이과 : 오랑우탄 고릴라 및 짐팬지 오랑우탄 Pan pyg maeus 은 아시아에 살고 있는 큰 유인원으로 성인 남자만큼이나 큰 몸집을 가지고 있다(사전 4. 29). 보르네오와 수마트 라에 걸쳐 숲속에서 살고 있는데, 현재 멸종의 위기에 처해 있다. 수 놈들은 시간의 대다수를 땅 위에서 보내며 몸무게는 135k gi끝 넘는다. 빙하기에는 지금 살고 있는 지역보다 더 널리 퍼져, 중국 지역에서도 살았다. 수놈 오랑우탄은 항상 혼자 있으며 새끼들과 같이 살고 있는 여러 마리의 암놈어미들의 영역을 어슬렁거리며 교미할 암놈과 음식을 구 하러 돌아다닌다. 짝을 지어 산다는 것은 아주 드물며, 서로 소리를
통해서 또는 냄새로써 자기의 영역을 지킨다. 오랑우탄은 아주 건 팔을 가지고 있는데 두 팔을 쳐들면 키가 약 2.5m 미터 정도가 되며 상당히 휘어진 손가락과 발가락을 가지고 있다. 수놈은 암놈에 비해 아주 큰 몸집을 가지고 있고, 나무를 천천히 기어 오르고 과일과 열매 그리고 잎사귀를 주식으로 한다.
 4. 29 늙은 수놈 오랑우탄
4. 29 늙은 수놈 오랑우탄
 4. 30 산악고릴라
4. 30 산악고릴라
고릴라 Gor i lla g or i lla 는 1800 년대 중반까지는 서유럽 세계에 알려져 있지 않았다. 이들은 아프리카 콩고의 우기 삼림지대와 리프트 골짝 서쪽인 중앙아프리카의 산악 기슭에서 산다(사진 4. 30). 고릴라는 땅 위에서 살며 영장류 중에서 가장 큰 몸집을 가지고 있는데, 성인 수 놈은 몸무게가 230kg 정도나 된다. 암놈은 수놈의 약 철반 가량의 몸 무게밖에 안 되는데 암수 사이의 몸무게가 이 정도로 크게 차이가 나 며, 두 발로 서 있는 시간도 많다. 고릴라는 건 팔을 갖고 있으며 손마 디 바깥면으로 땅을 짚고 걷는다. 늙은 수놈은 등의 털이 갈색과 흰색 으로 〈은빛등 S il ver Backs 고릴라〉라고도 불린다.
고릴라는 음식으로 야채만을 먹으며, 특히 〈대나무잎〉를 좋아한다. 가족은 환경에 따라 그 크기가 달라지기는 하지만, 가장 대표되는 구 성은 한 마리 또는 그 이상의 은빛등을 가전 수놈과 이보다는 젊은, 검은색의 등을 가진 2-3 마 리의 수놈, 최고 6 마리 정도의 암놈들과 새끼둘로 이루어져 있다. 고릴라는 전혀 자기 영역을 고집하지 않아 다른 그룹과 다두지 않는다. 고릴라는 밀림 속에서 살기 때문에 이들의 행위를 연구하기가 곤란 하다. 따라서 처음 오랫동안 고릴러는 난폭한 맹수로서 인식되었으나, 최근의 연구들에 따르면 이들은 아주 얌전한 동물임이 밝혀졌다. 침팬지는 팬속g enus Pan 으로 분류되며, 보통 침팬지 Pan trol og dy t - es 와 피그미침팬지 Pan pygm i es 두 종이 있다. 이들은 숲속이나 숲 가 장자리에서 살며 몸의 크기는 긴팔원숭이와 고릴라 몸집의 중간에 속 한다(사전 4. 31 ).
 4. 31 침팬지
4. 31 침팬지
 4. 32 제인 구달과 침팬지
4. 32 제인 구달과 침팬지
침팬지의 경우 암놈은 수놈보다 몸의 크기가 10% 정도가 작아 암 수 차이는 그리 큰 편이 아니다. 침팬지는 땅 위를 다닐 때는 손마디 바깥면으로 땅을 짚고 걸으며 숲속의 나무에서는 팔을 이용해서 움직 인다. 때로는 두 방법을 모두 쓴다. 침팬지는 대개가 과일울 주식으로 하나 때로는 작은 동물과 원숭이 그리고 작은 비비롤 잡아먹기도 한 다. 침팬지의 행위 연구로 이름난 제인 구달J ane Goodall 은 이들의 사 회행위를 20 년 이상 연구하였는데(사전 4. 32), 침팬지는 40 세 정도까 지 살기 때문에 구달처럼 오랫동안 연구를 하는 것이 매우 중요하다. 처음에 침팬지는 전원을 좋아하고 평화스러운 동물로서 인식되었으 나, 오늘날 이는 사실이 아님이 밝혀졌다• 침팬지는 50 - 100 마리가 8- 12km2 정도의 지역에서 모여 살며, 만일 다른 침팬지가 그들의 영 역에 들어오면 이를 잡아서 때리고 물고 때로는 죽여버리기조차 한 다. 영장류의 차이점 유인원과 원숭이와의 해부학상 차이점 유인원과 원숭이는 앤스로포이드아목에 속해 있으나 해부학상으로 서로를 구분할 수 있는 다음과 갇은 특칭둘을 가지고 있다(그립 4. 33 ; 그립 4. 34). 1. 원숭이와 유인원을 가장 쉽게 구분하는 방법은 몸체에 꼬리를 갖고 있는지에 있다. 대다수의 원숭이는 꼬리를 갖고 있는 반면에 유인원은꼬리가 없다. 2. 몸의 크기에 따른 비율을 가지고 살펴볼 때 원숭이는 유인원에 비해 몸통이 길며, 위에서 볼 때 유인원의 몸통은 사람의 몸통처럼
앞뒤보다는 양옆의 길이가 긴, 둥글납작한 모습을 지니고 있는 데 비해 원숭이의 몸통은 앞뒤 • 양옆의 길이가 비슷한 둥근형이다. 3. 원숭이의 주걱뼈는 둥근 몸통 양옆에 위치하고 있으나 유인원의 주걱뼈는 사람처럼 몸통 뒤쪽 양옆에 위치하고 있다. 4. 원숭이는 앞팔과 뒷다리의 길이가 같은 데 비해서 유인원은 앞 팔이 뒷다리에 비해 더 길다.
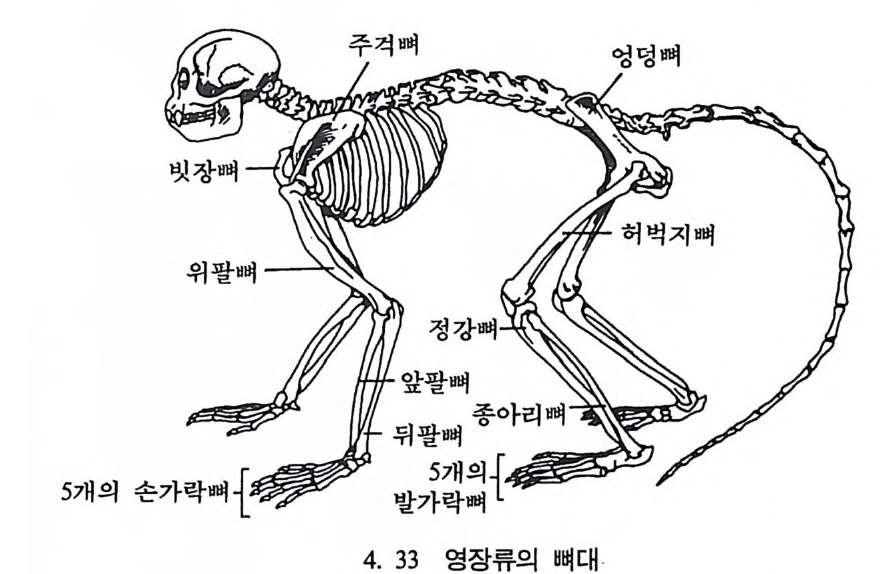 己뼈
己뼈
5. 빗장뼈의 경우, 유인원의 빗장뼈가 원숭이의 빗장뼈보다 길며, 주걱뼈를 뒤로 미는 구실을 한다. 6. 유인원의 엉덩뼈는 짧고 넓적한 편 Bo yv l-Sha pe인 데 비해 원숭 이의 골반은 길고 좁은 모양을 하고 있다. 7. 위팔뼈의 머리부분은 유인원의 경우 원숭이보다 더 둥글며, 원숭 이의 손목은 유인원의 손목보다 자유롭지 못하다. 8. 유인원의 몸체와 원숭이의 몸체는 둘 다 곧게 서 있으나 유인원 의 몸체가 원숭이의 몸체에 비해 더 곧게 서 있다.
9. 원숭이는 어금니의 봉오리가 두 개씩 서로 이어져 있는 데 Bi- lop” h odo nt 반해 유인원은 뭉뚝한 모양의 다섯 개의 봉우-리를 갖 고 있으며 이들 봉오리 사이에는 골이 져 있다. 짧은꼬리원숭이 고릴라 사람 4. 34 원숭이 • 고릴라 • 사람 뼈대구조의 차이는 주로 허리등뼈와 엉덩뼈 • 주걱뼈 • 빗장뼈의 위치에서 찾아진다. 이처럼 유인원과 원숭이를 구분하는 특칭들은 영장류의 진화과정에 서 나타나는 독특한 특칭들 Nove liti es 로, 이런 특칭들은 인류초과인
호미노이드에게서 나타나는, 공통으로 나누어 받은 특징들 Shared De- riv e d Charac t ers 이다. 위에서 언급한 특징의 많은 부분은 사람과 유 인원의 공통조상 Common Ances t er 의 걷기 방법 Locomoti on Patt er n 과 직접적으로 관계가 있으니, 이들의 공통조상은 팔을 이용하여 몸 을 이동하던 존재였다. 사람에게는 이렇게 몸을 이동했던 습관이 아 직도남아 있다. 유인원과 사람의 해부학상 • 기능상 차이점 유인원과 사람을 비교해 보면 다음과 같은 차이점을 찾아볼 수 있 다. 머리뼈의 비교(그림 4. 35 : 그림 4. 36)
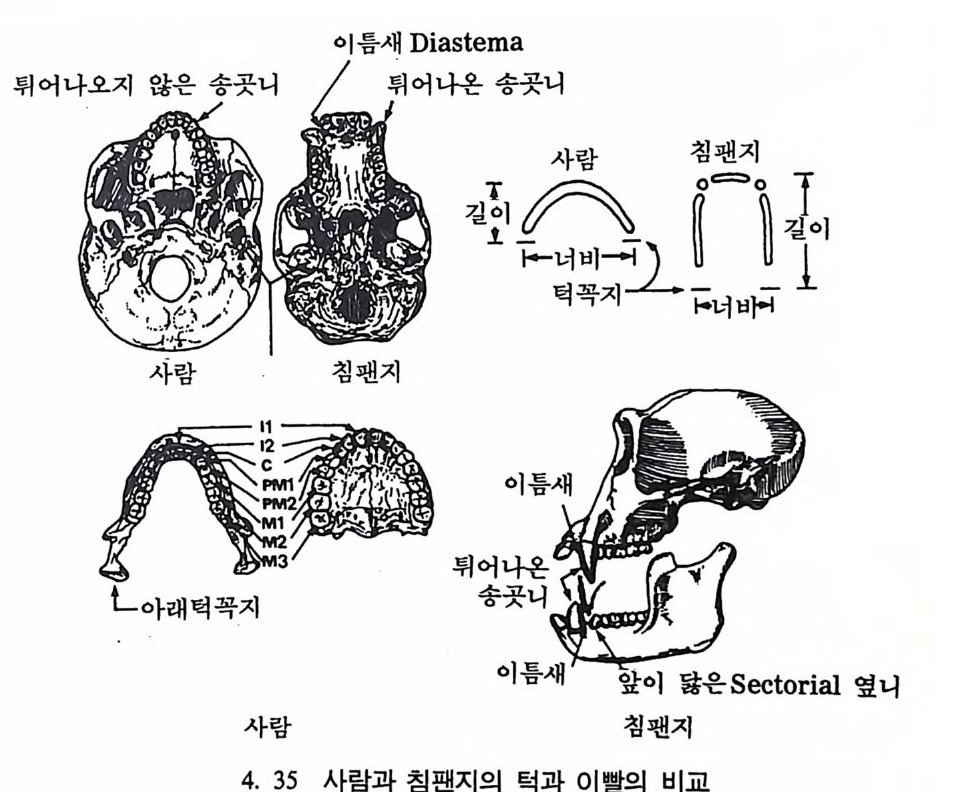 0,1 틈 새 Dias te m a
0,1 틈 새 Dias te m a
I. 유인원의 두뇌 크기는 사람 두뇌 크기의 3 분의 l 정도이다. 2. 사람의 얼굴은 편평하고, 몸의 전체 크기에서 볼 때 유인원에 비 해서는 아주 작은 얼굴을 지니고 있다.
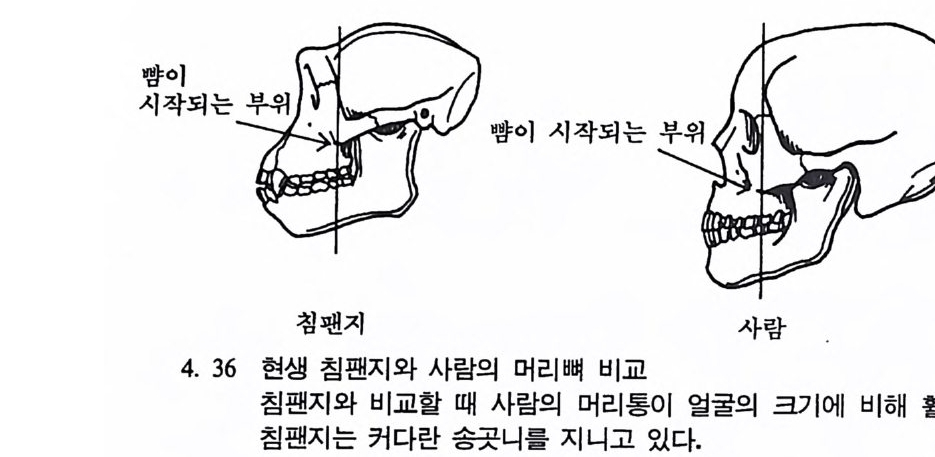 뺨이 시작되는
뺨이 시작되는
3. 사람의 잇몸과 입천장은 넓은 포물선형인 데 반해 유인원은 길 고 좁은 사각형 (U 형)으로 앞쪽 두 귀퉁이에 커다란 송곳니가 튀 어 나와 있다. 4. 중추신경굼은 유인원의 경우 머리 뒤쪽으로 나 있는 데 반해 사 람의 경우는 머리 밀에 위치하고 있다. 뼈대 각 부분의 비교 (그립 4. 37 : 4. 38 ; 4. 39 ; 4. 40) 1. 사람의 경우, 걸을 때 발뒤꿈치는 땅에서 받는 충격을 막는 구실 울 하고, 가운데가 움푹 파인 Axche~ shape d 발바닥이 충격을 줄 이는데 도움이 된다. 2. 유인원은 사람과 다르게 옆으로 비켜 나간 D i verg en t 엄지발가 락을가지고있다.
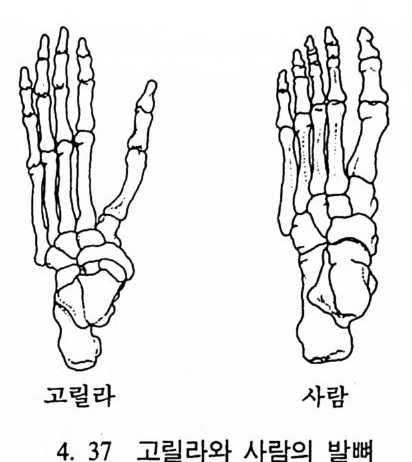 고릴라 사람
고릴라 사람
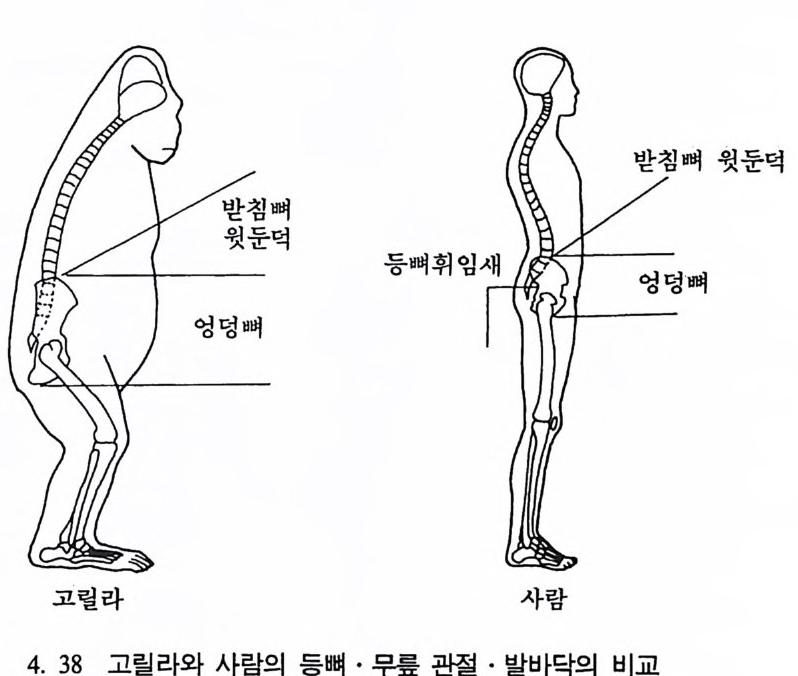 받침뼈 윗둔덕
받침뼈 윗둔덕
3. 무릎 관철의 경우, 사람과 유인원은 각기 다른 특칭을 지니고 있 다. 사람의 경우, 무릎뼈의 가장자리가 허벅지뼈 끝의 파인 부분 안에 자리하고 있고, 허벅지뼈는 몸 중앙선으로 향하여 각이 져
있다. 유인원의 경우 허벅지뼈는 납작하고 무릎뼈가 닿는 부분이 얇게 파여 있다. 다리는 밖으로 향하여 굽어 있다.
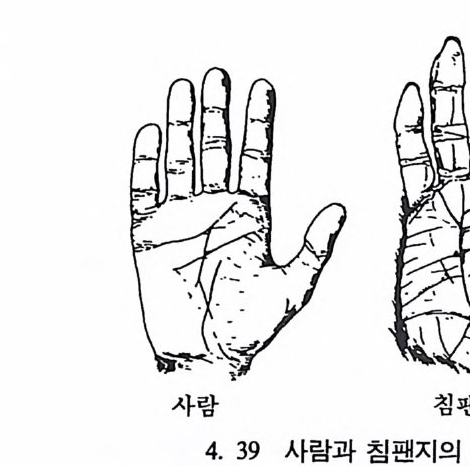 사람 `
사람 `
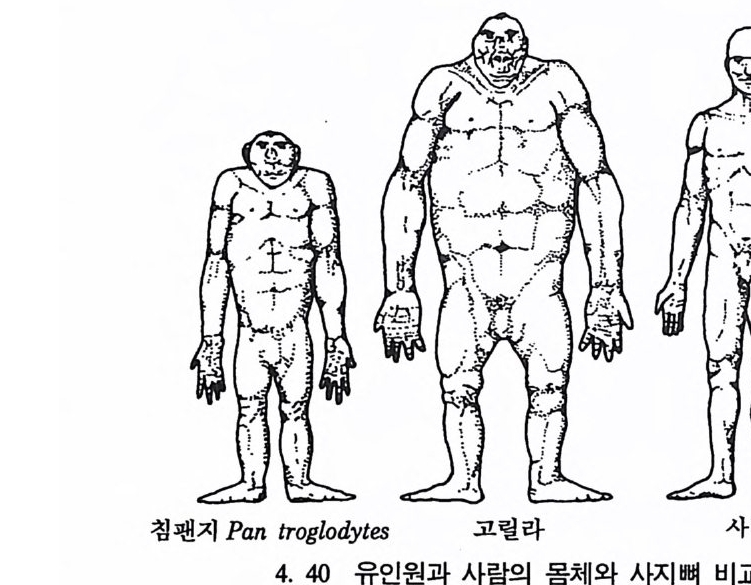 침팬지 Pa n. trog l o dy tes 고릴라 사람
침팬지 Pa n. trog l o dy tes 고릴라 사람
4. 사람의 엉덩뼈는 접시 모양으로 둥글고 짧은 데 비해 유인원은 길고 좁은 엉덩뼈를 지니고 있다. 등뼈는 사람이 S 자 모양인 데
비해 유인원은 등이 굽어져 마치 둥근 교각 같은 모양이다. 5. 사람은 긴 엄지손가락과 짧은 손가락을 가지고 있는 데 비해 유 인원은 긴 손가락에 짧은 엄지손가락을 가지고 있다.
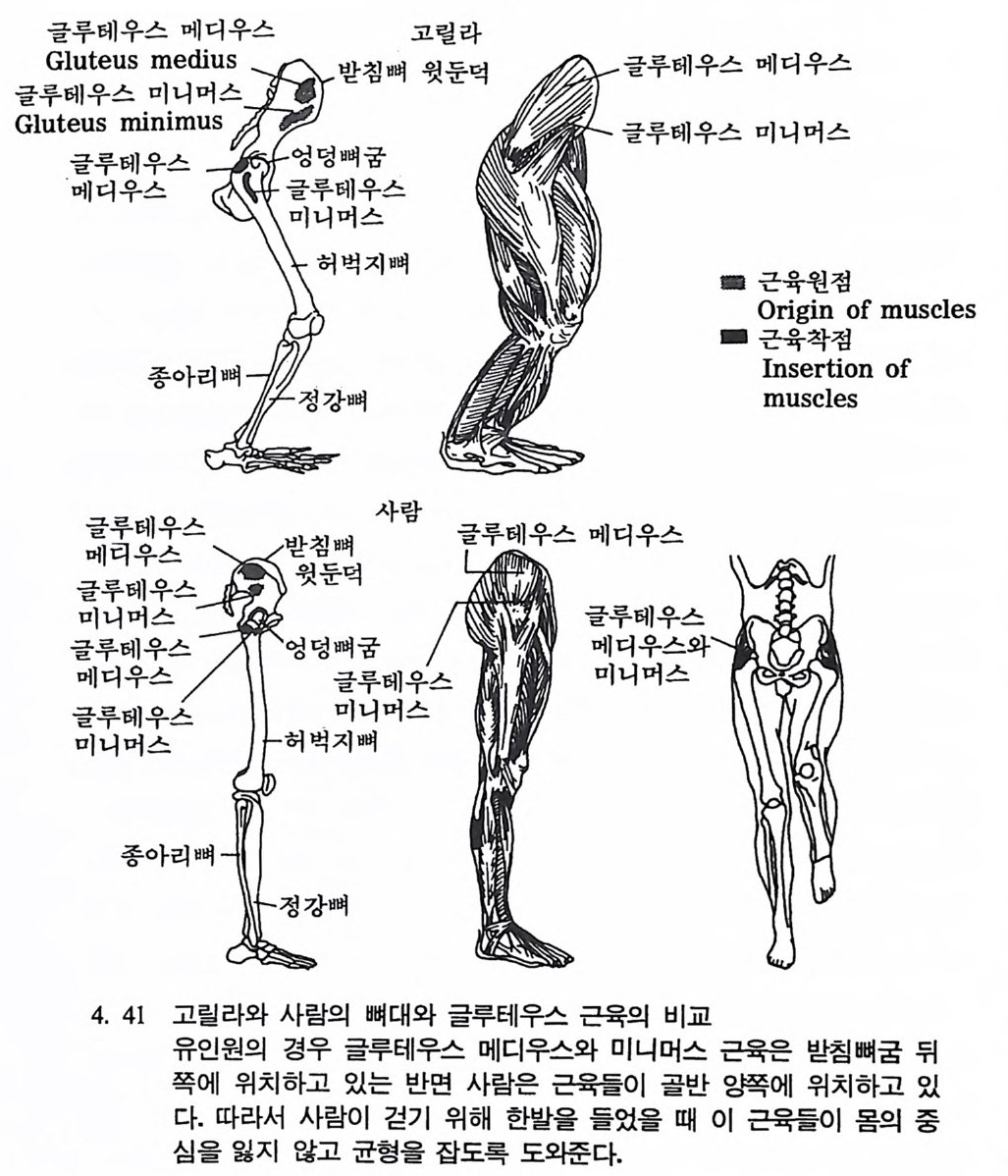 글루테우스 메디우스
글루테우스 메디우스
6. 사람의 경우 위팔뼈와 허벅지뼈의 길이가 거의 같은 데 비해 유 인원은 위팔뼈가 더 길다. 7. 유인원의 빗장뼈는 사람에 비해 더 크며 어깨쪽이 사람보다 울 라가있다. 위에서 언급한 차이점 이의에 다른 점들은 다음과 같다 ; 몸집의 크기에서 사람과 유인원을 구분할 수 있다. 성에 따른 몸집 의 차이는 유인원은 그 종류에 따라 차이가 난다. 예로 침팬지는 암 수 사이에 차이가 별로 없으나, 고릴라와 오랑우탄은 암수 사이의 차 이가 크게 나타난다. 사람은 성에 따른 남녀 차이가 . 크지 않다. 송곳니에서 볼 때 사람은 성에 따른 차이가 없으며, 가슴·과 음경은 다른 유인원과 비교해 볼 때 몸의 크기에 비해 매우 큰 편이다. 사람과 유인원의 차이는 엉덩이힘살 Glu t eus muscle 에서 뚜렷이 찾 아진다(그립 4. 41). 사람의 넓적하고 휘어져 있는 엉덩뼈에는 두 개 의 힘살 (Glu t eus me di us 와 Glute u s mini mus) 이 붙어 있는데, 이 힘 살둘은 다리를 밖에서 안으로(옆에서 옆으로) 잡아당긴다. 이를 밖돌 리기 Abduc ti on 라 한다. 이 힘살둘은 사람이 걷울 때 한 발이 땅에 닿기 전 중력의 중심을 잡아당겨 몸의 균형을 유지시키는 구실을 한 다. 유인원은 이와 같은 엉덩뼈와 힘살을 갖고 있지 않아서, 두 발로 걸을 때 온몸이 좌우로 움직인다 . 유인원에 관한 일반적인 이해 호미노이드 영장류의 수는 별로 많지 않으나 우리와 매우 가까운 관계에 있기에 중요한 연구대상의 하나가 된다. 그러나 지난 반세기 동안 유인원에 관한 우리의 지식은 부정확하였는데, 이에 관한 가장 연극상으로 나타나는 예로는 거대한 고릴라인 킹콩의 출현을 둘 수
있다. 킹콩은 지난 40 년간 유인원을 극도로 상칭화한 것이다. 그러나 재 미있는 사실은 실제로 고릴라는 킹콩을 닮지 않았다는 점이다. 오랑 우탄과 침팬지 또한 영화에서 그려지는 그런 이미지를 닮지 않았다. 우리는 지금까지 유인원에 대해 많은 연구를 진행시키고 있으나 이들 의 행위에 대해서 아직도 모르는 부분이 많다. 1 세기 전 유럽의 탐험가들은 아프리카의 숲속에 살고 있는 무서운 동물의 이야기를 가지고 유럽으로 돌아왔다. 그러나 오늘날 우리는 고릴라가 더 이상 공격을 좋아하는 동물이 아니라는 사실을 알고 있 고, 그들이 보여주는 행위는 다만 침입자가 놀라 도망가도록 하는 경 고용의 방법임을 알고 있다. 실제로 고릴라는 매우 평화로운 동물이 며 그들은 대부분의 시간을 먹고 놀고 잠자는 데 보낸다. 고릴라는 채식동물로서 커다란 몸집을 유지하기 위해서 많은 양의 잎사귀를 먹 는다. 오랑우탄에 대해서는 별로 알려져 있지 않은데, 오랑우탄이란 말은 말라야에서는 〈숲속의 사람〉이란 뜻이다. 그들은 대부분 나무에서 생 활하며, 긴 손가락둘은 오랑우탄이 나무에서 살기에 알맞은 생활습성 울 지니고 있다는 것을 보여준다. 이들은 나무에 참자리를 만들고는 대부분의 시간을 먹는 데 보낸다. 침팬지는 주어전 환경에 가장 잘 적응하고 모험을 좋아하는 유인원 이다. 이들은 아주 호기심이 많으며 매우 영리하다. 침팬지는 사람과 함께 살도록 훈련될 수 있는데, 한 예로 루시라는 이름을 가전 침팬 지는 일생을 사람과 함께 살아왔으며 200 개의 미국식 표현언어를 배 웠다. 호미노이드 영장류를 구성하는 인류와 유인원을 구분하는 가장 뚜 렷한 차이 중에 하나는 〈말〉을 할 수 있는지에 달려 있다. 말이란 단 어들을 사용하여 문장을 만들고 이를 가지고 시공을 초월하여서 우리
의 의사를 서로 소통하게 한다. 1970 년대에 침팬지에게 몸짓언어나 컴퓨터를 이용하여 말하는 법을 가르치려는 노력이 있었다. 미국 매사추세츠 공과대학 MIT 의 언어학 자인 놈 촘스키 Noam Chomsk y는 〈말이란 사람에게만 있다〉고 결 론을 내렸는데, 이에 대해 데러스 Terrace 는 촘스키의 주장이 일부 틀 렸다고 반박하며 님 촘스키란 침팬지가 사인으로 의사를- 표현하는 것 을 예로 들었다. 그의 주장에 따르면 이 침팬지가 사인(신호)으로 의 사를 전달할 수 있으며, 촘스키와 갇이 일하면 할수록 이 침팬지가 더욱더 신호를 익힐 수 있음을- 알 수 있었다고 보고하고 있다. 데러스는 다른 동물들도 말을 배울 수 있는지를 알기 위해 클레버 한스 Clever Hans 라는 〈영리한 말〉을 가르쳤는데, 이 말은 지난 세기 에 유럽과 미국에서 선을 보였다. 한스라는 이 말은 <2 + 2> 같은 질 문에 답하기 위해서 발굽으로 바닥을 두드렸으나, 실제로 이 질문에 답을 한 것이 아니라 단지 조련사가 주는 어떤 암시에 따라서 발을 두드렸을뿐이었다. 테러스는 침팬지가 이미 알고 있는 단어들을 사용하여 생각을 만들 어낼 수 있도록 문법의 법칙들을 사용할 수 있는지 알고자 하였다. 그러나 그는 〈침팬지는 사람처럼 문법을 사용하여 생각을 표현할 수 없으며, 또한 동물들은 대사를 가르쳐준 후에야 단지 가끔 신호를 보 낸다〉고 결론지었다. 한편 미국 스탠퍼드 대학교의 패터슨은 코코라는 고릴라를 연구하 여 데러스와는 반대되는 결론을 내렸는데, 말이란 더 이상 인간의 전 유물이 아니라고 주장한다. 또 다른 예로 미국 조지아 주의 요크 영장류 센터에 있는 라나라는 침팬지는 펀칭버튼을 이용하여 문장을 만들고 서로 의사를 교환하는 방법을 배웠다. 플로리다 주에 있는 한 야생동물·연구소에서는 일단 의 실험실 침펜지들을 섬에 풀어놓아 그들이 정상적인 생활로 되돌아
갈 수 있는지를 실험하였다. 위에서 살펴본 몇몇 실례에서 볼 수 있듯이, 오늘날 유인원이 말을 배울 수 있는 능력이 있는지에 대해서 많은 논란이 있다. 그러나 오 늘날 사람들은 유인원이 말을 배울 수 있다는 데에 1970 년대보다는 회의를 가지게 되었다. 왜냐하면 침팬지와 고릴라의 발성구조는 사람 의 발성구조와는 다르기 때문에 이들에게 말을 가르치는 것은 별로 소득이 없다는 것이 밝혀졌기 때문이다. 어떤 의미로는 유인원의 의사전달 연구는 유인원이 자신들 그대로 이기보다는 좀더 사람처럼 보인다는 점을 강조하기 위한 생각에서 비 롯된 것으로 보인다. 실례로 침팬지의 경우 사람과는· 다르게 기회에 따라 사냥을 하며 이들이 연모를 사용한다는 것은 갈라파고스 섬에 사는 핀치새들이 그들의 주둥이룰 사용하는 것과 비슷한 것이지 결코 사람이 연모를 사용하는 것과는 같지 않다는 것을 알 수 있다. 오늘날 35000 내지 105000 마리 정도의 오랑우탄과 5000 내지 9000 마리의 삼림 고릴라, 그리고 1000 마리가 채 못 되는 산악고릴라들이 살아 있을 뿐이다. 이들은 오늘날 지구에서 생태계가 파괴됨으로 인 해 멸종의 위기에 처해 있는 동물 중의 하나로, 이는 인류에게 모든 책임이 있다. 사람과 가장 가까운 유인원의 오늘날 상황은 어떠한지 생태학 측면 에서 살펴보자. 식물대는 열대우림 삼림지대로부터 사바나숲, 사바나, 그리고 사막순으로 변하는데 영장류는 이 지역에서 나뉘어 산다. 그 예로 콜로바인 원숭이는 우기 삼림지대에서 살고, 비비를 포함한 다 론 영장류들은 사바나 지대에서 산다. 따라서 이들의 지역분포에 따 라 대륙을 나눌 수 있으며, 또한 대륙을 2 4,,\]간의 활동 사이클에 따 라나누기도한다. 큰 유인원은 모두 열대 우림지대에서 살고 있는데, 오랑우탄은 수 마트라와 자바 등에 살며, 아프리카산 유인원은 아프리카 적도의 우
립지대에서 산다. 이들 모두는 서식지가 황폐되어서 멸종의 위기에 처해 있으며, 동남아시아의 숲은 자꾸 벌목되어가 여기서 살고 있는 오랑우탄둘 또한 다음 세기에는 아주 멸종될 운명에 처해 있다. 열대 우림지대는 지구에서 생물들이 가장 다양하게 살 수 있는 곳 인데 오늘날에는 1 시간당 약 1500km2 씩의 삼림이 개발이란 명목으 로 파괴되어가고 있다. 그 결과 지금까지 알려지지 않은 많은 생물들 이 매일 지구에서 영원히 사라져가고 있다. 대부분의 생태학자들은 중생대 말에 일어났던 생물의 대절멸 사건 과 같은 형태의 철멸이 앞으로 예견된다고 생각한다. 앞으로 일어날 생물의 멸종은 아스데로이드 혜성의 충돌 같은 자연에 의한 재난의 결과가 아니라 바로 사람에 의한 자연의 파괴행위에 기인한다고 생각 한다. 사람에 의한 자연파괴의 결과 무엇이 멸종될 것인가? 같은 규모의 지역에서 볼 때 온대 삼림지대에 살고 있는 나무에는 약 60 여 종의 동물들이 살고 있다. 또한 우기 삼림지대에 있는 한 그루의 나무에서 는 약 264 ;=- 43 종의 개미가 살고 있다. 이들은 영국에 살고 있는 모든 개미의 종류와 맞먹는다. 사실 지금까지 우리는 단지 이들의 종류가 다양하다는 점을 겨우 인식하고 연구를 시작하였을 뿐, 많은 생물들이 우리가 알기 전에 사 라져가고 있다. 잘 알려진 열대 생태학자인 단 얀젠 Dan J anzen 은 〈자연 파괴와 이에 따른 생명체의 멸종상태는 이미 심각한 수준에 도 달하고 있으며, 그리고 남아 있는 숲들도 이미 황폐화되어가는 중이 다 〉라고 문제의 심각성을 경고하고 있다. 우리 중 많은 사람들이 아런 사실들이 우리에게 별로 중요하지 않 다고 생각할지도 모른다. 왜냐하면 우리는 열대에 살고 있지 않기 때 문이다. 그러나 이런 숲둘은 아직 우리가 발견하지 못한 유용한 것들 을 간직하고 있다. 한 예로 미국 국립 암연구소에서는 암의 치료제를
우기 삼림지대에서 자라는 식물에서 구하는 연구를 진행중이다. 어떤 지역에서는 식물의 1% 정도만이 연구되었을 뿐이다. 오늘날 학자들이 지적하기를 숲의 파괴 속도가 너무 빨라서 어떤 식물이 유용하디는 것을 알았을 때는 이미 때가 늦어 벌써 그 식물은 다 멸종되어버릴 것이라고 경고하고 있다. 한편 브라질에서는 벌목을 원하는 목재회사나 목장들 그리고 광산들에 의해 이 지역에서 살고 있는 인디언들의 삼림터가 파괴되어간다. 오늘날 지구상에 살고 있는 영장류의 3 분의 l 이상이 멸종이라는 위험한 상태에 직면해 있다. 그 예로 브라질 동부지역의 낮은 지대 숲속에 사는 황금사자털 타마린 원숭이 Golden Li on Tamar i n 는 그 숫자가 겨우 300 마리 정도이다 . 이런 종류의 영장류는 대개 식량을 필요로 하는, 그리고 숲지대를 그대로 보존할 수 없는 그런 개발도상 국가들에 살고 있다. 따라서 이들의 보존문제는 국제사회 사이에 사 회, 정치, 경제 그리고 과학적인 방법으로 해결해야 할 문제들이다. 한편 유인원들은 아직도 음식으로 또 스포츠라는 취미를 위해서 사 냥되고 있으며, 의학상의 실험을 위해서 사로잡히고 있다. 특히 침팬 지는 사람과 생화학상으로 비슷한 구조를 갖고 있기에 여러 연구와 의학실험에 사용된다. 예로 이들은 오늘날 사람에게 치명적인 후천성 면역결핍증 AIDS 의 치료를 위해 인위적으로 균에 감염되는데, 이런 연구를 위해 이들을 희생시킨다는 것은 우리를 딜레마에 빠지게 한 다. 다시 말해 멸종되어가는 동물들을 희생시켜 가뜩이나 포화상태에 이른 사람이란 〈 종 〉 을 더욱더 포화상태로 이끌어간다는 것이 합리적 인 일일까 하는 비판이 일어난다. 동물원에서 동물을 잡는 것도 야생동물들에게는 하나의 위험이다 . 그러나 미국의 샌디에고 동물원 등에서는 야생동물을 위한 공원을 만 들어서 많은 수의 고릴라를 키우며 보호하고 있다. 그러나 이런 노력 에도 불구하고 고릴라와 오랑우탄은 지구상에서 거의 멸종상태에 이
르렀으며, 번식이 빠른 편인 침팬지도 위험한 상태에 이르고 있는 실 정이다. 유인원은 지구 어느 곳에서나 살 수 있는 사람과는 다르게 주로 숲 에서만 사는데, 이들은 사람처럼 그들 사회에 적응하며 제 몫을 할 수 있도록 사회의 많은 기술 그리고 살아 남을 수 있는 기술들도 배 워야한다. 비록 오늘날 살고 있는 유인원은 어느 정도 특수화되었지만, 만일 우리가 조심스럽게 이들의 해부학상 특징과 행위 및 사회조직 등을 연구한다면 사람의 조상과 유인원 사이에 서로 닮은 점을 찾을 수 있 는데 이 점에서 유인원에 관한 연구는 사람의 조상을 이해하는 데 철 대적으로 필요한 분야이다.
 제 5 장 뼈대와 이빨
제 5 장 뼈대와 이빨
제 5 장 뼈대와 이빨 뼈와 이빨 뼈의 얼개, 기능 그리고 성질 뼈의 얼개(구조) 뼈는 살아 있는 복잡한 유기체이다. 뼈는 2 분의 l 이 무기질이고, 4 분의 l 이 유기질, 그리고 나머지 4분 의 l 이 물로 되어 있으며 그 안 은 골수로 채워져 있다. 노란 골수는 지방울 함유하고 있고 붉은 골 수는 붉은 철세포를 만든다. 뼈는 살아 있는 물체로 자라나는 동안에 변하며, 심지어는 어른인 경우에도 모양이 바뀔 수 있다. 뼈는 한창 성장하는 나이일 때 많이 자라며 어른이 되었을 때는 성장이 멈춘다. 긴뼈의 경우, 뼈 양끝이 자란다(그림 5.1 ) . 연골인 끝부분은 세포들 로 형성되어 있다. 이 세포들은 광물질을 받아들여 그 수가 줄어들며 주위는 핏줄로 채워진다. 특별한 세포는 안으로 이동하여 점차 무기 질화된 연골로 변하면서 뼈 안으로 들어간다. 긴뼈들의 경우 뼈단위 (오스데온) 또는 하버시안 체계 Oste o n or Haversia n S y st em 라고 부 르는 세포 안에 하버시안관 Havers i an Canals 이 있는데 이 관을 통 해서 핏줄이 뼈와 다론 조직과 연결되어 있다(그립 5. 2).
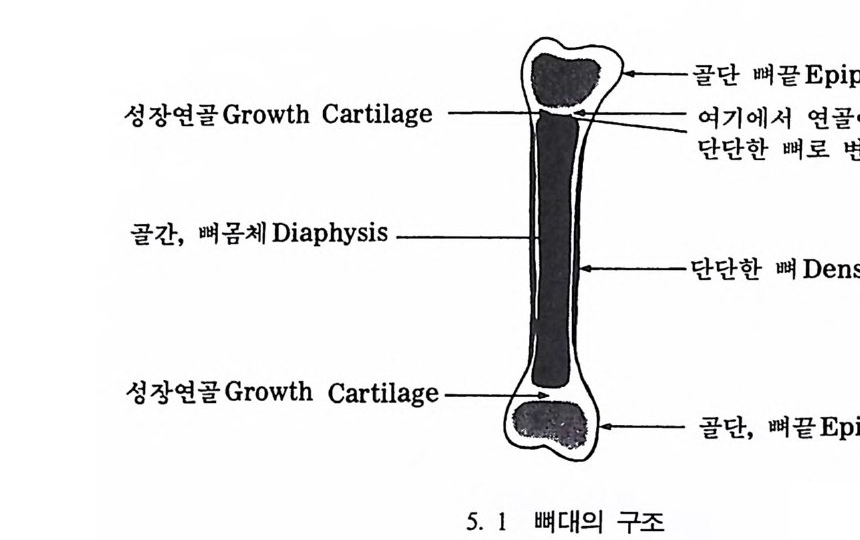 골단 뼈끝 Ep iph y si s
골단 뼈끝 Ep iph y si s
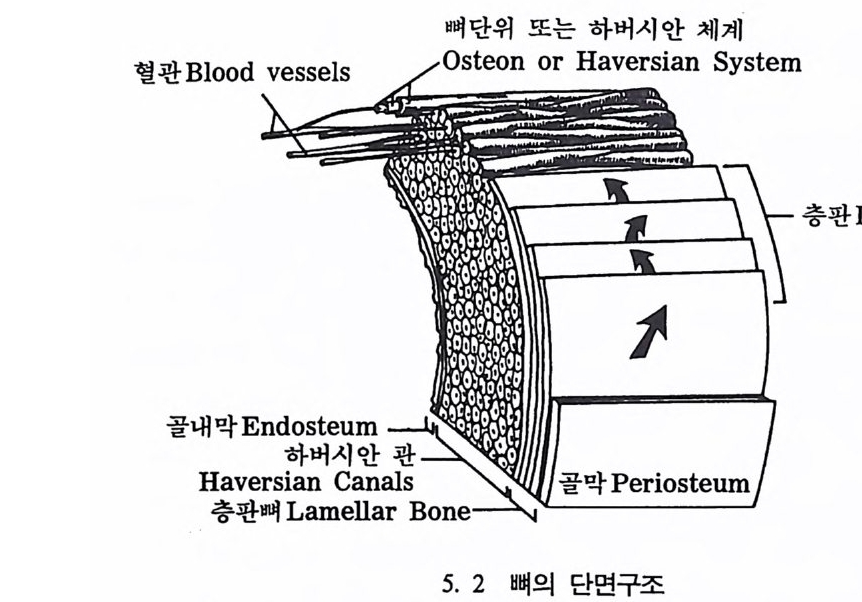 뼈단위 또는 하버시안 체계
뼈단위 또는 하버시안 체계
뼈대의 기능 뼈대는 동물의 몸체를 받쳐주고 보호하는 구실을 하며, 또한 동물 이 재빨리 그리고 효과있게 움직이게 하는 기능을 가지고 있다. 죽
뼈대는 몸체를 지탱 • 보호하고, 운동할 수 있는 기능을 지닌다. 해파리같이 뼈대가 없는 동물들은 천천히 움직일 뿐이다. 곤충들이 나 거미들과 게들은 갑옷걷뼈대를 가지고 있는데, 이는 단단하고 가 벼운 키틴질로 떼어질 수 있다. 몸 안 뼈대들은 몸을 자유롭게 구부 리며 빨리 움직일 수 있도록 도와준다. 사람의 경우 각 뼈둘은 특별 한 기능을 가지고 있는데, 머리뼈에서는 단지 아래턱만이 움직일 수 있다. 머리뼈는 모든 감각기관들을 감싸고 있기 때문에 중요하다. 특히 골을 감싸고 있고, 몸에 에너지를 공급하기 위해 절대적으로 필요한 입이 있다. 등뼈대는 머리등뼈, 가슴등뼈, 허리등뼈로 이루어진 기둥으로 중추 신경을 보호한다. 사람에게는 보통 12 쌍의 갈비뼈가 있으나 20 명 중 1 명은 13 쌍을 가지고 있으며, 갈비뼈통은 내장을 보호한다. 사람은 팔, 다리뼈들로써 몸체를 움직일 수 있고, 발뼈로 바로 설 수 있으며 또한 걸을 수 있다 . 팔꿈치, 무릎과 턱은 돌쩌귀같이 움직 인다. 손목은 부드럽게 움직이고 어깨는 볼과 소켓 형태로 만난다. 많 은 뼈둘은 언제나 함께 움직이는데 이것으로 사람은 부드러운 동작을 할수있다. 한편 머리뼈 부위에서 근육과 머리뼈가 어떻게 서로 작용하는지롤 살펴보기로 하자. 뼈들은 몸에 있어 기본구조의 역할을 하면서 동시 에 스스로가 움직일 수 있다. 뼈의 운동은 결국은 근육들의 운동에 달려 있다. 한 예로 음식이 입 안으로 들어오면 입은 씹는 운동을 하 게 되는데, 이는 음식을 잘게 부수어 위로 보내기 위한 운동이다. 옆머리힘살 Tem p or ali s muscle 은 아래턱과 옆머리뼈에 붙어 있다. 이 근육의 수축에 의해서 아래턱이 위턱에 당겨 붙어 있게 된다. 턱 의 바깥쪽에 붙어서 아래턱을 잡아당기는 역할을 하는 또 다론 근육 으로 턱모서리힘살 Masse t er m. 이 있다. 턱 안쪽으로는 또한 이들 의
에 턱을 잡아당기는 역할을 하는 턱 안힘살 P t er yg o i d m. 이 있다. 턱 모서리힘살과 턱안힘살은 함께 멜빵 같은 모양을 하고 있으며 턱의 안팎에서 턱을 잡아당기는 역할울 한다(그립 5. 3).
 턱 안힘 살 P t er yg o i d m.
턱 안힘 살 P t er yg o i d m.
뼈의 성질 뼈는 그 무게에 비해 매우 강하기 때문에 몸체를 유지하는 데 매우 유용하다. 한 예로 72k g의 몸무게가 나가는 사람은 약 13kg 정도의 뼈를 가지고 있을 뿐이다. 이에 비해서 같은 무게의 철근 구조물이 같은 강도를 유지하기 위해서는 사람에 비해 4 - 5 배 정도의 철근 무 게를 가져야 한다. 다론 예로 참나무로 된, 속이 빈 낚섯대와 속이 찬 낚섯대를 비교해 보면 속이 빈 낚섯대가 약 50% 정도 더 강하다. 뼈는 구성하고 있는 화학성분 때문에 또한 강하다• 뼈의 약 4 분의
l 을 이루고 있는 유기질인 단백질에 의해 뼈는 매우 탄력을 갖게 되 며, 뼈의 약 2 분의 l 정도를 이루는 무기질은 뼈를 강하게 하지만 뼈 롤 갈라지게 한다. 그러나 실제로 뼈는 유기질과 무기질의 성질이 합 쳐서 단단한 성질을 이루며, 이처럼 단단한 뼈의 성질과 뻣속의 빈 구조 때문에 뼈는 가벼우면서도 가장 강도가 센 특성을 지니고 있 다. 이빨의 얼개와 기능 이빨의 얼개 인류를 포함한 영장류의 이빨 얼개는 고인류학자들에게 인류 진화 에 관해 많은 정보를 제공한다. 인류의 이빨에서 찾을 수 있는 특칭 은 영장류를 비롯하여 젖먹이동물 모두에게 적용된다. 따라서 여기서 는 젖먹이동물의 이빨에서 공통으로 사용되는 용어들을 다루기로 한 다. 젖먹이동물은 앞니 Incis o r, 송곳니 Can ine , 옆니 Premolar, 어금니 Molar 의 모양이 서로 다른 모양의 이빨을 가지고 있다. 젖먹이동물 들은 종류에 따라 이빨틀에 박혀 있는 이빨의 수에 차이가 있는데, 이빨의 수를 계산할 때는 위 • 아래턱의 가운데선을 기준으로 나누어 이빨의 수를 센다. 예로 젖먹이동물의 표준 이빨틀은 I 123/123, C 1/1, P 1234(5) / 1234(5), M 123/123 으로 표시되는데, 이를 간단히 표 시하기 위 해서 가운데선을 기준으로 한쪽만 표시하여 3. 1. 4(5). 3/3. 1. 4(5). 무 표시한다. 인류는 2. 1. 2. 3 의 구조를 지니고 있다. 인류를 포함한 영장류의 이빨은 발달해 오는 과정에서 젖먹이동물 의 기본적인 이빨 얼개를 지니고 있다. 젖먹이동물의 앞니는 비교적 작고 편평한 모양을 하고 있다. 송곳니는 크고 이빨 끝이 날카로우며 다른 이빨보다 더 솟아나 있다. 옆니는 작은 편이며 씹는 면이 불록
솟아나 있는데 이룰 도드리 cus p라고 부른다. 또한 옆니는 이빨 표면 사기질에 돌아가면서 덧띠가 발달해 있었는데 전화과정중 이 덧띠는 옆니와 어금니에서 결도드리로 변하였다.
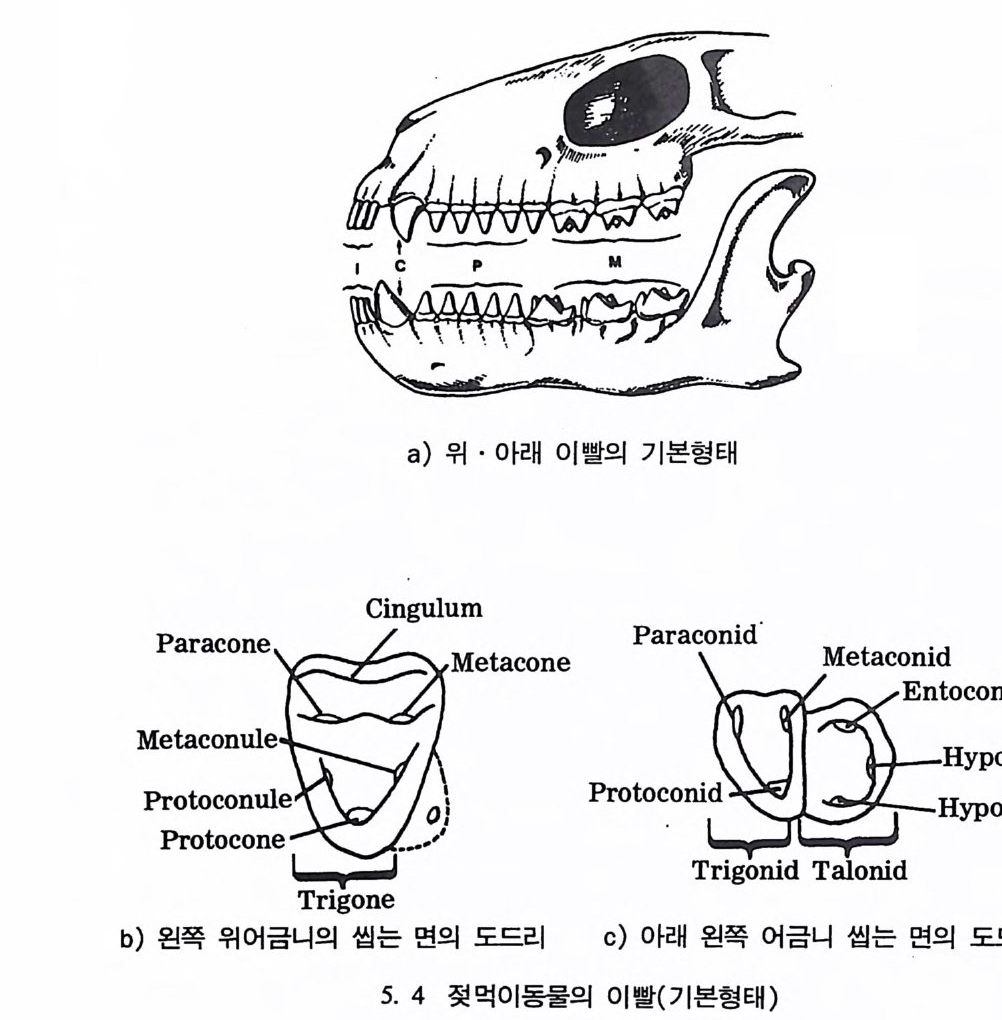 a) 위 • 아래 이빨의 기본형태
a) 위 • 아래 이빨의 기본형태
젖먹이동물의 위턱 어금니는 초기에 Proto cone, Paracone 그리고 Me t acone 이라고 불리는 세 개의 도드리를 지니고 있었으며, 이 세
개의 도드리가 있는 이빨 부위를 트라이곤 Tr ig one 이라고 부른다. 모 든 젖먹이동물둘 위어금니의 도드리 형태는 이 세 개의 도드리 형태 룰 바탕으로 하는데, 젖먹이동물의 이빨 전화를 말할 때 이를 〈 세 도 드리 이론 The Trit ub ercular Theor y 〉 이라고 한다(그림 5. 4). 위어금니는 도드리 이의에 이빨 표면에 사기질 능선들이 발달해 Pro t oconule 와 Me t aconule 를 이루고 있다.
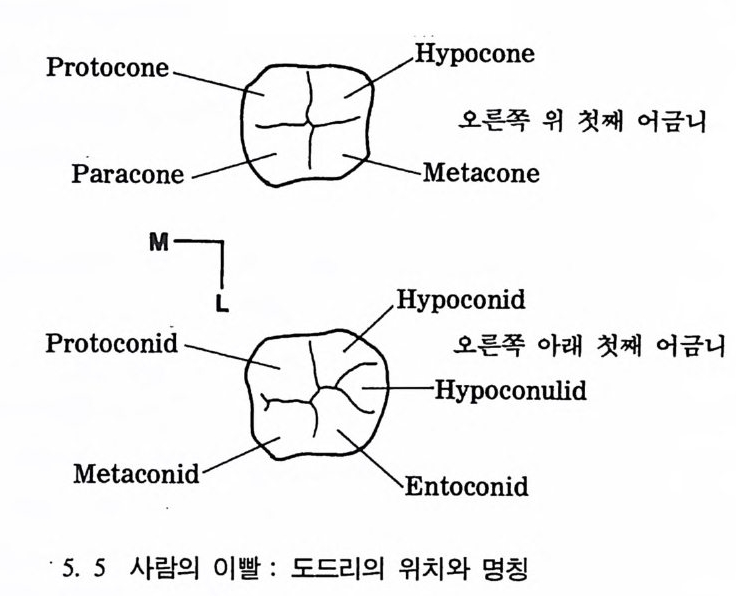 Proto c one
Proto c one
초기 젖먹이동물의 아래어금니는 Tr ig on i d 와 Talo ni d 라고 불리는 부분으로 나누어진다. 이때 앞부분인 Tr ig o ni d 는 뒷부분인 Talo ni d 보 다 더 발달해 있다. 세 개의 도드리 인 Proto c on id, Paracon id, Me- t aco ni d 는 T rig o ni d 에 자리하고 있다. 음식을 씹울 때 위어금니의 Tr ig one 부분은 아래어금니의 Tr ig o ni d 와 맞닿아 음식롤 자르는 기능 울 하며, 입을 다물고 있을 때 위어금니의 Pro t ocone 은 아래어금니의 Talo ni d 와 맞닿아 있다. 이런 형태의 젖먹이동물의 이빨울 트라이보
스페 닉 Trib o sph en ic 모양이 라고 한다. 이런 도드리의 모양은 영장류를 분류할 때의 특징으로 이용된다. 그 예로 원숭이는 도드리와 도드리가 서로 연결되는 바이로포돈트인 데 반해 사람과 유인원은 도드리가 이빨 표면에 팬 홈 Groove or Fis - sure 에 의해서 각기 분리된다. 또한 도드리는 낱개로 찾아진 이빨의 위치와 순서를 결정하는 데 이용된다. 도드리를 뜻하는 단어 끝에 -cone 이 붙이면 위이빨울, -co ni d 가 붙으면 아래이빨울 나타낸다. 사 람과 유인원의 경우, 위어금니의 경우 Pro t ocone 은 항상 이빨 앞 • 안 쪽 Mes i a!-L i n gu al 에 자리하며, 아래어금니의 경우는 Pro t ocon i d 는 이빨 앞 • 바깥쪽 Mes i a!-Lab i al 또는 Buccal 에 자리한다(그림 5. 5). 이빨의 기능 이빨의 기능을 이해하기 위해서 이빨울 작은 사각 모양으로 생각해
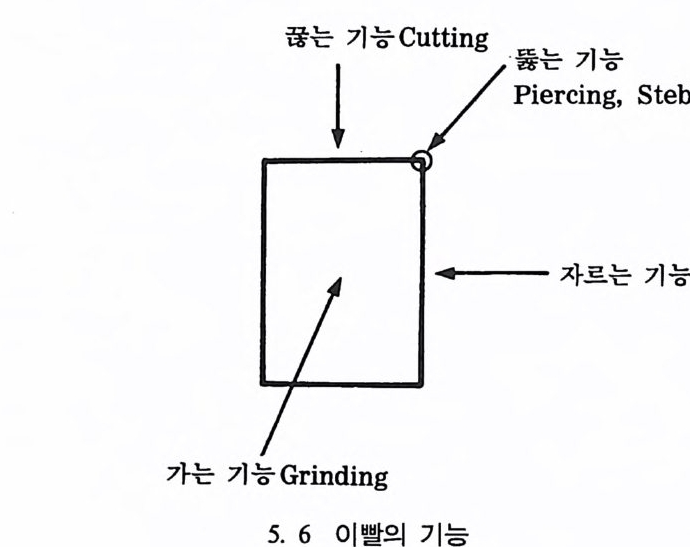 끊는! 기 능 Cutt ing
끊는! 기 능 Cutt ing
보는 것이 유용하다(그립 5.6). 이빨은 그 뿌리에 의해 위 • 아래턱 몸체에 박혀 있는데, 이빨의 모양은 섭취하는 음식과 밀접한 관계가
있다. 예로 사람의 앞니는 날을 사용하며, 송곳니는 뾰족한 면을, 그 리고 어금니는 음식을 갈기 위해 편평한 면을 사용한다. 서로 다론 형태의 동물들은 서로 다른 음식물을 취한다. 예로 많은 작은 영장류들은 곤충을 잡아먹는데, 곤충으로부터 영양을 얻기 위해 서 그들은 곤충을 감싸고 있는 단단한 각질을 부수어야 한다. 그래서 그들의 이빨은 날카롭고 뾰족하며 사각의 모서리가 강조된다. 나뭇잎을 먹는 동물은 잎을 소화하기 쉽게 작은 조각으로 잘라야 한다. 따라서 이빨은 편평하고 날과 같은 형태을 지니고 있다. 사각의 각을 주로 사용하는데, 이 위아랫니의 날은 마치 가위처럼 잎을 자론 다. 낱알을 먹는 동물은 편평한 이빨면을 가지고 있는데, 이는 낱알을 갈기 위한 것이다. 사람의 뼈대 사람 몸의 얼개는 매우 복잡하고 많은 부분으로 나누어지는데, 몸 의 얼개 중 뼈대의 모양과 기능은 해부학의 중요한 한 분야이다. 뼈 와 이빨은 자연 부패 과정에서 화석으로 찰 남을 수 있기 때문에 사 람이 죽어 땅에 묻힌 후 생기는 가장 중요한 칭표이다. 인류학자들은 이들 화석을 통해 이 주인공이 얼마나 큰 몸집을 가졌는지, 얼마나 오래 살았는지, 또 건강상태는 어떠했는지 등을 알 수 있다. 더욱 중 요한 것은 이들 화석의 연구를 통해서 화석 주인공들의 행위에 대한 실마리를 얻을 수 있는데, 가장 중요한 것으로는 걷기 방법과 이들의 행위 그리고 이들이 먹던 음식 등을 가늠할 수 있다. 복원은 기능과 관계 있는 이빨이나 뼈들의 모양과 . 형태를 관찰함 으로써 가능하다. 복원된 모습을 통해 유전 특칭을 알 수 있으며, 또 한 해부학상 모습은 이들이 살았던 환경과의 관계에서 형성되었기 때
문에 이들의 모습은 유전 특칭의 산물이자 또한 이들이 살았던 환경 의 산물이라고 말할 수 있다. 기본되는 해부학 용어 뼈대를 다루기 위해서는 해부학 용어에 관한 정확한 의미를 알아야 한다. 이 용어들은 사람뿐만 아니라 젖먹이동물 모두에게 적용된다. 해부학에서 사용되는 가장 기본되는 용어들은 아래와 갇다(그림 5.7; 5. 8 ; 5. 9). I. 해부학상 자세 Anato m ica l Posit ion . 해부학상 자세 란 사람이 똑바로 서서 두 발을 모은 차려 자세를 하고 오른손바닥이 앞을 향해 있는 모습을 말한다 . 이런 자세로 서 있으면 긴 뼈들이 서로 교차되 지를 않는다. 만일 손등이 앞으로 향하면 앞팔뼈와 뒤팔뼈가 서로 교 차되는데, 팔꿈치에서 앞팔뼈는 뒤팔뼈보다 밖에 있으나 손목에서는 반대 위치에 있다. 한편 뼈의 방향에 관해 말할 때, 뼈대는 항상 우리 자신이 서 있는 자세에서 시작된다. 사람이 이렇게 해부학적 자세로 서 있을 때는 아 래와 같은 3 개의 기본선 Planes 으로 나누어진다. I) 정중선 (Sa gitt al 또는 Mi ds ag ittal Plane) : 몸을 좌우로 이등분하는 선 2) 앞뒤선 (Fron t al 또는 Coronal Plane) : 몸을 앞뒤로 이등분하는 선 3) 위아래선 (Transverse 또는 Horiz o nta l Plane) : 몸을 위아래로 이등분 하는선 2. 몸체와 뼈대에서 쓰이는 방향에 관한 낱말 여기에서 방향을 나 타내는 낱말은 상대적인 뜻을 지닌다. I) 몸체
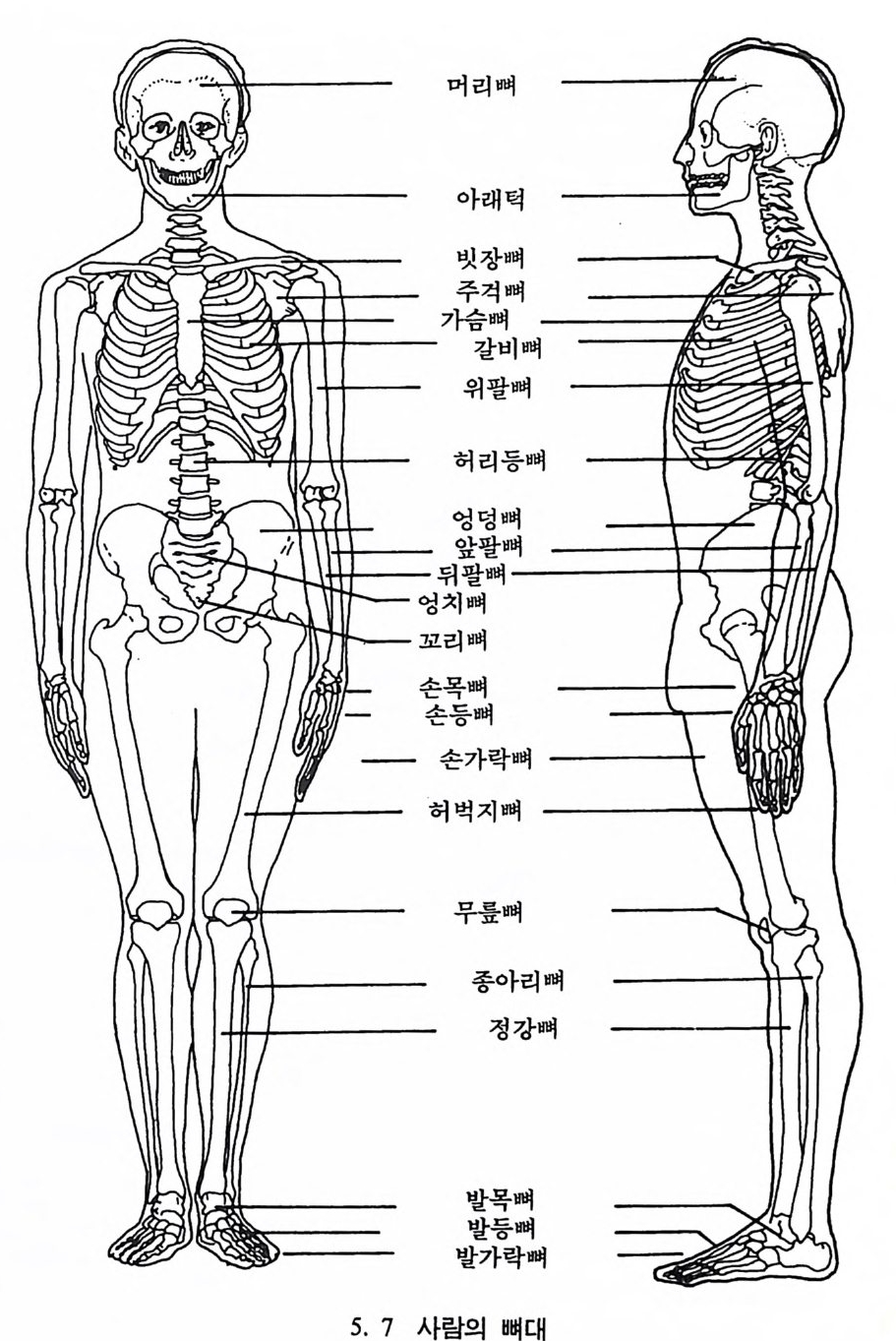 머리뼈
머리뼈
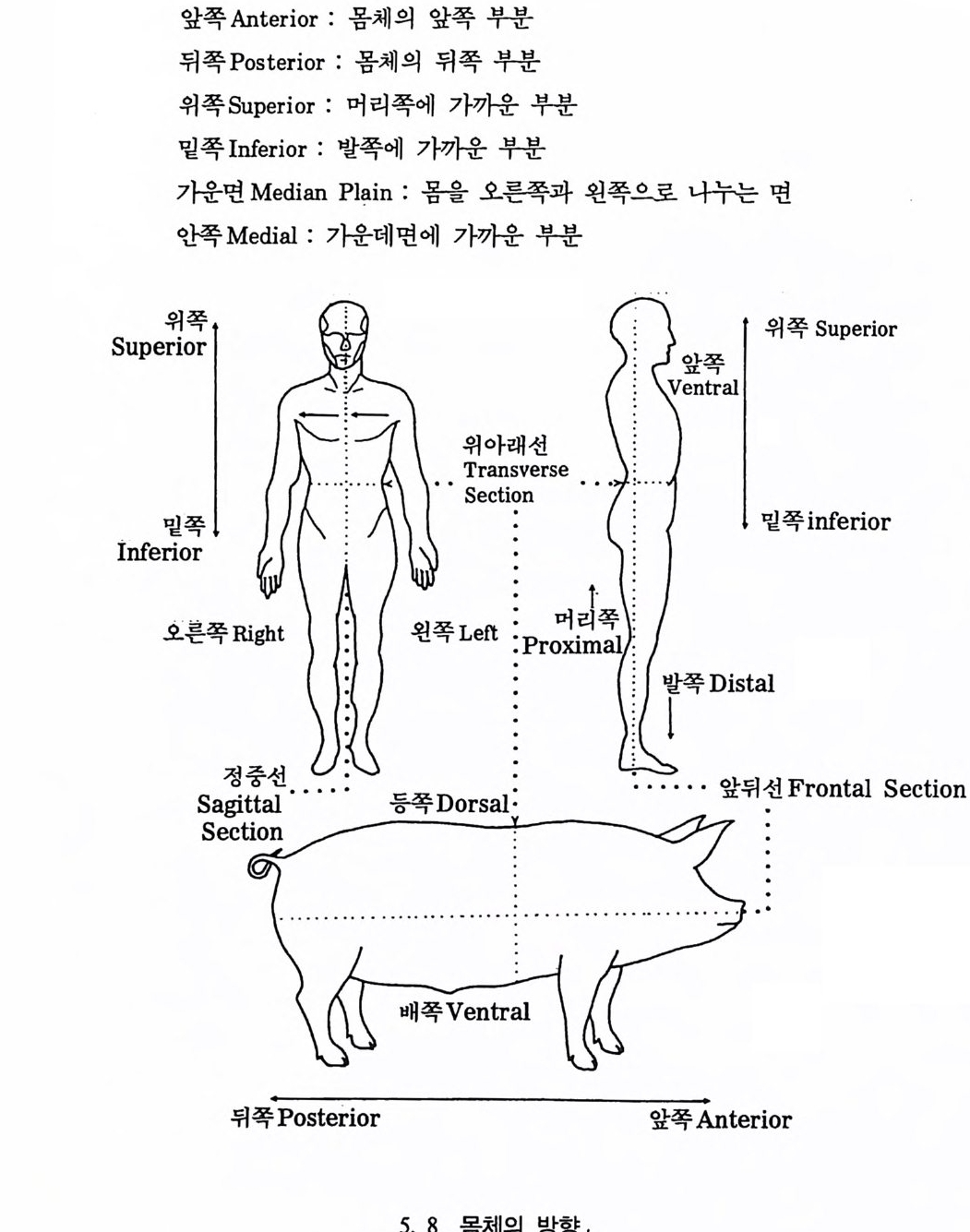 앞쪽 An t er i or : 몸체의 앞쪽 부분
앞쪽 An t er i or : 몸체의 앞쪽 부분
갓쪽 La t eral : 가운데면에서 먼 부분 2) 사지뼈 머 리쪽 Prox im al : 머 리쪽에 가까운 부분 발쪽 D i stal : 머리에서 먼 부분 3) 이빨의 방향 앞쪽 Mesia l : 이빨이 턱 앞쪽에 가까운 경우 뒤쪽Di stal : 이빨이 턱 앞쪽에서 먼 경우 입술쪽 (Lab i al 또는 Buccal) : 이빨 바깥쪽 혀쪽Li n g ual : 이빨 안쪽
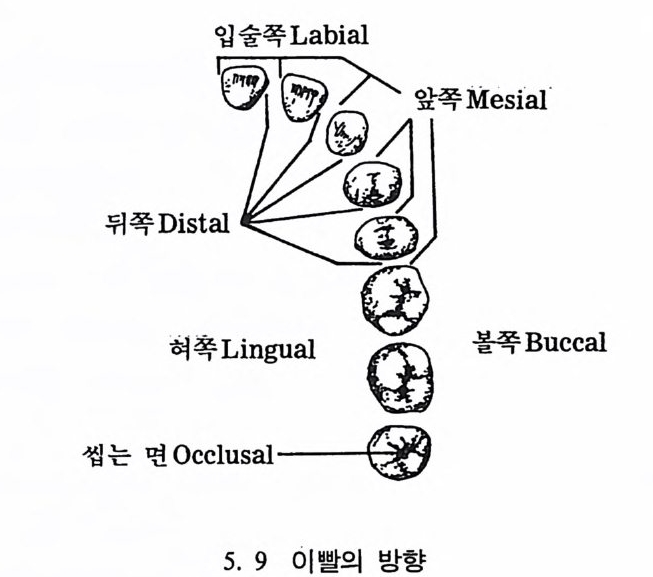 입lE술a쪽1尸 L`ab i、 al Mesia l
입lE술a쪽1尸 L`ab i、 al Mesia l
3. 모양에 따른 뼈의 구분 뼈는 모양에 따라 긴뼈, 짧은뼈, 납작뼈 그리고 불규칙뼈로 나누어 전다. 1) 긴뼈 : 사지뼈롤 이루고 있는 뼈로 몸무게가 일부 여기에 실리고, 근 육이 붙어 몸이 움직이기에 필요한 지레의 기능을 가지고 있다.
2) 짧은뼈 : 손등뼈와 발등뼈를 말한다. 뼈는 단단하고 신축성이 있다. 3) 납작뼈 : 머리뼈와 엉덩뼈 그리고 주걱뼈를 포함한다• 이 뼈들은 감싸 고 있는 기관을 보호하고, 근육이 와서 닿는다. 4) 불규칙뼈 : 등뼈와 손가락 • 발가락뼈, 머리뼈의 일부가 여기에 속한다. 이의에 뼈대에는 인대가 붙는 자리가 뼈에 남아 있게 된다. 이런 자리를 크기에 따라 볼록점 Tubercle, 불룩점 Tuberosit y, 솟은점 Tro- chante r , 솟기 Process 등으로 부른다. 뼈대의 이름 몸의 힘살과 힘살띠(인대) 그리고 두터운 덧(건)들은 모두 뼈대에 걸리는데, 뼈대를 이루고 있는 각 뼈들은 모두 이름을 갖고 있다. 어 론의 뼈대는 206 개의 뼈들로 구성되며·그 중 상당수가 쌍으로 되어 있다. 태아일 때는 800 개의 뼈를 만드는 부분이 있고 400 개는 태어날 때 뱃속에 남으며, 자라서 어른이 되면 206 개의 뼈만 남는다. 뼈대를 이해하는 데는 사람 자신이 훌륭한 길잡이가 된다. 즉 자신 의 몸의 구조를 생각하는 것이 가장 좋은 방법이다. 뼈대는 크게 머 리뼈대와 몸체뼈대로 구분된다. 1. 머리뼈 Ossa cran i 이음새 (維合 : Sutu ra : Su t ures) 라고 불리는 접합에 따라서 여러 조각으로 나누어진다. 머리뼈는 어느 부분은 한 개의 뼈로 이루어져 있으며, 어느 부분은 쌍으로 이루어져 있다. l) 머리뼈 이음새 앞머리 이음새 (Su t ura corona lis : 冠狀維合) : 앞머리뼈와 양윗머리뼈를 잇는이음새 윗머리 이음새 (Su t ura sag ittalis : 矢狀鍵合) : 윗머리뼈를 잇는 이음새 뒷머리 이음새 (Su t ura lambdoid e a : 人字維合) : 양윗머리뼈와 뒷머리뼈 를 잇는 이음새
옆머리 이음새 (Su t ura squ amosa : 麟狀維合) : 옆머리뼈와 윗머리뼈를 잇는이음새
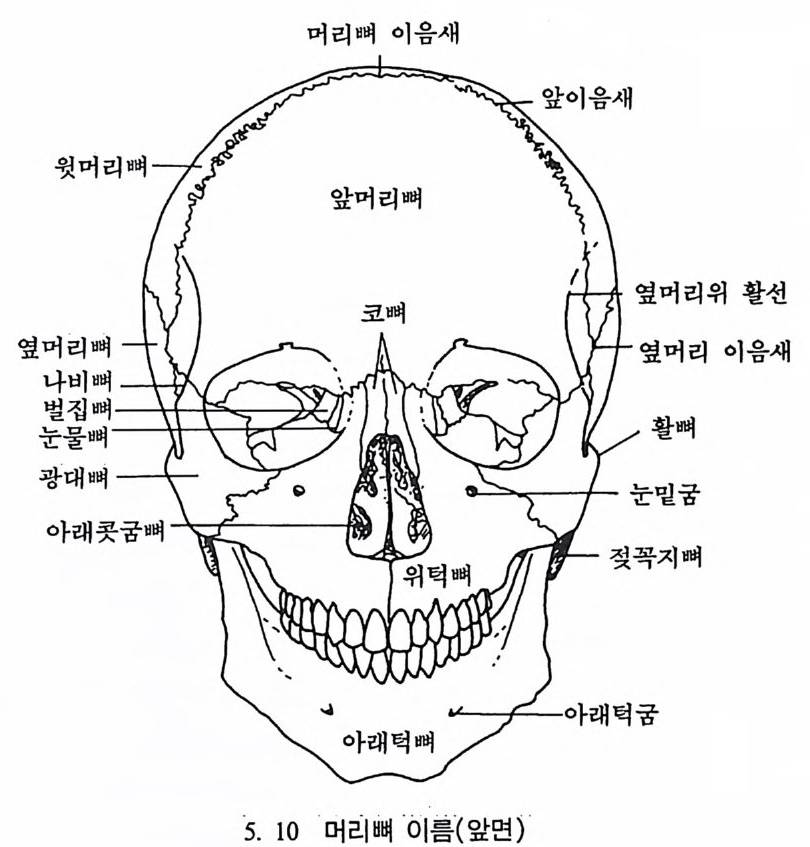 머리뼈 이음새
머리뼈 이음새
2) 머리뼈대 이름과 수(그립 5.1 0-5. 12 ) 하나로된 뼈 수
앞머 리 뼈 Os fron ta l e 前頭骨 뒷머리뼈 Os occip ital e 後頭骨 나비 뼈 Os sph enoid a le 蝶形骨 lll 아래턱 Mand ibu lar 下額骨 벌집 뼈 Os eth m oid a le 飾 骨보습뼈 Vomer 勘舌 骨骨 목젖뼈 Os hy oi d e um 7 모듬 쌍으로된 뼈 수
윗머리뼈 Os pa rie t a l e 頭頂骨 2 22222222 옆머리뼈 Os tem p o rale 側頭骨 코뼈 Os nasal 鼻 骨 위턱 Ma xilla 上類骨 광대뼈 Os zyg o mati cu m 類 骨 눈물뼈 Os lacrim ale 派 骨 입천장뼈 Os Palati nu m IJ蓋骨 아래뼈콧굼 Concha nasa lis infer io r 下鼻骨 말레우스 귀뼈 Malleus 인카스 귀뼈 Incus 스티에프 귀뼈 St ap es 모듬 222. 몸체 뼈대 하나로된 뼈 수
목등뼈 Verte b rae cervic a les 頭 椎 725 가엉허 슴치리등뼈등뼈 뼈O sVV seearrctteer ubb rmraa ee ltuhmo rbaaclaelse s 仙尾胸腹胸 椎骨椎椎椎 l 꼬리 뼈 Os coccy gis 가슴뼈 S t ernum모듬 27 쌍으로 된 뼈(몸체 윗부분) T^ 주걱뼈 Scap u la 周甲骨 2 빗장뼈 Clavic u la 鎖 骨 2 갈비뼈 Costa e 助 骨 24 위팔뼈 Humerus 上腕骨 2 앞팔뼈 Radi us 機 骨 2 뒤팔뼈 Ulna 尺 骨 2 손목뼈 Ossa carp i 手根骨 16 손등뼈 Ossa meta c arpi 中手骨 IO 손가락뼈 Ossa digitor um manus 基節骨 28 모듬 88 쌍으로 된 뼈(몸체 아래 부분) T^ 엉덩뼈 Os Coxae 骨 盤 2 받침뼈 Os ilium 腸 骨 감춤뼈 Os pu bis 恥 骨 앉음뼈 Os isc h ii 坐 骨 허벅지뼈 Femur 大旦骨 2 무릎뼈 Pate l la 藤蓋骨 2 정강뼈Ti b i a 歷 骨 2 종아리뼈 Fib u la 骨 2 발목뼈 Ossa tar si 足根骨 14 발등뼈 Ossa meta t a r si 中足骨 10 발가락뼈 Ossa digitor um pe d is 基節骨 28 모듬 62 모든모듬 206
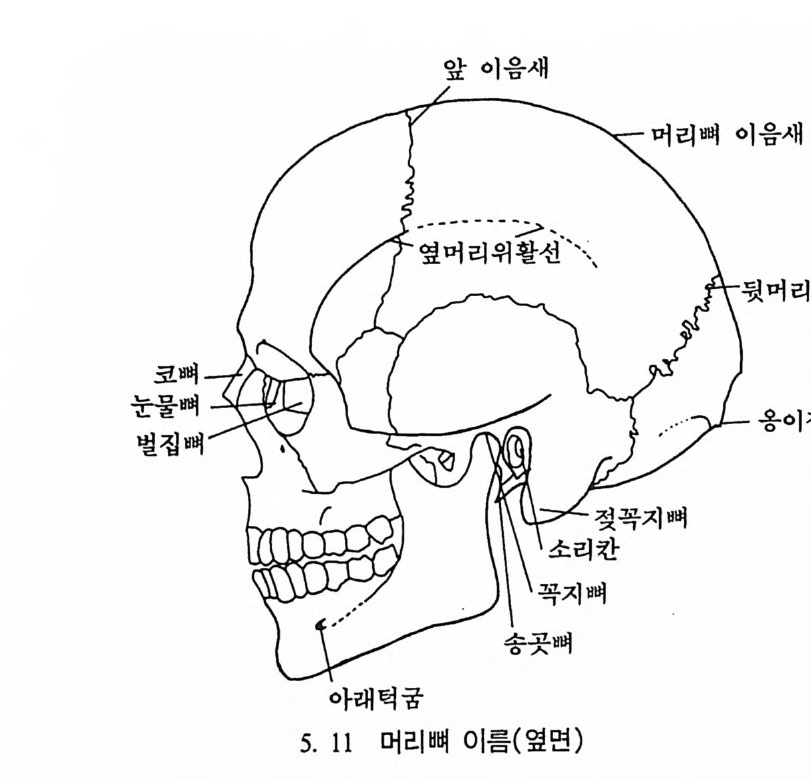 앞 이음새
앞 이음새
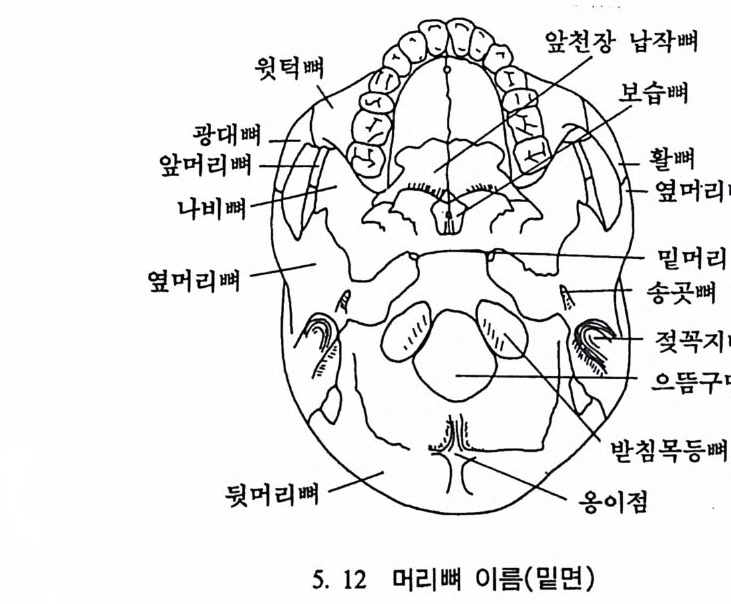 활뼈
활뼈
사람뼈 재기 사람의 체질 특칭을 비교하기 위한 방법으로 해부학상의 형태를 비 교하는 방법과 특징을 객관화하기 위해 수치로 표시하는 재기방법을 사용한다. 일부 학자들은 재기방법의 중요성에 의문을 제기하고 있으 나 이는 방법 자체보다는 오히려 단지 재기목록을 만둘기 위해 재기 를 하는 것을 지적한 것이다. 다시 말해 단순히 뼈의 크기와 각도 동 울 재서는 정확히 설명하는 데 도움을 주지 못한다는 점을 의미하는 것 0] 다. 재기방법은 오히려 정확하게 사람 뼈대의 특성에 관한 자료를 얻기 위한 객관적 수단으로, 다양한 통계학 방법을 이용하여 잰 값이 지니 는 의미를 설명할 수 있다. 따라서 정확한 사람뼈 재기와 잰 값은 개 인의 특칭뿐만 아니라 한 집단의 형태와 주변집단과의 천연성을 밝히 는 데 중요한 구실을 하며, 특히 옛 사람의 뼈는 전화과정에서의 관 계를 규명하는 한 방법으로 이용된다. 더욱이 머리뼈는 주민집단의 형태비교에 비교적 객관성을 띠기 때문에 인종의 구분이나 천연성을 연구하는 데 주로 이용된다. 그 동안 사람뼈 재기에 관한 다양한 방법들이 마틴 Mar ti n, 게일즈 Gi les, 게일즈와 엘리엇 Gi les & Ell iot 그리고 하웰스 Howells 등의 학 자들에 의해서 제시되었다. 이 책에서는 마틴의 Lehrb~h Der Anth ~ o- po log ie, 1928 를 바탕으로 하웰스의 연구를 함께 표시한다. 뼈를 재기 전에는 다음과 갈은 점에 주의해야 한다. 먼저 모든 뼈 의 정확한 재기점의 위치를 알아야 한다. 특히 머리뼈에는 많은 재기 점들이 있기 때문에 이들의 위치를 정확히 파악해야 한다. 다음에 모 든 뼈의 재기는 갇은 조건 아래서 전행되야 하는데, 무엇보다도 머리 뼈는 국제학계에서 정한 기준선 Frank fo rt Plane 의 위치에 놓고 재야 한다. 이 기준선은 오른쪽과 왼쪽 귓점 그리고 왼쪽 눈굼밀점의 세
점을 통과하는 수평면에 놓인다. 그립이나 사진 그리고 도판 등을 만 둘 때도 머리뼈는 이 기준선에 위치해야 한다. 여기서 사용되는 기준 점과 해부학상의 용어는 손보기의 우리말 연구(손, 1988) 를 바탕으로 한다. 재기를 통해 얻어진 자료들과 이 해석은 체질인류학의 다론 연구 분야(예 : 살아 있는 사람의 체질적 특칭과 뼈의 형태, 지문과 혈액형 의 비교, 그리고 유전학상 연구 등)와 결합하여 사람뼈에 관한 우리의 이해를도와준다. 사람뼈재기 An t hro p ome t r y는 살아 있는 사람뿐만 아니라 죽은 사 람의 뼈를 재는 기술로, 대상에 따라 아래와 같이 네 개의 분야로 나 누어전다.
Somato m etr y -살 아 있는 사람의 몸 재 기 Ce phalome tr y - 살아 있는 사람의 머리와 얼굴 재기 Oste o metr y -사 람뼈 재 기 Cranio m etr y -사 람의 머 리 뼈 재 기 사람뼈 재기 중 특히 머리뼈 재기에 필요한 기준점은 여러 곳에 있 는데, 크게 홀점과 마줏점의 두 개의 형태로 나눌 수 있다(그립 5. 13 ; 5. 14 ; 5. 15) . 첫째, 흩점은 머리나 몸체의 가운면에 있는 점으로 하나씩밖에 없 다. 예를 들어 코끝에는 오직 하나의 점이 있다. 둘째, 마줏점은 머리뼈의 가운데선을 중심으로 양쪽 같은 거리에 쌍으로 있는 점을 말한다. 예로 오른쪽 귀끝에 있는 한 접은 반대쪽 의 갇은 위치에 있는 점과 대칭된다. 재기방법은 재기점 사이에서 이루어지며 그 결과 두 접 사이의 거 리가 얻어진다(예 : 머리뼈 길이는 190mm). 이때 길이는 밀리미터 단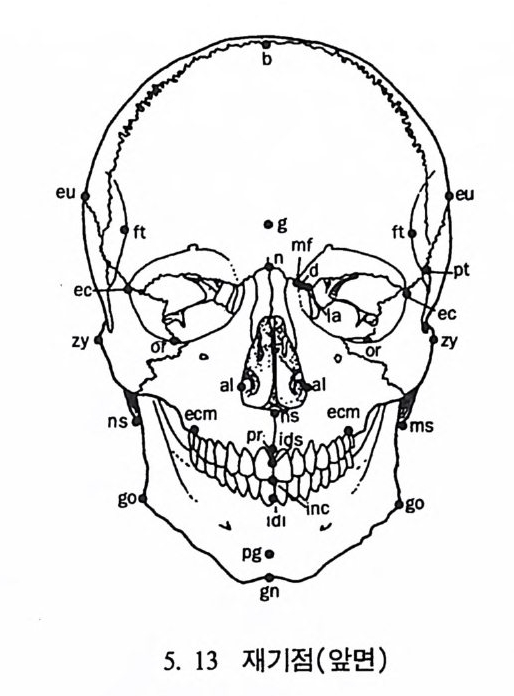 eu eu
eu eu
 b
b
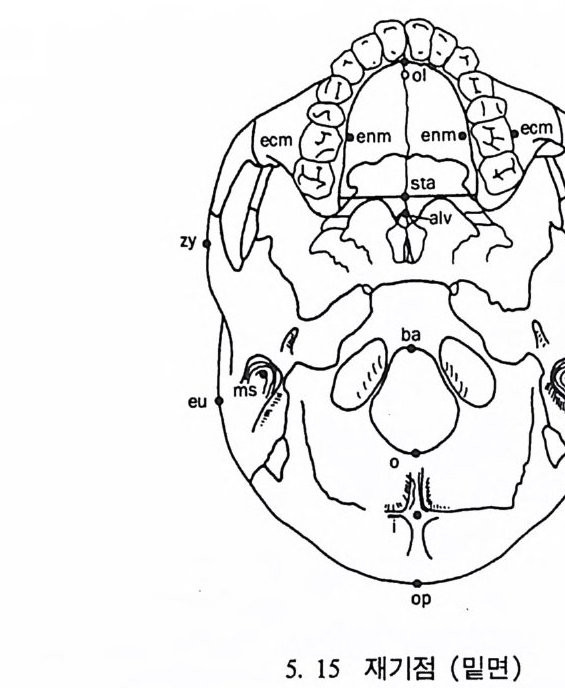 zy zy
zy zy
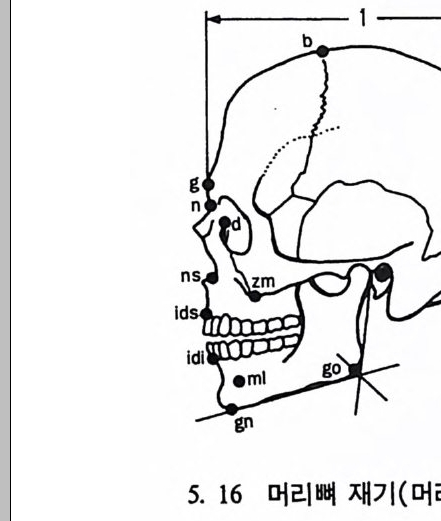 Id
Id
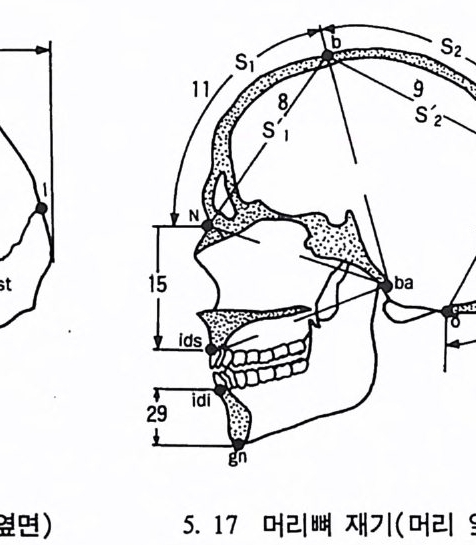 5. 17 머리뼈 재기(머리 옆면)
5. 17 머리뼈 재기(머리 옆면)
위로 표시되며, 오른쪽과 왼쪽에서 잰 값을 둘 다 얻을 때 비교는 같 은 쪽의 잰 값에서 이루어짐을 원칙으로 한다. 잰 값들은 뼈의 특칭 을 나타내기 위해 지수로 표시된다(예 : 머리뼈지수는 머리뼈 너비/ 길이의 비율을 나타낸다).
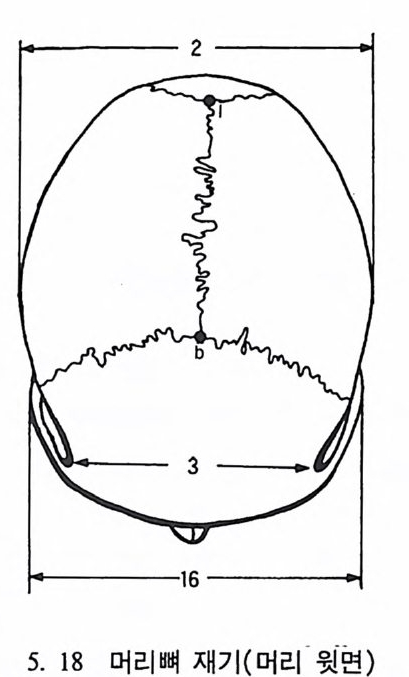 2
2
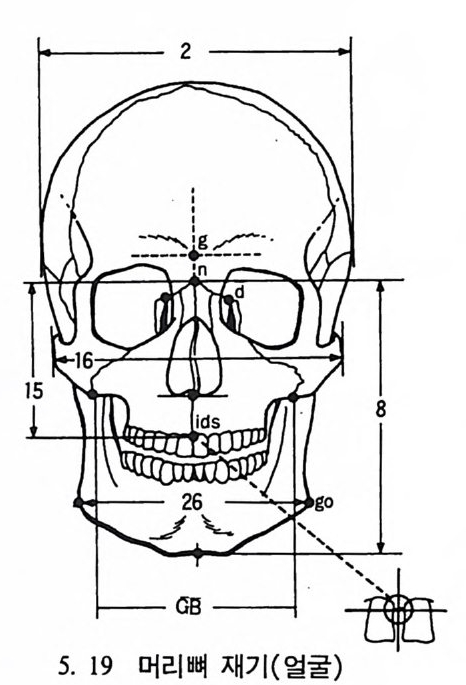 2
2
사람뼈 재기에는 재기에 이용되는 많은 기준점이 있지만 여기에서 는 가장 일반적으로 사용되는 기준점과 재기방법을 살펴보기로 한다. 머리뼈 재기방법과 잰 값에서 얻어지는 지수는 아래와 같다. 기준점 은 그림 5. 13-5. 21 에 설명한다. 머리뼈 재기 (그림 5.16-5. 21) 머리뼈 1) Ml(GOL ) 머리 죄대길이 : 섭 사이(g)~뒷머리정 (o p)
2) M8(XCB) 머리 조 1 대너비 : 옆불룩접 (eu) ~옆불룩점 (eu) 3) M9(WFB) 앞머리뼈 조|소너비 : 옆머리뼈갓점(ft)~옆머리뼈갓점(ft) 4) M17 머리 죄대높이 : 으뜸점 (ba)~ 정수리점 (br) 머리지수 : 머리 최대너비 X100/ 머리 최대길이 긴 머리 X- 7 4.99 가운 머리 75.00-79.99 짧은 머 리 80.00-84.99 꽤 짧은 머리 85.00-X 머리 모습 (Cr ani al Module) : 대강의 머리 크기를 알려준다 . (머리 최대길이+머리 최대너비+머리높이 )/3 머리길이 • 높이지수 : 으뜸점-정수리점 높이 XlOO/ 최대길이 낮은 머리 X- 6 9.9 9 가운 머 리 70.00-7 4.9 9 높은 머리 75.00-X 머리 너비 • 높이지수 : 으뜸점-정수리점 높이 XlOO/ 최대너비 낮은 머리 X-91.9 9 가운 머리 92.00- 9 7.99 높은 머리 98.0 0 - X 평균 높이지수 : 으뜸점-정수리점 높이 XlOO/ 머리길이 • 너비의 평균 낮은 머리 X-80.4 9 가운 머리 80.50-83.4 9 높은 머리 83.50-X(Hrdl ic ka, 1952) (스튜어트 (S t ewar t, 1962) 가 제시한 이 방법은 길이 • 높이지수와 너비 • 높이지수 대신 사용된다.) 평균 으뜸접-높이지수 : 으뜸점-정수리점 높이 XlOO/ 머리길이 • 너 비의 평균 낮은 머리 X-78.99 가운 머리 79.00- 8 5.99 높은 머리 86.0 0 -X
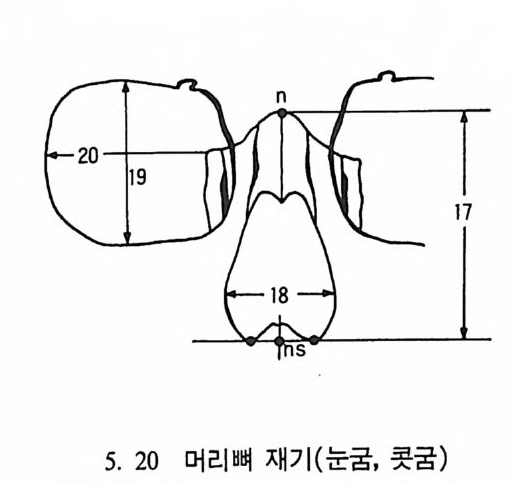 17
17
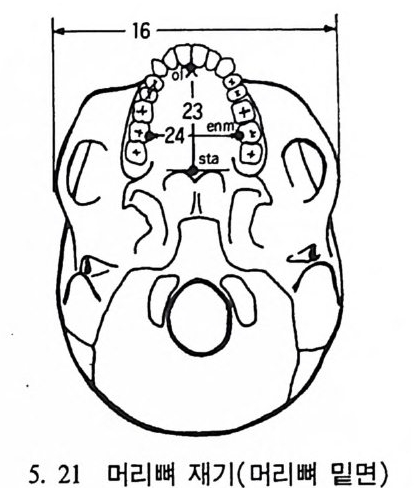 5. 21 머리뼈 재기(머리뼈 밑면)
5. 21 머리뼈 재기(머리뼈 밑면)
5) M20 머리높이 : 귓굼윗점(p o)- 정수리점 (br) 평균 귓굼윗점 • 높이 지수=귓굼윗점-정수리점 XlOO/ 머리 최대길이+머리 최대너비 /2 낮은 머리 X-66.99 가운 머리 67.00- 7 1 .99 높은 머리 72.0 0-X 6) 머리높이 : 으뜸점 (ba) ~귓굼윗점(p o) 7) M21 머리 수직높이 : 귓굼윗점(p o)~ 소리칸 수직점(p o) 8) M26(S,') 앞머리뼈 곧은길이 : 코맞점 (n)~ 정수리점 (br) 9) M27(S2') 윗머리뼈 곧은길이 : 정수리점 (br) -맞음목(I) IO) M28(S3') 뒷머리뼈 곧은길이 : 맞음목(I)-으뜸뒷점 (o) 11) M29(FRC) 앞머리뼈 굽은길이 : 코맞점 (n)- 정수리점 (br) 12) M30(PAC) 윗머리뼈 굽은길이 : 정수리점 (br)- 맞음목(I) 13) M31(OCC) 뒷머리뼈 굽은길이 : 맞음못(I)-으뜸뒷점 (o) 14) M45(ZYB) 얼굴너비(광대뼈 너비) : 광대정 (z y)~광대접 (z y) 15) M47(NGH) 얼굴 조 1 대길이 : 코맞접 (n) ~턱밀가운접(gn) 16) M48(NAH) 위얼굴높이 : 코맞접 (n) ~위잇몸접(i ds)
얼굴지수 : 얼굴 최대길이 X lOO/ 얼굴너비 꽤 넓은 얼굴 X- 7 9.9 9 넓은얼굴 80.00 -84.99 가운얼굴 85.00-89.99 좁은 얼굴 90.0 0 - 9 4.9 9 꽤 좁은 얼굴 95.00-X 위얼굴지수 : 위얼굴높이 XlOO/ 얼굴너비 꽤 넓은 이마 X-44.9 9 넓은 이마 45.0 0 -49.9 9 가운 이 마 50.00-5 4.9 9 좁은 이마 55.0 0 -5 9.9 9 꽤 좁은 이마 60.00-X (Marti n, 1928) 17) M5l( OB B) 눈굼너비 : 앞머리 위턱점(mf)~눈굼옆점 (ec) 18) M52(0BH) 눈굼높이 : 눈굼너비에 수직되는 최대높이 눈굼지수 : 눈굼높이 X lOO/ 눈굼너비 넓은눈굼 X-82.99 가운눈굼 83.00-89.99 좁은눈굼 90.00-X 19) M54(NLB) 코높이 : 코맞점 (n) ~코밀가운점 (ns) 20) M55(NLH) 코너비 : 코굼갓접 (al) ~코굼갓점 (al) 코지수 : 코너비 XlOO/ 코높이 좁은 콧굼 X-47 .9 9 가운콧굼 48.0 0 -5 2.99 넓은 콧굼 53.0 0 -X 21) M60 입전장길이 : 위잇몸가운점(p r) ~입천장뒤끝점 (alv) 22) M51(MAB) 입천장너비 : 위어금니안점 (ec t)~위어금니안점 (ec t) 입천장지수 : 입천장너비 X lOO/ 입천장길이 좁은 입천장 X-109.99
가운 임천장 110.00-I1 4.9 9 넓은 입천장 115.00 一 X 23) M62 임전장길이(안쪽) : 앞니가운접 (ol) ~입천장뒤점 (s t a) 24) M63 입전장너비(안쪽) : 위 어금니 양갓점 (erun) 사이 아래턱 (그립 5. 22, 5. 23)
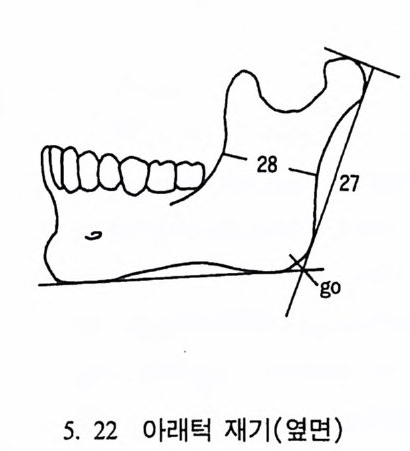 5. 22 아래턱 재기(옆면)
5. 22 아래턱 재기(옆면)
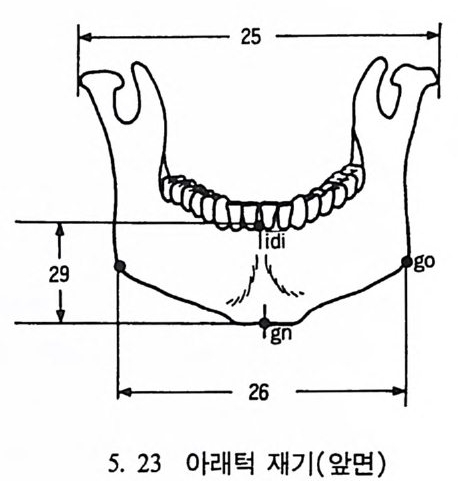 29
29
25) M65 아래턱 꼭지너바 :· 아 래턱 양꼭지점 (cdl) 사이 26) M66 아래턱 모서리너비 : 아래턱 양모서리(g o) 사이 27) M69 아래턱 이음새높이 : 아래턱밀가운접(gn)~아래잇몸가운점(idi) 28) M70 아래턱 가지높이 : 아래턱 모서리(g o) ~아래턱 꼭지점 (cdl) 29) M71 아래턱 가지 죄소너비 : 아래턱까지의 가장 좁은 곳의 너비 The Measurements of Human Bones The Measurements of the Skull Cran ial Bones I) Ml(GOL) Maxim um crania l leng th
2) M8(XCB) Maxim um cran ial breadt h 3) M9(WFB) Mi nim um fron t al breadth 4) Ml7 Maxim um heig h t Cran ial Index = Ma xim um crania l breadt h x 100 / Maxim um crania l leng th Doli ch ocrany X- 7 4.99 Mesocrany 75.00- 7 9.9 9 Brachy cr any 80.0 0 -8 4.9 9 Hy pe rbrachy c rany 85.0 0 -X Crania l Module = Leng th + breadth + heig h t / 3 Cran ial Leng th- H eig h t Index = Basio n - b regm a heig h t X 100 / Maxim um leng th Chamaecrany X-69 .9 9 Orth o crany 70.00-7 4.9 9 Hy psi c r any 75.0 0 -X Cran ial Breadth - H eig h t Index = Basio n - b reg m a heig h t X 100/Maxim um breadt h Tape i n o crany X- 9 1. 99 Metr i o crany 92.00 -9 7.9 9 Acrocrany 98.0 0 -X Mean Heig h t Index = Basio n-bregm a heig h t X 100/ Mean of crania l leng th+ breadth Low X-80.4 9 Med ium 80.5 0 - 8 3.4 9 High 83.5 0 - X (Ste w art, 1952) Mean Basio n-H eig h t Index = Basio n - b regm a heig h t X 100 / Cran ial leng th+ breadth /2 Low X-78.99
Mediu m 79.0 0 - 8 5.9 9 Hig h 86.00- X 5) M20 Porio n - b reg m a heig h t Mean Porio n - H eig h t Index=Pori on -bregm aX 100/ Cran ial leng th+ breadth /2 Low X-66.99 Mediu m 67.0 0 -71 . 99 High 72.0 0 -X(Ste w art, 1965) 6) Basio n - p o rio n heig h t 7) M21 Auric u lar heig h t 8) M26(S1') Fronta l chord 9) M27(S2') Parie t a l chord 10) M28(Si' ) Occip ital chord 11 ) M29( FRC ) Fronta l arc 12) M30(PAC) Parie t a l arc 13) M3l(OCC) Occip ital arc 14) M45(ZYB) Bi zy g o mati c breadth 15) M47(NGH) Tota l fac ia l heig h ~ 16) M48(NAH) Up pe r facial heig h t Tota l Faci al Index = Tota l fac ia l heig h t x I 00 / Bi zy g o mati c breadth Hy pe reuryp ro sop y X-79 .99 Euryp ro sop y 80.00-8 4.99 Mesop ro sop y 85.0 0 -8 9.99 Lep top ro sop y 90.00-9 4.99 Hy pe rlep top ro sop y 95.00-X Up pe r Faci al Index = Up pe r facial heig h t X 100 / Bi zy g o mati c breadth
Hy pe reurye n y X-44.9 9 Eurye n y 45.00- 4 9.99 Meseny 50.0 0 -5 4.99 Lep ten y 55.00-5 9.99 Hy pe rlep ten y 60.00-X(Marti n, 1928) 17) M5l(OBB) Orbit breadth 18) M52(0BH) Orbit heig h t Orbit al Index = Orbit al heig h t x 100 / 0rbit al breadth Chamaeconchy X-82 .99 Mesoconchy 83.0 0-8 9.99 Hy psi c o nchy 89.00-X 19) M54(NLB) Nasal breadth 20) M55(NLH) Nasal heig h t Nasal Index = Nasal breadth X 100 / Nasal heig h t Lep tor rhi ny X-47 .99 Mesorrhi ny 48.0 0 -5 2.99 Platy rr hi ny 53.00-X 21 ) M60 Maxil lo- a lveolar leng th 22) M61 (M AB) Ma xillo -alveolar breadth Ma xillo -Alveolar Index = Maxil lo- a lveolar breadth X 100 / Ma xillo -alveolar leng th Doli ch urany x-109. 99 Mesurany 110.00 기 14.99 Brachy uran y 115.00-X 23) M62 Palata l leng th (int e r nal) 24) M63 Palata l breadth (int e r nal) Mand ibu lar 25) M65 Inte r condy la r widt h
26) M66 Big o n ial breadt h 27) M69 Sy m p h y s eal heig h t 28 ) M70 Ascendin g ramus heig h t 29 ) M71 M ini m um ascendin g ramus breadth 몸체뼈 재기 1. 엉치뼈 (그림 5. 24) 1) 앞 조 1 대높이 Maxim um ante r io r heig h t : 엉 치뼈 가운몸체 sacral pro m- on t ar y에서 엉치뼈 끝까지의 길이 (A-B)
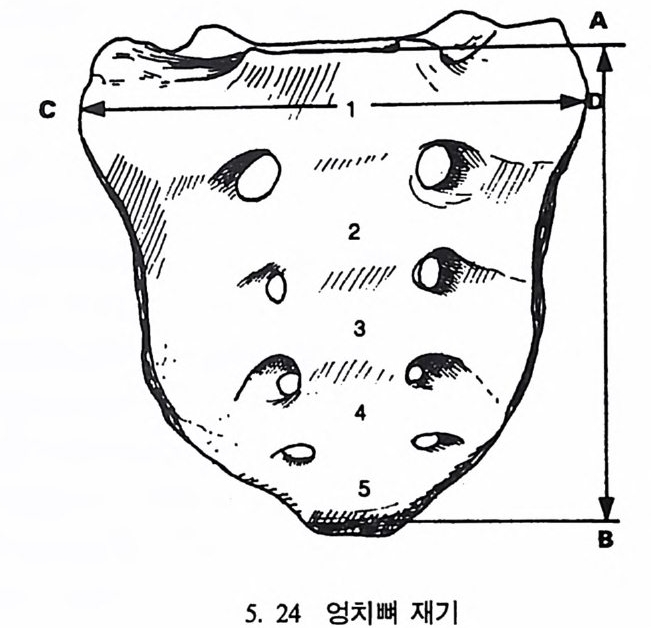 c
c
2) 앞 조|대너비 Maxim um ante r io r breadth : 엉치뼈 첫번째 날개부분 사 이의 가장 넓은 너비 (C-D)
2. 주걱뼈(그림 5. 25) 1) 조 1 대걸이 : 날개안각 Su p er i or Border 끝에서 날개밀각 In fer i or Bor- der 끝까지 직선거 리 (A- B ) 2) 조 1 대너비 : 주걱뼈굼 Gleno i d Fossa 의 가운데에서 주걱뼈마루 끝 Sp ina l A xi s 까지 거리 (C- D )
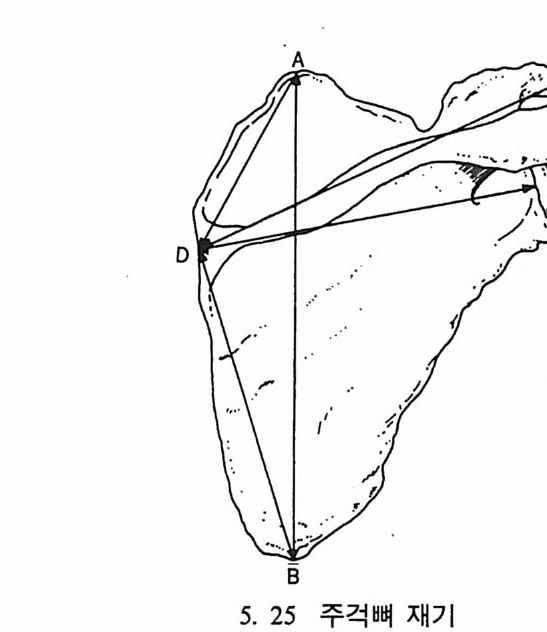 E
E
3) 주걱出 1 마루길이 : 주걱뼈마루 끝 S pi nal A xi s 에서 주걱뼈봉우리 Acr.- om ion Process 까지 거리 (D-E) 4) 마루우|우묵 0| 거리 : 날개안각 끝에서 주걱뼈마루 끝 S pi nal A xi s 까지 거리 (A-D) 5) 마루밑우묵이거리 : 주걱뼈마루 끝에서 날개밀각 끝까지 거리 (D-B) 주걱뼈지수 : 최대너비 XlOO/ 최대길이 3. 빗장뼈(그림 5. 26) 1) 조 1 대길이 : 가슴뼈맞자리와 주걱뼈맞자리 끝까지 거리 (A-B) 2) 몸체 가운 둘레 : 몸체 가운데 부분의 둘레
빗장 - 위팔뼈지수 : 빗장뼈최대길이 X !OO/ 위팔뼈 최대길이 빗장뼈 강건지수 : 몸체 가운 둘레 X!OO/ 최대길이
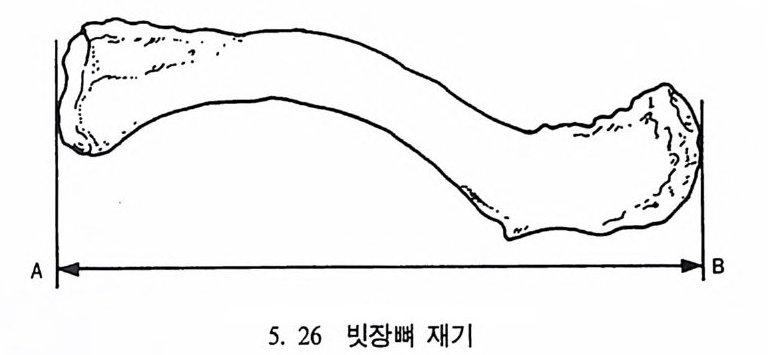 A B
A B
4. 위팔뼈(그림 5. 27) 1) 조|대길이 : 고정된 판에 뼈롤 놓고 위팔뼈머리와 위팔뼈도르래 사이 의 가장 큰 직선거리 (A-B) 2) 몸제 가운 조 1 대너비 : 뼈대 중간의 좌우 거리 (M-N) 3) 몸제 가운 죄소너비 : 뼈대 중간의 앞뒤 거리 (S-T) 최대너비와 수직으로 잰다. 4) 위팔뼈머리 죄대너비 : 머리 가장자리 사이의 최대너비 (C-D) 5) 몸제 조|소둘래 : 세모불룩이 Delto i d Tuberos ity에서 잰 둘레 위팔뼈 강건지수 : 몸체 최소둘레 X lOO/ 최대길이 앞팔 ― 위팔뼈지수 : 앞팔뼈 최대길이 XIOO/ 위팔뼈 최대길이 5. 앞팔뼈(그림 5. 28) I) 조 1 대길이 : 앞팔뼈머리에서 앞팔뼈끝솟기 Sty lo id Process 끝까지 거 리 (A-B) 위팔- 앞팔뼈지수 : 앞팔뼈 최대길이 X lOO/ 위팔뼈 최대길이 6. 뒤팔뼈(그림 5. 29) 1) 조 1 대길이 : 부리솟기 Olecranon Process 에서 뒤팔뼈끝솟기 Sty lo id Process 끝까지 거리 (C-D)
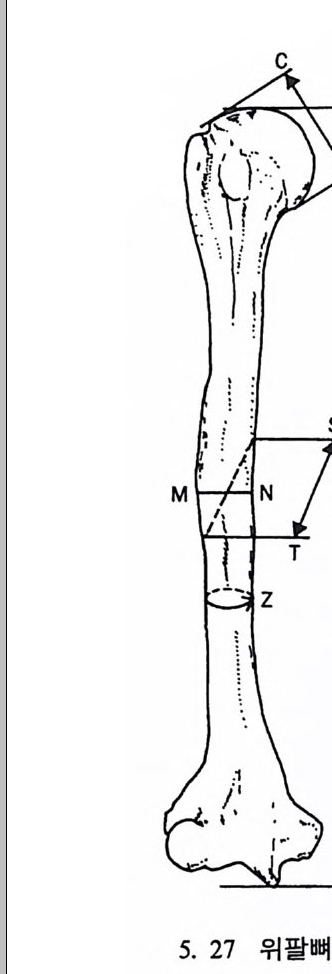 B
B
 A
A
 c
c
2) 뒤팔뼈길이 : 뒤팔뼈머리솟기 Semi lun ar No t ch 부터 뒤팔뼈머리까지 거리 (A-B) 3) 몸제 죄소둘래 : 팔 아래쪽 골단의 바로 윗부분에 비스듬하게 위치 한 점 (E) 에서 잰 둘레 7. 엉덩뼈(그림 5. 3 0) 1) 조 1 대길이 : 앉음뼈 lsc hi um 와 받침뼈윗둔덕 Iliac Crest 사이의 최대 직선거리 (A-B) 2) 조 1 대너비 : 받침뼈앞둔덕 Ante r io r-sup e rio r Iliac S pi ne 과 받침뼈뒷
둔덕 Poste r io r supe r io r Iliac Crest 사이의 거리 (C - D) 앉음뼈 · 감춤뼈지수 Isc hi um - Pub i s Index : (그림 5. 31) 감춤뼈 길이 (A - C)X100/ 앉음뼈 길이 (A-B)
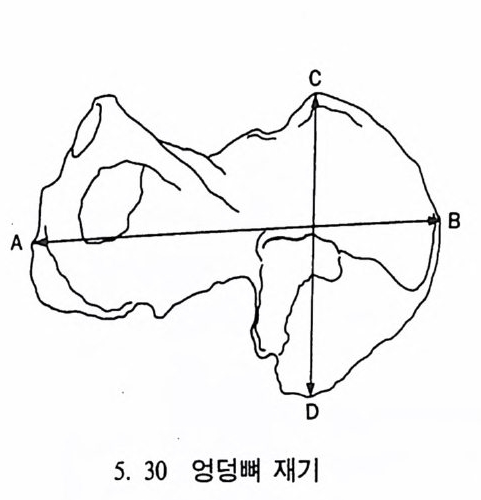 c
c
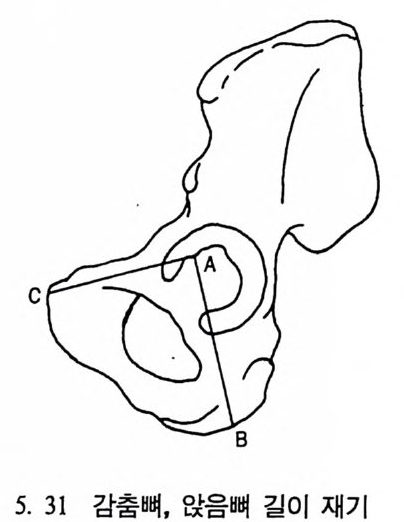 c
c
8. 허벅지뼈(그림 5. 3 2) l) 조 1 대길이 : 허벅지뼈 머리에서 안도르래 Med ial Cond y le 까지의 최대 직선 (A-B) 2) 도르래-머리길이 : 양도르래 Late r al-med ial Cond y le 와 머리 사이 길이 (C-D) 3) 몸제 가운 앞 • 뒤 너비 : 몸체의 가운데에서 앞 • 뒤 사이 너비 (S- T) 4) 몸제 가운 좌 • 우 너비 : 몸체의 가운데에서 좌우 사이 너비 (M-N) 로 3) 번과 직각으로 잰다. 5) 허벅지뼈 머리 조|대지름 : 허벅지뼈 머리의 최대너비 (E - F) 6) 몸체 가운 둘래 : 몸체 가운데의 둘레. 천으로 된 테이프를 가지고 뼈의 윤곽을 따라 돌려가며 잰다 . 7) 앞둔덕 앞 • 뒤 너비 : 앞둔덕 Lesser Trochan t er 의 바로 밀의 앞 • 뒤
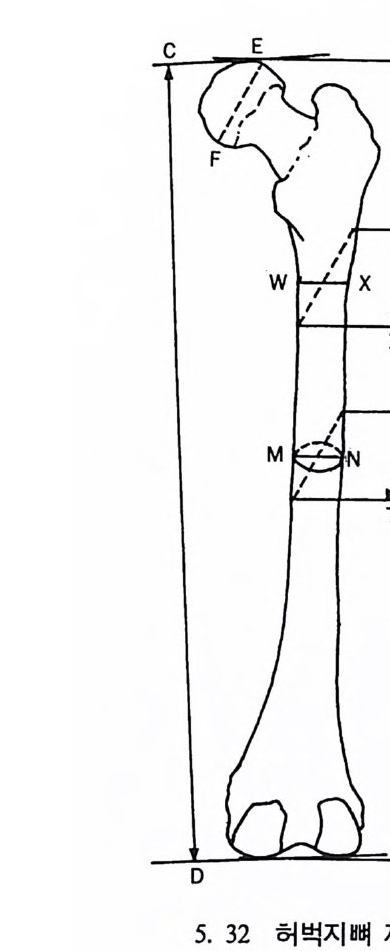 z
z
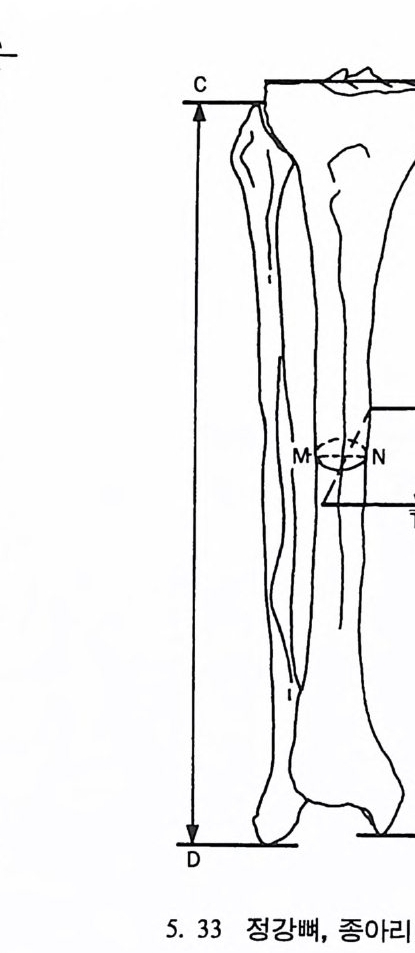 D
D
너비 (Y-Z) 8) 앞둔덕 좌 • 우 너비 : 앞둔덕의 바로 밀의 좌 • 우 너비 (W - X). 7) 번과직각으로잰다. 허벅지뼈 납작지수 Pla tym er i c Index : 앞둔덕 앞•뒤 너비 x • 100/ 앞둔덕 좌 • 우 너비 납작허벅지뼈 Platy me ric X-84.9 보통허벅지뼈 Eurym e ric 85.0-99.99 불허벅지뼈 Ste n omeric 100 . 00-X( 주로 병리학상의 경우임)
강전지수 Robus ti c ity : 몸체의 상대적 크기를 나타낸다. 몸체가운 앞•뒤 너비+좌·우너비 x 100/ 도르래-머리길이 9. 정강뼈 (그림 5. 3 3) l) 죄대길이 : 불룩이 Malleolus 끝에서 안도르래 Late r al Cond y le 의 가 장 높은 점까지의 직선길이 (A- B ) 2) 핏구멍 앞 • 뒤 너바 : 정강뼈 핏구멍에서 앞 • 뒤 사이 너비 (S-T) 3) 핏구멍 좌 • 우 너비 : 정강뼈 핏구멍에서 좌 • 우 사이 너비 (M-N). 2) 번과 직각으로 잰다. 역시 핏구멍의 높이에서 잰다. 4) 핏구멍 둘레 : 핏구멍 둘레를 풀라스틱 데이프로 잰다. 정강뼈 납작지수 Pla ty cne mi c Index : 핏구멍 좌 • 우 너비 XIOO/ 핏구멍 앞·뒤 너비 꽤 납작정 강뼈 Hy pe rpl a ty cn em ic X-54 .9 납작정 강뼈 Platy cn emi c 55.0-62.9 보통정강뼈 Mesocnemi c 63.0-69.9 불룩정 강뼈 Euryc n emi c 70.0-X 10. 종아리뼈(그림 5. 33) l) 조|대길이 : 위와 아래끝의 가장 먼 거리를 챈다 (C-D) 남녀 가늠하기 뼈대로써 남녀를 가늠하기에는 엉덩뼈가 가장 좋은 기준이 되며, 머리뼈와 사지뼈들도 이용된다. 머리뼈와 사지뼈의 경우 남녀 가늠은 뼈대의 일반 특징에 바탕을 둔다. 남성의 뼈는 여성보다 더 크고 굳 세 보이며 근육이 닿은 면이 두드러진다. 그러나 철대 차이가 있는 것은 아니어서 중간형이 제법 발견된다. 이들에서 보이는 두드러지는 특칭둘은다음과같다.
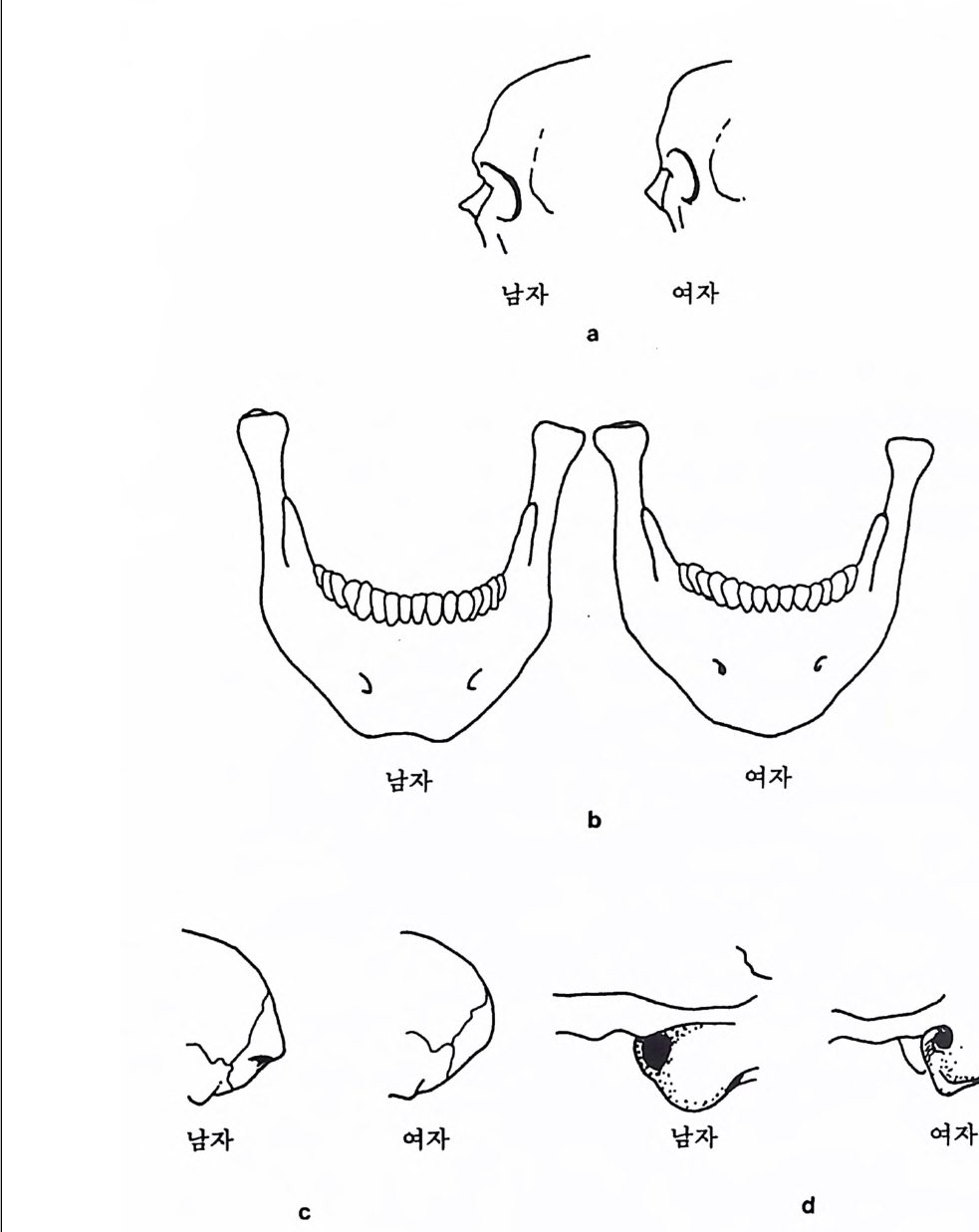 남자 a 여자
남자 a 여자
5. 34 남녀 가늠하기 남녀의 머리뼈 차이 a: 눈두덩과 앞이마 b: 아래턱 C: 응이점 d : 젖꼭지뼈
1. 머리뼈(그림 5. 34) I) 얼굴 • 남성의 눈두덩이 여성보다 더 튀어나왔다. •눈굼의 위 가장자리가 여성의 경우 날카롭고 남성은 둔하다• • 입천장뼈는 남성의 것이 여성에 비해 더 크다. ·이빨은 남성이 더 크다. 2) 머리통 ·여성의 머리뼈가 남성에 비해 더 작고 선이 부드럽다. 또한 여성의 머리뼈는 앞머리뼈, 윗머리뼈가 어린아이처럼 두드러지게 솟아 있 다. ·남성의 뒷머리뼈는 여성에 비해 크고 근육이 닿는 마루가 더 잘 발 달했다. ·남성의 광대뼈 솟기의 뒷부분이 여성에 비해 더 발달되어 있다. •젖꼭지뼈의 경우 남성이 여성의 것보다 크다. • 앞머리뼈 속의 빈 공간인 앞빈굼 S i nus Fron tali s 은 남성이 더 크다. 3) 아래틱 •남성의 경우 턱의 각이 져 있으며, 여성의 경우 턱 끝이 남성에 비 해 뾰족하며 둥글다• • 이빨은 남성이 더 크다. 2. 엉치뼈(그림 5. 35) • 일반적으로- 옆에서 볼 때 남성의 엉치뼈가 여성보다 더 휘어져 있다. ·남성의 경우 엉치뼈 날개가 몸체보다 작으며, 여성은 날개와 몸체가 갇은 크기를 보인다. 3. 엉덩뼈(그림 5.36) :남녀를 가늠하기에 가장 좋은 부위로 정확도 가가장높다. I) 감춤뼈 ·여성은 뼈가 가늘고, 길기 때문에 두 감춤뼈 아래각이 남성에 비해 크다. 또 집게손가락을 감춤뼈 이음새와 나란히 놓았을 때 여성의 것은 엄지손가락이 찰 움칙일 수 있는 데 비해 남성의 것은 약간
움직일 수 있다. • 활 모양 인대가 닿는 면의 형태 -감춤뼈와 앉음뼈 • 감춤뼈가지 Is-chi op u bic Ramus 에 남녀를 가늠해 주는 감춤뼈의 이음새 모양, 감 춤뼈 아래의 각도, 앉음뼈 • 감춤뼈가지의 옆모양 등 세 가지 대표되 는 특칭이 있는데, 이것으로 95% 이상 정확하게 남녀를 가늠할 수 있다(그림 5. 37).
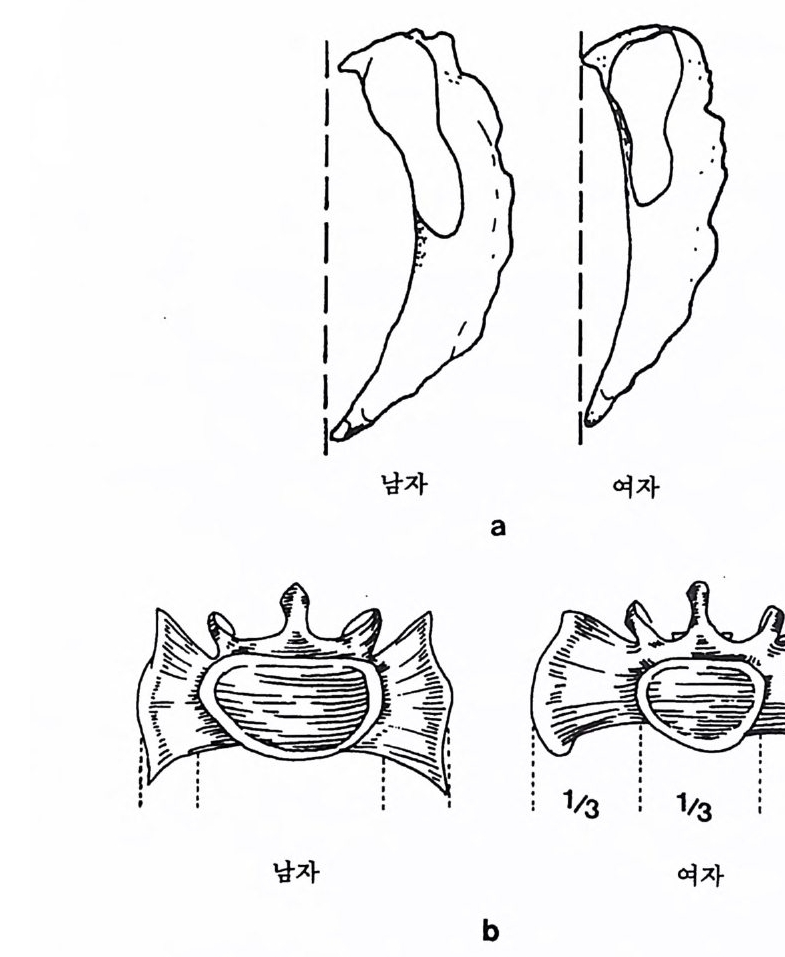 남자 a 여자
남자 a 여자
5. 35 엉치뼈에서 보이는 남녀 차이 a : 엉치뼈의 옆모습 b : 엉치뼈의 윗모습
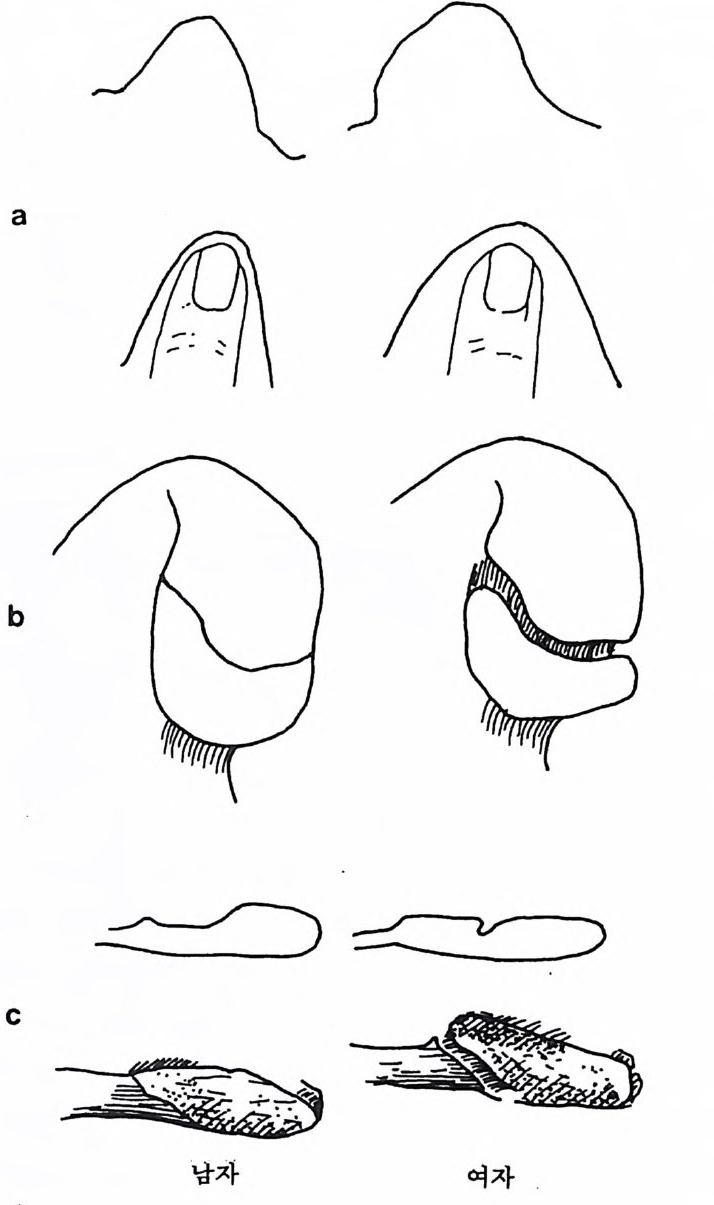 a <
a <
5. 36 엉덩뼈 a : 앉음뼈 큰활홈 Sc i a ti c notc h b : 엉치뼈 맞자리 Sacroil iac joi n t C : 엉덩뼈 홈 Preaur i cular sulcus
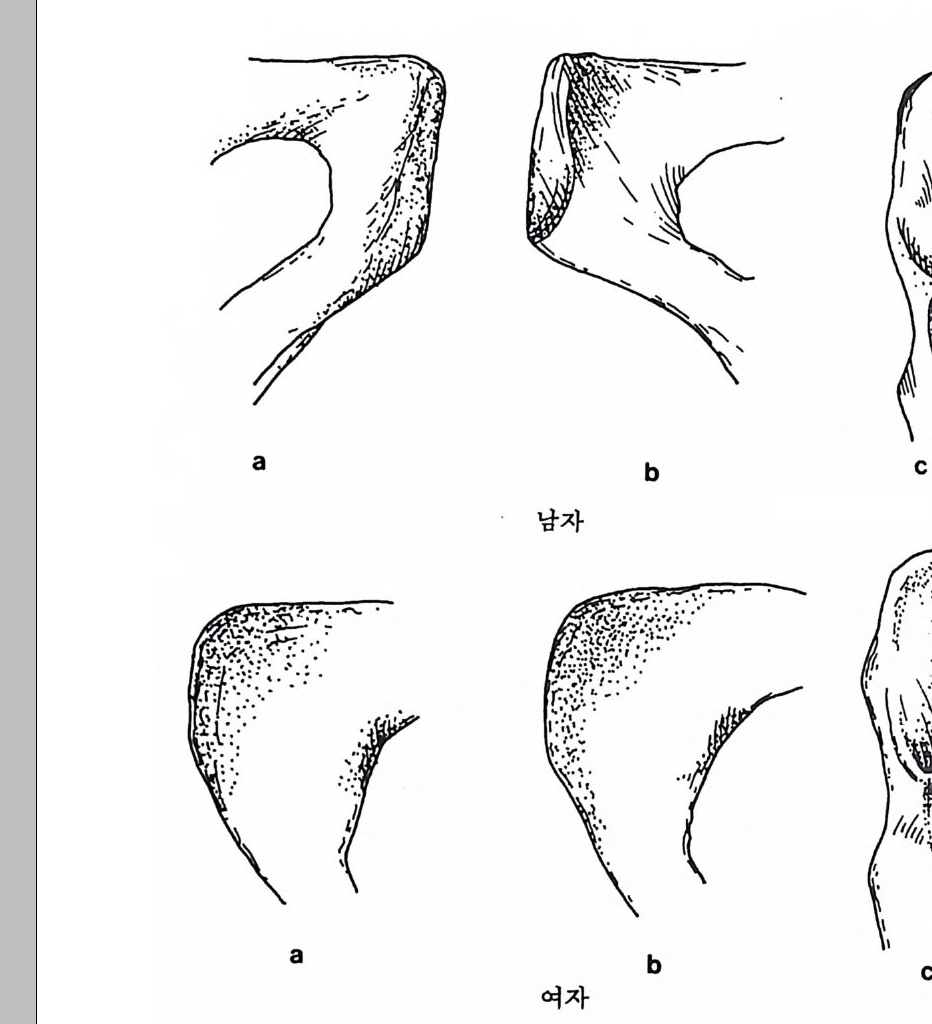 =..` ..
=..` ..
5. 37 감춤뼈 a : 감춤뼈 이음새 b : 감춤뼈 활 각 C : 앉음뼈 • 감춤뼈 가지의 옆모습 위의 세 특칭이 잘 관찰되지 않을 때는 각 표본의 가장 두드러지는 특칭에 기초를 두고 결정을 해야만 한다. 감춤뼈의 이음새 모양은 위 의 세 가지 기준 중 가장 중요한 것이다. 이의에도 남녀를 가늠하는
데 유용한 몇 개의 해부학상 특칭들이 있다. 며낮음뼈 큰활홈 Inc i sura Isch iad ic a Ma j or 이 여성은 넓고 남성은 좁다. 여기에 손가락을 넣었을 때 남성의 경우는 간신히 움직일 수 있으며 여성은 제법 움직임이 자유롭다. 성치뼈 • 받침뼈 닿는 면 Sacro- ili ac Ar ti cula ti on 이 여성에게서 두드러진다. 또한 앉음뼈 큰활홈과 엉치뼈 • 받침뼈 닿는 면 사이에 홈 Preaur i cular Sulcus 이 여성에게서 주로 발견된다. ·일반적으로 남성의 엉덩뼈가 굳세 보이고 근육 닿는 면이 두드러 전다. • 엉 덩 뼈구멍 Foramen Ob t ura t um 은 남성 이 더 크고 달걀형 이 며 , 여성은 작고 삼각형이다. • 여성의 골반은 분만을 해야 하기 때문에 골반통의 공간이 남성보 다더 넓다. • 엉덩뼈굼 Ace t abulum 은 남성 이 여성보다 더 크다. 나이 찾기 나이는 머리뼈 이음새, 이빨의 닳음새, 이빨 신경구멍 길이와 너비, 몸체뼈들의 붙음새(그립 5.3 8 ) 등을 종합해서 가늠해야 정확하게 찾 울 수 있다. 머리뼈에 이빨이 박혀 있는 경우 이빨이 솟아나는 정도 (그립 5. 39) 와 이빨 닳음새 정도가 좋은 기준이 된다. 사람의 이빨온 나이에 따라 나온 정도가 다르다. 태어날 때 잇몸에 이빨이 형성되어, 6 개월 내지 9 개월이 되면 잇몸 밖으로 이빨이 솟아 나기 시작하여 3 세까지는 어린니가 모두 나온다. 어 1 때 첫째어금니 가 나오며 어린니들을 갈기 시작한다. 12 세 정도에는 둘째어금니가, 18 세 내지 2M] 가 되면 셋째어금니가 나오는데 이렇게 어금니가 나오 는 정도를 바탕으로 발굴된 화석 인골의 나이를 추정할 수 있다.
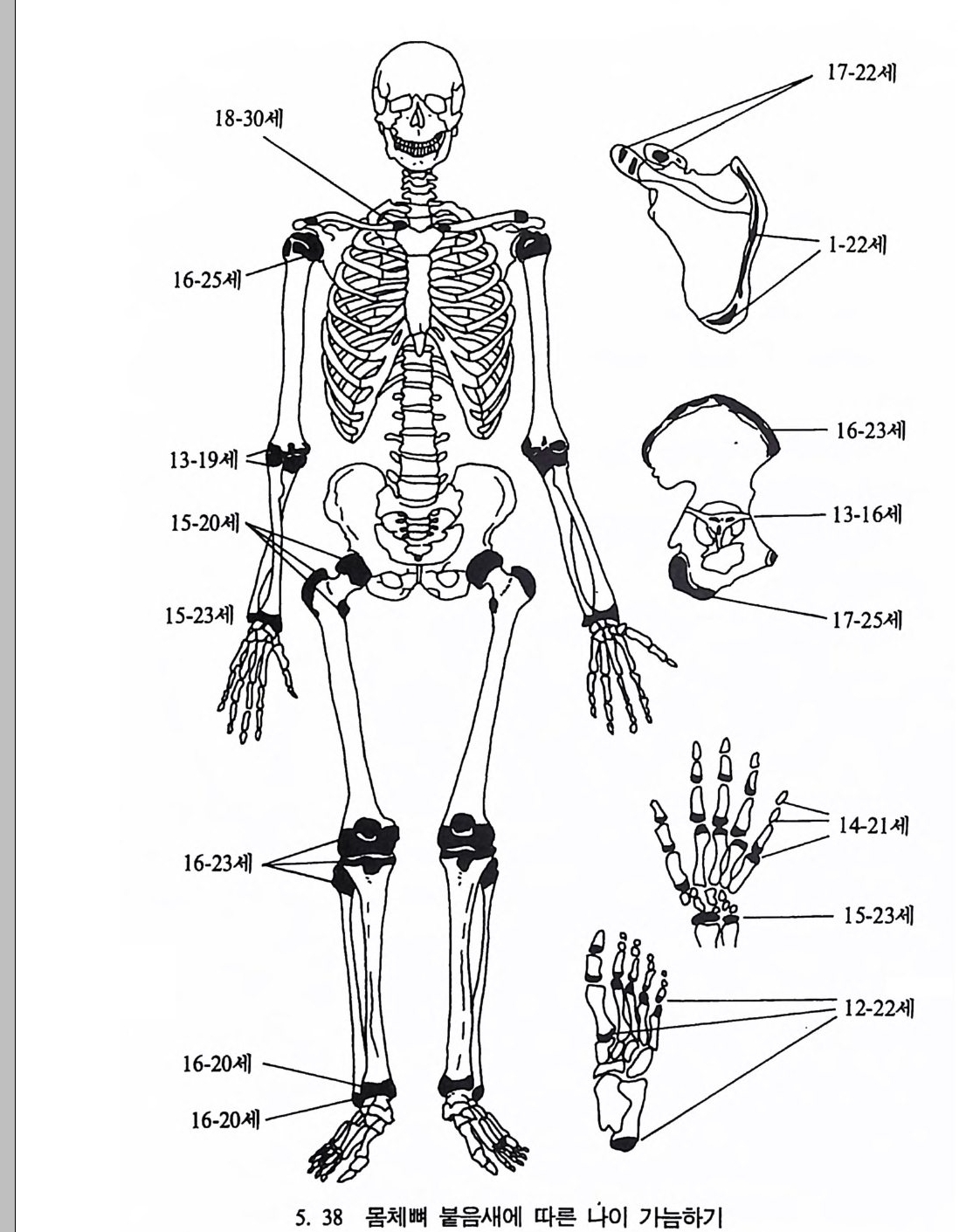 l7 - 22 세
l7 - 22 세
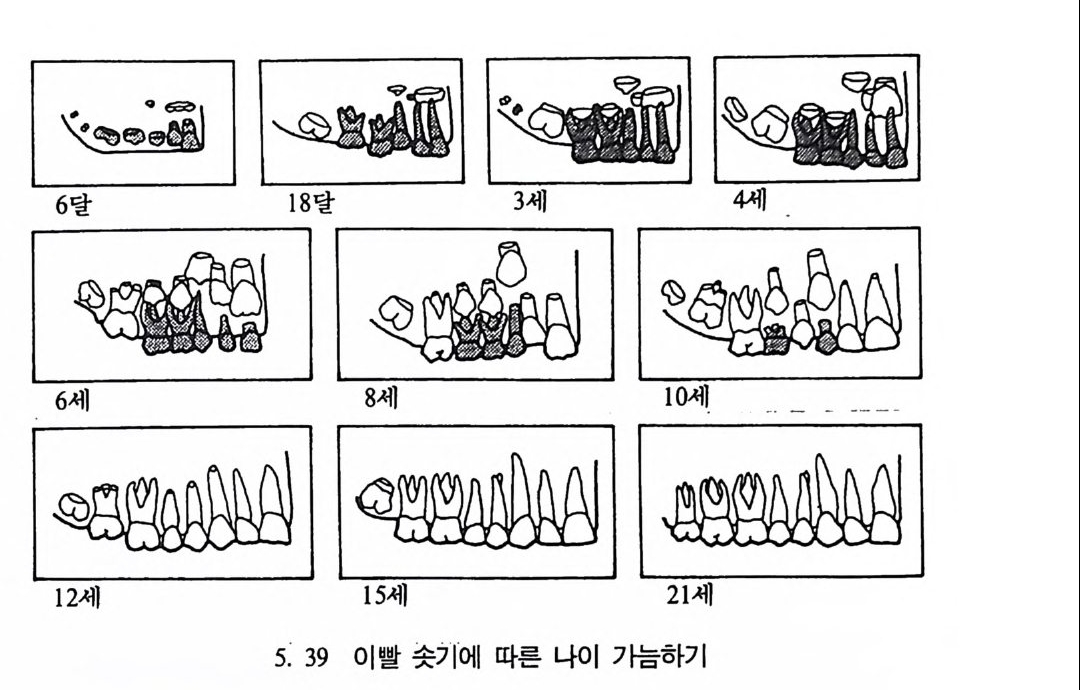 三6 세 一:8 세 一:<10 세一
三6 세 一:8 세 一:<10 세一
그러나 우리가 사람의 뼈를 통해 나이를 가늠할 때 위에서 언급한 기준들은 개인들과 집단들에게 똑같이 적용되지는 않는다는 점을 염 두에 두어야한다. 키 재기 사람의 키는 긴뼈의 잰 값을 바탕으로 헤아려내는데 이에 관한 많 은 연구들이 있다. 트로터와 글래서 Trott er & Glesser 의 연구가 주 로 이용되는데 이들의 방법은 긴뼈에 물렁뼈가 모두 붙어 있을 경우 에만 믿을 만한 값이 얻어지며, 특히 동북아시아 주민의 키를 가늠·할 때 자주 사용된다(표 5. 1 ). 물렁뼈들이 없을 때는 스듀어트 (S t ewar t, 1948) 와 울리버 (Ol ive r, 1969) 의 방법이 알맞다.
표 5. I 트로터와 글래서의 키재기 공식 (Trott er & Glesser, 1958) 동북아시아 갈래 l. 22X( 허벅지뼈 +종아리뼈) + 70.2 4 土 3.1 8 l. 22X( 허벅지뼈 +정강뼈) + 70.3 0 士 3.2 4 2 .4 0X( 종아리뼈) + 80.56 土 3.24 2 .3 9X( 정강뼈) + 81 .45 士 3.27 2.I SX (허벅지뼈 ) + 72.5 7 土 3.8 0 l.68X( 위팔뼈+뒤팔뼈) + 71 .18 士 4.1 4 l.67X (위팔뼈+앞팔뼈) + 74.83 土 4.l 6 2.68X( 위팔뼈) + 83.1 9 土 4.25 3 . 54X( 앞팔뼈) + 82.00 土 4.60 3 .4 8X( 뒤팔뼈) + 77.4 5 土 4.6 6 흑인종 l. 20X( 허벅지뼈 +종아리뼈) + 67.77 土 3.6 3 l.1 5X( 허벅지뼈+정강뼈) + 71 .75 土 3.6 8 2.IOX( 허벅지뼈) + 72.2 2 士 3.9 1 2 .1 9X( 정강뼈) + 80.56 土 3.96 2.3 4 X (종아리뼈 ) + 72.57 土 4.0 2 l.66 X (위팔뼈+앞팔뼈 ) + 73.08 土 4.1 8 l.65 X (위팔뼈+뒤팔뼈 ) + 70.67 士 4.2 3 2 . 88X( 위팔뼈) + 75.4 8 士 4.2 3 3 .3 2X( 앞팔뼈) + 85.43 土 4.57 3 . 20X( 뒤팔뼈) + 82.77 士 4.74 백인종 1.3 1X( 허벅지뼈 +종아리뼈) + 63.0 5 土 3.66 1. 26X( 허벅지뼈 +정강이뼈) + 67.09 土 3.74
2.60X (종아리뼈) + 75.50 土 3.86 2.32X (허벅지뼈) + 65.5 3 士 3.94 2.4 2 X (정강뼈) + 81 .93 士 4.00 l.82 X (위팔뼈 + 앞팔뼈 ) + 67.97 土 4.31 l . 78X( 위팔뼈 + 뒤팔뼈) + 66.98 士 4.37 2.89X( 위팔뼈) + 78.10 土 4.57 3.79X( 앞팔뼈) + 79.4 2 土 4.66 3.76X( 뒤팔뼈) + 75.5 5 土 4.72
이빨재기 사람 이 빨의 얼개는 그림 5. 41 과 같다. 사람은 2. 1. 2. 3/2. 1. 2. 3( 앞니 2, 송곳니 1, 옆니 2, 어금니 3 을 뜻함)의 여러 가지 이빨 He t erodon t을 지니고 있다. 어른의 경우 모두 327~ 의 이빨울 지니고 있 는데, 유인원도 사람과 같은 이빨틀을- 지니고 있다(그립 5. 40).
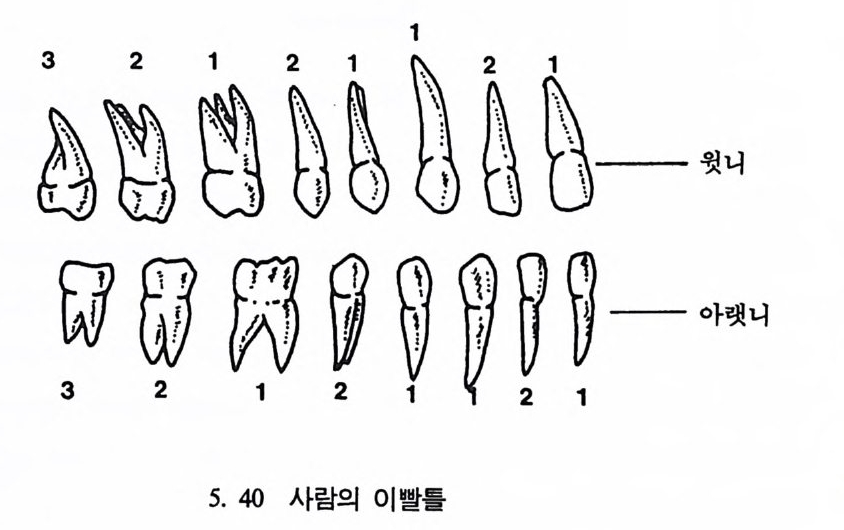 3\a》g凶3~ 2鬪난四 WW2 胃鬪1 2A料U 2 m내C1RN以니〉 HW\1emwV口 wue '멉2AHN묘3 1\11 갭|2— 아윗 —니랫 니
3\a》g凶3~ 2鬪난四 WW2 胃鬪1 2A料U 2 m내C1RN以니〉 HW\1emwV口 wue '멉2AHN묘3 1\11 갭|2— 아윗 —니랫 니
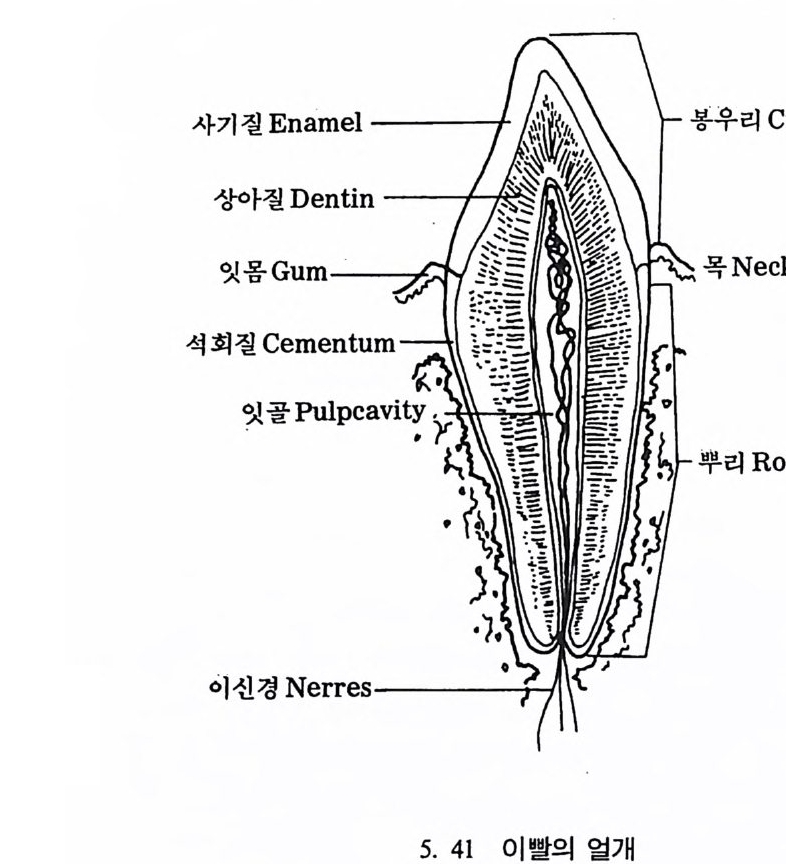 봉우리 Crown
봉우리 Crown
어금니의 경우, 도드리는 기본적으로 젖먹이동물과 같은 모양을 지 니고 있으나, 윗어금니는 Proto cone 뒤에 H yp ocone 이 발달해 있으 며, 아래어금니는 첫째어금니가 대표적인 모양을 하고 있는데 여기서 는 Trig o n id 부분에 Paraco ni d 가 없다(그립 5. 5 참조). 일반으로 사람의 이빨은 높이와 길이, 너비 그리고 전체 크기를 잰 다(그립 5. 42). 그러나 연구의 대상이 되는 옛 유적지에서 출토되는 이빨의 경우, 이빨이 심하게 닳아 높이를 정확히 잴 수 없기 때문에 이빨과 뿌리의 경계선에서 현재 남아 있는 씹는 면까지의 높이로 대 신한다.
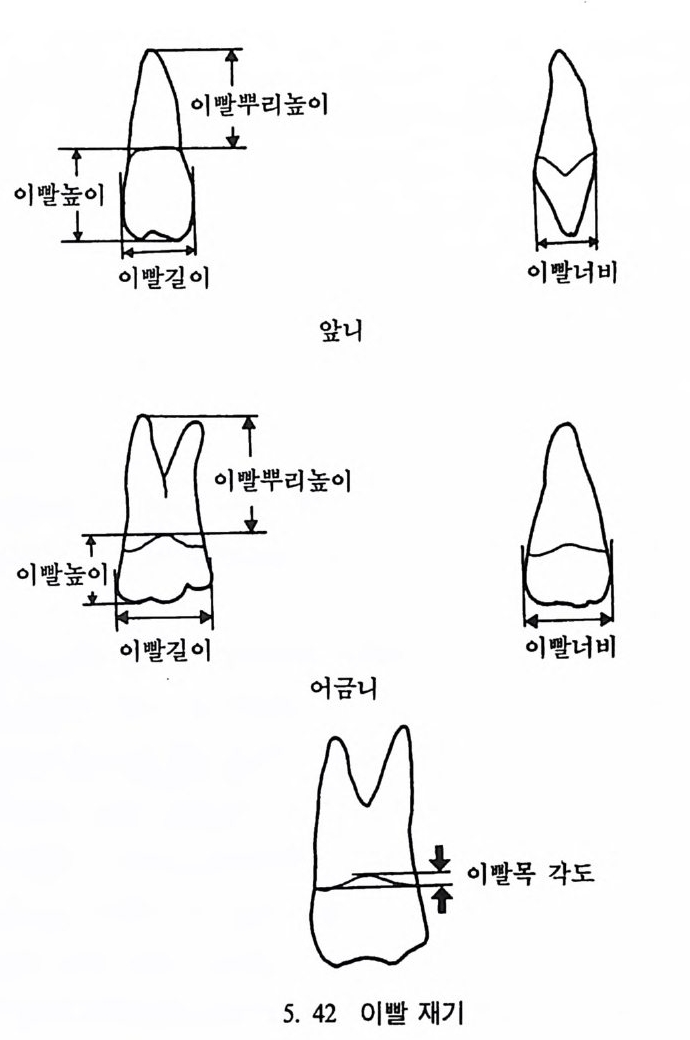 이빨길이 이빨너비
이빨길이 이빨너비
1) 이빨길이 Mesio d ist a l dia m ete r : 이빨길이는 때때로 이빨너비, 이빨폭이란 뜻으로도 사용된다. 이빨 앞 • 뒤의 다론 이빨과 맞닿 는 점 사이의 길이를 말한다. 2) 이빨너비 Buccolig ua l diam ete r : 1) 번과 수직되게 입술-혀 방 향의 길이 3) 이빨의 전체 크기 Crown module : 이빨길이 + 이빨너비 /2
이빨의 재기와 더불어 이빨의 형태들은 현재 사람의 집단에서의 차 이를 보여주며 또한 그들의 조상과의 관계를 알려줄 수 있다는 점에 서 매우 중요하다. 이빨의 형태는 크게 이빨의 수효와 자리에 따른 다양성과 이빨 형태의 다양성으로 나누어 설명되는데, 후자가 집단의 차이와 나아가 조상과의 관계를 알려주는 좋은 근거를 제공한다• 다양한 이빨 형태 이빨의 수효와 자리에 따른 다양성은 이빨이 여분으로 더 있는 경 우 Su p ernumerar y Teeth , 본래부터 이빨이 없는 경우 Con ge nit al Ab-sence, 이빨의 방향이 뒤틀려 있는 경우 Ro t a ti on, 그리고 이빨이 몰 려 있는 경우 Crow di n g로 나누어진다. 위의 네 가지 경우 중, 특히 이빨이 없을 경우, 아직 나지 않은 이 빨과 죽기 전에 잃어버린 이빨 그리고 선천성으로 나지 않는 이빨을 혼동하지 말아야 한다. 아직 나지 않은 이빨은 방사선촬영에서 쉽게 찾아지나 원래 있었으나 잃어버린 이빨의 경우에는 많은 문제가 있 다. 물론 죽은 후에 잃어버린 이빨은 이빨뿌리의 구멍이 잇몸부분에 서 나타나기 때문에 문제가 없으나, 살아 있을 때 이빨이 빠지면 이 빨뿌리 구멍은 잇몸에 다시 흡수되어 어떤 표시도 남아 있지 않게 된 다. 그러나 죽기 전에 이빨이 빠졌다면 잇몸이 뒤틀려 불룩해전 특칭 이 남는다. 또한 빠진 이빨이 다론 이빨과 맞닿아 있었다면 닿은 면 의 흔적이 인접한 이빨에서 찾아전다. 어금니 도드리 형태 어금니의 씹는 면에 있는 도드리와 홈의 형태는 인류집단과 그들의 조상에서 차이를 나타낸다. 위 • 아래 어금니는 각기 다른 도드리 형
태를 지니고 있다. 위어금니는 홈에 의해 분리된 3 - 4 개의 도드리를 가지고 있다. 첫 번째 위어금니는 같은 크기의 4 개 도드리가 있다. 두번째 어금니는 네번째 도드리인 H yp ocone 의 크기가 작아진다. 세번째 어금니는 H yp ocone 이 완전히 사라지거나 뒤쪽에 흔적 Cus p ule 이 남아 있다. 이 러한 도드리의 형태 분류는 주로 달베르그 (A.A.Dahlber g, 1951) 의 연구에 따른다(:그립 5. 43):. :: 4 4- 3+ 3 5. 43 위 어금니 도드리 유형 군군白田 Y-5 +5 v-4 +4 5. 44 아래 어금니 도드리 유형 정상의 아래어금니는 4 - 5 개의 도드리를 지니고 있다. 아래어금니 도드리는
은빈도율로나타난다. 곁도드리 결도드리는 위턱과 아래턱의 어금니에서 찾아지는데, 캐러벨리 도 드리 Carabell i's Cus p s 와 立 쿠 뚜人타일리드 Pro t os tyli d 가 가장 대표 되는 견도드리이다(그립 5. 45a , 그립 5. 45b).
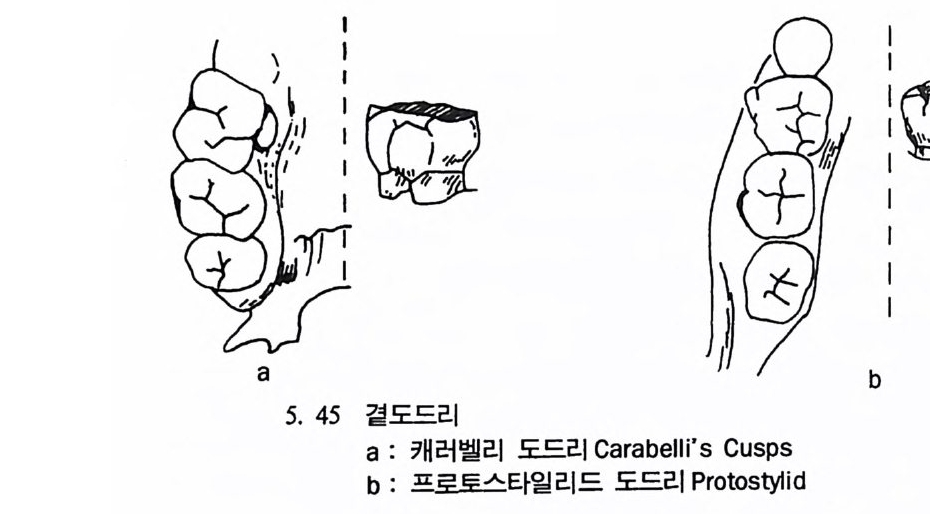 I덥i :
I덥i :
캐러벨리 도드리는 위어금니의 혀쪽 앞부분에서 찾아진다 .. 이 도 드리는 잘 발달된 도드리 형태로부터 흔적에 이르기까지 구분의 정도 가 다양하기 때문에 주로 크라우스 (Kraus, 1959) 의 방법을 사용한다. 이 결도드리는 현대 모든 집단에서 다양한 빈도로 나타난다. 立쿠두 아래어금니의 바깥면 앞쪽에서 나타난다. 이 결도드리는 오스트랄로피테쿠스와 아시아 곧선사람에게서 나타나며, 현대인의 경우는 피마 인디언을 제의하고는 거의 나타나지 않는다. 조사할 때는 이 도드리가 있는지, 없는지만을 기록한다.
부삽 모양 이 빨 Shovel-Shap ed Teet h 이빨에서 가장 빈번히 논의되는 유전상 특칭 중의 하나는 앞니의 씹는 면이 부삽 모양인지 아닌지에 있다. 엘드리카 (Hrdl i cka , I907) 에 의해 최초로 언급된 이후로, 동북아시아 주민에서 상대로 높은 출현 빈도수를 보여준다. 부삽 모양은 씹는 면의 모양에 따라 몇 가지로 분류된다(그립 5. 46).
 납작골 안쪽부삽 二안팎부삽 5둥군걸 一-입혀술쪽쪽
납작골 안쪽부삽 二안팎부삽 5둥군걸 一-입혀술쪽쪽
못 모양 이 빨 Peg -S hap ed Teet h 이빨이 비정상으로 작고 못대가리처럼 생겼다. 주로 셋째어금니와 바깥쪽 앞니 자리에 나타난다. 이런 형태의 이빨은 모든 현대 주민집 단에서 보고되고 있는데 선천성 결함으로 여겨진다. 타우로돈티즘 Taurodonti sm 어금니 속의 빈 공간 Pul p Cav ity이 커지고 이 뿌리가 줄어드는 것을 말한다. 이 특칭은 네안데르탈사람과 다른 인류화석에게서 자주 보이며 몇몇 현대 주민집단에서도 발견된다 . 타우로돈티즘은 정도에 따라 Cy no dont, Hy po ta urodont, Mesot a urodont, Hy pe rta urodont 로 구분된다 (Shaw, 1928) (그립 5. 47). 사기질 너더리와 혹 Enamel Exten sio n and Pearls 어금니와 옆니에서 사기질의 너더리가 뿌리 사이에서 발견되는데, 이런 너더리는 때때로 작은 덩어리(혹)로 나타나기도 한다. 이 특칭
은 많은 인류집단에서 발견되지만 전화상에서 얼마나 나타나는지는 알 수 없다(그립 5. 48).
 정상 타우로돈트
정상 타우로돈트
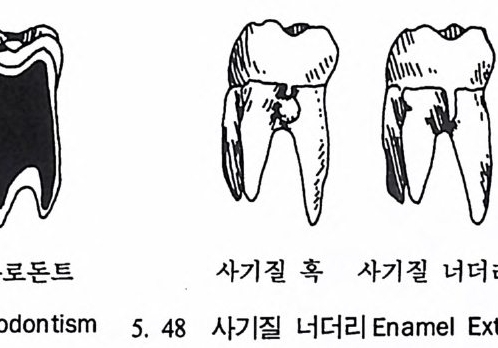 사기질 혹 사기질 너더리
사기질 혹 사기질 너더리
위 • 아래 이 빨 맞닿음새 Occlusio n 윗니와 아랫니가 맞닿는 모양은 인종을 구분하는 데 도움이 된다. 동북아시아 주민은 바로 맞닿으며, 백인종은 약간 윗니가 아랫니를 덮는다 . 이와 반대되는 모양은 모든 집단에서 잘 나타나지 않는다(그 림 5. 49).
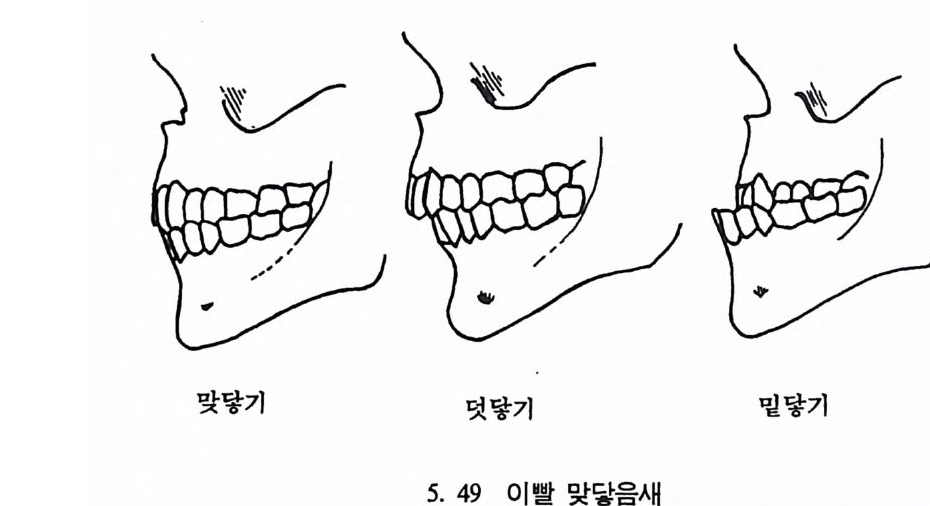 `` `
`` `
인종나누기 인종 분류는 린네가 1758 년 처음 시작한 이래 많은 학자들에 의해 거론되었다(표 5. 2). 오늘날 일반적으로 이용되는 피부색 등을 바탕 으로 분류된 백인종, 흑인종, 동북아시아 갈래(황인종) 분류는 후돈 (E. A. Hoot o n, 1926) 에 의해 제시되었다. 1960 년대에 인종 분 류 를 다 시 정 리한 쿤 (Carle t on S. Coon, 1962) 은 게이츠(R. R. Gate s , 1948) 의 연구를 밑받침으로 백인종, 흑인종, 동북아시아 갈래(황인종) 이의 에 오스프녁誌 1 이드와 카포이드를 더하고 있다. 그러나 현대 체질인류학에서는 인종을 분류하는 것에 별 의미를 두 지 않고 인종이란 단지 지역적인 차이만을 나타낼 뿐이라고 말한다. 이들은 인류의 변이를 등고선분포 C li nal D i s tr i bu ti on 라는 방법을 통 해 체질상의 특징이 어떻게 지역적으로 나타나는지를 연구한다. 인류 생물학자들은 인류집단 사이의 차이보다는 집단 안에서의 차이에 연 구를 집중하고 있다. 한 예로 하버드대학교의 집단유전학자인 레원론 은 대부분의 인류변이는 같은 무리 안에서의 차이로 설명되어야 하며 지역적으로 떨어전 집단 사이의 차이로 이해될 수 없다고 주장한다. (R. D. Lewonto n , 1972). . 많은 학자들이 인종 분류에 회의를 보이고 있지만 아직도 머리뼈를 바탕으로 인종을 규명하는 노력은 계속되고 있다. 머리뼈를 통해 인 종을 구분하기 위해서는 1) 머리뼈가 지닌 형태상 • 해부학상의 특칭 들, 2) 머리뼈의 잰 값에 연구의 초점을 맞춘다. 인종에 따른 형태상 해부학상의 차이는 그 동안많은 학자들에 의해 거론되었는데 여기서 는 바스 (W.M.Bass, 1987) 가 정리한 연구를 바탕으로 머리뼈의 앞모 습과 옆모습을 통해 크게 백인종, 혹인종, 동북아시아 갈래(황인종) 차이를 살펴보기로 한다.
백인종(그림 5. 50) 콧굼 둔덕 Nasal sill 콧굼 둔덕은 콧굼 입구에 있는데 콧굼 입구와 위턱 사이에 연필을 놓으면 연필은 콧굼 입구 쪽으로 서서히 구르다 가 콧굼 입구에 있는 둔덕에 걸려 멈춘다. 백인종은 이 둔덕이 잘 발 달해 있는 데 반해 혹인종은 이 둔덕이 없어 연필이 콧굼 가운데로 굴러든다. 동북아시아 갈래는 중간 정도를 보인다.
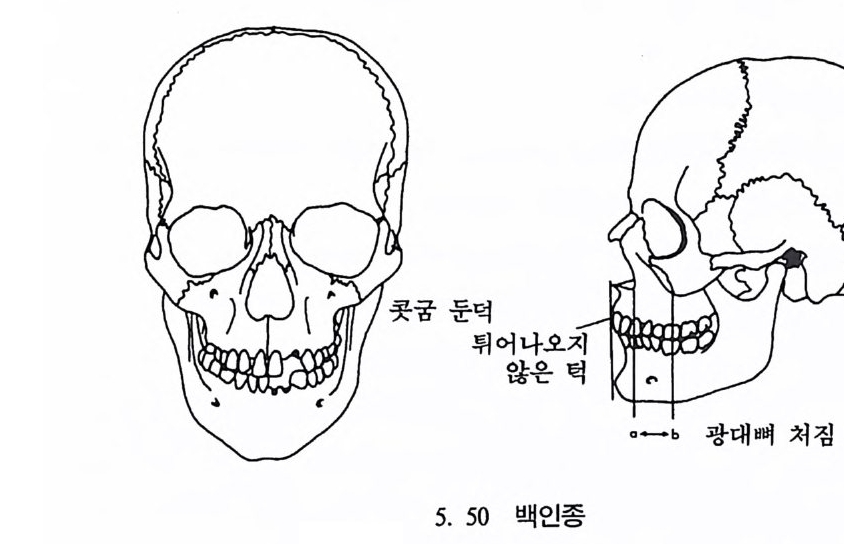 콧굼둔덕
콧굼둔덕
광대뼈 저짐 머리뼈의 얼굴이 위로 향하게 뒷머리뼈를 두 손으로 잡고 연필을 콧굼 입구에 놓은 다음 검지손가락을 광대뼈와 연필 사 이로 밀어 넣는다. 백인종의 경우 얼굴 가운선이 앞으로 나와 있고 광대뼈가 앞으로 나와 있지 않기 때문에 손가락을 넣었을 때 연필은 떨어지지 않고 제자리에 있다. 동북아시아 갈래는 넓적한 얼굴에 광 대뼈가 발달해 손가락을 넣으면 연필이 굴러 떨어전다. 이의에 백인종의 턱은 수직에 가까운 Or t ho g na t hous 모습을 지니 고 있는 데 반해 흑인종은 턱이 앞으로 나와 Pro gn a t hous 있다. 백인 종은 얼굴이 길고 좁으며, 콧굼 또한 좁다. 코맞점 부위가 꺼져 있으
며,코가오뚝하다. 흑인종(그림 5. 51 ) 흑인종은 코밑에 얕은 홈 Nasal Gu tt er i n g이 나 있으나 백인종에서 보이는 콧굼 둔덕이 없다. 흑인종의 위턱과 아래턱이 앞으로 많이 튀 어나와 있어 연필을 콧굼 입구에 놓으면 연필은 항상 콧굼 쪽으로 굴 러간다. 이의에 흑인종 머리뼈 특칭으로는 코맞점 부위가 편평하고, 앞이마 는 둥글며, 정수리점 부위가 약간 가라앉은 점 등을 둘 수 있다. 콧굼 은 넓적하며 뼈조직이 치밀하다.
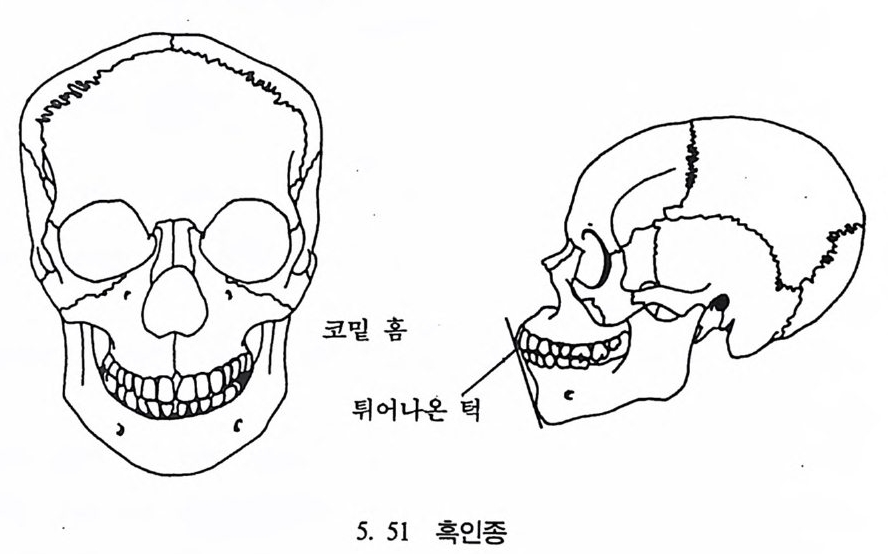 5. 51 호-, 이L..조C
5. 51 호-, 이L..조C
동북아시아 갈래(황인종)(그림 5. 52) 튀어나온 광대뼈 동북아시아 갈래의 머리뼈는 넓적하고 둥근 얼굴 을 지니고 있다. 이런 특칭은 광대뼈가 앞을 향해 발달해 있기 때문 이다.
위 • 아래 이빨 맞닿음새 동북아시아 갈래는 입을 다물었을 때 백인 종이나 흑인종과는 다르게 위 • 아래 앞니가 맞닿는다.
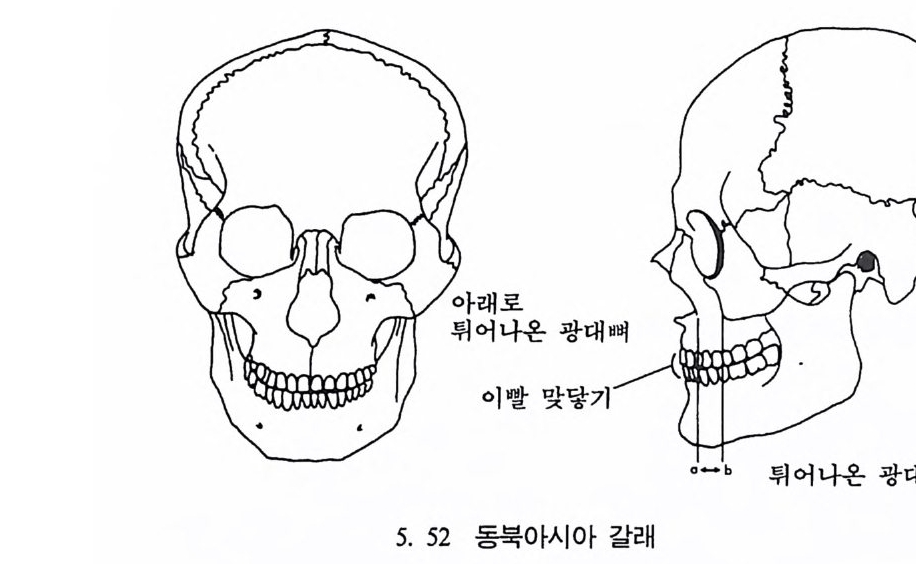 아래로
아래로
부삽 모양 앞니 동복-아시아 갈래 중 많은 사람의 경우 위앞니의 혀 쪽에 사기질이 발달해 부삽 모양처럼 보인다. 이의에 광대뼈의 아래 부분이 앞으로 향해 있고, 코뼈는 위턱보다 앞으로튀어나와 있다. 위에서 언급한 형태상 • 해부학상 특칭에 따론 인종 나누기와 함께 머리뼈 챈 값을 이용해서 인종을 나누는 방법은 여러 학자들에 의해 제시되었다 (G il es and El liot , 1962). 특히 길 (G ill, 1984) 은 머리뼈 재 기 중 세 개의 지수를 이용해서 북미대륙의 인디언을 구분하였는데, 90% 이상의 정확도를 보여주고 있다.
표 5. I 인종의 분류 Linn aeus, 1758 Blumenbach, 1781 Homo europ ae us Caucasia n Homo asia t i cu s Mong ol ia n Homo afer Malay Homo americ a nus Eth iop i an Americ a n E. A. Hoote n , 1926 Prim ary Prim ary Comp o sit e Comp o sit e Resid u al Race Subrace Race Subrace Mi xe d Type s Wh ite Medit err anean Austr a lia n Armenoid Nordi c- A lpi ne Ainu Indo-Dravid ian Di na ric Nordic - Kelti c Polyn e sia n Med iter ranean Nordic Alpi ne East Baltic Negr o id Afr ica n Neg ro Busrunan-H ott en to t Ni lo ti c Neg ro Neg ri t o Mong ol oid Classic Indonesia n - Mong ol oid Mong ol oid Arcti c Americ n a Ind ian Mong ol oid Secondary Subrace Malay - MIndoonnge osl oiia dn
Sta n ley M. Garn, l965 Geog ra ph ic a l Races : Amerin d ia n Melanesia n -P ap u an India n Polyn e sia n Austr a lian Europ ea n Mi cr onesia n Asia t i c Af rica n Local Races : There are examp le s of local races ; the re are many , many more. North w est Europ e an East Af rica n North Chrnese North e ast Europ e an Bantu Ex tre me Mong ol oid Alpi ne Tibe ta n Hi nd u Med iter ranean Mi cr o-R aces : Not well defi ne d bur app a rent ly refe r s to neig h -borhoods wi thin a cit y or cit y itsel f . (Nelson, et al. , 1992, p. 74 에서 인용)
 제 6 장 인류 조상의 발자취
제 6 장 인류 조상의 발자취
제 6 장 인류 조상의 발자취 이른시기의 유인원들 인류의 기원과 진화의 과정에 대해서는 많은 논쟁이 있는데, 그 논 쟁의 주요 쟁점은 인류가 진화를 했는지 아닌지를 규명하는 데 있는 것이 아니라 진화는 언제, 어디서, 어떻게 일어났느냐에 대한 학자들 의 각기 다른 설명과 해석에 있다 . 왜냐하면 고인류학자들은 부서진 뼈화석 조각들을 통해 자신들의 이론을 증명하고자 하기 때문에, 학 자들 사이에 서로 다른 해석이 나올 수 있으며 이에 관한 논쟁은 매 우 주관성을 지닐 수가 있다. 인류의 기원과 전화에 관한 여러 이론들을 이해하기 위해서는 먼저 고동물학상의 증거인 뼈화석에 초점을 맞추어 역사상의 기록들을 살 펴보아야 하며, 디론· 한편으로 오늘날 살아 있는 영장류 중 사람과 가까운 관계에 있는 영장류에 대한 연구는 이러한 여러 이론들을 이 해히는 데 있어서 중요한 방법 중의 하나이다. 인류의 기원과 전화에 관한 이론에 대해서는 많은 논쟁이 있는데, 이는 다음과 같은 이유에서 시작되었다고 볼 수 있다 ; 첫째, 지금까지 발견되는 화석기록이 단편적일 경우가 많기 때문에 이를 바탕으로 한 학자들 사이의 해석이 다를 수 있다. 그러나 이런
문재는 화석 증거가 보충됨으로써 올바른 해석이 이루어질 수 있다. 둘째, 고동물학자들은 때때로 그들이 연구하고자 하는 조상들의 후 손인 현재 살아 있는 동물들의 연구를 소홀히 한다. 그러나 현재 살 아 있는 후손들은 그들의 유전자 안에 전화의 정보들을 간직하고 있 으며, 해부학상 정보 또한 무시되어서는 안 된다 . 셋째, 많은 학자들은 개인 나름의 편견을 가질 수 있다. 그들은 자 신들이 가진 편견에서 벗어나고자 애를 쓰지만 때로는 이룰 극복하지 못하고 이런 편견들이 화석기록들을 설명하는 데 영향을 미친다.
 6. l 이집토피테쿠스의 복원 상상도
6. l 이집토피테쿠스의 복원 상상도
앞장에서 영장류의 전화를 말할 때 이집토피데쿠스가 발견된 아프 리카 대륙의 초기 올리고세 엘 화융 유적에 대해 언급하였다. 당시 그 지역은 삼림이 번성했던 지역으로 다양한 종류의 영장류가 살았는 데, 그 중 어떤 무리는 · 신대륙 원숭이의 조상일 가능성이 있고, 다른 · 종류의 화석은 당시에 이미 옆니가 없어져 우리와 닮은 이빨틀과 형
태를 가지고 있었다. 울리고세 당시에 유인원과 닮은 존재의 여부는 우리가 어떻게 유인 원을 정의하는가에 달려 있는데, 이집토피데쿠스의 적응도는 원숭이 와 같지만 오늘날 살고 있는 어떤 원숭이 종류와도 같지 않으므로 오 히려 유인원의 조상일 가능성이 크다(사전 6. l). 팔레오세와 이오세 중에 영장류가 어떻게 번성했는지는 앞에서 살 펴보았다. 여기서는 특별히 구대륙 원숭이와 침팬지, 고릴라, 오랑우 탄 및 하이로베이트 H y loba t es 같은 유인원과 사람에 관해 살펴보기 로 한다. 왜냐하면 이집토피데쿠스는 위에 언급한 영장류들의 조상일 가능성이 아주 높기 때문이다. 구대륙 원숭이는 이집토피데쿠스로부터 갈라져 나간, 마이오세 중 기에 살았던 빅토리아피테쿠스 V i c t or i ap it hecus 로부터 진화되었다고 알 려져 있고, 오늘날 살고 있는 유인원과 인류는 이집토피테쿠스로부터 갈라져 나간 다른 형태로부터 기원하였다고 알려져 있다. 영장류의 전화는 올리고세 중에 살던 영장류 화석들이 많이 발견되 지 않아 그 과정을 잘 알 수가 없다 . 그러나 마이오세에 오면 영장류 가 번성하여 많은 화석들이 남아 있어 인류와 다론 정도를 가늠할 수 있다. 드리 오피 데쿠스 Dry op ithe cus(= 프로컨슬 Proconsul) 아프리카의 케냐에 있는 빅토리아 호수 Lake V i c t or i a 는 아프리카 대협곡 Ri ft Valley 중 동쪽 협곡 Eas t ern Valle y과 서쪽 협곡 Wes t em Vally 사이에 위치하고 있다. 이 지역은 지질학상 매우 활동적인 지역이며 이 활동에 의해서 많은 동물화석들이 표면에 노출되었다. 마이오세 중 이 일대는 호수들과 홍수 퇴적지대, 화산 퇴적지대, 그리 고 산맥들이 발달해 있었다.
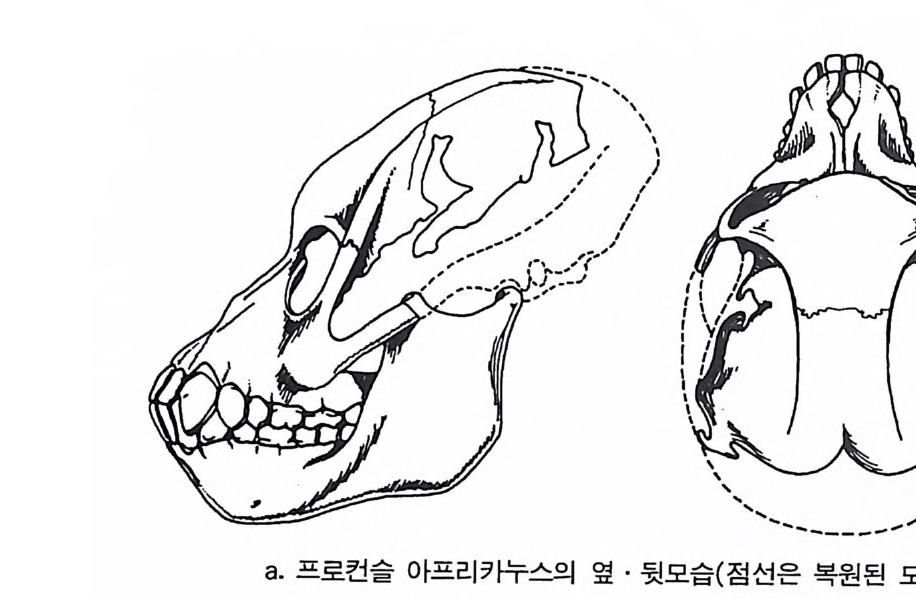 `` ``\\ `.\,/'/' ,, '
`` ``\\ `.\,/'/' ,, '
 ~델- 글•' ' J 。 討,, c.` ·:、 1
~델- 글•' ' J 。 討,, c.` ·:、 1
1940 년대에 루이스 리키 Louis Leake y는 이 지역에서 가장 오래 된 유인원의 조상에 속하는 화석을 발견하여 프로컨술이라고 이름 지 었다. 처음에 루이스 리키에 의해 턱조각들이 발견된 이후 많은 부분 들이 발견되어 지금은 하나의 완전한 골격을 이루고 있다. 프로컨술은 1856 년 이래로 유럽, 아시아 그리고 아프리카 등지에서 발견되어 드리오피테쿠스로 알려졌으나 별로 관심을 끌지 못하다가 루이스 리키가 아프리카 빅토리아 호수의 루성가 섬에서 발견한 화석 에 프로컨슬이란 이름을 붙이면서 다시 각광을 받게 되었다(사전 6.2). 프로컨술의 몸집은 오늘날 살고 있는 침팬지보다 작으며 유인원을 닮은 주둥이와 머리뼈를 가지고 있다. 크고 작은 턱뼈들이 함께 발견 되는 것으로 보아 여러 종의 프로컨슬이 있었던 것으로 여겨지며, 지 금까지는 3 종의 프로컨슬이 알려져 있다(표 6. I).
표 6. 1 지금까지 알려진 프로컨슬의 종류 프로컨슬 아프리카누스 Proconsul afr ic a nus 프로컨슬 니 안저 Proconsul ny an zae 프로컨슬 메이저 Proconsul majo r
프로컨슬의 사지뼈들 또한 많이 발견되었다. 프로컨슬은 밖으로 향 한 발가락 hallux 울 가지고 있으며 오늘날 살고 있는 유인원들에게서 보여지는 〈손마디 걷기〉 같은 특별한 형태의 걷기 증거는 볼 수 없고 아마도 네 발로 걸었을 것으로 여겨진다. 마이오세 초기중에 커다란 몸집을 한 영장류는 땅 위에서 살았으 며, 작은 몸집의 영장류는 나무 위에서 살았다. 이와 동시에 숲속에서 는 구대륙 영장류들이 살았다. 그러나 지금까지 발굴된 화석을 살펴 보면 유인원의 화석이 훨씬 더 많은 점으로 보아 마이오세 초기에는 유인원들이 구대륙 원숭이들보다 더 번성했음을 알 수 있다. 오늘날
은 이와 반대로 삼림이 바뀌어서 침팬지, 고릴라 등 대개의 호미노이 드 영장류를 제의한 구대륙 원숭이가 훨씬 더 번성하고 있다. 마이오세 중기가 되면 여러 가지 다른 형태의 영장류들이 나타난 다. 주로 유럽과 아시아에서는 드리오피데쿠스 D ry o pit hecus, 시바피테 쿠스 S i uap it hecus, 라마피 데쿠스 Ramap it hecus, 그리고 기 간토피 데쿠스 G ig an t op it hecus 와 갇은 형태들이 마이오세 중기 이후에 번성했으며, 이들 중 한 쌍이 당시 번성했던 지역 어느 곳에서 인류 조상으로서의 구실을 했을 것으로 가늠된다(그립 6. 3).
 6. 3 마이오세 영장류 화석의 분포(구대륙)
6. 3 마이오세 영장류 화석의 분포(구대륙)
마이오세 중기 이후에 진행된 지구의 변화는 당시에 번성했던 영장 류의 변화에 중요한 영향을 미쳤다. 먼저 유라시아 대륙을 살펴보면,
인도는 약 1000 만 년 전인 중기 마이오세에 동남아시아 대륙과 충돌 하기 전에는 〈섬〉이었다. 이 지각의 충돌로 히말라야 산맥이 형성되 었으며 그 결과 오늘날 동남아시아 지형이 이루어졌다. 이런 지형 변 화가 일어나자 새로 형성된 강물이 돌출된 산맥으로부터 홀러내렸는 데, 당시에 살았던 영장류들의 뼈가 이 산맥지대에 묻혀 있었다. 오늘 날 이 화석들이 다시 지표면에 나타났는데 파키스탄의 시와리크 언덕 Siw a lik Hi lls 등이 대표적 인 예이다. 시 바피 테쿠스 Siu a p ithe cus 시와리크 언덕에서 아래턱이 하나 발견되었는대 고릴라의 턱보다 작은 형태로 시바피테쿠스라고 불리었다. 그 후 197 01:i빈에 얼굴 부분 의 뼈가 발견되어 오늘날 살고 있는 침팬지, 오랑우탄의 얼굴들과 비 교되었다. 오랑우탄과 시바피데쿠스의 얼굴이 안쪽으로 파인 것에 반 해 침팬지의 얼굴은 편평하다. 또한 침펜지의 눈굼은 둥근형인데 시 바피데쿠스와 오랑우탄은 달걀형이다. 시바피데쿠스는 한때 라마피테 쿠스와 연관이 있는 것으로 알려지기도 했지만 오늘날에는 인도내시 아에서 살고 있는 오랑우탄의 조상으로 인정되고 있다. 기 간토피 데쿠스 Gig an to p i the cus 아시아는 호미노이드 영장류의 전화를 연구하기에 중요한 지역이 다. 최근에 중국 납부 유난성에서 작은 유인원의 부서전 얼굴과 턱뼈, 이빨들 그리고 사지뼈들이 석탄 속에 묻힌 채로 발견되었다. 이들 화 석뿐만 아니라 풀라이스토세에 살았던 기간토피데쿠스가 발견되었는 데 이들은 마이오세 중기에 인도에서 살았던 유인원 화석과 관련이 있는것으로 여겨진다.
중국의 전통 의약품으로 〈용골 Dra g on's Bone 〉이라는 뼈가 있는데, 이는 병을 치료하는 데 이용되었다. 이들은 실제로 여러 종류 젖먹이 동물의 뼈화석으로, 1935 년에 고생물학자인 퀘닉스발트 G. H. R. von Koe nig swald 는 홍콩의 거리에서 이 〈용골〉들을 파는 것을 보았다. 그는 한 유인원의 화석에서 나온 커다란 송곳니에 주목하였으며, 이 화석을 기간토피테쿠스라고 이름 지었다. 기간토피데쿠스는 오늘날에 살고 있는 어떤 유인원보다 상당히 큰 형태이나 그 이상 별로 알려진 것이 아직 없다. 중국 남부지방의 암반은 약한 산에도 잘 녹는 석회암지대로 이 석 회암은 산에 녹아 마치 탑과 같은 카르스트 Kars t 지형을 형성하는 데, 이 경관이 중국 남부지방의 대표되는 지형이다. 이 지역에 탑처럼 생긴 지형에는 많은 굴들이 형성되어 있어, 1950-1960 년에 걸쳐 이 중 한 굴에서 기간토피데쿠스의 이빨둘과 턱뼈 3 개가 발견되었다. 턱 들의 표면에는 포큐파인 Porcu pi ne 이 갑아놓은 흔적이 남아 있었다. 발견된 턱뼈와 뼈둘, 그리고 이빨만을 바탕으로 하여 한때 기간토 피테쿠스는 사람의 조상으로 분류되었다. 그러나 오늘날 기간토피테 쿠스는 약 900 만 년 전부터 100 만 년 전까지 살았던 것으로 추정되 며 이미 인류의 조상인 호미니드와 함께 살았던 것으로 여겨진다. 따 라서 기간토피테쿠스는 인류의 조상이라고 할 수 없다. 라마피테쿠스 Ramap it hecus : 인류의 직접 조상인가? 마이오세 중기에 살았던 또 다른 형태로는 라마피테쿠스가 있다. 이들은 동아프리카의 빅토리아 호수와 터키, 그리고 표준화석이 발견 된 인도에 이르기까지 아프리카와 유라시아 대륙에 걸쳐 넓게 퍼져 살았다(표 6. 2) (사진 6. 4).
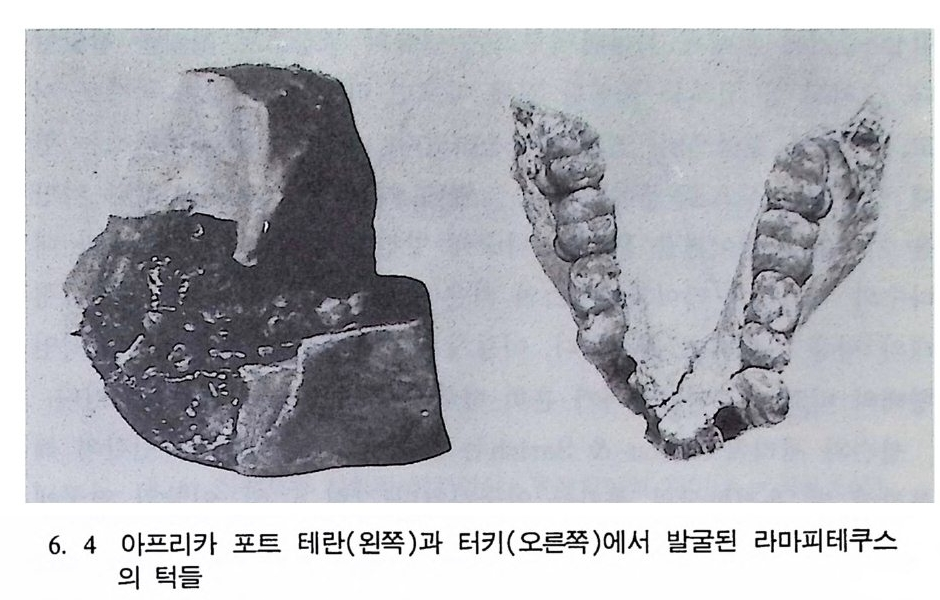 성'
성'
최초의 화석은 1932 년 루이스 G. E. Lew i s 에 의해 인도의 시와리크 언덕에서 발견되어 라마피테쿠스라고 불리었다. 루이스가 발견한 화 석은 가운데 부분이 일부 깨어져 나간 위턱으로, 그는 이 턱이 침팬 지 같은 유인원의 턱과는 다르게 아치형의 입천장을 가지고 있다고 생각하였다. 또한 라마피데쿠스의 송곳니는 침팬지가 지닌 송곳니 정 도의 크기이나 어금니는 매우 크다. 얼굴에는 뺨쪽 송곳니뿌리 끝 부위가 약간 움푹 팬 흔적인 송곳니파임 Ca nine Fossa 이 있다. 그러 나 침팬지의 얼굴에는 송곳니파임이 없다. 라마피테쿠스의 이빨에는 두터운 사기질이 있는데 이런 특칭은 현대 인류가 지니고 있는 이빨 의 특칭과 비슷하나 오늘날 살고 있는 유인원에서는 발견되지 않는 다. 1960 년대 사이몬스와 필빔 Sim ons & P ill beam 은 라마피테쿠스의 위덕을 복원하여 이를 인류와 오랑우탄의 턱들과 비교하였다. 그들의 복원에 따르면 라마피테쿠스의 턱은 현생인류의 턱과 너무도 닮았다 (그립 6. 5). 사이몬스와 필빔이 시도한 위턱의 복원에서 얻어전 해부
학상 중거에 의해서 라마피테쿠스는 인류의 조상으로 자리를 잡았었 다. 그리고 몸 전체를 복원한 것에 따르면 라마피테쿠스는 똑바로 서 고 두 발로 걸었으며, 연모를 사용하였다. 따라서 라마피테쿠스는 한 때 인류의 조상으로 분류되었다. 그런데 이 복원은 단지 이빨과 턱만 울 가지고 이루어졌을 뿐이다. 나중에 발견된 화석들의 연구 결과 대 다수의 학자들은 라마피테쿠스의 턱은 유인원도 사람의 턱도 아닌 형 태의 턱을 가졌다고 확신한다. 이렇게 현생인류와 화석과의 직접적인 형태의 비교는 고인류학자가 흔히 범할 수 있는 실수 중의 하나이다. 윌슨과 세리치 Wi lso n & Sar i ch 는 1960 년대에 침팬지와 인간의 해 부학상 및 유전학상의 분화를 연구하였다(그림 6. 6). 이들의 연구에 따르면 침팬지는 해부학상으로 우리의 공통조상으로부터 아주 많이 분화되지 않았으나 인류는 많이 떨어져 있다 . 그러나 유전학상으로 볼 때 침팬지와 인류는 똑갇은 정도로 분화되었다.
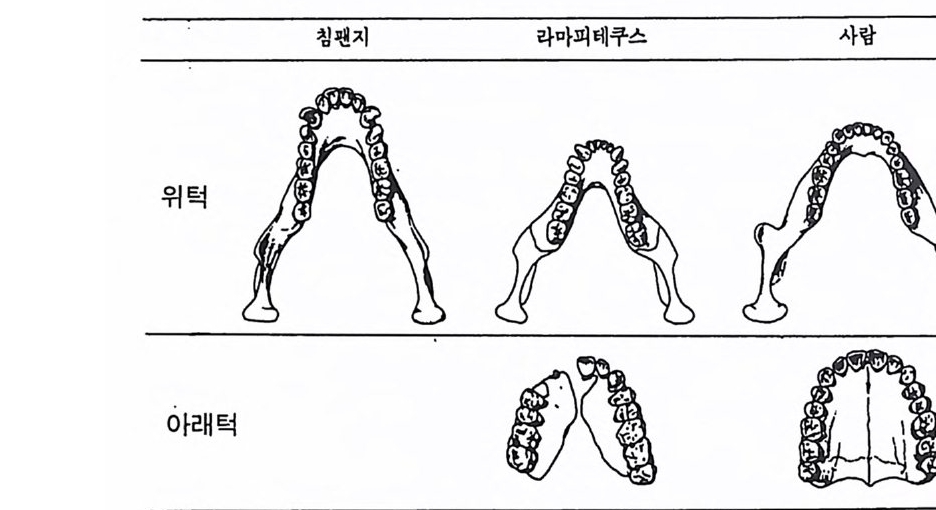 침팬지 라마피테쿠스 사람
침팬지 라마피테쿠스 사람
6. 5 침팬지, 라마피테쿠스, 사람의 턱 모양 비교 윗줄의 그립은 왼쪽부터 ~ 침팬지와 V 자형의 라마피테쿠스, 그 리고 사람의 아래턱이며 아랫줄 가운데 그림은 라마피테쿠스의 깨진 위턱으로 한때 사람의 턱과 같은 모양으로 복원되었다.
침팬지 사람 침팬지 사람 시入 l IIi
간 간 e사염=이한기d의편가+ e거다있 라리른으는 는면학 등침자9식8i,팬 들A4(이 개지 은공 가통와 성면조 립같상비역된)다비체다 와계(.98의 . 분 4거자%리)들. 와을이 같는이으 용놀며하랄 여신 만 대영한'II'I륙 장 i 二i〔일류원로들숭, 의 이왜 와유`냐, '전 비하 상해부학상 차이 유6. 6 침팬지와 사람의 해부학상 • 유전학상의 차이 (A =공통조상) 학상의 분화 거리를 측정하였다(그림 6. 7). 윌슨과 세리치는 인자에 는 규칙적인 변화의 비율이 있다는 것을 알았다. 따라서 사람과 비비 비 및 침팬지 또는 사람의 거리가 같다. 따라서 a+b+e=b+c+ 침팬지와 사람은 유전자 단계에서 아주 비슷하다. 평균 1000 개쌍의 면 침팬지와 사람 사이에는 신체와 행위상의 많은 차이가 있기 때문 이다. 형태상의 진화는 인자의 분자상 전화를 뜻하는데 인자는 동물들의 형태를 조절한다. 이때 형태에서 보이는 구조상의 차이는 모든 인자 중에 작은 부분인 조절유전자에 의해 일어난다. 인자의 다수는 복제 t ranscr ipti on 와 변형 t ransla ti on 중에 잘려져 나온 안마디로 이루어 져 있다. 그리고 이들은 개체에 영향을 주지 않고도 변할 수 있다.표 6. 2 라마피데쿠스 (Genus Ramap ithe cus) 화석이 발견된 유적지 유적지 화석 이름 연대(백만 년 전) 아시아 시와리크 언덕 , 라. 판자브쿠스 R. pa nja b cus 9-1 2 포트와 인도, 파키스탄 5 개의 아래턱과 2 개의 위턱 조각 (19 10-1 932) (19 73-1978) 아래턱을 포함한 43 개의 조각들 9- 1 2 칸디르, 터키 아래턱 • 약 14 (19 74) 파사라, 터키 이빨들 약 15 (19 69) 아프리카 포트 테란, 케냐 라 위케리 R. wi ck eri 14 (19 60) 이빨울 포함한 위 • 아래턱 조각 유럽 아데네,그리스 아래턱 8 (19 44) 루다반야,헝가리 라. 헝가리쿠스 R. hun ga ri cu s 약 13-14 (1975) 7 개의 턱조각과 3 개의 이빨
윌슨과 세리치는 육지에 사는 등뼈동물에 있는 알부민 분지를· 연구 하여 분자시계를 만들었다. 그들의 분자시계에 따르면 고릴라와 침팬 지, 그리고 인류는- 약 400 만 년 전에서 600 만 년 전에는 공통조상을 가지고 있었음을- 보여주고 있다. 다시 말해 고릴라와 침팬지 및 사람 은 위에서 말한 시기에 갈라져 나갔음을 뜻한다. 오랑우탄은 약 800 만 년 전에, 그리고 오늘날 살아 있는 원숭이들은 1000 만 년 전에서
1200 만 년 전에 갈라졌다는 주장이다. 윌슨과 세리치의 주장에 반해 사이몬스와 필빔은 약 1000 만 년 내 지 1500 만 년 전에 살았던 라마피데쿠스가 인류의 조상이라고 주장하 며 침팬지와 인류는 라마피테쿠스 이전에 갈라졌다고 주장하였다. 실 제로 1970 년대까지 고인류학 교과서에 라마피테쿠스는 인류의 조상으 로 기술되었다. 그러나 생화학자들은 인류와 침팬지는 약 600 만 년 전에 갈라졌다고 주장하고 있다. 화석중거를 바탕으로 한 고인류학자 둘은 생화학자들의 주장에 강한 이견을 제시하고 있으며, 일부 학자 둘은 라마피테쿠스를 인류의 조상으로 분류하지 않았다.
 사람
사람
1980 년대 초에 사이몬스와 필범은 새로이 발견된 화석자료들을 바 탕으로 라마피데쿠스는 더 이상 두 발로 걷지 않고 네 발로 걸었다고 인정하였다. 그 이후 다른 화석들의 발견으로 라마피데쿠스의 전화상 의 위치에 대한 설명에 보뎀이 되었다. 그 중 하나가 시바피테쿠스로 이는 라마피테쿠스와 아~느 닮았으며, 시바피테쿠스는 나중에 오랑우 탄의 조상으로 밝혀졌다. 또 다른 증거는 약 400 만 년 전에 살았던 호미니드의 화석으로 이는 라마피테쿠스가 복원되었을 때의 모습보다
더 유인원을 닮았다. 라마피테쿠스의 복원은 복원한 사람의 선입견에 의해서 이루어졌으 며, 이런 점에서 볼 때 학자들의 편견이 실제 연구에 어떠한 영향을 주는가에 관심을 두어야 한다. 라마피데쿠스는 여전히 사람의 조상일 가능성을 가지고 있으며, 또 한 유인원의 조상일 가능성도 가지고 있다. 왜냐하면 두 집단이 나타 나기 이전에 라마피데쿠스가 이미 분화되었을 것으로 여겨지기 때문 이다. 이 화석에서 보여지는 이빨의 두터운 사기질과 작은 송곳니 등 은 아마도 진화과정중에 음식물에 대한 적응으로부터 시작되었다고 가늠된다. 분자생물학의 연구 1969 년에 발표된 세리치와 윌슨의 연구에 대해 많은 논란이 제기되 었는데, 특히 화석을 전공하는 학자들로부터 비판이 제기되었다. 세리 치와 윌슨의 연구는 인류와 유인원이 500 만 년 전에 공통조상에서 분 화되었음을 보여주고 있다. 다음의 도표는 사람과 유인원의 해부학상 및 분자상의 관계를 나타내준다(그립 6. 8). 이 도표에 따르면 모든 유인원은 해부학상 많은 모양이 서로 닮았 으나, 이에 반해 사람과는 여러 면에서 닮지 않았다. 그러나 분자상으 로 사람과 침팬지, 고릴라는 서로 닮았고, 오랑우탄은 이들로부터 떨 어져 있는 것으로 보아 이들 정도는 닮지 않았음을 알 수 있다. 따라 서 침팬지와 고릴라, 침팬지와 사람, 사람과 고릴라의 분자상의 거리 는 모두 갇으며, 사람과 침팬지 또는 고릴라에서 오랑우탄까지는 모 두 같은 거리를 나타내주고 있다. 우리는 분자자료를 통해 공통조상에 대해 새로운 정보를 얻을 수
 l\ 우 탄
l\ 우 탄
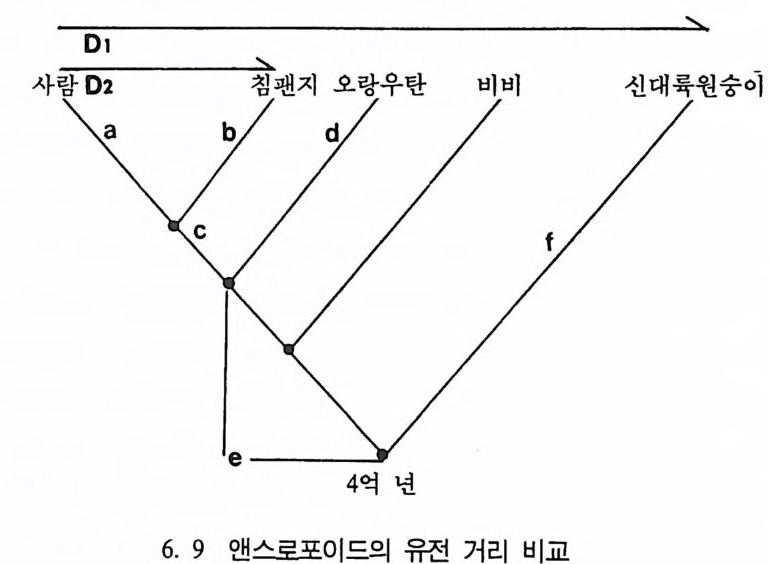 A Tr 溫i\ 비비 신대륙원숭이
A Tr 溫i\ 비비 신대륙원숭이
있으며 이 지식을 가지고 계통분화그립 Clado g ram 을 만들 수가 있고, 계속 자료들을 계통분화그립에 첨가하여 다른 영장류와 비교할 수 있 다. 그 예로 신대륙 원숭이와 비비들과의 관계를 아래의 계통분화그
립에서 볼 수 있다. 나아가 다른 〈 종 〉 들을 첨가할 때 분자상의 진화 에는 일정한 규칙이 성립됨을 알 수 있다• 죽 신대륙 원숭이와 비비, 침팬지, 고릴라, 오랑우탄 또는 사람과 갇은 앤스로포이드 사이의 분 자상의 거리는 항상 일정하다. 이는 사람과 유인원이 공통조상으로부 터 갈라진 이래, 해부학상의 전화는 크게 있었지만 분자상의 전화는 없었다는 것을 의미한다(그립 6. 9). D’ 와 D 놈 상대적으로 분자간의 거리를 나타낸다. D1 = a+c+e+f = 400 만 년 (D 戶 사람( H) 으로부터 신대륙 원숭이까지의 거리) D2 = a+b 이다. ( D2 : 침팬지로부터 사람에 이르는 거리이다) a = 사람에서 공통조상 1 까지의 거리 b= 공통조상 1 부터 침팬지까지의 거리 c= 공통조상 1 부터 공통조상 2 까지의 거리 d = 공통조상 2 부터 오랑우탄까지의 거리 e= 공통조상 2 에서 공통조상 . 3 까지의 거리 f=공통조상 3 에서 신대륙 원숭이까지의 거리 화석중거들은 이 신대륙 원숭이와 인류의 공통조상이 4000 만 년 전 에 살았음울 보여준다. 따라서 우리는 이것을 이용해서 사람과 다른 영장류들이 언제 갈라졌는지를 알 수 있다. 인류와 침팬지의 분화 =X my / 40 my = a+b/a+c+e+f = D1/D2 이 방법에 따르면 인류와 침팬지는 5~600 만 년 전에 이미 갈라졌 으며, 또 다른 분자물질을 사용한 결과 7~900 만 년이란 값을 얻었다. 앞에서 살펴보았던 화석자료들에 대한 해석을 다시 생각해 보자 . 필빔과 그 동료는 한때 1500 만 년 전에 살았던 라마피테쿠스가 유일
한 사람의 조상이라고 주장하였는데, 이는 다시 말해 사람과 유인원 의 갈라짐이 1500 만 년 이전에 일어났다는 것을 의미한다. 따라서 호 미노이드 화석자료의 해석과 분자상의 증거에는 명백하게 차이가 있 음을 알수 있다. 침팬지와 인간과의 차이 침팬지는 습관적으로 〈바깥 손마디 땅을 짚고 걷기〉를 하나 때때로 두 발로 걷기도 하는데, 그때는 몸의 중십이 좌우로 움직인다. 이는 침팬지의 근육, 뼈대 구조가 사람이 두 발로 서서 걸을 수 있는 근육과 뼈대 구조와 다르기 때문이다. 그러나 에너지의 사용이란 면 에서 볼 때 네 발로 걷는 침팬지에게 필요한 에너지는 두 발로 걷는 사람과 비교해 별 차이가 없다. 침팬지가 맹수로부터 도망가는 데는 사람만큼의 효과를 지닌다. 침팬지들은 과일과 채소, 그리고 비타민과 미네랄이 들어 있는 비 스킷을 먹는데, 그들이 쇠창살을 두드릴 때 보면 사람보다 5-7 배 정 도 더 힘이 세며 약 1kg 정도의 물체를 들어 울린다. 또한 특별한 훈 련을 받은 침팬지들은 많은 사람들 앞에서도 아무런 영향을 받지 않 는다. 사람들과 친숙해진 침팬지들이 다시 야생상태로 되는 데는 많은 시 간이 걸린다. 왜냐하면 그들에게 야생에서 살아 남는 기술을 가르치 고, 무엇을 먹을지, 무엇을 피해야 할지 등 생존에 필요한 적응력을 키우는 데는 많은 시간이 필요하기 때문이다. 또 다론 중요한 이유는 이 침팬지들을 위해서 남겨진 지역이 지구상에 거의 없다는 점이다. 이들이 살 수 있는 환경은 이미 살아 있는 야생 침팬지들이 점거하고 있으며, 어느 침팬지건 다시 야생상태로 되돌아가는 데는 이들에 의 해 방해를받는다.
침팬지들은 자주 사용되는 말들을 이해하며, 나이가 들면 사람처럼 목소리가 변하고, 지화법을 아는 침팬지가 디론· 침팬지들에게 이를 가르칠 수 있디는 주장도 있다. 지금까지 알려진 많은 침팬지의 행위는 실내에서도 조사되었다. 한 예로 미국 캘리포니아의 마린 월드에서 기르고 있는 침팬지들은 서로 찰 지내는데 왜냐하면 이들은 서로 같이 지낼 기회가 많기 때문이다. 이들은 몽둥이나 나뭇가지들을 무기처럼 사용하고 다른 침팬지에게 겁을 주기 위해 소리를 지르기도 한다• 침팬지가 어미로서의 행위인 새끼를 옮기는 일은 본능적이거나 아 니면 단순히 어미에 의해서 배우는 것으로, 어미는 어릴 때 배운 대 로 갓 태어난 새끼는 가슴에 안고 다른 새끼는 등에 업고 다닌다. 침팬지는 다른 소리들을 많이 낸다. 그들은 기쁠 때와 슬풀 때, 화 날 때와 즐거울 때, 그리고 음식을 취할 때 등등의 경우 각기 다른 소리를 낸다. 침팬지들은 사회성을 지니고 있으며 행동은 상냥하고 때로는 공격성을 지니기도 한다. 전에는 침팬지를 애완동물로 키우기 가 쉬운 일이었으나 오늘날에는 점점 어려운 일이 되고 있다. 왜냐하 면 침팬지는 사회동물로 홍미가 있는 것이지 혼자 있게 하는 것은 바 람직한 일이 아니라는 사실이 밝혀졌기 때문이다. 침팬지의 이빨 구성은 2: 1 : 2: 3 으로 사람과 갇고, 어린 침팬지는 5 세 내지 7 세가 되면 성인 침팬지가 된다. 침팬지는 감기, 수두, 홍역 과 사람들이 걸리는 병은 다 걸리며 치료약들도 사람에게 쓰이는 것 들과 똑같다. 침팬지의 수태기간은 8-9 개월이며 心1 ] 까지 부모들이 돌 봐준다.
 제 7 장 인류의 전화 I : 오스프로랄로피테쿠스
제 7 장 인류의 전화 I : 오스프로랄로피테쿠스
제 7 장 인류의 진화 I : 오스트랄로피테쿠스 Austr a lop ithe cus 인류의 기 원 : 플라이오 / 플라이스토세 Pli o- Pl e i s t ocene 의 호미니드 앞장에서는 6500 만 년 동안의 지질학상 구분과 영장류의 분류 및 영장류의 기원을 살펴보았다. 이오세 말에 지구의 기후가 변하면서 영장류의 생활은 적도지대로 한정되었다. 이 시기에 살던 초기 영장 류인 플레시아다피형에 속하는 영장류 중의 하나가 전화해 갔는데, 이들의 후예 중 하나가 올리고세 초기의 유적지인 이집트의 엘 화융 에서 발견된 이집토피테쿠스로, 이들은 유인원과 인류의 조상일 가능· 성이 높은 것으로 알려져 있다. 여기서는 이집토피테쿠스로부터 진화 한 호미노이드 영장류가 어떻게 오늘날의 인류까지 전화해 왔는지를 살펴보기로한다. 인류까지의 전화과정을 알기 위해서 하목 단계와 초과 단계의 프로 컨슬 영장류 화석까지 살펴보았다. 여기서는 프로컨슬이 번성했던 초 기 마이오세 이후에는 무슨 일이 일어났는지롤 알아보자. 프로컨술은 마이오세 초기에 살던 호미노이드 영장류로 오늘날 살 고 있는 호미니드의 멀고 먼 조상으로 알려져 있다• 그러나 이들이 호미니드의 조상이었다 하더라도 문제는 그들의 화석이 남아 있지 않
다는 것이다(그립 7. I). 위의 그림 7. I 에서 알 수 있듯이, 분자생물학 자료에 따르면 마이 오세 말기 (5-1000 만 년 전)는 유인원과 인류의 조상이 분리된 중요 한 시기인데, 불행히도 호미노이드 화석의 기록들이 가장 빈약한 시 기이기도 하다. 실제로 이 시기의 유적지 중 하나인 동아프리카의 로 타감 Lo t ha g arm 에서 턱조각들과 이빨이 하나 박혀 있는 작은 동물의 화석이 발견되었는데 이 화석들이 아프리카에서 발견된 5 - 600 만 년 전 호미니드에 관한 유일한 자료이다.
 123456789111111
123456789111111
한편 마이오세 말에 이르면 지중해가 메말라 아프리카 북부지역과 유럽 대륙이 서로 연결되었다. • 1 980 년대 초 북아프리카의 사하비 Sa habi 유적에서 이 시기에 속하는 많은 젖먹이동물의 뼈가 발견되었 다. 처음에 발견된 동물뼈 화석 중에 뻣조각 하나가 이 시기에 속하
는 인류화석의 빗장뼈라고 발표되었으나, 1980 년대 중반에 이 뼈는 돌고래의 갈비뼈로 밝혀졌다. 플라이오세 초기인 3 - 400 만 년 사이에 많은 호미노이드 화석자료 들이 발견되었는데, 이들 중 가장 유명한 사건으로 라마피테쿠스 화 석의 발견을 꼽을 수 있다. 화석자료를 살펴보면 이 시기의 라마피테 쿠스는 짧은 얼굴에 두껍고 큰 어금니를 가지고 있는데, 이 호미노이 드는 호미니드의 조상은 아니다 . 오늘날 라마피데쿠스는 마이오세와 플라이오세에 살던 호미노이드 영장류 중 아주 번성하였던 영장류의 한 〈 종 〉 으로 받아들여지고 있다 . 지금까지 아시아 대륙에서 이들에 관한 좋은 화석중거들이 발견되었지만 이들은 아프리카 대륙과 유럽 에도 널리 퍼져 살았던 것으로 알려지고 있다. 인류과에 속하는 오스트랄로피 테쿠스속g enus Aus t ralop i t hecus 과 인 류속g enus Homo 을 다른 영장류들과 구분할 때 화석에서 보여지는 해부학상 가장 중요한 기준의 하나는 〈 두발걷기〉 능력의 여부에 달려 있다. 1924 년 남아프리카 케이프 주에 있는 타웅 마을로부터 9.6 k m 떨어전 석회암 동굴에서 어린 개체의 머리뼈 화석 하나가 발견되었 다. 오스트랄로피 테쿠스 아프리카누스 Austr a lop ithe cus a fri canus 라고 아름 지어전 이 호미니드 화석이 발견된 이래 지금까지 인류의 기원 에 대한 많은 이론들이 있다. 이 이론들은 크게 오스트랄로피데쿠스 가 인류의 직접조상이라는 설과 우리의 먼 조상은 인류속g enus Ho- mo 의 한 종으로 아직까지 그 조상의 화석이 발견되지 않았다는 설로 나눌 수 있다. 1980 년대 후반 이후의 연구에 따르면 풀라이오세와 풀라이스토세에 걸쳐 아프리카 대륙에는 5 종의 오스트랄로피데쿠스가 살았다고 알려 져 있다(표 7. 1). 오늘날 일반적으로 이들 호미니드 중 5-400 만 년 전에 동아프리카에서 나타난 오스트랄로피테쿠스 아파렌시스 Aus tr a lop ithe cus afar ensis Aus t ralop it hec 盛를 인류의 가장 이른시기의 조상으
로 받아들이고 있다.
표 7. I 호미니드의 생물학상 분류 목 Order : 영 장류 Prim ate s 아목 Suborder : 유인원 아목 An t hro p o i dea 하목 In fra order : 좁은코원숭이 하목 Cata r rhi ni 초과 Su p er fami l y : 호미노이드 초과 Ho mi no i dea 과 Fa mily : 인류과 Homi nida e 속 Genus : + 오스트랄로피 데쿠스 Austr a lop ithe cus 종 Sp ec i es : 오스트랄로피데쿠스 아파렌시스 A . afa re nsis 오스트랄로피 테쿠스 아프리카누스 A . afr ica nus 오스트랄로피데쿠스 에티오피쿠스 A. eth i o p icu s 오스트랄로피데쿠스 로보스무스 A . robustu s 오스트랄로피데쿠스 보이세이 A. bois e i 속 Genus : 사람 Homo 종 Sp e ci es : + 손쓴사람 Homo habil is + 곧선사람 Homo erectu s + 네 안데르탈사람 Homo neanderth a lensis 슬기 사람 Homo sap ien s ·+ : 오늘날 철멸한 호미니드를 나타냄.
가장 이론시기 인류의 조상 : 오스트랄로피테쿠스 아파렌 시 스 Austr a lop ithe cus af a rens i s 의 발견 마이오세 말에 인류의 요람지로 알려져 있는 아프리카 대륙과 그 주위의 모습은 어떠했는지 살펴보자. 중동아시아 아키바 만과 사해, 그리고 이 지역을 연결하는 골짝은 오늘날 요르단과 이스라엘의 국경
선을 이루고 있는데, 이 지역은 지구의 얇은 지각층들이 서로 맞닿아 지질상 얼개를 이루고 있는 곳이다. 지구 내부의 대류작용에 의해 지각변동이 생기면 지각총이 맞닿는 부분이 움직여서 벌어지는데 그 결과 지각충이 맞닿는 부분에 골짝이 형성되며, 동시에 그 부분을 따라 많은 화산활동이 일어난다. 아키바 만과 사해는 바로 이런 지각운동에 의해 생간 골짝의 일부분이다. 요르단과 사해 근처의 퇴적은 공룡이 살았던 중생대에 물 밀에서 형성된 것으로 골짝도 같은 시기에 만들어졌다. 골짝이 형성된 뒤 골 짝 둘레로부터 물이 흘러 들어가는 분지는 물이 증발되고 소금만 남 게 되었는데 이런 작용에 의해 사해가 형성되었다. 중동아시아에 발달한 골짝은 남쪽으로 계속해 이어져 더 거대하고 오래 된 골짝과 맞닿아 있다. 이 연속된 계곡의 일부분에 물이 차 있 는데 홍해와 아덴만이 그러하다. 한편 홍해와 아덴만이 갈라지는 곳 에서 다론 갈래의 골짝은 에티오피아로 뻗어 나가는데 이 부분은 물이 차 있지 않고 소위 아프리카 큰 골짝을 이루고 있다. 에티오피아의 큰 골짝 안에는 강과 호수들이 형성되어 있으며 이 지역에서 강물이나 흐르는 물에 떨어진 것들은 호수로 쓸려가 퇴적된 다. 이런 작용은 수백만 년 동안 반복해서 전행되고 있는데 바로 이 런 자연환경의 원인에 의해 화석들이 이 지역에서 퇴적되고 발견된다 (그림 7. 2). 에티오피아에서 시작된 아프리카 큰 골짝은 동아프리카를 지나 남 쪽으로 수천 킬로미터를 따라 형성되어, 이 큰 골짝 안에서 가장 이 른시기의 호미니드 화석과 동물화석 및 인류 조상의 흔적들이 발견되 고 있다(표 7. 2). 이 유적지들 중에서 동아프리카에 있는 에티오피아 의 아파 분지에 위치한 하다 유적지와 탄자니아의 세렌게티 고원에 위치한 라에톨리 유적에서 가장 이론시기의 호미니드 화석과 그들의 발자국이 . 발견되었다.
 빅토리아
빅토리아
표 7. 2 동아프리카에서 가장 이른시기의 호미니드 화석이 발견된 유적지 유적지 발굴책임자 발굴연대(유적지 연대) 하다 (Hadar : 에티오피아) 도날드요한 슨 1973- 1 977 Donald Joh anson 마우리스타이브 (290 - 350 만 년 전) Mauris Taie b 라에톨리 (Lae t o li : 탄자니아) 메리 리키 1977- 1 980 Mary Leakey (350 만 년 전) 미들 아와쉬 (M idd le Awash : 데스몬드클라크 198 1( 400 만 년 전) 에티오피아) J.D . C lark
하다 Hadar : 〈 루시 〉 와 가장 이론시기 호미니드 화석의 발견 하다 유적지는 에티오피아의 북부지역인 아파 사막이라고 알려진 지역에 있는데, 아파란 이름은 아파 주민이 이 지역에서 살기 때문에 얻어진 이름이다(그립 7. 3). 이 지역은 옛날 호수와 강 바닥들이 수 백 평방킬로미터에 걸쳐 펼쳐져 있으며, 지질학자인 마우리스 타이브 가 이 지역에서 지질연구를 하다가 처음으로 호미니드 화석을 찾아냈 다. 이 지역은 습기가 많으며 과거에 하다 호수가 있었던 곳으로 동 물들이 살고, 죽고 그리고 호숫가에 묻혔다. 오늘날 하다 유적지 일대는 불모지로, 건조하고 사람이 살기에는 힘 든 지역으로 기온은 한낮에는 화씨 120-130 도를 오르내린다. 물을 얻는 것이 문제이나 때로는 땅을 파면 물이 나오는 곳도 있으며, 그 늘이라고는 자동차 때문에 생기는 것이 전부이다. 하다 유적지에서 동물들이 사지가 붙어 있는 채로 묻혀 뼈대들이 해체되지 않은 상태로 발견되는 점으로 보아 라에톨리 유적지와는 다
 /,'\ • 예멘 `` V ' ’
/,'\ • 예멘 `` V ' ’
르게 동물들은 죽은 후에 빠르게 땅 속에 묻혔다는 것을 알 수 있다. 하다 유적지에서는 코끼리뼈와 돼지의 긴머리뼈, 커다란 수달과 디세 노데리아 코끼리뼈 화석, 그리고 작은 식충류의 턱들과 화석화된 악 어알 등 풍부한 동물화석들이 발견되었다. 1973 년 미국 • 프랑스 합동조사단이 하다 유적지 일대를 조사하였는 데 당시 미국측 발굴단장은 클리블랜드 박물관의 젊은 고인류학자였 던 도날드 요한슨이었다(사전 7. 4). 첫해 조사에서 매우 중요한 호미
 7. 4 아파 지역에서 발굴중인 요한슨의 모습
7. 4 아파 지역에서 발굴중인 요한슨의 모습
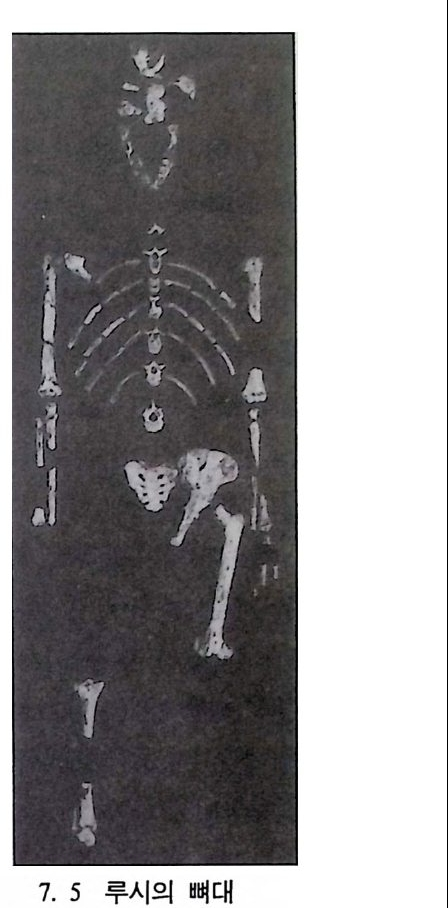 ,`i/,.‘i - '/b.tT:,^’.,i /) ' `1\ 4.\'·'\I
,`i/,.‘i - '/b.tT:,^’.,i /) ' `1\ 4.\'·'\I
니드의 입천장 뻣조각이 한 구릉에서 발견되었다 . 구릉에서 나온 흙 은 모두 체질을 하여 뻣조각들을 모았다. 이어서 무릎뼈 하나가 발견 되었다. 다음해에 팔뼈, 다리뼈, 머리뼈 조각들이 발견되었다 . 발견된 뼈화석들은 한 개체에 속하는 것으로 한 어른 개체의 거의 46% 에 달하는 양이었다. 이 뼈화석은 〈루시〉라고 이름 지어졌는데 발굴되기 2 년 전 유행하던 노래에서 따온 이름이었다. 약 300 만 년 전에 살았던 루시는 곧 세상에서 가장 유명한 화석 중의 하나가 되었 다. 루시는 약 90cm 의 작은 키와 침팬지보다 더 가벼운 몸무게를 가 지고 있으나 여자 어른으로 밝혀졌다(사진 7. 5). 루시 발견의 중요성
은 40% 가 넘는 뼈화석이 발견됐다든가, 키가 작다는 것에 있다기보 다는 이 호미니드가 두 발로 서서 걸은 인류의 가장 먼 조상의 선상 에 위치하고 있다는 데에 있다. 같은 시즌에 다른 이빨들과 턱들이 발견되었다. 다음해 아파 지역 333(AL 333) 으로 분류된 지역에서 13 개체의 호 미니드 화석이 함께 발견되었다. 이 발견은 루시에 버금 가는 대단한 발견이었다. 요한슨과 그의 동료들은 이 13 개체의 호미니드를 홍수와 같은 천재 지 변 Cata s tr o p hic Event 에 의 해 일시 에 파묻힌 한 사회 집 단이라고 추측한다. 따라서 이 화석 집단은 〈 최초의 인류 가족 〉 으로 알려져 있다. 아프리카 대협곡이 만들어질 때 진행되었던 화산활동은 고인류학자 와 지질학자들에게 매우 유용하다. 화산폭발로 생긴 화산재가 땅의 표면을 덮어 충을 이루고 있는데, 이 화산재가 쌓인 충은 칼륨/아르 곤과 분열 자국 방법에 의해 연대가 밝혀지며, 또한 지각운동에 의해 변화된 충위를 서로 비교함으로써 선후관계를 알 수 있다. 위의 연대 측정방법에 따라 하다 유적지에서 발견된 호미니드는 약 300 만 년 내 지 350 만 년 전에 살았던 것으로 밝혀졌다. 한편 아파 지역의 미들 아와쉬 유적 조사는 1981 년 버클리 대학교 의 인류학자인 데스몬드 클라크 등에 의해 전행되었다. 이 유적에서 400 만 년의 연대를 가전 충의 위 • 아래에서 호미니드 화석들이 발견 되었다. 화석은 침팬지와 비슷한 크기의 앞머리뼈를 포함하여 허벅지 뼈 등이 발견되었는데 넓적다리뼈는 이 호미니드가 곧바로 서서 걸었 음을 보여주며 하다에서 발견된 호미니드와 같은 종으로 알려지고 있 다.
라에톨리 : 가장 이론시기의 발자국 라에톨리 유적지는 탄자니아의 유명한 올드바이 고지로부터 남쪽으 로 약 32km 정도 떨어전 세렌게티 고원 남쪽 끝에 있는 경사면에 위치하고 있다 . 라에톨리는 아파 지역처럼 불모지가 아닌, 세상에서 가장 장관인 동물의 이동을 볼 수 있는 지역으로 사자와 하이에나, 기린 및 코끼리 등의 동물로부터 기니아 닭과 돼지, 토끼와 영양들, 그리고 독사와 전드기에 이르기까지 많은 종류의 동물들이 서식하고 있다. 이런 환경은 동아프리카의 사바나를 대표하는 진면목으로 가장 이른시기의 호미니드가 살던 350 만 년 전의 풍경과 별로 다르지 않 다. 그러나 칼날이빨 고양이과의 동물과 상아 끝이 밑으로 굽은 디노 세리움 코끼리와 칼리코세리 말과 난쟁이 기린 등은 오늘날 보이지 않는다. 라에톨리에서 호미니드 화석은 매우 귀하다. 1974 년에 메리 리키의 아프리카 조수인 멍게라 머카가 라에톨리를 횡단하면서 호미니드의 아래 셋째어금니 한 개를 찾아냈다. 이빨이 발견된 충을 덮고 있는 용암을 가지고 방사선동위원소 측정 K/Ar 을 한 결과, 240 만 년이란 연대를 얻은 메리 리키는 연구를 이 유적지에 집중하기로 결정하였 다. 1978 년 이 유적지롤 방문한 영국의 인류학자인 앤드루 힐 Andrew H ill을 포함한 학자들은 일부가 잘라져 나간 발자국이 단단하게 굳어 전 화산재 바닥에 남겨진 것을 발견하였다. 이 발자국을 조사해 본 결과 두 발로 걷는 세 명의 생물체가 남긴 것으로 판명되었다(사전 7. 6). 발자국 중 하나는 크고 또 다른 하나는 작은데, 이 발견은 대단히 중요한 의미를 지닌다 . 왜냐하면 이 발자국은 360 만 년 전에 살던 가 장 이른시기 호미니드의 발자국이고, 또한 이들이 두 발로 이미 걷고 있었다는 것을 증명해 주기 때문이다. 이 발자국을 남긴 호미니드는
 7. 6 라에톨리에서 발견된 370 만 년 전의 호미 니드 발자국
7. 6 라에톨리에서 발견된 370 만 년 전의 호미 니드 발자국
인류의 직접조상으로 보여지는 선상에 있음에 들림없다고 발굴자인 메리 리키는 주장하였다. 이 유적지의 연대는 350 만 년 전에서 375 만 년 전 사이로 알려졌는데, 이 연대는 지금까지 알려진 호미니드의 연 대로는가장 이르다. 360 만 년 전의 호미니드 발자국? 라에톨리 동쪽으로 42km 정도 떨어전 곳에 노몽고로 휴화산이 희미하
게 보인다. 거대한 분화구는 세렌게티 고원에서 살고 있는 초식동물에게 풍부한 물을 제공한다. 이 휴화산 주위 에는 여러 개의 작은 화산 봉우리 둘이 흩어져 있는데. 특히 마사이족들에 의해 〈신의 산〉으로 불리는 옹도 뇨렌가이 화산은 오늘날에도 때때로 증기구름과 화산재를 세렌게티 평원 하늘 높이 날려 보낸다.
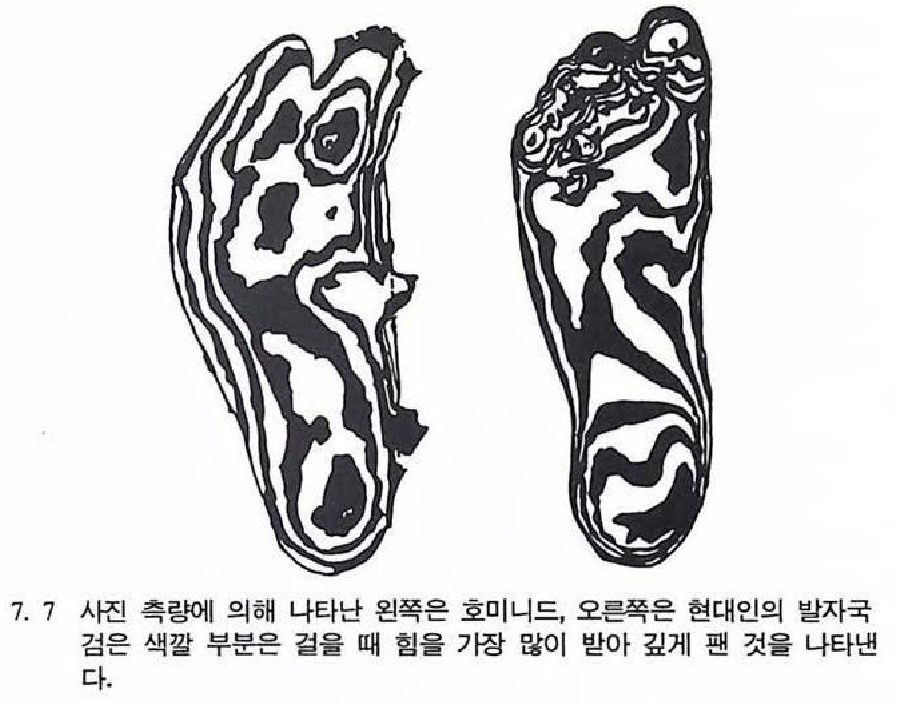 7. 7 사진 측량에 의해 나타난 왼쪽은 호미니드, 오른쪽은 현대인의 발자국
7. 7 사진 측량에 의해 나타난 왼쪽은 호미니드, 오른쪽은 현대인의 발자국
360 만 년 전 라에톨리 일대는 한때 화산재가 마치 눈처럼 온 지역을 덮은 적이 있었다 . 당시 화산의 활동법위는 지금과 어느 정도 비슷하였다. 라에돌리 동쪽에 위치한 사디만 화산은 그 당시 주기적으로 거대한 증기 구름과 화산재를 분출하였는데, 이것은 마치 오늘날 울도뇨렌가이 화산이 약하게 활동하는 것과 같다. 또한 울도뇨렌가이 화산처럽 사디만 화산은 특별한 성질을 지니고 있는 화산재를 분출하였는데, 이 화산재는 약간 젖 어 있을 때 시멘트처럼 단단하게 굳어버리는 특성이 있다. 이 성질이 360
만 년 전의 발자국을 만들고 보존하는 데 결정적인 구실을 하였다(그림 7. 7). 사디만 화산에서 분출된 화산재는 동물이 살아 남는 데 결정적인 영향 울 주지는 않았다. 화산의 폭발로 라에톨리 일대는 잠시 동안 갈색 해변 처럼 변하였는데, 여기 새로 쌓인 화산재 위에 발자국이 새겨졌다. 화산재 는 작은 나뭇가지들, 새알들, 벌들 그리고 심지어는 동물의 배설물까지도 마치 담요처럼 충충이 얇게 덮었다. 그때 빗물이 화산재에 작은 구멍을 뚫었고、 빗물에 의해 축축해진 카보나이트는 천천히 시멘트로 변하였으며, 동시에 화산재와 발자국의 기록들이 바위처럼 단단한 충으로 변하였다. 사디만 화산이 다시 폭발하자 화산재가 라에톨리의 발자국을 다시 덮어버 렸다. 그 위에 다른 동물들이 지나가면서 또 다른 발자국들이 찍혀지고, 그 위에 비를 뿌렸다. 이런 현상은 화산재가 약 15cm 정도의 두께가 될 때까지 계속적으로 반복되어, 마치 책 속에 끼워놓은 나뭇잎처럼 영원히 발자국이 보존되었는데, 이런 과정은 모두 2 주일 사이에 일어난 일이었다. (인류의 기원과 진화, 1992, 61 쪽) 1970 년대 후반에 버클리 대학교의 팀 화이트Ti m Whit e 가 요한슨 의 진영에 가담하였다. 하다에서 발견된 풍부한 화석을 바탕으로 요 한슨과 그의 발굴팀은 1978 년에 발표된 연구에서 루시와 다른 화석들 이 두 개의 서로 다른 종이 아닌 같은 단일종으로 보았다. 그들은 이 단일종을 오스트랄로피테쿠스 아파렌시스라고 이름지었으며 이 새로 운 이름의 호미니드는 제 3 기 말에 나타나며 모든 호미니드의 뿌리라 고 주장하였다(사진 7. 8). 그들은 탄자니아의 라에톨리에서 발견된 발자국은 에티오피아의 하다와 수천 킬로미터 이상 떨어져 있지만 하 다에서 발견된 화석과 같은 종인 오스트랄로피데쿠스 아파렌시스가 남긴 것이라고 주장하였다. 에티오피아의 하다에서 발견된 호미니드 화석은 서로 다른 두 종이
아닌 단일종이고, 인류과의 기원은 아주 오래 전이 아닌 제 3 기 말에 야 시작되며, 하다와 라에톨리에서 나온 화석은 같은 종이라는 주장 은 요한슨과 화이트 및 이를 지지하는 학자와 메리 리키와 리처드 리 키를 지지하는 학자들 사이에 격렬한 논쟁을 불러일으켰다. 이 문제 에 대한 논쟁은 적어도 20 여 년 동안이나 진행되었으나, 오늘날 많은 인류학자들은 오스트랄로피테쿠스 아파렌시스 한 〈 종 〉 만이 살았다는 주장에 견해를 같이하고 있다. 그러나 리키측은 아직도 이런 주장을 받아들이지 않고 있다.
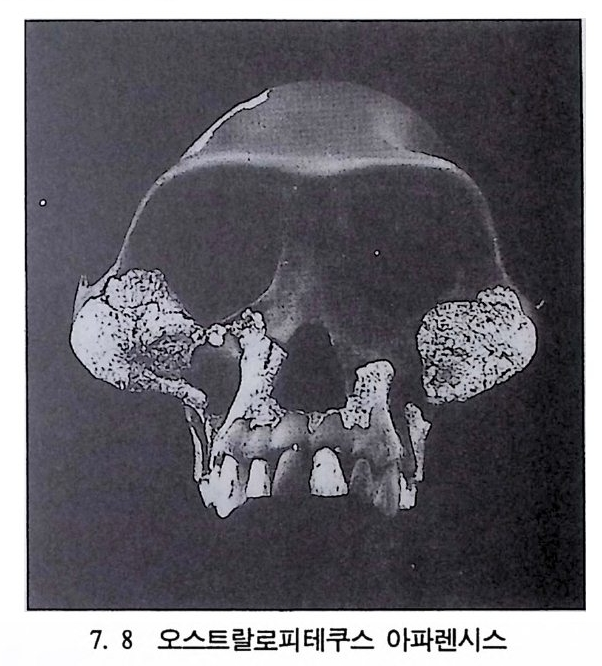 7. 8 오스트랄로피테쿠스 아파렌시스
7. 8 오스트랄로피테쿠스 아파렌시스
하다 호미니드 화석과 라에톨리 발자국은 둘 다 고인류학상 중요한 발견으로 우리에게 화석과 또 우리 자신에 대해 다음과 같은 것을 시 사해 준다. 첫째, 오스트랄로피테쿠스 아파렌시스가 인류의 가장 이른시기의
조상이라면 이는 다윈이 〈 아프리카가 인류의 기원지 〉 라는 가설이 옳 다는 것을증명해 준다. 19 세기 이래 학자들 사이에서는 인류가 어디서 처음으로 나타났는 지, 또 어떻게 퍼져 나갔는지에 대해 많은 논란이 있어 왔다. 찰스 다 윈과 토머스 헨리 헉슬리를 포함한 학자들은 아프리카가 인류의 기원 지일 가능성이 가장 높은 지역으로 꼽았다. 특히 다윈은 『 종의 기원 』 과 r 인류의 기원 』 에서 인류의 기원지를 아프리카로 추정하였는데, 다 윈이 살았던 당시 인류에 관계된 화석은 발견되지 않았으나 뛰어난 생물학자였던 다윈은 일차로 인간과 해부학상으로 가장 닮은 침팬지 가 아프리카에서만 산다는 점에 주목하였으며, 당시 유럽의 여러 학 자들처럼 다윈도 〈검은 대륙 〉 으로 인식되던 아프리카 대륙에 대한 호 기심이 그로 하여금 아프리카를 인류의 기원지로서 가장 타당한 장소 로 생각하게 하는 요인이었다. 그러나 다윈의 자연선택이론이 한때 인정을 받지 못했던 것처럼 아 프리카 대륙이 인류의 기원지라는 설 또한 거부당했으며, 대신 아시 아 대륙이 인류의 기원지로 각광을 받은 때도 있었다 . 다윈의 지지자 로 동물학자인 언스트 헤켈 Erns t Haekel 은 아시아에서 살고 있는 오 랑우탄과 긴팔원숭이가 인간과 가장 가깝다고 생각하여, 인류의 기원 지를 아시아에서 찾고자 하였다. 비록 헤켈의 생각이 정확하지는 않 았으나 그의 가설은 드보아 Eu g ene Dubo i s 를 위시한 많은 학자들로 하여금 아시아에서 〈인류의 화석〉을 찾게 하는 계기가 되었다. 따라 서 19 세기 후반부터 20Al] 기 초반까지 인류화석을 발견하고자 하는 노 력은 주로 아시아 대륙에 집중되었다. 따라서 인류학계는 1925 년 〈아프리카에서 최초의 유인원 - 사람〉으 로 보고된 레이몬드 다트 Ra y mond Dar t의 발견에 대해 알려고 하지 않았으며, 1931 년 루이스 리키가 캠브리지 대학의 인류학 담당연구원 들에게 선사인류를 찾기 위해 아프리카 대륙으로 가겠다고 말했을 때
그들은 루이스가 정신이 돌았거나 잘못된 지식을 가지고 있다고 생각 하였다. 그러나 20 세기 이후 수많은 인류화석이 아프리카에서 발견되 고, 인류 최초의 조상인 루시와 나머지 다른 오스트랄로피데쿠스 아 파렌시스의 화석이 발견됨으로써 다윈과 다트 그리고 리키 등의 가설 이 옳다는 것이 증명되었다. 아프리카는 정말로 인류의 기원지였다. 둘째, 루시와 라에톨리에서의 발자국 발견은 스티븐 제이 굴드가 언급한 〈 큰 골 우선 가설 〉 에 관한 검증이다. 영장류에서 네 발 대신 두 발로 걷기 시작한 전화상의 전환은 인류가 다론 영장류와 구분되 기 시작하여 인류의 길로 접어들게 한 여러 가지 적응의 한 부분으로 다윈은 생각하였다. 다윈의 이론에 따르면 칼날 같은 송곳니가 줄어 들면서 생기는 불이익은 바로 연모의 제작과 그것을 무기로써 사용하 는 것에 의해 보상되었다고 한다. 물론 연모를 만들고 사용함으로써 두 손은 자유롭게 되었으며 따라서 두 발로 걷는 이점을 가져왔다고 생각하였다. 복잡한 기술 발달은 지능 발달을 촉진시켰으며, 그 결과 두뇌가 더 욱 커졌다. 이와 함께 좀더 복잡한 사회생활이 요구되었고, 이런 사회 생활은 연모제작과 식량획득 방법에 관한 일들을 강화시켰다. 이 모 든 요소들은 철저한 〈피드백 작용〉의 범위에서 서로 연결되어, 옛인 류를 오늘날의 인류로 향하여 나아가게 하였다고 다윈은 생각하였다 (그림 7. 9). 이렇게 인류는 두 발로 걷기 전에 큰 머리를 먼저 가지 고 있었디는 다윈의 이론은 사실상 처음부터 인류의 조상을 동물의 세계와 분리하였다. 그의 피드백 이론은 모든 것을 설명할 수 있으면 서도 아무것도 설명하지롤 못했지만, 그의 이론은 인류의 진화에 관 한 생각에 오랫동안 많은 영향을 끼치면서 지속되었다. 그러나 1924 년 다트 교수가 타응에서 어린아이를 찾은 이래, 메리 리키의 라에톨리 발자국과 요한슨의 에티오피아 화석의 발견은 호미 니드가 인류처럼 사유할 수 있기 전, 죽 연모의 사용과 커다란 두뇌
의 발달 이전에 먼저 인류처럼 두 발로 걸을 수 있었다는 것을 밝혀 주고있다.
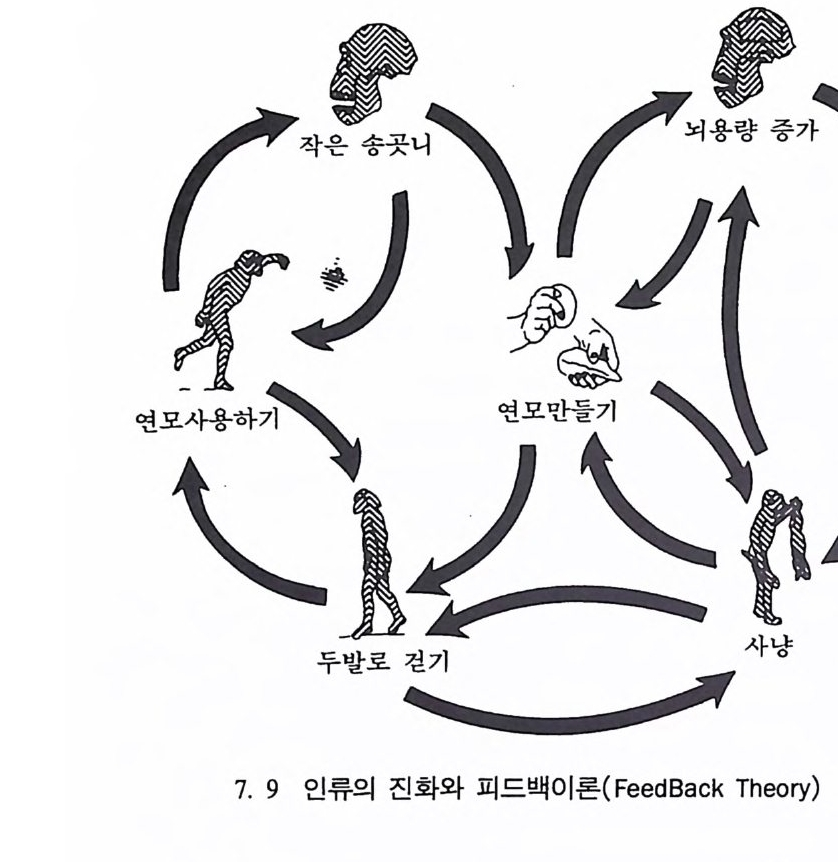 연 \\ 1L 〉、
연 \\ 1L 〉、
남아프리카 호미니드의 발견 사람은 인류과에서 유일하게 살아 남아 있는 〈종〉이다. 그러나 과 거에는 다론 존재들도 있었는데, 앞에서 살펴보았듯이 이들은 모두가 여러 오스트랄로피테쿠스속과 인류속 중의 한 종으로 알려져 있다. 이때 인류과에 속하는 호미니드와 다론 영장류와의 차이는 〈두 발로 걷기〉의 여부에 있다.
1973 년과 1978 년에 걸쳐 에티오피아의 하다 유적지와 탄자니아의 라에톨리 유적지에서 발견된 3 - 400 만 년 전의 화석들은 두 발로 걸 었으며, 이 특징을 바탕으로 오스트랄로피데쿠스 아파렌시스라는 새 로운 〈 종 〉 이 생겨났다. 그러나 이 화석들에 대한 해석은 고인류학계 에 많은 논쟁을 불러일으켰으니, 이 논쟁의 내용을 알기 위해서 오스 트랄로피데쿠스 화석의 발견과 이 호미니드에 대한 해석의 역사를 살 펴보기로 한다.
 7. 10 필트 E 듄 화석을 조사하는 영국 학자들
7. 10 필트 E 듄 화석을 조사하는 영국 학자들
이야기는 1912 년 영국의 필트다운이란 마을에서부터 시작된다 . 이 마울에서 찰스 도슨 Charles Dowson 이 선사시대 사람의 이빨과 아래 턱 및 머리뼈 부분울 찾아냈다(사진 7. 10). 그는 이 뼈화석을 〈이오 앤人쿠 立人- 도소니 (Eoa n.t hrop us dowson .i :여명의 사람 =dawn man)> 이라고 이름 지었다. 이 화석의 턱은 유인원를 닮았고 머리통은 현대
사람 것만큼이나 컸다. 이런 모습의 머리뼈는 당시에 유행하고 있던, 머리뼈가 커야만 인류의 조상이라고 생각했던 선입관에 딱 들어맞았 다 . 이오앤人쿠立人 화석이 발견되기 전에 독일과 프랑스 등지에서는 이미 많은 네안데르탈사람의 뼈둘이 동굴에서 발견되었으나 영국 내 에서는 선사시대 인류의 뼈화석이 발견된 적이 없었다. 이런 상황에 서 도슨의 발견은 영국의 자존심을 살려준 것으로 받아들여졌다. 따 라서 이 머리뼈는 1953 년 이 뼈의 발견이 사기라고 알려지기 전까지 교과서에 〈선사인〉이라고 실려왔었다• 그러나 도슨이 발견한 턱은 오 랑우탄의 것이며 머리통은 현대 사람의 것으로 화학적으로 처리해 옛 날 뼈처럼 보이게 조작하여 그 자리에 묻은 것이었다. 타웅 어린아이 화석(오스트랄로피테쿠스 아프리카누스 Aus t ra lop ithe cus africa nus) : 레이몬드 다트의 발견 네안데르탈사람의 발견과 필트다운 마울에서 발견된 뼈화석을 선사 인류의 표준으로 · 간주하는 생각이 유럽 학계를 풍미하는 동안에 아프 리카를 포함한 다른 지역들에서 인류의 조상을 찾고자 하는 노력이 계속되었다. 처음으로 두 발로 걸었던 영장류 화석의 발견은 사람들 이 기대하지 않았던 남아프리카의 트란스발 지역에서 시작되었다. 남아프리카의 요하네스버그에 있는 위트워터스랜드 대학교의 해부 학 교수였던 레이몬드 다트 교수는 학생들에게 근처에서 발견되는 뼈 화석이 들어 있는 바위조각들을 가져오도록 하였다. 1924 년에 한 학생 이 옛 비비의 머리뼈 화석을 보내왔는데, 이 화석은 요하네스버그 에서 320km 떨어전 타웅이라는 석회암 광산에서 발견된 것이었다. 다트는 이에 홍미를 갖고 다른 화석들을 보내도록 광산 주인을 설득 하였다. 그가 보내준 화석들을 조사하다가 다트는 머리통과 석회암으
로 덮인 어린 개체의 얼굴에 주목하게 되었다. 73 일간의 작업 끝에 머리뼈에 붙은 석회암이 모두 제거되자 다트의 눈앞에는 5 내지 g11 정도의 거의 완전한 얼굴과 머리통을 지닌 머리뼈가 나타났다(사진 7. 11 ) .
 -- — -_ ; 나函 r- '-- ._.
-- — -_ ; 나函 r- '-- ._.
〈두뇌 전화〉가 전문분야였던 다트는 죽각 이 어린 개체가 비비 화 석이 아님을 알게 되었다. 머리 크기는 어른 침팬지의 머리 정도였으 나 이 개체는 사람을 닮은 턱을 가지고 있었다. 위 • 아래턱에는 갓난 이가 있었고 어른 이빨도 솟아나고 있었는데 이빨도 사람의 이빨울 닮았다. 머리뼈굼의 위치는 침팬지와 고릴라와는 다르게 머리 앞쪽으 로 발달해 있어 타웅 아이가 두 발로 걸었음을 보여주고 있었다. 다트는 〈이 어린 개체가 사람의 조상〉이라고 단정하고 1925 '.1 이 롤 〈오스트랄로피데쿠스 아프리카누스 (Aus tra lop ith ecus africa nus, aus-
tral is :남쪽의 : pithe kos : 유인원 : 아프리카의 남쪽 원숭이, Af rica n south ern a p e) 〉라고 명명하였다. 그러나 이때는 유럽에서 필트다운사 람이 인류의 조상 중 하나로 받아들여지는 때였기 때문에 당시 대부 분의 유럽 인류학자들과 해부학자들은 다트의 주장을 묵살하였으며, 타웅 아이를 옛날에 살던 침팬지나 고릴라를 닮은 동물이라고 결론 지었다. 그러나 다트는 고동물학자이자 의사인 스코틀랜드 출신인 로버트 브룸 (Rober t Broom, 1866-1951) 의 도움을 받았다. 남아프리카의 여 러 지역에서 화석을 수집, 연구하던 브룸은 다트가 발견한 머리뼈를 보고 이는 〈인류 조상의 화석〉이라고 결론 지었다. 이에 힘입은 다트 는 남아프리카의 다른 지역들을 발굴하기 시작하였다. 스테르크폰테인 Ste r kf on te i n , 크롬드라이 Kromdraai, 스와 르트크란스 Swa rt krans 와 마카판스가트 Makap a nsga t : 오스트 랄로피테쿠스 아프리카누스와 오스트랄로피테쿠스 로보스두스 Austr a lop ithe cus robustu s 남아프리카의 프레토리아 지역의 트란스발 박물관의 연구원이 된 로버트 브룸은 1936 년부터 1950 년에 걸쳐 남아프리카의 스데르크폰테 인 유적의 발굴에 성공하여 완전한 형태의 어른 여자의 머리뼈 하나 와 거의 완전한 모습의 여자 엉덩뼈 하나를 포함하여 많은 화석들을 발굴하였다. 한편 1938 년 6 월 어느 날, 크롬드라이의 석회암 광산에서 한 국민 학교 학생이 어금니 한 개가 박혀 있는 호미니드 위턱의 일부를 발견 하였다. 브룸은 이 소년과 함께 호미니드 화석이 발견된 장소로 가서 많은 머리뼈 조각과 이빨울 찾아냈다. 이 조각들은 머리뼈 하나로부 터 부서져 나온 것이었다. 이 머리뼈는 잘 발달된 머리 가운능선
Sag ittal cres t을 가지고 있었으며, 오스트랄로피데쿠스 아프리카누스 의 머리뼈보다 더 큰 얼굴과 큰 어금니들을 갖고 있었다. 브룸은 이 머리뼈를 파란트로푸스 로보스두스 (Paran t hrop us robustu s , 인류의 옆 가지 : to the sid e of man) 라고 이름 지었다.
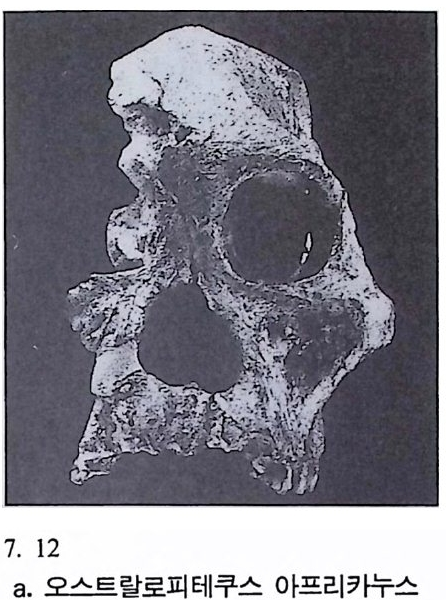 7. 12
7. 12
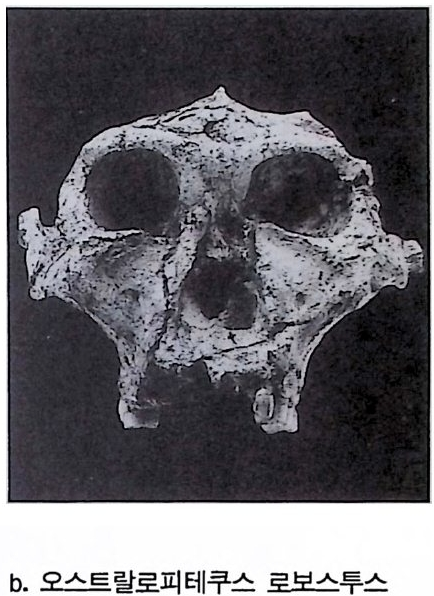 b. 오스트랄로피테쿠스 로보스투스
b. 오스트랄로피테쿠스 로보스투스
1948 년 다트는 스와르트크란스를 발굴하여 〈굳세 보이는 형태의 호 미니드 〉 의 커다란 턱 하나롤 포함하여 많은 화석들을 발견하였다. 이 호미니드 화석은 커다란 얼굴과 큰 이빨을 지니고 있는데, 학자들· 대 다수는 이 새로 발견된 화석이 오스트랄로피데쿠스 아프리카누스와는 다른 특칭을 지니고 있지만 새로운 속으로 분류하기에는 충분하지 않 다고 생각하였다. 그래서 이 화석을 오스프츠랄로피테쿠스 로보스투스 Austr a lop ithe cus robus tu s 라고 이름 지었다(사진 7. 12). 한편 브룸의 뒤를 이은 로빈슨J .TRob i nson 은 스와르트크란스 유적지에서 사람을 닮은 다른 화석을 찾아냈다.
1949 년까지 남아프리카의 동굴 유적지에서는 모두 30 개체 이상의 화석이 발견되었다. 이룰 바탕으로 영국 옥스포드 대학교의 해부학교 수인 윌프레드 르 그로스 클라크 Wi lf r ed Le Gros Clark 는 오스트랄 로피테쿠스가 사람의 조상이라는 설을 받아들였다. 1950 년대 중반까 지 모두 5 개의 유적지에서 많은 수의 호미니드 화석들이 발견되었는 데, 이들은 브룸과 로빈슨에 의해 모두 오스트랄로피테쿠스 아프리카 누스와 오스트랄로피데쿠스 로보스투스로 크게 분류되었다. 한편 동굴 내부의 퇴적을 연구하던 남아프리카의 브레인 C.K.Br ai n 은 호미니드가 발견된 퇴적충의 연대를 조사하였다. 그의 연구에 따르면 오스트랄로피데쿠스 아프리카누스의 연대가 오스트랄로피테쿠 스 로보스투스보다 오래 되었으며, 몸집이 작은 오스트랄로피데쿠스 아프리카누스는 더 큰 몸집으로 변해가는 데 비해 커다란 몸집의 오 스트랄로피테쿠스 로보스투스는 별로 변하지 않았음을 밝히고 있다. 오스트랄로피테쿠스는 사냥꾼인가? 남아프리카 일대에서 조사를 계속하던 다트는 1947 년 트란스발 부 근의 마카판스가트 농장 근처 동굴에서 부러전 동물들의 뼈와 함께 오스트랄로피데쿠스 아프리카누스의 화석들을 발굴하였다. 여기서 출 토된 동물들의 화석을 근거로 다트는 선사인류가 뼈와 이빨들을 연모. 로써 사용하였다고 결론 짓고 이룰 〈뼈 • 이빨 • 뿔의 문화 Bone • Too- th • Horn=Oste o donto kerati c Cul t ure 〉라고 이름 지었다• 다시 말해 서 다트는 이론시기의 인류는 동굴에서 살았으며 그들이 〈뼈 • 이빨 • 뿔의 문화〉의 기술을 가지고 있었던 사냥꾼들이었다고 주장하였다 (7. 13). 다윈의 〈피드백 이론〉의 영향을 받은 다트는 이른시기의 오스트 랄로피데쿠스는 이미 송곳니가 줄어들고 연모를 사용했다고 생각했 다. 그는 인류가 진화해 가는 중요한 원인은 바로 이 사냥행위에 의
해서라고 믿었다 . 이론시기의 인류가 사냥을 했으며 식인풍습 Carn ni bal i sm 이 있었다 는 가설의 근거는 스와르트크란스 동굴에서 찾아졌다 . 이 동굴에서 윗머리뼈 부분에 두 개의 구멍이 나 있는 머리뼈 조각이 발견되었는
 圈 -f 〈 · `之' : r广 크루 ·
圈 -f 〈 · `之' : r广 크루 ·
데, 다트는 이 구멍들을 이론시기 인류가 행한 살인의 증거로서 해석 하였다. 다트에 의해 많은 영향을 받았던 사람들 중의 하나로 『 아프 리카의 기 원 Af rica n Genes i s 』 를 쓴 아드레이 Ardre y를 꼽을 수 있다. 아드레이는 그의 책에서 타옹에서 발견된 머리뼈에 생긴 구멍에 대 해 〈어떤 동물도 이 구멍들을 뚫울 정도의 강력한 어금니를 갖고 있
지 않으므로 이 구멍들은 동물에 의해서 만들어지지 않았다 〉 고 설명 하고, 이어 말하길 〈 어떤 동물도 뼈둘을 주워 모으지 않는다. 따라서 동굴에 남은 뼈둘은 바로 인류의 행위에 의한 것이다. 〉 라고 기술하였 다. 그의 설명에 따르면 터옹에서 발견된 어린아이는 다른 사람들에 의해서 살해되었으며, 사람은 본능적으로 죽이려고 도구를 사용하는 살인본능을 가진 존재라고 단정 지었다. 한 걸음 더 나아가 다트는 오늘날 우리가 보는 〈폭력과 연모의 사용 〉 은 이미 이때부터 시작되었 다고 주장하며, 오스트랄로피데쿠스 아프리카누스를 연모를 사용하는 식육류로보았다 . 다트의 견해는 1950 년대에서부터 1970 년대 사이에 유행하던 인류 기원에 관한 해석인 〈사냥가설〉에 큰 영향을 미쳤다. 따라서 1960 년 대에 〈인류는 사냥꾼 Man the Hun t er 〉이란 견해가 인류학계의 주제 로 한때 유행했으며, 또한 〈연모 사용〉이라는 그의 설명은 우리 조상 울 진화의 길로 나아가게 한 요인으로 인식되어 한때 〈인류는 연모를 만드는 존재 Man the Tool Maker 〉라는 말로도 설명되 었다. 남아프리카에 있는 동굴들은 석회암지대에 만들어졌는데, 이는 남 중국의 카르스트 kars t 지형의 동굴들과 비슷하게 형성되었다. 많은 중국의 카르스트 Kars t 지형의 동굴들과 비슷하게 형성되었다. 많은 아 한데 엉켜 붙은 채로 발견되며 이렇게 만들어진 바위를 엉킨바위 Brecc i a 라고 부른다. 다트는 인류의 행위에 의하지 않고는 그렇게 많 은 수의 뼈둘이 동굴 안에 남아 있을 수 없으며, 인류를 제의한 다른 동물들은 뼈를 모으지 않는다고 단정지었다. 그러나 실제로 아프리카 사바나 지대나 요르단 사막에 사는 줄무늬 하이에나는 뼈를 물어 들여와 자신들이 살고 있는 곳에 모은다. 그리 고 모아전 뼈둘은 다트가 동굴에서 본 뼈화석처럼 깨어져 나갔는데, 이는 바로 하이에나 같은 식육류가 뼈둘을 씹어 생긴 결과이다. 1960 년대에 브레인은 스와르트크란스 동굴에서 발견된 머리뼈에 뚫

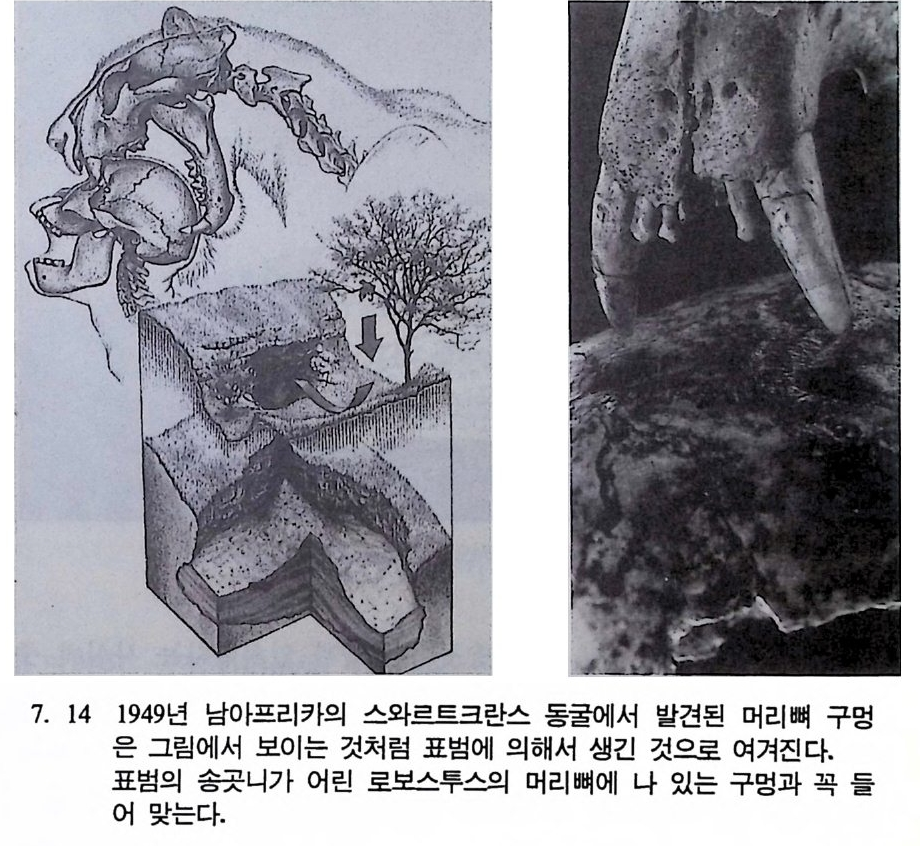 ~,·· ,鬱 ‘ > ·도., ,. ? , ;,,',· .` ·
~,·· ,鬱 ‘ > ·도., ,. ? , ;,,',· .` ·
 7. 15 복원된 남아프리카의 스와르트크란스 유적지
7. 15 복원된 남아프리카의 스와르트크란스 유적지
려 있는 구멍들이 표범의 아래 송곳니 모양과 일치한다는 사실에 주 목하였다. 브레인은 남아프리카의 석회암지대에 발달한 수직동굴의 형성과정을 연구한 끝에 동물화석들이 왜 동굴에서 되적되어 발견되 는지를 밝혀냈다(사전 7. 14). 이 석회암 동굴 내에는 갈라전 틈둘이
있는데 많은 동물들이 그 안에서 산다. 한편 동굴 밖 입구는 낮은 경 사를 이루고 있는 저지대로 나무들이 자라기에 좋은 환경을 만든다. 표범은 가끔 그들의 먹이룰 하이에나로부터 지키기 위해서 동굴 입 구에 자라고 있는 나무 위로 가지고 울라간다. 시간이 지나 먹이가 부패하면 그 뼈둘은 땅으로 떨어져서 동굴 안으로 쓸려 들어간다. 동 굴 바닥에 쌓인 뼈둘은 동굴 벽과 천장에서 떨어져 나간 바위 부스러 기와 밖에서 흘러 들어온 흙과 뒤섞인다. 수백만 년의 세월이 지나면 서 안에 뼈가 묻힌 채로 엉킨 석회암 엉킨바위가 되었다. 이것을 바 로 다트가 발굴한 것이다(그림 7. 15). 그러면 다트는 왜 그의 〈 뼈 • 이빨 • 뿔의 문화〉를 그렇게 강조했는 가? 다트도 다론 과학자들과 마찬가지로 자신의 야망과 편견을 가지 고 있었다 . 아마도 다트는 자신이 발견한 오스트랄로피데쿠스 아프리 카누스가 학계에서 받아들여지지 않자 이에 대한 실망과 학계에 대한 원망이 자신의 발견에 집착하게 하는 요인으로 작용했으리라 여겨진 다. 다트는 만일 그 자신이 발견한 화석의 주인공들이 연모를 사용했 다는 것을 증명할 수만 있다면 누구도 이 화석이 사람과 같은 선상에 위치한다는 것을 거부할 수 없다고 생각했다. 아마도 이갇은 이유 때 문에 다트는 자신의 이론울 강조했으리라 여겨지나, 그 전실은 알 수 없다. 중요한 것은 다트 자신에게 선임관이 존재했다는 사실이다. 동아프리카 호미니드의 발견 울드바이 골짝 Olduv ai Gorge : 오스트랄로피테쿠스 보이세이 Austr a / ,op ithe cus bois e i : 루이스 리키의 발견 1959 년에 인류화석에 대한 연구의 초점이 동아프리카로 옮겨졌다.
루이스 리키(1 903-1972) 는 선교사의 아들로 동아프리카에서 자랐다. 영국에서 학위를 끝낸 후 1930 년대에 인류의 조상을 찾기 위해 아프 리카로 돌아간 그는 탄자니아의 세렌게티 고원에 있는 올드바이 골짝 으로 갔다. 올드바이 골짝은 넓은 평원에 생긴, 약 40km 정도의 길이 에 90m 정도의 깊이룰 가지고 있는 골짝으로 골짝 단면은 과거에 흐 르던 강에 의해 찰려 나간 흔적이 남아 있다. 골짝 밑바닥에서 꼭대기까지는 네 개의 서로 다른 충으로 이루어져 있는 이 지역은 일찍이 1911 년에 빌헬름 카트빙클 W i lhelm Katt - wi nkel 과 1913 년 한스 렉 Hans Reck 이 이끄는 독일 탐험대에 의해 발견된 옛날 호수 바닥이었다. 한스 렉은 호수 바닥에서 호미니드 뼈 대 하나를 찾았다. 대다수의 사람들은 이 뼈대가 호수에 빠져 그 자 리에 묻혔을 뿐, 이 뼈대가 호수 바닥의 연대만큼 오래 되지는 않았 다고 생각했다. 그러나 이 바닥의 연대만큼이나 이 뼈대도 오래 되었 다고 믿는 한스 렉 등에 의해 설득된 리키는 그의 생각을 확인시켜 줄 증거를 찾고자 하였다. 아프리카로 돌아가야 한다는 리키의 꿈은 1931 년에 와서야 이루어 졌다. 아내 메리 리키와 어린 아들과 함께 올드바이로 돌아온 리키는 계곡 맨 밀충 Bed 1 에서 많은 동물 화석과 원시적인 돌연모를 찾아냈 다. 동물화석 중에 일부는 이미 절멸한 종이거나 아니면 학계에 소개 되지 않은 것들도 있었다. 그리고 리키가 찾은 돌연모들은 분명히 가 공된 것이었다. 그는 이들 연모를 그 지역의 이름을 따서 〈울도완 연 모 Oldowan Indus t r y〉라고 이름 지 었다. 고인류학사상 가장 끈질긴 조사가 올드바이 골짝에서 이루어졌다. 이 지역에서 조사를 시작한 지 28 년이라는 세월이 지난 1959 년 7 월 17 일 부인인 메리 리키가 골짝 맨 밀층 Bed 1 에서 호미니드의 머리뼈 한 개를 찾아냈다. 이 머리뼈가 바로 오스트랄로피테쿠스의 세번째 종인 오스트랄로피데쿠스 보이세이 Austr al op ithe cus bo i se i이다(사전 7.
16). 머리뼈는 사랑니가 마모되지 않은 점으로 보아 청년기 때 죽은 것으로 여겨지며, 비록 로보스두스의 이빨보다는 강전한 이빨을 지니 고 있지만 어금니의 형태와 넓적한 얼굴 모습 및 머리능선 등은 남아
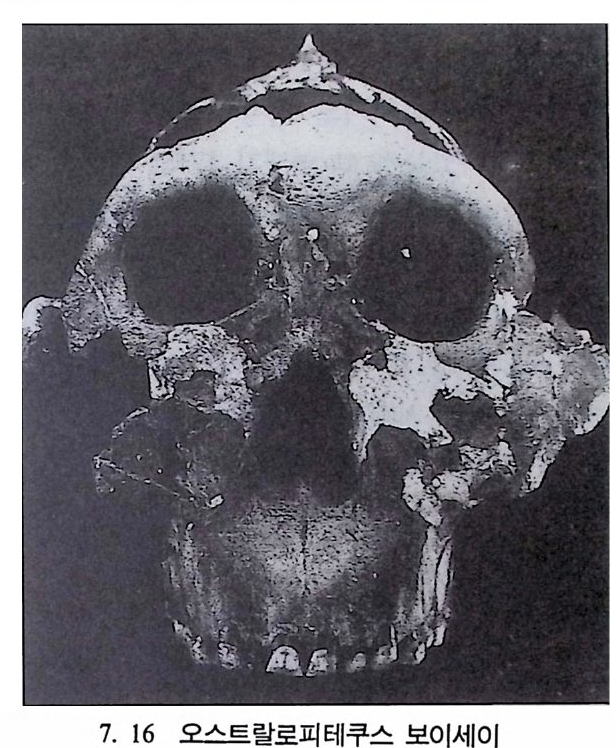 7. 16 오스트랄로피테쿠스 보이세이
7. 16 오스트랄로피테쿠스 보이세이
프리카에서 발견된 젊고 강건한 형의 호미니드와 매우 닮았다. 오스 트랄로피테쿠스 보이세이의 넓적한 얼굴은 씹는 운동과 관련이 있는 데, 이는 턱 모서리 힘살의 당김점이 머리 위와 앞쪽으로 이동하면 어금니를 가지고 음식을 씹는 데 편리하기 때문이다. 발견된 호미니드 화석은 남아프리카에서 발견된 강건한 형태의 화 석과 관계가 있었지만 리키는 이 화석에게 〈전잔트로푸스 보이세이 Zin z anth r op u s bo i se i〉라는 이름을 붙였으며 〈호도 까는 사람 Nu t Crack- er 〉이라고도 불린다. 전잔트로푸스라는 이름은 〈동아프리카 사람〉이
란 뜻으로, 리키는 이 호미니드가 동아프리카에서 기원한 인류의 직 접조상으로 믿었다. 올드바이 유적의 충위는 연대측정에 많은 문제를 지니고 있는 남아 프리카의 동굴 유적에 비해 옹회암이나 화산재 충으로 이루어져 있기 때문에 정확한 칼륨/아르곤과 고지자기 측정방법을 통해 제법 정확 한 연대측정이 가능하다. 호미니드 화석이 발견된 맨 밀충 Bed I 의 응회암울 가지고 측정하였는데 그 연대가 175 만 년으로 밝혀졌다. 이 로 인해 그때까지 알려졌던 인류기원의 연대가 100 만 년 이상이나 더 올라가게 되었다(그림 7. 17). 그러나 아마도 로보스두스처럼 거친 식물성 음식을 먹었으리라 여 겨지는 〈진즈〉는 〈동아프리카 사람〉이란 뜻보다는 〈호도 까는 사람〉 이란 뜻이 더 적절하다는 사실이 밝혀졌다. 나중에 발견된 증거들에 따라 고인류학계는 진잔트로푸스를 현생인류의 직접조상이 아닌 오스 트랄로피테쿠스의 한 종으로 분류하고 이를 오스트랄로피테쿠스 보이 세이로 고쳐 부르고 있다. 오스트랄로피데쿠스 보이세이는 1985 년 쿠비 포라에서 〈검은 머리 뼈〉로 불리는 오스트랄로피데쿠스 에티오피쿠스의 화석이 발견되기 전까지 남아프리카에서 살던 오스트랄로피데쿠스 로보스두스가 지리 적으로 떨어져서 생긴 변형으로 간주되었다. 아래에 보이는 도표들은 1925 년부터 1980 년대 초까지 발견된 화석 들과 이를 바탕으로 한 인류의 기원과 진화에 관한 고인류학계의 일 반적인 견해를 보여주고 있으며, 도표 7. 17 낡즌 1972 년 리키가 죽을 때까지 주장한 인류의 기원과 전화에 관한 의견이다.
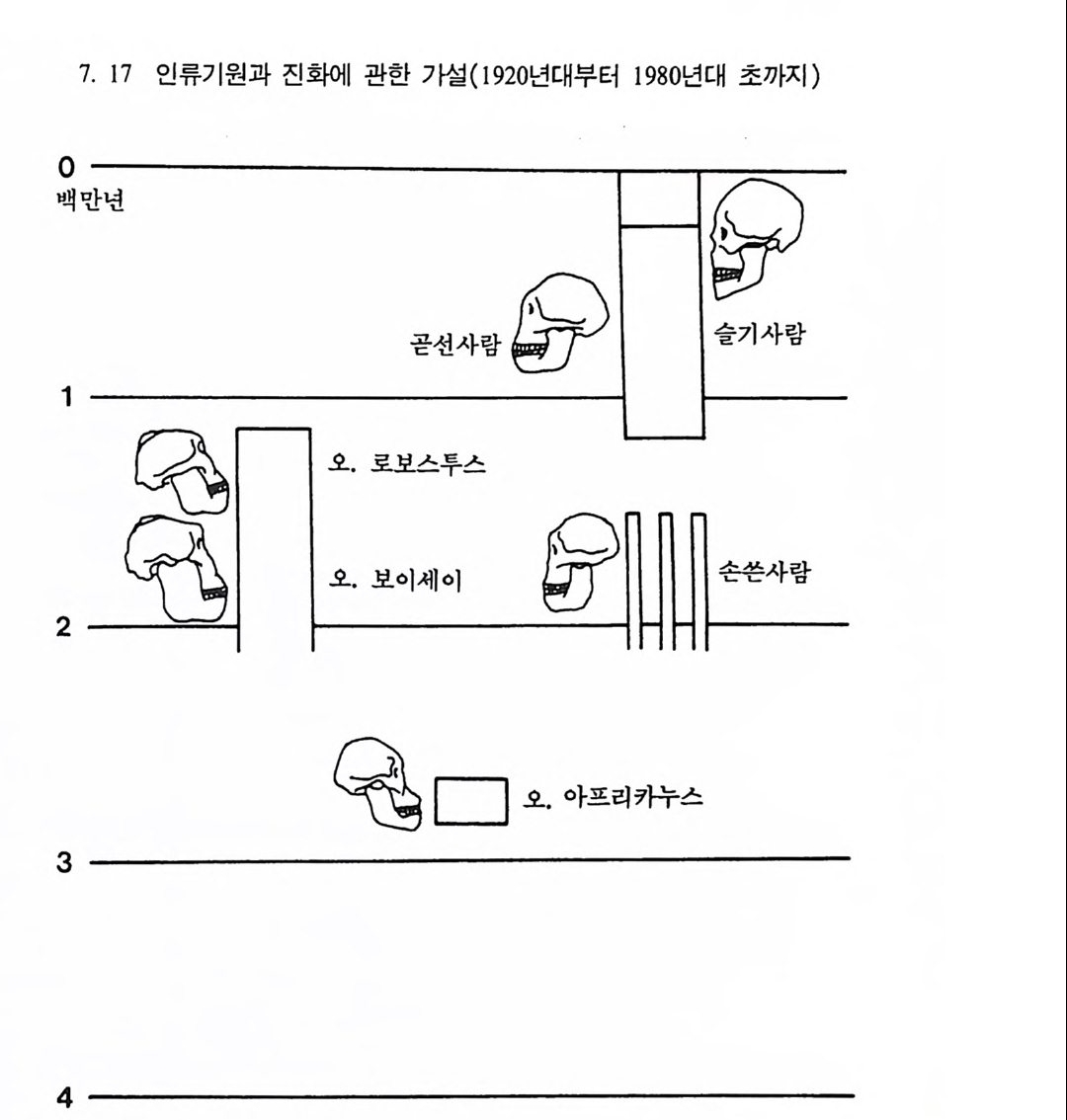 07 . 17 인류기원과 진화에 관한 가설 (1920 년대부터 1980 년대 초:까지)'
07 . 17 인류기원과 진화에 관한 가설 (1920 년대부터 1980 년대 초:까지)'
a. 1973 년까지 발견된 화석들이다. 이 그림에 따르면 3-200 만 년 사 이에는 오스트랄로피테쿠스 아프리카누스와 로보스투스의 화석은 별로 없으며 인류진화상 아프리카누스의 역할은 결정되지 않았다. 오스트랄로피테쿠스 보이세이는 로보스투스의 조상으로 알려졌으나 같은 시기에 살았던 손쓴사람과 곧선사람과의 관계는 밝혀지지 않 았다.
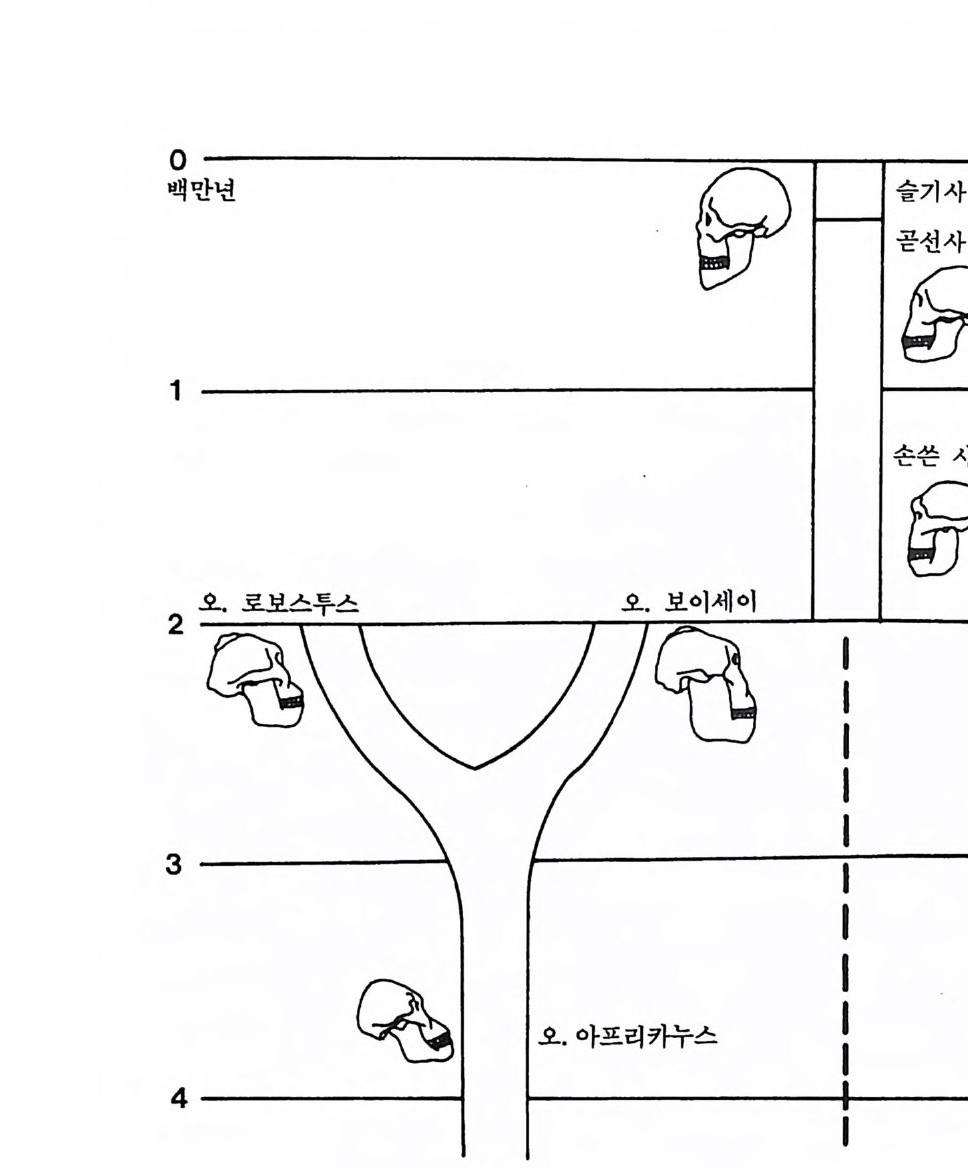 백0 만년:' 〔
백0 만년:' 〔
b. 루이스 리키(1 973) 의 가설. 이에 따르면 오스트랄로피테쿠스속은 인류진화 과정에서 호모속과는 아무런 관계가 없다고 하였다. 호모 속의 먼 조상은 호모속일 뿐이며 아직까지 이 호모의 화석이 발견 되지 않았다고 주장하였다. 오스트랄로피테쿠스의 화석은 3-200 만 년 사이에는 별로 나타나지 않았지만 이 호미니드가 로보스투스와 보이세이의 조상이었다고 주장하였다.
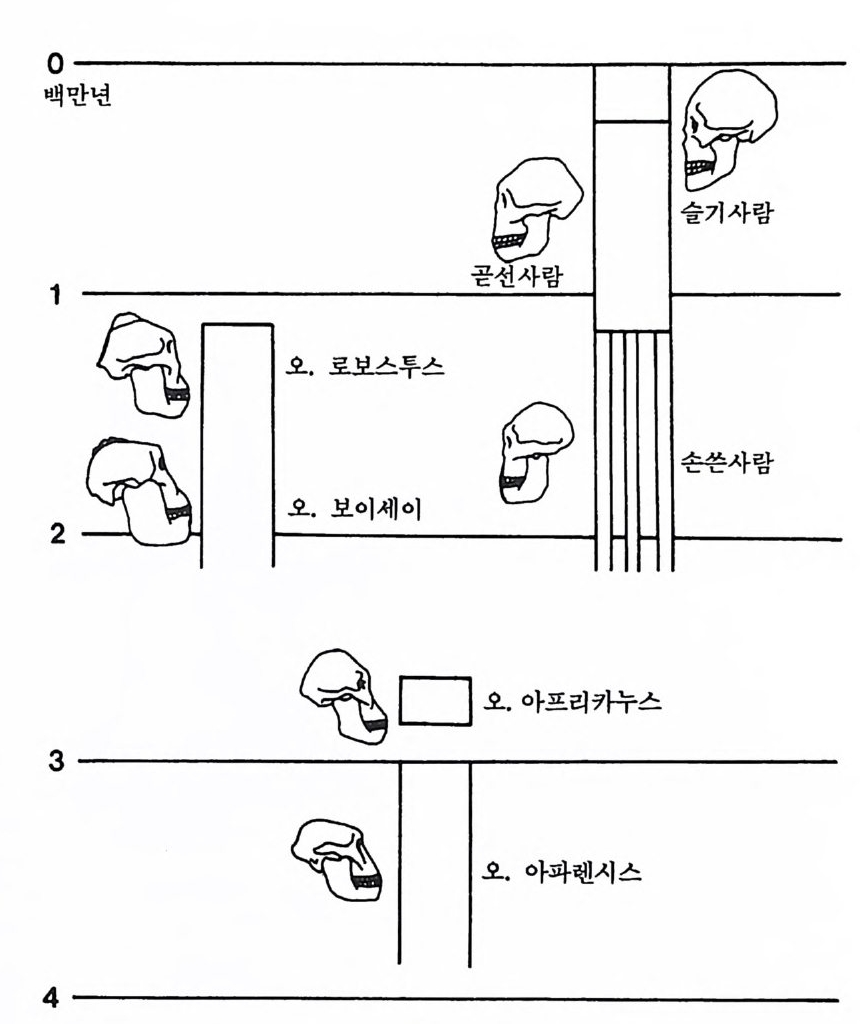 백。 만년 - -기 I~
백。 만년 - -기 I~
C.I 1978 년까지 발견된 호미니드 화석들에 근거하고 있다. 가장 오래 된 호미니드인 오스트랄로피테쿠스 아파렌시스(4- 300 만 년 전 )가 발견되었으나 아프리카누스와의 관계는 밝혀지지 않았다. 아프리카 누스의 역할 또한 밝혀지지 않았다. 더 많은 손쓴사람 화석들의 발 견으로 이 호미니드가 곧선사람의 조상이라는 주장이일반적으로 받아들여졌다.
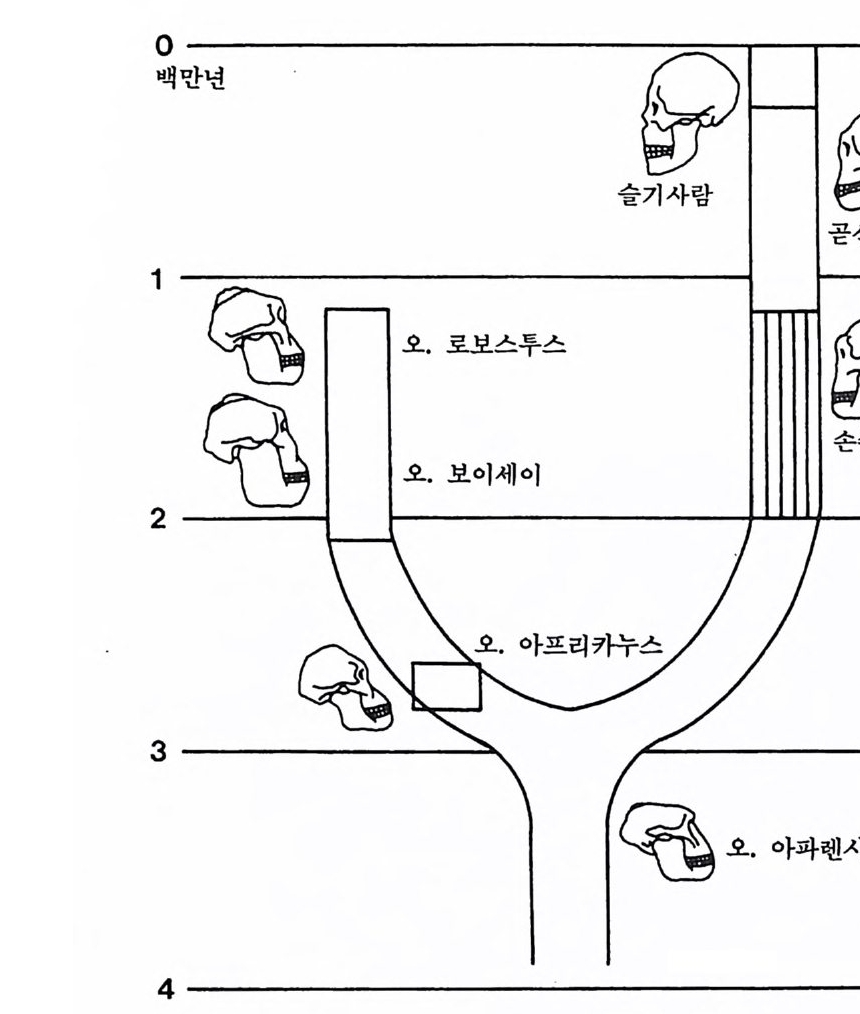 。백 만년 술E n g
。백 만년 술E n g
d. 1979 년에 들어와서 인류진화에 대한 새로운 이론이 주로 미국의 인류학자들에 의해 제기되었다.(.Jo hanson et al., 1979 : 1981). 이 들의 이론에 따르면 오스트랄로피테쿠스 아파렌시스가 손쓴사람의 직접조상이며 아프리카누스는 단지 보이세이와 로보스투스의 조상 일 뿐 호모속과는 진화상 아무런 관계가 없다고 한다.
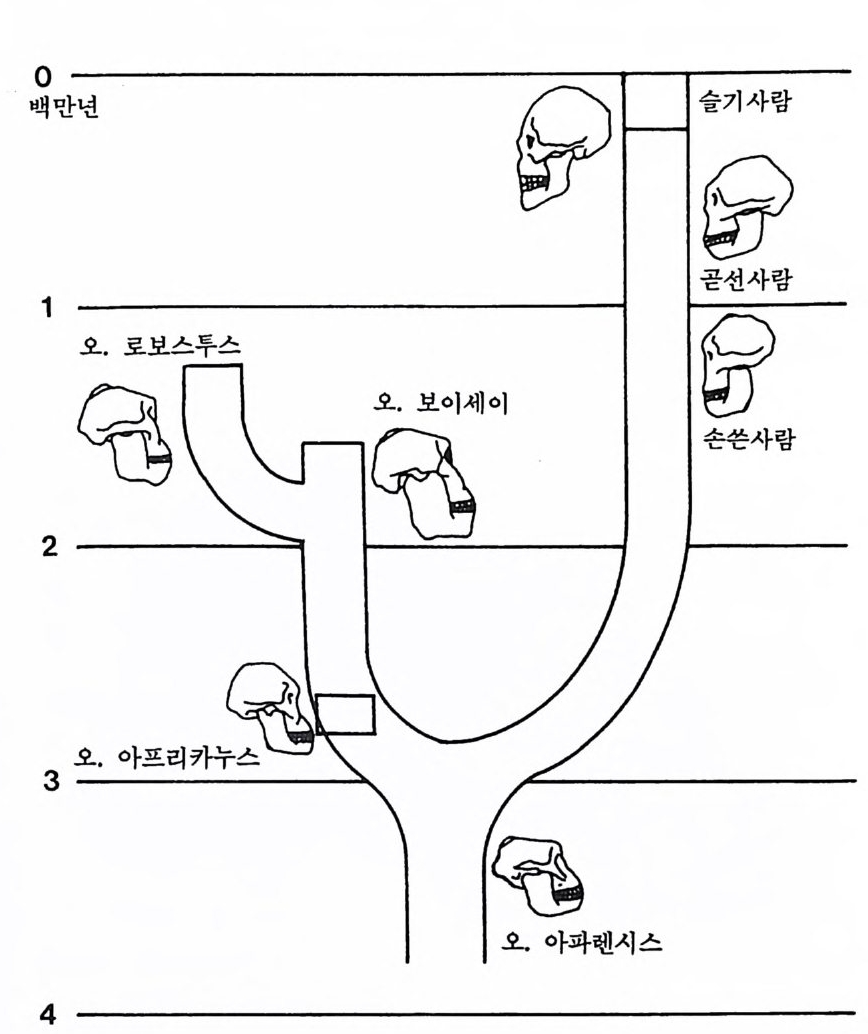 。 :; 낭
。 :; 낭
e. 1983 년에 발표된 요엘 락 Yael Rak 의 이론이다. 그는 오스트랄로 피테쿠스의 얼굴 구조 를 비교한 결과 오스트랄로피테쿠스 아파렌시 스롭 인류의 가장 먼 조상으로 받아들이며, 아프리카누스는 인류속 과는 아무런 관계가 없다고 주장한다. 7. 17 의 ct2j 이론과 기본 이 론은 같으나 단지 오스트랄로피테쿠스 보이세이와 로보스투스는 아 프리카누스로부터 갈라져 진화하였 E 는 · 점이 다르다.
두르카나 호수 서편 : 오스트랄로피테쿠스 에티오피쿠스 Aus t ralop ithe cus aeth i o p icu s 동아프리카와 남아프리카에서 발견되는 오스트랄로피테쿠스의 화석 울 조사할 때 가장 궁금한 문제 중의 하나는 오스트랄로피테쿠스 로 보스두스와 오스트랄로피데쿠스 보이세이에서 보여지는 커다란 어금 니들이 언제 나타났는지에 있다. 이에 대한 답은 1985 년 투르카나 호 수 서쪽에서 발견된 호미니드 화석에서 찾을 수 있었다.
 7. 18 오스트랄로피테쿠스 에티오피쿠스 (WT-17000) 머리뼈
7. 18 오스트랄로피테쿠스 에티오피쿠스 (WT-17000) 머리뼈
미국 볼티모어에 있는 존 홉킨스 대학교의 고동물학자인 패트 쉬프 만 Pat S hip man 은 남편 알렌 워 커 Alan Walker 와 리 처드 리 키 와 함 께 아프리카 북부 케냐의 두르카나 호수 서쪽 해안에 있는 로메퀴 일 대에서 발굴작업을 하고 있었다. 이 지역은 나무도 없는 황량한 퇴적 구릉이 호수가까지 이어진 곳이다. 1985 년 어느 여름날, 발굴단원인
카모야 Kamo y a 가 한 구릉에서 검은색을 떤 거의 완전하게 남겨진 호미니드 머리뼈를 찾아냈다. 이 머리뼈는 케냐 국립박물관 화석대장 에 KNM - WT - 17000 으로 등록되었으며, 일반적으로 〈 검은 머리뼈 Black skull 〉 로 알려져 있다(사전 7. I8).
표 7. 3 아프리카에서 발견된 오스트랄로피테쿠스와 유적지 (my a : 백만 년을 나타냄) 동아프리카 남아프리카 오 . 보이세이 (1. 3- 2.3 m y a ) 오 . 로보스투스( l.7m y a) 쿠비포라 스와르트크란스 올드바이 골짝 크롬드라 °l 오. 에티오피쿠스 ( 2. 6m y a) 오. 아프리카누스 (2- 3my a ) 오모 타웅 서 투르카나호수 스데르크폰테인 마카판스 7} 트 오. 아파렌시스 (3- 4 my a) 하다,라에몰리 미들아와쉬
이 새로운 호미니드 화석은 약 260 만 년 전에 살았던 것으로, 그 연대와 해부학상의 특칭으로 볼 때 이른시기의 오.아파렌시스와 오. 보이세이의 중간형으로 밝혀졌다. 이 화석의 머리뼈는 아파렌시스의 머리뼈처럼 머리 뒷부분에 마루가 있으며 또한 아파렌시스처럼 아래 턱이 튀어나와 있다. 반면에 어금니는 넓적하고 강건한 모양이 보이 세이의 어금니와 닮았다. 이런 특칭으로 볼 때 이 화석은 아파렌시스 와 보이세이의 완전한 진화상의 중간형이라 할 수 있으며, 미국 버클 리 대학교의 팀 화이트 등에 의해 오스트랄로피데쿠스 에티오피쿠스 A. ae t h i op i cus 로 분류되었다. 이 이름은 1930 년대에 프랑스 학자인 아
람버그 Arambor g가 이 지역에서 찾은 오스트랄로피테쿠스 화석의 아 래턱에 붙인 이름인데 , 화이트 등은 이들이 갇은 종에 속한다고 주장 한다. 리처드 리키는
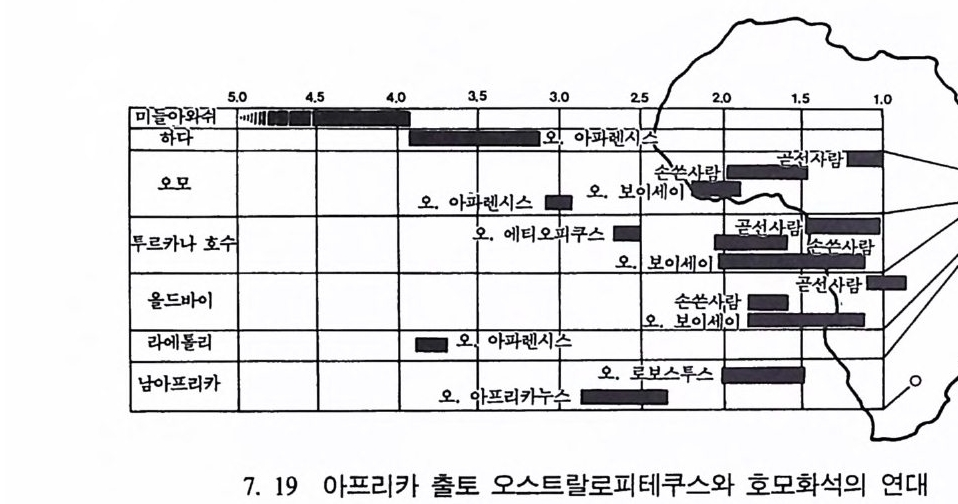 7. 19 아프리카 출토 오스트랄로피테쿠스와 호모화석의 연대
7. 19 아프리카 출토 오스트랄로피테쿠스와 호모화석의 연대
지금까지 아프리카에서 발견된 오스트랄로피데쿠스의 화석들을 살 펴보았다. 가장 이른시기의 인류 조상으로 알려진 오스트랄로피테쿠스 아파렌 시스의 머리뼈는 현생인류인 슬기사람의 머리뼈 모양보다는· 침팬지의 것에 가까우며 사지뼈둘은 인류를 닮았다. 이런 점에서 이 호미니드 는 침팬지와 인류의 특칭을 함께 지니고 있는 형태라고 말할 수 있 다.
오스트랄로피데쿠스 아파렌시스의 표본화석들은 다른 4 종의 오스트 랄로피데쿠스의 해부학상 기본 특징들을 모두 보여주고 있다. 이들은 모두 두 발로 걸었으며, 커다란 어금니와 줄어든 송곳니, 그리고 침팬 지 정도의 두뇌를 지니고 있고, 남자와 여자 사이는 몸의 크기에 따 른 차이가 있다. 오스트랄로피데쿠스들은 일반적인 특칭과 더불어 각기 종에 따른 차이가 있는데, 이빨과 머리뼈, 그리고 아래턱의 구조 등으로부터 구 별이 가능하다. 한 예로 가장 최초의 호미니드인 오스트랄로피데쿠스 아파렌시스는 침팬지와 가장 가까운· 모습을 지니고 있으며, 강건형의 호미니드들은 넓적한 얼굴을 하고 있다. 그러나 무엇보다도 가장 관 심을 끄는 호미니드는 가장 많이, 또는 가장 덜 특수화된 종들이다. 이런 관점에서 볼 때 오스트랄로피데쿠스 아파렌시스는 가장 덜 특 수화된 호미니드라고 할 수 있다. 같은 형태의 호미니드 화석들이 하 다와 라에톨리에서 발견되었는데, 이들이 이론 플라이오세 당시에 살 던 유일한 호미니드 형태였다고 여겨진다. 돌연모와 손질된 뼈들이 당시 유적지에서 발견되지 않았다는 점으로 그들은 연모를 사용할 줄 몰랐다고 생각되지만 각기 다른 환경 속에서 잘 적응해가며 살았던, 잡식성의 주워먹기짐승으로 알려져 있다. 한편 침팬지와 인류의 혼합 형태를 지닌 오스트랄로피데쿠스 아파 렌시스 같은 개체는 현대에 살고 있는 영장류나 분자상의 증거에 의 해서 단순히 예견될 수는 없고 유적 현장으로부터 얻어질 수밖에 없 다는 점을 인식해야 한다. 인류 기원에 관한 문제들 1980 년대 이후 고인류의 연구는 플라이오 • 플라이토세 (5-100 만 년 전)중 아프리카에는 5 종의 오스트랄로피테쿠스가 살았다고 밝히고 있
는데, 이들은 현생인류인 슬기사람과는 어떤 관계이며, 또 오스트랄로 피데쿠스 사이에는 서로 어떤 관계가 있는가에 대한 문제가 고인류학 계에 제기되었다. 인류과(오스트랄로피데쿠스속과 사람속을 포함)를 다론 영장류들과 구분할 때 적용되는 해부학상 가장 중요한 기준의 하나는 〈 두발걷기 〉 에 달려 있다. 1924 년 남아프리카의 타웅에서 오스트랄로피데쿠스 아 프리카누스가 발견된 이래로 1990 년대 초반까지 인류의 기원에 관한 많은 이론들이 있는데, 이 이론들은 크게 오스트랄로피테쿠스가 인류 의 칙접조상이라는 설과 우리의 먼 조상은 인류속g enus Homo 의 한 종으로 아직까지 그 조상의 화석이 발견되지 않았다는 설로 크게 나 누어진다. 리처드 리키 등을 대표로 하는 후자의 설은 그의 아버지인 루이스 리키의 이론을 전승하여 오스트랄로피테쿠스는 현생인류와 아무런 관 련이 없다고 주장한다. 그러나 오스트랄로피테쿠스 아파렌시스와 루 시 화석의 발견은 인류의 기원 문제에 있어 오스트랄로피데쿠스의 구 실에 커다란 논쟁을 불러일으켰다. 이들을 발굴하고 연구한 요한슨과 화이트 등은 인류 기원에 관한 리키의 지식은 정확하지 못하며 오스 트랄로피테쿠스 아파렌시스와 루시는 인류전화선상의 시작에 있다고 주장하며 리키 가족의 주장에 문제가 있음을 지적하고 있다. 1970 년대 말의 오스트랄로피데쿠스 아파렌시스 화석의 발견으로부 터 1980 년대 중반에 발견된 〈검은 머리뼈 WT-17000 〉 로 알려진 오 스트랄로피테쿠스 에티오피쿠스 화석의 발견은 인류의 기원에 관한 또 다른 문제를 제기하며 인류의 기원에 관한 논쟁을 학계에 불러일 으켰다. 10 여 년 동안의 격렬한 논쟁이 학계를 휩쓸고 지나간 후, 대 부분의 학자들은 4 - 500 만 년 전에 동아프리카에서 살았던 오스트랄 로피데쿠스 아파렌시스를 인류의 가장 이론조상으로 받아들이고 있 다 . 그러나 오스트랄로피테쿠스를 인류의 먼 조상으로 받아들일 때
다음과 같은 몇 가지 중요한 문제를 설명하여야 한다 ; 1. 오스트랄로피데쿠스 아파렌시스는 어떻게 걸었는가(방법 Loco moti on )? 2. 인류의 가장 이론조상으로 알려진 오스트랄로피테쿠스 아파렌시 스는 언제 지구상에 처음으로 나타났는가? 3. 에티오피아의 하다와 탄자니아의 라에톨리에서 발견된 오스트랄 로피데쿠스의 화석들은 서로 같은 종인가, 또는 다론 종인가? 4. 하다에서 발견된 다른 크기의 화석들은 무엇을 의미하는가? 5. 인류의 진화과정에서 오스트랄로피테쿠스 아프리카누스는 진화 계통상 무엇을 뜻하는가? 6. 〈 검은 머리뼈 WT-17000 〉로 알려진 오스트랄로피데쿠스 에티오 피쿠스는 인류진화상 어떤 구실을 하는가? 인류과에 속하는 호미니드와 다른 영장류를 구분하는 가장 중요한 기준은 두 발로 걷기에 있다고 앞에서 지적하였다. 따라서 호미니드 화석들 중에서 처음으로 두 발로 걸었던 호미니드 화석은 인류의 기 원으로 인정되며, 이런 점에서 하다에서 출토된 루시와 오스트랄로피 테쿠스 아파렌시스, 그리고 라에톨리에서 발자국을 남겨놓은 호미니 드가 두 발로 걸었는지를 살펴보기로 한다. 라에톨리에서 발견된 발자국을 루시의 발과 비교해 본 결과 서로가 아주 많이 닮았으며, 발의 구조는 이들이 두 발로 서서 걸을 수 있음 을 보여주고 있다. 또 다른 증거로 무릎구조를 들 수 있는데, 루시의 무릎 쪽에 있는 허벅지뼈 밑부분의 옆모양은 침팬지의 골반보다 사람 쪽에 더 가까우며, 아파렌시스의 무릎에 깊게 팬 홈 Groove 이 있으며, 사지뼈 중 허벅지뼈와 무릎이 이루는 각도도 침팬지보다는 사람 쪽에 가깝다. 또한 루시의 엉덩뼈는 침팬지의 것보다 사람의 엉덩뼈에 더 가까운 모습이다. 이런 특칭들이 바로 오스트랄로피데쿠스 아파렌시 스가 두 발로 걸었음을 보여주는 해부학상의 증거들이다(그림 7. 20).
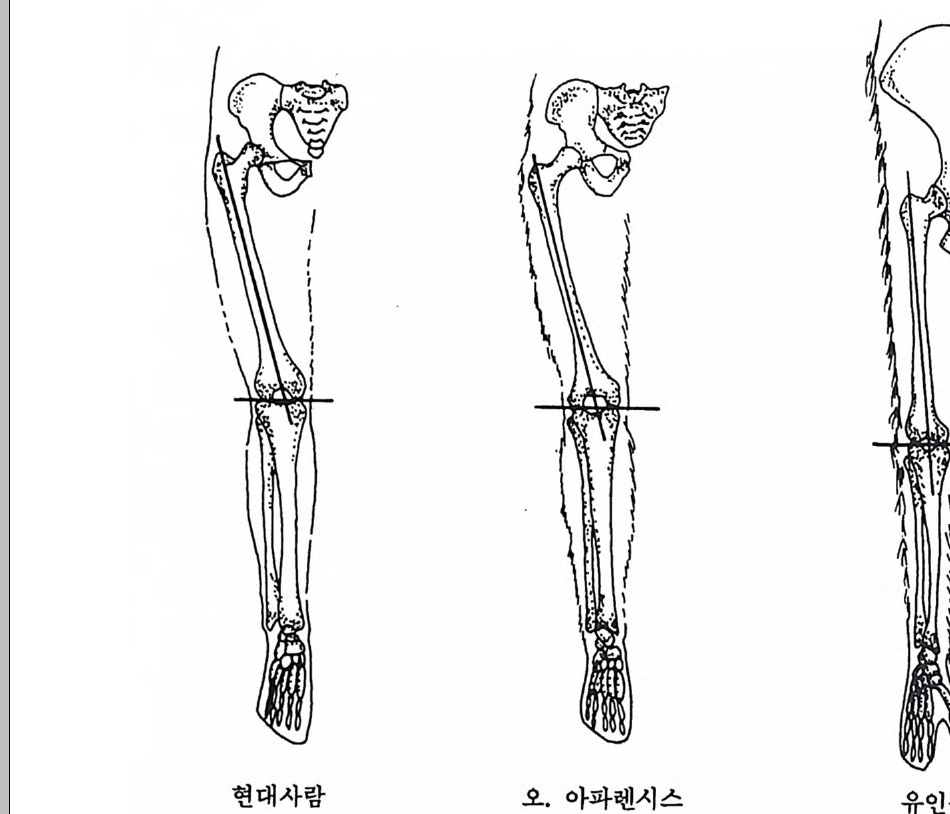 현대사람 오.아파렌시스 유인원
현대사람 오.아파렌시스 유인원
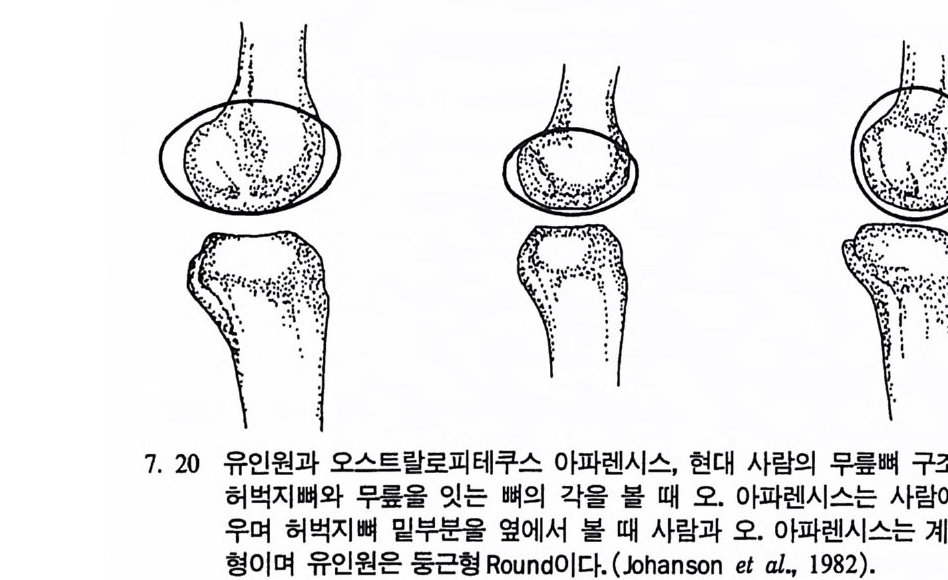 7\. 20 유우인며 원허과벅 지오스뼈 트밑랄부로분피율테 쿠옆스에 서아 파볼렌 시때스 사,람 현과대 오.사 람아의파 렌무시릎스뼈는 구 계조란 비Ov교al
7\. 20 유우인며 원허과벅 지오스뼈 트밑랄부로분피율테 쿠옆스에 서아 파볼렌 시때스 사,람 현과대 오.사 람아의파 렌무시릎스뼈는 구 계조란 비Ov교al
라에톨리와 하다에서 발견된 화석들의 연대는 각각 370 만 년과 330 만 년으로 알려져 있으며, 화석들의 뼈와 이빨들을 바교한 결과 이 두 지역은 수천 킬로미터가 떨어져 있어도 두 지역에서 발견된 화 석들은 갈은 종이며, 크기가 다론 호미니드의 화석들은 남자와 여자 사이에서 보이는 〈성별 차이 Sexual D i mor p h i sm 〉로 설명된다. 오스트랄로피테쿠스 아프리카누스의 인류진화상의 역할과 아파렌시 스와의 관계에 대해서 많은 논쟁이 있다. 1979 년 말 오스트랄로피테 쿠스 아파렌시스가 알려지기 전까지 아프리카누스는 인류의 직접조상 으로 알려져 왔었다. 오늘날에도 이 호미니드를 손쓴사람의 직접조상 으로 인정하고 아파렌시스를 아프리카누스의 아종으로 간주하고 있는 학자들도 있으며, 반면에 아프리카누스는 인류와 아무런 관계가 없고 아파렌시스룰 손쓴사람의 직접조상으로 간주하며 아프리카누스와 아 파렌시스를 각각 다른 종으로 주장하는 학자들도 있다. 오스트랄로피테쿠스를 연구할 때 중요한 과제 중의 하나는 언제부 터 커다란 어금니들이 강건형의 호미니드에게서 나타났는지에 있다. 리키팀이 1986 년에 동아프리카의 부르카나 호수 서쪽 지역에서 찾은 오스트랄로피테쿠스 에티오피쿠스로 이름 지어전 〈검은 머리뼈〉인 KNM-WT-17000 화석이 이 문제에 대한 해답을 주기에 충분한 해 부학상 특칭을 지니고 있다. 260 만 년 전이란 연대를 지닌 이 호미니 드의 머리뼈 화석은 오스트랄로피테쿠스 아파렌시스의 머리뼈처럼 윗 머리 뒷부분에 능선이 발달해 있으며 아래턱이 튀어나와 있으나 어금 니들은 오스트랄로피테쿠스 보이세이의 어금니를 닮았다. 이런 점에 서 이 머리뼈 화석은 전화상에서 볼 때 아파렌시스와 강건형 호미니 드의 중간형으로 알려져 있다. 위에서 살펴본 오스트랄로피테쿠스의 기원과 전화과정은 지금까지 알려진 여러 설들과 새로운 화석의 발견에 따른 해석 등을 종합해 볼 때 다음과 같은 세 개의 도표로 요약된다(그립 7. 21 ).
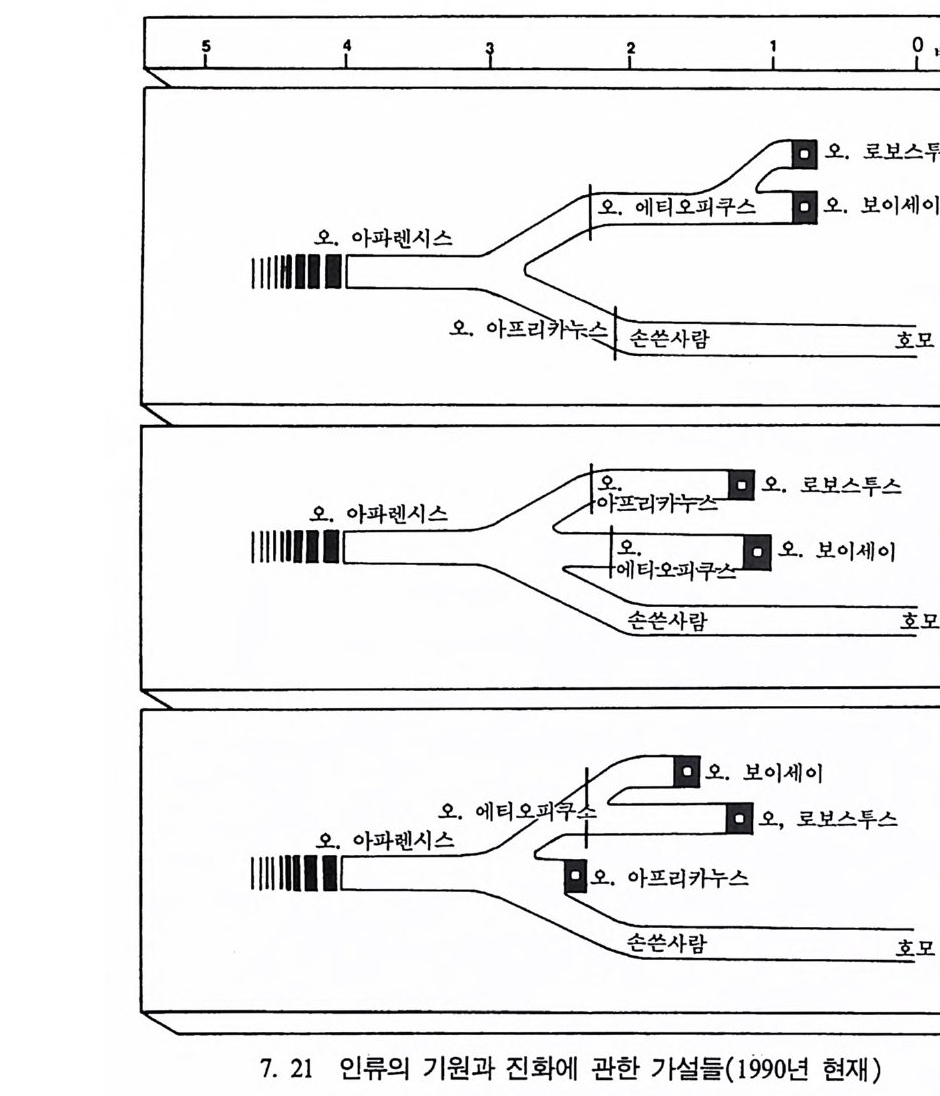 s 2 。 백만년
s 2 。 백만년
인류의 특징 : 〈두발걷기〉의 시작 오늘날 우리들은 인류의 전화에 관해 많은 자료와 지식을 가지고 있다. 그러나 홍미로운 사실은 인류전화에 대한 엄청난 자료와 지식
은 화석의 발견을 통해서 얻어전 것이 아니라 전화의 산물인 우리 자 신들과 오늘날 살고 있는 동물들로부터 얻어진 것들이다. 다윈이 인 류의 전화에 관해 거론할 때 당면했던 문제는 그가 살던 당시에는 인 류에 관한 화석기록이 없었다는 점이었다. 따라서 다윈이 알고 있던 지식은 살아 있는 유인원에 대한 해부학상의 특징들뿐이었으며, 유인 원에 관한 생리학이나 그들의 자연적인 행위 등은 알려져 있지 않았 거나 설사 알려진 지식이 있었다 하더라도 그것은 불확실한 지식이었 다. 다윈은 그 당시 자신이 알고 있던 해부학의 지식만을 가지고 과거 에 일어났던 일을 재구성하고자 노력하였다. 다윈은 인간과 침팬지롤 비롯한 유인원들을 관찰하고, 해부학상의 삼각측량 방법을 사용함으 로써 이들의 공통조상을 추론하였다. 그 결과 다윈은 인간을 땅에서 사는 유인원의 한 종류로 단정하였으며, 나아가 인간이 어떻게 땅 위 에서 살아 남을 수 있었는지, 또 어떻게 맹수들과 대항할 수 있었는 지를 규명하고자 하였다. 영장류들은 각기 나름대로 의부의 공격에 대항하는 방어수단을 가 지고 있다. 예로 어떤 영장류는 빠른 속도롤 가지고 맹수와 대항하며, 고릴라 같은 영장류는 커다란 몸집을 가지고 있는데 이 몸집 때문에 어떤 맹수들도 고릴라를 공격하려 하지 않는다. 비비 같은 영장류는 맹수의 공격에서 자신을 방어하기 위해서 복잡한 사회적 행위라든지 송곳니 같은 해부학상의 구조들을 발달시켰다. 그러나 인류는 다른 영장류가 가지고 있는 어떤 해부학상의 특칭도 지니고 있지 않다는 사실에 다윈은 주목했다. 그러나 인류는 다른 영장류들이 가지고 있 지 않은 한 가지 특칭, 죽 연모를 가지고 있다는 점에 착안해서 1871 년 다윈은 그가 본 인간울 진화적 방법을 가지고 비교 • 설명하고자 하였다.
〈 두발걷기〉에 관한 다윈의 시나리오 다윈은 인류가 두 발로 걷게 된 직접적인 이유를 연모의 사용 때문 이라고 설명한다. 그에 따르면 가장 이론시기에 살던 인류의 조상 중 남자들은 커다란 송곳니를 가지고 있었는데, 이들은 연모를 사용하게 되자 점점 송곳니가 줄어들고 턱을 덜 사용하게 되었다. 그들의 손과 팔은 걷는 데 사용되는 한 연모를 만드는 데 사용될 수 없었으므로 연모 사용의 필요성 때문에 두 발로 걷는 법이 발달되었으며, 연모를 사용하기 위해서 두뇌가 점점 커졌다고 가정하였다 . 그래서 다윈은 유인원과 인간의 다른 점을 연모를 만드는 기술의 소유라고 보고, 이에 따른 두 발로 걷기 발달과 그 결과 송곳니가 줄 어들고 머리가 발달했으며 이런 작용이 되풀이해서 일어났다고 생각 하였다. 다윈은 여기서 〈연모의 사용〉을 인류로 향한 핵심적인 전환 점으로 파악하고 있다. 이런 다윈의 전화에 대한 〈피드백 〉 시나리오 는 침팬지를 닮은 한 동물이 어떻게 인류로 전화되어갈 수 있었는지 를 설명하고 있다. 오늘날 우리는 우리 자신과 유인원들에 대해서 다윈 당시보다 더 많이 알고 있다. 인류 이의에 오늘날 살고 있는 유인원 중에 침팬지 도 초보의 도구 사용능력을 지니고 있다는 사실이 밝혀졌다. 그러나 침팬지는 여전히 네 발로 걷는다. 다윈은 침팬지가 도구를 사용한다 는 사실을 몰랐다. 그는 또한 돌연모는 두 발로 걷기가 시작된 한참 후인 250 만 년 전에 처음으로 만들어지기 시작했다는 사실을 알지 못 했다. 오늘날에 밝혀진 증거들은 다윈이 살았던 시대보다 더욱더 많이 축 적되었다. 가장 이론시기의 호미니드로 알려진 오스트랄로피데쿠스 아파렌시스는 이미 400 만 년 전에 두 발로 걸어 다녔다. 이는 두 발 로 걷기가 실제로 머리가 커지기 이전에 이미 시작되었음을 보여주고
있다. 에티오피아의 하다와 탄자니아의 라에톨리에서 발견된 호미니 드 화석들은 다윈의 시나리오가 잘못됐음을 밝혀주고 있다. 나머지 여러 가설들 인류의 조상이 왜 두 발로 걷게 되었는지를 설명해 주는 여러 가설 둘이 있는데, 그 대표되는 모델들 중의 하나로 〈유인원 도살자 가설 The Ki ller Ap e Theor y〉을 들 수 있다. 이 가설의 근본은 다윈과 같 은 논리로, 이 가설에 따르면 이론시기의 오스트랄로피데쿠스는 도살 자로서 이들은 동물을 죽이기에 필요한 연모를 쥐기 위해서 두 발로 걷게 되었다고한다. 다른 모델로는 〈정 찰 가설 The Reconnais s ance Theor y〉을 둘 수 있는데, 이 가설에 의하면 두 발로 걷기는 동물이 일어서서 풀을 넘 어 멀리 볼 수 있도록 하기 위해서 시작되었다고 한다. 그러나 이 가 설은 논리상 많은 문제를 가지고 있는데, 예로 만일 풀이 키를 넘는 경우에는 멀리 볼 수가 없으며, 또 풀이 초식동물에 의해 먹어 치워 졌다면 특별히 일어서서 볼 필요가 없다는 점이다. 보다 중요한 문제 는 180 여 종이 넘는 영장류들의 대다수는 가끔 땅 위에 내려와 일어 서서 단순히 풀 너머로 볼 수 있다는 점이다. 그들은 일어서기 위해 서, 그리고 주위를 둘러보기 위해서 특별히 두 발로 걷기를 발달시킬 필요가 없었다. 또 다른 모델로 〈과일 따기 가설 The Fru it Pic k i ng Theor y〉을 꼽 울 수 있다. 이 가설에 따르면 인류의 조상들이 나무에 열린 과일들 울 좀더 효과적으로 따기 위해 바로 일어서서 두 발로 걷게 되었다고 한다. 그러나 이 가설에서 주장하는 원인이 특별한 이유가 될 수 없 는 것이, 영장류들은 단순히 과일을 따기 위해서 나무 위로 기어오를 수 있다는점이다.
〈 딱딱한 열매 먹기 가설 The Seed Eati ng Model 〉은 영국의 인류학 자인 클리포드 졸리 Cli for d J oll y에 의해서 세워졌는데, 이 가설은 이 른시기의 호미니드와 침팬지에서 보이는 이빨과 턱 구조의 차이에 근 거를 두고 있다. 이른시기의 호미니드는 열매를 먹는 질라다 비비처 럼 음식을 먹는 중에는 똑바는 결과 두 발로 걷기가 발달되었다고한 다 . 그러나 중요한 문제는 질라다 비비가 여전히 두 발로 걷지 못하고 있다는사실이다. 지금까지 인류의 조상이 두 발로 걷게 된 이유를 설명하는 대표적 인 가설로 〈연모 사용 가설〉과 〈유인원 도살자 가설〉, 〈 정 찰 가 설〉, 〈과일 따기 가설〉 그리고 〈딱딱한 열매 먹기 가설 〉 등을 간단히 살펴보았는데, 이 중 어느 가설도 인류가 왜 두 발로 걷게 되었는지 를 설명하기에는 그 논리들의 타당성이 부족하다. 마이오세 말 • 플라이오세 초에 살았던 〈원 유인원 Pro t o-A p es 〉 을 두 발로 걷게 했던 요인이 무엇이었는지는 이들이 번성했던 마이오세 중기에 지구상에 일어난 변화를 살펴봄으로써 가능하다. 2000 만 년 전 아프리카 대륙에는 세 종류의 프로컨슬로부터 나온 많은 종류의 〈원 유인원〉들과 빅토리아피데쿠스 같은 몇 종류의 원숭 이들이 있었다. 그러나 당시에는 두 발로 걷는 동물은 물론, 침팬지와 고릴라 갇은 유인원도 없었다. 중기 마이오세(1 8 ― 1700 만 년 전)부터 1400 만 년 전으로 오면서 지구의 기온이 떨어져 추워지기 시작하며 환경이 변하였다. 지구상에는 메마론 지대가 증가하였고 지각의 변동 에 따라 지형의 모습이 변하였다. 이런 변화에 의해서 지구상에는 사 계절이 생겨나게 되었으며 그 결과 계절에 따라 음식이 변하였다. 이 시기에 살았던 호미노이드의 대다수는 어금니를 사용해서 음식 울 갇아 먹었다. 1000 만 년 전을 전후해서 호미노이드 중에 〈원 유인 원〉들은 계절화된 환경에 적응하지 못해 점차 다양성이 줄어들었으 며, 단지 한 종류의 〈원 유인원〉인 시바피테쿠스속이 새롭게 변한 사
바나 환경에 살아 남았다. 한편 계절화된 환경에 성공해서 적응한 구 대륙 원숭이들은 지구 도처에서 번성하였으며, 이 새로운 환경에서 시바피테쿠스 같은 〈 원 유인원 〉 들과 경쟁하였다. 400 만 년 전을 전후한 시기에 지금과는 다른 종류의 호미니드인 오 스트랄로피테쿠스 아파렌시스만이 사바나 환경에서 살았는데, 이들의 이빨과 턱은 침팬지롤 닮았으나 골반과 무릎 그리고 발의 모양들은 이 호미니드가 침팬지와는 다르게 두 발로 걸었음을 보여준다. 이 시 기에 다른 〈 원 유인원〉둘은 모두 절멸되었으나 오스트랄로피테쿠스 종류만이 살아 남는 데 성공해서 아프리카 대륙에 넓게 퍼져 서로 다 른 생활환경에서 살았다. 당시 어떤 기술이 있었는지는 알 수 없으나 머리뼈나 이빨 형태로 볼 때 이들은 아직도 마이오세에 살았던 〈원 유인원 〉 을 닮아 있다. 그러나 여러 종류의 오스트랄로피테쿠스는 다 른 마이오세 〈 원 유인원 〉 들에 비해서 날카로운 송곳니가 없다. 호미노이드의 생리상 • 해부학상의 특징 현생인류는 그들이 지구상에 퍼져 살고 있는 범위와, 생활환경에의 적응 그리고 인구의 증가라는 면에서 볼 때 전화상으로 아주 성공한 예이다. 이런 현생인류에 관한 생리학과 해부학의 연구성과는 인류진 화의 진화선상에 있던 다른 호미노이드와의 비교를 통해 전화과정의 여러 문제를 풀 수 있는 실마리를 제공한다(표 7. 4). 호미노이드를 비교할 때 먼저 생리학상의 특칭인 발정을 들 수 있 다. 발정이란 동물의 경우 암놈이 수놈에게 배란할 수 있음을 알리는 수단이다. 암놈이 발정기 때 암내를 풍겨 짝을 찾는 것은 적응도를 증가시키는 것이며 이 적응이야말로 전화의 기본이다. 동물들이 철멸하지 않고 살아 남으려면 어미의 유전자가 자식에게 로 전해져야 하며, 이때 유전자 전승은 암놈과 수놈 사이에 성적 결
합에 의해서만 가능하다. 이때 결합은 암놈의 배란시기에 일어난다. 문제는 침팬지 등의 유인원이 자신들의 배란기를 알고 있는지에 있 다. 실제로 침팬지 등은 배란기를 의식하지 않고 때가 되면 저절로 새끼를 가질 수 있게 되는데, 이때 침팬지는 엉덩이 뒷부분이 붉게 부풀어오르는 등 몸에 배란기를 알리는 증상이 나타난다.
표 7. 4 호미노이드의 생리상 • 해부학상의 차이 현생 유인원 시바피테쿠스 오스트랄로피테쿠스 현생인류 라마피테쿠스 아파렌시스 두걷뇌기 용량 4네00발cc ??큼?385두 一 발530 cc 1두400발cc 송곳니(남) 큼 ?작음 매우작음 기술 발달못함 ? 매우발달 의사전달 손짓 ·발짓 ? 말 음식 소얼량굴의표 육정식 ? ?. 30-40% 가 육식 나누어 먹기는 나누어 먹음 ?.? ? 거의 없음 . ?.? 남녀 결합 약함 . 강함 발정 있음 ?. . 숨겨진 배란
한편 호미노이드를 비교할 때 이용되는 해부학상의 특칭으로 두 발 로 걷기를 둘 수 있다. 침팬지와 사람은 둘 다 두 발로 걸을 수는 있 으나 실제 침팬지는 두 발로 걸을 때 금방 피곤울 느끼기 때문에 먼 거리를 두 발로 걸을 수가 없다. 그러나 두 발로 걸을 때와 네 발로 걸을 때에 소모되는 에너지는 대충 비슷하다. 하지만 두 발로 걷는다 는 것은 네 발로 걷기보다 달리는 속도와 민첩성이 떨어진다는 것을
뜻하며, 또한 몸의 균형감각도 감소하는 불리한 점을 지니고 있다. 이 렇게 두 발로 걸음으로써 속도와 민첩성, 그리고 몸의 균형 상실은 맹수에게 해를 입기가 그만큼 쉬워졌다는 것을 뜻하며, 두 발로 걷기 는 네 발로 걷기에 비해 몸의 무게중심이 등뼈와 무릎에 실려 그 결 과 이 부위에 문제를 가져왔다. 한편 영장류의 조상은 일찍이 전화과정중에 후각과 시각의 능력이 감소되었다. 그 결과 영장류의 일원인 사람은 맹수처럼 물체를 잘 보 거나 냄새를 맡을 수가 없으며, 동작은 느리고 또한 똑바로 서서 다 니기 때문에 맹수들의 눈에 찰 뜨인다. 이렇게 두 발로 걷는다는 것 은 맹수로부터 도망가는 데 별로 소용이 없을 뿐만 아니라 두 발로 걷기 그 자체는 무기로서도 사용되지 않는다. 물론 때때로 두 발로 걷기는 무기를 사용하는 데 효과적이긴 하지만……. 그러나 위에서 언급한 것처럼 네 발로 움직이는 동물들에 비해 여 러 불리한 점들이 있음에도 불구하고 인류가 〈두발걷기〉로 진화했다 면 분명히 두 발로 걷기에는 어떤 커다란 이점이 있음에 틀림없다. 여기서 인류의 조상이 전화과정중 두 발로 걸음으로써 얻어진 이점이 무엇인지를 이해하기 위해서 적응이라는 관점에서 살펴보는 것이 도 움이 된다. 오늘날 발달한 데모그래피 Demor gr a p h y란 학문은 살아 있는 생물 체의 출생 • 사망 • 질병 등 생명에 관계된 문제를 통계로 연구하는 분 야인데, 이 학문의 연구성과가 위에서 제기된 문제를 해결하는 중요 한 실마리를 제공한다. 이 학문의 성과가 인류의 조상으로 향한 호미 노이드의 전화를 아래와 같이 설명한다. 출산의 경우, 사람은 새로운 생명체를 출산하기 전에 38 주의 수태 기(임신)를 필요로 하며, 태어난 후 오랜 유아기와 어린 시절을 필요 로 한다. 한편 사람에 비해 짧은꼬리원숭이의 임신 • 출산 기간은 어 느 정도 빠른 편으로 6J\J l 면 새끼를 낳을 수 있다. 이에 반해 유인원
둘은 그 성장 • 발달과정이 원숭이보다는 사람에 더 가깝다. 고등영장류는 임신 • 출산 • 성장이라는- 생명의 진행단계가 원숭이 등의 영장류에 비해 느린 편이며, 출산 이후 성장기간이 길기 때문에 더욱더 많은 양의 부모의 에너지가 어린 새끼들을 돌보는 데 사용된 다. 한편 이들은 안정된 수의 집단의 유지를 위해서 최소한 2 마리의 새끼를 가져야 하며, 이렇게 적은 수의 새끼들을 돌보는 데는 많은 노력이 요구된다. 〈원 유인원〉은 자신들의 생명의 적응을 극대화하기 위해서 영장류 중 침팬지 같은 큰 유인원이 취하고 있는 적응전략을 취해 왔다. 오 늘날 살아 있는 침팬지들을 살펴보면 그들은 자식을 키우는 데 아주 많은 노력과 시간을 두자한다. 일반적으로 야생 동물의 경우 새끼들 간의 바람직한 평균 터울은 5-6 년으로 잡고 있는데, 침팬지의 경우가 이에 속한다. 침팬지 새끼들은 4-~l 까지 어미의 보살핌을 받고 5 세 까지 어미의 등을 타고 돌아다닌다. 유인원은 10 세가 되면 어른이 되는데, 만일 암놈이 처음 임신을 하 고 다시 임신을 하기까지는 5-6 년의 시간이 필요하다. 따라서 최소 2 마리의 새끼가 다 자라서 독립을 할 때가 되면 어미의 나이는 21 세 가 된다. 만일 지금까지 언급한 과정이 일정대로만 진행된다면 침팬 지 동의 큰 영장류는 2 마리의 새끼를 잃는 경우가 없다. 유인원에 비 해 원숭이의 경우 2 마리의 새끼가 출산해서 어론이 되는 이 모든 일 정은 9 년이란 세월을 필요로 하는데, 이 점이 바로 원숭이가 새로운 환경에서 잘 적응하여 번성하는 원인이다. 침팬지의 경우, 점점 늘어나는 유아기간은 어미 침팬지에게 그만큼 위험부담을 증가시켜주는 것이다. 이 증가된 위험을 극복하기 위해서 유인원들은 사회전략을 세운다. 예를 들어 지능이라든지 사회구조 및 배움 등을 발전시켜왔다. 이런 방법을 통해 유인원들은 자신들의 사 망률을 가능한 한 줄여왔다. 사망률이 감소하고 이에 따라 수명이 연
장되는데, 이렇게 수명이 연장된다는 것은 생명을 유지하는 데 필요 한 사회기술들의 발달이 증가했다는 것을 뜻하며, 여기에 피드백 작 용이 성립된다. 오늘날 살고 있는 침팬지가 취하고 있는 위와 같은 적응전략을 마 이오세에 살던 〈 원 유인원 〉 들도 취했으라고 생각되며, 중기 마이오세 까지 이 전략은 매우 성공하였다 . 중기 마이오세까지 구대륙 원숭이 와 〈 원 유인원〉들 모두에게 이로운 환경과 생활조건이 조성되었다 . 이 당시까지 〈 원 유인원〉은 환경에 찰 적응하여 널리 번성하였으나, 중기 마이오세를 경계로 이들은 후기 마이오세의 계철화된 새로운 환 경에 적응하는 데 실패하였다. 후기 마이오세부터 초기 풀라이오세에 걸쳐 〈 원 유인원〉들의 화석 이 원숭이들의 화석에 비해 아주 적게 발견되는데, 이는 실제로 이 기간중에 〈 원 유인원 〉 들이 별로 많지 않았기 때문이다. 이에 비해 구 대륙 원숭이는 아프리카, 남아시아, 동남아시아, 동아시아 그리고 더 나아가 일본에까지 널리 퍼져 있다. 이렇게 〈원 유인원〉들은 전화 초 기에는 아주 번성했었으나 지금은 사라지고 없으며, 이들의 후예인 유인원들도 지금 살고 있는 열대 밀림 등에서도 사라져가고 있다. 전화론자들은 생명체가 새로운 환경으로 퍼져 나가기 위해서는 같 은 〈 종 〉 안에서 경쟁을 거쳐야 한다고 말한다 . 〈원 유인원〉의 한 종 류가 오스트랄로피데쿠스로 전화해 가는 과정에서 이들은 · 빠른 속도 로 번식하는 원숭이들과 경쟁하기 위해서 자식을 빨리 가져야 하며, 생존성을.늘려야 하는 데모그래피상의 문제를 해결하여야 했다. 그러 나 〈원 유인원〉에서 갈라져 나간 대다수의 유인원들은 생리학상으로 그들의 오랜 생활유형을 고수하고 있어, 이 딜레마를 풀 수가 없었 다.
러 브조이 의 모델 미국 켄트 대학교의 해부학자이자 고인류학자인 러브조이 C.Owen Love j o y는 400 만 년 전 호미니드인 오스트랄로피데쿠스 아파렌시스 화석을 연구한 후 아파렌시스가 어떻게 이 딜레마를 해결했는지에 관 한 모델을 제시하였다. 그는 자신의 모델에서 〈 생산율을 증가시키기 위해서 호미니드의 조상들은 자식들 사이의 출산 간격울 줄였는데, 이는 우리의 조상들이 자식을 더 많이 낳았다는 것을 뜻하며, 이런 전략은 원숭이와 경쟁하는 데 더 효과적이다 〉 라고 가정하였다. 만일 어미가 새끼를 낳은 후에 곧 임신을 할 수 있다면 그 어미는 더 많은 자식을 가질 수 있다. 한편 계절이 변함으로써 식량자원 획 득에 제한을 받게 됨에 따라 〈원 유인원〉둘은 식량을 얻기 위해서 더 멀리 여행할 수밖에 없었다 . 암놈 어미는 출산 간격이 좁게 두 마 리의 자식을 가짐으로 인해서 행동의 제약을 받았으나 어미 자신과 새끼를 먹여 살리기 위해 먼 거리를 여행할 수밖에 없었다. 러브조이는 〈원 유인원〉이 당면했던 문제-죽 나이 차이가 별로 없는 2 마리의 새끼를 데리고 식량을 얻기 위해 먼 거리롤 여행해야 하는 일 __ 는 암놈의
어쨌든 이런 일은 수놈이 자기가 먹이를 가져다 주는 새끼가 자신 의 새끼라는 것을 모르는 경우에는 일어날 수 없었을 것이다. 여기서 우리는 수놈이 자기가 아비라는 사실을 알고 했는지에 대해서 말하고 자 하는 것이 아니라 수놈의 적응을 강하게 하는· 유인원 사이에서의 결합에 대해 말하고자 한다. 늘어난 성의 수용력(감수성)에 의해서 교미는 오랜 기간에 걸쳐서 일어날 수 있었으며 수놈은 새끼들이 자 신의 새끼들이라는 인식이 늘어났다고 러브조이는 가정하였다. 한편 암놈들은 수놈들 중에 가장 멀리서도 많은 식량을 가져올 수 있는 수놈을 선호하였을 것이며, 이때 두 발로 걷는 놈이 네 발로 걷 는 놈보다 더 많은 먹이를 나를 수 있었으라고 가정하였다. 따라서 두 발로 걸을 수 있는 수놈의 유전자만이 다음 세대로 전해질 기회가 주어졌으며, 많은 세대가 지나면서 두 발로 걷기는 더욱 완전해졌다. 이렇게 인류의 조상이 왜 두 발로 걷게 되었는지를 러브조이는 설명 하고 있다. 또한 하나의 암놈에 자기의 자식으로 구성된 일부일처의 사회조직은 집단 내에서 짝을 얻고자 하는 공격적인 노력을 감소시켰 울 것이고 수놈은 짝을 찾고자 하는 걱정 없이 식량을 찾으러 떠날 수 있었을 것이다. 이런 일들이 〈원 유인원〉의 원숭이에 대한 경쟁력 울증가시켰울것이다. 위와 같은 러브조이의 가설은 다윈의 이론에 반대되는 설로 그의 가설은 데모그래피의 연구성과와 400 만 년 전에 살았던 호미니드의 화석자료에 근거를 하고 있다. 로드만·맥켄리의 모델 한편 최근에 두 발로 걷기에 관한 중요한 연구가 캘리포니아 대학 교의 인류학자인 피터 로드만과 헨리 맥켄리에 의해 전행되었다. 그 들은 〈호미니드의 두 발로 걷기와 네 발로 걷기를 구분하는 데 에너
지가 철대적인 기준은 아니다. 〉 라고 말하며 인류의 두 발로 걷기는 침팬지가 두 발로 걷는 것보다 더 효과적이라고 지적하고 있다. 따라 서 두 발로 움직임으로써 이점이 있었다면 인류의 조상은 침팬지처럼 걷기보다는 인류처럼 두 발로 걷기로의 전환이 진화상에서 유리하였 을 것이라고주장한다. 과연 5-600 만 년 전에 두 발로 걷기는 어떤 점에서 유리하였을까? 로드만과 맥켄리에 따르면 중기 마이오세 이후 변화된 자연환경에 적 응하기 위해서 두 발로 걷기로의 전환이 일어났다고 주장한다. 다시 말해서 삼림지대가 드문드문 흩어져 있고 초원에 가끔· 나무들이 자라 는 환경으로 대체되자 〈원 유인원〉들은 식량을 얻고자 전보다 더 돌 아다녀야 했다. 이때 경제적으로 효과적인 걷기 방법, 두 발로 걷기가 이런 사바나 지역을 돌아다니기에 유리했다고 한다. 따라서 호미니드 의 두 발 걷기란 〈‘원 유인원’이 살 수 없었던 곳에서 이들이 살아가 기 위한 방법〉이라고 로드만과 맥켄리는 간략히 말한다 . 이들의 가설을 요약하면 두 발로 걷기는 식량의 운반이나 연모제작 의 결과, 또는 식성의 변화 때문이 아니라는 것이다. 두 발로 걷기는 단순히 〈원 유인원〉들이 자신들이 먹던 식량을 제공하던 환경이 변하 자, 이 변화에 맞추어 여기저기 돌아다니기에 좀더 효과적인 방법으 로 생태상의 적응에 따라 두 발로 걷게 되었을 뿐이라는 것이다. 지금까지 인류의 조상이 언제부터, 왜 두 발로 걷게 되었는지를 설 명하는 여러 모델들을 살펴보았다. 그러나 무엇보다 중요한 것은 어 떤 가설이 더 과학적인가를 검증할 수 있어야 하며, 이 과학적인 설 명은 증거에 의해서만 가능하다. 오늘날 우리는 인류의 기원에 관한 많은 증거둘을 가지고 있다. 다윈은 적은 증거를 가지고 연모의 사용에서부터 시작되는 피드백 시나리오를 만들어냈다. 다윈의 가설은 당시의 증거만으로는 훌륭했 으나 생리상이나 행위에 관한 연구가 이루어지지 않았으며 또한 고생
물상의 자료가 부족해서 그가 이용한 증거둘은 충분하지 못하였다. 동시에 그의 가설은 지난 세기에 발견된 호미니드 화석들에 의해 설 득력을 잃게 되었다. 이런 점에서 러브조이와 로드만 • 맥켄리의 가설은 이룰 검증할 수 있다는 점에서 매우 유용하다. 그러나 우리가 한 가지 염두에 두어야 할 점은, 이들의 이론은 현대 사회가 아닌, 우리들로부터 아주· 멀리 떨어진 몇백만 년 전의 일들에 적용된디는· 점이다. 그들의 가설은 과 거의 환경을 여러 각도에서 생각하게 하여 진화를 좀더 종합된 방법 으로 살펴볼 수 있게 한다. 이런 관점에서 우리가 인류의 전화를 비 교영장류학에만 근거하여 연구하는 것은 지양해야 할 일이다.
 제 8 장 인류의 전화 II •
제 8 장 인류의 전화 II •
제 8 장 인류의 전화 II : 호모 Homo 의 출현 손쓴사람 Homo hab ili s 의 출현 손쓴사람 화석의 발견 올드바이 골짝 메리 리키가 1959 년 올드바이 골짝에서 전잔트로푸스의 머리뼈를 발견한 지 얼마 지나지 않아 루이스 리키의 큰아들인 조나단은 이 유 적의 오래 된 아래충에서 〈전즈 Z i nz 〉보다 섬세한 형태의 호미니드 머리뼈 부분과 턱 및 손뼈들을 찾았다. 1960 년에 발견된 이 화석들은 오스트랄로피데쿠스 보이세이와 같은 충에서 발견되었지만 보이세이 와는 다른 호미니드 화석들이었다. 루이스 리키는 이 화석들이 우리 자신이 속해 있는 인류속g enus Homo 중 가장 이른시기에 속하는 것 이라고 믿었다. 1964 년에 루이스는 새로운 호미니드 화석들을 찾아 이론시기에 속하는 인류화석을 손쓴사람 Homo habil is =handy man 이라고 이름 지었는데, 1968 년까지 모두 4 개체의 손쓴사람 화석이 발 견되었다(표 8. 1). 올드바이 골짝에서 새로 찾아전 호미니드인 손쓴사람에 대해 고인 류학자들 사이에서 대단한 논쟁이 일어나게 되었다. 왜냐하면 화석들
이 매우 작은 조각들로 되어 있었으며, 또한 발달이 덜 된 머리 모양 과 작은 머리뼈를 가지고 있었기 때문이었다. 머리뼈의 모양은 오스 트랄로피테쿠스에 비해 눈뒤줄임이 약하며(그립 8. l) 이빨의 형태도 앞니는 어금니에 비해 상대적으로 커지고 옆니는 작아졌다.
표 8. l 1964-1968 년 사이에 올드바이에서 발견된 손쓴사람 화석 OH 7 : 부서진 머리뼈 조각과 뒤틀린 어린아이 아래턱 OH 13( 신데렐라 C i nderella) :머리뼈 일부와 완전한 위 • 아래 이 빨들 OH 16( 올드바이 꼴짝) : 여러 조각의 머리뼈 부분 OH 24( 트위기 Tw iggy) : 여러 조각 난 머리뼈
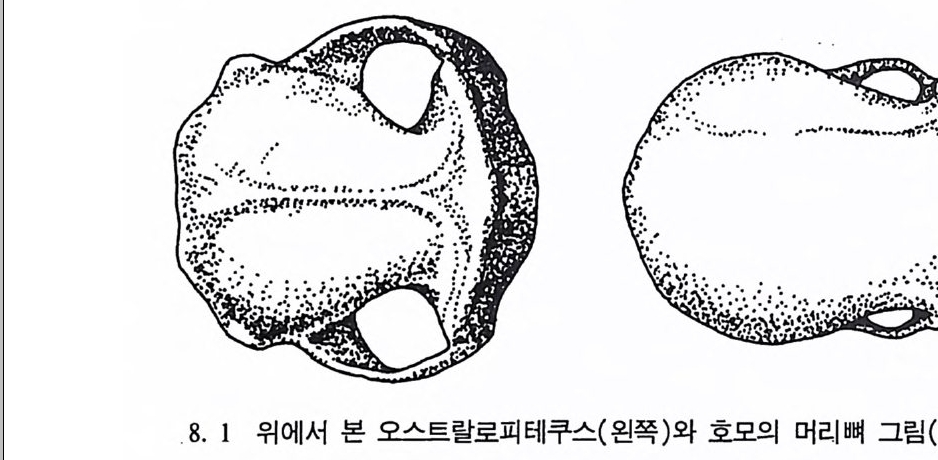 .... .
.... .
그러나 많은 학자들은 이 화석을 새로운 인류의 한 종으로 받아들 이기를 거부하였다. 루이스 리키가 〈기준 화석〉으로 정한 화석의 머 리 용량은 657c& 알려졌으며, 다른 손쓴사람의 머리뼈는 토바이아 스 P hilip V. Tob i as 와 홀로웨이 Ralph Hollowa y에 의해 평균 642cc
(600 - 684cc) 로 측정되었다. 올드바이 골짝에서 발견된 이 손쓴사람 머리뼈의 크기는 인류라고 부르기 위해서는 최소한 머리 크기가 700cc 이상은 되어야 한다는 클라크 S i r W ilf r id Le Gros Clark 의 최 소한의 크기 한계에도 미치지 못한다(현생인류인 슬기사람은 1200 - 1600cc 사이에 분포한다)(표 8. 2). 따라서 많은 학자들은 이 화석을 오스트랄로피테쿠스 아프리카누스 의 전화된 형태라고 생각했다. 그러나 실제로 손쓴사람의 이빨과 턱 그리고 얼굴 들은 강건한 오스트랄로피데쿠스에서 보이는 것처럼 특 수화되지 않았으며, 특히 오스트랄로피테쿠스와의 차이는 머리뼈 크 기에 있는데 이 호미니드의 머리는 몸에 비해 점차 팽창하기 시작했 다(그림 8. 2).
표 8. 2 머리 용량의 비교 머리뼈 용량의 범위 평균 용량 (cc) 쥐여우원숭이 10- 7 0 침팬지 282-500 383 고릴라 340-7 52 505 오랑우탄 276-540 405 오스트랄로피테쿠스 아파렌시스 385-450 440 오.아프리카누스 428-500 450 오 . 에티오피쿠스 410 오로보스두스 500 오.보이세이 506-5 30 450 손쓴사람 Homo habil is 509-752 632 곧선사람 Homo erectu s 727-1225 930 슬기 사람 Homo sap ien s 1000-2000 1350 *자료는 Tobia s , I972. 9.}- Cron in et. al., I981 에서 인용•
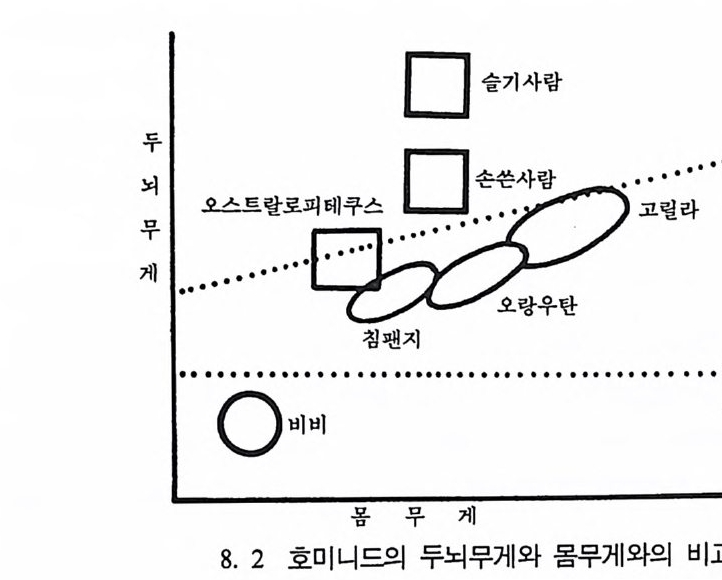 口슬기사람
口슬기사람
올드바이 골짝에서 발견된 손쓴사람 화석들은 185 만 년 전부터 160 만 년 전 사이의 연대를 가지고 있는데, 이 연대는 바로 〈전즈〉 가 살았던 시기와 일치한다. 올드바이 화석의 발견은 루이스 리키에 게 두 가지 중요한 생각의 근거를 제공하였다. 첫째는 손쓴사람이 일 찍이 올드바이에서 연모를 사용했다고 생각하여 같은 충에서 발견된 〈전즈〉는 손쓴사람의 먹이였으리라고 생각했다는 점이며, 둘째는 루 이스 리키가 손쓴사람 화석을 증거로 해서 오스트랄로피테쿠스는 절 대로 인류의 조상이 아니라는 주장을 내세웠다는 것이다. 올드바이 골짝에서 출토된 화석들이 오스트랄로피테쿠스의 한 종류 인지 아니면 인류속에 속하는 호미니드인지에 관한 오랜 논쟁은 1972 년 두르카나 호수에서 KNM ER-1470 으로 알려진 머리뼈가 발견되 어 루이스 리키의 손쓴사람이란 이름이 옳았다는 사실이 밝혀질 때까 지 계속되었다• 올드바이 골짝에서의 루이스 리키의 연구는 라에톨리 와 하다 그리고 두르카나 호수 주위에 있는 오모와 쿠비 포라 지역의 호미니드 발굴의 기폭제가 되었다.
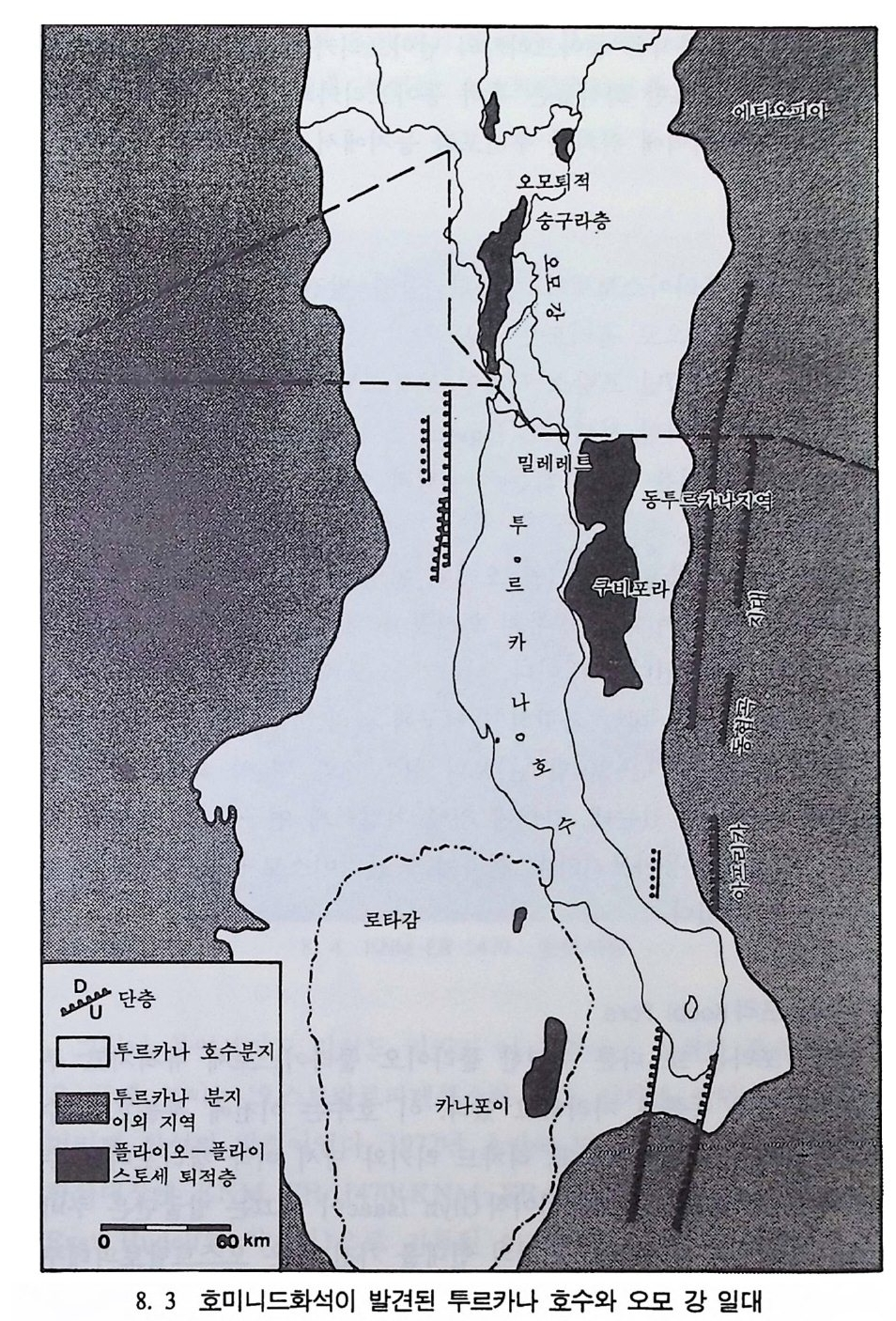 쇼단충 l('',,、,..I ',. I . J’' -로 I.타/ '감-\ r, < · -.· `· , .`- `·, \`.’. \. ., `.. ,,
쇼단충 l('',,、,..I ',. I . J’' -로 I.타/ '감-\ r, < · -.· `· , .`- `·, \`.’. \. ., `.. ,,
손쓴사람의 화석은 동아프리카와 남아프리카의 여러 유적지에서 발 견되었는데, 중요한 화석들은 특히 동아프리카의 오모 지역과 두르카 나 호수 동쪽지역에 위치한 쿠비포라 등지에서 발견되었다. 도 플라이오-풀라이스토세의 인류화석들이 발견된 오모 유적은 두르 카나 호수 북쪽으로 홀러드는 오모 강의 남쪽 골짝에 위치하고 있다 (그립 8. 3). 1967 년 프랑스 학자인 코팡스 Yves Co pp ens 와 아람버그 그리고 미국 학자인 하웰 F.C.Howell 등이 이 지역에서 많은 종류의 젖먹이동물과 인류속에 속하는 아래턱과 이빨 그리고 사지뼈 화석들 울발굴하였다. 화석 발견의 중요성은 가장 오래된 이빨 화석이 약 250 만 년 전의 지층에서 발견되며 같은 종류의 화석들이 계속해서 150 만 년 전의 지 층에서까지 발견된다는 접이다. 나아가 오모 지역의 중요성은 무엇보 다도 유적 충위에 대한 과학적인 연구에 있다. 이 지역에는 올드바이 골짝의 되적 (80 만 년 -200 만 년)보다 훨씬 오래 된 약 400 만 년 동안 퇴적된 지층들이 있는데, 하웰에 의해 치밀하게 연구된 이 지역의 지 충은 지금까지 동아프리카의 플라이오-플라이스토세 유적지의 표준 충위가되고 있다. 쿠비 포라 Koobi Fora 쿠비 포라는 또 다른 유명한 풀라이오-플라이스토세 유적지로 두 르카나 호수 동쪽에 자리하고 있다. 이 호수는 이전에 루돌프 호수 Lake Rudo lf로 알려졌는데, 리차드 리키와 당시 미국 캘리포니아 주 립 버클리 대학교의 글린 아이작 Gl yn Isaac 이 이끄는 발굴단은 쿠비 포라 유적에서 2-100 만 년 전의 연대를 가전 많은 오스트랄로피데쿠 스 보이세이 화석을 발굴하였다. 두르카나 호수 동쪽지역에서 출토된
보이세이 화석들은 오모 일대에서 발굴된 보이세이 화석들과 함께 동 아프리카에서 살던 굳세 보이는 오스트랄로피테쿠스 연구에 중요한 자료를제공하고 있다.
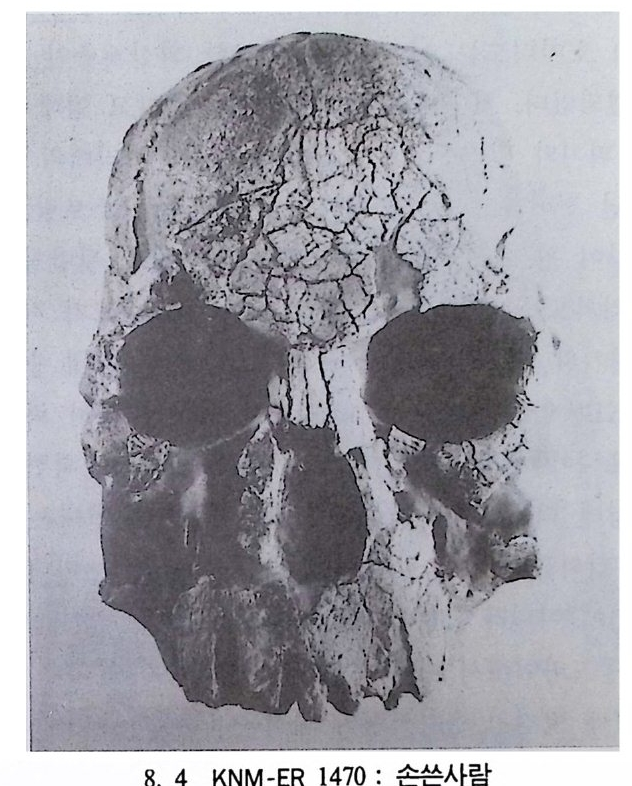 `
`
그러나 무엇보다도 리처드 리키가 이 지역에서 성취한 중요한 업적 은 굳세 보이는 오스트랄로피 테쿠스와 같은 시기에 살던 인류조상의 머리뼈 화석의 발견이었다. 1972 년 초반에 발굴되어 케냐 국립박물관 화석대장에 KNM ER - 1470(KNM-ER 은 Keny a Nati na l Museum East Rudo lf의 약자임 )으로 기록된 이 머 리뼈는 올드바이 에서 출토 된 어떤 손쓴사람의 화석보다 더 완전한 형태로 발견되었다(사전 8.
4). ER- 1 470 화석은 오스트랄로피테쿠스 아프리카누스를 일부 닮았으 나 머리뼈의 용량이 775cc 나 되는 큰 머리뼈를 지니고 있고, 또한 전 체적인 모습이 아프리카누스와는 달라 손쓴사람으로 분류되었다. 루 이스 리키 등이 주장한 손쓴사람의 존재가 사실로 증명되었다. ER-1470 호미니드는 이 지역에 발달한 화산재충인 KBS 충 바로 밀에서 발견되었다. 처음에 KBS 화산재를 가지고 연대측정을 한 영 국 학자인 피치와 밀러 Fit ch & M ill er 는 이 머리뼈의 연대가 최소 한 300 만 년 전이라고 발표하였다. 이 연대는 오스트랄로피테쿠스는 인류의 조상이 될 수 없다는 리키 가문의 주장을 뒷받침하기에 충분 하였으며, 리차드 리키로 하여금 ER-1470 호미니드가 자신들이 찾던 가장 이른시기의 인류의 직접조상이라고 믿게 하는 데 충분하였다. 그러나 KBS 지층에 대한 보다 정밀한 연구와 특히 화산재에 대한 미국 캘리포니아 대학교의 커티스 G. H. Cur ti s 의 정확한 칼륨 • 아르 곤 연대측정에 따르면 이 머리뼈가 발견된 충의 연대는 단지 180 만 년 전으로 밝혀졌다. 오늘날 학자들은 ER-1470 호미니드의 연대는 300 만 년 전이 아니라 180 만 년 전이라는 연대를 받아둘이고 있는데, 이 연대는 올드바이에서 나온 손쓴사람 화석들의 연대와 일치한다. 가장 최근에 발견된 손쓴사람의 화석은 1986 년에 미국 버클리 소재 인류기원 연구소팀에 의해 올드바이 골짝에서 찾아졌다. 화석들은 어 른 한 개체의 사지뼈둘로 손쓴사람의 키를 가늠할 수 있었는데, 이 개체는 키가 매우 작아 루시의 키보다도 작다(사전 8. 5). 오늘날 우리는 손쓴사람이 250 만 년 전부터 170 만 년 전 사이에 살았으며 매우 큰 머리부피 (530-800cc) 를 가지고 있었음울 알 수 있 다. 손쓴사람은 후에 철멸된 굳세 보이는 모습의 오스트랄로피데쿠스 들과 오랜 기간을 함께 아프리카에서 살았다고 알려졌다. 한편 지금 까지 알려진 손쓴사람의 화석에서 · 보이는 머리뼈 크기의 차이를 학자
둘은 남 • 녀 성별에 따른 차이로 설명해 왔는데, 이는 두르카나 호수 일대에서 250 만 년 전부터 170 만 년 전 사이에 인류속으로는 손쓴사 람 한 〈종〉만이 살았다는 〈단일종 가설〉을 수용한 결과이다.
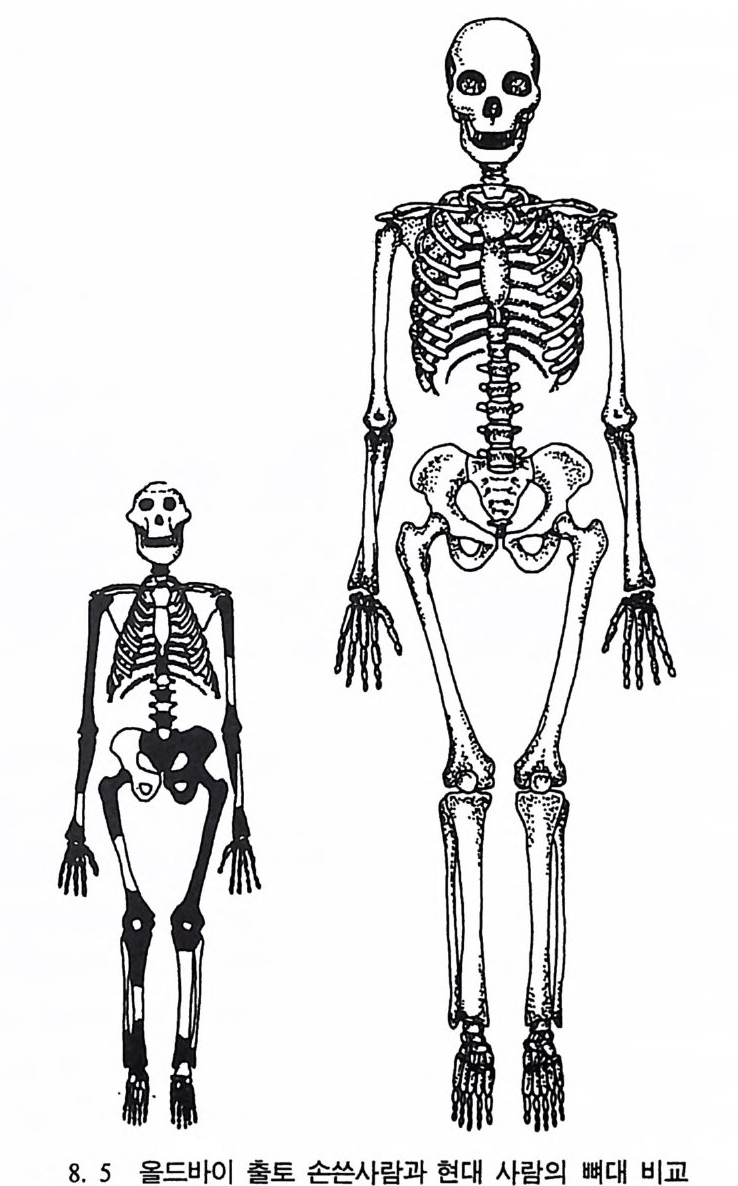 8. 5 올드바이 출토 손쓴사람과 현대 사람의 뼈대 비교
8. 5 올드바이 출토 손쓴사람과 현대 사람의 뼈대 비교
1992 년 영국의 인류학자인 우드 Bernard Wood 는 이런 단일종 가 설에 의문을 제기하고 이 지역에서 발견된 큰 머리뼈는 손쓴사람이 아닌 루돌프 사람 Homo rudol f ens i s 으로 분류하고 있으며, 작은 머리 뼈의 화석만이 손쓴사람으로 분류된다는 〈 복수종 가설 〉 을 세웠다. 그 러나 정말로 투르카나 호수 일대에는 여러 종의 인류의 조상과 사촌 들이 함께 어울려 살았었는지는 더 많은 화석의 발견과 연구가 필요 하다(사진 8. 6).
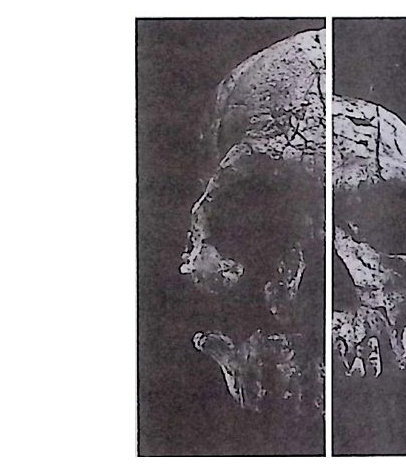
8. 6 왼쪽은 KNM-ER 1470 이고 오른쪽은 KNM -E R 1813 의 손쓴사람 화석으로 학자들은 이들이 성별차이 Sexual Di mor p h i sm 를 나타내기보 E 는· 머리 구조와 크기의 차이를 나타내고 있 다고주장한다.
이른시기의 호미니드의 문화와 행위 지금까지 우리는 단지 호미니드의 뼈화석에 대해서 살펴보았다. 그 러나 인류의 직접조상인 이들은 돌로 만든 연모들을 남겼는데, 바로 이 연모들이 구석기 고고학의 주된 연구대상이다. 우리가 구석기시대 유적지에 남겨진 뻣조각과 돌연모들만 가지고 이 당시에 살던 인류 조상의 행위를 결정한다는 것은 마치 탐정수사 와 같은 일이라 할 수 있다. 오랫동안 이들이 남긴 연모들의 연구는 미술사 연구처럼 매우 기술적 방법으로 이루어져 연대순으로 정리되
었기 때문에 마치 이 연모들이 진화했던 것처럼 여겨져왔다. 그러나 진화한 것은 사람이지 연모가 아니리는· 점이다. 1960 년대에 인류학자 들은 유적지에 남겨진 연모와 동 • 식물들을 연구하여 이를 남긴 인류 의 행위를 알고자 하였다. 오늘날 우리는 연모둘이 발견된 유적지에서 남겨진 호미니드 화석 과 동물화석 그리고 함께 남겨진 연모와의 모든 관계를 알고자 한다. 이렇게 풀라이오-풀라이스토세에 살던 호미니드의 행위는 올드바이 골짝의 연구에서 시작되었다. 울도완 돌문화 동아프리카의 세렌게티 고원 동쪽에 있는 올드바이 골짝은 가파르 기 때문에 골짝 위에서 밑바닥까지는 약 100m 정도에 달하며 약 200 만 년 전에 형성되었다. 밑바닥부터 골짝 위까지 옛날 호수 퇴적들과 화산재 및 응회암 등의 퇴적물로 쌓여 있다. 골짝 맨 밀총인 Bed I 은 약 200 만 년 전에 형성되었는데, 당시 주위에는 커다란 호수가 있 었으며 그 주위에서 오스트랄로피데쿠스와 손쓴사람들이 함께 살았 다. 올드바이 유적은 맨 밀총인 Bed l 에서부터 맨 위총인 내슈슈 Na isi u s iu 충까지 모두 7 층으로 구분되어 있는데 이 중 Bed I 에서 Bed 4 에 걸쳐 호미니드 화석과 젖먹이동물들의 전화기록들이 나타난다(그 림 8. 7). 이 지역은 처음에는 독일인이 찾았으며 후에 리키 등에 의 해 연구되다가, 1980 년 이후 미국 캘리포니아 버클리에 있는 〈인류기 원 연구소〉와 탄자니아의 고고문물부에 의해 연구되고 있다. 올드바이 골짝에는 도처에 동물의 뼈와 돌들이 흩어져 있는데 돌연 모는 몸돌 연모 Core Tools 와 격지 연모 Flake Tools 로 나누어전다. 가 장 오래 된 연모는 맨 밀총인 Bed l 에서 나온 울도완 연모로, 간단
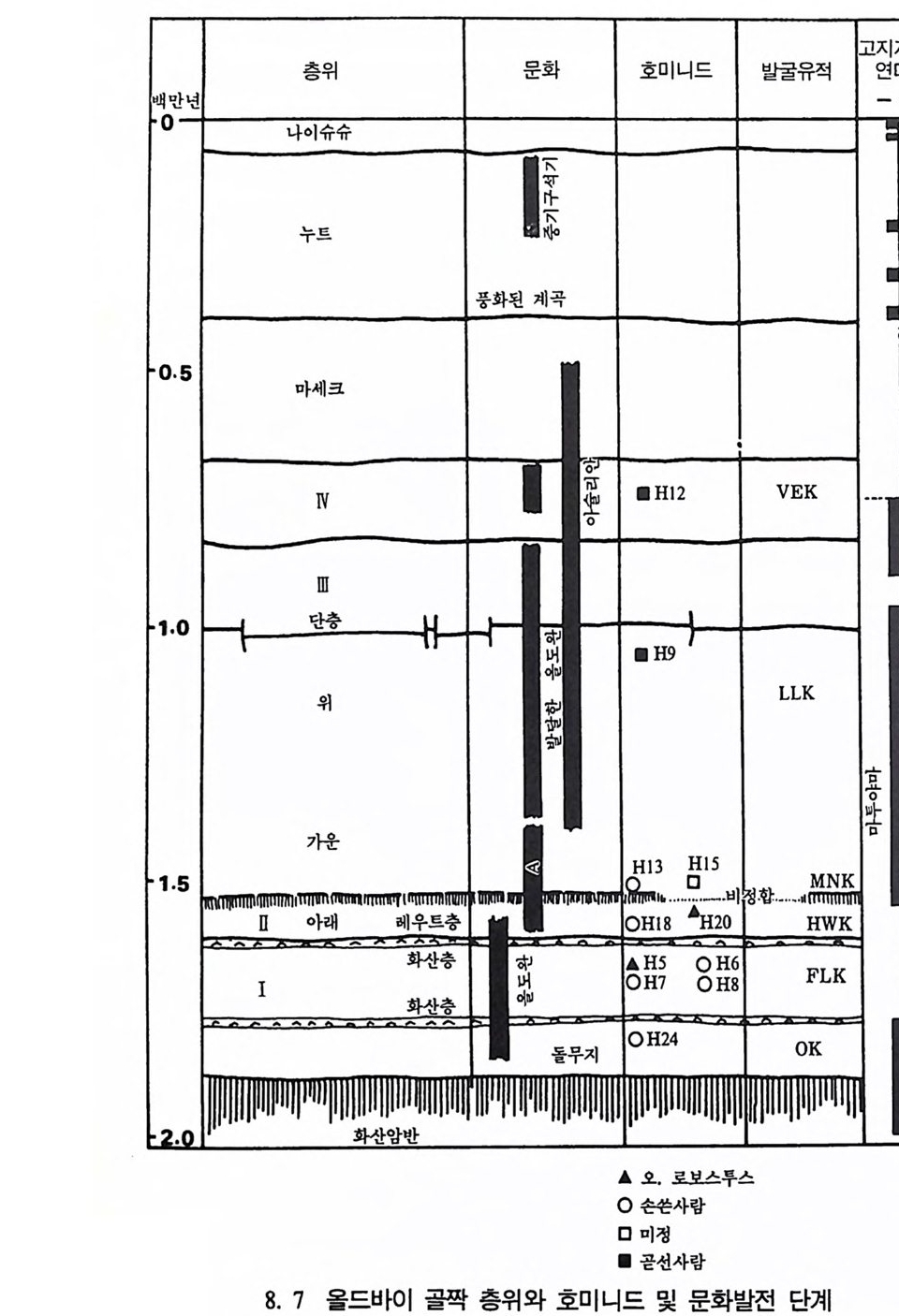 백만년 Iltz |고-지+자 7 1
백만년 Iltz |고-지+자 7 1
히 손질된 찍개가 대표 연모이다(그림 8. 8). 모든 연모들은 주변 화 산지대로부터 흘러내려오는 물에 쓸려 호수의 가장자리에 묻혀졌다. 이 유적에서 출토된 돌연모는 메리 리키 등의 고고학자들에 의해 자세 히 연구되었는데, 올드바이에서는 연모의 발달이 연속으로 나타나 인 류가 어떻게 연모를 만들고 이룰 발전시켰는지를 알 수 있게 해준다.
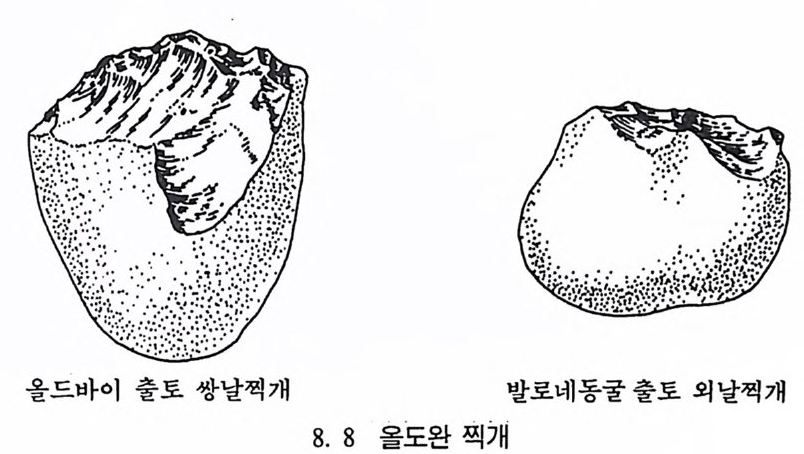 올드바이 출토 쌍날찍개 발로네동굴 출토 외날찍개
올드바이 출토 쌍날찍개 발로네동굴 출토 외날찍개
올드바이 골짝에서 살던 호미니드의 행위 올드바이 골짝에서는 돌연모와 함께 많은 동물의 뼈화석이 발견되 었다. 여기서 나온 뻣조각 표면에 자론 자국들이 나타나는데, 이 흔적 은 이 당시에 살던 우리의 조상이 고기를 얻기 위해서 연모를 사용했 음을 보여주는 증거로 해석되었다(사진 8. 9). 그리고 Bed 1 에서는 뻣조각들과 돌연모들이 커다랗게 함께 쌓여 있는 채로 발견되었는데, 이런 현상에 대한 설명은 1960 년대에 메리 리키와 글린 아이작에 의 해 시도되었다• 그들은 약 180 만 년 전부터 인류는 〈사냥을 하고 옮 겨온 돌들을 이용하여 연모를 만들고 그 자리에서 동물등 을 도살하였 으며, 먹다 남은 찌꺼기는 주위먹기짐승들이 먹었고, 발견된 뼈둘은 바로 호미니드와 주워먹기짐승들이 남긴 것〉이라고 해석하였다• 그
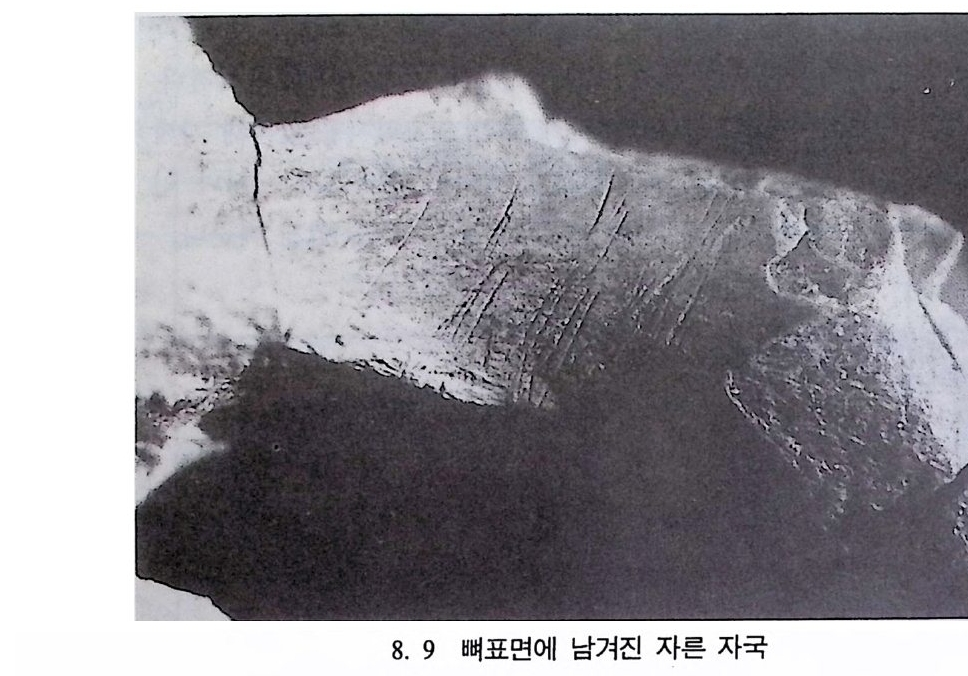 .‘; .'一, ·\-:.., ~ },.. s,C : ;. `· .` :.2••- ··
.‘; .'一, ·\-:.., ~ },.. s,C : ;. `· .` :.2••- ··
들에게 올드바이 유적지는 주거지로 인식되었다. 한 걸음 더 나아가 아이작에 의해 성립된 〈음식 분배행위 가설〉은 고고학계에 많은 영향을 끼쳤다. 아이작은 인류와 유인원을 구분하는 기준을 사냥 그 자체만으로 강조하지는 않았다. 그는 협동과, 특히 음 식의 분배행위가 인류진화의 중심이었다고 주장했기 때문에 그의 이 런 주장은 〈음식 분배행위 가설〉로 불리고 있다. 이 가설의 핵심은 남자들은 연모들을 가지고 지속해서 사냥을 하고 여자들은 식량을 채 집했다고 주장하며, 이런 행위를 위해 호미니드 사이에 협동과 의사 표현이 있었으리라고 가정하였다. 죽 동물성 음식과 식물성 음식은 남 • 녀에 의해 각각 얻어졌으며, 이들은 자기들이 얻은 음식을 주거 지로 가지고 돌아와 거기서 분배했다고 말한다. 이 가설은 오늘날 살고 있는 수렵 • 채집집단들의 생활유형을 과거 에 살았던 호미니드를 연구하는 데 적용한 결과이다. 리처드 리키는
이런 형태의 경제행위를 〈 최초의 혼합경제 〉 라고 불렀는데, 아이작은 음식 분배행위가 일어났던 사회환경의 복잡성이 인류전화의 추전력이 지 사냥에서 요구되는 지식과 기술의 요구가 전화의 추진력은 아니라 고 주장하였다. 이렇게 그의 가설은 사회환경에 초접을 맞춤으로써 주로 기술과 조직적인 면을 강조했던 이전의 〈 사냥 가설〉과는 구분된 다. 1980 년대에 들어와 아이작과 그의 제자들은 쿠비 포라에 있는 150 만 년 전 잠시 주거지로 이용됐던 50 번 유적지 Fxji 50 의 주거지를 복원 해서 이 음식 분배행위 가설을 검증하려는 노력을 진행시켰다(사전 8. 10). 50 번 유적지의 연구 결과 이 유적에 남겨진 돌들과 동물화석 및 식물성 음식들은 다른 곳으로부터 옮겨져 온 것이며, 당시의 호미 니드는 동물등을 사냥하기보다는 다른 동물들이 먹다 남긴 나머지를 주워먹었을 것이라고 아이작은 자신의 가설 일부를 수정하였다.
 8. IO 50 번 유적지 전경
8. IO 50 번 유적지 전경
아이작은 유적지에 남겨진 뻣조각들과 연모(사진 8. 11) 들은 주거 지 주위에 있던 한 선사인류 집단의 행위를 보여주는 것으로 이런 형 태의 행위를 〈 인류행위 중심지 이론 〉 이라고 부르는 가설로 설명하였 다. 이 가설에 따르면 손쓴사람들은 〈 주거지를 중심으로 식량을 획득 하여 분배하는 생활행위 〉 로 설명할 수 있는 사회조칙 유형을 가지고 있었다고한다.
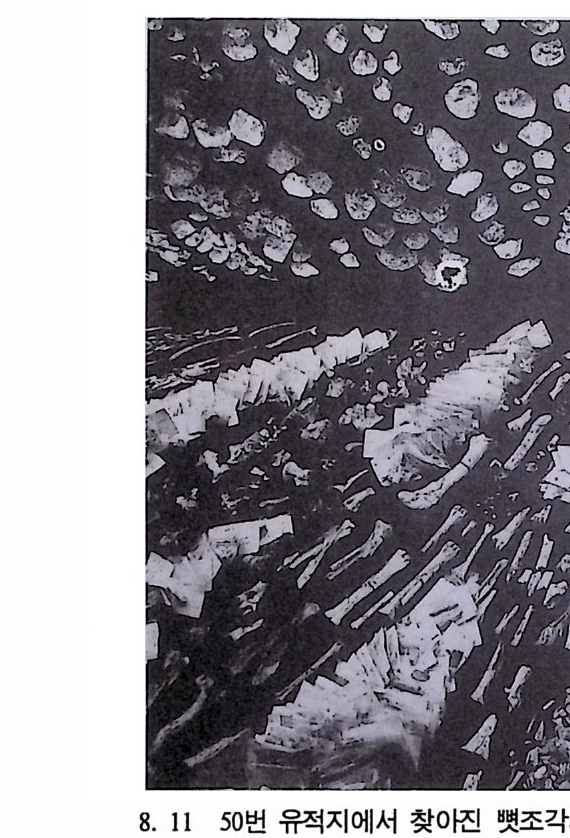 8. 11 50 번 유적지에서 찾아진 뻣조각과 돌연모
8. 11 50 번 유적지에서 찾아진 뻣조각과 돌연모
이렇게 아이작의 변화된 생각과 같은 의견이 이미 1970 년대 중반부 터 몇몇 고고학자들에 의해 제기되기 시작했다. 오늘날 고고학자들은 그들이 본래 생각했던 것만큼 알지 못한다는 사실을 인정한다. 과거
에 고고학자들은 유적지에서 얻은 증거를 일정한 모델에 적용하여 과 거 인류의 행위와 사회를 재구성하고자 하였는데 이는 정확한 연구 방법은 아니다. 우리가 과거에 일어난 사건들을 해결하고자 할 때 우 리는 과거의 일어난 일은 단지 한 방법으로 행하여졌는데, 오늘날 이 에 대한 해석은 여러 가지 다른 결과로 나타난다는 사실을 우리는 알 고 있으며, 동아프리카에서 발견된 손쓴사람들의 행위에 관한 가설들 이 그 좋은 실례라 할 수 있다• 인류 조상의 행위에 관한 1970 년대 초반까지의 연구에 관한 새로운 비평 • 평가는 루이스 빈포드 Le wi s B inf ord 로부터 시작되었다. 그 논 〈 사냥 가설〉뿐만 아니라 〈음식 분배행위 가설〉에 회의를 나타내었 다. 그는 고고학 유적지에서 뼈들을 발견했을 때 이 뼈둘이 인류의 행위에 의해서 거기에 있디는` 생각을 거부했다. 심지어 연모 가까이 에서 나온 화석들도 연모들과 상관없이 거기에 도달하였을 수도 있 고, 또 그들이 서로 가까이 있는 것도 단지 우연일 수도 있다고 그는 주장한다. 빈포드는 유적지에 퇴적된 뼈들이 어떻게 운반되었는가를 알기 위 해 현재에 일어나는 상황을 연구하여 사람의 영향을 받지 않는 자연 조건 아래서 어떻게 뼈들이 서로 분리되고 흩어지는가를· 관찰하였다. 그 결과 그는 과거를 보는 우리들의 시야가 매우 좁기 때문에 오류가 생겨났다는 결론에 도달하게 되었다. 빈포드는 인류전화상 늦은시기에 살았던 호미니드 중 특히 네안데 르탈사람의 행위에 대한 재검토 이후 이론시기 호미니드의 행위에 관 한 그 동안의 해석에 의문을 제기하여 리키와 아이작이 재구성한 이 른시기 호미니드의 행위는 논리상으로 가능하지 않다고 주장하였다. 따라서 빈포드는 가장 이론시기의 유적지인 올드바이 골짝에 관해 이미 발표된 고고학기록들에 대한 검토 끝에 〈올드바이 유적지는 사 냥을 하던 식육류와 이들이 남긴 고기를 얻어먹던 호미니드들이 죽은
장소 〉 라고 자신의 견해롤 밝혔다. 빈포드에게는 올드바이 골짝에서 발견된 〈 알 수 없는 뼈들과 돌더미들 〉 은 단지 알 수 없는 더미일 뿐 이며, 남겨진 뼈둘은 골수를 얻으려고 몇 개의 하잘것없는 뼈들을 깨 뜨렸을 때 남겨진 뻣조각일 뿐이었다. 그에게 올드바이 골짝은 더 이 상 호미니드가 살던 주거지가 아니었다. 이론시기의 호미니드는 더 이상 고귀한 사냥꾼도 아니고 음식을 분배하던 사회 존재도 아니었 다. 많은 양의 고기를 먹던 존재도 아닌, 남겨진 고기나 주워 먹으며 가끔 사냥을 하던 존재였거나 아니면 식육류가 먹다 남긴 음식이나 훔쳐먹는 그런 존재였다. 빈포드에게 사냥채집의 생활방식은 생각보 다 늦게 호미니드에게 나타난 행위였다 . 아이작과 빈포드의 논쟁 에 자극을 받은 리 처드 포츠Ri chard Pott s 가 최근에 올드바이 유적지에서 발굴된 뼈화석들을 재조사하였다. 이 작업에는 존 홉킨스 대학의 패트 쉬프만과 아이작의 제자인 위스콘신 대학의 헨리 번 Henr y Bunn 과 그의 아내 엘렌 크롤 Elen Croll 도 이 조사에 일익을 담당하였다 . 이들의 연구는 두 개의 질문, 죽 돌들과 뼈화석이 어떻게 같은 장소에서 나타나는가? 하는 질문과 올드바이 유적지는 주거지인가? 하는 질문에 집중되었다. 연구 결과 포츠는 올 드바이 유적지에서 발견된 돌감둘은 특별한 지점으로부터 계획성있게 운반되었다고 주장하고 있다. 한편 여기 남겨진 뼈화석들에서는 인류 가 행한 자론 자국과 동물들이 씹은 자국이 (사전 8. 12) 함께 나타난 다고 쉬프만 등은 밝히고 있다. 그리고 두번째 질문에 대해서 그는 〈아니다〉라고 답하고 있다. 왜냐하면 주거지란 짧은 기간중에 집중적 으로 사회활동과 음식분배 행위가 일어나야 하며 나중에는 그 자리를 떠나는 그런 장소가 주거지로 정의되는데, 올드바이 유적지는 이와는 반대로 오 년에서 십 년이란 기간 동안에 가끔씩 사용되었기 때문이 다. 따라서 포츠는 올드바이 유적지를 〈선 주거지〉란 용어로 표현하 였다.
 a) 하이에나
a) 하이에나
 `.'. ,, 「. ``내
`.'. ,, 「. ``내
 c) 하이에나가
c) 하이에나가
지금까지 200 만 년 전에 살았던 이론시기 호미니드인 손쓴사람의 행위를 설명하는 여러 가지 연구를 살펴보았는데, 이들의 행위는 단 한 가지 행위유형으로는 설명될 수 없다는 점을 알 수 있다. 그러나 글린 아이작 등에 의해 시도된 호미니드 행위에 관한 새로운 증거의 발견과 동시에 수반되는 의문은 우리를 옛 유적지에 대한 재조사와 새로운 연구의 장으로 안내하였다는 데 의의가 있다. 곧선사람 Homo erectu s : 直立猿人의 출현 이른시기의 인류들에 관한 우리의 지식은 화석에 바탕을 두고 있기 때문에 호미니드 화석의 발견의 역사를 살펴보는 것은 인류의 조상에 관한 생각이 어떻게 변해 왔는지를 이해하는 데 도움이 된다. 지금까 지 고인류학계에서 진행되어온 인류화석에 관한 연구는 크게 두 개의 범주로 나누어진다. 첫째는 전에 알려지지 않았던 화석의 발견을 통 해 인류의 전화관계를 살펴보는 것이고, 둘째는 이미 발견된 화석을 새로운 지식과 방법으로 설명하는 것이다. 이 장에서는 전화상 손쓴사람의 후예로 알려진 곧선사람의 이른 발 견의 역사를 뒤돌아보고, 이 호미니드에 관해서 고인류학계에서 연구 대상이 되는 내용을 살펴보기로 한다. 오늘날 고인류학계는 지금까지 발견된 호미니드 화석을 바탕으로 인류의 전화과정을 복원하는 연구 를 해왔는데 이렇게 인류의 전화과정을 연구하는 〈인류계통학 Human Ph y lo g en y〉에 따르면 인류의 전화는 일반적으로 오스트랄로피데쿠스 아파렌시스가 손쓴사람으로 전화하였고, 손쓴사람은 곧선사람으로, 이 어 곧선사람은 슬기사람으로 전화했다고 밝히고 있다. 곧선사람은 해부학상으로는 우리와 꼭 갇지는 않다. 그러나 인류속 안에서는 우리 자신과 가장 가까운 종이라 할 수 있다. 오늘날 이들
은 존재하지 않으나 이들의 후손인 슬기사람은 그들의 유전인자 안에 곧선사람의 유전인자를 포함하고 있다. 160 만 년 전부터 10 만 년 전 까지 아시아, 아프리카 그리고 유럽의 구대륙에서 살던 곧선사람들의 발견의 역사는 19 세기로 울라간다. 오스트랄로피테쿠스 아프리카누스 가 발견되기 35 년 전의 일이었다.
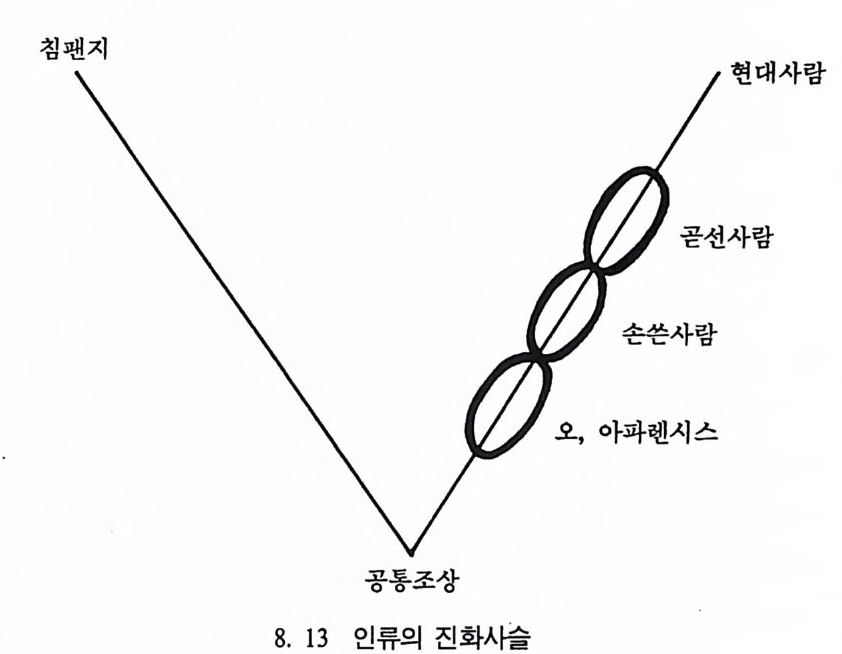 침팬지
침팬지
1800 년대 중반, 유럽에서는 지구의 현재와 미래에 대한 연구에 관 심이 모아졌다. 다윈의 『종의 기원』과 『인류의 기원』이 출판된 이후, 일단의 학자들은 인류와 유인원 양쪽을 잇는 중간형의 존재가 있었다 고 가정하였다. 여기서 〈잃어버린 사슬〉이라는 개념이 생겨났는데, 이 개념은 인류와 침팬지의 공통조상이 과거 어느 시점에 있었다는 것을 암시한다. 호미니드 화석 기록들을 통해서 볼 때 우리의 공통조상부 터 우리 자신에 이르기까지 사슬이 있음울 알 수 있다(그립 8. 13). 위의 그림에서 볼 수 있는 공통조상에서 침팬지에 이르는 사슬은
현재까지는 알려져 있지 않으며, 공통조상에서 절멸한 오스트랄로피 테쿠스로 가는 다론 사슬이 있음을 알 수 있다 . 오늘날 호미니드 화 석에 관한 증거는 아주 훌륭하나, 1 세기 전인 1800 년대에는 어느 것 도 가능했던 증거가 아니었고 상상으로 채워져 있었다(사전 8. 14).
 `:
`:
이 당시 다윈의 지지자였던 독일의 동물학자인 헤켈은 화석의 증거가 없음에도 불구하고 〈잃어버린 사슬〉에게 〈피테칸트로푸스 아라루스 Pit hec an thr op u s alalus(Pit he canth r op u ~ 단어는 그리스 말에서 온 pithe kos, 죽 유인원 a pe과 anth rop o s, 죽 인류 man 라는 말이 결합된 단 어이며, ala~ 말을 할 수 없는 존재 : witho ut s pe ech 라는 뜻임)〉 라는이름을붙였다.
곧선사람의 발견 자바 곧선사람 P it hecan t hrop us erectu s 1859년 에 네덜란드에서 태어난 드보아 Eu gene Dubo i s 는 의과대학 을 졸업한 후 암스테르담 의과대학의 해부학교수가 되었다. 당시에 알려져 있던 인류의 기원과 전화에 관한 여러 가설과 네안데르탈 화 석에 많은 관심을 가지고 있던 드보아는 특히 헤켈로부터 커다란 영 향을 받았다. 네안데르탈사람이 정말로 오래 된 인류라고 믿었던 드 보아는 유럽의 기후가 네안데르탈사람 이전에 살던 〈잃어버린 사슬〉 인 유인원과 인류를 잇는 중간형의 존재가 살기에는 너무 추웠기 때 문에 이 존재는 유럽 이의의 지역에 있었으리라는 생각에 영향을 받 았다. 또 다론 영향은 다윈과 월리스로부터 왔다. 다윈과 헉슬리를 포함 한 대다수의 학자들은 아프리카 대륙이 인류의 기원지일 가능성이 가 장 높은 지역으로 꼽았다. 이런 견해는 일차적으로 인류가 머리칼이 없는 점으로 보아 열대지방에서 기원했음이 틀림없다고 생각했으며, 인류와 가장 가깝게 닮은 침팬지가 아프리카에서 살고 있고, 끝으로 〈 검은 대륙 〉 으로 인식되던 아프리카 대륙에 관한 호기심이 이들로 하 여금 아프리카를 인류의 기원지로 보게 하는 요인이었다. 한편 말레이시아에서 오랫동안 살았던 월리스는 수마트라와 보르네 오에서 살고 있는 기본과 오랑우탄에 주목하여 만일 인류의 조상을 찾고자 한다면 유인원이 사는 열대지역에서 찾을 수 있을 것이며 그 지역을 아시아로 가늠하였다. 이에 따라 드보아는 인류 조상의 화석 울 찾기 위해서 네덜란드령 동인도네시아 지역으로 갔다. 이는 훈련 울 받은 연구자가 호미니드 화석을 찾기 위해서 알려지지 않은 지역 으로 가서 실제로 화석을 찾은 역사상 몇 안 되는 예 중의 하나이다. 가장 중요한 화석들이 1891 년에 자바 섬의 솔로 Solo 강가에 있는
트리닐 Trin il 지역에서 찾아졌다(사전 8. 15). 드보아는 여기서 몇 개 의 이빨과 머리뼈 하나를 찾아냈다. 이 머리뼈는 현생인류의 머리뼈 에 비해 머리가 낮고 머리통이 편평한 모양을 하고 있으며, 머리부피
 8. 15 자바, 트리닐 지역의 솔로 강. 여기서 드보아가 피테칸트로푸스 에렉
8. 15 자바, 트리닐 지역의 솔로 강. 여기서 드보아가 피테칸트로푸스 에렉
는 850cc 로 유인원 (275-750cc) 과 슬기사람 (1000 一 2 000cc) 의 중간에 속했다. 그 이듬해 같은 지역에서 넓적다리뼈 하나를 발견하였는데 이는 현생인류의 허벅지뼈와 매우 닮았다. 드보아는 헤켈의 직관을 기념하는 뜻에서 이 화석을 피데칸 E 쿠 立스라고 불렀다. 그리고 이 호미니드 화석이 말을 할 수 있는 능력이 있었는지에 상관없이 그는 〈에렉두스 erectu s 〉라는 이름을 선택했는데, 이는 발견된 허벅지뼈가 두 발로 걸었음을 보여주었기 때문이었다. 나중에 〈자바 곧선사람〉으 로 알려진 피데칸트쿠프스 에렉투스의 탄생이 이루어전 셈이다(사전 8. 16).
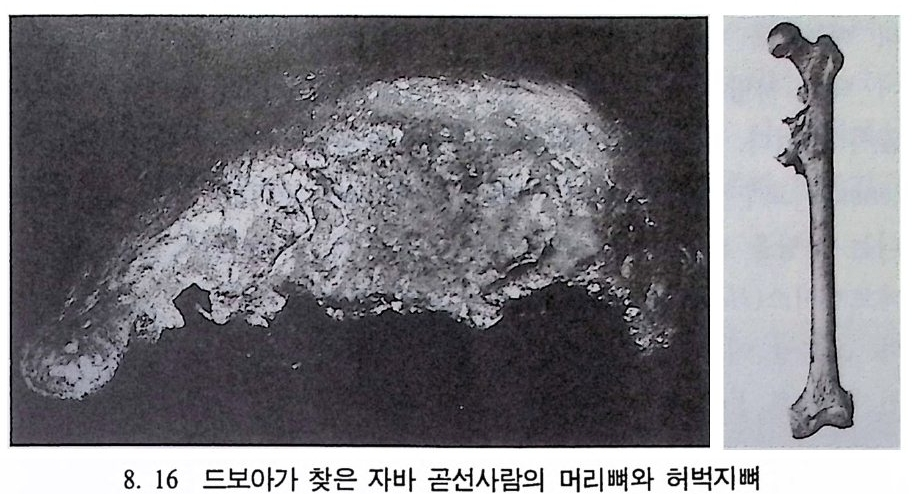 8. 16 드보아가 찾은 자바 곧선사람의 머리뼈와 허벅지뼈
8. 16 드보아가 찾은 자바 곧선사람의 머리뼈와 허벅지뼈
드보아는 피데칸 E 쿠立人를 1895 년에 네덜란드의 라이덴에서 열린 제 3 차 국제동물학회에 보고하자 곧바로 자바 곧선사람에 관한 논란이 일어났다. 독일 계통의 학자들은 피데칸뜨쿠立人가 인류의 특칭을 가 진 유인원으로 믿었고, 대다수의 영국 학자들은 이 화석을 유인원의 특칭을 지닌 인류로 생각하였다. 미국 학자들만이 드보아가 생각한 것처럼 유인원에서 인류로 진행되어가는 중간형의 존재로 받아들였 다. 드보아는 피데칸 E 쿠立스에 대한 자신의 생각이 옳다는 것을 증 명하기 위해 많은 노력을 기울였으나 피테칸트로프스에 대한 논란은 계속되었다. 이에 실망을 느낀 드보아는 이 화석들을 자신의 거실 바 닥 밑에 묻어버렸으며 다시 이 화석이 세상에 나오기까지는 30 여 년 의 세월이 걸려야만 했다. 하이델베르크사람 Homo heid e lberge n sis 자바 곧선사람에 이어 유럽에서도 곧선사람의 화석이 발견되었다. 1907 년 독일의 마우어에서 가까운 곳에 있는 모래구덩이에서 여러 개
의 뼈화석을 발굴하였는데 그 중에 커다란 아래턱을 하나 찾아냈다. 이 턱은 사람의 턱을 닮아 있었으나, 크기는 커다란 유인원의 턱과 닮아 있었다. 이를 연구한 하이델베르크 대학의 쉐텐삭 O tt o Schoe- ten sack 교수는 이 아래턱이 매우 크기는 하지만 작은 송곳니와 어금 니는 사람을 닮은 데에 주목하였다. 그는 이 화석에게 〈 호모 하이델 베르겐시스 (Homo heid e lberg e nsis = 하이델베르크사람) 〉 란 이름을 붙였 다. 오늘날 이 호미니드는 중기 풀라이스토세에 유럽에서 살았던 곧 선사람으로 분류되고 있다. 마우어에서 발견된 곧선사람의 턱이 인류의 전화과정에서 자기 자 리를 잡는 데에는 아시아 대륙 중국에서 새로운 화석이 발견되는 20 년 후까지 기다려야만 했다. 베 이 징 곧선사람 Sin a nth r op u s pe kin e nsis , Beij ing 베이칭 사람으로 알려진 곧선사람의 화석은 중국의 수도 베이칭에 서 남서쪽으로 40km 떨어전 저우커우덴(周口店 Zhoukoud i an) 이라 불리는 곳에서 발견되었다. 중국에서 용골로 알려진 동물뼈둘은 한약 방에서 구할 수 있었는데, 이미 인류를 닮은 이빨이 1900 년 초에 독 일의 고동물학자인 슐뢰서 Max Schlosser 에 의해 보고되었다. 저우커 우뎬 일대는 석회암이 발달한 지대로서 석회암 광산과 동굴이 있었는 데, 1920 년대 초에 스웨덴의 지질학자인 앤더슨J ohn Gunnar Ander- sson 은 용골산으로 알려진 동굴에서 많은 동물화석을 발견하였다(사 진 8. 17). 1927 년에 이 지역을 조사하던 미국의 보린 B i r gi r Bohlin 등은 이 론시기에 살던 호미니드의 어금니 세 개를 찾아냈다. 그는 당시 북경 의과대학에 와 있던 캐나다 출신 해부학자인 데이비드슨 블랙 Da- vids on Black 에게 이 이빨들을 감정해 줄 것을 요청했다. 이빨울 감
정한 블랙은 이 이빨을 가전 존재는 현재 살고 있는 사람과는 다른 옛 사람으로 단정하여 이 이빨을 가졌던 호미니드에게 〈 시난트로푸스 페키넨시스 S i nan t hro p us p ek i nens i s 〉 라고 이름을 지었다. 이 유적에서 발굴이 계속되었는데 1927 년과 1937 년 사이에 40 개체분의 뻣조각들 이 발굴되었다.
 8. 17 저우커우옌
8. 17 저우커우옌
1934 년에 블랙이 갑작스럽게 죽은 후 연구는 미국 시카고 대학 해 부학 교수인 바이덴라이히 Franz We i denre i ch 가 대신하였다. 발굴은 제 2 차 세계대전까지 계속되었으며, 제 2 차 세계대전이 발발하자 저우 커우뎬 화석들을 해의로 반출하려는 시도가 있었다. 1941 년 5 월 일본 군이 베이징에 전주하기 직전 아홉 명의 미국 해병들이 이것을 호송 할 책임을 지고 있었으며, 이 유물들은 미국 잠수함에 실려지기 위해 톈진항으로 옮겨졌다. 그러나 이들 해병들은 모두 일본군에 의해 체 포되었고 배 또한 여류되었다. 동시에 화석들도 사라져버렸고 오늘날
까지 아직 찾아지지 않고 있다. 그러나 일본이 베이칭에 전주하기 전 에 바이덴라이히는 저우커우뎬 화석의 연구 결과와 복제품을 미리 미 국으로 보냈고 본인은 가까스로 중국을 탈출하였다. 오늘날 남아 있 는 화석들은 바이덴라이히가 중국에서 만든 복제품들이다. 최근 아프리카와 유라시아에서 발견된 화석 자바에서 처음으로 곧선사람이 발견된 이후 약 1 세기 동안에 아프 리카와 유럽 및 아시아에서 많은 곧선사람의 새로운 화석들이 발견되 었다. 1950 년대 중반부터 60 년대에 걸쳐 아프리카에서는 알제리의 터니파 인을 시작으로 올드바이 골짝의 Bed 2 에서 곧선사람의 화석이 발견 되었다. 1970 년대에 오면서 모로코의 살레를 시작으로 아프리카에서 의 곧선사람 화석의 발견은 동아프리카의 쿠비 포라에서 절정을 이룬 다. 1974 년 리처드 리키는 이 지역에서 160 만 년 전에 살던 거의 완전 한 머리뼈를 포함해서 많은 곧선사람의 화석을 발견하였다. KNM ER - 3733 으로 알려진 이 머리뼈 화석은 커다란 머리통과 가날폰 머 리선 및 옆니를 가지고 있지만 전체 모습은 저우커우덴에서 출토된 화석과 매우 닮았다. 한편 남아프리카의 트란스발 지역의 스와르트크 란스에서 브레인은 굳세 보이는 모양의 아래턱을 하나 발견하였다. 델란트로푸스 카펜시스'le lan t hrop us cap ens i s 로 알려졌던 이 화석은 지 금은 곧선사람으로 분류되고 있는데, 정확한 연대는 알 수 없다. 최근에 동아프리카의 두르카나 호수 일대를 조사하던 리처드 리키 팀은 1985 년에 호수 서편 지역의 나리오코몸에서 165 만 년 전에 살 던 곧선사람의 화석을 발굴하였다. 나이는 대략 9 세로 키는 160cm 정도이다(처음에 이 인류화석은 1 양] 정도로 보고되었으나 신체 성장
속도가 슬기사람과 어느 정도 차이가 있었다는 연구에 따라 9 세로 정정 되었으며, 이 소년 화석의 발견으로 곧선사람의 키는 슬기사람보다 작 았으리라는 생각은 잘못된 것으로 밝혀짐). 뼈대의 특칭으로 보아 남 자 아이로 판명된 이 호미니드는 지금까지 발견된 이른시기 호미니드 중 가장 완전하게 남은 화석이다(사전 8. 18).
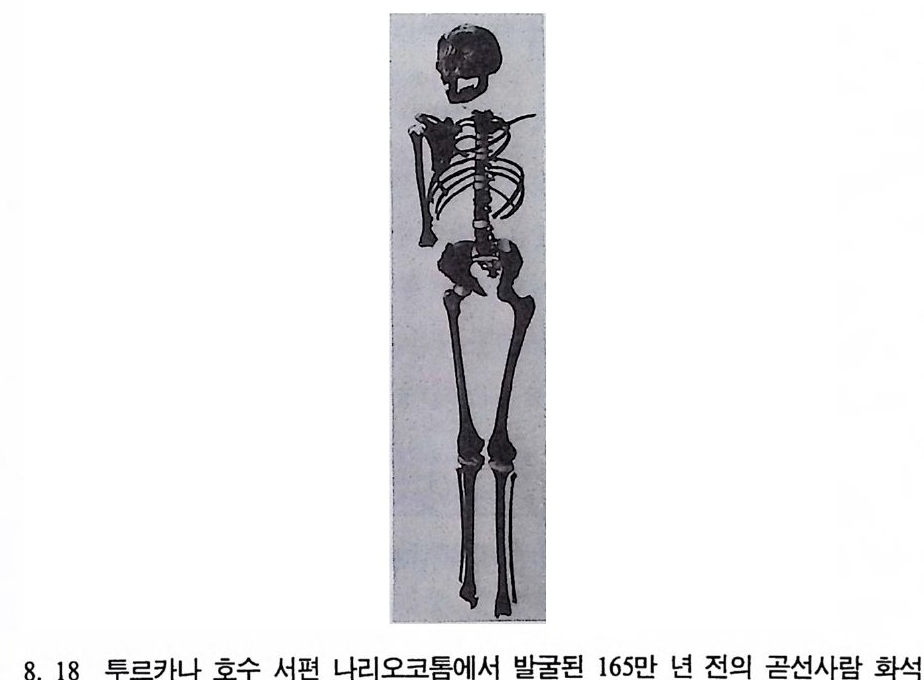 8. 18 투르카나 호수 서편 나리오코톰에서 발굴된 165 만 년 전의 곧선사람 화석
8. 18 투르카나 호수 서편 나리오코톰에서 발굴된 165 만 년 전의 곧선사람 화석
한편 유럽에서는 중기 플라이스토세에 속하는 여러 유적지에서 곧 선사람의 화석이 출토되었는데 이들은 주로 머리뼈 조각들로 이루어 져 있으나,스타인하임,스완스콤,베르데스쾰러스,빌정스레벤,아라 고 및 페트라로나 등지에서 발견된 화석들은 보존상태가 좋아 곧선사 람이나 옛슬기사람 Archa i c Homo sap i ens 으로 분류되었다. 아시아에서는 중국의 위엔모우(元謀)와 란덴(藍田), 그리고 저우커 우뎬 위층과 최근에 뮈샨(巫山)과 윤지안(限縣)에서 곧선사람의 화석
이 발견되었으며, 인도네시아에서는 자바의 상기란과 모조케르토, 삼 붕마찬 등에서도 많은 곧선사람의 화석이 발견되었다(그립 8. 19).
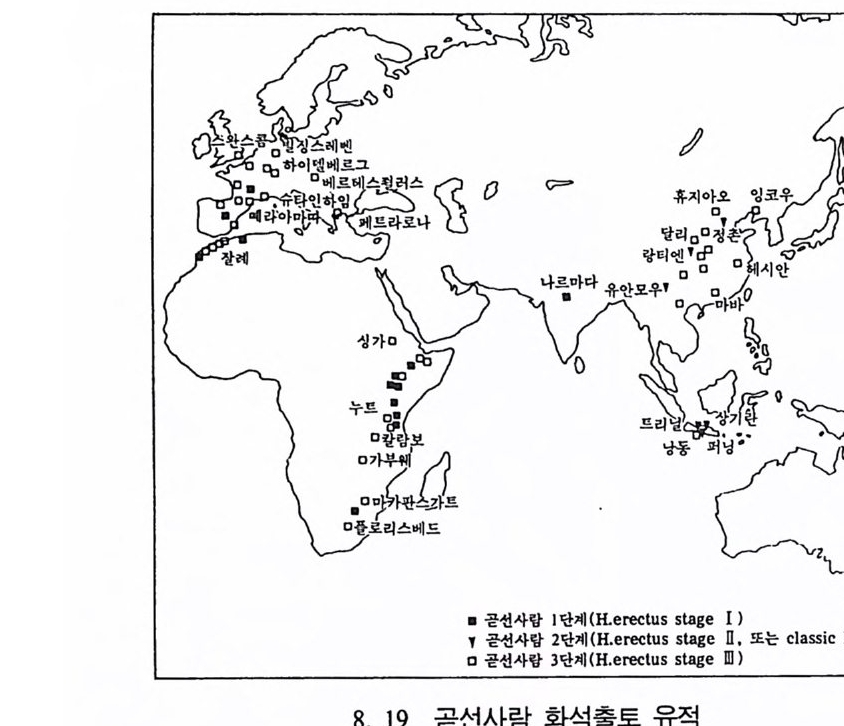 。 휴/ 지 쨩P잉 :/
。 휴/ 지 쨩P잉 :/
곧선사람의 해부학상 특징 곧선사람의 해부학상 특칭은 슬기사람과의 비교에서 얻어졌는데, 바이덴라이히, 마이어 E. Maye r, 르 글로스 클라크, 하웰 및 하웰스 W.W.Howells 같은 많은 학자들이 곧선사람의 특칭을 거론하였다. 비록 하웰이 곧선사람의 해부학상 특칭을 정의하는 데 있어서 란덴 과 아프리카에서 발견된 ER-3733 등의 머리뼈 특칭을 추가하였지만, 대개의 학자들은 오래 전에 아시아에서 발견된 화석을 근거로 하고 있다.
이들의 해부학상 특칭 중 머리뼈 용량은 저우커우덴에서 발견된 화 석들에 의해 매우 잘 알려졌는데, 이들의 머리 크기는 약 800- 1 200cc 에 이른다. 머리뼈 앞면에서 볼 때 앞이마는 낮고, 눈두덩이 뚜렷이 나타나 있으며, 뒷머리에는 옹이점이 두텁게 발달해 있다. 위에서 볼 때 곧선사람의 머리뼈는 넓적하며, 머리뼈 최대너비는, 현생인류인 슬 기사람의 경우 머리 위쪽에 최대너비가 있는 데 비해, 곧선사람은 귓 구멍 높이에서 얻어진다. 머리뼈는 매우 두꺼우며 얼굴은 앞으로 뒤 어나왔다. 이빨은 손쓴사람의 이빨만큼 크지는 않으나 슬기사람의 것 보다는 매우 크다. 사지뼈에서 볼 때 곧선사람은 슬기사람과 비슷하며, 루시 같은 오 스트랄로피테쿠스 아파렌시스나 손쓴사람에서처럼 긴 팔을 가지고 있 지 않다. 전체 몸의 크기로 볼 때 곧선사람은 손쓴사람보다 크며 거 의 현대 인류만큼 크다. 그러나 곧선사람이 슬기사람으로 전환되기 위해서는 머리뼈의 크기가 증가하고, 또한 턱이 발달해야 하며 눈두 덩과 머리 뒷두덩이 줄어들어야 한다. 우리가 곧선사람을 거론할 때 풀어야 할 문제 중 하나는 이들의 해 부학상 특칭이 언제 처음 나타났느냐 하는 점이다. 이 문제에 관한 가능한 답은 인류의 전화과정중 손쓴사람에서 곧선사람으로 전화해 가는 과정에서 전화의 방법과 속도를 설명해 주는 모델에서 찾을 수 있는데, 지금까지 가능한 두 개의 가설이 이를 설명해 주고 있다(그 립 8. 20). 두 개의 가설 중 하나는 전통으로서의 다윈의 견해를 따르는 것으 로서 손쓴사람에서 곧선사람으로 접전적으로 변하였다는 가설이며, 다른 하나는 엘드리지와 쿨드에 의해 제시된 계단식 평형 가설로, 이 에 따르면 곧선사람은 고립된 작은 손쓴사람 집단으로부터 짧은 시간 내에 빠른 속도로 발달했다고 주장된다. 1980 년대 초반까지 알려진 화석들을 바탕으로 한 연구들은 양쪽의 가설이 손쓴사람으로부터 곧
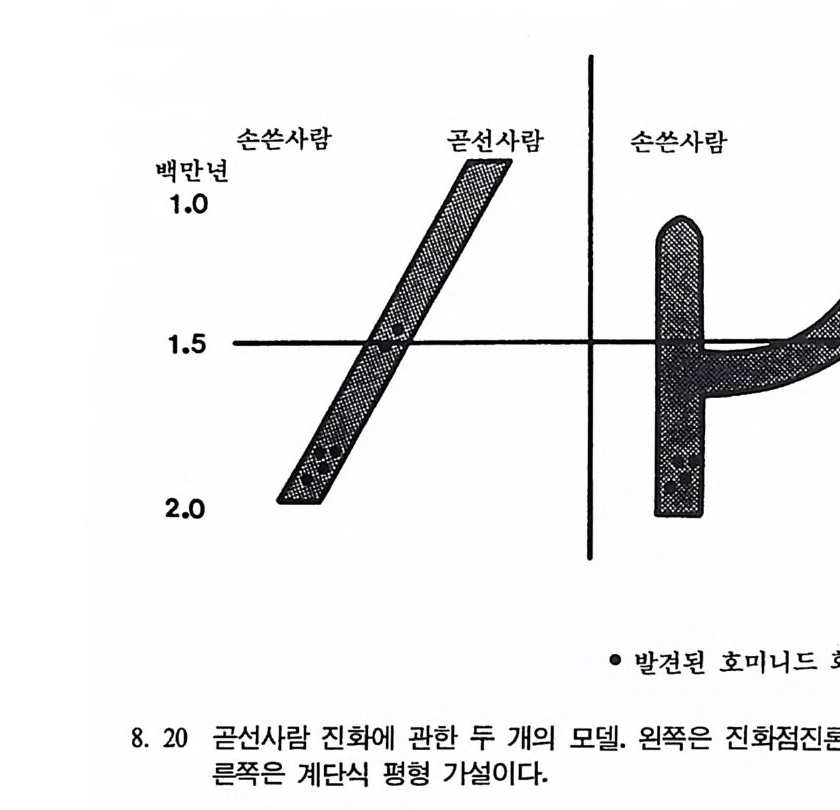 손쓴사람 곧선사람 손쓴사람 곧선사람
손쓴사람 곧선사람 손쓴사람 곧선사람
선사람으로 전화과정을 설명하는 데 적합하다고 생각되었다. 그러나 최근에 아프리카에서의 두 개의 호미니드 화석의 발견은 두 개의 가설 중 후자가 더 가능한 모델임을 보여주고 있다. 발견된 화 석 중 하나는 1980 년대 초에 올드바이 골짝에서 발견된 180 만 년 전 의 손쓴사람 화석이다. 과거에 손쓴사람의 사지뼈들은 상대적으로 모 두가 크다고 믿었다. 또 많은 학자들은 손쓴사람은 상대적으로 작은 몸집의 오스트랄로피테쿠스 아파렌시스와 커다란 몸집을 가전 곧선사 람의 전화상의 존재로서 인정되었기 때문에 손쓴사람의 몸집은 이들 의 중간 크기였으리라고 가늠하였다. 그러나 올드바이 골짝에서 출토 된 손쓴사람의 사지뼈는 과거의 해석과는 달리 루시의 사지뼈 크기 정도로 작다는 것이 밝혀졌다.
아주 중요한 또 다른 화석은 1985 년에 두르카나 호수 서쪽지역에서 발견된 160 만 년 전의 곧선사람 뼈대의 발견이다. 이 어린 곧선사람 의 뼈대는 팔다리의 비율이 우리와 거의 같고, 다 자랐을 때는 약 165~ 181cm 정도의 키를 가졌으리라고 가늠된다. 새로 발견된 이 두 개의 화석은 짧은 시간 내에 엄청난 변화가 호미니드 사이에서 일어 났음을보여주고 있다. 곧선사람의 경우에 아프리카에서 출토된 화석들과 아시아에서 출토된 화석을 비교해 볼 때, 160 만 년 전의 아프리카 출토 머리뼈와 50 만 년 전의 저우커우뎬 출토 머리뼈는 단지 머리 용량이 시간이 지 남에 따라 증가하는 경향이 있으나 모든 화석들이 기본적으로 같은 특칭을 지니고 있는 것으로 보아, 곧선사람은 오랜 기간 동안 거의 변하지 않았다고 말할 수 있다. 손쓴사람의 진화과정은 화석의 발견을 시간과 공간 내에서 이를 그 려보는 방법으로 살펴볼 수 있다(그립 8. 21). 아프리카, 아시아 그리
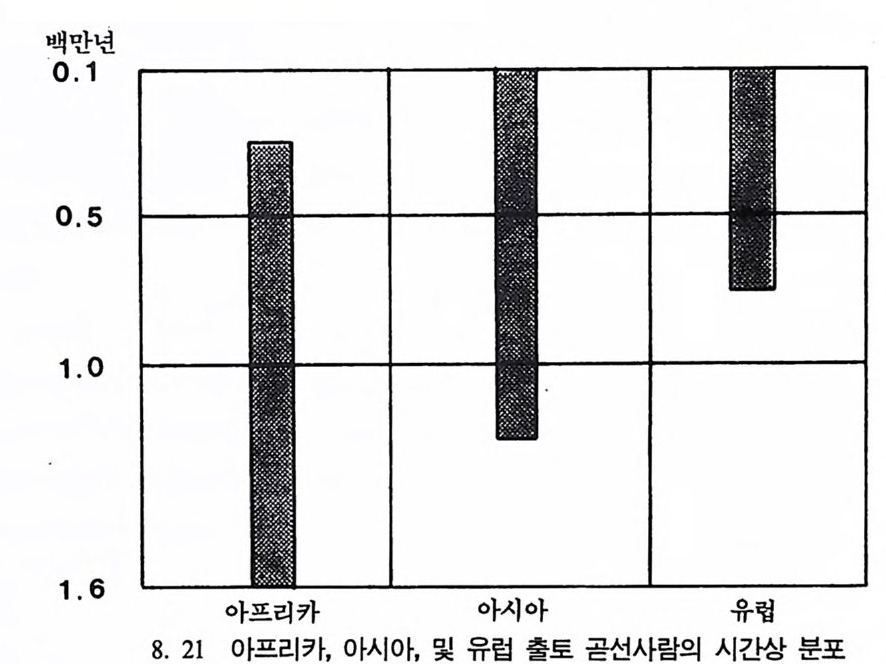 백만년
백만년
고 유럽에서 출토된 곧선사람의 연대는 절대연대 측정방법과 고지자 기측정법, 동물충위학 및 바다퇴적 분석법 동의 도움으로 알려져 있 는데, 지금까지의 연구에 의하면 아프리카 곧선사람의 시작은 170 만 년 전으로 밝혀져 있고, 아시아에서는 100 만 년이 조금 넘는 연대를 보여주며, 유럽의 호미니드는 50 만 년을 넘지 못하는 연대를 가지고 있다. 곧선사람은 풀라이스토세 중 빙하가 발달한 상당 기간 동안 살았는 데 이 시기 동안 모든 시기가 다 춥지는 않았다. 지구는 이 기간 중 에 몇 차례의 기후 변동이 있었으며 이런 기후에 적응해서 살던 동물 군과 식물군은 언제 기후가 춥고 언제 따뜻했나를· 알려준다. 기후의 변화는 단지 기온에 영향을 미친 것만이 아니다. 중기 플라이스토세 빙하기 중에 얼음량의 증가는 지구상의 많은 물을 끌어들이는 효과를 가져와 그 결과 바다 수위가 떨어져 현재 물밀에 잠겨 있는 거대한 지역이 육지로 나타났다. 과거에는 유럽의 알프스 빙하를 연구한 펜크와 브루커 Albrecht Penck and Edward Brucker 의 연구를 따라 플라이 스토세 에는 네 번 의 중요한 기온의 변동이 있었다고 생각되었으나, 오늘날은 샤크레톤 N. J. Schakle t on 과 옵다이크 N. D. O pd y ke 에 의 한 「바다퇴 적 물 연구」* 에 따르면 적어도 아홉 번의 중요한 기온 변화가 이 시기 중에 있었 던 것으로 알려지고 있다. 바다되적 연구는 바다 해저에 있는 퇴적으로부터 얻어전 지식 으로 바닷물 속에는 0/ 018 의 두 동위원소가 있는데, 바다 퇴적 물에 함유되어 있는 0 과 018 의 양을 비교해서 학자들은- 풀라이 스토세에 빙하기가 몇 번이나 있었는지 또 언제 시작되었는지를 밝혀낼 수 있다.
곧선사람 화석의 분류와 진화 늦은 초기 폴라이스토세인 160 만 년 전부터 시작해서 중기 풀라이 스토세에 아프리카와 아시아 , 유럽에서 출토된 호미니드 화석의 분류 에는 많은 문제가 있다. 아프리카 출토 호미니드는 시간의 선후에 따 라 곧선사람과 옛슬기사람으로 구분되었으며, 중기 풀라이스토세에 유럽에서 출토된 호미니드는 곧선사람과 슬기사람, 또는 곧선사람이 아닌 옛슬기사람만으로 분류되었다. 아시아의 경우, 전기 • 중기 풀라이스토세에 살던 호미니드의 전화 상의 위치와 분류는 중국과 인도네시아에서 발견된 화석을 중심으로 이루어졌다• 중국에서 발견된 호미니드는 모두 곧선사람으로 분류되 고 있지만, 인도네시아 출토 화석의 경우 〈종의 단위〉에서 볼 때 생 물학상 차이를 찾아볼 수 없으나 일부 인도네시아 학자들은 100 만 년 전부터 슬기사람이 나타나기 전까지 자바에서는 시기에 따라 곧선사 람을 포함해서 여러 종류의 호미니드가 살았다고 주장하였다(표 8. 3). 그러나 최근의 아프리카의 하다와 라에톨리에서 발견된 호미니드 화석의 연구에 따르면 이론시기의 호미니드 화석은 그 크기의 범위 가 매우 넓고, 성에 의한 차이가 있음을 밝혀주고 있어 인도네시아 호미니드 화석은 모두가 단일종인 곧선사람으로 분류되는 것이 타당 하다. 오늘날 고인류학자들은 호미니드 화석의 분류에 관해 많은 논쟁을 벌이고 있는데, 특히 전화상 중간형의 화석이 주요 논쟁의 대상이다. 1965 년에 캠벨 Bernard Cam p bell 이 호미니드를 분류하는 방법을 제시하면서 〈종〉보다 더 세분된 단계로 〈아종 subs p ec i es 〉 단계를 설 정하였다. 이후 많은 학자들이 캠벨의 분류를 받아들였다. 그러나 아 종 단계의 구분은 화석들 사이의 관계뿐만 아니라 전화상의 선후관계 를 설명할 수 없는 단점을 가지고 있으며, 실제로 〈아종〉이란 지역
표 8. 3 인도네시아 출토 호미니드 분류 1. 사르토노 Sar t ono, 1982 : 머리뼈 크기와 충위를 기준한 표 큰머리 늦은시기 곧선사람 낭동 곧선사람 H. e. nga ndong e nis Homo erectu s 트리 닐 곧선사람 H. e. trin i le n sis 옛자바사람 옛자바 로보스투스사람 H. p. robustu s Homo pal eoja v anic u s 옛자바 상기란사람 H. p. sang ira nsis 작은머리 Ol 른시기 2 퀘닉스발트 von Koen igsw ald, 1976 : 이빨 형태를 비교한 표 피데칸 E 쿠立人 곧선 피테칸드쿠~ P.erectu s Genus Pithe canth r op u s. 모조케르텐 피테칸트로프스 P. mojo k ert en sis 듀비우스 피테칸 E 쿠 立~ P. dubiu s 매간트로프스 옛자바 매간 E 쿠 立~ M. pal eoja u anic u s Genus Ma ga nth r op u s 3. 야곱J acob, 1975 솔로 피테칸드쿠 立~ P. soloensis 피데칸 E쿠 立人 곧선 피데칸'E쿠立~ P.erectu s 모조케르토 피데칸드 쿠 ~ P. mojo k ert en sis 4. 산타 루카 Santa Luca, 1980 곧선사람 곧선 곧선사람 H. e. erectu s (인도네시아) 베이칭 곧선사람 H. e. pek in e nsis (중국) 5. 포프 Pope , 1982 곧선사람 곧선사람 H. erectu s
사이의 차이만을 뜻한다• 한편 이 당시의 호미니드를 설명할 때 〈이론시기 또는 옛슬기사람 〉 이라는 용어와 소위 〈발달한 형태의 곧선사람 Develop ed Homo ere- c tus 〉 이라는 용어를 사용하는데, 이런 용어들은 단지 일반용어일 뿐 진화상 중간형을 의마하는 학술용어가 아니다. 따라서 가장 이론시기 에 속하는 아프리카 호미니드를 곧선사람 I 기로, 처음 찾아진 아시아 의 대표 호미니드를 곧선사람 11 기로 분류하고, 〈이른시기나 옛슬기 사람 또는 발달한 형태〉 등의 용어로 표현되는 호미니드를 모두 곧선 사람 冊기로 분류하는 방법이 최근에 제시되었다. 아프리카, 아시아 및 유럽에서 발견된 곧선사람 m 기에 속하는 호미니드 화석은 서로 아주 닮았으며, 슬기사람의 조건을 일부 보여주는 경향이 있으나 아 직 슬기사람의 단계에 도달하지는 못했다(사전 8. 22).
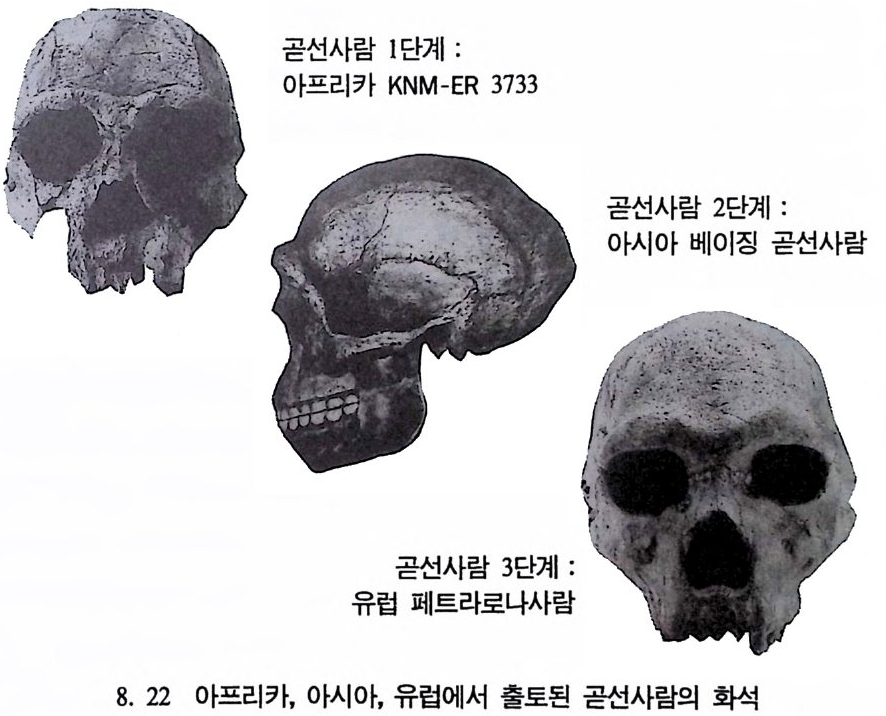 곧선사람 1 단계:
곧선사람 1 단계:
곧선사람의 기원지와 이동 19 세기 이래 학자들 사이에서는 인류가 어디서 처음으로 나타났는 지, 또 어떻게 퍼져 나갔는지에 대해 많은 논란이 있어왔다. 그간에 이루어전 호미니드 화석 출토 유적지의 연대와 화석의 해부학상 비교 연구 및 당시 환경에 관한 연구에 힘입어서 이 문제에 관한 설명이 가능하게 되었다. 19 세기중에 찰스 다윈울 포함한 많은 학자들은 아프리카를 인류의 기원지일 가능성이 가장 높은 지역으로 꼽았으며, 반면에 월리스 등 은 인류의 기원지를 아시아에서 찾고자 하였다. 따라서 19 세기 후반 부터 20 세기 초반까지 인류 조상의 화석의 발견은 아시아에서 주로 이루어졌으나, 2 伊1 ] 기 중반에 이르러 많은 수의 호미니드 화석들이 아프리카에서 발견되기 시작하면서 아프리카가 인류의 기원지로 받아 들여지기 시작하였다. 어떤 한 지역이 왜 인류의 기원지로 인식되는가? 고동물학상의 연 구에 따르면 동물들은 환경에 적응해야만 번성하게 된다고 한다. 이 때에 〈적응하여 번성〉이란 뜻은 바로 한 동물이 한 지역에서 지형상 으로 고립되어 있을 때 일어나는데, 고립은 기후라든가 산맥, 물의 조 건 및 사막 등에 의해 일어난다. 그리고 동물집단은 뚜렷한 지형상의 기원지를 가지고 있다는 고동물학상의 원리가 오랫동안 학자들에 의 해서 인류의 기원과 전화라는 문제에 적용되어왔다. 이때에 어떻게 지역의 집단이 시작되는지를 결정하는 것이 중요한 문제인데, 일반적 으로 이 문제의 해답은 어떤 지역에서 그 집단들이 나타나는가에 따 라서 결정된다. 최근의 연구에 따르면 중기 마이오세 이후, 인류의 전단계에 속하 는 〈원호미니드 Pro t o-Ho mi n i d 〉둘은 아프리카에서 번성하였으며, 마 이오세 말에 오면서 기후의 변화에 의해 이루어전 환경의 변화는 아
프리카 대륙에 사바나라는 독특한 환경을 제공하였다. 인류의 조상이 살기에 좋은 온도(임계온도)는 약 27°C 로 알려져 있는데, 이론시기 호미니드가 살던 시기에 이런 조건을 가진 지역은 아프리카의 사바나 지대뿐임이 밝혀졌다. 오늘날 고인류학자들은 오스트랄로피테쿠스 아파렌시스를 인류의 가장 먼 조상으로 받아들이고 있는데, 이 호미니드 화석은 동아프리 카의 아파 지역에서 나온 것으로 상한 연대가 500 만 년 전까지 울라 가고 있다. 따라서 오늘날 대부분의 학자들은 지금까지 발견된 호미 니드 화석과 고고학 유물들의 연구를 바탕으로 아프리카를 인류의 기 원지로 받아둘이고 있다. 풀라이오-풀라이스토세에 살던 인류 조상의 기원은 아프리카 기원설 과 아시아 기원설로 크게 나눌 수 있는데, 아프리카 기원설에 따른 이동 울 가상하면 다음과 같다. 첫째, 아프리카 대륙 기원의 오스트랄로피테쿠스가 유라시아로 퍼져 나간 후, 한 갈래가 아프리카로 되돌아와서 이른시기의 호미니드가 되었 울 경우와 둘째, 아프리카에서 살던 오스트랄로피데쿠스로부터 진화한 손쓴사람의 한 갈래가 유라시아로 이동하여 곧선사람의 조싱이 되었을 경우와 셋째, 아프리카에서 살던 손쓴사람에서 전화한 곧선사람이 유라 시아로 이동했을 경우로 요약해 볼 수 있다. 그런데 지금까지의 연구들 (예 : 호미니드 화석 및 고고학 유물들, 지형 및 동물)에 따르면 세번째 가설이 가장 타당한 것으로 받아들여지며, 이 세번째 가설을 바탕으로 세 갈래의 이동 경로가 추정될 수 있다(그림 8. 23). 첫번째 이동경로는 약 160 만 년 전에 동아프리카에 처음 출현한 곧 선사람의 한 갈래로부터 시작되었다고 추정할 수 있다. 이 갈래는 전 기 풀라이스토세 중반에 이미 아프리카 대륙을- 벗어나 아라비아 남단 을 거쳐 인도를 횡단하는 경로를 따라 이동하였다고 여겨진다. 이 경 로는 버마를 지나 한 갈래는 동쪽으로, 또 다른 갈래는 인도네시아의
자바로 이어졌으리라 추정된다. 이들을 아프리카로부터 떠나게 한 요인은 정확히 알 수 없으나, 당 시의 아프리카 • 아시아의 지형변화는」 이 전파 모델의 가능성을 뒷받 침해 준다. 지질학상 플라이오세 말에서 플라이스토세 초기까지 홍해
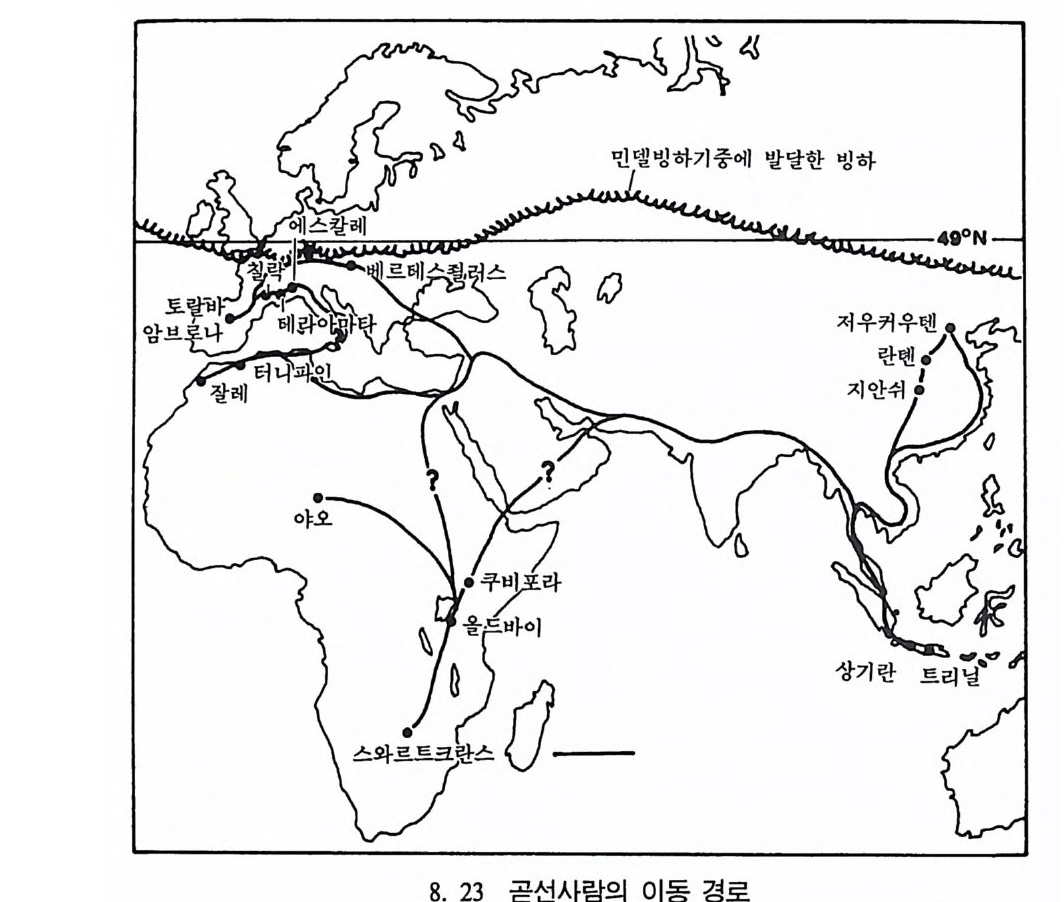 달
달
는 육지로 변해 에티오피아와 아라비아 반도는 붙어 있었던 것으로 밝혀져, 동물과 호미니드의 이동이 가능하였다고 여겨진다. 한편 인도 북부와 중국의 남부에 경계를 이루는 히말라야 산맥은 동물군과 호미 니드 이동에 결정적인 방해요인이었다. 따라서 버마를 거쳐 동북아시
아에 이르는 경로와 말레이시아를 거쳐 인도네시아에 이르는 경로만 이 가능한 이동경로로 여겨진다. 동남아시아의 경우, 대륙과 인도네시아 사이의 순다 해협 Sunda Shel f은 약 300 만 년 전과 125 만 년 전, 그리고 65 만 년 내지 45 만 년 전 사이에 육지로 변하였다. 순다 해협이 육지로 변했던 125 만 년, 65 만 년 내지 45 만 년 전에 동남아시아에 퍼져 있던 호미니드와 동물 들이 버마와 말라야 반도를 걸쳐 인도네시아로 이동하였으리라고 추 정된다. 두번째는 곧선사람이 유럽에 도달했던 경로이다. 이 경우는 다시 세 갈래의 가능한 이동경로로 나눌 수 있다. 그 첫번째가 중동아시아 를 거쳐 발칸 반도를 돌아 중부 유럽에 이르는 경로이다. 이때 아시 아 대륙에 위치한 자이고러스 산맥과 코카서스 산맥, 그리고 중부 유 럽에 위치한 카파치안 산맥과 카스피 바다는 곧선사람들이 이동하는 데 방해가 되었으리라 짐작된다. 유럽에 이르는 또 다른 길로는 아프리카 북서쪽의 해안을 따라 난 통로를 들 수 있다. 이 중 한 갈래가 아프리카 북서지역에서 지중해 의 시실리 섬을 거쳐 이탈리아 반도를 지나고, 프랑스 남부지역을 거 쳐 해안선을 따라 스페인에 도달했으리라고 추정된다. 이때 남부 프 랑스에서 북부 이탈리아 서쪽지역에 걸쳐 있는 알프스 산맥과 프랑스 와 스페인을 가르는 피레네 산맥이 이들의 이동에 장애가 되었으리라 여겨진다. 한편 곧선사람이 아프리카 북서지역과 유럽의 이베리아 반도 사이 롤 이동했다는 흔적은 보이지 않는다. 이는 아마도 두 대륙을 가르는 지브롤터 해협이 곧선사람의 이동을 가로막는 장애 역할을 했기 때문 이라고 여겨진다. 중기 풀라이스토세에 아프리카 북서지역에서 살던 동물들이 동서 방향으로는 이동을 하였지만, 남북 방향으로는 이동을 한 흔적이 없다는 연구는 곧선사람이 남북 방향으로 이동하지 못했으
리라는 가정을 뒷받침해 준다. 세번째 이동경로는 아프리카 대륙에서 찾아진다. 동아프리카에서 출발한 곧선사람은 아프리카 동부지역에서 길게 남북으로 이어지는 대협곡을 따라 남아프리카에 도달했으리라고 추정된다. 인류의 먼 조상은 아프리카에서 기원했다는 가설에 비해서, 아시아 기원설은 인류의 조상이 아시아와 아프리카 대륙에서 전화하여 서로 간에 유전자 교환이 있었다고 주장한다. 이 가설은 가장 오래된 호미 노이드가 인도와 파키스탄 일대에서 살았으며, 이 중의 한 갈래가 서 쪽방향으로 이동하여 아라비아 반도를 거쳐 700 내지 400 만 년 전에 아프리카에 이르렀다고 가정한다. 이들이 아프리카에서 가장 이론시 기의 인류의 조상이 되었을 것이며, 아파렌시스 화석이 아프리카와 아시아 두 대륙의 경계에서 발견되는 점이 이를 뒷받침하고 있다고 주장한다. 한편 또 다른 늦은 갈래는 동쪽 방향으로 향하여 버마를 지나 일부 는 중국으로 향하고, 일부는 인도네시아에 도달했으리라 여겨진다. 아 프리카로 이동한 같은 단계의 호미노이드가 남아시아와 동남아시아, 그리고 남중국으로 퍼져 나갔으며, 이 호미노이드의 후손인 오스트랄 로피테쿠스와 호모는 아시아로 퍼져 나갔고, 약 100 만 년 전을 조금 지난 시기에 곧선사람이 육지로 변한 순다 해협을 통해 인도네시아로 이동했으리라고 추정한다. 현재 대다수의 고인류학자들은 인류의 기원지에 관해서는 아프리카 기원설을 받아들이고 있다. 그러나 최근 남중국에서 발견된 새로운 호미니드의 화석은 아시아 대륙에서도 아프리카 대륙에서처럼 플라이 오-풀라이스토세에 세 종류의 인류 조상이 있었을 가능성을 제시해 준다.
곧선사람의 문화와 행위 곧선사람은 매우 성공적으로 살아 남았으며, 최초로 아프리카 대륙 울 벗어나 당시 환경이 나쁜 북남미 대륙과 오스트레일리아를 제의하 고 아시아와 유럽으로 퍼져 나갔다. 곧선사람들은 자신들보다 먼저 지구상에 살았던 손쓴사람보다는 더 발전된 문화를 가졌으며, 문화와 기술은 그들의 생존에 중요한 요소 가 되었고, 또한 문화는 그들 환경의 일부분이 되었다. 이들의 문화는 신체변화와는 관련없이 이루어졌고 비생물학적인 방법으로 다음 세대 에 전달되었다. 문화는 세대를 거쳐 계승되어갔으며 생물학상 변화보 다 더 빠르게 진행된 문화발달은 인류 생활에 즉각적인 변화를 불러 일으켰다. 문화의 발달에 따라 기술이 축적되자 곧선사람들의 신체상 에 변화가 일어났다. 어금니와 얼굴의 근육들 및 얼굴 등이 작아졌으 며 머리부피는 점차 커져갔다. 구대륙에 널리 퍼져 나간 곧선사람들은 많은 유적과 유물을 남겨놓 았다. 이들이 남긴 돌연모문화는 찍개문화와 주먹도끼로 대표되는 아 슐리안 문화로 나누어전다. 찍개문화는 아프리카의 〈발달한 울도완 문화〉의 전통을 지닌 문화로 곧선사람에 의해 아프리카와 유럽 그리 고 아시아로 전파되었다. 한편 아슐리안 문화는 올드바이 골짝에서 〈발달한 울도완 연모〉보다 늦은 시기에 발견된다. 아프리카 대륙 이의에 찰 알려진 가장 오래된 곧선사람의 유적지 중에 하나인 우베이디야 유적은 아프리카 큰골짝이 시작되는 이스라 엘의 골짝에서 발견되었다. 이 유적에서는 옛날에는 호수였으나 지금 은 메말라버린 바닥에서 돌연모들이 발견되었는데 100 만 년이 넘는 연대를 가지고 있다. 이 유적에서 나온 연모들은 소위 아슐리안 전동 으로 알려진 연모들로 올드바이 골짝에서 나온 연모들과 같은 시기이 다.
아슐리안 연모들은 사용하기에 매우 효과적이며, 대표적인 연모는 주먹도끼와 자르개이다.(그림 8. 24). 이 연모들은 주로 모롯돌떼기나 직접떼기 방법 등에 의해 제작되었는데, 아슐리안 연모를 대표하는 주먹도끼가 어떻게 쓰였는지는 정확히 알 수 없다. 지금까지 대부분 의 학자둘은 이 아슐리안 연모들이 아프리카와 유럽 그리고 인도 서 쪽지역에서만 발견되며, 인도 동쪽지역의 자바와 중국 등의 아시아 대륙에서는 이 전통이 나타나지 않는 대신 찍개문화만이 나타난다고 주장하고 있다. 한편 남중국과 안도네시아 동지에서 살던 곧선사람들 은 대나무로 만든 연모를 지닌 문화가 있었으리라고 추정된다. 오늘날 우리가 알고 있는 곧선사람의 행위와 그들의 기술에 관한 지식은 주로 저우커우뎬 유적지로부터 얻어졌다. 저우커우뎬 유적지
 8. 24 아슐리안 주먹도끼와 자르개
8. 24 아슐리안 주먹도끼와 자르개
에서는 많은 곧선사람의 화석과 돌연모, 풍부한 동물 화석 및 불에 탄 뼈와 재 등의 자료들이 나왔다. 불의 사용은 인류를 특정짓는 중 요한 요소인데 가장 오래된 불의 사용은 250 만 년 전에 아프리카 케 냐의 체소완자 유적에서 그 흔적을 찾을 수 있다. 그러나 일반적으로 고고학자들이 불의 사용에 관해 말할 때는 불에 탄 뼈들과 재의 발견 울 바탕으로 해서 곧선사람 단계에서 불을 사용하였으며, 여기서 발 견된 뼈둘은 사냥행위의 결과라고 오랫동안 믿어왔다. 불의 사용은 여러 면에서 인류에게 유익한데, 우선 불은 인류가 살 던 환경을 따뜻하게 해주어 환경의 적응도를 높여주며, 맹수를 물리 치는 데 사용되었다. 또한 불로 음식을 조리하는 것은 특히 식물성 음식이 지닌 독성을 중화시키는 작용을 함으로써 식량의 사용폭을 넓 혀주는 이점을 가졌다. 그러나 저우커우덴에서 발견된 불의 사용 혼 적은 곧선사람의 행위라기보다는 자연발화라는 주장과 여기서 남겨진 뼈둘이 과연 곧선사람들의 사냥행위에 의한 것인지에 대한 의문이 빈 포드 등에 의해 제기되고 있다. 인류의 두뇌 : 얼개와 그 기능 인류학자들은 인류의 전화과정에서 호미니드의 두뇌(골)는 어떻게 변했을까에 대해서 많은 관심을 가지고 있다. 골의 크기는 지난 300 만 년 동안에 빠르게 변하였는데 (사전 8. 25), 여기서는 두뇌의 얼개 와그기능을살펴본다. 두뇌의 일개 두뇌는 사람에 있어 가장 복잡한 구조로 되어 있지만 연구할 수 있는 얼개를 가지고 있다• 두뇌는 단백질과 지방 그리고 단수화물로 된, 마치 말랑말랑한 두부 같은 형태로 1.12 -1. 35 kg 정도 의 무게가 나간다. 두뇌논 100 억 개의 신경세포를 가지고 있는데, 이
 8. 25 두뇌 측정 입체 플로터를 사용하여 인류 두뇌 윤곽을 조사하고 있는
8. 25 두뇌 측정 입체 플로터를 사용하여 인류 두뇌 윤곽을 조사하고 있는
신경세포들이 신경에 충동을 보내고 받아들이는 역할을 한다. 이때 신경세포와 세포 사이의 신경충동의 전달은 〈전기작용〉과 〈화학작 용〉에 의해 조절된다. 대표되는 신경세포는 세포 자신이 몸체를 가지고 있는데, 신경충동 울 가지로 보내고 받아들이는 역할을 한다. 신경충동은 세포 사이에 있는 신경세포연접을 신경전달물질이라고 하는 화학물질을 통해 통과 한다. 신경전달물질에는 많은 종류가 있는데 그 중 하나가 도파민이 라 불리는 물질로 이 도파민이 부족하면 나이 먹은 사람에게 주로 나 타나는 파킨슨병에 걸리게 된다. 의사들은 환자들의 두뇌를 수술하면서 뇌에 대해 많은 것을 배우
나, 우리는 이것에 대해 완전히 이해하지 못한다. 신경세포에 대해서 알기 시작한 것은 불과 100 여 년 전부터이며, 특히 신경전달물질에 관한 연구가 시작된 것은 불과 50 여 년이 되지 못한다. 실제로 두뇌 는 영양을 공급하고 보호해 주는 다른 조직들에 의해 둘러싸여 있기 때문에 관찰하기가 매우 힘들다. 그러나 두뇌에 착색을 한다든지 또 는 다른 방법에 의해서 볼 수 있게 되었다. 두뇌의 갈색 바깥 부분은 세포 몸체들을 감싸고 흰색의 안쪽 부분은 세포를 감싸고 있다. 두뇌의 구조상으로 볼 때 인류의 골은 특별하지 않으며 오랜 진화 상의 계열과 일치한다. 해파리 같은 생명체에도 신경세포가 있으며, 모든 젖먹이동물은 인류와 같은 기본구조의 골을 가지고 있다. 골은 골간과 작은골, 그리고 큰골을 가지고 있는데, 동물의 지능은 크게 큰 골의 크기와 몸의 크기에 비례해서 결정된다. 사람은 다른 동물과 비교 해 볼 때 몸의 크기에 비해 큰 골을 가지고 있다. 그러나 사람 사이 에 있어서 골의 크기와 지능간에는 관련이 없다. 두뇌의 기능 두뇌는 매우 섬세하며 유액충과 세 겹의 조직 그리고 뼈로 둘러싸여 있다. 만약 두뇌가 심하게 타격을 받으면 의식을 잃지 만 죽지는 않는다. 죽 숨쉬고 심장이 박동하고 또 다른 주요 기능들 은 계속되는데, 이런 행동들은 두뇌의 가장 깊은 곳에 있는 골간에 의해서 조정된다. 골간에 의해 조정되는 중추신경은 마음의 상태나 밖으로부터의 조 철 없이 두뇌의 기본행동들을 조절한다. 이는 두뇌의 남은 부분을 일 깨우고 혈압을 조절하며, 침을 만들어내고, 기타 살아가는 데 필요한 기능들을 조철한다. 만일 이 중추신경에 손상이 오면 중풍에 걸리거 나 아니면 혼수상태에 빠지게 되며 심하면 죽음의 원인이 된다. 작은골은 근육을 조절하며 배고품과 다른 행동둘을 조절한다. 작은 골은 우리가 원하는 행동을 통해 근육을 부드럽게 조절하고 몸의 평
형을 유지하게 한다. 따라서 우리가 한 가지 행동을 반복하면 작은골 은 그만큼 기능이 더 좋아전다. 동맥경화는 작은골이 몸의 근육과 함 께 운동하는 것을 방해한다. 한편 작은골은 골의 중추신경처럼 그 기 능이 자동으로 작동하나 이런 기능은 밖으로부터의 반응에 의해서 일 어난다. 사람들의 자발적인 행동들은 큰골에 의해서 조절되는데, 큰골은 세 부분으로 나누어진다. 가장 안쪽 부분은 자동적으로 체온과 배고품, 졸음과 다론 기능들을 조철한다. 가운데 부분은 감정이라든지 배움, 그리고 냄새를 맡는 기능을 조절한다• 다른 감각기능들은 가장 바깥 충 Cor t ex 에 모여 있다. 이성이라든지, 말, 그리고' 생각을 담당하는 부분으로 나누어져 있다. 과학자들은 수술을 통해서 골을 배워왔으며 맛과 감각 갇은 것을 담당하는 곳을 나타내는 두뇌 지도를 만들었다. 또한 골 손상의 효과 로 많은 지식을 알게 되었다. 몸 왼쪽에 있는 근육과 감각은 머릿골 바른쪽에 있는 신경세포에 의해 조절된다. 골 중추신경에서 서로 교 차되는 길은 없다. 비록 골의 양쪽 둥근 무 분들은 똑같은 모양으로 보이지만 기능에서는 같지가 않다. 두뇌는 언어기능을 담당하는 브로 카 부위 (령 : Broca's Area) 와 베르니크 부위 (령 : Wern ick e's Area) 라는 두 개의 주요 부위를 가지고 있는데 둘 다 골 바깥충의 왼쪽에 있다(그립 8. 26). 이 부위들의 손상은 곧 언어상의 문제를 가지고 오 나, 오른쪽에 있는 감은 부위의 손상으로는 아무런 문제가 일어나지 않는다. 인류의 전화과정중 두뇌에서 언어를 담당한 브로카와 베르니크 부 위는 손쓴사람인 KNM-ER-1470 에서 희미하지만 이미 형성되었다 고 화석을 연구한 홀로웨이가 보고하고 있다.
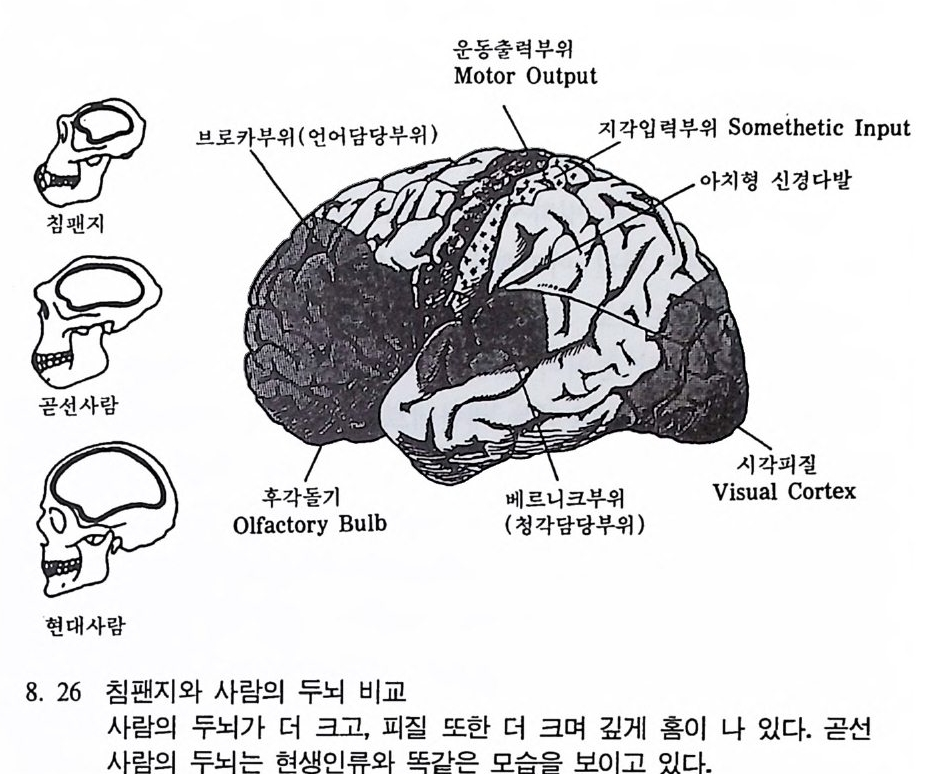 운동 출 력부위
운동 출 력부위
네안데르탈사람 Homo neander t halens i s 의 출현 오늘날 고인류학계에서 가장 열띤 논쟁을 벌이는 주제 중의 하나는 네안데르탈사람이 인류의 전화과정에서 차지하는 위치에 관한 것이 다. 킹 K i n g이 1864 년에 네안데르탈사람을 〈호모 네안데르탈렌시스 Homo neande rt halens i s 〉로 분류하여 우리와는 다른 종으로 구분하였 다. 그러나 1964 년에 캠벨이 이들을 우리와 같은 〈종〉에 속하는 〈호모 사피엔스 네안데르탈렌시스 Homo sap ien s neanderth a lensis > 로 분류한 이후 고인류학계에서는 지금까지 이 낱말을 사용해 왔는데,
최근에 네안데르탈사람의 명칭에 관한 논쟁이 재연되고 있다. 최초의 네안데르탈사람 화석은 일찍이 1800 년대 중반에 발견되었기 때문에, 오늘날 이들은 선사인류의 대명사처럼 사용되어져 왔다. 실제 로 네안데르탈사람의 화석이 처음 발견된 후 여러 해 동안 네안데르 탈사람은 구석기시대의 빙하시대에 살던 호미니드로, 튀어나온 눈두 덩과 둔탁한 턱을 가전 머리형태에 굳센 근육을 지녔고, 동이 굽고 휘청거리며 걷는 존재로 묘사되었다. 이들은 인간으로서 마땅히 지녀 야 할 마음을 조절할 능력이 없는 흉포한 존재로 인식되었다. 우리가 일반적으로 사용하는 네안데르탈사람이란 용어는 인류의 진 화과정중 그들이 차지하는 위치와 모습을 나타내는 학명이 아니라 단 지 그들이 지닌 인간성과 우리들 자신의 안간성을 식별하고자 하는 의미를 내포하고 있다• 최근에 새로운 화석의 발견과 연구는 이런 인 식이 잘못되었다는 사실을 밝혀주고 있다. 10 만 년 전부터 4 만 년 전 사이에 이들은 슬기사람이 살았던 지역에서 함께 살았으며 자신들 의 독특한 연모와 사회조직 및 세계관 등을 지니고 있었다. 분명히 이들은 지식을 지닌 인류 조상의 한 갈래였다. 그러면 왜 네안데르탈사람에 관한 잘못된 인식이 생겨나게 됐는가? 그 원인으로는 당시에 알려진 호미니드 화석이 거의 없었다는 점과 복원이 찰못된 화석을 바탕으로 한 지식 및 인류전화에 대한 부족한 이해 등을 들 수 있다. 그러나 무엇보다도 중요한 원인은 이 호미니 드 화석들이 발견된 시기에 있다고 하겠다. 유럽에서 최초로 발견된 선사시대 호미니드는 네안데르탈사람의 화 석이었는데 『종의 기원』이 출간되기 이전의 유럽 사회의 지적 분위기 는 네안데르탈사람을 인류의 조상으로 인정할 수가 없었다. 오늘날 학자들은 네안데르탈사람이 그들의 조상인 곧선사람에 비해 전화했다 는 사실을 알고 있다. 그러나 네안데르탈사람이 처음 발견된 당시 이 호미니드를 슬기사람과 비교했던 학자들은 이들이 서로 닮은 점보다
는 차이가 더 많다는 사실에 충격을 받았다. 오늘날 학자들은 네안데 르탈사람이 등이 굽고 다리가 흰 모습 대신에 근육이 발달하고 현대 사람의 평균 키보다는 작은 땅딸막한 몸집을 지니고 있었다고 얘기한 다. 최초의 네안데르탈사람 최초의 네안데르탈사람은 1848 년 지브롤터에서 발견되었으나 이후 50 여 년간 주목을 받지 못했다. 사람들이 관심을 두기 시작한 것은 1856 년 독일의 라인 강 지류인 뒤셀 강 근처에 있는 뒤셀도르프 시에 서 멀지 않은 곳에 있는 네안더 골짝에서 발견된 화석인류에 대해서 였다. 인부들이 이 골짝에서 석회암울 캐던 중, 머리통과 갈비뼈들 그 리고 엉덩뼈의 일부와 사지뼈들 일부를 발견하였다. 아마도 완전한 형태의 뼈대가 남아 있었으나 석회암을 캐기 위한 발파중에 많은 부 분이 유실되었으리라 여겨진다. 처음에 뼈둘은 고등학교 과학선생인 풀로트J.K. Fuhlro tt에게 보내졌는데, 그는 이를 노아의 홍수 때 동 굴로 쓸려 들어간 〈 이른사람〉으로 생각하였다. 그러나 이 화석을 본 일단의 학자들은 이 화석의 주인공은 나폴레옹을 추격하던 병든 코사 크 사람으로 판정하거나, 아니면 유럽 북쪽에서 살던 야만인 또는 에 이레 사람 등으로 설명하였다. 이후 이와 비슷한 화석들이 1886 년에 벨기에의 스피 근처의 한 동 굴에서 발견되었다. 한편 네안데르탈사람에 관한 연구는 프랑스에서 주로 이루어졌는데, 그 이유는 유적지들이 잘 보존되었고 또한 연구 에 많은 돈을 두자하였기 때문이다. 가장 중요한 네안데르탈사람의 발견은 1908 년에 프랑스 중부의 도르도뉴 지방의 여러 유적지에서 이 루어졌다(사진 8. 27). 나이가 든 네안데르탈 남자의 머리뼈가 라 샤 펠 오 솅 마울 근처의 동굴에서 발견되었으며, 르 무스티에 근처 동
굴에서는 어린 개체의 뼈대가 돌연모와 함께 발견되었다• 라 펠라제 바위그늘에서는 어른 남자 및 여자와 여러 개체의 어린아이 뼈둘이 함께 발견되었으며, 라퀴나 근처의 바위그늘에서도 여러 개체의 네안 데르탈사람의 뼈대가 발견되었다 .
 . ·- , ·-'. ,
. ·- , ·-'. ,
프랑스에서 출토된 화석들은 완전한 형태로 남아 있다는 점에서 네 안데르탈사람을 연구하는 데 무엇보다도 중요한 근거를 제공하였다. 그 결과 앞에서 언급된 네안데르탈사람에 대한 설명은 점차 설득력을 잃게 되었으며, 네안데르탈사람이 인류의 진화과정중에서 차지하는 중요성이 점차 인식되기 시작하였다. 네안데르탈사람에 관한 복원연구는 라 샤펠 오 솅에서 출토된 화석 으로부터 시작되었다. 이 유적에서 출토된 화석은 1908 년에 당시 프 랑스가 배출한 뛰어난 고동물학자인 마르셰린 부울 Marce lli n Boule 에 의해 네안데르탈사람을 대표하는 화석으로 복원되었다. 부울은 네 안데르탈사람을 털복숭이에 엄지발가락은 밖으로 향하고 등뼈는 유인
원을 닮았으며 다리는 안쪽으로 휘었다고 묘사하였으며, 네안테르탈 사람은 이들을 계승한 크로마뇽사람에 비해서 야만적인 의모와 지적 으로 열등하다고 주장하였다. 여러 면에서 부울이 복원한 네안데르탈 사람은 인류와는 다른 모습이었다(그림 8. 28). 아마도 오늘날 우리들 머릿속에 박혀 있는 네안데르탈사람에 대한 부정적인 인상은 부울의 설명에 원인이 있는 것으로 알려져 있다.
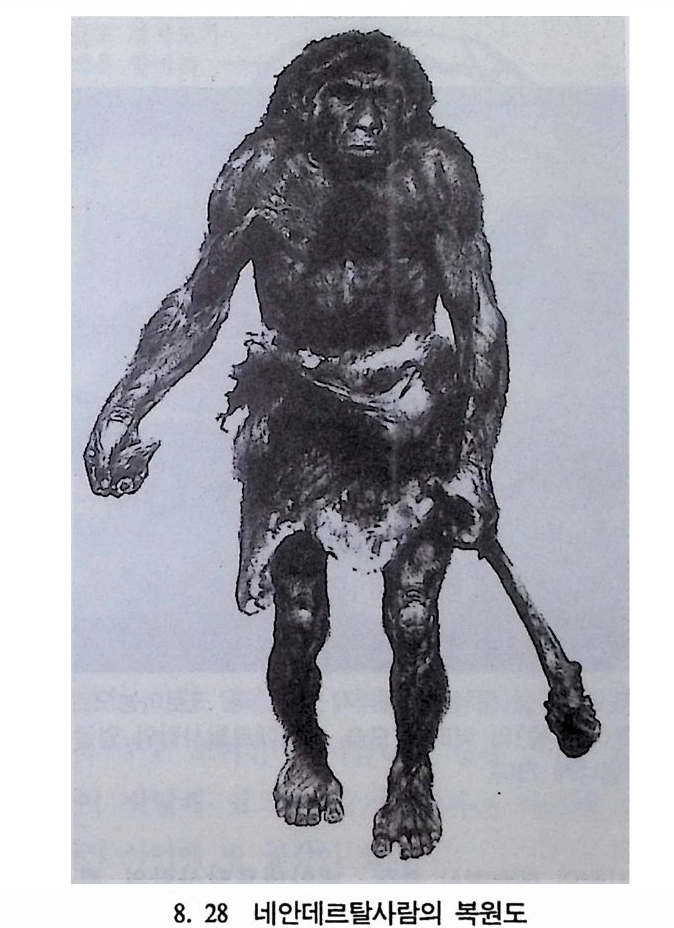 8. 28 네안데르탈사람의 복원도
8. 28 네안데르탈사람의 복원도
부울은 화석이 완전한 상태로 발견되었음에도 불구하고 네안데르탈 사람을 해석하는 데 많은 실수를 범하였다. 네안데르탈사람의 엄지발
가락은 밖으로 향해 있지 않으며, 무릎뼈에 관한 그의 견해는 부정확 하였다. 부울이 범한 또 다른 중요한 실수는 네안데르탈사람의 골 크 기를 잘못 해석한 데 있다. 그는 네안데르탈사람의 특칭인 긴 머리에 낮은 앞머리는 이들이 지적으로 열등하다는 것을 보여주는 증거로 판 단하였다. 그러나 부울 이후 최근까지 진행된 네안데르탈사람에 대한 많은 연구는 네안데르탈사람의 지능이 슬기사람과 비교할 때 크게 차 이가 나지 않음을- 보여주고 있다.
 8. 29 네안데르탈사람(왼쪽 ; 라펠라제 출토)과 크로 □ ~으로 대표되는 현
8. 29 네안데르탈사람(왼쪽 ; 라펠라제 출토)과 크로 □ ~으로 대표되는 현
네안데르탈사람의 해부학상 특징 네안데르탈사람의 해부학상의 특칭 은 현생인류인 슬기사람에 비해 여러 면에서 비교가 된다(사진 8. 29). 네안데르탈사람의 콧궁과 눈굼은 매우 크다. 전체적으로 두텁게 발달해 있는 눈두덩은 미간 부위와 양눈두덩 끝에서 두께가 약해진
다. 옆에서 보았을 때 앞머리뼈는 낮고 얼굴은 앞으로 튀어나와 있다. 뒷머리뼈의 가운데 부위가 약간 불룩 나와 있다(이 부분을 이라고 부른다). 뒤에서 본 머리형태는 슬기사람이 5 각형인 데 비해 달걀형이다. 머리 최대너비는 슬기사람보다는 밑으로 처져 있어 머리 통 중간 부분에서 얻어진다. 위에서 본 머리뼈는 앞뒤가 길고 뒷머리 뼈로 가면서 밖으로 향하여 편편해진다(그립 8. 30).
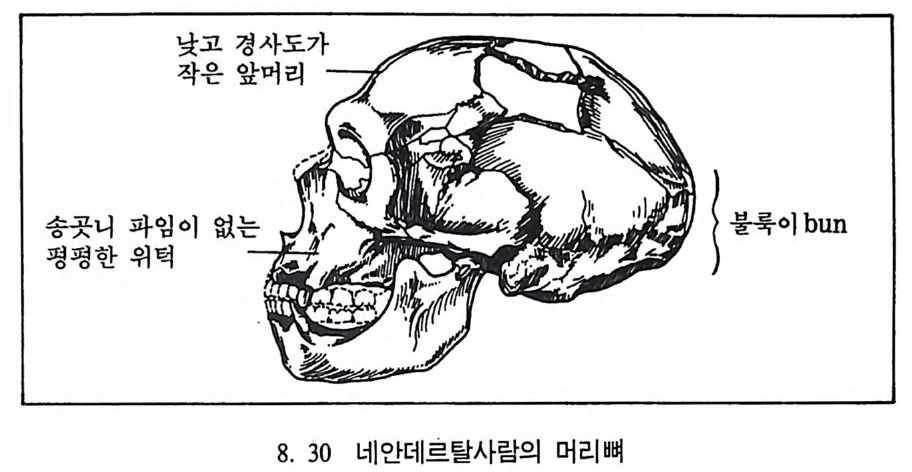 송곳니 파임이 없는 }탸J bun
송곳니 파임이 없는 }탸J bun
아래턱의 경우 〈턱 가운데 부위〉가 위아래로 밋밋한데 이런 모양 은 곧선사람과 닮았다. 슬기사람의 경우 이 부위의 가운데부분이 안 쪽으로 휘어 들어갔다. 이빨의 경우 앞이빨은 어떤 경우에 매우 닳아 있는데, 앞이빨의 이런 닳음새는 마치 오늘날 가죽을 가지고 일을 하 는 사람들에서 종종 보여진다. 어금니의 경우 〈이빨 내부의 공간(타 우로돈티즘)〉이 발달해 있으며, 슬기사람과는 다르게 마지막 어금니 와 아래턱 가지 사이에 빈 공간이 있다. 사지뼈는 굳세 보이고 두터우며, 특히 뼈의 밑부분들은 짧고 손가 락과발가락은크다. 위에서 살펴본 네안데르탈사람의 해부학상 특칭을 바탕으로 일련의
복원작업이 이루어졌는데, 부울 등에 의해 시도된 최초의 복원에 따 르면 네안데르탈사람은 땅딸막한 몸집에 큰 손과 발을 가전 야만인으 로 묘사되었다. 그러나 오늘날의 복원은 유인원들과 현대사람에 관한 뼈대와 근육들에 관한 지식을 바탕으로 전문가들에 의해 이루어지고 있는데, 이 새로운 복원에 따르면 네안데르탈사람들은 현재 추운 지 방에서 살고 있는 우리들과 비슷한 해부학상 특징을 지니고 있다고 한다. 네안데르탈사람의 기원과 인류진화상 위치 네안데르탈사람이 언제, 어디서 시작했는가 하는 문제와 현생인류 인 슬기사람이 출현하는 시기와 같은 시기에 살았던 이들이 슬기사람 으로의 전화과정에서 어떤 역할을 했는지는 슬기사람의 기원을 이해 하는데 매우중요하다.
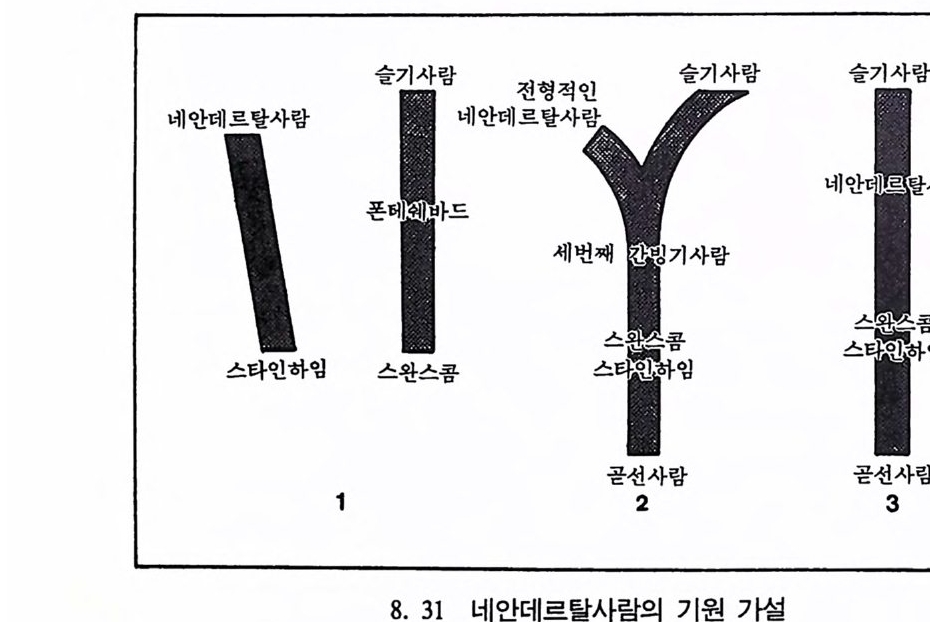 I 술기사람전형적인 슬기사람 술기사람
I 술기사람전형적인 슬기사람 술기사람
1980 년대 중반까지 고인류학자들은 네안데르탈사람과 40000- 3 5000 년 전에 나타나는 슬기사람과의 관계를 아래와 같은 3 개의 가설로 요 약하였다(그림 8. 31 ). 1. 선 슬기 사람 가설 Pre-S ap ien s Hy po th e sis 2. 선 네안데르탈 가설 Pre- N eanderth a l Hy po th e sis 3. 네안데르탈 조상 가설 Neanderth a l Phase of Man Hy po th e sis 첫번째 가설은 네안데르탈사람을 독일의 스타인하임에서 출토된 곧 선사람의 후손으로 인정한다• 이 가설에 따르면 네안데르탈사람은 해 부학상으로 너무 특수화되었으며 바이첼 빙하기 때 절멸했다고 한다. 한편 슬기사람은 스완스콤에서 시작하여 폰테쉐바드 단계를 거쳐 후 기구석기 문화를 담당한, 해부학상으로 현대사람과 닮은 슬기사람으 로 이어졌다고한다. 두번째 가설에 따르면 곧선사람에서 진화한 호미니드가 스완스콤과 스타인하임 단계를 거쳐 세번째 간빙기에 살았던 호미니드로 진화하 였다고 한다. 네번째 빙하기인 바이첼 빙하기중 이들은 추운 환경에 고립되었으며, 살아 남기 위해 여러 차례의 자연선택을 거치며 특수 화된 해부학적 특성을 지니게 되었다고 한다. 오늘날 서부 유럽에서 발견되는 대표적인 네안데르탈사람이 바로 이들로, 이때 이들 사이에 는 제한된 유전자 교환이 있었다고 한다. 최근에 학자들은 아프리카 의 오모와 라에톨리, 그리고 보더 동굴 등지에서 발굴된 해부학상 슬 기사람 화석을 근거로 네안데르탈사람의 조상은 유럽 밖의 지역, 죽 아프리카 대륙을 고향으로 하고 있다고 주장한다. 세번째 가설은 단일종 가설로 네안데르탈사람은 슬기사람과는 다른 가지가 아닌, 슬기사람의 직접조상으로 진화해 왔다고 인정하고 있다. 그러나 1985 년 이후 네안데르탈사람과 슬기사람 화석의 발견과 이 들에 관한 새로운 연구는 이들의 기원과 진화상의 위치에 관해 새로 운 해석의 증거를 제공하고 있다. 오늘날 고인류학자들의 견해는 〈네
안데르탈사람을 네안데르탈 술기사람 Homo sap ien s neanderth a lensis > 이라 부르고, 아주 먼 세대를 올라가는 슬기사람의 직접조상으로 인 정해야 하는가, 아니면 그들을 〈 호모 네안데르탈렌시스 Homo nean- de rt halens i s 〉라는 독립된 종으로 분류하고 우리의 직접 조상과는 거 리 룰 두고 갈라져 나간 다른 갈래 인가로 크게 구분된다. 최근에 이루어전 일련의 연대측정 결과는 슬기사람이 네안데르탈사 람과는 다른 갈래라는 점을 강하게 시사하며, 위의 두번째 가설에서 주장하는 네안데르탈사람의 조상은 유럽 밖의 지역, 죽 아프리카대륙 에서 시작됐으리라는 가설이 지지를 받고 있다. 특히 네안데르탈사람 들이 중동아시아 지역에서 이론 슬기사람보다 시간적으로 앞서기도 하며 또 늦게도 나타나는데, 이는 이론 슬기사람들이 다른 지역으로 부터 이 지역으로 이주해 왔기 때문이다. 새로운 연대와 더불어 고생물학적인 증거는 네안데르탈사람을 독립 종인 호모 네안데르탈렌시스라고 분류해야 한다는 견해를 뒷받침해 준다. 중동아시아의 게바라 동굴에서 발견된 화석은 슬기사람과 네안 데르탈사람의 혼혈 흔적이 보이지 않으며 오직 네안데르탈사람의 전 형적 인 특정만이 나타난다. 한편 30000 년 내지 40000 년 전에 이스라 엘과 레바논 지역에서 살았던 슬기사람의 화석에서도 네안데르탈사람 의 유전자 교환 흔적이 나타나지 않는다. 따라서 네안데르탈사람과 슬기사람은 약 20 만 년 전에 살았던 공통조상에서 갈라져 진화하여 네안데르탈사람은 유럽에서, 슬기사람은 아프리카에서 각각 다른 방 향으로 전화해 서로 사이에 유전자 교환이 없었으며, 고고학상 다소 문화교류가 있었다는 견해가 고인류학자들 사이에서 일반적으로 받아 들여지고 있다.
네안데르탈사람의 문화와 행위 무스테리앙 돌연모 문화 네안데르탈사람의 화석은 독일, 프랑스, 이탈리아, 스페인 그리고 지중해 근처에서 발견된다(그림 8. 32). 이들에 관한 연대기는 잘 연 구되어 있는데, 이는 그들이 방사선 탄소 동위원소 연대측정이 가능 한 시기까지 살았기 때문이다. 네안데르탈사람이 살던 세계는 오늘날 의 세계와는 아주 다르다. 빙하가 스칸디나비아와 북부 독일 및 영국 의 대부분을 덮고 있었으며, 그 의 유럽의 나머지 지역은 둔드라 지 대의 특칭을 가지고 있었다.
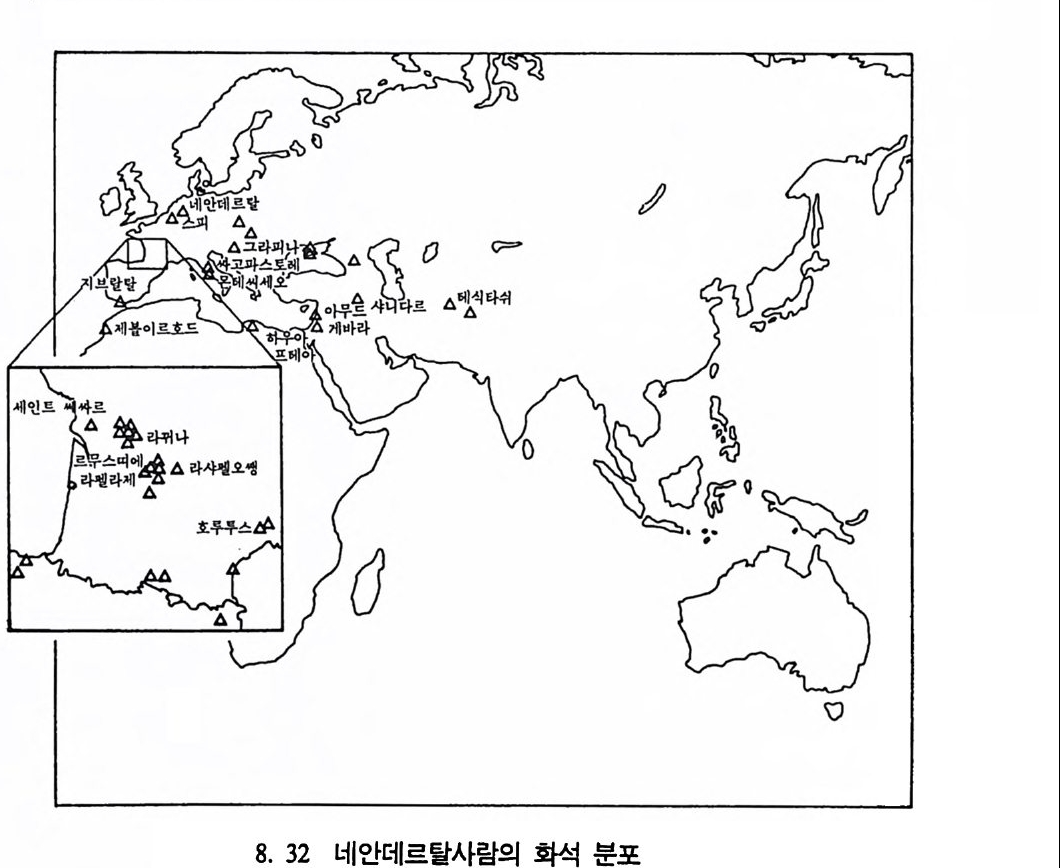 / _:
/ _:
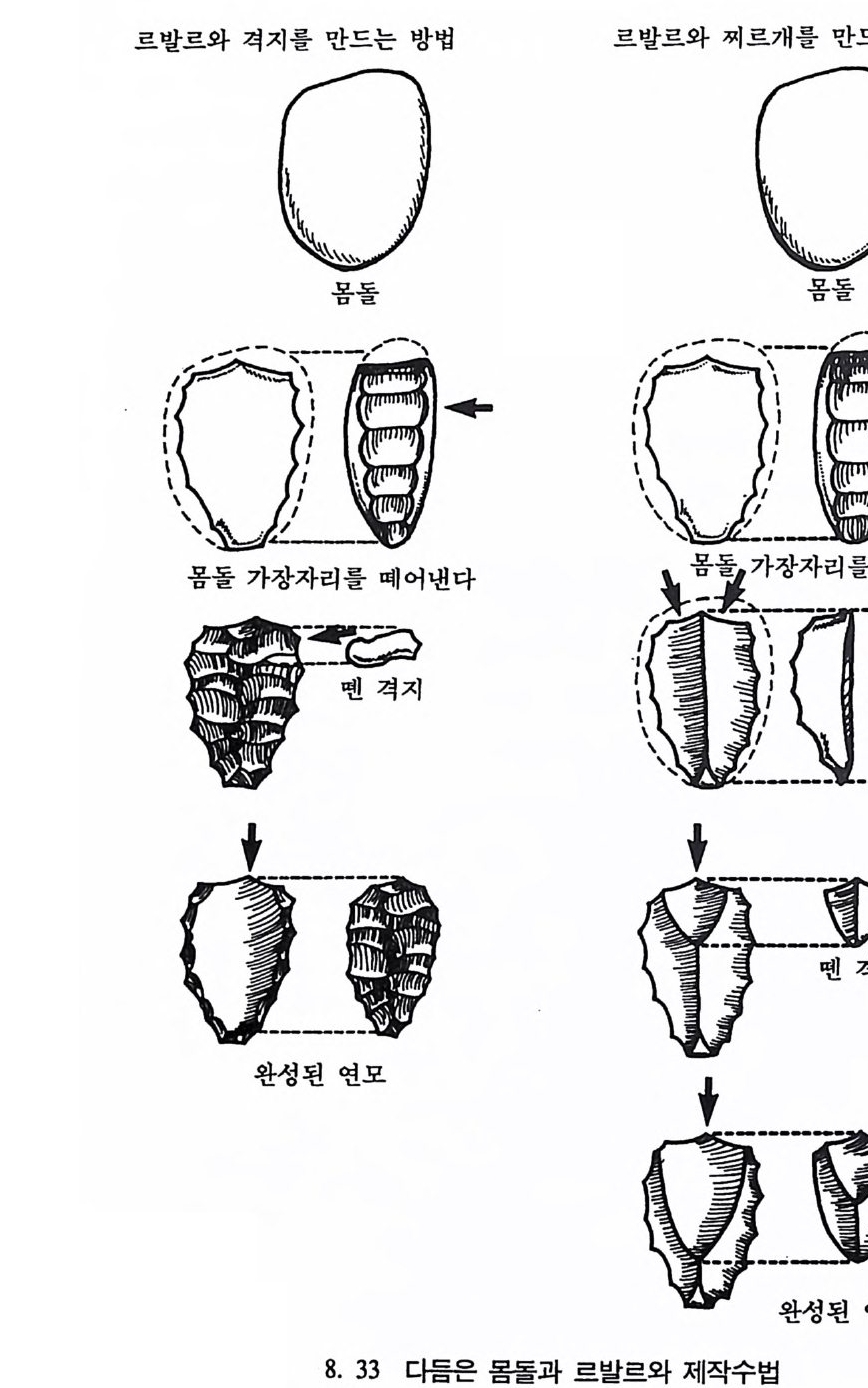 르발르와 격지를 만드는 방법 르발르와 찌르개 를 만드는 방법
르발르와 격지를 만드는 방법 르발르와 찌르개 를 만드는 방법
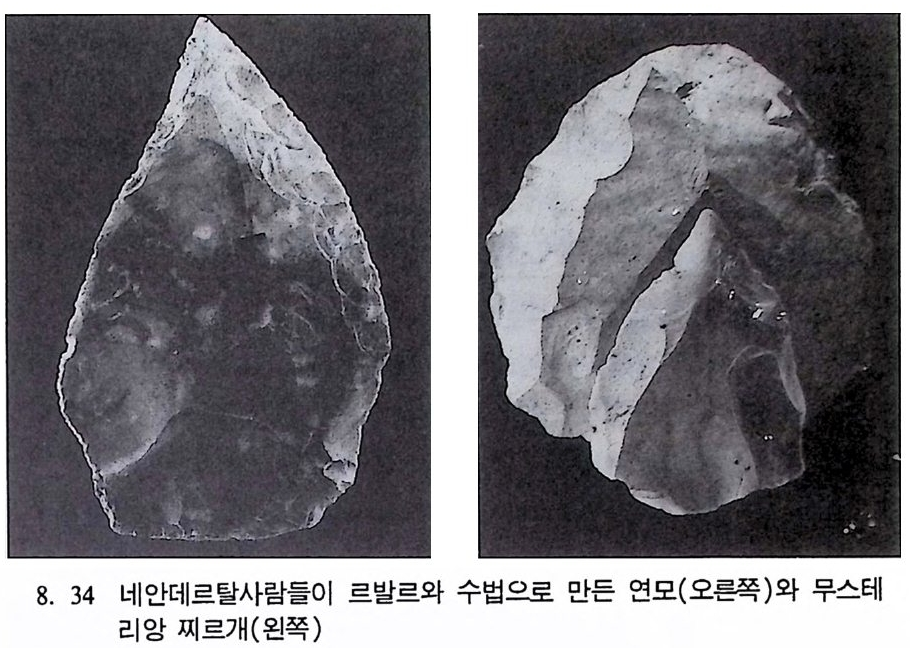 8. 34 네안데르탈사람들이 르발르와 수법으로 만든 연모(오른쪽)와 무스테
8. 34 네안데르탈사람들이 르발르와 수법으로 만든 연모(오른쪽)와 무스테
네안데르탈사람은 추운 빙하기 동안에 척박한 환경에서 살았고, 또 살아 남기 위해서도 그들은 아마도 영리했을 것이다. 네안데르탈사람 은 돌을 재료로 찰 사용하였고 사냥을 했던 것으로 알려져 있다. 그 둘은 무스테리앙이라는 돌연모 문화의 담당자들이었다. 이 무스데리 앙 연모 제작기술의 특칭은 날카로운 격지롤 만들기 위해서 몸돌을 미리 다듬는 것이다. 거북동날떼기 또는 르발루와 수법으로 만들어전 연모들은 사냥 등의 행위에 사용되었다(그림 8. 33, 사진 8. 34). 그러 나 네안데르탈사람들이 능동적으로 사냥을 했었는지 아니면 단지 기 회가 주어져서 사냥을 했었는지는 알 수 없다. 주로 네안데르탈사람에 의해 만들어전 무스테리앙 연모들은 프랑스 의 페리고르드- 일대에서 풍부히 발견되었다. 오늘날 이 일대는 풍부 한 돌연모와 자세한 지층의 연구를 통해 이 시기 문화연구의 중십지 로 알려져 있다. 이렇게 풍부한 자료를 가지고 보르드 F. Bordes 는 구석기시대의 연모를 63 개의 돌연모로 분류하였다. 나아가 돌연모와
그 형태들을 연구한 끝에 보르드는 이 지역의 동굴 및 한데 유적, 바 위그늘 유적지에서 반복해서 나타나는 돌연모들을 긁개의 출현 정도 에 따라 크게 다섯 유형으로 나누었다(표 8. 4).
표 8. 4 굵개를 기준으로 분류한 무스테리앙 돌연모의 분류 l. 대표되는 무스테리앙 연모 Ty pica l Mouste r ia n 2. 라 퀴나/라 펠라제 무스데리앙 연모 Charenti an La Quina ~ La Ferrassie Mouste r ia n 3. 톱니날 무스테 리 앙 연모 Denti cu late Mouste r ia n 4. 아슐리 앙 전통의 무스테 리 앙 연모 Mouste r ia n of Acheulia n Tradit ion A 전통 Type A B 전통 Type B
무스테리 앙 연모를 다섯 개의 유형으로 나눈 보르드의 분류는 구석 기 유적지에서 유물들이 왜 다르게 나타나는가에 대한 해석에 논쟁을 불러일으켰다. 논쟁은 주로 보르드와 빈포드 사이에 일어났다. 보르드 부부는 무스테리앙 연모에서 보이는 차이는 문화전통의 차이에서 기 인하는 것으로, 이는 아마도 〈서로 다른 집단 E thni c Grou p 〉 이 이 일 대에서 각기 다른 돌연모 문화를 지녔기 때문이라고 설명하며, 이런 전통들은 오랜 기간 동안 서로 섞이지 않고 지속되었다고 주장하였다 . 이에 반해 빈포드는 다섯 개로 구분되는 연모 유형은 서로 다론 기 능에 의한 결과라고 주장한다. 그는 당시에 살았던 수렵-채집인들은 주거지와 일하는 장소를 따로 가지고 있었는데, 각각의 장소에서는 각기 디른 행위가 있었기 때문에 이에 따라 다른 연모들이 출토된다 고주장한다.
네안데르탈사람의 행위 네안데르탈사람은 언어와 종교 그리고 내세에 관한 개념 등을 지니 고 있었는지 또한 그들은 식인풍습 행위를 가졌었는지 등에 관한 연 구는 그들 자신의 화석과 유적지 그리고 남겨진 유물을 바탕으로 이 루어졌다(사진 8. 35). 오늘날 세상의 여러 지역에서는 사람들의 죽음 울 서로 다르게 처리하고 있다. 우리들은 시체를 땅에 묻으나, 어떤 사람들은 화장을 하며 또 다른 사람들은 납골당으로 가져간다. 티벳 에서는 시체를 잘라서 새 등에게 먹이는데, 이는 죽은 자의 영혼을 자유롭게 하기 위해서라고 해석된다.
 8. 35 네안데르탈사람의 생활 복원도. 남부지역의 여름 캠프에서 네안데르
8. 35 네안데르탈사람의 생활 복원도. 남부지역의 여름 캠프에서 네안데르
네안데르탈사람들은 의도를 가지고 죽은 자를· 묻었다 . 네안더 골짝 과 스피 유적지롤 위시해서 태식 타쉬와 샤니다르 및 라 샤펠 오 솅 과 라 펠라제에서 발견된 화석들은 네안데르탈사람들이 죽은 후에 동 굴에 묻혔음을 보여준다. 특히 약 60000 년 전의 유적지로 알려진 라 펠라제의 바위그늘 유적에서는 가족의 묘로 보여지는 매장지가 발견 되었는데, 부모들은 머리를 맞대고 묻혔으며 근처에 한 개체의 갓 태 어난 아이와 27n 체의 5 세 된 아이, 그리고 한 개체의 @1l 된 아이의 뼈가함께 묻혀 있었다. 한편 옛 유고슬로바키아의 크라피나 유적지에서는 약 20 - 30 개체의 뼈가 부서지고 흩어전 채로 발견되었다. 이 뻣조각들에는 연모를 사 용해서 사지를 잘라내는 과정에서 생긴 자른 자국들이 뼈 표면에 나 타나고 있다. 한편 주먹도끼를 쓴 흔적도 나타나는데 이는 뼈롤 깨서 속을 열려고 했던 것으로 추정된다. 왜냐하면 뼈들은 좋은 영양 공급 원이며 이 동굴에서 살던 네안데르탈사람들은 식인자들이었다. 그러 나 크라피나 유적지에서 보이는 식인풍속이 신앙 이유인지 아니면 영 양분을 얻기 위해서인지는 아직 알 수 없다. 한때 식인풍속은 네안데르탈사람들의 일반적인 행위로 알려졌었다. 이는 네안데르탈사람의 유적지에서 이들의 머리뼈가 부서전 채로 발 견되었기 때문으로, 초기에 이들을 연구한 학자들은 네안데르탈사람 은 서로 잡아먹었으며 이를 식인풍속이라고 해석하였다. 그러나 최근 의 연구는 많은 네안데르탈사람의 부서진 머리뼈는-특히 얼굴부분 이 없어지고 머리통만 남은 것—―당시에 살고 있던 크로쿠타를 포함 한 많은 식육류들의 행위에 의해서라는 주장이 대두되고 있다.
 제 9 장 인류의 전화 III :
제 9 장 인류의 전화 III :
제 9 장 인류의 전화 m: 슬기 사람 Homo sap ien s : 현생인류의 시작 최초의 슬기사람 : 크로마뇽사람의 발견 지금부터 40000 년 전 유럽에서는 바이첼 빙하기가 발달해 있었다. 이 빙하기중 따뜻한 시기에 인류의 전화과정에 중요한 변화가 일어났 다. 오늘날의 우리를 꼭 닮은, 슬기사람으로 알려진 현생인류가 나타 나 인류의 문화발달 과정에 주목할만한 변화를 가져왔기 때문이다. 돌연모와 뼈연모의 발달 및 예술의 시작이 바로 그 변화이다. 오늘날 슬기사람에 관한 연구는 지구상의 여러 지역에서 진행되고 있다(그림 9. 1). 그러나 최초의 현생인류에 관한 증거는 1868 년 프랑 스의 도르도뉴 지방의 레 제지 마을 근처 유적에서 발견되었다. 바로 네안데르탈사람의 화석이 발견된 지 12 년 후의 일이었다. 도르도뉴 지역은 중생대 말에 형성된 석회암이 발달한 지대로 강과 물에 의해 생긴 동굴과 바위그늘이 발달해 있으며, 사냥감이 풍부하여 후기 구 석기시대에 인류가 살기에 좋은 환경을 제공하였다. 마을 근처에서 철로작업을 하던 일단의 인부들이 크로마농이라고 부르는 석회암 바위그늘에서 적어도 5 개체에 달하는 사람의 뼈대를 찾아냈다. 이들은 중년 남자 한 명과 한 명 내지 두 명의 젊은 남자,
한 명의 젊은 여자, 그리고 한 명의 어린이와 2 주 내지 3 주 정도 된 아기의 뼈둘로 이루어져 있었다. 이 뼈대들은 현대인과 닮았으며, 프 린트로 만든 돌연모와 구멍이 뚫린 바다조개 및 장식으로 쓰인 동물 이빨들이 함께 발견되었다 .
 접
접
이 호미니드 화석들은 발견된 유적지의 이름을 따서 크로마뇽사람 으로 부르게 되었다. 고고학과 고인류학에서 사용하는 크로마뇽이란 이름은 엄밀히 말하면 35000 년부터 10000 년 사이에 남서프랑스 지역 에서 후기 구석기문화를 담당했던 선사인류를 의미하나, 일반적으로 는 40000 년을 전후한 시기에 지구상에 나타난 늦은시기의 술기사람을 총칭하는 의미로 통용된다(사전 9. 2).
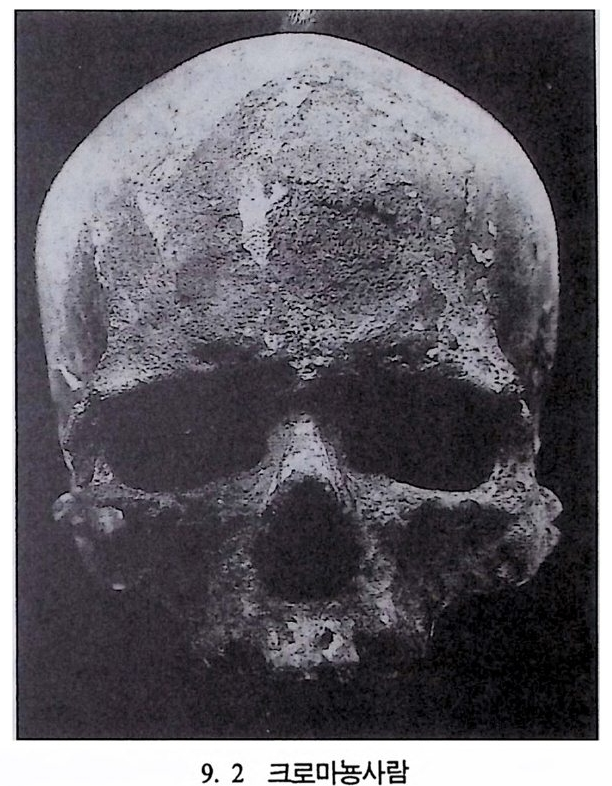 9. 2 크로 □ 悟사람
9. 2 크로 □ 悟사람
프랑스에서 발견된 화석을 시작으로 최근까지 크로마농사람을 닮은 화석들이 러시아를 포함한 유럽과 중동아시아, 북아프리카, 남아프리 카, 중국과 동남아시아, 그리고 오스트레일리아와 북남미 대륙 등 세 계 전 지역에서 발견되었다. 크로마농사람은 곧선사람과 네안데르탈
표 9. l 네안데르탈사람과 술기사람과의 해부학상 특징 비교 네안데르탈사람 l. 머리뼈의 평균 용량은 1600cc 정도이다. 2. 뒷머리뼈는 둥글지만 머리뼈 끝이 튀어나와 있다(이 부분을 〈 Bun 〉이라고 부른다)• 3. 눈두덩은 두럽게 발달해 있는데, 눈썹 사이에서 끊어지며, 눈 가장자리에서 점차 약해진다. 4. 뒷머리뼈-젖꼭지뼈 마루가 찰 발달해 있다. 5. 머리뼈 최대너비는 곧선사람보다는 위쪽에 있다. 6. 아래 셋째어금니와 아래턱 턱가지 사이의 빈 공간 Re t romolar Sulcus 이 있다. 7. 뒤에서 본 머리뼈는 달걀형이다. 8. 머리뼈의 두께는 곧선사람보다 얇다. 9. 골반뼈는 현생인류보다 길다. 10. 사지뼈는 짧고 굳세 보인다. 슬기사람 I. 머리뼈의 평균 용량은 1400cc 정도이다. 2. 머리뼈에는 근육이 닿는 부위가 약하게 보인다. 3. 곧은 앞머리뼈를 가지고 있다. 4. 눈두덩은 미약하며, 눈썹 사이는 편편하다. 5. 뒷머리뼈는 둥글며, 근육이 닿는 면이 약하게 발달하고 있다. 6. 이복파임(二復寫)과 뒷머리뼈홈이 뚜렷이 보이는 피라미드 모 양의 젖꼭지돌기가 있다. 7. 머리뼈 최대너비는 옆머리뼈에서 얻어전다. & 아래덕과 이빨둘은 작으며, 셋째어금니와 턱가지 사이에 빈 공 간이 거의 없다. 9. 아래턱에 턱이 뚜렷하게 발달해 있다. 10. 뒤에서 본 머리뼈는 오각형이다.
사람과는 해부학상으로 다른 특징을 지녔다(표 9. I). 크로마뇽사람들은 파여 들어간 턱과 높은 앞이마 그리고 작은 이빨들을 지니고 있으며, 머 리뼈의 크기는 현대인의 것과 차이가 없다. 해부학상의 전체 특징으로 볼 때, 이들은 오늘날 살고 있는 인류와 똑같은 특징을 보이고 있다. 크로마뇽사람에서 무엇보다 중요한 점은 이들에게서 처음으로 우리 들이 사용하는 것과 같은 유형의 언어와 자의식의 표현인 미적 감각 능력이 나타났다는 점이다. 바로 오늘날의 우리와 같이 생각하고 행 동하는 인류가 최초로 출현하여 후기 구석기문화를 꽃피운 것이다. 슬기사람 기원과 발달에 관한 최근의 연구들 슬기사람인 현생인류의 연구는 오랫동안 유럽 중심의 사고에 의해 지배되어 유럽이 술기사람의 기원지로 인식되었다. 여기에는 몇 가지 이유가 있는데, 첫째, 선사인류 화석의 연구는 1800 년대에 프랑스와 독일 등의 유럽 국가에서 처음 시작되었고, 둘째, 인류는 체질상 일찍 이 현대인이 되었음에도 불구하고 인류의 전파과정중 늦게 아메리카 와 오스트레일리아에 도달했으며, 끝으로 선사인류의 화석은 유럽국 가들에서 보존상태가 아주 좋은 채로 발견되었고 또한 집중해서 연구 가 진행되었으나 아프리카나 아시아에서는 화석들이 별로 나타나지 않았기 때문이다. 1970 년대와 1980 년대에 걸쳐 동아프리카 케냐의 쿠비 포라에서 발견 된 손쓴사람과 곧선사람의 화석들, 그리고 탄자니아의 라에톨리에서 발견된 370 만 년 전의 호미니드의 발자국과 에티오피아의 하다에서 발견된 〈루시〉를 포함하여 오스트랄로피데쿠스 아파렌시스 등의 발견 은 인류 기원과 전화에 관한 그간의 고인류학상 연구에 많은 논란울 불러일으켰다.
현재 아프리카 사 람 유럽사람 동아시아사람 오스트레일리아사람 100.0 00 300.OOO 살단하 페트라로나 저우커우뎬 삼봉마찬 700,000 올드바이 유럽 란덴 자 II~ (년) 곧선사람 현재 아프리카사람 유럽사람 동아시아사람 오스트레일리아사람 30 0.00 0 TI\ 달리 I ' ? 100,000 I [ ?+ 700,0 00 (년) 곧선사람 9. 3 슬기사람의 다지역 기원설과 단일지역 기원설. 위 그림은 다지역기원 설로, 지역적으로 나타나는 호미니드의 해부학상 특성은 아프리카, 유 럽, 동아시아, 그리고 오스트레일리아에서 살던 옛 사람들로부터 각기 그 특성을 물려받았기 때문이며, 진화과정중 같은 시기에 살았던 집단 사이에 유전자 교환이 있었다고 주장한다. 아래 그림은 단일지역기원설로 지역적인 특성은 아프리카에서 기원한 슬기사람들이 유전자 교환 없이, 옛 사람들을 대신하였다고 밝히고 있 다. 이때 오스트레일리아에서만은 어느 정도의 유전자 교환을 인정하 고있다.
같은 시기에 오늘날 지구상에 퍼져 살고 있는 슬기사람의 기원에 대한 중요한 연구가 진행되었다. 슬기사람이 지니고 있는 해부학상의 특칭인 눈두덩이 밋밋하고 머리뼈가 얇은 것은 현생인류가 어떤 한 지역에서 살던 조상으로부터 전화해 왔기 때문인지, 아니면 지구상에 퍼져 살고 있는 모든 슬기사람들이 진화해 오는 과정중에 지니게 된 해부학상의 특칭인지가 논쟁의 초점이 되어왔다. 그간에 발견된 새로 운 증거들에 따라서 양쪽의 가설들이 서로 엇갈리며 부각되었고, 또 한 중간형의 가설들이 제기되었다. 그러나 최근에 고인류학자들과 유전학자들의 연구에 의해 슬기사람 의 기원에 관한 새로운 해석이 가능·하게 되었다. 특히 현재 전행되고 있는 유전학상의 연구는 모든 슬기사람이 최근에 아프리카에서 살던 한 주민에서 시작되었음을 밝히고 있으며, 고고학상 • 고인류학상의 증거 또한 이를 뒷받침해 주고 있다. 슬기사람 기원에 관한 연구의 초점은 이들의 지리적인 차이에 따른 전화의 유형에 집중되었으며, 이에 대한 연구는 크게 다지역 기원설 과 단일지역 기원설로 나누어진다(그림 9. 3). 하웰스 W. W. Howells 는 다지역 기원설을 촛대형 모형설로, 그리고 단일지역 기원설을 노 아의 방주 모형설로 부르고 있다(그림 9. 4). 다지역 기원설 이 설에 따르면 현재 인류가 지니고 있는 인종의 특 징은 이미 그들이 오늘날 발견되는 지역에서 오랜 세월 동안 진화해 온 결과라고 주장한다. 슬기사람은 여러 지역에서 살던 곧선사람으로 부터 전화하여, 유전자 교환이나, 유사전화의 과정을 거쳤기에 슬기사 람은 아주 깊은 뿌리를 가지고 있다고 한다. 이 설을 지지하는 학자 들의 주장에 따르면 특히 유럽 사람들과 아프리카 사람들은 20000 년 전부터 그들만이 지닌 해부학상의 특칭을 지녀 내려왔다고 한다. 이 다지역 기원설은 여러 해 동안 미시건 대학의 월포프 M.H.
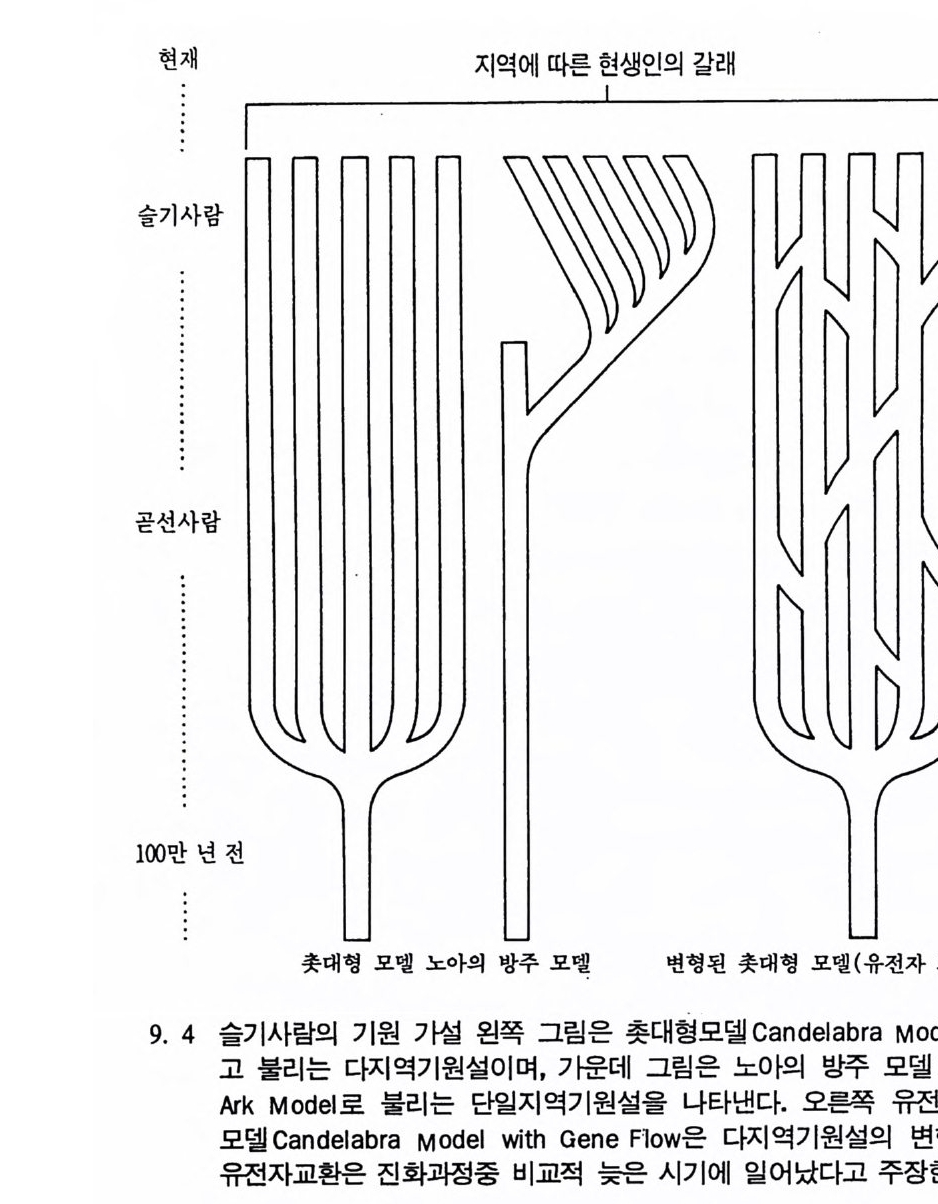 현재 지역에 따른 현생인의 갈래
현재 지역에 따른 현생인의 갈래
Wol p o ff를 위시한 고인류학자들에 의해 주창되어오고 있다. 만일 이 둘의 가설이 옳다면 인종을 분류하는 데 이용되는 체질상의 여러 특 칭들은 오래 전에 형성되었음에 틀림없다. 예를 들어 유럽 사람들의
커다란 코라든지, 오스트레일리아 원주민들에게서 나타나는 굳세 보 이는 광대뼈둘은 20 만 년 전부터 3 만 년 전에 유럽에서 살았던 네안 데르탈사람과 약 중기 홍적세에 인도네시아에서 살았던 곧선사람 중 의 하나인 자바사람들로부터 이어받았다고 할 수 있다. 다지역 기원설의 변형인 다지역 기원 유전자교환 모델은 옛 사람 계동이 지역에 따라 다르긴 하지만, 슬기사람으로 진화해 오는 과정 에서 유전자 교환 비율이 점차적으로 증가하였음을- 설명하여 준다. 이 설은 뉴멕시코 대학의 트링카우스 Er i ck Tr i nkaus 와 북일노이 대 학교의 스미스 F.H.S mith 등에 의해 지지를 받고 있다. 단일지역 기원설 이 설에 따르면 현생인류의 뿌리는 매우 짧으며, 이들은 지구상의 한 지역-아프리카-에서 옛슬기사람 (Arch ai c Homo sap i ens 로 곧선사람 Sta g e III 울 뜻함)으로부터 진화한, 인종상 별다른 차이가 없는 일단의 슬기사람의 조상이 전 세계로 퍼져 나갔 으며, 먼저 살던 옛슬기사람들을 몰아내고 그 자리를 대신하였다고 주장한다. 그 과정중에 슬기사람은 새로운 환경에 적응해 가며 차차 인종적인 특칭이 발달했다고 설명하고 있다. 단일지역 기원설은 다지역 기원설보다논리적으로 슬기사람은옛슬 기사람으로부터 단지 한 번 진화한 후, 적어도 어느 지역(예 :유럽)에 서는 빠른 속도로 슬기사람의 모습으로 나타났다고 주장한다. 이 설 은 또한 모든 슬기사람은 최근의 공통조상(약 10 만 년 전 내외)으로 부터 시작됐다고 생각되는 멘델의 유전적인 특칭을 가지고 있다는 자 료에 지지를 받고 있다. 최근에 단일지역 기원설은 미국 주립 캘리포니아 버클리 대학교의 윌슨A. W il son 과 영국 자연사박물관의 스트링거 C.S t r i n g er 등이 이 끄는 일단의 유전학자들에 의해 확고한 지지를 받고 있는데, 이들의 연구에 의하면 유럽에서 살던 네안데르탈사람과 인도네시아에서 살던
솔로사람(이들은 아마도 자바사람들의 지역상 후예로 인정된다) 같은 옛슬기사람은 이론시기의 슬기사람과는- 유전자 교환이 없었으며, 이 론시기의 슬기사람이 옛슬기사람들을 몰아내고 그 자리를 대신하였다 고주장한다. 단일지역 기원설의 변형으로 슬기사람이 전화해 가는- 과정중 특히 아시아의 경우, 어느 정도 그 지역에서 살던 선주민들과 유전자 교환 이 있어 지역상 • 해부학상 특징이 슬기사람을 통해 전해졌으리라는 주장인 교배 및 교체 설 Hy b rid iza ti on and Rep la cement Model 이 제 기되고 있다. 위에서 슬기사람의 기원과 전화에 관한 대표되는 2 개의 가설들과 그 변형된 가설들을 간단히 살펴보았다. 슬기사람 기원에 관한 가설 둘은 오늘날 슬기사람이 지닌 체질상 특칭이 늦게 전화되었으며, 인 종상 특칭들은 지역에 따라 전화하였다는 데는 의견의 일치를 보이고 있으나, 인종의 차이들이 언제 시작되었는지에 대해서는 근본적으로 차이가 있다. 다지역 기원설은 인종의 특칭은 슬기사람이 나타나기 이전에 형성되었다고 주장하며, 아프리카 단일지역 기원 모형설은 그 반대의 입장을 견지하고 있다. 오늘날 우리는 이들 두 가설을 평가할 수 있는 충분한 증거를 가지 고 있다. 그 중 하나가 미토콘드리아 핵산 연구로, 이에 따르면 모든 인간은 최근의 한 공통조상으로부터 갈라져 나왔다는 것이다. 또 다 론 증거는 일련의 호미니드 화석들의 발견으로, 하나는 남아프리카의 클라시에스 강 어구에서 나온 화석들이고 다론 하나는 이스라엘의 카 프체 유적에서 발견된 화석이다. 이 두 유적지에서 발굴된 화석들은 모두 〈해부학상 슬기사람 Ana t o mica lly Modern Man 또는 Homo sap i e 짜인데, 이들은 최소한 5 만 년이 넘는 연대를 갖고 있으며, 가장 오래 된 화석은 10 만 년이 넘는
연대를 갖고 있는 것으로 알려지고 있다. 따라서 이스라엘에서 나온 화석중거에 따르면 네안데르탈사람이 사라지기 이전에 이미 현생인류 인 슬기사람이 존재했다는 것으로, 이는 네안데르탈사람이 슬기사람 의 출현에 어떤 역할도 하지 못했다는 확실한 증거 중의 하나이다. 미토콘드리아 핵산 (m t DNA) : 유전학자들의 연구 유전학자들은 처음에 눈의 색깔이나 단백질의 구조 등과 관련된 유 전자 연구를 시직하였으나, 지난 20 여 년간의 연구가 발전하면서 핵 산을 구성하는 뉴클레오티드 자체를 분석할 수 있는 단계까지 발전하 였다. 유전학 연구대상의 하나인 인류와 유인원이 언제 갈라졌는가에 관 한 대답에는 수백만 년이라는 연대가 필요하나, 슬기사람 기원에 대 한 논쟁은 100 만 년보다는 작은 범위 내의 연대가 필요할 뿐이다. 이 처럼 시간의 범위가 좁기 때문에 미국 주립 캘리포니아 버클리 대학 교의 윌슨 교수와 그의 동료인 레베카 칸 Rebecca Kahn 등은 인류와 유인원이 갈라전 시기를 추정하는데 사용되는 분자시계 대신, 극히 짧은 시간을 알려주는 보다 정밀한 분자시계를 찾고자 하였다. 그들 은 이 시계를 세포 안에 있는 미토콘드리아에서 찾았는데, 이 미토콘 드리아는 동물의 각 세포조직에서 글리코즈를 에너지로 전환시키는 기능과 유전물질인 핵산울 가지고 있으며, 이 핵산은 세포핵에 있는 핵산보다 열 배나 빠른 속도로 돌연변이를 일으킨다. 따라서 핵산의 변이는 극히 짧은 시간에 일어나며 그 시간을 측정할 수 있다. 미토콘드리아가 지니고 있는 또 다른 기능은 무엇인가? 여성의 난 자가 남성의 정자와 결합하여 태아를 형성할 때, 정자가 난자 속으로 자신의 염색체를 주입하지만, 자궁 안에서 정자가 활발히 움직이도록 동력을 공급하는 미토콘드리아는 뒤에 남는다. 아이가 만들어질 때는
언제나 어머니의 미토콘드리아만이 아이에게 전달된다. 따라서 우리 가 지니고 있는 미토콘드리아의 핵산은 오직 여성 쪽에서 전해 받은 유전물질로 우리들의 어머니, 할머니, 증조모 등을 거쳐 우리들 자신 의 역사 중 가장 먼 시기까지 거슬러 올라갈 수 있다. 따라서 미토콘 드리아 유전학을 이용함으로써 지구상에서 누가 누구와 서로 관계가 있는지를 규명하는 일이 가능하다. 이것을 바탕으로 윌슨 등은 세계를 지리 구분에 따라 아프리카, 아 시아, 유럽, 오스트레일리아 및 뉴기니아 등으로 나누어 이 지역들에 서 온 150 명의 여성들로부터 미토콘드리아 핵산을 찾을- 수 있었다. 그 결과는 1987 년에 발표되었다. 그러나 미토콘드리아 핵산에서 일어나는 돌연변이룰 이용해서 만 든, 가계혈통이 갈라져 나가는 시간을 측정할 수 있는 분자시계는 아 래와 갇은 가정이 사실일 때에만 믿을 만하다. 죽 생물체의 유전 동 일성이 서로 연관을 갖고, 동시에 공통조상으로부터 분화되는 시간의 비례와 직접적으로 관련되어 기능을 발휘해야 한다. 이때에 연구대상 의 유전물질들은 일정한 비율로 돌연변이가 일어나며, 돌연변이가 어 떤 자연선택을 선호하지 않는다는 것을 전제조건으로 한다. 분자시계가 유용하기 위한 또 다른 전제조건으로는 생물체가 분화 된 후, 전화과정중에 많은 정도의 유전자 교환이 있어서는 안 된다는 점이다. 그렇지 않으면 그들이 실제 분화된 시기보다 더 늦은 시기에 갈라져 나간 것처럼 보이기 때문이다. 다행히도 미토콘드리아 핵산의 일부가 바로 이런 전제조건에 알맞다고 할 수 있다. 위에서 언급된 전제조건을 바탕으로 여러 종류의 동물 사이에서의 유전 차이와 또 동물 안에서의 다양성을 비교하여 그들의 선후관계를 밝히려는 노력이 진행되고 있다. 이 중 갈라져 나간 연대를 알고 있 는 집단들을 비교함으로써 이들의 선후 연대와 분화가 발생한 시기를 추정한다. 물론 이렇게 얻어진 결과가 실제 인류의 가계를 밝힌다기
보다는 문제가 되는 집단들의 유사성과 차이를 바탕으로 분류하는 데 도움이 된다. 많은 유전학 연구에 따르면 슬기사람은 사하라 이남지역과 다른 지 역의 모든 인류집단으로 구성된 두 개의 기본단위로 이루어져 있음을 알 수 있다(그립 9. 5). 그러나 이 유전 가계로부터 전화상의 연대는 문제가 있을 수 있다는 접을 유의해야 한다. 어떤 학자들의 연구에 따르면 살아 있는 슬기사람 중에서 나타나는 유전 차이는 10 만 년 전 부터 20 만 년 전 사이에 이루어졌던 집단간의 유전자 이동 때문에 그 들에게 보이는 차이가 실제보다 더 오래 된 것처럼 보인다고 주장한 다. 물론 인류진화 과정중에 언제 술기사람이 시작되었는지를 정확하게 말한다는 것은 불가능한 일이다. 예를 들어 미토콘드리아 핵산에 관 한 이른 연구에 따르면 아시아나 아프리카가 슬기사람의 기원지로서 타당하다고 설명한다. 만일 누가 심정상 아시아 대륙을· 선호할 때 만 일 그 대륙에서 사는 집단에서 나타나는 미토콘드리아 핵산의 형태가 모든 집단에서 평균에 가장 가까운 형태로 나타나면 그때는 아시아 대륙을 인류의 고향으로 인정할 것이다. 그런 경우에 부시먼 등에서 보이는 커다란 유전 차이는 지역적으로 큰 비율로 진행된 돌연변이가 원인이었다고 해석할 것이다. 한편 모든 인류집단 내에서 미토콘드리 아 핵산이 일정한 비율로서 돌연변이를 일으켰다면 그때는 아프리카 가 인류의 기원지가 될 것이다. 이런 결론은 침팬지가 오래 전에 인 류의 뿌리에서 벗어난 집단이었음을 증명하는 데 사용되던 미토콘드 리아에 관한 연구가 더 진행되면 확실히 알 수 있다. 인류의 공통조상 이후 나타나는 호미니드들에서 보이는 변이는 바 로 비율의 차이를 의미하는 것으로, 죽 낮고 불규칙한 비율은 조상들 이 곧선사람 시기인 50 만 년 이전에 있었음을 의미하며, 반면에 높고 일정한 비율은 공통조상이 슬기사람이 이미 존재했던 시기인 15 만 년
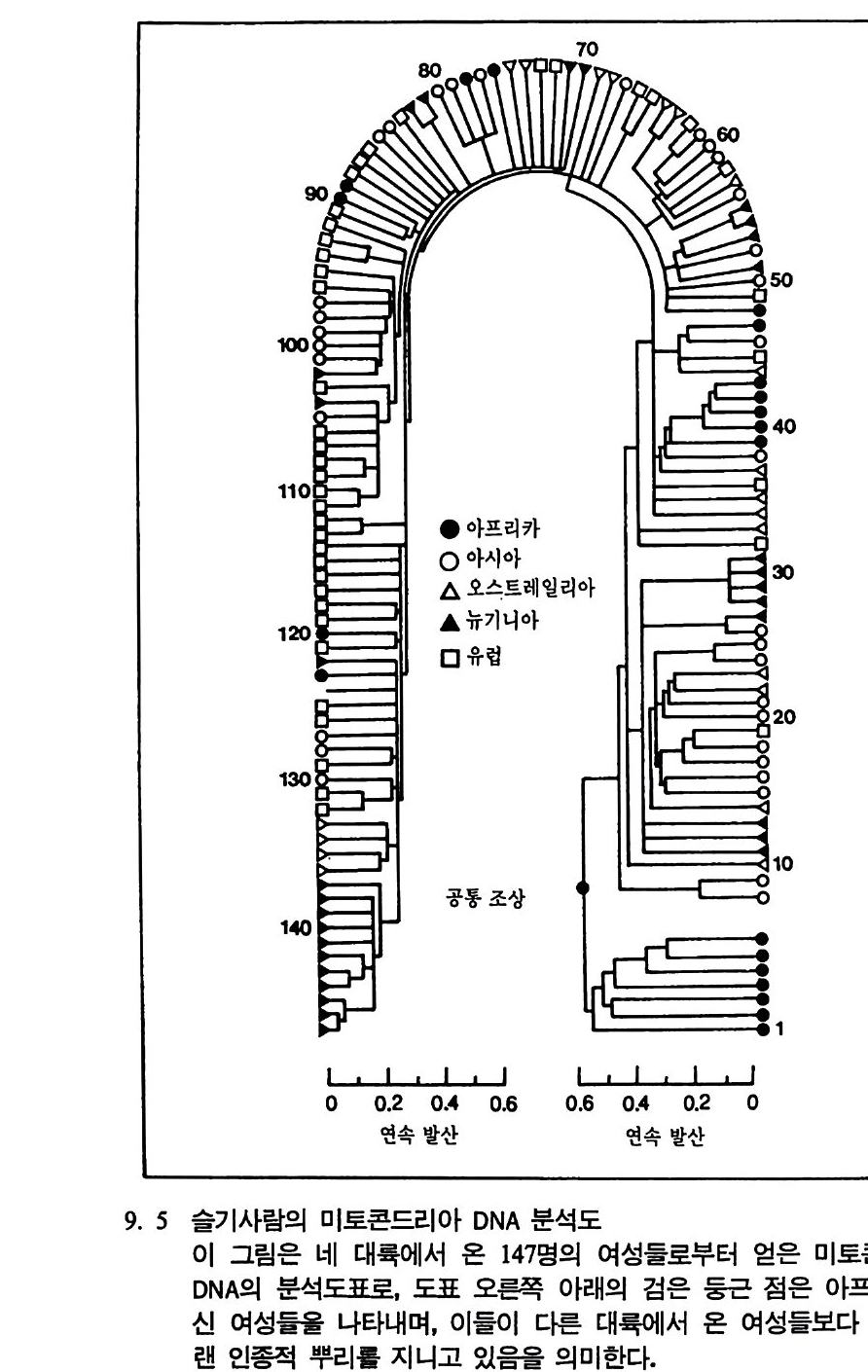 40
40
내에 있었음을뜻한다. 만일 슬기사람이 곧선사람이 살던 시기에 갈라지기 시작했다면, 이 들의 갈래 중 일부는 슬기사람들이 지니고 있는 해부학상 특칭들을 독자로 발전시켰음에 틀림없다. 그러나 미토콘드리아 핵산 연구는 술 기사람이 지니고 있는 특칭들은 이미 그 특칭이 형성된 집단으로부터 갈라져 나간 것을 보여주고 있다. 나아가 미토콘드리아 핵산 연구에 바탕을 둔 유전학 연구는 현생 유럽 사람과 아시아 사람 그리고 오스트레일리아 사람들이 사하라 이 남의 아프리카 사람들에 비해 유전상으로 서로 아주 가깝다는 사실을 밝혀주고 있다. 이러한 사실은 30 만 년 전부터 유럽과 아프리카 집단 의 서로 가까운 관계를 나타내고, 한편으로는 중국과 인도네시아 집 단이 가깝다는 사실을 보여주는 인류 화석자료들과는 일치하지 않는 다. 더욱이 10 만 년 전까지 그 차이는 더욱 증가하여 각 지역에서 호 미니드 집단들은 아주 다른 모습을 갖추게 되었다. 죽 유럽과 서아시 아에는 네안데르탈사람이 있었으며, 아프리카와 서아시아에는 현생인 류가, 그리고 중국과 자바에는 옛슬기사람으로 알려진 곧선사람 중 늦은시기의 형태 (H. erectu s S t a g e ffi)가 존재했었다. 그러나 오늘날 유럽과 아시아, 그리고 오스트레일리아에서 살고 있는 집단들은 유전 상 아주 가깝다는 사실과 2 만 년 내지 3 만 년 전에 이 지역에서 살았 던 인류 또한 가깝게 닮았다는 사실을 우리는 알고 있다. 고고학상 • 고인류학상의 새로운 증거들 아프리카 • 중동아시아 지역에서 나온 증거둘 위에서 언급한 사실을 토대로 단일지역 기원설이 성립되었는데, 이 설에 따르면 슬기사람은 아마도 늦어도 지난 10 만 년 전에 아프리카 로부터 시작된 이른시기 슬기사람의 전파에서 시작되었다. 이런 단일
지역 기원설은 10 년 전에 이미 제시되었다. 그러나 아프리카에서 시 작되었다는 결론은 최근까지 받아들여지지 않았는데 이는 방사성 탄 소 연대 측정으로는 3-4 만 년 이전의 연대를 얻을 수가 없었기 때문 이었다. 그러나 최근에 발달한 우라늄계열 원소, 가열발광, 그리고 전 자 회전반응 등의 연대 측정방법에 의해 연대상의 문제가 해결되었 다. 어느 경우에든 호미니드의 연대는 이와 관련된 자료들의 연대에 의 해서 결정된다. 비록 위에서 언급한 세 개의 측정방법이 각기 한계가 있기는 하지만 이 방법들의 결과는 슬기사람이, 네안데르탈사람들이 유럽과 서아시아에서 그리고 솔로사람들이 자바에서 살고 있을 당시 에, 중동지방과 아프리카에서 살고 있었음을 밝혀주고 있다. 이런 사 실들은 남아프리카의 클라시에스 리버 마우스 동굴과 보더 동굴에서 발견된 머리뼈와 턱, 그리고 사지뼈둘의 발견에 의해서 확인되고 있 다 이 유적지들에서는 호미니드 화석과 함께 13 만 년 전부터 35000 년 전 사이에 아프리카 중기 구석기시대의 돌연모들이 발굴되었다. 클라 시에스 유적지에서 실시된 우라늄 원소 측정방법과 전자 회전반응 측 정에 따르면 중기 구석기시대의 이른 시기충과 대부분의 호미니드 화 석의 연대는 90000 년이 넘는 것으로 밝혀졌다. 한편 보더 동굴의 경우, 처음 두 개체의 호미니드 화석이 체계있는 발굴을 통해 찾아지지 않아 문제가 있으나, 학자들은 3 개체의 화석은 75000 년의 연대를 지닌 충에서 발굴되었고, 또 다른 1 개체는 60000 년 의 연대를 가전 충에서 발견되었다고 주장한다. 남아프리카의 풀로리 스베드와 동아프리카의 아이시 호수가, 오모, 라에톨리, 성가, 그리고 칸제라 등에서 해부학상 현대인인 슬기사람들의 뼈화석을 찾을 수 있 었는데(그립 9. 6), 일부학자들은 이 뼈화석의 연대가 남아프리카 유 적지들의 연대보다 더 오래 되었을지도 모른다고 말하고 있다.
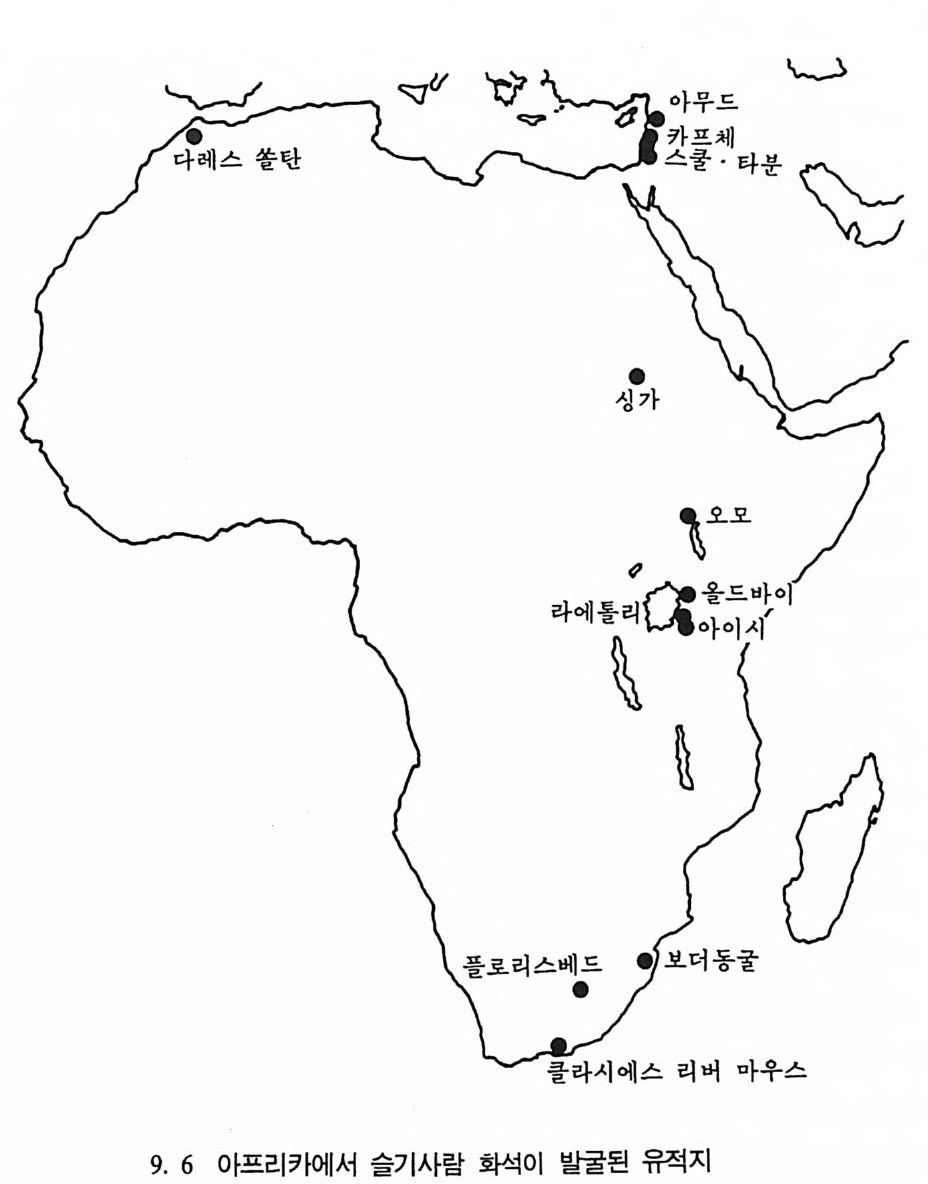 노
노
이론시기의 슬기사람은 늦은시기 유럽의 옛슬기사람(곧선사람 Stag e m) 으로부터 시작되지 않았다는 새로운 증거가 이스라엘의 동굴 유적 지들에 대한 재해석 끝에 처]시되었다. 이들 유적지에서 발견된 증거 둘은 지금까지 통용되던 인류진화에 관한 새로운 이해를 도와준다.
1930 년대 레반트 지역에 있는 카멜 산의 스쿨과 타분 동굴에서 인 류화석 뼈대의 일부가 발견되었는데, 이 뼈들은 당시에 살던 호미니 드의 뜻을 가전 행위의 결과였다고 해석되었다. 이후 두 동굴 유적에 서 살았던 주민들에 관해 많은 가설들이 제기되었다. 이 가설들 중 타분 유적지에서는 네안데르탈사람이 살았으며 스쿨 유적에서는 원시 적인 모습의 슬기사람이 살았다는 가설이 점차 받아들여졌다. 1935 년과 1975 년 사이에 나사렛 근처의 제불 카프체 유적에서 많 은 인류화석이 발굴되었으며, 갇은 시기에 카멜 산의 케바라 동굴과 갈릴리 호수 근처의 아무드 동굴에서 네안데르탈사람의 화석이 발굴 되었다. 따라서 유럽에서처럼 중동아시아에서도 현생인류는 네안데르 탈사람들에 이어 나타났으며 현생인류는 네안데르탈사람으로부터 시 작되었다고 가정하였다. 이런 견해는 198 71;빈과 1988 년에 무너지기 시작했으며, 가열발광 과 전자 회전반응 연대측정은 케바라 출토 네안데르탈사람의 연대가 60000 년임을 알려주며 동시에 카프체 유적은 이보다 40000 년이 더 이론시기로 밝혀지고 있으니, 이는 두 개의 다른 인류진화 갈래가 중 동아시아에 있었음을- 의미한다. 죽 한 갈래는 카프체 주민의 조상으 로 이들은 아프리카나 알려지지 않은 지역에서 시작하여, 이후 현생 인류가 되었다는 것과, 또 다른 갈래는 70000 년 전에 나타나기 시작 한 네안데르탈사람의 갈래로 이들은 아마도 빙하가 발달한 유럽에서 중동아시아로 이주하였으리라는 것이다. 이 네안데르탈사람들은 40000 년 전 스쿨 유적 주민이 다시 나타나기 시작하는 때까지 이 지 역에서 살았다고 여겨졌다. 그러나 최근 1988-1989 년에 이루어진 전자 회전반응 연대측정 결 과는 지금까지 스쿨 주민이 살았던 연대에 대해 의문을 제기한다. 죽 스쿨과 타분 유적지의 인류가 묻힌 충에서 발굴된 젖먹이동물의 이빨 의 연대를 측정한 결과, 이 두 유적지에서는 IO 만 년에서 12 만 년 전
사이에 인류가 살았음이 밝혀졌다. 이 새로운 연대는 현생인류가 일 찍이 이 지역에서 살았다는 것을 시사하고 있으며, 동시에 이론시기 의 네안데르탈사람 또한 여기서 살았다는 것을 의미한다. 그리고 타 분 유적지 주민들은 카프체와 스쿨 주민들보다 조금 앞서 살았든지 아니면 거의 같은 시대에 살았다고 해석된다(그립 9. 7).
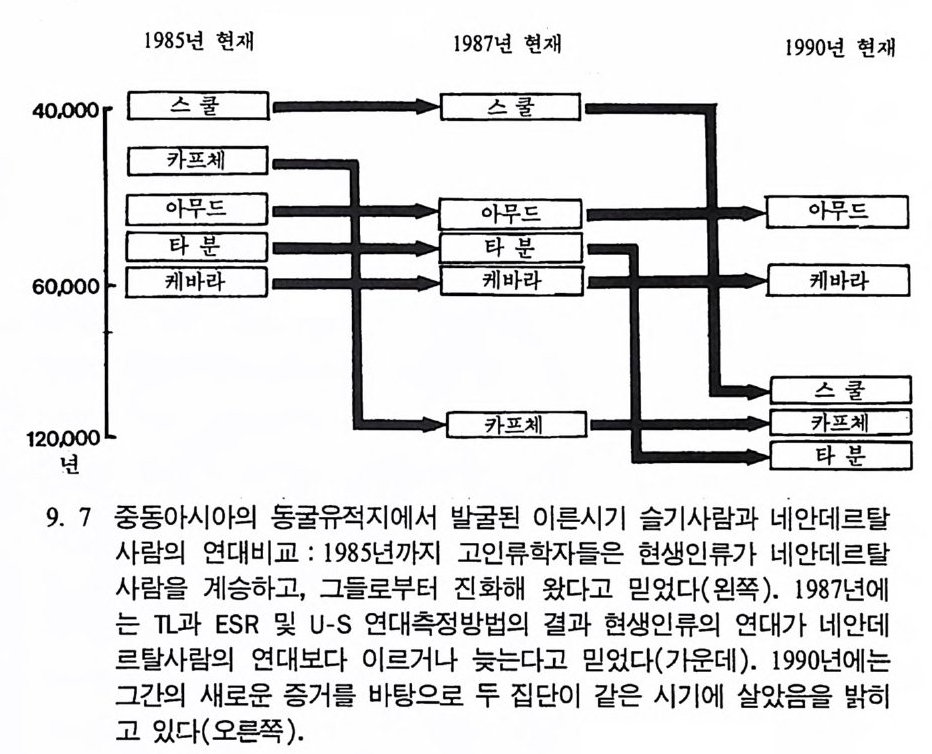 1985 년 현재 1987 년 현재 1990 년 현재
1985 년 현재 1987 년 현재 1990 년 현재
동아시아 • 오스트레일리아의 슬기사람 동아시아에서 발견된 인류화석들(그림 9. 8) 또한 유럽과 중동아시 아에서처럼 단일지역 기원설을 지지해 줄 수 있을까 하는 의문이 제 기된다. 동아시아의 슬기사람 기원 문제는 아래와 같은 2 개의 상반되 는 가설로 설명되고 있다. 첫째, 동아시아의 해부학상 현생인류는 유럽과 아프리카 대륙에서
 찌 / \一’_ \양,~〈;-三미;}皇;l ?-~;;n타; :} 타
찌 / \一’_ \양,~〈;-三미;}皇;l ?-~;;n타; :} 타
처럼 아시아에서 살던 곧선사람에서 독립하여 옛슬기사람을 거쳐 슬 기사람으로 전화하였다는 가설과, 둘째, 동아시아에서 살던 옛슬기사람은 아프리카에서 시작한 슬기 사람에 의해 대치되었다는 설로 크게 나누어진다.
중국의 저우커우덴(周口店) 유적지의 윗동굴(山頂洞)에서 발견된 화석과 슈지아요(許家察), 츠양(資陽), 류지앙(柳江) 등지에서 발견된 인류화석은 같은 시기에 살던 네안데르탈사람과 베이칭 곧선사람(北 京猿人)들같이 먼저 살던 호미니드와는 분명히 다른 계열임을 나타내 고 있다. 이 지역에서 약 20000 년을 전후한 시기에 살았던 인류화석 은 아프리카 기원설을 뒷받침해 주고 있으니, 이는 아시아에서 살던 호미니드들의 해부학상 모습들이 계속 발전하고 있을 때 이들과 다론 모습은 지닌 채 발견되기 때문이라고 주장한다. 만일 이 가설이 옳다 면 이들이 지닌 해부학상 모습은 베이칭 곧선사람으로부터 물려받았 다고할 수는 없다. 한편 이론 후기 홍적세에 동아시아에서 발견된 옛술기사람(마바, 달리)과 늦은 후기 홍적세의 슬기사람(슈지아요, 츠양, 류지앙, 산명 뚱)의 해부학상의 특칭을 비교한 결과 이론시기와 늦은시기의 화석을 연결해 주는 중간형은 없으나, 늦은시기의 화석에서 이른시기 인류 화석의 특칭이 보이며, 또한 유럽과 중동아시아에서 발견되는 술기사 람 화석들과도 닮은 점으로 보아 단일지역 기원설 중 교배 및 교체설 이 더 타당하다고 생각된다. 이 교배 및 교체설은 한국을 포함하여 동북아시아에서 발견된 늦은시기 슬기사람(예 : 용곡, 만달, 홍수아이, 미나토가와 등) 화석들이 왜 동북아시아 출토 곧선사람들의 해부학상 특칭을 지니고 있는지를 설명해 줄 수 있다. 슬기사람의 이빨 형태 비교에 따르면, 동남아시아 주민들의 이빨 형태는 인류의 평균 모습과 아주 닮았다고 한다. 그러나 미토콘드리 아 핵산울 비교해 볼 때 아시아 대륙은 인류의 고향으로 증명될 수가 없다. 아마도 인류가 아프리카에서 기원했다면 아시아와 유럽으로, 이 어 오스트레일리아 그리고 북미 대륙으로 퍼져 나갔다고 여겨진다. 왜냐하면 일찍이 북미 대륙에서 살았던 주민들의 화석은 의모에서 뚜 렷한 다양성을 보여주고 있기 때문이다(그림 9. 9, 9. 10).
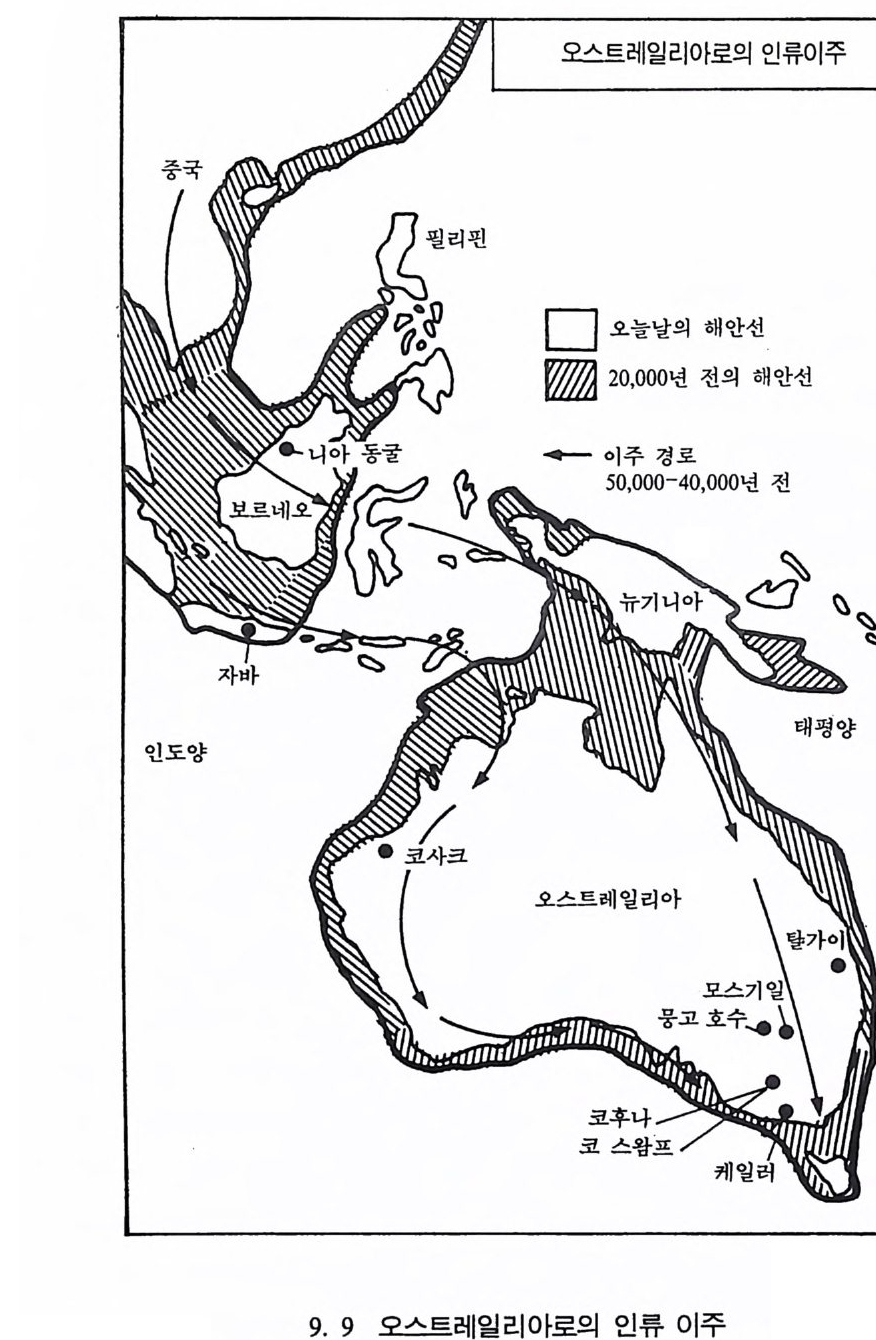 오스트레일리아로의 인류이주
오스트레일리아로의 인류이주
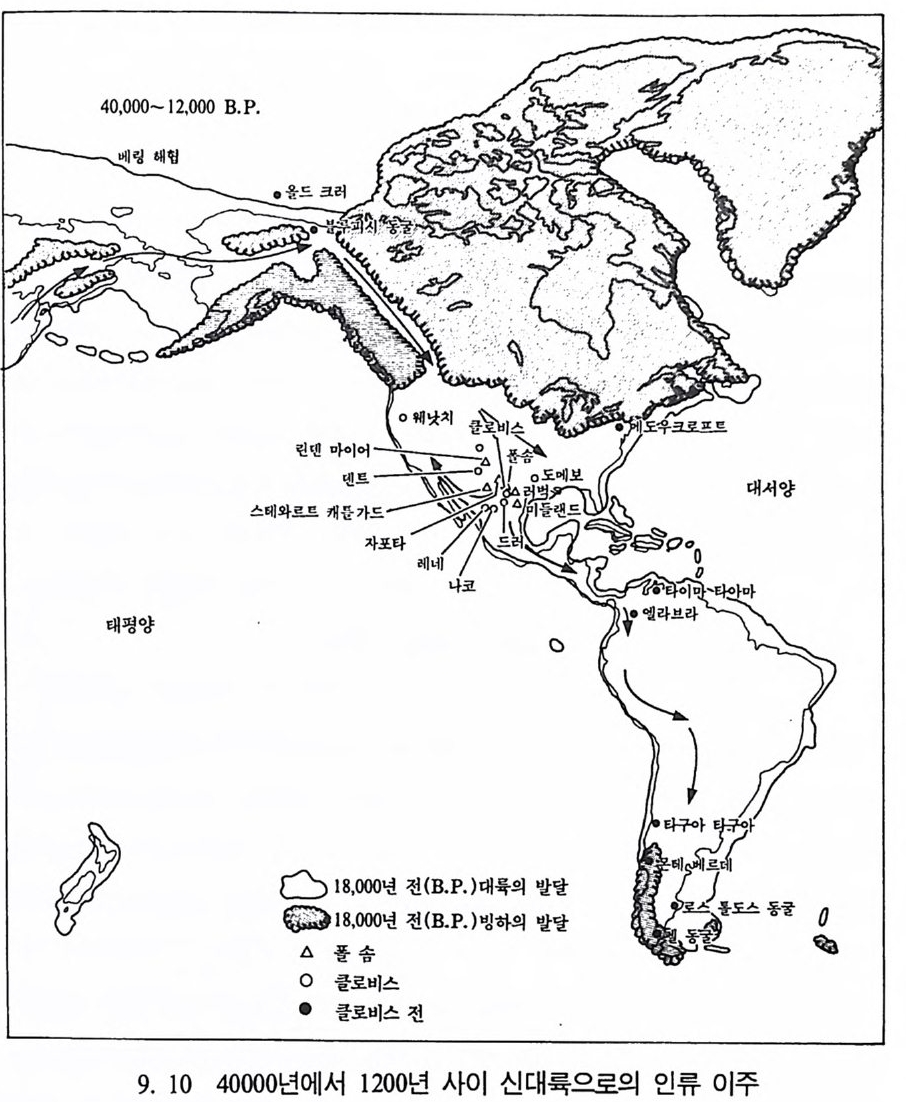 40,000~ 12,0 0 0 B. P.
40,000~ 12,0 0 0 B. P.
오스트레일리아로의 이동은 특별히 설명하기 어려운 문제이다. 그 러나 최초의 오스트레일리아 사람은 동남아시아 어느 지역에서 배를 타고 50000 년 전경에 오스트레일리아 대륙에 도착했다고 여겨진다. 그러나 가장 이른시기에 주민의 체질 형태는 아직 알려져 있지 않다. 25000 년 내에 발견된 인류화석 머리뼈의 경우, 어떤 머리뼈는 같은
시기 유럽과 아시아에서 살던 호미니드의 머리뼈보다 가볍기도 하며, 어느 머리뼈는 더 투박한 특칭을 지니고 있다. 일반적으로 인류진화 중 투박한 인류의 머리뼈는 늦은시기로 갈수록 섬세해지는 경향을 띠 고있다 이렇게 섬세한 머리뼈로부터 투박한 머리뼈에 이르기까지 다양한 머리뼈 특칭의 발견은 오스트레일리아 대륙 내에서 환경과 행위상의 요인들에 의한 〈지역 작은 전화〉의 결과로 설명될 수 있다. 이에 반 해서 월포프 같은 다지역 기원 주장자들에 따르면 이런 다양성은 자 바에서 살았던 솔로사람(낭동사람)들의 영향을 받았기 때문이며, 이론 시기 슬기사람과의 교배 흔적이 오스트레일리아 원주민의 유전자에 남아 있다고주장한다. 슬기사람의 아프리카 기원 모델의 문제 최근에 발견된 화석인류와 고고학상, 유전학상, 그리고 언어학상의 증거들은 슬기사람의 아프리카 단일지역 기원설을 지지하며, 학자들 의 의견은 단일지역 기원설로 모아지고는 있으나, 이 가설이 더욱 설 득력을 갖기 위해서는 아래와 같은 몇 가지 중요한 의문이 해결되어 야한다. 아프리카 대륙에서 슬기사람이 전화했다는 사실을 설명하기 위해서 는 궁극적으로 아프리카 대륙만이 인류를 위한 어떤 조건들이 존재했 는지를 설명해야 한다. 이 조건들은 인류의 행위와 이에 관련된 생태 상의 것들로, 우리는 실제 이 시기의 아프리카 대륙의 생태조건에 대 해 별로 알지 못한다. 아프리카 대륙으로부터 서아시아로, 그리고 이어 지구상의 기타 지 역으로 왜 이른시기 현생인류의 확산이 일어났는지를 설명해야 한다. 아마도 이른시기 슬기사람의 이주는 기후상의 변화나 인구증가 등과
함께 점차 환경에 대한 인류의 적응능력 때문이라고 여겨진다. 실제 로 인류를 확산시킨 중요한 〈 선택적 요인 〉 이 무엇이었는지는 아직 밝 혀 지 지 않았으나, 그 요인들은 50000 년부터 15 만 년 전 사이 에 작용 했음에 틀림없으며, 이 요인들이 술기사람들로 하여금 약 10000 년 전 까지 계속해서 이주하도록 영향을 미쳤을 것으로 보인다. 언어의 출현과 이에 따른 행위가 지니고 있는 이점이 아마도 슬기 사람들로 하여금 먼저 살던 호미니드의 자리를 대신하게 하였다고 가 늠된다. 학자들은 변화가 대개 70000 년 전 전후에 일어났다고 가늠하 며, 왜 이런 이점이 이때에 일어났는지롤 알고자 한다. 많은 고고학자 들은 이른시기의 슬기사람이 다론 호미니드에 비해 뚜렷한 어떤 이점 울 가지고 있지 않으며, 슬기사람이 지닌 문화 특성은 이론시기의 슬 기사람을 대신한 후인 약 30000 년 전 전후에 발달했다고 말하고 있 다. 다지역 기원설과 아프리카 단일지역 기원설 및 교배 • 교체설의 충 돌을 해결하기 위해서는 늦은시기의 곧선사람 (S t a g e ill)과 이른시기 슬기사람 사이의 유전자 교환문제가 해결되어야 한다. 미토콘드리아 핵산 연구에 바탕을 둔 아프리카 단일지역 기원설은 전적으로 슬기사 람이 먼저 살던 호미니드를 대신하였다고 주장한다. 그러나 어떤 수 렵 /채집집단이 지구의 도처에서 조금도 유전자 교환없이 다른 호니 미드를 대신할 수 있었을까 하는 문제가 제기된다. 일단의 학자들은 동유럽 같은 지역에서는 늦은시기의 곧선사람과 가장 이론시기의 슬기사람 사이에 유전자 교환 흔적이 있다고 주장한 다. 또한 네안데르탈사람은 같은 시기에 살던 이른 호미니드들이 사 용했던 연모 제작기술과 함께 나란히 사용했던 그들의 오랜 연모 제 작기술들을 변화시켰다고 일반적으로 받아들여지고 있다. 네안데르탈 사람들이 사용했던 연모 제작기술의 변화는 새로이 이주해 온 주민들 로부터 배워서인지 아니면 유전자 교환을 이루면서 얻어진 것인지에
대한 문제가 해결되어야 한다. 인류화석에서 유전자 교환의 흔적이라고 설명될 수 있는 특칭들이 찾아질 수 있는가 하는 문제 또한 설명되어야 한다. 더 나아가 설사 유전자 교환이 일어났다 하더라도 유전자 교환은 지역에 한정되어 일 어났을 것이며, 따라서 이종간의 유전자 교환에서 생긴 자손들의 유 전자는 슬기사람에게 아주 낮은 빈도로 전해지거나 아니면 전혀 전달 되지 못했을 것이다. 그런 경우, 잡종의 특칭을 지닌 화석을 발굴하였 다 하더라도 현대 유전학상 연구에서는 그 경로를 추적할 수가 없다 는 점을 염두에 두어야 한다 . 아프리카 단일지역 기원설은 위에 제기된 여러 문제들을 논리를 세 워 설명해야 하는 과제를 가지고 있다. 수년 내로 고인류학자들과 유 전학자들을 놀라게 할 많은 기술과 연구가 진행될 것이다. 그러나 점 차 슬기사람은 아프리카와 중동아시아 지방에서 존재했으며, 이후 유 럽과 오스트레일리아로 전파하였다는 가설로 의견이 모아지고 있다. 또한 비록 기원연대가 언제인지는 논쟁이 되고 있지만, 유전학 연구 로 슬기사람이 궁극적으로 아프리카에서 기원하였다는 데에 점차 동 의하고 있다. 멀지 않은 장래에 새로운 인류화석의 발견과 뼈화석과 돌연모들, 그리고 유전학 연구롤 통해 새로운 사실이 밝혀질 것이다. 인류화석 으로부터 미토콘드리아 핵을 추출해 넬 수 있는 기술이 발달할 것이 며, 그 결과 현생인류에서 얻을 수 있는 모든 결과와의 비교가 가능 해질 것이다. 과거에는 알지 못했던 새로운 방법과 기술의 발달 결과, 우리는 슬기사람의 기원과 그들이 지구를 정복해 가는 과정에 필연적 으로 관계가 있는 인류 이동에 관해 이해할 수 있으며 또 그 과정을 재구성할 수 있을 것이다.
후기 구석기문화 연모의 특징과 발달 네안데르탈사람들이 담당한 문화 시기를 중기 구석기라고 부르는 데, 구석기는 그리스의 〈 p ala i(옛적에 : long ag o) 〉 란 말과 〈 l it hos( 돌 : s t one) 〉 이란 말의 복합어이다. 이 시기의 돌연모 문화는 무스테리앙 연모와 동아시아, 아프리카에서 출토된 연모들 중 상고안과 파우레스 미스에서 출토된 유형의 연모를 모두 포함하는 개념으로 쓰인다. 한 편 전기 구석기시대 돌연모는 울도완과 아슐리앙 연모들을 의미한다. 약 40000 년 전에 유럽에서 나타난 슬기사람인 크로마뇽사람들이 담 당했던 문화를 후기 구석기 Up pe r Paleol ithi c 라고 부르는데, 이 말은 유적의 지층 위쪽에서 나온 연모라는 뜻이다 . 후기 구석기문화 담당 자들은 네안데르탈사람들이 지녔던 무스데리앙 연모와는 디론· 돌연모 를 사용하였다.
 9. 11 돌날 격지로 만든 후기 구석기 연모인 앵무새 부리 모양의 새기개와
9. 11 돌날 격지로 만든 후기 구석기 연모인 앵무새 부리 모양의 새기개와
대다수의 중기 구석기 연모들은 르발루와 방법으로 뗀 격지에 날을 다듬어 원하는 연모를 만들었다 . 후기 구석기시대 연모들도 뗀 격지 로 만들었으나, 이들 연모는 네안데르탈사람들이 격지를 만드는 데 사용한 방법과는 다른 새로운 방법을 사용하여 돌날 격지라고 불리는 특별한 격지를 만들었다(사전 9. 11). 돌날 격지는 양쪽의 날이 나란 하며 길이가 너비에 비해 두 배 이상이 되는 얇은 격지를 말하는데, 이때 돌날 격지는 대고 떼기 방법으로 만들어졌고 이 방법으로 많은 격지들을 하나의 돌감에서 얻을 수 있다. 각기 다른 4 개의 돌연모 문화를 비교해 볼 때, 같은 무게의 돌감에 서 얻어진 연모의 자르는 면의 전체 길이는 아래와 같다.
450 g의 돌감에서 얻어지는 날의 전체 길이 후기 구석기 305cm 무스테리앙 102 아슐리앙 20 올도완 5
위에서 볼 수 있듯이 후기 구석기시대에서는 돌감을 아주 아껴 썼 으며, 많은 격지를 얻기 위해 다양한 연모 제작기술이 발달했다. 무스 데리앙 연모의 경우 지역에 따라 다른 형태의 돌연모들이 나타난다는 사실을 앞장에서 살펴보았는데, 후기 구석기시대에는 더 많은 돌연모 문화들이 나타난다 . 프랑소와 보르드 Franso i r Bord ei근 200 만 년 동안 의 전기와 중기 구석기문화에서 사용된 연모들을 모두 62 개의 유형으 로 분류하였다. 한편 손누빌 보르드 M. Den ise de Sonnevil le Bordes 는 단지 30000 년 동안의 후기 구석기시대에 나타난 연모를 모두 112 개의 유형으로 분류하였다. 이 유형의 수는 전기 • 중기 구석기시대에 비해 엄청난 증가를 보여주고 있다.
한편 뼈와 뿔 그리고 이빨들이 연모의 재질로 사용되었는데, 이는 후기 구석기시대의 연모를 대표하는 특칭 중의 하나이다. 중기 구석 기시대 무스데리앙 문화에서는 일부 손질되지 않은 뻣조각들이 연모 로 사용된 경우는 있어도, 뼈를 손질하여 만든 연모들은 나타나지 않 는다. 후기 구석기시대의 대표되는 연모로는 새기개를 둘 수 있다. 이 연 모는 비스듬히 잘려져 나간 단면의 끝을 사용한 연모로 오늘날의 끌 과 같은 기능과 모양을 가지고 있다. 새기개는 뼈나 돌에 그립를 새 기거나 연모의 손잡이로 사용되던 나무나 뼈에 홈을 파 넣는 데 사용 되었다. 후기 구석기시대에 사용된 연모들에 관한 연구는 유럽에서 출토된 연모들을 바탕으로 이루어졌는데, 돌칼과 찌르개, 홈날연모, 뚜르개, 밀개와 창던지개와 창, 그리고 창의 몸체를 바르게 펴는 연모 등이 특칭으로 나타난다. 이들 연모들은 주로 프랑스의 도르도뉴 지역의 여러 유적지에서 발견되었다. 이 지역의 20km2 범위 안에서 약 200 개 가 넘는 후기 구석기 유적들이 발견되었으며, 어떤 유적의 퇴적층은 60-80cm 에 달한다• 그 결과 후기 구석기 연구는 이 지역을 중심으로 진행되었으며 , 따라서 오늘날 우리가 사용하는 용어의 대부분이 프랑 스말로표시되어 있다. 후기 구석기 사람의 행위 후기 구석기시대에 들어오면서 이 문화를 담당한 사람들의 행위와 사회조직 동에 많은 변화가 일어났다. 먼저 다양한 연모의 출현과 더 불어 사냥감의 대상인 동물에서도 변화가 나타난다. 중기 구석기시대 에는 사냥감의 50% 정도가 단일종의 동물임에 반하여 후기 구석기시 대에는 사냥된 단일종의 동물인 사슴의 출현 빈도가 80% 이상이 된
다. 이와 더불어 물고기와 물가에서 사는 동물 및 새들의 사냥이 증 가하였다. 중기 구석기시대에 사람이 살던 유적지의 크기는 평균 4m2 가 넘지 않았으며 그 수도 제한되어 나타나고, 후기 구석기시대에는 작은 규 모의 유적지도 많으나 큰 유적지는 약 6000m2 가 넘는 경우도 있다. 이는 바로 후기 구석기시대에 인구가 중기 구석기시대에 비해 크게 증가했음울 나타내는 것이다. 프랑스의 펭스벵에서 발견된 주거지를 보면 이들이 살던 주거지의 구조는 대개가 문이 남쪽이나 남서쪽을 향해나있음을알수있다. 한편 후기 구석기 사람들은 먼 거리에 살던 집단과 접촉을 하거나 아니면 먼 거리를 여행했다는 사실을 알 수 있는데, 이는 도르도뉴 지방에서 약 500km 나 멀리 떨어진 지중해에서 살고 있는 조가비들의 발견이 이를 증명해 준다. 이런 종류의 조가비는 같은 지역에서 발견 되는 중기 구석기시대의 무스데리앙 유적지에서는 발견되지 않는다. 유럽의 후기 구석기문화들 유럽에서 후기 구석기문화는 약 30000 년 정도 (40000-10000 년 전) 지속되는데 이 기간 중에 유럽의 각 지역에서는 여러 다른 문화들이 지속해서 나타난다(그립 9. 12). 이 시기 중에 유럽은 빙간기로 따뜻한 기후가 시작되었으나 30000 년 전을 전후하여 추운 시기로 바뀌었다가 20000 년 전에 다시 따뜻한 기후가 참시 지속되었다. 그리고 18000 년 전을 전후해서 유럽에는 마지막 빙하기가 도래하여 혹심한 추위를 몰 고온 바이첼 빙하기가 시작되며, 11000 년 전을 전후해서 현재 시기인 홀로세가시작된다. 유럽에서 살던 후기 구석기시대 인류는 중기 구석기문화를 담당했 던 네안데르탈사람에 비해서 아주 발달된 문화를 발전시켰다. 유럽에
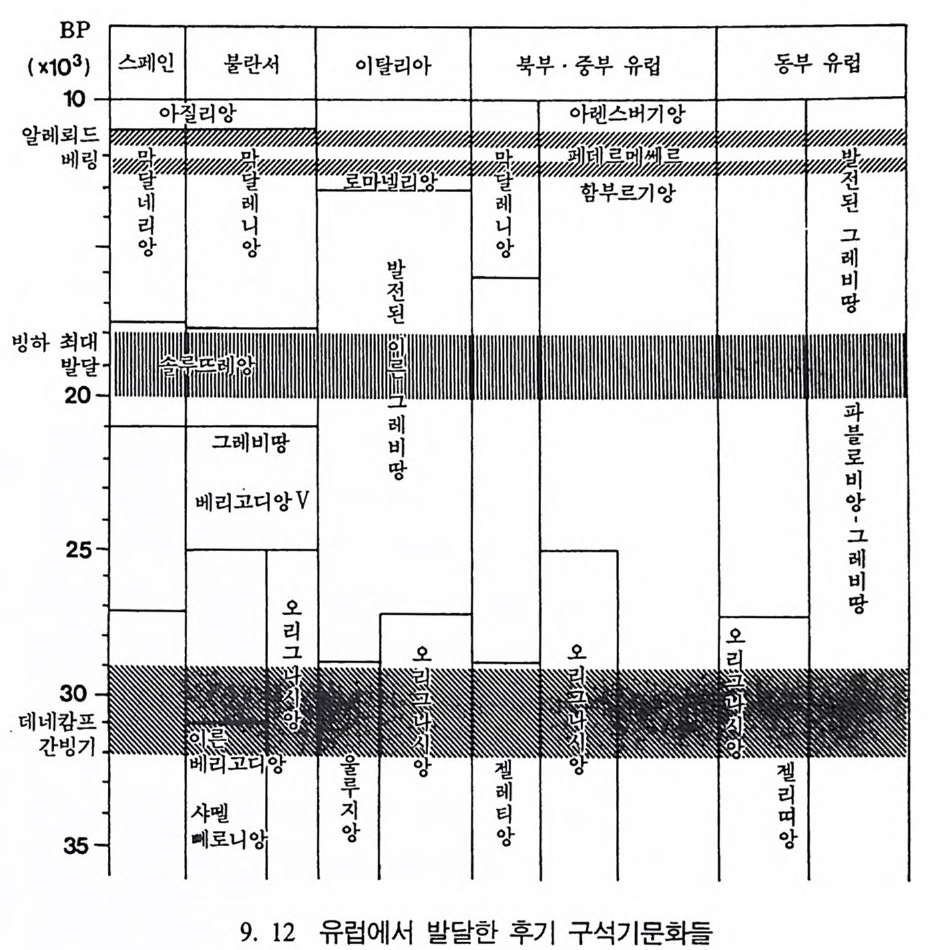 BP
BP
서 대표되는 후기 구석기문화를 남긴 유적지들은 주로 프랑스 남서부 지방에서 발견되어 집중 연구가 이루어졌다. 오늘날 선사고고학에서 사용되는 후기 구석기문화의 이름들은 이 지역을 중심으로 한 연구 결과이다. 여기서는 이 지역을 중심으로 연구된 후기 구석기문화들을 연대기순으로 간단히 살펴본다. 샤텔페로니앙 Cha t el p erron i an 오늘날 고인류학계가 가지고 있는 의 문 중의 하나는 바로 네안데르탈사람들의 운명에 관한 것이다. 네안
데르탈사람은 술기사람에 의해 제거되었을까? 아니면 그들에게 동화 되었을까? 또는 우리가 알지 못하는 종말을 맞았을까? 이런 의문이 해결되지는 않았지만 네안데르탈사람들이 지닌 돌연모 제작기술의 전 통은 얼마 동안 후기 구석기 연모에서 찾아전다. 32000 년 전에 시작 되는 가장 이른시기의 후기 구석기문화인 샤텔페로니앙 문화는 무스 데리앙 문화와 직접적인 관계가 있는데, 다양한 밀개와 무스테리앙 찌르개 및 무스테리앙 전통의 흔적이 남아 있다. 따라서 샤텔페로니 앙은 아마도 중기 구석기시대에서 후기 구석기시대로 넘어가는 과도 기적인 시기에 발달한 문화로 간주되며 이 시기를 대표하는 연모로는 〈등을 죽인 자르개〉를 들 수 있다. 오리나시앙 Aur ig nac i an 30000 년 전에 유럽의 기후는 점차 추워지기 시작했다. 이 시기에 중부와 동부 유럽에서 일단의 주민들이 문화적 으로 뒤떨어진 지역인 서부 유럽으로 이주를 하였다. 크로마뇽사람으 로 알려진 슬기사람인 이들은 뛰어난 수렵인들로 오리나시앙이라 불 리는 문화를 지니고 있었다. 이 문화는 유럽 전역에 퍼진 문화로, 자 르개가 가장 대표되는 연모이며 뼈와 뿔로 만든 연모가 처음 나타난 다. 오리나시앙 문화를 가진 크로마뇽사람들은 예술가로, 이들은 비너 스로 알려진 여인의 조각상을 남기었다. 이 조각상은 풍요와 임신을 기원하는 그들의 염원이 상칭적으로 담겨 있다고 해석된다. 최근에 새로운 해석은 추운 시기에 유럽 전역에서 발견되는 비너스상은 혹독 한 추위에서 살아 남기 위해 작은 집단의 크로마뇽사람들이 사용한 정보의 수단으로 설명되고 있다. 그라베티앙 Grave tt i an 오리나시앙 문화는 뒤이어 일어난 그라베티앙 문화에 의해 대체된다. 그라베티앙 문화는 약 25000 년 전부터 프랑스
납부지역에 퍼져 나갔다. 이 문화가 시작될 시기에 유럽의 기후는 제 법 따뜻하였으나 곧이어 혹독한 추위로 바뀌었다. 그라베티앙 연모에서는 샤텔페로니앙 연모의 전통이 나타나고 있 다. 〈등이 뭉뚝한 찌르개〉가 대표되는 연모이며, 뼈의 사용이 증가하 여 뼈로 만든 뚜르개와 펀치 및 간단한 찌르개 등이 나타난다• 후기 구석기 예술의 대표되는 유적지인 프랑스 라스코 동굴의 채색벽화는 이 시기중에 만들어지기 시작했다. 솔루트레앙 Solu t rean 그라베티앙과 오리나시앙 문화는 20000 년 전 에 갑자기 사라지고 솔루트레앙 문화가 이들을 대신한다. 솔루트레앙 문화 담당지들이 누구이며 어디서 왔는지는 아직 수수께끼로 남아 있 지만 파이퍼 P feiffe r 는 이들이 프랑스 남동지역에 고립되어 살던 네 안데르탈사람의 후예라고 추측하고 있다. 솔루트레앙 문화는 유럽에서 빙하가 최고에 달했을 시기에 발달한 문화로 이 문화의 존속기간은 불과 2-3000 년에 불과하다. 그러나 솔 루트레앙 사람들은 부싯돌을 재료로 눌러떼기 수법으로 만든 월계수 잎 모양의 정교한 자르개를 만들었다. 막달래니앙 Ma g dalen i an 부싯돌을 재질로 정교한 연모를 만들던 솔 루트레앙 문화 담당자들 또한 알 수 없는 이유로 갑자기 사라지고 이 자리는 막달레니앙 문화 담당자들에 의해 대신된다. 솔루트레앙 문화 담당자들은 뛰어난 돌연모 제작자들임에 반해 막달레니앙 문화 담당 자들은 돌감을 잘 다루지 못하였다. 따라서 막달레니앙 문화 담당자 둘을 솔루트레앙 문화 담당자들과 관련지을 수 없다. 막달레니앙 문화 담당자들이 어디서 왔는지는 알 수 없지만 이들은 제 4 기 마지막 빙하기의 여러 문화 중 가장 뛰어난 연모의 제작기술을 지니고 있었다. 서부 유럽의 툰드라 환경에서 살던 순록은- 이들의 주
요 식량자원이었으며, 털코끼리와 야생말, 들소 그리고 작은 몸집의 동물들이 사냥의 대상이었다• 막달레니앙 문화 담당자들은- 유럽에 널 리 퍼져 동굴과 한데 유적에서 살았으며, 이전에 살던 주민들에 비해 인구밀도가 증가하였다.
 9. 13 후기 구석시대 (18000-10000) 에 뼈로 만든 H 海t
9. 13 후기 구석시대 (18000-10000) 에 뼈로 만든 H 海t
막달레니앙 문화 담당자들은 뼈와 뿔을 사용하여 다양한 모양의 연 모를 만들었는데(사전 9. 13), 연모둘은 아름답게 치장되었다. 한편 그라베티앙 문화에서 시작된 후기 구석기시대의 예술인 동굴벽화는 막달레니앙기에 와서 절정에 달하였다. 이런 동굴벽화와 조각품들은 프랑스 남서지역과 스페인의 북동쪽의 여러 유적지에서 발견되는데, 유명한 스페인의 알타미라 동굴의 채색벽화들도 이 시기에 완성되었 다(사전 9. 14, 9. 15). 지금까지 현생인류인 술기사람이 언제, 어디서 처음 나타나기 시작 했는지에 대해서 살펴보았다. 최근의 유전학상의 연구와 고고학 및 고인류학 연구들은 이른시기의 슬기사람인 해부학상 현대인은 약 20
 。 rt’ •• • ’· ' I • •• • • • .,.
。 rt’ •• • ’· ' I • •• • • • .,.
 、 /, J A``
、 /, J A``
9. 14 후기 구석기시대의 동굴벽화들
 스 불란서 라우셀에 있는 바위그늘 유적에 새겨진 비너스상으로 손으로 물
스 불란서 라우셀에 있는 바위그늘 유적에 새겨진 비너스상으로 손으로 물
 • 1900~ 전부터 16000 년 전 사이에 순록의 • 19000 \1 전에서 16000 년
• 1900~ 전부터 16000 년 전 사이에 순록의 • 19000 \1 전에서 16000 년
9. 15 후기 구석기시대에 만들어진 조각품들
만 년 전에 남아프리카에서 시작하여 10 만 년 전을 전후한 시기에 중 동아시아에 도달하였음을 밝히고 있다. 소위 노아의 방주설이 슬기사 람의 이동 모델로 더 지지를 받고 있다. 그러나 20 만 년 전에 출현한 이들이 오늘날의 우리와 똑같은 존재 를 의미하는 것은 아니다. 우리와 똑같은 존재의 기준을 자의식을 갖 춘 존재로 삼는다면, 자의식을 가진 인류가 나타나는 시기가 바로 우 리와 똑같은 인류가 나타나는 때라고 생각하는 것이 타당하다(그립 9. 16). 선사인류가 자의식을 지녔는가에 대한 여부는 이들에게서 예 술의 행위가 언제 나타났는가에 달려 있다. 선사인류에 의한 예술행 위의 증거들은 35000 년 전에 소위 크로마뇽사람으로 알려진 슬기사람 에 와서야 비로소 나타난다. 또한 복잡하고 정교한 기술을 바탕으로 한 문화가 이 짧은 시기중에 발달하였으며, 이 문화를 지닌 슬기사람 들은 전 세계로 퍼져 나갔다 . 진정한 우리의 모습인 슬기사람이 출현 한것이다.
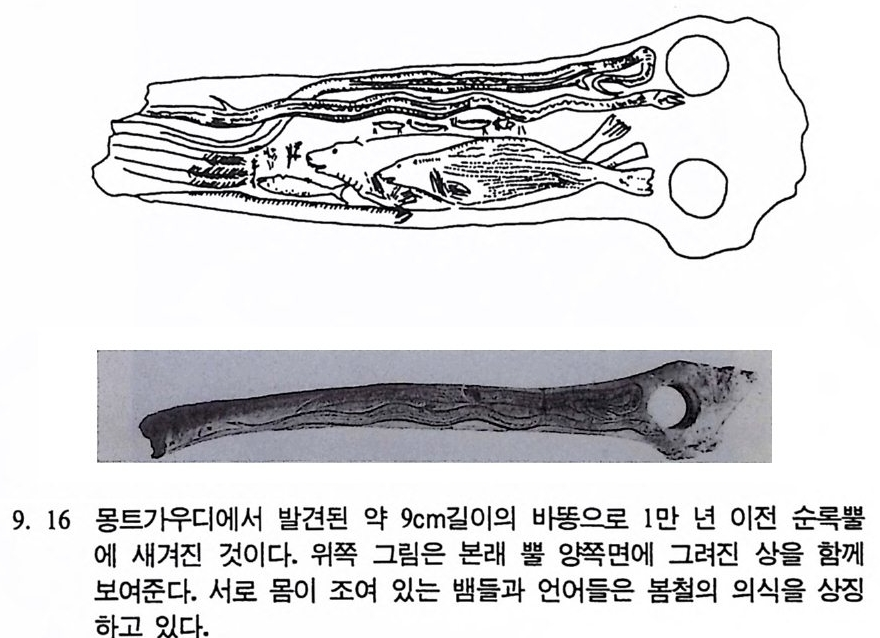 9. 16 몽트가우디에서 발견된 약 9cm 길이의 H 追옵으로 1 만 년 이전 순록뿔
9. 16 몽트가우디에서 발견된 약 9cm 길이의 H 追옵으로 1 만 년 이전 순록뿔
 제 10 장 인류의 적응과 사회생물학
제 10 장 인류의 적응과 사회생물학
제 10 장 인류의 적응과 사회생물학 인류 : 어떻게 자연에 적응하나? 지난 400 만 년 동안 호미니드는 전화과정 중, 생물학상의 변화와 정 체를 되풀이하면서 기술상 발달을 보여주고 있다. 이와 더불어 호미 니드는 성에 따른 특징이 점차 줄어들었으며, 이런 과정은 다른 동물 들에 비해 아주 빠른 속도로 일어난 것이다. 머리 크기는 200 만 년 사이에 세 배로 늘어났으며 이들이 퍼져 살던 지리적인 범위도 급속 히 확대되었다. 가장 이른시기의 인류 조상인 오스트랄로피데쿠스와 손쓴사람은 아프리카에서 생겨나서 대륙을 벗어나지 못했다. 곧선사 람에 와서야 비로소 유라시아 대륙으로 퍼져 나갔으며 오스트레일리 아와 신대륙으로의 이주는 현생인류인 술기사람에 의해서 이루어진다. 예술은 30000 년 전 동굴벽화에서 시작하여 막스 헤드럼 Max Head- room 까지 발달하였다. 인류의 조상과 인류가 지녔던 경제체계는 기 회주의인 주워먹기침승으로부터 수렵 • 채집단계를 거쳐 10000 \1 전 울 전후해서 생산단계로 집어들기 시작했다. 오늘날 슬기사람인 현생 인류는 지구 곳곳에 퍼져 살고 있다. 이들은 각기 채식, 육식 또는 잡 식을 하면서 다른 환경에 널리 퍼져 번성하며 살아가고 있다. 이렇게 인류가 번성할 수 있는 주요 열쇠는 인간이 갖고 있는 행동
의 유연성 때문이라고 할 수 있다. 인간은 조상들이 축적해 온 지식 을 배우기 때문에 오늘날의 우리가 있다. 한 세대에서 얻어진 문화의 득칭은 라마키안의 이론처럼 다음 세대로 전달된다. 오늘날의 인간은 그들의 조상보다 더 영리하지는 않지만 지식은 빠르게 확대 및 축적 되어간다. 이렇게 축적된 지식은 바로 다음 세대로 전해지며, 즉각 유 용하게 쓰여질 수 있다. 이 장에서는 지구의 각기 다른 환경에서 사는 현대인류 집단이 어 떻게 환경에 생물학상 적응을 하는가 살펴보자. 환경에 따른 인간의 적응을 연구하는 분야는 〈인류생물학〉의 주된 연구분야이다. 이 분야 의 연구는 단순히 환경에서 오는 추위와 더위, 그리고 높은 지대의 생활 등이 현대인에게 미치는 선택되는 영향의 관찰에서 시작된다• 인간에게 영향을 주는 갈등의 요인들을 살펴보기 전에 사람의 몸은 압박을 받을 때 몸을 일정한 상태로 유지하기 위해서 다양한 방법으 로 반응한다는 점을 알아야 한다. 인간은 주어진 환경에 〈적응 Ada pta ti on 〉과 〈생리상 조절 Ph y s i o log ica l Adju s tm ents > 방법으로 자신과 환경과의 관계를 조절한다. 일반적으로 적응이란 말은 유전상, 생리상 그리고 문화상 반응을 나 타낼 때 쓰인다. 그러나 엄밀히 말하면 〈적응〉이란 개념은 〈생물학상 적응 Gene ti c Ada pta ti on 〉과 〈문화 적응 Cul t ural Ada pta ti on 〉을 의 미한다. 사람이 살아 있는 동안에 차용하는 생리상 조절로는 〈순응 Acc li ma ti za ti on 〉과 〈순화 Acc li ma ti on 〉란 조철방법이 있다. 생물학상 또는 유전학상 〈적응〉이란 자연선택을 동해서 얻어진 유 전인자가 개인에 있어 오랜 세월에 걸쳐 진행되는 진화과정을 의미한 다. 이에 대해 〈순응〉은 오랜 기간 동안 환경에 대한 개인의 비유전 적 조절방법을 뜻한다. 이에 반해 짧은 기간 동안의 조철방법은 〈순 화〉라고 부른다. 한편 인간은 생물학상 〈적응〉과 생리상 순응 • 순화 이의에 문화 〈적응〉으로 주어전 환경에 맞추어 살아간다.
추위에 의한 갈등 사람은 항온동물이기 때문에 항상 일정한 체온을 유지해야만 한다. 기온이 낮아지면 사람은 몸 안에서 더 많은 열을 만들어 체온을 일정 하게 유지하고, 만일 기온이 울라가 더워지면 스스로 체온을 떨어뜨 린다. 추위에 대한 적응, 죽 날씨가 추울 때 사람은 체온을 일정하게 유지하기 위해 유전상 적응과 생리상 조절 그리고 문화 적응 등의 방 법에 의존한다.
 10. 1 추운 지 방의 몸떨림 Hy po sia
10. 1 추운 지 방의 몸떨림 Hy po sia
추위를 덜기 위한 비의도적인 운동으로는 유전상 적응인 몸떨림이 일어난다(사전 10. 1). 이때 몸떨림은 사람에 따라 그 정도가 다르다. 한 예로 미국의 생태학자인 풀 베이커 Paul Baker 는 퀘추아Q uechuas 인디언과 백인 학생들을 대상으로 손가락이 추위에 노출됐을 때의 적 응 정도를 비교하였다. 그는 조사를 통해 〈인디언과 백인 학생들의
추위에 대한 적응은 환경과 생리상의 조절보다는 유전 차이에서 오는 것 〉 이라고 설명하고 있다. 한편 의도있는 운동으로는 생리상의 조절과 문화 적응을 둘 수 있 다. 사람은 손발이 원활히 움칙일 수 있도록 근육을 항상 따뜻하게 유지해야 하며, 특히 손가락과 발가락에 따뜻한 피가 원활히 공급되 어야 한다. 날씨가 추워지면 손가락과 발가락을 통해 열의 손실이 일 어나 몸의 내부 온도가 떨어지게 되는데, 체온의 급격한 하락은 죽음 의 원인이 되므로 열 손실을 막아 손가락과 발가락을 따뜻하게 유지 하여야 한다. 이렇게 사람이 추위에 노출되면 제일 먼저 손가락과 발 가락에 혈관수축이 일어난다. 이런 현상은 열 손실을 막기 위해 사지 끝으로 가는 피의 흐름을 줄이기 때문이다. 다시 따뜻해지면 혈관확 장이 일어난다.
 10. 2 에스키모의 방한복
10. 2 에스키모의 방한복
이렇게 기온이 변했을 때 사지에서 주기적으로 반복되는 혈관수축 과 확장은 사람이 체온을 급격히 떨어뜨리는 것을 예방하는 생리반응 이다. 사람들은 추위에 노출되는 몸의 면적을 줄이기 위해서 몸을 구 부리는데, 이런 자세도 생리상 적응방법이라고 할 수 있다. 한편 문화 적응은 추위를 이겨내는 가장 좋은 방법이다. 여기에는 여러 가지 방법이 있는데, 그 중 사람이 사는 건물 주변에 불을 피워 기온을 따뜻하게 하는 방법이 있다. 그러나 무엇보다도 중요한 것은 옷을 입어 추위를 막는 단순한 행위상의 적응이다 . 한 예로 에스키모 사람들이 입는 파커는 옷 안의 열이 전혀 밖으로 새나가지 않도록 고 안되어 있어 열의 손실을 최소화한다(그림 10. 2). 더위에 의한 갈등 더운 기후에 적응하는 것도 추위에 적응하는 것만큼 복잡한 조절작 용을 필요로 한다. 밖의 온도가 섭씨 30° 정도 되면 사람은 땀을 홀 려서 체온을 적철하게 유지한다. 그러나 실제로 사람에게 스트레스를 주는 것은 열 그 자체보다는 열과 습도가 함께 나타날 때이다. 열과 습도는 사람의 몸에서 땀이 나는 데 영향을 미친다. 죽 덥지만 건조 한 환경에서는 땀이 잘 증발하지만 습도가 높은 공기에서는 수분의 증발이 잘 일어나지 않아 몸이 끈적거린다. 어느 동물도 사람처럼 더위에 대한 조철작용으로 땀울 홀리지는 않 는다. 이렇게 땀을 홀리는 행위는 건조하고 더운 환경에 매우 효과있 게 순응하는 생리상의 조절방법이다. 사람은 1 시간에 1 리터 정도까지 땀을 흘릴 수 있다. 그러나 이렇게 땀을 홀리는 것은 체온을 조절하는 데 효과적이기는 하나, 몸의 수분과 소금을 모자라게 하는 원인이 된다. 만일 우리가 사하라 사막에서 물 한 병만 가지고 가다가 길을 잃었 다고 가정해 보자. 곧 우리의 몸에서는 땀이 나기 시작할 것이다. 땀
은 참시 동안 우리의 생명을 연장해 줄 수 있으나 곧이어 우리는 이 땀에 의한 탈수 때문에 죽어갈 것이다. 진화상에서 볼 때 적응이란 생존에 있어서의 이로움을 뜻한다. 이 이로움은 행동일 수도 있고, 또 는 해부상의 특징으로 나타날 수도 있으며, 아니면 생리상의 이로움 으로나타날수도 있다.
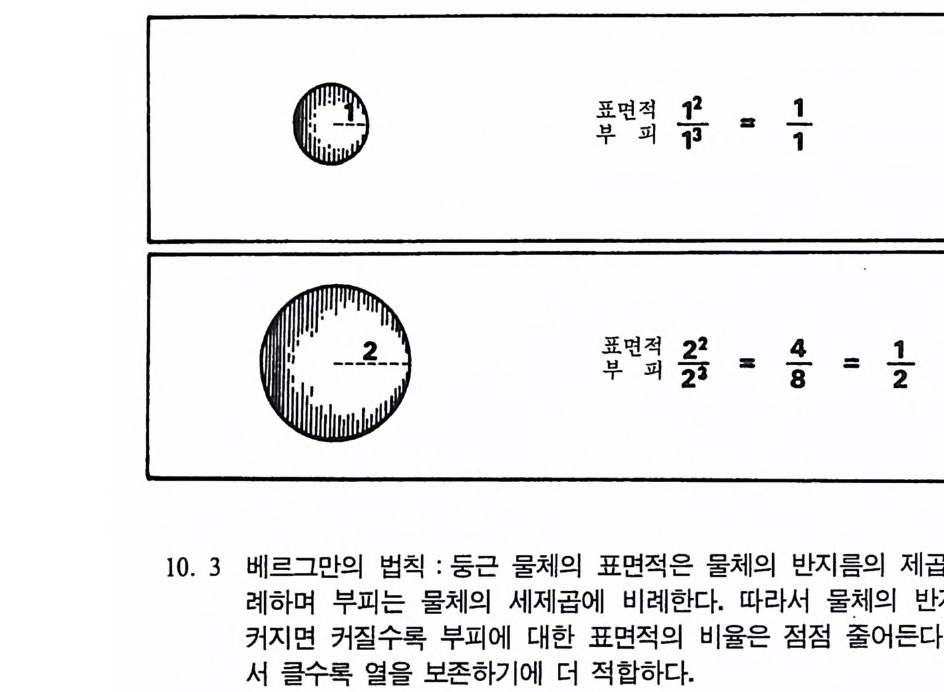 e 부표면피적 곱 Z T1
e 부표면피적 곱 Z T1
현대인들이 땀울 훌리는 데에는 어떤 차이가 있는가? 모든 사람은 lm2 의 면적에 같은 수 (200 만 개)의 땀구멍을 가지고 있다. 땀을 홀리 는 것은 바로 땀구멍의 운동에 달렸다. 땀홀립에 대한 연구 결과에 따르면 개인에 따라 그 정도가 다르지만 유전적인 차이는 없다고 한 다? 또 다른 더위에 대한 적응으로 몸에서 나타나는 해부상 반응을 들 수 있다. 19 세기의 생태학자인 베르그만 Ber g mann 과 알렌 Allen 은 젖
먹이동물을 해부하여 몸의 크기와 형태에 따른 열 손실의 관계를 공 식화하였다. 베르그만의 법칙 Bergm ann's Rule 에 따르면 같은 모양을 지닌 동물의 경우, 몸집이 큰 놈은 몸의 부피에 비해 표면적이 적어 서 열을 잘 보존할 수 있기 때문에 추위에 더 잘 적응한다고 한다. 이와 반대로 몸집이 작은 놈은 그 크기에 비해 아주 큰 표면적을 가 지고 있기 때문에 더 많은 열을 잃는다고 한다. 이는 두 개체의 몸집 울 둥근 물체(구)라고 가정하면 부피가 큰 개체일수록 표면적이 줄어 든다는 기하학의 이론을 응용한 것이다(그림 10. 3). 베르그만의 법칙은 일반적으로 젖먹이동물의 경우에는 실제로 적용 된다. 한 예로 아메리카 대륙의 표범은 살고 있는 지역의 기온이 떨어 질수록 몸집이 커짐을 알 수 있다(그림 10. 4). 한편 환경과 사람의 몸 무게에 관한 여러 연구는 더운 지역에서 사는 사람의 몸무게는 추운 지방에서 사는 사람의 몸무게보다 작다는 것을 보여준다(그림 10. 5). 몸의 크기 이의에 몸의 형태도 기후에 따라 변한다고 베르그만의 법칙은 말하고 있다. 기하학적으로 볼 때 원은 고정된 표면적에 최대 의 부피를 지니고 있다. 따라서 길고 가는 형태는 표면적에 따른 부 피가 적어지기 때문에 열을 분산시키기에 가장 좋은 형태이다. 반대 로 단단하고 퉁퉁한 몸집은 열울 잘 보존한다. 이렇게 기후에 따라 각기 다론 형태의 몸집을 지닌 예는 사하라 사막 남쪽지역에서 사는 사람들은 가늘고 긴 몸집을 지닌 것과, 북극지역에서 사는 에스키모 사람들은 키가 작고 단단한 몸집을 지녔다는 데서 찾아진다. 또 다른 생태상의 법칙으로 몸과 사지 부분과의 비율을 다룬 알렌 의 법칙 Allen's Rule 을 들 수 있다. 그의 법칙에 따르면 더운 지역에 서 사는 사람들이 긴 팔과 다리를 지니고 있는 것은 열을 분산시키기 위해 몸의 표면적을 국대화하기 위해서라고 설명하고 있다. 추운 지 방에서 사는 사람은 이와 반대로 표면적을 줄여 열 손실을 방지할 수 있도록 팔다리가 짧다고 한다. 실제로 긴 몸집과 팔 다리를 가전 사
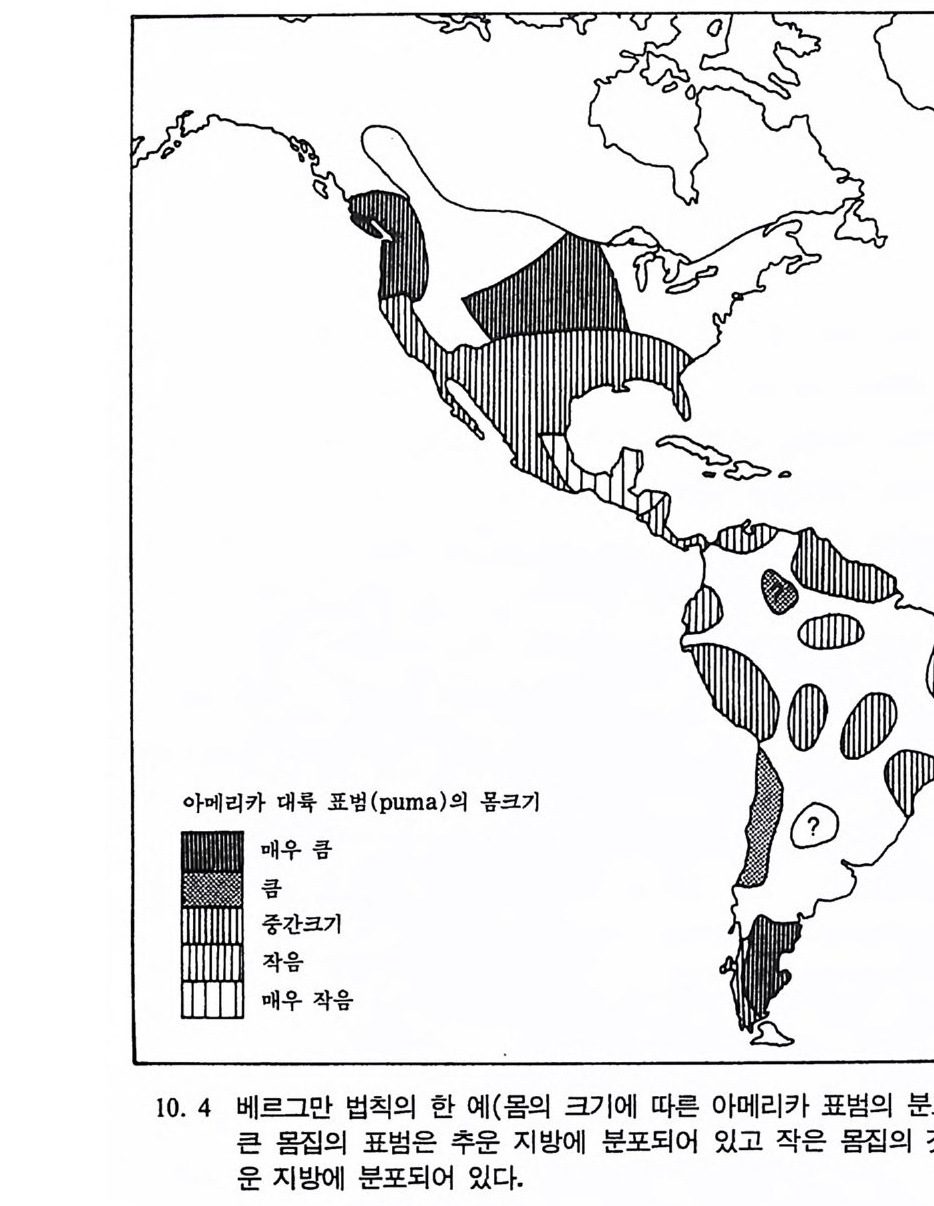 아메’리카 대륙 표범 (p uma) 의 몸크기
아메’리카 대륙 표범 (p uma) 의 몸크기
하라 사막 이남에 사는 주민과 땅딸막한 몸매에 짧은 팔다리를 지닌 에스키모 주민들에게 베르그만과 알렌의 법칙이 모두 적용됨을 알 수 있다.
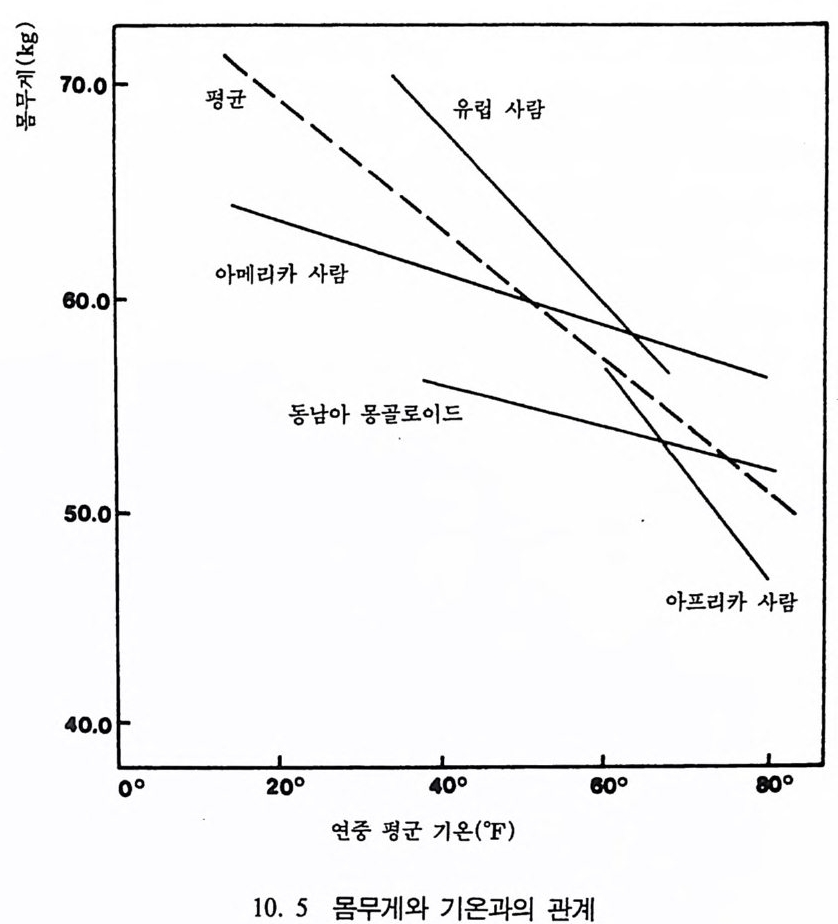 (
(
위에서 살펴본 생태상의 법칙들은, 물론 예의도 있으나, 사람의 몸 이 어떻게 생리적 방법으로 더위라는 압박을 조철하는지를 보여준다. 이의에도 음식이라든지 혈관의 확장 같은 생리조절로 더위에 의한 압 박을조철한다. 한편 사람은 문화방법으로 더위에 적응한다. 옷을 입는 것은 그 중 의 한 방법인데, 옷을 입는 행위는 그 지역의 습도 정도에 따라 효과 가 다르다. 특히 덥지만 습도가 높지 않은 사막에서 옷은 아주 훌륭 한 조절효과를 가져다 준다. 옷은 태양으로부터 받는 열이 직접 피부 에 닿는 것을 차단하는 역할을 하여 마치 그늘에 있는 것과 같은 효
과를 가져온다. 이때 증발작용에 의해 몸의 열을 잃게 하는 데는 헐 렁한 옷이 더 효과적이다. 그러나 습도가 높은 지역에서 옷은 도움이 되지 못한다. 더위에 적응하는 또 다른 문화방법으로 집의 역할을 들 수 있다. 고산지대에서 흔히 볼 수 있는 햇빛에 말린 벽돌집(아도브)과 돌로 지은 집들은 추운 지역에서뿐만 아니라 덥고 건조한 지역에서 효과가 있다(사전 10. 6). 태양열이 비치면 벽돌은 열이 내부에 도달하기 전 에 많은 열을 자체에 흡수시킨다. 따라서 낮에는 시원하고 밤이 되면 방출된 열에 의해 집안이 따뜻해진다. 이에 곁들여 창문의 구조라든 지, 칠이라든지 또는 집 주변에 심는 나무들은 모두 더위에 적응하는· 문화방법이 된다.
 10. 6 안데스 고산지대에 아도브 Adobe 벽돌집
10. 6 안데스 고산지대에 아도브 Adobe 벽돌집
한편 열대 삼림지역처럼 습기가 많고 더운 지역의 경우, 나무들에 의해 햇빛은 충분히 차단되나 습기가 문제가 된다. 따라서 공기의 순 환작용이 매우 중요한 이런 환경에서는 풀처럼 가볍고 공기의 유통이 자유로운 재료로 집을 짓는다(사전 10. 7). 때때로 공기의 순환을 극 대화하기 위해 땅 위에 얼마간의 공간을 두고 집을 짓기도 한다.
 ?T;r• ,`
?T;r• ,`
고산지대에서 오는 갈등 페루와 히말라야 고원지대 같은 높은 지대에서 사는 사람에 관한 연구는 주로 페루의 안데스 고원에서 사는 주민을 대상으로 전행되었 다. 페루의 안데스 고원은 해발이 평균 2.5km7} 넘으며 어떤 인디언 주민들은 5.2km7} 넘는 곳에서 산다(1 0. 8). 비록 짧은 시간이지만 어떤 인디언들은 6.l km 7} 넘는 곳에 있는 광산에서 일한다. 이렇게
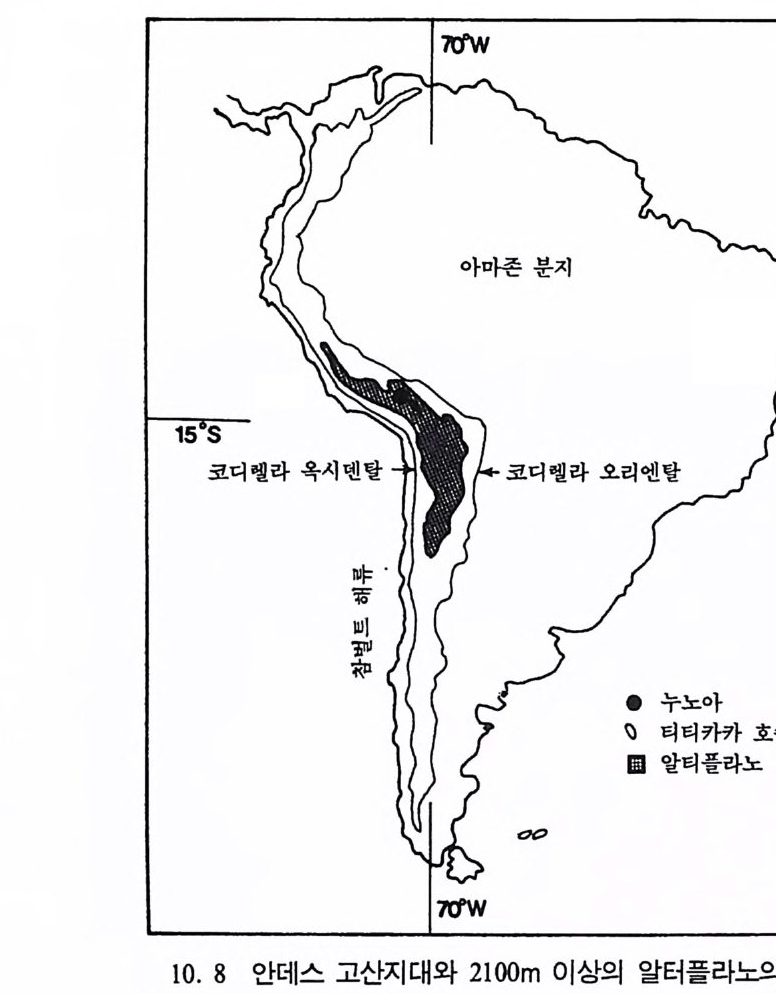 아마존분지
아마존분지
높은 지대에서 사는 주민들은 낮은 지대에서 사는 주민과는 다른 압 박을 받게 된다. 이들이 당면하는 가장 큰 압박으로 산소의 부족을 들 수 있다. 사람은 5.5 k m 높이에서까지 살 수 있는데, 4.5km(15000 피트)의 높이에서는 해수면 높이에서 호흡할 수 있는 산소의 58% 정 도만이 가능하다. 이는 대기 중 산소의 양이 줄어들기 때문이 아니라 고산지대에서는 기압이 줄어들기 때문이다. 기압의 감소는 산소를 폐 와 핏줄로 보내는 힘을 감소시킨다.
기온 또한 감은 위도상의 낮은 지역보다 더 떨어진다. 따라서 추위 또한 고산지대에서 사는 주민들의 생활을 위협하는 스트레스이다. 오늘날 우리는 환경의 변화에서 오는 기압의 변화를 조절함으로써 산소부족에서 오는 문제를 해결한다. 예로 비행기 내부가 높은 고도 에서 정상적인 기압을 유지하고 적당한 산소를 공급한·다든지, 높은 산울 등반할 때 사용하는 산소통 등은 기압의 변화에 따른 산소부족 울 해결하는 방법들로, 이런 방법들은 최근의 기술발달에서 나온 결 과이다. 그러나 고산지대에서 사는 주만들은 기술방법이 없이 오랫동 안 이런 조건에서 살아왔다. 보통 사람은 산소탱크나 기압을 정상으로 조절해 주는 장치 없이 산소부족을 견디어낼 수 있는 방법이 없다. 따라서 학자들은 고산지 대에서 사는 주민들에게 맞는 생리적응이 있으리라고 생각해 왔다. 고산지대에서 살고 있는 인디언을 연구한 폴 베이커 등의 연구 결과, 고산지대에 사는 사람들은 평지에 사는 사람들에 비해서 더 많은 양 의 적혈구와 모세관, 그리고 아주 큰 폐활량을 가지고 있음이 밝혀졌 다. 그러나 이 모든 현상은 환경에 대한 생리상 조절이지, 유전상의 차이는 아니다. 모든 사람은 고산지대에 적응할 수 있는 능력을 가지 고있다. 고산지대에서 사는 사람에게서 태어나는 아이는 낮은 지대에서 사 는 사람의 아이들보다 적은 몸무게를 가지고 태어나는데 이는 어린아 이의 생존률을 높이는 구실을 한다. 그 결과 고산지대에서는 작은 몸 무게를 지닌 유전인자가 큰 몸무게를 지닌 유전인자보다 유리하리라 고 가늠되어 생물학상 적응의 한 예로 한때 인용되었다. 그러나 최근 의 연구에 따르면 고산지대에서 사는 사람은 생물학상의 적응에 필요 한 유전인자를 거의 가지고 있지 않은 것으로 밝혀지고 있다. 이는 아마도 고산지대라는 환경에서 살아온 지 불과 수천 년밖에 되지 않 아 진화가 일어나기에는 충분한 시간이 아니기 때문으로 여겨진다.
낮은 지대에서 사는 사람이 고산지대라는 환경에 집했을 때 그는 두 단계의 생리적인 조절방법으로 압박에 반응한다. 먼저 고산지대의 주민들이 겪는 압박 중의 하나인 추위를 극복하기 위해 이들에게는 숨쉬기와 맥박이 빨라지고, 혈압과 심장의 박동이 증가하며, 혈관의 팽창이 일어난다. 그러나 이런 생리상의 조절은 세포에 필요한 산소 룰 더 얻고자 하는 작용 때문에 심장에 무리를 가져다 준다. 보다 효과있는 방법으로는 더 많은 적혈구와 헤모글로빈의 생산울 통해 심장의 부담을 줄이는 방법이다. 따라서 고산지대에서 사는 사 람들에게는 더 많은 산소를 각 조직에 운반을 할 수 있도록 피의 양 이 증가한다. 한편 산소가 부족한 지역에서 성장한 사람은 평지에서 사는 사람보다 더 커진 폐와 폐활량을 가지게 되어 이런 생리상 조절 이 고산지대에서 사는 사람들로 하여금 그들이 처한 압박을 극복하게 해준다. 그러나 이런 순응의 조철방법은 오랜 시간을 필요로 한다. 위에서 언급한 생리상의 조절과 생물학상 적응과 함께 문화 적응의 연구는 고산지대에서 사는 메스티조스(스페인 사람과 인디언 사이의 뒤기를 말함)들에 관한 많은 연구로부터 알려졌다. 메스티조스의 임 신한 여성이 유산울 할 빈도는 평지에서 임신한 사람의 빈도보다 높 다. 따라서 이들은 임신한 여성을 평지로 내려보내 출산율을 높인다. 또한 메스티조스 여성들은 평지에 사는 여성보다 일찍 결혼하며, 오 랜 기간 동안 아이룰 생산한다 . 이런 문화상의 적응은 이들의 인구수 를 일정하게 유지하는 방법 중의 하나이다. 이들 여성의 경우 초경은 평지의 여성보다 늦게 시작하나 첫번째 임신은 평지에 사는 여성보다 빠르다. 결혼 후 고산지대의 여성은 오랜 기간 동안 아이를 가지는 데 반해, 평지에 사는 여성은 결혼한 지 몇 해가 지나면 출산율이 뚝 떨어진다. 고산지대의 주민들은 위에서 언급한 방법으로 추위 때문에 높아전 태아 사망률을 상쇄한다. 더위와 추위 그리고 고산지대라는 환경이 인류에게 주는 압박을 어
떻게 극복하는가 하는 연구는 지구의 도처에서 광범위하게 진행되었 다. 많은 연구들에 따르면 사람은 서로 다른 환경에 생리상 순응보다 는 주로 문화 적응을 한다는 사실을 밝혀주고 있다. 다시 말해 기술 에 의해 환경과 사람 사이의 관계가 조절되는 것이다. 그럼에도 불구 하고 어쨌든 다른 환경에서 사는 현대인들에게는 해부상 차이들이 있 음울알수있다. 남· 녀의 성에 따른 차이 여기서 우리는 집단이 아닌 개인 사이에서 가장 중요한 차이는 어 디에 있는지를 살펴보기로 한다. 개인 사이에 있어 가장 중요한 차이 는 바로 성(性 ; Sex) 이 다르다는 사실이다. 이론 시기의 생물은 암수 구분이 없었으며, 오늘날도 암수 구별이 없는 생물들이 많다. 모든 생 물은 본래가 여성이었으며, 남성은 겉으로 보아서는 기능장애를 가지 고 있다. 암수 양성에 의해 생산을 하는 생물의 경우, 이들 전체의 절 반은 남성이 차지하고 있다. 이 절반을 차지하는 남성은 다음 세대로 계속 전할 알을 생산하지 못하기 때문에, 결과적으로 미래의 〈종〉에 대해 별로 공헌을 하지 못 한다. 암수 양성에 의한 생산은 다양성을 증가시키거나, 아니면 남성 은 새끼들을 증가시키는 데 도움이 된다는 가정은 별로 받아들일 수 없다. 영장류는 남 • 녀라는 두 개의 다른 성을 가지고 있다. 사람의 경우 는 언제, 그리고 왜 성의 차이가 생겨났는지가 연구의 초점인데, 이를 위해서 다윈의 전화론을 더 깊이 살펴보아야 한다. 다윈의 전화론은 일반적으로 생물은 자식을 번식시켜주는 방법으로 행동한다는 점을 밝히고 있다. 남성과 여성은 생리상으로 구조가 다르며 적응도를 높 이기 위해서 각기 다른 방법을 사용한다. 사람의 경우 여성은 임신을
할 수 있는 기회가 일 년에 단지 12 번인 데 반하여 남성은 어느 때나 여성을 임신시킬 수 있다. 임신을 하고 아이를 낳고 또 아이를 키우 는 과정에서 여성의 투자는 생물학상으로 볼 때 근본적인 것이지만 남성의 두자는 생물학상의 것이 아니다. 여성은 그 일생에 있어 한정된 수의 자식만을 만들기 때문에 한 남 성이 한 여성이 어머니가 될 수 있는 것보다 더 많은 자식의 아버지 가 될 수 있다. 한편 남성은 위험성은 많으나 반면에 많은 자식을 얻 울 수 있는 조건을 가지고 있다. 여기서 위험은 남성 사이에 일어나 는 경쟁으로부터 오는 것이며, 이때 경쟁에서 승리한 남성은 적응도 가 증가하는 데 반해 경쟁에 패한 남성의 적응도는 감소한다. 여성의 경우 적응도는 조심스런 배우자의 선택에 의해 높아전다. 일반적으로 남성과 여성의 행위가 서로 다른 까닭은 이들이 문화에 서 배우는 것이 다르기 때문이라고 한다. 사이몬스라는 인류학자는 이런 일반적인 통념이 옳은지를 시험하기 위해서 각 문화들을 서로 비교하고자 하였다. 특히 그는 각기 다론 문화에서 여성의 매력 At - t rac ti veness 의 기준이 무엇인지를 밝히고자 하였다. 이 연구는 임신 할 수 있는 나이에 찬 여성 모두가 고르게 남성에게 매력을 주는 것 은 아님을 보여주고 있다. 나아가 그의 연구는 어떤 문화권에서 사는 남자를 막론하고 매력을 주는 이상화되는 짝은 공통으로 아래와 같은 조건을 충족시켜야 한다고 결론짓고 있다 ; 1. 막 결혼 적령기에 든 여자일 것 2. 몸이 건강하며 특히 피부가 고운 여자일 것 3. 몸매가 비교적 통통한 여자(몸의 지방이 몸무게의 3 분의 l 정도) 일것 4. 좋은 가문의 여자일 것 5. 적극적으로 사랑을 표현하는 여자일 것 6. 헌신해서 자식을 키우는 여자일 것
만일 위에서 언급한 조건을 충족시키는 여자가 남성들에게 매력을 주는 여자라는 그의 이론이 옳다면, 각기 다른 문화권에 사는 남성들 이 어떻게 이런 조건을 가전 여자가 매력이 있는 여자라는 것을 공통 적으로 느끼게 했을까? 기존의 통념처럼 그들은 무엇이 매력인지를 그들의 문화 안에서 배워서 인식할까? 아니면 남성의 유전자 중에는 매력을 알게 하는 유전인자가 있을까? 또는 미리 남성이 그런 여성들 로부터 매력을 느끼게 하는 유전인자가 선별되어서 적응하는가? 우리 는 어떤 설명이 더 타당한지는 아직 알 수 없으나 지구상의 여러 사 회가 지니고 있는 결혼제도와 관습은 이 문제를 푸는 실마리를 제공 해 준다. 오늘날 지구상의 대다수 사회는 완전하지는 않지만 일부일처제의 결혼제도를 가지고 있다. 그러나 각기 다론 사회의 결혼제도와 관습 에 관한 연구에 따르면, 지구상에 있는 약 849 개의 사회 중 83% 에 달하는 708 개의 사회가 과거에 일부다처제를 경험했었고, 아직도 이 사회 중 40% 정도의 사회가 일부다처제를 유지하고 있으며, 나머지 의 절반 정도의 사회는 때때로 일부다처제의 관습을 갖고 있다. 일부 다처제 관습은 때때로 수렵과 채집경제 사회에서 나타난다. 일부일처 제 사회는 모두 137 개로 전체의 약 13% 에 달하며, 일처다부제 사회 는 모두 4 개의 사회로 전체의 1.5% 정도일 뿐이다. 남성과 여성의 차이를 거론할 때 앞으로 우리가 집중해서 연구를 전행해야 할 과제는 아래와 같은 문제를 야기시키는 근본 원인을 밝 히는 데에 있다; 1. 왜 남성이 여성보다 더 공격성을 지니는가? 2. 대입 학력고사를 보면 남학생의 성적이 여학생보다 더 높은데 그 까닭은 무엇일까? 3. 오늘날 사회문제로 등장하는 혼음은 여성보다 남성에게서 더 높 은 비율로 나타나는 까닭은 무엇 때문일까?
4. 일반적으로 모든 사회에서는 왜 남창보다 창녀가 더 많은가? 5. 왜 모든 사람이 성 Sex 에 홍미있어하는기.? 6. 왜 남성이 절대로 포르노 산업을 지탱하는 데 큰 역할을 하는· 가? 7. 왜 형무소의 죄인들은 처음에 두옥될 때보다 남성호르몬 Tes t os t erone 의 양이 증가하는가? 8. 사람이 저지르는 범죄는 사회의 영향 때문일까, 아니면 범죄를 유발시키는 유전인자가 있기 때문일까? 9. 사람이 가지는 느낌 Fee li n g s 은 무엇이며, 왜 우리는 느낌을 가지 고있는가? 끝으로 우리가 매일 대하는 학생들의 행위를 하나 살펴보자. 강의 시간에 교실에 들어온 학생들은 대개가 언제나 같은 의자에 앉는다. 이때 여학생은 대개 앞자리에 앉고 남학생은 주로 뒷자리에 앉는다. 이들은 자기의 이름이 어떤 의자에 특별히 새겨져 있기 때문에 같은 자리를 매번 찾아갈까? 아니면 그들은 어려서부터 정해전 의자에 앉 도록 교육을 받았기 때문일까? 혹은 본능적으로 같은 자리를 택하도 록 하는 유전인자의 역할 때문일까? 아니면 생리상 환경이 학생들을 편안하게 만드는 무엇이 있기 때문일까? 우리는 사람의 몸에서 일어니는· 신전대사와 인체 내부에서 생겨나 는 호르몬은 단백질을 구성하는 유전자 코드에 의해 조절된다는 사실 을 알고 있다. 유전자 코드는 사람의 행동에 영향을 미친다는 사실 또한 우리는 알고 있다. 그렇다면 우리는 유전자 코드를 조절하여 위 와 같은 문제들을 해결할 수가 있지 않을까? 위에서 제기된 질문들은 사람의 구성과 행동을 조철하는 유전인자 롤 규명하고자 하는 인류의 야심찬 연구인 게놈 계획이 순조로이 진 행된다면 이에 관한 해답을 얻을 수 있다.
사회생물학의 소개 1975 년 미국 하버드 대학의 동물학자인 윌슨 E. 0 W il son 이 『 사회 생물학 : 그 새로운 이론 Soc i ob i olog y The New Sy n t hes i s .!l 이라는 제목 의 저서를 출판하였다. 이 책의 내용 중 90% 는 동물의 행동에 관한 연구를 요약한 것으로 이 부분은 세계에서 격찬을 받았다. 그러나 그 의 저서 중 나머지 10% 는 사람에 관한 것인데 이 부분은 많은 학자 둘로부터 비난과 함께 많은 논쟁을 불러일으켰다. 사람에 관한 이 부 분의 연구는 윌슨에게 인종주의자, 남녀차별주의자, 엘리트 양성주의 자, 생물학 결정론자 그리고 환원지지자(과학에서 복잡한 이론 따위를 하나의 통일된 원리로 단일화하는 사람을 말함)라는 명칭을 가져다 주 었다. 대다수의 사회과학자들이 윌슨의 연구에 반대한 보다 중요한 이유는 동물학자가 그들의 분야에 침입해 온 것을 환영하지 않았기 때문이다. 사회과학지들에 따르면 사람의 행위는 어떤 강요 없이 수없이 많은 문화 방법으로 배워서 얻는다고 한다. 이에 반해 사회생물학은 동물 과 사람에 있어 나타나는 모든 형태의 행위에 영향을 미치는 것을 생 물학상 요인(유전)으로 파악한다. 윌슨은 전화의 이론에 근거해서 동 물과 사람의 행위에 관한 어떤 면을 체계있게 설명하고자 하였다. 다 시 말해 사회생물학은 동물집단에서 관찰되는 행위들을 설명할 수 있 는 일반 법칙을 발전시켜 이 법칙을 인간에게 적응시키고자 한다. 이 런 윌슨의 연구는 생물학자들과 사회과학자들에게 동물과 사람의 행 위에 대한 연구를 전행하게끔 커다란 영향을 주었으며, 동시에 사회 과학자들로부터 많은 비난을 불러일으켰다.
생물의 집단에서 관찰되는 행위들 윌슨은 집단사회의 진화를 가장 찰 보여주는 동물들로 곤충을 꼽았 다. 곤충의 세계는 사람들의 이해를 넘어 자기희생과 협동의 행위에 근거를 두고 있다. 이들 곤충의 사회는 매우 복잡하나 개개인의 행위 는 상대적으로 단순하다. 예로 레몬이나 오렌지 등의 감귤류 C it rus 나무에 살고 있는 위버 개미 Weaver An t들의 집단을 보면 각각의 개미들은 아주 단순하게 행동하며 환경에 대한 적응능력이 거의 없으나 집단 그 자체는 뛰어 난 적응능력이 있다. 이들 집단이 다론 나뭇가지로 옮겨가거나 또는 어느 곳으로 이동할 때 개미들은 자신들의 몸으로 다리를 만든다. 많 은 개미들이 잎사귀를 함께 운반하며, 이때 한 개미가 실크를 생산하 기 위해서 유충개미에게 자극을 주며 또한 잎사귀 가장자리에 실크망 울 짜기 위해서 유충개미를 조정한다. 이런 일들은 아주 고도의 작업 이라 할 수 있는데, 비록 이런 행위가 복잡하기는 하나 각각의 개미 돌은 기계갇이 행동한다. 그들은 자신들이 행하는 협동이란 행위를 알지 못하고 단지 그들이 갖고 있는 유전인자의 명령에 복종하는 것 이다(그립 10. 9). 한 무리의 개미가 밤을 지낼 때는 약 70 만 마리의 개미들이 함께 있다. 이들이 밤에 잠을 잘 때는 모두가 단단히 얽혀 있어 마치 거의 한 마리의 생물체와 같다. 이때 개미들은 개인이 아닌 집단의 관계를 이루고 있다고 할 수 있다. 이런 경우 집단을 구성하는 구성원 사이 에는 계속되는 전달이 있으나 집단 안에서 개인이라는 인식은 없다. 새벽이 되면 얽혀 있던 상태가 깨지고 개미들은 움직인다• 이렇게 자 신을 생각지 않고 집단 행위를 유발하는 유전인자는 특히 새끼를 못 낳는 개미들에게 어떻게 전달되는가? 개미들의 이런 행위에 대한 답은 여왕개미에서 찾아진다. 각 일개
미들은 여왕개미 유전인자의 일부분을 가지고 있기 때문에 여왕개미 와 일개미들은 아주 가까운 관계가 있다고 할 수 있다 . 결과적으로 집단을 돌보는 일개미들은 그들 자신의 유전인자를 증가시킨다.
 10. 9 오스트레일리아 불독개미의 땅속 집
10. 9 오스트레일리아 불독개미의 땅속 집
꿀벌도 그들의 세계에서 개미와 비슷한 관계를 가지고 있다. 개미 들은 그들의 생활주기의 과정에서 새로운 여왕개미가 필요할 때는 완 전히 자기를 버리고 남을 생각하는 마음과 행동인 자기회생이 조금 약해진다. 두 마리 내지 더 많은 새로운 여왕벌들이 그들의 집단 내 에서 나와 한 마리가 남을 때까지 싸우는 것이 한 본보기이다. 협동과 자기회생 그리고 결집력 등은 사회생활을 하는 동물들에게 서 나타나는 행위인데, 이런 종류의 행위는 단순한 동물보다는 복잡
한 동물일수록 약하게 나타난다. 키티웨이크라는 새는 암초 위에서 새끼를 키운다. 이 새들의 행위는 개미보다는 아주 복잡하지만 곤충 들의 사회생활에서 보이는 협동과 자기희생 그리고 결집력 등의 어떤 행위도 이들에게서는 나타나지 않는다. 그럼에도 불구하고 개미와는· 다르게 어른 키티웨이크 새는 살아 남아서 전화한다. 이때에 이웃에 있는 키티웨이크들은 서로간에 아무런 관계가 없다. 이에 반해서 이웃을 잘 도와주는 놈은 자기의 유전인지를· 자식에게 제대로 전하지 못하므로 남을 잘 도와주는 행위의 특칭이 다음 세대 로 잘 전달되지 못한다. 물개의 경우에도 이들의 사회행위는 개인에 게 달려 있기 때문에 대개 공격성을 띤 수놈의 유전인자가 주로 디옴 세대로 전달된다. 젖먹이동물의 세계에서는 먼저 개인 사이에 결합이 일어나고 그 다 음에 집단 내에서 개인의 인식이 생겨난다. 한 예로 질라다 비비의 경우 수놈은 자기의 암놈모듬을 유지하고자 하며 또 모든 새끼들이 자기의 새끼임을 확인하고자 한다. 여러 수놈 중에 뛰어난 수놈은 자 기의 자리를 지킬 것이며, 이들 사이에서 일어나는 갈등은 적절한 행 위에 의해서 둔화된다. 실제로 많은 젖먹이동물의 경우 이들의 행위 는 곤충에서처럼 협동하기보다는· 질라다 비비처럼 갈등을 둔화시키게 끔고안되어 있다 . 사회생물학자들은 질라다 비비의 이런 행위를 가능·한 한 많은 새끼 를 만들어 내려는 생물학상의 전략으로 인정하고 있다. 우리는 이렇 게 비비에 적용되는 생각을 사람에게 적용할 수 있다. 그런데 문제는 사람을 다른 동물들과 같은 방법으로 다루는 것이 과연 타당한가 하 는 데에 있다. 과연 사람의 부모는 가능한 한 많은 유전인자를 다음 세대에게 전하려 한다고 인정할 수 있을까? 동물이 자식을 생산하여 키우는 데는 크게 두 가지 전략을 사용한 다. 하나는 가능한 한 많은 자식을 생산하여 그 중 얼마가 살아 남아
어른이 되기를 바라는 것이며, 또 다른 전략으로는 적은 수의 새끼를 낳고 이들이 살아 남게 하기 위해서 많은 투자롤 하여 돌보는 방법 01 다. 사람의 경우 자식을 생산하여 키우는 데는 한 가지 전략에 한정되 어 있지 않다. 인간은 영양의 공급이 충분하지 않은 지역에서는 일부 다처제를 지니고 있는데, 이는 한 여자로 하여금 자식을 적게 갖게 하는 역할을 한다. 그렇게 함으로써 그들의 생존은 보다 확실하게 된 다. 한편 일처다부제인 사회에서는 두 형제가 부인 하나를 공유하는 데, 이때 모든 아이들의 유전인자는 양쪽(아버지가 누구인지 상관없 이) 남자로부터 물려받는다. 남아메리카에서 야나마토 원주민을 연구하는 한 인류학자는 이들의 미 을들은 가까운 가족들끼리로 이루어져 있음울 알았다. 그들은 자원 이 풍부한 처녀림에서 살고 있었는데, 마울 하나의 규모는 매우 작 아 주민은 120 명이 넘지 않으며 모든 마울은 본래 하나의 마울로부터 갈라져 나왔음을 알았다. 야나마토 부족은 다른 사람에 대해 공격성 행위를 보여주는데 자기 들과 혈연관계가 없는 사람들에게 더욱 그러하다. 그리고 마울은 그 둘이 충분히 커졌다고 생각되면 갈라져 새로운 마을을 만들며, 이때 이들이 지녔던 혈연관계 또한 깨져버린다. 야나마토 부족들에게 있어 서 선물의 교환은 다른 부락들간에 협동을 가져올 수 있다. 사회생물학자에 의하면 인간은 자신의 유전인자를 공유하고 있는 사람만을 돕고자 하는 경향을 지니고 있는데, 이런 경향은 야나마토 부족들에게서 보이는 선물교환 같은 사회활동을 통해서 극복할 수 있 다고 믿는다. 또한 야나마토 사회는 오랫동안의 진화를 통해 이루어 전 사회들과 같은 조건을 가지고 있으므로 이들에 대한 자세한 연구 롤 통해서 우리는 우리 자신들의 행위에 관한 생물상의 근거를 더 명 확히 그려낼 수 있다.
윌슨의 연구에 따르면 남성과 여성 사이에서의 노동의 분화는 유전 요인에 의해 일어난다고 한다. 예를 들어 사냥에서 필요한 협동을 위 한 유전인자는 자식을 생산하는 데 이로움을 갖고 있어 이 인자는 집 단에 퍼진다고 한다. 사회생물학자들은 남성과 여성이 다르게 행동하 는 것은 그들의 행위를 일으키는 유전인자가 각기 다르기 때문이라고 주장하고 있다. 그러나 이런 사회생물학지들의 주장에 대해 많은 비 판이 제기되고 있다 . 비판자들은 어느 행위도 사회생물학자들이 주장 하는 유전요인에 의해서 일어난다는 증거가 없다고 지적하고 . 있다. 인간은 수백만 년 동안 작은 무리를 이루고 살아왔다. 만일 무리에 있는 어떤 사람들이 서로 짝을 구하지 못하면 그들의 유전인자는· 다 음 세대로 전달되지 않을 것이다. 이렇게 부모의 특칭은 세포의 염색 체 안에 있는 유전인자라는 유전물질 안에 저장되어 자식에게로 전해 진다. 유전인자의 기본구조는 뉴클레오티드로 불리며 나선형으로 나 타나며, 이때 각각의 염색체는 수많은 유전인자를 가지고 있다. 그러 나 어떤 유전인자 하나가 직접적으로 인간의 행위에 영향을 주지는 못한다. 윌슨이 지적하듯이 하나의 유전자가 한 행위에 책임이 있다 는 일부 사회생물학자둘의 주장은 잘못된 생각으로, 오히려 많은 유 전자들이 서로 엉키어 각 행위의 범주에 영향을 미친다. 이 유전자들 은 하나의 행위를 만드는 것이 아니라 가능한 범위를 짜는 것이다. 따라서 개인은 환경 자체가 변할 때 그 변한 환경에 다른 방법으로 반응할수 있다. 그러나 문제는 이런 내용을 과학적으로 증명하기가 불가능하다는 점이다. 비판자들에 따르면 사회생물학자들은 그둘 자신의 관념에 의 해 인간의 행위를 잘못 이해하고 있다고 주장한다. 예로 개인들 사이 에서 보이는 공격성은 때때로 전쟁을 설명하는 데에 이용된다. 그러 나 실제로 전쟁이란 정치성 이유에서 일어나는 것이지, 개인이 지니 고 있는 공격성과는 하등 상관이 없다는 점이다. 전쟁에서 사람들이
싸우는 것은 어떤 이데올로기에 대한 신념이나 아니면 군대에 정집을 당했기 때문이지, 적군 개개인에 대한 적개심을 가지고 있기 때문은 아니다. 사회생물학자를 비판하는 학자들은 배워서 얻어진 행위의 전달방법 은 생의 시작만큼이나 중요하다고 말한다. 한 예로 지난 25 년간 뉴기 니아의 부족은 트랙터를 운전하고 비행기를 조종하며, 의사가 되고 국회를 만돌고 하는 것을 배웠다. 이들 중 어느 것도 유전인자 안에 있는 것이 아니라 배워서 알게 된 것이다. 그러나 사회생물학자들은 비록 행위에는 문화 요소들이 있지만 전 세계에 살고 있는 사람들의 행위에는 서로 공통되는 특성이 있으며 이에 대한 설명이 필요하다고 믿는다. 한 예로 이스라엘의 키브츠에 사는 사람들은 인간행위에 관해 놀랄 만한 면을 보여주고 있다. 키브 츠에서 대등한 조건을 가전 아이들끼리 모인 집단에서 아이들은 부모 와 떨어져서 자랐다. 아이들은 서로가 오빠와 누이들 같았으며 이렇 게 자라난 아이들은 비록 그들이 서로 인척이 아니라도 결코 서로 결 혼을하지 않는다. 모든 인간사회는 근친상간을 금기로 한다는 점을 일반으로 인류학 자들은 문화 측면에서 규명하고자 한다. 그러나 사회생물학자들은 키 브츠에서 자란 사람들은 대등한 집단에서 자란 사람과 결혼을 하지 않는다는 사실에 근거해서 근친 사이에 결혼을 피하고자 하는 유전 요인이 있다고 생각하였다. 실제로 가까운 인척 사이에 결혼을 기피 하는 것은 유전문제를 야기할 수 있는 근친교배를 막는 데 도움이 될 것이다. 조셉 쉐퍼 Jo sep h She ffe r 라는 이스라엘의 한 교수는 원래 키브츠 구성원의 다수는 주로 개인주의 교육을 받은 유럽 사회에서 이주해 왔다고 지적한다. 키브츠에서 이주민들은 본래가 매우 신앙성이 있기 도 하였지만, 함께 모여 배우기 시작하면서 불가지론자가 되었다고
한다. 이런 변화는 강력한 이념에 의해서 이루어졌다. 키브츠의 특칭 은 아이를 기르는 데 있어서 경험을 중요시했을 뿐만 아니라, 성에 따른 일의 구분이 없다는 데에 있다. 다시 말해 키브츠에서는 이론상 남 • 녀라는 성에 따른 노동의 구분이 없다. 그러나 여성들은 여전히 부엌이나 세탁장에서 일하며, 남자들은 들판에서 일하거나 경영 또는 경찰업무 같은 일들을 주로 담당한다. 키브츠의 남 • 녀에서 관찰되는 이런 행위들은 유전상 근거에 바탕을 두고 있는 것일까, 아니면 대다 수의 사람들이 믿고 있는 것처럼 단지 강제성의 사회규범 때문일까 하는 의문이 제기된다. 심지어 여자가 남자들과 함께 일하는 곳에서 도 여자들은 낮에 아이들을 방문할 권리를 위해 투쟁했으며, 또한 밤 에 자식들과 떨어져 있는 것에 불만을 터뜨렸다. 쉐퍼는 어떤 강력한 이념에 의한 효과도 아직은 우리들의 유전인자 의 효과만큼 강력하지 못하다고 주장한다. 그러나 인간은 자신의 운 명을 변하게 할 수는 있지만 한계가 있음울 알 수 있다. 키브츠 사회 는 사회, 정치 그리고 생물학의 측면에서 볼 때 일반 사회와는 달리 제한된 사회이다. 그러나 키브츠가 아닌 일반 사회구조를 사회, 정치 그리고 생물학인 측면으로 따로따로 나누어 본다는 것은 불가능하다. 사회생물학 연구의 문제와 전망 윌슨 등은 사람의 행위를 곤충의 행위를 통해 살펴보고자 하였다. 그러나 인간은 곤충과는 다르게 사회 • 윤리의 관점에 근거해서 자신 둘의 행동을 선택하는 자유를 가지고 있다. 그러나 윌슨과 그에 동조 하는 학자둘은 인간으로부터 그들만이 누리고 있는 이런 선택의 자유 라는 특성을 빼앗아갔다. 따라서 인간을 생물학 측면에서 분석하는 학자들은 인간의 윤리 면을 강조하는 철학자들에게 인간 개개인들이 보여주는 특별한 행동에 관해서 그런 행동들이 왜 일어나는지, 또 그
행동들을 어떻게 생각하는지를 설명해야 할 과제를 가진다. The Selfis h Gene 이라는 책의 저자인 리처드 도킨스 R i chard Daw- k i ns 는 복잡한 원리를 하나의 통일된 원리로 단일화하기를 좋아하는 사회생물학자로, 유전인자란 단지 〈 복제를 조절하는 유전 요소들 〉 이 라고 기술하였다. 그에 따르면 지금 이들 유전인자는 인간의 몸 안에 있으며, 이들이 인간을 만들었고, 인간은 유전인자의 생존을 위한 기 계일 뿐이라고 지적한다. 그리고 성이란 유전인자가 한 세대에서 다 른 세대로 전해지는 수단일 뿐이라고 한다. 지금까지 위에서 거론된 내용들은 우리들에게 인간의 행위를 전화 라는 입장에서 살펴보아야 한다는 점을 암시하고 있다. 그러나 인간 을 오직 생물학적인 측면에서 보는 걸 비판하는 측에서 두려워하는 것은 인간이 저지르는 나쁜 행위의 근거는 유전 요인에 기인하기 때 문에, 따라서 이를 방지하고 근철하기 위한 사회제도와 노력은 소용 없다는 결론을 내리지나 않을까 하는 데 있다 . 오늘날 발달한 유전학의 연구 결과 우리는 어떤 유전성 질환들을 치료할 수 있다. 그러나 인간의 어떤 유전성 질환을 완전히 치료하기 위해서는 인간의 유전 특성을 완전히 이해한 후에야 치료가 바람직하 다 . 발달한 유전학의 지식은 인간으로 하여금 그들이 싫어하는 자신 들의 유전인자들을 제거할 수 있는 시기가 곧 현실로 다가오고 있다 는 사실을 알려주고 있다. 그러나 보다 중요한 것은 인간이 자신들의 유전인자들에 관해 충분히 이해한 후에야만 바람직하다는 점이다•
체질인류학의 오늘과 내일 인류학은 인문과학과 자연과학 양쪽을 포괄하고 있는 학문이다. 과 학은
에게 더 많은 지능 자극을 주어왔다. 그러나 오늘날 자연과학의 발달은 인간에게 더 이상 이 분야에 대 한 이해 없이는 인간의 과거와 현재 그리고 미래에 대해 말할 수 없 다는 사실을 일깨워주고 있다. 특히 유전학이나 분자생물학 둥의 발 달은 우리가 아직 경험하지 못했던 방법으로 인류의 전화에 영향을 끼치고 있다. 이 분야의 연구성과는 우리가 겪었던 고통과 고뇌를 누 그러뜨리기도 하지만 또한 여기서 얻은 지식이 잘못 이용될 수도 있 다는 점을 인식해야 한다. 이제 우리는 유전학 등의 연구성과에 따라 파생되는, 우리 사회가 지금까지 경험하지 못했던 여러 문제들에 대 해 윤리상의 결정을 위한 사회의 가치기준을 세워야 한다. 우리가 인간을 다룰 때 그 대상이 생물학 또는 사회의 문제이든 간 에, 무엇보다도 머릿속에 두어야 할 점은 인간이란 존재는 생물학 특 성을 배제한 문화의 산물이라거나 아니면 문화 능력은 무시한 채 동 물 특성만을 지닌 존재로 이해되어서는 안 된다는 것이다. 오히려 인 간이란 존재는 생물학 특성과 문화 특성을 함께 지닌 복합된 산물이 란점을기억해야한다.
 [부록]
[부록]
이 책에서 인용된 중요 외래 낱말들 제 1 장 체질인류학의 전망 유럽 중심주의 또는 유럽 우월주의 : Et hn ocentr i s m or Cultu r al Self cen - ter edness 전화 : Evoluti on 종합적 방법 론 : Multid isc ip li n a ry Meth o d 인류의 진화 : Human Evoluti on 영 장류학 : Prim ato l og y
영 장류 진화 : Prim ate Evoluti on 고인류학 : Paleoanth r op o log y / Human Paleonto l og y 인류의 적 응성 : Human Adap tab il ity 사회생물학 : Socio b io l og y 작은 진화 : Mi cr oevoluti on 큰 진화 : Macroevoluti on 인류유전학 : Human Geneti cs 집단유전학 : Pop u lati on Geneti cs 인류의 다양성 : Human Dive rsit y 현장방법 : Fie l d Tech niqu es 인종전형성 : Racia l St er eoty pe 제 2 장 진화란 무엇인가 자연체계 : Sy st e m a Natu r ae(Sy s te m of Natu r e)전화설 : Transfo r m ism 획득형질 : The Acq uire d Characte r is t i cs 철멸 : Exti nc ti on 카타스트로피즘 : Cata s tr o p h is m 균일설 : Unif orm i ter ia nism 생 존경 쟁 : Str u g gle of Ex ist e n ce 자연선택 : Natu ral Selecti on 서로 다른 재생산 : Di ffere nt Rep ro ducti on 전화점 진론 : Phy le ti c Gradualis m 변이 : Varia t i on 돌연변이 : Muta t i on 유전자 표류 : Geneti c Drif t 유전자 이동 : Gene Flow 급격한 변화 : Salta t i on Chang e 계 단식 평 형설 : Punctu ate d Eq u il ibr iu m 다른 지 역 분화 : Allop a tr i c Sp ec ia t i on 전체 유전특칭 : Gene Pool 정통 그리스도교인 : Chris t i an Fundermenta l i st 정통 그리스도교 : Chris t i an Fundermenta l i sm 우주론 : Cosmolog y 제 3 장 생명의바탕:유전과진화 유전 : Heri tab il ity 적 응 : Fit tne ss 염 색 체 : Chromosome 핵산 : DNA(Deox yribo nucleic acid ) 생 명 의 날 : The Thread of Li fe 단백 질 합성 : Prote i n Sy nthe sis
구조단백 질 : Str u ctu r al Prote i n 효소단백 질 : Enzym e Prote i n 뉴클레오티드 : Nucleoti de 리보솜 : Ri bo some 코드 : Code 코돈 : Codon 트리플레트코드 : Trip le tc o de 프로카리오트 : Procaryo t e s 유카리오트 : Eucaryo te s 안마디 : Intr o n 사이마디 : Exon 인자 : Genome 유전자 재 결합 : Recombin a ti on of Gene 헌팅톤병 : Hunti ng ton Di se ase 과다 콜레스테롤 : Hy pe rchokeste r olem ia 인위 선택 : Arti ficia l Selecti on 갑은 질의 : Homozyg o us (동형 의 ) 다른 질의 : He t eroz yg ous( 이형의 ) 좌위 (유전자자리 ) : Locus( 복수는 Loci) 표현형 : Phy no ty pe 유전형 : Genoty pe 혈액체계 : Blood Sy s te m 게 발 : Lobste r Foot 근육영 양실조 : Chondody st r o p h y 색소결핍 : Albin ism 산업에 따른 검은색화 : Industr i a l Melan ism 분가유전 : Founder Ef fect 우유 소화 : Mi lk Consump tion 평 행다형현상 : Balanced Polym o rph ism
주머니섬유증 : Cy st i c Fib r osis 낫꼴(=경상)적혈구 빈혈증 : Sic k le-Cell Anem ia 조철유전자 : Reg u rato r y Gene 구조유전자 : Str u ctu r af Gene 락타제 : Lact as e 락토제 : Lacto s e 공우성 : Codom ina nt 스포로자이트 : Sp o rozit e 메로자이트 : Merozit e 스위던 농업(화전농업) : Sw idd en Ag ri c u ltu re(= Slash and Burn Ag ri ~ cultu re) 계통학 : Sy st e m ati cs 분류학 : Taxonomy 린네 또는 이중 이름 명명법 : Lin n ean or Bin o mi al Nomenclatu r e 등 결동물민척색동물문) : Chordata 고종 : Paleospe c i es 분화 : Sp ec ia t i on 본원화 : Prim itive 서 행 전화 : Anage n esis 파행진화 : Cladoge n esis 같은 지역 분화 : Sy mpa tr i c Sp ec ia t i on 짝짓기 전 : Premati ng 짝전 뒤 : Postm ati ng 퇴적의 법칙 : Prin c iple of Sup e rpo sit ion 화석 연속의 법 칙 : Prin c ip le of Faunal Successio n 암석 층위 학 : Lit ho str a ti grap h y 분열자국 : Fis s io n Track 가열발광 : Thermolum ine scence 전자 회전반옹 : Electr o n Sp in Resonance
고지 자기 : Paleomagn e ti sm 부석 : Pum ice 계 통분화그립 : Cladogr a m 전화계 통족보 : Phy lo g en eti c Tree 상동관계 : Homolog y 유사관계 : Analog y 주워 먹 기짐승 : Scaveng er 동물화석 학 : Tap h onomy 제 4 장 영장류 프로시미안 : Prosim i an 앤스로포이드 : Anth r op o id 편편한 손톱/발톱 : Nail 뼈로 된 눈굴레 : Posto rbit al Bar 여우원숭이 : Lemur 나무 위 생 활 가설 : The Arboreal Theory 사냥을 위한 시각발달 가설 : The Vi du al Predati on Theory 나무뒤 쥐 : Tree Shrew 본고장 : Habit at 삶터 : Nic h e 분포구역 : Rang e 한낮의 활동영 역 : Day Rang e 주거활동영 역 : Home Range 지 역 : Territ or y 중십 영 역 : Core Area 대비성장측정학 : Allometr y 식충을 먹는 : Insecti vo ry 과일을 먹는 : Frug ivo ry
잎사귀를 먹는 : Foliv o ry 늘보원숭이 : Loris 긴팔원숭이 : Gi bb on 나무가지타기 : Brachia t o r 나무 위에서 네 발로 다니기 : Arboreal Qu adrup e d 땅바닥에서 네 발로 다니기 : Terrestr i a l Qu adrup ed 바깥 손마디 땅 짚고 걷기 : Knuckle Walk ing 두발걷기 : Bip e dal Locomoti on 공간분포 : Sp a ti al Dist r i b u ti on 무리구성 : Group Comp o sit ion 암수짝 : Consort Pa ir 떼 : Cong er y 암수 섞 인 무리 : Troop s 숫왕암무리 : Harem 의쪽무리: Band 흰개미 : Term ite 안경 원숭이 : Tarsid 쥐여우원숭이 : Mouse Lemur 일반특칭 : Grade 가지 치 기 : Clad. ist i cs 뒤쥐 : Sorex 난쟁이 여우원숭이 .: Dwarf Lemur 알락여우원숭이 : Ri ng - t a il ed Lemur 붉은 여우원숭이 : Brown Lemur 머리빗이발 : Tooth Comb 바로서서 뛰어오르는 걷기 : Verti ca l Cli ng and Leap ing Locomoti on 늘보원숭이 : Loris 스트랩시라이니 : S t re p s i r hini(프로시미에서 안경원숭이는 제의) 하프로라이니 : Ha p lor hini(안경원숭이를 포함한 영장류)
바다 퇴적물 : Deep - S ea Cores 퓨가토리 언덕 : Pug at o r y Hi ll 이 빨틈새 : Di as te m a 짝굽동물 : Ar ti oda ty lar( 우제류) 훌굽동물 : Per i ssodac ty lar( 기제류) 아래 턱 이 음새 : Mand ibu lar Sy m p h y si s 굳은 엉 덩 이 판 : Isch ial Callosit ies 비 비 원숭이 : Baboon 긴코원숭이 : Proboscis Monkey 엘파융 저지대 : El Fayu m Dep r essio n 칼날주머 니고양이 : Saber Tooth e d Masup ial Cat 회랑 : Land Brid g e 제 5 장 뼈대와 이빨 얼개 ( =구조) : Str u ctu r e 하버시안 체계 : Haversia n Sy st e m 하버시안관 : Haversia n canals 갑옷겉뼈대 : Exoskeleto n 몸안뼈대 : Endoskeleto n 등뼈대 : Ax ial Skeleto n 갈비뼈통 : Ri b Cage 옆머리힘살 : Temp or al is Muscle 턱모서 리 힘 살 : Mussete r Muscle 턱 안힘 살 : Pt er ig o id Muscle 앞니 : Incis o r 송곳니 : Canin e 옆니 : Premolar 어금니 : Molar
서로 다른 모양 이 빨 : Hete r odont 이 빨툴 : Denta l Formula 도드리 : Cusp 덧띠 : Cin g ul um 세 도드리 이 론 : The Trit ub ercular Theory 사기 질 능선 : Enamel Crest 팬 홈 : Groove / Fis s ure 이 빨 앞 • 안쪽 : Mesia l - L in g ua l 이빨 앞 • 바깥쪽 : Mesilt l-B uccal(or Labia l ) 해부학상 자세 : Anato m ica l Posit ion 볼록점 : Tubercle 불룩점 : Tuberosit y 솟 : Trochante r 솟기 : Process 힘 살띠 ( 인대 ) : Lig a ment 두터운 덧(건) : Tendon 머리 : Cran ium : Cran ial Bone : Skull( 아래턱울 포함) 머리뼈 : Ossa cr anii(아래턱을 제외한 머리 부위) 몸체뼈 : Postc r an ial Bone 이음새 : Sutu ra 재 기접 : Landmark 재 기 기 준선 : Fran kfor t Plane 사람뼈재 기 : Anth r op o metr y 주걱뼈마루 : Sp ine of Scapu la 주걱뼈 마루위 우복이 : Sup ra sp ino us Lin e of Scap u la 주걱뼈 마루밀 우묵이 : In fras p ino us Lin e of Scap u la 앞빈굼 : Sin u s Fronta lis 엉덩뼈구멍 : Foramen Obtu ratw n 엉덩뼈굼 : Aceta b ulum
부삽 모양 이 빨 : Shovel- s hap e d Teeth 못 모양 이 빨 : Peg - sh ap ed Teeth 이 빨 속 빈 공간 : Pulp Cavit y 사기질 너더리와 흑 : Enamel Ext en ti on and Peals 등고선분포 : Cl ina l Di st r i b u ti on 콧굼 둔덕 : Nasal Sil l 수칙 인 ( 얼굴) : Orgn a th o us (fac e) 뒤어나온(얼굴) : Prog n ath o us(fa c e) 코밑 의 얕은 홈 : Nasal Gutt er in g 이빨 맞닿음새 : Occulusio n 머리뼈 재기점 gn( Gnath ion • 턱밀가운점) pg( Pog on ion • 턱불룩점 ) idi ( l nf rad enta l e • 아래잇몸가운접) inc (Incis i o n ) ids (Alveolare • 위 잇몸가운점 ) pr( Prosth ion • 위턱앞점) ns(Nasosp ina le • 코밀가운점 ) n(Nasio n • 코맞점 ) g(G labella • 섭사이 ) b(Bregm a • 정수리점 ) v(Verte x • 으뜸봉우리점 ) ap ( Ap ex • 소리 칸- 수리접 ) l(L ambda • 맞음목) op (O p ist h o cran ion • 뒷머리점) i(l ni on • 옹이점) o(Op ist h ion • 으뜸뒷접 ) ba(Basio n • 으뜸점 )
alv(Alveolon • 입천정뒤끝점 ) sta ( St ap h y lion • ) ol( O rale • ) eu(Euryo n • 옆불룩점) po (Porio n • 귓굼점 ) ms(Masto i d a le • 젖꼭지점 ) d(Dacron • 눈물뼈맞음목) la(Lacrim ale • 눈물뼈 갓점 ) mf(M axi llo Fronta l e • 앞머 리 -위 턱 접 ) al(Alare • 콧굼갓점 ) or(Orbit ale • 눈굼밀점 ) zy( Z y gion • 광대점 ) ec(Ecto c onch ion • 눈굼옆점 ) ecm(Ecto m olare • 위어금니갓점) pt(P t er io n • 관자놀이 ) enm(Endomolare • ) cdi ( Condy lion late r ale • 아래 턱 꼭지 점 ) go( Gon ion • 아래턱모서리점) ft(F ronto t e m p o rale • 옆머리뼈갓점 ) 제 6 장 인류조상의 발자취 큰 골짝 : The Ri ft Valley 동쪽 골짝 : Easte r n Valley 서쪽 골짝 : Weste r n Valley 시와리크 언덕 : Siw alk Hill 카르스트: Karst 송곳니 파임 : Ca nine Fossa 칸디르 : Cand ir 파사라 : Pasalar
포트 테란 : Fort Teman 루다반야 : Rudabany a 아테네 : At he ns 포트와 : Potw ar 복제 : Transcrip tion 변형 : Translati on 제 7 장 인류의 진화 I : 오스트랄로피 테쿠스 타웅, 케이프 주 : Taung, Cape Provin c e 아파 분지 : Afar Basin 하다 : Hadar 세 렌게 티 평 원 : Serenge t i •P lai n 라에 톨리 : Laeto l i 머리뼈굼 : Foramen Magn u m 식 인풍습 : Ca nniba lism 석회암 엉킨 바위 : Breccia 울도완 돌문화 : Oldowan Sto n e Industr y 짧은꼬리원숭이 : Macaqu e 제 8 장 인류의 진화 11 : 호모의 출현 눈뒤줄임 : Posto rbit al Constr i c t i on 올드바이 골짝 : Olduva i Gorge 오모: Omo 몸돌 연모 : Core Tool 격 지 연모 : Flake Tool 찍 개 : Chopp er Core Tool 자른 자국 : Cutm ark
주거 지 : Liv i n g Floor 음식 분배 행 위 가설 : Food Sharin g Theory 인류행위 중심지 이론 : Centr a l Place Forag ing Theory 선 주거 지 : Prehabit ati on 인류계 통학 : Hwnan Phy lo g en y 잃어버린 사슬 : Mi ss in g Lin k 용골 : Drago n Bone 용골산 : Drago n Bone Hi ll 터 니파인 : Terfin e 살레 : Sale 나리오코톰 : Nario k ot om 옛술기사람 : Archaic Homo sap ien s 스타인하임 : Ste i n h eim 베르테스쾰러스 : Verte s zollos 빌칭스레벤 : Bil zin g sl eben 아라고 : Arago 페트라로나 : Petr a lona 상기란 : Sang ira n 모조케르토 : Modjo k erto 삼봉마찬 : Sambung ma chan 눈두덩 : Sup r aorbit al Torus 옹이 점 : Occ ipital Torus 전화의 방법과 속도 : Mode and Temp o of Evoluti on 크기의 범위 : Rang e of Varia t i on 발달한 형 태의 곧선사람 : Develop ed Homo erectu s 임계온도 : Crit ical Tem p era t ure( 인류가 의복을 입지 않고 살 수 있는 온 도) 찍개문화 : Chop pe r Core Industr y 주먹도끼 : Hand Axe
아슐리안 문화 : Achulia n Industr y 발달한 울도완 문화 : Develop ed Oldowan Industr y 우베이디야 : Ubeid iya 자르개 : Cleaver 대나무문화 : Bamboo Industr y 도파민 : Dop amine 브로카 부위 (령 ) : Broca's Area 베르니크 부위 (령 ) : Werni ke 's Area 뒤 셀도르프 : Di isse ldorf 네 안더 골짝 : Neander Valley 스피 : Spy 도르도뉴 : Dordog ne 라 샤펠 오 솅 : La Chape lle-Aux-Sa int s 르 무스티 에 : Le Mousti er 라 펠라제 : La Ferrassie 라 퀴나 : La Quina 크로마뇽 : Crog Magn o n 오각형 : Penta g on 계란형 : Oval 턱 가운데 부위 : Ch in 바이첼 빙하기 : Weis c hel Glacia t i on 폰테쉐바드 : Fonte c hevade 대표되는 네안데르탈사람 . : Classic Neanderth a l (또는 Neander t라로 사용; Trik a us & Sh ipm an, 1993) 보더 동굴 : Boder Cave 게바라 동굴 : Gebara Cave 무스데 리 앙 : Mouste r ia n 페리고르드 : Perig o rd
긁개 : Sid e Scrap e r 태 식 타쉬 : Tesik Tash 샤니다르 : Sha nida r 크라피나 : Krap ina 크로쿠타 : Crocu t a( 하이에나 속의 한 종) 주거 지 : Base Camp 일하는 장소 : Worki ng Camp 제 9 장 인류의 진화 m: 슬기사람 Homo sap ien s : 현생인류의 시작 레 저 지 : Les Ey z ie s 뒷머리뼈-젖꼭지뼈 마루 : Occ ipito- Masto i d Crest 아래 셋째어금니와 턱가지 사이의 공간 : Retr o molar Sulcus 이복파임 (二復常) : Diga tr i c Fossa 뒷머리뼈홈 : Occip ital Groove 제불 카프체 : Jab el Qafze h 풀로리스베드 : Floris b ad 아이시 호수 : Lake Ey as i 싱가 : Sin g a 칸제라 : Kanje r a 스쿨 동굴 : Skuhl Cave 터분 동굴 : Tabun Cave 케바라 : Kebara 아무드: Amud 상고안 : Sang oa n 파우레스미스 : Fauresm ith 펭스벵 : Pin c event 등을 죽인 자르개 : Backed Kn ife 등이 뭉뚝한 자르개 : Blunte d Backed Kn ife
월계수잎 모양 자르개 : Wi llow Leaf Blade 제 10 장 인류의 적응과 사회생물학 적 응 : Adap tat i on 생리상 조철 : Phy si o l og ica l Adju s tm ents 생 물학상 적 응 : Geneti c Adap tat i on 문화 적 응 : Cultu r al Adap tat i on 순응 : Accli m ati za ti on 순화 : Acclim ati on 갈등 : Str e ss
항온동물 : Homeoth e me An im al 몸떨림 : Hy po sia 혈관수축 : Vasoconstr i c t i on 혈관확장 : Vasodil at a ti on 일부다처제 : Polya n dry 남성호르몬 : Testo s te r one 위버개미 : Weaver Ant 자기 회 생 : Altr u ism찾아보 71
7 가슴등뼈 187 가운능선 289 가운면 196 갈비뼈 187 갈비뼈통 187 감귤류 나무 454 감비모기 93 감수분열 63 갑옷 겉뼈대 187 갓쪽 197 같은 지역 분화 103 개체변이 36 거북등날떼기 389 걷기방법 132, 171 검은 머리뼈 305, 308, 311 게놈 73, 116 게놈 계획 452 게이츠 239 게일즈 203 게일즈와 엘리엇 203 격 지 339, 422 결도드리 236 계단식 평형설 51, 106, 107, 359, 360 계통분화그림 112, 261 계통학 97
고고인류학 26 고고학자 26, 27 고릴라 163-167, 170, 173-177, 181, 249, 252, 253, 258, 260, 262, 264 331 고지자기 측정법 362 곤충먹는 영장류 133 곧선 곧선사람 364 곧선사람(직립원인) 270, 348, 348- 364, 367, 369, 371, 371-373, 397, 399, 401, 407, 409, 414, 415, 435 곧선 피데칸드쿠立~ 364 골간(뼈몸체 ) 186, 375 골내막 186 골단(뼈끝) 186 골막 186 공간분포 133 공우성 91, 123 과다 콜레스테롤 76 과일을 먹는 영장류 133 과일따기 가설 315, 316 과학성 창조론 55, 56 광대뼈 처짐 240 교배 및 교체설 404, 415, 419 구대륙 원숭이 152-154, 158, 161, 249, 251, 252, 317 구아닌 65
구조 단백질 67 구조 유전자 86 굴드, 스티븐 제이 51, 106, 107, 283 균일설 43, 46 그라베티 앙 426-4 28 근친상간 459 글루데우스 메디우스 175 글루테우스 미니머스 175-176 글리코즈 405 기간토피 테두스 252-254 기준선 203 긴뼈 197 긴팔원숭이 131, 135, 163, 164, 167, 282 긴팔원숭이과 136, 163, 164 길, G.W. 242 끊는 기능 192 L 나리오코톰 356, 357 나무뒤쥐 137 나무잎사귀 먹는 영장류 133 나무위 생활가설 124, 125 난자 62, 62, 405 남성 호르몬 452 납작뼈 197, 198
납작코원숭이 152, 155 낫꼴(경상)적혈구 빈혈증 86, 88, 90, 93, 94 낭동 400 낭동곧선사람 364 내슈슈충 339 네 안더 골짝 379, 392 네안데르탈사람 2)7, 270, 286, 345, 351, 377-392, 395, 397, 404, 405, 409, 412, 421, 422, 424, 425 네안데르탈 슬기사람 386 네안데르탈 조상 가설 385 노롱고로화산 278 눈굼 382 논 굼둘레뼈 123, 150 눈두덩 359, 378, 382 눈뒤줄임 329 뉴클레오티드 65, 67, 81, 458 느낌 (Feeli ng s) 452 늘보원숭이 136, 137, 141, 154 니메시스 항성 147 C 다레스 쏠탄 411 다른 지역 분화 103, 104 다른질 77 다우벤토니과 136 一 138, 140
다윈, 찰스 19, 28, 35, 40, 41, 43, 44, 48, 49, 51, 282- 28 3, 313, 314, 351, 359, 366 다윈, 에라스무스 38, 40 다지역 기원설 400, 401, 403, 419 다트, 레이몬드 282, 283, 286-289, 292, 295 단백질 합성 67, 68, 69 단일종 가설 337, 338 단일지역 기원설 400-404, 409, 418-420 달리 400 달베르그, A 235 닳음새 383 당 65 대고 떼기방법 422 대립유전자 77 대 립 형 질 78, 80, 82, 84, 86, 88, 89, 91 대비성장측정학 130 대표되는 무스테리앙 연모 390 대표적 표본 99 대표형태의 규칙 99 대협곡 249 덧닿기 238 데모그래피 319, 321, 323 도드리 190-192, 232, 234, 235 도르도뉴 379, 380, 395, 423, 430
도브잔스키, 데오도시우스 50 도순 찰스 285 도킨스, 리처드 461 도파민 375 돈 칼 423 돌날격지 421, 422 돌연변이 51, 81, 82, 88, 91 동물계 98 동물층위학 362 동북아시아 갈래(황인종) 239, 241 동쪽 협곡 249 동형집합체 89 두뇌 진화 287 두발걷기 269, 284, 308, 312, 314, 319, 323, 324 뒤셀 강 379 뒤셀도르프 379 뒤팔뼈 169, 194 뒷머리 이음새 198 듀엣 165 드리오피테쿠스 249, 250, 252 드보아, 유진 282, 351-353 드 뷔풍, 죠지 21, 38, 40 드브리,휴고 49 등고선 분포 239 등공동절문 98, 100 등을죽인 자르개 426 등이 뭉뚝한 찌르개 427
디세노데리아 274 딱딱한 열매 먹기 가설 315, 316 떼 134 뚜르개 423, 427 뚫는 기능 192 三 라마르크 38, 40, 49 라마피 데쿠스 252 , 254-2 56, 258- 260, 262, 269, 318 라마피테쿠스 판자브쿠스 라마피데쿠스위케리 라마피데쿠스 헝거리쿠스 라 샤펠 오 솅 379, 380, 392 라에톨리 271, 273, 277, 279-2 81, 283; 285, 305, 309, 311, 332, 363, 385, 399, 410, 411 라우셀 430 라이 엘, 찰스 19, 38, 42, 43 라이트, 서웰 50 라이트푸트, 존 37 라 퀴나/라 펠라제 무스떼리앙 연 모 390 라 펠라제 380, 382, 392 락타제 86-88 락토제 86-88 란텐 357, 358, 400
러브조이, 오웬 322, 325 레 제지 395 렉, 한스 296 로드만, 멕헨리 323, 325 로메퀴 304 로빈손, J. T 289, 290 로사이(로커스) 77 로타감 268 루 게릭 질병 73 루다반야 258 루돌프 사람 338 루돌프 호수 334 루소, J 39 루시 274, 280, 283, 308, 309, 336, 359, 360399 루싱가 섬 251 류지앙 415 르 무스티에 379 르발루와 422, 388 리보솜 65, 68 리키 루이스 251, 255, 282, 283, 286-2 89, 292, 295, 329, 330, 332, 336 리키 리처드 281, 304, 306, 308, 335, 342, 345, 356 리키 메리 273, 277, 278, 281, 296, 329, 341 리키, 조나단 329
린네, C 38, 39, 239 口 마르모세트 122 마사이족 279 마수피얼 125, 145 마우어 353 마이어, E. 358 마이오세 144, 249, 250, 252-25 4, 267- 2 69, 316, 317, 324 마줏정 204 마카판스가트 288, 290, 305 마틴, R 203 막달레 니 앙 427, 428 막스헤드럼 435 만달 415 말라리아 93 맞닿기 238 매력 4~0 맬서스 38, 43, 48 MN 혈액체계 머리 뒷두덩 359 머리등뼈 187 머리뼈와 몸체뼈대 이름 199-202 참조 머리뼈 이음새 198 머리뼈 재기 204
멍게라, 머카 277 메로조이트 93 메스티조스 448 면역학 115 모조케르토 358 모조케르토 피데칸드 쿠 立~ 364 몸떨림 437 몸돌 연모 339 못 모양 이빨 237 몽티가우디 431 무리구성 133 무스데리앙 421-426 무스데리앙 돌연모 389 문화인류학 24, 27 문화방법 443 문화 적응 436-43 9, 448, 449 뮈산 357 미나토가와 415 미들 아와쉬 273, 276, 305 미토콘드리아 405, 406 미토콘드리아 핵산 404, 405, 407, 409, 415 밀개 423, 426 믿닿기 238 t::I 바다퇴적 분석법 362
바똥 431 바스, W. M 239 바이덴라이히, 프란쯔 355, 356, 358 바이첼 빙하기 385, 395 바일로포돈트 도드리 158 바깥 손마디 땅 짚고 걷기 263 밖돌리기 176 밖으로 향한 발가락 251 반전화론자 53 받침뼈 닿는 면 227 받침뼈 윗둔덕 173, 175 발달한 곧선사람 365 발달한 울도완 연모 371 발로네 동굴 341 방주 모형설 401, 402 백악기 143-146, 148 백인종 239, 240 버클리 39 번, 헨리 346 베르그만 440 베르그만의 법칙 440-442 베르니크 부위 375, 377 베르데스쾰러스 357 베이칭 곧선사람 354, 364, 365 변이 48, 62, 78, 407 변형 257 보더 동굴 410, 411
보르드, 프랑소와 389, 390, 422 보르드, 손 루빌 422 보린, 비거 354 복수종 가설 338 복제 257 본고장 128 본원화 101 부삽 모양 이빨 237 부싯돌 427 부쉬베비 135, 142 부울, 마르셰 린 380-38 2 분류학 97 분포구역 128 불가지론자 459 불규칙 뼈 197, 198 불독개미 455 불룩이 383 붉은여우원승이 138 브레 인, C. K 2901 292, 294, 356 브로카 부위 376, 377 브루커, 에드워드 362 브룸, 로버트 288, 290 블랙, 데이비슨 354, 355 비교해부학 24 비글호 45 비너스 426 비비 159, 160, 179, 257, 261, 262, 286, 287
빅토리아피데쿠스 249, 316 빅토리아 호수 249, 254 빈포드, 루이스 345, 346, 373, 390 빌칭스레벤 357 빗장뼈 169, 176 뼈단위 185 뼈 • 이빨 • 뿔의 문화 290, 295 人 사기질 232 사냥을 위한 시각발달 가설 124, 125 사냥가설 292, 343, 345 사람의 머리와 얼굴재기 204 사람의 몸 재기 204 사람뼈대학 24 사람뼈 재기 204 사르투x., S 364 사바나 367 사이마디 70, 71 사이몬스, E. L 255, 259 사이 안 화산 279, 280 사이토신 65 사하비 268 사해 270, 271 사회 생물학 24, 27, 453 산업에 따른 겁은색화 85
산타 루카, A. P 364 살단하 400 살레 356 삶터 128 삼봉마찬 400, 358 상기란 358 상대연대측정 110 상동관계 114 상아질 232 색소결핍 82 생리상 조절 436-438 생물학상 적응 436 생물학상 종의 개념 99 생식세포 62, 63 생존경쟁 43 생태학 127, 128 샤니다르 392 샤크레몬, N. J. 362 샤텔페로니앙 425-42 7 샨명뚱(山頂洞) 414, 415 서로 다른 재생산 51, 95 서로 다론 집단 390 서행전화 103 서쪽협곡 249 석회질 232 선사고고학 26, 27 선 네안데르탈 가설 385 선 술기사람 가설 385
선 유인원 31 선 주거지 346 설치류 140, 149, 151 성별차이 311, 338 성성원숭이과 136, 163-165 세 도드리 이론 191 세렌게티 고원 271, 277, 279, 296, 339 세리치, V 256-260 세비드과 153, 155, 156 손마디 걷기 251 손쓴사람 270, 329-332, 334-339, 344, 345, 348, 359-361, 367, 371, 389, 435 솔로 강 351 솔로사람 404 솔로 피테칸트로프스 364 솔루트레앙 427 송곳니 123, 171, 172, 189 송곳니 파임 255 순다 해협 369, 370 순응 436, 449 순화 436 쉐텐삭, 오토. 354 쉐퍼 조셉 459, 460 쉬프만, 패트 346 슈지아요 415 술리서; 막스 354
스미스, 윌리암 109 스미스, F. H. 403 스미스, G. E 124, 125 스와르트크란스 288, 289, 291-293, 305, 356 스완스콤 357, 384, 385 스위던농업 94 스쿨 411-413 스타인하임 357, 384, 385 스데노, 월리암 108 스테르크폰데인 288, 305 스태빈스, 레드야드 50 스듀어트 229 스트렙시라이니 136, 142 스트링거, C. 403 스포로자이트 93 스피 379, 392 슬기사람 270, 308, 331, 332, 352, 357, 358, 359_, 365, 378, 382, 385, 393, 395-421, 426, 428, 431, 435 시각피질 377 시바피테쿠스 252, 253, 259, 316, 318 시난트로푸스 페키넨시스 355 시와리크 언덕 253, 255, 258 식 인풍습 291, 391, 392 신경세포 374-376
신경세포연접 374 신경 전달물질 374, 375 신경충동 374 신대륙원숭이 152-155, 157, 158, 248, 261, 262 신생대 144, 145 신화 51, 52 심프슨, 조지 게일로드 50 싱가 410, 411 쌍날찍개 341 。 아가지즈 L. 109 아다피스 150 아덴만 271 아도브 벽돌집 444 아드레이, R 291 아람버그, C 306 아래턱이음새 152 아래턱 꼭지 171 아무드 411, 412 아미노산 67-70 아라고 357 아슐리앙 문화 371 아슐리앙 연모 372 아슐리앙 전동 372 아슐리앙전통의무스테리앙연모
390 아스테로이드 146, 180 아시아 기원설 367 아이시 호수 410, 411 아이-아이원숭이 140 아이작, 글린 334, 341, 342-346 아종(亞種) 363 아치형 신경다발 377 아키바 만 270, 271 아데네 258 아데닌 65 아파 271, 277 아프리카의 기원 291 아프리카 기원설 367, 370 안경원숭이 136, 137, 142, 151, 154 안데스 고원 445 안마디 70, 71, 116, 257 앉음뼈 • 감춤뼈 가지 224 앉음뼈 큰 활흥 225, 227 알락여우원숭이 138 알렌 430 알렌의 법칙 441, 442 알타미라 428 알터플라노 446 알프스 산맥 369 앞니 123, 189 앞뒤선 194
앞머리 이음새 198 앞빈굼 223 앞 • 바깥쪽 192 앞 • 안쪽 192 앞팔뼈 169, 194 앤더슨, 존 가너 354 앤스로포이드 121, 127-128, 136, 142, 151, 152, 160, 161, 262 야곱, T 364 야냐마토 원주민 457 야오족 52 얀젠, 단 180 어금니 123, 189 어금니 속의 빈 공간 237 어셔, 제임스 37 언스트 마이어 50, 51 얼개 185, 189, 193, 231, 271, 373, 374 엉덩뼈 169, 173, 174 엉덩뼈 구멍 227 엉덩뼈 굼 175, 227 엉덩뼈 홈 225 엉 킨바위 292, 295 엉치뼈 맞자리 225 워커, 알렌 304 에티오피아 271, 273, 309, 399 엘드리지, 니일 51, 106, 107 엘드리카, A 237
엘 화융 248, 267 여우원숭이 123, 137-140, 151, 154 역사고고학 26, 27 연모사용 가설 316 열성 78, 82 염기 65 염색체 62, 65, 77, 81, 405, 458 영양실조 82 영장류학 23 옆니 123, 171, 189, 248, 249 옆머리 이음새 198 옆머리 힘살 187, 188 옛날종 100 옛술기사람 357, 363, 365, 403, 411, 414, 415 옛영장류 148 옛자바 로보스두스사람 364 옛자바사람 364 옛자바 상기란사람 364 오랑우탄 163, 165, 166, 176, 177, 181, 249, 255, 258, 260, 262, 282, 286, 331, 351 오리나시 앙 426, 427 오모 305, 332, 334, 385, 410, 411 오스데은 185 오스트랄로이드 239 오스트랄로피데쿠스 269, 270, 290, 296, 305, 306, 309, 315, 317,
435, 330-332, 335, 336, 339, 350, 367, 370 오스트랄로피테쿠스 속 269, 284 오스트랄로피 데쿠스 로보스두두스 270, 288-290, 298, 299, 304, 331 오스트랄로피데쿠스 보이세이 270, 295, 297, 298, 304, 305, 311, 329, 331, 334 오스트랄로피테쿠스 아파렌시스 269, 270, 280, 281, 283, 285, 305- 311, 314, 318, 322, 331, 348, 359, 360, 367, 370 오스트랄로피테쿠스 아프리카누스 269, 270, 286, 288-292, 295, i99 , 305, 308, 309, 311, 331, 336, 349 오스트랄로피테쿠스 에티오피쿠스 270, 298, 304, 305, 308, 309, 311, 331 50 번 유적지 343 오포섭 101 울도뇨렌가이 화산 279 올드바이 골짝 295, 296, 329, 331- 336, 339, 340, 341, 345, 346, 356, 371, 360 올드완 문화 339 올드완 연모 296, 339 울리고세 144, l60-162, 248, 267 올리버, G 229
옵다이크, N. D. 362 옹이접 359 의날찍개 341 요하네스버그 286 요한슨 도날드 273, 274, 276, 281, 283 용곡 415 용골 254 우드 버나드 338 우베이디야 371 우선의 규칙 99 우성 77, 82, 89 우월주의 22 우유소화 86 운동출력부위 377 원-영장류 128 원-유인원 316, 317, 320, 322, 324 원 호미니드 366 월러스, 알프레드 러셀 19, 35, 43, 351, 366 월포프, M. H 401 웨이크, 데이비드 107 위비 개미 454 위아래선 194 위엔모우 357 위트워터스랜드 286 윌슨, A. C. 260, 403, 405 윌슨, E. 0. 453, 458, 460
윗머리 이음새 198 유럽중심주의 22 유사관계 115 유사분열 63 유인원 23, 31 유인원 도살자 가설 315, 316 유인원 아목 270 유전상 적응 437 유전자 교환 모델 401-403 유전자의 빈도 79 유전자 이동 51, 81 유전자 재결합 71, 72 유전자 표류 51, 81, 82, 84 유전형 77, 81, 86, 91 유카리오트 70, 71 윤지안 357 음식분배행위 가설 342, 345 이복파임 399 이빨 맞닿음새 242 이빨를 189 이스라엘 404, 405 이신경 232 이오세 144, 149-151, 155, 160, 249, 267 이오앤人쿠立~ 286 이오앤人쿠 立人 도소니 285 이중 이름명명법 99 이집토피데쿠스 161, 248, 249
이틈새 171 이형집합체 88 인류계동학 348 인류기원 연구소 336 인류생물학 436 인류의 기원 282, 349 인류행위 중심지 이론 344 인구론 43, 48 인드리 원숭이 140 인류과 31, 270, 284, 308 인류속 269, 284, 308 인류의 다양성 23 인류 유전학 23 인산(염) 65 인위선택 76, 85 일부다처제 451, 457 일부일처제 451 잃어버린 사슬 349-351 임계온도 367 잇골 232 x: 자르개 372, 427 자르는 기능 192 자론 자국 392 자바 곧선사람 351-353 자연선택 35, 46, 48, 51, 61, 81,
84-86, 282, 385 자연체계 38, 97 자이고러스 산맥 369 작은골 376 작은 전화 23, 96 저우커우덴 354, 356, 357, 359, 361, 373, 400 적응 436 전기작용 374 전달 RNA(mRNA) 68, 69 전환 RNA(tR NA) 68, 69 절멸 41, 102 접합자 62, 63 정자 62, 63, 405 정중선 194 정찰 가설 315, 316 정통 그리스교주의 53 젖먹이동물강 98, 100 절대연대측정 11 조철유전자 257 졸리 클리포드 315 좁은코원숭이 152, 157 좁은코원숭이 하목 270 종의 기원 28, 46, 51, 62, 106, 282, 378 종의 분화 103 좌위들 78 주거활동영역 129
주걱뼈 169 조절유전자 86, 88 적자생존 95 주대오-크리스천 36 주머니섬유증 86, 88, 89 주머니쥐 • 101 주먹도끼 371, 372, 392 주워먹기 짐승 307, 341, 343, 435 줄리안 헉슬리 50 중기 풀라이스토세 370 중생대 143-145, 271, 395 중십영역 129 중추신경 375, 376 중합체 고리 반응법 73 쥐안경원숭이 130 쥐여우원숭이 131, 138, 139, 331 지각입력부위 377 지브롤터 369, 379 지질학 원론 42, 43, 46 지질학 원리 109 진잔트로푸스 298, 329 전즈 298, 329, 332 진화계통그립 112 전화계통족보 113 전화계통학 103 진화론 36, 49, 51, 53 진화 생물학 23, 36 진화설 40
전화점진론 51, 104, 105, 107, 360 질리다 비비 135, 315, 456 집단유전 79 집단유전학 23 짧은꼬리여우원숭이 136- 1 38, 140 짧은꼬리원숭이 122, 151, 153, 319 짧은꼬리원숭이과 153, 157 짧은뼈 197, 198 찌르개 426, 427 찍개 341 찍개문화 372, 371 * 창던지개 423, 430 창조론 51, 53, 54 천재지변 276 체로키족 52 체소완자 373 초인류과 31 촘스키, 놈 178 촛대형 모형설 401, 402 츠양 415 충판 186 층판뼈 186 침 팬지 249, 252, 253, 256-258, 260, 262, 263
구 카르스트 254, 292 카멜산 412 카모야 305 카보나이트 280 카스피 바다 369 카타스트로피즘 41, 42 카트밀, M 125, 126, 148 카트빙클, 빌헬름 296 카파치안 산맥 369 카포이드 239 카프체 411- 41 3, 404 칸, 레베카 405 칸제라 410 칸트 39 칼륨-아르곤 방법 111 칼리스리스드과 153, 155, 157 캐러벨리 도드리 236 캠벨, 버나드 363, 377 커디스, G 336 케바라 412 KBS 충 336 케이프 주 270 코돈 70 코밑에 얕은 홈 241 코카서스 산백 369 코팡스 334
콜로바엔원숭이 153, 158 콧굼 382 콧굼 둔덕 240 쿠비포라 332, 334, 343, 356, 389 쿤 239 퀘 닉 스발트, 폰 G. H. R. 254, 364 퀴비에 38, 41, 42 크라우스, B. S 236 크라피나 392 크로마뇽 381, 395, 397, 398, 421, 426, 431 크로쿠타 392 크롤, 엘렌 346 크롬드라이 288, 305 큰골 376 큰골 우선가설 283 큰긴팔원숭이 163-165 큰 진화 23, 95, 97 쿨라시에스 400, 404, 410, 4ll 클라크, 데스몬드 273, 276 클라크, 월프레드 르 그로스 290, 331, 358 클레버 한스 178 키브츠 459, 460 키티웨이크 456 킹 377
E 타분 411-413 타오로돈티즘 237, 238 타웅 269, 283, 286, 288, 291, 292, 305, 308 타이브, 마우리스 273 타이슨, 에드워드 28 타지역분화 51 타키두스 22 탄자니아 271, 273, 280, 309, 399 터니파인 356 턱모서리 힘살 187, 188, 297 턱안 힘살 188 데숴 타쉬 392 덱사 98 텍손 98 토바이아스, 필립 330 톱니날 무스데리앙 연모 390 되적의 법칙 108, 110 두르카나 호수 304, 332, 334, 338, 356, 357, 361 튀어나온 광대뼈 241 트로터, M 과 글래서, G. C 229 트라이곤 191 트라이보스페닉 192 트란스발 286, 288, 290, 356 트리닐 곧선사람 364
트리플리트코드 70 트링카우스, E. 403 특수화 101, 102 티민 65 고 파란드로푸스 로보스두스 289 파사라 258 파웰, 제리 54 파이퍼, 존 427 파킨스병 375 파행진화 103 팔레오세 143-146, 148-150, 155, 161, 249 패트 쉬프만 304 펀치 427 페리고르드 389 페트라로나 357, 365, 400 펜크, 알브레크트 362 펭스벵 424 평형다형현상 86, 88, 89 포츠, 리처드 346 포큐파인 254 포트 와 258 포트 데란 258 포프, 제프리 364 폰테쉬바드 384, 385
표본화석 307 표준화석 254 표현형 77, 81 퓨가토리우스 148 프로시미안 121, 126, 128-130, 133, 136-138, 140-143, 150, 151 프로스타일리드 236 프로카리오트 70 프로컨슬 249, 250, 267 프로컨슬 니안저 251 프로컨술 메이저 251 프로컨슬 아프리카누스 251 프로토플라즘 350 풀라스모디움 기생충 93 풀라스미드 71 풀라이스토세 144, 161, 253, 354, 362, 363, 368 풀라이오一플라이스토세 334, 339, 370, 367 풀라이오세 144, 269, 316, 368 플레시아다피스 149, 150, 267 풀로리스베드 410, 411 立 쿠 :§.., J. K. 379 피드백 이론 283, 290, 314, 320, 324 피레네 산맥 369 피데칸뜨쿠立~ 352, 353, 364 피데칸 E 쿠 立人 아라루스 350
피데칸드쿠立人― 에렉두스 352 피츠로이 45 피치, F. J 와 밀러, F. A 336 필빔 , D. R 255, 259 필트다운 285 一 287 굴 하다 271, 273, 274, 280, 281, 305, 309, 311, 363, 399 하디-와인버그 80, 81 하버시안 관 185, 186 하버시안 체계 185, 186 하웰, F. C. 334, 358 하웰스, w. w. 203, 358, 401 하이델베르크사람 353 하일로베이트 249 하프로라이니 136, 142 한낮의 활동영역 129 항온동물 437 해부학상 술기사람 385, 404 해부학상 자세 194 핵 68 핵산 (DNA) 65-67, 69-71, 76, 77, 96, 405 핵산 교배 115 허리등뼈 187 허벅지뼈 169, 173-176
헉슬리, 토마스 헨리 47, 48, 282 , 351 헌팅톤병 76 헤모글로빈 67, 90, 91 헤 켈 , 언스트 282, 350, 351, 352 헤두르아 프레르 동굴 429 헨슬로, J 45 현대진화론 50, 51 혈관 수축 438, 439 혈관 확장 438, 439 호모 370 호모 네안데르탈렌시스 377, 386 호모 사피엔스 네안데르탈렌시스 377 호모 하이델베르겐시스 354 호미노이드 31, 176, 177, 252-254, 259, 316-319 호미노이드초과 270 호미니드 24, 31, 36, 163, 267- 269, 271-277, 283, 284, 288-290, 296-298, 309, 325, 329, 332 , 338, 339,_ 341, 342, 345, 348, 355, 357, 360, 362-369, 378, 385, 397, 410, 435 호미노이드 370 홀로웨이, 랄프 330, 374 홀로세 144 홈 192 , 234, 235
홈날연모 423 홍해 271, 368 홀점 204 화석연속의 법칙 109, 110 화이트, 팀 280, 281, 305 화전농업 94 화학작용 374 환경생물학 127 환원주의자 453 효소단백질 67 후각돌기 377 후돈, 제임스 108, 239 혹인종 239-241 홍수아이 415 힐, 앤드류 277 A Arboreal Qu adrup e d 131 At trac ti on Provis i o n ing 322, 323 Bip ed al Locomoti on 132 Band 134 Brach iat o r 13I Bun 383, 398 Cong en ital Absence 234 Consort Pair 134 Crowd ing 234 Cy no dont 237 Ento c on id 190, 191
Harem 134 Hy pe rta u rodont 237 Hy po cone 191, 232, 235 Hy po con id 190, 191 Hy po conuli d 190, 191 Knuckle Walki ng 132 Leap er 131 Mesota u rodont 237 Meta c one 190, 191 Meta c on id 190, 191 Meta c onule 190 Orth o g na th o us 240 Paracone 190, 191 Paracon id 190, 191, 232 Progn a th o us 240 Proto c one 190-192, 232 Proto c on id 190-192 Retr o molar Sulcus 398 Rota t i on 234 Sollta r y 134 Sup e rnumerary Teeth 234 Suspe n sory 131 Talon id 190, 191 Terrestr i a l Qu adrup e d 132 Tri~ o n id 190, 191, 232 Troop s 134
*卜善周 연세대학교 사학과 졸업 동 대학 석사, 박사과정 미국 캘리포니아 주립 버클리 대학교 인류학과 박사과정 (체질 인류학 전공) 현재 충북대학교 고고미술사학과 교수 역서 『인류의 기원과 진화』(1 992) 논문 「인류기원에 관한 연구-오스트랄로피테쿠스에 대한 최근의 연구를 중심으로」 「유라시아대륙의 중기홍적세 호미니드 화석」 둥 20 여 편 체질인류학 대우학술충서 • 인문사회과학 78 1 판 1 쇄 찍음 • 1994 년 9 월 1 일 1 판 1 쇄 펴냄 • 1994 년 9 월 15 일 지은이-朴善周 펴낸이-朴孟浩 펴낸곳-(주)民音社 출판등록 1 9<)1 년 12 월 20 일 제 16-490 호 서울 강남구 신사등 5% 번지 강남출판문화센터 5 층 대표전화 • 515-20 0) 팩시밀리 • 515- 幻 m 값 __ 14,000 원 편집-임인기 조판-위드뱅크 KDC 생물과학, 체질인류학 /47 1. 5 ® 박선주, 1994 Pri nt e d in Seoul, Korea ISBN 89-3 7 4 -30 78-9 94490 ISBN 89-374 -30 00-2 (세트)
대우학술총서 ( 인문사회과학 ) 1 한국어의 계통 김방한 46 어원론 김방한 2 문학사회학 김현 47 조선의 서학사 강재언 3 商周史 윤내현 48 한국음악학 이강숙 4 인간의 지능 황정규 49 중국언어학 문선규 5 중국고대문학사 김학주 50 스포츠 심리학 6 일본의 만엽집 김사엽 류정무 ·이강 헌 7 현대의미론 이익환 51 원시유교 김승혜 8 베트남사 유인선 52 전쟁론 김홍철 9 인도철학사 길희성 53 삼국시대의 漠字音 111120 중사한상국회의주과의 학풍 과김수 광사수수상학 최이창승조훈 외 5554 아베유날르창균학고 파송 의김 응철종학 김형효 13 방언학 이익섭 56 자유주의의 원리와 역사 노명식 14 구조주의 소두영 57 국 민투표 구병삭 외 15 외교제 도사 김홍철 58 한국서지학 천혜봉 16 아 동심리학 최경숙 59 조선통신사 이원식 17 언어심 리학 조명한 60 명말청초사상 배 영동 18 법사회학 양건 61 한국의場市 이재하 · 홍순완 19 해양법 박준호 유병화 62 고려시대의 后妃 정용숙 20 한국의 정원 정동오 63 포스트모더니즘의 이론 21 현대도시론 강대기 김욱동 22 이슬람 사상사 김정위 64 한국의 복식미 김영자 23 동북아시아의 岩刻 畵 황용훈 65 정신신경증 이현수 24 자연법사상 박은정 66 한국주거사 홍형옥
