
데이비 드 틸 만 Da vid Ti lm an 데이비드 틸만은 〈 종간 경쟁의 기작 〉 과 〈 생물 다양성과 생태계의 동태 〉 그리고 이러한 주제와 관련된 수학적인 이론에 대해 연구하고 있다. 그는 미국생태학회로부터 쿠퍼상(1 989 ) 과 맥아더상(1 995) 을 수상하였 고, 미 국생태학회에서 발간하는 학술지의 창립 편집자이며, « 사이언 스»의 편집위원이 다 . 현재 맥나이트 대학 생태학 교수겸 미네소타 대학의 CedarCreekNa t u ra l A r ea 의 연구책임자이다. 문 형태 서울 대 학교 생물교육학과를 졸업했고. 서울대 대학원에서 식물학 박사 학위를 받았으며 , 뉴멕시코 주립대학에서 박사후 연구원을 역임했다. 현재 공주대학교 생물학과 교수이다.
식물의 전략과군집 구조
PI. ANT STRATEGIES AND THE Dy na mi cs and Str u ctu r e of P lant Communit ies by David Til m an Cop yrigh t © 1988 by Princ eto n Un ive rsity Press All righ ts reserved. Korean Translatio n Cop yrigh t © 1998 Minu msa Publi sh i ng Co., Ltd . Korean tran slatio n edit ion is p ub li sh ed by arrang em ent wit h Pri nc eto n Un ive rsity Press. 이 책의 한국어 판 저작권은 Princ eto n Un ive rsity Press 와 독접 계 약한 (주) 민음사에 있습니 다. 저작권법으로 한국에서 보호를 받는 저작물이므로 무단전재와 무단복제를 금합니다•
 식물의 전략과 군집 구조
식물의 전략과 군집 구조
서문 소비자와 이들이 이용하는 자원 사이의 상호 작용은 자연계의 질서 를 결정하는 중요한 요인이다. 이러한 상호 작용은 자원의 이용도와 소 비자의 자원 이용 방법, 죽 섭식 방법에 의해 영향을 받는다. 섭식 행동 이라는 표현은 동물의 경우 혼히 사용하는 용어이지만 식물의 경우에도 섭식 행동이 있댜 식물이 필요한 자원을 얻는 능력은 식물의 형태에 의해 영향을 받는댜 식물의 생장이 자원의 이용도에 얼마나 의존적인 가는 그 식물의 생리적인 특성과 형태적인 특성의 상호 작용을 통해 결 정된다. 식물은 고착 생활을 하기 때문에(동물에 비해) 조사하기가 쉽다 는 것 외에도 식물이 가지고 있는 주요 장점은 섭식 활동(광합성)을 주 로 하는 지상부의 형태가 시각적으로 분명하다는 것이다. 유감스럽게도 지하부의 섭식 활동(수분이나 양분 흡수 등)은 관찰하기가 더 어렵다. 식물은 진화 과정을 통해 놀랄 만큼 다양한 형태와 생활사를 갖게 되 었으며, 식물 군집에는 공간적으로 반복되는 많은 동적인 패턴이 있다 . 이러한 것들을 파악하기 위하여 이 책을 집필하게 되었다. 나는 2 년 전 부터 이러한 생각들을 구체화시키기 시작하였다 . 처음에는 그 당시에 기정 사실로 인정되고 있던 편견들을 보완하는 것에서부터 출발하였다. 토양에 있는 자원과 햇빛에 대하여 식물 개체군간의 경쟁 기작에 관한 논리적인 관계가 밝혀짐에 따라 일부의 편견은 더욱 강화되었고 일부는 배척되었다. 책을 쓴다는 것은 피곤할 정도로 시간이 많이 소요되고, 아직 밝혀지 지 않은 수많은 과학적인 사실들과 수시로 부딪쳐야 하기 때문에 때에
따라서는 지적인 면에서 두려운 일이기도 하다. 그러나 완전히 다른 생 각들이 하나로 통합될 때나. 혼돈과 같은 무질서 속에서 어떤 패턴을 찾아내게 되면 기분이 상쾌해지기도 한다. 지난 2 년 동안의 결과를 회 고해 보면 많은 것들이 미진한 상태로 남아 있음을 알 수 있다. 그러나 그 동안의 일을 통해 몇 가지 기본적인 과정들을 간파할 수 있었고. 이 를 통해 자연 식생에서 볼 수 있는 일반적인 패턴을 추론해 낼 수 있었 다. 나는 이것들을 이 책에서 여러분과 함께 나누고 싶은 생각입니다. 우리가 자연을 진실로 이해하기 위해서는 그 전에 알아야 할 일이 많이 있다는 것을 인정하며 이 일을 하려고 합니다. 나는 이 책에 제시된 생 각들을 통해 여러분이 식물 속성의 진화와 식물 군락의 구조와 동적인 상태를 결정하는 힘을 이해하는 데 도움이 되었으면 합니다. 이 책은 많은 사람들의 도움과 격려에 의해 완성되었습니다. 내가 집 필에만 몰두하는 동안 나를 도와준 아내 Ca t h i e 에게 먼저 감사를 드리 고 싶습니다. 그 다음에는 이 책에 있는 모든 그림을 준비해 주고 본문 을 교정해 주었으며, 인용 문헌을 정리하고 원고 준비의 거의 모든 면 에 도움을 준 AndreaLarsen 에게 깊은 감사를 드립니다. 또한 자료 분석 과 컴퓨터 시뮬레이션에 도움을 준 Robert Buck 과 Abderrahman El Hadd i에게도 감사를 드립니다. 이 책에 제시된 생각들은 내가 수년간에 걸쳐 관계를 맺어온 많은 사람들로부터 영향을 받은 것입니다. 많은 사 람들 중에 서 특히 Nancy Huntl y, Ric h ard Inouy e, Joh n Pasto r , Edward Cushi ng, Joh n Teste r , Evil le G orham, Hal Mooney, Marga r et Davis , Pete r Abrams, Lauri Oksanen, David Gr iga l, Deborah Goldberg, Norma Fowler, Joh n Ha rpe r, Pete r Vi tou sek, 그리 고 David We di n 에 게 고마움을 표합니 다. 또한 이 책 의 초고를 읽고 중요한 조언을 해준 Hal Mooney, Joh n Ha rpe r, Terry Chap in, Joh n Pasto r , Deborah Goldberg, Norma Fowler, Ste v e Pacala, Jim Grace, Scott Gleeson, Ste v e Fif ield , Barb Delaney, Bob McKane, Jen ny Edg er t on , 그리 고 미 시간 대 학의 대 학원생 들에 게 감사를
드립니댜 원고 편집을 도와준 Jud it h Ma y에게 고마움을 표합니다. 그러 나 이 책에 있을지도 모르는 모든 잘못된 점은 본인에게 책임이 있음을 밝혀두는 바입니다. 내가 특별 연구원 신분으로 이 일을 시작하도록 시 간을 허 락해 준 Joh n S. Gug ge nheim Memoria l Founda ti on 에 감사를 드 리 고. 또한 Cray 2 를 사용하도록 허 락해 주신 Mi nn esota Sup er comp ut e r Ins tit u t e 에도 심심한 사의를 표하는 바입니다. 미네소타의 Cedar Creek Natu r al Hi sto r y Area 에 있는 Long -T erm Ecolog ica l Research 에 재 정 적 인 도움을 준 Nati on al Scie n ce Foundati on (BSR-81143202, BSR-8612104) 에 깊은 감사를 드립니댜 만약 NSF 의 도움이 없었다면 이 책에서 언급된 일들은 수행할 수 없었을 것입니다. 1987 년 미네소타 대학에서
차례
서문 .5제 1 장 서론 11제 2 장 자원 경쟁에 대한 등구배 접근 방법 27제 3 장 영양염류와 햇빛에 대한 경쟁의 기작 63제 4 장 분배 패턴과 생활사 109제 5 장 생산력 구배와 손실률 구배상에서의 식생 패턴 149제 6 장 식물 경쟁의 동태 197제 7 장 천이 227제 8 장 미네소타 모래평원에서의 2차 천이 255제 9 장 문제점과 결론 317부록 : 모델 ALLOCATE를 이해하기 위한 수학적 기초 • 343옮긴이 해제 • 351참고문헌 • 359찾아보기 • 379제 1 장 서론 특정한 영양염류의 첨가나 p H 의 변화, 지하수위의 변화, 수분 스트레스 나 피음을 가하면 혼생하고 있는 한 쌍의 종 사이에 형성되는 경쟁적인 평형이 변화될 수 있음을 많은 사람들의 연구 결과를 통해 알 수 있다. 이러한 연구 결과들은 한 쌍의 종 사이의 상호 작용은 그 상호 작용이 일어나고 있는 환경의 함수이며, 환경과 종의 독특한 조건에서 특정한 변 화의 효과를 가져온다는 것을 강조한다는 점에서 역사적인 중요성이 있 다. 그러나 이러한 실험이 경쟁의 기작을 이해하는 데나 경쟁의 효과를 일반화시키는 데 중요한 기여를 하였는지의 여부는 분명하지 않다. —J.L . Ha rpe r(1 9 77, p. 369) 생태학의 가장 중요한 목표는 우리가 자연에서 관찰할 수 있는 패턴의 원인을 이해하는 것이다. 서식처간에 비슷한 패턴이 나타나 는 것은 이들 서식처에서 비슷한 힘이 작용하고 있음을 의미한다. 이 책의 내용은 육상식물에서 보고된 바 있는 좀더 광범위하고 일
반적인 패턴과 식물의 형태. 생활사 , 생리 등에 영향을 주어 식물군 집의 구조와 동태를 결정하는 몇 가지 힘에 관한 것이다 . 예를 들면, 조건이 매우 다른 서식처에서 각각 진행되고 있는 2 차 천이 과정 중 비슷한 천이 단계에서는 생리적. 형태적, 그리고 생활사의 특징 이 비슷한 종들이 우점하는 이유는 무엇인가 ?(B illi n g 1938, Bazzaz 1979, Chris te n sen & Peet 1981, MacMahon 1981, Coop er 1981. Inouy e et al. M19a8j7o ar ,1 T95il 5m) .a n 그 1리98고7a )오 스인트디레 애 일나리 (C아ow (lWeasl k1e8r9 e9t) a, l. 알19래81 )스 에카서 ( Cr진oc행ke되r 는& 1 차 천이가 적어도 처음 200 년 동안 그 패턴이 비슷한 원인은 무엇 인가? 한 지역 내에 있는 식생의 종 구성이 지소에 따라 많은 변화를 보이는 것은 지소에 따른 토양형――특히 토양이 형성되는 모재 과 그 지역에 형성되는 식생의 생산력과 관계가 있다 (L i ndse y 1961, Hole 1976, Rabin o vit ch -Vi n 1979, 1983, Jen ny 1980). 식물군계에서 식물 의 최고 높이, 상대 생장률, 분배 패턴, 그리고 생활사가 바슷한 종 들이 생산력 구배상의 비슷한 지점에서 우점종이 되는 이유는 무엇 인가? 생산력을 일정하게 하면 초식, 교란, 또는 다른 손실률(l oss ra t e) 들이 식생의 구조를 결정하는 주요인이 된다 (Lubchenco 1978. Gr im e 1979, W 血 ney 1986). 서식처에 따라 환경 조건이 서로 다르지 만 손실률의 구배에 따른 식생의 변화가 비슷한 이유는 무엇인가? 더욱 두드러진 패턴으로는 지리적으로 멀리 떨어져 있지만 물리적 인 환경 조건이 비슷한 서식처에서는 서로 관련없는 종들이 생리적, 형태적, 그리고 생활사의 특징이 비슷하게 수렴된다는 것이다 (Mooney 1977, Cody & Mooney 1978, Or ian s & Pa ine 1983, Walte r 1985). 또한 육상의 관속식 물은 대 부분 그들의 modul arity와 형 태 적 인 가변 성이 크다는 점에서 서로 비슷하다 (H arp er 1977). 이와 같이 유사한 패턴이 나타나는 원인을 밝혀내는 것이 식물
생태학자들의 당면 과제라고 할 수 있다. 이러한 유사성은 육상식물 의 진화와 군집의 구조가 몇 가지 일반적인 과정에 의해 크게 영향 울 받는다는 것을 의미하는가? 간단한 방법을 통해 이러한 모든 패 턴의 대략적인 윤곽이라도 설명이 가능한가? 아니면 이러한 패턴은 서로 관련이 없기 때문에 각각의 패턴에 따라 독자적인 설명이 필 요한 것인가? l 제약과 대응책 생태학에서 패턴은 물리적, 생물적인 환경 제약 (cons trai n t s) 과 그 러한 환경 제약을 해소시키기 위해 생물이 갖는 대응책(tr adeo ff s) 에 의해 유발된다. 이러한 제약과 대응책이 일반적이고 반복되는 것일 수록 일반적인 패턴이 형성된다. 종이 오랜 기간 동안 유지되기 위 해서는 분화가 필요하다. 즉, 자신의 환경에 의한 제약에 적절히 반 응할 수 있는 대응책을 갖어야 한다. 이 책의 내용은 육상식물에서 볼 수 있는 광범위한 분화 패턴의 원인과, 이러한 분화가 식물군집 의 동태와 구조에 미치는 영향에 관한 것이다. 그러면 육상식물이 직면하고 있는 주요 제약은 무엇인가? 몇 가 지 일반적인 제약은 먹이그물에서 그 식물이 차지하고 있는 위치에 서 비롯된다. 모든 종은 여러 가지 자원을 소비하는데, 자원 중에는 공급이 부족한 것도 있다. 관속식물은 영양염류, 물, 이산화탄소, 그 리고 햇빛을 필요로 한다. 이러한 자원을 이용할 수 있는 능력은 온 도, pH , 습도, 그리고 토양의 통기성(산소) 등에 따라 달라진다. 이 밖에 식물은 생태계에 있는 다양한 소비자, 초식동물, 기생자, 병원 성 생물, 포식자들의 자원이 되며, 다양한 유형의 환경 교란에 의해 일부가 소실되거나 사망하기도 한다. 따라서 주어진 서식처 내에서
식물은 자원 이용도, 그리고 교란과 초식으로 인한 손실과 사망에 의해 제약을 받는다. 또 하나의 제약은 식물에게 필수적인 자원이 물리적(공간적)으로 분리되어 있어 생긴다. 육상의 관속식물은 필요한 햇빛은 지상부에 서 얻고 영양염류와 물은 토양으로부터 얻는다. 이 자원들은 광합성 에 필수적이기 때문에 각 식물은 최적 생장을 위해 특정한 햇빛:영 양염류의 비를 필요로 한다. 햇빛이 제한되어 있는 식물은 더 많은 햇빛을 받기 위하여 줄기나 잎의 생장에 더 많은 에너지를 분배해 야 한다. 따라서 뿌리의 생장에 분배되는 에너지는 적어진다 . 반대 로 영양염류의 제한울 받는 식물은 뿌리의 생장에 많은 에너지를 분배해야 하기 때문에 줄기나 잎의 생장에 분배되는 에너지가 적어 진다. 따라서 식물체가 특정한 자원의 소비를 증가시키는 쪽으로 자신 의 에너지 분배를 조절한다면 그것이 얻을 수 있는 다른 자원의 상 대적인 양이 필연적으로 줄어든다. 이것은 육상식물의 형태와, 그리 고 햇빛과 토양 자원이 물리적으로 분리되어 있기 때문에 생기는 것으로 육상식물에게는 피할 수 없는 대응책이다. 이 책에서 전개시 킨 이론으로 예측할 수 있는 가장 중요한 것 중의 하나는 이러한 대응책이 우리가 자연의 식물 군집에서 볼 수 있는 패턴을 유발시 키는 주원인이라는 것이다. 서식처의 토양 자원과 광량의 수준에 따 라 각각의 서식처는 독특한 형태, 생리, 생활사를 갖는 식물에게 유 리하기 때문이다. 따라서 서식처 내에서 식생의 상관은 햇빛의 수직 적 구배와 토양 자원의 부족되는 수준을 조절하는 힘에 의해 큰 영 향을 받는다. 만약 일반적이고 반복되는 몇 가지 과정이 어느 서식 처의 자원 이용도의 패턴을 조절한다면 그 서식처에 있는 식물의 진 화와 군집 구조에서도 일반적이고 반복적인 패턴이 나타날 것이다.
1_l 생산력 구배 서식처에서 제한되는 토양 자원과 햇빛의 이용도를 결정하는 2 가 지의 주요인은 토양 자원의 공급률과 식물의 손실률 혹은 사망률이 다 . 제 9 장에서 설명한 바와 같이 손실률 혹은 사망률과 토양 자원의 공급률은 자연 서식처와 관계가 있으며. 이러한 관계는 자연적인 패 턴을 유발시킬 수 있다. 그러나 처음에는 어느 한 쪽을 일정하게 할 때 다른 것의 영향을 살펴보는 것이 도움이 된다. 손실률 혹은 사망률을 일정하게 할 때 식물의 서식처는 다음과 같은 두 가지 양극단의 서식처 사이의 어느 한 곳에 위치하게 된다. 죽 부족되는 토양 자원의 수준과 공급률이 낮고, 식물 생물량이 적 고, 지표면에 이입되는 햇빛이 많은 서식처로부터 제한되는 토양 자 원의 수준과 공급률이 높고, 식물 생물량이 많고 지표면에 도달되는 햇빛의 양이 적은 서식처로 이어지는 구배상의 어느 한 점에 위치 하는 것으로 볼 수 있다. 나는 이러한 구배를 〈 생산력 구배(p roduc tivi t y grad ie n ts ) > 혹은 〈토양 자원 :햇빛 구배 (soil r esource:lig h t gradi- en t s) 〉 라고 부르겠다. 새로 정착하는 식물의 실생과 줄기는 키가 작 고 , 이들의 생장은 햇빛의 강도에 영향을 받기 때문에 지표면의 햇 빛의 강도는 중요하다. 생산력 구배상에서 볼 수 있는 부족되는 토 양 자원의 공급률과 지표면에서의 햇빛의 강도 사이의 역상관 관계 는 육상 서식처에서 주요 제약이 된다. 이 책에서 나는 토양 자원을 자원의 수준과 자원의 공급률로 구 분하고 있다. 나는 자원의 수준을 이용 가능한 형태로 토양에 존재 하는 자원의 농도라고 정의한다. 때에 따라서는 자원의 수준과 같은 의미로 자원 이용도를 사용하기도 하였다. 자원의 공급률은 이용 가 능한 형태의 자원이 토양으로 방출되는 비율로 정의하였다. 모든 식 생의 수관층에서는 전광량을 받기 때문에 햇빛의 공급률에 대해서
는 정의를 하지 않았다. 오히려 나는 식생이 어떻게 햇빛의 수직 구 배에, 특히 식생이 지표면에서의 햇빛의 강도에 영향을 주는지 고려 하였다. 생산력 구배는 다양한 공간적인 규모에서 나타나는 것으로 알려 져 있다. 예를 들면, 미네소타, 위스콘신, 미시간에 있는 모래평원 또 는 플로리다 북부의 모래평원은 영양염류가 적고, 현존량이 적으며 지표면의 햇빛의 강도가 높은 지역이다. 반면에 미네소타, 위스콘신, 그리고 미시간에 있는 빙퇴석에 의해 형성된 토양은 영양염류 공급 률이 높고, 식물 생물량이 많으며 지표면에 입사되는 광량이 적다. 위스콘신의 Blackhawk Island 에서는 토양 모재의 차이에 의해 자연 적인 생산력 구배가 형성되는 토양이 발달되어 있다 (Pas t or et al. 1984). 또한 모든 서식처에는 1 차 생산력과 현존량에서 소규모의 공 간적 변이가 있다. 이러한 변이의 대부분은 지역에 따라 토양 영양 염류 공급률이 다르기 때문에 나타나는 것이다. 토양자원 공급률의 지역적인 변이는 수분 침투력의 차이, 질소 또 는 인의 치환 부위의 차이, 초식동물 배설의 영향, 토양 침식, 지형 적안 변이, 그리고 식물에 의한 피드백의 차이 등 여러 가지 요인에 의해 야기될 수 있다(J enn y 1980). 사면에 따라 나타나는 토양형의 차이와 같이 생산력 구배가 공간적으로 분명한 경우도 있다. 그러나 토양 자원:햇빛 구배는 공간적으로 뚜렷할 필요는 없으며, 제한되는 토양 자원의 공급률이 지점에 따라 변화가 있는 곳이면 어디서나 이러한 구배가 있을 수 있다. 이러한 크고 작은 규모의 구배는 모두 식물이 진화하고 분화되는 서식처에서 일반적이며 반복될 수 있는 중요한 서식처의 특징으로 작용하는 것 같다
1-2 손실 혹은 교란 구배 두번째로 중요한 서식처 제약은 교란 , 초식, 포식 그리고 식물의 부 분적 손실 혹은 사망을 유발시키는 비선택적인 요인에서 비롯된다 . 편 의상 나는 이 모든 것 들을 〈 손실(l oss) 〉 혹은 〈 교란 (d i s t ur-bance) 〉 이라고 부르겠다. 가상적인 손실률 구배(즉 토양의 자원 공급률을 일정하게 하고 손실률만 변화하는)상에서 부족되는 토양 자원의 이용도와 지표면에서의 햇빛 이용도가 어떻게 변하는지 살펴보자. 이러한 구배상에서는 서식처 의 토양 자원과 지표면에서의 햇빛의 강도가 모두 낮고 손실률도 낮다. 손실률이 높은 서식처는 토양 자원의 수준과 지표면에서의 햇빛 의 강도가 비교적 높다 . 따라서 토양 자원의 수준과 지표면의 햇빛 수준은 손실률 구배상에서는 정의 상관이 있지만 생산력 구배상에 서는 역상관 관계에 있다. 생산력 구배에 비해 손실 혹은 교란 구배 에 관한 연구가 적지만 토양 자원과 햇빛 수준은 이러한 구배상에 서 정의 상관을 보여준다 . 예를 들어 Hubbard Brook 에서의 실험을 살펴보도록 하자 (L ik ens et al. 1977, Boronann & Lik en s 1979). 교란되지 않은 삼림은 식물의 생 물량이 많고 지표면에 도달하는 햇빛의 강도가 낮으며, 추출할 수 있는 토양 영양염류의 수준이 낮다. 완전벌목은 지표면의 햇빛의 강 도를 증가시키며, 추출할 수 있는 토양의 영양염류의 양을 현저히 중가시킨다. 이것은 벌목지의 토양으로부터 분수역으로 홀러나오는 물 속에 영양염류가 증가되는 것으로 알 수 있다. 식물체가 고사하 면 이용 가능한 영양염류의 소비가 줄어들기 때문에 토양의 영양염 류 수준이 증가한다 (V it ousek et al. 1979, 1982). 모든 삼림은 그들 내부에 형성되는 다양한 크기의 군반에 영향을 주는 자연적인 교란이 있다. 삼림 전체에서 교란된 군반과 교란되지 않은 군반의 평균, 추출 가능한 토양 자원과 지표면에서의 햇빛의
평균 수준은 평균 손실률에 따라 증가한다 (Swank et al. 1981, Vi tou sek et a l. 1979, Vi tou sek & Mats o n 1985). 이 책에서 나는 식물의 고사나 생물량의 손실을 가져오는 모든 밀도 독립적. 그리고 비선택적인 과정을 하나로 통합하였다. 그 이 유는 식물의 고사나 생물량 손실의 원인이 무엇이든 간에 자원의 수준에 미치는 정성적인 영향은 바슷하며, 따라서 식물 형태나 생활 사에 미치는 영향도 비슷하기 때문이다. 나는 일반적인 원칙을 찾기 위해 이와 같이 단순화시켰다 . 그러나 특정한 초식동물의 효과나 특 정한 유형의 교란 효과가 포함되는 좀더 세밀하고 복잡한 연구를 통해서 더 많은 것을 알 수 있을 것이다. 그러나 이것은 이 책이 의 도하는 바가 아니다 . 이 책에서 나는 손실 혹은 교란을 초식동물, 종 자 혹은 실생 포식자, 불, 산사태, 나무의 고사 그리고 다른 과정에 의해 일어날 수 있는 밀도 독립적이며 비선택적인 손실과 같은 의 미로 사용할 것이다. 2 기계론적 이론에 대하여 손실률을 일정하게 유지할 때 토양 자원과 햇빛 수준이 역상관 관계가 되는 경향은 야외와 온실 실험을 통해 알 수 있는데 , 스웨덴 에 있는 가문비나무의 최적 영양에 관한 야외 실험에서 잘 나타나 있다(그림 1.1, Tamm et a l. 1974, Tamm & Aronsson 1982, Tamm 1985). H arp er(1977) 는 영양염류 시비 실험에 관한 종설에서 공급된 영양 염류가 식생에 미치는 주영향은 단순히 햇빛이 제한되는 시기를 단 축시키는 것이기 때문에 실험 결과를 해석하기가 어렵다는 것을 강 조한 바 있다(p. 340). Ha rpe r(1 9 77, p. 361) 는 벼과식물과 토끼풀 사 이의 경쟁 실험을 설명하는 과정에서 (S t em & Donald 1962), 다음과
같이 기술하였다. 〈 적당량의 질소가 공급된 벼과식물은 토끼풀보다 더 높이 자라 점차적으로 토끼풀을 억제하였다. 처음에는 이러한 실 험 결과가 질소영양에 관한 문제인 것으로 설명될 수 있다 . 질소를 공급하지 않으면 뿌리혹을 형성하는 질소고정 콩과식물이 유리하지

그림 1.1 스웨덴의 S tr asan 에서 가문비나무의 최적 영양을 결정하기 위해 만든 실 험 설계를 1975 년 7 월에 찍은 항공사진 . 질소 시비는 가문비나무의 생물 량을 증가시켜(검게 나타나는 실험구) 지표면에 이입되는 광량을 감소시 킨다. 영양염류 시비는 1967 년에 시작되었다. 실험구의 크기는 일반적으 로 30mx30m 였다. 최적 생장은 N,P,K , M g를 첨가하였을 때 나타났으 며 . 침엽의 영양염류 함량 및 뿌리, 침엽, 줄기로의 분배 패턴과 관련이 있 었다 (Tamm 1985). 자세한 내용이 필요하면 Tamm et a l.(1 974) 과 Tamm( 1985) 을 참조하기 바람 항공 사진은 Jo urnal oft he Roy al Swedis h Academy of A g ri c u ltu re and Forestr y, Sup ple ment 17, 12 쪽에 있는 Tarnm(1985) 의 논문에서 저자와 출판사의 허락하에 게재한 것이다 . 이 책에 실을 수 있도록 사진의 원본을 제공하여 준 Carl Tamm 교수에게 고 마움을표한다 .
만 적당량의 질소가 공급되면 벼과식물이 승자가 된다. 그러나 질소 원의 영향을 입사하는 햇빛의 영향으로부터 분리한다는 것은 비현 실적이댜 실험은 처음에 단일 요인의 영향을 파악하는 실험으로 시 작되었지만 곧 요인들간의 상호 작용에 관한 연구로 바뀌게 되었 댜〉 토양 자원과 햇빛 사이의 이러한 상호 작용을 통하여 식물의 경 쟁에 관한 실험의 설계, 실행 및 결과 해석이 어렵다는 H arp er 의 생 각이 타당함을 알 수 있다. 그러나 이것들은 식물 경쟁의 기본적인 제약이자 기작인 것이기 때문에 식물 경쟁에 관한 기계론적 이론에 는 이러한 것들이 포함되어야 한다. 이 책의 3, 4, 6, 그리고 7 장에서 전개되는 이론은 광합성 산물을 줄기, 뿌리, 잎의 생물량으로 분배 하는 패턴에 의해 식물의 영양생장이 결정되는 것으로 모델화한 것 이다. 생 장을 계 속하고 있는 크기 구조를 갖는 (s i ze-s tru c tu red) 식 물 개 체 군에 관한 영양염류와 햇빛의 경쟁 모델은 가능한 단순하게 구성되 어 있다. 그러나 그 속에는 토양 자원과 햇빛을 획득하기 위한 육상 식물의 경쟁 능력에 영향을 주는 주요 형태적, 생활사적 특징이 포 함되어 있다. 이 모델은 식물의 형태(분배 패턴)와 생활사의 진화에 대하여, 그리고 이러한 식물의 특징이 식물 군집의 동적인 구조와 평형구조에 미치는 영향에 대한 다양한 의문점들을 탐구하는 데 사 용된다. 이 책의 가장 중요한 목표는 영양염류와 햇빛에 대한 식물 의 경쟁 기작의 논리적인 관계를 탐구하는 것이다. 대부분의 생태학적 이론은 현상학적인 것이었다. 이러한 이론은 경쟁이나 공생과 같은 상호 작용에서 어느 한 종이 다른 종에게 영 향을 주는 실제적인 기작에 대해서는 전혀 언급이 없이 단순히 한 종의 밀도가 다른 종의 생장률에 어떻게 영향을 준다는 것으로 기 술하고 있다. 이러한 이론으로는 종 특성의 진화나, 개체군, 군집, 그
리고 생태계의 구조와 동태를 유발시키는 기작을 탐구할 수 없다 (Til m an 1987a). 따라서 이 책에서는 다른 접근 방법을 택하였다 . 즉 한 종의 개체들이 같은 종의 개체나 다른 종의 개체에 영향을 주는 과정을 분명히 제시하는 방법을 택하였다. 종의 형태와 생리, 생활사를 결정짓는 것과 각 종이 우점종 혹은 희귀종이 되는 조건 에 영향을 주는 것은 바로 이러한 기작이다. 생태학 분야에 만약 어 떤 단순성이나 일반성이 있다면, 그것은 이러한 기작을 무시하는 단 순한 이론에 있는 것이 아니고 환경적 제약과 상호 작용의 기작에 서 발견할 수 있을 것이다. 기계론적 접근 방법의 주요 장점은 처음 에는 좁은 범위에 초점을 맞출 수 있지만 필요에 따라 다른 기작과 먹이그물의 거의 모든 부분과 비생물 환경이 포함되도록 그 범위를 확장시킬 수 있다는 것이다. 이 책의 범위는 제한되어 있다. 이 책의 내용은 육상식물에서 종 내 그리고 종간 경쟁의 몇 가지 기본적인 기작과. 이러한 기작들이 식물 특성의 진화와 식물 군집의 동태 및 구조와 어떤 관련이 있는 가에 초점을 맞추고 있다. 앞에서 언급된 구성원 이외에 식물과 초 식동물의 상호 작용은 다루지 않았다. 그 이유는 초식의 중요성을 경시해서가 아니고 자원 경쟁 기작의 논리적인 결과를 탐구하기 위 한 것이다. 이 책은 또한 식물 경쟁에서 Pacala 와 S il ander(1985), 그 리고 Pacala(1986) 에 의해 제기된 바 있는 서로 이웃하고 있는 개체 간의 공간 효과에 대하여 자세히 설명하지 않았다. 토양 영양염류 공급의 기본적인 기작과 식물이 토양에 미치는 반작용 효과에 대해 서도 깊이 있게 다루지 않았다. 이들 각각은 이 책에서 제시한 아이 디어로 더 깊이 탐구할 가치가 있는 중요한 영역이다. 그러나 어느 하나의 연구 논문이나 책이 생태학의 모든 분야를 다룰 수는 없다. 자연 생태계는 매우 복잡하며, 그것을 구성하는 주 요 기본 과정을 모두 이해한다는 것은 어려운 일이다. 우리는 경쟁,
포식. 상리공생. 교란만이 자연 생태계를 구성하는 중요한 과정이 아니라는 것을 알고 있다(Q u i nn&dunham 1983). 물론 이 모든 것들 은 중요하며, 또한 모두 상호작용을 하고 있다. 그러나 아직도 단순 한 시각을 통해 우리가 알아야 할 것이 많고, 분석을 용이하게 하기 위하여 잠재적으로 중요한 다른 요인들을 일정하게 유지한 상태에 서 몇 가지 요인들의 연관을 탐구함으로써 얻어야 할 것들이 많이 있다. 그렇게 얻어진 지식이 모여 다른 단순화된 가정을 제시할 수 있는 기회를 주게 된다. 다세포식물 사이의 종내 그리고 종간 경쟁의 실질적인 기작은 단 순하지 않다. 다세포식물은 크기와 연령 의존적인 과정들을 가지고 있는데. 이것들이 식물에 대한 이해를 더욱 복잡하게 한다. 식물은 형태적으로나 생리적으로 가변성을 가지고 있다 . 식물은 상당히 고 정된 소단위(잎, 종자, 뿌리, 줄기)로 구성된 모듈 형태를 가지고 있다 (Ha rpe r 1977). 그러나 식물은 이러한 소단위에 대한 상대적인 자원 분배량을 형태적으로나 유전적으로 변형시킬 수 있다. 제 2 장과 9 장 에 설명되어 있는 바와 같이 이러한 형태적 가변성은 식물의 종간 그리고 종내 경쟁 능력에 영향을 줄 수 있다. 이렇게 복잡한 상태에 서 식물의 경쟁을 어떻게 보아야 할 것인가? 한 가지 방법은 기작을 무시하고 경쟁 현상을 설명하여 단순함을 찾는 것이다. 이것은 Lotk a -Volte r ra 방정식에서 사용한 방법이다. 이 와 유사하게 de W it(1 960) 은 식물 경쟁은 너무 복잡하고 각 생물과 서식처에 따라 너무 독특하여 일반적이고 기계론적인 식물 경쟁 이 론식을 만들어낼 가망성이 없다고 주장한 바 있다. 대신에 그는 현 상학적인 접근 방법을 제시하였다. 이와는 대조적으로 이 책에서는 식물 경쟁의 일반적인 특징 몇 가지를 탐구하는 데 이용할 수 있는 단순하지만 기계론적인 경쟁의 이론을 개발하려는 것이다. 기계론적인 접근 방법은 식물 생태학에서 큰 도움이 될 수 있는
사물을 보는 시각에 제약을 준다 . 식물 생태학에서 고전적인 몇 가 지 논쟁 예를 들어 식물 군집이 하나의 실체 단위로 존재하느냐 그 렇지 않으면 개별적 반응에 의한 종의 집합체이냐에 관한 논쟁에 많은 생태학자들이 너무 오랜 기간 매달려 있었다. 그러한 문제는 기계론적인 접근 방법에서 보면 별로 중요한 것이 아니다. 기계론적 인 접근 방법은 식물의 형태를 결정하는 진화적인 힘에 기초를 두 지 않은 그럴 듯한 대규모의 군집 일반화를 검증할 필요가 없다. 대 신에 종 생물학에서 기본적인 대응책과 물리적, 생물적 환경의 제약 때문에 예견되는 좀더 정량적인 패턴에 관심을 두고 있다. 3 자원부족 식물 경쟁을 이해하기 위한 중요한 단계는 주어진 상황에서 종간 상호 작용의 실제 기작이 무엇을 위한 것인가를 결정하는 일이다. 식물 경쟁의 가장 그럴 듯한 두 가지 기작은 자원 경쟁과 타감 효 과적 경쟁이다. 자원 경쟁은 부족한 자원의 소비를 통해 한 식물이 다른 식물의 생장을 저해할 때 나타난다. 타감 효과적 경쟁은 한 개 체가 환경에 방출하는 물질이 다른 식물의 성장을 저해하거나 사망 률을 증가시킬 때 일어난다. 이 기작들이 모두 다른 종의 생장률에 대한 어느 종의 밀도의 직접적인 효과를 나타내는 것은 아니다. 두 가지 경우 모두 각 종의 밀도가 어떤 중간 매개체에 직접 영향을 주고 이 중간 매개체가 다른 종의 생장률에 실제로 영향을 준다. 따 라서 자연 상태에서 종이 자원에 대해 실제로 경쟁하는지를 보여주 기 위해서는 야의에서 실험적으로 자원의 수준을 조절할 필요가 있 댜 이와 마찬가지로 야의에서 타감 효과적 경쟁이 일어난다는 것을 보여주기 위해서는 야외에서 타감 효과가 있는 물질의 농도를 실험
적으로 조절할 필요가 있다 . 이 책에서 개발된 이론은 자원 경쟁의 경우에만 적용할 수 있다 . 이 이론을 특정한 군집에 적용하기 전에 식물이 자원에 대한 경쟁을 하고 있다는 것과, 어떤 자원이 부족된 다는 것을 보여주는 야외 실험이 선행되어야 한다. 식물 군집에 대한 기계론적인 접근 방법이 갖는 강점은 이 방법 이 자연계에 있는 광범위한 패턴과 과정에 대하여 분명한 예측을 할 수 있다는 것이다. 그러나 기계론적인 이론은 다른 이론과 마찬 가지로 잘못 적용될 수도 있다. 질소와 햇빛에 대한 기계론적인 경 쟁 이론의 예측과 자연계에서 실제로 볼 수 있는 패턴 사이의 일치 성은 그 군집에서 질소와 햇빛이 제한 요인이 아니라면 별로 중요 하지 않다. 제한 요인이란 증거도 없으면서 일치성을 기대한다는 것 은 참재적으로 위험성이 크다. 이 책에서 나는 현존하는 중거를 바 탕으로 우리가 자연계에서 볼 수 있는 패턴을 설명하기 위한 기작 을 제시하겠지만 그러한 경우의 대부분이 단순히 그럴 듯한 것만을 보여준다는 점을 강조하고 싶다. 나는 이론의 증거로서가 아니고 다 른 사람들이 여기에서 개발된 생각들을 검증할 수 있도록 하기 위 해 그러한 기작들을 제시하였다. 마지막으로, 이 책에서 택한 접근 방법은 개념적인 면에서 Gr ime (1 979) 의 방법과 매우 다르며, G ri me 의 결론과 직접 상충되는 결론 에 이른다는 점을 지적하고 싶다. 나는 여러 가지 다양한 상호 작용 과정에 대하여 Grim e 이 제시한 방법에 동의하진 않지만 경쟁, 손실 률(Grim e 의 교란), 그리고 자원의 이용도(낮은 이용도가 G ri me 의 스 트레스에 가장 중요한 구성원임)가 식물 군집 구조에 중요한 영향 을 준다는 그의 견해에는 동의하고 있다 . 나는 Gri me 의 견해와 나 의 견해 사이의 같은 점과 다른점을 이 책의 관련된 부분에서 설명 하였다.
4 개관 Til m an(1980, 1982) 은 부족한 토양의 영양염류와 지표면의 햇빛 이용도의 장기간 평균치가 영양염류 공급률과 서식처에서의 손실률 에 의존적이라는 것을 보여주기 위해 경쟁에 대한 자원 의존적 등 구배 생장 방법을 발전시켰는데, 이 책에서는 바로 이 방법이 사용 되었댜 식물은 지상부의 자원인 햇빛과 지하부의 자원인 영양염류 와 물 모두를 필요로 하기 때문에 여기에 대한 대응책을 가지게 된 다. 한 자원을 더 얻기 위해서는 다른 자원을 상대적으로 덜 얻어야 한댜 따라서 지상부와 지하부 생장에 대한 식물의 분배 패턴은 주 어진 서식처에서 식물의 경쟁력에 영향을 준다(제 4 장과 제 5 장). 그러나, 줄기나 뿌리와 같은 비광합성 기관에 대한 분배가 항상 식물의 최대 영양생장률을 감소시키는 것은 아니며(제 3 장), 따라서 식물 개체군 동태에 큰 영항을 줄 수 있다(제 6 장). 최대 생장률의 차이 때문에 나타나는 일시적인 개체군 동태는 2 차 천이 패턴의 주 원인이 될 수 있으며, 단기간의 야외 실험 결과를 해석하는 데 어려 움을 줄 수 있다(제 7 잣) . 미네소타의 Cedar Creek Natu ra l Histo r y Area 에서 5 년 동안 식물의 분포와 천이의 동태를 조사한 실험 연구 는 많은 정보를 제공하고 있다 . 이들 중 많은 부분이 전에 발표되지 않은 것들이다. 그 정보를 가지고 이 책에서 개발된 이론의 예측을 평가해야 한다(제 8 장). 이 책의 마지막 장에는 여기에서 개발된 이 론이 가지고 있는 몇 가지 부수적인 결과에 대한 탐구와 앞으로의 연구 방향이 제시되어 있다(제 9 장).
제 2 장 자원 경쟁에 대한 등구배 접근 방법 1 서론 개체군의 크기 구조, 영양염류와 햇빛과의 연관, 부족되는 토양 자원과 햇빛을 확보하기 위한 식물의 대응책 둥에 의해 나타나는 복잡성 때문에 식물 경쟁에 관련된 모든 내용을 하나의 단순한 이 론 속에 다 포함시킬 수는 없다. 그렇다면 이것은 경쟁에 관한 단순 한 접근 방법이 아무 쓸모가 없다는 것을 뜻하는 것일까? 복잡한 모델들은 때로는 그 내용을 몇 개의 식으로 적절히 요약할 수 있다 (Scha fter 1981). 모델에 포함되어 있는 여러 가지 복잡한 과정 중 일부는 전체 과정을 통제하게 되는데, 이러한 과정들은 우리가 관찰 할 수 있는 패턴을 결정하는 주요인이 된다. 우리는 아직 식물 경쟁 의 기작에 대한 단순한 모델과 복잡한 모델의 장점과 단점을 모두 다 알 수 없다. 이 장에서 나는 자원에 대한 식물 경쟁의 단순한 이 론을 요약하려고 한다. 이 책의 나머지 부분에서 나는 식물 경쟁의
기작에 대한 좀더 복잡하고 실질적인 모델을 개발하여. 이 복잡한 모델에 의한 예측과 이 장에서 전개시킨 단순한 이론에 기초를 둔 예측을 서로 비교하게 될 것이다. 단순한 이론에서는 자원에 대한 각 개체군의 종간 경쟁의 결과를 예측하기 위해 각 개체군이 평형 상태에 도달하였을 때의 조건을 이용한다. 이것은 생활사의 여러 단계에서의 자원 요구는 그 종의 자원요구 평형점에 미치는 각 단계 자원 요구의 영향으로 집약될 수 있다고 가정한다. 평형 상태에 도달하지 않는 개체군의 경우, 나 는 자원 이용도의 장기간의 평균값이 평형 조건에 가장 알맞은 근 사값이 될 것이라고 가정한다 . 자원 요구의 평형점은 그 종의 자원 의존적 생장 등구배곡선에 의해 주어진다 (T i lman 1980, 1982). 이 장 내용의 상당 부분은 그 전에 내가 발표한 내용의 설명을 되풀이하고 있지만 (T i lman 1980, 1982), 여기에서 나는 4 가지의 기본 개념을 발전시키려고 한다. 첫째, 모든 부족되는 자원의 이용도는 손실률 구배와 정의 상관을 갖어야 하며, 둘째 부족되는 토양 자원 과 햇빛의 이용도는 생산력 구배상에서 부의 상관을 갖어야 한다. 셋째 식물체는 그들의 자원 요구에 의해 결정된 방식으로 이러한 구배상에서 서로 분리되어야 하며, 넷째, 형태적으로 가변적인 식물 이 필수 영양 자원을 최적으로 얻을 때는 자원 의존적 생장 둥구배 곡선이 되어야 한다. 이때 형태적으로 가변적인 식물은 형태가 고정 된 식물에 비해 경쟁에 유리하다. 이전에 발표된 내용은 다시 언급하지 않으려고 했지만 어느 정도 의 반복은 불가피하였다. 전에 내가 발표한 내용을 잘 알고 있는 사 람들은 이 장을 그냥 대강 훑어보고 지나갈 수 있다. 나는 전에 내 가 발표한 내용을 잘 알지 못하는 사람들도 이해할 수 있게 쓰려고 노력하였다. 이 장에서 제시하는 단순한 모델들은 제 3 장과 제 4 장에 서 제시된 크기 구조를 갖는 식물의 분배와 생장을 위한 좀더 복잡
한 모델과 대조를 이룰 수가 있다. 그 모델은 뿌리, 잎, 줄기, 그리고 종자로의 에너지 분배 패턴에 의해 식물의 생장이 결정된다고 가정 한다. 각 식물종은 그것의 분배 패턴, 종자의 크기, 성숙기의 키, 최 대 광합성률, 영양염류와 햇빛에 대한 광합성의 의존성, 조직에 따 른 호홉률, 상대 생장적인 제약과 구조적 제약 그리고 다른 변수들 에 의해 설명된다. 2 단일 제한 자원에 대한 경쟁 먼저 몇 종의 서로 다른 식물이 같은 자원에 의해 모두 제한을 받는 경우를 생각해 보자. 상호 작용의 결과로 결국 평형 상태에 이 른다고 가정하면 종간 경쟁의 결과는 어떻게 될까? 단일 제한 자원 에 대한 경쟁의 결과를 예측하기 위해서는 한 종 개체군의 순변화 율이 0 일 때의 자원의 수준을 알아야 한다. 이것은 주어진 서식처에 서 종의 영양 생장과 생식이 손실률과 균형을 유지할 때 나타난다. 나는 이때의 자원 수준을 그 서식처에서 제한되는 자원에 대한 그 종의 R* 라고 지칭하였다 . 이에 관한 정보를 얻기 위해서는 두 가지 방법이 있댜 주어진 서식처에서 한 종의 R* 롤 결정하는 첫번째 방법은 그 서 식처의 단일종 지소 (monos p ec ifi c s t and) 에서 생물량이 평형 상태에 이르도록 하는 것이다. 이와 같은 평형 상태에서 그 종이 제한되는 자원을 감소시키는 수준이 그 종의 R* 가 되는 것이다. 평형 상태에 서 자원의 소바율은 자원의 공급률과 같을 것이다. 만약 실제 자원 수준이 R* 보다 큰 서식처에 어떤 종이 살고 있다면, 그 개체군의 크 기(그 개체군이 안정한 연령 상태 혹은 안정된 크기 분포에 도달하였을 때 단위 면적당 그것의 무게를 의미한다)는 증가할 것이다. 따라서 자
원 수준은 R * 까지 낮아진다. 자원 수준이 R * 보다 낮으면 개체군의 크 기는 감소하여 소비율이 감소하기 때문에 자원 수준은 증가할 것이다. 자원 수준이 R * 인 서식처에서만 개체군의 크기가 일정하게 유지된다 . 나는 R * 를 부족되는 자원에 대한 그 종의 자원 요구량이라고 부른다. R * 를 결정하기 위한 두번째 방법은 그림 2.1 의 자원 의존적 생장 곡선에서 나타난 바와 같이 자원 수준에 대한 종의 생장률의 의존도 를 결정하는 것이다. 이 그림의 y 축은 개체군의 장기간의 생장률 혹 은 손실률을 나타낸다 (dB!d t • l!B, B 는 단위 면적당 생물량). 만약 개 체군의 크기가 단위 면적당 생물량으로 표시되는 크기 구조를 갖는 개체군이라면 생장률은 각 자원 수준에서 결정된 개체군 투영행렬의 우세한 고유치의 자연대수가 될 것이다. 그러나 이 경우에는 자원 독 립적 사망이나 다른 손실이 없어야 한다 (Hubbell & Werner 1979, Vandermeer 1980 ) . 개체군의 전체 손실률은 비슷한 방법으로 계산되어야 한다. 크기 구조를 갖는 개체군에서 이러한 손실률은 자원 의존적인 생장 항목을 빼고 모든 자원 독립적인 손실 항목을 포함하는 개체군 투영 행렬의 우세한 고유치의 자연대수로 측정될 것이다. 생식과 영양 생장을 통한 이득과 교란, 초식동물, 포식. 기타 다른 사망원을 통한 손실이 균형을 이룰 때의 자원의 이용도가 R * 이다(그림 2.1). 몇 가지 종이 모두 한 가지 자원에 의해 제한울 받을 때 가장 낮 은 R* 를 갖는 종은 평형 상태에서 다른 모든 종을 경쟁적으로 교체 할 것으로 예측된다 (O' Br ien 1974, Ti lm an 1976, Hsu et al. 1977, Armstr on g & McGehee 1980). 경쟁적 교체의 기작은 자원 소비이댜 가장 낮은 R* 를 갖는 종의 개체군 크기는 그 종이 자원의 수준(농 도)을 자신의 R* 수준으로 감소시킬 때까지 계속 증가할 수 있을 것 이다. 이 시점에 이르면 다른 종이 생존하기에 충분한 자원이 없게 된다. 단일 제한 자원에 대한 경쟁 기준으로 R * 를 실험적으로 시험
 A aj
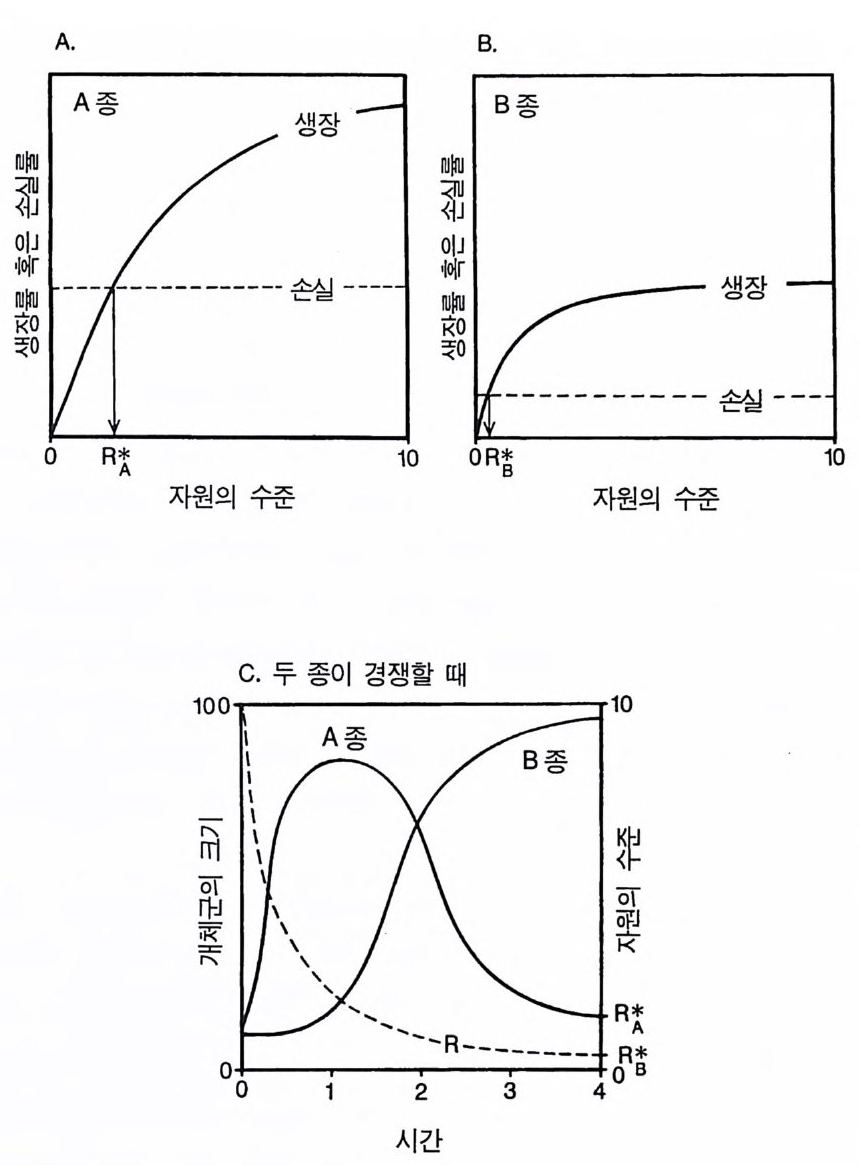
A aj
그림 2.1 (A) 종 A 의 자원 의존적인 생장(실선)과 손실(점선). RA* 는 종 A 가 이 서식처에서 생존하는 데 필요한 자원의 양이다. (B) 종 B 에 대한 곡선. (C) 두 종이 단일 자원 (R) 에 대해 경쟁할 때 일단 평형 상태에 이르면 자원 요구량이 낮은 B 종이 A 종을 완전히 교체하게 된다.
그림 2.2 18L 용량의 화분에 8 종의 휴경지 식물을 심었을 때 질소 구배에 따른 고밀도(화분당 약 100 개체) 혹은 저밀도(화분당 7 개체)에서 식물 개체당 평균 무게. 질소 구배는 비옥한 표층토를 하층에 있는 모래와 혼합 비율 을 달리하여 만들었다 . 질소 무기화율은 전질소 함량과 높은 상관이 있었 다 . 질소의 모든 수준에서 식물체당 평균 두게는 밀도가 낮은 화분에 비 해 밀도가 높은 화분에서 적었다. 가장 척박한 토양(전질소 함량이 125m g/kg)은 Cedar Creek 의 휴경지에서 자연 상태로 있는 토양에 비해 더 척박하였고` 가장 비옥한 토양 (1,825m g/kg)은 Cedar Creek 에 있는 가 장 비옥한 토양과 비슷하였다 . Cowan( 1986) 에서 인용.
한 결과가 T il man(1982) 에 요약되어 있댜 이론적으로 가능한 한 가 지 경우가 그림 2 .1에 나타나 있다. 여기에서 제시된 이론에 의하면 식물은 자원 수준이 낮은 서식처 에서 경쟁을 심하게 한다는 것이다 . 이러한 견해는 G ri me(1979) 의 주장과 상치된다. Gr i me 의 주장에 따르면 식물은 영양염류가 적은 환경, 또는 교란의 빈도가 높은 서식처 등과 같이 스트레스가 있는 환경에 살 때 경쟁을 하지 않는다는 것이다. 그러나 그의 주장은 종 간 경쟁이나 종내 경쟁에 관한 많은 다른 연구 결과와 일치하지 않 는다. 만약 영양염류가 빈약한 토양에서 식물이 경쟁을 하지 않는다 면 그런 토양에서 단일종 재배시 식물의 밀도를 증가시키더라도 상 대생장률 (dB/Bd t, B 는 식물 생물량)과 식물 개체당 평균 무게가 감소 하지 않을 것이다. 그러나 많은 연구 결과에 의하면 영양염류가 빈 약한 토양이나 풍부한 토양에서 초기의 식물 밀도를 증가시키면 상 대생장률과 개체당 무게가 감소하는 것으로 나타나 있다 (Donald 1951, Clatw ort hy 1960, Ha rpe r 1961, 1977). 실제로 밀도 증가에 따른 생장률 감소는 〈최종 생산량 일정의 법칙 〉 에 선행 조건이다 (K ir ae t al. 1960, Ha rpe r 1961, 1977). Cowan(1986) 은 토양의 질소 함량 구배상에서 8 종의 초본을 고밀 도와 저밀도로 생육시켰다. 전체 구배상에서 모든 종의 식물 개체당 최종 평균 무게(그림 2 , 2) 와 상대 생장률은 고밀도에서 현저히 낮았
다 . 이것은 질소 함량이 매우 낮은 토양을 포함하여 모든 구배에서 종내 경쟁이 심함을 보여주는 것이다. Inouy e 등 (1980) 과 Inouy e (1 980) 의 연구 결과에 의하면 사막 1 년생 식물 사이에서 경쟁이 심 함을 알 수 있다. S t em 과 Donald(1962) 의 실험에 의하면 질소 함량 이 적은 토양에서 토끼풀은 133 일 동안의 경쟁 후에 벼과식물을 교 체함을 알 수 있다. 질소 공급이 많은 지역에서는 이러한 경쟁 결과가 역전된다. Mahmoud 와 Gr i me(1976) 은 3 종의 벼과식물 (Arrhenatl z erum elati us , Festu c a ovin a , Ag ro stis t enu i s) 을 재 료로 비 옥한 토양과 척박한 토양에서 이들 종 사이의 경쟁적 상호 작용을 조사 하였댜 단일종 지소의 생육 실험과 비교하면 척박한 토양에서 두번 째 종이 있음으로 식물 개체당 무게가 감소되었다 . A g ros ti s 는 Arrhena t herum 의 줄기당 무게를 30% 감소시켰고 Fes t uca 의 경우 43% 가 감소되었다 . 그리고 Arrhena t herum 은 낮은 질소 처리구에서 A g ros ti s 를 59% 나 감소시켰다 . 따라서 이러한 실험 결과들은 모두 영양염류가 부족한 토양에서 종간 경쟁과 종내 경쟁이 있음을 입증 하는 것들이다. 3 R* 와 손실률 혹은 교란율 평형 상태에서 부족되는 자원이 감소되는 최종 수준은 식물 생장 의 자원 의존도와 손실에 의해 결정된다. 개체군의 손실률은 교란, 종자 포식, 초식 등 여러 가지 요인에 의해 야기된다. 특정한 서식처 에서 손실을 야기시키는 요인이나 자원에 대해 경쟁하고 있는 종 수 및 종의 경쟁 능력과는 무관하게 평균 자원 수준 (R * )은 손실률 에 따라 증가할 것이다(그림 2.3). 손실률에 따른 R * 의 증가는 식물 의 생장률이 자원 수준에 의존적이기 때문이다. 평형이 유지되기 위해
 A.
A.
그림 2.3 (A) 손실률이 평균 자원 이용도에 미치는 영향 . 실선으로 된 곡선은 자원 이용도에 대한 dB/Bd t의 의존도를 나타낸다. 접선으로 표시된 D1-D5 는 교 란 . 사망 . 초식 기타 손실 률 이 다른 경우이다. R * 들은 각 손실률과 연관된 자원 이용도 를 나타낸다. 개체군은 서식처에서 자신의 생장률이 적어도 손실률과 균형을 이 룰 때만 유지될 수 있다 . 생장률 (dB/Bd t, B 는 생물량) 은 자원의 이용도 (R) 에 따라 중가하기 때문에 높은 손실률에서 개체군이 생존하기 위해서는 자원 이용도가 높아야 한다. (B) 많은 종이 경쟁할 때 에도 R* 는 손실률에 따라 증가한다. D 1 에서는 A 종이 다른 모든 종을 교 체하게 되고, C 종은 D2 에서」 3 종은 D3 에서. 그리고 G 종은 D 4 에서 경쟁에 이긴다. (C) A 종이 항상 경쟁에 우세한 경우 R* 는 D에 따라 증가한다 . (D) 이 경우들은 R * 가 손실률이나 교란율에 따라 항상 증가하여야 한다 는 것을 보여준다 . B 의 일부에서는 R* 가 D에 따라 거의 직선적으로 증가 한다(곡선 #2). C 의 일부에서는 R * 가 D 에 따라 거의 지수함수적으로 증
서는 생장은 손실과 균형을 이루어야 한다. 평형 상태에서 손실률이 높으면 생장률도 높아야 하고, 생장률을 높이기 위해서는 자원 수준 이 높아야 한다. 따라서, 평균 자원 수준은 손실률의 증가에 따라 증 가되어야 한다(그림 2.3A). 손실률이 높을 때 자원 수준이 높으면 소비자 생물량이 낮게 유지된다. 생장률이 자원 이용도에 따라 증가하고. 모든 종의 손실률이 증가 한다면 경쟁하고 있는 종 수나 그 종 개체들의 생장이 자원 수준에 얼마나 의존적이냐에 관계없이 이러한 패턴은 유지된다. 최대 생장 률이 높은 종이 자원이 빈약한 서식처에서 생장률이 낮은 경우를 생각해 보자. 손실률이 증가함에 따라 평형 상태의 자원 수준은 증 가하고 연속적인 종의 교체가 있을 것이다(그림 2.3B). 종이 그러한 대응책을 가지고 있지 않다면(그림 2.3C), 모든 손실률에서 한 종이 우점종이 될 것이다. 그러나 R* 는 손실률에 따라 계속 증가할 것이 다. 모든 종의 손실률이 증가하는 한 평형 상태의 자원 수준은 그 공급원이 무엇이든 간에 정도의 차이는 있울지언정 손실률에 따라 증가할 것이댜 평형 상태의 자원 이용도와 손실률 사이의 곡선 모 양은 여러 가지 요인에 의해 결정되지만 자원 수준은 손실률의 층 가함수임에 틀림없다. 그림 2.3C 의 관계가 그림 2.3D 의 곡선 1 을 유도하고, 그림 2.3B 의 관계들은 그림 2.3D 의 곡선 2 를 유도한다. 평형 상태의 자원 수준이 서식처의 손실률에 따라 증가하는 경향 은 서식처의 구조를 결정하는 주요소가 된다. 더욱이 이러한 경향은 경쟁과 모든 손실원(교란과 초식 포함) 사이에 불가분의 관계가 있음 을 보여주고 있다. 많은 사람들이 주장하는 것처 럼 (Grim e 1979, Ha rris 1986, Wi en s 1984), 손실률이 높은 서식처에서는 경쟁이 중요 하다. 그러한 서식처는 모든 부족되는 자원의 수준이 높은 반면에 이러한 지역에서 종이 생존하기 위해서는 요구하는 자원 수준도 높 다. 자원에 대해 경쟁하고 있는 식물이 초식의 영향도 받는다면, 초
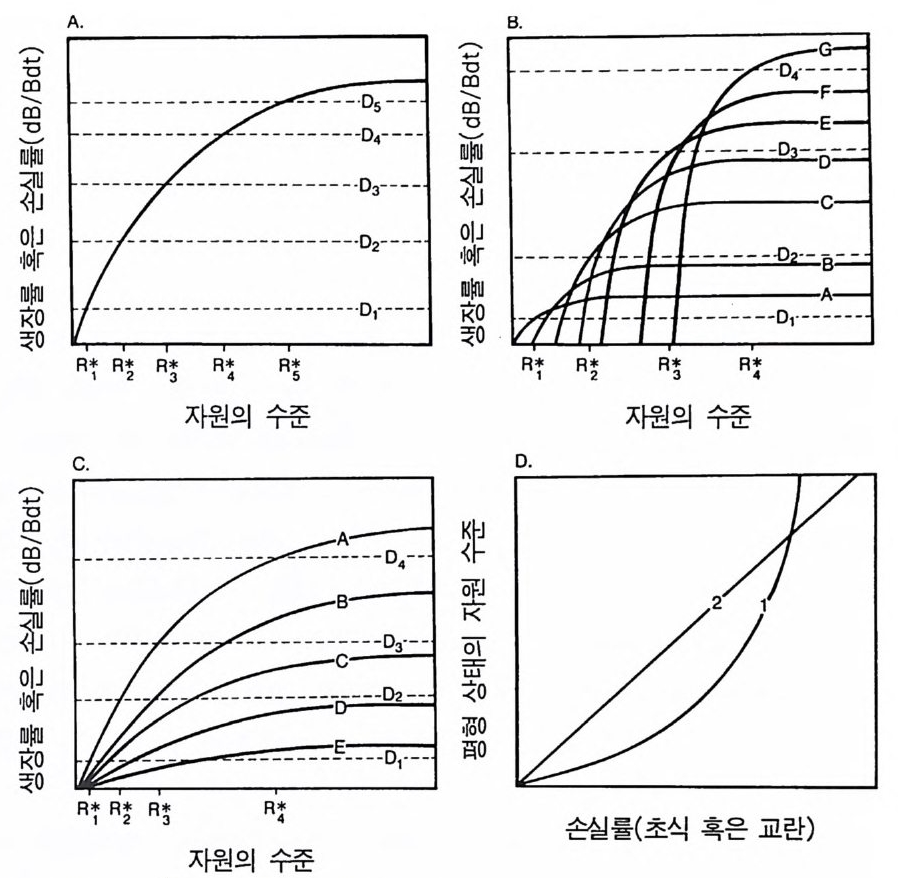
식동물의 밀도가 높은 서식처에서는 초식이 중요하고 경쟁은 중요 하지 않다고 말하는 것은 논리적으로 타당하지 않다(Q u i n & Dunham 1983). 종의 손실률에 영향을 주는 모든 과정들은 그들의 경쟁적 상호 작용에도 영향을 준다. 경쟁은 교란이나 초식과 불가분의 관계에 있 댜 손실률과 생장 사이의 이와 같은 관계로 인하여 식물이 분화할 수 있는 주요 구배가 형성된다. 이것은 손실률이 낮고 평형 상태의 자원 수준이 낮은 서식처로부터 손실률이 높고 자원 수준이 높은 서식처로 이어지는 구배이다. 이러한 정성적인 패턴은 평형 상태에 도달하지 않고 변동하는 서식처에서도 똑같이 유지된다. 즉 장기간 의 평균 손실률이 높은 서식처에서는 모든 제한되는 자원의 장기간 에 걸친 평균 수준이 높아야 한다. 평균이란 말은 시간에 따른 평균 과 공간에 따른 평균 모두를 의미하는데, 그 이유는 개개의 식물체 와 그들의 사망원 혹은 손실원이 시간과 공간에 따라 불연속적인 점으로 나타나기 때문이다. 4 자원 의존적 인 생장 등구배곡선 어느 한 종이 2-3 가지의 자원을 소비할 때는 자원이 그 종의 생 장에 미치는 전체적인 효과를 알아야 한다. 평형 상태에서 이러한 효과는 그 종의 제로 순생장 둥구배곡선 (zero net grow t h i socl in e) 으 로 요약할 수 있다 (T il man 1980). 그 종의 단위 생물량당 생장률이 그것의 손실률과 균형을 유지할 때 이 등구배곡선은 두 가지 이상 의 자원 수준을 보여준다. 종의 둥구배곡선 모양은 자원의 유형을 정의하는 데 이용될 수 있다. 따라서 한 쌍의 자원은 완전 대체가 가능한 경우, 상호 보완적인 경우, 길항 관계인 경우, 교환이 가능한
그림 2.4 각 그림에서 실선으로 된 곡선은 개체군의 제로 순생장 등구배곡선을 나 타낸다. 제로 순생장 둥구배곡선은 생장과 생식이 모든 손실원과 정확히 균형을 유지하도록 하는 자원 l 과 자원 2 의 이용도를 보여준다. R1 과 R 2 는 서식처의 자원 수준임을 주목하기 바란다. 죽 각 자원의 이용 가능한 형태의 측정 농도이다. 자원 수준이 어두운 부분에서는 개체군의 크기가 증가하고 밝은 부분에서는 감소될 것이다.
경우. 완전히 필수적인 경우, 상호 작용에 의해 필수적인 경우나 반 필수적인 관계에 있을 수 있다(그림 2.4 . Til m an 1980, 1982 참조). 페 쇄된 곡선을 포함하여 다른 모양도 이론적으로 가능하다. 광합성을 하는 식물은 모두 빛, 물. 여러 가지 형태의 C. N, P. K, Ca, Mg , S 그 리고 약 20 가지의 다른 영양염류를 필요로 한다. 이러한 자원들은 영양 면에서 서로 필수적이다. 동물에게 필요한 자원은 대부분 대체가 가능하거나 치환이 가능 하댜 이러한 성질은 식물과 동물 경쟁 사이에 차이를 유발시키고 그들의 다양성 패턴에도 차이를 가져온다 (T i lman 1982). 두 자원이 모두 완전히 필수적이라면 한 자원이 부족할 때 다른 자원이 증가 되더라도 그 부족을 상쇄시킬 수 없다(그림 2.5A). 따라서 햇빛과 영양염류가 모두 필수적일 때 식물체가 햇빛에 의해 제약을 받는다 면 P. K, N 등의 수준을 증가시키더라도 생장률이 증가될 수 없다 . 만약 두 자원이 상호 작용적 필수 자원인 경우라면 등구배곡선은 코너가 구부러진 곡선을 갖을 수 있고, 두 자원 중 어느 한 자원이 증가되더라고 생장률이 증가된다(그림 2.5B). 그립 2 .4 E 의 둥구배곡선에서 직각은 R1 과 R2 가 완전히 필수적 임 을 의미한댜 R1 과 R 2 의 이용도가 점 x 와 같은 등구배곡선의 수평부 분에 있는 서식처에서는 R 의 감소 혹은 증가가 이 종의 생장률에는 아무런 효과가 없다. 그러나 R 2 의 증가는 그 종의 손실률에 비해 생 장률이 크기 때문에 밀도가 증가하게 될 것이다. R2 의 감소는 반대 로 밀도의 감소를 초래하게 된다 . 따라서 등구배곡선의 수평 부분에
 A. 완전 대체가 가능한 경우 B. 상호 보완적인 경우
A. 완전 대체가 가능한 경우 B. 상호 보완적인 경우
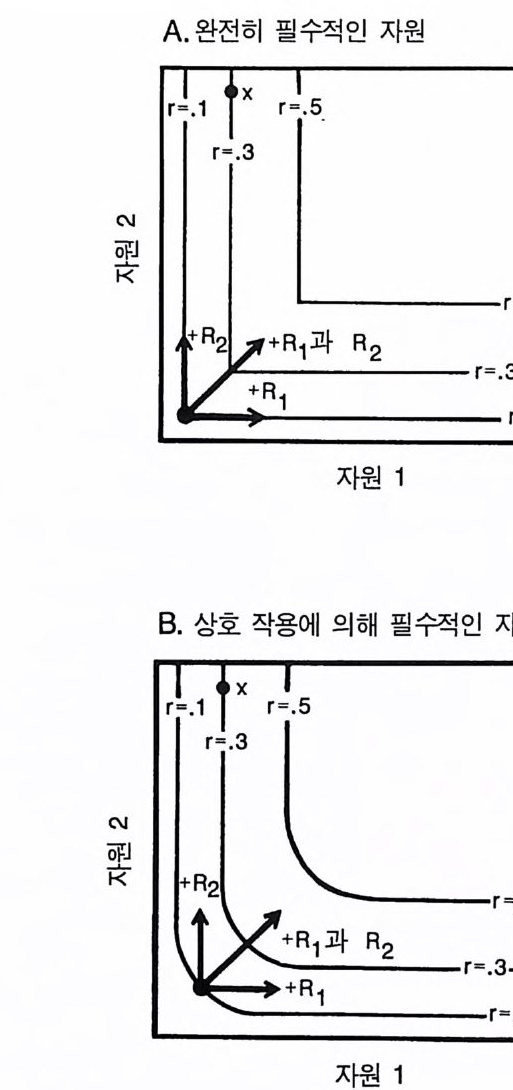 A. 완전히 필수적인 자원
A. 완전히 필수적인 자원
그림 2.5 (A) 완전히 필수적인 자원을 갖는 개체군의 자원 의존적 생장 등구배곡 선. (B) 상호 작용적 필수 자원을 갖는 개체군의 생장 등구배곡선 각 그 립에 나타나 있는 3 개의 생장 등구배곡선은 단위 생식률 r 가 0.1, 0.3 , 혹 은 0 .5/tim e 에서의 자원 수준을 나타낸다. 화살표는 R1 만을 첨가할 때, R2 만을 첨가할 때, R, 과 R2 를 같이 첨가할 때의 생식률의 효과를 보여준다 .
서 이 종은 R 2 에 의해 제한울 받고 R1 에 의해서는 영향을 받지 않는 다. 이와 비슷하게 R1 과 R2 의 이용도가 점 y와 같은 부분에 위치할 때에는 R 2 에 의해서는 영향을 받지 않고 R1 에 의해 제한울 받는다. 등구배곡선의 코너에서는(그림 2.5 A ), 두 자원에 의해 모두 제한울 받기 때문에 이 종의 생장률을 증가시키려면 두 가지 자원을 모두 증가시켜야 한다. 그러나 생장률을 감소시키려면 한 가지 자원만 감 소시켜도 된다. 두 자원이 상호 작용적 필수 자원이라면 등구배곡선은 코너가 둥 글게 된다(그림 2.4 F ). 이 등구배곡선의 점 x 또는 점 y에서 R1 혹은 R 2 를 제거 혹은 첨가하면 이 종의 반응은 완전히 · 필수적인 자원에 서와 거의 같다. 그러나 등구배곡선의 구부러진 부분 근처에서 R1 또는 R2 를 첨가하면 이 종의 개체군 밀도가 증가하게 된다(그림 2.5B). 따라서 두 자원에 의한 이중 제한이 생기게 되는데, 이것은 자원 수준의 전범위에 걸쳐 기능적으로 서로 대체가 가능하다 . 완전 한 필수 자원과 상호 작용에 의한 필수 자원 사이의 정성적인 차이 는 거의 없다 (T i lman 1982). 이 책에서 혹은 그 이전에 제시된 아이 디어 중에서도 그것들이 완전한 필수 자원과 상호 작용에 의한 필 수 자원을 근거로 한 것이라면 정성적으로 차이가 없다. 그러나 다 음 항에서 설명한 바와 같이 최적 획득 이론을 통한 예측에 의하면 다세포식물은 둥근 등구배곡선을 갖어야 하기 때문에 실험을 통해 그 차이를 입증할 가치가 있는 것이다. 4-1 최적 획득과 동구배곡선의 모양 어느 한 쌍의 자원에 대한 자원 의존적인 둥구배 생장곡선의 모 양은 단지 자원의 영양적인 성질보다도 다른 많은 요인에 의해 결 정된다 (T i lman 1982). 예를 들어 영양 면에서 완전히 대체가 가능한
자원은 교체 방식으로 소비되어야 하는데, 그 결과 코너가 직각이 며, 밖으로 구부러진 등구배곡선이 형성된다 (T i lman 1982). 그러나 다음에 설명된 바와 같이 영양 면에서 필수적인 자원의 최적 획득 에 관한 이론에 의하면 자원 의존적 생장 등구배곡선은 코너가 직 각이 아니고 둥글어야 한다 . 식물체는 현재 자신의 생장에 제한 요 인이 아닌 필수 원소의 획득을 위해 소비하는 에너지를 감소시켜 다른 필수 원소의 획득을 증가시킬 수 있기 때문에 코너가 둥근 등 구배곡선이 나타난다. 이것이 육상식물의 진화와 어떤 연관이 있는 지를 설명하기 전에 먼저 영양 면에서 모두 필수적인 두 가지 자원 의 최적 획득에 대하여 살펴보도록 하자. 내가 최적 획득설을 사용 하는 이유는 모든 생물들이 자원을 최적으로 획득하는 경향이 있어 서가 아니라 이것이 생물이 갖는 특성 중 어떤 것이 적합도를 높이 는가를 결정하는 간단한 방법이기 때문이다. Ra pp o rt (1971) 와 Cov i ch(1972) 는 최적 영양에 관한 여러 가지 면 을 쉽게 파악할 수 있는 도식적 방법을 개발하였다. 이 방법은 영양 에 관한 두 가지 제약을 고려하고 있다. 하나는 개체가 단위 시간당 소비할 수 있는 두 자원의 양에 대한 억제(소비억제라 불린다)이고, 다른 하나는 자원의 영양적인 면과 소비되는 두 자원의 양에 따른 개체의 적합도에 대한 억제(영양적 억제라고 불린다)이다. 영양 면에서 완전히 필수적인 자원의 경우, 영양적 억제는 그림 2.6 의 1 번 곡선과 2 번 곡선에서와 같이 동일한 적합도(생장률)의 등 구배곡선으로 나타난다. 직각의 코너는 영양 면에서 완전히 필수적 인 자원에 의해 야기된다. 기점에서 멀리 떨어진 영양 등구배곡선은 적합도가 높은 곡선인데, 이것은 생장률이 단위 시간당 소비되는 자 원의 양에 따라 증가함을 나타낸다(이 그림들의 축은 자원의 수준이 아니고 소비되는 R1 과 R2 의 양이다). 소비 억제곡선 (consum pti on constr aintc urve, 그림 2.6 의 a 혹은 b 로 표시된 것)은 개체가 주어진 시
간에 소비할 수 있는 두 자원의 최대량을 나타낸다 . 소비억제곡선에는 4 가지 유형이 있다. 이 중 3 가지(직선. 볼록형 . 오목형)는 다른 자원에 대한 어느 한 가지 자원의 획득에 대응책이 있음울 가정하고 있다. 이 3 가지 소비억제곡선의 경우 다른 자원의 소비가 감소되면 어느 한 자원의 소비 증가가 가능하다. 식물의 경 우. 이러한 대응책은 형태적 혹은 생리적인 가변성으로 반영될 수 있다. 예를 들어 , 잎과 줄기의 생물량이 상대적으로 많은 식물은 필 연적으로 뿌리의 비율이 낮다. 소비억제곡선의 실제 모양(직선, 볼 록형. 오목형)은 생리적, 형태적, 환경적 요인에 의해 결정된다. 이 3 가지 유형이 갖는 중요한 면은 그것들이 석물의 자원 획득에 가변 성이 있음을 가정하고 있는 점이다. 죽 식물체는 한 가지 자원의 획 득을 줄임으로써 다른 자원을 더 많이 획득할 수 있다는 것이다. 이 와는 반대로 완전한 직각 코너를 갖는 소비억제곡선(그림 2.6C 와 E) 은 식물체가 가변성은 없지만 두 가지 자원을 소비할 수 있는 독자 적이고 고정된 능력이 있는 것으로 가정한다. 개체의 최적 영양은 그것의 소비억제곡선에서 종의 적합도를 최 대로 하는 점이다. 이 점은 소비억제곡선과 영양억제곡선 사이의 접 촉점으로 적합도는 가장 높지만(기점에서 가장 멀리 있음) 아직도 소 바억제곡선에 접촉되어 있댜 직선형, 볼록형 혹은 오목형 소비억제 곡선의 경우 최적 영양은 항상 두 자원이 균등하게 생장을 제한하 도록 하는 비율로 자원을 소비하는 것이다(그림 2.6A,B.D). 따라서 자원을 최적으로 획득하는 식물이 형태적 혹은 생리적으로 가변성 이 있다면 자기들의 형태와 생리를 모든 자원에 의해 균등하게 제 한될 수 있도록 조절해 야 한다 (Iwasa & Roug hg ar den 1984, Bloom , Chapi n & Mooney 1985). 이것이 어떻게 자원 의존적인 등구배 생장 곡선의 모양에 영향을 주는가? 그림 2.6A 에 나와 있는 소비억제곡선인 a 와 b 를 살펴보자. 소비
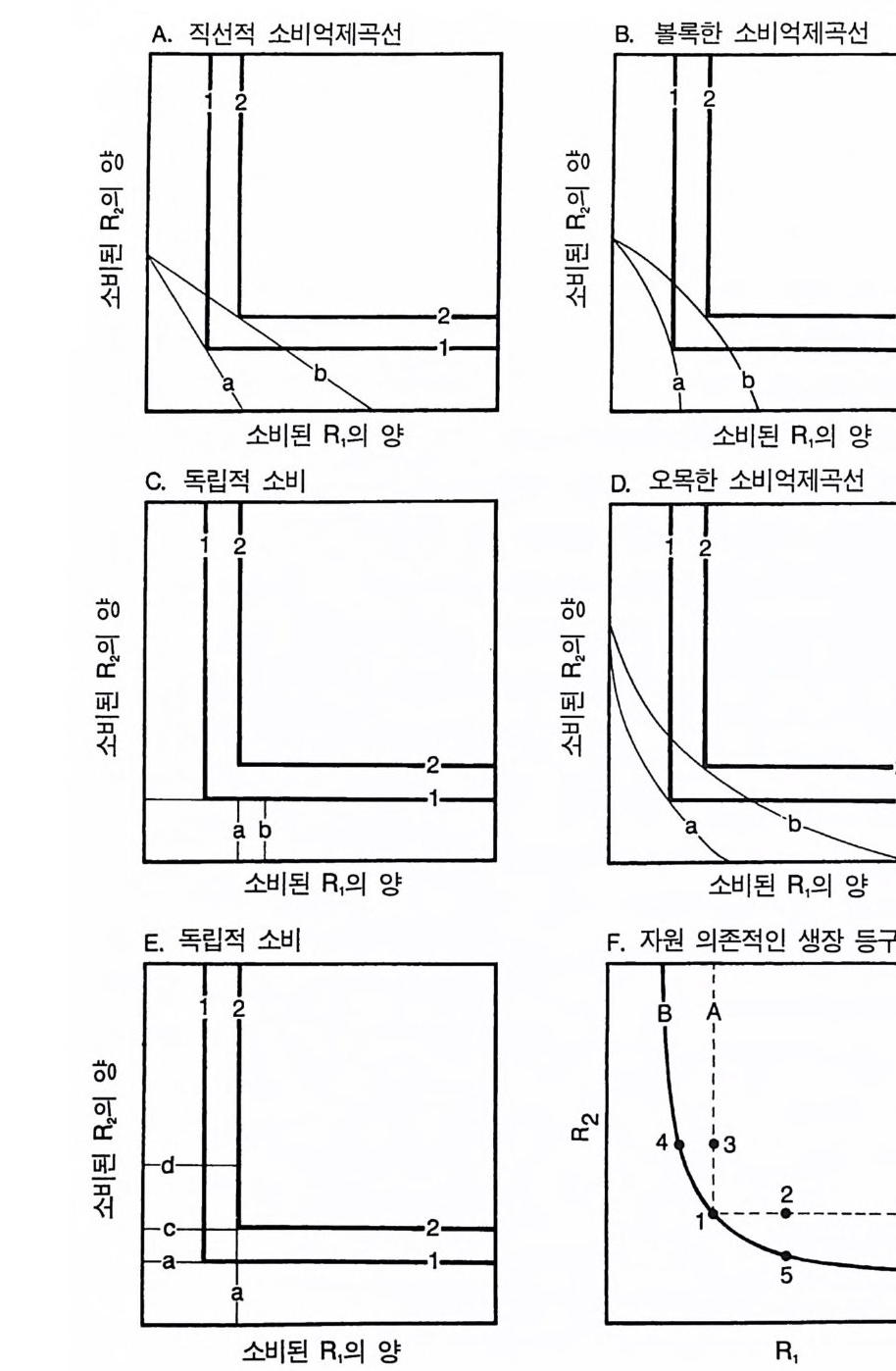 A. 직선적 소비억제곡선 B 볼록한 소비억제곡선
A. 직선적 소비억제곡선 B 볼록한 소비억제곡선
그림 2.6 (A-E) 이 그림들은 환경에 있는 자원의 양이 아니고 소비된 자원의 양에 대한 생장 의존도를 보여주는 것이기 때문에 이 책에 있는 다른 둥구배곡 선 그림들과는 다르다. 굵은 선들은 주어진 시간 동안에 개체 혹은 단위 생물량당 소비된 두 가지 자원의 양에 대한 개체 혹은 단위 생물량당 생 장 의존도를 나타내는 영양 등구배곡선들이다. 가는 선은 주어진 시간 동 안에 개체 혹은 단위 생물량당 소비될 수 있는 자원의 최대량을 나타내는 소비억제곡선들이다. 적합도(생장률)가 가장 높은 영양 둥구배곡선이 소 비억제곡선과 만나는 점이 최적 영양이다 . 소비억제곡선에 대응책이 있다 면 (A, B 그리고 D), 식물은 자신이 모든 자원에 의해 균등하게 제한 받 을 수 있는 비율로 자원을 소비하도록 자원 획득을 조정해야 만한다. (F) 그러한 최적 자원 획득은 이 그림에 있는 실선으로 된 곡선과 같은 자원 의존적 생장 등구배곡선을 초래하여 코너가 직각이 아닌 곡선이 된다. 점 선으로 표시되어 있는 코너가 직각인 둥구배곡선은 내재하고 있는 생리 적 제약을 보여주는 것이다 . 식물이 가변성을 갖는 데 따로 경비가 소요 되지 않는다고 가정하면 형태적 또는 생리적 가변성은 구부러진 등구배 곡선을 나타낼 것이고, 만약 가변성을 갖는 데 여분의 경비가 필요하다면 등구배곡선 A 가 등구배곡선 B 와 교차할 것이다.
억제곡선 a 의 경우. 자원을 최적으로 획득하는 개체는 적합도 등구 배곡선 l 을 갖는댜 만약 R1 의 이용도가 증가한다면 어떤 현상이 일 어날까? R 의 이용도가 층가하면 R2 의 어떤 주어진 소비율에서도 이 개체는 전보다 더 높은 비율로 R1 을 소비할 수 있음을 의미한다. 따라서 그것의 소비억제곡선은 b 로 전환될 것이다. 그러나 곡선 b 는 개체가 R 과 R2 를 모두 더 많이 소비하고 높은 적합도(생장률)를 갖 는 새로운 최적 영양점으로 유도한다. 그 결과 R1 과 R2 가 영양 면에 서 완전히 필수적인 자원일 경우에도 그것들은 자원 획득의 가변성 때문에 생태학적으로 상호 작용적 필수 원소로 행동한다. 죽 두 자원 중 어느 하나를 첨가하면 생장률이 증가한다. 볼록형 혹은 오목형 소 비억제곡선일 경우에도 비슷한 패턴이 나타난다(그림 2 . 6B 와 D). 이러한 모든 소비억제곡선은 식물이 한 자원을 소비하는 데 소모 하는 시간과 노력을 감소시켜 다른 자원의 소비를 위한 시간과 노
력을 증가시킬 수 있는 쪽으로 자신의 소비 패턴을 변형시킬 수 있 다는 것을 가정하고 있다. 이러한 자원 획득의 가변성은 영양 면에 서 완전히 필수적인 두 개의 자원은 둥근 코너를 갖는 자원 의존적 생장 등구배곡선을 유도한다는 것을 의미한다(이 점에 착안할 수 있 게 해준 Joe l Brown 에게 감사한다). 영양 면에서 완전히 필수적인 두 개의 자원이 상호 작용적 필수 자원의 생장 등구배곡선을 유도하지 않는 유일한 경우는 식물이 자 원 획득의 가변성이 없을 때이다(그림 2.6C 와 E). 이 경우에는 식물 의 자원 의존적 생장 등구배곡선은 직각 코너를 갖는다 . 죽 완전한 필수자원이 될 것이다. 4-2 형태적 가변성의 진화 육상 관속식물에서 가장 일반적인 특징 중의 하나는 형태적인 가 변성이다 (Waller 1986). H arp er(1977) 가 주장한 것처럼 식물은 제각 기 고유한 기능을 갖는 다소 고정적인 소단위로 구성되어 있지만, 이러한 소단위들(잎, 줄기, 뿌리 등)은 환경 조건에 따라 생장 정도가 서로 다르다. 대부분의 식물체들이 이러한 형태적인 가변성을 갖게 된 것은 무엇 때문인가? 식물의 형태는 그것의 자원 획득 노력이 체형으로 구체화된 것이 다. 뿌리는 영양염류를 획득하는 역할을 한다. 다른 모든 것이 균등 하다면 뿌리의 양이 많은 식물은 더 많은 토양 자원을 획득할 수 있다. 잎은 햇빛을 획득한다. 줄기도 잎을 더 높이 유지하기 때문에 햇빛을 획득하는 기능이 있고 일부 줄기는 광합성을 하는 것도 있 다. 햇빛 그리고 물과 같은 토양 자원은 영양 면에서 필수적이기 때 문에 어느 주어진 시점에서 식물 개체의 생장률은 이것들 중 어느 하나에 의해 결정된다. 따라서 현재 자신을 제약하고 있는 자원의
획득량을 증가시킬 수 있는 식물은 자신의 생장률을 증가시킬 수 있다. 형태적으로 가변성인 식물은 잎, 줄기. 뿌리의 생산을 변화시 킬 수 있으며. 따라서 제한되는 자원 획득을 증가시키는 쪽으로 자 신들의 자원 획득 노력을 조절한다. 그들은 모든 자원에 의해 균등 하게 제한울 받도록 자신의 형태를 조절해야 한다(I wasa & Roug ha rden 1984, Bloom, Chapi n, Mooney 1985). 가변성이 없는 식물은 그림 2.6F 의 점선으로 나타낸 것과 같이 직각의 코너를 갖는 자원 의존적 생장 둥구배곡선울 갖는다. 그러나 형태적인 가변성을 갖는 똑같은 개체는 실선과 같은 등구배곡선을 갖는다. 형태적으로 가변성이 있는 개체는 1 로 표시된 점을 제외한 모든 서식처에서 가변성이 없는 개체와의 경쟁에서 이길 수 있다. 따라서 두 개의 필수 자원을 최적으로 획득하고 있는 개체들은 모 든 자원에 의해 균등하게 제한을 받도록 자신들의 생리와 형태를 조절한다. 가변성 유지에 에너지가 소요되지 않는다고 가정하면 이 러한 개체들은 자신들의 생리와 생태를 최적으로 조절하지 않는 개 체들을 경쟁을 통해 교체한다. 그림 2.6F 에 있는 두 개의 등구배곡 선은 식물에서 형태적 가변성이 혼히 있음을 보여주는 것이다. 5 두 가지 필수 자원에 대한 종간 경쟁 육상식물은 형태적, 생리적으로 가변성이 있기 때문에 (H arp er 1977, Waller 1986, Bloom, Chapi n, Mooney 1985), 나는 햇빛과 토양의 영양염류와 같은 지상부와 지하부 자원에 대한 자원 의존적 생장 등구배곡선은 상호 작용적 필수 자원, 죽 둥근 코너를 갖는 것으로 가정하려 한다. 자원 의존적 생장 둥구배곡선은 종간 경쟁의 결과를 예측하는 데 필요한 정보의 하나이다. 평형 상태가 되기 위해서는
각 종의 생장이 그것의 손실과 같아야 한다 . 이것은 종의 제로 순생 장 둥구배곡선상의 어떤 점에서도 나타날 수 있다 . 이밖에 평형 상 태가 되기 위해서는 자원의 농도가 일정하여야 한다. 즉 각 자원의 소비율이 공급률과 균형을 이루어야 한다. 따라서 둥구배상의 실제 점은 특정한 서식처에서 자원의 공급과 소비의 동태에 따라 달라지 는 평형점이 될 것이다 . 5-1 자원 소비 각 종의 자원 소비율은 벡터로 나타낼 수 있다(그림 2.7 A ). 주어진 종은 단위 시간 그리고 단위 생물량당 각 자원의 일정량을 소비할 것이다. C1 을 단위 시간. 단위 생물량당 소비된 자원 1(RI) 의 양이라 고 하고 C2 를 소비된 자원 2(R 시의 양이라고 하면 두 자원의 전체 소비율은 벡터 c 로 나타낼 수 있다. 벡터 c 는 C1 과 C2 두 가지 요소 로구성된댜 그림 2.6 에 나타나 있는 바와 같이, 영양 면에서 완전히 필수적안 두 가지 자원은 그들이 식물 생장을 똑같이 제한하는 비율로 소비 되어야 한다. 생장에 제한 요인이 되지 않는 자원을 과다하게 소비 하는 것은 변동하는 환경 조건에서 경쟁 방해의 수단 이외에는 자연 선택의 장점이 될 수 없다. 변동하는 환경 조건에서도 장기간의 자원 소비율의 평균값은 그 식물이 자원에 의해 균등하게 제한되는 비율 에 근접해야 된다. 이러한 비율이 그림 2.7B 에 점선으로 나타나 있 는데, 기준점에서부터 등구배곡선의 코너를 통하는 것으로 되어 있 다. 둥구배곡선상에 있는 한 점에서의 소비 벡터는 그 선이 둥구배곡 선과 교차하는 점에서 이 점선의 접선과 평행하여야 한다. 그러한 소 비 벡터의 몇 개가 그림 2.7B 에 나와 있다. 이 그림은 D1-D4 로 표시 된 4 가지 상이한 서식처와 연관된 등구배곡선을 보여주고 있다. 서
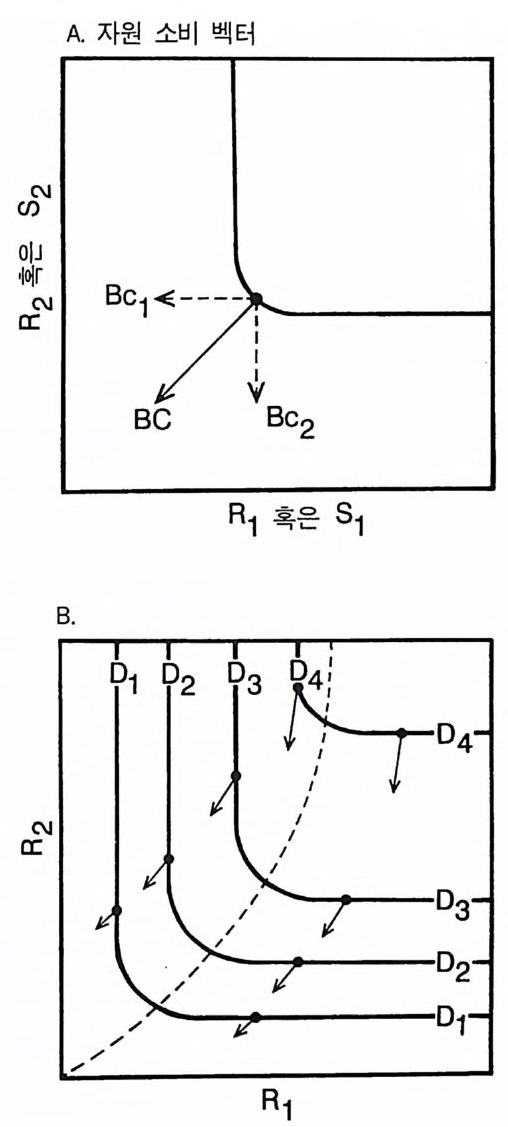 A. 자원 소비 벡터
A. 자원 소비 벡터
그림 2.7 (A) 실선으로 된 곡선은 자원 의존적 생장 등구배곡선이다 . 화살표는 자 원 소비 벡터이다. 소비 벡터인 BC 는 두 가지 구성원을 갖는데. c1 은 단 위 생물량당 R1 의 소비율이고, C2 는 단위 생물량당 R2 의 소비율이다. 전체 홉수율을 얻기 위해서는 이들 구성원 각각에 전체 식물 생물량 B 를 곱한 다 . (B) 4 개의 상이한 손실률 (D1-D4) 과 연관된 자원 의존적 생장 등구배 곡선(실선). 점선은 이러한 상이한 손실률에서 이 종에 의해 소비되어야
식처 D1-D4 는 사망률 혹은 손실률이 서로 다른데. D1 은 손실률이 가장 낮고 D4 는 가장 높다. 단일 제한 자원에서와 똑같이 손실률 혹 은 교란율이 높은 서식처는 모든 제한되는 자원의 평형 수준이 높다. 5-2 자원 공급 경쟁의 결과를 예측하는 데 필요한 또 하나의 정보는 자원 공급 의 동태이댜 실제 서식처에서 자원 공급의 동태는 직접 관찰되어야 만 한댜 그러나 경쟁에 대한 상이한 자원 공급률의 효과를 나타내 기 위해서는 이론상 자원이 공급되는 단순한 규칙만 정의하면 된다 . 특정한 서식처에서 S1 과 &룰 각각 자원 l 과 자원 2 의 최대량이라고 하자. 점 (S i, S2) 은 자원 공급점이라고 불린다 . 각 서식처는 고정된 자원 공급점을 갖으며, 자원 공급률은 현재 이용 가능한 형태가 아 닌 그 자원의 양에 비례한다고 가정하자. Rj 는 이용 가능한 상태에 있는 자원 j의 양이고, sj 는 자원 j의 총량이라 할 때 dR/dt = a(Si - R) 로 표시되며, 이때 a 는 비율상수이다. 이 식은 특정한 공급점 (S1,S2) 과 자원의 이용도 (R i, R2) 를 갖는 서 식처에서 자원 1 과 자원 2 의 공급률을 정의하고 있다. 이러한 공급 률은 벡터로 표시할 수 있다 . 그림 2.8A 에서 자원 1 의 공급률은 벡 터 U1 으로, 자원 2 의 공급률은 벡터 U2 로 나타나 있다. 두 자원의 전 체 공급률 U 는 이 두 벡터의 합이다. 자원 공급률을 규정하는 이 식 은 자원 공급 벡터가 항상 공급점 쪽을 가리키고 있음을 의미한다. 그림 2.8B 에 있는 등구배곡선의 경우 자원 공급점은 둥구배곡선 상에 점으로 표시된 점에서 평형에 이를 것이다. 등구배곡선상의 그 점에서 개체군의 밀도는 일정하고(생장과 손실이 동일함), 자원 공급
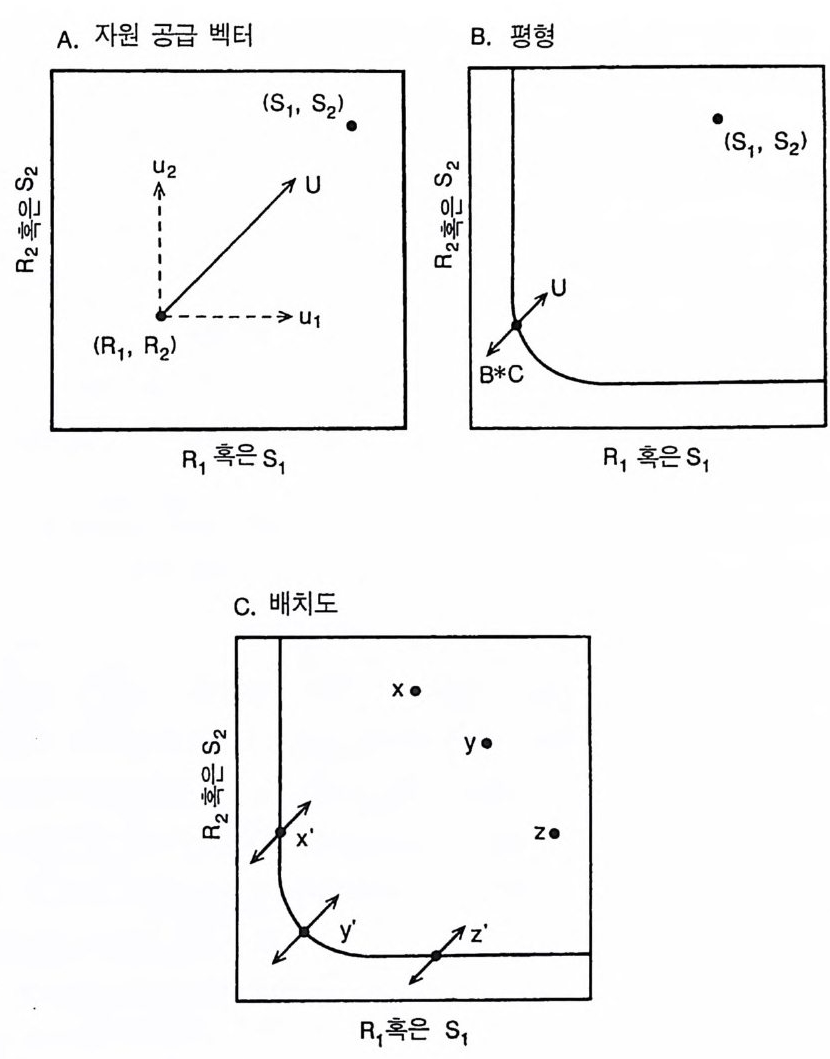 A. 자원 공급 벡터 B 펴0 형
A. 자원 공급 벡터 B 펴0 형
그림 2.8 (A) 자원 공급 벡터인 U 는 자원 공급점이 (Si, S 2 ) 인 서식처에서 이용 · 가 능한 R1 과 R2 의 공급률을 보여준다. (B) 평형 상태에 이르기 위해서는 B*C 로 표시되어 있는 전체 자원 소비율이 U 로 표시되어 있는 전체 공급 과 균형을 이루어야 한다. B* 는 평형 상태에서 전체 식물 생물량이다. (C) 자원 공급 벡터와 자원 소비 벡터는 특유한 자원 공급률을 갖는 각 서식 처 (x, y, 혹은 z: x, y 그리고 z 는 상이한 자원 공급점을 나타낸다)로 하여 금 자원 의존적 둥구배곡선상의 평형점 (x' , y' , z' )에 위치하도록 한다.
량 u 는 자원 소비량인 C 와 방향이 반대가 될 것이다. 개체군의 크 기(생물량)가 B* 수준에 도달하면 평형이 이루어질 것이다. B* 수준 에서는 개체군에 의한 전체 소비율이 전체 공급률과 정확히 균형을 유지한다 . 전체 소비율은 C·B * 로 표시된다. 그 이유는 C 가 단위 생 물량당 소비 벡터이기 때문이다. 따라서 평형 상태에서 C·B* +U=O 이 된다. 각 공급점은 그림 2.8C 에 나타난 바와 같이 생장 등구배곡선상의 특정한 점과 소비자 종의 평형 상태에 있는 생물량 과 관련이 있다. 특정한 서식처에서 종이 살아남기 위해서는(즉 자 신의 B * 를 0 보다 크게 하기 위해서는) 공급점이 그것의 등구배곡선의 바깥쪽에 있어야 한다. 5-3 종내 경쟁 부족되는 두 가지 자원에 대한 두 종의 경쟁에는 4 가지 경우가 있다(그림 2.9). A 종의 등구배곡선이 항상 B 종 안쪽에 있으면, A 종은 B 종에 비해 R1 과 R2 의 요구량이 적다. 따라서 A 종은 두 종이 모두 생존할 수 있는 모든 서식처에서 경쟁을 통해 B 종을 교체할 것이다 (그립 2.9A). B 종의 등구배곡선이 A 종 안쪽에 있으면(그림 2.9 B ), 모 든 서식처에서 B 종은 A 종을 교체할 것이다. 등구배곡선들이 교차할 때만 두 종의 평형점이 생긴다. 이 점은 그림 2 . 9C 상에 두 등구배곡 선이 만나는 점으로 나타난다. 만약 각 종이 모든 자원에 의해 균등하게 제한을 받을 수 있는 비율로 자원을 소비한다면 평형점은 안정할 것이다 . 서식처 유형은 동구배곡선과 각 종의 소비 벡터와 평행한 기울기로 평형점을 통과 하는 선에 의해 결정되는데, 죽 A 종이 우점하는 서식처, 두 종이 안 정을 유지하며 공존하는 서식처, B 종이 우점하는 서식처로 구분된 다. 이에 대한 설명은 다음과 같다 .
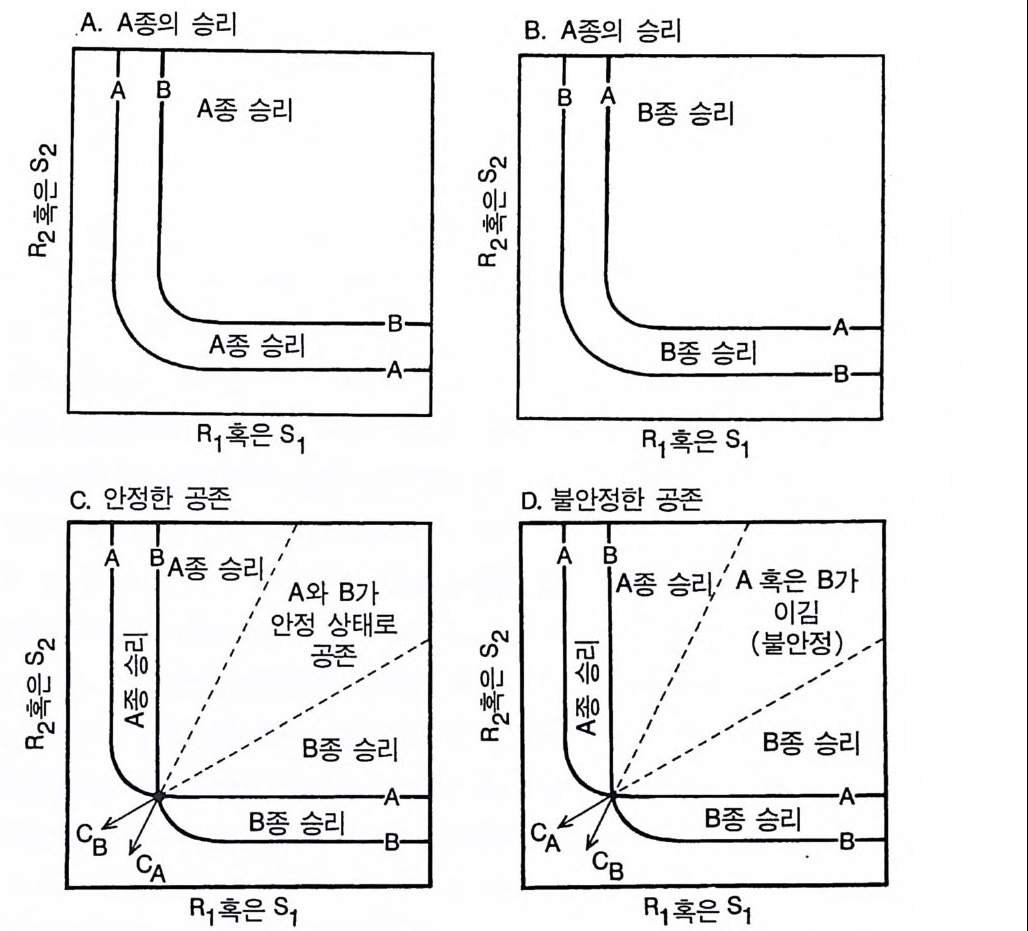 .A. A 종의 승리 B. A 종의 승리
.A. A 종의 승리 B. A 종의 승리
그림 2.9 A 와 B 로 표시된 굵은 실선은 A 종과 B 종의 자원 의존적 제로 순생장 등 구배곡선이다 . 이들 둥구배곡선의 위치와 소비 벡터의 위치가 각 서식처 에서 경쟁의 결과를 결정한다. (A) A 종은 두 종 모두 생존할 수 있는 모 든 서식처에서(자원 공급점) 두 자원에 대한 경쟁 우세종이며. B 종을 교 체한다. (B) B 종은 두 자원에 대한 경쟁 우세종이며. A 종을 교체한다 . (C) 둥구배곡선들이 두 종의 평형점에서 교차한다. 두 종의 자원 소비 벡 터와 조합하여 이 등구배곡선들은 종 A 가 이기는. 두 종이 공존하는. 종 B 가 이기는 서식지 조건을 결정한다. 각 서식처는 그것의 자원 공급점 (S1, S2) 에 의해 특징지어진다. 부호가 붙은 지역은 각 지역에 해당되는 공급점에서 예상되는 경쟁 결과를 보여준다. 이 경우 평형점은 안정하다. (D) 여기에서는 소비 벡터가 반대로 되어 있다 . 이것은 두 종의 평형접을 불안정하게 만든다. 이 지역에서는 두 종 중 어느 한 종이 이기는데. 이때 초기 조건에 의해 승자가 결정된다.
자원 R1 에 대하여 경쟁력이 큰 A 종은 두 종이 모두 R1 에 의해 제 한을 받는 서식처에서 우점한다. R 전] 대한 경쟁력이 큰 B 종은 두 종이 모두 자원 R 2 에 의해 제한을 받는 서식처에서 우점한다 . 중간 정도의 서식처에서 각 종은 서로 다른 자원에 의해 더 많은 제한을 받아 두 종이 모두 공존할 수 있다. 각 종은 평형 상태에서 자신을 더 많이 제한하는 자원을 더 많이 소비하기 때문에 안정 상태의 공 존이 유지된다 . 달리 표현하면, 종간 경쟁에 비해 종내 경쟁이 더 심 하고 그 결과 공존이 가능하다. 각 종이 제한 요인으로 작용하지 않 는 자원을 과량으로 소비할 때 4 번째 경우가 나타난다(그림 2.9D). 이처럼 자원 획득이 최적이 아니면 두 종의 평형점은 불안정하다. 자원 공급점이
양과 위치를 추정하는 데 유용하다. 실제로 이러한 자료는 지소의 성격이 다른 지역에 오래전에 목재용이나 펄프용으로 조성한 단일 수종 재배 지소에서 얻을 수 있다 . 영양염류 흡수율과 광 차단을 분 석하면 필요한 소비 벡터를 얻을 수 있으며, 이때 그림 3 .1 1 에서 제 시된 분석 방법이 사용될 수 있다. 동구배곡선이 추정된 종을 포함하고 있는 서식처의 경우 이 방법 에 의하면. 평형 상태의 식생 구성을 예측하기 위해서는 자원 공급 률의 측정이 필요하다. 그러한 예측은 자원 공급률이 일정하고, 초 식, 교란 그리고 다른 손실률이 일정하며, 또한 이들 요인들이 각 종 의 자원 의존적 생장 등구배곡선을 추정하기 위해 사용된 서식처에 서의 그것들과 동일하다는 가정을 기본으로 하고 있다. 또한 지표면 의 광량이 광 수직구배상의 어떤 지점의 광량보다도 중요하다는 가 정을 기초로 하고 있다 . 어떤 종이 우점종으로 될 것인지. 어떤 쌍이 공존하게 될 것인지 를 결정하기 위해서는 등구배곡선상에서 소비 벡터와 공급 벡터의 방향이 서로 다른 점을 찾아야 한다. 그림 2.9C 의 경우 자원 공급 벡터의 기울기가 A 종과 B 종의 소비 벡터 사이의 중간이라면 두 종 은 공존한다. 공급 벡터의 기울기가 두 종의 소비 벡터 사이에 위치 하지 않는다면 A 종이나 B 종 중 어느 하나가 경쟁에서 이기게 된다. 두 종의 평형점에서는 자원 공급 벡터가 A 종이 이기는 지역을 향 하면 A 종이 이기고, B 가 이기는 지역을 향하면 B 종이 이긴다. 이러 한 근사치는 여러 야외 조건에서 유용하다는 것이 입증되고 갔다. 다른 하나의 방법은 햇빛이 제한 자원이 될 때 오히려 바람직한데. 그림 3 . 11 에 제시되어 있다.
5-4 다종간의 경쟁 등구배곡선 방법에 의하면 다종간의 경쟁에서 어떤 패턴이 예상 되는가? 토양 자원과 햇빛에 대한 5 종의 경쟁을 생각해 보자(그림 2.1 0 ). 이 종들의 등구배곡선에 의하면 자원에 대한 이들의 경쟁 능 력이 분화되어 있음을 알 수 있다. 각 종은 자원의 특정한 상대적인 이용도에서 각기 경쟁력이 가장 높다. 각 종은 모든 자원에 의해 균 둥하게 제한을 받는 비율로 자원을 소비한다. 이러한 종들은 토양 자원의 공급률이 낮은 서식처로부터 높은 서식처에 이르는 구배상 에서 분리된다(그림 2.10). 평형 상태에서 자원의 수준은 등구배곡선상의 점까지 감소되어야 한다. 이것은 평형에 도달하였을 때 부족되는 토양 자원의 수준과 지표면에서의 햇빛의 강도 사이에 역상관 관계가 있음을 의미한다. 자원이 빈약한 토양(낮은 공급률)을 갖는 서식처는 영양염류의 수준 (농도)이 낮고, 식물의 생물량도 낮아 부족되는 토양 자원의 공급률 이 높은 서식처보다 지표면에 투과되는 햇빛이 많다(그림 2.1 0 ). 각 종은 자신의 동구배곡선의 코너에서 자원 공급률이 평형 수준에 이 르는 서식처에서 최대 생물량을 갖는다. 손실률이 동일한 서식처의 경우, 각 서식처는 영양염류의 수준은 낮지만 햇빛의 이용도는 높은 곳에서부터 영양염류 수준은 높지만 햇빛 이용도는 낮은 곳으로 이어지는 구배상의 어느 지점에 위치하 게 된다. 각각의 손실률에서 토양 자원:햇빛이 정성적으로 비슷한 구배가 나타난다. 그러나 부족되는 토양 자원을 일정하게 하고 손실 률을 변화시키면 영양염류와 햇빛에 대한 이 경쟁 모델은 영양염류 와 햇빛이 정의 상관을 갖는 구배를 예측할 것이다. 손실률이 낮은 서식처는 생물량이 많고. 영양염류와 햇빛의 이용도가 낮다. 손실률 이 높은 서식처는 두 자원의 이용도가 높다.
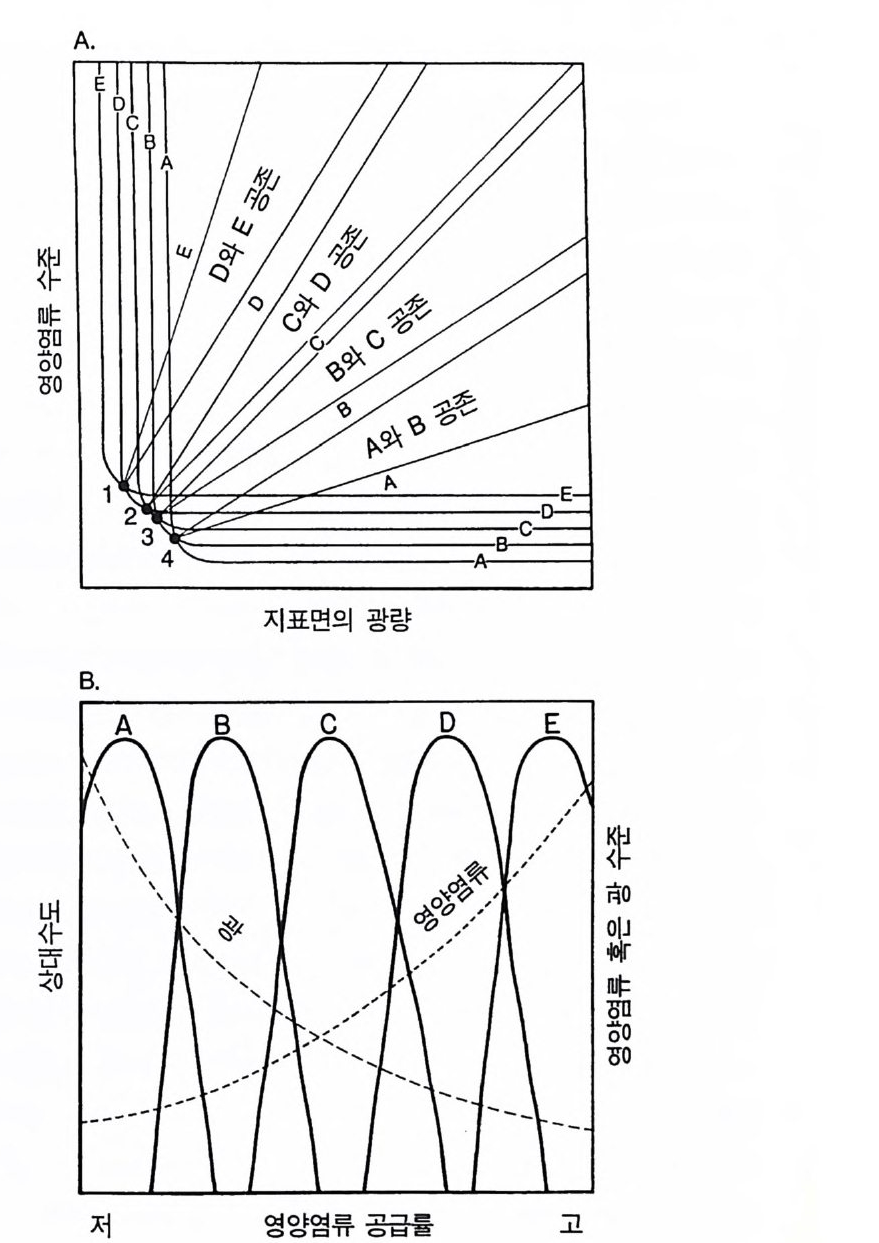 A
A
그립 2.10 (A) A 에서 E 까지 표시된 선들은 A-E 종의 제로 순생장 등구배곡선들이다. 번호가 붙어 있는 4 개의 점은 두 종의 평형접이다. 여러 종이 우점하거나 공존할 서식처의 유형이 나와 있는데. 이것은 자원의 최적 획득의 가정에 기초를 둔 것이다. (B) 이들 5 종은 평형 상태에서 토양의 영양염류 구배를 따라 그립에서 보는 것처럼 분리되어 야 한다 . 영양염류 공급률이 낮은 서식처는 A 종에 의해 우점되어야 하며, 영양염류 수준은 낮지만 지표면에 입사되는 광량은 많다. 영양염류 공급률이 높은 서식처에는 E 종이 우점하여야 하며. 영양염류 수준은 높지만 지표면에 입사되는 광량은 적다.
5-5 초강력 경쟁종과 대응책 영양염류와 햇빛에 대한 단순한 등구배곡선 경쟁 모델은 개체군 의 장기간의 생장률이 지표면에서의 햇빛 강도에 가장 큰 영향을 받는 것으로 가정된다. 그 이유는 지표면의 햇빛의 강도는 새롭게 정착하는 식물의 실생이나 가지에 영향을 주기 때문이다. 어느 지역 에 다수의 종들이 장기간 존속하는 이유를 이러한 방법으로 설명하 기 위해서는 다음과 같은 가정이 필요하다. (1) 서식처의 토양 영양 염류 공급률은 지점에 따라 공간적 변이가 있다. (2) 각 종은 부족 되는 토양 영양염류와 햇빛에 대한 요구 조건에서 부족되는 토양 영양염류에 대해 경쟁력이 강한 종은 다른 종에 비해 햇빛에 대한 경쟁력이 약한 쪽으로 분화되어 있다 . 이러한 가정 대신에, 일부의 종은 모든 자원에 대해 경쟁력이 약 하다거나 혹은 어느 한 종이 모든 자원에 대해 경쟁력이 강하다는 식으로 가정할 수도 있다. 모든 자원에 대해 일부 종의 경쟁력이 약 하다면 이들은 모든 서식처에서 자원 공급률과 무관하게 다른 종에 의해 교체될 것이다. 그런 경우에는 이 종들이 공간적으로 이질적인 서식처에서도 존속할 수 없을 것이다. 그 서식처에서 존속하는 종들 은 특정한 자원 공급률에 경쟁력이 강한 종들일 것이다. 다시 말하 면, 그림 2.10 에 나타난 바와 같이 대응책을 가진 종들이다. 만약 어느 한 종이 다른 종에 비하여 모든 자원에 대한 경쟁력이 크면, 이 초강력 경쟁종은 모든 서식처에서 다른 모든 종을 교체할 것이다 (T i lman 1982, 1986a). 그리고 종 사이에 분화가 없을 것이다. 그러나 그러한 초강력 경쟁종이 나타나면, 자연 선택은 그 초강력 경쟁종 내에서도 그들이 살고 있는 특정한 서식처 유형에 특수화된 개체들에게 유리하게 작용하여, 그 결과 새로운 초강력 경쟁종을 형 성하는 쪽이나. 그림 2.10 에서 가정한 것과 똑같이 특정한 자원비
(resource ra t e) 에 한정되는 다양한 종 혹은 유전자형을 형성하는 쪽 으로 진화가 일어날 것이다. 이러한 분화는 각 개체가 자신의 서식 처에서 자신을 제한하는 자원의 획득이나 사용 능력을 증가시키기 위하여 자신을 제한하지 않는 자원의 사용 능력의 일부를 포기함으 로 생길 수 있다 (T i lman 1982). 따라서 그들의 공존 이유를 설명할 수 있는 생물적 혹은 비생물적 제약이 없다고 가정하면 공존하는 종들은 부족되는 자원에 대한 그들의 경쟁 능력에 대응책을 가진 것으로 보아야 한다. 이 책에서 나는 자원에 대한 개체 혹은 종의 경쟁 능력응 설명하 면서 대응책을 몇 번 언급하였다. 각 경우에서 대응책을 가정하기 위한 논리적인 기초가 없다는 반론이 있을 수 있다. 그러나 그러한 모든 경우에 초경쟁적인 종의 출현을 기대하는 것은 가능한 일이다 . 그리고 초경쟁적인 종 개체군 내에서도 자신이 살고 있는 서식처에 서 경쟁력을 향상시킬 수 있는 형태적, 생리적, 생활사적인 특성(비 록 이러한 특성이 다른 서식처에서는 그 개체의 경쟁력을 감소시키는 경 우도 있겠지만)을 갖는 개체에게 자연 선택은 유리할 것이다. 나는 이 책 전체에서 이 점을 반복하여 거론하는 대신에, 내가 제시한 경 쟁적 대응책을 반대하기 전에 먼저 논리를 개발하도록 독자들에게 권하고 싶댜 다음 장에서 설명한 바와 같이 대응책을 유발시켜 현 재 지구상에 있는 다양한 종의 진화를 가능케 한 형태적, 생리적, 생 화학적 과정들을 우리는 이해할 수 있어야 한다. 영양염류와 햇빛에 대한 식물의 경쟁을 이와 같이 단순화시키는 것에 반대할 만한 여러 가지 이유가 있다. 이 이론에서 현실성이 가 장 적은 면 중의 하나는 지표면에서의 광량이 지상부의 다른 어느 지역의 광량보다 더 중요하다는 가정이다. 고착성인 다세포식물은 수직적인 광구배 속에 살고 있다. 햇빛의 강도는 일반적으로 지표면 에서 가장 낮고 위로 올라갈수록 증가한다 . 따라서 식물은 성장하면
서 그들의 잎이 위치하는 높이에 따라 상이한 광량을 받게 된다 . 게 다가 식물에 따라 성숙하였을 때의 높이가 다르다. 더구나 같은 종 내에서도 개개의 식물은 형태 • 생리 • 생활사의 특성이 가변적일 수 도 있다. 3 장과 4 장은 식물 생장과 경쟁에 관하여 형태적으로 분명 한 모델을 제시하고 있다. 이 모델은 더 복잡하고 따라서 좀더 폭넓 고 다양한 문제들(종자 크기의 진화, 최대 높이의 진화. 분배 패턴의 진 화. 최대 생장률의 진화 등)을 설명할 수 있지만, 이 장에서 제시된 광 범위한 일반적인 이론과 유사하다. 6 요약 이 장에서는 부족한 자원에 대한 식물의 경쟁을 이해하기 위하여 단순한 평형 상태의 접근 방법을 제시하고 있다. 식물이 필요로 하 는 많은 자원들은 영양 면에서 필수적이다. 필수 자원에 대한 최적 획득의 그래프 이론은 그러한 자원이 개체의 생장을 균등하게 제한 하는 비율로 소비되어야 한다는 것을 예측하고 있다. 식물은 비제한 적 필수 자원의 소비를 줄이는 쪽으로 자신의 형태를 조절함으로써 자원의 획득 노력을 조절할 수 있고, 그 결과 제한되는 자원의 소비 를 증가시킬 수 있기 때문에 식물의 자원 의존적 생장 둥구배곡선 은 직각인 코너가 아니라 둥근 코너를 갖어야 한다. 자원 획득 노력 을 조절하는 데 경비가 들지 않는 한 형태적으로 가변성이 있는 식 물 개체는 그렇지 않은 개체에 비해 경쟁력이 강하다. 식물에서 형 태적인 가변성이 혼한 것은 영양 면에서 필수적인 자원의 최적 획 득이론과 일치한다. 이 간단한 이론은 또한 육상 서식처에서 자원 이용도의 평형 패 턴에 중요한 영향을 주는 두 가지 요인, 즉 손실률(교란, 초식, 사망
에 의한)과 제한되는 토양 자원의 공급률을 보여주고 있다. 식물 생 장률은 자원의 농도에 따라 증가하기 때문에 손실률이 높은 서식처 는 손실률이 낮은 서식처에 비해 평형 상태에서 모든 제한되는 자 원의 수준이 높아야 한다. 따라서 식물이 분화될 수 있는 하나의 주 요 환경 구배는 손실률 혹은 교란율이 낮고 토양 자원과 햇빛 자원 의 수준이 낮은 서식처로부터 손실률 혹은 교란율이 높고 자원의 수준이 높은 서식처에 이르는 구배이다. 두번째의 주요 구배는 부족 되는 토양 자원의 공급률이 낮은 서식처로부터 높은 서식처에 이르 는 생산력 구배이다. 토양 자원의 공급률이 낮은 서식처는 영양염류 수준이 낮고, 식물의 생물량이 적으며, 지표면에 입사되는 광량은 많다. 반면에 공급률이 높은 서식처는 영양염류 이용도가 높고, 식 물의 생물량이 많으며, 지표면에 입사되는 광량은 적다. 이것은 식 물의 진화 과정에서 그들이 경험하는 두 가지 중요한 구배이다. 만 약 이것들이 중요한 구배라면 우리가 육상식물에서 볼 수 있는 대 부분의 형태적, 생리적, 생활사적인 패턴은 이 두 가지 축상에서의 경쟁에 대한 적응으로 설명할 수 있다. 이 문제는 3 장과 4 장에서 설 명하겠다.
제 3 장 영양염류와 햇빛에 대한 경쟁의 기작 1 서론 제 2 장에서 제시된 둥구배곡선 이론은 육상식물 사이의 자원 경쟁 과정을 개괄하고 있지만. 식물이 햇빛과 토양 자원에 대해 경쟁하는 형태적, 생리적. 생활사적인 기작은 포함되어 있지 않다. 등구배곡선 이론은 이러한 기작을 포함하고 있지 않기 때문에 부족되는 토양 자원의 특정한 공급률과 특정한 손실률을 갖는 서식처에서 식물의 어떠한 형태, 생활사 그리고 생리가 경쟁에 유리한지를 결정하는 데 이용할 수 없다. 이 장에서 나는 크기 구조를 갖는 식물 개체군 사이의 경쟁에 관 한 기계론적인 모델을 제시하고 경쟁의 평형과 동적인 특성 중 몇 가지를 설명하겠다. 다음 장에서 나는 식물의 특성이 손실률과 자원 공급률에 의해 어떻게 영향을 받는지를 결정하기 위해 이 모델을 사용하였다. 이 모델에서는 식물 개체의 생장 패턴이 현재의 광합성 산물을 뿌리, 잎, 줄기 혹은 종자의 생산에 분배하는 패턴에 의해 결
정된다는 것을 기본 전재로 하고 있다 . 이 모델을 설명하기 전에 분 배 패턴과 최대 생장률 사이의 일반적이면서도 매우 중요한 관계를 먼저 제시하겠다. 2 분배와 최대 생장률 4 억 년 전까지만 해도 육상의 광합성 식물은 남조류와 녹조류밖 에 없었다. 그때부터 현재까지 수백만 종 이상의 다세포 육상식물이 번창하였는데, 이 모든 종은 조상형인 일부의 조류에서부터 비롯되 었다 (S t ebb i ns&H ill 1980) . 이러한 육상식물 종은 형태나 생리, 생장 률, 그리고 생활사 면에서 매우 다양하댜 그러나 어떤 의미에서는 토양에 있는 조류와 거대한 세코이아 사이의 차이가 광합성 산물의 분배 패턴에 의해 야기되는 형태적인 차이라고 할 수 있다. 단세포 조류가 자신의 크기의 두 배에 해당되는 충분한 자원을 획득하면 그것은 두 개의 개체로 나뉜다 . 이러한 이분법은 이상적인 조건에서는 하루에 두 번도 일어날 수 있다. 반면에 세코이아의 실 생은 자신의 생장 능력의 대부분을 뿌리, 줄기, 잎에 분배하여 그것 의 전체 크기가 수백년 동안에 걸쳐 엄청나게 중가한다 . 결국 나중 에는 생산량의 일부를 종자에 분배한다. 식물은 종에 따라 생태학적 으로 중요한 생화학적, 생리적인 특성이 다르지만 단세포 조류로부 터 관속식물로의 진화 과정에서 가장 두드러진 특징은 뿌리, 줄기, 잎, 종자와 같은 식물의 기관에 대한 분배 패턴의 차이이다. 2-1 최대 생장률 현대의 관속식물에서 볼 수 있는 생활사의 패턴을 설명하기 위해
서는 그들의 조상과 그리고 조류와 관속식물 사이의 주요 차이점을 알 필요가 있댜 조류는 최대 생장률이 매우 높아 생물량이 하루에 자신의 생물량의 두 배까지 층가될 수 있다 (RGR ~ =1 .4 /da y, RGRm ax 는 이상적이고 제한이 없는 서식처에서의 최대 상대 생장률을 말하며. (dB/dt) x ( 1/ B) 로 계산된댜 이때 B 는 식물의 생물량이다) 육상의 관속식물 실생 중 가장 빨리 자라는 것은 1 년생 새포아풀 (Paa annua) 인데. 영양 기관의 최대 상대 생장률이 조류의 1/ 4 에 불 과하다 (RGR max = 0.3 8 /day )( Gr im e & Hunt 1975). 가장 천천히 자라는 것은 가문비나무 실생으로 최대 상대 생장률 RGRmax= 0 . 03/da y이다. 관속식물 진화의 초기에는 단세포조류에서 키가 작은 다세포식물로 발전되고 점차 키가 크고 가지가 있는 식물로 발전되었다 (S t ebb in s & Hi ll 1980). 어떻게 생장이 빠른 생물에서 생장이 느린 생물이 진화되는 것인 가? 식물 영양 기관의 최대 생장률이 이처럼 현저히 감소된 상태에 서 어떤 선택적인 장점이 식물에게 이득을 주는 것일까? 이 질문에 대답하기 위해서는 식물의 최대 생장률을 결정하는 힘을 알아봐야 한댜 그림 3.1 에 있는 식물 생장에 관한 간단한 모델을 살펴보자. 이론 적으로 식물체는 광합성 조직, 몸을 구성하며 양분을 수송하는 조 직, 영양분을 흡수하는 조직, 그리고 생식에 관계하는 조직에 대한 다양한 분배 패턴을 가질 수 있다. 관속식물의 경우 이러한 조직은 잎, 줄기, 뿌리, 그리고 종자에 해당된다. 이처럼 각 조직들로의 분배 패턴은 어떻게 식물의 영양 기관의 최대 생장률에 영향을 주는가? 최대 생장률은 식물이 이상적인 서식처, 즉 어떠한 자원에 의해서 도 제한을 받지 않는 서식처에서 보이는 가장 높은 생장률이다. 이 러한 서식처에서 식물의 생장은 지수함수적으로 나타난다. 생산된 것의 재투자를 통해 계속 생장 물질을 합성하기 때문에 지수함수적
 줄기 (S)
줄기 (S)
인 과정이 된다. 현재의 모든 생장이 새로운 광합성 조직의 생산에 분배된다면(영양염류와 광이 풍부한 서식처에서 살고 있는 단세포조류 의 경우) 최대 광합성률을 갖는 식물은 가장 빠른 영양 생장률에 도 달할 수 있을 것 이 다 (Mons i 1968). 그러 나 식 물이 광합성 산물의 일 부를 광합성 조직의 생산에 사용하지 않는다면 최대 생장률은 감소 할 것이다. 따라서 자신의 생장의 일부를 줄기나 뿌리, 혹은 다른 비 광합성 조직에 분배하는 식물은 생장의 전부를 광합성 조직의 생산 에 분배하는 식물에 비해 최대 영양 생장률이 낮다. 이것은 다음과 같은 미분방정식으로 설명할 수 있다. 이 미분방정 식은 그림 3 .1에 있는 흐름도를 수학적으로 간단히 구체화시킨 것이 다. 먼저 식물체가 자신의 생장을 모두 광합성 조직에 분배하는 경 우를 알아보자. B 를 식물의 전체 생물량이라 하고 L 은 광합성 기관
(잎)의 생물량이라고 하자. 식물의 생물량 변화, 죽 dB/d t는 dB/dt = LPm -r B ( 3. I ) 로 주어진댜 이때 P,,, 은 최적 조건하에서 단위 광합성 조직당 최 대 광합성률이며, r 는 단위 생물량당 호흡률이다. 여기에서 식물체는 오직 광합성 조직으로만 되어 있기 때문에 B=L 이 된다. 따라서 이 식은 다음과 같이 변형된다. RGRmax = dB/Bdt = pm - r (3.2 ) dB/Bd t는 이 식물의 최대 영양 생장률이다. 이 방정식에 의하면 식 물의 최대 생장률은 자원이 포화되었을 때의 광합성률과 호홉률의 차이와 같다는 것을 나타내고 있다. p n, 과 r 가 주어진 식물에서 생산 량의 일부를 줄기나 뿌리와 같은 비광합성 조직에 분배하면 최대 영양 생장률은 어떤 영향을 받는가? 우리는 최대 생장률에 관심이 있기 때문에 다시 자원의 제한이 없는 서식처에 살고 있는 식물체를 생각해 보자. S 를 식물의 전체 줄기의 생물량, R 를 뿌리의 양이라고 하자. 그러면 B=R+S+L 이 된 댜 식물의 전체 생물량인 B 중에서 L 은 광합성 조직의 양인데. L=B ― R ― S 이다. B-R-S 를 L 대신에 대입하면 방정식 3.1 은 dB/dt= (B-S-R)Pm-rB (3.3) 가 된다. 그리고 RGRmax= dB/Bdt = Pm[ 1-(R +S)/B] 규 (3. 4 ) 또는 RGRmax = (UB)Pm-r (3.5)
로 된댜 방정식 3 .4는 무엇을 의미하는가? (R+S ) /B 항은 전체 생물 량 중 비광합성 조직인 줄기와 뿌리로 분배되는 비율이다. 따라서 이상적인 환경 조건하에서 이 식물의 최대 영양 생장률은 전체 생 물량 중 비광합성 조직으로 분배되는 비율에 의해 결정된다(방정식 3.5). 방정식 3.4 에서 도출해 낼 수 있는 간단하지만 중요한 결론은 광합성 조직이 아닌 다른 어떤 조직으로의 분배는 필연적으로 식물 의 최대 생장률의 감소를 초래한다는 것이다. 이러한 예측에 대한 실험적인 증거가 있는가? G ri me 과 Hun t (1975) 는 영국의 Sheff iel d 지역에서 132 종의 현화식 물 실생의 최대 생장률을 조사하였다 . 그들은 강광하에서 영양염류 룰 충분히 공급하는 사경 재배를 하였는데 , 하나의 용기에 1 개체씩 심어 5 주 동안 키웠다. 이들의 실험 결과를 그림 3.2 에 요약하였는 데, 나는 Claph a m 둥 (1962) 이 제안한 생장형(협엽초본, 광엽초본, 콩 과식물 혹은 목본식물) 및 생활사(1년생 혹은 다년생)와 관련하여 RGRmax 값을 사용하였다. 다년생 기관(목본 줄기, 목본 뿌리 , 에너지와 영양염류 저장기관, 눈)에 자원을 분배하지 않는 1 년생 식물의 실생 은 다년생 초본에 비해 최대 생장률이 현저히 높았다. 그리고 다년 생 초본은 다년생 목본에 비해 최대 생장률이 높았다. 이와 같이 1 년생 초본 —’ 다년생 초본 —> 다년생 목본 순으로 최 대 생장률이 변화하는 것은 식물에 따라 광합성 산물을 비광합성 조직에 분배하는 데 차이가 있음을 의미한다. 생장이 가장 느린 다 년생 목본식물은 광합성 산물의 대부분을 목본성 뿌리와 줄기의 생 산, 다년생 조직의 생산 , 그리고 생육계절의 초기에 생장을 위한 에 너지 저장에 분배한다. 뿌리, 잎, 그리고 줄기의 손실률과 호홉률의 차이는 식물의 분배 패턴이 이러한 기관에서의 생물량 비율과 매우 다를 수 있음을 의미하지만 , 목본식물은 전체 생물량이 줄기에 70- 90%, 뿌리에 9-25%, 잎에 1-6% 가 분배되어 있다 (K i ra 1964,
 2.52.0.15
2.52.0.15
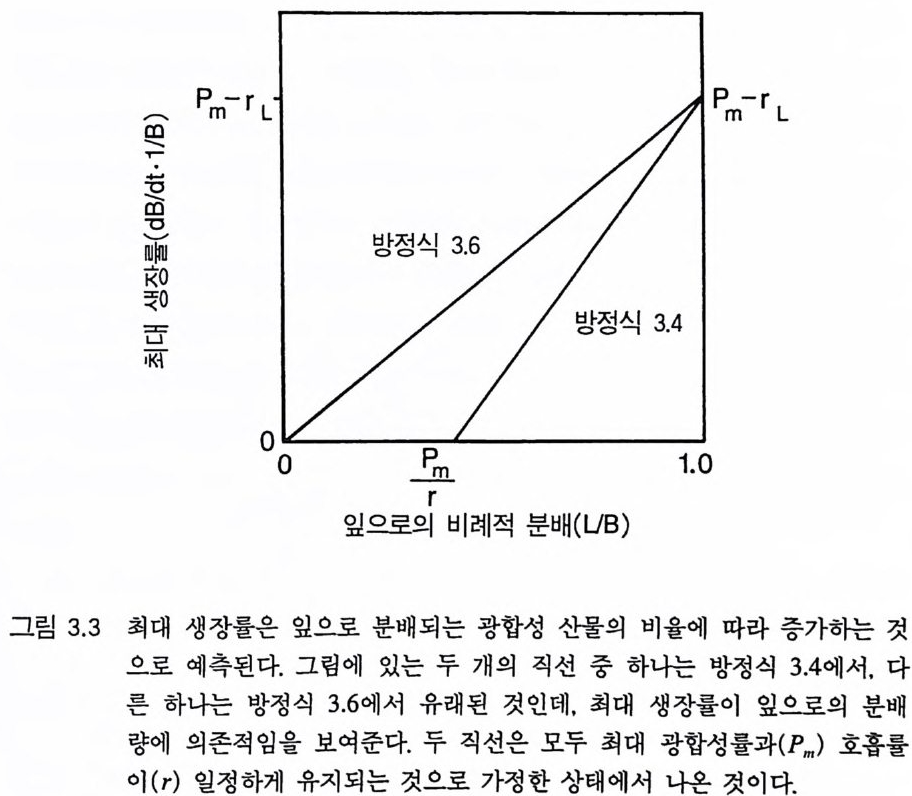
Whit tak er 1975). 이러한 형태를 갖는 식물은 상대 생장률이 낮다. Tamm 과 Aronsson(1982) 은 가문비나무에 시비를 한 후 생장률이 증가하는 것은 잎에 분배되는 광합성 산물의 양이 증가하고 세근에 분배되는 양이 감소하는 것과 일치함을 알았다 . 가장 빨리 자라는 식물군인 1 년생 식물은 실생 때 생산량 중 잎에 분배하는 비율 (50- 70%) 이 높았다 (Ha rpe r & Og de n 1970, Howrath & Wi lliam s 1972). 따라 서 G ri me 과 Hun t에 의하여 제시된 자료는 방정식 3 .5에서 예측되는 것과 정성적으로 일치한다. 방정식 3 .5에 의한 예측을 직접 정량적 으로 검증하는 데에는 이러한 종들의 분배 패턴에 대하여 현재 이 용 가능한 것보다 더 많은 정보가 필요하다. 그러나 Gri me 과 Hun t (1975) 는 한 그룹 내에서 생장이 느린 종은 뿌리에 분배량이 크다는 것을 알았다. 더구나 다년생 콩과식물은 다
른 다년생 초본에 비해 최대 생장률이 현저히 낮았다(그림 3.2). 콩 과식물은 질소고정세균과 공생관계를 유지하기 위해 이 세균에게 광합성 산물을 제공하여야 한다. 이와 같이 광합성 산물을 광합성 조직의 생산이 아닌 다른 기능에 분배하기 때문에 콩과식물의 최대 생장률이 낮고 비옥한 토양에서 진행되는 2 차 천이 과정에서는 이 들을 볼 수 없다(제 7 장). Gr i me 과 Hun t의 자료뿐만이 아니 라 이 보다 전에 발표된 Ja rv i s 와 Jar vis ( 1964), Monsi( 1 968), Loach(1970) 의 자료를 분석해 보면 광합 성 산물을 잎이 아닌 다른 기관에 분배하는 것은 일반적으로 식물 의 최대 생장률을 감소시킨다는 것을 알 수 있다. 최대 생장률의 차 이는 생태학적으로 매우 중요하다. 그 이유는 제 6 장에 설명되어 있 는 것과 같이 식물의 최대 생장률은 실험적인 조작이나 생물적 혹 은 비생물적 교란과 같은 그들의 환경 변화에 대한 단기간의 반응 동태를 결정하는 주요 인자이기 때문이다. 방정식 3.5 는 식물의 최대생장률이 그 식물의 형태에 의존적임을 보 여주고 있다. 이 식에서는 잎, 뿌리, 그리고 줄기의 단위 생물량당 호흡 률이 같고 이들의 순환률이 비슷하다고 가정하고 있다. 이것은 현실적 인 가정은 아니다. 그러나 방정식 3 .4의 정성적인 예측은 아주 완벽하 기 때문에 이러한 요인들이 포함되더라도 예측 결과에는 변함이 없다. 예를 들어 뿌리와 줄기는 잎에 비해 호흡률이 낮다. 잎의 호홉률 이 rL 이고 줄기와 뿌리의 호흡률이 0 인 극단적인 경우를 가정해 보 자. 자원이 충분한 서식지의 식물은 이 경우 RGRmax= dB/Bdt = LIB(Pm-r> (3.6 ) 이 되는데, 잎에 분배되는 모든 비율에서 방정식 3.5 보다 RGRmax 가 크다. 잎은 줄기에 비해 순환율이 높다. 이러한 순환율은 호흡과 같 이 지출경비로 작용하기 때문에 잎당 순생산량이 감소된다. 방정식
3 .4를 방정식 3.6 과 비교해 보면(그림 3.3), 모든 조직의 호홉률 혹은 순환율이 같다는 가정과(방정식 3.4 ), 잎사귀만이 호흡이나 순환에 필요한 지출경비가 든다는 가정(방정식 3.6) 은 RGR ~ 가 분배 패턴 에 의존적인 경계를 제공함을 알 수 있다. 기관에 따른 호흡과 순환 경비가 다르기 때문에 직선적인 관계를 기대하기는 어렵다. 그러나 실질적인 기능은 그림 3.3 의 두 선 사이에 경계가 그어져 최대 생장 률이 잎에 분배되는 비율에 따라 증가한다는 것을 보여주어야 한다. 그림 3.3 의 관계를 다른 종에 적용하기 위해서는 영양염류와 햇 빛이 포함된 환경에서 종에 따른 형태의 차이가 단위 생물량당 광 합성의 차이 또는 단위 생물량당 호홉률의 차이보다 훨씬 크다는
 ..-,. •P m - r' L Pm - r• L
..-,. •P m - r' L Pm - r• L
가정이 필요하다. 육상식물은 기능적으로 거의 100% 잎으로 되어 있는 토양의 조류에서부터 잎이 1% 이하인 수관층 수에 이르기까 지 그 범위가 매우 넓다. 주어진 지리적인 지역범위 내에서 야외 조 건하의 최대 광합성률의 차이는 10 배 이하이다 (Chabo t & Mooney 1985). 그러나 형태 다음으로 최대 생장률에 중요한 영향을 주는 요 인은 최대 광합성률이라고 할 수 있다. 생리적인 면보다는 형태적인 면에서 유전적인 변이가 더 많은 것처럼 보인다. 다양한 곡류 중 수 천년 동안의 선택 과정을 통해 형성된 생산력이 높은 재배종은 생 리에는 별로 영향이 없었고 분배 패턴과 형태에 주로 영향을 받았 다 (F itt er 1986). G iffo rd 와 Je nki ns (1 981) 는 식물의 최대 광합성률보 다 잎으로 분배되는 양이 곡류의 생장률을 결정하는 데 더 중요하 다는 것을 알았다. 다른 조건이 모두 똑같다면 최대 생장률이 높은 식물이 낮은 식 물에 비해 유리할 것이다. 그러면 생장이 빠른 식물로부터 어떻게 생장이 느린 식물이 진화되었는가? 광합성 조직이 아닌 다른 기관 에 자원을 분배하는 식물 형태의 진화를 설명하기 위해서는 자원이 제한되는 환경에서 그 식물이 생활할 때 비광합성 기관으로부터 어 떤 이득을 얻을 수 있는지를 알아볼 필요가 있다. 자원이 제한되지 않는 환경의 경우 다세포성이나 비광합성 조직의 진화는 그러한 조 직이 손실이나 사망을 감소시키지 않는 한 자연 선택에 유리하지 않다. 3 ALLOCATE: 크기 구조롤 갖는 식물 개체군에서 경쟁의 모델 이러한 문제들을 설명하기 위한 유일한 방법은 식물 환경의 복잡
성뿐만이 아니라 식물의 형태, 생리, 생활사의 중요한 면이 포함되 는 모델을 만드는 것이다. 그러나 식물이 갖는 이러한 특성들은 매 우 복잡하기 때문에 이들을 다 포함시키려면 수학적으로 복잡한 모 델이 필요한데, 이러한 모델은 아주 특이한 경우에서만 분석적으로 다룰 수가 있다. 나는 ALLOCATE 라고 부르는 동적인 모델을 개발 하였는데, 이 모델은 식물에게 매우 중요한 것으로 여겨지는 몇 가 지 특징을 포함하고 있다. 모델에 포함되어 있는 요소들은 (1) 실생으로부터 출발하여 계속 적인 영양기관의 생장을 통해 그들의 최대높이에 도달하는 크기 구 조, (2) 뿌리, 잎 줄기에 분배되는 광합성 산물의 양에 의해 결정되 는 영양 생장, (3) 식물의 햇빛 차단에 의해 형성되는 수직적인 광 구배, (4) 영양염류 및 햇빛 의존적인 광합성, (5) 무기화율과 식물에 의한 영양염류 흡수율로 결정되는 토양의 영양염류 이용도, (6) 식 물이 생산하는 종자의 수와, 발아 후 실생의 키, 뿌리의 양, 잎의 양, 줄기의 양에 영향을 주는 종자의 크기, (7) 뿌리, 잎, 줄기의 상이한 호흡률, (8) 밀도 독립적인 손실률(특히 사망) 등이다. 같은 종 또는 다른 종이든 간에 모델에서 가정하는 식물 개체 사이의 상호 작용 에 대한 유일한 기작은 이들이 자원을 소비함으로써 야기된다. 따라 서 이 모델에서 전재하고 있는 직접적인 밀도 의존적인 과정은 없 다. 오히려 각 개체는 햇빛의 수직 분포와 토양의 영양염류 이용 도에 영향을 주어 자기 자신의 생장과 다른 개체의 생장에 영향을 준댜 이 모델은 개념적인 면에서 피음에 의한 경쟁과 영양염류 소비에 의한 경쟁을 구별하지 않는다. H arp er {1 977) 가 지적한 바와 같이 이 두 과정 사이에는 차이가 있지만 이러한 차이는 이 모델에 직접 포 함되지 않았다. 그 이유는 각 식물의 높이와 그 식물이 살고 있는 광 환경이 모델의 일부이기 때문이다. 이 모델을 단순화시키면 서식
처에 수직적인 구조와 수평적인 구조가 없다는 것이다. 따라서 어느 지점에서든지 모든 식물은 똑같은 토양 영양염류의 농도에 접하고 있고. 주어진 높이에 있는 모든 식물은 똑같은 광량에 노출된다는 것이댜 이것만으로 본다면 이 모델은 토양 조건이 균일하고 숲톰이 없으며, 이웃과의 경쟁이 없는 작은 군반 내의 식물 동태에 관한 것 으로 생각할 수 있다 (Pacala & Sil a nder 1985). 이 모델은 다른 모델들 과 마찬가지로 현실을 묘사한 것이다. 이러한 모델을 만드는 목적 중의 하나는 등구배곡선 모델에서와 같이 복잡성을 무시한 단순한 모델이 얼마나 타당성이 있는지를 알 아보고 그것으로부터 얻을 수 있는 일반적인 패턴을 찾는 것이다. 또 하나의 목적은 식물이 성숙하였을 때의 형태, 종자의 크기, 그리 고 높이가 주요 환경 구배에 따라 어떻게 변하는지를 알아보는 것 이다 . 세번째 목적은 식물의 형태 및 생활사가 자연적인 교란 혹은 실험적으로 가해진 조작 후에 일어나는 군집 변화의 동태와 어떤 관련성을 갖는지를 알아보는 것이다 . 그러나 모의 모델 (s i mula ti on models) 은 결코 세련된 것이 아니다. 이 모델들은 유능한 수학자가 분석적안 방법으로 해결할 수 있는 영역을 넘어서고 있다. 이 책의 제한된 범위 내에서 그리고 내가 슈 퍼컴퓨터를 이용할 수 있는 제한된 시간 범위 내에서 이 모델에 있 는 모든 변수들 사이의 상호 작용을 찾아내고 그것들의 동적인 기 능을 상세히 밝혀내기란 불가능하다. 그러나 이 모델을 통해 얻을 수 있는 것도 많이 있다. 내가 생물 학적으로 현실성이 있는 쪽으로 ALLOCATE 의 실제 형태를 변형시 키더라도 다음 장에서 제시된 일반화가 변하리라고는 생각하지 않 는다. 그 이유는 ALLOCATE 가 균일한 토양, 하나의 부족되는 토양 자원, 식물 자신에 의해 형성되는 수직적인 광 구배, 그리고 밀도 및 크기 독립적인 사망률을 갖는 서식처의 크기 구조를 갖는 식물 사
이에 일어나는 경쟁의 중요한 면을 포함하고 있기 때문이다. 제 9 장 다음에 실린 부록에 식물의 분배와 생장 모델의 수학적인 면을 자세히 제시하였다. 나는 이 모델의 주요 가정들을 요약하고 이러한 가정들의 논리적인 결과를 제시하겠다. 이런 과정을 통해 나 는 독자들로 하여금 모델의 목적을 상기하도록 하고 싶다 . 모든 모델은 일련의 가정을 예측으로 전환시키는 논리적인 도구 이다. 수학은 이러한 전환 과정에 사용되는 도구일 뿐이다. 수학이 정확하다고 가정하면 모델의 중요한 특성은 그것의 가정과 예측이 다. 만약 모델이 예측한 내용이 실험이나 관찰을 통해 틀린 것으로 밝혀진다면 그 모델의 기본 가정 중 하나 이상이 잘못된 것이다. 하 나의 종 내에서도 개체에 따라 높이가 다르고 따라서 상이한 광량 에 접하는 복잡성 때문에 나는 이 모델을 분석적으로 해결하는 방 법을 찾지 못하였다 . 그러나 내가 제시한 생물학적인 가정과 연관이 있는 예측을 결정하기 위해 모의를 이용하였다 . Ha rp er(1977) 가 누차 강조한 바와 같이 식물은 토양 영양염류와 햇빛을 얻기 위해 경쟁한다. 식물의 키는 햇빛에 대한 경쟁력을 결 정하는 주요 인자이기 때문에 식물 경쟁의 기계론적 모델은 식물의 키를 다르게 하여 그들이 받는 햇빛의 양이 다르도록 하여야 한다 . 식물은 실생이나 줄기 혹은 포복경으로 생활을 시작하여 영양 기관 의 생장을 통해 자란다. 식물은 키가 커짐에 따라 더 많은 햇빛을 받을 수 있다. 이와 유사하게 근계가 큰 식물은 부족되는 토양 자원 올 더 많이 획득할 수 있다. ALLOCATE 모델에서는 각 식물 개체가 종자로부터 시작된다 (그림 3. . 4). 발아가 되면 종자에 있던 에너지와 영양염류가 뿌리. 줄 기, 잎을 생산하는 데 사용된다. 종자가 뿌리, 줄기. 잎에 분배하는 생물량은 환경 조건과 무관하게 고정되어 있고, 영양 생장을 하는 동안에 광합성 산물을 이러한 기관에 분배하는 비율과 같다. 식물은
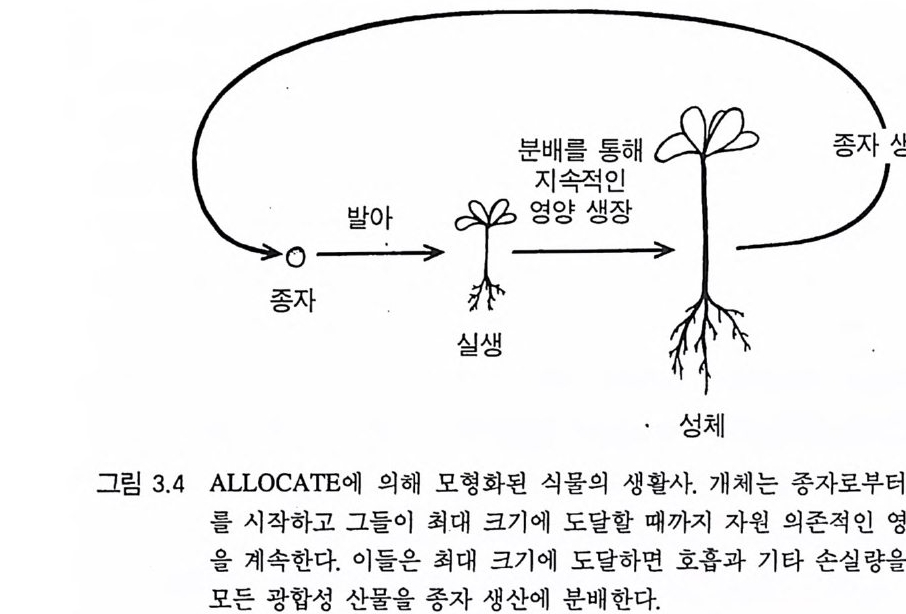 종 - 二 露 二雪• 성체 一종자생산
종 - 二 露 二雪• 성체 一종자생산
최대 높이에 도달할 때까지 영양 생장을 계속한다 . 최대 높이에 도 달하면 이 모델은 광합성 산물 중 호홉에 사용되고 남은 양은 모두 종자 생산에 분배되는 것으로 가정한다. 이와 같이 영양 생장으로부 . 터 생식으로의 전환 과정은 많은 식물에서 볼 수 있다. 1 년생 식물 의 최적 생장 모델에서도 이와 같은 전환 과정이 나타나는 것으로 예측되 었다 (Iwasa & Roug hg ar den 1984). ALLOCATE 는 생물의 모든 생활사 단계에서 밀도 독립적인 사망 이 있는 것으로 가정하고 있다. 밀도 독립적인 사망은 크기 독립적 이며 자원 독립적인 사망을 의미한다. 이 가정은 단순하기 때문에 유용성이 있다. 영양염류 이용도가 높은 조건하에서는 광합성이 광 의 제한을 받을 수 있다. 이때 광합성은 광량의 Mich aelis - Mente n 함수로 가정된다. 강광 조건하에서 광합성은 영양염류에 의해 제한 을 받는데, 이 때 광합성은 상대 적 인 영 양염 류 이용도의 Mich aeli s- Mente n 함수로 가정된다. 상대적 영양염류 이용도는 토양에 있는
영양염류의 농도에 잎 무게에 대한 뿌리 무게의 비를 곱한 것으로 정의된다. 더구나 부족되는 영양염류와 햇빛은 광합성률을 조절하는 완전히 필수적인 자원으로 가정하고 있다 . 이것은 부족되는 영양염 류의 수준이나 광량의 수준에 의해 광합성률이 결정됨을 의미하는 데. 어느 것이든 광합성률을 낮게 한다. 모델에서는 주어진 기간 동안에 생산된 광합성 산물이 그 기간 동안에 존재하는 조직의 호홉에 맨 먼저 사용되고, 광합성 산물의 나머지가 새로운 뿌리, 줄기 , 잎(혹은 그 식물이 최대 높이에 달했을 경우에는 종자)의 생산에 분배된 것이라고 가정하고 있다. 호흡량이 광합성량보다 많으면 식물은 휴면하는 것으로 간주된다(Fitt er 1986). 휴면중인 식물도 여전히 사망할 수 있다. 여러 가지 기관으로 분배 되 는 광합성 산물은 뿌리 로의 분배 (AR), 잎으로의 분배 (AL), 줄기 로의 분배 (AS) 비에 의해 결정된다. 잎이나 줄기로 광합성 산물을 새로 분배하면 잎과 줄기의 생물량이 증가함으로 키가 작은 모든 개체들의 광 환경도 변화한다. ALLOCATE 에서 한 개체가 받는 광량은 그 개체의 잎의 높이와 다른 모든 식물체 생물량의 수직적 분포에 의해 결정된다 . 모든 잎 은 줄기 꼭대기에서 한 층으로 형성되는 것으로 가정된다. 잎의 높 이는 줄기의 높이로 결정되고, 줄기의 높이는 줄기의 생물량의 함수 로 가정된다. 사용된 함수는 구조적 제약 (McMahon 1973), 식물의 높 이와 줄기의 생물량 사이의 상대 생장 관계 (K ir a 1964) 에 근거를 두 고 있댜 또한 초본식물에 관한 것 중에는 내 자신이 관찰한 자료도 포함되어 있다. 각 조직은(뿌리, 줄기, 잎, 종자) 각기 특유한 영양염류 함량을 가진 댜 영양염류는 생산되는 뿌리, 줄기. 잎, 종자로 인하여 새로 생기는 필요량을 충족시킬 수 있는 양만큼 흡수된다. 이것은 현재 필요한 양보다 영양염류가 과도하게 소비되는 것을 막는다 . 각 조직의 영양
염류 요구량은 그 조직의 단위 생물량당 필요로 하는 영양염류의 양과 새로 생산된 조직의 양을 곱하여 얻는다. 부족되는 토양의 영양염류는 균등한 방법으로 공급되는 것으로 가정된다 (T i lman 1982). 죽 부족되는 영양염류의 공급률은 그 영양염 류의 무기화가 가능한 전체량에 비례한다. 이것을 전체 영양염류 (TN) 로 부르겠댜 영양염류 공급률이 부족되는 영양염류의 전체량 에 항상 비례적이기를 바라는 것은 아니다 (Pas t ore t a l. 1984). 이것은 질소나 인과 같은 토양 영양염류의 공급 동태를 파악하기 위한 유용 한 가정으로 이용되어야 한다. 왜냐하면 이러한 영양염류는 이용 가 능한 형태로 전환될 수 있는 불용성 형태를 가지고 있기 때문이다. 물의 경우에도 유용한 가정이 될 수 있다. 물의 공급률은 강우량 과 토양의 전체 포장용수량 중 사용되지 않은 부분에 의해 결정되 기 때문이다. 식물이 많은 양의 토양 수분을 감소시킬수록 재충전되 어야 할 토양 수분의 양은 많아질 것이다. 이러한 영향은 질소나 인 산의 공급률이 전질소와 전인산량에 의존적인 것과 똑같이 토양 수 분의 재충전율이 강수율에 의존적이도록 하는 원인이 된다. 영양염 류의 공급률이 대체적으로 전체 영양염류의 양에 비례한다고 생각 하면 전체 영양염류의 양은 부족되는 토양 자원의 공급률 측정치로 간주될 수 있다. 모델은 각 종 혹은 표현형의 동시 출생 집단이 동시에 생장하는 것을 전제로 한다. 그들이 생장함에 따라 영양염류의 이용도는 감소 된다. 광은 식물의 잎과 줄기에 의해 Beer 의 법칙에 따라 감소된다. 수관층 밑의 주어진 지점에서 잎에 도달하는 광합성 유효방사량의 비율은 전체 식물의 생물량과 음의 지수함수 관계에 있다. 매일매일 증가되는 시간을 사용하여 수리적으로 모델을 . 설명할 수 있다. 미네소타의 기후에서 연중 생육 기간은 140 일이다. 주어진 기간 동안에 각 종의 동시 출생 집단은 그 서식처의 햇빛과 영양염류 환
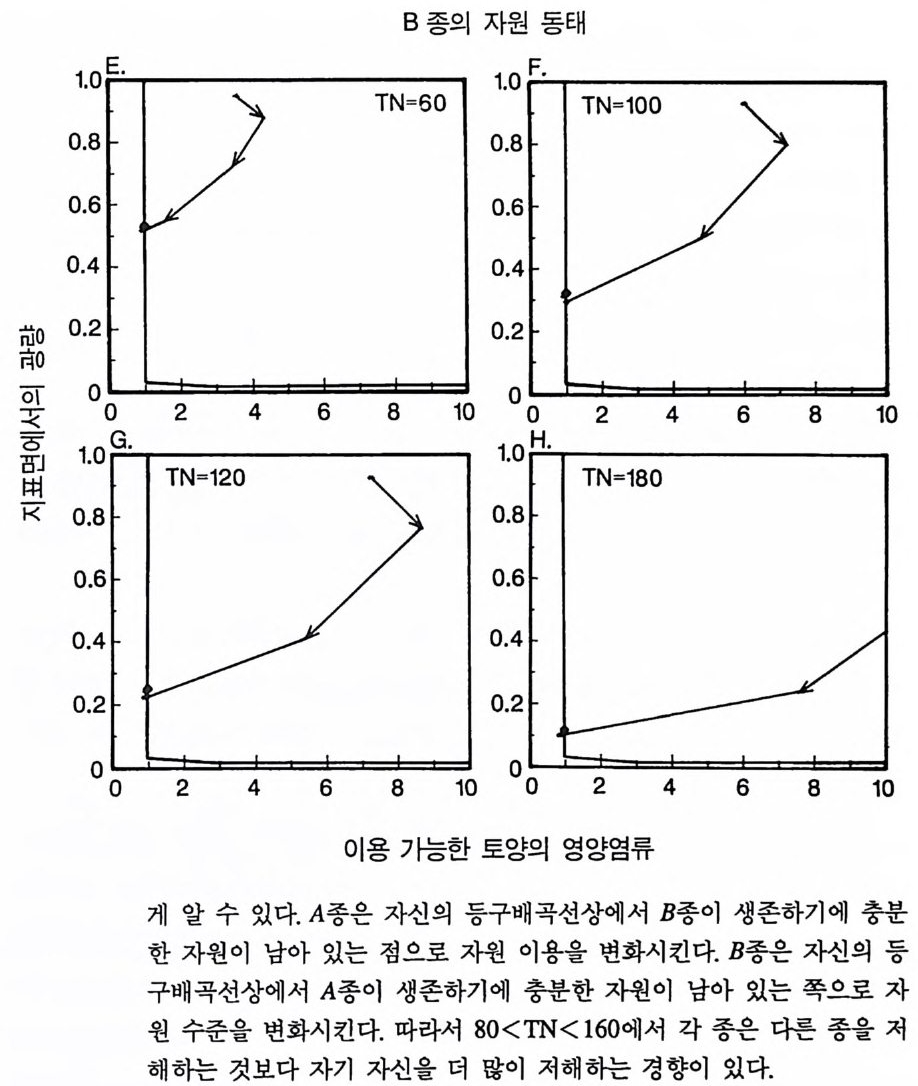
경에 따라 생장한다. 다음 기간의 햇빛과 영양염류의 환경은 그 식 물들이 홉수한 영양염류와 줄기 및 잎 생산의 누적적인 효과에 의 해 설정된댜 이러한 과정은 평형에 도달할 때까지 반복된다. 모델 에는 계절성이 포함되지 않고 오히려 연속적인 생장을 전제로 한다. 따라서 종간 상호 작용과 종내 상호 작용의 유일한 기작은 자원의 소비를 통해서 일어난다는 것을 가정하고 있다. 모델은 일반성을 갖도록 만들어졌다. 각 종은 자기 자신의 분배 패 턴, 성숙하였을 때의 높이, 종자 크기, 최대 광합성률, 광합성을 위한 햇빛과 영양염류의 반포화상수, 그리고 조직의 영양염류 농도가 주어 진다. 대부분의 모의에서 나는 토양 영양염류 중 질소가 부족한 초지 종에 대표적인 변수들을 사용하였다. 따라서 최대 높이는 2.5m, 최소 높이는 0.6cm 를 사용하였고, 최대 생장률은 G ri me 과 Hun t(1 975) 가 초본종에서 보고한 범위를, 영양염류 제한 생장률은 Til m an(1986b) 에 의해 보고된 것과 비교할 만하다. 그러나 이러한 변수의 선택은 모델에 의해 예측되는 패턴의 성질에 영향을 주지 않는 것이다. 3-1 단일종의 생장 먼저 특정한 환경에 처해 있는 단일종의 생장 동태를 살펴보자. 선별된 변수에 따라 두 가지의 서로 다른 생장 반응이 나타날 수 있다. 첫째, 다른 변수들이 평행 상태에 점근하면 개체군도 평형 상 태의 밀도에 점근할 수 있다. 둘째, 개체군은 전체 밀도와 자원 수준 에서 일련의 변동을 보일 수 있다. 먼저 영양염류 공급률이 매우 낮은 (1N=50) 서식처에서 평형 상 태에 이른 경우를 생각해 보자(그림 3.5A-C). 개체군이 생장함에 따 라 식물의 전체 생물량은 증가하고 토양 영양염류의 이용도가 감소 되는 것과 똑같이 식생을 통해 투과되는 햇빛의 비율도 감소된다.
 0 330
0 330
해용처식치
의이식서정에 고 서한 측 류리한부률염그부풍급 양이영없 량(B).광 가풍류염더 씬훨 류공 염양 관와계크 의기가 자신든 체개모)C,B 테장A동( 생. A의 종 처에서이 식한서 의상개3 의 에서표 면지A),량(물생체 전에서의 TN0)=5식서(처 한약빈가엽류 는양영받 을한제 영양 )다EF된D(, ,.지유 가태그 상달하도여 에형평준 히속신 는)C(엽양류한 영능가 ) (G가I, 류양, H염영 .다된속지 이동 아받변 을한제광 이들체개 부일에 는서)001=(TN 영서처식의하 .데는미의 를류염양영체전 은N T.다된속지 이동변 )도서에TN005=(처 다. 있 어되명설 에절소 는라이〉쟁경 간종〈 의장 이특 은성 의종A 다.이 .3 5 그림1 년 정도 후에 개체군의 전체 생물량은 평형에 이르게 된다 . 지표면 에 입사되는 광량과 이용 가능한 토양 영양염류도 일정한 값에 근 접한다. 전체 생물량과 지표면에서의 광량 , 이용 가능한 영양염류가 일정 하게 된다 하더라도 개체군이 필연 적 으로 안정한 크기 분포를 하는 것은 아니댜 예를 들어 그림 3.5A - C 의 경우 크기의 구조는 일정한 변동을 보이는데, 그 이유는 일단의 실생이 자라서 성체로 되고, 이 들이 다시 새로운 실생에 의해 교체되기 때문이다. 그러나 전체 생 물량 , . 햇빛의 투과, 그리고 이용 가능한 영양염류의 수준은 필히 일 정하게 유지된다. 그 이유는 모든 식물체가 그들의 크기와는 무관하 게 이 척박한 토양에서 영양염류의 제한울 받기 때문이다. 영양염류 가 제한되기 때문에 전체 생물량은 일정하게 유지된다. 두번째 상황은 결코 평형 상태에 도달하지 못하는 경우이다. 대신 에 전체 개체군의 밀도와(각 연령군의 상대수도) 영양염류 및 햇빛의 이용도가 일정한 변동을 보인다(그림 3.5 D-I) . 내가 많은 모의를 실 행한 바에 의하면 영양염류 공급률이 충분히 높은 토양에서 이러한 변동은 흔히 있는 일이댜 이때 일부 개체들(키가 작은 것들)은 햇 빛에 의하여 제한을 받고 일부 개체들(키가 큰 것)은 영양염류에 의 하여 제한을 받는다. 이러한 두 가지 상황이 그림 3 . 5 의 D-1 에 나와 있댜 개체군이 영양염류의 제한을 받을 때에는 개체군의 크기 구조가 개체군의 생장에 미치는 전체 효과는 거의 없다. 그러나 햇빛에 의 하여 제한을 받으면 동시 출생 집단의 개체들은 성체 크기까지 자 라는 경향이 있고, 많은 자손을 생산한다. 이 자손들은 수관층을 구 성하는 개체들이 만든 그늘 때문에 생장이 저해된다. 따라서 대부분 실생의 생장이 억제되기 때문에 현재의 수관층 개체가 죽기 시작할 때에야 이들 실생들은 수관총 개체로 자라게 된다.
변동은 개체군 생장의 연령 구조 모델이나 크기 구조 모델에서 흔히 볼 수 있는데 (Sonleit ne r 1977), 그 이유는 연령 구조나 크기 구 조가 밀도 의존적인 과정에 시간지연 개념을 도입하기 때문이다. Ma y (1973) 가 지적한 바와 같이 시간지연은 개체군 안정성을 감소 시키는 경향이 있다. 나는 종의 최대 생장률이 커지거나 모델을 숫 자적으로 해석하는 데 걸리는 시간 간격이 더 길어지면 개체군의 변동이 커진다는 것을 지적하고자 한다. 시간 간격이 길어지면 모델 이 미분방정식 모델보다는 계차방정식 모델로 작용하게 만든다 (Ma y 1973). 나는 이 책에 있는 대부분의 모의에서 두 종 사이의 경쟁에 대한 몇 가지 경우에 다양한 시간 간격을 적용하였다. 그러나 컴퓨 터 사용상의 문제로 다른 모든 경우에도 이 시간 간격이 가장 최적 인지를 확인할 수 없었다. 이 시간 간격은 대략 하루에 1 회 반복하 는 것이었댜 그림 3.5 는 부족되는 토양 영양염류의 공급률이 서로 다른 서식 처에서 크기 구조를 갖는 A 종의 반응을 보여주고 있다. 이 종은 생 산력 구배상의 다른 지점에서는 어떻게 반응하는가? 특히 영양염류 공급률이 높을 때 반응에 변동이 있는 경향이 있기 때문예 각 영양 염류 공급률에서 나는 지속적인 변동에 접어들었을 때 식물 생물량, 이용 가능한 토양 영양엽류, 다양한 높이에서의 광량 등과 같은 각 변수의 장기간 평균값을 계산하였다(그림 3.6 ). A 종 단독으로 된 지 소에서 개체군의 평균 밀도는 영양염류 공급률에 따라 증가하여 공 급률이 높은 점에서 점근값이 된다 . A 종에 의해 감소되는 부족되는 토양 영양염류의 평균 수준은 영양염류 공급률이 낮을 때 일정하며, 영양염류 공급률에 따라 증가한다. 수관층 아래의 주어진 지점에 입사되는 광량은 반대의 패턴을 보 인댜 예를 들어 영양염류 공급률이 낮은 서식처에서는 지표면과 지 상 17cm 높이에 입사되는 광량이 대체적으로 높다. 영양염류가 점
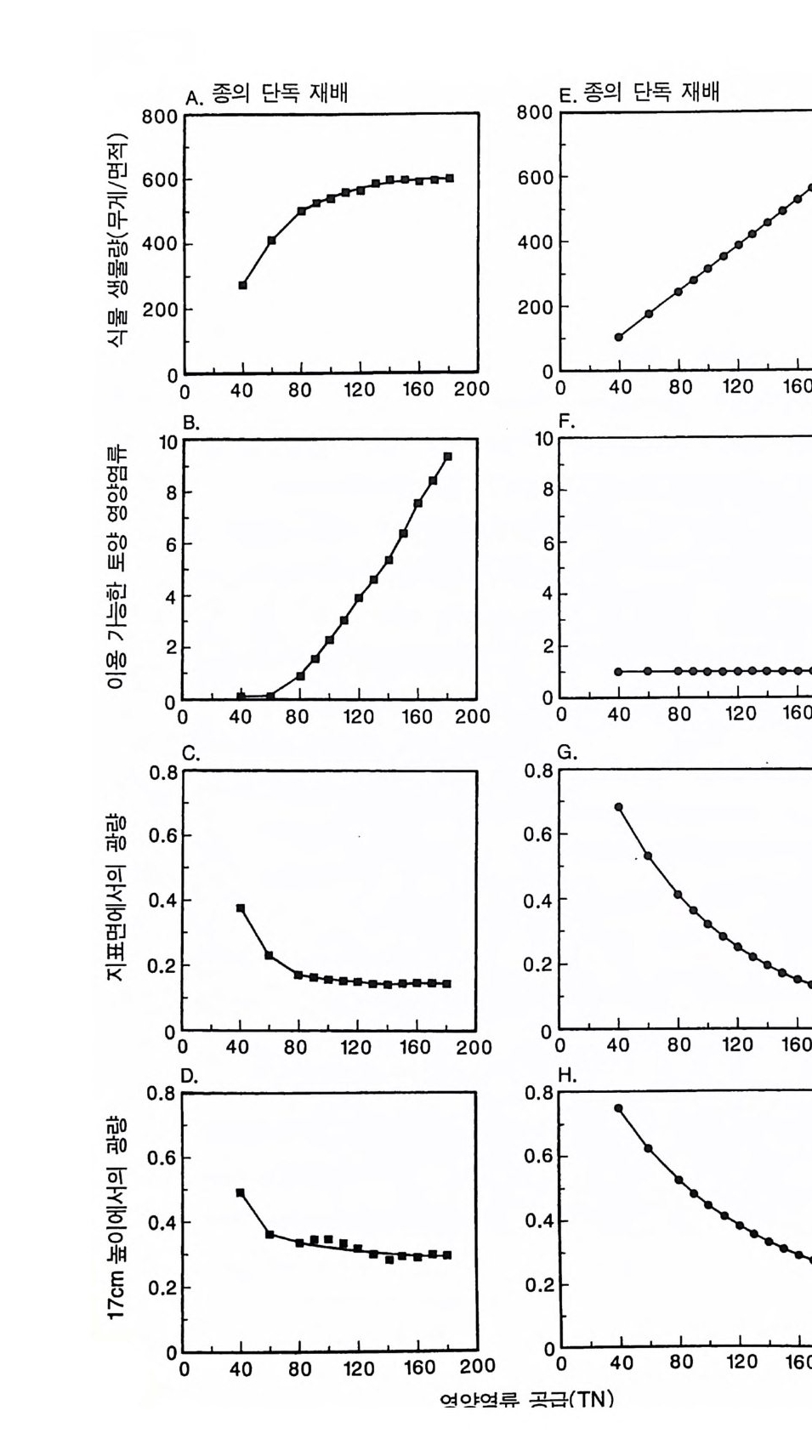 8Oo6A Oo종 4 으Oo 단2 독Oox, u卜 800 E . 종의 단독 재배
8Oo6A Oo종 4 으Oo 단2 독Oox, u卜 800 E . 종의 단독 재배
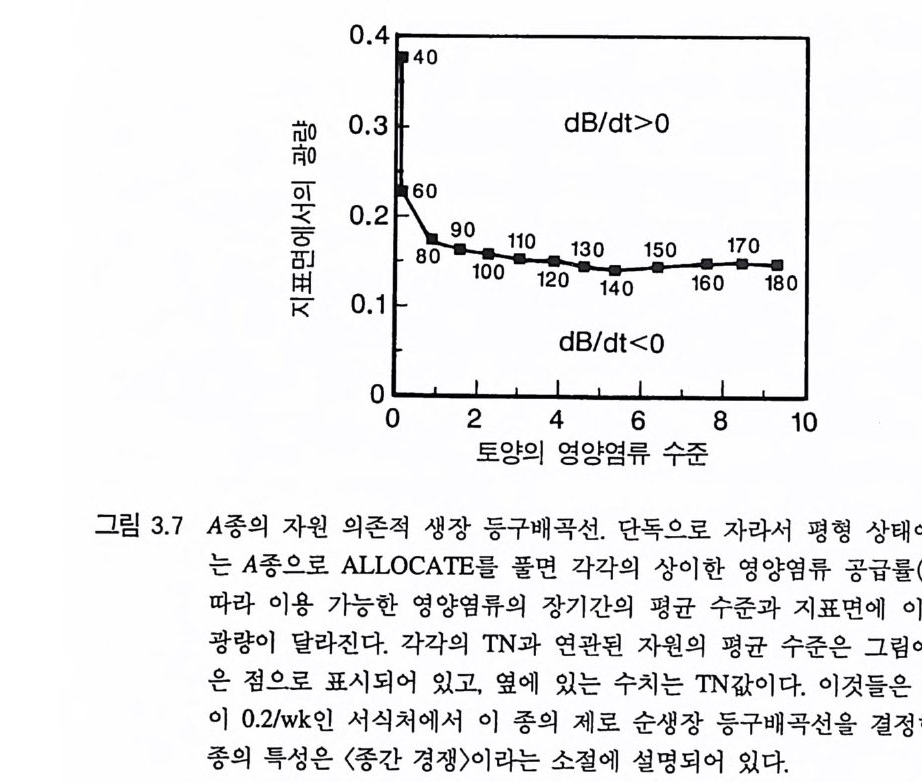
그림 3.6 (A-D) 손실률 0 . 2/wk 인 단일 종 지소에서 식물이 자랄 때(컴퓨터 모델 인 ALLOCATE 에서) , A 종의 장기간의 평균 생물량과 A 종이 감소시키는 부족한 토양 영양염류의 평균 수준, 그리고 지표면에 이입되는 광량 혹은 지표면으로부터 17cm 위의 광량은 영양염류 공급률에 의존적이다. (E-H ) B 종은 단일 종 지소에서 자랄 때 영양염류 구배에 유사하게 반응한다. A 종과 B 종의 특성은 〈종간 경쟁〉이라는 소절에 설명되어 있다.
진적으로 증가되는 서식처로 감에 따라 지표면과 지상 17cm 높이 에 도달하는 광량이 적어진다. 영양염류가 매우 풍부한 서식처에서 지표면에 도달하는 광량은 전체 식물 생물량에서와 같이 영양염류 공급률과는 무관하게 일정한 값에 이른다. 3-2 자원 의존적인 생장 동구배곡선 각각의 영양염류 공급률에는 부족한 토양 영양염류가 감소되는 평균 수준이 있으며, 이 개체군이 단일 재배로 오랫동안 성장한 후 에 식생에 의해 형성되는 수직적인 광 구배가 있다. 당분간 지표면 에 입사되는 광량을 종의 생장에 대한 햇빛의 효과를 나타내는 것 으로 간주하자. 그러면, 각 영양염류 공급률에서(죽 각각의 TN) 이 종이 단일 지소에서 자랄 때 부족한 토양 영양염류의 농도와 지표 면에서 광량을 감소시킬 수 있는 수준을 나타내는 한 쌍의 숫자가 존재하게 된다 . 각 영양염류 공급률에서 이 숫자들을 도시하면, 이 종의 자원 의존적인 제로 순생장 등구배곡선을 만들 수 있다(그림 3.7 ) . 실제로 이렇게 얻어질 수 있는 등구배곡선은 많은데. 각각의 둥구배곡선은 지표면으로부터 상이한 높이에서 광량의 수준을 나타 낸다. 이러한 모든 등구배곡선은 이 종이 부족한 토양 영양염류와 지상부의 모든 높이에서 광량을 감소시킬 수 있는 수준을 나타내는 3 차원적인 면을 구성하는 데 사용될 수 있다. 먼저 지표면에서의 광량이 어떤 높이에서의 광량보다 중요하다는
제 3 장 영양엽류와 햇빛에 대한 경쟁의 기작 았i
가정을 전제로 한 등구배곡선을 생각해 보자. 그림 3.7 의 등구배곡 선은 이 종의 장기간의 생식률이 장기간의 사망률과 정확히 균형을 유지하는 데 필요한 광량과 토양 영양염류의 수준을 보여주고 있다. 영양염류와 광량의 수준이 등구배곡선 밖에 있으면 이 종의 개체군 크기는 증가한다. 그러나 안쪽에 있으면 개체군의 크기는 감소한다. 등구배곡선에서 볼 수 있는 불규칙성은 영양염류와 광량의 수준이 변동하는 경향이 있기 때문이다. 등구배곡선은 장기간의 평균 자원 이용도를 기초로 하고 있다는 것을 기억하기 바란다.
 0.4
0.4
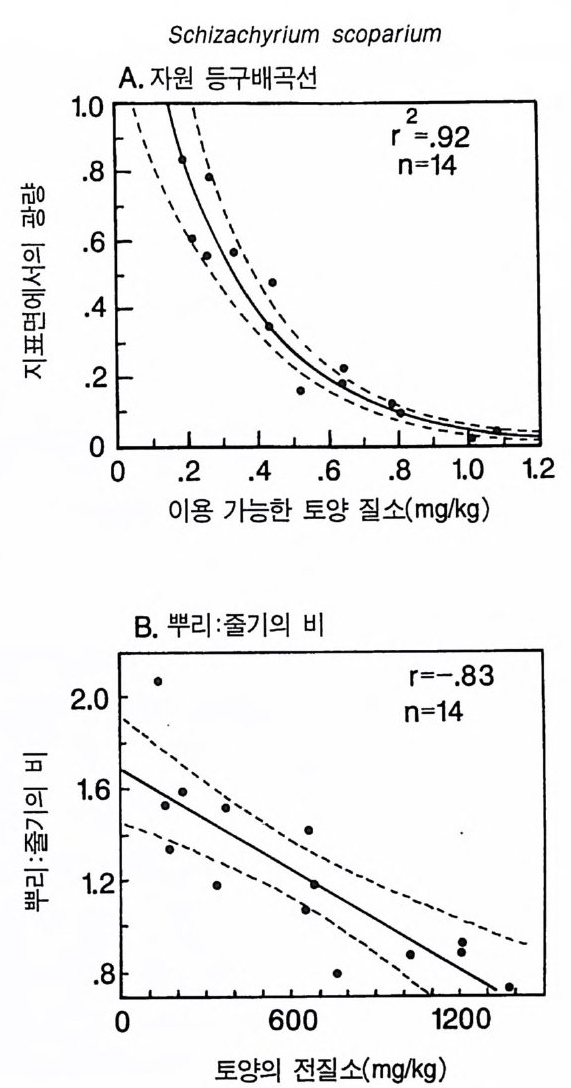
3-3 구부러진 동구배곡선 이 둥구배곡선은 코너가 완전한 직각이 아니고 곡선으로 되어 있 댜 이것은 부족한 토양 영양염류와 햇빛이 상호작용적 필수자원으 로 작용하고 있음을 의미한다. 영양염류 혹은 햇빛만이 광합성률을 조절하고. 이 종은 광합성 산물을 뿌리, 줄기, 잎으로 분배하는 고정 된 패턴을 갖기 때문에 자신의 형태를 두 자원에 의해 똑같이 제한 될 수 있도록 조절할 수 없다는 가정에도 불구하고 이러한 결과가 나타난댜 어떻게 이러한 현상이 나타날 수 있는가? 이러한 자원 의존적 생장 등구배곡선은 크기 구조가 완전한 개체 군에서 볼 수 있는 종합적인 곡선이다. 개개의 식물은 키가 다르고, 수직적인 광 구배가 있는 서식처에서 살고 있다. 따라서, 일부 개체 는 키가 작아 햇빛을 덜 받고 반면에 키가 큰 다른 개체들은 더 많 은 햇빛을 받는댜 모든 개체들은 균일한 토양 환경에 있다고 가정 한다. 영양염류 공급률이 중간 정도인 서식처에서 키가 작은 개체들 은 광량의 제한을 받고 키가 큰 개체들은 영양염류의 제한울 받는 댜 이러한 2 중 제한의 전체적인 효과에 의해 구부러진 등구배곡선 이 형성된다. 공간적으로 이질적인 서식처에서 각 식물체가 공간적인 위치에 따라 상이한 자원에 의해 제한을 받는다면 영양 면에서 완전히 필 수적 인 두 가지 자원도 상호 작용적 필수 자원으로 작용할 수 있다. 따라서 햇빛 이용도의 수직구조, 부족한 토양 영양염류의 공간적 이 질성, 혹은 자원을 획득하는 데 있어서 형태적 혹은 생리적 가변성 이 영양 면에서 필수적인 자원을 상호 작용적 필수 자원으로 작용 하게 할 수 있다. CedarCreekNa tur al Hi s t o ry Area 에서 2 년 동안의 생장 실험을 통해 얻은 자료는 토착 초지종인 little blueste m (Schiz a chy rium scop a ri um) 이
구부러진 둥구배곡선을 형성함을 보여주고 있다(그림 3.8 ) . 이 등구 배곡선의 자료는 이 종이 감소시키는 NH 4 와 N0 3 의 계절적인 평균 수준과 이 식물의 잎을 통과하여 지표면에 도달하는 햇빛의 수준에 근거한 것이다. 이 종의 형태적인 가변성에 관하여 조사된 자료에 의하면 뿌리 :줄기의 비가 질소가 가장 빈약한 토양에서 1.6, 가장 풍 부한 토양에서 0 . 7 을 갖는 것으로 나타났다(그림 3.8B). 따라서, 이 종의 구부러진 등구배곡선은 질소와 햇빛의 이용도에 따라 자신의 형태를 조절할 수 있는 이 식물의 능력과 일치한다. 3-4 종간 경쟁 이 모델을 이용하여 두 종 또는 그 이상의 종 사이의 경쟁에 대 하여 탐구할 수 있는 경우가 많이 있지만 먼저 두 종 사이의 경쟁 에 관한 몇 가지 경우를 살펴보도록 하자. 만약, 위에서 논의한 바 있는 A 종이 두번째 종인 B 종과 경쟁 관계에 있고, 두 종은 영양염 류와 햇빛의 제한을 받는 생장에서 단지 반포화상수만 다를 때 이 들 사이의 경쟁에서는 어떤 결과가 나타날까? A 종과 B 종은 잎과 줄 기 그리고 뿌리에 대한 분배 패턴이 각각 AL=0.7, AS=0.1, AR=0.2 로 똑같다. 두 종의 최대 높이는 200cm 이고 종자의 무게는 0.001g , 최대 광합성률은 2.0 / wk, 뿌리, 줄기, 잎의 호흡률은 0.3/wk 이다. 따라서 두 종의 RGRm ” 는 똑같댜 두 종은 영양염류와 광 의존적인 광합성에서 반포화율이 다르다 (영양염류의 반포화상수는 A 종이 KN(A)=0.05, B 종 KN(B)=0.50, 광의 반포화상수는 KL(A)=0.5, KL(B)=0.0 5 ). 잎, 줄기, 뿌리 그리고 종자 의 영양염류 농도도 다르다 (A 종은 0 . 01 이고, B 종은 0 . 02 이다). 선택된 변수들은 A 종이 B 종보다 영양염류 요구량은 적고 광 요구량은 높게 한다(그림 3.9A).
 Schlza c hy ri um scop a riu m
Schlza c hy ri um scop a riu m
그림 3.8 (A) 실험을 통해 얻은 Schiz a chy ri u m scop ar i um 의 제로 순생장 등구배곡선 각 접은 18L 의 화분에 Sch i zachy r i um 을 심어 야외조건에서 2 년 동안 키운 상태에서 이용 가능 한 질소와 지표면에 입사되는 광량의 평균 수준이다 . 14 개의 화분은 토양의 질소 공급 률 이 달랐다. 이들 자료에는 L=ex p (0.05-3.68N) 이란 지수함수 모델이 적합하였는 데 . 이때 규=0 . 92 , P< 0.001 이었다. L 은 지표면에 입사되는 광량의 비율, N 은 KCl 로 추출되는 암모니아와 가용성 질산의 합으로 단위는 mg /kg dry so il이다. 접선은 희귀직 선의 95% 신뢰구간을 나타낸다 . (B) 14 개의 화분에 있는 식물을 생육기간 말기에 수 95% 신뢰구간이 나와 있다 . M. L
 0.4 A.
0.4 A.
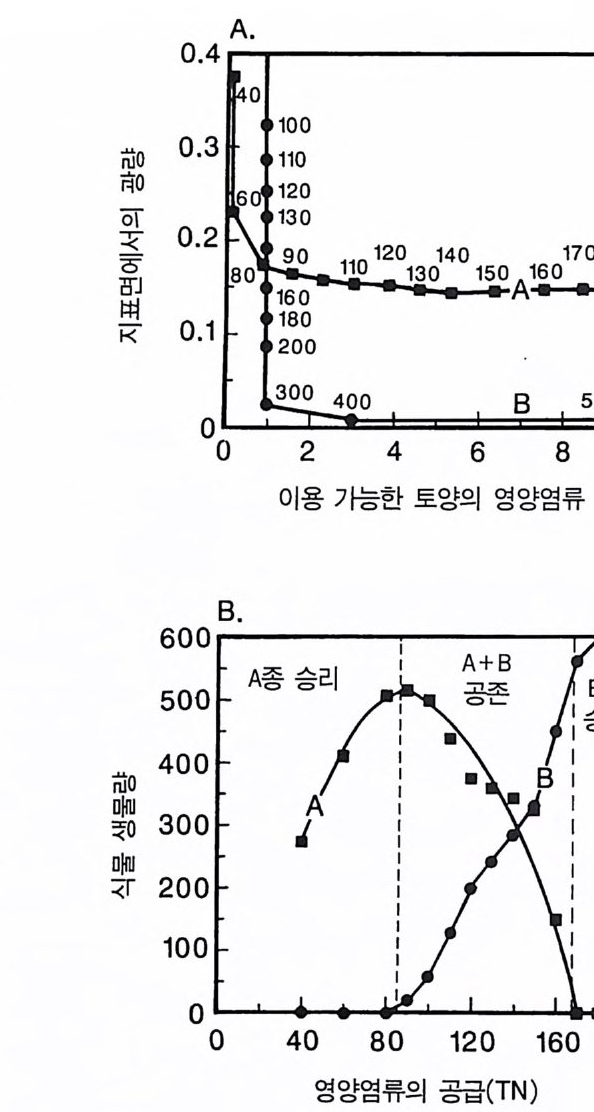
그림 3.9 (A) A 종과 B 종의 등구배곡선 각 곡선은 각각의 영 양염류 공급률 (TN) 에서 각 종이 감소시키는 자원의 평균 수준을 보여준다. 영양염류 공급률은 해당되는 점 옆에 표 시되어 있다. 등구배곡선은 ALLOCATE 를 이용하여 얻은 것이며, 이때 각 종은 영양 염류 공급률이 서로 다른 단일 종 지소에서 생장하는 것이다. (B) A 종과 B 종 사이 의 장기간에 걸친 종간 경쟁의 평균 결과. TN<80 에서 A 종은 경쟁을 통해 B 종을 교체하며. 80
이 두 종 사이의 경쟁의 최종 결과는 서식처의 영양염류 공급률 인 TN 에 의해 결정된다. 영양염류 공급률이 낮을 때에는 (TN<80) 그림 3 . 9B 에 나타난 바와 같이 A 종이 B 종을 경쟁적으로 교체한다. 영 양염류 공급률이 중간 정도에서는 (80
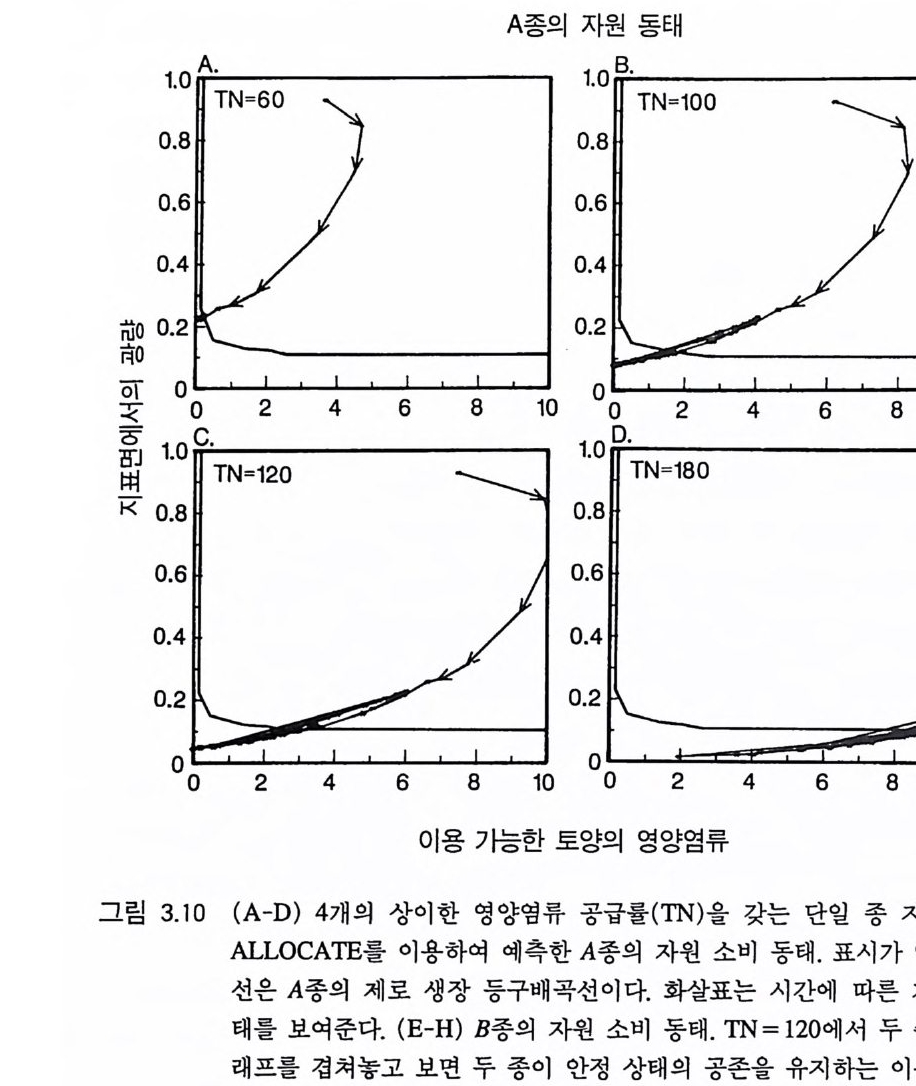 A 종의 자원 동태
A 종의 자원 동태
만 그것에 의한 예측은 제 2 장에서 제시된 등구배 접근 방법에서 얻 울 수 있는 예측과 정성적인 면에서 비슷하다. 이와 같이 두 방법을 통한 예측이 비슷하다는 것은 단일종 재배 를 통해 얻을 수 있는 장기간의 자원 요구량에 관한 정보를 이용하
 B 종의 자원 동태
B 종의 자원 동태
여 이론상 종간 경쟁의 결과를 예측할 수 있음을 의미한다. 이것이 일반적인 결과라면 모델 ALLOCATE 에 포함되어 있는 모든 생리적. 형태적인 세부 항목을 얻을 필요가 없음을 의미한다. 대신에 일련의 단일종 재배를 통해 실험적으로 얻을 수 있는 등구배곡선으로 식물
의 형태적 그리고 생리적 복잡성을 요약할 수 있음을 의미한다 . 그 러나 등구배곡선 접근 방법이 얼마나 유용성이 있는지를 결정하기 위해서는 더 많은 이론적 . 실험적 연구가 필요하다. 단일종 재배를 통한 예측과 모델 ALLOCATE 로부터 얻을 수 있 는 종간 경쟁의 결과는 정량적으로 서로 일치하는가? 각 종의 단독 재배시 영양염류와 지표면에서 광량을 감소시키는 평균 수준은 각 영양염류 공급률 (TN) 에서 종간 경쟁의 결과를 예측하는 데 이용될 수 있댜 다음에 설명하는 방법은 내가 알고 있는 단독 재배시의 영 양염류와 광 자료 중 경쟁 결과를 예측하기 위해 사용하는 가장 좋 은 방법이댜 그 이유는 햇빛이 방향성을 갖기 때문에 햇빛의 공급 률과 소비율이 제 2 장에서 제시된 수학적인 정의와 맞지 않기 때문 이다. 자원 경쟁에 관하여 제 2 장에서 그림으로 제시한 평형 이론은 단 독 재배에서 얻은 등구배곡선과 평형시의 자원 수준으로 어떻게 경 쟁의 결과를 예측할 수 있는지를 보여주고 있다. 이것은 그림 3.11 에 나와 있다. X, Y, z 로 표시된 3 개의 상이한 공급점을 생각해 보자. 공급점 x 는 R1 의 공급률이 낮고 R 2 의 공급률이 높은 서식처를 나타내며, 공급점 y는 두 자원의 공급률이 중간 정도이고, 공급점 z 는 R1 의 공급률이 높고 R2 의 공급률이 낮은 서식처를 나타낸다. 공급점 x 에서(그림 3.I IA ) L 종의 단일 지소는 평형 상태의 자원 수준을 그것의 등구배 곡선상의 점 R*L .x 까지 감소시킬 것이다. 이와 비슷하게 종 M 의 단 일 지소는 자원 수준을 R* M . X 까지 감소시킨다. 공급점 x 에서 각 종 의 평형시 자원 수준은 두 종 평형점을 기준할 때 같은 쪽에 위치 하게 된다. 이러한 서식처에서 M 종은 L 종에 의해 경쟁적으로 교체될 것으로 예상되는데, 그 이유는 L 종이 자원 R1 을 M 종의 생존에 필요한 수준
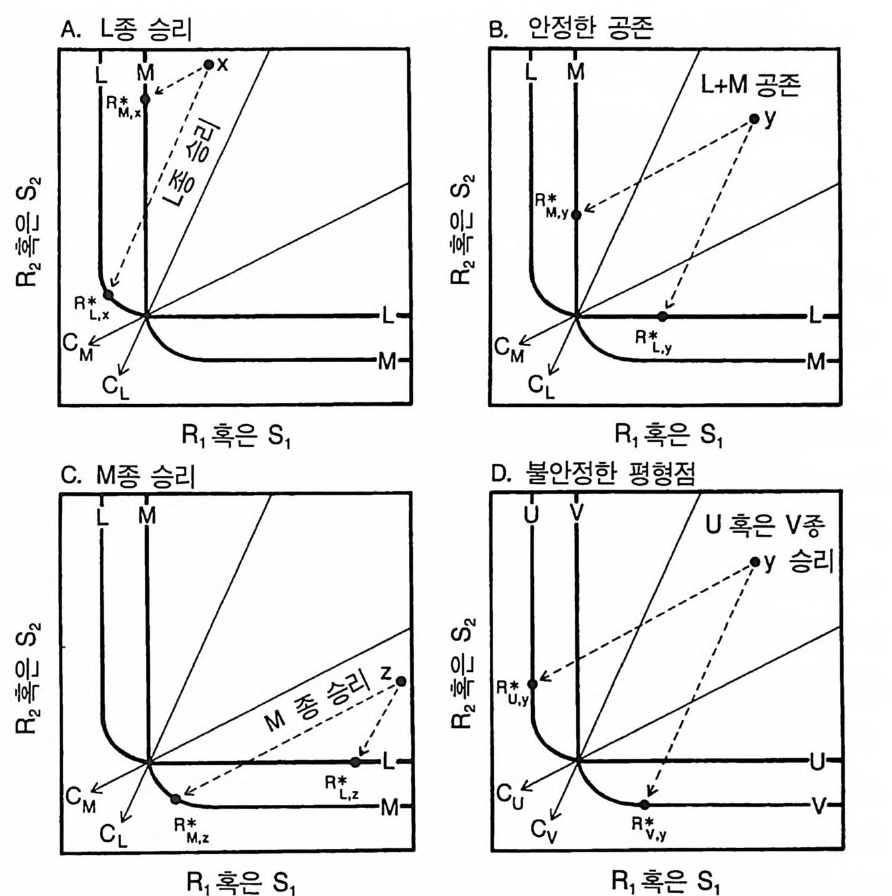 A. L 종 승리 B. 안정한 공존
A. L 종 승리 B. 안정한 공존
그림 3.1 1 L 과 M으로 표시된 굵은 곡선은 L 종과 M 종의 제로 순생장 등구배곡선들이다. (A) 평형 상태에서 공급점 x 에서는 L 종이 자원 수준을 R* l.초: 감소시킨다. 이 것은 점선 화살표에 의해 표시되어 있다. M 종은 자원의 수준을 R*M , ;. 로 감소시 킨다. 이들이 경쟁을 하면 〈 L 종 승리〉라고 표시된 지역에 있는 모든 공급점에서 L 종은 M 종을 교체한다. (B) 이 경우 평형 상태에서 각 종은 자원의 수준을 다 른 종의 순생장 범위 내의 점까지 감소시킨다. L 종과 M 종은 점 y와 같은 공급접 에서만 공존할 수 있다. M 종의 경우 R*M. y, L 종의 경우 R*l. y 둥이 그런 접에 해 당되는데. 각각은 자원 공급점 y와 관련이 있다 . (C) 〈 M종 승리〉라고 표시된 지 역에 있는 모든 공급점에서 M종은 경쟁에 의해 L 종을 교체한다. Z 와 같은 공급 점은 R* 들이 두 종의 평형접과 같은 쪽에 있도록 한다. (D) 만약 각 종이 다른 종이 살 수 없는 지역에 R* 를 가지고 있다면 두 종의 평형점이 불안정하다. 이 경우와 그림 B 사이의 차이점은 두 종의 소비 백터 그리고 그것과 관련된 R* 값 이다 . 〈 U 혹은 V종 승리〉로 표시된 지역에 있는 y와 같은 모든 공급접에서는 u 혹은 V 에 의해 우점되며. 이때 다른 종은 U 나 V 에 의해 경쟁적으로 교체된다. 경쟁에서 이기는 종은 초기 조건에 의해 결정될 것이다.
이하로 감소시킬 수 있기 때문이다 . 이와 유사하게 L 종이 M 종에 의 해 교체될 것으로 예측되는 지역의 점 z 와 같은 공급점은 R * l .Z 와 R * M . Z 가 두 종 평형점을 기준할 때 같은 쪽에 위치하여, M 종이 경 쟁에서 이길 것으로 예측된다(그림 3.I IC ). 그 이유는 M종이 L 종의 생존에 필요한 것보다 낮은 수준으로 자원 R 2 를 감소시킬 수 있기 때문이댜 각 종의 단독 재배시 평형 상태의 자원 수준 (R * l . ) 와 R * M .). )이 두 종 평형점울 기준하여 서로 반대편에 위치하는 공급점 y와 같은 경 우는 두 종이 공존할 수 있는 것으로 예측되는 지역이다(그림 3.1 l B). 이 경우는 안정 상태의 공존이 가능하며. 단독 재배시 평형 상태의 자원 수준은 L 종이 M 종의 생존에 필요한 수준 이하로 자원을 감소 시키지 않고 M 종은 L 종의 생존에 필요한 수준 이하로 자원을 감소 시키지 않는 지점(자원 공급점 y)에 위치한다. 따라서 R * l , y 는 M 종의 순생장 지역에 위치하고 R*M, y는 L 종의 순생장 지역에 위치한다. 마지막 경우는 평형점이 불안정한 경우이다(그림 3.1 1 D). 이 경우 를 그림 3.11 의 A-C 와 구분하기 위하여 나는 관련된 두 종을 각각 U, V로 표시하였다. 평형점이 불안정하면 각 종은 중간 정도의 서식 지에서 단독 재배시 두 종 평형점의 반대편 점까지 자원 수준을 감 소시킬 수 있지만, 이러한 자원 수준은 각 종이 다른 종의 생존에 필요한 충분한 자원을 남기지 않는 수준이다. 예를 들어 그림 3.11D 의 공급점 y는 R*u. y와 R*v. y 를 유도한다. R*u . y 에서는 U 종만이 생존 할 수 있고, R*v. y에서는 V종 만이 생존할 수 있다. U 종과 V종 이 단 독 재배시 여기에 나타난 것과 같은 둥구배곡선과 R* 를 갖는다면, 이들 사이의 경쟁에서 예상되는 결과는 불안정한 공존이 될 것이다. 죽 처음의 출발 조건에 따라 U 종 혹은 V종 이 완전히 상대종을 교 체하게 된다. 전체적으로 그림 3.11 에 제시된 분석을 통해 특정한 자원 공급
성질을 갖는 서식처에서 각 종의 단독 재배를 통해 실험적으로 관 찰된 등구배곡선과 R * 들은 종간 경쟁의 결과를 예측하는 데 충분한 정보를 가지고 있음을 알 수 있다. 자원의 공급과 소비의 기본적인 동태는 그것들이 등구배곡선의 모양, 위치 및 R* 에 어떤 영향을 주 느냐를 제외하고는 중요하지 않게 된다. 따라서 이 방법은 햇빛에 대한 경쟁을 분석하는 데 좋은 방법이다. 왜냐하면 이 방법은 햇빛 의 공급률을 정의할 필요가 없으며. 전체 식물 생물량이 햇빛의 소 비에 의존적이라는 내용이 포함되어 있기 때문이다. 자원 경쟁에 대한 평형 이론을 통해 나올 수 있는 이러한 정량적 인 예측은 그림 3.9 에 나와 있는 여 러 가지 TN 값에서 ALLOCATE 룰 사용하여 수행된 종 A 와 B 의 경쟁의 경우에도 적용할 수 있다. 두 종의 등구배곡선과 몇 가지 상이한 영양염류 공급률 (TN 수준)에 서 각 종이 감소시킬 수 있는 지표면에서의 이용 가능한 영양염류 와 햇빛이 그림 3.9 에 나와 있다. TN 이 40-80 에서는 A 종과 B 종 모 두 두 종의 평형점 이상의 수준까지 자원을 감소시킨다. 두 종은 주 로 토양 영양염류의 제한울 받으며, 이때 영양염류의 요구량이 적은 A 종이 경쟁에서 이길 것으로 예측된다. 둥구배곡선들은 A 종의 등구 배곡선상의 TN=82 인 지점에서 교차한다. 이와 유사하게 등구배곡 선들은 B 종의 동구배곡선상의 TN=150 인 점에서 교차한다. TN 값 이 82-150 사이에서는 단일종 재배시 영양염류와 광량의 수준이 두 종 평형점을 기준할 때 서로 반대편에 있으며. 이런 상태의 공존은 안정한 것으로 예측된다. 이것을 알아보기 위해서는 그림 3.9A 를 그 림 3 .1 1 과 비교해 보아라. 이러한 예측은 지표면에서의 광량이 전체 광 구배의 좋은 척도가 된다는 가정을 기본으로 하고 있다. TN=90, 100, 110, 120, 130, 140, 그리고 150 일 때 경쟁의 모의는 모두 A 종과 B 종이 안정한 상태로 지속됨을 보여주고 있다. 지속 상 태의 안정성은 각 TN 에서 많은 모의를 행하여 판단할 수 있는데.
각 모의는 두 종의 초기 수도를 달리하여 시작한다. TN>150 인 모든 서식처에서 등구배곡선은 B 종이 A 종을 교체할 것으로 예측한다. 이러한 서석처에서 두 종은 모두 광에 의해 제한 을 받으며. B 종은 A 종의 생존에 필요한 수준 이하로 광량을 감소시 킨댜 이러한 현상은 TN=160 에서는 일어나지 않는다. TN=160 에 서 A 종과 B 종은 안정하게 공존한다. 그러나 TN>160 에서는 예측한 대로 B 종이 A 종을 교체한다. 따라서 이러한 모의는 단순히 평형을 기초로 한 둥구배곡선 방법을 통해 예측되는 경쟁 결과와, 토양 영 양염류와 햇빛에 대한 좀더 현실적이고 좀더 복잡한 경쟁 모델에 의해 예측되는 경쟁 결과가 서로 완전히 일치되지는 않지만 그래도 상당한 일치성이 있음을 보여주고 있다. 단일종 재배에서 식물이 영양염류와 지표면에서의 광량을 감소시 키는 장기간의 평균 수준을 근거로 하는 등구배곡선은 두 종간의 장기간 경쟁 결과의 패턴을 예측할 수 있다. 이것은 수직적인 광 구 배에서 지표면에서의 광 수준을 광 이용도의 정도를 나타내는 지수 로서 사용할 수 있다는 타당성을 인정하는 것이긴 하지만. 앞의 설 명에서는 중요한 가정이 포함되어 있다. 죽, 두 종이 성숙하였을 때 의 최대 높이가 200cm 로 같다는 가정이댜 이들의 최대 높이가 다르면 어떤 일이 일어날까? 이것을 알아보기 위해서 앞의 경우에서 사용한 모든 변수를 일정하게 두고 단지 A 종의 최대 높이룰 50cm 로 바꾸고, 이후부터 이것을 C 종이라 불렀댜 B 종 의 최대 높이는 200cm 로 그림 3.5 에서 3.11 에 이르기까지 모두 똑 같다. 최대 높이룰 변화시킨 결과 A 종의 등구배곡선이 그림 3 .1 2 에 있는 C 종의 둥구배곡선 위치로 옮겨졌고, B 종과의 경쟁적 상호 작 용이 변화되었다. 그러나 경쟁의 정성적인 면에는 영향이 없었다. C 종은 영양염류 공급률이 낮은 토양에서 우점하였고, 두 종은 영 양염류가 중간 정도인 토양에서 공존하였으며, 영양염류가 풍부한
 0.4 E A.
0.4 E A.
그림 3.12 (A) B 종과(그림 3.6, 3.9 그리고 3.10 에 있는 B 종과 동일함) C 종(그립 3.6, 3.9 그리고 3 . 10 에 있는 A 종과 동일함 단지 C 종의 최대 높이는 200cm 가 아닌 50cm 임)의 자원 등구배곡선. (B) ALLOCATE 에 의해 예측된 B 종과 C 종 간의 경쟁 결과.
토양에서는 B 종이 우점하였다. 그러나 지표면에서의 광량을 기초로 한 등구배곡선은 그림 3 . 9 에서와 같이 이 경우에서도 안정한 공존 지소와 B 종이 우점하는 지소 사이의 실제적인 경계의 예측에는 적 절하지 않았다. 지표면에서의 광량을 기초로 예측된 공존 지역은 TN=90 부터 TN=165 까지였다(그림 3.1 2 A 참조). 그러나 모의에서 관찰된 실제 공존 지역은 TN=90 부터 TN= 130 까지였다(그림 3.12B). TN= 140 이상에서는 B 종이 C 종을 교체하였다. 지표면의 광량을 기준한 등구배곡선에 의한 예측이 모의에 의한 예측과 차이가 있는 것은 개체군의 크기 구조에서 비롯된다. B 종은 성숙하였을 때 C 종에 비해 키가 크다. B 종의 장기간 생물량 평균치 는 TN 에 따라 증가한다. TN 이 130 보다 클 때에는 B 종이 광량을 감 소시켜 키가 작은 C 종은 광 제한울 받는다. 그 결과 C 종은 자신을 교체할 충분한 종자를 생산할 수 없다. 따라서 성숙하였을 때 크기 가 다르면 지표면의 광량이 아닌 다른 높이에서의 광량이 경쟁의 결과를 결정하는 데 매우 중요할 수 있다. 이 경우에 지표면보다는 위이지만 키가 작은 종의 수관(최대 높이 50cm) 보다는 아래에 있는 어떤 지점에서 평균 광량을 기초로 한 등구배곡선이 종이 공존하는 데 필요한 영양염류의 공급률을 예측하는 데 훨씬 유용하다. 지상부 17cm 와 33cm 에서의 광량을 기초로 한 둥구배곡선(그림 3 .1 3) 은 공존이 일어나는 TN 의 범위를 약 90-130 으로 예측하는데. 이것은 모의를 통해 관찰되는 공존 범위와 매우 근접한다. 그러나 나는 이것이 일반적인 결과인지를 확신할 만큼 충분한 모의를 하지 못하였다. 나는 ALLOCATE 모델을 사용하여 두 종 이상의 경쟁에 대한 수백 번의 모의를 수행하였다. 그 결과 단일종의 재배를 기초 로한 둥구배곡선은 경쟁의 정성적인 결과를 예측하는 데 일반적으 로 타당성 이 있다는 것을 알았다.
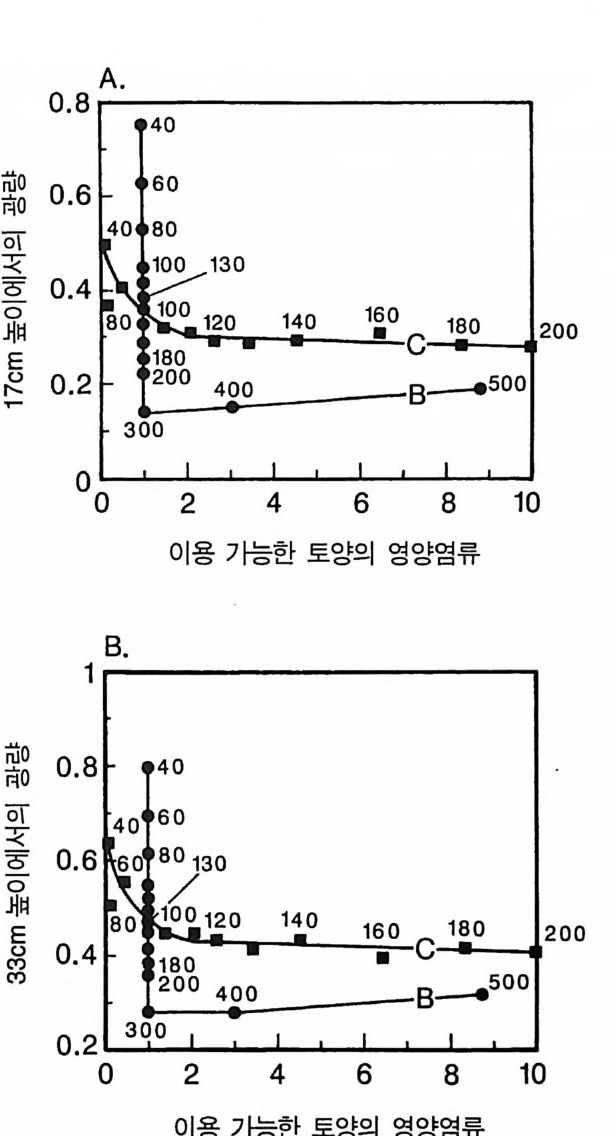 0.8[A .
0.8[A .
그림 3.13 (A) 17cm 높이에 입사되는 광량을 기초로 한 B 종과 C 종의 등구배곡선 . (B) 33cm 높이에 입사되는 광량을 기준으로 한 B 종과 C 종의 등구배곡 선. 두 경우에서 이 등구배곡선들은 지표면에 입사되는 광량을 기준으로 한 둥구배곡선에 비해 키가 큰 종 (B 종)과 키가 작은 종 (C 종) 사이의 장 기간에 걸친 경쟁 결과를 예측하는 데 더 적합하다. 이것을 이해하려면 이 그림을 그림 3 .1 2 와 비교하여라.
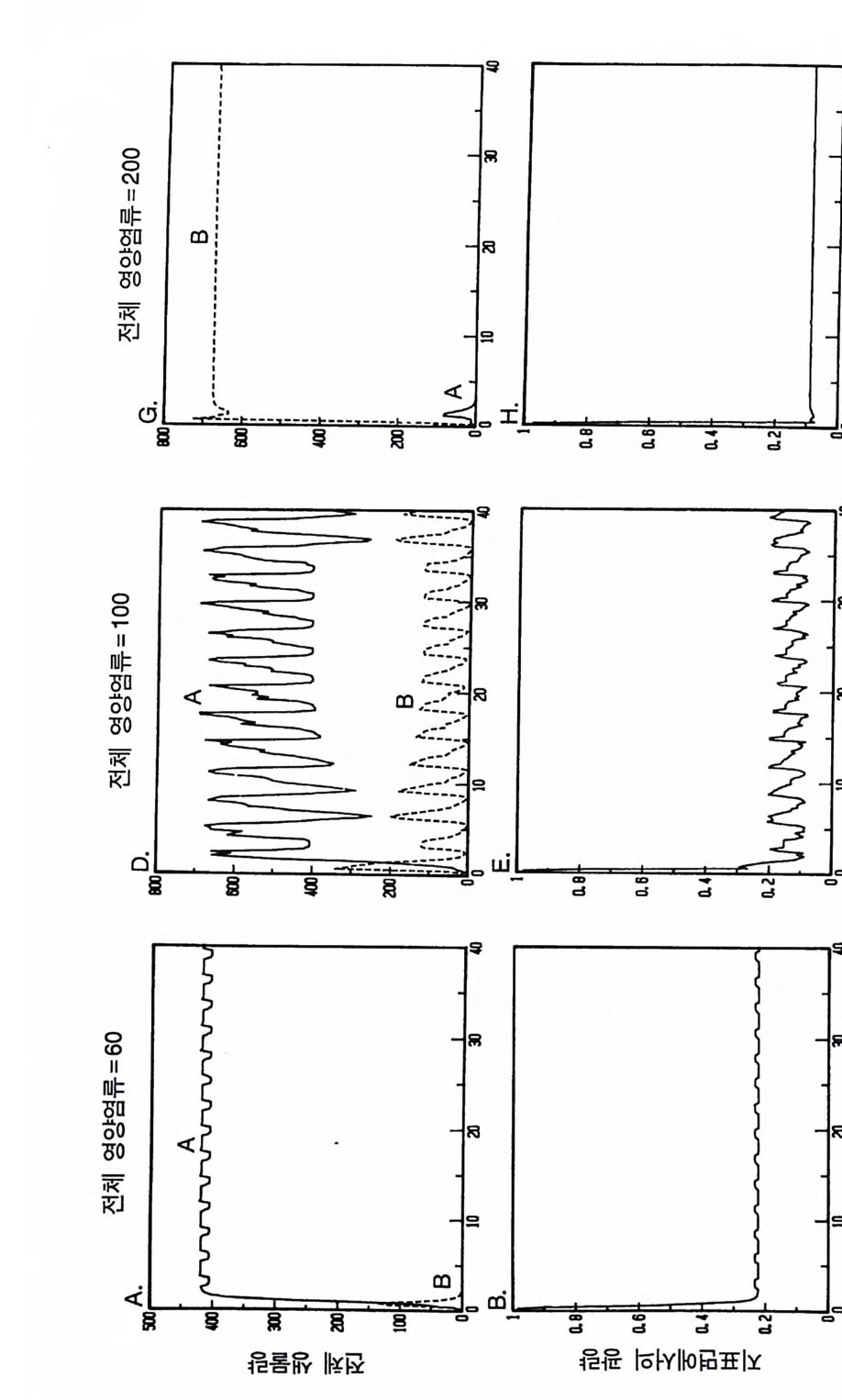
앞에서 논의한 등구배곡선은 각 종의 단독 재배시 자원을 감소시 키는 평균 수준을 근거로 한 것이다. 그림 3.5 에 나타난 바와 같이, 지표면에서의 광 이용도와 영양염류 이용도는 실제로 많은 경우에 서 일련의 변동을 보이고 있다. 앞에서 논의된 장기간의 경쟁 결과 는 둥구배곡선 모델을 통해 정성적으로 예측할 수 있는데, 25 년간의 경쟁이 있은 후 각 종의 평균 밀도에도 근거를 두고 있다. 그러나 이러한 경쟁적 상호 작용에는 몇 가지 재미있는 동태가 있다(그림 3.1 4 ). TN=60 에서 A 종은 B 종을 교체하는데, 최종 평형에 이르는 곡선은 다소 평탄하며 일정한 점근값에 접근한다. TN=200 에서 B 종은 A 종을 교체하는데 역시 곡선이 평탄하며 평형점에 접근한다. 이 두 경우 경쟁에서 이기는 종은 영양염류에 의해 제한을 받는다. B 종은 TN=60 에서 상호 작용의 초기에 급격히 증가하는 기간이 있고, 첫해 동안에는 A 종에 비해 수도가 높다. TN=100 에서와 같이 두 종의 공존은 밀도가 고정된 것이 아니다. 두 종과 두 자원은 모 두 일련의 변동을 보이는데, 영양염류, 햇빛, 그리고 두 종의 밀도 변동이 일치하지 않는다. 자원의 수준은 처음에는 A 종이 유리하게 그 다음에는 B 종이 유리하게 교대로 계속 변동하였다. 그러나 각 종 은 수도가 낮을 때에는 증가할 수 있는데, 그 이유는 경쟁 관계에 있는 다른 종의 수도가 높으면 자원의 수준을 첫번째 종에게 유리 한 지역으로 변형시키기 때문이다. 그러한 변동은 뚜렷한 크기 구조 가 없는 비슷한 모델에서는 나타나지 않았다. 수평적으로 균질한 지역에 살고 있는, 크기 구조를 갖는 개체군 사이의 경쟁 모델인 ALLOCATE 는 공존하는 두 종의 식물 사이에 서 혼히 볼 수 있는 동적인 상호 작용을 예측하는데, 그 중 하나는 각 종이 다른 종에게 유리하도록 자원 이용도를 변화시켜 아주 좁 은 지역에서 각 종은 다른 종을 교체할 수 있다는 것이다. 이러한 소지역에서의 교체 패턴은 미시간 남서부 너도밤나무와 당단풍나무
에서 보고된 바 있다 (Woods 1979, Woods & Whit tak er 1981 ). Woods(1979) 는 당단풍나무림의 경우 그 밑에 당단풍나무 실생에 비해 너도밤나무 실생을 더 많이 가지고 있고, 너도밤나무림 밑에는 반대로 당단풍나무의 실생이 많은 것을 알았다. Fox(1977) 는 Great Smoky Moun t a i ns 에서 프레저전나무와 붉은가문비나무에서도 비슷 한 패턴이 있음을 보고하였고, 플로리다에 있는 떡갈나무와 물참나 무, 펜실베이니아에 있는 너도밤나무와 솔송나무, 그리고 Medic ine Bow Moun t a i ns 에 있는 아고산전나무와 엥겔만가문비나무 사이에도 비 슷한 패 턴 이 있다 (Woods & Whit tak er 1981 에 있는 표 19.1 참조) . 이러한 모든 패턴은 자원 공급률이 공존을 유도하는 서식처에서 연령 구조를 갖는 두 종류의 개체군이 변동 모드로 경쟁할 때 일어 날 것이라고 이 모델이 예측하고 있는 바로 그것이다. 이와 같은 변 동이 없이 공존하는 종의 경우에도 이 모델의 영역에 포함된다. 그 러나 육상의 관속식물에서는 크기 구조가 있기 때문에 어떤 지소에 서도 장기간의 변동이 있게 마련이다. 이것은 공존을 유도하는 전체 적인 기작이 바로 이 종들의 영양염류와 광 요구량의 분화이며, 상 호 교체와 같은 지역적인 동태는 이러한 기작과 이 개체군들의 크 기 구조 사이의 상호 작용의 결과임을 나타내 주는 것이다. 이 장에서 제시된 경쟁에 관한 이론적인 예는 영양염류 및 광 의 존적인 광합성과, 성숙했을 때의 식물의 높이가 경쟁 능력에 영향을 줄 수 있음울 보여주고 있다. 제 4 장에서 설명된 바와 같이 분배 패 턴, 종자의 크기, 그리고 첫번 생식을 시작할 때의 크기도 크기 구조 를 갖는 식물의 경쟁에 영향을 준다. 광합성 효율, 뿌리로의 분배, 기타 식물이 가지고 있는 어느 한 가지의 생리적 혹은 형태적인 특 징만으로는 크기 구조를 갖는 둘 이상의 식물 개체군이 경쟁할 때 어떤 결과가 나타날 것인지를 예측할 수 없다. 특정한 자원 공급률 과 특정한 손실률을 갖는 서식처에서 식물이 생장하고 경쟁하는 능

력은 여러 가지 생리적. 형태적. 그리고 생활사적인 특징들 사이의 상호 작용에 의해 결정된다. 이러한 분석은 실험적으로 얻어진 자원 의존적인 생장 등구배곡 선을 종의 생리적. 형태적. 생활사적인 특징의 복합적인 효과로 받 아들일 수 있다는 것을 제시하고 있다. 둥구배곡선의 모양과 위치는 식물이 갖는 이러한 모든 특징에 의해 영향을 받는다 . 또한 이러한 결과는 각 종의 자원 의존적안 생장 등구배곡선을 육상 관속식물 사이의 종간 경쟁의 결과를 정성적으로 예측하는 데 사용할 수 있 음을 제시하고 있다. 둥구배곡선 방법은 햇빛이 복합변수이기 때문에 근사치이다. 지표 면에서의 광량에 의해 더 많은 제한을 받는 종이 있는가 하면 지표 면이 아닌 다른 높이에서의 광량에 의해 더 많은 제한을 받는 종들 도 있다. 식물 개체군의 각 개체는 어릴 때에는 어느 한 가지 자원 에 의해 제한울 받지만 어느 정도 생장하면 다른 자원의 제한울 받 는댜 그렇지만 ALLOCATE 같은 모델을 사용하는 데 있어서 모델 의 복잡성, 측정해야 할 변수의 수를 고려하면 등구배곡선 방법이 다양한 야의 조건과 일반적인 이론에 잘 맞는 것이라고 할 수 있다. 자원 의존적인 둥구배곡선 방법이 갖는 유용성은 직접 실험에 의해 서만 결정될 수 있다. 모든 모델은 추상적인 것이며, 자연의 복잡성을 단순화시킨 것이 다. 주어진 모델 내에 들어 있는 단순화를 위한 가정이 그 모델을 통해 설명할 수 있는 범위를 결정한다. 형태적으로 뚜렷하지 않은 모델은 형태적 패턴의 원인에 대해 예측할 수 없다. 따라서 이러한 모델들을 사용하여 형태적으로 매우 상이한 종 사이의 상호 작용의 성질을 연구할 때에는 조심하여야 한다. 다음 장에서는 서식처에 우 점하는 식물의 형태와 생활사가 그 서식처의 자원 공급률과 손실률 혹은 교란율에 얼마나 영향을 받는지를 결정하기 위해 이 장에서
제시된 크기 구조를 갖는 식물 사이의 영양염류와 광에 대한 경쟁 모델을 사용할 것이다. ALLOCATE 는 크기 구조를 갖는 식물들 사이의 경쟁에 관한 많 은 모델 중의 하나이다. ALLOCATE 모델 속에는 또다른 제약, 대응 책, 가정 등이 들어 있으며, 이것을 다른 모델들과 비교함으로써 우 리는 새로운 것을 얻을 수도 있다. 나는 독자들이 ALLOCATE 와 친 숙해지고 그것이 가지고 있는 효과에 대하여 좀더 알도록 하기 위 해 이 장에서 ALLOCATE 의 몇 가지 특징을 설명하였다 . 이 모델은 식물경쟁의 기작에 대하여 완전히 실제적인 모델로 작성된 것이 아 니고 영양 생장을 계속할 수 있는 크기 구조를 갖는 식물 사이의 경쟁에서 참재적으로 중요성을 갖는 몇 가지를 포함하고 있다. 이러 한 기능을 갖는 모델을 만들 수 있는 방법들이 많이 있지만, 나는 ALLOCATE 를 이용하여 얻은 예측이 타당성이 있으며, 뿌리, 줄기, 그리고 잎으로의 분배 패턴에 의해 결정되는 생장의 제약을 포함하 고 있는 다른 크기 구조를 갖는 모델과 상치되지 않을 것으로 믿고 있다. 4 요약 자원의 제한이 없는 서식처에서 dB/dt • 1/ B 로 측정되는 식물의 최대 영양 생장률은, 혼히 최대 상대 생장률로 불리는데, 식물의 형 태와 관련이 많다. 영양염류와 광 포화 상태의 광합성률과 주어진 호흡률을 갖는 식물은 잎으로의 분배를 최대로 할 때 영양 생장률 이 최대가 된다. 잎, 뿌리 그리고 줄기의 호흡량과 순환율을 서로 비 슷한 것으로 가정하면 식물의 최대 생장률은 잎으로의 분배량과 직 접적인 비례 관계가 있다. 뿌리와 줄기의 진화는 자원 부족에 대한
적응으로 설명되어야 한다 . 크기 구조를 갖는 식물 사이의 생장과 경쟁의 모델인 ALLOCATE 를 제시하였고, 이 모델의 몇 가지 특징을 설명하였다. ALLOCATE 가 다음과 같은 복잡성, 죽 (1) 영양염류와 광 의존적인 광합성, (2) 연속적이고 자원 의존적이며 분배에 의해 결정되는 식 물의 영양 생장, (3) 뿌리, 잎, 줄기의 생물량과 호흡량이 다르고, (4) 식물 자신에 의해 야기되는 수직적인 광 구배 둥을 포함하고 있 지만 이 모델의 장기간의 결과는 대부분 영양염류와 광 의존적인 생장 등구배곡선을 이용하여 정성적으로 예측될 수 있다. 이러한 등구배곡선은 각 종의 단독 재배를 통한 모델에서 얻는다 . 모델에 포함되어 있는 크기 구조는 많은 개체군에서 일련의 변동 이 있는 생장을 유도하는데, 햇빛이 부족할 때는 특히 그렇다. 이 모 델은 손실률이 일정하고 토양이 균일한 서식처에서 각 종의 실생이 다른 종의 성체 믿에서 더 잘 자라게 됨으로 두 식물이 공존할 수 있다는 것을 시사하고 있다. 한 종은 토양 자원에 대한 경쟁력이 강 하고 다른 종은 햇빛에 대한 경쟁력이 강하여 두 종의 공존이 가능 하다. 상호 작용의 변동성은 개체군의 크기 구조에서 비롯된다. 상 이한 서식처에서 경쟁적 상호 작용에 대한 식물의 형태와 생활사의 영향을 조사하기 위해 이 장에서 제시된 모델을 제 4 장에서도 사용 하였다.
제 4 장 분배 패턴과 생활사 식물의 자원 획득과 분배에 관한 정량적인 이해를 통해 경쟁자나 포식 자가 함께 있는 주어진 환경 속에서 그 식물이 생존에 성공할지의 여부 를 예측할수 있다. —H. A . Mooney ( 1972, p. 315) 1 서론 제 3 장에서 설명한 바와 같이 식물이 뿌리와 줄기로 자원을 분배 하면 최대 영양 생장률이 감소된다. 그럼에도 불구하고 식물의 형태 는 다양하게 진화되어 왔다. 이 장에서 나는 식물의 형태와 생활사 의 진화에서 토양의 자원 공급률과 손실률의 역할을 연구하기 위하 여 3 장에서 개발한 영양염류와 광에 대한 식물의 경쟁 모델을 사용 하였다. 식물의 상관과 서식처의 토양형 및 기후와 같은 물리적 특
성 간에는 밀접한 관련이 있다는 것이 오래전부터 알려져 왔다 (Raunkie r 1934, Bi lling s 1938, Mooney 1972). 이러한 상관 관계를 식물 이 가지고 있는 잎-줄기-뿌리의 대응책으로 설명할 수 있는가? 이 러한 대응책은 영양염류 공급률과 손실률이 정해진 소규모 지역에 서만 그 식물이 경쟁에 이킬 수 있도록 각 식물의 형태를 필연적으 로 제약하는가? 이 장에서 나는 모델 ALLOCATE 를 사용하여 수많은 경쟁 관계 를 조사하였다. 나는 이 중 일부를 등구배곡선 방법과 비교하긴 하 였지만 다음 절의 주목적은 토양 자원:광 구배 그리고 손실 구배에 따라 식물 형태와 식물의 경쟁 능력 사이에 존재할 수도 있는 광범 위하고 일반적인 관계를 결정하기 위한 것이다. 경쟁의 동태에 대한 분배 패턴의 효과를 포함하여 ALLOCATE 가 가지고 있는 다른 특 성은 이 책의 뒷부분에 설명되어 있다 . 이 장에서는 잎, 줄기, 뿌리 로 분배되는 광합성 산물의 양을 제의하고 다른 모든 특징이 같은 다종경쟁의 경우만을 고려하려 한다. 이것은 모든 식물이 갖어야 할 대응책을 나타내 주기 때문에 중요하다. 2 분배와 형태 식물의 분배 패턴은 자신의 형태에 많은 영향을 준다. 식물의 잎, 뿌리, 줄기의 상대적인 양은 이것들이 생산되고 소실되는 상대적인 비율에 의하여 결정된다. 이 장에서 나는 식물체의 손실은 호흡에 의해서만 일어나며, 잎. 뿌리. 줄기의 호홉률은 똑같다고 가정하였다. 이러한 제한을 두면 분배와 식물의 형태는 동의어가 된다. 그러나 대부분의 실제 식물에서는 분배와 식물의 형태가 같은 것은 아니다. 식물의 형태가 분배 패턴에 의해 많은 영향을 받는다는 것을 알면
이 장에서 제시된 결과는 식물 형태의 차이에서 비롯되는 것으로 더 잘 설명될 수 있을 것이다 . 이 장에서 설명된 식물의 분배 패턴 혹은 형태는 두 가지의 다른 관점에서 생각해 볼 수 있다 . 첫째, 각 식물의 형태(즉 각각의 독특한 분배 패턴)는 서로 상이한 종을 대표하는 것으로 볼 수 있다. 같은 종 내의 개체들이 형태가 다를 경우도 있지만 이러한 종 내의 형태 적인 변이는 종간에 비해 아주 적다. 따라서 첫번째 가정으로 각 종 은 실생으로부터 성체에 이르기까지 영양 생장을 하는 동안 뿌리, 앞 줄기로 일정한 분배율을 갖는 형태적으로 고정된 생활형을 갖는 다고 생각할 수 있다 . 두번째 각각의 식물 형태는 주어진 종내에서 상이한 유전자형으 로 보아야 하며, 이 장에서 제시된 결과는 각각의 유전자형에게 유 리한 서식처(자원 공급률과 손실률로 정의된)를 설명하는 것으로 생 각할 수 있다. 이것은 상이한 종 사이에서와 마찬가지로 특정한 종 의 유전자형 사이에서도 자원 공급률 및 손실률과 관련된 똑같은 형태적인 변이 패턴을 예상할 수 있다는 것을 제시하고 있다. 또한 각 식물 개체 내의 형태적인 가변성 패턴이 자원의 공급률과 손실 률에 대해서도 비슷한 의존성을 보일 것이라고 기대하는 것은 논리 적으로 타당성이 있는 것으로 보인다. 그러나 이 후자의 가능성은 제 9 장에 설명되어 있다. 3 분배 패턴과 동구배곡선의 위치 먼저 분배 패턴을 제외한 모든 것이 동일한 6 종의 식물을 생각해 보자. 그림 4.1 에는 이 식물들의 분배 패턴과 ALLOCATE 로부터 나 온 자원 의존적 생장 등구배곡선이 나와 있다 . 분배 패턴은 등구배
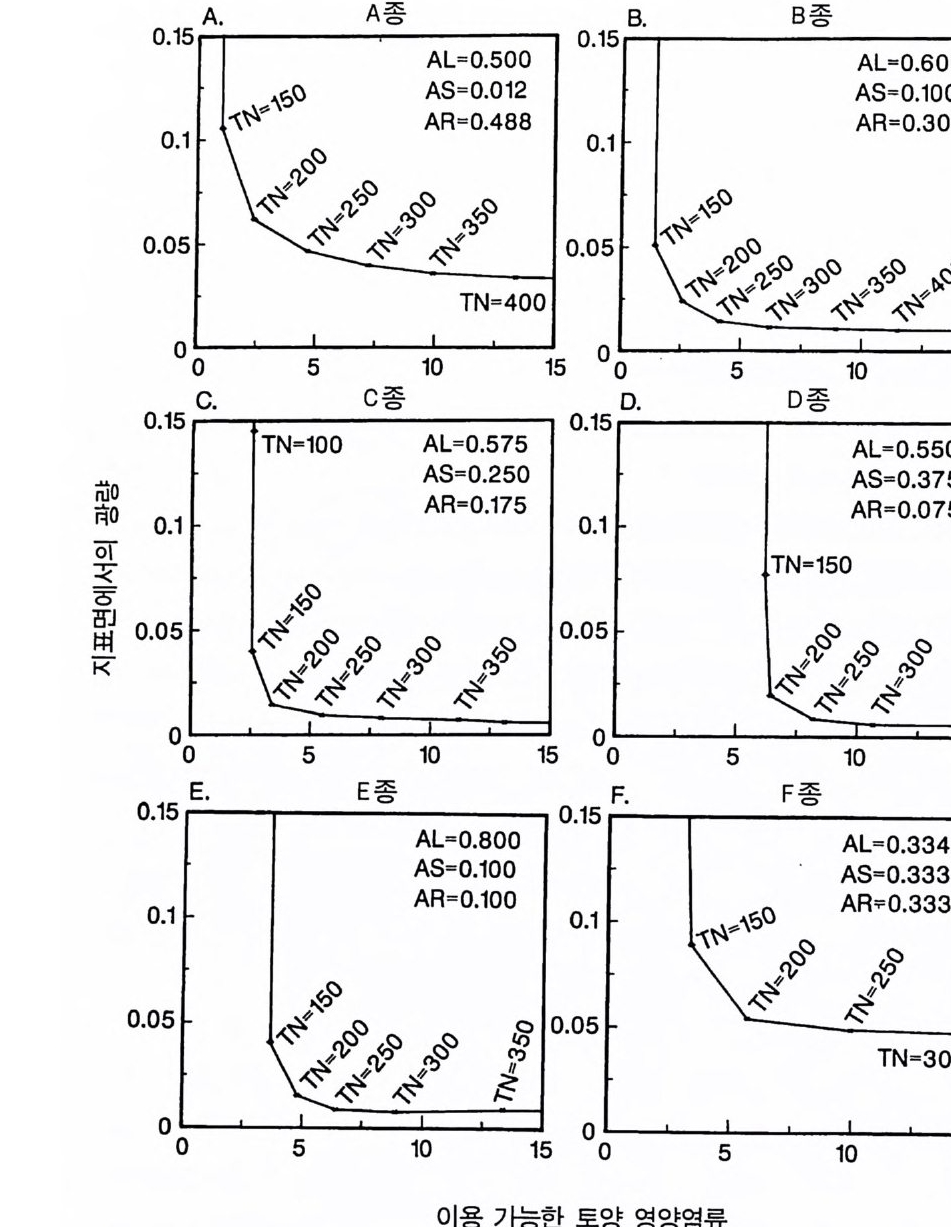 0.15 A. A 종 0.15 8. B 종
0.15 A. A 종 0.15 8. B 종
그림 4.1 잎 (AL) , 뿌리 (AR) , 줄기 (AS) 로의 분배 패턴만 다르고 나머지 특성은 모 두 동일한 6 종의 자원 의존적 인 등구배곡선들. 이들 각 등구배곡선상의 다양한 점을 유도하는 영양염류 공급률 (TN) 과 함께 분배 패턴이 열거되 어 있다. 등구배곡선은 지표면의 광량을 이용하였다. 등구배곡선의 위치는 뿌리, 잎, 줄기로의 분배 패턴에 의해 영향을 받는다. 뿌리로의 분배량이 증가하면 등구배곡선이 왼쪽 윗부분으로 이동하여 종이 토양의 영양엽류 에 대한 경쟁력은 우세하나 햇빛에 대한 경쟁력은 약하게 된다 . 잎과 줄 기로의 분배가 증가하면 등구배곡선이 반대 방향으로 이동하여 일반적으 로 빛에 대한 경쟁력은 증가하나 영양염류에 대한 경쟁력은 감소한다. 6 가지의 분배 패턴은 4 종(종 A-D) 에게는 경쟁력이 강하고 두 종(종 E, F) 에게는 경쟁력이 약한 분배 패턴을 갖도록 선정되었다. 이들 등구배곡 선들은 불안정한 평형접을 유도한다.
곡선의 모양과 위치에 뚜렷한 영향을 준다는 것을 알 수 있다. 일반 적으로 토양 영양염류가 낮은 조건에서는 뿌리로 분배가 큰 식물이 잎과 줄기로 더 많은 양을 분배하는 식물에 비해 더 잘 생존할 수 있댜 이것을 알아보기 위해서는 뿌리에 분배량이 가장 높은 A 종의 등구배곡선의 위치를 다른 종의 등구배곡선의 위치와 비교해 보기 바란다(그림 4.2A). 잎과 줄기로의 분배량은 개체가 지표면의 광량 이 낮은 서식처에서 생존할 수 있는 능력에 영향을 준다. 이러한 6 개의 서로 다른 분배 패턴을 갖는 종들이 부족한 토양 자 원의 공급률만을 제의하고 다른 조건이 동일한 다양한 서식처에서 경쟁할 때 A 종은 영양염류가 가장 적은 서식처에서 우점하고, B 종 은 영양염류가 많은 지역에서, C 종은 더 많은 지역에서, 그리고 D 종 은 영양염류 공급률이 가장 높은 지역에서 우점하게 된다(그림 4.2B). E 종과 F 종은 모든 서식처에서 부족한 토양 자원의 공급률과 는 무관하게 다른 종으로 교체된다. 이러한 결과는 장기간의 경쟁 결과에 근거를 둔 것이며, 일련의 변동을 평균한 것이다. 한편 크기 구조를 갖는 개체군에서의 이러한 경쟁 모델은 장기적으로 생산력 구배를 따라 토양 영양염류의 평균 수준은 증가하고 지표면의 광량
 겁 O 0.15(A 1. 1 幡 古
겁 O 0.15(A 1. 1 幡 古
그림 4.2 (A) 그림 4 .1의 종 A-E 의 둥구배곡선을 같은 그림에 표시한 것. 점들은 두 종 평형점을 나타내는데 , 이들 각각은 불안정하다. (B) 그림 4.1에 있 는 6 종 사이의 장기간 경쟁 결과를 ALLOCATE 로 예측한 것. E 종과 F종 은 이 손실률에서 영양염류 공급률 구배상의 모든 서식처에서 교체됨을 주목하기 바란다. A, B, C, D 종은 이 구배상에서 자신들의 둥구배곡선에 의해 결정된 순서로 분리되었다 . 이들 종들이 안정적으로 공존하는 공급 률 (TN) 은 없었다. (C) 이 6 종 사이의 경쟁이 진행되는 동안 지표면에서 토양 자원과 광량이 감소되는 장기간의 평균 수준이다.
은 감소함을 보여주고 있다(그림 4.2C). 이 6 종 사이의 분배 패턴에 차이가 있다고 하여 생산력 구배상에 서 서로 다른 두 종이 안정한 상태로 공존하는 것은 아니다(그림 4.2B). 오히려 그림 4 . 1 의 다양한 TN 수준에서 나타나는 R * 를 사용 하는 그림 3.11D 에서 볼 수 있는 바와 같이 각각의 두 종 평형점은 지역적으로 불안정하다. 예를 들면 TN= 150 에서 A 종의 R* 는 그림 4.2A 에 l 로 표시되어 있는 평형점 위에 있고, B 종은 아래에 있다. A 종의 R * 는 그것이 지표면의 광량과 토양의 영양엽류 수준을 충분히 감소시킬 수 있어 TN=150 인 서식처에서 A 종이 다른 모든 종울 교 체할 수 있다는 것을 보여주고 있다. 그러나 B 종의 R* 도 TN=150 인 서식처에서 역시 다른 종을 교체할 수 있음을 보여주고 있다 . 따라 서 이 종들은 두 종 사이에 안정된 평형점을 가지고 있지 않으며, TN= 150 에서 그들의 상호 작용의 결과는 초기 조건에 따라 달라진다. 그림 4.2B 를 도출하는 데 사용된 모의에서 각 종은 동일한 밀도 를 갖는 실생으로부터 출발하였으며, TN=150 에서 B 종이 A 종울 교 체하였다. B 종과 C 종도 C 종과 D 종에서처럼 두 종 사이에 안정된 평형점이 없다 . 그러나 각 종은 출발 조건과는 무관하게 자신의 경 쟁력이 가장 우세한 토양 자원 공급률 (TN) 의 범위를 가지고 있다. 예를 들면 A 종은 TN= 100 이하에서 다른 모든 종을 교체할 수 있 으며, B 종은 TN 이 160-180 사이에서, C 종은 220-250 사이에서, 그
리고 D 종은 500 이상에서 다른 종을 교체할 수 있다 . 이러한 결과는 토양 자원과 광이 제한 자원이 되고 있는 균일한 서식처에서 분배 패턴의 차이가 안정 상태의 공존을 유도하지 못한 다는 것을 보여주고 있다. 이것은 분배 패턴과 자원 소비율 사이의 강한 연관 때문이다. 식물이 두 가지의 자원에 대하여 경쟁할 때 두 종의 평형점에서 각 종이 자신을 더 제한하는 자원울 비교적 더 많 이 소비할 때만 안정 상태의 공존을 유지할 수 있다 (T i lman 1982, 그림 2.8 혹은 3.1 1). 그러 나 하나의 기 관에 더 많은 자원을 분배 하는 것은 그 기관이 획득하는 자원이 부족하게 될 가능성이 감소된다. 왜냐하면 분배가 많으면 다른 자원의 소비량에 비해 그 자원의 소 비량이 증가하기 때문이다. 분배 패턴만이 서로 다른 종의 경우 한 종은 어느 한 특정한 자 원의 소비율을 높힘으로써 그 자원에 대한 경쟁 능력을 얻는다 . 이 것은 두 종의 평형점에서 그 종이 그 특정한 자원에 의해 제한되지 않고 또한 그 특정한 자원을 더 많이 소비할 것임을 의미한다. 따라 서 각 종이 두 종의 평형점에서 자신을 제한하지 않는 자원을 비교 적 더 많이 소비하려는 경향이 있으면 평형점은 불안정하게 될 수 있다 . 영양염류에 대한 경쟁력이 큰 종은 조직의 영양염류 요구 수준을 낮추어 토양 요인과 광이 제한되는 서식처에서 여러 가지 종 쌍이 안정 상태로 공존할 수 있도록 해준다 . 그러나 광합성 산물이 뿌리, 잎, 그리고 줄기로 차등적으로 분배되는 것은 안정 상태의 공존과 관계가 없다. 이러한 모의는 두 가지 중요한 점을 보여준다. 첫째, E 나 F 와 같 은 분배 패턴을 갖는 종은 주어진 토양 자원:광 구배의 어느 점에 서든지 생존할 수 있는 경쟁자는 아니다. 둘째, 토양 자원:광 구배의 어느 한 지역에서 경쟁력이 강한 분배 패턴은 구배상의 다른 점에
서는 경쟁력이 약하다 . 이러한 결과들은 또한 경쟁의 정성적인 결과 와 평형점의 안정성이 그 종들의 자원 의존적인 등구배곡선과 R * 에 의해 예측될 수 있음을 시사하고 있다 . 4 경쟁력이 강한 형태 특정한 서식처에서 어떤 분배 패턴이 경쟁에 유리한지를 결정하 기 위해서는 다른 조건은 똑같고 분배 패턴(형태)만 다른 많은 개체 들울 경쟁하도록 하는 것이다. 분배 패턴에 의해 결정되는 각 개체 의 서로 다른 형태는 상이한 종으로 간주할 수 있고, 아니면 단성생 식을 하는 개체군 내의 유전자형으로 간주할 수도 있다. 이러한 모 의를 수행하기 위한 또 하나의 간단한 방법으로는 Iwasa 와 Rou g hg arden(1984) 이 한 것과 같은 최적화 과정을 사용하는 것이다. 그들의 연구는 형태와 서식처 사이의 관계에 관한 몇 가지 중요한 내용을 제공한다. 그러나 이러한 최적화 방법은 자원 수준이 고정된 것으로 전재하기 때문에 자원 경쟁의 과정이 포함되어 있지 않다. Iwasa 와 Rou gh g arden(1984) 이 지적한 것처럼 최적 조건은 자원 수준에 따라 다르다. 경쟁 조건하에서는 경쟁하고 있는 종에 의해 자원 수준이 결정된다. 최적화 방법이 몇 가지 내용을 제공하긴 하 지만 이 방법은 경쟁을 유발시키는 원동력인 자원 수준에 대한 생 장의 피드백 효과가 포함되어 있지 않다. 따라서 나는 어떤 형태가 특정한 환경 조건에서 경쟁에 가장 유리한가룰 결정하기 위해 동적 인 자원 경쟁 모델을 이용하기로 결정하였다. 이러한 모의에서는 분배를 제외한 모든 변수를 일정하게 한다. 따 라서 두 종의 안정된 평형점이 없다. 나는 먼저 종간 경쟁이 없을 때 어떤 분배 패턴이 생존할 수 있는지를 결정하였다. 죽 다른 분배
패턴을 갖는 종과의 경쟁이 없을 때 어떤 분배 패턴이 개체군을 유 지할 수 있는지(생물량 >O) 를 결정하였다. 이를 위해 나는 서로 다른 171 종에서 잎 줄기, 그리고 뿌리로의 분배 패턴의 전체 범위에 그 종들이 균일하게 위치하도록 분배변수들을 선정하였다. 각각의 상이 한 분배 패턴은 그림 4.3 의 분배 삼각형에 점으로 표시될 수 있다. 이 그림에서 하나의 점은 잎 (x 축)과 줄기(y축)로의 분배비를 나타낸 댜 식물이 성숙할 때까지 새로운 생장은 잎이나 줄기 혹은 뿌리로 분배되기 때문에(성숙하면 호흡을 하고 남은 모든 광합성 산물을 종자 로 분배한다) 잎 . 줄기, 뿌리로의 분배비를 모두 합하면 1. 0 이 되어야
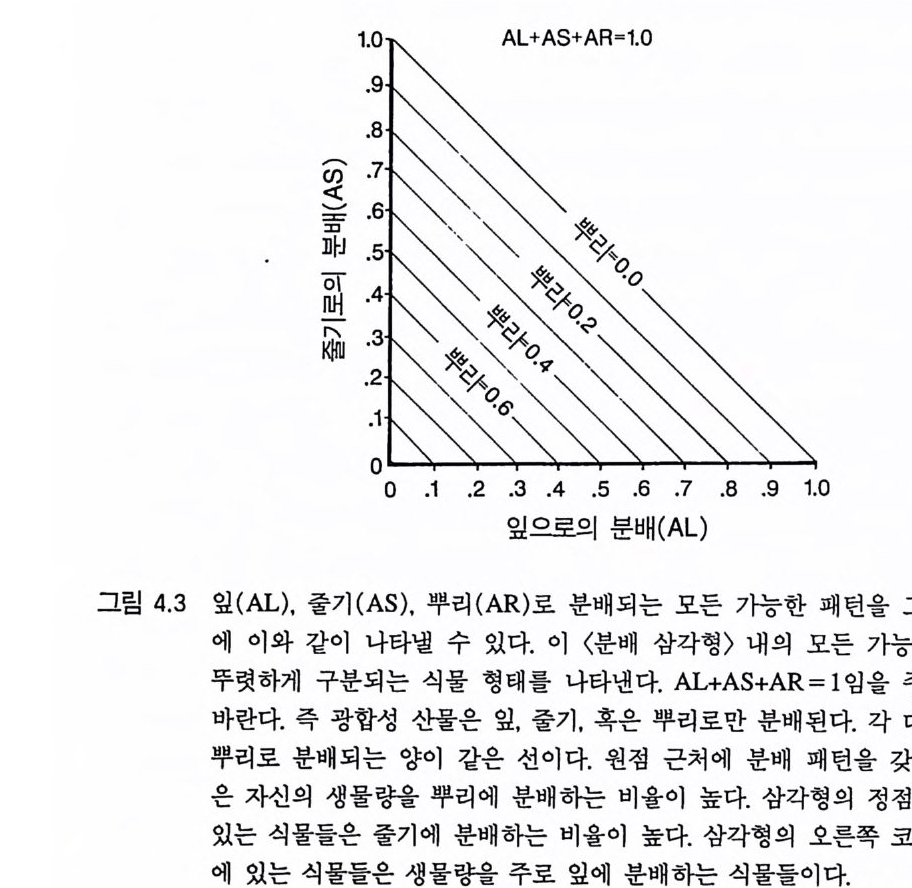 993rsAL5+ A SA+AR3= 1 .20
993rsAL5+ A SA+AR3= 1 .20
한다. 이것은 잎과 줄기로의 분배비를 합하여 이 값을 l 에서 빼면 뿌리로의 분배비를 알 수 있음을 의미한다. 이러한 내용을 간단히 그래프로 나타낼 수 있다. 그림 4.3 의 분배 삼각형에서 각각의 대각선은 뿌리로의 분배가 일정한 선이다 . 그림 4.3 의 삼각형상에 있는 각 점은 하나하나가 서 로 다른 식물 형태이다. 줄기로의 분배가 적은 식물은 로제트형을 갖고, 뿌리:잎의 비가 큰 로제트(왼쪽으로)로부터 뿌리:잎의 비가 낮 은 로제트(오른쪽으로)의 범위를 갖는다. 줄기로의 분배가 큰 식물은 (삼각형의 정점 쪽) 키가 크고 직립성인 생장형을 갖는다. 줄기로의 분배는 식물이 얼마나 빨리 길이 생장을 할 수 있는지를 결정하며, 따라서 식물의 평균 높이를 결정한다. 이 171 종의 식물을 각각 특정한 서식처, 죽 특정한 손실률과 영양 염류 공급률을 갖는 서식처에서 단독으로 평형 상태에 이를 때까지 생장하도록 하면 생존력이 없는 식물이 나타난다. 이 생존력이 없는 식물은 종간 경쟁이 없는 서식처에서도 자체 유지를 할 수 없다 . 이 와같이 생존력이 없는 형태들은 그림 4 .4에서 빗금찬 부분으로 표시 되어 있다. 이 식물들은 호흡과 사망에 의한 손실률이 생장률보다 크기 때문에 생존할 수 없다. 종간 경쟁이 없는 비옥한 토양에서는 넓은 범위의 분배 패턴이 생존 가능하였다. 척박한 토양 (TN=50) 에서 생존력이 있는 식물은 단독 재배시 줄기로의 분배가 생존에 그리 중요하지 않지만 뿌리와 잎으로의 최소 분배량이 있음을 나타내고 있다. 이러한 최소 분배 수준은 토양의 비옥도가 증가함에 따라 감소한다. 비옥한 토양 (TN=lOO) 에서 경계는 거의 직선이 되는데, 잎으로의 분배 비율이 가장 중요한 변수가 된다. 이 모의에 사용된 변수에서, 잎으로의 분배가 20% 미만인 식물은 호흡이나 다른 소실량을 능가 할 수 있는 광합성을 하기에 충분한 잎 면적을 갖지 못한다. 생존력
 전체 토양 영양염류 =125
전체 토양 영양염류 =125
그림 4.4 특정한 분배 패턴을 갖는 식물을 단일 종 지소에서 자라게 하면 일부 식 물은 살지 못한다 . 즉 , 종간 경쟁이 없을지라도 특정한 서식처에서 식물이 자신을 유지할 수 없다 . 개체군이 생존하지 못하도록 하는 모든 분배 패 턴들은 이 그림의 빗금천 부분에 위치한다 . 영 양염류 공급 률 이 높은 서식 처에서는 생존할 수 있는 형태의 범위가 증가한다. 모든 경우에서 손실률 은 0.2/wk 였다 . 결과들은 ALLOCATE 를 이용한 모의에 근거를 두고 있 다. 광합성률이 높거나 뿌리와 줄기의 호흡률이 낮으면 생존할 수 없는 형태의 범위가 더 좁을 것이다. 이들 그림들은 그림 4.5-4.8 의 모의에 사 용된 특정한 일단의 변수를 갖는 환경에서 어떤 분배 패턴이 생존 가능한 지를 보여준다 .
이 있는 분배 패턴과 생존력이 없는 분배 패턴 사이의 실제 경계는 각 조직의 호흡률, 사망률 혹은 손실률, 그리고 광합성이 영양염류 와 광에 얼마나 의존적이냐에 따라 달라진다. 이러한 분석을 통해 뿌리, 잎 줄기로의 분배와 뿌리 및 줄기의 호흡을 보상하기 위해 충 분한 양의 잎이 필요하다는 것 사이에 정성적인 관계가 있음을 알 수 있다. 더구나 이러한 분석은 생존력이 있는 형태의 범위가 넓다 는 것을 시사하고 있다. 이러한 생존력 있는 형태들이 서로 상대편 과 경쟁할 때는 어떻게 살아갈 수 있을까? 4-1 생산력 구배상에서 분배와 경쟁 단지 형태(분배 패턴)만이 다른 식물 사이의 경쟁 결과를 결정하 기 위해 , 생존력이 있는 분배 패턴 중에서 200 개의 서로 다른 분배 패 턴을 선정 하였다. 그런 다음 ALLOCATE 모델을 사용하여 200 개 의 서로 다른 형태를 부족한 영양염류와 광 조건에서 동시에 경쟁하 도록 하였다. 모델의 크기와 복잡성 때문에 Cray 2 슈퍼컴퓨터를 사용 해서만 이것을 풀 수 있었다. 모든 모의는 40 년 동안으로 계산되었다. 주어진 영양염류 공급률 (TN) 과 손실률에서 하나 혹은 두 개의 형 태가 경쟁적으로 우세하여 나머지 200 종 모두를 교체하였다. 두 종 이 우점할 경우 이들은 매우 유사한 분배 패턴을 갖는 것으로 나타 났다. 이러한 사실은 하나의 분배 패턴이 어떤 주어진 서식처에서 높은 경쟁 능력을 유도한다는 것을 시사하고 있다. 이것은 또한 생 태적으로 비슷한 종이 경쟁에 의한 교체가 일어나기 전에 오랫동안 공존할 수 있음을 보여주는 것이다 (Hubbel & Foste r 1986). 나는 경쟁적으로 우세한 형태, 즉 주어진 서식처에서 다른 모든 개체를 교체할 수 있는 형태를 그 서식처에서 우세한 형태 또는 우 세한 분배 패턴이라고 부른다. 나는 생산력 구배 혹은 손실률 구배
롤 따라 하나의 우점 형태에서 그 다음의 형태로 매끄럽게 넘어가 기 위해서는 많은 종 (200) 을 사용해야 함을 알았다 . 다양한 영양염류 공급률에서 다종 경쟁의 모의를 수행함으로써 , 나는 우세한 분배 패턴이 서식처의 생산력에 얼마나 의존적인가를 결정하였다 . 그림 4.5 에 있는 곡선은 광에 대한 토양 영양염류의 구 배를 따라 상이한 점에서 우세한 경쟁 능력을 갖는 분배 패턴들을 보여주고 있다. 생산력이 매우 낮은 서식처 (TN=25 와 50) 에서, 우세 한 분배 패턴은 뿌리와 잎에 분배량이 많고 줄기에 분배되는 양이
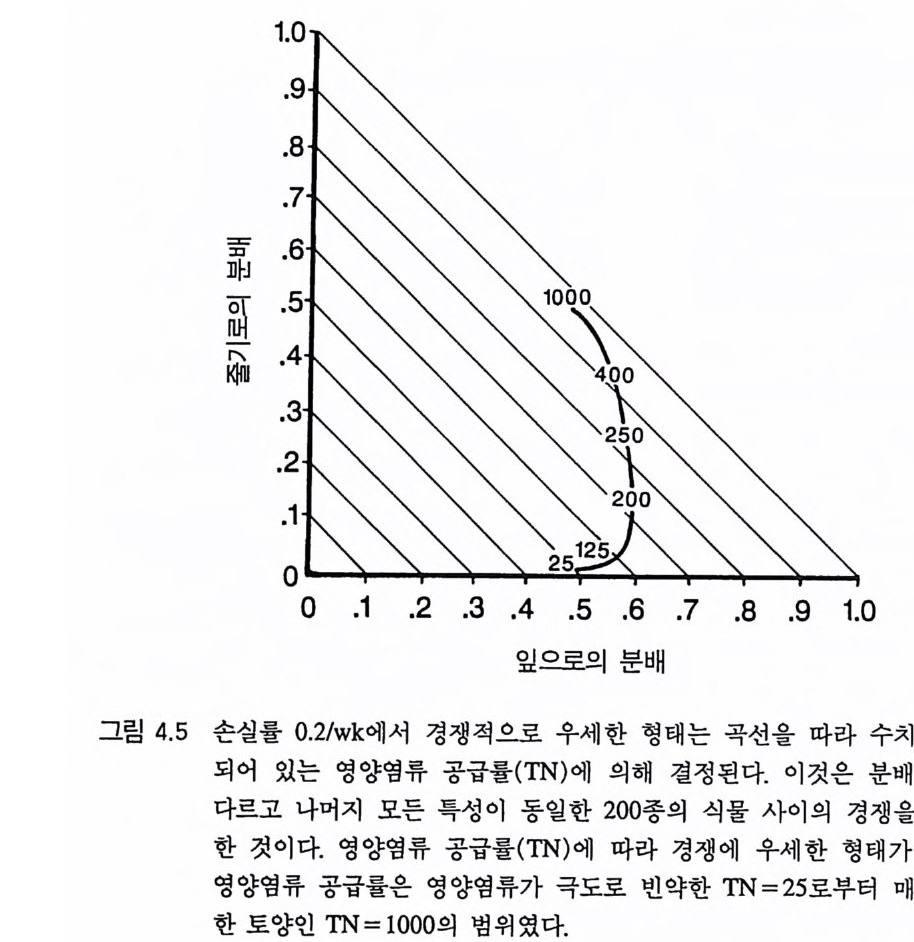 1.0
1.0
적댜 영양염류 공급률이 높은 서식처 (TN=150) 에서는 줄기에 대한 분배량이 약간 많은 것이 유리하지만 주요한 변화는 뿌리의 분배량 이 적고 잎으로의 분배량은 더 적다 . 따라서 영양염류가 빈약한 서 식처에서는 영양염류 공급률을 증가시키면 잎을 위해 뿌리를 포기 하는 식물에게 유리하다. 영양염류를 더욱 증가시키면 주로 뿌리로 분배되는 양을 줄여 줄기로 더 많이 분배하는 것이 유리하고 매우 비옥한 토양에서는 생산력이 더욱 증가되면 잎으로 분배될 양을 줄 여 주로 줄기로 분배되는 양이 증가된다. 이러한 결과는 토양 자원과 광에 대한 식물의 경쟁 능력이 식물 의 형태, 죽 앞 줄기 혹은 뿌리로 분배되는 생물량의 비율에 매우 의존도가 높음을 보여주고 있다 . 모든 가능한 형태 중에서 단지 하 나의 특정한 분배 패턴만이 특정한 서식처에서 우세한 경쟁 능력을 갖는다. 따라서 하나의 영양염류 공급률에서 경쟁력이 높은 식물은 이보다 높거나 낮은 공급률에서는 다른 형태를 갖는 식물에 비해 경쟁력이 낮다. 부족한 토양 영양염류의 상이한 공급률에 대한 식물의 반응은 (그림 4.5 ) , 이용 가능한 토양 영양염류와 지표면에서의 광량 사이 에서 역구배를 유도하였다(그림 4.6A). 전체 식물량과 토양의 영양 염류 수준은 영양염류 공급률에 따라 증가하지만 지표면의 광량은 감소한다. 이 구배에서 뿌리로의 분배량은 감소되고 줄기로의 분배 량은 증가한다. 그리고 잎으로의 분배량은 영양염류 공급률이 중간 정도일 때 최대에 달한다(그림 4.6 B ). 그림 4.5 와 그림 4.6 에 요약되어 있는 결과들은 조사 대상으로 한 200 종의 식물들이 갖는 일단의 생리적 상수들, 일단의 손실률, 성숙 했을 때의 키 등으로부터 유도되었다. 나는 이것과 똑같은 손실률에 서 이와 비슷한 두 가지의 다른 모의를 수행하였는데(변수는 다르지 만 분배 패턴을 제외하고 각 모의의 종 구성은 동일함) 역시 분배 패턴
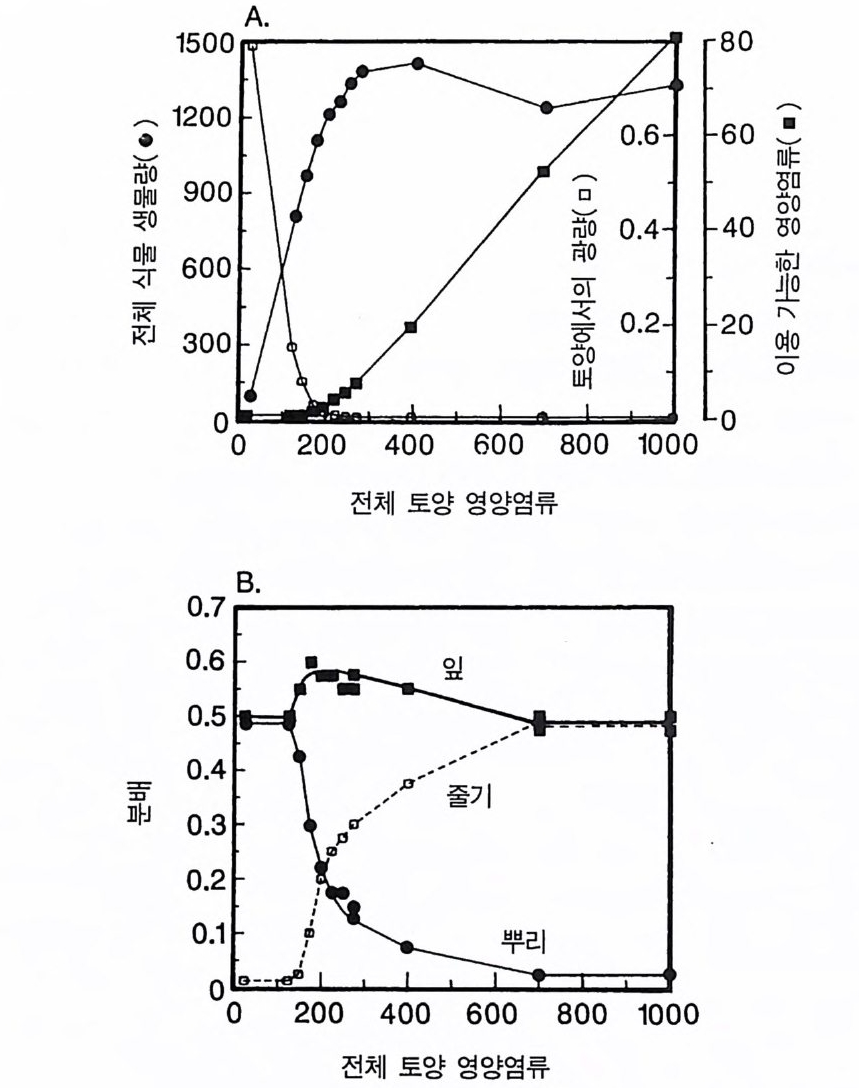 80
80
그림 4.6 (A) 그림 4.5에 예시된 경우에서 장기간의 평균 생물량( •), 토양 영양염 류 수준( • ). 그리고 지표면에 입사되는 광량( □ ) 이 영 양염류 공급률에 대한 의존도. (B) 그립 4.5 에 예시된 경우에서 서식처의 영양염류 공급률 의 함수로 주어지는 경쟁에 우세한 형태를 갖는 식물의 잎. 뿌리, 그리고 줄기로의 분배.
이 영양염류의 공급률에 의존적임을 알 수 있었다 . 각 조직을 생산하고 유지하는다] 필요한 경비, 구배상의 각 점에 서 우점하는 식물이 성숙하였을 때의 키, 그 식물의 줄기 및 잎의 무게에 관련된 구조적 제약, 그리고 광합성에 영향을 주는 햇빛과 영양염류를 포함하는 다른 생리적 변수들의 차이에 따라 정량적인 분배 패턴은 달랐다. 그러나 이들 각각의 요인들이 생산력 구배상에 서 예측되는 정량적인 분배 패턴에 영향을 줄 수 있긴 하지만 동일 한 일반적인 힘이 모든 생산력 구배상에서 분배 패턴에 영향을 준 다. 영양염류 공급률이 증가하면 뿌리로의 분배는 감소한다. 영양염 류가 풍부하고 광량이 적은 서식처에서 비광합성 조직으로의 분배 는 증가하는데, 그 이유는 광량이 적은 서식처에서는 식물의 키가 신속히 자라는 것이 생존에 매우 중요하기 때문이다. 이 두가지 힘 에 의해 영양염류의 공급률이 중간 정도인 서식처에서 잎으로의 분 배가 최대가 되기도 한다. 토양 자원:광 구배상의 각 점에서 예상되는 상이한 분배 패턴은 부족되는 자원에 대한 종의 경쟁력에 영향을 줄 뿐만 아니라 그들 의 최대 생장률에도 영향을 준다. 각각의 분배 패턴과 연관된 예상 되는 최대 생장률이 그림 4.7 에 나와 있다. 제 3 장에서 언급한 간단 한 이론(식 3.5) 에 의한 예측대로 이러한 최대 영양 생장률은 새로 운 광합성 기관의 생산에 분배되는 광합성 산물의 비율과 거의 일 치한다. 최대 생장률의 차이는 이러한 식물의 개체군 동태에 중대한 영 향을 준다(제 6 장 참조). 4-2 손실률 구배상에서의 식물 형태 앞에서 언급한 식물과 동일한 식물을 이용하여 손실률이 서로 다 른 서식처에서 비슷한 분석을 실시하였다. 손실은 개체의 크기나 분
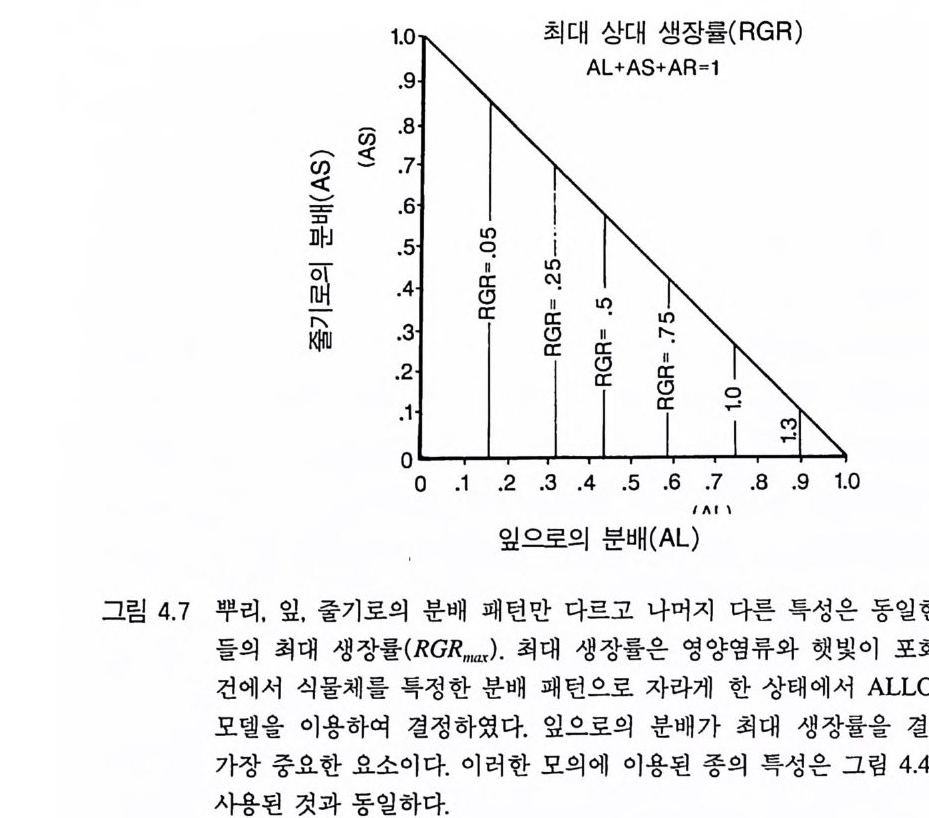 1.0 ~ 최대 상대 생장률 (RGR)
1.0 ~ 최대 상대 생장률 (RGR)
배 패턴과는 무관하게 모든 개체에 균등하게 적용되는 밀도 독립적 인 사망률이었다. 각각의 손실률에 대해 나는 200 개의 전체 패턴 중 에서 어떤 분배 패턴이 토양 영양염류:광 구배상의 각 점에서 경쟁 에 가장 우세한 분배 패턴인지를 결정하였다. 분석은 ALLOCATE 모델을 사용하여 모든 200 종(형태)이 영양염류와 햇빛에 대하여 경 쟁을 하도록 이루어졌다. 그 결과 각각의 손실률에서 유리한 분배 패턴이 서로 달랐다(그림 4.8). 높은 손실률은 잎으로의 분배량이 증 가하고 뿌리와 줄기로의 분배량울 감소시키는 쪽으로 경쟁에 유리 한 분배 패턴을 변화시켰다. 그림 4.8 에 제시된 자료는 다른 측면에서 고려할 수도 있다 . 죽
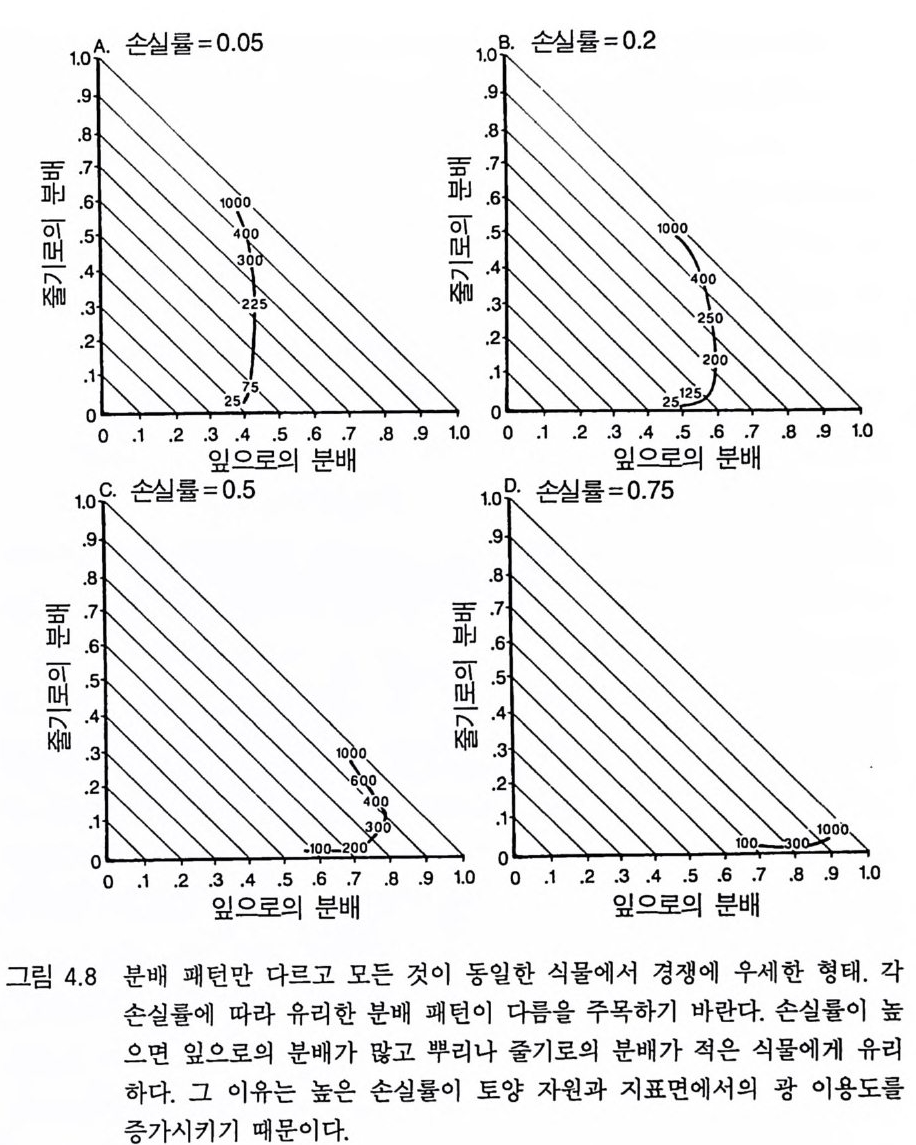 1.0 A. 손실률 =0.05 1 9 요93찍7률 6=I 5。 2A 32A
1.0 A. 손실률 =0.05 1 9 요93찍7률 6=I 5。 2A 32A
주어진 토양형에서 유리한 분배 패턴에 대한 손실률의 효과를 나타 내는 것으로 생각할 수 있다(그림 4.9). 높은 손실률은 생물량을 잎 에 더 많이 분배하고 뿌리와 줄기에 더 적게 분배하는 식물에게 유 리하다. 이것은 높은 손실률이 토양 자원과 광의 이용도를 높게 하
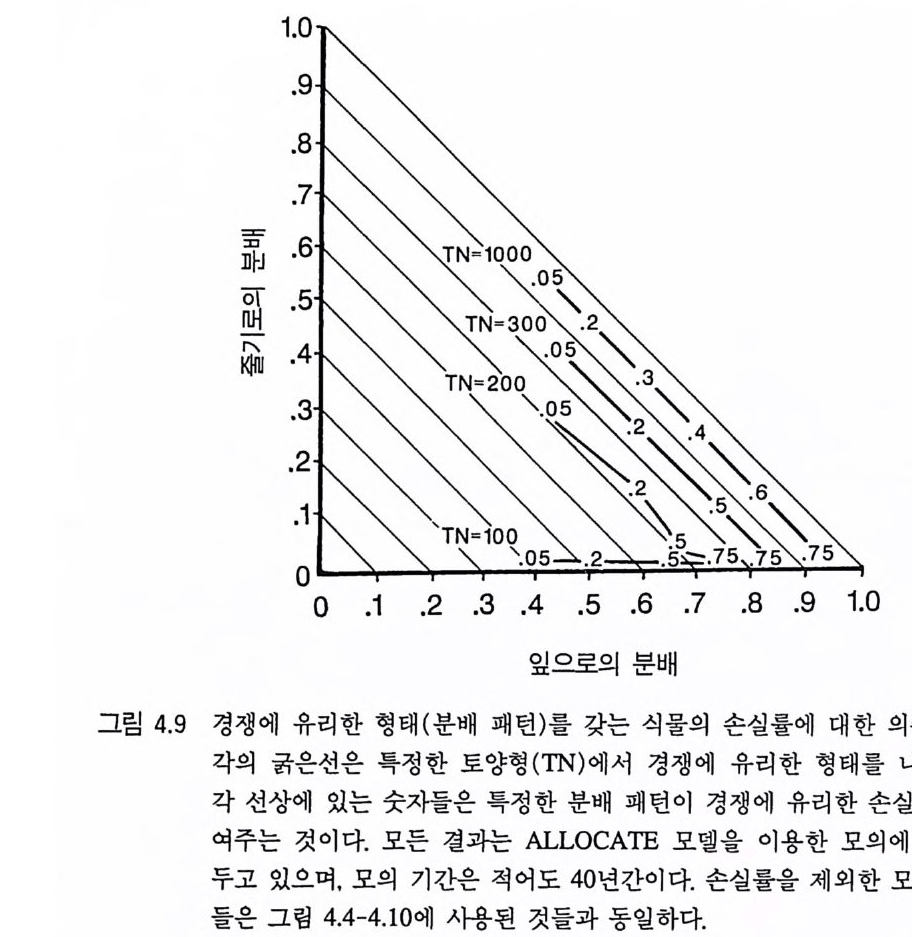 1.o
1.o
여 뿌리와 줄기에 더 많은 생물량을 분배하는 개체들이 받는 선택 적 장점을 감소시키기 때문이다. 손실률이 낮은 서식처(교란되지 않은 서식처)에서부터 손실률이 높 은 서식처에 이르는 연속적인 구배가 있다 . 이 구배상의 주어진 토 양형에서 손실률 증가의 주효과는 뿌리나 줄기에 비해 잎에 더 많 이 분배하는 개체에게 유리하다는 것이다(그림 4.9). 척박한 토양에 서는 줄기로의 분배가 항상 적고, 손실률이 높은 서식처에서는 잎의 성장을 위해 뿌리의 성장을 포기하는 개체가 유리하다(그림 4.9). 비
옥한 토양에서는 손실률이 낮을 때 뿌리로의 분배가 적다. 높은 손 실률에서는 잎의 생물량 증가를 위해 줄기의 생물량을 포기하는 개 체가 유리하다(그림 4.9 ) . 모든 토양형에서 손실률의 증가는 최대 생장률이 높은 개체에게 유리하다. 이것은 손실률 층가가 생존 가능한 분배 패턴의 폭을 감 소시키는 데 일부의 원인이 있다 . 예를 들어 손실률이 0 .5 /wk 인 경 우 그림 4.7 에서 RGR=0 .5 /wk 를 나타내는 선의 오른쪽에 분배패턴 을 갖는 것들만이 생존할 수 있다 . 그러나 설명은 이것보다 더 복잡 하다. 손실률이 높은 서식처는 영양염류와 햇빛의 이용도가 높다. 그 이유는 이들 식물의 생물량이 적기 때문이다. 제 3 장에서 설명한 바와 같이 모든 자원의 이용도가 높은 조건하에서는 생장률이 가장 높은 식물체가 경쟁력이 가장 클 수 있다. 그림 4.7 의 가장 우측 구석에 있는 식물들은 뿌리나 줄기로는 전 혀 분배하지 않고 모든 것을 잎으로만 분배하는 것들이다 . 이러한 식 물에는 토양에 있는 녹조류가 속하는데, 모든 가능한 형태 중에서 상 대 생장률이 가장 높다. 극도로 높은 손실률을 갖거나 극도로 비옥한 토양을 갖는 서식처를 제외한 모든 서식처에서 이러한 식물은 좀더 복잡한 형태를 갖는 종에 의해 경쟁적으로 교체될 것으로 기대된다. 복잡한 형태라는 것은 식물체가 자신의 생장을 제한하는 자원을 더 많이 획득하도록 하는 형태를 의미한다. 경쟁적으로 유리한 형태와 손실률 및 영양염류 공급률 사이의 관계가 그림 4 .1 0 에 나와 있다 . 그림 4 . 9 와 4 .1 0 은 손실률과 토양의 영양염류 공급률의 각 조합에 서 상이한 형태가 우점하는 것을 보여주고 있다. 또한 환경 구배를 따라 우점하는 형태의 점진적인 구배가 있음을 보여주고 있다. 더구 나 광합성량이 호흡이나 사망으로 인한 손실량보다 항상 적어서 생 존할 수 없는 식물 형태를 제외하고는, 특정한 서식처 유형에 따라 가장 경쟁력이 높은 식물 종이 존재한다 . 자연 상태의 서식처는 지
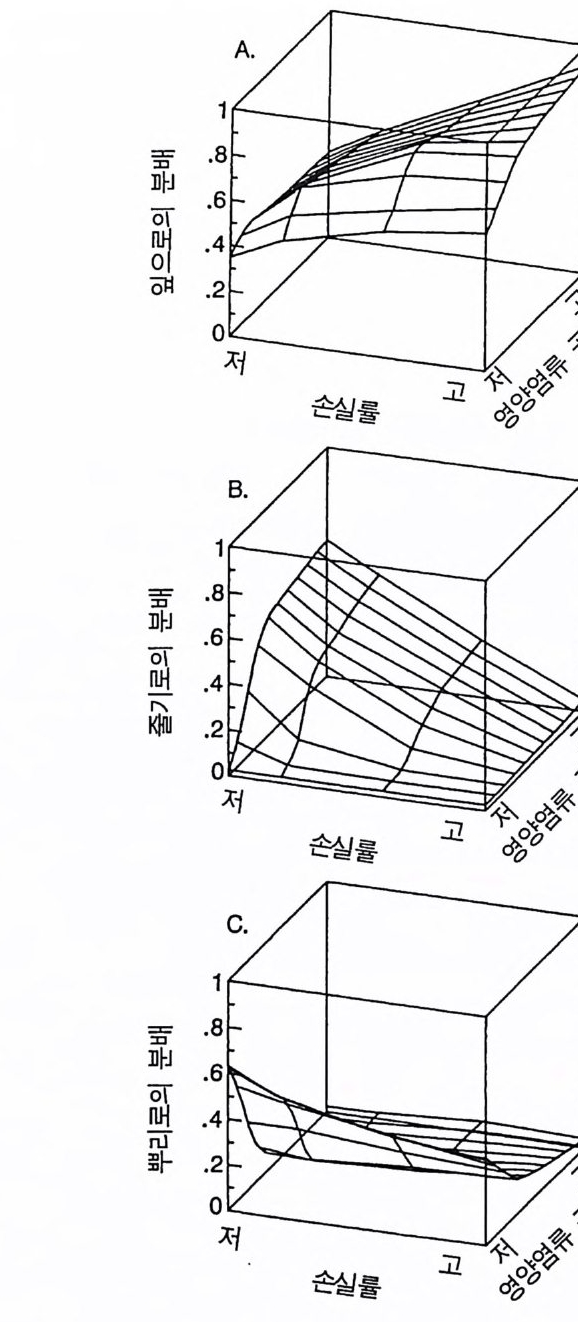 A
A
1 림 4.10 경쟁에 유리한 형태를 갖는 식물이 서식처의 손실률과 토양 영양염류 공급률에 대 한 의존도 (A) 손실률과 영양염류 공급률에 대한 잎으로의 분배가 갖는 의존도. (B) 손실률과 영양엽류 공급률에 대한 줄기로의 분배가 갖는 의존도. (C) 손실률과 영양염류 공급률에 대한 뿌리로의 분배가 갖는 의존도.
소에 따라 영양염류 공급률이 다양하고, 손실(교란, 초식 등)은 우연 적이고 공간적으로 이질적일 수 있기 때문에(서식처 전역에 걸쳐 분 산되어 있는 숲톰 혹은 고퍼 마운드), 그림 4.9 에 제시된 결과는 어떤 주어진 서식처에서 관찰될 수 있는 식물 형태의 범위를 의미한다. 실제 식물의 형태 범위는 평균 영양염류의 공급률과 그 속에 있는 식물의 변이에 의해, 그리고 평균 손실률과 그 손실률에서의 식물의 변이에 의해 결정되어야 한다 . 주어진 식물 군집은 분배 수평면상의 어떤 지역에 속하는 것으로 생각할 수 있다. 그러나 여기에서 제시된 것은 식물의 형태, 토양 영양염류의 풍부 도 그리고 손실률 사이의 관계를 분석하기 위한 최초의 근사치에 지나지 않는다. 다른 여러 종류의 구조적, 생리적, 그리고 환경적인 제약이 이 모델에 포함될 수 있다. 그러나 이 모델은 제한된 서식처 조건의 범위에서 모든 식물체가 우세한 경쟁자가 되기 위해 가지고 있는 대응책을 보여주고 있다. 4-3 생리적인 대응책 식물은 광합성 산물을 뿌리, 잎, 혹은 줄기로 분배하는 대응책을 가지고 있는 것과 똑같이 생리적인 대응책도 가지고 있다. 생리적인 과정은 효소에 의해 조절되는 반응인데, 효소의 수준이 증가하면 생 리적인 반응도 현저히 증가한다. 그러나 하나의 생리적인 반응에만 관련이 있는 어느 하나의 효소 생산량이 증가하는 것은 영양염류의 흡수와 같은 다른 기능에 관련이 있는 효소를 만드는 데 필요한 아 미노산이 줄어든다는 것을 의미한다. 이 책에서 논의된 분배 패턴은 단순히 앞 뿌리, 줄기 등과 같은 여러 가지 기관으로의 분배량의 차 이를 의미할 수도 있다. 그러나 잎, 뿌리, 줄기의 무게에 차이가 없는 상태에서도 식물은
자신의 생리적 특성을 변화시켜 부족되는 토양 자원과 광 조건에서 자신의 경쟁 능력을 변화시킬 수 있다. 식물은 어느 하나의 생리적 반응의 효율을 감소시켜 다른 생리적 반응의 효율을 증가시키기 때 문에 이러한 생리의 변화도 하나의 대응책일 수가 있다 . 이러한 대 응책이 없다면 어느 하나의 식물이 〈 초경쟁적인 종 〉 이 될 수 있을 것이다 . 대응책이 없는 경우 방향성 선택은 더 이상의 증가가 없을 때까지 모든 과정의 효율을 증가시키는 식물에게 유리할 것이다. 여기에서 개발된 모델과 그 모델이 기초하고 있는 대응책은 처음 에 예상했던 것보다 훨씬 일반적이다 . 비록 이 모델에는 생리적인 대응책이 뚜렷하게 포함되어 있지 않지만, 이 모델의 예측 결과는 생리적 대응책이 포함된 모델에 의해 예측되는 결과와 비슷하다. 형 태를 일정하게 유지하면 식물 개체는 영양염류의 효율적인 흡수와 보유 기작 대신에 효율적인 광합성 기작에 단백질(질소)을 분배하는 대응책을 갖는다. 이러한 생각은 Cha pi n(1980) 에 의해 입증된 바 있 는데. 그는 다양한 식물종의 최대 광합성률이 잎의 질소(단백질) 함 량에 비례함을 밝혔다. 5 식물의 키 그림 4 .3에서부터 4.10 까지에 제시된 결과는 줄기의 분배량으로 측정된 식물의 평균 키가 토양의 영양염류 수준에 따라 증가함을 보여주고 있다. 그러나 이 모든 경우에서 모든 식물이 성숙하였을 때의 키는 모두 같았다. 햇빛은 위쪽으로부터 오는 방향성이 있는 자원이기 때문에 키가 큰 개체는 더 많은 햇빛을 받는다 (Hom 1971, Ha rpe r 1977, Gr im e 1979, Giv n is h 1982). 식물의 키에 약간만 차이가 있어도 식물이 받는 햇빛의 양에는 큰 차이가 있을 수 있다 (H arp er
1977). 영양염류가 더 풍부한 서식처에서(주어진 손실 률 에서 ) 햇빛의 이용도는 더 낮기 때문에 자신의 생장을 키를 크게 하거나 잎면적 울 증가시키는 쪽으로 분배하는 개체는 이미 앞에서 살펴본 바와 같이 토양 자원 : 광의 비가 높은 곳에서 유리하다(그림 4.1 0 ). 그러나 키의 증가에 분배하는 식물은 구조상의 비용이 따른다. 직 립한 식물은 기본적으로 그 기부를 땅에 내린 장대와 같다. 키가 커 지면 필연적으로 나무를 지탱할 지하부의 조직에 더 많은 분배가 있어야 한댜 더구나 수직으로 서 있는 개체는 구조상으로 불안정하 다 (Greenh ill 1881, Hom 1971, McMahon 1973, Gi vn is h 1982). 외부로부 터 받는 힘에 의해 줄기는 한쪽으로 굽어지게 되고, 그 결과 무게 중심이 이동하여 줄기는 더 굽는다. 이렇게 줄기가 굽으면 줄기가 뒤틀리게 된다. 또한 줄기는 압축하는 힘 때문에 자신의 무게를 지 탱하지 못하게 되면 넘어질 수 있다 . 압축하는 힘에 의해 넘어지거 나 뒤틀리지 않으려면 줄기의 직경 (D) 이 자신의 높이 (H) 의 3/2 승으 로 증가되 어 야 한다 (McMahon 1973). 줄기가 원통형이면 그것의 부피 또는 무게는 HD2 에 비례할 것이 다. 줄기가 안정되기 위해서는 D 가 J-f3 12 에 비례하여야 한다. 따라서 원통형 줄기의 무게는 H( J-f3 1 2 ) 라 혹은 H4 에 비례하여야 한다 . 이것은 줄기의 끝이 가늘지 않은 식물이 뒤틀리지 않고 자신의 키를 2 배로 하기 위해서는 줄기의 생물량을 16 배로 증가시켜야 함을 의미한다. 그러나 식물의 줄기는 그 끝이 가늘기 때문에 (Waller 1986), 줄기의 부피가 키의 4 승으로 신속히 증가하지는 않는다. 이 책에서는 이 관 계를 좀더 완화시켜 식물의 키가 줄기 무게의 제곱근에 비례하는 것으로 하였다. 키와 줄기의 무게 사이의 정확한 관계가 어떻든 간 에 키의 증가는 줄기 조직에 많은 양의 분배를 필요로 하고 있다. 키의 증가에 따른 에너지 비용을 고려한다면 성숙하였을 때의 최 대 키는 어떻게 토양의 영양염류 공급률과 손실률에 의존적이게 되
는가? 나는 이 생각을 입증하기 위해 ALLOCATE 모델을 사용하였 고. 두 가지 경우를 고려하였다. 두 경우 모두 성숙하였을 때의 키만 제외하고 모든 것이 동일한 20 종을 부족한 토양 영양염류와 광 조 건에서 경쟁하도록 하였다. 그 결과 성숙하였을 때의 키가 5-240cm 의 범위로 나타났다. 첫번째 경우. 식물의 사망률은 식물의 키에 무 관하게 0 . 2 로 일정하였는데, 모든 크기의 개체들이 똑같은 밀도 독 립적인 사망률을 보였다. 두번째 경우. 사망률이 밀도 의존적이었는 데, 모든 형태의 개체들의 사망률이 그들의 키에 따라 직선적으로 증가하였다 . 즉 키가 1cm 이하인 식물은 사망률이 0 이었는데 키가 240cm 인 식물은 사망률 D=0.2 로 증가하였다. 첫번째의 경우 식물이 척박한 토양에서 경쟁할 때에는 키가 별로 영향을 주지 않았다(그림 4.1 1 ). 즉 식생의 평균키와 각 종의 상대수 도는 초기값과 같았다. 그러나 비옥한 토양에서는 키가 큰 식물이 유리하였는데, 가장 비옥한 토양에서는 키가 가장 큰 3 가지 형태의 식물이 키가 작은 식물을 교체하였다 . 이것은 식물이 항상 영양염류 의 제한을 받는 척박한 토양에서는 성숙하였을 때의 식물의 키가 그 식물의 경쟁력에 별다른 영향력이 없지만 광이 제한 요인이 되 는 비옥한 토양에서는 성숙하였울 때 키가 큰 식물이 유리하다는 것을 의미한다. 비옥한 토양에서 키의 증가로 인한 선택압의 크기는 두번째 경우에서 볼 수 있다. 키 의존적인 사망률이 있는 두번째의 경우에서 키가 작은 식물은 척박한 토양에서 유리하지만 비옥한 토 양에서는 키가 큰 식물에 의해 경쟁적으로 교체되고 있다(그림 4.1 1). 성숙하였을 때 식물의 최적 키는 영양염류. 광합성의 광 의존성, 식물의 형태, 식물의 상대 생장에 영향을 주는 구조적인 제약 둥 여 러 가지 요인에 의해 결정된다. 그림 4.11 에 나와 있는 결과는 성숙 하였을 때의 식물의 키가 영양염류 공급률에 따라 증가하여야 함을
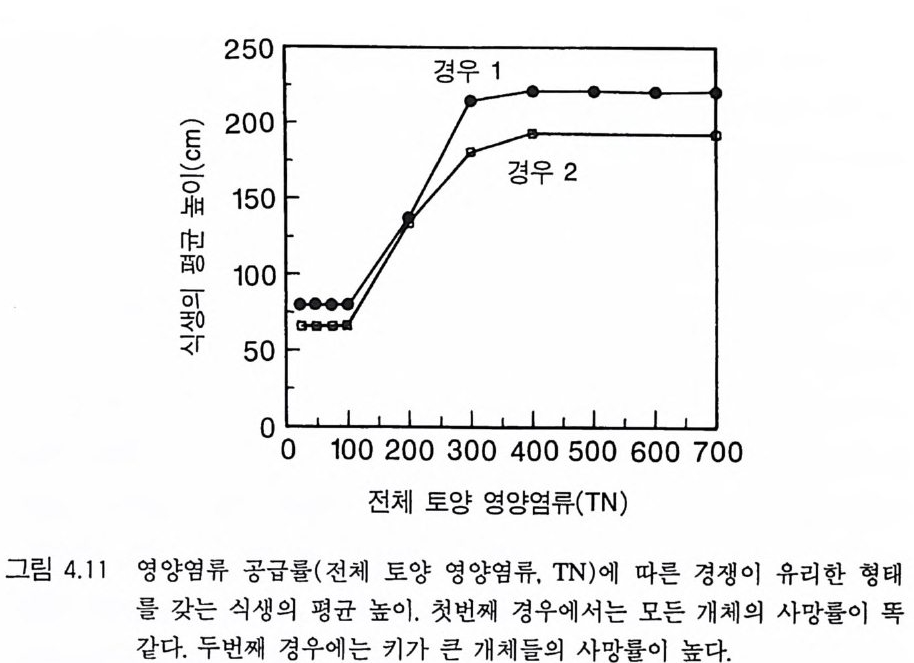 250200
250200
보여주고 있다. 식물이 성숙하였을 때의 최적 키는 그것과 경쟁을 하고 있는 다른 식물의 특성에 의해서도 영향을 받는다 (G ivni sh 1982). 6 종자의 크기 종자의 크기는 토양 자원:광 구배에 의해 영향을 받는 또 하나의 성질이다. 자원을 종자 생산에 분배할 때에도 식물은 대응책을 갖는 다. 식물이 종자를 많이 생산할수록 개개의 종자는 크기가 작아야 한다 (Werner & Platt 1976, Ha rpe r 1977). 종자의 크기가 생존, 생장 혹 은 생식에 영향을 주지 않을 경우 자연 선택은 많은 수의 작은 종 자를 생산하는 개체에게 유리할 것이다. 큰 종자는 실생 상태의 식
물 생장을 빠르게 하고 , 식물의 키의 증가는 수광량의 지수함수적인 증가와 관련이 있기 때문에 실생이 햇빛의 제약을 받는 환경에서는 종자가 큰 것이 유리할 것으로 보인다. 그러나 큰 종자는 다른 비용 이 수반된다. 일반적으로 종자는 영양분이 많아 동물이 잘 먹는다. 또한 큰 종자는 동물의 눈에 쉽게 띄기 때문에 먹힐 확률이 크다. 종자의 크기가 커서 생기는 유리한 점이 종자포식으로 인한 손실을 능가할 수 있을까? 나는 영양염류와 광에 대한 두 가지 서로 다른 경쟁의 경우를 조 사하기 위하여 ALLOCATE 모델을 이용하였다. 두 경우 모두 경쟁 관계에 있는 20 종의 서로 다른 식물들은 그들의 종자 크기를 제외 하고는 모든 성질이 동일하였다. 사용된 종자의 크기는 0.0 0 1- 0.090 g/종자의 범위였다. 첫번째 경우, 종자의 사망률을 0 으로 하였 고, 두번째 경우는 종자의 사망률이 종자의 크기와 비례하도록 하였 다. 즉, 가장 큰 종자는 발아하기 전에 40% 가 죽는 것으로 하였다. 종자는 생산된 직후에 발아가 되었다 . 종자의 사망률이 0 일 때(경우 1), 종자의 크기는 척박한 토양에서 는 식물의 경쟁력에 별 영향이 없었다. 형태가 서로 다른 20 종의 식 물의 상대수도는 시간이 지나도 변하지 않았다. TN=140 일 때까지 종자의 크기는 식물의 경쟁력에 별 영향이 없었다(그림 4.1 2 ). 이후 부터 큰 종자를 생산하는 개체가 경쟁에 유리하게 되었는데, TN= 160 에서 가장 큰 종자를 생산하는 종이 다른 종들을 교체하였 다 . 이러한 교체 현상은 새로 발아하는 종자가 광에 의해 제약을 받 는 영양염류 공급률에서 나타나기 시작하였다. 실생이 광의 제약을 받자마자 실생의 키가 큰 상태로 발아하는 큰 종자를 생산하는 개체들은 매우 유리하게 되었다 . TN=160 에서 모든 실생은 광의 제한울 받았는데, 큰 종자에서 생긴 키가 큰 개체 들이 다른 개체들을 교체하였다. 가장 작은 종자가 척박한 서식처에
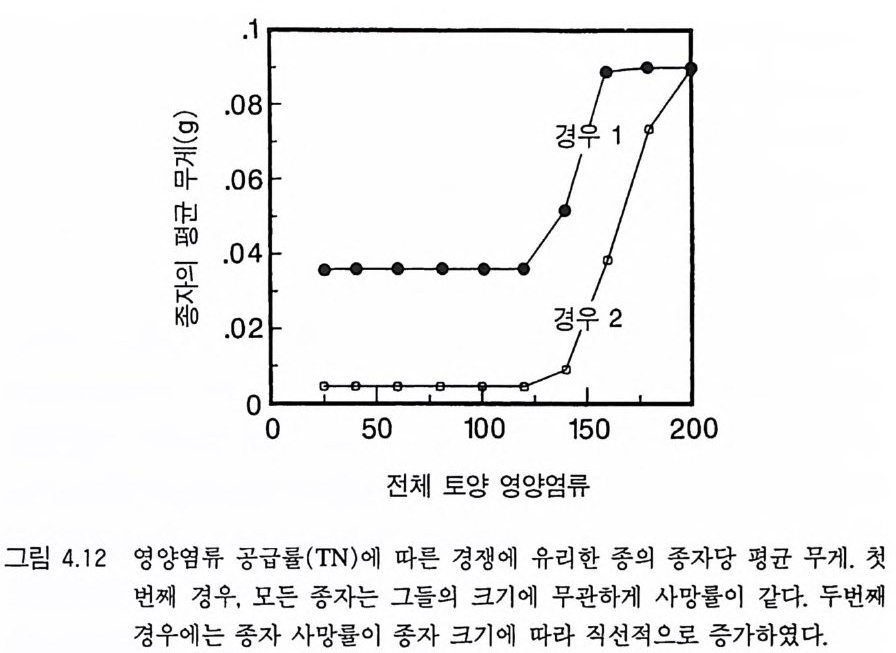 .1
.1
서 유리한 것을 제외하고는 종자의 크기에 따라 사망률을 증가시켰 을 때에도 비슷한 양상이 나타났다(경우 2, 그림 4.1 2 ). 가장 큰 종자를 생산하는 개체의 종자 사망률은 작은 종자를 생 산하는 개체의 90 배이고, 이 식물들은 종자가 크기 때문에 작은 종 자를 생산하는 개체에 비해 그 수가 1/ 90 에 불과하지만 TN=200 에 서 큰 종자를 생산하는 식물이 다른 식물들을 교체하였다. 따라서 종자의 크기는 토양 자원:광 구배에 따라 증가하여야 한다. 종자 크 기에 따른 포식의 차이가 없는 경우, 식물은 영양염류와 광에 의해 똑같이 제한을 받도록 큰 실생을 형성할 수 있는 크기의 종자를 갖 도록 진화할 것이다. 종자 크기에 따른 포식의 차이가 있을 경우에 는 일반적으로 크기가 작은 종자가 유리하지만 , 종자의 크기는 토양 영양염류 공급률에 따라 증가할 것으로 예상된다 . 크기가 작은 종자는 식물로 하여금 새롭게 교란된 지역에 신속히
정착할 수 있도록 하는 능력을 갖게 하는 것으로 설명되고 있다 (Werner&Platt 1 976), 교란은 식생을 제거하여 지표면에 광량을 증 가시키기 때문에 교란이 잦은 서식처에 살고 있는 식물 중 작은 종 자를 생산하는 식물이 갖는 이로운 점은 단지 이러한 종자가 그런 서식처에 이입될 수 있는 기회가 증가한다는 것만은 아니며, 광 이 용도가 크기 때문에 큰 종자가 불리하다는 것도 아니다. 이러한 모의는 또한 소수의 큰 종자를 생산하는 개체가 다수의 작은 종자를 생산하는 개체만큼 성공적일 수 있음을 보여주고 있다. 그 이유는 작은 종자로부터 시작되는 식물은 적어도 이 모델의 범 주 내에서는 큰 종자로부터 시작되는 개체에 비해 영양 기관의 생 장 기간 내내 크기가 작기 때문이다. 종자가 큰 식물은 필연적으로 적은 수의 종자를 생산하지만 이들의 실생은 작은 종자로부터 출발 하는 실생에 비해 신속하게 성장하여 성숙한 크기에 이른다 . 크기 구조를 가지고 있는 개체 사이의 이 경쟁 모델에서는 종자의 크기 가 종자 사망률이나 정착률과 관련이 없는한 영양염류가 부족한 서 식처에서 종자의 크기는 식물의 경쟁력에 별 영향이 없다. 그러나 건조한 서식처에서의 수분 이용도처럼 자원의 이용도가 토양의 깊이에 따라 증가하는 서식처에서는 큰 종자가 경쟁에 유리 할 수 있댜 이러한 조건하에서는 큰 종자를 생산하는 개체들은 근 계가 커 토양 자원을 더 많이 이용할 수 있다 (Baker 1972). 이 경우 토양 자원은 광의 수직 구배와 같이 깊이에 따른 구배를 갖는 것으 로 간주되며, 광량이 적은 서식처에서는 큰 종자가 경쟁에 유리할 것이다.
7 처음 생식시의 연령과 다년성 생식이 처음 시작되는 연령은 개체의 적합도에 큰 영향을 준다 (Cole 1954). 생식이 처음 시작되는 연령이 약간 증가하면 개체의 생 식률이 크게 저하될 수 있다. 다른 모든 조건이 동일할 때 생식을 일찍 시작하는 개체는 생식률이 크기 때문에 자연 선택에 유리하다. 그러나 모든 조건이 동일하기는 매우 어렵다. ALLOCATE 모델의 범주 내에서 생식이 처음 시작되는 연령은 척박한 토양에서는 자연 선택에 별 영향이 없다. 그 이유는 식물이 두 가지 상이한 생식 방 법, 즉 영양 기관을 통한 생식과 종자를 통한 생식을 할 수 있기 때 문이다 . 종자를 일찍 생산하는 식물은 영양 기관의 생장을 위한 에 너지를 그 해의 종자 생산을 위해 사용하는 것이다 . 실생과 성체의 사망률이 같고 특정한 서식처에서 단위 생물량당 생장률이 같다면 종자의 생산이나 영양 기관의 생장이 기능상 동일하고 경쟁력에 별 영향이 없을 것이다. 그러나 실생과 성체의 사망률이 같고 이들의 상대 생장률이 같은 서식처는 거의 없을 것이다. 초식이 많고 교란이 빈번하며, 혹독한 환경 조건은 종자에 비해 성체를 더 많이 죽게 할 것이다. 그 이유 는, 종자가 작고 휴면 상태에 있기 때문이다. 만약 이러한 손실이 사 망률의 요인이 되면 성체가 종자에 비해 높은 손실률을 갖게 되며, 손실률의 증가는 다년생보다는 1 년생 혹은 수명이 짧은 식물에 유 리할 것이다 (Chamov & Scha ffer 1973). 손실률이 고정되어 있는 상태에서는 부족한 토양 자원의 공급률 이 생식이 처음 시작되는 연령에도 영향을 줄 수 있다. 이를 알아보 기 위하여 같은 종내의 1 년생 생장형과 다년생 생장형이 서로 어떻 게 경쟁하는지를 생각해 보자. 1 년생 식물은 생육기가 끝나가면 가 용성인 모든 화합물을 종자 생산에 재분배한다 (Moone y 1972). 그러
나 다년생 식물은 그 해 생장중의 일부를 다년생 조직과 저장에 분 배해야 하기 때문에 생장 첫해 동안에는 종자 생산량이 적다. 다음 생육기가 시작될 때 1 년생 식물과 다년생 식물로부터 생산된 종자 는 실생으로 자라기 시작하지만 이미 존재하고 있는 다년생 식물은 기존의 근계와 분열 조직. 그리고 저장된 에너지와 영양염류를 가지 고 생장을 시작할 것이다. 따라서 다년생 식물은 종자로부터 자라는 식물에 비해 지상부 생물량의 초기 생장이 더 빠르다. 저장된 물질 이 많을수록 초기의 지상부 생장은 더 빠르다. 서식처가 광이 쉽게 제한 요인이 되는 비옥한 토양이면 초기에 식물의 키가 약간 차이 가 생겨도 수광량에 큰 차이가 나기 때문에 기존의 다년생 식물은 크게 유리하게 된다 (Black 1958, 1960, Harpe r 1977, Newberry & Newman 1978 ) . 종자로부터 시작하는 1 년생 식물은 토양이 비옥한 지역에서는 다 년생 식물에 의해 경쟁적으로 교체될 것이다. 왜냐하면 생육계절 초 기에 다년생 식물이 갖는 큰 키로 인한 이점이 그 다음해에도 계속 되기 때문이다. 계절이 있는 환경에서 식물의 다년생 부분은 크기가 큰 종자와 같은 기능이 있다. 큰 종자가 영양염류가 풍부한 서식처 에서 유리한 것과 마찬가지로 다년성도 그런 서식처에서 유리하다. 자원이 풍부한 서식처에서 다년성에 유리한 또 하나의 힘은 솜음 질이다 (H arp er 1977). Wes t ab y (1984) 가 설명한 바와 같이 키가 큰 식 물에 비해 키가 작은 식물이 죽을 확률이 더 높다 . 따라서 자원이 풍부한 토양에서 1 년생 생장형은 생육 초기의 길이 생장이 느려 다 년생 생장형보다 사망률이 높다. 따라서 이러한 토양에서는 다년성 이 매우 유리하다. 영양염류가 풍부한 서식처에서 모든 것이 동일하면 다년성이 유 리하지만 연령 의존적인 사망률에 차이가 없다면 1 년생 생장형과 다년생 생장형은 자원이 빈약한 토양에서는 별 차이가 없다. 연령
의존적인 사망률의 원인이 될 수 있는 것으로 계절성을 들 수 있다. 계절성은 어느 면에서 보면 종자나 다년생 뿌리. 줄기와 같은 휴면 기관에 유리한 일종의 교란이라고 할 수 있다. 계절성이 매우 극단 적이면 다년생 기관보다는 종자가 유리하며, 자원이 부족한 서식처 에서는 1 년생 식물이 유리하다. 전반적으로, 이것은 영양염류가 빈 약하고 계절성이 있는 서식처에서 1 년생 식물의 비율이 가장 높고. 교란이 없고 영양염류가 풍부한 토양에서 가장 낮음을 의미한다. 또 한 초식. 교란 혹은 다른 손실원이 종자보다는 성체의 사망률을 높 게 한다면 1 년생 식물의 수도가 손실률에 따라 증가한다는 것을 암 시하고 있다. 그러나 다년성에게 유리할 수 있는 특성도 있다. 이러한 특성에는 (1) 계절성이 큰 서식처의 다년생 식물은 자원을 저장할 수 있어 (Mooney 1972), 1 년생 식물이 자원 이용도의 변이에 따라 달라지는 데 비해 다년생 식물은 자원 이용도의 평균치에 반응할 수 있다는 가능성 (Til m an 1982, 그림 89-90), 그리고 (2) 기존의 다년생 식물은 생육계절의 초기에 나타나는 토양 자원의 변화를 더 잘 이용할 수 있는 가능성 등이 포함되어 있다. 여기에서 말로 설명한 내용은 서로 다른 크기의 종자를 생산하는 식물에서 수학적으로 설명한 내용과 성질상 유사하다. 종자 크기에 의존적인 사망률이 없고 안정한 서식처의 경우 실생이 햇빛의 제한 을 받지 않을 정도의 척박한 토양에서는 종자의 크기가 그 식물의 경쟁력에 아무런 영향을 주지 않는다. 그러나 실생이 햇빛의 제한울 받을 정도로 충분히 비옥한 토양에서는 큰 종자를 생산하는 식물이 실생의 키가 크기 때문에 유리하다. 식물의 다년생 기관은 크기가 큰 종자와 유사하다. 생육계절 말에 다년생 기관에 저장된 영양염류와 에너지는 다음 생육계절 초에 식 물의 생장을 빠르게 한다. 햇빛이 신속하게 제한되는 비옥한 서식처
에서는 이러한 빠른 생장이 가장 중요하다. Rau nki aer(1934) 에 의해 언급된 것처럼 다년생 식물의 또 하나의 특징인 눈의 위치도 중요하다(그림 4.1 3 ). Raun ki aer 에 의해 지중식 물로 분류된 1 년생 식물과 초본성 다년생 식물은 지표면이나 그 바 로 밑에 다년생 기관을 가지고 있다. 지표식물로 분류된 관목은 지 상부에 있는 목질 줄기에 눈이 있다. 지상식물로 분류된 교목은 지 표면에서 훨씬 위쪽에 눈이 있다. 식물의 눈이 지표면으로부터 멀리 있을수록 다음 생육기간 초기에 다른 식물에 의해 그늘이 질 확률 이 낮다. 이것은 계절성이 있는 서식처에서 지표면 위에 눈을 가지 고 있는 식물이 비옥한 토양에서 유리하다는 것을 의미한다. 그러나 목질 구조는 몇 가지 손실도 수반한다. 즉, 겨울에 초식동 물에게 먹히고, 목질 조직을 만들고 유지하는 데 광합성 산물과 영 양염류가 필요하다. 이러한 손실은 이들이 척박한 토양에서는 유리 하지 않음을 의미한다. 따라서 척박한 토양에서 비옥한 토양 구배를 따라 지중식물에서 지표식물 그리고 지상식물로, 키가 작은 식물에
 A. 지중식물 B 반지중식물 C. 지표식물 D. 지상식물
A. 지중식물 B 반지중식물 C. 지표식물 D. 지상식물
서 키가 큰 식물로, 눈이 낮은 곳에 위치한 식물에서 높은 곳에 위 치한 식물로 이어지는 단계적인 변화가 있다. 8 요약 이 장에서 제시된 이론은 광합성 산물을 뿌리, 잎, 그리고 줄기로 분배하는 패턴이 식물의 최대 생장률과 경쟁 능력에 크게 영향을 줄 수 있다는 것을 보여주고 있다. 환경의 제한이 없는 곳에서 사는 식물은 광합성 산물 모두를 여분의 광합성 조직의 생산에 분배하는 개체에서 영양 생장의 최대 생장률을 볼 수 있다. 최대 생장률은 비 광합성 기관에 분배되는 광합성 산물의 양에 비례하여 감소한다. 생 산량의 일부를 뿌리와 줄기에 분배한 식물은 환경이 제한되지 않는 서식처에서 생장률이 저하되지만 자원이 부족한 환경에서는 유리하 댜 영양염류의 공급률과 손실률이 고정된 특정한 서식처에 사는 식 물의 경우, 생리나 구조적인 제약이 동일한 개체들 사이에서 경쟁 능력이 우세하도록 해주는 특정한 형태가 있다. 그러나 각 서식처에 따라 뿌리, 잎, 줄기의 분배 패턴이 다른 식물, 죽 상이한 형태의 식 물이 우세하다. 서식처의 손실률과 토양 자원:광 구배상의 위치는 모두 경쟁에 유리한 형태를 결정하는 중요한 인자들이다. 손실률을 일정하게 하 면, 척박한 토양에서 우세한 경쟁자는 생산량의 대부분을 뿌리와 잎 에 분배하고 줄기에는 분배량이 적다. 비옥한 토양에서는 잎에 분배 되는 양이 많고 뿌리에 분배하는 양이 적은 개체가 경쟁에 유리하 다 . 이보다 더 비옥한 토양에서는 뿌리로 분배되는 양을 줄이고 줄 기로 분배되는 양을 늘리면 유리하다. 더욱 비옥한 토양에서는 뿌리 와 잎으로 분배되는 양을 줄이고 줄기로 분배량을 늘이는 개체가
유리하댜 따라서 척박하지만 햇빛이 풍부한 서식처로부터 비옥하지 만 햇빛이 부족한 서식처로 이어지는 구배상에서 줄기로의 분배량 은 증가하고 뿌리로의 분배량은 감소하며, 잎으로의 분배량은 영양 염류의 공급률이 중간 정도에서 최대가 될 것으로 예상된다. 이러한 분배 패턴은 매우 척박하거나 비옥한 토양에 우점하는 식 물보다 중간 정도의 비옥도를 갖는 토양에서 우점하는 식물이 다소 높은 최대 생장률을 갖도록 만든다. 또한 광량은 많지만 토양이 척 박한 서식처로부터 광량은 적지만 비옥한 서식처에 이르는 구배를 따라 종자의 크기, 성숙하였을 때의 키, 생식이 처음 시작되는 연령 은 모두 증가한다. 토양 자원 : 광 구배상의 각 지점에서 우점하는 식 물의 일반적인 특징이 그림 4.14 에 종합되어 있다. 식물의 생리적 특성도 생산력 구배를 따라 달라져야 한다. 예를 들면, 척박한 토양에서는 광합성의 광포화율이 감소될지라도 조직의 영양염류 농도가 낮은 식물이 유리하다. 비옥한 토양에서는 광합성 의 광포화율이 높고 광보상점이 낮은 개체가 유리할 것이다. 형태 적, 생리적, 그리고 생활사적인 특징 모두가 상호 작용을 통해 식물 의 경쟁력을 결정한다. 각 서식처에서 특정한 식물이 우세한 경쟁자가 되도록 해주는 이 러한 성질들의 특정한 조합이 있다. 그러나 서식처에 따라 영양염류 공급률이나 손실률이 서로 다르기 때문에 각 서식처에 알맞는 조합 도 다르다. 나는 식물의 생리가 마치 고정된 것처럼 식물 형태의 진 화를 설명하였지만 식물의 형태, 생리, 그리고 생활사는 상호 연관 되어 있고 상호 의존적이다. 또한 3 가지 모두가 생산력과 손실률 구 배를 따라 변화한다. 손실률(초식이나 교란 둥에 의한)도 특정한 토양에서 경쟁력이 우 세한 식물의 성질에 영향을 준다. 비옥한 토양에서는 손실률이 낮은 곳에서 우점하는 식물이 손실률이 높은 곳에서 우점하는 식물에 비
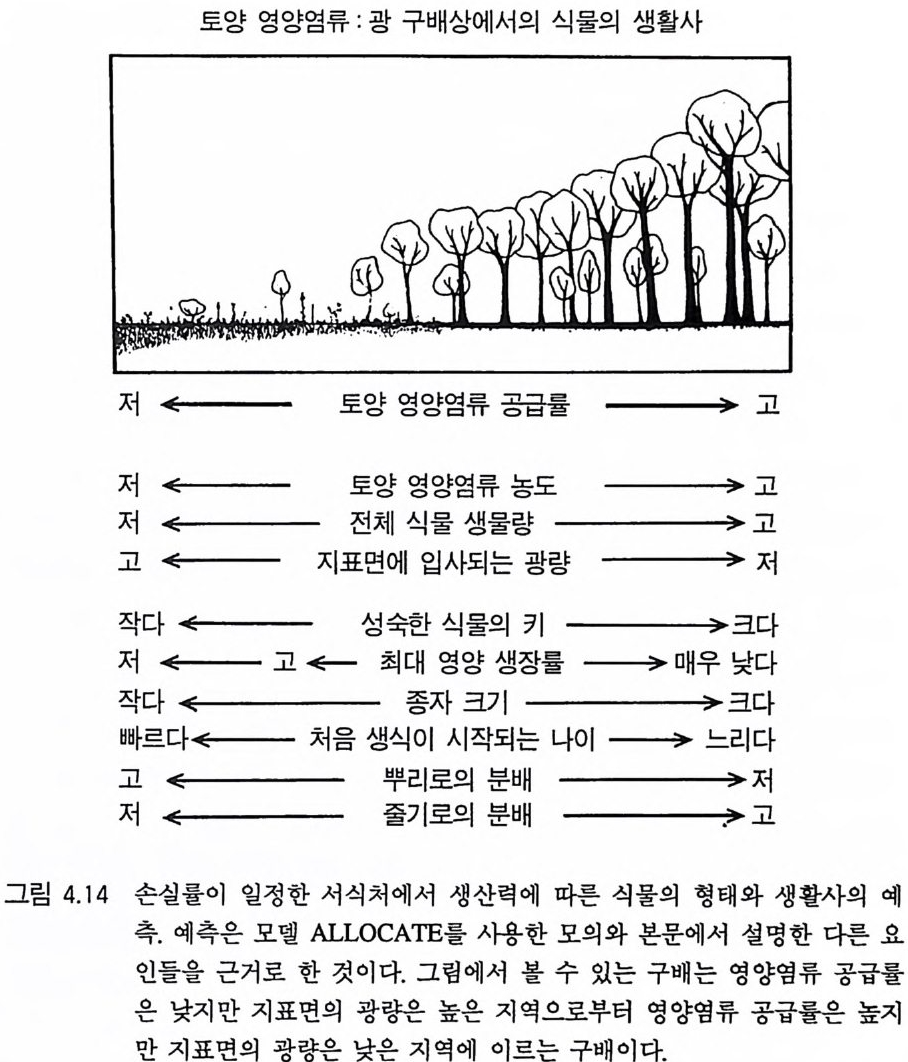 토양 영양염류 : 광 구배상에서의 식물의 생활사
토양 영양염류 : 광 구배상에서의 식물의 생활사
해 키가 크고, 영양 기관의 최대 생장률이 낮고. 수명이 길며, 생활 사의 후반부에 종자를 생산한다. 또한 줄기로의 분배 량이 많고 잎으 로의 분배 량이 적다(그림 4.1 5 ). 이 구배상에서 뿌리로의 분배 량은 상당히 일정하다. 척박한 토양의 손실률 구배상에서 식물의 키는 손실률에 크게 의
 비옥한 토양에서 손실률 구배에 따른 생활사
비옥한 토양에서 손실률 구배에 따른 생활사
존적이지는 않지만 손실률이 낮은 곳에서 우점하는 식물은 높은 곳 에서 우점하는 식물에 비해 뿌리로의 분배량이 많고 잎으로의 분배 량이 적다. 척박한 토양이나 비옥한 토양의 손실률이 높은 서식처에 서 잎으로의 분배량이 높은 것은 손실률이 높은 곳에서 우점하는 식물이 높은 최대 생장률 (RGRmax) 을 갖는다는 것을 의미한다. 높은 손실률은 전체 식물량을 감소시켜 영양염류와 광의 이용도를 증가
시키기 때문에 최대 생장률이 나타나게 된다. 제한되는 자원의 수준 이 높으면 생장률이 높은 개체들이 유리하다. 손실률 구배상에서 유리한 식물의 특징과 생산력 구배상에서 유 리한 특징 사이에 차이가 있는 것은 이들 구배상에서 자원 이용도 의 패턴이 다르기 때문이다. 손실률을 일정하게 유지하면 부족한 토 양 자원의 농도는 생산력 구배상에서 지표면에 이입되는 광량과 음 의 상관이 있다. 반대로, 영양염류의 이용도와 지표면에 이입되는 광량은 모두 손실률 구배를 따라(낮은 손실률에서 높은 손실률) 증가 한댜 죽 손실률과 양의 상관이 있다. 식물의 형태는 그 식물이 우세한 경쟁자가 되는 지소의 토양 자 원의 수준과 지표면의 광량(혹은 다른 높이에서의 광량)을 결정하기 때문에 이러한 상이한 자원 구배는 종 교체의 양상을 변화시킨다. 생산력 구배나 손실률 구배는 영양염류 공급률과 손실률로 형성되 는 평면상의 가능한 수많은 괘적 중 두 가지에 불과하다(그림 4.1 0 ). 각 괘적에 따라 식물의 형태와 생활사가 달라진다. 부족한 영양염류 의 공급률과 손실률은 모두 특정한 서식처에서 경쟁력이 우세하도 록 유도하는 식물의 형태에 영향을 준다. 따라서 이 두 가지는 서식 처의 식생 구성에 영향을 주는 중요한 요소이며, 서로 불가분의 관 계에 있다.
제 5 장 생산력 구배와 손실률 구배상에서의 식생 패턴 l 서론 제 4 장에서 설명한 바와 같이 식물의 형태와 생활사의 여러 가지 특징은 서식처의 영양염류 공급률과 손실률에 따라 다를 것으로 예 상된댜 예를 들어 식물들은 그 종의 수도가 최대로 되는 점과 그 종의 형태, 생활사, 그리고 생리적인 특성이 일치하도록 주요 환경 구배상에서 분리될 것으로 예상된다. 손실률이 낮은 서식처에 우점 하는 식물은 높은 서식처에 우점하는 식물에 비해 최대 상대 생장 률이 낮을 것으로 예상된다. 종자의 크기와 줄기로의 분배는 생산력 구배를 따라 중가할 것으로 예상된다. 수관이 밀폐된 서식처는 개방된 서식처에 비해 부족한 토양 자원 의 공급률이 높고 키가 큰 식물들이 우점할 것으로 예상된다. 관찰 과 실험연구 자료를 통해 이러한 예측이 얼마나 잘 입증될 수 있는 가? 우선 서식처의 손실률과 영양염류 공급률에 대한 최대 생장률
의 의존도를 살펴보고, 다음에는 식물 군집 내에서 그리고 군집 사 이에서 볼 수 있는 몇 가지 광범위한 패턴을 알아보도록 하겠다. 2 최대 생장률 식물의 최대 영양 생장률은 여분의 광합성 조직에 분배되는 생산 량의 비에 따라 증가하여야 한다(제 3 장 . Monsi 1968). 따라서 비질소 고정 토양 조류는 최대 생장률이 가장 높아야 하며, 뿌리와 줄기로 가장 많은 양을 분배하는 식물은 최대 생장률이 가장 낮아야 한다. ALLOCATE 모델은 광합성 산물을 잎으로 분배하는 양의 비율을 예측하고, 따라서 최대 생장률은 서식처의 손실률과 영양염류 공급 률에 의존적 임을 예측하고 있다(그림 4 . 7 과 4.8 참조). 그림 5 .1에는 경쟁에서 우세한 · 형태를 갖는 식물의 최대 생장률이 서식처 손실 률과 서식처의 영양염류 공급률에 따라 다르다는 것을 보여주고 있다. 영양염류 공급률을 일정하게 유지하면 RGR,,,ax 는 손실률에 따라 증가하여야 한다. 손실률을 일정하게 유지하면 RGR,,,ax 는 손실률이 낮거나 중간인 서식처에서는 중간 정도의 생산력을 갖는 서식처에 서 최대가 되어야 하며, 손실률이 높은 서식처의 경우 생산력에 따 라 RGRmax 는 증가하여야 한댜 일반적으로 서식처의 영양염류 공급 률에 비해 서식처의 손실률은 우점종의 RGRmax 를 결정하는 데 더 중요한 요인인 것으로 간주된다. 생산력 구배상에서 생장이 가장 빠 른 식물과 가장 느린 식물 사이의 차이는 손실률 구배상(낮은 서식 처에서 높은 서식처로 이어지는)에서의 차이에 비해 작다. 이러한 예측은 제 4 장에서 사용된 ALLOCATE 의 형태에 기초를 둔 것이다. 이러한 모의에서 사용된 변수를 변형시키면, 생산력 구
 상대 생장률 등치선
상대 생장률 등치선
배상의 영양엽류 공급률이 중간 정도에서 RGR,n” 가 최대에 이르고 그 이외의 서식처에서는 RGR~ 가 더욱 신속하고 극적으로 감소하 게 된다. 그 이유는 줄기 무게에 대한 식물의 키의 의존도를 설명하 는 데 사용된 상대 생장 관계가 식물의 키 (H) 는 줄기의 생물량 (S) 과 H=c§.5 (5.1 ) 로 주어지기 때문이다. 즉, 키가 두 배로 되기 위해서는 줄기의 무게 가 4 배가 되어야 한다. 나는 이러한 관계를 몇 종의 벼과식물에서 알아냈기 때문에 이 관계를 선택하였다.
그러나 제 4 장에서 설명한 바와 같이 줄기가 안정되기 위해서는 그것 의 높이 가 줄기 무게 의 4 승근으로 증가하여 야 한다 (McMahon 1973). H=cSJ .2 5 (5.2) 즉 키가 두 배로 되기 위해서는 줄기의 무게가 16 배로 증가하여 야 한다. 이끼와 같이 키가 작은 식물에서부터 세코이아와 같이 키 가 큰 식물을 고려할 때. 큰 키를 위해서는 예상했던 것보다는 줄기 에 더 많은 분배를 하여야 한다. 그러나 줄기에 많은 분배를 하면 잎이나 뿌리로의 분배량이 감소되어야 한다. 이것은 영양염류가 매 우 풍부한 토양에 우점하는 종은 예상되는 최대 생장률이 매우 낮 아야 한다는 것을 의미하며, 그림 5 .1에 나타난 것보다 생산력 구배 상에서 최대 생장률이 더 신속히 감소한다는 것을 의미한다. G ri me(1979) 과 Cha pin (1980) 은 매우 척박한 서식처에 있는 식물 들의 최대 상대 생장률이 낮다는 실험적인 증거를 제시하였다. 또한 두 사람은 모두 비옥한 서식처에 있는 식물들의 최대 상대 생장률 이 높다고 주장하고 있다. 가장 비옥한 곳에 우점하는 종의 RGR,’'g 가 가장 크다는 G ri me 과 Cha pi n 의 주장이 맞다면 그들의 자료는 이 이론에 의한 주요 예측과 상반된다 . 더구나 이 책에서 제시하고 있 는 견해와 G ri me(1979) 이 주장하고 있는 견해 사이에는 많은 차이 가 있다. 나는 이 기회를 이용하여 깊이 있는 실험적인 증거를 찾아 내고 식물 진화와 군집 구조에 대한 서로 다른 접근 방법을 비교해 보려고 한다. G rim e (1 979) 은 영국 Sheff iel d 지방에서 132 종의 현화식물 실생의 최대 상대 생장률에 관한 연구와, 각 종이 발견되는 서식처의 비옥 도에 대한 주관적인 판단 기준으로부터 자료를 얻었다 (G ri me&Hun t 1975). 제 3 장에서 설명한 바와 같이 G ri me 과 Hun t (1975) 는 최대 상
대 생장률은 1 년생 식물이 가장 높고. 초본성 다년생 식물이 낮으 며, 목본성 다년생 식물이 가장 낮다는 것을 알았다(그림 3.2 참조). 그러나 이 132 종의 최대 상대 생장률과 각 종이 자연 상태에서 출 현하고 있는 서식처의 비옥도 사이에는 어떤 관계가 있는가? RGR 과 서식처 비옥도 사이의 관계를 결정하기 위해서는 이 식물 군의 자연 상태의 수도가 토양 비옥도에 얼마나 의존적인지를 알아 야 한다. 그러나 Tansle y (1949) 에 의해 여러 번 강조된 것처럼 이것 은 쉬운 문제가 아니다. 왜냐하면 영국의 식생이 벌목과 과방목에 의해 많이 변화되었기 때문이다. Tansley (l94 9, p. 129) 는 〈삼림에서 기원한 목야지에서는 계속적인 방목으로 인해 목본식물이 재정착할 수 없댜 그리고 현재 우리가 보는 식물 군집은 한편으로는 기후와 토양에 의해 조절되고 다른 한편으로는 방목에 의해 조절되고 있다〉 고 언급한 바 있다. 사람이 벌목하기 전에는 삼림이었지만 현재는 목야지로 된 지역에 삼림을 조성하기 위해서는 울타리를 만들어 초 식 동물을 막아야만 한다 (Farrow 1916, 1917, Summerhaye s 1941, Tansley 1949). 벌목이나 과방목 이전의 식생은 어떠하였을까? 가장 척박한 사토 에는 철쭉과의 관목에 의해 우점하는 황원이 형성되어 있었거나 (Tansley 1949), 아니면 사토에 형성되어 있는 삼림이나 관목림 내의 일부에 황원이 형성되어 있었을 것이다. Tansle y는 신석기시대나 현 대와 같은 방목이 있기 이전에는 목본식생이 없는 지역, 특히 척박 한 사토나 최근에 형성된 자갈로 된 하안 단구, 언덕 꼭대기, 토심이 얕은 지역에만 초지가 형성되어 있었다고 주장하고 있다 (Tansle y 1949, p. 164). 이에 비해 삼림은 토심이 깊고 비옥한 식토나 양토로 된 지 역 에 형 성 되 었다 (Tansle y 1949, p. 163, Godwin 1956). 따라서 벌목과 방목이 있기 이전에는 영국의 대부분의 비옥한 지 역이 교목에 의해 우점되어 있었다. G ri me 과 Hunt( 1 975. 본서의 그림
3. 2) 가 고려한 모든 식물군 중에서 목본성 다년생 식물의 RGR,,, a. ,가 가장 낮았다. 실제로 G ri me 과 Hun t가 연구한 132 종의 식물 중 가장 생장이 느린 6 개 중의 5 개는 교목이었다 (Acer p seudop la t anus, Pic e a abie s , P. nig r a, P. sit ch e ns is , P. syl v estr is ) . 매우 비옥한 토양의 우점 생 활형 인 교목은 모든 생 활형 중 RGR,, UL t가 가장 낮댜 Gr i me 과 Hunt 의 자료에 의하면 그 다음으로 RGRm a., 가 낮은 생활형은 황원에 혼한 목본성 다년생 식물이 다. 월귤나무인 Vacc ini u m v iti s- i daea 는 132 종 중에서 두번째로 최대 생장률이 낮았고. 다른 두 종의 황원식물 (Calluna vulga ris, Vacc ini u m m y r till us) 은 생장이 느린 10 종 중에 포함 되었다. 따라서 영양염류가 빈약한 서식처 중의 하나인 황원에 혼한 식물 과 가장 비옥한 서식처인 삼림에 우점하는 식물이 Gr i me 과 Hun t (1975) 가 조사한 모든 종 중에서 생장률이 가장 낮았다. 영양염 류가 빈약한 토양에서 최대 수도를 보이는 콩과식물도 RGRm ax 가 매 우 낮았다 (T i lman 1982). 벌목이나 방목 이전에 중간 정도의 비옥한 토양에 우점하던 협엽초본, 광엽초본, 목본성 관목은 최대 상대 생 장률이 훨씬 높았다. G ri me 과 Hun t가 수집한 자료를 벌목이나 방목 과 같은 인간에 의한 주요 교란이 있기 전의 서식처에 혼한 종과 연결시켜 설명하면 이 자료들은 제 4 장에서의 예측과 부합된다. 그러 나 비옥한 서식처에 우점하는 식물의 최대 생장률이 가장 높다는 G rirn e (1 979) 의 일반적 인 주장과는 완전히 상치된댜 G ri me 과 Hun t의 자료가 왜 그들이 내 린 결론과 상치 되 는가? 문제 는 자신들이 연구한 식물들의 현재의 분포를 너무 믿은 데서 비롯 되었다. 신석기 이전에는 숲이 형성되었던 비옥한 땅이 현재는 영국 에서 가장 비옥한 농경지 와 목야지로 되 었다 (Godw i n 1956). 목야지 는 그것이 부양할 수 있는 가장 많은 수의 양과 소들을 방목하기 때문에 비옥한 지역일수록 더 심하게 방목이 이루어진다. 따라서
Gr i me 과 Hun t가 보고한 현존식생의 구성과 토양 비옥도 사이의 관 계는 토양 비옥도와 초식동물의 밀도 사이의 상관 관계에 의해 혼 동되고 있댜 또한 완전벌목과 과방목에 의해 야기된 차이는 방목을 몇 년 금한다고 제거되는 것이 아니다. 목본식물은 생장이 느리고, 성숙시 그 크기가 엄청나며, 현재에는 이러한 지역에 드물게 나타난 댜 토양 영양염류와 식물의 특성 사이의 자연적인 상관 관계(교란 이전의 관계)가 다시 형성되기 위해서는 1 세기 혹은 그 이상이 아니 더라도 몇 십년의 기간이 필요하다. 또한 제 6 장에서 설명하겠지만 처음 몇 십년 동안 관찰되는 여러 가지 식물종의 동태는 장기간의 패턴과 반대로 나타날 수도 있다. 이러한 개체군의 일시적인 동태는 토양 비옥도와 식물 분포 사이의 관계를 설명하는 데 많은 어려움 올 준다 . 특히 비옥한 토양의 천이 계열 식생에서는 더욱 그렇다 (Til m an 1985, 이 책의 제 6 장). 가장 비옥한 지역인 경작지에 현재 가장 생장이 빠른 초본성 다 년생 식물과 1 년생 식물이 우점하고 있다는 Gr i me(l979) 의 주장은 맞다. G ri me 과 Hun t(1 975) 는 자신들의 분석에서 1 년생 식물을 제거 함으로써 비옥한 토양에서 방목의 영향을 고려하였지만 그들은 방 목 효과의 정도를 인식하지 못하였다. 현재는 양이 방목되지 않는 목야지도 개체군의 일시적인 동태 뿐 만 아니라 Tansle y (1949) 가 주장한 것처럼 초식동물의 포식자들이 줄어들자 토끼 등과 같은 작은 초식동물의 밀도가 증가되어 방목이 심했을 수도 있다. 이러한 경작 가능한 목야지는 벌목과 방목이 있 기 전에는 임지 였고 (Tansle y 1949, Godwi n 1956), 초식동물을 배제시 키면 삼림이 재형성되기 때문에 이 비옥한 토양에 생장이 빠른 종 이 존재하는 것은 과방목에 의한 손실, 초식동물에 의한 교란, 그리 고 토양 비옥도의 복합 효과에 기인하는 것으로 보아야 한다. 영국 의 원식생에 관한 Tansle y와 Godw i n 의 기재에서도 분명히 밝혀진
바와 같이 현재의 식물 분포를 토양의 높은 비옥도 하나에만 기인 하는 것으로 돌릴 수는 없다 . 나는 Gr i me 과 Hunt( l97 5), 그리고 Gr i me (l 979) 이 방목과 교란으 로 인한 손실률이 낮은 척박한 토양으로부터 손실률이 높은 바옥한 토양에 이르는 구배를 연구한 것으로 해석한다. 손실률이 가장 낮은 지역에는 RGR111 a.,가 가장 낮은 식물이 우점하고. 반면에 손실률이 가 장 높은 지역에는 RGR111 a. , 가 가장 높은 종이 우점하였다. 이런 측면 에서 볼 때 . 그들이 내린 결론은 제 4 장의 예측과 완전히 일치한다. 영양염류가 빈약하며 손실률이 낮은 서식처로부터 영양염류가 풍부 하고 손실률이 높은 서식처로의 구배상에서 그림 5.1 에서 보는 바와 같이 RGRm a:c는 거의 직선적으로 증가할 것으로 예상된다. 방목된 목야지의 식생과 벌목과 방목이 있기 전의 식생 사이의 뚜렷한 대비는 이러한 군집의 구조와 유지에 대한 초식. 다른 손실 원 혹은 교란의 역할을 분명히 보여주는 것이다. 비옥한 토양에서 자연 상태의 초식동물 밀도를 갖는 지역은 수관이 밀폐된 삼림으로 되어 있다. 그러나 가축에 의한 손실률이 높은 지역은 키가 작고 빨 리 자라는 종에 의해 우점되는 목야지 식생이 형성된다 . 따라서 자 연 상태의 목본식생(비록 느리지만 방목을 멈추면 원상으로 복귀하는) 과 목야지는 손실률 구배의 서로 반대편 끝에 위치한다. 이 구배상 에서의 식생의 구조, 식물의 생활사, 그리고 최대 생장률은 그림 4 .1 5 에서 예측한 것과 일치 한다. 그러나 방목된 지역과 방목되지 않은 지역 간의 식생 대비를 통 해 우리가 알아야 할 중요한 점이 있다. 이러한 지역을 목야지로 유 지하고 있는 초식동물은 포식자, 기생충, 질병 등으로부터 보호되기 때문에 그 밀도가 대단히 높다. Tansle y (1949) 에 의하면 이 지역에 서 포식자를 제거하기 이전에는 포식자들이 야생의 초식동물을 훨 씬 낮은 밀도로 유지시켰다. 그러한 자연밀도에서는 초식동물이 비
옥한 지역에 형성되어 유지되는 삼림을 방해할 수 없었다 . 따라서 본질적으로 문제를 혼동시키는 것은 초식동물의 존재가 아니라 포 식자의 조절 혹은 포식자의 감소로 인하여 초식동물의 밀도가 대 단히 높다는 데 있다. 이것은 모든 자연 군집의 구조가 다영양 단 계 (mul ti-t ro ph i c-level) 의 상호 작용에 의존적 임을 시사하고 있는 것 이댜 다영양 단계 상호 작용에 대하여 알아야 할 것이 많지만 유감스 럽게도 전세계적으로 자연 서식처의 파괴와 포식자의 계속적인 감 소로 이 분야를 연구할 시간이 많지 않다. 각 종은 전체 먹이그물의 범위 내에서 생활한다. 초식동물의 밀도는 식물에게 큰 영향을 주지 않을 정도로 항상 포식자나 기생자에 의해 억제되고 있다고 믿을 만한 근거가 없으며, 생산력이 서로 다른 군집에서 초식과 포식의 상대적인 역할에 대하여 확고한 결론을 내릴 만한 뚜렷한 경험적, 실험적, 혹은 이론적인 근거도 없다. Ha irs to n 둥 (1960) 과 Oksanen 등 (1981) 이 각각 이 문제에 대한 의견을 발표한 바 있다. Hair s to n 등(1 960) 은 많은 식 물이 그들이 필요로 하는 자원에 의 해 제한을 받을 수 있고, 많은 초식동물들이 그들의 포식자와 기생 자에 의해 제한을 받을 수 있다고 주장하였다. 그들은 경쟁이 식물 군집의 구조를 결정하는 주요 생물적 상호 작용이며, 포식은 초식동 물 군집의 구조를 결정하는 주요 상호 작용이라고 주장하였다. 나는 생물이 그들의 자원에 의해 또는 그들을 포식하는 생물에 의해 제 한을 받는다는 그들의 단순한 주장에 동의하지 않는다 . 그 이유는 제 2 장에서 설명한 바와 같이 자원 의존적인 생장과 사망률 혹은 손 실은 항상 개체군의 크기를 조절하기 위해 상호 작용을 해야 하기 때문이댜 그러나 그들의 주장을 바꾸어 말하면 〈포식자는 초식동물 의 밀도를 포식자가 없을 때보다 훨씬 낮게 유지한다〉라고 할 수 있다. 초식동물의 밀도가 낮으면 식물에게는 손실률과 교란율이 낮
고, 따라서 자원의 이용도가 낮은 상태에서 생장이 효율적인 식물종 에게 유리하댜 이런 측면에서 설명하면, Tansle y (1949) 에 의해 언급 된 많은 예들이 그들의 주장을 지지하는 것처럼 보이며. 특히 포식 자가 절멸한 후에 형성된 영국 식생의 패턴과 비교할 때 더욱 그 렇다. 이러한 생각은 Oksanen 등 (1981) 에 의해 다시 연구되어 발전되었 다. 그들은 먹이그물의 구조가 1 차 생산력에 따라 변할 수 있다고 주장하였댜 자원이 매우 빈약한 토양을 갖는 서식처에서 초식동물 은 그들이 이용할 수 있는 먹이의 양이 적고 질이 낮기 때문에 밀 도가 매우 낮은 수준에 머물 것이라고 주장하였다 . 이보다 생산력이 높은 서식처에는 식물의 생물량이 많기 때문에 이에 비례하여 높은 밀도의 초식동물을 부양할 수 있을 것이다. 그러나 포식자의 수가 적게 유지될 정도로 초식동물의 밀도가 낮게 유지될 수 있다. 가장 비옥한 서식처에서는 초식동물의 밀도가 높아 포식자의 밀도를 높 게 유지할 수 있으며, 식물에 대한 초식동물의 효과는 낮을 것이다. 이러한 다영양 단계의 상호 작용에 관한 내용 중의 일부가 9 장에 설명되어 있댜 이러한 패턴은 한 생태계에 있는 전체 영양 관계를 무시하였을 때 나타날 수 있는 잠재적인 위험을 보여주는 것이다. 토양 비옥도 와 생장률 사이의 관계를 재분석함으로써 우리가 알게 되는 것은 토양 비옥도와 초식 사이의 상관 관계를 통해 토양이 식물 군집의 구조에 간접적으로 영향을 줄 뿐만 아니라 직접적으로 영향을 줄 수 있다는 점이다. 자연 서식처에서의 상관 관계의 연구는 눈에 보 이는 어떤 패턴을 야기시키는 이유가 무엇인지를 명확하게 결정할 수 없다. 자연계에서 우리가 관찰할 수 있는 어떤 패턴을 야기시키 는 원인이 무엇인지를 밝혀내기 위해서는 영양염류, 초식동물 그리 고 포식자의 밀도를 다르게 하는 실험적인 접근이 필요하다.
모든 생물은 먹이그물 속에 묻혀서 살고 있다. 모든 종은 소비자 이면서 동시에 포식자, 기생자. 초식동물, 분해자의 자원이 된다. 이 책에서 설명하는 기계론적안 접근은 생물이 그들의 자원과 상호 작 용하는 방법을 자세히 알 수 있도록 하며. 다른 영양적 상호 작용을 포함할 수 있도록 쉽게 확장시킬 수 있다. 이러한 방법을 통해 군집 을 구성하는 힘을 이해할 수 있다. 3 식물상관 전세계의 다양한 식생에서 관찰할 수 있는 가장 일반적인 것 중 의 하나는 어느 식물종이 성숙하였을 때의 평균키는 그 종의 상대 수도가 최대가 되는 서식처의 비옥도와 일치한다는 점이다 (Beard 1944, 1955, 1983, Whit tak er 1975, Mooney 1977). 이 러 한 패 턴은 비옥도 구배상에서 각 종 내에서 볼 수 있는 키의 변이와 유사하여 하찮은 것으로 여겨왔기 때문에 그 중요성이 간과되어 왔다. 그러나 영양염 류를 첨가한 후에 식물체의 키가 자라는 것과 비옥한 토양에서 성 숙하였을 때의 키가 큰 종이 키가 작은 종을 교체하는 것 사이에는 차이가 있다. 전자는 형태적인 필요성으로 간단히 해결할 수 있지만 후자의 경우에는 생태학적인 설명이 필요하다. 영양염류가 증가하면 식물이 더 빨리 자랄 수 있지만 왜 그러한 생장이 식물의 키를 크게 하는 쪽으로 분배되는가? 생산력 구배상 에서 햇빛은 토양의 영양염류 수준이 증가함에 따라 더 제한되는 자원이다. 키의 증가는 개체로 하여금 더 많은 햇빛을 받도록 해주 는 적응 현상으로 설명될 수 있다. 이와 유사하게 생산력 구배상에 서 키가 작은 종이 키가 큰 종에 의해 교체되는 것은 키가 큰 식물 이 햇빛에 대한 경쟁에서 더 유리하기 때문일 것이다.
먼저 자연 상태의 생산력 구배상에서 식물의 키를 생각해 보자. 예를 들어 지중해성 기후를 갖는 캘리포니아, 칠레, 남아프리카, 그 리고 지중해 지역에서는 계통학적으로 관련이 없는 식물들의 형태 적, 생리적 특징이 현저하게 수렴되는 것을 볼 수 있다 (Moone y 1977, Cody & Mooney 1978). 이 지역들은 모두 상록성 관목식생이 우 점하고 있댜 토성이 거칠어 배수가 양호한 토양으로부터 미세한 토 양에 이르는 구배상에서 식생은 성숙하였을 때의 키가 0.5m 이하인 식물들이 드문드문 홑어져 있는 상태에서부터 키가 5m 이상되는 종 으로 구성된 뻑뻑한 관목림으로 변화된다. 최근에 교란이 없었던 비 옥한 토양에는 키가 10m 나 되는 교목으로 구성된 수관층이 밀폐된 삼림이 우점하고 있는 지역이 많다. 예 를 들어 , 오스트레 일 리 아 남서 부의 Coolga r die Bota n ic a l Di st r ic t 내에서 Beard(1983) 는 그 지역 식생이 사토에 형성되어 있는 키가 3m 정도인 관목림으로부터 가장 비옥한 지역의 키가 25m 나 되는 숲에 이르기까지 다양하다는 것을 알았다. 또한 Beard( 1983) 는 이 전체 지역 식생의 지리적인 면을 고려하면서 토양의 모재를 일정하 게 하면 연간 강우량이 200mm 인 지 역으로부터 1,200mm 에 이르는 기후 구배를 따라 3m 높이의 황원식물에서부터 25m 높이의 삼림에 이르는 똑같은 식생 계열이 존재함을 알았다. 따라서 토양 모재에 의해 비롯되는 소규모의 식생 패턴과 기후와 관련된 광범위한 지리 적인 식생 패턴 사이에는 유사성이 높았다. 이러한 유사성은 두 경우 모두에서 제한 요인이 되고 있는 수분 에 의해 비롯될 수도 있다. 토양형의 차이는 수분 이용도에 영향을 줄 수 있고, 수분 이용도가 지리적인 기후 구배상에서 생산력을 조 절할 수 있는 것과 똑같이 지역적인 생산력을 조절할 수 있다. 다른 한편으로는 질소와 인과 같은 상이한 토양 자원이 생장을 제한할 수 있다. 물은 유기물의 축적과 미생물의 분해에 영향을 주어 부족
되는 영양염류의 무기화율에 영향을 주기 때문에 이러한 상관을 보 일 수 있다. 토양의 모재도 서식처 내에서 부족되는 토양 자원의 축 적률이나 무기화율에 영향을 줄 수 있다. 그러나 Beard 의 분석에 의 하면 부족한 토양 자원이 무엇이든 간에 생산력이 낮은 지역에는 밀도가 낮고 키가 작은 식물종이 우점되어 있고, 반면에 생산력이 증가하는 지역에는 밀도가 높고 키가 큰 식물들이 우점하는 것으로 나타난댜 위스콘신. 미시간. 미네소타의 경우 가장 생산력이 낮은 일부 지 역은 조사토로 되어 있다. 가장 생산력이 높은 장소는 빙퇴석이나 과거 호소의 퇴적물에서 유래된 토양이다. 생산력이 가장 낮은 지역 은 토양 조류. 지의류. 이끼, 양치식물. 협엽초본 그리고 광엽초본이 우점하는 경향이 있고, 여기에 pin che rry나 choke che rry가 드문드문 흩어져 있댜 비옥한 사토에는 참나무사바나 식생이 형성되어 있다. 더 비옥한 토양에는 수관층이 밀폐된 참나무림이 형성되어 있고. 가 장 비옥한 토양에는 당단풍나무와 참피나무로 구성된 밀폐된 삼림 이 형성되어 있다 (Hole 1976, Olson 1958, Whit ne y 1986). 척박한 토양 에 우점하는 식물은 성숙하였을 때의 키가 작고, 식생의 키는 토양 의 비옥도에 따라 증가하는 경향이 있다. 지중해 지역에서와 같이 위스콘신 지역에서는 토양 모재를 포함한 토양과 식생 사이의 일치 성 이 매 우 뚜렷 하다 (Hole 1976). G i vn i sh(1982) 는 척박한 토양의 삼림 지역에서 하층 초본의 밀도 가 가장 높고 비옥한 토양에 형성된 삼림에서 가장 낮은 것으로 보 고하였다. 이것은 비옥한 지역의 삼림이 햇빛을 더 많이 차단하여 하층 초본식물의 밀도를 낮게 하기 때문이다. 그는 또한 표면상으로 위에서 언급한 일반화된 내용과는 상치되는 패턴을 발표하였다. 하 층 초본의 경우(수관층 교목은 제외), 키가 가장 큰 식물종이 가장 척박한 토양에서 우점하였고. 키가 가장 작은 종이 가장 비옥한 토
양에서 우점하였다. 그러나 G i vn i sh 는 하층 식물은 주로 수관층 식물이 차단하고 남 은 햇빛에 대하여 서로 경쟁한다고 주장하였다. 이러한 상황에서는 초본식물의 밀도가 높을 때만 광 차단이 키에 의존적이다. 수관층이 밀폐되어 지표면에 도달하는 광량이 적으면 하층 초본은 빈약하고, 하나의 초본식물이 다른 초본식물에게 그늘을 드리울 기회가 별로 없다. 이러한 상황에서는 식물의 키가 증가함에 따라 수광량이 거의 증가하지 않는다. 따라서 키가 큰 식물이 유리하지 않다 (G i vn i sh 1982). G i vn i sh 의 관찰은 이론에 의한 예측과 정성적으로 일치한다. 죽 식물이 햇빛의 제한을 받고 키의 증가가 수광량을 중가시킬 때 에는 식물의 키가 큰 것이 유리하다. Beard(1 9 44, 1955) 는 신대륙의 열대 지역에 발달된 극상 식생에서 주요 변이 패턴 중의 하나는 연간 강우량에 의해 결정된다고 발표 하였다(그림 5.2A). 그는 일년중 한달 동안만 비가 오는 지역에서부 터 연중 모든 달에 충분한 강우량을 갖는 지역을 비교하면서 점진 적인 종 구성의 변화를 관찰하였다(그림 5.2A). 이것을 〈계절적 군계 계 열 ( seasonal form ati on seri es )> 이 라고 이 름지 었 다. 설 명 을 쉽 게 하기 위해서 Beard 는 이 식생의 연속체를 몇 개의 다른 군계로 나누었다. 그가 조사한 것 중 생산력이 가장 낮은 지역은 사막으로, 작고 가시 가 있는 관목과 다육성 인 초본이 우점 하였다 (Beard 1944, p. 141 ). 이 강우량 구배상에서 그 다음으로 생산력이 낮은 지역은 선인장 관목 림이었는데, 원주상 선인장과 2-4m 의 주걱선인장(pri ckl y p ear) 이 우 점하였고, 여기에 가시가 있는 관목이 산재되어 있었다. 관목상의 가시나무로 된 숲은 3-lOm 에 이르는 상록성 가시나무가 우점하는 다소 개방된 식생으로 강우량이 많은 지역에 나타났다. 그 다음으로 는 높이가 15-20m 인 나무들이 우점하는 낙엽성 계절림인데, 덩굴식 물이나 착생식물이 거의 없었다.
계절적인 강우량 구배상의 다음 식생은 반상록성 계절림으로 키 가 20-26m 인 나무들이 밀폐된 수관층을 형성하고. 덩굴식물은 많지 만 착생식물은 거의 없었다. 그 다음에는 상록성 계절림이 출현하였 는데. 지역에 따라서는 높이가 35m 이상 되고 중간 정도의 층 높이 가 14-30m 의 범위였다 . 착생식물이 잘 발달되어 있고 덩굴식물도 혼하댜 매달 충분한 비가 내리는 지역은 연강우량도 가장 많았는 데. 40m 나 되는 교목층과 하나 혹은 두 개 이상의 하층을 갖는 식 생에 의해 우점된 열대 다우림이 형성되어 있다. 덩굴식물은 별로 없고 착생 식 물은 수관층 근처 에 만 나타났다 (Beard 1944). Beard(1944, 1955) 는 신대륙 열대 지방의 식생은 그것의 상관과 종 구성이 토양 자원과 수분 이용도에 의해 결정되는 연속체를 형 성하는 것으로 보았다. 이 연속체상에서 가장 건조한 지소는 생산력 이 가장 낮고 지표면에 이입되는 광량이 가장 많았다. 이러한 지소 에는 성숙하였을 때 키가 작은 식물이 우점하였다. 강우량이 많은 지소는 생물량이 많고. 지표면에 이입되는 광량이 적었으며, 점점 키가 큰 식물에 의해 우점되었다. Beard(1944) 에 의해 기술된 계절적인 군계 계열 구배가 토양 자 원과 햇빛의 상대적인 이용도 구배라는 것을 믿는다. 그러나 Beard 에 의해 기술된 패턴이 수분 이용도의 계절성과 일치한다고 해도. 이 구배상의 위치에는 수분이 실제로 부족한 중요한 토양 자원이라 는 것에 대한 실험적인 증거가 전혀 없다. 열대 지방에 있는 임학자 들은 질소가 흔히 제한 요인이 된다고 주장하고 있다 (V it ousek 1982). 이 구배상이나 다른 생산력 구배상의 다양한 위치에서 어떤 자원이 제한요인이 되는지를 결정하기 위해서는 직접적인 실험이 필요하댜 그러한 연구는 첨가하는 자원에 대한 여러 종의 반응에 관한 정보를 제공해 주기도 한다. 그러나 부족한 토양 자원이 무엇 이든 간에 Beard(1 9 44, 1955) 에 의해 기술된 열대의 계절적 군계 계
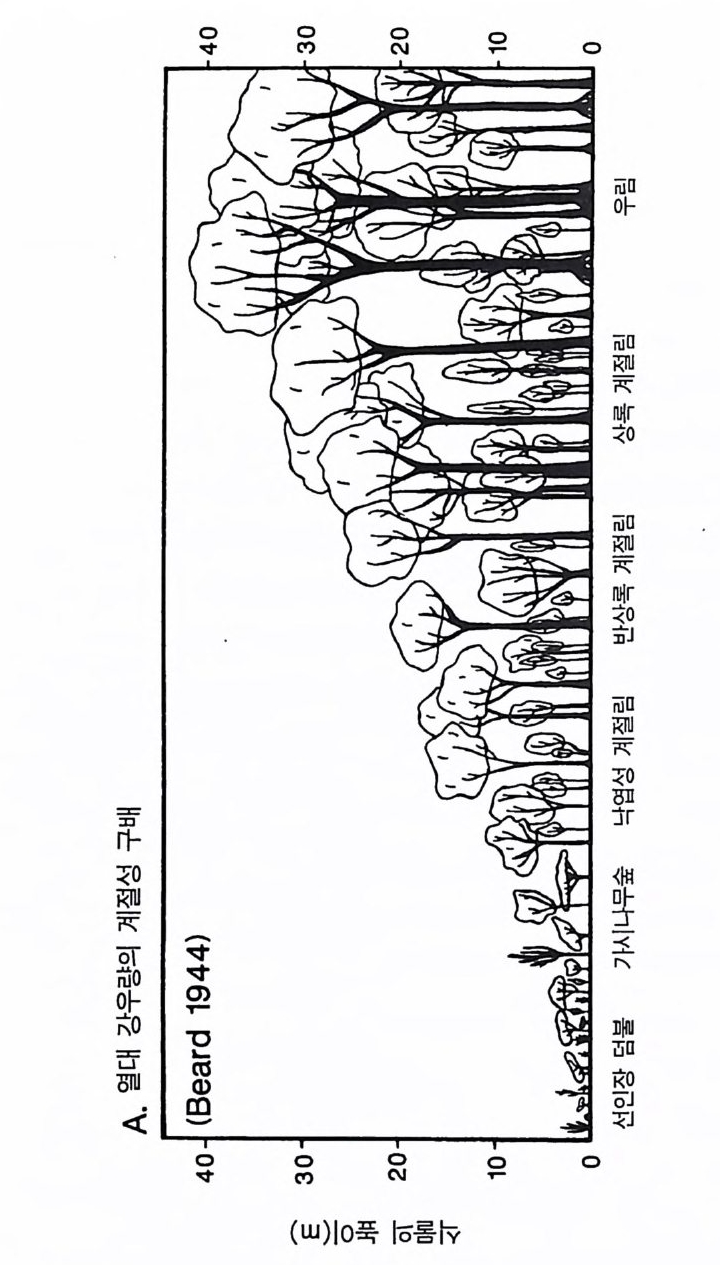 04 3 002 10。 12
04 3 002 10。 12
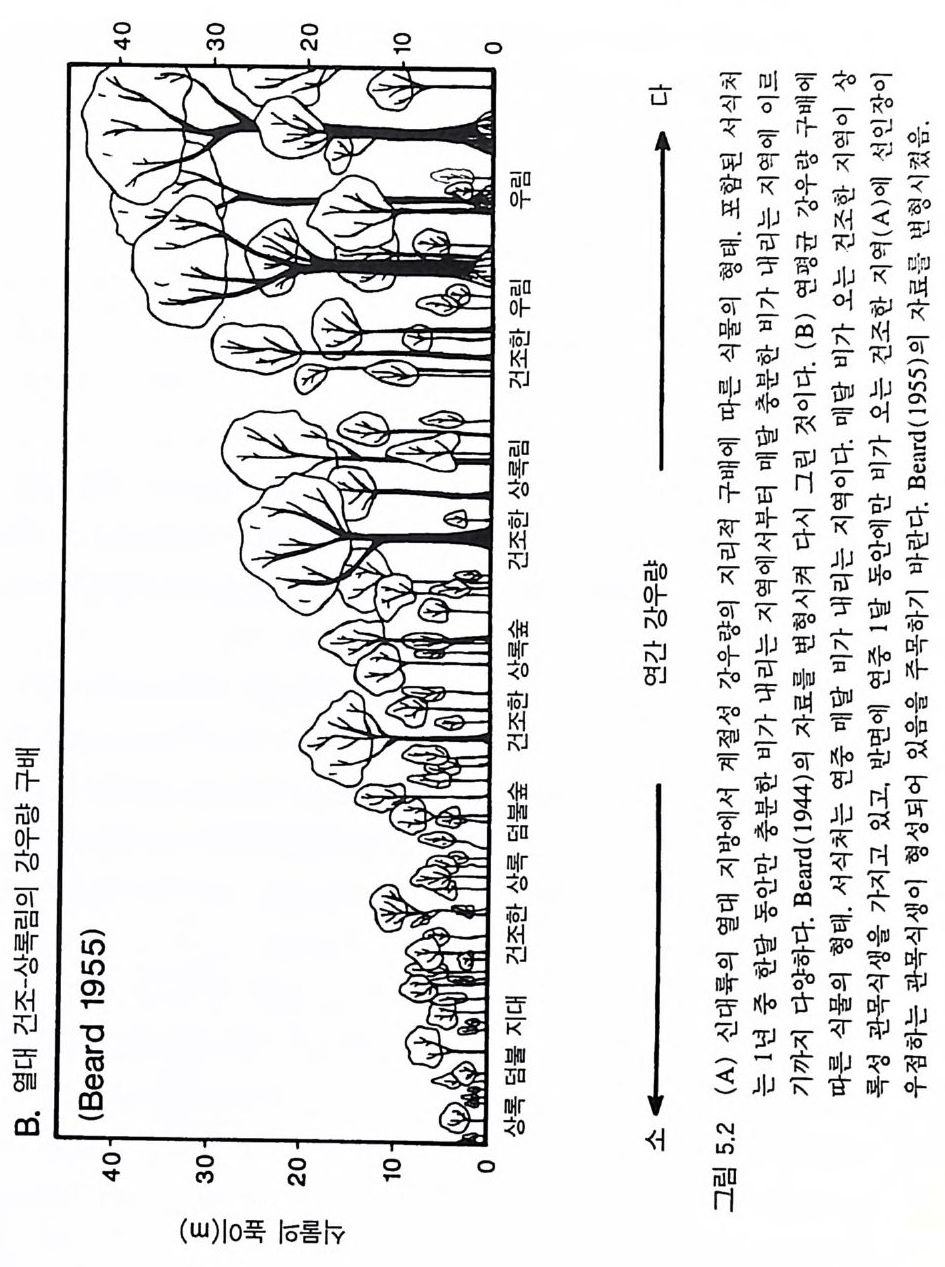
열은 제 4 장에서 전개시킨 이론과 일치하는 토양 자원:광 구배상에 서 식물의 상관이 변화됨을 분명히 보여주고 있다. 따라서 이 이론 은 광범위한 군계 전이 패턴을 설명하고 있는 것 같다. Beard(}944, I955) 는 또한 몇 개의 다른 환경 구배도 인식하였는 데, 그 구배상에서 신대륙 열대 지방의 우점종이 뚜렷하게 구분되었 다(그림 5.2 , 5.3 ). 이러한 모든 구배는 생산력 구배이며, 각 구배는 저지대 열대 우림의 생산력과 비교해 생산력을 감소시키는 특정한 하나의 환경변수나 일단의 변수들을 나타낸다. 각 구배에서 생산력 의 감소는 종 구성의 변화를 초래하는데. 생산력이 낮은 서식처에는 키가 작은 종이 우점한다. 〈 건조한 상록성 군계 계열 〉 이 계절 내내 수분 이용도가 일정노r 서식처, 죽 모든 달의 강우량이 거의 같은 서 식처에서 나타난다(그림 5.2 B ). 가장 습한 지역에는 열대 우림이 우 세하고, 가장 건조한 지역은 바위로 되어 있다(그림 5.2B). Beard(1 9 44, 1955) 가 인식한 또 하나의 생산력 구배는 고도 증가 로 인한 온도 감소와 생육계절의 길이가 짧아지는 것인데, 그는 이 것을 〈산록 군계 계열 (mon t ane fo rma ti onser i es) 〉 이라고 불렀으며, 열 대 우림에서부터 툰드라까지 나타난다(그림 5.3A). Beard(1955) 는 두 개의 늪지 군계도 인식하였는데, 식생의 구성은 그 지역의 침수 빈 도와 고인 물의 깊이로 결정 된다(그림 5.38). Wh itt aker(1975) 는 자신이 ecoc li ne 이라고 이름 붙인 두 개의 또 다른 구배를 설명하였다. 첫번째는 북미 남서부의 사막으로부터 애 팔래치아 산맥의 중습림에 이르는 습도 구배였다(그림 5.4 A ). 이 구 배상에서 선인장과 사막관목림은 건조초지로, 건조초지는 단경초지 와 장경초지롤 거쳐 참나무사바나. 참나무숲, 참나무-히코리림으로 변화되고. 마지막에는 당단풍나무. 참피나무 등이 우점하는 삼림으 로 변한다. 두번째는 위도에 의한 온도 구배로 계절적 열대림으로부 터 북방침엽수림과 툰드라까지 나타난다(그림 5.4 B ).
그림 5.2 와 5 .4에 나와 있는 6 개의 구배와 이 장의 앞에서 설명한 바 있는 다른 구배들은 비록 지리적으로 상이한 서식처에 살고 있 는 서로 무관한 종들에 관한 것이지만 이들은 매우 비슷하다. 이러 한 모든 구배상에서 생산력이 가장 높은 서식처에는 키가 크고 잎 면적 지수가 크며 수관층이 밀폐된 삼림이 우점되어 있다. 생산력이 나 현존량을 감소시키는 환경 요인들은 키가 작은 종에게 유리하다. 이러한 구배 중에서 실제로 부족한 토양 자원이 무엇인지를 밝히는 실험적인 증거는 별로 없다 . 또한 이 모든 것은 복잡한 환경 구배로 써, 이 구배를 따라 많은 물리적인 특징이 변화된다. 이러한 물리적 특징 중의 일부는 잠재적으로 부족한 토양 자원의 공급률에 영향을 준다 . 예를 들어 산록 계열의 경우 고도가 높으면 온도가 더 낮다. 낮은 온도는 미생물의 분해 활동을 감소시켜 영양염류의 무기화를 감소시킨다. 이와 유사하게 계절적으로 형성되는 늪지와 늪지 군계 구배상에 서처럼 토양이 혐기 상태로 됨에 따라 분해가 느려진다. 토양의 수 분함량이 낮으면 역시 미생물에 의한 분해가 느려지고, 그 결과 영 양염류의 순환과 공급률이 느려진다 . 따라서 이 모든 구배는 공통점 울 가지고 있는데, 구배를 따라 하나 이상의 부족되는 토양 자원의 공급률이 증가하고 지표면이나 수관층 아래의 어느 부분에서의 광 이용도가 감소하는 점이다 . 따라서 이들 구배에는 토양 자원:광 구 배가 내재되어 있다. 그러나 온도와 같이 구배를 따라 변화하는 일부 요인들은 부족되 는 토양 자원의 공급률에 대한 효과와는 별개로 직접 식물생장에 영향을 준다. 따라서 이러한 광범위한 환경 구배는 비록 그것이 군 집구조를 결정하는 가장 중요한 요인이더라도 토양 자원:광 구배로 뚜렷하게 설명할 수 없다. 전체적으로, 이러한 구배상에서 식생의 정성적인 상관 특성은 제 4 장에서 예측한 것과 일치한다. 죽 각각의 구
 04 30 02 01 。 저
04 30 02 01 。 저
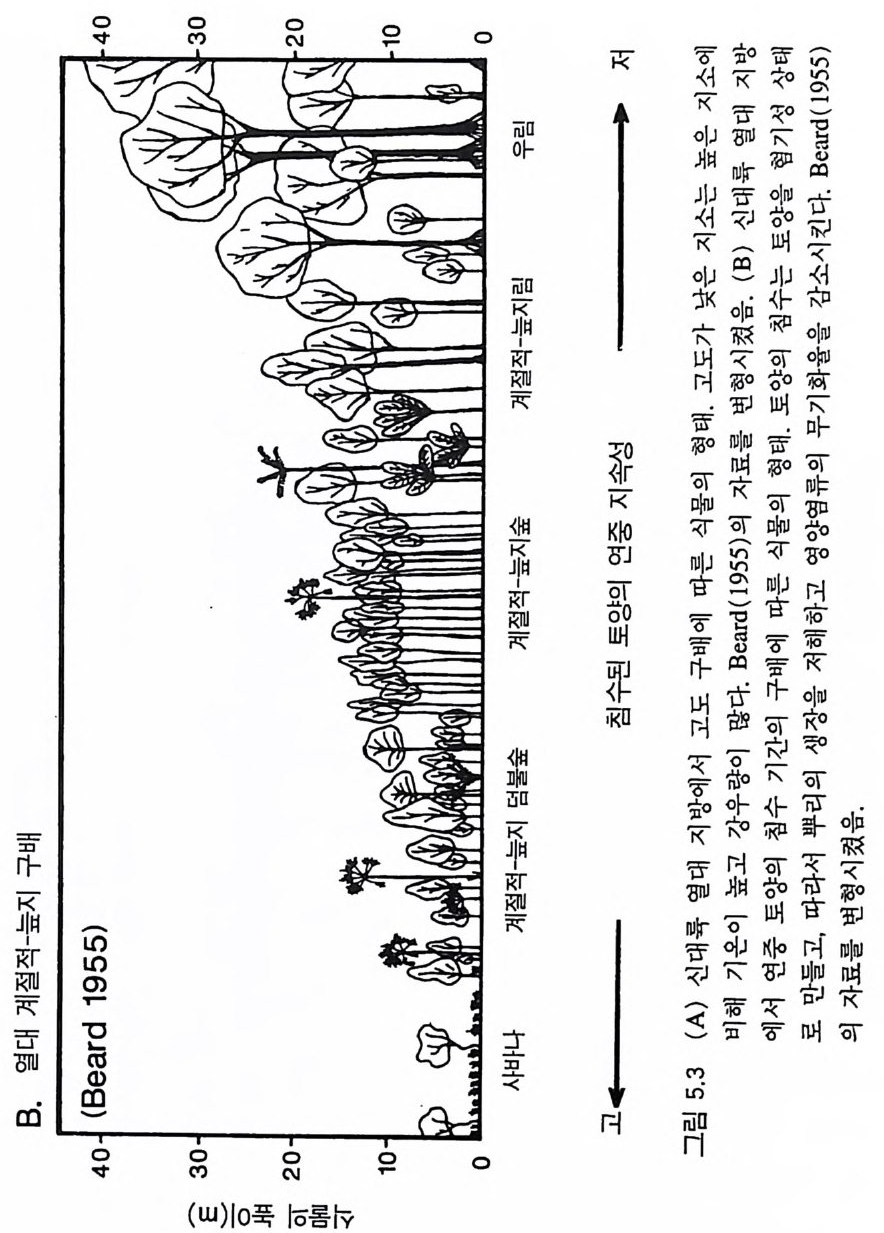
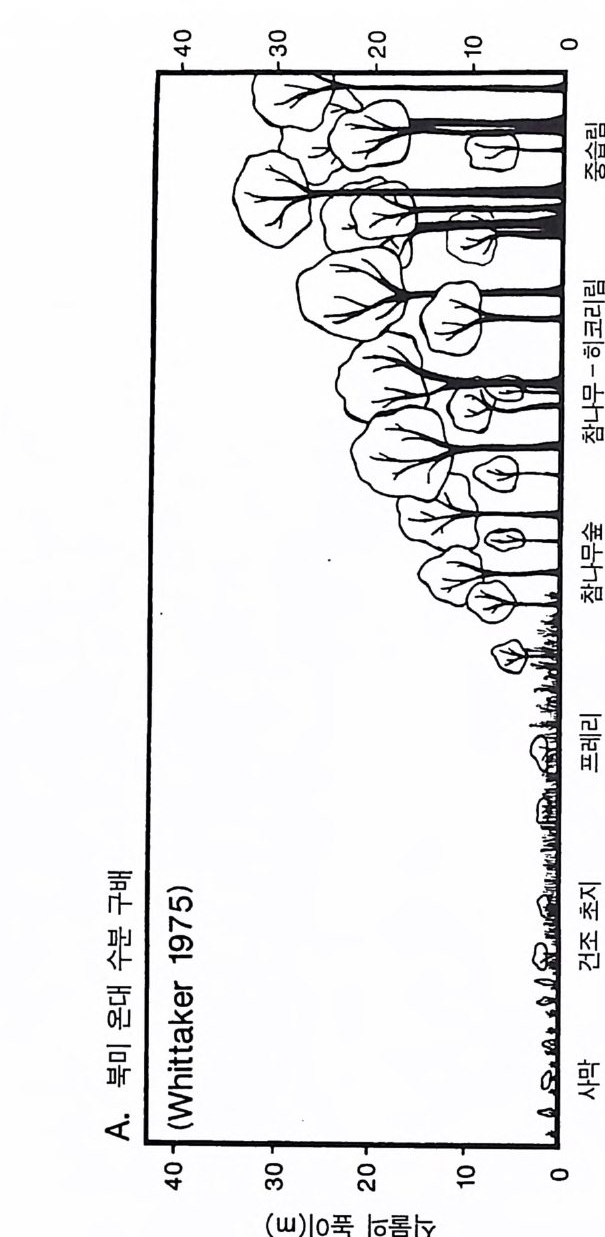

배상에서 식생의 상관적인 특성이 점진적으로 변화된다 . 토양 자원:광 구배상에서 종의 교체와 식물의 키의 분포 패턴은 전세계적으로 동일하다 . 이와 같이 전세계적으로 일정한 분포 패턴 이 존재하는 것은 식물의 진화와 육상 생태계의 구조를 결정하는 중요한 환경 제약이 있음을 의미하는 것이다. 관속식물의 경우 이러 한 패턴은 토양 자원:광 구배상의 상이한 지역에 알맞도록 식물이 분화하는 것과 일치하며, 이러한 구배상에서 경쟁 능력을 결정하는 주요 인자인 식물의 형태와도 일치한다. 4 영양염류공급률 Pasto r 등(1 984) 은 BlackhawkIsland 에서 측정한 토양 영얌염류의 실제 공급률을 토양극상 식생의 생산력과 그것의 구성에 관련시킨 연구를 수행한 바 있다. W i scons i nR i ver 에 있는 70 헥타르 넓이의 이 섬에는 과거에 전혀 벌목이 없었던 일단의 삼림이 사토, 식양토, 유 기토양 등 다양한 토양에 형성되어 있다 .Red pi ne, 스트로브잣나무, red oak, wh ite oak, 단풍나무, 솔송나무 둥에 의 해 우점 되 는 상이 한 삼림은 일단의 토양극상을 나타낸다. 이 모든 지소에서 질소는 가장 부족한 토양 영양염류로 여겨진다 (Pas t ore t a l. 1984). Pasto r 둥 (1984) 은 현장에 비닐백을 묻는 방법을 이용하여 8 개의 서로 다른 지소에서 질소 무기화율과 지상부 1 차 순생산력의 계절 변화를 측정하였다. 그들의 조사 결과에 의하면 1 차 생산량은 연간 질소 무기화량과 높은 상관이 있었고(그림 5.5A), 각 지소의 하층토 에 있는 미사와 점토의 비율과도 높은 상관이 있었다(그림 5.5 B ). 하 층토의 미사와 점토의 함량은 이들 토양이 형성된 모재를 나타내는 것이다. 따라서 미사와 점토의 함량이 높은 모재는 질소 무기화율이
 A.
A.
높은 토양을 형성하고 1 차 생산력도 높다. 반면에 사질성 모재는 질 소 무기화율이 낮고 생산력이 낮은 토양을 형성한다. 하층토의 모래 함량이 가장 많은 곳에 형성된 식생에서부터 모래 함량이 가장 적은 곳에 형성된 식생에 이르기까지 이들을 순서적으 로 생각해 보자. 그림 5.5B 에는 솔송나무가 포함되지 않았는데, 그 이유는 솔송나무가 자라는 지역은 무기토양이 없고 유기물 함량이
59% 나 된다 (Pas t or et al. I984). Red pine (Pin u s res i nosa) 은 임 학자들 (Powells 1965) 에 의해 내음성 이 없는 식물로 분류되는데, 모래가 가장 많은 지역과 질소 무기화 율이 가장 낮은 지 역 에 우점 한다. 나는 Blackhawk Island 에 서 이 식 물의 키가 얼마인지 잘 모르지만, 이 식물은 성숙하였을 때의 키가 16-22m 에 이른다 . 그 다음으로 질소 무기화율이 높고 모래가 가장 적은 토양에는 스트로브잣나무가 우점하였다. 스트로브잣나무는 중 간 정도의 내음성을 갖는 것으로 여겨진다. 죽 햇빛에 대한 경쟁력 이 중간 정도이다. 스트로브잣나무는 매우 크게 자랄 수 있지만 대 부분이 18-24m 이 댜 그 다음에는 redoak 에 의해 우점된 두 지역이 있는데, 질소 무기 화율이 높은 사질식양토에 발달되어 있다 . Redoak 는 내음성이 중간 정도로 분류된다 (Powells 1965). Whit e oak 가 우점하는 지역은 red oak 가 우점하는 지역에 비해 하층토에 미사와 점토 함량이 높고 질 소 무기화율이 높다. Whit e oak 은 내음성 식물로 분류된다. 죽 햇빛 에 대한 경쟁력이 높은 식물이다. 당단풍나무가 우점하고 있는 두 지역은 연간 질소 무기화율이 가 장 높고 하층토에 미사와 점토 함량이 가장 많다. 당단풍나무는 그 지역에서 내음성이 가장 강한 식물로 분류된다. 죽 햇빛에 대한 경 쟁력이 가장 큰 식물이다 . 나무의 키는 22-30m 에 달한다. Pasto r 등 (1984) 은 이 지소에서 지표면에 이입되는 광량을 언급하 지 않았다. 그러나 Joh n Pas t or 는 친절하게도 그가 각 지소의 지표면 에서 측정한 광량을 나에게 제공하였다(그림 5.6). Red pi ne 이 우점하 며, 질소 무기화율이 가장 낮은 곳은 지표면에 도달하는 광량이 가 장 높았고 , 당단풍나무가 우점하며 질소 무기화율이 가장 높은 지역 은 지표면의 광량이 가장 낮았다. 실제로 Blackhawk Island 에 있는 이 7 개 지역에서 연간 질소 무기화율과 지표면의 광량 사
 160 A.
160 A.
그림 5.6 (A) 위스콘신의 Blackhawk Island 에 있는 8 곳의 삼림지에서 지표면에서 측정 된 광량과 연간 질소 무기화율 사이의 관계. 연간 질소 무기화율은 표충토에서 만 측정한 것입 (B) 표충토와 하층토의 연간 질소 무기화율을 사용하였을 때 에도 기본적으로 동일한 관계가 있음을 알 수 있다 . Pas t or 가 자신의 미발표된 광량 자료를 본인이 이용할 수 있도록 허락해 준 것을 매우 고맙게 생각한다.
이에는 현저한 음의 상관이 있었다 . 표층토의 질소 무기화율이나(그 림 5.6A). 표층토와 하층토의 전체 질소 무기화율의 어느 것을 사용 해도 이러한 음의 상관이 나타난다(그림 5.6 B ). 따라서 이 7 개의 삼 림지소는 토양 자원:광 구배상의 위치가 각각 다르다. 이 구배상에서 각 삼림지소의 위치는 토양이 형성되는 초기 모재 에 따라 다르지만 임학자들에 의해 분류된 내음성 순위에서 보는 것처럼 광에 대한 그들의 경쟁 능력 순서와 잘 일치한다. 이러한 결 과는 우점하는 나무의 질소에 대한 경쟁 능력과 광에 대한 경쟁 능 력 사이에 대응책이 있다는 가설과 일치한다 . 이것은 〈 통합된 경쟁 능력〉이란 G ri me(1979) 의 개념과는 일치하지 않는다. G ri me 이 말하 는 통합된 경쟁 능력이란 광에 대한 경쟁력이 우세한 종이 질소나 인 등에 대한 경쟁력도 우세하다는 것이다. 이 패턴과 일치하지 않는 하나의 지소는 솔송나무가 우점하는 지 소이댜 솔송나무 (Tsu g a canadens i s) 는 내음성이 강한 종이다 (Powells 1965). 그러나 이 종이 우점하는 지소의 토양은 질소 무기 화율이 매우 낮다. Pasto r 등(1 984) 은 그림 5.5B 에서 이 종을 생 략하 였는데, 그 이유는 이 지소가 하층토가 없고 유기물 함량이 매우 높 기 때문이다. 솔송나무는 껍질과 잎의 타닌 함량이 높은 것을 포함 하여 몇 가지 면에서 특이한 종이다 (Powells 1965). 실제로 솔송나무 껍질의 타닌 함량은 매우 높아 타닌산을 생산하기 위해 솔송나무를 선별적으로 벌목하여 껍질을 벗겼었다. 타닌 함량이 높아 이 지소 토양의 질소 무기화가 저해를 받을 수도 있다. 솔송나무는 내음성이 크기 때문에 토양의 영양염류가 풍부한 지 역에도 침입할 수 있다. 그러나 질소 무기화율이 낮은 토양에 솔송 나무가 우점하는 것은 성체가 척박한 토양에서도 살 수 있다는 것 을 의미한다. 겉으로 보기에 상치되는 이러한 현상은 솔송나무에서 생산되는 낙엽의 타닌 함량에 의해 비롯된다 . 솔송나무는 실생이
정착하기에 알맞은 지역에 침입하여 우점종으로 자랄 수 있다. 그 러나 솔송나무 낙엽의 타닌 함량이 매우 높아 결국 하층토의 질소 무기화율을 현저히 감소시켜 자신의 재생이 저해된다. 이러한 효과 는 솔송나무를 벌목한 후 재생이 안되는 것으로 설명할 수 있으며, 이러한 이유로 그 지역에서는 장기간에 걸친 솔송나무 순환이 야 기된댜 솔송나무의 예는(생태계를 구성하는 모든 종과 생태계에서 진행되는 과정들은 서로간에 영향을 주고 상호 작용을 할 수 있는 잠재력을 가지 고 있음을 보여주는 것) 개체군 생태학자들은 항상 인정하지 않지만 생태계 생태학자들에게는 거의 신조와 같은 다음과 같은 점을 강조 하는 것이다. 죽 생태계 내에서 고립된 종은 없고. 모든 종은 서로간 에 영향을 주고 상호 작용을 할 수 있는 잠재력을 가지고 있다. 이 러한 피드백은 주어진 상호 작용을 안정한 혹은 불안정한 상태로 만들 수 있다. Pasto r 등 (1984) 이 연구한 8 개의 지소 중 7 개에서는 토양의 무기화에 대한 낙엽 생산의 장기간의 피드백이 상호 작용을 안정되게 하여 각 토양이 형성되는 모재와 밀접한 관련이 있는 안 정된 식생 구성을 유도하는 것으로 보인다. 따라서 이러한 지소는 토양극상으로 간주된다. 이러한 토양 모재와 식생 구성 간의 의존 관계는 하층 토양의 미 사 및 점토 함량과 최종 우점식생의 구성 간의 상관을 통해 분명히 알 수 있다. 하층 토양에 미사와 점토가 적은 사토는 배수가 잘되고 NH4 와 결합할 수 있는 양이온 치환 부위가 없다. 따라서 그러한 모 재는 세탈에 의한 질소 소실이 많고 결국에는 질소 수준이 훨씬 낮 은 상태에서 평형에 이른다 (Olson 1958). 점토와 미사의 함량이 높은 모재는 세탈에 의한 질소 소실이 적어 결국 BlackhawkIsland 에서처 럼 질소 이용도가 높은 토양이 형성된다. 그러나 이 패턴은 그 지역에 살고 있는 식물에 의해서도 영향을
받고, 그들이 생산하는 낙엽의 질에 의해서도 영향을 받는다. c:N 비 가 낮고 쉽게 분해되는 낙엽을 생산하는 식물은 더 풍부한 분해자 군집을 유지할 수 있다. 따라서 질소 무기화율이 더 빠르다. 만약 그 러한 식물이 비옥한 토양에 우점하는 종이라면 이것이 낙엽 생산을 통해 자신의 생존에 유리한 토양을 형성하도록 하는 안정된 피드백 환(fe edback loo p)을 유도할 수 있다. 이러한 현상은 질소 함량이 높 고 쉽게 분해되는 낙엽울 생산하는 당단풍나무에서 볼 수 있다 (Pasto r et al. 1984). 척박한 토양에 우점하는 식물이 C:N 비가 높고 천천히 분해되는 낙엽을 생산하면 무기화율이 낮을 수 있으며, 그러한 종은 자신들의 생존에 유리한 환경 조건을 유지하려는 경향이 있다. Red pi ne 이 바 로 이러한 경향이 있는 식물이다 (Pas t ore t a l. 1984). 솔송나무의 생물 학적 특성을 파악하려면 더 많은 연구를 해야 하지만, 이것이 정착 하기 위해서는 비교적 비옥한 토양이 필요하다. 그러나 솔송나무는 결국 질소 무기화율이 낮고 척박한 산성 토양을 형성하는데, 그 이 유는 솔송나무가 생산하는 낙엽이 산성이고 반응성이 낮은 유기화 합물을 함유하고 있기 때문이다. 이러한 패턴이 그저 우연에 의한 것인가? 아니면 식물 낙엽의 질 이 진화된 특성인가? 이 질문에 대답하기 전에 우리가 알아야 할 것이 많이 있지만 개체 사이의 선택적인 차이는 그들이 생산하는 낙엽의 질을 변화시키는 것으로 판단된다. 그러나 이러한 주장은 각 개체가 자신의 직접적인 환경과 자신의 실생이 경험할 환경에 어떤 영향을 주느냐에 달려 있다. 생산된 낙엽은 대부분 그 나무 밑에 떨어지고 그것과 그것의 실 생이 뿌리를 내리고 있는 토양에 영향을 준다. 그렇다면 극상을 형 성하는 종은 자신이 우세한 경쟁자가 되도록 토양 영양염류 공급률 을 강화시키는 낙엽을 생산하여야 한다는 주장이 가능하다. 그러나
어느 지역의 최초의 모재와 마지막 토양극상 사이의 일치는 이러한 낙엽 피드백 효과가 최종적인 식생 구성을 결정하는 주요 인자가 아님을 보여주고 있다 . Blackhawk Islands 에 있는 소나무림과 참나무 림에 지난 수천 년 동안 당단풍나무 종자가 분명히 여러 번 침입 하였고, 당단풍나무가 생산하는 낙엽의 영양염류 이용도가 높지만 아직 소나무나 참나무를 교체하지 못하고 있다. 다른 한편으로는 솔송나무와 같이 자신의 경쟁 능력과 일치하지 않는 낙엽을 생산 하는 종은 한 지역에 장기간의 지속적인 삼림을 유지하지 못할 수 있다. 우점종이 생산하는 낙엽의 질과 토양의 질소 공급률 사이의 상관 관계에 대하여 다르게 설명할 수도 있다 . 예를 들어 척박한 토양에 우점하는 식물의 경우 질소의 손실이 가장 적은 개체가 그렇지 않 은 개체들보다 유리할 것이다 (Cha pi n 1980). 질소 손실을 가장 적게 하는 간단한 기작은 잎이 떨어지기 전에 잎으로부터 질소를 전이시 키는 것이다. 식물에게 질소가 더 중요하면 할수록 그와 같은 질소 전이를 통해 얻는 장점은 더 클 것이다. 질소 전이에 비용이 든다고 가정하면 질소가 빈약한 토양에 우점하는 종은 질소가 더 많은 서 식처에 우점하는 종에 비해 C:N 비가 더 높은 낙엽을 생산할 것이 댜 토양의 질소 공급률에 대한 낙엽의 상이한 C:N 비의 영향은 식 물 개체에서 최적 질소 보존을 위한 자연 선택의 간접적인 효과일 수 있다. 진화적인 설명이야 어떻든 간에 BlackhawkIslands 에 있는 식생에 대한 연구는 초기 모재와 식생 사이의 뚜렷한 연관을 보여 주고 있으며, 제 4 장에서 자연식생에 실제로 존재하는 것으로 가정한 바 있는 토양 영양염류 공급률과 지표면에서의 광량 사이의 관계룰 보여주고 있다 . 내음성: 앞에서 언급한 바와 같이 내음성에 대한 임학적인 개념은 낮은 광량에서 실생과 유묘가 생존하고 자랄 수 있는 능력을 측정
하는 것이다. 교목은 생활사 중 이 시기에 가장 낮은 광 이용도를 경험하며. 따라서 내음성은 햇빛에 대한 경쟁 능력의 측정일 수도 있댜 유감스럽게도 내성이란 용어는 종내 혹은 종간 상호 작용에 대한 의미가 포함되어 있지 않고 어떤 혹독한 환경에서 생존할 수 있는 능력의 차이만을 제시하는 것이다. 그러나 수목이 견뎌야 하는 상이한 피음 수준은 생물량과 다른 식물의 키 분포에 의해 결정되 는 광의 실질적인 이용도 수준인 것이다. 수직 구배에 따른 광량의 감소는 다른 자원의 소비 패턴과 정성적으로 같다. 그것만으로도 내 음성이 광에 대한 경쟁 과정의 한 구성원임을 알 수 있다. 식물의 경쟁을 이해하기 위해서는 다양한 크기를 갖는 식물의 생리적 그리 고 형태적인 특징이 어떻게 종의 경쟁적 상호 작용을 결정하는지를 연구함으로써 내성의 개념을 넘어서야 한다. 5 종자의 크기 종자의 크기는 토양 자원:광과 손실률 구배상에서 식물의 위치에 의해 영향을 받는다. 손실률이 낮고 영양염류가 풍부한 서식처에서 는 큰 종자가 유리하다. 그 이유는 큰 종자는 키가 큰 실생을 만들 고 키가 큰 실생은 더 많은 햇빛을 받을 수 있기 때문이다. 작은 종 자는 영양염류가 빈약한 서식처나 손실률이 높은 서식처에서 유리 하다. 모델에 포함되어 있지는 않지만 다른 많은 요인들이 종자의 크기에 영향을 줄 수 있다. 예를 들어 종자 혹은 실생을 먹는 포식 자가 실생의 밀도를 충분히 낮게 유지하여 지표면 근처에서의 광량 의 수직 구배가 거의 없을 정도라면 비옥한 토양에서 큰 종자가 유 리하지 않을 것이다. 수분이 부족한 사막 환경이나 식물이 성장함에 따라 광이 부족하
게 되는 지역에서 생육계절 초기에 생장이 빠른 개체가 짧은 기간 이지만 부족한 자원을 더 많이 얻을 수 있다면 계절성은 종자가 작 은 개체보다 큰 개체들에게 유리하게 작용할 것이다. 두꺼운 낙엽층 도 큰 종자가 유리하도록 한다. 그 이유는 큰 종자는 뿌리가 토양층 까지 자라고 잎이 햇빛이 있는 낙엽층 위로 자라는 데 필요한 양분 을 더 많이 가지고 있기 때문이다. Gr i me 과 J e ffe r y(l 964) 는 북미에 있는 9 종의 교목 실생을 피음이 강한 조건에서 12 주 동안 키운 후 실생의 최대 키가 종자의 무게와 상관이 매우 높음을 알았다. 큰 종자는 ALLOCATE 에 의해 가정한 바와 같이 키가 큰 실생을 생산한다. 해바라기 실생 사이의 종간 경 쟁 실험에서, 피음이 높은 지소에서 키울 때 큰 종자로부터 나온 실 생은 키가 더 빨리 자라서 작은 종자로부터 나온 실생과의 경쟁에 서 이 겼다 (Ku ri owa 1960). Sa li sbu ry (1942) 는 영국에 있는 식물 중 377 종의 종자 크기를 보 고한 바 있다. 그는 이 식물들을 광량이 높은 개방된 서식처로부터 수관층이 밀폐된 광량이 낮은 서식처에 이르는 구배상의 위치에 따 라 분류하였다. 그가 사용한 7 개의 그룹은 (1) 개방된 서식처에 나 타나는 종, (2) 반 밀폐된(비피음) 서식처에 나타나는 종, (3) 목야지 종, (4) 관목지나 숲의 가장자리에 나타나는 초본, (5) 숲의 임상식 물, (6) 관목, (7) 교목이 었다. 그림 5.7A 에 요약되 어 있는 그의 연구 결과는 햇빛의 제한이 증가함에 따라 종자의 평균 크기가 커지는 것을 보여주고 있다. 그는 수관층이 밀폐된 지역을 천이의 후기와 연관시키고, 모수에 의해 종자나 혹은 다른 생식체에 제공되는 영양 물질의 공급량이 많으면 그 종이 우점하는 식생의 천이가 더 진행 된 것이라고 주장하였다 (Sal i sbu ry 1942, p. 18). Baker(1972) 는 캘리포니아 고유식물 2,160 종의 종자 크기를 분석 하였다. 더 비옥한 토양과 더 중습한 기후를 갖는 서식처에 우점하
 A. 영국 식물들
A. 영국 식물들
그림 5.7 (A) 다양한 식물종의 종자 크기와 각 종의 수도가 가장 높은 토양 자원: 광 구배상의 점 사이의 관계. 영국에 있는 287 종의 식물을 이용하였다. 종 자 크기의 평균과 표준오차는 Sal i sb ury(1 942) 의 자료에 기준한 것이다. 종은 Sa li sbu ry에 의해 서식처의 개방 정도(광 이용도)에 따라 구분되었 다. (B) 캘리포니아에 있는 2,160 종의 식물에 관한 유사한 자료 (Baker 1972). 종은 Baker 에 의해 생활형으로 구분되었다. 종자의 크기가 대수눈 금임을 주목하기 바란다.
는 교목은 큰 종자를 갖는데. 건조하고 영양염류가 빈약한 초지나 사막 그리고 관목지역에 우점하는 1 년생 초본은 작은 종자를 갖는 다(그림 5.7 B ). 중간 정도의 서식처에 가장 혼히 나타나는 다년생 초 본과 관목은 종자 크기가 중간 정도였다. Baker(1972) 는 수분과 고 도를 포함한 다른 환경변수들도 종자 크기에 영향이 있음을 보여주 었다. 따라서 척박한 토양과 광량이 많은 서식처에 특징적인 종은 토양이 비옥하고 지표면에서의 광량이 낮은 서식처에 특징적인 식 물에 비해 평균적으로 종자의 크기가 적다. 6 분배 패턴 제 4 장의 이론으로 예측한 바에 의하면 식물의 뿌리, 줄기, 잎으로 의 분배는 중간 정도의 생산력 구배상에서 잎으로의 분배량이 최대 가 되고, 척박한 토양에서 비옥한 토양으로의 구배를 따라 뿌리로의 분배는 줄어들고 줄기로의 분배는 늘어나는 쪽으로 변해야 된다. 많 은 관찰 결과들이 일반적으로 이러한 예측과 일치한다. 예를 들어 Cha pi n(1980) 은 척박한 토양의 서식처에 우점하는 식물은 비옥한 토양의 서식처에 사는 식물에 비해 균근 연합체(뿌리의 기능이 있음) 와 뿌리 에 더 많은 투자를 한다는 것을 알았다. Mooney ( 1972) 는 이 와 유사하게 1 년생과 다년생 초본식물은 관목에 비해 뿌리:줄기의
비가 더 큰 경향이 있고. 관목은 교목에 비해 뿌리:줄기의 비가 더 큰 경향이 있다고 주장한 바 있다. 만약 초본으로부터 관목을 거처 교목에 이르는 계열이 척박한 토양에 우점하는 종으로부터 바옥한 토양에 우점하는 종의 계열을 나타내는 것이라면. 이것은 더 척박한 토양에 우점하는 식물은 뿌리로의 분배량이 크고 줄기로의 분배량 이 적다는 것과 일치하는 것이다. 애팔래치아의 중습림에서부터 사막에 이르는 구배를 생각해 보자 (Whit tak er 1975. 그림 5.4 A ). 튤립나무가 우점하는 골짜기의 중습림은 순생산력이 1 , 300 g /m 2 /y r 안데. 뿌리 :줄기의 비가 0.16:I 이다 (Whit tak er 1975, 그의 표 5.1 ). 생산력이 약간 낮은 참나무-소나무림 (1,060 g /m 2 / y r) 은 뿌리:줄기의 비가 0.52:1 이었다 (Wh itt aker 1975). 평균 생산력이 600 g /m2/ y r 인 (Wh itt aker 1975. 그의 표 5.2) 온대 초지 (프레리)는 뿌리 :줄기의 비가 6.5:1 이었다 (Barbour et al. 1980). (이와 같이 뿌리:줄기의 비가 높은 것은 부분적으로 주기적인 불과 포유동물의 방목에 의해 야기될 수 있는데, 이 두 가지 요인은 모두 지하부로의 분배 량을 증가시킨다.) 비록 나는 사막식물에서 정확한 뿌리 : 줄기의 비를 조사하지는 못하였지만 Cod y (1986) 가 조사한 사막식물의 지상부와 지하부 생활형 패턴에 의하면, 수명이 긴 많은 사막식물들은 뿌리: 줄기의 비가 2 이상인 것으로 되어 있다. Wh itt aker 는 사막의 평균 생산력 이 90 g /m2/y r 인 것으로 보고하고 있다. 따라서 그림 5 .4 A 의 구배는 그 구배상에서 자원이 부족한 토양에서는 뿌리에 더 많은 분배를 하고 자원이 풍부한 토양에서는 줄기에 더 많은 분배를 하 는 생산력 구배이다. 동일종 내에서 물과 무기염류에 의해 제약을 받는 서식처에 살고 있는 개체들은 뿌리 : 줄기의 비가 높고. 영양염류가 풍부하고 광량이 낮은 서식처에 있는 개체들은 그 비가 낮다 (Moone y 1972, Gr im e & Hunt 1975, Shamsi & Whit eh ead 1977, Grim e 1979, Parris h & Bazzaz
1982. Bloom et a l. 1985, Chapi n & Shaver 1985). 따라서 종간 그리 고 종 내 모두의 경우에서 줄기로 분배되는 양에 대한 뿌리로의 분배량은 토양 자원:광 구배상에서 필수 자원의 최적 획득 이론이 예상하는 것과 일치하면서 다양한 경향을 보이고 있다 (Ra pp o rt &Turner 1975, Til m an 1982, Bloom et al. 1985, 이 책의 제 2 장). 그러나 뿌리:줄기의 비는 지상부 자원에 대한 지하부 자원의 획 득에 관한 분배 패턴의 추정지수에 불과하다. 뿌리는 토양 자원을 획득하는 구조물로서의 기능이 있을 뿐만 아니라 지지 기관이나 저 장 기관의 역할도 한다. 지상부가 매년 다시 형성되는 다년생 협엽 그리고 광엽 초본과 같은 식물의 경우 줄기의 재형성에 사용되는 자원은 줄기의 기부와 뿌리에 저장되어 있다 (Moone y 1972). 이러한 종이 다음 계절에 생장하기 위해서는 광합성 산물을 저장화합물에 분배하여야 한다 . 온대 초지에 있는 많은 종은 뿌리:줄기의 비가 높 은데. 이것은 영양염류 흡수를 위한 뿌리로의 분배량 증가뿐만 아니 라 물질저장의 결과이기도 하다. 현존량은 기능 면에서 순환율이나 전체 투자를 나타내는 것이 아 니댜 뿌리는 줄기에 비해 순환율이 훨씬 빠르고, 식물은 균근 연합 체에 상당량의 생산량을 투자하기 때문에 뿌리의 생물량은 지하부 자원의 획득을 위한 분배량의 추정치로서는 적당하지 않다. 더구나 줄기는 잎을 지탱하는 데 이용되는 구조적인 조직과 광합성 조직을 같이 가지고 있다. 키가 큰 식물은 작은 식물에 비해 더 많은 지지 가 필요하며 (Greenh ill1 881, McMahon 1973), 키가 작은 식물에 비해 지하부의 구조적 조직의 생산에 더 많이 분배하여야 한다. 이러한 이유 때문에 뿌리:줄기의 비는 광 획득에 대한 영양염류 획득을 위 한 분배 패턴에 대한 근사치에 지나지 않는다. 이러한 문제점들이 있지만 뿌리:줄기의 비는 서식처에 따라 차이가 크고, 패턴이 잘 일 치하기 때문에 식물 군집의 구조에서 분배 대응책의 역할을 하는
것으로 간주되어야 한다. 7 실험상의 구배에서 식물의 수도와 형태 그림 5.2-5.6 에 요약된 구배들은 지리적으로 넓은 범위를 갖는 다 양한 환경 복합체의 구배상에서 식생 구성과 식물 형태의 주요 변 화를 보여준다. 이러한 광범위하고 상관 관계적인 패턴들이 매우 유 사하기 때문에 매우 상이한 서식처에서 서로 비슷한 기작이 작용하 리라고 판단할 수 있지만, 모든 상관 관계적인 패턴은 또다른 설명 이 가능하댜 이러한 패턴이 광과 토양 자원을 획득하기 위한 경쟁 에 의해 야기되었다는 가설을 검증하기 위해서는 실험적 구배를 확 립할 필요가 있다. 실험적인 구배에 대한 식물의 반응이 좀더 복잡 한 자연환경 구배상에서 관찰되는 패턴과 유사하면 이론적인 예측 이 지지를 받을 것이다. 실험적인 구배를 설정하는 데 사용되는 방법이 몇 가지 있다. 첫 번째 방법은 주어진 서식처에서 부족한 토양 자원이 무엇인지를 실 험적으로 알아낸 다음 이 부족한 자원의 처리량을 달리하여 반복구 에 공급하는 것이다. 제 4 장에서 제시한 이론이 자연 현상을 단순화 시킨 것과 잘 일치한다면 영양염류의 공급률이 높으면 생산량이 중 가하고 토양 자원의 수준이 높아지며, 지표면에서의 광량은 감소되 고 분배 패턴과 생활사에 변화가 초래될 것이다. 두번째 방법은 그늘을 지우거나 인위적으로 광량을 증가시키는 것이다. 세번째 방법은 햇빛과 영양염류를 함께 자연식생에 직접 처 리하는 것이다. 이론상 영양염류의 첨가는 식물의 형태와 식생 구성 에서 입사되는 광량을 감소시키는 것과 비슷한 변화를 초래하여야 한다. 세번째 방법은 제 4 장에서 전개시킨 이론을 직접 실험을 통해
검증할 수 있는 가장 좋은 방법이다. 불행하게도 아직까지는 토양 자원:광 구배의 최종 효과를 측정할 수 있을 정도로 반복구가 많고 오랫동안 계속되어 온 연구가 별로 없다. 영국의 Harp endon 에 있는 Roth a mste d Exp er i m enta l S t a ti on 의 Park Grass Ex p er i men t s 는 대 부분의 처 리 에 반복구가 없다. 그러 나 이 연 구는 1856 년부터 시작되어 130 년 이상 계속되고 있다 (Lawes & Gi lb ert 1880, Thursto n 1969, Til m an 1982 참조). 20 개의 상이한 실험구 는 각각 130 년 동안 독특한 조합의 영양염류를 공급받아 왔는데, 영 양염류의 공급 패턴과 토양 p H 를 다양하게 조작한 후 종 구성상에 현저한 변화가 있었다. 이러한 변화는 주요 제한 자원이 햇빛과 토 양염류 특히 질소와 인이라는 가설과 일치한다 (T i lman 1982). 그러나 각 실험구는 매년 두 번씩 건초를 얻기 위해 풀을 베기 때문에 이 토양에서 일반적으로 진행될 수 있는 삼림으로의 천이가 억제되어 왔다. 이러한 손실은 제 4 장에서 분석된 손실과 완전히 동등한 것은 아니다. 그 이유는 이 손실이 모든 종에게 균일하게 부과되는 밀도 독립적 사망률이기 때문이다. 풀베기는 지상부 생물량을 제거하지만 필연적으로 식물을 죽이는 것은 아니다. 풀베기는 키가 낮은 초본식물에 비해 키가 큰 목본종 울 훨씬 많이 제거하는 효과가 있다. 토양 p H 가 4.5 이상인 실험구 만을 고려하면, 질소 공급률이 가장 높고 완전한 영양염류를 공급받 는 구는 키 가 가장 큰 종인 Alop ecurus 와 Arrhena t herum 이 우점 하였 댜 이들은 모두 튼튼하고 직립성안 다년생 벼과식물로 . 평균 키가 60-80cm 였다. 반대로 비료를 공급받지 않은 대조구에는 Festu c a rubra, Ag ro sti s ten uis , Leonto d on his p idu s 등 키 가 작고 총생 하며 가 지가 많은 포복성 혹은 로제트형의 식물이 우점하였다. 이들의 평균 키는 15-30cm 였댜 ParkGrassPlo t s 의 실험 결과는 부족한 토양 영양염류를 공급하면
전체 식물량을 증가시키고 지표면에서의 광량을 감소시키기 때문에 직립성이며, 키가 더 높이 자라는 종에게 유리하다는 것을 보여준 다. Snayd o n(1970), Sna y don 과 Davie s (1972), 그리고 Snay do n (19 76) 은 이러한 힘이 동일종의 개체 내에서 유전적 변이의 유사한 패턴 에도 유리하다는 것을 보여주었다. 비료를 많이 준 구에서 채집한 Anth o xa nth u m 개체군은 비료를 주지 않았거나 영양염류 공급량이 낮은 구에서 채집한 식물에 비해 한 개체당 지상부 생물량도 많고 키도 큰 쪽으로 진화되었다(그림 5.8 ) . 따라서 부족한 토양 영양염류 의 공급을 증가시키면 키가 큰 종이 우점하는 것과 똑같이 특정한 종내에서 유전적으로 키가 큰 개체에게 유리하였다. 이들 개체들은 키가 크고 개체당 지상부 생물량이 많았다. 비록 지하부 생물량은 보고되지 않았지만 , 이러한 사실은 영양염류 수준이 높고 현존량이 많은 서식처에서 120 년 동안 생장하면 비료를 공급하기 전에 살았 던 개체들에 비해 뿌리에는 분배량이 적고 잎과 줄기에 분배량이 많은 개체들이 유리하다는 것을 증명하는 것이다. 이 개체군에서 지역적인 토양 조건과 관련하여 진화의 역할에 관 한 또다른 검정으로 Sna ydon(1976) 은 이식 실험의 결과를 발표하였 다. 6 개의 서로 다른 Park Grass 실험구로부터 채집한 식물을 Rea ding에 있는 공동포장에 수년 동안 키운 다음 각각 원래의 실 험구와 그리고 원래의 실험구와는 다른 실험구에 이식하였다. 원 실험구에 다시 이식된 식물은 대조적인 상이한 실험구에 이식된 식물에 비해 생존율이 두 배였고, 생존한 식물의 건물생산도 원실 험구에서 50% 나 많았다. 이러한 사실은 식물이 자신들의 원실험 구의 조건에 더 적합하다는 것을 입증하는 것이다. 이것은 또한 자연 선택이 각 실험구에서 부적합한 식물을 제거하고 가장 적합 한 식물에 유리하게 작용한다는 것을 의미한다 (Sna y don 1976, pp. 16-17).
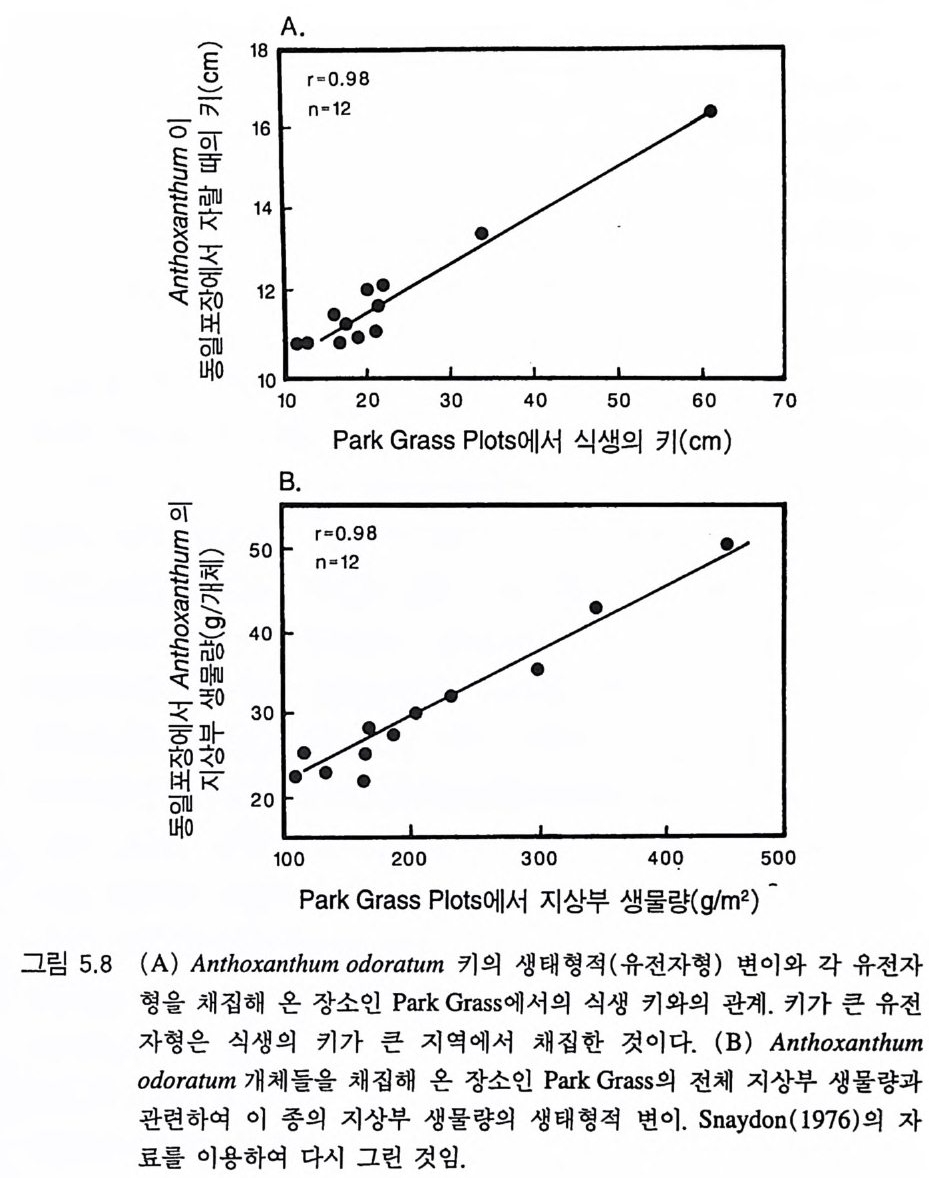 A.
A.
이러한 결과는 식물 군집의 구조에 제약을 주는 주요 환경변수들 은 4 가지의 다른 수준에서 작용한다는 것을 제시하는 것이다. 식물 개체의 수준에서 이러한 구배는 개체로 하여금 자신의 자원 요구
조건에 적응하도록 해주어 다양한 서식처에서 더 효율적으로 경쟁 하도록 하는 형태적 가소성이 있는 개체에게 유리하게 작용할 것이 댜 개체군 수준에서 이러한 구배는 서식처에 따라 상이한 유전자형 이 유리하게 할 것이다. 군집 수준에서 이러한 구배는 서식처에 따 라 일부 종이 다른 종보다 유리하게 할 것이다. 군계 수준에서 이러 한 힘은 초원, 관목림, 사바나, 그리고 수관이 밀폐된 삼림이 형성되 도록 한다. 모든 환경 요인이 이 4 가지의 모든 수준에서 비슷하게 작용하지는 않는다. 그러나 전세계에 있는 각양각색의 서식처에 형 성된 군집들 사이에서 볼 수 있는 유사성은 바로 이 4 가지 수준에 서 유사하게 작용하는 그러한 변수들이다. 1982 년에 미네소타의 Cedar Creek Natu r al H i s t or y에 있는 4 개의 상이한 천이 단계에서 일단의 질소 구배 반복구가 만들어졌다. 1986 년 이 되자 대부분의 보편종은 이 실험 구배상에서 뚜렷하게 구분되었 는데, 질소가 적은 곳에는 키가 작고 로제트롤 형성하는 초본식물이 우점하고 질소가 많은 곳에는 일부 목본종을 포함하여 키가 크고 직립성인 초본이 우점하였다 (T i lman 1987a). 이 실험의 내용은 8 장에 설명되어 있다. 8 식물이 갖는 특성간의 상관 관계 우점종 식물의 서식처와 관련된 모든 형태적, 생리적, 그리고 생활사적인 특성을 측정하고, 이 특성들을 토양 자원:광 구배상에 서 각 종이 최대 수도에 이르는 위치와 관련시킬 수 있는 연구는 아직 없는 것으로 알고 있다. 그러나 상이한 천이 단계에 있는 식 물의 종자에 대한 Sa li sb ury (1942) 의 연구는 G ri me 과 Hun t(1 975) 의 최대 상대생장률, 그리고 Claph a m, Toti n, Warbur g (1962) 가 보고한
생활사, 식물의 키. 그리고 서식처 천화성과 그 내용이 유사하다 . 이 러한 분석은 다년생 식물에 비해 1 년생 식물은 RGR 이 높고 (1.7: 1. 1/ wk), 종자의 크기가 작고 (0.00l6 : 0.93 g /seed), 성숙하였을 때 의 키가 작다 (38.5:339cm). 목본성 다년생 식물은 모든 일년생이나 다년생 식물에 비해 평균적으로 종자가 크고 (0 .3 2 g /seed), 키가 크며 (1, 200cm), 생장률이 가장 느렸다 (0.60/wk). 이러한 패턴은 제 4 장에 서의 예측과 일치한다. 이밖에 이러한 종들의 서식처에 대한 Sal i sbu ry의 구분은 Clap ha m 등 (1962) 이 한 구분과 함께 성숙하였을 때의 식물의 키와 최대 생장률이 자연 상태에서 그 종이 출현하는 서식처에 얼마나 의존적인가를 결정하는 데 이용될 수 있음을 보여 준다(그림 5.9). Sal i sbu ry에 의한 서식처 구분이 그가 주장하는 것처럼 천이의 단 계나 지표면에서의 햇빛의 이용도를 나타내는 것이라고 가정하면 식물의 키와 종자의 크기는 햇빛의 이용도가 감소함에 따라 증가하 고 최대 생장률은 감소한다. 그림 5.9 의 구배가 나타내는 생산력 구배 대 손실률 구배의 범 위에 대하여 나는 알지 못한다 . 나는 그것을 천이 초기 단계 서식 지의 심하게 교란된 상태로부터 천이 후기 단계 서식지의 교란이 적은 상태로 진행되는 손실률 구배로 설명하고 싶다. 이러한 나의 견해는 Tansley (19 49), Godwi n(19 56), G ri me (1 979) 의 주장에 기초 를 두고 있다. 만약 나의 설명이 맞다면, 그림 5.9B 는 그림 5.1 에서 예측한 바와 같이 RGRmax 가 손실률에 따라 증가함을 보여준다. 모든 구분을 하나로 통합하면 몇 가지의 재미있는 패턴이 있음을 알 수 있다. 종자의 무게와 성숙하였을 때의 키는 상관 관계가 매우 높다(그림 5.I OA ). 최대 상대 생장률은 종자의 무게와 음의 상관 관 계가 있다(그림 5.I OB ). RGRmax 는 성숙하였을 때의 키와 음의 상관 이 있지만 그 관계가 크게 유의하지는 않는다(그림 5.I O C). 이들 자
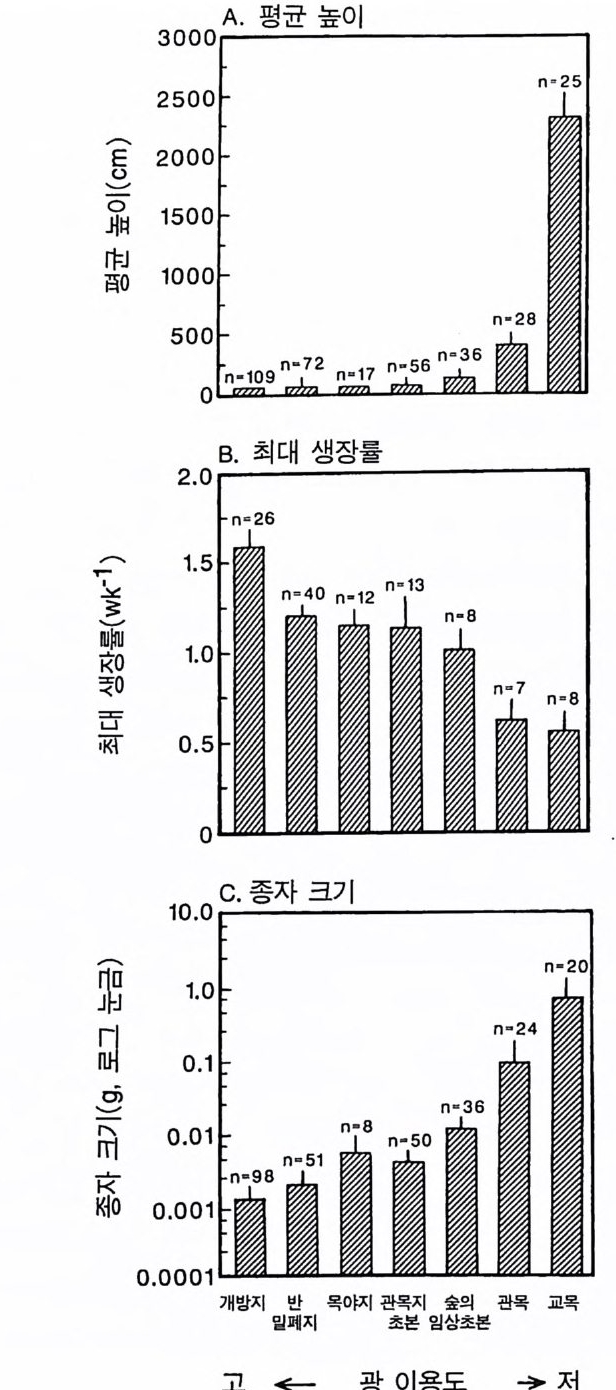 A. 평균 높이
A. 평균 높이
5.9 (A) 각 종의 수도가 가장 높은 서식처 유형에 따른 식물의 키의 의존도 (Cla p ham et al. 1962). (B) 서식처 유형에 따른 최대 상대 생장률 (G ri me & Hunt 1975). (C) 각 종의 수도가 가장 높은 서식처 유형에 따른 종자 크기의 의존도 (S ali sb ury 1942). 서
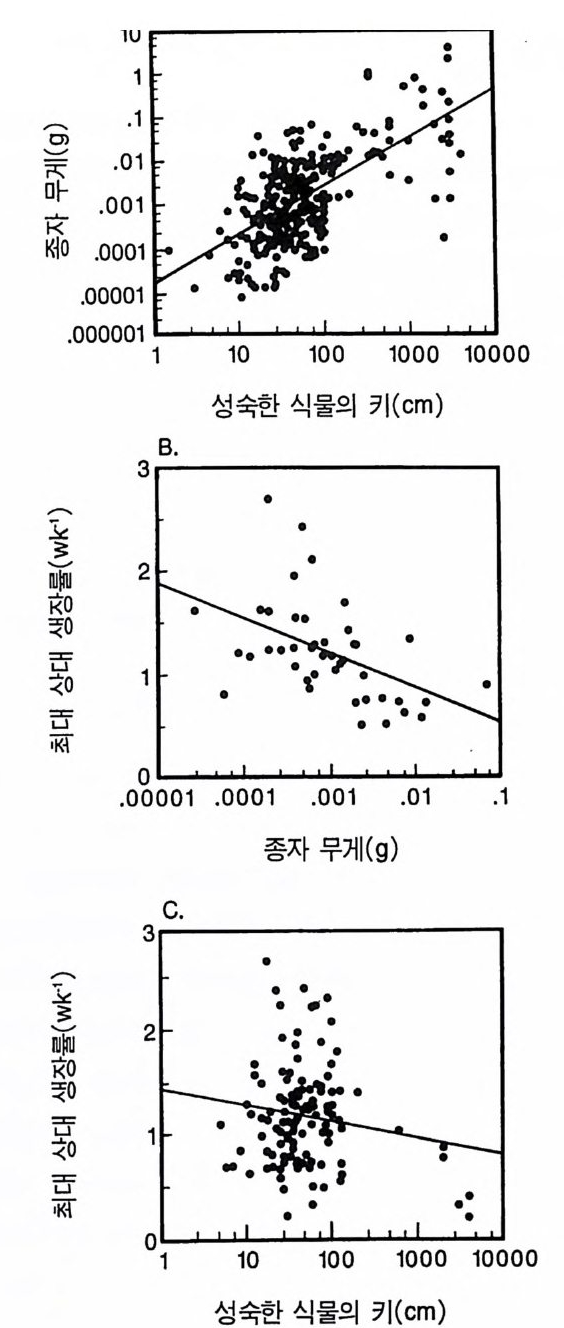 lU ••
lU ••
그림 5.10 (A) 종자당 무게의 대수값과 성숙시 식물의 키의 대수값은 유의한 상관관계가 있다 (r=0.6 4 , n=274, P
료에는 많은 변이가 있지만 이러한 상관 관계는 제 4 장에서 예측한 것과 똑같이 성숙하였을 때의 키가 더 큰 식물이 키가 작은 식물에 비해 종자가 크고 최대 생장률이 낮은 경향이 있음을 보여주고 있다. 9 형태적 가소성 식물의 키, 모양, 처음 생식시의 나이, 가지가 뻗은 모양, 그밖의 다른 형태적인 특징은 그들이 자라는 서식처의 환경 조건에 매우 의존적이다. H arp er(1977) 가 설명한 바와 같이 이러한 형태적인 차 이는 잎, 꽃 등 여러 가지 식물 단위의 형태적인 변화가 아닌 수의 변화에서 비롯된다. 토양의 영양염류가 풍부하고 밀도가 낮은 상태 에서 자라는 식물은 잎이 다층으로 된 수관을 갖고 키가 작은 덤불 모양을 한다. 반대로 비옥한 토양에서 밀도가 높은 상태로 자라는 식물은 키가 크고 잎이 대부분 식물 윗부분에 붙어 있다 (Horn 1971). 이러한 이유 중의 하나는 밀도가 높은 상태에서 자라는 식물 의 아래에 있는 잎은 광합성량이 호흡량을 능가할 정도로 충분한 광을 받지 못하여 떨어진다 (H arp er 1977). 따라서 햇빛의 수직적 감 소가 크고 밀도가 높은 조건에서 살고 있는 식물은 키와 관련된 구 조적인 조직에 더 많은 분배를 한다. 이 외에도 밀도가 높고 광이 제한되는 조건은 제 4 장에서 예측한 것과 똑같이 생식력의 감소, 생 장률의 감소, 성숙과 생식 지연 등을 초래한다 (H arp er 1977, p.x vi) . 대부분의 식물에서 개체 전체의 형태적인 변화에 비해 잎은 비교 적 안정된 형태적 단위이긴 하지만 잎의 광합성 능력은 햇빛에 노 출되는 정도에 따라 다르다. 우리가 양엽이라고 부르는 완전광을 받 는 잎은 음지에 있는 잎에 비해 광포화점이 높은 경향이 있다. 그러 나 음엽은 낮은 광도에서 광합성률이 최대에 이른다 (Moone y 1972,
Harp e r 1977, Lassoie et al. 1985 ) . 유전적으로 동일한 식물 개체들 내에서 볼 수 있는 이러한 모든 형태적, 생리적 가소성의 패턴은 이러한 가소성이 햇빛과 부족한 토 양 자원을 얻기 위한 경쟁의 한 가지 적응이라는 가설과 일치한다. 식물들 사이의 상호 작용을 예측하는 데 주요 문제 중의 하나는 식 물의 형태적인 변이가 커서 그들의 경쟁 능력을 명확히 측정하기가 어렵다는 것이다. 실제로 부족되는 여러 가지 자원의 획득률이 서식 처에 따라 다르듯이 식물의 경쟁 능력도 서식처에 따라 다르다. 그 러나 원인이 무엇이든 간에 개체들 사이의 변이 범위는 한 종의 다 른 구성원들 사이의 변이폭에 비해 좁고, 한 종 내의 변이 범위는 종 간의 변이 범위에 비해 훨씬 좁다. 10 요약 전체적으로 이 종설은 식물의 생활사와 형태적 특징의 많은 것들 이 토양 자원:광의 구배 혹은 손실률 구배상에서 그 종이 최대 수 도를 얻는 위치에 따라 다르다는 것을 보여주고 있다. 앞에서 설명 한 다양한 생산력 구배의 경우, 그것이 질소 공급률이 낮은 서식처 에서 높은 서식처로의 구배이거나 고도가 높은 곳에서 낮은 곳으로 의 구배 혹은 고도가 낮은 곳에서 높은 곳으로의 구배, 또는 강우량 이 적은 곳에서 많은 곳으로의 구배이든 간에 우점종의 형태와 생 활사의 변화가 매우 비슷하다. 이와 같이 넓은 규모의 환경 구배상 에서 한 종으로부터 다음 종으로 나타나는 같은 유형의 변화가 규 모는 작지만 각 종 내에서 유전적인 변이로 나타난다. 또한 각각의 식물 개체는 형태적으로 그리고 생리적으로 가소성을 갖는데. 이러 한 가소성은 그러한 구배상에서 종내 그리고 종간의 유전적인 변이
에서 볼 수 있는 것과 같은 바슷한 변이 패턴을 갖는다 . 따라서 키, 뿌리나 잎 그리고 줄기로의 분배 패턴, 종자의 크기, 생장률, 광합성 효율 둥과 같은 다양한 식물의 특성을 결정하는 것은 토양 자원과 광의 이용도를 결정하는 토양 자원 공급률과 손실률이 주축이 되는 것으로 보인다. 토양 자원 공급률과 손실률에 따라 동일종 내의 유 전적 분화인 종 분화와, 전세계적으로 기후와 토양이 비슷한 서식처 에 살고 있는 계통학적으로 먼 식물들 사이의 수렴진화가 나타난다. 이와 같이 규모가 큰 다양한 구배상에서 볼 수 있는 식생의 유사성 은 서식처 내의 토양 자원과 광의 평균 이용도, 그리고 광에 대한 토양 자원의 획득을 위해 식물이 가지고 있는 대응책이 식생 패턴 울 결정하는 주요 인자임을 제시하는 것이다.
제 6 장 식물 경쟁의 동태 1 서론 지금까지는 주로 식물 경쟁의 장기적인 결과, 즉 경쟁적 상호 관 계에 있는 여러 종들이 평형 상태에 도달하였을 때의 결과에 대해 서만 설명하였댜 예를 들면, 특정한 환경 조건하에서 경쟁하고 있 는 두 종을 설명하면서 나는 평형 상태에서 한 종이 다른 종을 완 전히 교체할 때 그것을 경쟁 우세종이라고 표현하였다. 나는 경쟁의 장기적인 결과를 강조하였는데, 그 이유는 인간에 의한 교란이 없는 자연 식물 군집에서 볼 수 있는 지리적인 분포 패턴과 지역적인 수 도 패턴에 가장 중요한 영향을 줄 수 있는 것은 바로 경쟁의 장기 적인 결과이기 때문이다.
과거에 인간에 의한 교란이 없었던 군집들은 현존하는 식물종이 진화해 온 조건을 알아내기 위한 가장 좋은 참고 자료가 되고, 또 이러한 군집들은 35 억 년의 진화의 결과 풍부한 유전적 다양성을지니고 있기 때문에 자원의 보고라고 할 수 있다. 그러나 생태학자 들의 생각은 그들이 자주 접하는 서식처에 너무 편중되어 있다. 인 간에 의한 교란이 없는 자연 식물 군집은 지구상에서 점점 사라지 고 있고. 나머지는 임학, 농경. 방목 , 기타 인위적인 교란에 의해 심 하게 변형되어 있댜 더구나 모든 생태 실험은 필연적으로 자연을 교란시키게 된댜 따라서 생태학자들이 관찰할 수 있는 것은 대부분 교란 후에 특정한 종이 나타내는 단기간의 매우 동적인 반응들이다. 이들 종들이 과거에 거쳐온 진화의 과정에 중요하게 작용하였던 요 인들을 알아내기 위해서는 이러한 단기간 동안에 관찰되는 동적인 반응들을 이용하여 추론하여야 한다. 그러나 이러한 추론은 상당히 위험하댜 그 이유는 실험적인 조작에 대한 즉각적인 반응이 장기간 의 효과와는 무관한 일시적안 동태일 수 있기 때문이다. 이 장의 목적은 최근에 교란된 군집에서 우리가 볼 수 있는 패턴 과 인간의 간섭이 전혀 없었던 자연 군집에서 볼 수 있는 패턴 사 이의 유사성과 차이점을 찾아내는 일이다 . 이 장에서 전개시킨 주제 중의 하나는 특정한 교란 혹은 실험적인 조작에 대한 즉각적이고 동적인 반응은 그것의 장기적인 효과와는 현저히 다를 수 있다는 것이댜 따라서 최근에 교란된 생태계에서 가장 중요하게 여겨지는 패턴과 과정들이 교란되지 않은 생태계에서는 별로 중요하지 않을 수 있다. 여러 가지 교란이나 실험적 조작에 대한 종의 죽각적이고 동적인 반응은 다른 선택적인 힘의 간접적인 효과일 수 있다. 경쟁에서 예측되는 결과가 무엇이 되어야 하는지를 결정하는 것 은 그것이 분석적이든 아니면 계산적이든 간에 특정한 모델의 범주 내에서는 간단한 과정이다. 그러나 특정한 자연 환경에서 실제로 경 쟁력이 가장 강한 종이 어떤 것인지를 결정하는 가장 좋은 실험 혹 은 관찰 방법이 무엇인지는 쉽게 알 수 없다. 식물성 플랭크톤과 같 이 수명이 짧은 생물은 10 세대 혹은 20 세대 동안 경쟁이 계속되도
록 하여 그 결과를 분명히 알 수 있다 . 그러나 많은 육상식물의 수 명은 연구자가 연구비를 받는 기간이나 과학자들의 관심이 지속되 는 기간 혹은 과학자들의 수명보다 훨씬 길다. 따라서 육상식물의 경우 경쟁에 관한 대부분의 연구는 단기간의 실험이나 관찰 자료를 이용하여 장기간의 경쟁 결과를 추정할 수 있도록 고안된 여러 가지 손쉬운 방법들이 이용된다. 이러한 손쉬운 방법들, 특히 de W it의 교체계열에 기초를 둔 방법들은 내가 설명한 것과는 경쟁 능력에 대한 의미가 다르다. 이러한 방법들은 상대밀도 가 서로 다른 상태에서 자라는 두 종의 단기간의 상대적인 생산성 (혼히 지상부 생물량만을 사용함)을 경쟁 능력과 같은 의미로 사용하 고 있다. 또다른 방법은 여러 종이 혼합된 상태에서 어떤 종이 다른 종들에 비해 빨리 자라면 그 종을 경쟁 우세종이라고 부르는 것이 다. 따라서 만약 종 A, B, C 를 포장에 심어 놓았는데 관찰기간 동안 에 A 종의 수도가 B 종이나 C 종에 비해 증가하는 것으로 관찰되었다 면 그 포장의 조건에서는 A 종이 경쟁 우세종이라고 불릴 것이다. 이 와는 대조적으로 나는 어떤 종이 그 서식처에서 다른 종을 경쟁적 으로 교체할 때 그 종을 경쟁 우세종이라고 부른다. 이러한 용어상의 차이는 몇 가지 중요한 문제를 야기한다. 경쟁적 상호 작용의 단기적인 동태를 장기간의 경쟁 결과를 예측하는 데 이용할 수 있는가? 경쟁의 초기에 상대적 생산성이 높은 종이 결국 다른 종을 경쟁적으로 교체할 수 있는가? 경쟁의 동태에 관하여 예 측되는 패턴이 있는가? 2 경쟁에 관한 간단한 모델 먼저 크기 구조나 연령 구조가 없는 이상적인 3 종이 부족한 단일
자원에 대하여 경쟁하는 경우를 생각해 보자(그림 6.1) . 모든 종이 같은 손실률인 D1 을 갖으면, A 종의 R 가’ 가 가장 낮아 그 자원에 대하 여 경쟁 우세종이다. 그러나 B 종과 C 종에 비하여 A 종은 최대 생장 률이 낮기 때문에 B 종과 C 종이 A 종에 비해 초기에 더 빨리 성장할 것이댜 따라서 이 3 종 사이의 경쟁 동태는 C 종에 의해 우점하는 기간이 있고 그 다움에는 B 종에 의해 우점하는 기간이 있으며, A 종 이 결국 B 종과 C 종을 교체하게 될 것이다 . A 종이 B 종과 C 종을 교체하는 데 필요한 시간은 초기의 밀도 , 최 대 생장률, 부족한 자원의 소비율, 초기의 자원 이용도, 그리고 자원 공급의 동태에 따라 다르다. 예를 들어, 모든 토양 자원이 과량으로 존재하고 햇빛만 부족한 상황을 생각해 보자. C 종이 1 년생 초본이 고 B 종은 벼과의 다년생 식물이며, A 종은 교목이라면 생장이 훨씬 느린 교목이 1 년생이나 다년생 초본이 햇빛이 부족하여 완전히 제 거될 정도로 밀폐된 수관층을 형성하는 데에는 많은 시간이 소요될 것이다 . 아쉽게도 대부분의 관찰이나 실험 연구의 기간은 매우 짧아 B 종이나 C 종과 같이 생장이 빠른 식물이 일시적으로 우점하는 기간 만 볼 수 있고 마지막 우점종이 우점하는 것은 볼 수 없다. C 종에 의한 일시적인 우점은 그것의 최대 생장률에 의해 비롯된 댜 어느 환경 조건이나 실험적인 조작이 그곳에서 자원의 이용도를 증가시키면 C 종이 초기 우점종이 될 것이다. 부족한 자원의 소비로 인해 자원이 낮은 수준으로 유지되는 것이기 때문에 하나 혹은 그 이상의 우점종을 제거하는 것처럼 자원의 수준을 높일 수 있는 조 작은 최대 생장률이 높은 C 종이나 다른 종이 일시적으로 우점하도 록 한다. 손실률 D2 에서는 C 종이 경쟁 우세종이 되는데, 높은 손실 률에 의해 자원의 수준이 높게 유지될 수 있다. 교란율 D2 에서는 경 쟁력이 낮은 종에 의한 일시적인 우점 기간이 없이 C 종이 우점종이 될 것이다. 따라서 실험적 조작 혹은 교란 후의 개체군 동태는 후에
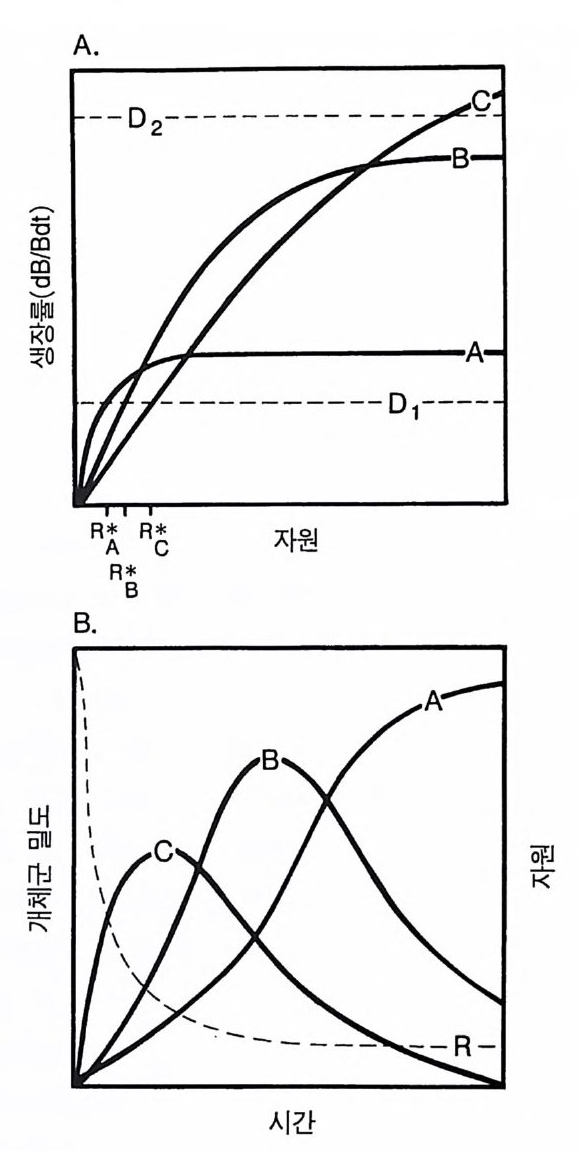 A. c
A. c
그림 6.1 (A) A, B 그리고 C 로 표시된 곡선들은 A 종, B 종 , 그리고 C 종의 상대 생 장률 (dB/Bd t , B 는 식물의 생물량)이 자원의 이용도 R 에 대한 의존도를 보여주는 것이다 . D, 과 D 2 는 두 개의 상이한 손실률이다. 손실률 D, 을 갖 는 환경에서 자랄 때 3 종의 평형적 자원 요구량은 그들의 R * 에 의해 표 시된다. (B) 손실률 D, 에서 이들 3 종 사이의 경쟁 동태.
교체되는 종에 의한 일시적인 우점 기간을 포함할 수 있고. 그렇지 않을 수도 있다. 따라서 일시적인 동태는 이러한 실험의 결과가 시 간 규모에 의존적임을 의미한다. 짧은 기간 동안에 수행된 반복구가 많은 야외 실험을 통해 통계적으로 유의성이 매우 높은 경쟁 결과 를 얻을 수 있지만 이 실험은 부족한 자원에 대한 경쟁 능력보다는 최대 생장률을 측정한 결과에 불과하다. 3 크기 구조를갖는모델 평형 상태에 이르기 훨씬 전에 시작되는 경쟁적 상호 작용에서 최대 생장률은 어떤 종이 초기에 우점하게 될 것인지를 결정하는 주 요인이 된댜 최대 생장률은 새로운 광합성 기관으로 분배되는 광합성 산물의 비율에 따라 증가한다. 육상식물처럼 크기 구조를 갖 는 개체군의 경우 식물이 햇빛에 대해 경쟁할 때 개체의 키가 증가 되는 비율은 경쟁에 매우 중요하다. 초기 생장 기간 동안 키가 더 빨리 자라는 개체는 입사되는 햇빛을 더 많이 받을 수 있어 다른 개체들의 생장을 억제한다. 초기에 키가 크게 된 식물이 성숙하였을 때의 키가 작다면 초기 경쟁 단계에서 일시적인 우점 기간이 나타 난다. 식물 경쟁의 동태에 영향을 줄 수 있는 다른 요인으로는 자원의 동태를 들 수 있다. 특히 크기 구조를 갖는 개체군의 경우 토양의 영양염류와 햇빛은 단순하게 그들의 평형 상태의 수준으로 감소되 지는 않을 것이다. 이것은 출발 조건에 따라 자원의 수준이 처음에 는 이 종에 유리하고 다음에는 다른 종에게 또 그 다음에는 다른 종에 유리하게 되는 등의 복잡한 괘적을 따라 자원의 수준이 평형 상태에 이를 때까지 움직일 수 있다는 것을 . 의미한다. 자원의 수준
이 결코 평형 상태에 도달하지 않는다면, 하나 이상의 식물종이 지 속적인 변동을 하거나, 아니면 처음에는 이 종에게 그 다음에는 다 음 종에게 유리한 무질서한 동태를 지속할 수 있다. 나는 크기 구조를 갖는 개체군에서 자원 경쟁에 관한 모의를 통 해 이러한 모든 동적안 패턴을 보았다. 이 다음 절에서는 이러한 모 델에서 볼 수 있는 몇 가지 유형의 동적인 반응을 제시하려고 한다. 이것들은 크기 구조를 갖는 개체군에서 볼 수 있는 경쟁의 동태에 관한 몇 가지 예가 된다. 내가 이러한 예를 제시하는 것은 식물 경 쟁의 초기 동태와 이러한 경쟁적 상호 작용의 궁극적인 결과 사이 에는 아무런 관계도 없다는 것을 강조하기 위한 것이다. 또한 이것 들은 단기간(휴경지의 경우 1-10 년, 삼림이나 숲톰의 경우 1-25 년)의 동적인 반응을 설명해 주기 때문이다. 다음 장에서도 설명하겠지만 이러한 모델들은 분명히 천이 동태에 관한 모델이다. 3-1 다종 경쟁 100 개의 서로 다른 종이 부족한 토양 자원과 햇빛에 대하여 경쟁 하는 몇 가지 경우를 ALLOCATE 모델을 이용하여 알아 보자. 100 종은 모두 뿌리나 줄기, 잎으로의 분배 패턴을 제외하고는 모든 것 이 동일하댜 이 100 종은 식물에서 볼 수 있는 모든 분배 패턴이 포 함되도록 선별되었다. 모든 경우에서 손실률은 0.2/wk 였다. 영양엽류가 빈약한 경우를 생각해 보자 (TN=100). 경쟁하고 있는 100 종 중 대부분은 결코 우점종이 되지 못한다. 실제로 첫해에 8 종 이 전체 식물량의 55% 를 차지하였으며. 10 년이 지난 후에는 92% 를 차지하였다. 처음 10 년 동안의 평균을 보면 이 8 종이 전체 식물량의 84% 를 차지하였다. 이 8 종의 동태와 부족한 토양 영양염류 및 지표 면에 이입되는 햇빛의 동태가 그림 6.2 에 나와 있다.
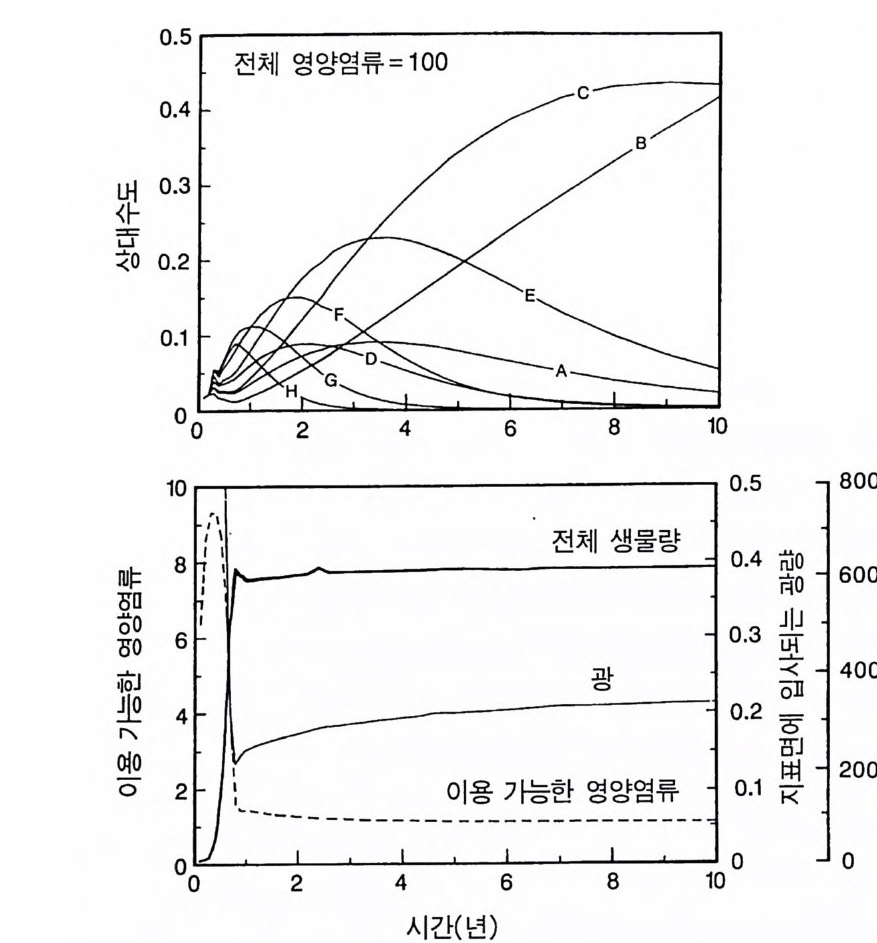 0.5
0.5
그림 6.2 형태만 다르고 다른 특성은 동일한 100 종이 손실률이 0.2/wk 이고 영양염 류가 빈약한 토양 (TN=lOO) 에서 경쟁하는 경우의 개체군 동태 . 여기서는 100 종 중 8 종만 나와 있다 . A 에서 H 까지 표시된 이들 8 종은 각각 다음과 같은 잎, 줄기. 뿌리로의 분배 를 가지고 있다. A 종 : 0.55, 0.05, 0.4 0 , B 종: 0.5 0 , 0.025, 0.4 7 5, C 종: 0.55, 0.025, 0.4 2 5, D 종 : 0.60, 0.0 5 , 0.3 5 , E 종: 0.60, 0.025, 0.375, F 종 : 0.65 , 0.0 2 5, 0.325, G 종: 0.70, 0.0 2 5, 0.275, H 종: 0.7 5 , 0.025, 0.225. 이들 8 종만이 이러한 모의 과정의 어떤 시점에서 우점 종의 위치를 차지한다 . 40 년 동안을 모의하였지만 초기의 동태 를 좀더 자 세히 보여주기 위해 처음 10 년 동안의 결과만 나타냈다. 40 년째에는 B 종 이 다른 모든 종을 교체하였다 .
경쟁 과정을 거쳐 최종 우점종이 되는 B 종은 오랜 기간에 걸쳐 서서히 절대수도를 중가시켜 생물량의 평형에 접근한다. 그러나 이 종의 상대수도는 그것이 증가를 시작하기 전에 수년 동안 감소한다. 이러한 경쟁적 상호 작용 과정 동안에 일시적으로 우점 기간을 갖 다가 경쟁적으로 교체되는 종이 5 종 있었다. 이와 같이 영양염류가 빈약한 서식처에서 경쟁의 동태는 상당히 복잡하고, 이기는 종이 우 점되기까지는 상당한 기간이 필요하다. 대부분의 경쟁적 교체는 전체 식물량(지상부 생물량과 지하부 생물 량의 합)이 일정하게 되는 기간에 일어났다. 초기에 지표면에 이입 되는 햇빛의 비율은 0.13 정도였으나 생물량의 상당 부분이 지하부 에 분배됨에 따라 약 0.22 로 서서히 증가하였다. 이 과정에서 이용 가능한 영양염류의 수준은 감소하였다. 이러한 상호 작용이 있는 동안의 각 시기에는 식물의 어떤 형태 적인 특징이 유리한가? 이것을 알아보기 위한 쉬운 방법은 상호 작 용이 있는 동안의 각 시기에 우점하는 종의 분배 패턴을 조사하는 것이다(그림 6.3 A ). 이 100 종 중에서 종 H 같이 가장 일찍 최대 수도 에 이르는 종은 잎으로의 분배량이 많아 상대 생장률이 큰 것이다. 이러한 종들은 점차적으로 잎에 분배되는 양이 적고 뿌리에 분배되 는 양이 많은 종에 의해 교체된다(그림 6.3 A ). 이러한 과정은 B 종이 우점할 때까지 계속된다. 언젠가 한번은 우점하게 되는 8 종은 모두 줄기로의 분배량이 낮 았다(실제로 이 모의에서 사용된 종 중 줄기로의 분배량이 가장 낮은 것 은 2.5% 였다). 따라서 이 종들은 모두 키가 매우 서서히 증가하는 로제트형의 식물이다. 이러한 모의는 정성적으로 그림 6.1 의 결과와 비슷한 결과를 낳는다. 그러나 이들 사이에는 중요한 차이점이 있 다. 절대 우점종이 되지 못하는 종 H 에 비해 최대 생장률이 큰 종 이 많이 있다. 이러한 종은 뿌리로의 분배량이 너무 적어 처음부터
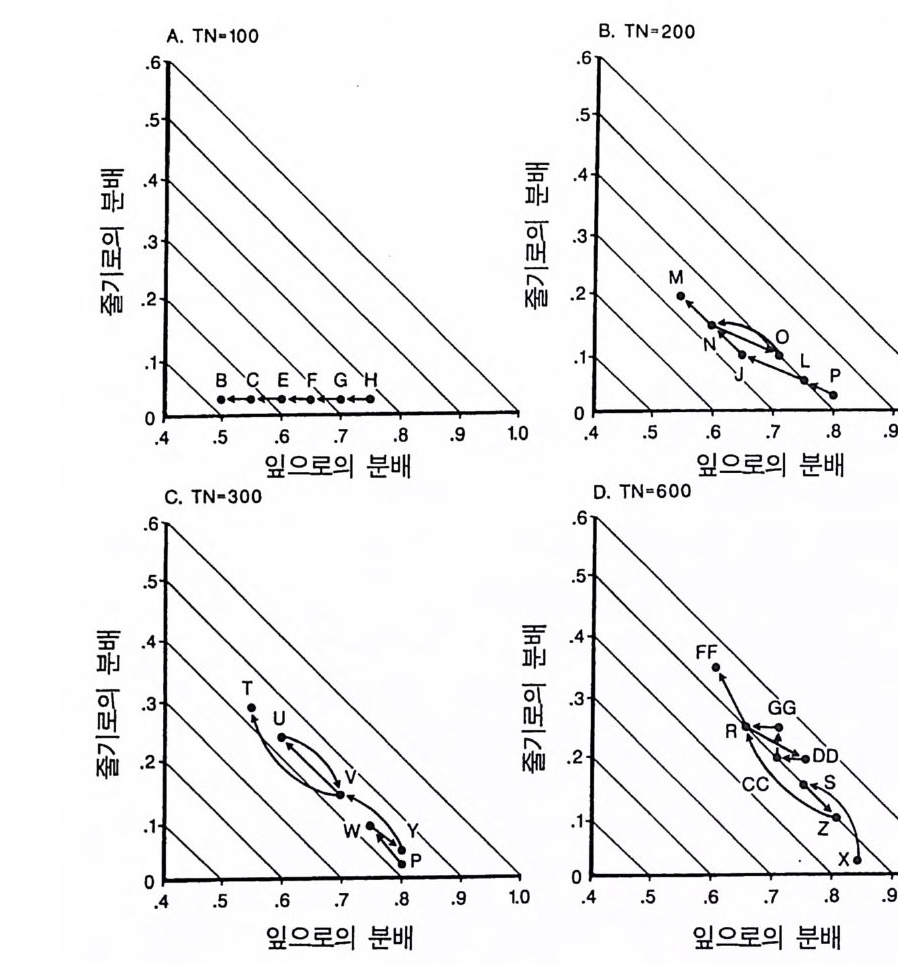 A. TN•100 B. TN• 2 00
A. TN•100 B. TN• 2 00
그림 6.3 (A) 경쟁적 교체 과정 동안에 우접종의 분배 패턴의 괘적 . 각 점은 특정 한 시기에 우점종의 분배 패턴을 보여준다. 각 점에 있는 글자는 그 종의 이름이다. 화살표는 TN= lOO 인 토양에서 경쟁적 교체 과정 중에 우접종 분배 패턴의 순서를 나타내는 것이다. (B) TN=200 에서 우점종 분배 패 턴의 괘적. (C) TN=300 에서 우접종 분배 패턴의 괘적 . (D) TN=600 에 서 우점종 분배 패턴의 괘적 . 경우에 따라서 같은 종이 한 번 이상 우접 종이 됨을 주목하기 바란다. 이러한 괘적들은 그림 6.2 , 6.4 , 6.5 , 그리고 6.6 에 나와 있는 개체군 동태와 일치한다. 4 가지 경우 모두에서 동일한 100 종이 경쟁하였고, 이들 100 종은 분배 패턴을 제외하고 모든 것이 동일 하였다. 모든 결과는 ALLOCATE 를 사용한 모의에 근거를 둔 것이다. 축 의 범위가 0-1 이 아닌 것을 주목하기 바란다.
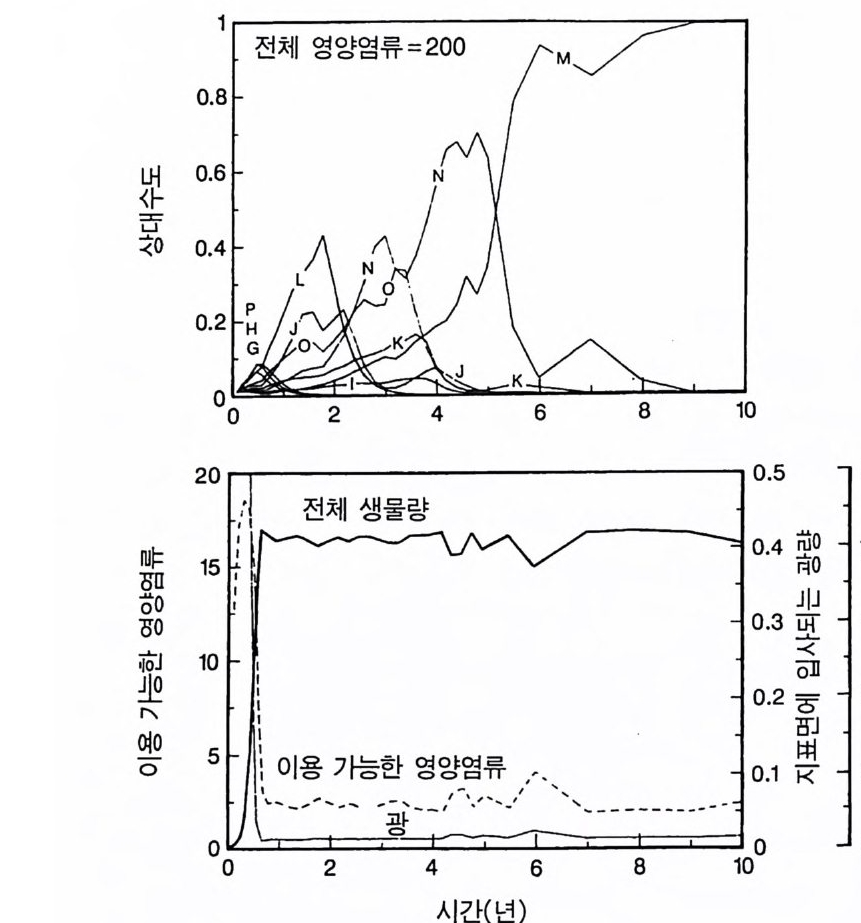 0.8 I 전체 영양염류 =200
0.8 I 전체 영양염류 =200
그립 6.4 토양의 영양염 류 공급 률 만 더 높고 (TN=200) 나머지는 그림 6 . 2 에 서 사 용한 것과 갇은 100 종의 식물의 경쟁 적 교체 동 태 . 언 젠 가는 우 점종 이 되 는 10 종만 나와 있으며. 이들 10 종의 잎 줄기 , 뿌리로의 분 배 패턴은 각 각 다음과 같다. G 종 : 0.7 0 , 0.0 2 5, 0.2 7 5, H 종 : 0.75, 0.0 2 5, 0.2 2 5, I 종: 0.6 0 , 0.2 0 , 0.2 0 , J종 : 0.6 5 , 0.10, 0.25, K 종: 0.6 5 , 0.1 5 , 0.2 0 , L 종 : 0.7 5 . 0.0 5 , 0.2 0 , M종 : 0.5 5 , 0.20, 0.2 5 , N 종 : 0.6 0 , 0.1 5 , 0.2 5 , 0 종 : 0.7 0 , 0.10, 0.20, P 종: 0.80, 0.025, 0.1 7 5.
는 형태로부터 광량이 적은 서식처에서 가장 빨리 자라는 형태로 식물이 점진적으로 교체되는 것이다 . 이 100 종의 식물이 좀더 비옥한 서식처(손실 =0.2. TN=300) 에서 경쟁할 때. 마지막 우점종인 T 의 절대수도와 상대수도는 이 값들이 증가하기 전 4 년 동안 감소하였다(그림 6.5 ) . 이 종의 절대수도는 자 신이 우점종이 되었을 때 진동이 계속되었다. 종 T 가 우점종이 되기 전에 다른 종에 의한 일시적인 우점 기간이 6 번이나 있었다. 대부분 의 종은 경쟁하는 동안에 변동적인 반응을 보였으며, U 종과 V 종은 각각 2 년 간격으로 2-3 개의 뚜렷한 피크를 보였다. 전체 식물량. 이용 가능한 영양염류, 그리고 지표면에 입사되는 광량도 변동하였다. 이러한 상호 작용이 있는 동안 각 시기에 우점 하는 종의 형태는(그림 6.3 C ). 잎으로의 분배량이 높은 종이 초기에 우점하고 그 다음에는 잎으로의 분배량은 적고 줄기로의 분배량이 많은 종이 우점하는 일반적인 경향을 보였다. 이 과정 동안에 뿌리 로의 분배량은 상당히 일정하였다. 그러나 이미 설명한 바와 같이 일부 종은 교체 기간 동안에 한 번 이상의 우점 기간이 있었는데, 그 결과 식물 형태에서 변동이 나타난다. 마지막 경우로 이 100 종의 식물을 매우 비옥한 서식처 (TN=600) 에서 경쟁하도록 하였다. 마지막 우점종인 FF 의 절대 및 상대수도 는 처음 2 년 동안 감소한 후 느리고 불안정한 중가가 시작되었다 (그림 6.6). 최초의 우점종인 X 는 잎으로의 분배 량이 높고 (85%) 줄 기 로의 분배 량이 낮았다(그림 6.6. 그림 6.3D). X 종 다음에 일시 적 인 우점 기간을 갖는 종의 계열은 줄기로의 분배량이 증가하고 잎으로 의 분배량이 감소하는 종들이었다(그림 6.3D). 이들은 또한 최초의 우점종에 비하여 뿌리로의 분배량이 낮았다. 모든 종이 변동하는 경향을 보였는데, R 종은 두 시기에 최대 수 도를 보였다. 이러한 변동은 우점하는 식물 형태의 괘적이 본래 변
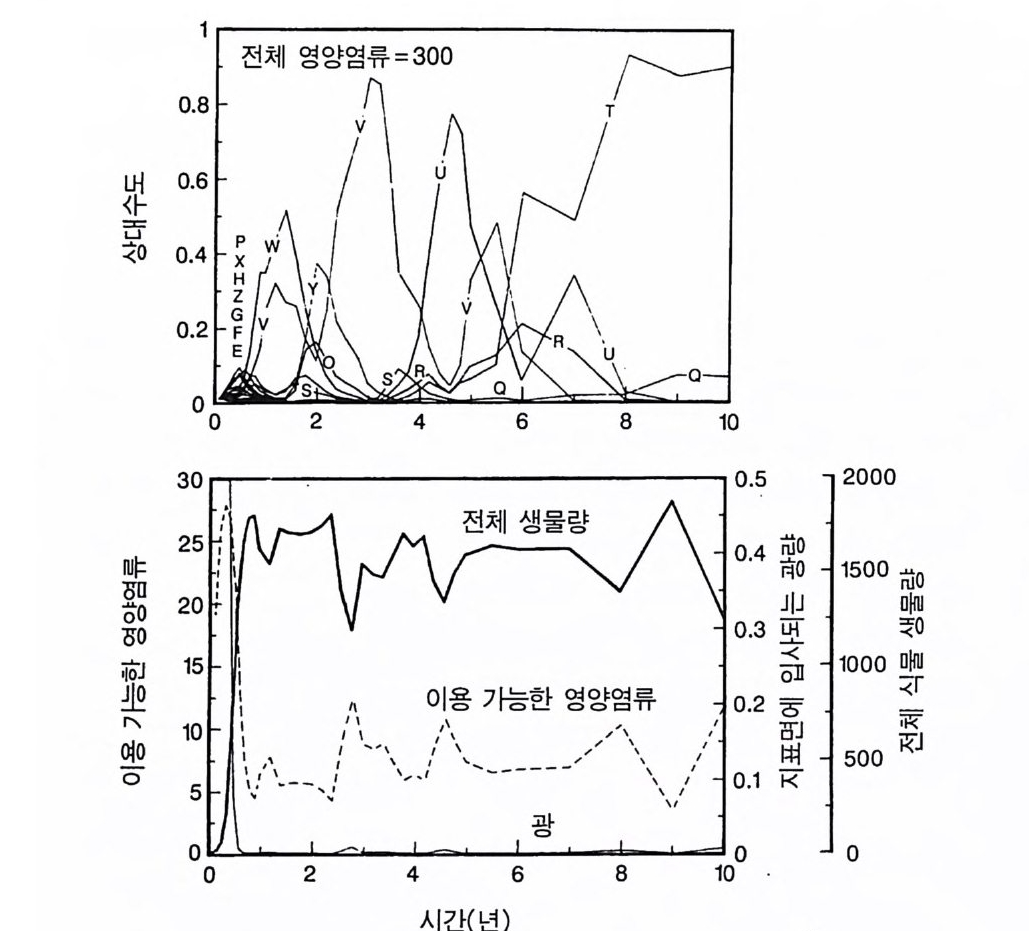 전체 영양염류 =300
전체 영양염류 =300
그림 6.5 토양의 영양염류 공급률만 더 높고 (TN=300) 나머지는 그립 6.2 , 6 .4에 서 사용된 것과 동일한 100 종의 경쟁적 교체 동태. 언젠가는 우점종이 되 는 16 종만 나와 있다. 이들 종의 잎, 줄기 . 뿌리로의 분배는 각각 다음과 같다 . Q종: 0.60, 0.30, 0.1 0 , R 종 : 0.65, 0.25, 0.10, S 종: 0.7 5 , 0.15, 0.1 0 , T 종 : 0.5 5 , 0.3 0 , 0.15, U 종 : 0.6 0 , 0.2 5 , 0.1 5 , V 종 :0 . 70, 0.15, 0.15, W 종: 0.75, 0.10, 0.1 5 , X 종: 0.85, 0.025, 0.125, Y종 : 0.80, 0.05, 0.1 5 , Z종 : 0.80, 0.10, 0.10. 다른 6 종의 분배 패턴은 앞의 그림 설명에 나와 있다.
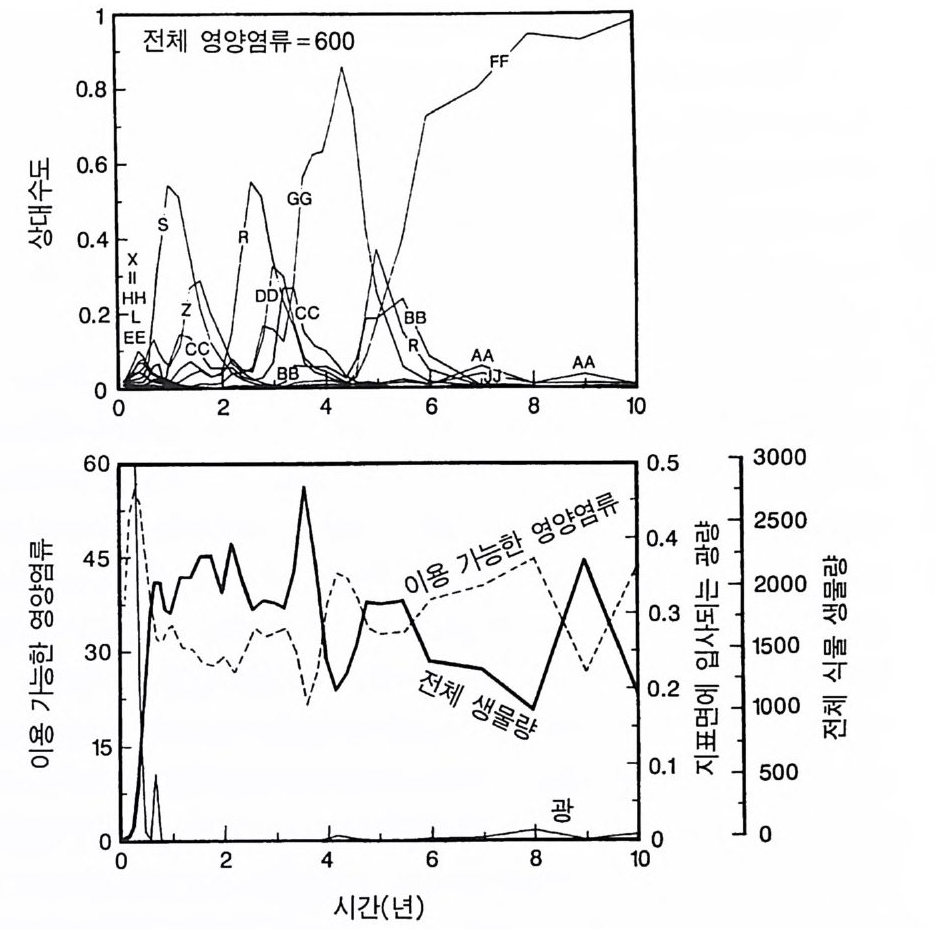 전체 영양염류 =600
전체 영양염류 =600
그립 6.6 토양의 영양염류 공급률만 더 높고 (1N=600) 다른 것은 그립 6.2. 6.4. 6 .5에 나온 것과 동일한 100 종의 경쟁적 교체 동태. 언젠가 우점종이 되 는 15 종만 나타나 있다. AA- JJ까지 표시되어 있는 종의 잎 줄기 . 뿌리로 의 분배는 각각 다음과 같다. AA 종: 0.55, 0.4 0 , 0.05, BB 종: 0.65, 0.30, 0.05, cc 종: 0.70, 0.20, 0.1 0 , DD 종 : 0.7 5 , 0.20, 0.0 5 , EE 종 : 0.50, 0.4 5 : 0.05, FF 종: 0.60, 0.3 5 , 0.05, GG 종: 0.70, 0.25, 0.05, HH 종: 0.90, 0.025, O.Q 75 , I/ 종: 0.8 5 , 0.05, 0.10, JJ종: 0.65, 0.3 2 5, 0.025. 다른 종의 분배 패 턴은 앞의 그림 설명에 나와 있다.
동하는 성질이 있기 때문이다(그림 6.3D). 전체 생물량, 영양염류 이 용도, 그리고 투과되는 광량은 모의 과정 전체에 걸쳐 변동하였다. 앞의 두 경우에서처럼, 최종 우점종은 줄기로의 분배량이 많아 더 빨리 키를 증가시켜 햇빛에 대한 경쟁에서 다른 종을 능가하기 때 문에 다른 종을 교체한다. 이러한 성질은 그것이 최종 우점종으로 하여금 최초의 우점종보다 훨씬 낮은 최대 생장률을 갖도록 할 경 우에도 그 종에게 유리하다. 이러한 모든 모의는 특정한 환경조건에서 결국 우세한 경쟁자에 의해 교체되는 종들에 의한 장기간에 걸친 일시작인 우점이 있음을 보여주고 있다. 이 각각의 모의는 비교적 높은 종자 밀도로 시작되 었으나 전체 식물량은 평형 상태에서 얻는 수준보다는 훨씬 적은 상태에서 시작되었다. 따라서 각 모의 과정 초기에는 개체들이 햇빛 과 영양염류가 비교적 많은 상황에서 자라고 있었다. 이러한 조건은 줄기에 분배량이 적고 잎에 분배량이 많은 종에게 유리하다. 식물량의 중가로 영양염류의 이용도와 광량이 감소함에 따라 다 른 형태가 유리하게 되었다. 그러나 일시적인 우점 기간을 갖는 종 의 종류는 각 서식처에 따라 달랐다(그림 6.3). 평형 상태에서 손실 률 D=0.2/wk 를 갖는 서식처의 최종 우점종은 그림 4.5 의 곡선에 맞는 분배 패턴을 갖는다. 그림 6.3 은 그림 4.5 와 중요한 대조를 보 여주고 있다. 모든 가능한 분배 패턴 중에서 항상 일시적인 우점 기 간을 얻는 유일한 종은 최종 평형 상태의 우점종에 비해 잎으로의 분배량이 많은 종들이었다. 최종 우점종에 비해 잎으로의 분배량이 적은 종들이 많이 있다 (그림 4.4 참조) .• 그 러나 이 종들은 모든 모의에서 결코 일시적인 우 점 기간을 갖지 못하였다. 그림 6 . 3 의 괘적들은 최종 우점종보다 잎 으로의 분배량이 큰 거의 모든 식물 형태가 적당한 토양에서 언젠 가는 일시적인 우점 기간을 갖을 수 있음을 보여주는 것이다.
이 패턴과 그림 4.8 및 그림 4 . 9 에 나타나 있는 결과를 비교해 보 면 재미있는 사실을 알 수 있다 . 그림 4 . 8 과 4.9 는 평형 상태에서 각 각의 식물 형태에게 유리한 환경 조건이 있음을 보여주고 있다 . 그 림 6.3 은 상당히 넓은 범위의 식물 형태가 비평형 조건에서 유리함 울 보여주고 있다. 실제로 최근에 교란된 서식처에서 일시적인 우점 상태에 있는 종은 손실률이 높은 서식처에서 최종 평형 상태의 우 점종들이다 . 이 패턴은 한 단계 더 진행된다 . 주어진 토양형에서 (ALLOCATE 모델에서 주어진 TN 값) 일시적인 우점 기간을 갖는 종 (형태)의 계열은 그 토양형에서 손실률이 높은 상황에서 평형 상태 의 우점종이 되는 종으로부터 그 토양에서 점차 손실률이 낮아지는 상황에서 평형 상태의 우점종이 되는 종으로 진행되는 경향이 있다. 이러한 교체는 그 서식처의 실제 평형 상태의 손실률에 도달할 때 까지 계속된다. 손실률이 서로 다른 서식처에서 경쟁적 교체의 일시적인 동태와 경쟁의 평형적 결과 사이의 유사성은 자원의 이용도 패턴 때문에 나타난다. 주어진 토양형에서 손실률이 높은 서식처는 부족한 토양 영양염류의 수준이 높고, 지표면에서의 광량이 많고, 수관층이 낮아 광의 수직 구배가 완만하다. 이것과 비슷한 조건이 경쟁적 교체가 일어나는 초기 과정에서 나타난다 . 개체군 성장 과정의 초기에 식물 량이 적고 식물의 키가 작은 것은 부족한 토양 영양염류의 수준이 높고, 지표면에서의 광량이 많고 수관층이 낮다는 것을 의미한다. 따라서 식물 생장의 초기에 볼 수 있는 자원 수준은 종으로 하여금 손실률이 큰 서식처에서 우점종이 되도록 해주는 경향이 있다. 개체 군이 성장함에 따라 부족한 토양 자원의 평형 수준과 광의 평형적 수직 구배가 형성되는데, 이러한 상태는 최초의 우점종과는 형태가 다른 종에게 유리하다. 서식처 내에서 볼 수 있는 일시적인 동태는 천이의 동태와 매우
유사하다(제 7 장). 이러한 일시적인 동태는 최대 생장률과 자원 이용 도에 의해 많은 영향을 받는다. 최대 생장률은 분배 패턴에 의해 결 정되고, 각 서식처에 따라 유리한 분배 패턴이 다르기 때문에 일시 적인 동태는 육상식생에서는 흔히 볼 수 있는 특징이어야 한다. 식물의 밀도를 감소시키거나 자원 이용도를 평형 수준 이상으로 증가시키는 교란이나 실험적인 조작은 천이 계열 즉 일시적인 우점 기간을 유도한다. 이러한 일시적인 동태는 영국의 Ro t hams t ed 에서 지난 130 년 동안 수행된 ParkGrass 의 시비 실험에서도 관찰되었다. 모든 영양염류를 공급한 두 경우에서 질산태의 형태로 질소를 첨가 하였는데, 최종 우점종은 Alop e curus p ra t ens i s 와 Arrhenath e rum avenaceum 이 었다(그림 6.7 의 B 와 C). Alo p ecurus 는 실험 기간 내내 절대수도가 증가하였다 . 그러나 Arrhenath e rum avenaceum 의 절대 수도는 질산태 질소를 공급받는 두 실험구 모두에서 처음 14 년 동안 7 배와 100 배로 감소하였다 . 그 이 후에는 이 식물의 수도가 서서히 증가하기 시작하였다. 시비를 하지 않은 대조구(그림 6.7A) 와 암모니아태질소를 질소원으로 공급받는 구(그림 6.7D-F) 에서는 수도가 감소하거나 증가하지 않았다. A g ros tis 와 Fes t uca 는 수도가 감소하기 전에 수년 동안 수도가 안정 상태에 있거나 중가하였다(그림 6.7 B-C). 암모니아태질소와 모든 종류의 무기질 비료를 공급받는 실험구도 일시적인 우점 기간이 있었다. 예를 들어, 어떤 실험구는 최종 우점 종인 Holcus lana tu s 가 다른 종을 교체하기 전에 처음에는 Dacty lis 가 우점하였으며 그 다음에는 A g ros ti s 가 우점하였다(그림 6.7E). 암 모니아태질소를 적게 받는 구에서도 이와 비슷한 우점종의 천이가 있었다(그림 6.7D). Cedar Creek 에서의 시비 실험 (제 8 장, 그림 8.23~ 8.26), 삼림의 tren chin g 실험 (Toumet & Kie n holz 1931), 그리고 포유 류를 제거하는 실험 (Browne t al . 1986) 을 포함하여 많은 다른 실험에
서도 이러한 일시적인 동태가 관찰되었다 . 4 일시적인 동태와 생태학적인 이해 일시적인 개체군 동태는 생태학자들이 자연 현상을 이해하는 데 어려움을 주는 주요인일 수 있다. 단기간의 실험 결과로부터 장기간 의 결과를 추측하여야 하기 때문에 현재로서는 그 종이 우점하고 있는 서식처를 결정하는 데 별로 중요하지도 않고. 그 종의 진화 과 정에서 간접적으로 선택된 특징을 너무 강조하는 경향이 있다. 생태 학에서는 아직도 실험적 조작(혹은 교란)에 대한 생물의 직접적인 반응을 조사하는 연구들이 많이 수행되고 있고, 이러한 직접적인 반 응들이 평형 상태의 군집 구조를 파악할 수 있는 지식을 제공하는 것으로 가정한다. 그러나 이러한 방법은 매우 부적합한 결론에 도달 하기 쉽다. 예를 들어 부족한 영양염류가 식생 구성에 미치는 영향을 연구한 다고 가정해 보자. 부족한 영양염류의 ·효과를 결정하기 위한 확실한 방법은영양염류의 첨가율을 달리하고 많은 반복구가 있는 야외 실 험을 하는 것이다. 몇 년이 지난 후에 실험 결과는 처리구에 따른 종 구성에서 통계적으로 매우 유의한 변화가 있는 것으로 나타난다. 부족한 영양염류가 이 서식처에서 볼 수 있는 자연식생의 패턴을 유발시키는 원인이 되는지를 결정하기 위해서 보통 사용되는 방법 은 실험적 구배상에서 나타나는 그 종의 수도가 야외의 자연 상태 에 있는 평형 구배상에서의 수도와 비슷한지의 여부를 알아보는 것 이다. 그러나 이러한 방법이 영양염류가 군집의 구성에 미치는 영향 을 판정하는 데 타당성이 있는가? 그림 6 . 8 에 나와 있는 예의 경우 를 생각해 보자.
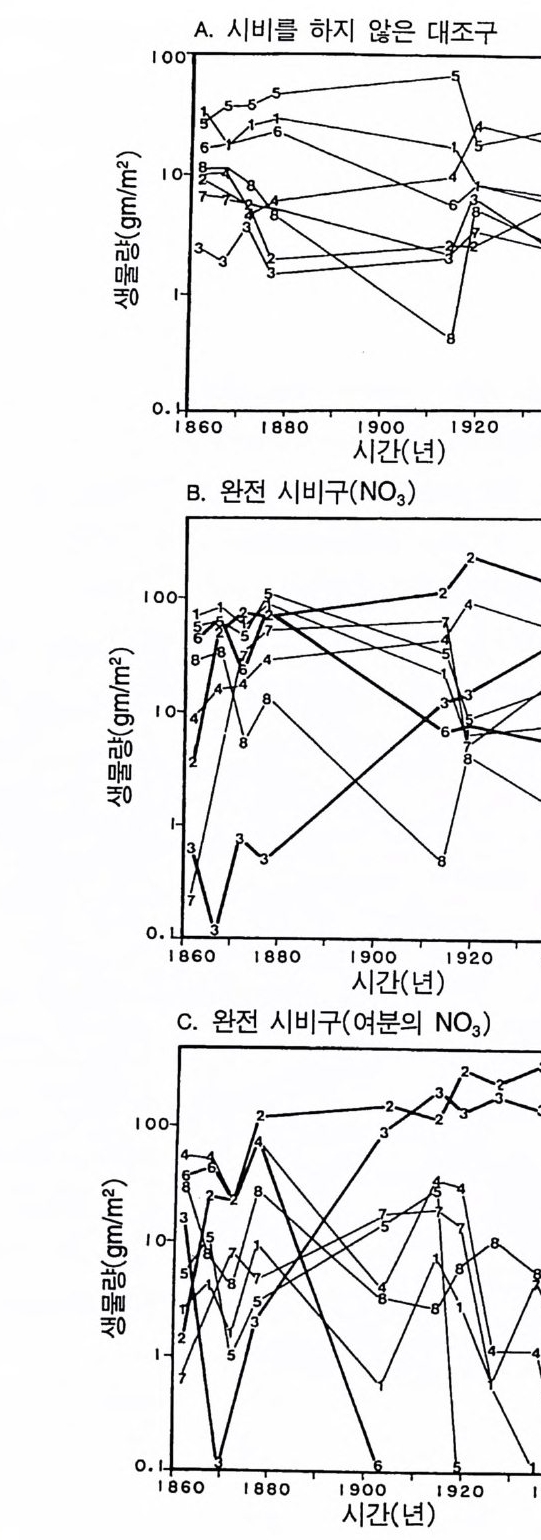 亡A 르
亡A 르
 뀜(e6팽E順nl)o 0DI,L18人,`8~A| H 二0函二{ o회1k92) 무 祠J I/ H1 \
뀜(e6팽E順nl)o 0DI,L18人,`8~A| H 二0函二{ o회1k92) 무 祠J I/ H1 \
그림 6.7 영국 Roth a mste d Exp er i me nta l S tati on 의 Park Grass Ex perim en t s 의 처음 100 년 동안 주요 식물종의 동태 . 후에 교체되는 종들에 의한 일시적인 우 점 기간이 있음을 주목하기 바란다. 일부 일시적인 우접은 특정한 비료와 연관된 토양 p H 의 점진적 인 변화에 의해 비롯될 수 있다. 생물량은 대수 눈금으로 표시되어 있다. T i lman (1 982) 의 자료를 변형시킨 것임.
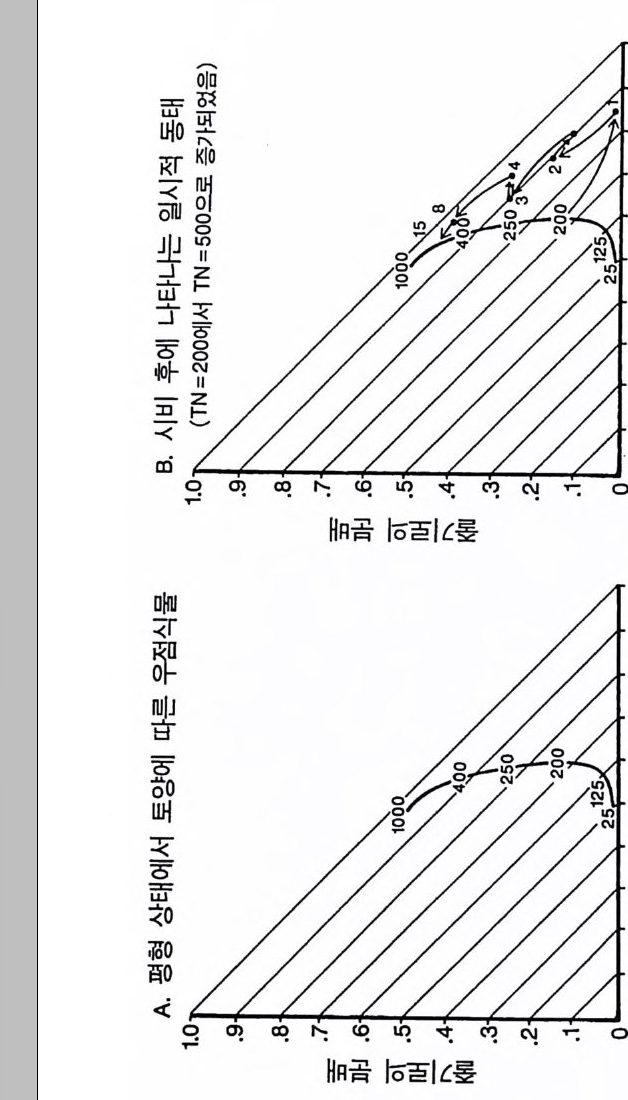
태처여의작물종
염손 실영양률류급이 률같()공A고 일정 되유게지 하평식 서 의처는는에 상 형서태TN 떤 형어이 종 경세적쟁우가 인를지될것 이결그이다 정한. 인림5T N은2=서 에서터처식부서 Ol인OO T=식 N 범에위서 우 쟁에세경준여태형보 한를 다N.2T= 처식서 B)인0 (0에실을 토험서 양조작로하으적 N게 0=되5T0이인시일적 하 면. 점우 획를위지종는하순 의득 서. 이다긴생 가 들 은그이종들 림 분A평 의부 라따 을선곡형고않 지하재존잎 분으 배로가의쪽 종 큰 편 로으존 어 되향할재이. 다것 은(들자숫 ,1 과, 43,,8 2 1COLLA 은)5TA한측예 이델모E 로으것 의각각 형 한를이 상태갖는식 인적시일 이얻 을간기 점우 한요필 데 는 우경 이 .다이수햇=NT는 인002서에처형식서평 우적점 다 .며되 이에턴패 배분 서른이량물생 는적 하작 시로으 종은 .다였.6 8 립그그림 6.8A 는 특정한 손실률을 갖는 서식처에서 평형 상태일 때 어떤 형태의 식물이 경쟁적 우세종이 될 것인지에 대한 예측을 보 여주고 있다 (ALLOCATE 의 모델을 기초로). 위의 논리는 TN=200 인 서식처를 TN=500 이 되도록 비옥하게 하면 그림 6 . 8A 의 곡선상의 그 점에 해당되는 형태를 갖는 종이 최종 우점종이 된다는 것을 제 시한댜 그러나 영양염류가 증가한 후 이 군집이 보이는 괘적은 단 순히 그림 6.8A 의 곡선상의 초기 우점종으로부터 최종 우점종으로 이동하는 것은 아니다 . 오히려 영양염류의 첨가는 이 곡선상에 속하 지 않는 종이 일시 적 인 우점 기 간을 갖도록 한다(그림 6.8 B ). 이 모의에서 평형 상태의 우점종이 일시적으로 우점하는 여러 가 지 종을 교체하는 데 약 15 년이 소요되었다 . 더구나 모델은 일시적 인 우점종의 계열이 규칙적이고, 경쟁 동태에서 반복되어 나타날 수 있는 특징으로 예측하고 있다. 이것만으로도 반복구가 잘 되어 있는 실험은 한 해가 지난 후에 영양염류 첨가가 어느 한 종에 유리하다 는 것을 보여주는 통계적으로 유의성이 높은 결과를 낳기 쉽다. 2 년 후에도 다른 종이 유리하다는 것을 보여주는 결과가 나올 수 있으 며, 이러한 과정은 평형 상태의 우점종이 다른 종들을 교체할 때까 지 계속된다. 실험 조작의 장기적인 결과를 파악하기 위해 15 년 이 상이 필요하다는 것을 알지 못하는 사람들은 중간에 나오는 결과를 잘못 해석할 수 있다. 삼림식생에서 수행되는 실험에는 이보다 더 긴 기간이 필요할 수 있다. 나는 삼림의 숲틈, 초지에서 볼 수 있는 소규모의 교란, 그리고 일반적인 재생전략에 관한 대부분의 연구에 근시안적인 요소가 포 함되어 있다고 믿는데, 그것은 바로 이런 이유 때문이다. 그런 연구 들의 대부분은 단기간의 일시적인 동태에 초점을 두고 있으며, 여기 에서 얻은 정보를 가지고 외삽하여 전체 군집의 구조를 설명하려고 한다. 그러나 이러한 단기간의 연구는 최대 생장률의 차이에 너무
큰 비중을 두고 군집의 구조에 중요한 요인이 될 수 있는 부족한 토양 자원과 햇빛에 대한 경쟁 능력의 차이를 모호하게 하는 경향 이 있다 . 단기간의 연구에서 매우 중요하게 여겨지는 최대 생장률의 차이 는 진화적인 면에서 보면 상이한 식물 형태의 선택에 간접적인 영 향을 줄 뿐이며. 군집의 평형 구조에는 직접적인 영향이 거의 없을 수 있댜 수관층을 구성하는 교목에 의해 형성되는 평균 조건에 대 한 실생과 유묘의 반응은 교란에 대한 반응만큼이나 중요할 수 있 다. 이러한 교란은 삼림의 다양성을 유지하는 데 분명히 중요한 요 인이 되며. 장기간의 평균 교란율이 서로 다르면 종 구성도 달라야 한다. 이러한 종 구성상의 변화는 내재하는 기작에 의해 설명되어야 한다. 높은 교란율은 자원의 평균 이용도를 변화시키기 때문에 교란 의 빈도가 높으면 낮을 때에 비해 상이한 생리와 형태를 가진 종이 유리하게 된다. 생태 연구를 하는 과정에서 보다 어려운 것 중 하나는 많은 인내 가 필요하다는 점이다 . 사람의 수명, 직업, 야망 그리고 호기심이 지 속되는 기간은 군집이 실험적인 조작이나 자연적인 교란에 완전히 반응하는 데 필요한 시간보다 보통 짧기 때문에 인내가 필요한 것 이다. 따라서 육상의 식물 군집에서 수행되고 있는 대부분의 생태 실험은 어느 중간 종에 의한 일시적인 우점 기간에 해당되는 것들 이다. 유감스럽게도 최초의 우점종이 갖는 특성은 자연 군집을 구성 하는 요소에 관하여 우리에게 별로 정보를 주지 못한다. 또한 어떤 종이 마지막 승자가 될 것인지를 결정하는 데 초기 우점종 개체군 의 동태를 이용하는 것도 만만한 일은 아니다. 그림 6.5 와 6.6 에서는 비옥한 토양에서의 최종 우점종이 그들의 상대수도가 증가하기 전에 오랫동안 상대수도(그리고 절대수도까지 도)가 감소할 수 있음을 보여주었다. 예를 들어, 그림 6.6 의 종 FF 는
처음에 수도가 감소하였고 다른 종들이 밀집되고 키가 큰 숲을 형 성하기 전에는 수도가 증가되지 않았다. 이 종은 줄기로의 분배량이 많기 때문에 다른 종들이 밀집되고 키가 큰 숲을 형성하면 오히려 경쟁에 유리하다. 나는 주어진 토양형 혹은 손실률에서 경쟁의 초기 (2-3 년) 동태를 이용하여 어느 종이 최종 우점종이 될 것인지를 예 측하는 방법을 찾아내지 못하였다(그림 6.2, 6.4 , 6.5 , 6.6 ) . 그러나 야 외에서 수행된 대부분의 경쟁 실험은 단지 1-2 년 동안만 지속되었 다 (Schoener 1983, Connell1983). 만약 이러한 모의가 육상의 식물 군집에서 경쟁의 기작을 나타내 는 것이라면. 이러한 결과는 성숙한 평형 상태의 군집을 직접 관찰 하거나 장기간의 실험을 통해 서식처 특성에 따른 경쟁 능력의 차 이를 가장 잘 추정할 수 있음을 제시하는 것으로 볼 수 있다. 이 두 방법 모두가 생태학에서 최근에 유행하는 단기간의 실험보다 더 나 은 방법이라고 할 수 있다. 그러나 이 두 방법은 모두 중요한 결점 울 가지고 있다. 성숙한 군집에서 패턴에 기초를 둔 상관 연구는 그 저 상관일 뿐이다. 이러한 연구 결과는 종과 종 사이의 인과 관계를 보여주지 않기 때문에 쉽게 인과 관계에 관한 오해를 야기할 수 있 댜 생태학에서는 종과 종 사이의 상호 작용이 직접 또는 간접적인 경로를 통해 일어나기 때문에 특히 더 그렇다 (Lev i ne 1976, Lawlor 1979, Vandermeer 1980). 이러한 간접적인 효과 때문에 다영양 단계가 포함되어 있는 계의 종간 상호 작용의 기작을 하나의 장기간의 실험으로 명확히 설명하 는 데 어려움이 있다. 예를 들어 영양염류를 첨가하는 실험은 특정 한 종이 최종 우점종이 되도록 유도할 수 있는데. 그것은 이 종의 경쟁 능력 때문이 아니고 그 종이 영양염류 첨가 후에 생산력이 증 가함에 따라 수도가 증가한 초식동물에 대하여 저항력이 가장 크기 때문일 수가 있다. 이러한 어려움 때문에 생태학자들이 자연 군집에
서 실험을 수행할 때 여러 가지 경쟁 가설을 분명하게 설명해 줄 수 있는 유일한 실험울 결코 고안해 낼 수 없을 것이다. 그러나 자 연 군집에서의 상관 연구나 장기 혹은 단기간의 조절된 실험, 그리 고 조절된 실험 조건하에서 종간 상호 작용의 기작에 대한 연구들 은 모두 자연이 어떻게 작용하는가를 이해하는 데 중요한 정보를 제공할 수 있다. 따라서 자연계를 이해하기 위해서는 실험, 관찰, 그 리고 이론적안 접근 방법이 포함된 다면적인 연구 방법이 필요하다. 나는 순전히 현상학적 인 방법 (ph enomenolog ica l a pp roaches) 을 사용 하는 것은 바람직하지 않다고 본다 . 최근에 식물 경쟁에 대한 연구 의 두 가지 주요 접근 방법은 모두 현상학적인 것으로, (1) 제거 실 험이나 이식 실험과 같은 밀도 조작 실험 (Schoener 1983, Connell 1983 의 종설 참조)과, (2) de W it의 교체 계열이 있다. 두 가지 방법 은 모두 중요한 개념상의 문제가 있다. 밀도 조작 방법은 Lo tk a-Vol t era 에서 유래된 한 쌍의 경쟁, 즉 한 종의 밀도의 증가가 다른 종의 밀도(혹은 생장률)를 감소시킬 때 두 종이 경쟁한다는 정의에 기초를 두고 있다. 그러나 이 정의는 다른 종이 없는 상태에서 두 종 사이의 상호작용에만 적용된다. 두 종 이 상이 존재하면 다른 종을 통한 간접적인 효과가 이들 사이의 상호 작용 효과를 완전히 변형시킬 수 있다. Lawlor(1979) 가 보여준 바와 같이, 두 종의 직접적인 상호 작용의 기작이 경쟁인 경우에도 많은 종이 있는 군집에서 두 종은 상리공생의 기능을 할 수 있다. 밀도 조작 실험은 두 종간에 직접적인 경쟁이 아닌 분명한 상리공생 관 계 가 있음을 알아낼 것 이 다 (T il man 1987b). Fowler(1981) 는 노스캐롤라이나의 초원에서 수행한 일련의 제거 실험에서 이러한 분명한 상리공생 관계를 발견하였고, 이 상리공생 을 다종 경쟁의 간접적인 효과로 설명하였다. Bender 등 (1984) 은 주 어진 종 한 쌍이 직접적인 경쟁자인지의 여부를 결정하기 위해서는
별개의 반복되는 실험에서 모든 종이 제거되어야 한다는 것을 보여 주었다. 대 안으로 Bender 등( 1984) 은 한 종의 밀도 변화가 모든 종 의 밀도와 생장률에 미치는 즉각적이면서 일시적인 영향을 파악할 수 있도록 실험이 계획되어야 한다고 주장하고 있다. 이러한 방법에 는 Lotk a -Volte r ra 모델에서 가정한 것처럼 모든 상호 작용이 직접 적인 밀도 효과에 의해 결정될 수 있다고 가정하면 한 쌍의 종이 어떻게 상호 작용을 하는지를 결정하기 위해서는 2 번의 반복 실험 만이 필요하다. 그러나 자원의 이용도가 밀도 조작에 반응하여 서서히 변화하고, 자원 수준에 대한 생장과 사망의 반응에 지연시간이 있다면, 그리고 그림 6.5 와 그림 6.6 에 나타난 것처럼 자원의 동태가 복잡하다면 이 러한 실험은 두 종 사이의 경쟁을 보여주지는 못한다. 따라서 밀도 조작 실험은 특히 단기간의 실험일 경우 자연 상태의 경쟁을 기술 하거나 설명하는 데 그 범위가 제한되어 있다. 그러나 Bender 둥 (1984) 의 방법은 그것이 단기간의 실험일지라도 적당한 동적인 모 델의 범주 내에서 설명된다면 경쟁 관계를 설명하는 데 유용할 수 도 있다는 것을 보여주기 때문에 중요성이 있다. de W it(1 960) 의 교체 계열 방법은 이론적으로 결점이 있어 종간 경쟁의 장기적인 결과를 예측할 수 없다. Inou y e 와 Sch affe r(1981) 는 de Wi t 방법을 통한 예측은 실험에 사용하는 전체 개체군의 밀도에 의존적임을 밝혔다. Sp itter s(1983), Connoll y (1986) 도 교체 계열 방법 이 전체 밀도에 대한 경쟁적인 효과의 의존도를 무시하기 때문에 이론상 종간 경쟁의 결과를 예측하지 못한다는 것을 밝혔다. 더구나 de W it (1960) 의 방법은 경쟁의 장기적인 결과를 예측하는 데 초기 의 동태를 이용하려고 한다. 그러나 이미 설명한 바와 같이 경쟁의 초기 동태는 경쟁의 장기적인 결과와 거의 관계가 없다. 따라서 식물들 사이의 종간 경쟁 현상을 설명하기 위해 사용되어
온 두 가지 중요한 방법은 매우 신중하게 고려되어야 한다. 나는 자 연을 이해하려는 일념으로 우리가 종간 상호 작용의 기작. 이러한 상호 작용을 변화시키는 선택적인 힘의 진화적 근거. 그리고 이것들 과 서식처의 주요 비생물 및 생물적 제약들 사이의 관계를 연구하 는 것이라고 믿고 있다. 이러한 연구를 하는 과정에서 최대 생장률의 차이 . 간접적인 효과 의 존재, 연령 구조의 존재. 그리고 다른 많은 요인들이 자연 군집들 로 하여금 실험조작 후에 일시적인 동태의 기간을 갖도록 할 수 있 다는 것을 기억하여야 한다. 하나의 실험이 평형 상태의 결과를 관 찰할 수 있을 정도로 오랜 기간 동안 수행되지 않는다면. 그 실험이 입증하고자 하는 내용과 거의 관계가 없지만 통계적으로 매우 유의 한 실험 결과를 얻을 수 있다. 단기간의 실험은 필요하다. 그러나 이 러한 실험은 동적인 모델의 단기간의 예측을 시험하는 경우에만 유 용하댜 5 요약 경쟁의 동태는 매우 복잡한 것으로 그 군집이 최종 우점종에 의 해 우점되기 전에 여러 종이 일시적인 우점 기간을 갖는다(그림 6.2-6.6). 그러나 이러한 일시적인 동태에도 일정한 패턴이 있다. 최 초의 우점종은 최대 생장률이 높은 키가 작은 식물인데, 잎으로의 분배량은 많고 줄기로의 분배량이 적다. 이러한 종들은 최대 생장률 이 점차 낮아지는 종들에 의해 교체되는데, 그 이유는 뿌리나 줄기 로의 분배량이 많아지기 때문이다. 영양염류가 빈약한 토양에서 맨 처음 우점종은 척박한 토양의 손실률이 높은 상태에서 경쟁에 이기 는 종들이다(그림 6.3 과 그림 4 .1 8 올 비교). 이러한 종들은 자신과
비슷하게 역시 영양염류가 빈약한 토양에서 경쟁력이 큰 종에 의해 교체되지만 이러한 종들은 점점 손실률이 낮아진다. 따라서 영양염 류가 빈약한 토양에서 진행되는 일시적인 우점 과정은 그러한 토양 에서 우세한 경쟁자가 되는 일단의 종을 나타내는 것이지만 이들 종은 점차적으로 손실률이 낮아진다. 비옥한 토양에서도 이와 비슷한 점진적인 교체 패턴을 볼 수 있 댜 주어진 토양에서 일시적인 우점 기간을 얻는 종의 계열은 같은 토양에서 평형 상태의 손실률이 높은 쪽에서 낮은 쪽으로 이어지는 구배상에서 볼 수 있는 종의 계열과 비슷하다. 손실률이 다른 평형 상태를 경쟁의 일시적인 동태와 정확히 연계시키지는 못하지만, 이 들 사이의 유사성은 각각의 영양염류 공급률과 손실률에서 특정한 종을 우점하게 하는 것과 똑같은 과정이 경쟁의 단기간의 동태에도 영향을 준다는 것을 의미하는 것이다. 이 과정은 자원의 경쟁이다. 경쟁하는 동안에 볼 수 있는 최초의 동태는 경쟁의 최종 결과와 직접적으로 관련이 없는 일시적인 동태일 수 있다. 이러한 동태가 많은 생태학자들이 연구비를 받는 기간 혹은 관심을 갖는 기간 내 에 일어나기 때문에 생태학의 일반적인 지식은 종의 생활사를 변화 시키는 기본적인 힘, 그리고 자연군집의 모양을 결정짓는 주요 제약 과는 직접적으로 관계가 없는 일시적인 동태에 기초를 갖는 경우가 많다 .
제 7 장 천이 1 서론 육상의 식생 천이는 나지에서의 식물 개체군의 동태로 정의되는 데, 19 세기 말부터 20 세기초에 생태학의 주요 연구 과제였고 논쟁의 대상이 되어온 분야이다 (Cowles 1899, Coop er 1913, Clements 1916, Gleason 1917, 1927). 그 동안 천이 에 관한 많은 논문과 책 이 나왔지 만 아직도 천이의 패턴을 결정짓는 원인에 대해서는 생태학자들 사 이에 의견이 일치하지 않고 있다. 이것은 천이라는 주제가 너무 광 범위하기 때문에 어느 하나의 요인이나 하나의 과정으로 천이의 모 든 것을 설명하기가 불가능하다는 데에 그 원인의 일부가 있다. 군집을 구성하는 개개의 종은 특유의 생리적, 형태적. 그리고 생 활사적인 특징을 가지고 있다. 또한 각각의 서식처는 독특한 토양. 지형. 기후. 그리고 과거의 역사를 가지고 있다. 따라서 초기 개척자 식물의 밀도나 그 지역에 정착할 수 있는 식물의 종은 각각의 천이
과정에 따라 다르다. 더구나 식물의 정착이 확률적인 것이며 , 정착 에 성공한 식물이 그 서식처의 식물 생장에 많은 영향을 주기 때문 에 천이는 우연적이라고 볼 수 있다. 그럼에도 불구하고 천이는 지 역적으로 반복되는 과정이며, 각 지역에서 진행되는 천이 과정에는 많은 공통점이 있다 (Wes t et a l. 1 981 참조). 많은 생태학자들이 천이 에 관한 연구에 관심을 갖는 이유는 바로 이러한 유사성 때문이다. 그러나 천이의 원인에 대한 실마리를 제공해 주는 것은 바로 여러 지역의 천이에서 관찰할 수 있는 차이점들이라고 할 수 있다. 2 1 차 천이 천이는 나지에서부터 시작된다. 빙하가 후퇴하여 생긴 지역, 새로 형성된 사구 . 산사태 지역, 화산 폭발이나 침식 지역 등에서 진행되 는 1 차 천이의 경우에는 나지의 기질 속에 유기물이나 식물의 생식 체가 들어 있지 않다. 완전벌목 지역이나 휴경지 등에서 진행되는 2 차 천이는 초기에 이미 유기물이 풍부한 토양과 여러 종류의 식물 의 생식체가 들어 있다. 대부분의 천이는 이러한 극단의 두 가지 천 이 패턴 사이에 위치한다. 이러한 지역에서 천이를 유발시키는 힘은 무엇인가? 천이를 유발시키는 힘에는 많은 것이 있을 수 있으며, 이들 요인 들은 상호 배타적인 것이 아니다 . 나는 1 차 천이의 패턴을 결정하는 데 있어서 부족되는 자원의 역할을 먼저 설명하고, 2 차 천이에서 자 원의 역할을 설명한 후 1 차 천이와 2 차 천이 모두에서 볼 수 있는 기타 다른 과정들의 역할을 설명하도록 하겠다. 질소를 제외하고 식물이 필요로 하는 모든 무기염류는 토양이 형 성되는 모암에서 기인된다(J enn y 1980). 따라서 빙하가 후퇴한 지역
이나 중습 (mes i c) 한 서식처에 형성된 사구에서 식물의 생장을 제한 하는 주요 토양 요인은 질소이다. 예를 들어 알래스카의 Glacie r Bay 에서는 소빙기 (L itt le Ice Ag e) 말기 이후 230 년 동안 빙하의 후퇴가 빠르게 진행되었다 . 따라서 이러한 지역에서는 1 차 천이 과정의 식 생 변화사를 분명 하게 알 수 있다 (Coo p er 1923, 1939, Crocker & Majo r 1955, Lawrence 1958). 미류나무나 가문비나무처럼 Glaci er Bay 에서 진행되는 천이의 후기 우점종들의 대부분은 새로 퇴적되는 빙 퇴석에 초기에 정착한 식물들이지만 초기에는 대부분 생존하지 못 하거나 질소 부족으로 왜소한 형태를 갖거나 백화 현상을 나타낸다 ( Lawrence 1979 ) . 천이 초기의 우점종들은 대부분 질소를 고정할 수 있다 (Law-rence 1979). 예를 들어 일부 지역에는 초기에 질소를 고정하는 남조 류인 blackcrus t가 우점하였다 (Worle y 1973). 질소를 고정할 수 있는 초기 우점종에는 지의류인 S t ereocaulon 과 Lemp h olemma, 장미과인 Drya s drummondii , 오리 나무 (A/nus crisp a ), 기 타 여 러 가지 루핀과 콩과식 물이 있다 (Lawrence et al. 1967, Rein e rs et al. 1971, Lawrence 1979). 이러한 초기 우점종이 잎이나 뿌리의 일부를 소실하거나 개체 전 체가 죽으면 질소를 포함하고 있는 유기물들이 여러 가지 분해자들 에게 이용된다. 이 분해자들은 유기물 속에 있는 질소의 상당량과 대기중으로부터 공급되는 질소를 토양에 유지시켜 결국 토양의 질소 함량이 증가하게 된댜 분해자들도 함질소 노폐물을 분비하는데 이 것을 관속식물이 이용할 수 있다. 따라서 질소를 고정할 수 있는 생 물이 초기예 우점하고, 대기중으로부터 이입되는 질소의 축적으로 Glaci er Bay 지역 토양의 전질소 함량이 점진적으로 증가한다. Crocker 와 Maj or (1 955) 는 Glaci er Ba y에서 1 차 천이의 처음 100 년 동안에 토양의 질소 함량이 5-10 배 증가한 사실을 관찰하였으며.
Crocker 와 D i ckson(1957 ) 도 알래스카의 다른 지역에서 이와 비슷한 질소 증가를 보고한 바 있다. Ol s on ( 1958) 은 미시간 호 남부에 형성 된 사구에서 천이가 진행됨에 따라 질소 함량이 증가하는 것을 관 찰한 바 있다 . 이와 같이 토양의 전질소 함량의 증가는 질소 공급률 을 증가시킨다. 토양 내에 무기 상태의 질소인 암모니아태질소 (NH 4 ) 와 질산태질 소 (N0 3 ) 가 가용성 상태로 만들어지는 비율을 토양의 질소 무기화율 이라고 한다. 질소의 무기화율은 토양의 pH , 산소량, 수분, 온도 , 유 기물 함량 등 많은 요인과 관련이 있는 것으로 알려져 있다 ( Chic h este r et al. 1975, Melill o et al. 1982, Vi tou sek et al. 1982, Pasto r et al. 1982). Robe rt son 과 Vit ou sek( 1981 ), 그리고 Robert so n( 1982) 은 미 시간호 남부에 있는 사구 토양의 질소 무기화능이 전질소 함량, 곧 사구의 나이에 비례하여 증가함을 알았다. 이것은 1 차 천이의 처음 100 년 혹은 200 년 동안에 질소가 매우 결핍되어 있는 무기토양이 시간이 지남에 따라 질소 공급률이 증가되는 질소가 풍부한 유기토 양으로 전환됨을 의미한다. 토양의 질소 함량이 증가함에 따라 전체 식물의 생물량도 증가한 다 . 예를 들어 Gla ci erBa y의 Mu i rInle t에서는 초기의 우점종이 토양 조류와 지의류이었는데, 그 이후에는 질소를 고정하는 관목인 Dr ya s drummond ii로 바뀌었다. 이들은 높이가 3-lOm 까지 자라는 오리나 무로 대체되었다 . 그 이후에는 오리나무의 그늘에서 자라다가 오리 나무보다 키가 커지는 미류나무가 우점종이 된다. Lawrence(1 9 79, p. 217) 에 의하면 빙하가 녹은 후 50 년이 경과하면 Mu ir Po i n t에는 10 여 m 까지 자라는 오리나무 덤불숲이 형성되며, 이 덤불숲으로부터 미류나무가 나타나기 시작한다. 이 미류나무는 빙하가 후퇴한 후에 바로 나타나지만 직립하지 못하고(지역에 따라서는 거의 40 년 동안) 오리나무가 적당한 양의 질소를 공급한 후에야 신속한 직립생장이
일어난다는 것이다 . 이 미류나무는 그보다 키가 큰 가문비나무와 솔송나무에 의해 교체되는데, 이 식물들은 미류나무의 그늘을 뚫고 자랄수 있댜 이와 비슷한 현상을 미시간 호에서도 볼 수 있는데 (Cowles 1899, Olson 1958), 초기 우점종의 극히 일부만이 질소 고정식물이라는 점 이 다르다 (Olson 1958). Olson 은 사구 토양에 유기질소가 축적되는 이유를 대기로부터 이입되는 질소량은 같고 소실되는 질소량이 적 기 때문인 것으로 설명하고 있다. 사구는 처음엔 질소 함량은 낮지 만 광량은 많은 서식지이다. 토양의 질소 함량이 중가함에 따라 전 체 식물의 생물량이 증가하고 키가 큰 식물이 유리하게 된다. 일부 지역에서는 초기에 marram g rass 와 기타 사구를 고정하는 협엽초본 들이 우점하지만, 시간이 지나면 다양한 광엽초본이 들어오고, 그 이후에는 관목이 침입한다. 마지막에는 방크스소나무, 스트로브잣나 무, black oak 등의 교목이 우점한다. 사면과 경사가 다르고 점토의 함량이 많은 지역에서는 (Olson 1958), 초기의 협엽초본 다음에 광엽 초본, 관목, 참피나무, redoak, 그리고 당단풍나무가 차례로 우점하 게 된다. Olson 은 이러한 천이 과정에서 단일극상은 없다는 점을 강 조하였다. 사구에서 마지막으로 형성되는 군집의 종 구성은 사구의 경사, 사면, 그리고 토양의 모재에 따라 달랐다. 이 러한 연구들은 다양한 1 차 천이에 적용할 수 있는 특징을 제시 하고 있다 . 1 차 천이의 초기에 침입한 실생들은 토양의 영양염류 함 량은 낮고 광량은 높은 환경 조건에 놓이게 된다. 다양한 분해생물 에 의해 진행되는 토양 형성 과정을 통해 부족한 영양염류의 이용 도가 증가함에 따라 지상부의 생물량은 증가하고 그 결과 지표면에 이입되는 광량은 감소한다. 따라서 1 차 천이는 적어도 처음 몇 백년 동안에는 하나 이상의 부족한 토양 자원과 지표면에서의 햇빛의 상 대적인 이용도의 시간에 따른 구배로 생각할 수 있다. 부족한 토양
자원의 축적률이 경쟁적 교체율에 비해 느리면 1 차 천이는 평형 상 태의 식물 군집에 이르는 점진적인 과정으로 설명할 수 있다. 이때 어느 시점에서의 식생 구성은 주로 부족한 토양 자원과 햇빛의 상 대적인 이용도에 의해 결정된다. 나는 이것을 천이의 〈 자원비 가설 (resource rati o h yp o t hes i s) 〉이라고 부른다 (T i lman 1982, 1985). 그러나 이 장의 후반부에서 설명하겠지만 일시적인 동태도 1 차 천이에 영 향을 줄 수 있다. 자원비 가설은 자원 의존적인 생장 둥구배곡선을 이용하여 쉽게 설명할 수 있다(그림 7.1 ). A 에서부터 E 로 표시된 5 종은 부족한 토 양 영양염류와 햇빛의 요구량에서 분화되어 있다 . A 종은 영양염류 의 요구량은 가장 낮고 지표면에서의 햇빛 요구량은 가장 높다. 이 종은 짧은 포복성 생장형을 갖으며 뿌리:지상부의 비가 높아, 줄기 가 적고 조직의 영양염류 농도는 낮다. 이 종은 천이 초기에 흔히 볼 수 있는 영양염류 함량이 적고 광량이 높은 서식처에 우점한다. E 종은 영양염류 함량이 높고 광량이 낮은 서식처에서 경쟁력이 탁 월한 종으로 햇빛의 요구량은 가장 낮고 영양염류의 요구량은 가장 높다. 이 종은 키가 크고 직립성이며, 뿌리:지상부의 비가 낮다. 또 한 줄기에 분배하는 에너지량이 많다. 종 B, C, D 는 영양염류와 햇 빛의 요구량 면에서 중간 정도에 속한다. 만약 식물종들이 이 5 종의 식물에서 가정한 것처럼 부족한 토양 영양염류와 햇빛에 대한 경쟁 능력에 차이가 있다면 각 종은 토양 영양염류:광 구배상의 서로 다 른 위치에서 최대 수도를 갖을 것이다(그림 7.l B) . 1 차 천이의 어떤 주어진 시점에서, 경쟁적인 상호 작용은 평형 상 태에 도달하려는 경향이 있으나 토양의 영양염류 수준은 시간에 따 라 점진적으로 변화하는 경향이 있다면 서식처 0 에서부터 서식처 4 까지의 구배는 천이 과정의 시간을 나타내는 것으로 볼 수 있다. 천 이 초기종은 부족한 영양염류에 대한 경쟁력이 우세하여 영양염류
 버ot0o 前―O lJ韓輯O部O鄧
버ot0o 前―O lJ韓輯O部O鄧
종원된영이발
이 들 자한 안정건 이 의빛를유 .다이 러이 종의의조 햇 의 체교()A 서곡존시구 A의의에 된 들표들곡- 종생둥E장A터로선배부 적원 E 자선 은 과 부의다자빛 서햇 각이원로으것 요토 양구 량된 종주한간은 은다른족 들로. 의비들 율들점소는개 해의 4울제하비한원에 자을받 등은두게이로균다 .한 준천여보 을점평형 데는1나조 서 구을인처과의 식것타 양차.모걀낸달이. 정다 하표약태 는류상 염양비은서높 지 는은도면용나 이양 토 의빛옥 서햇빈하에에나 )B( 도용 자종 러 . 다천이적된화변 한 의로원은으률는건이조급의적 은공낮 괘 이 〉다부원자 고천키시을가것이고 .비다라킨원 른시화설변자.이 〈를 도이의용 .17 림그수준이 낮은 때인 1 차 천이의 초기에 우점하게 될 것이다 . 영양염류 수준이 증가하고 지표면의 햇빛 이용도가 감소됨에 따라 이 종들은 햇빛에 대한 경쟁력은 높지만 부족한 영양염류에 대한 경쟁력은 낮 은 종들에 의해 교체될 것이다. 따라서 우점종의 변화는 부족한 토 양자원의 공급률을 조절하는 과정에 의해 결정되는 것으로 예측할 수 있다. 둘 이상의 부족한 자원에 대한 요구량이 서로 다를 때는 자원비 구배상에서 항상 종이 분리될 것으로 예상된다. 식물종에 따라 햇빛 과 토양 자원에 대한 요구량이 다른 것은 뿌리-줄기-잎으로의 분배 대응책에서 비롯될 수 있다. 자원비 가설은 천이룰 이해하기 위해서 는 부족한 자원의 비만을 알아야 한다든지, 천이는 직선적인 과정이 어야 한다든지, 혹은 천이가 안정된 극상을 갖어야 한다는 의미는 아니다. 이 자원비 가설의 요점은 부족한 자원의 상대적인 공급률의 변화가 식물군집 구성의 변화를 초래한다는 것이다. 식물의 소비. 교란, 그리고 다양한 생물지화학 과정들이 모두 부족한 자원의 상대 적인 이용도 변화에 영향을 줄 수 있다 . 이 가설은 부족한 자원의 공급을 조절하는 요인들이 방향성이 있고 반복되는 성질이 있다면 천이가 방향성이 있고 반복될 수 있 는 과정임을 가정하고 있다. 만약 자원 공급률이 순환성이라면 천이 는 진행천이가 아닌 퇴행천이가 된다. 예를 들면, Gla ci erBa y에서의 고전적인 1 차 천이는 처음 몇 백년 동안의 식생 변화를 기술하고 있다. 그러나 삼림지에 Sp hag num 이 침입하거나 철분에 의한 경질지 층 (hard p an) 이 형성되면 토양이 산성화되고 통기성이 낮아진다. 그 결과 질소의 무기화율이 저하되고 식물 생물량이 감소되어 햇빛의 입사량이 중가된다. 그러면 밀폐된 수관이 소택지와 같이 키가 작고 수관층이 개방된 식생에 의해 교체된다 . 동부 오스트레일리아의 사구에서도 이와 유사한 퇴행천이를 볼
수 있다 (Walkere t a l. 1981). 사구가 형성된 후 최초의 천이는 북미에 서 기술된 바 있는 고전적인 경우. 즉 수관층이 폐쇄된 삼림이 사구 에 형성되는 것과 매우 유사하다. 그러나 인과 칼슘이 서서히 세탈 되고 대기로부터 이입되는 양이 매우 적으면 사구가 형성된 지 100,000-400,000 년 동안에 이러한 영양염류의 심한 결핍이 일어난 댜 영양염류의 수준이 감소됨에 따라 식물량이 감소하고, 지표면의 광량은 증가한다. 그리고 30m 정도의 키가 큰 나무들이 밀폐된 수 관을 형성하였던 곳에 키작은 교목이나 관목이 우점하게 된다. 자원 비 가설은 자원 이용도에서 방향성이 있거나 반복될 수 있는 변화 를 가정하지 않기 때문에 천이에 관한 연구, 죽 교란 후 식물 동태 룰 극상 개념의 한계로부터 구분하고 있다 (Clemen t, 1916). 토양 자원의 공급이 부족한 육상 서식처에서는 1 차 천이의 초기 단계 기간에서와 마찬가지로 토양 자원의 이용도 증가가 식물량의 중가를 초래하여 지표면에 이입되는 광량을 감소시킨다. 이러한 현 상은 그림 7 .1에서 볼 수 있는 것과 같이 각 종이 분리될 수 있는 일시적인 토양 자원:광 구배를 형성하게 된다. 이 가설은 토양의 영양염류보다 광이 더 부족하게 됨에 따라 천 이가 진행된다고 주장하는 것이 아니다. 오히려, 이 가설은 그러한 일시적인 구배가 특정한 토양 자원 : 광량의 비에서 경쟁력이 큰 일 련의 종에게 순차적으로 유리하다는 점을 강조하고 있다. 이 이론에 의하면 어떤 서식처에서 두 종이 공존하는 것은 한 종이 어느 한 자원에 의해 더 제한울 받고 다른 한 종은 또다른 자원에 의해 더 제한을 받기 때문에 가능하다. 토양 자원:광 구배상의 특정한 위치 에서 경쟁력이 큰 종은 그것의 생리, 형태, 생활사가 그 위치에서 두 자원에 의해 거의 똑같이 제한을 받도록 하기 때문에 우위를 차지 할 수 있게 된다. 토양 조류는 영양염류가 극도로 빈약한 토양에 우점하는데, 그것
은 이 조류들이 영양염류는 매우 적지만 광량이 많은 조건에서 경 쟁력이 우세하기 때문이다. 이 조류는 약간 비옥해진 토양에서는 키 가 작은 1 년생 식물들과 공존하는데, 그 이유는 영양염류에 대해서 는 경쟁력이 큰 조류가 햇빛의 제한을 받기 때문이다 . 반면에 1 년생 식물은 키가 커 토양 조류에 비해 햇빛에 대한 경쟁력이 우세하지 만 영양염류의 제한울 받는다 . 더 비옥한 토양에서 1 년생 식물과 토 양 조류가 모두 햇빛의 제한을 받으면 1 년생 식물이 토양 조류를 교체하게 된다. 비옥한 토양에서 1 년생 식물은 자신보다 키가 더 큰 다년생 식물과 공존할 수 있는데, 그 이유는 각각의 식물이 서로 다 른 자원, 즉 1 년생 식물은 햇빛에 의해 그리고 다년생 식물은 영양 염류에 의해 제한을 받기 때문이다. 영양염류 공급의 전범위에서 햇 빛과 토양 자원 모두는 우점종에게 부족된 상태일 것으로 예측된다. 지표면에서 햇빛의 이용도가 감소되고 영양염류의 이용도가 증가 하는 경우에도 자원비 가설은 우점종 식물의 일부가 가장 비옥한 토양을 제외한 모든 토양에서 영양염류에 의해 제한 받을 것임을 예측한다. 이 종들이 햇빛에 대한 경쟁력을 얻기 위하여 부족한 토 양 자원에 대한 경쟁력을 포기함으로써 영양염류 함량은 높지만 광 량이 적은 토양에서 경쟁에 우세할 수 있기 때문에 이러한 현상이 나타난다. 이와 같은 예측은 세계에서 가장 비옥한 토양에 있는 평 형 상태의 삼림에서 우점종 교목이 여전히 영양염류에 의해 제한울 받는다는 사실과 일치한다 (M ill er & Mi ller 1976, Albrekts o n et al. 1977, Wa rning et al. 1978, Br ix 1983). 이와 유사하게 다양한 생태계에서 조사된 영양염류의 순환과 영 양염류 이용 효율에 대한 종설에서도 비옥한 토양에 형성되어 있는 성숙림이 토양 영양염류, 특히 질소에 의하여 제한된다는 것이 지적 된 바 있다 (V it ousek 1982). 또한 자연 식생에 처리한 시비 실험에 의해서도 이러한 사실이 입증된다. 예를 들어 Cedar Creek 의 시비
실험은 (T i lman, 1983, 1984, 1987a) 천이 초기의 극도로 척박한 토양 에서부터 훨씬 비옥한 참나무사바나에 이르는 상이한 5 개 지역에서 오직 질소만이 부족한 토양 자원임을 보여주고 있다. 그러나 모든 지소에서 공통종의 절반 정도만이 질소 시비에 반응하여 증가하였 다 . 나머지 반은 질소 시비 후에 감소되었다 . 이러한 패턴은 질소 이 외의 또 하나의 다른 자원인 햇빛에 의하여 거의 균등하게 제한을 받는 지소에 있는 종들의 반응 결과와도 일치하였다. 3 토양요인과군집 구성 그림 7 .1에 요약되어 있는 이론은 평형 상태에서 식물 군집의 구 성이 부족한 토양 자원의 공급률에 의해 크게 영향을 받을 것이라 고 예측한다. 식생 구성에 따른 토양 요인의 변이에 관해서는 많은 사람들이 보고한 바 있다. Whit tak er( 1953, p. 44) 는 〈토양 요인에 대 한 언급은 거의 모든 문헌에서 볼 수 있으며, 토양 요인의 효과는 모든 군집에서 뚜렷하게 관찰된다〉고 언급하였다. J enn y (l980) 가 지 적한 바와 같이 지리적으로 광범위한 규모에서 식생 구성을 결정하 는 주요 요인들은 기후와 토양이다. 지역의 규모가 작을 때에는 기 후의 변이가 훨씬 적은데 비해 토양은 매우 이질적일 수 있다. 토양 의 이질성은 모재, 경사, 사면, 그리고 교란의 차이에서 비롯된다. 또 한 퇴적과 침식 과정들이 토양의 이질성에 영향을 준다. Pasto r 등 (1982) 의 연구 결과는 지소에 따른 질소 무기화율과 그 지소에 우점하게 되는 식생의 종 구성 사이의 관계를 잘 보여주고 있댜 죽 이 연구는 초기의 토양 모재(특히 점토의 함량), 그 모재 에서 형성된 토양의 질소 무기화율과 그 지소에 최종적으로 형성되 는 식생 사이에 뚜렷한 상관이 있음을 보여주고 있다. Zak 둥(1 986)
은 Pasto r 등 (1982) 이 위스콘신의 Blackhawk Island 에서 행한 것과 비슷한 연구를 미시간에서 수행하였다. 이들은 Blackhawk Island 보 다는 훨씬 넓은 지역에 걸쳐 있는 12 개의 서로 다른 지소를 조사하 였댜 이 12 개 지소 중 4 개 지소는 큰떡갈나무와 whit e oak 가 혼합 되어 있고 하층에는 Vac ci n i um 이 우점하고 있었다. 나머지 중 4 개 지소는 당단풍나무와 red oak 가 우점하고 하층에는 Ma i an t hemum 이 우점하였다. 나머지 4 개 지소는 당단풍나무와 피나무가 우점하고 있 고 하층에는 Osmorh i za 가 주로 분포하고 있었다. 이 지소들은 앞에서 언급된 순서대로 비교적 수관이 개방된 삼림 으로부터 더 밀폐된 수관을 갖는 삼림으로, 그리고 수관층이 현저히 밀폐된 삼림에 이르는 구배를 가지고 있다. 그들은 각 지소에서 토 양을 채취하여 각 토양의 질소 무기화율을 측정하기 위하여 일정한 온도와 습도에 토양을 배 양하였다. 이들은 black oak 와 whit e oak 가 우점된 지소의 토양이 다른 두 군집보다 질소 무기화율이 현저히 낮음을 알았다. 그리고 당단풍나무와 redoak 가 우점하고 있는 지소 는 당단풍나무와 피나무가 우점하고 있는 지소에 비해 질소 무기화 율이 현저 히 낮았다(그림 7.2). 따라서 적어도 6km 정도 떨어져 있는 지소들간의 토양의 질소 무기화율은 그것의 식생 구성의 좋은 지표가 된다 (Zak et al.1986). 하나 예외적인 것은 12 개의 지소 중 질소 무기화율이 가장 낮은 당 단풍나무 지소였다(그림 7.2). 이 지소는 처음 채집된 토양들이 잘못 된 것일 수도 있다는 생각에 다시 토양을 채집하여 실험하였는데 그 결과도 역시 똑같았다 (Zak, 개인적인 정보 교환). 그러나 이 지소 는 특이하게 식물의 생물량과 현존량이 낮았고, 지표면에 입사되는 광량이 많았다. Zak 은 이 지소의 상층 토양이 완전벌목된 기간에 소 실되었으며, 당단풍나무와 피나무는 종자량이 많아 재형성되었을 것 이고 black oak 와 wh ite oak 는 종자량이 많지 않아 재형성되지 못하
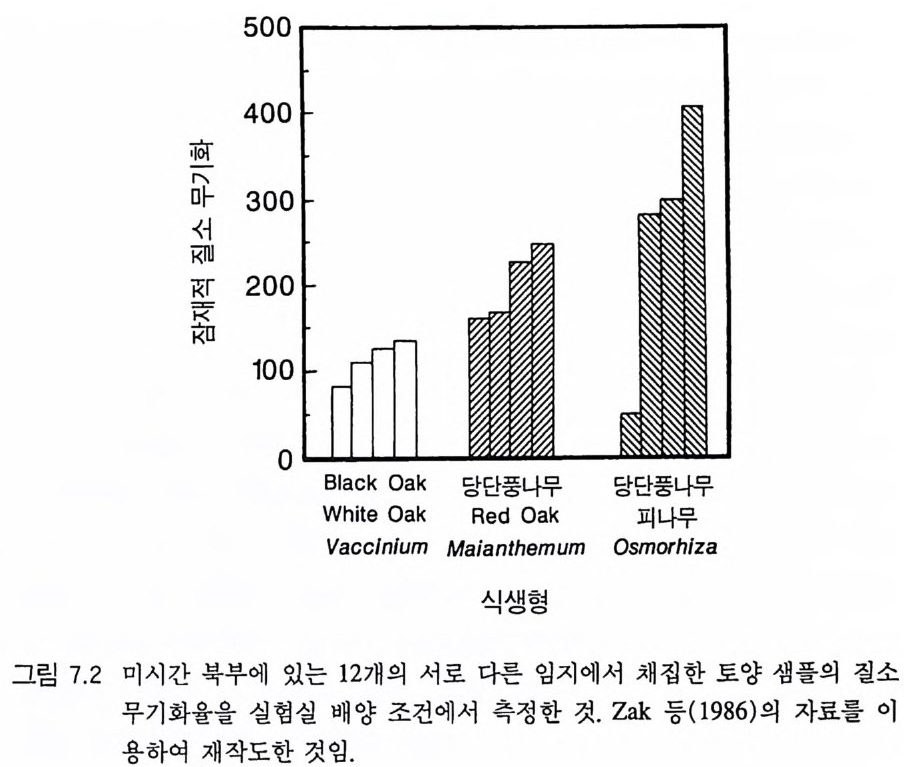 500
500
였을 것이라고 믿었다. 만약 그렇다면 이 지소는 참나무의 침입이 가능하며, 결국 참나무에 의해 당단풍나무와 피나무의 밀도가 감소 될 것이댜 당단풍나무-피나무가 우점한 지소를 포함하여 모든 지소 에서 질소 무기화율이 높은 지소는 Pas t or 가 Blackhawk Island 에서 밝힌 것처럼(그림 5.6) 수관층이 더 밀폐되고 지표면에 도달하는 광 량이 적었다. 이러한 결과들은 Olson (1 958) 이 미시간호의 남부 인디애나 사구 에서 관찰한 결과와도 놀랍도록 잘 부합된다. Olson 은 이 지역의 천 이 후기의 식생이 토양의 질소 함량과 관련이 있음을 보고하였다. 질소 함량이 적은 토양에 발달된 식생은 피나무-당단풍나무가 우점 하였다.
Robe rt son 과 Vit ou sek(1981). Robe rt son(1982) 은 이 사구 토양에서 질소 무기화능을 조사한 결과 전질소 함량이 낮은 토양은 무기화율 도 낮음을 알았댜 이 러 한 결과는 Zak 등( 1986) 의 연구와 인디 애 나 사구에서 수행된 연구 결과 사이의 유사성을 입증하는 것이다 . 따라 서 수백 km 의 거리가 떨어져 있어도 특정한 지역에 마지막으로 우점 하는 식생의 구성은 모든 지역에서 바슷하게 토양의 부족한 자원의 공급률에 의해 결정되는 것 같다 . 토양의 질소와 임상에 이입되는 광량에 의해 상층 수관수의 종류 가 분리되는 것은 이 수종들의 질소와 광 요구량이 다름을 의미한 다. 이 종들 사이에 예상되는 차이는 자원 의존적인 생장 등구배곡 선을 이용하여 나타낼 수 있다(그림 7.3). 그림 7 . 3 에 나타나 있는 관계는 이 종들을 서로 상이한 토양에서 단독 재배를 할 때 이용 가능한 토양의 질소와 햇빛이 감소되는 수준을 측정하여 검증할 수 있는 가설이다. 이러한 자료를 얻는 것은 생각보다 그렇게 어렵지 않다. 이 종 중 일부는 임학에서 매우 중요하기 때문에 많은 단독 재배 지소가 이미 조성되어 있다. Pasto r 등(1 984), Olsen( l95 8), Rober t son 과 Vi tou sek(1 9 81), Robert so n(1982), 그리고 Zak 등 (1986) 의 연구 결과들이 식생의 구 성과 질소 공급률 사이의 관계를 보여주긴 하지만 이러한 상관 관 계의 연구는 인과 관계를 잘 설명하지는 못한다. 이들의 결과는 식 생에 대한 질소 공급 효과보다는 질소 공급에 대한 식생의 효과를 나타내는 것으로 설명될 수 있다. 이미 설명한 바와 같이 이러한 양 자택일의 해석이 가능한 이유는 토양은 식물에게 영향을 주고 또한 식물은 토양에게 영향을 주기 때문이다. 이러한 문제를 해결하는 가 장 좋은 방법은 햇빛을 포함하여 부족한 자원을 직접 실험적으로 조절하는 것이다. 그러나 삼림식생을 이해하는 데 이러한 실험을 이용하기 위해서는 실험이 오랜 기간 계속되어야 한다.
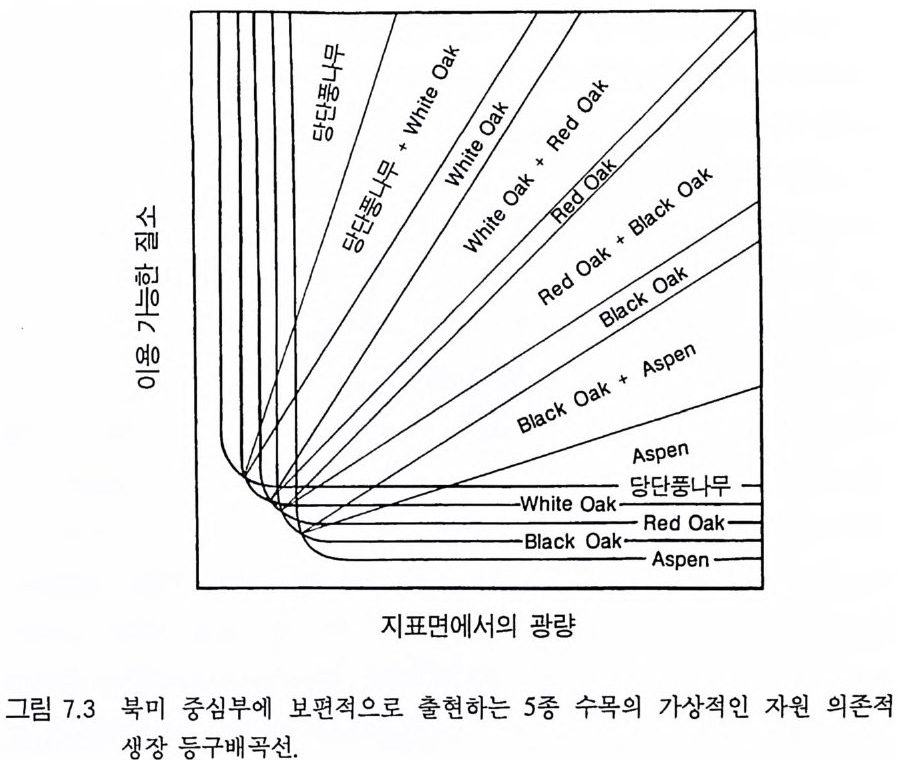 서
서
다른 방법으로는 식생과 그 지역 토양의 모재 사이의 관계를 살 펴봄으로써 어떤 관련성을 알아낼 수 있다. 이미 언급한 바와 같이 Blackhawk Island 에서는 표충토 아래의 점토 함량과 최종적으로 형 성되는 토양 및 식생 사이에는 일치하는 점이 많았다. Jen ny (1 980) 는 토양 형성 요인 중에서 모재의 중요성을 강조한 바 있다. 따라서, 한 지역의 토양 모재와 그 지역에 마지막으로 우점하는 식생의 구 성 사이에는 강한 상관 관계가 있다. 물론 모재가 식생 구성을 결정 하는 유일한 요인은 아니다. 교란의 체제, 산불의 빈도, 지역에 따른 초식동물의 밀도, 그리고 기후가 한 지역의 식생 구성에 영향을 준 다. 그러나 이러한 요인들을 일정하게 할 경우 식생 구성은 토양의 모재와 밀접한 관련이 있다.
예를 들면, 위스콘신 주의 토양은 깊이가 lm 이상이고 산성이며, 빙하 퇴적물인 모래로 되어 있는데, 정착 이전에는 주로 소나무사바 나(pi ne barrens) 가 우점하였다(그림 7.4 . Hole 1976). 다른 식생형으로 는 참나무사바나, 초지, 소나무림 이 발달되 어 있었다 (Hole 1973). Hole 이 만든 위스콘신의 토양과 식생지도에 의하면 토양 모재와 식 생 사이에 뚜렷한 유사성을 볼 수 있고, 대부분 Pasto r 등(1 984). Olson (19 58 ) , Zak 둥 (19 86 ) 그 리 고 Roberts o n 과 Vit ou sek ( 1981 ) 에 의 해 보고된 패턴을 입증하고 있다. Wh itn e y (1986) 는 미시간의 몇 지역에서 원식생과 토양을 조사하 였는데, 이 조사 결과도 이들의 주장과 부합되었다 .• 미 네소타의 토 양과 식생 지도에서도 토양 모재와 그 지역의 우점 식생 사이에 강 한 상관이 있음을 볼 수 있다. 위스콘신에서와 마찬가지로 미네소타 의 넓은 모래평원에는 소나무사바나와 참나무사바나 혹은 초지가 형성되어 있다. 두 지역 모두 빙퇴석에는 참나무림이나 혹은 당단풍 나무-피나무림과 같은 밀폐된 수관층을 갖는 식생이 형성되어 있다. Lin d sey (1 961, p. 434) 는 북부 인디애나의 교란되지 않은 식생의 경우 서로 다른 빙하의 작용에 의해 만들어진 모재에 따라 그 패턴 이 다름을 알았다. 이와 유사하게 이스라엘의 Up pe r Gal i lee 에서 Rabin o ri tch -Vi n( 1 9 79. 1983) 은 비슷한 기후 조건하에 있는 같은 위 도대에 상이한 극상군락이 형성되어 있음을 발견하였다. Rabin - ov it ch-V i n 은 이러한 식생 구성의 차이룰 토양의 모재에서 비롯된 것으로 설명하고 있다. 식생은 토양에 의존적이라는 많은 연구들이 있다. 이들 연구 중에는 불과 몇 m 혹은 몇 km 떨어져 있는 지역의 식생을 비교한 것도 있고 서로 다른 대륙에 형성된 식생을 비교한 것도 있다 (Beard 1944, 1955, Box 1961, Snayd o n 1962, Pig o tt & Tayl o r 1964, Zecller & Zecller 1969, Hanawalt & Whit tak er 1976, Cody & Mooney 1978, Beard 1983).
 A. 빙하에 씻겨나온 모래가 밑에 있는 사토
A. 빙하에 씻겨나온 모래가 밑에 있는 사토
그림 7.4 (A) 위스콘신에서 빙하에 씻겨나온 모래가 밑에 깔려 있는 깊은 사토가 있는 지역. (B) 위스콘신의 원식생에서 소나무사바나의 위치. Hole(1976) 의 자료를 이용하여 재작도한 것임 .
전반적으로 이러한 연구 결과둘은 초기의 모재를 포함하여 토양 의 특성이 1 차 천이 과정 중에 종의 수도를 조절하는 주요인임을 보여주고 있다. 또한 토양 자원과 햇빛이 공간과 시간상으로 역상관 관계에 있고, 식물은 토양 자원:광 구배상의 특정한 지점에 적응한 다는 견해를 뒷받침하는 것이다. 이러한 증거는 경쟁력이 좋은 종은 모든 자원에 대한 경쟁력이 좋다는 Gr i me(1979) 의 주장과 상반되는 것이다. 게다가 이러한 연구들은 토양 자원의 공급률에 영향을 주는 여러 가지 요인들이 자원 공급률을 평형 상태에 이르도록 할 경우 에만 1 차 천이가 안정된 극상에 이를 것임을 제시하고 있다. 4 자원과 2 차 천이 앞에서 설명한 바와 같이 1 차 천이의 동태는 토양 발달에 의해 크 게 영향을 받으며, 토양의 형성은 경쟁에 의한 식물의 교체율과 바 슷하게 진행된다. 이와 같이 토양 형성이 느리게 진행되기 때문에 비옥한 토양에서 진행되는 2 차 천이 과정에 영향을 줄 수 있는 여 러 가지 요인들의 중요성이 감소된다. 비옥한 토양에서 진행되는 2 차 천이 중에는 천이가 진행되는 동안 영양염류가 축적되지 않고 소실되는 경우도 있다 (Aarsen & Turki ng ton 1985). 영 양염류가 빈약한 토양에서 진행되는 2 차 천이의 경우에는 1 차 천이에서처럼 천이가 진행되는 동안 영양염류의 축적이 일어난다 (B illi n g s 1985, Odum 1960, Rice et a l. 1960, Inouye et al. 1987a). 영양염류의 상태에 차이가 있음에도 불구하고 1 차 천이, 척박한 토양에서 진행되는 2 차 천이, 그리고 비옥한 토양에서 진행되는 2 차 천이 사이에는 많은 공통점이 있다. 척박한 토양이나 비옥한 토양에 서 진행되는 2 차 천이는 모두 초기의 우점종이 초본식물이다. 이들
은 생장이 빠르며, 성숙한 개체의 키가 작고 수명이 짧은 것들이다. 반면에 천이의 후기에 우점하는 종은 보통 생장이 느리고 수명이 길며, 키가 큰 목본성 다년생 식물이다 . 1 차 천이에서 천이 후기에 우점하는 종은 보통 생장이 느리고 수명이 길며, 키가 큰 목본성 다 년생 식물이다 . 1 차 천이에서도 키가 작고 수명이 짧은 초본성 식물 에서 키가 큰 목본식물로 변화된다. 그러나 이와 같은 유사성은 모든 천이과정을 일반적으로 설명하 는 데 사용되는 토양 자원 : 광 구배의 역할에 위배되는 것이다. 비옥 한 토양에서의 2 차 천이는 영양염류와 햇빛이 모두 풍부한 상태에 서 시작된다 . 그림 7 .1의 A 종이나 B 종과 같이 1 차 천이의 초기종들 이 경쟁력이 높은 조건, 죽 광 이용도는 높지만 영양염류의 이용도 는 낮은 조건은 결코 나타나지 않는다. 그러나 제 6 장에서 설명한 바 와 같이 주어진 서식처(자원의 공급률과 소실률이 고정된) 내에서 경 쟁적 교체의 동태는 경쟁종들의 최대 생장률에 의해 크게 영향을 받는댜 부족한 자원의 이용도 범위 내에서 교란에 의해 식물의 밀도가 낮은 새로운 서식처에서의 초기 우점종은 생장률이 빠른 것들이다. 이 우점종들은 자신들보다 최대 생장률은 낮지만 그 서식처의 조건 에서 경쟁력이 좋아지는 일단의 종에 의해 교체된다(그림 6.2-6.6). 따라서 각 종의 최대 생장률이 다르다면 경쟁적 교체 과정 중에 우 점종이 점진적으로 변화되리라는 것을 예상할 수 있다. 이것은 비옥 한 토양에서 진행되는 2 차 천이 과정 중 관찰되는 천이 계열이 경 쟁적 교체의 일시적인 동태임을 의미하는 것이다. 만약 그렇다면, 정착이나 초식 등과 같은 다른 과정들을 비슷하다고 간주할 때 비 옥한 토양과 척박한 토양에서 진행되는 2 차 천이 과정의 유사점과 차이점은 무엇인가?
4-1 일시적인 동태 2 차 천이 과정 중의 특정한 시기에 어떤 종이 우점종이 될 것인 가를 결정하는 데에는 두 가지 요인이 관여한다. 그 중 하나는 일시 적인 동태인데, 이것은 최대 생장률 , 분배 패턴, 그리고 천이 초기의 개체군의 밀도가 낮을 때 모든 자원의 비교적 높은 초기 이용도에 의해 영향을 받는다. 다른 하나는 장기적인 토양 변화 과정으로 척 박한 토양에서 진행되는 2 차 천이 과정에서 관찰되는 장기간에 걸 친 질소의 축적과 같은 것이다. 척박한 토양이나 비옥한 토양을 막론하고 2 차 천이 과정의 모든 경우에서 일시적인 동태에 의해 주로 조절되는 기간이 있음을 예상 할 수 있다 . 예를 들어 불모지인 척박한 토양 (TN=100) 에 100 종이 서로 경쟁하는 경우를 생각해 보자. 100 년 동안에 토양의 영양염류 량이 점차 중가하여 TN=1000 이 되었다. 이 2 차 천이 과정의 처음 10 년 동안의 동태는 종의 최대 생장률의 차이에 가장 크게 영향을 받는 주로 일시적인 동태이다(그림 7.5A). 이 기간 후에 관찰되는 동 태는 점진적으로 축적되는 영양염류에 의해 더 많은 영향을 받는데, 어떤 주어진 시점에서의 종 구성은 최근의 토양 영양염류 풍부도에 의해 결정된다. 척박한 토양에서 진행되는 이 2 차 천이 과정의 각 시점에서 우점 종의 특성을 추적해 보면 최대 생장률이 높은 종에서 낮은 종으로 이행된다. 또한 잎에 분배량이 많고 줄기에 분배량이 적으며, 뿌리 에는 중간 정도의 분배량을 갖는 키가 작은 종에서 뿌리로의 분배 량이 많고 잎으로의 분배량이 적은 키가 작은 종을 거쳐, 줄기로의 분배량은 증가되고 뿌리로의 분배량이 점차 감소되는 키가 큰 식물 로 변화된다(그림 7.5A). 초기 우점종 식물에서 볼 수 있는 잎이 많고 생장이 빠른 특징은
 .6 A 척박한 토양
.6 A 척박한 토양
그림 7.5 초기의 영양염류 풍부도가 서로 다른 토양에서 천이 룰 조절하는 과정에서 일시적인 동태와 장기적인 영양염류 축적의 역할. 모든 경우에서 토양은 100 년 후에 TN=lOOO 이 되는 것으로 간주되었다. 빗금천 타원형은 일시적인 동태에 의해 야기되는 우점종 의 순서를 나타낸다. 빗금이 없는 타원형은 100 년 동안에 영양엽류 축적에 의해 야기 되는 우접종의 순서를 나타낸다. (A) 척박한 토양 (TN=lOO) 에서는 일시적인 우접 기간이 약 10 년간 지속되며, 그 이후 90 년 동안은 영양염류가 축적됨으로써 우접종의 변화가 일어난다 . (B) 비옥도가 중간 정도의 토양(초기의 TN=200) 에서는 일시적인 동태가 7 년 정도 지속된다. (C) 비옥한 토양(초기의 TN= lOOO) 에서는 전체 순서가 순전히 일시적인 동태이다. 이러한 일시적인 동태는 약 6 년 동안 지속되었다. 이 토양 은 TN=lOOO 으로 출발하여 더 이상의 양분 축적이 일어나지 않았음을 주목하기 바 란다. 이러한 모의는 모든 토양이 마지막에 TN=lOOO 이 되는 것으로 가정하며 . 따라 서 똑같은 평형 군집을 갖는 것으로 간주한다. 이것은 분명히 모든 천이에 해당되는 경 우는 아니다. 이것은 순전히 이 3 가지 경우의 비교를 쉽게 하기 위해 가정된 것이다.
평형 상태의 군집에서는 볼 수 없다 . 초기에 우점하는 식물종은 생 장이 빠르고 새로 생긴 휴경지에 조성되는 영양염류와 광 조건에서 다른 종과의 경쟁에서 일시적으로 우세하기 때문에 우점 기간을 획 득하는 일시적인 종들이다. 척박한 토양에서 일시적인 우점 기간이 있을 수 있다는 예측은 T i lman(1985) 과는 다르다. 즉 , 나는 비옥한 토양에서 진행되는 2 차 천이 과정에서만 일시적인 동태가 중요하다 고 말한 바 있다. 그러나 그와 같은 결론이 나오도록 한 모델은 형 태적으로 그 종을 분명히 규정하지 않았고 단지 천이의 초기종을 영양염류가 빈약한 서식처의 종으로 국한시킨 것이었다 . 좀더 비옥한 토양(초기의 TN=200 에서 100 년 후에는 TN= 1000) 에 서 진행되는 2 차 천이 과정의 우점종을 추적해 보면 일시적인 동태 가 중요성을 갖는 기간이 더 길다(그림 7.5 B ). 더욱 비옥한 토양(초 기의 TN=1000, 100 년 후의 TN=1000) 에서 전체 천이 계열은 일시적 인 동태에 의해 결정된다(그림 7 . 5C) . 이 모든 것들이 마지막에는 같 은 토양을 갖는 것으로 제한하고, 손실률이 같은 것으로 제한하였기 때문에 이들 2 차 천이의 모든 경우에서 마지막 우점종은 모두 같다. 더구나 모든 경우의 2 차 천이에서 초기 우점종은 모두 키가 작고 잎이 많으며 생장이 빠르다는 특징을 갖는다. 따라서 이 모델은 천 이 초기에 영양염류 함량이 서로 다른 토양에서 시작되는 2 차 천이 사이에 정성적인 유사성이 있음을 예상하고 있다. 그러나 실제 우점종의 계열은 다르게 예측된다. 즉 영양염류가 빈 약하고 손실률이 낮은 척박한 토양에서 진행되는 2 차 천이 과정에 서 우점하는 종은 비옥한 토양에서의 2 차 천이 과정에서는 결코 우 점종이 되지 못한다(그림 7.5 의 A, B, C 부분을 바교하기 바람). 반대 로 영양염류가 풍부하고 손실률이 높은 서식처에 우점할 수 있는 종은 비옥한 토양에서 진행되는 2 차 천이 과정에서 일시적인 우점 기간을 확보할 수 있지만 척박한 토양에서의 2 차 천이 과정에서는
그렇지 못하댜 이러한 결과들은 종이 천이 초기종이 되는 데 필요한 여러 가지 성질이 있음을 제시하는 것이다. 손실률이 중간 정도인 서식처에서 1 차 천이 과정 중에 초기 우점종은 최대 생장률이 낮은 종이 될 것 으로 예상된다. 그 이유는 이 식물이 영양염류를 획득하기 위한 구 조(뿌리와 균근) 혹은 과정(질소 고정 혹은 매우 효율적인 흡수계)에 분배량이 많아 잎으로의 분배량이 적기 때문이다. 영양염류 요구량 이 많고 생장이 빠른 종은 1 차 천이 초기에는 생장이 불가능하여 일시적인 우점 기간을 확보하지 못할 것이다. 이와는 반대로 비옥한 토양에서의 2 차 천이 과정의 경우 초기 우점종은 생장률이 빠르기 때문에 일시적인 우점 기간을 얻을 수 있다. 식물이 우점종이 되는 서식처의 조건을 고려하지 않고 단순히 천이 초기종 혹은 천이 후 기종으로 구분하는 것은 별 의미가 없는 것으로 판단된다. 4-2 야초의 진화 이 책에서는 식물의 분화를 유도하는 두 가지 중요한 요인이 토 양 자원의 공급률 구배와 손실률 구배라고 주장하고 있다. 식물 개 체의 생리적인 특성과 형태적인 특성이 특정한 영양염류 공급률과 손실률에서 경쟁에 이길 수 있는지의 여부를 결정하게 된다. 농경의 시작과 기타 인위적인 교란의 시작으로 손실률이 높은 서 식처가 급격히 증가하게 되었다. 신석기시대로부터 현대까지 농업이 확장되는 동안 많은 종들이 손실률이 높은 서식처에서 경쟁에 유리 하도록 진화되었울 것이다. 이들 중 많은 종이 생활사가 짧고 비 목 본성이며 줄기로의 분배량이 비교적 적은 종으로부터 파생되었을 것이댜 현재 볼 수 있는 야초의 가장 그럴듯한 조상은 토양의 영양 염류 함량이 비교적 낮고 개방된 곳에서 경쟁력이 큰 종들일 것이
댜 이들은 사구나 자갈이 덮인 지역, 암반이 노출된 지역 , 황무지 둥지에서 우점하는 종일 것이다. 현재 천이 초기 과정에 우점하고 있는 종들은 인간에 의한 대대 적인 교란이 있기 전에는 어떤 과정을 통해 존속되어 왔을까? 가장 일반적인 설명은 그 종들은 개척종 또는 주변종(fugiti ves p ec i es) 적인 생활사를 갖기 때문에 유지되어 왔다는 것이다. 즉 정착률이 우세하 고 생장률이 빠르기 때문에 원시 서식처에서 소규모의 교란지를 활 용할 수 있었다는 것이다 . Marks (1 983) 는 북미 동부의 원시 경관에서 현재 천이 초기의 우 점종들은 농경이 시작되기 전에 어디에서 잔존하였는가를 조사하는 과정에서 이 문제를 설명하였다. 그는 구시대의 식생에서부터 현재 삼림의 개방지역에 있는 식생 구성, 그리고 북미에 식민지가 형성된 후 교란되지 않은 많은 임지의 서식처에 대한 기술 내용을 연구의 기본 자료로 사용하였다. 그는 오늘날에는 새로 형성되는 휴경지와 관련된 고유식물이 일시적으로 형성되는 삼림 공간이 아닌 지속적 인 개방지에 일차적으로 출현한다는 것을 알았다 (Marks 1983 p. 225). 이것은 오늘날 천이 초기종 역할을 하는 이 식물들이 삼림 내에 소 규모의 지역적으로 교란된 곳에 특수하게 적응되도록 진화된 것이 아님을 의미한다. 그는 대부분의 고유식물이 수백년 혹은 수천 년간 개방된 주변 서식처에서 진화되었다고 주장하고 있다(p . 225). 이러한 지속적인 서식처로는 석회암 노출 지역, 자갈로 된 언덕, 모래평원, 애추사면, 암반의 톱 산둥성이, 바위가 있는 불모지의 토양, 강의 하중도, 가파 른 침식강변 둥이 속한다. 또한 이러한 모든 지역은 광량이 많은 서 식처이다. 이들 중 대부분은 토양이 척박하기 때문에 광량이 많지 만, 일부 지역은 손실률도 높다. Marks 가 천이 초기종으로 생각한 바랭이새류, 망초, 몇 종의 미역
취, 몇 종의 목본 관목 등은 성숙하였을 때 키가 작고, 북미 동부의 비옥한 토양에 우점하는 당단풍나무와 너도밤나무보다는 최대 생장 률이 큰 식물들이다. 따라서 Marks 의 관찰에 의하면 2 차 천이의 초 기에 혼히 나타나는 많은 종들은 비옥한 토양을 갖는 극상식생의 교란된 지역에 일시적으로 존재하도록 생활사가 진화된 것이 아니 고 척박한 토양을 갖는 지속적이며 개방된 서식지, 그리고 손실률이 높은 서식지에 적응한 종들이다. 5 천이에 관한 다른 견해들 나는 천이에서 자원 경쟁의 역할을 강조하였는데. 그 이유는 자원 경쟁이 가장 중요하기 때문만은 아니다. 자원 경쟁 이외에도 몇 가 지 다른 과정들이 있는데, 경쟁적 교체의 일시적 동태도 천이의 패 턴을 결정할 수 있는 중요 요인 중의 하나이다. 이러한 요인 중의 하나가 천이 과정의 첫단계인 정착이다. 만약 어떤 종이 그 서식처에 도달할 수 없다면 결코 그 지역의 우점종이 될 수 없다. 그 지역에 도달한다하더라도 그곳에서 생존하여 생장할 수 없으면 역시 우점종이 될 수 없다. 그러나 종간 경쟁이 없는 서 식처에서 우점할 수 있는 잠재적인 정착식물 모두를 고려한다 하더 라도 정착 그 자체로는 천이를 설명할 수 없다. 나는 지금까지 연구 된 것 중에서 그 지역에 처음 정착하여 자라는 종이 영원히 그 지 역을 점유할 수 있다는 결과를 보지 못하였다. 〈정착-경쟁 가설 (coloniz a ti on -comp et i tion h yp o th es i s) 〉로 천이 과정을 설명 할 수 있기 위해서는 식물의 정착 능력과 생존하고 생장하기 위한 식물의 다른 능력들 사이에 대응책이 있어야 한다. 예를 들어 Werner 와 Pla tt (1979) 의 연구에서 제시된 것처럼 정착-
경쟁 가설은 정착 능력이 좋은 종은 경쟁력이 낮은 것으로 가정하 고 있댜 그렇다면. 나지에서 진행되는 천이는 처음에는 신속히 정 착할 수 있는 종들이 우점하다 후에는 정착 능력은 낮지만 경쟁력 이 높은 종에 의해 대체될 것이다. 그 지역은 결국 정착력은 가장 낮고 경쟁력은 가장 큰 종이 우점하게 된다. 〈정 착-초식 가설 (Coloniz a ti on herbiv o ry h yp o t hes i s) 은 정 착력 이 좋 은 종이 초식에 더 민감한 것으로 가정하여, 천이 후기에는 초식에 더 저항성이 있는 종들로 교체될 것이다. 영양염류가 풍부한 지소에 서는 식물의 생장이 빠르기 때문에 정착은 그 중요성이 증가할 수 있댜 초기 정착자의 생장이 빠르면 그 조건에 더 알맞은 종들이 그 지역에 들어오기 전에 우점하는 기간을 갖게 된다. 그러나 모든 1 차 천이와 일부 2 차 천이는 영양염류가 빈약한 토 양에서 시작된다. 척박한 토양에서는 식물의 생장이 느리기 때문에 어떤 종이 다른 종을 능동적으로 교체할 수 있는 기회를 갖기 이전 에 긴 정착 기간이 있게 된다. 예를 들어 알래스카의 Glacie r Ba y에 서 천이 과정의 후기 우점종 중 많은 수가 새로 노출된 빙퇴석에 맨 처음 정착하는 종들이다 (Lawrence 1979). 그러나 이와 같이 정착 이 빠른 종은 첫번째 우점종이 되지 못한다. 그 이유는 이 종들은 극도의 척박한 토양에서 생장할 수 없기 때문이다 (Lawrence 1958, Crocker & Majo r 1955). 따라서 정착은 비옥한 토양에서 진행되는 2 차 천이 과정보다는 척박한 토양에서 진행되는 1 차 천이와 2 차 천 이 과정에서 오히려 중요성이 적은 것 같다. 천이의 패턴에 관한 또 하나의 가능한 설명은 〈경쟁-초식 가설〉 이다. 이 가설은 종의 경쟁 능력과 초식에 대한 민감도 사이에 대응 책이 있다고 가정한다. 죽 경쟁력이 큰 종은 초식에 더 민감하다는 것이다. 만약 그렇다면, 초식률의 변화가 일정한 방향성이 있으면 식생 구성의 변화도 어떤 방향성을 갖게 될 것이다. 예를 들어 천이
초기에 초식률이 낮고 천이가 진행됨에 따라 초식률이 증가한다면 경쟁력은 높지만 초식에 민감한 종으로부터 경쟁력은 낮지만 초식 에 저항성이 높은 종으로의 천이 계열이 형성되어야 한다. 천이가 진행되는 동안 초식률이 감소된다면 경쟁력은 낮지만 초식에 저항 성이 높은 종으로부터 경쟁력은 높지만 초식에 저항성이 적은 종으 로 식물의 천이 계열이 형성될 것이다. 6 요약 정착. 경쟁, 일시적인 동태, 초식, 그리고 그 밖의 다른 과정들이 천이 과정의 동태에 영향을 줄 수 있다. 이러한 과정들의 상대적인 중요성은 서식처에 따라 다를 수 있다. 이러한 이유만으로도 모든 천이 과정을 설명할 수 있는 어느 하나의 이론을 찾아내기는 불가 능하댜 그러나 서식처에 따라 이렇게 차이가 있음에도 불구하고 모 든 천이 과정에 공통적인 몇 가지 특징이 있다. 최초의 우점종은 키 가 작은 초본식물이며, 이들은 점점 키가 큰 목본성 식물에 의해 연 속적으로 교체된다. 이러한 유사성은 모든 천이 과정이 광량이 많은 서식처에서 시작되고, 광량이 많은 서식처에는 키가 작은 식물의 경 쟁력이 높은 데 그 원인이 있는 것 같다. 이러한 유사성이 있지만 이 장에서 제시된 이론은 천이 과정 중 에 우점하는 실제 종 계열은 1 차 천이, 척박한 토양에서 진행되는 2 차 천이. 그리고 비옥한 토양에서 진행되는 2 차 천이 사이에 차이가 있어야 하는 것으로 예측된다. 그 이유는 이러한 각각의 천이 과정 이 영양염류와 햇빛의 이용도 면에서 차이가 있고, 각 종의 형태와 생리가 특정한 영양염류와 광 이용도를 갖는 서식처에서 경쟁력이 높도록 제한하기 때문이다. 더구나 자원 이용도가 장기적으로 서서
히 변화하는 것은 1 차 천이에 더 중요한 원인이 될 수 있으며, 경쟁 적 교체의 일시적인 동태는 초기에 영양염류가 풍부한 기질에서 더 중요한 원인이 될 수 있다. 매우 비옥한 토양에서 진행되는 전체적인 천이 계열은 경쟁적 교 체의 일시적인 동태일 수가 있다. 척박한 토양에서는 초기의 동태가 주로 일시적인 동태일 수 있으며, 후기의 동태는 토양에 영양염류가 축적되는 기간에 의해 더 큰 영향을 받을 수 있다. 그러나 천이의 동태를 결정짓는 이러한 힘은 실제 모든 천이 과정에서 정착이나 초식 등과 같은 다른 과정들과 상호 작용을 한다. 서식처에 정착할 가능성이 있는 종 자원도 천이 과정의 동태를 결정하는 중요한 요 소가된다 . 천이는 다원인에 의한 과정이지만 서로 다른 지역에서의 천이 과 정이 유사성을 갖는 것은 식물의 분배 패턴을 결정하는 제약과 이 러한 제약이 경쟁의 동태에 미치는 효과 때문인 것 같다. 이 제약들 을 통해 모든 천이 과정은 적어도 초기에는 키가 작은 식물에서 키 가 큰 식물로 변화되는 것으로 예측할 수 있다. 또한 2 차 천이 과정 은 초기에는 신속히 생장하는 종에서 생장이 느린 종으로 변화되는 것으로 예측된다.
제 8 장 미네소타 모래평원에서의 2 차 천이 1 서론 1982 년에 나는 Cedar Creek Natu r al Histo r y Area 에 있는 휴경지의 식생 구성과 종 다양성 그리고 식생의 동태를 조절하는 요인을 규 명하기 위한 일련의 관찰과 실험 연구를 시작하였다. 이 장에서는 지금까지 밝혀진 이 연구의 결과를 이용하여 이 책에서 제시된 많 은 아이디어들을 평가하려고 한다. 이 장에서 제시된 자료 중에는 이 미 부분적 으로 발표된 것도 있지 만 (T il man 1983, 1984, 1986a, 1986b, 1987a, Inouy e et al. 1987a, 1987b), 대부분의 자료는 여기에서 처음 제시되는 것들이다. 본인이 수행하고 있는 실험은 1,000 개 이 상의 지소와(대부분 4mx4m) 다양한 처리구가 포함되어 있고, 그 중 극히 일부만을 여기에서 설명한다는 것을 먼저 밝혀두고 싶다. CedarCreek 은 대략 13,000 년 전에 굽이쳐 흐르는 빙하천에 의해 퇴적된 넓은 모래평원에 위치한다(그립 8.1 ). 1800 년대 중엽 유럽인
 A. 미네소타 동중부의 토양
A. 미네소타 동중부의 토양
그림 8.1 (A) 미네소타 동중부의 토양 . 그림에 나타나 있는 지역은 대략 85kmx 85km 이다. 밝은 색으로 된 넓은 지역은 Anoka 모래평원으로 여기에 Cedar Creek Natu ral Histo r y Area 가 위치하고 있다. Cedar Creek Natu ra l Histo r y Area 의 위치는 〈+〉로 표시되어 있다.
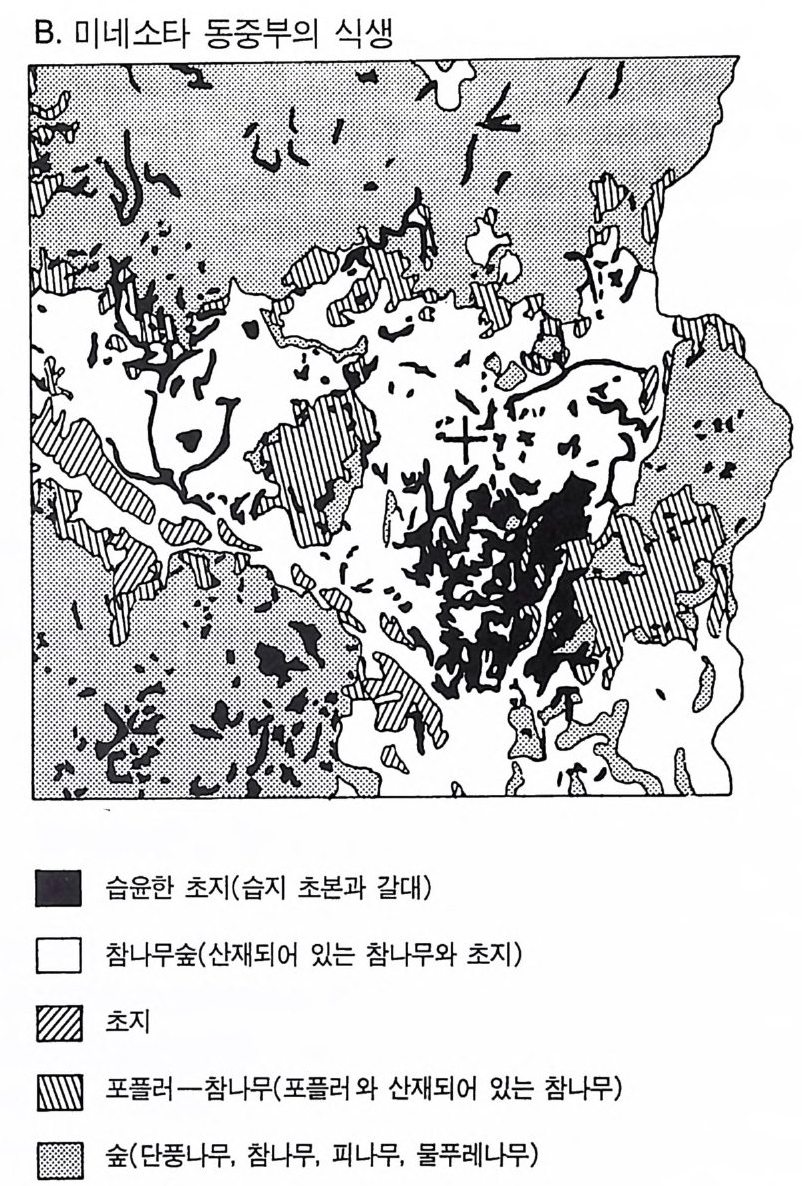 B. 미네소타 동중부의 식생
B. 미네소타 동중부의 식생
그림 8.1 (B) F.J . Marchner (l 930) 의 〈미네소타의 원식생〉의 지도에 근거를 둔 정 착 이전의 이 지역 원식생 .
들이 정착하였을 때 이 모래평원 고지의 식생은 초지와 참나무사바 나, 그리고 참나무림이 혼합되어 있었다 (Gr ig al et al. 1974, Til m an 1983, I984, 1987a). 이 고지대에 형성된 토양은 배수가 잘되는 사토 로 되어 있어 유기물 함량이 낮고 질소 함량도 낮았다. 근처에 있는 빙력토에서 형성된 토양과 남쪽 및 서쪽에 있는 바람에 의해 퇴적 된 황토에서 형성된 초지 토양은 유기물 함량과 질소 함량이 2-5 배 나 많았다. 1890 년대에서 1910 년 사이에 CedarCreek 의 고지대 식생 은 대부분 완전벌목이 되고 농경지가 형성되었다. 1930 년부터 현재 까지 이 농경지의 대부분은 휴경지로 전환되어 2 차 천이가 진행되 었으며 , 그 동안에는 산불이 없었다. 정착 이전에 이 지역은 1-4 년 마다 한 번씩 산불이 발생했었다 (Gr i mm 1984). 정착과 함께 산불이 억제되어 전에는 초지이던 지역에 참나무사바나가 확산되었고, 참나 무사바나 지역은 수관층이 더 밀폐된 참나무숲으로 변화되었다 (Whit e 1983). 2 천이 계열 새로 형성된 휴경지의 처음 우점종은 돼지풀, 마디풀, 금강아지풀, 망초, 명아주와 같은 1 년생 식물들이다. 이러한 1 년생 식물들은 보 통 지표면의 10-40% 를 점유하였고, 수명이 짧은 다년생 식물인 긴 겨이삭과 같은 초본이 5-10% 를 점유하였다. 나머지는 식물이 없는 맨땅이었다. 천이 과정의 처음 60 년 동안에 1 년생 식물의 피도는 감 소하고 다년생 식물의 피도는 증가하였으며, 식물이 덮이지 않은 맨 땅의 면적은 줄어들었다(그림 8.2, Inouy e et al. 1 987). 우점도가 점차 중가하는 다년생 식물은 Schiz a chy ri u m scop a riu m (little blueste m ), 바랭이새류, 인디안그라스와 같은 초지 고유의 벼과
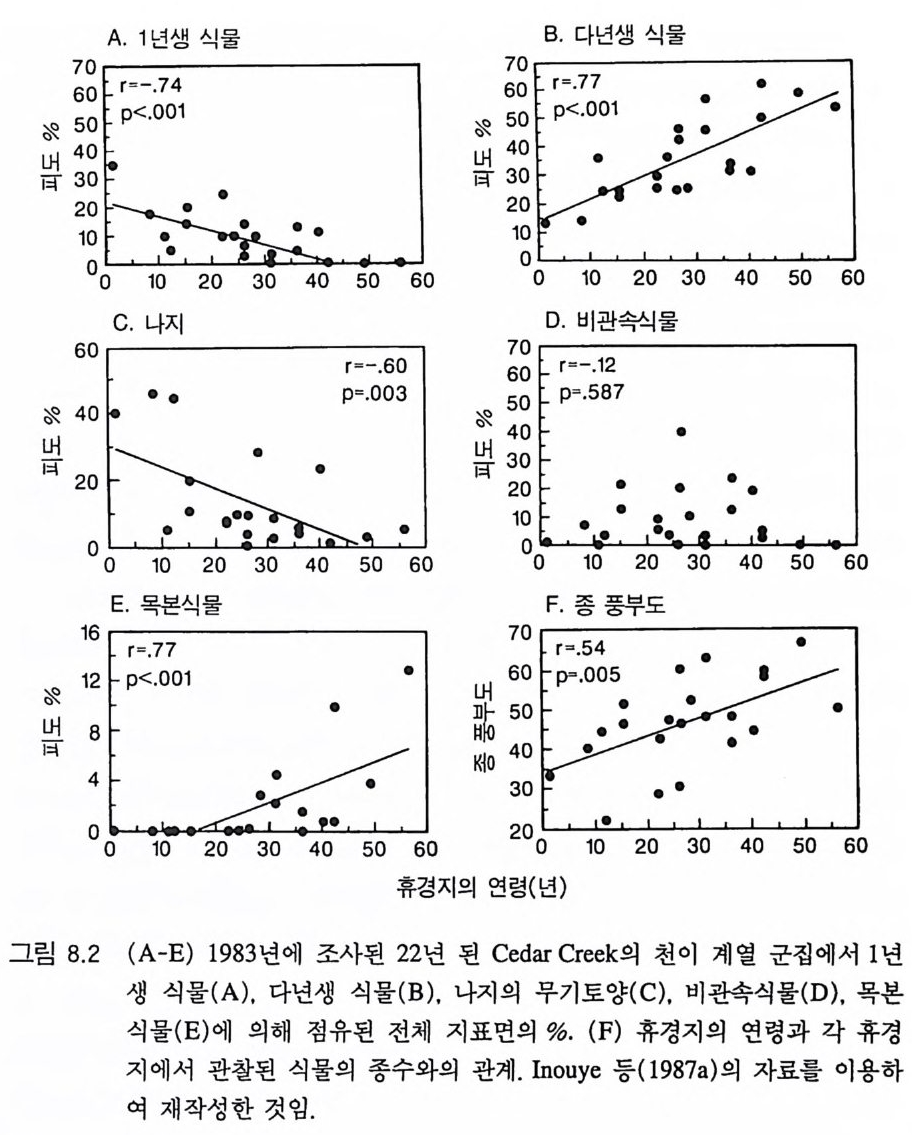 ~ 6500 Ar.p =<1- . 0년.7 0생41 식물 殺 6so0 E B『p .= < .다 .7 07년0 1생 삭울 • ·. /• •
~ 6500 Ar.p =<1- . 0년.7 0생41 식물 殺 6so0 E B『p .= < .다 .7 07년0 1생 삭울 • ·. /• •
식물과 Carex muhlenberg ii와 같은 사초과식물, 그리고 타지역에서 이주한 왕포아풀 등이었다. 쌍자엽식물은 천이 과정에서 우점도가 낮았지만, 초지의 쌍자엽식물은 휴경지에서 수도가 증가하였다. 비 관속식물은 주로 지의류이었는데 새로 형성된 휴경지나 오래된 휴
경지 모두에서 매우 적었고, 20-30 년 된 휴경지에서 최대 수도를 기 록하였다(그림 8.2). 검은딸기와 같은 덩굴성. Rosa arkansana 와 같은 작은 덤불 관목, bur oak, red oak 그리 고 스트로브잣나무와 같은 목 본성 식물은 15 년 이하의 휴경지에는 나타나지 않고 시간이 지남에 따라 피도가 서서히 증가하였다(그림 8.2). 60 년 된 휴경지에서도 목 본식물의 피도는 기껏해야 13% 정도였고, 이것도 대부분 교목이 아 닌 덩굴성 그리고 관목에 의한 것이었다. 대부분의 휴경지가 그 규 모가 작고 (5ha 혹은 lOha 이하), 벌목이 안된 숲으로 둘러싸여 있는 데도 이와 같이 오래된 휴경지에서 목본식물의 피도가 낮은 것은 매우 이상한 현상이다. 이러한 휴경지에서는 종 구성의 변화와 함께 종수도 증가하였는데, 새로 형성된 휴경지에는 평균 약 30 종이던 것 이 가장 오래된 휴경지에서는 약 60 종으로 증가하였다(그림 8.2). CedarCreek 의 휴경지에서 이러한 종 구성의 변화나 다양성의 변 화가 일어나는 원인은 무엇인가? 여기에는 분명히 여러 가지 기작 이 관련되어 있다. 휴경지에서 관찰되는 천이의 동태는 식물종에 따 른 정착률 또는 생장률의 차이에서 비롯되는 일시적인 동태일 수도 있다. 식물은 종에 따라 경쟁력이 서로 다를 수 있으며, 이들의 경쟁 력은 서식처의 조건이 변화됨에 따라 달라질 수 있다. 또한 초식동 물의 밀도가 천이 과정에 따라 변할 수 있으며. 이것이 천이의 패턴 에 영향을 준다. 1 년생 식물이 2 차 천이 초기에 우점하는 이유는 그것들이 농경지 가 휴경지로 될 때 이미 높은 밀도로 존재하고 있었기 때문이다. 흙 을 자주 갈아엎고 농사를 지은 결과 생장률이 빠르고 생활사가 짧 은 식물들이 선택된 것이다. 따라서, 휴경지가 형성될 때 1 년생 식 물의 종자가 이미 토양 속에 많이 존재하고 있었다. 나는 갓 형성된 휴경지에서 1 년생 식물의 실생 밀도가 1,000 개체 /m2 이상인 것을 흔 히 관찰하였다. 그러나 다른 지역에서의 천이와는 달리 1 년생 식물
들이 수십년 동안 식생의 주요 구성원으로 남아 있었다. 20 년 된 휴 경지에서 1 년생 식물은 전체 식피 면적의 40% 이상을 차지하였고, 40 년 된 휴경지에서는 20% 이상을 차지하였다(그림 8.2). 교란이 잦으면 1 년생 식물의 피도가 증가하는데, 이 지역의 휴경 지에서는 큰 교란도 없었다. 따라서 천이 초기에 1 년생 식물이 우점 하는 것은 처음부터 이들의 종자가 많았고 생장률이 높은 것으로 설명이 가능하지만 40 년 이상이 지날 때까지 식생의 주요 구성원으 로 남아 있다는 사실은 설명이 쉽지 않다. 40 년 동안에 많은 다른 종들이 침입하여 정착하였지만 1 년생 식물을 교체하지는 못하였다. 1 년생 식물이 교체되지 않았을 뿐만이 아니라 전체 식물의 피도는 60 년의 천이가 있은 후에도 60% 정도밖에 되지 않았다. 가장 오래 된 휴경지에서도 지표면의 약 40% 는 맨땅이거나 얇은 낙엽층으로 덮여 있었다. 천이 초기 과정인 5-l0 년 된 휴경지에서 목본식물이 드문 것은 이들의 분산율이 느리고 생장률이 느리기 때문에 나타나는 일시적 인 효과로 설명할 수 있지만 오래된 휴경지에서 목본식물이 우점하 지 못하는 원인을 설명하기는 어 렵다. Cedar Creek 휴경지에서 참나 무 실생은 혼하지만 유식물은 드물다. Wh it e(1983) 는 Cedar Creek 사바나에 있는 buroak 가 대부분 40 년 이내에 성숙한다는 것을 알았 다. 따라서 정착률과 생장률의 차이를 가지고 CedarCreek 에서의 천 이 초기 패턴의 일부는 설명할 수 있지만, 이곳에서의 천이 변화율 이 너무 느려서 이 두 가지만으로는 우리가 관찰할 수 있는 패턴을 다 설명할 수는 없다. Cedar Creek 에 있는 22 개 의 휴경지 에서 우리 가 조사한 자료들을 분석한 결과 우리는 휴경지의 나이와 토양의 질소가 모두 천이 패 턴과 중요한 관련이 있음을 알았다. 우리는 시간이 식물의 분산율과 정착률의 차이에 영향을 주어 특정한 휴경지에 존재하는 종을 결정
하는 데 중요한 역할을 하며. 반면에 자원을 기초로 한 상호 작용은 휴경지 내의 특정한 지소에 존재하는 종을 결정하는 것으로 결론지 었댜 따라서 이 연구의 결과는 미네소타 모래평원에서의 2 차 천이 의 패턴이 종의 분산과 기타 시간 의존적인 과정들. 그리고 주요 토 양 자원, 특히 질소 이용도의 변화에서 비롯된다는 것을 시사하고 있 다(I nou y e et a l. 1987a. p. 25 ) . 여 러 가지 과정 중에 서 어 느 하나만으 로는 우리가 관찰한 패턴을 충분히 설명할 수 없다. 이러한 천이 패턴의 일부를 초식동물에 의한 영향으로 설명할 수 있다. Cedar Creek 의 주요 초식동물로는 poc ket gop he r. 흰꼬리사슴, 들쥐, 그리고 몇 종의 메뚜기 등이 있다. CedarCreek 에서 수행된 초식 동물 배제 실험에 의하면, 이들 각각의 초식동물이 지상부 식물량에 영향을 줌을 알 수 있다 (Hun t l y et al., 원고 준비중). 그러나 모든 초식 동물을 4 년 동안 배제한 경우에서도 식물량의 증가는 10% 정도였다. 대부분의 초식동물이 먹이에 대한 선택성을 가지고 있었지만 초식동물 의 밀도가 너무 낮아 토양 영양염류 구배에 따른 식물종의 상대적인 수도에 영향을 주진 못하였다. 사슴에 의한 새싹 소비가 많았지만 사슴 을 배제한 실험에서도 목본식물의 길이 생장에 약간의 영향만 미치는 것으로 밝혀졌다. 따라서 초식동물들이 식물의 생물량에 영향을 주긴 하지만 그것들이 천이 과정에 주영향력을 갖는 것은 아닌 것 같다. 예를 들어, pocke tg o p hers 는 자신들의 섭식굴을 파는 과정에서 맨 땅에 둔덕을 만든다. Go p her 가 만든 둔덕에는 처음에 1 년생 식물이 우점한다 (T il man 1983). 그러나 go p her 에 의해 교란되는 면적은 정상 적인 휴경지에서 지표면의 1-2% 정도로 그 교란율이 낮기 때문에 장기간에 걸쳐 1 년생 식물이 높은 밀도로 유지되고 목본성 다년생 식물이 적은 이유를 토양의 교란율만으로 설명할 수는 없다. 또 하나의 가능한 설명으로는 토양의 영양염류 수준을 들 수 있 다. Cedar Creek 에 있는 휴경 지 와 원래 부터 있던 참나무사바나에 서
수행된 영양염류를 첨가하는 많은 실험에 의하면 이 지역의 전체 천이 과정에서 가장 부족한 토양 자원은 질소라는 것이 밝혀졌다 (Til m an 1983, 1984, 1987a, 그림 8.3 ). 질소 첨가는 지상부 생물량을 현저히 증가시켰지만 질소를 제의한 다른 모든 영양염류를 첨가하 였을 때에는 대조구에 비해 생물량의 증가가 결코 현저하게 증가하 지 않았다 (T i lman 1987a). Cedar Creek 휴경지의 시간에 따른 조사에서 우리는 22 개의 상이 한 지역에서 2,300 개의 방형구를 이용하여 토양과 식생을 채집하였 다(I nou y e et al. 1987a). 이 토양들을 분석 한 결과 토양의 질소 함량 이 지소의 나이에 따라 현저히 증가하였다(그림 8.4 ). 새로 형성된 휴경지의 토양 질소는 평균 370m g/kg이었는데, 이 값은 휴경지 주 변에 있는 삼림토양 질소 함량의 1/3 정도였다. 휴경지의 나이에 따 른 질소 함량의 증가율로 보면. 휴경지의 질소 함량이 주변에 있는 교란되지 않은 참나무사바나나 참나무림 수준으로 회복되는 데에는 100 년 정도가 필요한 것으로 판단된다. 휴경지 토양의 질소 함량이 증가함에 따라 관속식물의 전체 피도 는 증가하고 비관속식물의 피도는 감소되었다(그림 8.5). 1 년생 식물 과 이입된 식물의 수도는 이 일시적인 질소 구배에 따라 감소하였 고. 다년생 초본과 목본의 피도는 증가하였다(그림 8.5). 따라서 휴 경지의 나이와 뚜렷한 상관을 갖는 많은 변수들이 휴경지의 질소량 과도 상관이 있었다. 1984 년에 이 휴경지에서 채취한 19 개의 표본 은 전체 지상부 생물량이 토양의 질소량에 따라 현저히 증가하였고. 지표면에 입사되는 광량은 전체 지상부 생물량과 음의 상관이 있음 울 보여주었다(I n?u y e et a l. 1987b). 또한 1986 년에 이 지역의 16 개 지소에서 같은 트랜섹트를 따라 채집한 토양의 이용 가능한 질소의 수준이 (NH4+N03) 가장 나이가 많고 질소가 풍부한 휴경지에서 높았다(그림 8.6, Til ma n 1987b).
 400
400
그립 8.3 4 개의 서로 다른 휴경지에서 수행된 영양염류 첨가실험에서 9 개의 각 처 리구로부터 수확한 평균 지상부 생물량 . 각 휴경지에서 처리구당 4 반복을 두어 전체 실험구는 144 구였다. 휴경지는 T i lman(1987a) 에 의해 자세히 설명된 바 있는데 A 는 14 년 된 휴경지. B 는 25 년 된 휴경지, C 는 48 년 된 휴경지. 그리고 D 는 토착 참나무사바나이다 . 처리구는 여러 가지 영양엽 류를 한 번에 1 가지씩 첨가한 것이다 . 질소는 질산암모늄, 인은 인산나트 륨, K 는 KCl. Ca 은 CaC03, M g은 Mg S 04, S 는 Na2S04, Tr 은 미 량 금속의 혼합체 (CO, Mn, Mo, Cu, Zn), HP 는 물로 6 월, 7 월, 8 월 동안에 1 주일에 2cm 의 비율로 처리하였다 . Ct!은 대조구이다. 이들 휴경지에서 지상부 생 물량을 항상 증가시키는 영양염류는 질소였다 .
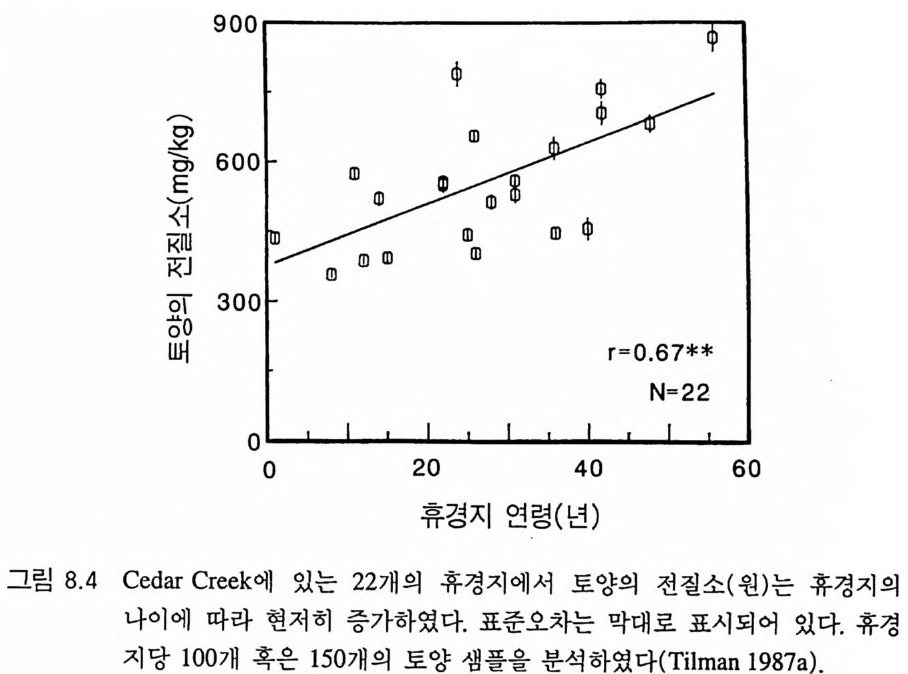 /--` 900 Ir ¢
/--` 900 Ir ¢
더구나, Pasto r 등(1 987) 은 휴경지 4 개 지역에서 채취한 토양의 질소 무기화율이 토양의 전질소 함량과 양의 상관이 있음을 밝혔다. 따라 서 CedarCreek 에서 2 차 천이의 한 요인은 시간에 따른 질소 공급률 의 증가이다. 질소 공급이 증가함에 따라 가장 오래된 휴경지에서는 전체 생물량은 증가하고 지표면에 입사하는 광량은 감소하며, 토양 의 이용 가능한 질소량은 증가한다. 식물종이 질소와 햇빛에 대한 경쟁력에서 분화가 일어나고 이 분 화가 천이의 원인이라면 질소가 풍부한 오래된 휴경지에서 이용 가 능한(추출 가능한) 질소량의 증가와 입사되는 광량의 감소는 예측할 수 있는 것이다 (T il man 1982, 1985 그림 7.1 ). 만약 햇빛과 같은 다른 자원이 제한되지 않는다면 질소가 더 많은 휴경지에서 이용 가능한 질소의 수준이 더 높은 것을 설명하기가 어렵다. 질소가 유일한 제 한 자원이라면, 이용 가능한 질소량이 적은 신생 휴경지에 생존할
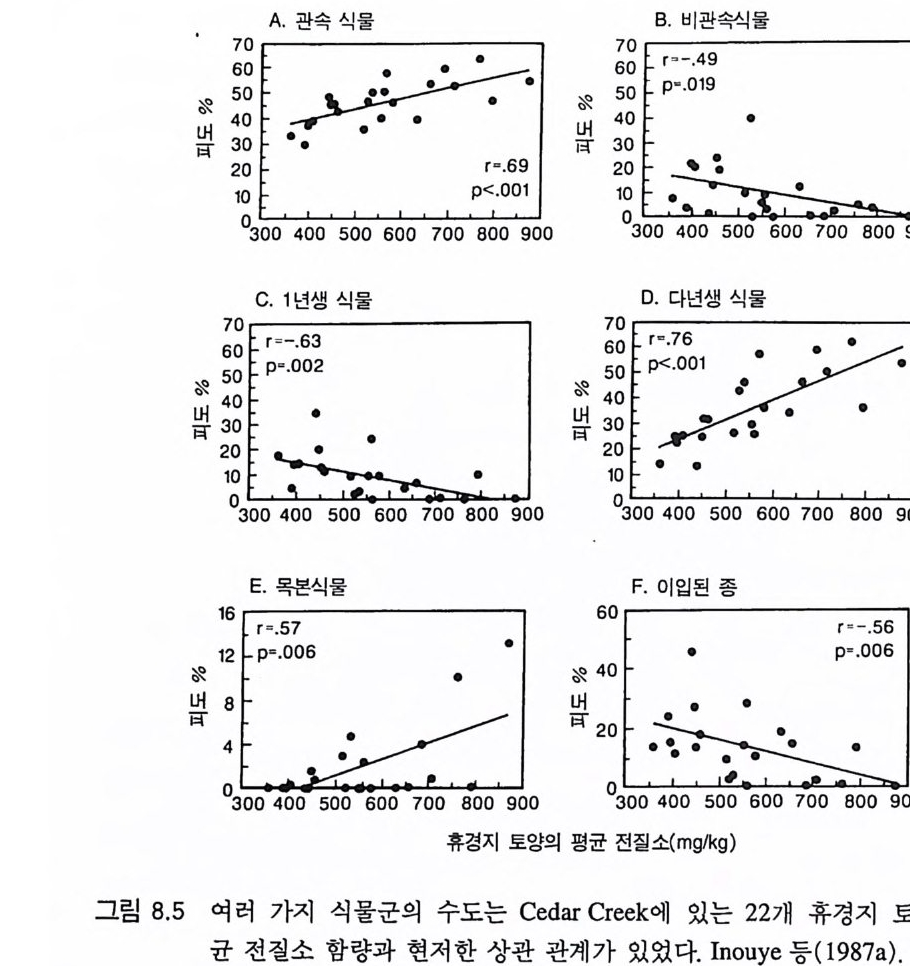 〈A 관속 식물二 0 0 버603 0i 08r . -4 -비0. 40관9 속50 식0• 물 600 700 800 900
〈A 관속 식물二 0 0 버603 0i 08r . -4 -비0. 40관9 속50 식0• 물 600 700 800 900
수 있는 종은 이용 가능한 질소량이 많은 오래된 휴경지에서 수도 가 증가하여야 하며, 토양의 질소 수준을 자신들이 못살 정도의 낮 은 수준으로 감소시킬 때까지 수도가 계속 증가하여야 한다(그림 2.1 참조) . 이용 가능한 토양 질소의 증가는 지표면에 이입되는 광량의 감소 를 수반한다. 지표면에 입사되는 광량이 전광량의 10% 정도만 낮아 져도 제한요인이 될 수 있다. 그러나 우리는 Cedar Creek 의 휴경지
 11..58 rrA . nr== 01 .67 3 •
11..58 rrA . nr== 01 .67 3 •
그림 8.6 (A) Cedar Creek 에 있는 16 개의 휴경지에서 이용 가능한 질소(NH.i -N 과 N03-N) 는 휴경지의 토양 전질소 함량에 따라 증가하였다. 가장 비옥한 토양과 가장 오래된 휴경지에서 가장 많은 증가가 일어났다 . 이 회귀식이 유의성을 갖도록 한 것은 바로 가장 오래된 두 휴경지였다. (B) 이용 가 능한 질소는 휴경지의 나이에 따라서도 증가하였다. 각 접은 휴경지의 평 균 전질소 함량 혹은 특정한 휴경지에서 이용 가능한 질소의 평균을 나타 낸다. 1986 년 여름에 T ilm an 에 의해 수집된 미발표 자료`
에 서 피 음 효과에 대 한 실 험 을 하지 않았다. 따라서 Ced 紅 Creek 에 서 햇빛의 잠재적인 역할에 대한 다음의 설명은 잠정적인 것으로 간주되어야 한다. 3 이론을 통한 검증 가능한 예측들 CedarCreek 휴경지에서 토양과 식생의 조사 결과는 시간뿐만이 아니라 질소와 햇빛에 대한 식물의 경쟁도 천이 패턴의 많은 부분 울 설명할 수 있음을 보여주고 있다 . CedarCreek 에서 질소와 햇빛 이 가장 중요한 제한 자원이라고 가정하면 이 책에서 제시된 이론 으로 다음과 같은 예측이 가능하다. (1) 질소 공급률의 중가는 전체 식물량의 증가와 지표면에 이입되 는 광량의 감소를 초래 한다. (2) 피음에 의해 광량이 감소되면 전체 식물량이 감소되고 이용 가능한 토양의 질소 수준이 증가된다 . . (3) 질소나 햇빛 이용도의 변화는 식물종의 상대수도의 변화를 초 래한다 . (4) 질소 첨가에 대한 초기 반응은 뿌리:줄기의 비가 낮은 종, 즉 최종 우점종에 비해 최대 생장률이 높은 종에 의한 일시적인 우점 기간이어야 한다. (5) 결국, 식물은 실험적인 질소 구배에 따라 분리되어야 하는데, 식물이 자연 상태의 질소 구배에 따라 나타나는 순서와 인위적으로 조성한 질소 구배에 따라 나타나는 순서가 같아야 한다. (6) 자연적 혹은 인위적으로 조성된 질소 구배상에서, 질소가 풍 부한 쪽에서 우점하는 종은 질소가 적은 쪽에서 우점하는 종에 비 해 성숙하였을 때 키가 크고, 줄기에 대한 분배량이 많고 뿌리에 대
한 분배량이 적어야 한다. (7) 처음에는 종 구성이 서로 다른 식물 군집들도 질소 공급률이 비슷해지면 종 구성도 비슷하게 수렴되어야 한다. (8) 각 식물종이 질소가 부족한 토양에서 단독으로 자랄 때, 생산 력 구배상의 질소가 부족한 쪽에서 우점하는 종은 질소가 많은 쪽 에서 우점하는 종에 비해 질소가 낮울 때 더 신속히 자라야 하며, 평형 상태에서 이용 가능한 토양 질소를 더 낮은 수준으로 감소시 켜야한댜 (9) 평형에 도달되면, 질소 첨가시 나타나는 식물종의 상대수도의 변화는 광량을 감소시킬 때 나타나는 변화와 같아야 하며, 광량을 증가시키면 질소를 제거할 때와 같은 변화가 나타나야 한다. 4 실험에 의한 검증 우리는 위에 열거한 예측들을 검증하기 위한 일련의 실험을 수행 하였다. 주요 실험 중 하나는 7 가지로 질소 구배를 달리하는 것이었 다(그림 8.7, 8.8, Til m an 1987a). 각 구배는 9 가지 서로 다른 처리구에 5-6 개의 반복구를 설치하였다. 처리구는 대조구(처리구 I) 와 8 개의 처리구로 되어 있는데 모두 기본적으로 P, K, Ca,Mg , S 그리고 미 량원소를 공급하였고 연간 첨가하는 질소만 달리하였다(그림 8.9). A 에서 H 까지 기호를 붙인 이 8 개의 처리구에 0-27g /m2 • y r 의 질소 를 공급하였는데, A 는 질소를 공급하지 않았고 H 에는 가장 많은 질 소를 공급하였다. 이 실험은 휴경 후 15, 25 그리고 48 년 된 3 곳의 휴경지와 토착 참나무사바나에서 1982 년에 시작되었다• 각기 다른 천이 과정에 있 는 휴경지에 두 개의 상이한 질소 구배를 만들었는데, 하나는 현존
 . ' ..-.
. ' ..-.
그림 8.7 1983 년에 14 년 된 휴경지 A 에서 여러 가지 실험구를 찍은 항공사진. 작 은 실험구는 4mx4m 이며 lm 넓이의 보도로 분리되어 있다. 중앙부의 왼 쪽에 일부가 보이는 큰 실험구는 20mx50m 이다. 이러한 실험구간의 주 요 차이점은 질소 첨가율이다. 이 책에서 설명된 두 개의 실험적 질소 구 배는 울타리 안쪽에 있다. 맨 위쪽에 있는 54 개의 실험구는 각 실험구로 나누고 비료를 주기 전인 1982 년에 교란되었다. 맨 밑에 있는 54 개의 구 는 교란되지 않았지만 1982 년에 비료를 주기 시작하였다. 두 지역은 모두 포유 초식동물의 침입을 막기 위해 울타리를 설치하였다. 이 지역은 삼림 에 의해 둘러싸여 있지만 재생된 나무는 거의 없었다 . 이 지역 토양의 전 질소 함량은 주변의 교란되지 않은 숲의 약 40% 수준이었다.

그림 8.8 48 년 된 휴경지 C 에 있는 실험구를 1983 년에 찍은 항공사진. 작은 실험구 는 4mx4m, 큰 실험구는 20mx50m 이다 . 이 지역에서 재생된 나무는 거 의 없었고, 이 지 역 토양의 전질소 함량은 주변 숲의 약 65% 수준이 었다 .
 30
30
하는 휴경지 식생에서, 다른 하나는 1982 년 봄에 가래로 토양을 철 저히 갈아엎어 새로 교란시킨 곳에 만들었다. 토착 참나무사바나 지 역에 있는 초지의 현존하는 식생에 하나의 질소 구배구를 설정하였 다 . 이러한 각각의 구배구에는 그 내부에 포유 초식동물을 배제시킨 구가 포함되 어 있다 (T i lman 1987a). 4-1 생물량, 광 그리고 질소 모든 휴경지의 교란된 곳과 교란되지 않은 곳에 설정한 질소 구
배 에 서 식 물의 반응에 공통점 이 많았다 . 1982 년부터 1986 년까지 매 년 지상부 식물량이 증가하였으며(그림 8.10), 지표면에 이입되는 광 량은 감소하였다 ( 그림 8.11). 그리고 식생의 키는 질소 구배의 낮은 곳에서부터 높은 곳으로 갈수록 증가하였다(그림 8.12). 따라서 이 러 한 실험 결과는 질소가 제한 자원이라는 것과, 질소 공급률의 증가 는 식물의 생물량 증가를 초래하여 햇빛의 이용도를 감소시킨다는 것을 보여준다. 이러한 변화는 이것과 동시에 나타나는 식생의 키의 변화와 함께 예측 l 과 6 에 잘 일치된다 . 이 이론이 예측하고 있는 또 하나의 중요한 사항은 이용 가능한 질소의 양이 질소 공급 구배를 따라 증가해야 한다는 것이다. 이용 가능한 질소의 양은 질소를 첨가하는 기간인 매년 이른 봄과 늦은 봄 후에 즉시 증가하여야 한다. 그러나 이론은 이용 가능한 질소의 양이 생육기간 내내 높게 유지되는 것으로 예측하고 있다. 이것을 검증하기 위해서 1986 년에 각 질소 구배구에서 생물량이 최고에 이 르는 하절기에 토양을 채집하였다. 이용 가능한 NH 4 +N03 는 토양을 채집한 직 후 야외에서 2MKC1 로 추출하였다. 이용 가능한 질소는 이 7 개의 실험 구배에서 질소 공급률에 따라 현저히 증가하였다. 7 개의 실험구 결과를 평균할 때 토양의 이용 가 능한 질소 (N) 는 질소를 첨가하지 않은 구의 0 ,4 m g/kg에서 질소를 가장 많이 공급한 구의 5 .1 m g/kg으로 증가하였다. 예측한 바와 같이 이용 가능한 질소와 지표면에서 이용 가능한 광량은 7 개의 구에서 모두 음의 상관을 보였다 . 25 년 된 휴경지와 토착 참나무사바나 지 역의 교란되지 않은 실험구의 결과가 그림 8 . 13 에 나타나 있다. 처리 구배에 따른 결과나 모든 지역의 결과를 평균할 경우 모두 질소 첨 가율의 중가는 이용 가능한 질소의 양을 증가시키고 지표면에 입사 되는 광량을 감소시켰다(그림 8.14).
 1982 1983 1984 1985 1986
1982 1983 1984 1985 1986
그림 8.1 0 1982 년부터 1986 년까지 매 년 각각의 7 개의 실 험 구배에서 전체 지상부 식 물량과 질 소 첨가율과의 관계. 각 막대 그래프의 값은 처리구에서 6 반복구의 근평균이:다 . 막군대 그\ 래프\의\ 첫:번4째 :것:은 곳이며, D 는 1982 년 이 연구를 시작할 때 토착 사바나였다. 교란되지 않은 구 배는 기존의 식생에 만들어졌고, 교란 된 구배는 1982 년 실험을 시작할 때 철저히 가래로 갈아엎은 토양에 만들 어졌다. 처리구의 순서는 그림 8 . 9 와
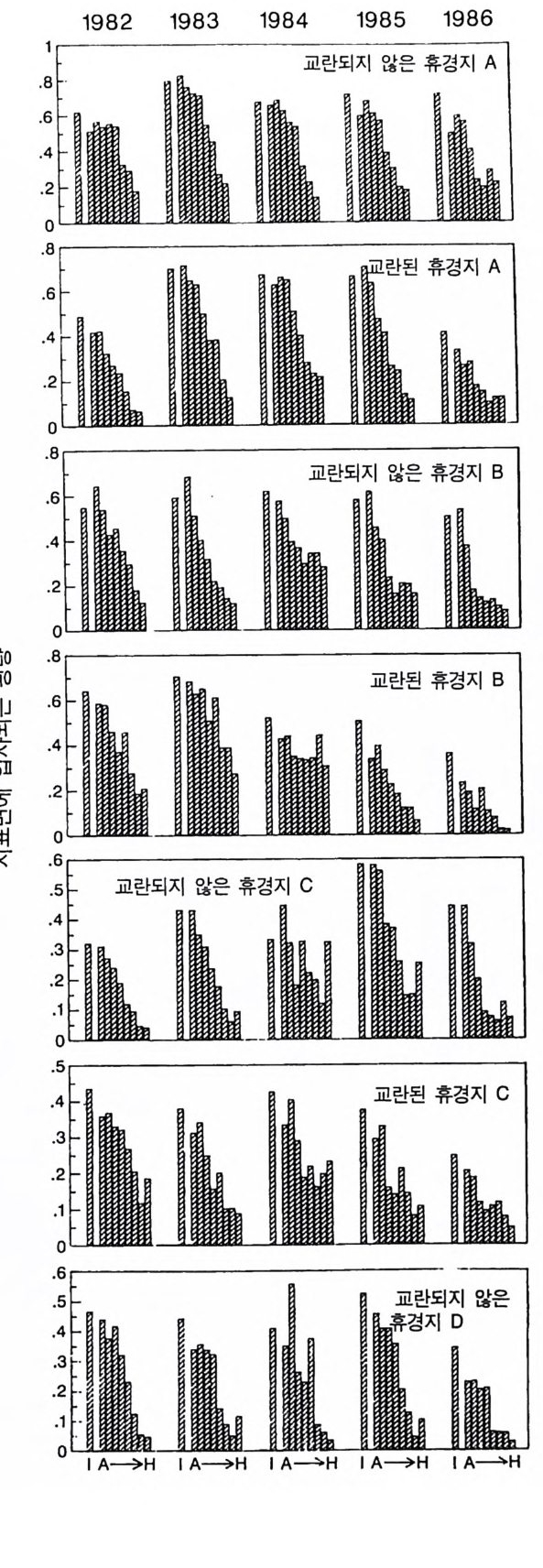 1982 1983 1984 1985 1986
1982 1983 1984 1985 1986
그림 8.11 1982 년부터 1986 년까지 Cedar Cree 에 있는 7 개의 질소 첨가 구배에서 l 년 지표면에 입사되는 광량과 질소 ·: 배 사이의 관계 . 막대 그래프의 첫번 1 것은 1982 년에 질소 첨가에 대한 입, 광량을 보여주는 것이고, 그 다음 것? 1983 년의 결과이다. 교란되지 않은 • 배는 기존의 식생에 만들었고` 교란: 구배는 새로 갈아엎은 토양에 만들! 다. 지표면에 입사되는 광량은 지표' 에 도달하는 광합성 유효방사량의 1
 1982 1983 1984 1985 1986
1982 1983 1984 1985 1986
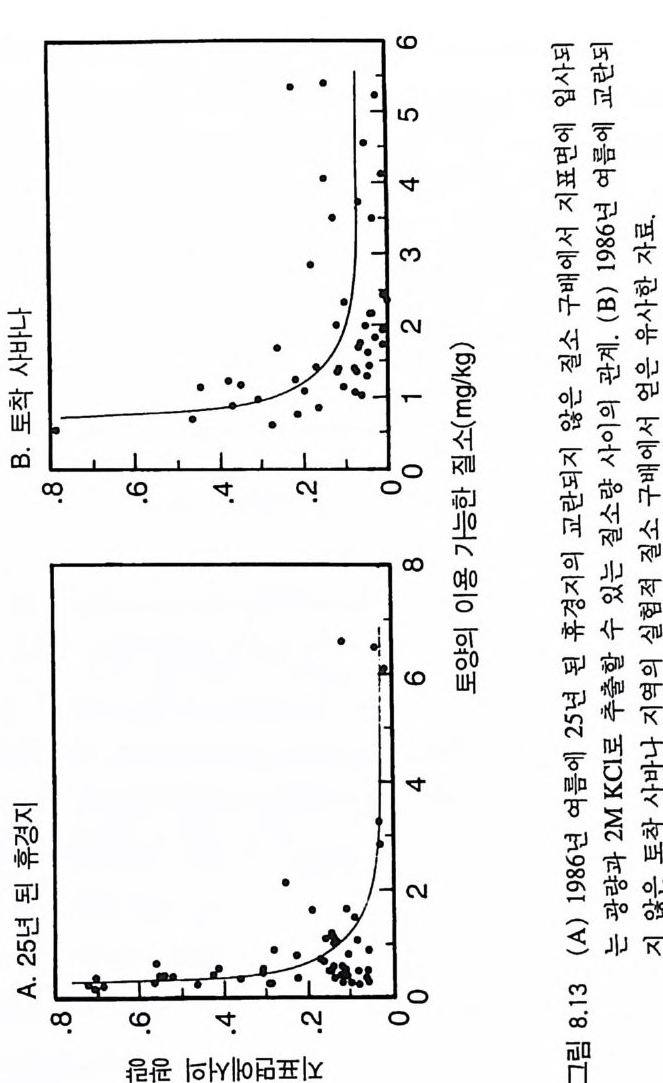
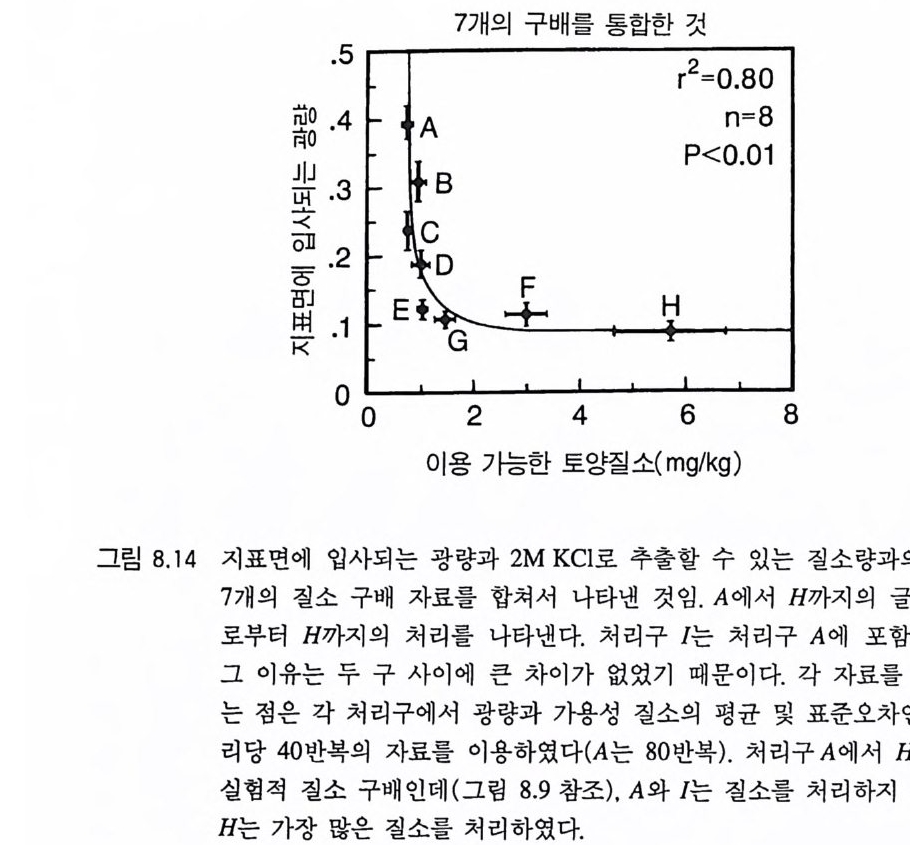 5437 개의2 구J배를 통합한 것
5437 개의2 구J배를 통합한 것
4-2 구배에 대한 종의 반응 앞에서 설명한 것과 같은 구배 실험에 대한 각 식물종의 반응도 이 이론의 검증에 이용된다. 실험이 시작된 첫해부터 보편종(전체 식물량의 2% 이상되는 종들)의 절대수도와 상대수도에 대한 질소 첨 가율의 효과가 매우 두드러지게 나타나기 시작하였다. 실험이 시작 된 첫해에 처음 시비 후 2-3 개월 후에 채집한 결과 57 종 중에서 29 종이 질소 첨 가율에 따라 생 물량이 현저 히 증가하였다 (T i lman 1987a). 첫해에 절대수도가 감소된 종은 하나도 없었다. 따라서 이들
종 중 절반 정도가 질소에 의해 제한되고 있었음을 알 수 있다. 나머지 절반에 해당되는 종이 무엇에 의해 제한되고 있다는 직접 적인 실험적 증거는 없지만, 이들이 제한을 받지 않았다는 증거는 많이 있다 이 실험에서 설정한 구배에는 영양염류를 전혀 받지 않 는 구(처리구 I) 와 질소를 제외한 모든 영양염류를 공급받는 구(처 리구 A) 가 있다. 1982 년부터 1985 년까지 매년 종 대 종 (s p ec i es-b y s p ec i es) 을 기준으로 수행한 이 두 처리구 사이의 분산 분석에 의하 면 시비를 하지 않은 대조구에 비해 P, K, Ca, Mg , S 그리고 미량원 소인 Cu, Co, Mo, Zn, Mn 의 혼합비료를 처리한 구에서 생물량이 증 가한 종은 전체 가능한 232 회 중 단지 7 회 밖에 없었다 (T i lman 1987a). 이 일곱 번의 경우도 지소나 연도 및 종이 각각 달랐다. 이 경 우들은 Ty pe I 오류일 가능성 이 높다. 이 러 한 자료들은 P, K, Ca, Mg , S 그리고 미량원소들이 보편종의 생장을 제한하지 않음을 시사 하고 있댜 이것은 물을 포함하여 여러 가지 자원을 단독으로 첨가 한 실험 결과와도 일치하였다(그림 8.3 ). 따라서 이러한 결과는 질소 에 의해 제한을 받지 않는 종은 물을 포함한 다른 자원에 의해서도 제한을 받지 않는다는 증거가 된다. 다른 부류의 증거는 햇빛이 이러한 구배상에서 중요한 제한 자원 임울 제시하고 있다. 질소 공급률의 증가는 지표면에 이입되는 광량 울 전체 입사광의 10% 로 감소시키는데, 시비를 하지 않은 대조구에 서 지표면에 도달하는 광량은 평균 40% 이다(그림 8.1 4 ). 이러한 광 량에서는 새로 정착하는 식물의 실생과 줄기가 햇빛의 제한을 받을 수 있다 (Bazzaz 1979). 더구나 생육계절 초기에 키가 커지는 종이 질 소가 많은 구에서 우점종이 되는 경향성이 강하다. 시간이 지남에 따라 보편종, 특히 다년생 식물은 7 단계의 질소 구 배상에서 분포 영역이 점점 뚜렷해진다 (T il man 1987a). 더구나, 각각 의 다년생 식물은 그것의 초기 수도에 관계없이 7 단계의 구배상에
서 그것의 반응이 점점 유사해진다(그림 8.21). I 년생 식물도 구배를 따라 분화되는데. 이들의 반응은 해마다 변이가 더 크다. 이 실험이 시작될 당시 14 년이 경과된 가장 나이가 어린 휴경지에는 1982 년에 많은 1 년생 식물과 소수의 다년생 식물이 있었다. 1984 년부터 토착 1 년생 식물인 돼지풀과 박하가 교란된 지역과 교란되지 않은 지역 의 질소 구배상의 질소 함량이 가장 낮은 쪽에서 상대수도가 가장 높았으며. 질소 구배를 따라 감소하였다 (T i lman 1987a). 장대냉이. 마 디풀, 망초 등과 같은 이입된 1 년생 식물들이 구배의 중간 부분에서 상대수도의 최대치를 보였다. 1986 년에는 두 종의 다년생 식물이 이 신생 휴경지의 교란된 지소와 교란되지 않은 지소에서 모두 우점하 였는데, Ag ro p yro n rep e ns(qu a ck gr ass) 는 질소가 많은 쪽에서, 왕포아 풀은 질소가 적은 쪽에서 우점하였다. 25 년 된 휴경지에 혼한 1 년생 식물은 교란된 지소에서는 일부가 출현하였으며, 1986 년에는 매우 드물었다. 가장 많은 다년생 식물로 는 Schiz a chyr i u m scop a rium , 왕포아풀, Ag ro p yro n rep e ns 둥이 었는데. 교란된 지소와 비교란된 지소의 질소 구배에 대한 반응이 비슷하였 다. Schiz a chy ri u m scop ar i um 은 25 년 된 휴경지에서 질소 구배에 따라 모두 감소하였다. 왕포아풀은 중간 정도의 질소 공급률에서 최대 수 도를 보였다. 질소가 많은 구에서는 A g rop y ronrep ens 가 우점하였다. 이 실험이 5 년째 되던 해에 48 년 된 휴경지에서도 비슷한 패턴이 나타났다. Schiz a chy ri u m scop ar i um 이 질소를 적 게 처 리 한 구에 서 우 점하였으며, 왕포아폴, Panic u m o lig osan t hes 는 중간 정도의 질소구배 에서, 그리고 A g rop y ronrep ens 는 질소를 많이 처리한 구에서 우점하 였댜 이밖에도 몇 종의 식물이 구배상에서 뚜렷한 분포역을 갖는 것으로 나타났다. Soli da g o nemora li s 는 질소를 적게 처리한 구에서 가장 많았고, Arte m i si a ludov ici ana 는 중간 정도의 처리구에서 가장 많았다(그림 8.1 5 ).
토착 참나무사바나 내에 있는 초지에는 교란되지 않은 식생에 하 나의 구배만 설정하였었다 . 보편종들은 분포역이 뚜렷하게 구분되었 는데, Sorgh astr u m nu t an s 는 질소가 적은 쪽에서 최대 수도를 보였다 (그림 8.1 5 ). 중간 정도의 질소를 처리한 구에는 질소량이 적은 쪽에 서 많은 쪽으로 콩과식물인 Lath y ru s venosus 와 Panic u m pe rlong um , 왕포아풀 , Ambrosia coronop ifoli a 그리고 Arte m i si a ludov i c i ana 의 순으 로 우점하였다. 질소를 가장 많이 첨가한 구에는 덩굴성 목본인 검 은딸기 (Rubus s p.)와 미 역취 (Soli da g o g ram i n ifo l i a) 가 우점하였다(그림 8.1 6 ). 전체적으로 이러한 자료들은 실험적으로 설정된 질소 구배를 따 라 식물이 분화됨을 분명히 보여주는 것이다. 더구나, 이 종들은 초 기의 수도가 각각 크게 다름에도 불구하고 질소 구배에 따른 반응 이 유사한 경향이 있었댜 예를 들어 A g rop y ronrep ens 는 25 년 된 휴 경지와 48 년 된 휴경지의 많은 구에서, 그리고 참나무사바나의 모든 구에서 매우 희귀하거나 아예 존재하지도 않았다. 그러나, 이 종은 25 년 된 휴경지와 48 년 된 휴경지의 질소를 많이 처리한 구에 침입 하여 그 수도가 급격히 증가하였으며, 7 개 지소의 처리 구배 중 6 개 지소에서 유사한 반응을 보였다. 질소 구배에 따라 이 종의 수도가 증가하지 않은 곳은 참나무사바나 지역이었는데, 이 종은 이곳에 출 현하지 않았었다. 왕포아풀는 7 개 지소 모두에서 출현하였는데, 질소를 적게 처리한 곳과 중간 정도 처리한 구배에서 최대 수도를 보였다 . 가장 나이가 적은 휴경지에서 Schiz a chy ri u m sco p ar i um 이 없을 때 왕포아풀은 질 소 공급이 가장 적은 곳에서 최대 수도를 보였다. Sch i zach y r i um 이 존재하면 왕포아풀의 최대 수도가 질소 공급률이 중간 정도의 지소 로 이동되고 Sch i zach y r iy m 은 질소 공급이 낮은 지소에서 우점하였 다. 기타 많은 종들의 질소 구배에 따른 반응이 T il man(1987a) 에 의
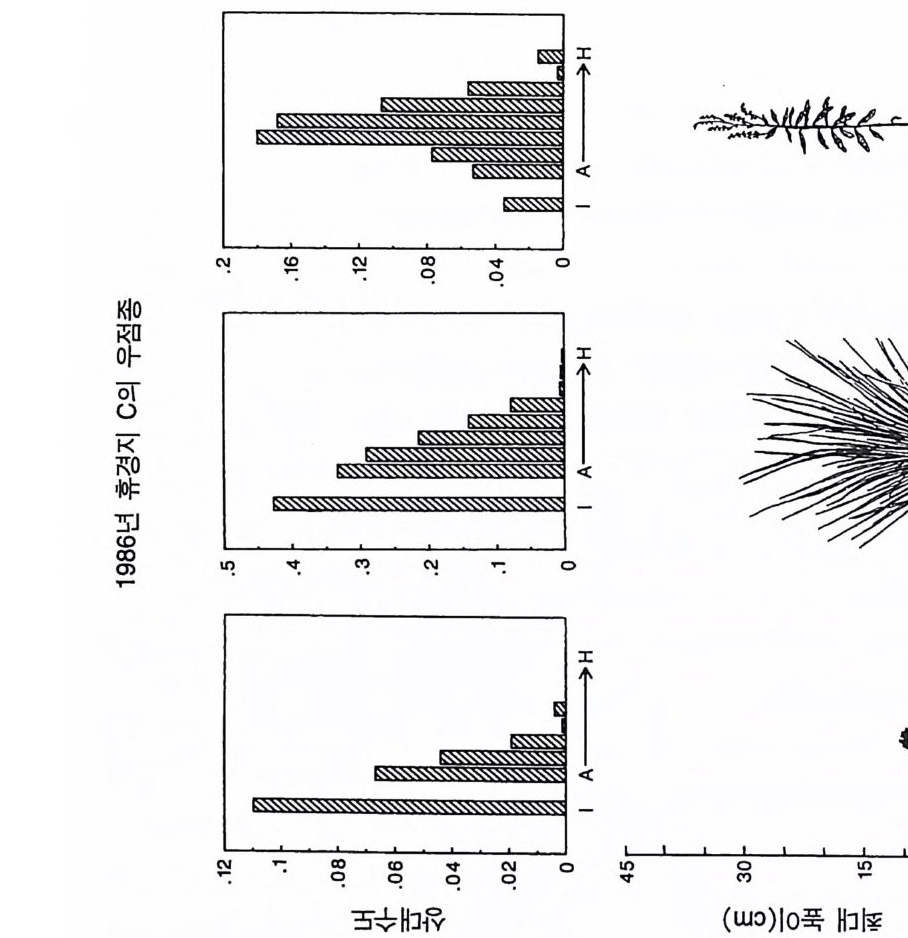 I
I
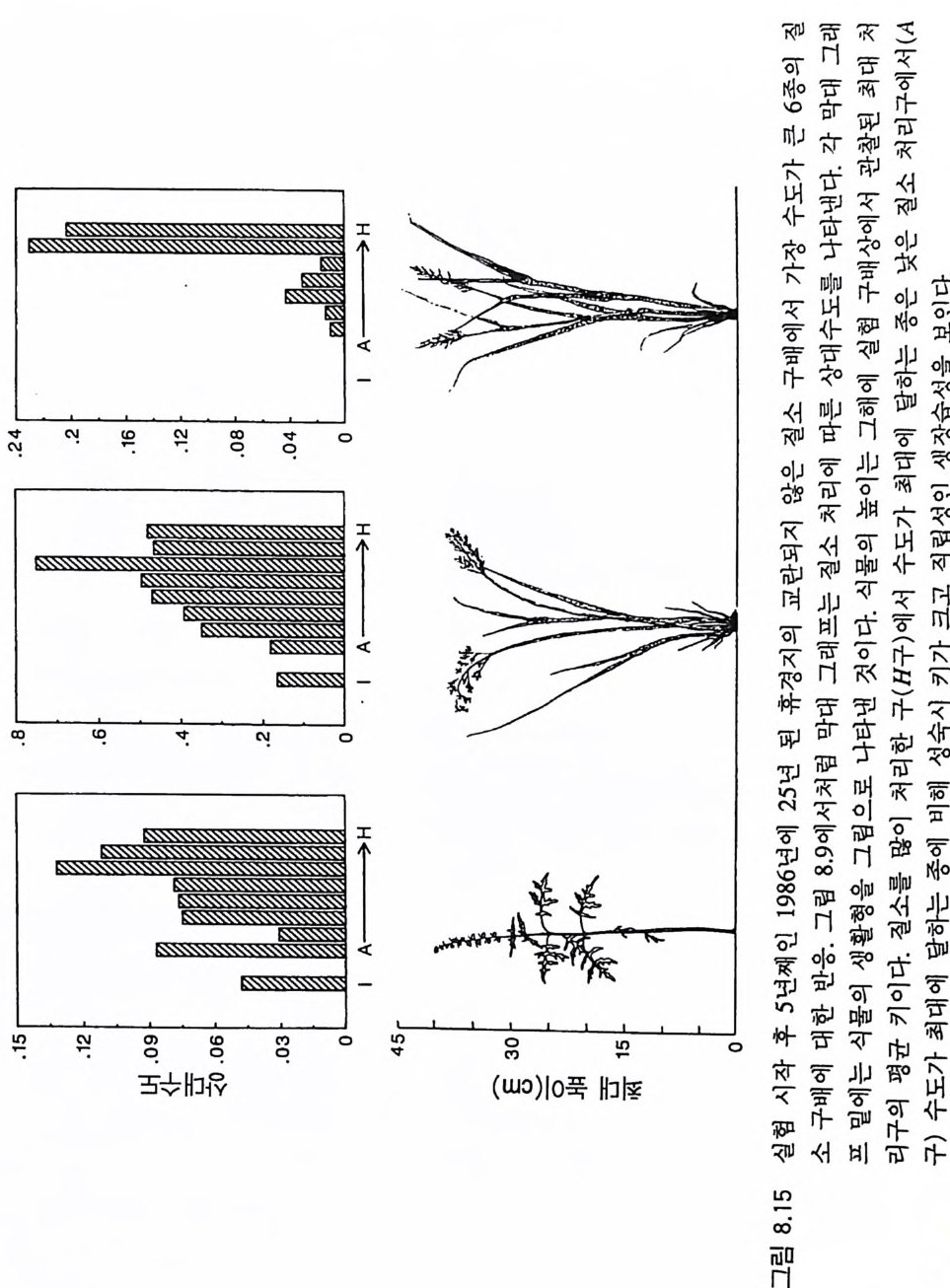 질 래 처 A (
질 래 처 A (
 I<工C
I<工C

해 제시되어 있댜 극히 예외적인 것이지만 각 종은 1982 년 실험을 시작할 당시의 수도가 낮거나 높은 것에 관계없이 1986 년에는 그 종 이 존재하는 모든 구배상에서 같은 수도를 갖는 것으로 수렴하였다. G ri me (l 979, p .16) 은 광, 수분, 영양염류 및 공간에 대한 전체적인 경 쟁 능 력 으로 〈 경 쟁 능력 의 통합 개 념 (unif ied concept of comp e tit ive ab il ity) 〉 을 제안하였댜 그는 일부 종은 모든 자원(질소, 물 광)에 대 한 경쟁력이 우세하고, 일부 종은 모든 자원에 대한 경쟁력이 열등 하다고 주장하였다. 만약 Gr i me 의 주장이 맞다면 식 물은 Cedar Creek 에서의 질소 구배와 같은 자원 구배상에서 그 분포역이 분화 되지 않을 것이다. 따라서, 이러한 실험은 G ri me 의 견해를 반박하는 강력한 증거가 된댜 이러한 실험적인 구배상에서 종의 분리는 질소와 햇빛에 대한 식물의 경쟁 능력이 분화되어 있다는 가설과 일치한다. 그러나 , 그 가설이 맞다면 키가 작고 생활사가 짧은 종은 질소 공급률이 낮은 곳에서 우점하고, 키가 크고 수명이 긴 다년생 식물은 질소 공급률 이 높은 곳에서 우점하여야 한다. 이 실험의 결과는 비록 그 패턴이 결코 완전한 것은 아니지만 이러한 예측을 뒷받침해 주고 있다 (Til m an 198.7 a ). 성숙하였을 때의 식물의 키는 질소 공급률에 따라 증가하는 경향 이었다(그림 8.1 5 , 8.1 6 ). 1984 년에 각각의 질소 구배에서 각 보편종 의 최대 수도가 나타나는 위치와 성숙했을 때의 키 사이의 순위상 관의 결과를 보면 예상한 바대로 7 개의 상관 . 중 6 개가 양의 상관을 보였으며 , 이 중 두 개는 유의성 이 P<0.05 였다. 1985 년의 자료를 이용하여 비슷한 분석을 한 결과도 7 개 상관 모두가 양의 상관을 보였고, 이 중 두 개는 유의성 이 있었다 (T ilm an 1987a).
4-3 휴경지 내에서의 공간 패턴 이러한 실험 결과는 질소가 중요한 제한 자원이며 , 질소 구배상에 서 최대 수도를 보이는 지역이 종에 따라 다름을 보여주고 있다. 그 러면 이와 같이 실험적으로 설정된 질소 구배상에 나타나는 종의 수도의 패턴을 각각의 휴경지 내의 지소에 따른 질소의 공간적 이 질성에 대한 식물의 반응 혹은 천이 과정에 따른 토양 질소의 차이 에 대한 반응과 어떻게 비교할 수 있는가? 만약 질소가 이러한 종 들의 자연 상태에서의 분포 패턴을 결정하는 중요한 요인이라면, 그 리고 실험이 시작된 후 5 년째 해인 1986 에 질소 구배에 대한 식물 의 반응이 질소 구배의 장기적인 영향을 나타내는 것이라면 질소 구배에 대한 식물종의 반응은 한 휴경지 내나 휴경지들 사이에서 관찰되는 상관적인 패턴과 일치하여야 한다 . 7 개의 상이한 질소 구배에 보편적으로 나타나는 모든 종 중 15 종 이 휴경지에서 가장 풍부한 20 종 속에 포함되어 있었다 (T i lman 1987a). 실험 후 5 년째 해에 이들 대부분이 개개의 휴경지 내의 질 소의 공간적인 변이에 대하여 비슷한 반응을 보였고, 7 개의 질소 구 배에도 비슷한 반응을 보였다 . 예를 들어 Ag ro sti s scabra 는 우리가 조사한 22 개의 휴경지 중 21 개 지소에 출현하였다. 각각의 휴경지에 서 나는 0.5X1.0m 방형구 내에 있는 A g ros ti s 의 피도%가 그 방형 구 내의 중앙부에서 채취한 토양의 전질소 함량과 상관이 있는지의 여부를 조사하였다. 각 휴경지마다 100-150 개의 방형구를 조사하였 댜 그 결과 A g ros ti s 의 수도는 토양의 전질소 함량과 17 개의 휴경지 에서 음의 상관을 보였으며, 이 중 8 개는 유의 수준이 P<0 . 05 였다. 이것들 중 6 개의 결과가 그림 8 .1 7 에 나와 있다. 4 개의 휴경지에서 는 양의 상관을 보였으나 5% 수준에서 유의성이 있는 것은 없었다 . 양의 상관이 가장 높은 휴경지의 상관계수는 r=0.06 이었다.
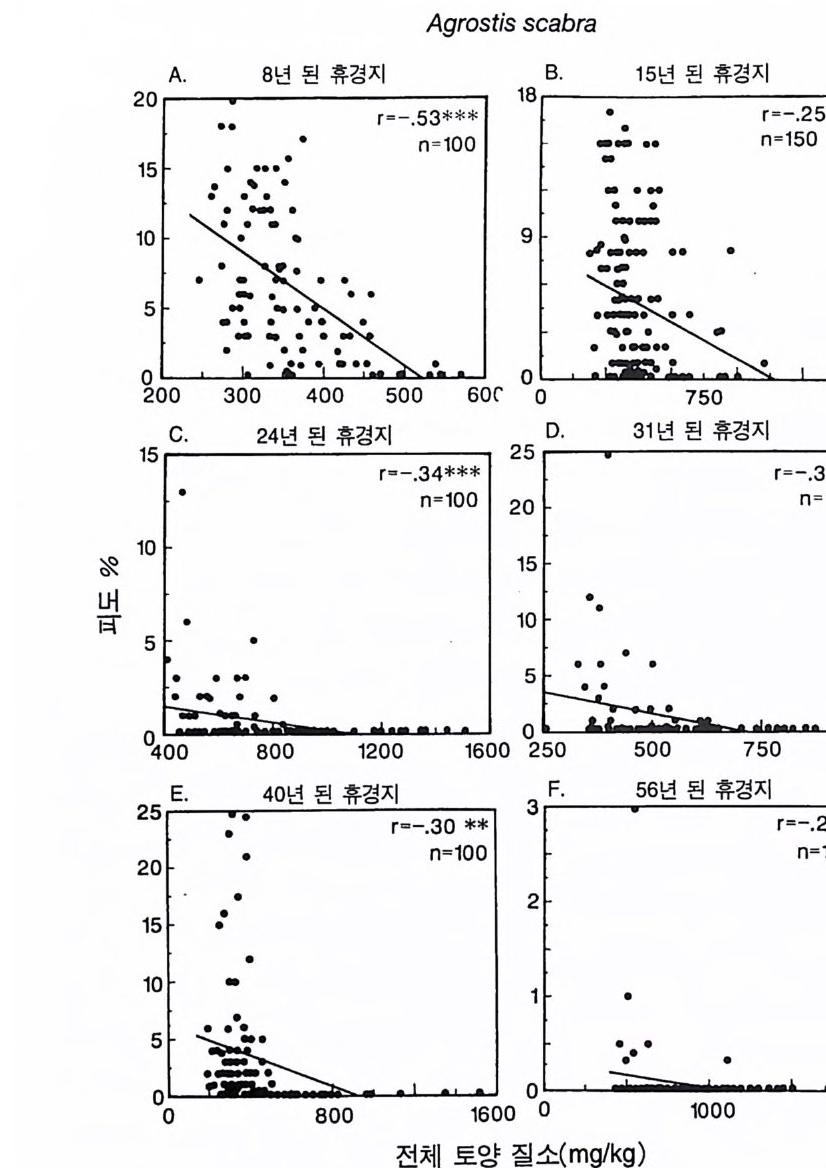 Ag ro sti s s cabra
Ag ro sti s s cabra
그림 8.17 각 그래프는 Cedar Creek 의 특정한 휴경지 내에서 토양의 전질소에 대 한 A g ros ti s 의 피도를 나타낸 것이다. 직선은 희귀직선이며. r 는 상관계수 이다. 각 그래프는 휴경지당 100 혹은 150 개의 방형구에서 조사된 식생 의 피도와 토양의 전질소 자료를 이용하여 작성된 것이다.
그림 8 .1 7 에 나와 있는 바와 같이 각 종의 수도에는 토양 질소에 의해 설명할 수 없는 많은 분산이 있음을 알 수 있다. 가장 유의성 이 있는 상관도 A g ros ti s 의 경우 실험구 대 실험구(p lo t-t o- p lo t) 변이 의 25% 만을 설명할 수 있었다. 그러나 그와 같은 소규모의 변이지 만 A g ros ti sscabra 는 이 휴경지에서 질소가 적은 지역에 더 많이 분 포하는 일정한 경향성이 있었다. 모든 휴경지를 함께 고려할 때에도 이러한 패턴은 여전하였다(그림 8.1 8 ). A g ros ti s 가 출현하는 21 개 휴 경지의 모든 자료를 사용하면 이 종의 피도는 토양의 질소와 뚜렷 한 음의 상관을 보였다 (r= -0.28, n=2,162, P<0.001 ). 모든 자료를 40 개의 상이한 질소 범위로 나누고. 각각에서 A g ros ti s 의 피도를 계 산하여 나온 종합곡선은 A g ros ti s 가 휴경지의 질소가 빈약한 지역에 서 수도가 가장 높고, 질소 수준이 증가함에 따라 수도가 감소함을 보여주고 있다(그림 8.1 8 ). A g ros ti sscabra 의 수도가 질소가 적은 지역에서 높고 질소가 많은 곳에서 낮은 이러한 패턴은 질소 시비에 대한 이 종의 반응과 일치 한댜 실험 을 시 작한 지 2 년째 부터 A g ros ti s 의 수도는 모든 휴경 지 에 서 질소 함량의 증가에 따라 감소하였다(그림 8.1 9 ). 따라서 질소 첨 가에 대한 A g ros ti s 의 반응은 공간적으로 이질적인 휴경지 내에서 그리고 이 종이 출현하는 모든 휴경지에서 자연적인 분포 패턴과 일치하였다. 몇몇 다른 종도 각각의 휴경지 내에서 토양 질소가 증가함에 따 라 그 수도가 감소하는 경향이 있었으며. 이들은 또한 실험적으로 설정한 질소 구배상에서도 수도가 감소하였다. 예를 들어 박하인 Hedeoma h i sp i da 는 22 개 휴경지 가운데 16 개의 휴경지에서 출현하 였는데, 14 개의 휴경지에서 질소가 증가함에 따라 수도가 감소되었 고. 이 중 9 개의 휴경지에서는 뚜렷한 음의 상관을 보였다. 이 종은 2 개의 휴경지에서는 질소가 증가함에 따라 수도가 증가하였는데. 이
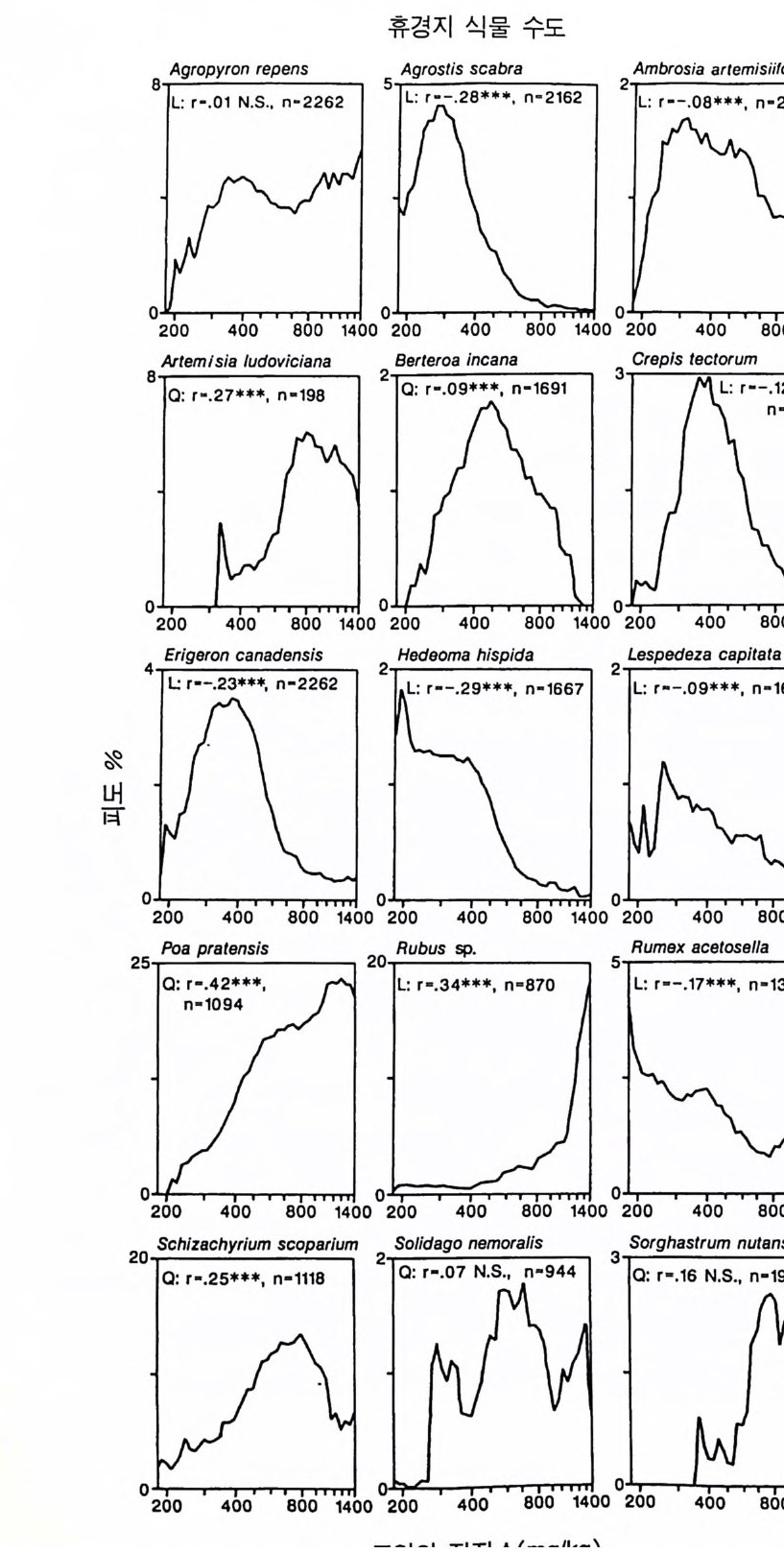 휴경지 식물 수도
휴경지 식물 수도
그림 8.1 8 Cedar Creek 에 있는 22 개의 휴경지에서 흔히 출현하는 15 종 식물의 피 도와 토양의 전질소 함량 사이의 관계를 보여주는 그립 각 종의 그립은 그 종이 존재하는 모든 휴경지의 전체 자료를 이용한 것이다 . 각 그립은 또한 식물의 피도치와 토양의 전질소 함량 사이에 직선적 (L) 혹은 2 차 함수적(Q) 회귀 관계가 있는지를 보여주고 있다. 평균을 사용하여 막대 그래프롤 곡선으로 표시하였다 (T il man 1987a).
중 하나의 휴경지에서는 뚜렷한 양의 상관을 보였다. 이 종은 1983, 1984, 1985 년 그리고 1986 년에는 실험적으로 설정한 질소 구배에 따라 수도가 감소하였다 (T i lman 1987a). Ambrosia artem i si i foli a, Crep is t ec to r um, 망초, Lespe deza cap itat a , 애기수영은 휴경지마다 수도와 토양의 질소 함량 사이에 일치된 상 관을 보이지 않았다. 그러나 모든 휴경지의 자료를 통합하여 고려할 때 단순상관은 이들 각각의 종이 토양의 전질소 함량에 따라 수도 가 감소하고, 토양의 전질소 함량이 낮은 곳에서 최대 수도에 도달 하는 것으로 나타났다(그림 8.1 8 ). 이 종들은 각각 질소 첨가에 따라 수도가 감소하는 경향을 보였지만 몇 종은 예외적인 경향을 보였다. 특히 망초와 Crep ist ec to r um 등의 1 년생 식물은 다른 경향을 보였다 (Til m an 1987a). 다른 종들은 토양 질소의 증가에 따라 수도가 증가하였다. 예를 들어 Ag ro p yro n rep ens 는 22 개의 휴경지 모두에 출현하였는데, 이 종 의 피도는 16 개의 휴경지에서 토양의 질소 함량과 양의 상관이 있 었으며, 이 중 8 개의 휴경지에서는 P<0.05 의 유의성이 있었다. 이 종의 피도는 6 개의 휴경지에서 질소와 음의 상관을 보였으며, 이 중 하나만이 유의성이 있었다(그림 8.20). 이미 설명한 바와 같이, 이 종 은 자신이 출현하는 6 개의 구배 중 질소 함량이 많은 구에서 우점 하였다. 따라서 질소를 많이 첨가한 구에서 이 종이 우점하는 것은 각 휴경지의 질소가 더 많은 지소에서 이 종의 우점도가 높은 자연 적인 분포 패턴과 일치한다.
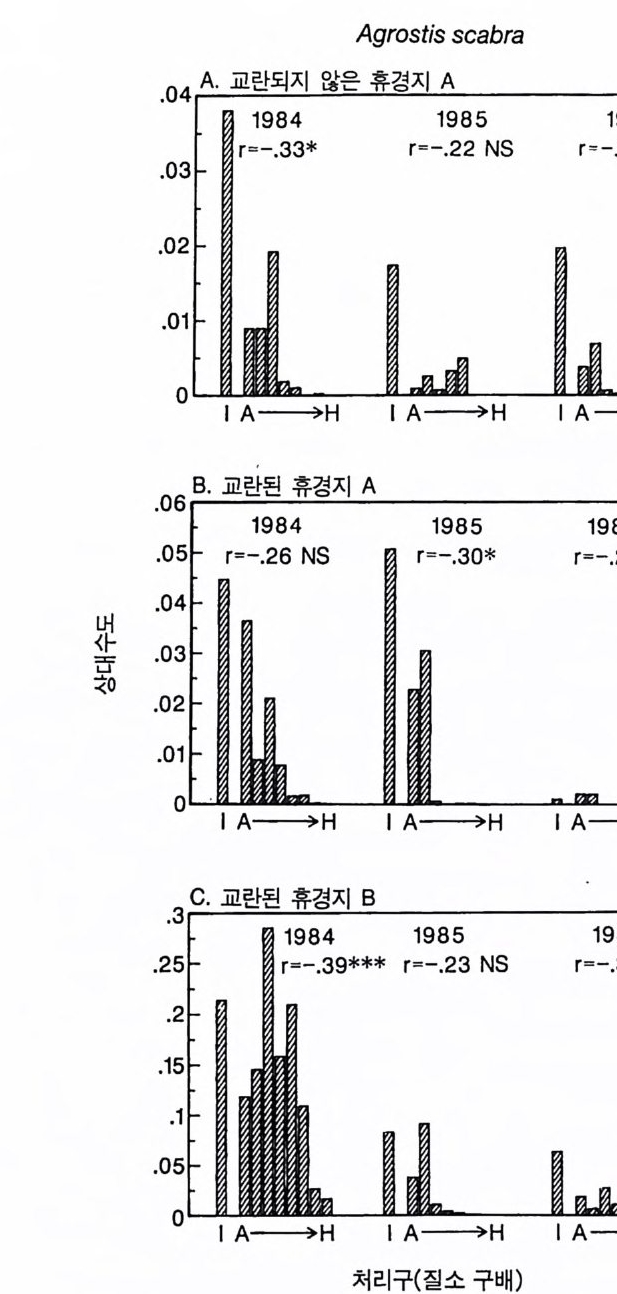 Ag ro stis s cabra
Ag ro stis s cabra
그림 8.19 (A) 1984, 1985 그리고 1986 년에 휴경지 A 의 교란되지 않은 실험구에서 질소구배 에 대한 Ag ro stis s cabra 의 반응. (B) 휴경지 A 의 교란된 지소에서의 Ag ro stis s cabra 의 반응. (C) 휴경지 B 의 교란된 지소에서 A g ros ti sscabra 의 반응. 다른 4 개의 질소 구배구에서는 이 식물이 없거나 아주 드물게 출현하였다 . 그림에 나와 있는 상관계 수 r 는 A g ros ti s 의 상대수도와 질소 첨가율 사이의 단순상관에 근거한 것이다 .
그러나 22 개 휴경지의 자료를 함께 분석하면 토양의 질소와 Ag ro py ro n re p ens 의 수도 사이 에 유의 한 상관이 나타나지 않았다 (그림 8.18). 이것은 A g rop y ron 이 신생 휴경지보다 오래된 휴경지에 서 수도가 낮기 때문이다. 이 종은 각 휴경지의 질소가 많은 지소에 서 수도가 일률적으로 높긴 하지만, 오래된 휴경지가 질소 함량이 더 많아도 오래된 휴경지에서는 분명히 이 종이 다른 종으로 교체 되고 있었다 . 이러한 사실은 천이 과정 중 A g ro py ron 의 동태를 결정 하는 요인으로서의 질소의 역할과 위배된다. 다음에 설명하겠지만, 이 종이 천이 초기에 우점하는 것은 일시적인 동태일 수 있다. A g rop y ron 은 질소 함량이 많은 교란된 서식처에 특별히 분화된 종 일 수 있댜 각 휴경지에서 질소 함량에 따라 수도가 증가한 또 하나의 종으 로는 목본성 덩굴식물인 검은딸기 (blackbe rry, Rubuss p.)가 있다. 검 은딸기는 9 개의 휴경지에서 출현하였다. 이 중 5 개의 휴경지에서 이 종의 피도는 토양 질소와 뚜렷한 양의 상관이 있었고, 나머지 중 하 나에서는 질소 함량에 대한 피도치가 2 차 함수로 나타났고, 또 하나 에서는 질소 함량과 음의 상관을 보였다. 이 9 개의 휴경지에서 얻은 모든 자료를 한 단위로 분석하면 검은딸기의 수도와 토양의 질소 함량 사이에 고도의 양의 상관이 있었으며. 이 종의 수도는 토양의 질소 함량에 따라 거의 지수함수적으로 증가하였다(그림 8.1 8 ). 이 종은 토착 참나무사바나 지역에 보편적으로 출현하였는데, 이 종의 상대수도와 절대수도는 1983 년 이후부터 실험적으로 설정된 질소 구배를 따라 현저히 증가하였다(그림 8.1 6 , Til ma n 1987a). 각 휴경지 내의 질소 함량이 높은 지소에서 수도가 높은 또 하나 의 종은 왕포아풀이었다. 이 종은 22 개의 휴경지 중 11 개의 휴경지 에서 출현하였으며, 이 종의 수도는 9 개 휴경지에서 질소 함량과 양 의 상관이 있었고 이 중 6 개가 유의한 상관을 보였다• 이 밖에 이
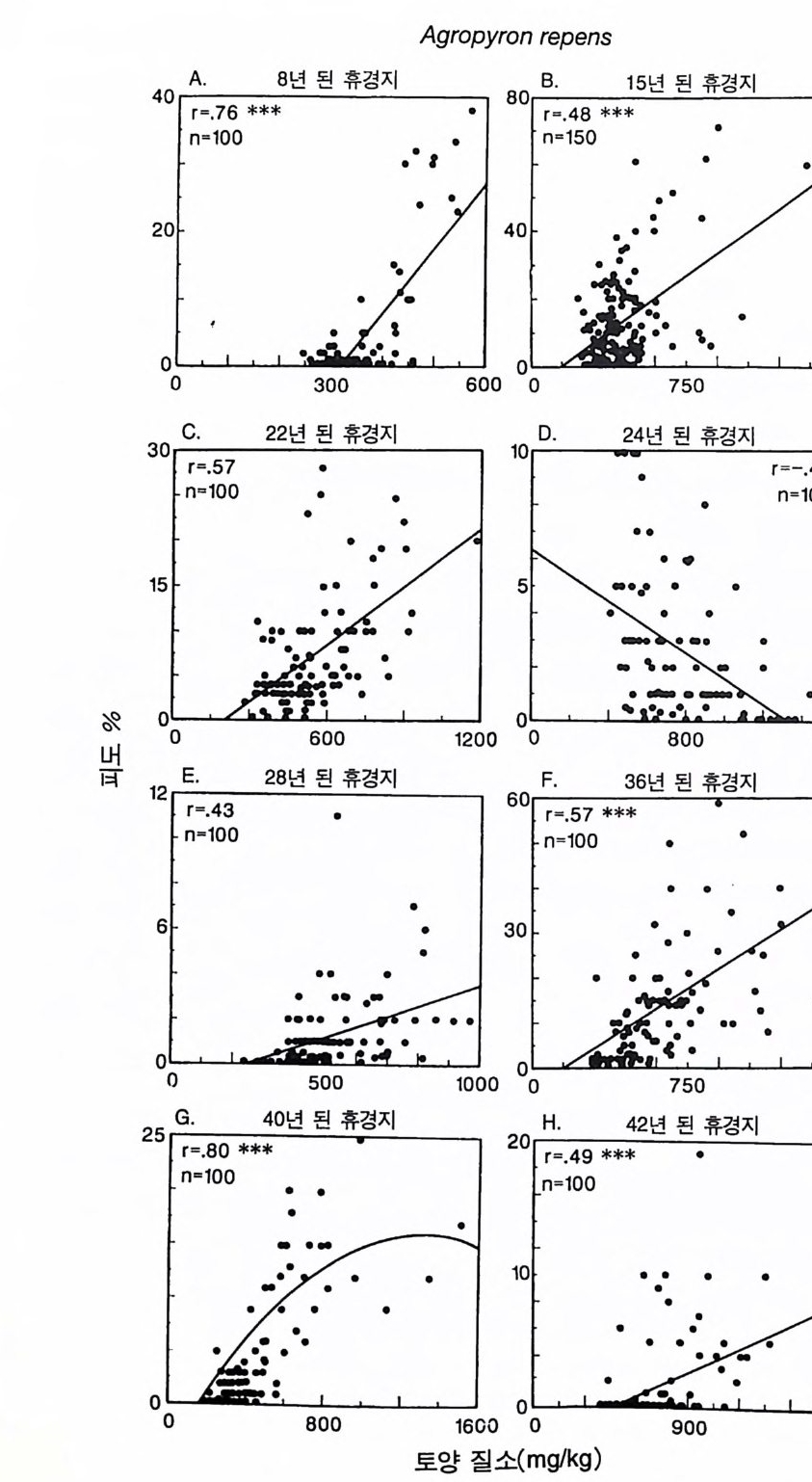 Ag ro p yro n rep e ns
Ag ro p yro n rep e ns
그림 8.20 Cedar Creek 에 있는 8 개의 상이한 휴경지에서 토양 전질소 함량과 Ag ro py ro n rep ens 의 피도와의 관계. 이들 8 개의 휴경지는 A g rop y ron 의 수도가 가장 높고 , 그것의 수도와 토양의 질소 함량 사이에 유의한 직선 적 혹은 2 차 함수적 상관이 있는 지소이다. 각각의 휴경지에서 각 점은 0.5 m x l.O m 방형구 내의 A g rop y ro11 의 피도와 각 지소의 중심부에서 채 취한 토양의 전질소 함량을 나타난다 .
종의 수도는 두 개의 휴경지에서 질소 함량에 대하여 2 차 함수로 나타났댜 이 종이 출현하는 11 개의 휴경지의 자료를 합쳐서 그룹으 로 분석하면 이것의 수도는 토양의 전질소 함량에 대하여 2 차 함수 로 나타난다(그림 8.18). 이러한 현상은 왕포아풀이 CedarCreek 내 에 있는 휴경지 토양의 대부분의 질소 범위에서 토양의 질소 함량 에 따라 그 수도가 증가하지만 질소 함량이 매우 높은 지역에서는 그 수도가 감소함을 의미한다. 질소 첨가에 대한 이 종의 반응이 분 포 패턴과 일치하기 위해서는 질소 첨가율이 낮을 때 이 종의 수도 가 증가하여야 하고 질소 첨가율이 높을 때 수도가 감소되어야 한 다. 이러한 패턴이 휴경지 A 를 제외한 모든 휴경지에서 관찰되었다 (그림 8.21 ). 따라서 휴경지 조사에서 토양의 질소 함량에 따라 그 수도가 증 가하는 경향이 있거나 감소하는 경향이 있는 많은 보편종의 경우 실험적으로 설정된 질소 구배에 대한 이러한 반응은 휴경지에서 그 들의 공간적인 분포를 결정하는 데 질소의 역할이 크다는 것을 입 증하는 것이다. 왕포아풀 이외에 다른 몇 종은 휴경지 조사 결과에 의하면 토양의 전질소 함량이 중간 정도에서 최대 수도를 보였다 (그림 8.1 8 ). 이들 중 실험적으로 설정된 질소 구배에 대한 반응 결 과는 이 종의 자연 상태의 분포 패턴을 결정하는 데 질소의 역할이 중요함을 입증하는 것이다 (T il man 1987a). 그러나 Schiz a chy ri u m scop ar i um 은 달랐댜 실험적으로 설정된 질소 구배에 대한 이 종의
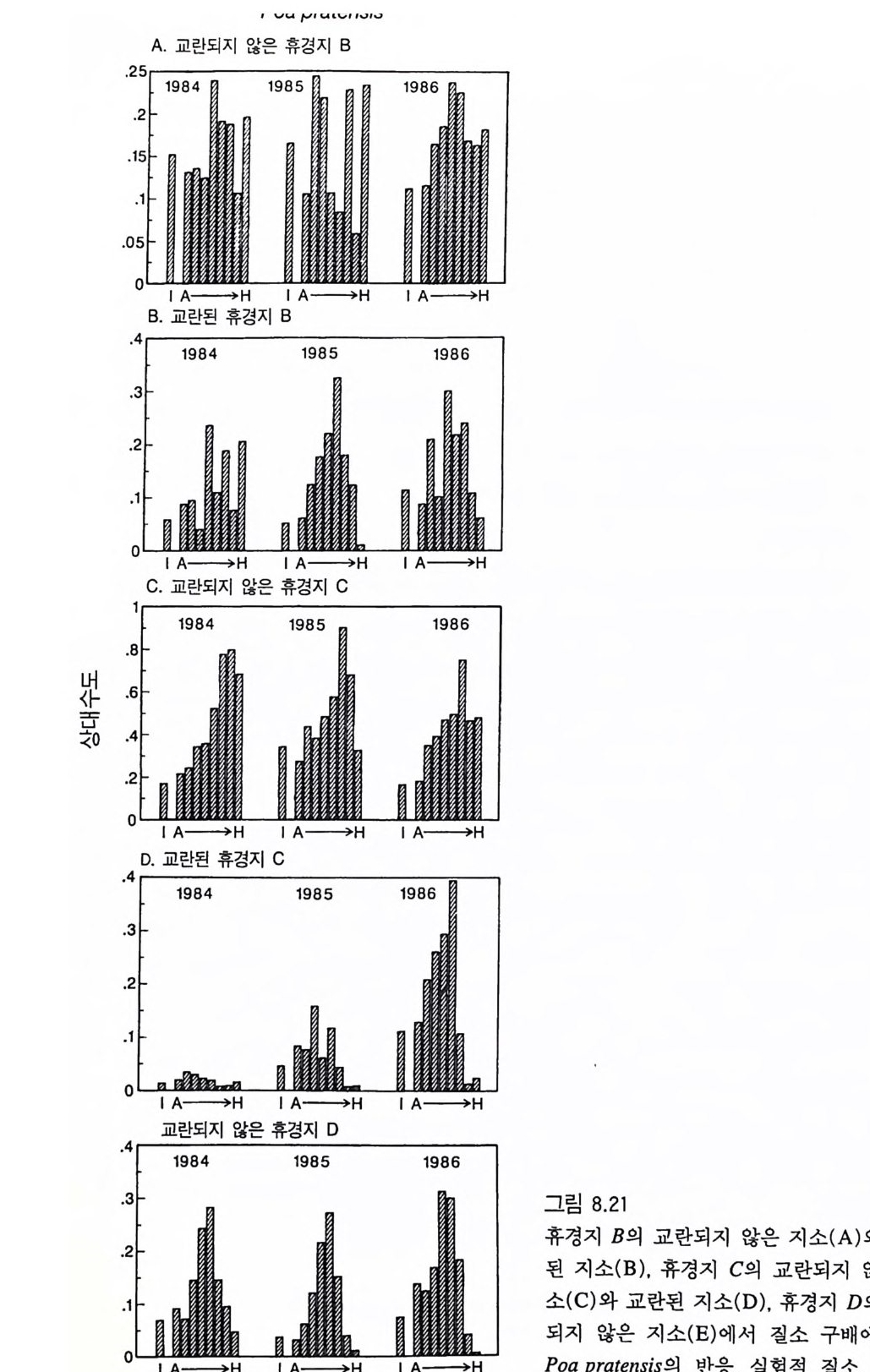 , v F''V''>
, v F''V''>
반응은(그림 8.1 5 ), 자연 상태의 토양의 질소 구배에 따라 감소할 것 으로 예상되었지만 실제로는 Gaussia n 반응을 보였다. 이상을 종합해 보면, 휴경지 조사에서 피도가 토양의 전질소 함량 에 의존적인 12 개의 식물 중(그림 8.18) 실험적으로 설정된 9 개의 질 소 구배에 대하여 보인 식물의 반응은 토양의 질소 함량이 휴경지 에서 이들 종의 분포를 결정하는 주요인이라는 가설을 뒷받침하는 것이었다. 5 천이의 동태 제 7 장에서 제시된 이론은 척박한 토양에서의 2 차 천이가 진행되 는 동안 초기의 종 계열은 주로 일시적인 동태에 의해 결정되며, 토 양 영양염류의 점진적인 증가는 주로 천이의 후기 단계에 영향을 주는 것으로 예측하였다. CedarCreek 에서의 실험을 통해 이러한 예 측을 다른 대안들과 비교하여 검증할 수 있는 기회를 갖게 되었다. 대안 중의 하나는 일시적인 동태가 중요하지 않으며, 전체 천이 과정은 오직 토양의 질소 함량의 증가와 그것이 경쟁에 미치는 효 과만으로 설명이 가능하다는 것이다. Cedar Creek 에서 질소의 축적 이 천이의 주된 요인이라면 초기 단계에서는 질소 함량이 낮은 곳 에서 최대 수도에 이르는 종이 우점해야 되고, 중간 단계의 휴경지 에서는 질소 함량이 중간 정도인 곳에서 최대 수도에 이르는 종이, 그리고 후기 단계의 휴경지에서는 질소 함량이 많은 곳에서 최대 수도에 이르는 종이 우점해야 된다. 또다른 대안은 전체 천이 과정이 일시적인 동태에 의해 결정된다 는 것이다. 만약 그렇다면, 천이 초기종은 최대 생장률이 가장 커야 하고, 이들은 점차적으로 최대 생장률이 낮고 줄기로의 분배량이 점
그림 8.22 연도가 다른 휴경지의 식생 자료와 일부 영구 조사구의 관찰을 통해 나 온 Cedar Creek 에서의 천이 동태 . 휴경지에 흔히 출현하는 17 종 각각의 수도(피도치로 추정한 것)를 천이 단계에 따라 도시하였다. 이 그림은 22 개의 휴경지를 조사하여 나온 각 종의 평균 피도와 (Inou y e et al. 1987a), Barbara Delane y가 Cedar Creek 에 있는 11 개의 천이 계열 군집을 반복하 여 조사한 자료를 사용하여 작성되었다 . 나는 이 33 개의 자료점을 이용하 여 이들 17 종 각각의 천이 동태를 보여주는 곡선을 만들었다. 천이 후기 종 중 일부는 휴경지의 나이가 40 년 이하인 곳에서는 나타나지 않았다.
진적으로 증가하는 종에 의해 교체되어야 한다(그림 7.5 참조) . 3 번째 대안은 천이의 전체 과정이 정착률과 경쟁적 상호 작용에 의해 결정된다는 것이다. 그러기 위해서는 천이 초기종은 휴경지가 될 때 그곳에 존재해야 하거나 종자의 분산력이 좋아야 하며, 후기 종은 경쟁력은 강하지만 종자의 분산력이 나빠야 한다. 이러한 대안 들을 평 가하려 면 Cedar Creek 에 서 천 이 의 실제 동태 를 알아야 한다 . 이를 위해 나는 두 개의 서로 다른 자료들을 합쳤다. 하나는 앞에서 설명한 22 개의 휴경지를 조사한 것이고(I nou y e eta l. 1987a), 다른 하 나의 자료는 1974 년부터 1984 년까지 순차적으로 폐경된 휴경지에서 11 개의 20mx 100m 의 식생을 조사한 것이댜 이 계획은 Don Lawrence 에 의해 시작되었으며, 자료는 Barbara Delane y에 의해 수집되었다. 이 두 분 모두에게 감사드리며, 특히 Barbara 는 미발표 자료를 이용하도록 배려해 주었다 . 각 종의 평균 수도는 각 휴경지마다 계산되거나 각 천이 단계에 따라 계산되었고, 이 33 개의 점을 천이 단계에 따라 도시하였다. 이 곡선들은 그림 8.22 에 나와 있다. 모든 식물을 총괄한 피도를 보면 100% 보다 훨씬 적은데, 이것은 각 휴경지의 12-50% 가 낙엽으로 덮여 있고 (Inou y e eta l. 1987a), 나머지 중 0-30% 가 맨땅이기 때문이다(그림 8.2).
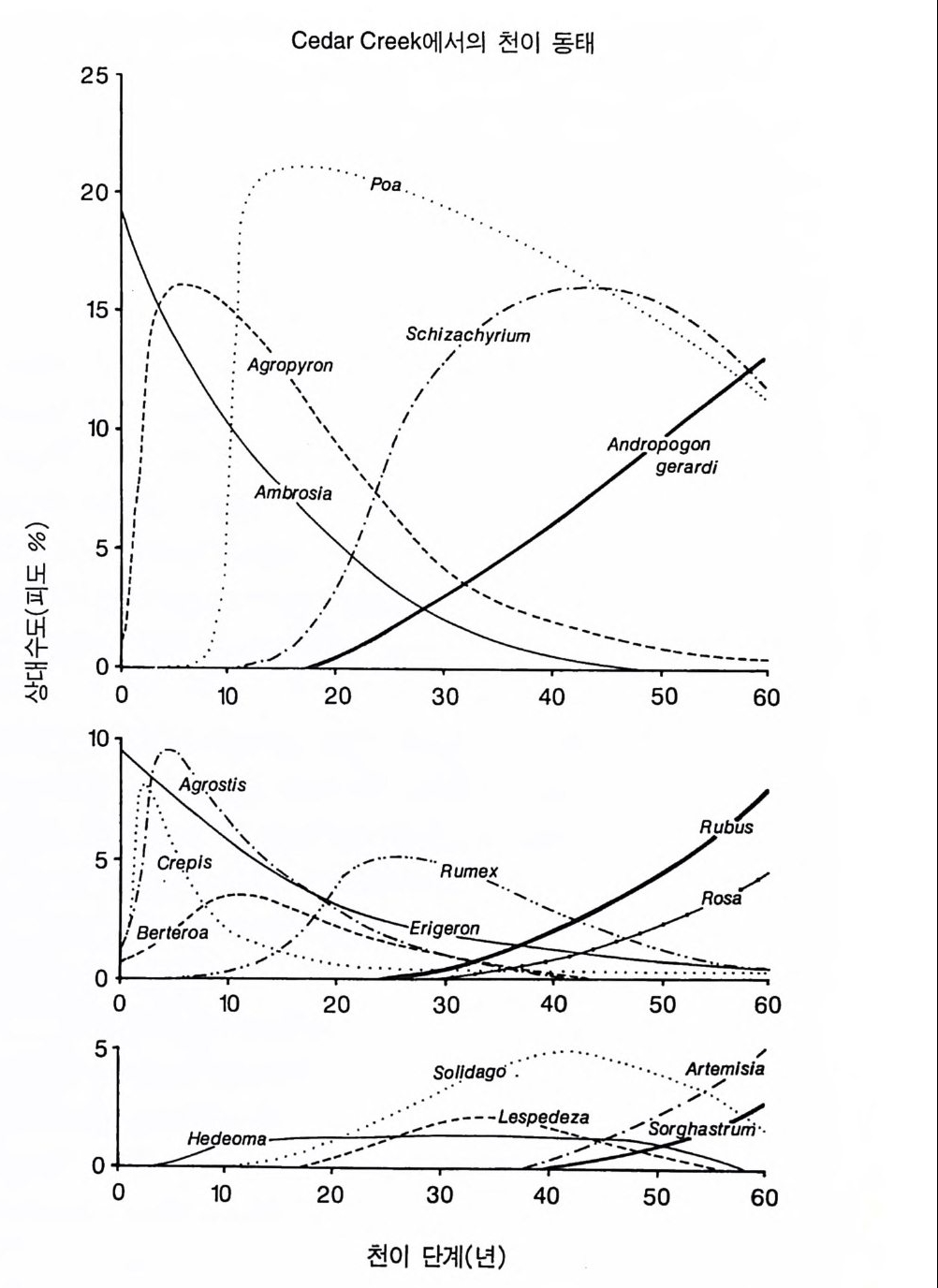 Cedar Creek 에서의 천이 동태
Cedar Creek 에서의 천이 동태
5-1 질소 축적 개략적으로 살펴보면 Cedar Creek 에서 실험적으로 설정한 질소 구배로부터 얻은 자료는 천이의 패턴이 일시적인 질소 구배에 의해 영향을 받는다는 가설을 뒷받침하고 있다 . 천이 초기의 휴경지에는 1 년생 식물이 우점하는데(그림 8.2 ) , 이 식물들은 질소 공급률이 낮 은 곳에서 최대 수도에 이르며, 질소 공급률이 증가하면 수도가 감 소된다 (T il man 1987a). 수명이 긴 협엽초본과 광엽초본, 특히 초지의 고유종은 천이가 진행되는 동안 그리고 질소 구배에 따라 수도가 증가한다. 목본 식물종도 천이가 진행되는 동안 그리고 질소 구배에 따라 수도가 증가한다. 이러한 결과로 볼 때 질소의 점진적인 축적 은 Cedar Creek 에서 볼 수 있는 2 차 천이 패턴의 원인이 된다는 가 설과 부합된다. 그러나, 이러한 자료들을 좀더 세분된 시간이나, 종 대 종의 기준에서 볼 때 다른 과정도 중요하다는 것을 분명히 알 수 있댜 몇몇 보편종들은 천이가 진행되는 동안 일시적인 수도 패턴을 갖 는데, 이것은 천이 동태를 결정하는 주요인이 질소라는 사실과 일치 한다. 예를 들어 Ambrosia ar t em i s iifo li a 는 질소를 시비한 후 수도가 감소되었는데, 휴경지 내의 질소가 풍부한 지소와 질소 함량이 많은 휴경지에서 수도가 낮고, 천이가 진행됨에 따라 수도가 감소하였다 (그림 8.22). 망초는 휴경지 조사에서 일반적으로 토양의 질소 함량 이 증가함에 따라 수도가 감소하였고(그림 8.1 8 ). 천이 과정 중에서 도 수도가 감소하였으나(그림 8.22), 질소 첨가에 대해서는 일치된 반응이 없었댜 검은딸기 (Rubus s p . )는 질소 첨가에 따라 수도가 증 가하였는데, 휴경지 내의 질소가 많은 지소와 질소가 풍부한 휴경지 에서 수도가 높았으며(그림 8.1 8 ), 천이가 진행되는 동안 수도가 증 가하였다(그림 8.22). Berte r oa i ncana 와 Arte m i si a ludov ici ana 는 휴경
지 조사에서 질소에 대하여 Gaussia n 반응을 보였고(그림 8.1 8 ), 그 들이 최대 수도에 이르는 전질소의 수준과 CedarCreek 에서 천이 과 정 중 관찰되는 실제 토양의 전질소 수준을 고려할 때 이것과 일치 되는 천이 동태를 보였다. 그러나 Ag ro stis s cabra, Hedeoma his p i da , Crep is t ec to r um, 애기수영, 그리고 Lespe deza cap it a t a 와 같은 종은 휴 경지 내부나 휴경지 간에 질소 구배에 따라 수도가 감소하였고, 실 험적으로 설정한 질소 구배상에서도 수도가 감소하였다. 그러나 천 이 과정의 3-30 년 기간에서는 시간에 따라 이들의 수도가 증가하였 다(그림 8.22). 따라서 이 각각의 종은 질소에 대한 반응과 천이 과정 중 질소 증가의 경향을 기준으로 할 때 매우 수도가 높을 것으로 예상되는 천이 초기 과정 중에 전혀 나타나지 않거나 수도가 매우 낮다. 이와 유사하게 Solid a g o nemora li s 는(질소 첨가에 따라 수도가 감소하였지 만) 천이 초기의 질소 함량이 가장 낮은 토양에 나타나지 않았고, 시간이 지남에 따라 서서히 수도가 증가하였다. 왕포아풀과 Schiz a -chy ri u m sco p ar i um 의 경우 만약 시간에 따른 질소 중가에 의해 천이 의 동태가 결정된다면, 질소 첨가 실험이나 토양 질소에 대한 종의 의존도 면에서 볼 때 Sch i zach y r i um 이 왕포아풀 이전에 우점종이 되 어야 하지만 왕포아풀이 Schiz a chy ri u m scop ar i um 에 비해 일찍 우점 하였다. 더구나 Ag ro py ro n re p ens 는 실험적으로 설정한 7 개의 질소 구배구 중 6 개에서 질소 첨가에 따라 현저히 증가하였지만 천이의 4 번째에서 8 번째 해에 수도가 최고치에 달한 다음 감소하였다. 따라서 천이 동태의 많은 부분이 천이 과정 중에 접진적으로 증 가하는 토양 질소에 대한 경쟁의 평형 결과로 설명될 수 없다. 실제 로, 천이 과정 중 가장 많이 출현하는 5 종 중 Ag ro p yro n, 왕포아풀 그리고 Schiz a chy ri u m 3 종의 경우 이들의 수도가 최고에 달하는 순 서가 질소 축적 가설에 의해 예상되는 것과는 정반대였다(그림 8.1 8
울 그림 8.22 와 비교). 그러면 정착률 혹은 일시적인 동태로 이러한 것을 설명할 수 있을까? 5-2 정착 어느 서식처에서 종이 생장하기 위해서는 그 서식처에 그 종이 존재하여야 한다. 영양염류가 반약한 토양에서 경쟁력이 낮은 종은 초기의 척박한 토양에서 느린 정착률이 그 종의 천이 동태에 크게 영향을 주지 않는다 . 그러나 영양염류가 적은 서식처에서 경쟁력이 좋은 종에게는 느린 정착률이 종자의 분산거리가 짧아서건, 새로 교 란된 지역에 종자의 밀도가 낮아서건 간에 천이 동태에 중요한 영 향을 줄 수 있다. Cedar Creek 에 서 천 이 초기 에 우점 하는 Ambrosia ar t em i s iifo li a 는 매토종자가 많은 1 년생 식물이다. 이 종은 또한 농경지에 혼한 야초 이댜 이 식물의 종자는 휴경지가 조성될 때 이미 그 지역에 존재하 며, 초기에 이 종이 우점하는 것은 처음부터 종자가 많기 때문이다. 그러나 이 식물은 20 년 이상 이 지역에서 높은 수도를 유지하고 특 히 휴경지 내의 질소가 적은 지역에서 질소 첨가에 따라 현저히 감 소하는데, 이것은 초기에 이 종의 수도가 높다는 것 하나만이 천이 초기 과정에서 이 종의 우점도 패턴을 결정하는 유일한 요인이 아 님을 의미한다. 종자가 바람에 의해 쉽게 분산되는 월년초인 망초는 새로 교란된 지역의 또 하나의 우점종이다 . 이 종은 매토종자가 많지는 않지만, 농경지의 야초이기 때문에 휴경지가 조성될 때 종자는 존재한다. 천 이의 초기 과정에 이 종이 나타나는 것은 그것의 야초로서의 역할 및 종자 분산이 양호하다는 사실과 밀접한 연관이 있다. 이 종은 질 소 첨가실험에서는 일관성을 보이지 않았다.
휴경지의 질소 분포와(그림 8.18), 질소 첨가에 대한 Crep is tec to r um, Ag ro stis scabra, 애 기 수 영 , 그 리 고 Lespe deza capi tat a 의 반 응을 기준으로 할 때 이 종들은 새로 형성된 질소가 적은 휴경지에 서 우점종이 될 것으로 기대된다. 그러나 이 종들이 최대 수도에 이 르려면 2-30 년이 필요하다(그림 8.2 2 ). 이렇게 2-30 년이 지연되는 것은 이들의 정착이 느리기 때문인 것 같다. 이러한 가설은 이 종들 의 가능한 분산율과 각 종이 최대 수도에 도달하기 전에 지연되는 기간이 서로 일치하는 것으로 보아 타당성이 입증된다. A g ros ti s 는 종자 분산 방식 이 t umbleweed 의 기 작과 같고, Crep i s 는 분산을 돕는 낙하산 모양의 깃털을 가진 작은 종자를 생산한다. 이 두 종은 2-5 년 내에 최대 수도에 이른다(그림 8.2 2 ). 이에 비해 애기수영과 Lesp edeza 는 분산 구조가 없는 큰 종자를 생 산하기 때문에 정 착이 느릴 수 있다. 이들은 최대 생물량에 도달하는 데 20-30 년이 필요하 다 . 또한 이들의 최대 생물량은 낮은데, 그 이유는 이 종들이 정착할 때까지 그 지역 토양의 평균 질소 함량이 낮아 생장에 불리하기 때 문인 것 같댜 그러나 그런 지역에서도 이들은 지역적으로 수도가 높게 유지될 수 있다. 5-3 일시적인 동태 CedarCreek 에서 관찰되는 천이 동태의 일부를 분산 기작 이외에 도 분배 패턴의 차이에 의해 야기될 수 있는 최대 영양 생장률의 차이로 설명할 수 있다 . 예를 들어 질소를 고정하는 콩과식물인 Lespe deza cap it a t a 는 최대 생장률이 낮을 수 있다 (G ri me & Hunt 1975, 그림 3.2 ) . 이와 같이 최대 생장률이 낮으면 질소에 대한 경쟁 력은 낮지만 최대 생장률이 높은 종에 의해 초기에 이 식물의 생장 이 억제될 수 있다. 큰 종자를 생산하여 분산이 느리다는 사실과 함
께 최대 생장률이 낮다는 것은 이 종이 최대 수도에 이르는 데 30 년이 필요한 이유를 설명할 수 있다. 반대로 경쟁력이 약하지만 최대 생장률이 높으면 일시적으로 우 점할 수 있다 . 이러한 현상은 질소 첨가 실험에서 관찰되었다 . 이미 설명한 바와 같이 1 년생 식물은 다년생 식물에 비해 최대 생장률이 더 높다(제 3 장, 그림 3 . 2).1 년생 식물은 14 년 된 휴경지에서 실험적 으로 설정한 질소 구배에 따라 실험 첫해에 그 수도가 현저히 증가 하였으나 반면에 다년생 식물의 수도는 현저히 감소하였다 (T i lman 1987a). 따라서, 질소 첨가는 1 년생 식물에 의한 일시적인 우점 현상 울 초래하였고, 1 년생 식물은 실험 후 3-4 년 만에 생장이 느린 다년 생 식물로 교체되었다. Cedar Creek 에서 소규모로 교란된 지 소에서 는 바슷한 패 턴 이 나 타났다. 예 를 들어 poc ket g o p her 에 의 해 형 성 된 맨 땅의 mound 와 같 이 소규모의 교란된 장소에 처음 우점하는 식물은 매토종자가 풍부 한 1 년생 식물이다 (T i lman 1983, 1987a). 몇 년 이내에 이 식물들은 다년생 식물로 교체되는데, 다년생 식물 중 많은 수가 주변으로부터 영양 기관으로 침입한다. 이와 유사하게 실험적으로 새로 교란시킨 질소 구배구에는 첫해에 1 년생 식물이 우점하였고 다음 해에는 다 년생 식물이 우점하였다. 일년생 식물 중 일부 개체는 질소 함량이 많은 구에서 다년생 식물의 억압을 피하는데, 그 이유는 다년생 식 물이 드문 지소에서 이들이 발아하기 때문인 것 같다 . 이 일년생 식 물은 생장률이 빠르고 이들의 최대 크기에 가변성이 있기 때문에 매우 큰 개체로 발달하여 이웃하고 있는 다년생 식물을 억압하기도 한다. 그러나 이러한 현상은 드문 일이고 , 이 미소 지소에 다년생 식 물이 침입하여 1 년 혹은 2 년 이내에 1 년생 식물을 억압한다. 질소 함량이 높은 구에서 이런 기작을 보이는 가장 보편적인 종은 망초 와 금강아지풀이다.
교란되지 않은 질소 구배 실험구에서 많은 종들이 일시적으로 우 점하는 극적인 패턴을 보였다 . 14 년 된 휴경지의 기존 식생에 질소 를 첨가하면 첫해의 생육 기간에 Ambrosia ar t em i s iifo li a 의 상대수도 가 질소를 가장 많이 처리한 구에서 12 배나 증가하였다 . 그러나 두 번째 생육계절에는 Ambros i a 가 거의 사려져버렸다(그림 8.23). Bert e roa i ncana 도 이와 유사한 반응을 보였다. 이 두 종은 모두 질 소를 가장 많이 처리한 구에서 A g ro py ronrep ens 에 의해 교체되었다. 25 년 된 휴경지의 질소가 많은 구에서(그림 8.24), 애기수영은 첫 해에 상대수도가 증가한 후 감소하였으며, 1986 년에 다시 증가하였 댜 Panic u m o lig osan t hes 는 3 년 동안 수도가 증가한 후 감소하였고, 왕포아풀은 처음 4 년 동안 점진적으로 증가하였다. Ag ro p yro n rep en s 는 처음 5 년 동안 수도가 증가하였다. Schiz a chy ri u m sco p ar i um 의 상 대수도는 처음 2 년 동안은 질소 처리에 영향을 받지 않았고 그 이 후 감소하였댜 콩과인 Lespe deza cap it a t a 는 질소를 처리한 첫해에 상대수도가 감소하였으며, 그 이후 값이 그대로 유지되었다. 48 년 된 휴경지(그림 8.25) 와 토착 사바나 지역(그림 8.26) 에서도 이와 유사하게 보편종에 의한 일시적인 우점 패턴이 나타났다. 첫해 에 최대 수도를 보인 종도 있고 종에 따라서는 두번째, 3 번째, 4 번 째 혹은 5 번째 해에 최대 수도를 보였다. 따라서 천이 초기 상태인 지소에서부터 토착 사바나까지 그 범위가 걸쳐 있는 4 개의 모든 지 역에서 질소 첨가에 대한 죽각적인 반응은 3 년 후에 관찰된 패턴과 현저히 달랐고, 5 년 후에 볼 수 있는 반응과도 달랐다. 실제로 질소 첨가는 다양한 종들에 의해 연속적으로 일시적인 우점 현상을 초래 하였댜 실험이 시작된 뒤 5 년이 지날 때까지도 그러한 전이가 계속 되었다. Rubus, Rosa, 그리 고 Rhus 와 같은 목본식 물의 반응과, 휴경 지 에 서 토양 질소와 목본식물의 생물량 사이의 상관에 의하면, 질소 함량이
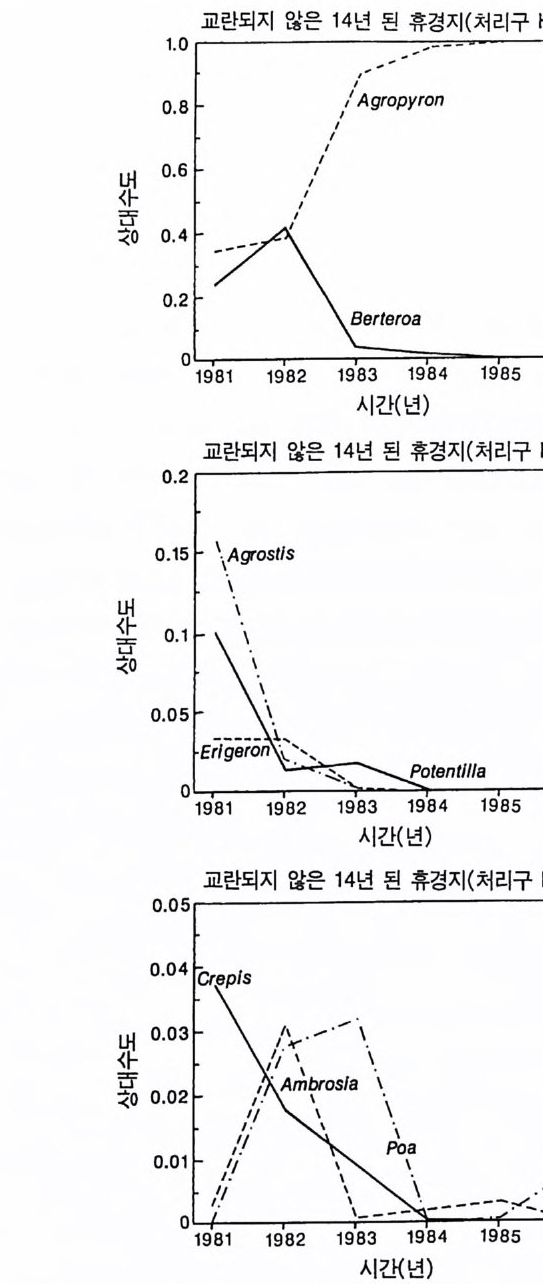 교란되지 않은 14 년 된 휴경지(처리구 H)
교란되지 않은 14 년 된 휴경지(처리구 H)
그림 8.23 CedarCreek 에 있는 교란되지 않은 14 년 된 휴경지의 H 구(질소 첨가가 많은 구)에 서 2 차 천이의 동테 1981 년의 자료는 실제로는 1982 년의 [구의 평균치이다. 이들 자료는 영양염류를 첨가하기 전의 식생에 대한 가장 근접한 추정값이다. 1982 년과 그 이후의 자료는 생육 기간의 말경에 영양염류 첨가 효과를 보여주는 것이다. 수도 가 가장 높은 종만을 표시하였다.
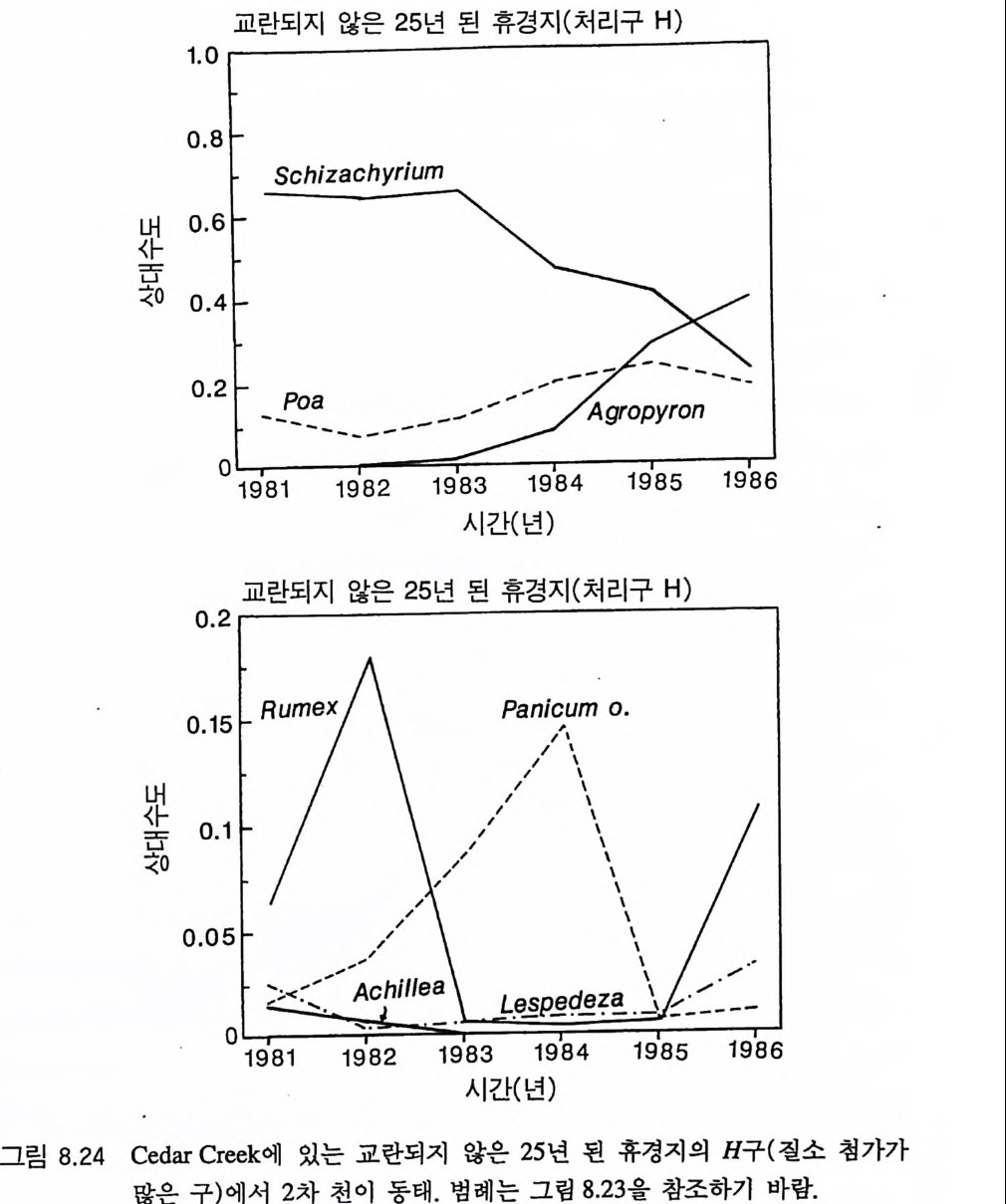 교란되지 않은 25 년 된 휴 경지(처리구 H)
교란되지 않은 25 년 된 휴 경지(처리구 H)
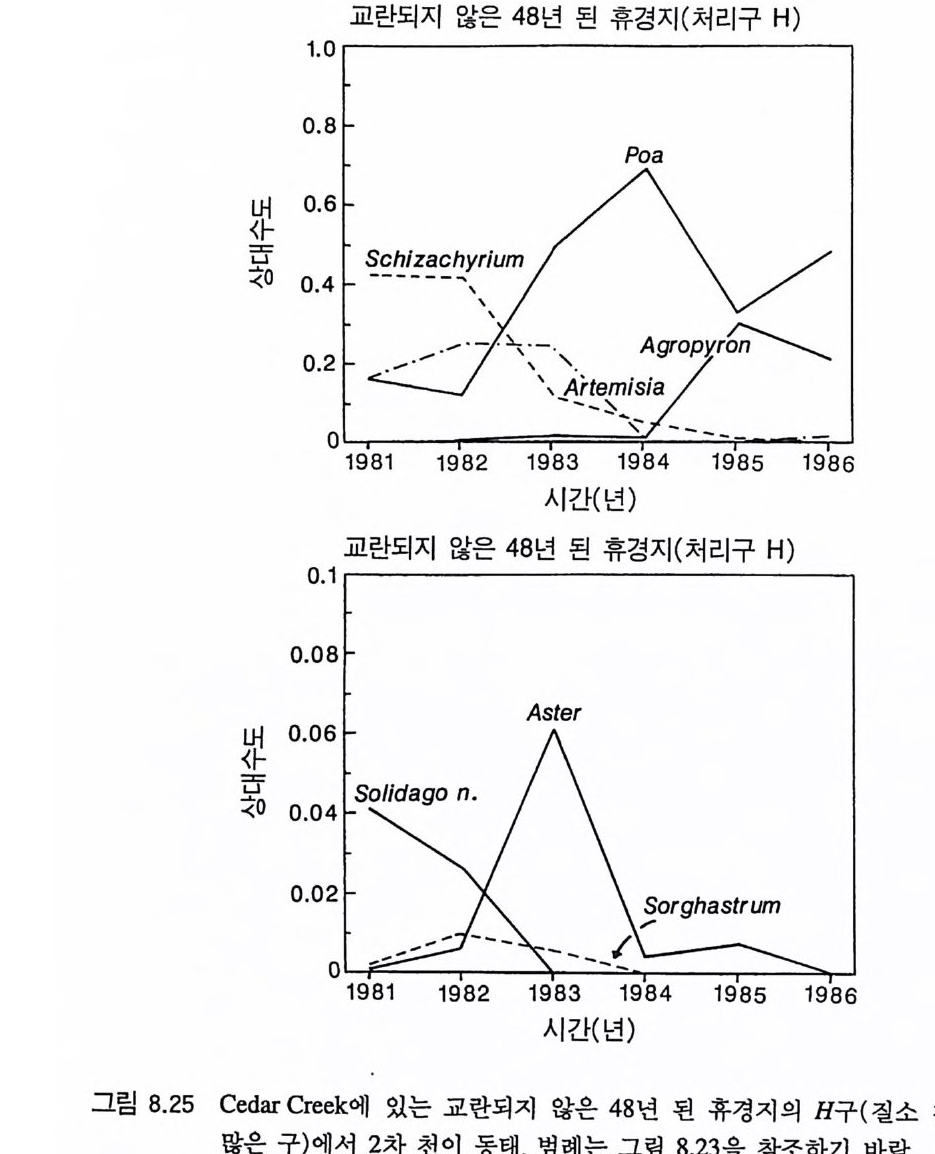 교란되지 않은 48 년 된 휴경지(처리구 H)
교란되지 않은 48 년 된 휴경지(처리구 H)
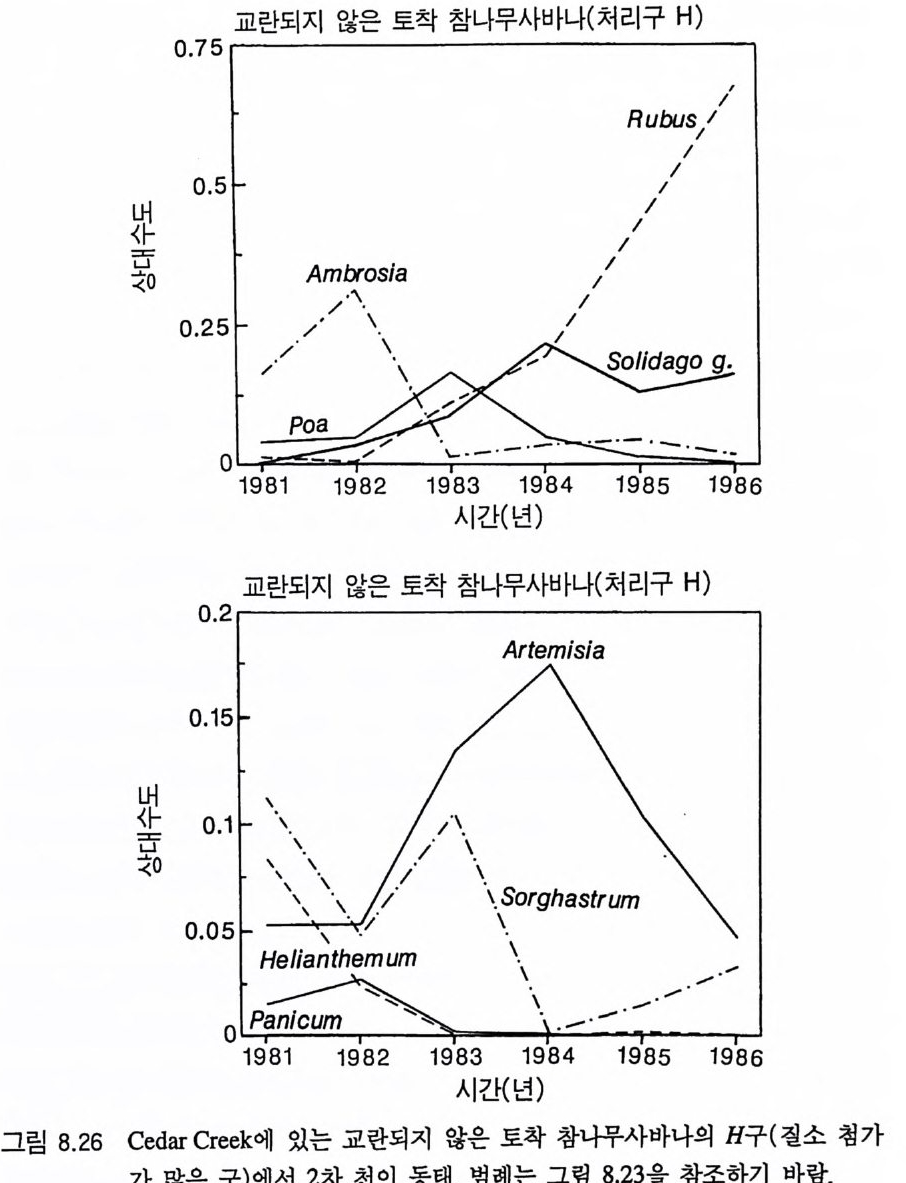 교란되지 않은 토착 참나무사바나(처리구 H)
교란되지 않은 토착 참나무사바나(처리구 H)
높은 구에서 최종적인 평형 군집은 목본식물. 특히 교목에 의해 우 점될 것으로 보인다. 만약 그렇다면, 이 실험이 평형 상태에 도달하 는데는 수십년이 필요할 것이며. 5 년째에 우점하는 종의 거의 대부 분은 일시적인 것으로 간주할 수 있다 . 실제로 6 년째에 수집된 자료 에 의하면 질소 함량이 높은 구에서 A g rop y ron 이 Rubus 에 의해 교 체되는 것을 알 수 있다 . 이러한 일시적인 동태는 제 6 장과 7 장에서 예측된 내용과 정성적 인 면에서 일관성이 있다. 최대 수도를 갖는 첫번째 종은 1 년생 식 물이나 수명이 짧은 다년생 식물이며. 이들은 최대 영양 생장률이 높다 . 그러나 5 년째에 우점하는 종은 생장률이 느리고 실험의 초기 에는 없었거나 드문 것들이었다. 14 년 된 휴경지에서는 실험 초기에 Ag ro py ro n rep ens 가 전체 생물량의 약 30% 를 차지하였는데, 생장이 빠른 종에 의한 일시적인 우점 기간이 1 년뿐이었다. 실험 초기에 A g rop y ron 이 드물거나 (<1%) 전혀 없는 지소도 포함되어 있었던 25 년 된 휴경지와 48 년 된 휴경지에서는 처음 3 년 동안 다른 종들 이 일시적으로 우점하였다. 이와 비슷하게 토착 참나무사바나에서는 비교적 생장이 느린 목본성 덩굴식물이 5 년 후에 우점하였는데, 실 험 중 처음 3 년 혹은 4 년 동안에는 다른 종들이 일시적으로 우점하 였다. 나는 CedarCreek 에 있는 9 종의 식물에서 토양 질소 수준에 대한 생장률의 의존도뿐만이 아니라 영양 생장률도 측정하였다 (T i lman 1986b). 이러한 영양 생장률은 일시적인 동태의 역할에 관하여 더 많은 증거룰 제공해 준다. 14 년 된 휴경지의 맨 처음 우점종안 Ambrosia ar t em i s iifo li a 는 비옥한 토양에서 12 주 동안 생장하는 중 Ag ro py ro n rep ens 보다 생물량이 2.5 배 많았다. 더구나 그 시기에 Ambros i a 의 키는 30cm 로 A g rop y ron 의 2 배 이상이 었댜 따라서 질소 구배 실험의 첫해 동안에 Ambros i a 의 생장률이 빨라 A g rop y ron 보다
키가 커 A g rop y ron 을 억압할 수 있다(그림 8.23). 그러나 2 년째부터 는 A g rop y ron 이 Ambros i a 를 교체하였는데, 그 이유는 A g rop y ron 의 뿌리와 지하경에 저장된 에너지와 영양염류가 생육계절 초기에 1 년 생 식물보다 더 빨리 자랄수 있도록 하기 때문인 것 같다. 질소 첨가 후에 절대수도는 증가하나 상대수도는 증가하지 않는 Schiz a chy ri u m scop ar i um 은 (T i lman 1987a) 온실에서의 생장 실험에서 9 종 중 생물량이 가장 적었다 (T il man 1986b). 질소 첨가 후에 상대수 도 면에서 이 종에 의한 일시적인 우점 기간이 없는 이유는 이와 같이 생장률이 낮기 때문인 것 같다. 그러나 이와 똑같은 온실 실험 자료들은 질소에 대한 평형 상태의 경쟁 능력이 천이 과정을 결정 하는 주요인이라는 가설을 뒷받침하기도 한다. 이러한 자료는 질소 가 적은 토양에서의 종의 생물량과 천이 과정 중 그 종의 출현 순 서 사이 에 매 우 유의 한 순위 상관이 있음을 보여 준다 (T i lman 1986b). 천이 초기종은 후기종에 비해 척박한 토양에서 더 많은 생물량을 생산한다. 교란된 구에서 관찰된 결과도 2 차 천이에서 일시적인 동태의 역 할을 뒷받침한다. 예를 들어 25 년 된 휴경지에서 교란되었지만 시비 를 하지 않은 구의 처음 우점종은 1 년생 식물인 금강아지풀과 . Polyg o num convolvulus 였는데, Poly g onum 이 금강아지풀에 비해 훨씬 풍부하였다. 이 두 종은 수명이 짧은 다년생 식물인 Ag ro stis s cabra 와 애기수영에 의해 교체되었고, 그 다음에는 생장이 느린 Schiz a - ch y r i umscop ar i um 에 의해 교체되었다. 질소 함량이 많은 구는 첫해 에 Poly g onum 이 금강아지풀에 비해 더 풍부하고, 금강아지풀이 2 번 째 해 에 최 대 상대 수도를 가지 며 왕포아풀과 Panic u m oli go santh e s 가 3 년째 해에 최대 상대수도에 이르고, A g rop y ron 이 4-5 년째에 우 점종이 되는 매우 다른 동태를 보였다. 척박한 토양이나 비옥한 토 양에서 모두 교란 후의 종의 교체 패턴은 생장이 빠른 종에서 생장
이 느린 종으로 전이됨을 알 수 있다 (T i lman 1986b, Gr im e & Hunt 1975). 또한 그림 7.5 에 요약된 이론에 의해 예측되는 바대로, 비옥 한 토양에서보다 척박한 토양의 2 차 천이에서 우점종의 괘적이 서 로 매우 다르다. 6 천이의 원인 Cedar Creek 에서의 이러한 천이 연구는 척박한 토양에서 2 차 천 이의 동태가 천이 과정 중 영양염류의 축적 패턴, 영양염류 구배상 에서 경쟁 능력의 분화, 종의 최대 생장률의 차이. 그리고 정착의 동 태에 따라 영향을 받음을 보여주고 있다. 이들 각각의 과정은 Cedar Creek 에서 천이 동태의 일부를 설명할 수 있지만 천이 과정 전체를 설명할 수는 없다. CedarCreek 의 영양염류가 빈약한 토양에서도 천이 동태의 대부 분은 일시적인 동태인 것 같다. 우점종의 순서를 보면 처음에는 Ambros i a 이지만 그 다음에는 Ag ro p yro n, 왕포아풀, Sch i zachy r i am 을 거쳐 마지막에는 Androp o g on 이 우점종이 되는데(그림 8.22), 이러한 현상은 중간에 있는 3 종이 질소 첨가 실험에서 보인 반응이나 각 휴경지 내에서 토양의 전질소에 대한 이들 종의 분포 결과와도 일 치하지 않는댜 생장이 빠른 A g rop y ron 이 일찍 우점하고 그 다음에 는 생장이 약간 느린 왕포아풀, 그리고 그 이후에는 생장이 느린 Sch i zachy r i am 이 우점하는데(그림 8.22), 이것은 이들 종들의 최대 생장률의 차이와 일치한다 (T i lman 1986b, Cowan 1986). 따라서 일시 적인 동태의 예가 될 수 있다. 만약 이것이 맞다면 일시적인 동태는 Cedar Creek 에서 처 음 30-40 년 동안 천이 과정 을 결정 하는 중요한 요인이 될 수 있다(그림 8.22).
CedarCreek 의 2 차 천이 동안 일시적인 동태의 중요성은 천이 과 정 중 뿌리:줄기 동태에 의해 뒷받침된다. 그림 7.5A 와 B 에 나와 있는 바와 같이 척박한 토양에서 일시적인 동태의 기간은 뿌리:줄 기의 비가 낮은 종이 뿌리 : 줄기의 비가 점차 커지는 종으로 교체되 는 기간으로 예상된다. Cedar Creek 의 혼합 식생에서 뿌리 : 줄기의 비를 측정하면 바로 이 패턴을 볼 수 있다. 1 년 된 휴경지에서 뿌 리 :줄기의 비는 약 1:1 이다. 15 년 된 휴경지에서 이 비는 5.2:1 이고. 50 년 된 휴경지에서는 11:1 이다(R. Mckane, 개인적인 연락). 이러한 사실은 A g rop y ron 과 같은 종은 뿌리:줄기의 비가 낮아 최대 생장률 이 높기 때문에 천이 초기에 우점하게 되고, 생장률은 느리지만 토 양 질소에 대 한 장기 간의 경 쟁 력 이 우세 한 Sch i zachy ri um 과 같은 종 에 의해 교체된다는 가설을 입증하는 것이다. 각 종의 뿌리:줄기의 비롤 고려할 때에도 이것과 같은 패턴이 유지된다. 야의의 서로 다른 토양에서 단독 재배할 때 CedarCreek 에서 가 장 많이 출현하는 5 종의 · 벼과식물은 척박한 토양에서의 뿌리:줄기 의 비를 갖는데 , 이것은 그들의 천이 계열이 일시적인 동태에 의해 결정된다는 가설과 완전히 일치한다(표 8.1 , Wed in & Til ma n, 미발표 자료). 이 5 종 중 가장 먼저 우-점종이 되는 Ag ro stis s cabra 는(그림 8.22) 뿌리:줄기의 비가 가장 낮다 . 가장 늦게 우점종이 되는 Androp o g on gerad i의 뿌리 :줄기의 비는 가장 높다(표 8.1 ). 또한 Cedar Creek 에 있는 8 종의 초본에 관하여 2 번째 해 에 수집 된 자료도 비슷한 패턴을 보였다 (T ilm an & Cowan, 미발표 자료). 각 종에서 관 찰된 뿌리:줄기의 비와 천이 과정에서 그 종이 우점하는 시간 사이 에 유의한 순위상관이 있었다 (r,=0.81, n=8, P<0.05). 이러한 여러 가지 증거들은 CedarCreek 에서 천이 계열의 상당 부분이 경쟁적 교 체의 일시적 동태임을 입증하는 것이다.
7 요약 미네소타의 Cedar Creek Natu r al Hi st o r y Area 의 휴경지는 경작을 그만둔 뒤 2 차 천이가 시작되었는데, 그 과정은 다음과 같은 요인에 의해 결정되는 것 같다. 첫째, 질소와 햇빛에 대한 종의 경쟁 능력. 둘째, 질소가 빈약한 토양에 질소의 점진적인 축적률, 셋째, 종의 최 대 생장률에 영향을 주는 분배 패턴 등이다. 정착률도 중요한 역할 을 할 수 있다 . 이와 똑같은 요인들, 특히 질소와 햇빛에 대한 경쟁 능력이 주어진 휴경지 내에서 종의 지역적인 수도에 영향을 준다 . Cedar Creek 에서 2 차 천이의 초기 과정 중에 우점하는 Ambrosia artem i si i foli a, Ag ro stis s cabra, 그리 고 Ag ro p yro n rep ens 와 같은 종은 영양 기관의 최대 생장률이 높고, 뿌리 : 줄기의 비가 낮아 천이 과정 의 후기에 우점하는 Androp o g on ger ardi, Rubus sp. , Sorgh a str u m nuta n s, 그리고 Ar tem i si a ludov ici ana 와 같은 종에 비해 질소가부족 표 8.1 C edarCreek 의 2 차 천이에서 중요한 협엽초본 5 종의 뿌리 : 줄기의 비는 각 종 울 질소 구배상에서 단독 재배하여 결정하였다. 토양의 질소가 낮은 상태에 서의 뿌리 : 줄기의 비는 토양의 전 질소 ( x ) 에 대한 뿌리 : 줄기의 비(y)의 회귀 식이 戶죽과 만나는 점이다 . 이것은 18 개의 1 년 된 단독 재배구의 자료를 이 용한 것이다. 천이의 순서는 그림 8 . 22 에서 자료를 얻었고 , 뿌리 : 줄기의 자료 는 D. We din과 D. T i lman 의 미발표 자료이다. 천이의 순서와 뿌리 : 줄기의 비 사이의 Sp ea nnan 순위상관은 r,=1 .0, N=5, P < 0 . 05 이다. 1986 년 자료를 근 거로 하였음. 종 천이순서 낮은 질소 함량에서 뿌리 :줄기 Agr o stis s cabra 12345 0.4 2 :1 Agr o py ro n repe n s 1.0 :1 Poa pra te n sis 1.2 :1 Schiz a chy rium scop a riu m 1.5: 1 Androp og o n ger adi 2.3 : 1
한 토양에서 더 빨리 자랄 수 있다. 따라서 척박한 토양에서의 2 차 천이는 많은 요인에 의해 주도되는 과정이다. 이 연구 결과에 의하면 척박한 토양에서 진행되는 2 차 천이는 1 차 천이와 비옥한 토양에서 진행되는 2 차 천이의 중간에 해당된다. 모든 천이 과정이 많은 요인에 의해 주도되지만 1 차 천이에서부터 비옥한 토양에서 진행되는 2 차 천이에 이르기까지의 연속선상에서 각 요인의 상대적인 중요성은 변화될 수 있다 .1 차 천이는 토양의 영양염류 축적 과정과 지표면에 입사되는 광량 및 광의 수직적 분 포에 크게 영향을 받는다. 토양의 발달은 바교적 완만한 과정이고 1 차 천이가 시작되는 기질은 영양염류가 극히 제한적이기 때문에 정착률과 경쟁적 교체의 일시적인 동태가 1 차 천이에는 큰 영향을 주지 못한다 . 반면에 비옥한 토양에서 진행되는 2 차 천이는 장기적 인 토양 동태에 의해서는 영향을 받지 않지만 다양한 종의 정착률 과 최대 생장률, 즉 일시적인 동태의 차이에 많은 영향을 받는다. CedarCreek 에서의 2 차 천이는 중간적인 경우에 해당된다. Cedar Creek 토양의 질소 함량이 낮고 질소의 축적률이 완만한 것이 이 지역에서 천이 초기종이 오래 잔존하고 천이 후기종 특히 목본식물의 우점을 지연시키는 중요한 요인인 것 같다. 많은 종들은 질소 축적 가설로는 설명할 수 없는 우점 기간울 갖지만, 일시적인 동태 혹은 정착이 천이에 영향을 준다는 가설과 일치한다. 이것은 천이 과정에서 식물의 정착이나 일시적인 동태, 그리고 영양염류의 축적이 갖는 상대적인 중요성을 결정하는 데 있어서 토양의 초기 영양염류 상태가 중요한 요인이라는 것을 의미한다.
제 9 장 문제점과 결론 앞의 여러 장에서 설명한 내용들은 나름대로 많은 문제점들을 가 지고 있다. 이 책의 전체 내용을 요약하기 전에 그 문제점들을 살펴 보도록 하겠다. 1 표현형의 가소성 제 2 장에서 설명한 바와 같이 식물이 갖는 형태적인 가소성은 식 물의 적응이란 측면에서 가치가 있다. 예를 들어 토양의 영양염류에 의해 제한을 받는 식물은 잎과 줄기로의 분배량을 줄이고 뿌리의 생물량 증가를 통한 영양염류의 흡수율 증가로 자신의 생장률을 증 가시킬 수 있다 . 이와 유사하게 햇빛에 의해 제한을 받는 식물은 자 신의 잎면적이나 키를 증가시켜 더 많은 햇빛을 흡수하여 생장률을 더 빠르게 할 것이다 .
생리적인 기작도 중요하다. 인에 의해 제한울 받는 식물은 뿌리에 있는 세포 표면의 흡수지점의 수를 증가시켜 더 많은 인을 흡수하 여 더 빨리 자랄 수 있다. 햇빛에 의해 제한울 받는 식물은 잎의 엽 록소나 단백질의 증가를 통해 자신의 광합성 효율을 증가시켜 더 빨리 자랄 수 있댜 비록 이러한 조정이 다른 자원의 이용 능력에 영향을 줄 경우에도 이들 자원이 현재 그 식물의 생장을 제한하지 않는 한 이러한 형태적 그리고 생리적 조정 능력은 그 식물에게 유 리 하다(I wasa & Roug hg ar den 1984, Bloom et al. 1985). 식물의 자원 획득 행동은 식물이 자신의 생리와 형태를 조절할 수 있는 능력으로부터 나온다. 필수 자원에 대한 최적 획득 이론에 의하면 식물은 모든 필수 자원에 의해 균등하게 제한을 받도록 자 신의 잠재적인 생장을 분배하고 생리를 조절해야 한다 (Ra pp o rt 1971, Covic h 1972, Til m an 1982, Iwasa & Roug hg ar den 1984, Bloom et al. 1985). Bloom 등(1 985) 은 식물의 생리적 가소성에 관한 많은 문헌을 검토 한 후 식물은 자신들이 여러 가지 자원에 의해 더 균등하게 제한을 받는 쪽으로 자신들의 생리를 조정한다는 것을 알았다. 그러나 다양 한 자연식생에서 수행된 영양염류 첨가 실험에 의하면 대부분의 종 은 여러 가지 영양염류들에 의한 복합적인 제한울 받는 것이 아니 고 오직 하나의 영양염류에 의해 제한을 받는 것으로 나타나고 있 다 (Lawes & Gi lb ert 1880, Mi lton 1947, Brenchley & Warin g ton 1958, Wi llis 1 963, Til m an 1980, 1982, 1983, 1987b). 그러나 현재까지의 연구 결과로는 자원 획득 이론에 의한 예측 사항들을 시험하는 데에는 충분하지 않다. 균등제한 가설을 완벽하 게 시험하기 위해서는 동시에 두 가지, 세 가지 그리고 그 이상의 영양염류를 첨가하는 모든 가능한 조합에서의 실험이 필요하다. 이 책에서 전개시킨 대부분의 이론에는 분명히 가변성이 포함되어 있 지 않다. 지금까지 이 책에서 내린 결론들은 표현형적으로 가소성이
있는 식물에게 적용할 수 있는 것들인가? 만약 식물이 표현형적으 로 가소성이 있다면 왜 단일종이 모든 서식처에 최적인 형태나 생 리를 갖지 않는가? 가소성으로 인하여 종간 경쟁의 과정이 너무 복 잡하게 되기 때문에 식물의 상호 작용을 설명할 수 있는 단순한 이 론은 나올 수 없는 것인가? 단일종의 생장이나 종간 경쟁에 미치는 형태적 가소성의 효과를 알아보기 위하여 나는 주어진 지점에서 각각의 식물이 현재 자신의 자원 제한 패턴에 의해 영향을 받는 분배 패턴을 갖도록 ALLOCATE 모델을 변형시켰다. 만약 식물이 현재 토양의 영양염류 에 의해 제한을 받는다면 그 식물은 뿌리로의 분배 비율을 증가시 키고. 잎과 줄기로의 분배비를 감소시킬 것이다. 만약 식물이 현재 햇빛에 의해 제한울 받는다면 그 식물은 잎이나 줄기로의 분배 비 율을 증가시켜 현 조건하에서 순생장률을 높게 하고 뿌리로의 분배 비율을 감소시킬 것이다. 이와 같은 방법을 통해 그 식물은 분배 패 턴이 고정된 개체보다 토양 자원과 햇빛을 더 최적인 상태로 이용 할 수 있을 것이다 . 생리적으로 동일한 식물들이 형태적으로는 가변적일 때 어떤 일 이 일어날까? 만약 특이한 서식처에서(예를 들면 영양염류의 공급률 이 일정하고 손실률이 일정한 서식처) 하나의 개체군이 단독으로 자란 다면 평균 분배 패턴은 식물의 크기(높이)에 의존한다(그림 9.1 ) . 또 한 분배는 그 서식처의 영양염류 공급률에도 의존적이다. 영양염류 가 빈약한 서식처에서 (TN=SO), 잎으로의 분배는 45-75% 범위이 며, 뿌리로의 분배는 10-55% 범위이다. 비록 이 값은 식물의 키에 따라 상당한 변이가 있지만 이러한 범위는 비옥한 토양 (TN=200) 에 서 자라고 있는 같은 종의 키 의존적 변이 범위와는 거의 겹치지 않는다. 주어진 생리적 조건이 일정할 경우 더 비옥한 토양에서는 뿌리로의 분배율이 낮고 잎이나 줄기로의 분배율이 높은 것이 유리
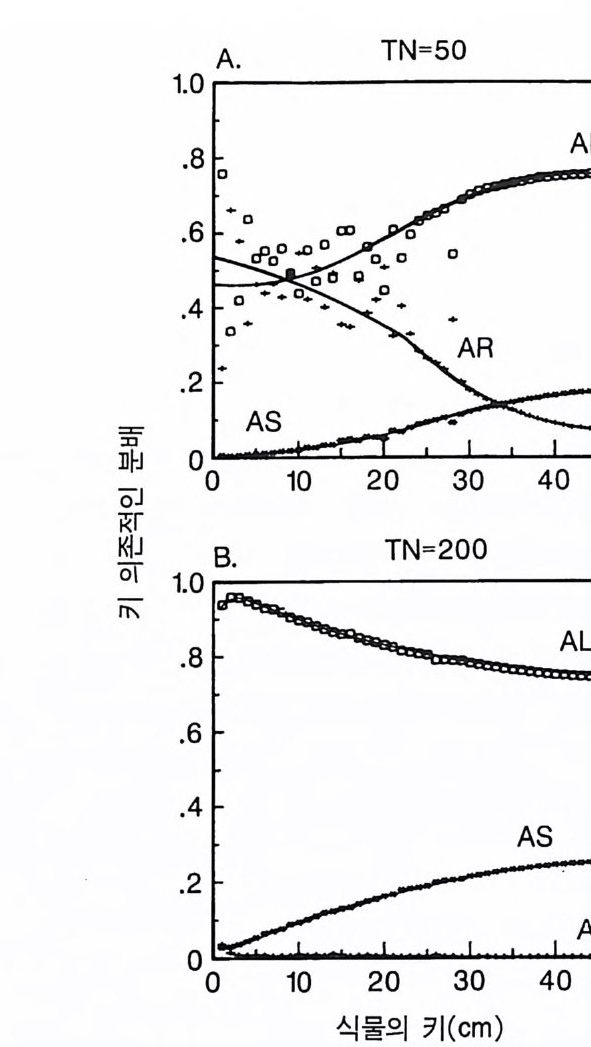 1.0 |A . TN=50
1.0 |A . TN=50
그림 9.1 (A) 형태적으로 가소성이 있어 자원 이용도에 따라 자신들의 분배 패턴 을 변형시키는 종이 영양염류가 반약한 토양에서 (1N=50) 보이는 키 의 존적인 분배. 키 의존적인 잎으로의 분배 (AL), 뿌리로의 분배 (AR), 줄기 로의 분배 (AS) 는 장기간의 평균 분배 패턴이다 . (B) 비옥한 토양 (1N=200) 에서 같은 종의 분배 패턴 척박한 토양에서의 분배 패턴과 비 교할 때 이들 개체들은 잎에 훨씬 많이 분배하며 . 줄기로는 다소 많이 , 그 리고 뿌리로는 훨씬 적게 분배함을 주목하기 바란다. 결과들은 개체들이 현재의 생물량 증가율을 최대가 되도록 유도하는 기관에 비례적으로 더 많은 분배를 하도록 변형시킨 ALLOCATE 모델을 근거로 한 것이다.
하댜 형태적으로 가소성이 있는 두 식물이 경쟁을 한다면 어떤 일이 일어날까? 만약 두 식물의 생리가 같고 같은 방법으로 형태적 적응 울 하고 있다면 그들은 기능상으로 동일할 것이다. 그러나 이 두 식 물체의 생리가 다르다면 어떤 일이 일어날까? 두 식물 중 하나는 새로운 조직을 생산하는 데 영양염류가 적게 필요하기 때문에 토양 영양염류의 요구량이 낮다고 해두자. 이 종을 A 라고 하자. 이 종의 다양한 분배 반응이 그림 9 .1에 나와 있다. 다른 종 B 는 광 요구량 이 낮댜 즉 낮은 광 수준에서 광합성률이 높다. 이 두 종은 모두 동 일한 영양염류 및 광포화 광합성률을 가지고 있고, 종자의 크기가 같고 , 성숙하였을 때 키가 같다. 또한 호흡률이 동일하고 줄기 무게 와 키 사이의 상대 생장 관계가 같다 . 이 두 식물이 경쟁을 하게 되 면 가소성이 없을 때 일어날 수 있는 것과 똑같이 영양염류가 적은 서식처에서는 A 종이 가장 많고, 영양염류가 풍부한 서식처에서는 B 종이 많다(그림 9.2). 종간 경쟁이 두 종 모두에서 형태적인 변화의 폭을 축소시켰다. 종간 경쟁이 없을 때 그것의 형태와 비교해 보면, 종간 경쟁은 각 종으로 하여금 두 종 사이에 내재하고 있는 생리적인 차이룰 강화 하는 분배 패턴을 갖도록 한다. 예를 들어 영양염류 면에서 경쟁력 이 좋은 A 종은 종간 경쟁에 의해 뿌리로의 분배가 많은 쪽으로 분 배 패턴이 변화된다. 경쟁하는 종의 수가 많을수록 종들이 더 비슷 해지고, 종간 경쟁에 의해 더 많은 제한울 받으면 각 종은 더 좁은 서식처 범위로 제한되어 분배 패턴도 더 제한될 것이다. 영양염류 공급 구배에 따라 형태적인 가소성이 경쟁에 영향을 줄 수 있지만 다음과 같은 몇 가지 이유에서 앞의 여러 장에서 설명한 결론에는 큰 영향을 주지 못할 것이다. 첫째, 가소성이 종의 생존을 지속시켜 영양염류 구배에 따른 종의 상대수도를 변형시킬지라도
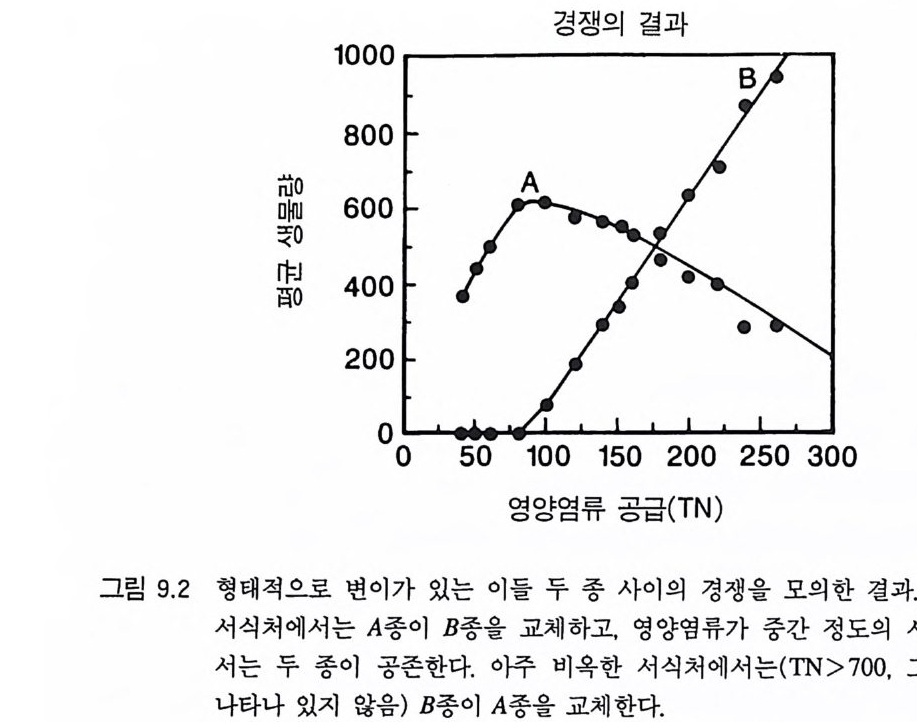 경쟁의 결과
경쟁의 결과
만약 모든 종이 똑같은 가소성을 갖는다면 이러한 구배상에서 가소 성이 종 분포의 정성적인 패턴을 변화시키지는 않을 것으로 보인다. 둘째, 가소성 그 자체는 손실이 따른다. 위에서 언급한 모델은 손실 이 없는 것으로 간주되어 있다. 환경 조건을 계속 모니터하고 환경 변화에 반응하여 형태적 혹은 생리적 적응을 하는 데 손실이 따른 다면 그러한 손실은 가소성이 갖는 이점을 감소시키고 가소성의 범 위를 제한시킬 것이다. 셋째, 경쟁하는 종의 수가 많을수록 각 종 내 에서 생존 가능한 형태의 범위는 줄어들 것이다. 넷째, 모든 종은 그 들의 형태적인 가소성에 구조적으로 부여된 한계를 가지고 있다. 그 러나 위에서 제시된 가소성의 모델은 단일 유전자형이 모든 가능한 형태를 생산하는 데 충분한 가소성을 갖는 것으로 가정하고 있다. 따라서 단일 유전자형이 단세포인 토양 조류나 이끼와 같은 식물 ,
초본. 목본성 덩굴식물 그리고 교목이 될 수 있는 것으로 간주된다. 식물은 형태적 그리고 생리적으로 가소성을 갖지만 주어진 종 내 에서의 가소성의 범위는 서로 다른 종간의 형태적 그리고 생리적 변 이의 범위에 비교하면 미미한 것이다. 토양 조류는 절대로 이끼와 같 은 형태를 갖을 수 없다. 협엽초본은 관목의 형태를 갖울 수 없으며. 관목은 교목이 될 수 없다. Ambrosia ar t em i s iifo l i a 나 Chenop o diu m album 과 같은 천이 초기의 1 년생 식물들은 생육계절 말기의 키가 생육장소에 따라 5 배 정도의 차이가 있다. 그러나 이들을 구성하는 물질과 이 구성 물질들이 배열하는 방법은 관찰된 최대높이보다 훨 씬 큰 개체를 지탱할 수 없다. 예를 들어 키가 5m 나 되는 Ambrosia ar t em i s iifo li a 는 구조적으로 불가능하다. 그렇게 키가 크면 자신의 무게 때문에 구부러질 것이다. 뿌리:줄기의 비율도 형태적으로 가소성이 있지만 질소 이용도에 서 15 배의 차이가 있는 Cedar Creek 의 질소 구배상에서 자라는 8 종 의 식물에서는 그 비가 2.5 배 이상 차이가 있지 않다 (Cowan 1986, Cowan & Til ma n, 원고 준비중). 이와 비슷하게 Chapi n(19 80, 236 쪽)은 〈부족한 영양염류의 이용도를 100 배 낮추면 뿌리:줄기의 비가 1.5 - 12 배 증가하지만, 증가된 뿌리:줄기의 비와 뿌리의 홉수능 증가의 상보적인 효과는 감소된 영양염류 이용도를 완전히 보상하지 못한 다〉고 주장하였다. 따라서 형태적 그리고 생리적 가소성에는 한계가 있다. 이러한 한계 때문에 비교적 좁은 범위의 서식처 유형에서만 식물이 경쟁 우세종이 되도록 제한된다. 키가 Im 인 참나무는 분명히 구조적으로 불가능한 것은 아니다. 그러나 주어진 서식처에서 참나무가 성숙하였을 때의 최대 키가 Im 라면 참나무 실생의 정상적인 목본 줄기를 생산하는 개체들은 좀더 경비가 적게 드는 줄기를 생산하는 개체와의 경쟁에서 패배하게 될 것이댜 실생이나 유목일지라도 리그닌화가 훨씬 잘 되고 밀도가 높
은 줄기는 식물의 키가 크는 데 필수적이다. 왜냐하면 그 식물이 성 숙하였을 때 자신을 지탱하기 위해서는 식물의 줄기 전체에 그와 같이 강한 구조적인 조직이 형성되어야 하기 때문이다 . 식물이 자신의 키를 Im 혹은 2m 까지 성장시키는 데는 가늘고 팽 압에 의해 유지되는 줄기를 생산하고, 자신의 키가 증가함에 따라 이 줄기를 밀도가 높고 고도로 리그닌화된 물질을 갖는 줄기로 바 꾸는 것은 불가능하다. 따라서 줄기와 가지를 구성하는 데 사용하는 물질의 성질은 그 개체가 갖는 최대 크기에 따라 처음부터 결정되 어야 한다. 성숙하였을 때 키가 작고 수명이 짧은 식물은 성숙시 키 가 큰 식물에게는 적당하지 않은 조직과 방법(팽압과 같은)을 사용 하여 최대 키를 유지할 것이다 . 현존하는 육상 관속식물은 약 4 억 년 전에 단세포 조류로부터 유 래되었다. 토양의 영양이 풍부해짐에 따라 토양의 조류층은 두꺼워 지고, 그러자 키가 큰 개체가 경쟁에 유리하게 되었다. 예를 들어 초 기의 토양 조류 중에는 몇 개의 세포는 토양 속에 있고 몇 개의 세 포는 지상부로 돌출된 형태를 갖는 것이 있을 수 있다. 그러나 지상 부에 있는 세포와 지하부에 있는 세포 사이에 영양염류와 광합성 산물의 확산이 너무 느려 불과 몇 개의 세포 크기 이상으로 클 수 는 없었다. 따라서 키가 현저히 증가하기 위해서는 구조적인 물질과 뿌리와 잎 사이에 영양염류 및 광합성 산물을 수송하는 관속계가 필요하게 되었다 . 그러나 키가 작은 식물에게 알맞는 기능을 하는 관속계는 키가 큰 식물에게는 그 기능이 충분하지 않을 수 있다. 식 물체로 하여금 키가 10cm 에 이르도록 할 수 있는 다양한 관속계가 있지만 . 이들 관속계 중에는 키가 40m 인 식물에게 알맞는 것은 거 의 없었다 . 지지 작용과 수송 작용을 하는 각각의 관속계는 각자 경비가 소 요된다. 주어진 어느 개체군에서 자연 선택은 각 관속계를 생산하는
비용과 각 관속계와 연관된 이익(관속계의 존재로 인한 사망률 저하. 효율적인 수송을 통한 생장 증가 등)의 범주 내에서 가장 큰 순생식률 을 갖도록 하는 지지 및 수송계를 갖는 개체에게 유리하게 작용하 였을 것이댜 주어진 개체군에서 이러한 최적 관속계를 결정하는 가장 주요한 요인은 개체가 성숙하였을 때의 최대 키일 것이다. 그 이유는 줄기 에 미치는 하중과 줄기가 자신의 하중으로 인해 구부러지는 경향을 결정하는 것이 식물의 키이기 때문이다. 일반적으로 성숙하였을 때 의 키가 비교적 작은 식물들은 경비가 적게 드는 줄기를 갖는 것이 유리할 것이다. 경비가 덜 드는 줄기란 줄기를 구성하는 물질의 단 위 건량당 소요되는 경비가 아니고 줄기의 단위 길이 (m) 당 경비를 말한다. 줄기의 단위 길이당 첨가되는 구조 물질을 위한 경바는 육상식물 에서 형태적인 가소성의 범위를 제한하는 중요한 요인이다. 자신이 생산할 수 있는 구조 물질을 저장하고 있는 각 개체는 비교적 좁은 범위의 최대 키에서만 경쟁 우세종이 되도록 제한을 받는다. 자신의 최적보다 작은 키에서는 그 높이에 적당한 저렴한 줄기를 생산하는 개체와의 경쟁에서 지게 될 것이다. 키가 큰 상태에서는 하중 증가 로 인한 손실 혹은 비효율적인 수송으로 인한 손실을 견뎌야 한다. 따라서 형태적으로나 생리적으로 가소성이 있는 개체가 가소성이 없는 개체들보다 유리하긴 하지만, 그러한 가소성의 잠재적 범위는 식물이 갖을 수 있는 가능한 형태적 그리고 생리적 범위에 비교하 면 좁다. 이것은 서식처의 전범위에서 다른 모든 유전자형을 교체할 수 있는 형태적으로 가소성이 있는 단 하나의 유전자형이 진화한다 는 것이 거의 불가능함을 의미한다. 더구나 가소성이 포함될 경우에 도 경쟁의 정성적인 결과는 변해야 할 필요가 없다. 따라서 식물이 가소성이 있건 없건 간에 토양 자원:광 구배, 그리고 손실률 구배상
의 위치에 따라 형태 적 으로나 생리적으로 유리한 형이 서로 다를 것이다. 앞 장에 제시된 주요 결론들이 모두 가소성에 의하여 영향 을 받을 것 같지는 않다. 가소성에는 두 가지의 중요한 장점이 있다. 첫째, 식물이 고정된 형태를 갖을 경우 경쟁에서 불리한 서식처, 죽 그 식물의 자원 의존 적 생장 등구배곡선의 코너 부분에서 멀리 떨어져 있는 서식처에서 가소성은 그 식물에게 경쟁 능력을 증가시킬 수 있도록 해준다. 식 물의 경쟁력이 가장 큰 서식처 , 즉 그것의 생장 등구배곡선의 코너 에 자원 이용도를 갖는 서식처에서는 가소성이 그 식물의 경쟁력을 증가시키지 않는다는 것을 기억할 필요가 있다(그림 2.5 , 그림 9.2). 만약 식물의 서식처가 평형 상태에 있고 , 공간적으로 균일하다면, 각 식물종의 분포는 그것의 경쟁력이 좋은 지역, 즉 그것의 생장 둥 구배곡선의 코너 근처에 자원 이용도를 갖는 지역에만 국한될 것이 댜 식물의 서식처는 공간과 시간에 따라 매우 이질적이기 때문에 식물 한 개체가 경험할 수 있는 서식처는 상당한 변이가 있다. 만약 종자가 그 식물에게 최적 조건이 아닌 서식처에 떨어진다면 가소성 은 그 식물체로 하여금 나쁜 조건에 순응하도록 할 것이다 . 가소성의 두번째 장점은 식물이 사는 서식처의 장기간에 걸친 평 균 자원 수준과는 동떨어진 수준의 자원을 이용하게 한다는 것이다. 가소성은 일시적으로 모든 자원의 수준이 높은 서식처에서 식물의 생장률이 극대가 되도록 자신의 형태를 조정할 수 있도록 하여 자 신의 적합도를 증가시킨다 . 따라서 가소성은 식물체로 하여금 좋은 상황이 되도록 해준다. 종내에서 가소성의 범위를 측정하는 것은 그 종이 진화 과정을 거치는 동안 경험한 자원 이용도의 변이를 측정 하는 것일 수도 있다. 형태적인 가소성의 효과는 야초의 진화에 대한 단서를 제공할 수 있다 . 제 7 장에서 설명한 바와 같이 현재 농업에서 야초로 . 간주되거
나 2 차 천이의 초기 단계에서 흔히 볼 수 있는 많은 종들은 영양염 류가 빈약한 서식처에서 진화된 것으로 보인다. 그러한 종은 비옥한 토양에서 진화된 종들에 비해 부족한 토양 영양염류에 대한 생리적 인 요구량이 낮아야 하지만 햇빛에 대한 생리적인 요구량은 많아야 한댜 만약 이들이 햇빛이 많고 개방된 비옥한 토양에서 출현한다면 이들은 영양염류 요구량이 큰 식물에 비해 뿌리는 적고 잎과 줄기 가 많게 자신의 형태를 조절할 수 있을 것이다. 이와 같이 영양염류 가 풍부한 토양에서 잎으로의 분배가 많은 종은 햇빛이 많고 비옥 한 토양에서 햇빛에 대한 경쟁력은 우수하지만 영양염류에 대한 경 쟁력은 불량한 종보다 최대 생장률이 더 높도록 할 것이다. 형태적 그리고 생리적 가소성에 대하여 앞으로 밝혀야 할 것이 많이 있으며, 그러한 가소성이 종간 그리고 종내 경쟁에 미치는 효 과에 대해서도 많은 연구가 필요하다. 이 논의에서는 다른 종의 존 재가 한 종의 개체들 내에서 생리적 그리고 형태적인 가소성의 범 위를 제한하는 것으로 작용한다고 주장하지만 이러한 논의는 이론 적인 것이댜 우리는 식물 가소성의 범위와 환경과의 상관에 대한 더 많은 실험자료가 필요하다. 또한 종내와 종간에서 이러한 가소성 의 실험 연구가 더 필요하다. 그리고 형태적, 생리적인 가소성이 식 물 군집의 구조와 동태에 미치는 영향을 결정하기 전에 좀더 확고 한 이론적인 근거를 확보해야 한다. 2 공간적 구조 이 책에 제시된 모델은 서식처가 수평적으로 균일하여 어느 한 서식처에 사는 모든 식물은 토양 자원의 이용도가 같고 동일한 수 직적 광구배를 경험하는 것으로 가정하고 있다. 그러나 실제 서식처
는 공간적으로 패치를 형성한다. 그러한 공간적인 패치는 식물이 살 고 있는 장소의 토양 모재의 차이에서 비롯되는 토양 영양염류의 차이에 의해 야기될 수 있으며, 지소에 따른 영양염류 이용도나 햇 빛의 입사량에 대한 각 식물의 반응이 서로 달라 야기될 수도 있으 며 (Til m an 1982, Pacala & Sil a nder 1985, Pacala 1986), 지 역 적 인 교란의 차이에 의해서 야기될 수도 있다. 이러한 이질성이 식물 군집의 구 조에 어떤 영향을 주는지 생각해 보자. 토양의 영양염류 풍부도가 지소에 따라 이질적이기 때문에 각 지 소마다 유리한 식물종이 다룰 것이다. 시간이 지나면 각 식물은(기 후와 교란 체제가 고정되어 있다면) 자신이 경쟁에 유리한 토양형과 밀접한 관련을 갖게 될 것이다. 그러나 이러한 모델이 공간적으로 타당성이 있다면, 각 미소 서식처는 그 속에 그 조건에서 경쟁에 우 세한 한 두 종의 식물뿐만이 아니라 다른 일시적인 종들도 포함할 수 있다(도서 생물지리학적인 관점에서). 이러한 일시적인 종은 바로 인접해 있는 상이한 토양에서 경쟁 우세종이 되는 종들일 가능성이 크다. 이들은 그 지소에 이입되는 비율과 경쟁적으로 교체되는 비율 이 평형을 유지하기 때문에 그 서식처에서 지속할 수 있을 것이다 (Rick lefs 1987). 제 6 장에서 제시한 바와 같이 만약 경쟁에 의한 교체가 서서히 일 어나는 과정이라면 상당히 균일한 서식처는 자원이 부족한 서식처 에 비해 훨씬 많은 종을 포함할 수 있을 것이다. 이러한 서식처에서 는 희귀종이 일시적인 종으로 유지되는데, 그 이유는 그들의 이입률 이 지역적으로 소멸되는 비율과 균형을 유지하기 때문이다. 그러나 전체 경관 내에서 이러한 일시적인 종이 장기간 유지되는 것은 각 종이 우세한 경쟁자가 되는 서식처 조건이 존재하기 때문일 것이다. 두번째로는 서식처의 토양이 공간적으로 균일하지만 햇빛을 차단 하는 면이나 토양의 영양염류를 흡수하는 면에서 식물의 특성이 달
라 자원 이용도에서 지소에 따른 이질성을 유발하는 경우이다. 이러 한 견해에 의하면 서식처의 이질성은 각 식물의 성질이 다르기 떄 문에 나타나는 직접적이고 피할 수 없는 현상이란 것이다. 각 식물 은 시간과 공간상으로 서로 다른 특정한 지역에 존재한다. 식물은 생활사 기간 중에 자신이 있는 지역과 그 지역 내에 있는 다른 식 물에게만 영향을 준다. 또한 식물의 특성이 다르기 때문에 공간적으 로 이질적인 서식처가 만들어지는 것은 필연적이다. 한 그루의 나무가 죽으면 햇빛과 영양염류 이용도가 높은 지소인 숲톰이 형성된다. 따라서 모든 서식처는 그 내부에 햇빛과 영양염류 이용도가 그 서식처의 평균보다 높은 숲톰들을 항상 포함하고 있다. 이것은 햇빛과 영양엽류 이용도가 평균보다 높을 때 생장이 더 양 호한 식물(평형 상태의 손실률이 높은 서식처에서 경쟁력이 더 좋은 식 물들)로 하여금 서식처의 평균 조건에서 경쟁이 우세한 종들과 오랫 동안 공존할 수 있도록 해준다. 따라서 서식처는 하나 혹은 여러 그 루의 나무가 죽어서 갓 형성된 숲툼에서부터 폐쇄 단계에 있는 오 래된 숲툼에 이르기까지 천이의 단계가 상이한 미소 서식처들의 모 자이크로 볼 수 있다. 각 식물의 성질이 다르기 때문에 그리고 각 식물이 공간적으로 고정된 지역에 위치함으로써 나타나는 어쩔 수 없는 결과로 자원 이용도에서 공간적인 이질성을 낳게 된다. 따라서 부족한 자원이 있는 지역보다 더 많은 종이 공존할 수 있도록 한다. 주어진 자연 서식처는 지소에 따라 이질적인 토양울 갖울 수 있 는데, 이것은 각 식물체의 성질이 달라서 야기될 뿐만 아니라 식물 체가 직접 원인이 되지 않는 경우도 있다. 이 두 과정은 모두 서식 처의 종 다양성과 식생의 구조를 설명하는 데 중요하다 . 영양염류의 수준은 낮지만 햇빛 이용도는 높은 서식처로부터 영양염류의 수준 은 높지만 광량은 낮은 서식처에 이르는 구배상에서 지점에 따른 토양의 이질성 때문에 미소 서식처들의 위치가 서로 다르다 . 공간적
이질성이 있기 때문에 서식처는 손실률 구배를 갖는데, 영양염류 수 준과 광 이용도가 높은 최근에 교란된 지역으로부터 영양염류와 광 이용도가 낮은 오래된 지역에 이르는 구배상에 미소 서식처들이 위 치하게 한다. 모든 서식처에서 볼 수 있는 영양염류 공급률과 손실 률 구배는 부족한 자원이 있는 지역보다 더 많은 종을 유지할 수 있도록 해준다. 이 러 한 간단한 설 명 은 〈 이 웃식 물과의 경 쟁 ( neig h borhood comp et i -tion )> 개념의 효과를 이 모델에 포함시키는 두 가지 방법을 제시하 고 있다. 가장 간단한 방법은 식물 개체의 성질이 다르기 때문에 자 원 이용도와 손실률 면에서 공간적 이질성이 초래된다고 간주하는 것이댜 이러한 이질성은 식물 경쟁의 평형 모델과 함께 주어진 서 식처에서 지속 가능한 종 특성의 범위를 나타내는 데 사용될 수 있 다. 다른 하나의 방법은 각 종의 위치가 주어지고 (x, y 좌표상에서), 그 식물이 경험하는 자원 이용도가 그것의 이웃에 있는 모든 식물 의 위치, 크기 그리고 소비율에 의해 영향을 받는 뚜렷한 공간적 모 델을 만들 수 있다. 두번째 방법은 숫자상으로 복잡하고 귀찮지만 이 방법을 통해 중요한 지식을 얻을 수 있다. 3 교란과 영양염류 공급률 사이의 상관 나는 서식처의 손실률과 그 토양의 영양염류 공급률을 마치 이들 이 독립적이고 관련이 없는 과정인 것처럼 다루었다. 그것은 이 책 에서 개발된 이론을 자연적 혹은 실험적으로 조작된 식물 군집에서 의 패턴과 쉽게 비교하도록 하기 위한 것이었다. 서식처의 손실률과 그 토양의 영양염류 풍부도 사이에는 어떤 상관이 있어야 할 필요 가 없지만 손실률과 토양의 영양염류 공급률은 자연 상태의 군집이
나 인위적인 군집에서 서로 연관성이 있기 쉽다. 서식처에 따라 어 느 정도까지는 일관된 상호의존 패턴이 있으며, 그러한 연관은 전 세계의 자연 식생에서 패턴을 유발시키는 또 하나의 요인이 될 수 있다. Oksanen 등 (1981) 은 초식동물에 의해 야기되는 단위 식물량당 손 실률은 생산력이 극히 낮은 서식처에서 낮아야 한다고 가정하였다. 그 이유는 그 지역에는 지속적으로 초식동물을 유지하는 데 필요한 충분한 식물량이 없기 때문이다. 생산성이 중간 정도인 서식처에서 는 단위 식물량당 손실률이 가장 높은데, 그 이유는 높은 생산성이 초식동물을 유지할 수 있지만 초식동물의 양이 적어 포식자가 드물 기 때문이다. 생산력이 매우 높은 서식처에서는 초식동물을 많이 유 지할 수 있어 지속적으로 포식자 개체군을 유지할 수 있다. 그러나 포식자는 초식동물을 잡아먹음으로써 이들을 포식자가 없을 때보다 낮은 밀도로 제한하게 될 것이다. 따라서 생산력이 높은 서식처에 서는 생산력이 중간 정도인 서식처에 비해 · 단위 식물량당 손실률 이 더 낮게 된다. 식물이 경험하는 특정한 손실률과(단위 생물량당 손실률) 서식처의 생산력 사이의 정성적인 관계가 그림 9.3A 에 나와 있다 . 자연 군집에서의 영양 구조가 Oksanen 등이 가정한 것과 같다면, 이것은 손실률이 생산력과 관계가 없는 경우에 비해 자연식생에서 생산력과 식생 구조 사이에 매우 상이한 관계를 유발할 수 있다. Oksanen 등(1 981) 에 의하면 식물의 형태는 그림 9.3B 에 나와 있는 것과 같이 생산력에 의존적이어야 한다. 이것은 영양염류가 중간 정 도로 풍부한 토양에서 우점하는 식물은 척박한 토양이나 비옥한 토 양에서 우점하는 식물에 비해 식생의 최대 생장률이 훨씬 높아야 함을 의미한다(그림 9.3 C ). 또한 그 식물들은 손실률이 일정하고 생 산력과 무관하다는 가정에 근거하였을 때 예상되는 형태와는 매우
 A 초식과생산성 D 교란과생산성
A 초식과생산성 D 교란과생산성
그림 9.3 (A) Oksanen 등(1 981) 에 의해 가정된 서식처의 생산력(서식처의 영양엽 류 공급률)과 초식동물에 의해 야기되는 손실률(단위 생물량을 기준함) 사이의 관계. (B) 이 그림의 A 부분에서 나타난 것처럼 손실률과 토양의 영양염류 공급률이 상호 의존적이라면 식물의 형태는 토양의 영양염류 풍부도에 의존적일 것이다. 죽 작고 뿌리가 많은 식물은 척박한 토양에서 유리하고 , 키가 다소 크고 잎이 많은 식물은 중간 정도의 비옥도를 갖는 토양에서 유리하며. 줄기에 분배량이 많은 키가 큰 식물은 비옥한 토양에 서 유리하다. (C) 이들 토양에 유리한 분배 패턴은 중간 정도의 비옥도를 갖은 토양에서 우점하는 종이 척박하거나 비옥한 토양에서 우점하는 종 보다 훨씬 높은 최대 생장률을 갖도록 한다. (D) 교란 혹은 손실률과 영 양염류 공급률의 상호 의존도에 대한 또다른 가정 . (E) 특정한 서식처에 서 경쟁 능력을 우세하게 하는 분배 패턴에 대한 D 부분 관계의 영향. (F) 영양염류 공급률 구배를 따라 최대 생장률에 대한 이들 분배 패턴의 효과. 이것들은 ALLOCATE 를 이용한 모의에 근거한 것이다.
다른 형태를 갖울 것으로 예상된다. 식물의 손실률과 토양의 영양염류 공급률은 몇 가지 이유에서 상 호 의존적이라고 할 수 있다. 예를 들어 산불의 빈도는 생산력에 의 존적이다 . Wh it ney (1986) 는 미시간 주의 Roscommon 과 Crawf ord 군 에 있는 정착 전의 삼림에서 토양, 삼림형, 그리고 산불이나 숲틈을 형성하는 교란 사이의 관계를 알아보기 위해 초기 관찰 자료를 이 용하였다. 거친 모래로 된 꼭대기의 가장 척박한 토양에는 방크스소 나무와 red pi ne 이 우점하였는데, 이런 지역에서는 산불이 80 년 주기 로 발생하여 빈도가 가장 높다는 것을 알았다. 입자의 크기가 중간 정도인 모래로 되어 있는 토양에는 생산력이 더 높은 스트로브잣나 무와 참나무림이 형성되어 있었는데, 산불이 170 년 주기로 발생하여 산불의 발생빈도가 낮았댜 가장 생산력이 높은 지소는 스트로브잣 나무와 북방활엽수림(너도밤나무와 당단풍니~ 형성된 지소인데, 이 지역에는 입자가 미세한 빙퇴석에서 유래된 토양이 발달되어 있 었댜 이 지역의 산불 빈도는 더욱 낮아 1400 년 주기로 발생하는 것
으로 추정되었다. 따라서 이 지역 내에서 Wh it ney (1986) 는 산불에 의한 교란율과 생산력 사이에 음의 상관이 있음을 알았으며, 산불의 빈도는 토양이 형성되는 초기 모물질과 관련이 있었다. 이와는 대조적으로 Wh it ne y는 산불과는 관련없이 나무가 쓰러져 서 야기되는 교란은 생산력과 양의 상관이 있음을 알았다. 그 이유 는 키가 큰 나무는 상대적으로 근계가 작아 강한 바람에 더 잘 쓰 러지기 쉽기 때문인 것 같다. 그러나 나무가 쓰러져서 생기는 교란 율은 불에 의한 교란율보다 훨씬 낮았다. 이들 두 가지의 손실원을 합했을 때 손실률은 생산력과 음의 상관이 있었다. 손실률과 생산력 사이의 이러한 음의 상관은 다른 다양한 서식처 에서도 예상되는 일이다. 예를 들어 교란율을 증가시키는 노출된 사 면에는 척박한 토양이 발달하기 쉬운데, 그 이유는 교란과 연관된 침식과 세탈에 의한 영양염류 손실이 크기 때문이다 (L i kens et al. 1977, Bonnann&Lik en s 1979). 지면이 수평일 경우에도 교란율이 높 은 지역에는 척박한 토양이 발달되는데, 그 이유는 교란된 지소에서 영양염류의 세탈과 바람에 의한 침식이 크기 때문이다. 경사가 급한 사면에서 주요 침식원이 되는 산사태와 같은 교란들이 이러한 지역 에 척박한 토양이 형성되도록 한다. 따라서 다른 모든 것이 같다면 교란율이 낮은 지역이 높은 지역에 비해 비옥한 토양이 형성된다. W i lson 과 Kedd y (1986) 는 온타리오의 AxeLake 에 있는 보호된 만 에서 뿌리가 있는 대형 식물이 있는 지역이 손실률이 더 낮고 유기 물이 더 풍부한 기질로 되어 있음을 알았다. 반면에 파도에 완전 노 출된 해변은 손실률이 더 높고 모래가 훨씬 많은 기질로 되어 있었 다. 따라서 교란과 기질의 영양염류 풍부도 사이에는 음의 상관이 있었다. 손실률과 토양의 영양염류 공급률이 음의 상관을 갖는 서식처에 서 토양의 영양염류 공급률 구배에 따른 종(형태)의 분포는(그림 9.3
D-F) 손실률과 토양 자원의 공급률이 서로 독립적인 경우(그림 8.4 C), 혹은 Oksanen 등 (1981) 의 경우(그림 9.3 A-C) 와는 매우 다를 것 이다. 영양염류 공급률이 증가함에 따라 손실률이 감소하는 경우에 척박한 토양이 발달된 지소에는 뿌리로의 분배가 적고, 키가 작고 잎이 많으며, 생장이 빠른 식물들이 우점할 것이다 . 반면에 초식동 물-생산력 구배상에서 가장 척박한 토양을 갖는 지소에는 뿌리로 분배량이 많고, 잎이 적고 생장이 느린 식물들이 우점할 것이다. 자연 군집에서 토양 자원과 손실률에 관한 여러 가지 가능한 괘 적들이 있다. 이미 설명한 바와 같이 (G ri me 1979), 방목되는 초지의 손실률은 영양염류 공급률과 양의 상관을 갖기 쉽다. 이러한 각 괘 적은 생산력에 대한 식물의 형태와 생활사의 의존도 면에서 다른 패턴을 보일 수 있다. 이러한 패턴을 이해하기 위해서는 자연식생의 패턴 연구에서 자원 공급률과 손실률을 같이 측정할 필요가 있으며. 적어도 이 둘 중 어느 하나가 상당히 안정한 지소를 조사 지소로 선택하여야 한다. 4 요약과종합 이 책을 집필하게 된 동기는 전세계적으로 육상식물 군집에서 관 찰되는 유사성을 식물에게 가해지는 보편적인 제약과 이러한 제약 들에 대처하기 위해 식물이 가지고 있는 대응책으로 가장 잘 설명 할 수 있다는 신념에서였다. 내가 가장 관심을 갖은 주요 제약은 식 물이 필요로 하는 지상부와 지하부의 자원, 죽 햇빛과 토양의 영양 염류였다. 이러한 필수 자원들이 물리적으로 분리되어 있기 때문에 이들을 획득하기 위하여 식물은 어쩔 수 없이 대응책을 갖어야 한 다. 하나의 자원을 더 많이 획득하기 위해서는 식물이 그 자원을 획
득하는 데 관여하는 기관의 생장에 더 많은 분배를 하여야 한다. 따 라서 필연적으로 다른 자원을 적게 획득하게 된다. 이 책에서는 이러한 제약과 대응책의 논리적인 중요성을 알아보 고, 그 결과로 예측되는 형태, 생활사, 그리고 군집의 패턴을 결정하 였다. 그 이유는 이러한 제약과 대응책은 자연에서 관찰할 수 있는 것들과 일치하기 때문이다. 이론에 의해 예측되는 패턴과 자연계에 서 볼 수 있는 광범위한 패턴 사이에는 뚜렷한 유사성이 있다. 식물은 생활형, 키, 그리고 습성 면에서 사막의 1 년생 식물과 다 년생 식물로부터 (Gu ti errez & Whit ford 1986). 열대림의 수관층 수 (Vi tou sek 1982), 대평원이나 사바나 혹은 휴경지의 협엽초본과 광엽 초본 (T i lman 1984, 1987a), 토양 조류(개인적인 관찰), 더글러스전나무 (Wa ring et al.1 9 78), 툰드라의 초본과 관목 (Shaver & Chapi n 1980) 에 이르기까지 그들의 자연 서식처에서 자원의 제한울 받는다는 것이 (알T려il m졌 a다n .1 98지2)하. 부이 자책원은의 식 비물율 경에쟁 따의라 이 경러쟁한 우면세에종 이초 점다을르 긴맞 추하지지 만않 고 부족한 토양 자원과 햇빛에 대한 경쟁 효과에 초점을 두었다. 식 물의 필수 자원이 지상부와 지하부로 분리되어 있기 때문에 지하부 자원을 획득하는 데 사용되는 식물의 구조물은 지상부 자원을 획득 하는 데 이용될 수 없고 그 역도 마찬가지이다. 그러나 토양 자원과 햇빛은 영양 면에서 필수적이기 때문에 식물 이 균형 있는 최적 생장을 위해서는 특정한 비율로 이들 자원을 획 득해야 한다. 이 자체만으로도 식물 개체는 자신의 생장을 줄기, 뿌 리. 혹은 잎의 생장에 분배하는 대응책을 갖게 된다. 각 분배 패턴, 그리고 그 결과 비롯되는 형태가 식물이 획득할 수 있는 토양 자원 과 햇빛의 상대적인 양을 결정한다. 식물의 생리적인 특성과 함께 이것은 각 종으로 하여금 부족한 토양 자원과 광 이용도의 좁은 범 위에서만 경쟁 우세종이 되도록 제한한다.
각 식물종은 특정한 자원 수준에서만 경쟁 우세종이 되도록 제한 되기 때문에 자원의 수준을 결정하는 힘은 식생의 패턴을 결정하는 데 매우 중요하다 . 한 서식처에서 토양 자원과 햇빛의 평균 이용도 룰 결정하는 두 가지 주요인은 그 서식처의 토양 자원 공급률과 식 물이 그 서식처에서 경험하는 손실률 혹은 교란율이다. 이들 각각은 상이한 자원 이용도의 패턴을 유도한다. 손실률을 일정하게 유지하면 부족한 토양 자원의 공급률이 낮아 식물의 생물량이 적고 햇빛의 이입량이 많은 서식처로부터 토양 자 원의 공급률이 높아 식물의 생물량이 많고 입사되는 광량이 적은 서식처에 이르는 생산력 구배상의 위치에 따라 서식처가 구분된다. 이러한 생산력 구배상의 점의 위치에 따라 식물의 형태, 생리, 생활 사 둥이 다를 것으로 예측된다(그림 4.1 4 ). 정확한 패턴은 손실률에 따라 다르지만, 영양염류의 공급률이 낮은 토양에 우점하는 식물은 영양염류 공급률이 높은 토양에 우점하는 종들에 비해 뿌리로의 분 배량이 많고 줄기로의 분배량은 적어야 한다 . 이러한 예측과 부합되 는 패턴들이 전세계적으로 다양한 기후 지역, 고도, 그리고 토양 구 배에서 보고되었고(그림 5.2 - 5.5 ) , 각 삼림 내에서 볼 수 있는 소규 모의 생산력 구배에서도 보고되고 있다. 토양의 영양염류 공급률을 일정하게 유지한다면 손실률이 낮은 지역은 식물의 생물량이 가장 높고 손실률이 높은 지역은 생물량이 가장 낮을 것이다 . 식물 생물량은 영양염류 홉수율과 식물에 의해 차단되는 광량을 결정한다. 손실률이 낮은 지역에서부터 손실률이 높은 지역으로 이어지는 구배를 따라 토양 영양염류와 지표면에서 의 햇빛의 이용도는 증가하고 수관총의 높이는 낮아질 것이다. 손실 률이 높을 때 토양 영양염류와 햇빛의 이용도가 높은 것은 토양 자 원 공급률이 일정하게 유지될 때 잎으로 더 많은 분배를 하고 뿌리 와 줄기로 분배가 적은 식물이 유리하도록 한다. 이러한 식물은 최
대 영양 생장률 (RGR) 이 높은데. 그 이유는 잎에 분배하는 양이 많 기 때문이댜 따라서 부족한 토양 자원과 햇빛의 이용도는 손실률 구배상에서는 양의 상관을 갖어야 하고. 생산력 구배상에서는 음의 상관을 갖어야 한다. 실제 서식처는 그것의 손실률과 토양 자원의 공급률에 의해 특징지어지며. 이들 두 가지 모두는 식물이 그 서식 처에서 부족한 토양 자원과 햇빛을 감소시키는 장기간의 평균 수준 을 결정하는 중요한 요인들이다. 식물이 가지고 있는 많은 상이한 특성들이 영양염류의 공급률과 서식처의 손실률에 의존적인 것으로 예측된다 . 성숙하였을 때의 식 물의 키, 죽 줄기에 분배되는 양은 생산력 구배를 따라 증가하여야 하며. 손실률 구배를 따라 감소되어야 한다. 교란되지 않고 영양염 류가 풍부한 서식처에서 식물의 키는 최대가 되어야 하고, 교란이 심하고 영양염류가 빈약한 서식처에서 최소가 되어야 한다 . 종자의 크기는 생산력 구배를 따라 중가하여야 하고. 손실률 구배를 따라 감소하여야 한다. 더구나 생물 군계의 광범위한 지리적 차이를 유도 하는 것과 똑같은 힘이 소규모의 토양 구배상에서도 종의 분리를 유도하여야 하며, 단일 개체군 내의 개체들 사이에서 비슷한 유전적 인 변이체를 선발하여야 한다. 따라서 식물 개체 내의 가소성의 패 턴, 주어진 종내의 개체들 중 유전 가능한 변이체의 패턴, 서식처 내 에서 종 사이의 차이. 그리고 서식처에 따른 종의 분리가 모두 같은 제약, 같은 형태적 그리고 생리적 대응책, 그리고 같은 자원 이용도 패턴에 의해 영향을 받는다. 서식처에서 특정한 식물을 지속시키게 하는 성질은 그 서식처에 있는 식물의 장기간의 평균 수도뿐만 아니라 군집 동태에도 큰 영 향을 준다. 손실률이 높은 서식처에서 경쟁에 우세한 식물은 최대 영양 생장률이 높아야 한다. 그 이유는 잎으로 분배되는 양이 많기 때문이다. 이와 같이 높은 최대 생장률은 이들 종이 교란 후나 실험
적 조작 후에 서식처에서 맨 먼저 우점종이 될 수 있음을 의미한다. 그러나 이들 종은 그 서식처의 영양염류 공급률과 손실률에 의해 결정된 장기간의 평균 자원 이용도에서 경쟁력이 더 좋은 일련의 종들에 의해 경쟁적으로 교체되어야 한다. 특히 영양염류가 풍부한 토양에서는 천이 동태의 대부분이 줄기로의 분배와 최대 생장률 사 이의 역상관 관계에 의해 조절되는 일시적인 효과로 설명될 수 있 다(그림 4.1 0 ). 1 차 천이와 같이 극도로 척박한 토양에서 진행되는 천이는 비옥 한 토양에서 진행되는 천이와 일부 정성적인 면은 같지만 잎이 많 은 식물의 일시적인 우점보다는 토양의 영양염류 공급률의 점진적 인 변화에 의해 그 과정이 결정될 수 있다. 비옥한 토양에서 진행되 는 천이의 초가 우점종은 뿌리에 분배되는 양은 적고 잎에 분배되 는 양은 많으며, 줄기에 분배되는 양은 중간 정도인 종이어야 한다 . 이와는 대조적으로 매우 척박한 토양에서 진행되는 1 차 천이 혹은 2 차 천이 과정에서 초기의 우점종은 뿌리(혹은 질소 고정과 같은 뿌 리의 기능)에 분배량이 많고 줄기에 분배되는 양이 적어야 하며, 잎 에 분배되는 양은 중간 정도인 종이어야 한다. 형태적인 제약과 이에 대한 대응책이 있는 것과 마찬가지로 생리 적인 제약과 그것에 대한 대응책이 있다. 예를 들어 Cha pin (1980) 은 최대 광합성률이 잎 단백질 함량에 따라 증가한다고 보고하였다. 그 러나 광합성 기관에 분배된 단백질은 뿌리의 홉수능을 증가시키고 초식을 감소시키는 기능이 있는 2 차 대사산물의 농도를 중가시키는 데 사용할 수 없다. 제 4 장에서 설명한 바와 같이 모든 종이 같은 생 리적 특성을 갖는다고 가정하면 각각의 독특한 서식처(즉 손실률, 교 란율, 영양염류 공급률의 독특한 조합을 갖는 서식처)는 특정한 식물 형 태에게 유리하게 작용하여야 한다. 식물의 생리적 특성이 동일하지 않고 다양한 생화학적 기능에 분
배되는 패턴으로 결정되는 식물생리의 범위가 있는 것으로 가정하 면 생리와 형태 사이에 일치하는 점이 있어야 한다. 척박하고 교란 이 되지 않은 서식처는 뿌리의 흡수가 효율적이고 뿌리 생물량이 많은 식물에게 유리할 것이다. 비옥하고 교란이 되지 않은 서식처는 뿌리의 홉수효율이 낮거나 뿌리의 생물량이 적지만 광합성 효율이 높고 줄기와 잎의 생물량이 많은 종에게 유리할 것이다. 주요 생리 적인 과정들 사이 그리고 형태적인 특징과 생리적인 과정들 사이의 대응책과 이들에 대한 제약을 밝히기 위해서는 더 많은 연구가 필 요하댜 형태적 그리고 생리적인 가소성이 생산력과 손설률 구배에서 식 물의 분포에 영향을 주지만 이러한 가소성의 효과는 생리적으로 상 이한 경쟁자. 줄기를 구성하는 물질의 특성에 의해 나타나는 구조적 인 한계. 그리고 각 개체의 한정된 생화학적 저장 물질에 의한 생리 적인 한계에 의해 제한될 것이다. 토양 자원과 햇빛의 최적 획득은 식물 개체들 내에서 형태적 그 리고 생리적 가소성을 갖는 개체들에게 유리하게 작용하겠지만, 생 산력 혹은 손실률 구배상에서 종의 분포 패턴에 정성적인 효과를 갖지는 않을 것으로 보인다. 따라서 육상의 관속식물들에게 공통으 로 필요한 지상부와 지하부 자원은 각 종이 이들 자원 이용도의 좁 은 범위에서만 경쟁적 우세종이 되도록 제한한다. 식물의 생리. 형태. 생활사와 그 식물이 우점하는 서식처 사이에 는 강한 일치성이 있어야 한다. 식물의 크기 구조와 이에 상응하는 광의 수직 구배는 추정치에 불과하지만 자원 의존적인 생장 둥구배 곡선은 특정한 서식처에서 식물의 경쟁 능력에 대한 이러한 모든 면의 효과를 종합한 것이라고 볼 수 있다. 이 책을 집필하게 된 목적은 식물 경쟁의 주요 기작과 식물 형태 에 기본적인 제약들이 자연계에서 우리가 볼 수 있는 패턴을 얼마
나 잘 설명할 수 있는지를 알아보기 위해 논리적인 관계를 파악하 기 위한 것이었다. 이 책에서 제시된 이론에 의한 예측과 자연 상태 의 패턴 사이에는 대체적으로 일치하는 점이 있었다. 이 이론을 좀 더 실제적이 되도록 만들 수 있지만 그러면 이론이 더 복잡해진다. 초식을 감소시키는 화합물이나 구조에 대한 분배를 포함하도록 하 거나, 경쟁에서 이웃하고 있는 식물에 의한 영향을 명백히 포함하도 록, 혹은 식물체로 하여금 형태적 그리고 생리적으로 가변성을 갖도 록 이 이론을 변형시키면 이 모든 것들이 식물의 분포 패턴에 관한 중요한 지식을 제공할 수 있다. 따라서 앞으로 해결해야 할 일들이 많이 남아 있다. 생태학자들이 잘 밝혀낼 수 없을 정도로 자연계는 그 구조와 기 능이 매우 복잡하지만 우리는 자연계에서 관찰할 수 있는 패턴을 설명할 수 있는 방법을 찾아내야 한다. 이 책은 식물 상호 작용의 몇 가지 주요 기작을 이용하여 식물 군집의 구조와 동태에서 우리 가 볼 수 있는 대부분의 패턴을 설명할 수 있다고 제시하고 있다. 신중하면서도 장기적인 실험과 관찰을 통해서만 여기에서 제시된 아이디어들이 실제적으로 얼마나 유용한지를 결정할 수 있다. 그러 나 이 책과 T i lman(1982) 의 논문에서 제시된 관찰과 실험적인 연구 를 통해 자연 상태 또는 관리되고 있는 개체군에서 종의 분포와 수 도를 결정하는 데 있어서 소비자와 자원 사이의 상호 작용이 중요 한 요인이란 사실을 분명히 알 수 있다. 소비자와 자원 사이의 상호 작용의 기작을 좀더 깊이 있게 연구하게 되면 오늘날 지구상에서 생활하고 있는 종의 진화를 유도한 많은 힘(기작), 그리고 이들 종 을 존속시키는 힘들을 이해할 수 있고, 또한 자연계에서 패턴을 유 발시키는 힘도 이해할 수 있게 된다.
부록: 모델 ALLOCATE 를 이해하기 위한 수학적 기초 1 서론 크기 구조를 갖는 식물 사이의 경쟁 모델에서 언급하고 있는 많은 가 정들을 제 3 장에서 설명한 바 있다 . 여기에서 나는 모의 모델인 ALLOCATE 에 내재하고 있는 모델의 수학적 형태를 제시하려고 한다. 입출력, 그리고 시스템 운용에 관한 여러 가지 프로그램 때문에 5 쪽 이 상이나 되는 전체적인 FORTRAN 기호를 제시하는 대신에 나는 이 모 델의 주요한 수학적 요인들을 제시하고 그것들이 갖는 의미를 설명하려 고 한댜 나는 FORTRAN 모델에서 일반적으로 사용하는 부호와, 예를 들면 * 는 곱하기, 여러 개의 글자를 갖는 독립변수를 이용하려고 한다. 예를 들어 BR 은 〈 뿌리의 생물량 (b i omass o f roo t s) 〉을 의미한다. 이들 방정식을 연속 미분방정식으로 나타낼 수도 있지만 모델을 설명하는 데 수학을 너무 자세히 하면 모델의 동태에 영향을 줄 수 있고, 특히 여러 단계의 모델일 경우에는 더욱 그렇다. 따라서 나는 모델을 반복된 형태 (it era t ed fo rm) 로 제시하고 싶다. 각 식물 개체는 연속적인 방법으로 영양 생장을 하며 , 그 식물의 생 장률은 그것이 처해 있는 서식처의 영양염류와 햇빛에 의해 결정되는 것으로 가정한다 . 주어진 시간에 발아하는 종 혹은 유전자형의 모든 식 물체는 동일한 코호트 (coho rt)로 취급된다. 모든 식물은 소비를 통해 자 원이용도에 영향을 주며, 따라서 생장은 자원 의존적이다. 종 I 를 생각 해 보자. 종 I 는 별개의 코호트로 구성되며 , 각 코호트는 특정한 시간에
발아한댜 종 I 의 J번째 코호트는 다음과 같은 속성을 갖는다. BR (I,J)는 /종의 J코호트 개체당 뿌리의 생물량 . BL (I,J)는/종의 J코호트 개체당 잎의 생물량. BS (I , J)는 /종의 J코호트 개체당 줄기의 생물량. AL( I), AR( I), 그리고 AS (I)는 /종의 모든 코호트에서 각각 잎, 뿌리, 줄기로의 분배율 QN L( I), QN R( I), QN S( I), 그리고 Q NSD (I)은 /종의 잎 뿌리, 줄기, 종자의 단위 생물량당 영양염류의 함량. R (I)는 /종의 단위 잎량당 최대 광합성률. RR(I) , RS(I) , 그리고 RL (I)는 /종의 뿌리, 줄기, 잎의 단위 생물량 당호홉률. KN (I)와 KL (I)는 1 종에서 각각 영양염류와 광 제한 광합성의 반포화 A0l -기~-. N (I,J)는 J코호트에 있는 1 종의 개체수. D (I,J)는 1 종에 있는 J코호트의 개체의 사망률. NSEED (I)는[종의 종자수. BSEED (I)는 1 종의 종자(종자 크기)당 생물량 . H 먀 (I, J)는 [종의 J코호트에 속하는 모든 개체의 키(높이). L(X) 는 지표면으로부터 X 만큼 떨어져 있는 곳의 광량(전광량에 대 한 비율). S 는 부족한 토양 자원의 이용도. TN 은 모든 토양 영양염류의 총량이며, 영양염류 공급률을 결정하는 주요 인자이다.
2 풀이 고陸 모든 식물은 종자로부터 생 활 을 시작한다. 발아를 하면 종자의 생물 량 (BSEED) 은 다음 방정식에 따라 뿌리. 잎 그리고 줄기로 분배된다. 새 로운코호트는J이다 . BR( I, J) = AR( I)* B SEED(J) BL(I,J) = AL(I) *B SEED(J) BS( I, J) =AS( I)* B SEED(J) 새로 발아한 이 식물의 높이와 모든 식물의 높이는 다음의 관계식에 따라 개체당 줄기의 생물량 BS( I, J)에 의해 결정된다. HITE( I, J) =4* [2 500* B S( I, J )] 0-5 이 식물이 받는 광량은 이 개체의 키보다 더 높이 있는 전체 잎량에 의해 결정된다. 이것은 모든 잎이 줄기 위쪽에서 한 층으로 존재한다는 가정을 기초로 한 것이다. 식물체의 높이가 X 이고 . 그 위에 있는 전체 잎량을 T(X) 라 하자. 그러면 높이 X 인 식물체에게 입사되는 광량은 T(X) 의 음의 지수함수로 가정된다 . L(X) =exp ( 一 0.0045*T(X)) 부족한 토양 자원의 이용도는 S 이다. 따라서 S 는 토양에 있는 부족한 영양염류의 측정 가능한 가용성 형태의 농도이다. 식물의 광합성률에 대한 주어진 S 의 영향은 그 식물체의 잎량에 대한 뿌리량의 비에 의해 결정되는 것으로 가정한다. 따라서 C (I,J)를 토양 자원의 효율적인 이용 도로 정의하면 C( I, J) = S*BR( I, J )/BL (I, J)
1 종에 속하는 J코호트의 한 개체에 의해 DT 기간 동안에 생산된 광 합성 산물의 양을 DB( I, J)라고 하자. DT 는 모델을 해결하는 과정의 시 간 단계이다. 그러면, DB(I,J) =MIN[(DT* R { I)*B L( I,J) *C { I)) /(C( I,J) + KN( I))) 혹은 (DT*R{ I)*B L( I, J)* L(HITE( I, J)) /(KL(I , J) + L(HITE(I,J )))]-R ESP 최소 함수 (MIN) 의 존재는 개체당 새로운 광합성 산물의 생산율이 영양염류나 혹은 햇빛에 의해 결정됨을 의미한다. 어느 것이든지 낮은 비율을 유도한다. 1 종의 J코호트 각 개체에 의해 생산된 생물량은 먼저 식물의 호흡에 사용된다 . 이것은 현재 생산량에서 호홉 요구량을 빼면 된다. 호흡 요구량 RESP 는 다음과 같이 결정된다. RESP=DT*[RR(J) * BR( I,J) + RS(J) * BS( I,J) + RL(J) * B L( I,J)] 만약 DB(I, J )>O 이라면 개체는 시간 간격 내에 순생산울 갖게 되며, 이 개체가 자신의 최대 높이에 도달하지 않았으면 (sub-adu lt), 이 생산 량은 뿌리, 잎, 줄기로 분배되고, 최대 높이에 도달했다면 종자로 분배 된다. 만약 DB (I,J )
모델의 특별한 공식을 제외하고 종자는 사망하지 않는다. 시간 간격 의 마지막까지 살아남은 코호트 내의 식물 개체의 수(따라서 다음 시간 간격의 시작점에서 개체의 수)는 다음과 같이 결정된다. SURVIVORS( I,J) =N( I,J)* ( l-D T* D ( I,J) 이것은 다음 시간 간격의 출발점에서 이 코호트의 개체수가 된다. 이러한 영양염류 의존적인 생장. 분배, 종자 생산 그리고 생존은 주어 진 시간 간격 동안에 모든 종의 모든 코호트에 있는 모든 개체에서 반 복된댜 이러한 모든 개체는 이 시간 간격 동안에 토양 영양염류 이용 도가 똑같고 동일한 수직적 광 구배를 경험하는 것으로 간주된다. 모든 종의 모든 코호트가 일단 주어진 시간 간격 동안에 생장, 생식, 사망이 허용된다면 자원 이용도에 대한 이들 생장의 전반적인 효과는 결정된 다. 햇빛의 이용도는 위에서와 같이, 즉 모든 개체에 대하여 새로운 높 이와 개체당 새로운 잎량을 사용하여 결정된다. 영양염류 이용도 S 는 두 가지 과정, 죽 영양염류 흡수와 영양염류 공급의 결과로 결정된다. 새로운 생물량과 종자의 생산에 필요한 영양염류에 균형을 맞출 수 있 을 만큼 영양염류 흡수는 충분한 것으로 간주된다. 따라서 /종과 J코호 트의 각 개체에 의해 흡수되는 영양염류는 그것이 성체인지 아니면 유 식물인지에 의해 달라진다 . 그것이 유식물일 경우(더 많은 잎과 뿌리 그 리고 줄기를 생산) 개체당 흡수는 UPTAKE(!, J) = DB (I, J)* [ AR(J) * Q N R(J )+ AL(J) *Q N L( J) +AS (J)*Q N S (J) ] 가 될 것이다. 그것이 성체라면 개체당 흡수는 종자의 영양염류 함량에 의해 결정될 것이다. UPTAKE(!, J) = DB (I, J)*QN SD(J) .
/종과 J코호트의 각 개체당 각각의 흡수율을 시간 간격의 마지막에 살아남은 개체수로 곱하고, 그런 다음 모든 종의 모든 코호트의 값을 합한다. 이것이 군집 전체의 흡수율이댜 영양염류의 공급률은 SUPPLY = DT*A* (TN -B N -S ) 로 계산된다. BN 은 식물체 속에 들어 있는 전체 영양염류의 양이다. 따 라서 영양염류의 공급은 토양에 남아 있는 전체 영양염류의 비율에 비 례한다. 길이 DT 인 시간 간격 동안 전체 흡수량과 전체 공급량이 주어 지면 영양염류의 새로운 이용도는 이전의 이용도 (S) 에 SUPPLY 를 더 하고 전체 흡수량을 뺀 것과 같다. 이 과정이 한 번 완결된 다음에 새로운 출발 조건으로 사용되는 모든 변수에 새로운 값을 주어 이 과정을 반복한다. 이것은 크기 구조를 갖 는 식물들 사이의 생장과 경쟁의 모델이다. 식물의 생장률은 영양염류 와 햇빛의 이용도에 의해 결정된다. 식물은 생장을 통해 수직적인 광구 배를 형성한다. 식물의 생장과 영양염류 소비 과정은 반복적인 방법으 로 풀 수 있다. 죽 모든 종의 모든 코호트에서 현재의 자원 수준에 대한 반응으로 식물이 생장하면 다음 반복을 위한 식물의 크기와 개체수가 결정된다. 이것이 종결된 후 영양염류 이용도와 수직적인 광 구배에 대 한 이 생장의 효과가 결정된다. 그 다음 이러한 새 값들이 다음 반복을 위한 생장 패턴을 결정한다. 종자는 생산된 후 즉시 발아하는 것으로 허용되었기 때문에 식물의 새로운 코호트는 매일 생산될 수 있다. 따라서 코호트의 수가 너무 많 아져서 Cray 2 를 가지고도 이 모델을 수학적으로 해결할 수 없게 된다. 이 문제를 극복하기 위하여 나는 비슷한 코호트를 묶어 종당 40 개의 코 호트가 넘지 않도록 하였다. 한 종이 20 개 이상의 상이한 코호트를 갖 으면 매 20 회 반복을 통해 이 코호트들 중에서 그 종의 개체당 생물량 의 가능한 범위의 1/ 40 보다 작은 것들이 어느 것인지를 결정하여 그것
들을합했댜 나는 Cray 2 슈퍼컴퓨터에서 CRAY FORTRAN 을 사용하여 모델울 수학적으로 해결하였다 . 대부분의 모의에서 나는 약 하루(실제로는 0.7 일)의 시간 단계룰 사용하였다 . 무게의 95% 를 잎에 분배하는 식물이 갖 는 최대 생장률 0.2/da y는 여러번 반복하지 않고도 모델 해결이 시간 단계-크기 독립적이라는 것임을 알 수 있었다. 전체 모의를 위해 140 일 간의 기간을 하나의 생장 기간, 즉 1 년으로 간주하였다. 수백 종이 경쟁 할 때 에는 ALLOCATE 를 슈퍼 컴퓨터로 풀어 야 하지 만 2-20 종이 경쟁 할 경우의 ALLOCATE 모델은 IBM-XT 와 같은 마이크로컴퓨터로 풀 수 있다. 마이크로컴퓨터 사용을 위한 ALLOCATE 의 전체 FORTRAN 코드의 복사본은 요청하면 받을 수 있다.
옮긴이 해제 생태학의 흐름은 종래의 기재적인 차원에서 생태계의 구조와 기능을 과학적인 체계하에서 실험을 통해 밝히려는 실험 생태학적인 방향으로 바뀌고 있다. 이번에 역자가 번역한 이 책은 1988 년에 David T il man 이 집필한 개체군 생물학에 관한 많은 연구 논문 중의 하나이다. 이 책에 는 우리가 자연 생태계에서 관찰할 수 있는 식물 군락의 천이 과정을 자원 이용도의 패턴에 의해 설명하려고 하고 있다. 이 책의 저자인 T i lman 이 생태계의 종 다양성에 관한 문제를 영양염류 이용도의 수준 에 따라 모델화한 것은 여러 사람들의 야외 실험의 결과에서도 그 타당 성이 입증되고 있다. 또한 토양이나 수중의 부영양화에 의하여 육상식 물 군집이나 수중의 식물성 플랑크톤의 종 구성이 단순하게 바뀌는 것 도 그의 모델로 설명이 가능하다. 역자는 이 책을 읽기 전에 이 책의 저자가 발표한 몇 편의 논문을 읽 게 되었는데. 미국의 뉴멕시코 주에 있는 JO RNADA Long Term Ecolog ica l Research(LTER) S it e 에서 질소를 첨가한 tr ansec t와 첨가하지 않은 tr ansec t에서 초본식물의 종 다양성의 변화를 조사하는 동안 그의 이론과 접하게 되었다. 역자가 J ORNADALTER 에서 조사한 결과도 그 의 이론과 일치하는 점이 많았다. 1988 년 귀국한 후에 이 책을 보게 되 었는데, 많은 부분은 종래의 자기 주장을 다듬고 보완하였으며 그의 자 원 이용 패턴에 관한 모델을 생태적인 현상에 광범위하게 적용하려는 의도를 엿볼 수 있었다. 이 책에서 T i lman 이 주장하는 이론들은 그 나름대로 독창성이 있는
것이 많이 있지만 그가 이러한 이론을 제시할 수 있었던 것은 이전부터 많은 사람들이 실험을 통해 발표한 귀중한 자료들이 뒷받침되고 있다. 자원 이용 패턴을 근간으로 한 그의 천이 모델은 종래의 편의 모델이나 내성 모델 또는 촉진 모델과는 상당히 다른 차원이라고 할 수 있다. 다 른 사람들이 실험한 내용들을 분석하고 자신의 결과와 비교하며, 이것 을 토대로 새로운 이론을 창출해 내는 그의 학문 스타일은 우리 나라의 젊은 생태학도들에게 많은 자극이 될 것으로 생각된다. 1982 년에 T il man 은 Cedar Creek Natu r al Histo r y Area 에 있는 휴경지 의 식생 구성과 종 다양성 그리고 식생의 동태를 조절하는 요인을 규명 하기 위한 일련의 관찰과 실험 연구를 시작하였다. T i lman 은 이 연구의 결과를 이용하여 이 책에서 자신이 제시하고 있는 많은 아이디어들을 평가하려고 시도하였다. 그는 이 지역에 형성되어 있는 휴경 연도가 다 른 다양한 휴경지에서 식생의 형성 과정을 오랫동안 관찰하고 시간에 따른 종 구성의 변화나 다양성의 변화가 일어나는 원인을 밝히기 위한 실험을 하였댜 T i lman 이 Cedar Creek 의 휴경지에서 토양과 식생을 조사한 바에 의 하면 이 지역에서의 천이 패턴이 시간뿐만이 아니라 질소와 광에 대한 식물의 경쟁에도 많은 영향을 받는다는 것을 알게 되었다. 그는 질소 공급량의 증가는 식물의 지상부 생산량을 증가시켜 결국 지표면에 도달 하는 광량의 저하를 초래하게 되며, 이러한 환경 제약에 대응하기 위해 식물의 종에 따라 특이한 자원 분배 패턴을 갖는다고 주장하고 있다. 죽 지표면의 광량이 풍부한 지역에서는 토양의 질소 함량이 제한 요인 이 되어 식물은 광합성 산물을 지하부에 많이 분배하게 된다. 지상부 생물량이 증가하여 광이 제한 요인이 되면 줄기나 잎에 광합성 산물을 많이 분배하여 수광량을 많게 한다는 것이다. Cedar Creek 에 있는 22 개 의 휴경지 에서 그가 조사한 바에 의 하면 휴 경지의 나이와 토양의 질소가 모두 천이 패턴과 중요한 관련이 있음을
알 수 있다. 죽, 시간은 식물의 분산율과 정착률의 차이에 영향을 주어 특정한 휴경지에 존재하는 종을 결정하는 데 중요한 역할을 하는 반면 에 자원을 기초로 한 상호 작용은 휴경지 내의 특정한 지소에 존재하는 종을 결정한댜 따라서 T i lman 의 연구 결과는 미네소타 모래평원에서의 2 차 천이의 패턴이 종의 분산과 기타 시간 의존적인 과정들. 그리고 주 요 토양 자원, 특히 질소 이용도의 변화에서 비롯된다는 것을 시사하고 있다. 천이에 영향을 주는 여러 가지 과정 중에서 어느 하나만으로는 2 차 천이의 패턴을 충분히 설명할 수는 없다. Cedar Creek 에서 2 차 천이의 한 요인은 시간에 따른 질소 공급률의 증가이다. 질소 공급이 증가함에 따라 가장 오래된 휴경지에서는 전체 생물량은 증가하고 지표면에 입사 하는 광량은 감소하며. 토양의 이용 가능한 질소량은 증가한다 . Cedar Creek 에서 질소와 광이 가장 중요한 제한 자원이라고 가정하면 이 책에 서 제시된 이론을 통해 다음과 같은 예측이 가능하다. 죽 질소 공급률 의 증가는 군집 전체의 식물량 증가와 지표면에 이입되는 광량의 감소 를 초래하고 피움에 의해 광량이 감소되면 군집 전체의 식물량이 감소 되고 이용 가능한 토양의 질소량은 증가된다. 질소나 광 이용도의 변화 에 따라 이들 자원의 이용 효율이 높은 종과 낮은 종 사이에 상대수도 의 변화가 생기기 때문에 시간에 따른 우점종의 변화가 나타나게 된다. 그는 이러한 예측을 7 가지로 질소 구배를 달리하는 실험을 통해 확인하 고 있는데, 제 7 장에 그의 실험 내용들이 자세히 설명되어 있다. 이 책은 모두 9 장으로 구성되어 있다. 제 1 장에서는 생태학의 가장 중 요한 목표가 자연에서 관찰할 수 있는 특정한 패턴을 이해하는 것임을 강조하고 이러한 패턴은 식물이 받는 환경 제약과 그 환경 제약을 해소 하기 위해 식물이 갖는 대응책에서 비롯됨을 지적하고 있다. 그는 육상 식물이 직면하고 있는 여러 가지 제약과 대응책에 대하여 간략하게 설 명하고 있다. 제 2 장에서는 부족한 자원에 대한 식물의 경쟁을 이해하기
위해 단순한 평형 상태의 접근 방법을 제시하고 있다 . 식물이 필요로 하는 많은 자원들은 영양 면에서 필수적이며. 필수 자원에 대한 최적 획득의 그래프 이론은 그러한 자원이 개체의 생장을 균등하게 제한하는 비율로 소비되어야 한다는 것을 예측하고 있다. 식물은 비제한적 필수 자원의 소비를 줄이는 쪽으로 자신의 형태를 조절함으로써 자원의 획득 노력을 조절할 수 있고. 그 결과 제한되는 자원의 소비를 증가시킬 수 있기 때문에 식물의 자원 의존적 생장 등구배곡선은 직각인 코너가 아 니라 둥근 코너를 갖어야 한다는 것이다. 제 3 장에서는 부족한 영양염류와 광을 획득하기 위한 식물의 경쟁 기 작을 설명하기 위해 크기 구조를 갖는 식물 사이의 생장과 경쟁의 모델 인 ALLOCATE 를 제시하였고. 이 모델의 몇 가지 특징을 설명하고 있 다. ALLOCATE 모델은 영양염류와 광 의존적인 생장 등구배곡선을 이 용하여 정성적으로 예측될 수 있으며, 이러한 등구배곡선은 각 종의 단 독 재배를 통한 모델에서 얻을 수 있음울 설명하고 있다. 제 4 장에서는 식물이 자신의 광합성 산물을 뿌리. 잎. 그리고 줄기로 분배하는 패턴을 달리함으로써 최대 생장률과 경쟁 능력에 크게 영향을 줄 수 있다는 것 을 설명하고 있댜 환경이 제한되지 않는 서식처에서 광합성 산물 모두 룰 여분의 광합성 조직의 생산에 분배하는 개체에서 영양 생장의 최대 생장률을 볼 수 있으며, 식물의 최대 생장률은 비광합성 기관에 분배되 는 생산량에 비례하여 감소한다. 생산량의 일부를 뿌리와 줄기에 분배 하는 식물은 환경이 제한되지 않는 서식처에서는 생장률이 저하되지만 자원이 부족한 환경에서는 오히려 유리하다 . 척박하지만 광이 풍부한 서식처로부터 비옥하지만 광이 부족한 서식처로 이어지는 구배상에서 줄기로의 분배량은 증가하고 뿌리로의 분배량은 감소하며, 잎으로의 분 배량은 영양염류의 공급률이 중간 정도에서 최대가 될 것으로 예상된 다. 식물의 생리적 특성도 생산력 구배에 따라 달라져야 하는데 척박한 토양에서는 광합성의 광포화율이 감소될지라도 조직의 영양염류 농도
가 낮은 식물이 유리하며` 비옥한 토양에서는 광합성의 광포화율이 높 고 광보상점이 낮은 개체가 유리할 것이다. 식물의 형태는 그 식물이 우세한 경쟁자가 되는 지소의 토양 자원 수준과 지표면의 광량을 결정 하기 때문에 이러한 상이한 자원 구배는 천이 과정에서 종 교체의 양상 올 변화시킨다 . 제 5 장에서는 식물의 생활사와 형태적 특징의 많은 것들이 토양 자 원:광의 구배 혹은 손실률 구배상에서 그 종이 최대 수도를 얻는 위치 에 따라 다르다는 것을 설명하고 있다 각각의 식물 개체는 형태적으로 그리고 생리적으로 가소성을 갖는데, 이러한 가소성은 그러한 구배상예 서 종내 그리고 종간의 유전적인 변이에서 볼 수 있는 것과 같은 비슷 한 변이 패턴을 갖기 때문에 키, 뿌리나 잎 그리고 줄기로의 분배 패턴, 종자의 크기, 생장률 , 광합성 효율 등과 같은 다양한 식물의 특성을 결 정하는 것은 토양 자원 공급률과 손실률이 주축이 되는 것으로 설명하 고 있다. 제 6 장에서는 식물 경쟁의 동태에 관하여 설명하고 있다. 이전까지는 주로 식물 경쟁의 장기적인 결과, 죽 경쟁적 상호 관계에 있는 여러 종 들이 평형 상태에 도달하였을 때의 결과에 대해서만 설명하였다. 모든 생태 실험은 필연적으로 자연을 교란시키게 되며, 따라서 생태학자들이 관찰할 수 있는 것은 대부분 교란 후에 특정한 종이 나타내는 단기간의 매우 동적인 반응들이다. 이들 종들이 과거에 거쳐온 진화의 과정에 중 요하게 작용하였던 요인들을 알아내기 위해서는 이러한 단기간 동안에 관찰되는 동적인 반응들을 이용하여 추론하여야 하는데, 실험적인 조작 에 대한 즉각적인 반응은 장기간의 효과와는 무관한 일시적인 동태일 수 있기 때문에 이러한 추론은 상당히 위험하다. 따라서 최근에 교란된 군집에서 볼 수 있는 패턴과 인간의 간섭이 전혀 없었던 자연 군집에서 볼 수 있는 패턴 사이의 유사성과 차이점에 대하여 설명하고 있다. 또 한 어떤 종이 특정한 서식처에서 다른 종을 경쟁적으로 교체할 때 그
종을 경쟁 우세종이라고 지칭하고 있다. 제 7 장에서는 육상식물 군집의 1 차 천이와 2 차 천이에 대한 것으로 정착, 경쟁, 일시적인 동태, 초식, 그리고 그 밖의 다른 과정들이 천이 과정의 동태에 영향을 줄 수 있으며, 이러한 과정들의 상대적인 중요성 은 서식처에 따라 다를 수 있음을 언급하고 있다 . 모든 천이 과정에 몇 가지 공통적인 특징으로 최초의 우점종은 키가 작은 초본식물이며, 이 들은 점점 키가 큰 목본성 식물에 의해 연속적으로 교체된다. 그러나 7 장에서 제시된 이론에서는 천이 과정 중에 우점하는 실제 종 계열이 1 차 천이, 척박한 토양에서 진행되는 2 차 천이, 그리고 비옥한 토양에서 진행되는 2 차 천이 사이에 차이가 있어야 하는 것으로 예측하고 있다. 그는 그 이유를 각각의 천이 과정이 영양염류와 광의 이용도 면에서 차 이가 있고, 각 종의 형태와 생리가 특정한 영양염류와 광 이용도를 갖 는 서식처에서 경쟁력이 높도록 제한하기 때문이라고 설명하고 있다 . 제 8 장에서는 그가 실제로 미네소타의 Cedar Creek Natu r al Histo r y Area 의 휴경지에서 실시한 실험 과정과 결과를 설명한 것으로 휴경지 에서의 2 차 천이 과정은 질소와 광에 대한 종의 경쟁 능력, 질소가 빈 약한 토양에 질소의 점진적인 축적률, 종의 최대 생장률에 영향을 주는 분배 패턴, 정착률 둥에 의해 결정되는 것으로 설명하고 있다. 제 9 장에 서는 앞에서 자신이 제시한 여러 가지 이론적인 주장이 가지고 있는 나 름대로의 문제점들을 열거하고 그에 대한 자신의 의견을 피력하였으며, 이 책을 집필하게 된 목적도 간략하게 언급하고 있다. 마지막 부록에서 는 자신이 3 장에서 설명한 ALLOCATE 모델의 수학적인 성질을 소개 하는 것으로 책을 마무리하고 있다. 이 책은 생태학이 궁극적으로 추구하고 있는 자연에 내재하고 있는 질서를 밝히는 것을 목적으로 하고 있으며, 식물들 사이의 상호 작용의 몇 가지 주요 기작을 이용하여 식물 군집의 구조와 동태에서 볼 수 있 는 많은 패턴을 설명할 수 있다고 주장하고 있다. 그러나 본문에서 저
자가 지적한 바와 같이 많은 부분에서 Gr i me 의 생각과 다른 점을 강조 하고 있댜 이것은 생태 현상에 대한 이해가 사람에 따라 다를 수 있음 울 보여주는 것이다 . 생물과 생물 그리고 생물과 환경과의 상호 작용을 통한 군집 발달 과정이 어느 한 가지 요안에 의해 주도되지 않고 수많 은 요인들의 상호 작용에 의하여 진행되기 때문이다. 그러나 이 책에서 주장하고 있는 내용들은 현대 생태학의 새로운 연구 방향을 제시하는 것 이기 때문에 국내의 생태학도들에게 많은 도움이 될 것으로 판단된다. 1998 년 1 월 문형태
참고문헌 Aarsen, L. W., and R. Turkin g ton . 1985. Wi thi n -spe cie s div e rsit y in natu r al po pu lati on s of Holcus lanatu s , Loli um pe renne and Trifo li u m rep e ns from fou r dif fere nt- a g e d pas tu r es. J o urnal of E colog y 73:869-886. Albrekts o n, A. , A . Aronsson, and C. 0. Tamm. 1977. The eff ec t of fore st fer t iliz a ti on on pri m a ry pro ducti on and nutr ien t cyc lin g in the fore st ecosys t e m . Sil va Fenn. 11:233-239. Armstr o ng , R. A., and R. McGehee. 1980. Comp e ti tive exclusio n . Americ a n Natu rali st 115:151- 1 70. Baker, H. G. 1972. Seed weig h t in relati on to envir on menta l condit ion in Calif orn ia . E colog y 53:9 9 7-1010. Barbour, M., J . Burk, and W. Pit ts. 1980. Terrestr i a l Plant Ecolog y. Menlo Park, CA: Benja m in/C um ming s Publi sh i ng Comp an y. Bazzaz, F. A. 1979. The ph y s io l og ica l ecolog y of pla nt successio n . Annual Revie w of E colog y and Sys te m ati cs 10:351-371. Beals, E. W., and J. B. Cop e . 1964. Veg e ta t i on and soil s i n an easte r n India n a woods. Ecolog y 45:777-792. Beard J. S. 1944. Cl ima x veg e ta t i on in tro p ica l Americ a . Ecolog y 25:127-158. Beard, J. S. 1955. The classif ica ti on of trop ica l Ameri ca n veg e ta t i on type s. Ecolog y 36:89-100.
Beard, J. S. 1983. Ecolog ica l contr o l of the veg e ta t io n of South w este r n Austr al ia : m ois t u r e versus nutr i e n ts . In F. J. Krug e r, D . T. Mi tch ell and J. V. M. Jarv is , eds., M edit er ranean-Typ e Ecosys te m s, 66-73. New York: Sp ring er -Verlag. Bender, E. A., T. J. Case, and M. E. Gi lp i n. 1984. Pertu rbati on exp e rim ents in communit y ecolog y: the ory and pr acti ce . Ecolog y 65:1-13. Bil ling s, W. D. 1938. The stru c tu r e and develop m ent of old field short le af pine sta n ds and certa i n associa t e d ph y s ic a l pr op e rti es of the soil . Ecolog ica l Monog r ap h s 8:437-499. Black, J. N. 1958. Comp e ti tion betw een pla nts of dif fere nt ini t ial seed siz e s in swards of subte r ranean clover(Trifo li um subte r raneum L.) wi th pa rtic u lar refe r ence to leaf area and the lig h t mi cr oclim ate . Austr al i an Jo urnal of A g ri c u ltu r al Research 9:299-318 . Black, J. N. 1960. An assessment of the role of pla nti ng densit y in comp a ris o n betw een red clover(Trif oli u m pr ate n se L.) and lucerne(Medic a g o sati va L.) in the early veg e ta t i ve sta g e . Oi ko s 11:26-42. Bloom, A. J., F . S. Chap in III, and H. A. Mooney . 1985. Resource limitation in pla nts — an economi c analog y. Annual Revie w of Ecolog y and Sys t e m ati cs 16:363- 3 92. Bormann, F. H., and G. E. Like ns. 1979. Patt er n and Process in a Foreste d Ecosys t e m . New York: Sp ring er -Verlag. Box, T. W. 1961 . Relati on shi ps betw een pla nts and soil s o f fou r rang e pla nt commun ities in south Texas. Ecolog y 42:794-810. Brenchley, W., and K. Warin g ton . 1958. The Park Grass Plots at Roth a mste d . Ha rpe ndon, U. K.: Roth a mste d Exp e ri m enta l Sta t i on .
Brix , H. 1983. Eff ec ts of thi n n in g fertiliza ti on on gro wt h of Doug la s fir: relati ve contr i b u ti on of fol ia g e qua nti ty and eff ici e n cy. Canadia n Jo urnal of F orest R esearch 13: 167-175. Brown, J. H., D. W. David s on, J. C. Mung er , and R. S. Inouy e. 1986. Exp e rim enta l communit y ecolog y: the desert gra niv o re sys te m . In J. Di am ond and T. J. Case, eds., Communit y Ecolog y, 41-62. New York: Ha rpe r and Row. Chabot, B. F., and H. A. Mooney (e ds.). 1985. Phys io l og ica l Ecolog y of North Americ a n Plant Communit ies . L ondon: Chapm an and Hall. Chap in , F. S . Ill. 1980 . The mi ne ral nutr i t ion of wi ld pla nts . Annual Revie w of E colog y and Sys te m ati cs 11:233-260. Chapi n, F. S. IlI, and G. R. Shaver. 1985. Arcti c. In B. F. Chabot and H. A. Mooney, eds . , Phy s io l og ica l Ecolog y of North Americ a n Plant Communit ies , 16-40. London: Chapm an and Hall. Chamov, E. L., ~nd W. M. Schaff er. 1973. Life- his t o r y consequ e nces of natu r al seledti on : Cole' s result revis i t ed . Americ a n Natu rali st 107:791-793. Chic h este r , F. W,,; J . 0. Leg g, and G. St an fo r d. 1975. Relati ve mi ne raliz a ti on r~te s of ind ig e nous and recentl y inc orp o rate d N- labelled nit rog e n . Soil Sc ien ce 120:455-460. Ch riste n sen, N. L., and R. K. Peet. 1981 . Seconda ry fore st successio n on the Nort h Caroli na Pie d mont. In D. C. West, H. H. Shug art, and D. B. Botk in, eds., Forest S uccessio n : Concep ts a nd Ap plica tio n s, 230-244. New York: Sp ring e r -Verlag. Claph am, A. R., T. G. Toti n, and E. F. Warburg. 1962. Flora oft he B riti sh Isles, 2nd ed. Cambri dg e: Cambri dg e Un ive rsit y Press. Clatw ort hy , J. N. 1960. Stu d ie s on the natu r e of comp e ti tion betw een
closely relate d spe cie s . Ph. D. Thesis , Univ e rsit y of Oxfo rd . Clements , F. E. 1916. Plant Successio n . Carneg ie Insti tut e Washin g ton Public a ti on 242. 512 pa ge s. Cody, M. L. 1986. Str uc tu r al nic h es in pla nt communit ies . In J. Di am ond and T. Case, eds., Communit y Ecolog y, 381-405. New York: Ha rpe r andRow. Cody , M. L., and H. A. Mooney. 1978 . Converge nce versus nonconverge nce in Medit er ranean-clim ate ecosyt em s. Annual Revie w ofE colog y and Sys te m ati cs 9:265-321. Cole, L. C. 1954. The po p u lati on conseq u ences of lif e h is t o ry ph enomena. Qu arte r ly R evie w of B io l og y 29: 103-137. Connell, J. 1983 . On th e pr evalence and relati ve im p o rta n ce of int e r spe cif ic c omp e ti tion : evid e nce from fiel d exp e ri m ents . A meric a n Natu r ali st 122:661-696. Connolly, J. 1986. On dif ficu ltie s wit h rep la cement- s eri es meth o dolog y in mixt u re exp e ri me nts . Jo urnal of Ap plied Ecolog y 23: 125-137. Coop e r, A. W. 1981 . Above-g r ound bio m ass accumulati on and net prim a ry pro ducti on durin g the firs t 70 ye ars of successio n in Pop u lus gr andid e nta t a sta nd s on po or site s in nort he rn lower Mi ch ig a n . In D . C. West, H. H. Shug art, and D. B. Botk in, eds., Forest Successio n : Concep ts a ,u J Ap plica ti on s, 339-360. New York: Sp ri n g e r-Verlag. Coop er , W. S. 1913. The clim a x fore st of Isle Roy al e, Lake Sup er i or , and its d evelop m ent. Bota n ic a l Gazett e 55:1-44, 115-140, 189-235. Coop er , W. S. 1923. The recent ecolog ica l histo r y of Glaci er Bay, Alaska. Ecolog y 4:93-128, 223-246, 355-365. Coop e r, W. S. 1939. A fourth exp e dit ion to Glacie r Bay, Alaska. Ecolog y 20:130-155.
Covic h , A. 1972. Ecolog ica l economi cs of seed consump tion by Peromy sc us -a g ra p h ic a l model of resource substi tut i on . Transacti on s of the Connecti cu t Academ y of A rts and Scie n ce 44:71-93 . Cowan, M. L. 1986. Growt h respo nses of old-fi el d pla nts to a nit rog e n gra die n t. M .S . Thesis , Univ e rsit y of Mi nn esota . Cowles, H. C. 1899. The ecolog ica l relati on s of the vege ta t i on on the sand dunes of Lake Mi ch ig a n. Bota n ic a l Gazatt e 27:95-117, 167-202, 281-308,361-391 .I Crocker, R. L., and B. A. Di ck son. 1957. Soil develop m ent on the recessio nal morain e s of the Herbert and Mendenhall gla ci er s of south e aste r n Alaska. Jo urnal of E colog y 45:169-185. Crocker, R. L., and J. Majo r . 1955. Soil develop m ent in relati on to vege ta t io n and sur fac e age at Glacie r Bay, Alaska. Jo urnal of E colog y 43:427-448. de Wi t, C . T. 1960. On comp e ti tion . Ag r ic u ltu r al Research Rep o rt s(V ersl. landbouwk. Onderz.) 6 6.8, Wage nin g e n, The Neth e rlands. 88 pa ge s. Donald, C. M. 1951 . Comp e ti tion among pa stu re pla nts . I. Intr a- spe cif ic comp e ti tion among annual pa stu r e pl ants . Austr a li an Jo urnal of Ag r ic u ltu r al Research 2:355- 3 76. Farrow, E. P. 1916. On the ecolog y of the veg e ta t i on of Breckland. IL Jo urnal of E colog y 4:57-64. Farrow, E. P. 1917, On the ecolog y of the vege ta t i on of Breckland. lY. Jo urnal of E colog y 5:104- 1 13. Fit ter , A. H. 1986. Acq ui s i t ion and uti liza ti on of resources. In M. J. Crawley, ed., Plant Ecolog y, 375-405. Oxfo rd : Blackwell Scie n ti fic Publi ca ti on s . Powells, H. A. 1965. Sil vic s of Forest Trees of the Unit ed Sta t e s . U. S.
Dep a rt m ent of Ag ri c u ltu r e, Washin g ton , D. C. Fowler, N. 1981. Comp e ti tion and coexis t e n ce in a North Carolin a gr assland. II. The eff ec ts of the exp e rim enta l removal of spe cie s . Jo urnal of E colog y 69:843-854. Fox, J. F. 1977. Alte r nati on and coexis t e n ce of tre e spe cie s . Americ a n Natu rali st 111:68-89. Gi fford , R. M., and C. L. Jen kins . 1981 . Prospe cts of app ly i ng knowledg e of ph oto s yn th e sis tow ard im p r ovin g crop pr oducti on . In Gorin d je e , ed., Photo s yn th e sis : CO2 Assim i lat i on and Plant Producti vit y , Vol. 2. New York: Academi c Press. Gi vn is h , T. J. 1982. On the adap tive sig n if ica nce of leaf heig h t in fore st herbs. Americ a n Natu r ali st 120:353-381 Gleason, H. A. 1917. The str uc tu re and develop m ent of pla nt assoc iat i on . Bulletin oft he Torrey Bota n y Club 44:463-481. Gleason, H. A. 1927. Furt he r vie w s on the successio n concept . Ecolog y 8:299-326. Gleason, H. A. and A. Cronq ui s t . 1963. Manual of Vascular Plants of North e aste r n Unit ed Sta t e s and Adja c ent Canada. New York: D. Van Nostr an d Comp an y. Godwi n , H. 1956. The Hi st o ry of th e Brit ish Flora. Cambrid g e : Cambri dg e Univ e rsity Press. Green hill, G. 1881 . Dete r mi na ti on of the gre ate s t heig h t consis t e n t wi th sta b il ity tha t a vert ica l po le of mast can be made, and of the grea te s t heig h t to whic h a tree of give n pro p o rt ion s can grow . Proceedin g s of the Cambrid g e Phil oso p h ic a l Soc iet y 4:65-73. Gr iga l, D. F., L. M. Chamberlai n, H. R. Fin n ey, D. V. Wroblewski and E. R. Gross. 1974. Soil s o f the Cedar Creek Natu ral Hi st o ry Area.
Mi sc ellaneous Rep o rt No. 123. St. Paul, MN: Univ e rsity of Mi nn esota Ag ri c u ltu r al Exp e rim enta l Sta t i on . Grim e, J. P. 1979. Plant Str a te g i es and Veg e ta t i on Processes. Chic h este r : Joh n Wi ley & Sons. Grim e, J. P., and R. Hunt. 1975. Relati ve gro wt h rate : its rang e and adap tive sig n if ica nce in a local flor a. Jo urnal of E colog y 63:393-422. Gr im e, J. P., and D. W. Jef f ery . 1964. Seedli ng esta b lish ment in vert ica l gra die n ts of sunlig h t. Jo urnal of E colog y 53:621-642. Grim m, E. C. 1984. Fir e and oth e r fac to r s contr o llin g the big woods veg e ta t i on of Mi nn esota in the mi d -ni n e te e nth centu r y . Ecolog ica l Monog ra p h s 54:291-311. Guti er rez, J. R., and W. G. Whit ford . 1986. Respo nses of Chih u ahuan Desert herbaceous annuals to rain f a ll aug m enta t i on . Jo urnal of Arid Envir o nments , in pre ss. Hair s to n , N. G., F. E. Smi th, and L. B. Slobodkin . 1960. Communit y str u ctu r e, po p u lati on contr ol , and comp e ti tion . Americ a n Natu rali st 94:4 2 1-425. Hanawalt, R. B., and R. H. Whit tak er. 1976. Altit ud in a lly coordi na te d pa tt er ns of soil s and veg e ta t i on in th e San Jac in to Mounta i n s , Calif ornia. Soil S c ien ce 121 : 114-124. Ha rpe r, J. L. 1961. Ap pro aches to the stu d y of pla nt comp e ti ton . In F. L. Mi lth o rp e , ed., Mechanis m s in Bio l og ica l Comp e ti ton . Sym p . Soc. exp. Bio l . 15:1-39. Ha rpe r, J. L. 1977. Pop u lati on Bio l og y of P lants . London: Academ ic Press. Ha rpe r, J. L. and J. Og d en. 1970. The repr oducti ve str at e g y of high er pla nts . 1. The concep t of str at e g y wi th spe cia l refe r ence to Senec io
vulga ris L. Jo urnal of E colog y 58:681-698. Harris , G. P. 1986. Phy top l ankto n Ecolog y: Str u ctu r e, Functi on and Fluctu a ti on . London: Chap m an and Hall. Hole, F. D. 1976. Soil s o f Wi sc onsin . Madis o n: Univ e rsit y of Wi sc onsin Press. Horn, H. S. 1971 . The Adap tive Geometr y of Trees. Prin c eto n : Prin c eto n Univ e rsit y Press . Howrath , S. E., and J. T. Wi lliam s. 1972. Chry s an the mum seg et u m . In Bi ol og ica l Flora of the Briti sh Isles, Jo urnal of Ecolog y 60:573-584. Hsu, S. B., S. P. Hubbell, and P. Waltm an. 1977. A math e mati ca l the ory for sin g l e-nutr i e n t comp e ti tion in conti nu ous cultu res of micr oorga nis m s. SIAM Jo urnal of Ap plied Math e mati cs 32:366-383. Hubbell, S. P., and R. B. Foste r . 1 986. Bi ol og y, chance, and his t o r y and the stru c tu re of trop ica l rain fore st tree communit ies . In J. Di am ond and T. J. Case, eds., Communit y Ecolog y, 314-330. New York: Ha rpe r andRow. Hubbell, S. P., and P. A. Werner. 1979. On measur ing the i n 떠 ns i c rate of inc rease of po p u lati on s wi th hete r og e neous life h is t o r ie s . Americ a n Natu r ali st 1 13:277-293. Inouy e , R . S. 1980. Densit y- depe ndent ge rm ina ti on respo nse by seeds of desert annuals. Oecolog ia 4 6:235-238. Inouy e , R. S., G. S. By e rs, and J. H. Brown. 1980. Ef fec ts of pre dati on and comp e ti tion on surviv o rshi p , fec undit y, and communit y str uc tu r e of d esert annuals. Ecolog y 61:1344-1351 . Inouy e , R. S., and W. M. Scha ffer. 1981 . On the ecolog ica l meanin g of rati o (de Wi t) dia g r am s in pla nt ecolog y. Ecolog y 62:1 6 79-1681 .
Inouy e , R. S., N. J. Huntl y , D. Til m an, J. R. Teste r , M. A. Sti llw ell, a nd K. C. Zin n el. 1987a. Old field successio n on a Mi nn esota sand pla in . Ecolog y 68:12-26. Inouy e, R. S., N. J. Huntl y, D. Til m an, and J. R. Teste r . 1987b. Pocket go p h ers, vege ta t i on and soil nit ro g e n along a successio n sere in east centr a l Mi nn esota . Oecolog ia 72: 178-184. Iwasa, Y., and J. Roug h g a rden. 1984. Shoot/ r oot balance of pla nts : op tim al gro wt h of a sys t e m wi th many vege ta t i ve orga ns . Theoreti ca l Pop u lati on Bio l og y 25 : 78-104. Jan zen, D. H. 1970. Herbiv o res and the number of tre es in tro p ica l fore sts . Americ a n Natu r ali st 104:501-528 . Jarv is , P. G., and M. S. Jarv is . 1964. Growt h rate s of woody pla nts . Phys i o l og ia Pl. 17:654-666. Jen ny, H. 1980. Soil ge nesis wi th ecolog ica l pe rspe cti ve s. Ecolog ica l Stu die s , vol. 37. New York: Sp ring er -Verlag. Ki ra , T. 1964. Meta b olic asp e cts of th e tro p ica l rain for est. Natu r e(Shiz e n) 19:22-29. Ki ra , T., H. Og a wa, and K. Shin o zaki. 1953. Intr a spe cif ic c omp e ti tion among hig h er pla nts . 1. C omp e ti tion -densit y-yield int e r -relati on shi ps in regu larly disp e rsed po p u lati on s. J. Inst. Polyt ec h. Osaka Cy . Univ . D.4:1-16. Kurio w a , S . 1960 . Intr a spe cif ic c omp e ti tion in a arti fici a l sunfl ow er communit y. Bot. Mag. Tok yo 73:300-309. Lassoie , J. P., T. M. Hinc kley, and C. C. Gr ier . 1985. Con ifero us fore sts in the Pacif ic N orth w est. In B. F. Chabot and H. A. Mooney, eds., Phy s io l og ica l Ecolog y of North Americ a n Plant Communit ies , 127- 161 . London: Chapm an and Hall.
Lawes, J., and J. Gi lbe rt. 1880. Ag ri c u ltu r al, bota n ic a l and chemi ca l results of exp er i m ents on the mi xe d herbage of per manent gra ssland, conducte d for many yea rs in successio n on the same land . I. Phil os op h ic a l Transacti on s oft he Roy a l Soc iety 171:1 8 9-514. Lawlor, L. R. 1979. Di re ct and ind ir e ct eff ec ts of n-spe c i es comp et i tion . Oecolog ia 43:355-364. Lawrence, D. B. 1958. Glaci er s and vege ta t i on in South e aste r n Alaska. Americ a n Sc ien ti st 4 6:89-122. Lawrence, D. B. 1979. Pri m a ry versus seconda ry successio n at Glacie r Bay Nati on al Monument, south e aste r n Alaska. In R. M. Lin n , ed., Proceedin g s of the Fir s t Confe rence on Sc ien ti fic Research in the Nati on al Parks, 213-224. Lawrence, D. B., R . E. Schoenik e, A. Qu is p e l , and G. Bond. 1967. The role of Dr ya s drummondi in veg e ta t i on develop m ent foll owi ng ice recessio n at G laci er Bay, Alaska, wit h spe c i al refe r ence to its n itrog e n fixa ti on by root nodules. Jo urnal of E colog y 55:793-813. Levin e , S. H. 1976. Comp et it ive int e r acti on s in ecosys te m s. Americ a n Natu r alist 1 10:903-910. Lik e ns, G. E., F. H. Bormann, R. S. Pie r ce, J. S. Eato n , and N. M. Joh nson. 1977. Bio -ge o-chemi st r y of a Foreste d Ecosys te m . New York: Sp ringe r-Verlag. Lind sey, A. A. 1961. Veg e ta t i on of the drain a g e - a erati on classes of nort he rn India n a soil s in 1830. Ecolog y 42:432-436. Loach, K. 1970. Shade tol erance in tree seedli ngs: II. Growt h analys i s of pla nts rai se d under artificial shade. New Phyt ol og ist 6 9:273-286. Lubchenco, J. 1978. Plant spe c ie s div e rsit y in a marin e int e r ti da l communit y: im p o rta n ce of herbiv o re foo d pre fe r ence and alga l
comp e ti tive abil it i es . A meric a n Natu r ali st 112:23-39. MacMahon, J. A. 1981. Successio n al pro cesses: comp ar is o ns among bio m es wi th spe cia l refe r ence to pro bable roles of and inf l ue nces on anim als. In D. C. West, H. H. Shug a rt , and D. B. Botk i n , eds., Forest Successio n: Concep ts and Ap pl ic a ti on s, 227-304. New York: Sp ri n g e r -Verlag. Mahmoud, A., and J. P. Gr im e. 1976. An analys is of comp e ti tive abil ity in thr ee pe rennia l gra sses . New Phy tol og ist 7 7:4 3 1-435 . Marks, P. L. 1983. On the ori gin of the field pla nts of the nort he aste r n Unit ed Sta t e s . Americ a n Natu r ali st 122:210-228. May, R. M. 1973. Sta b il ity and Comp l exit y in Model Ecosys te m s. Prin c eto n : Pri nc eto n Univ e rsit y Press. McMahon, T. A. 1973. Siz e and shap e in bio l og y. Scie n ce 179:1201-1204. Melill o, J. M., J. D. Aber, and J. F. Murato r e. 1982. Ni trog en and lignin contr o l of hardwood leaf litt er decomp o sit ion dy n ami cs . Ecolog y 63:621-626. Mi ller, H. G., and J. D. Mi ller. 1976. Ef fec t of nitrog en sup ply on net prim ary pro ducti on in Corsic a n pine . Jo urnal of Ap pli e d Ecolog y 13:249-256. Mi lton , W. 1947. The yiel d, bota n ic a l and chemi ca l comp o sit ion of natu r al hil l herbag e under manurin g , contr o lled gra zin g and hay condit ion s. I. Yie l d and bota n ic al . J o urnal of E colog y 35:65-89. Monsi, M. 1968. Math e mati ca l models of pla nt communit ies . In F. E. Eckardt, ed., Functi on in g of Terrestr i a l Ecosys te m s at the Prim ary Producti on Level, 131-149. Lie g e , Belgi um : Va illa nt- C armanne, S. A. Mooney, H. A. 1972. The carbon balance of pla nts . Annual Revie w of
Ecolog y and Sys te m ati cs 3 :3 15-346 . Mooney, H. A. 1977. Converge nt Evoluti on in Chil e a nd Cali forn ia . Str ou dsburg, P A: Dowden, Hutc h in s on & Ross. Newberry, D. M., and E. I. Newman. 1978. Comp e ti tion betw een gras sland pla nts of dif fere nt ini t ial siz e s. Oecolog ia 33:361-380. O' Brie n, W. J. 1974. The dy n ami cs of nut rien t lim i tat i on of ph y top la nkto n alga e: a model reconsid e red. Ecolog y 50:9 3 0-938. Odum , E . P. 1960 . Orga nic pr oducti on and tu rnover in old fiel d successio n . Ecolog y 41 :34-49. Oksanen, L., S. D. Fretw ell, J. Arrud, and P. Ni em ala. l-98 1. Exp lo it at i on ecosys t e m s in grad ie n ts of prim ary pro ducti vi t y. Americ a n Natu r ali st 118:240-261 . Olson, J. S. 1958. Rate s of successio n and soil chang es on south e rn Lake Mi ch ig a n sand dunes . Bota n c ial Gazett e 119: 125-169 . Oria n s, G. H., and R. T. Pain e . 1983. Converge nt evoluti on at the commun ity level. In D. J. Futu ym a and M. Slatk in, eds., Coevoluti on , 431-458. Suth e rland, MA: Sin a uer Assoc iat e s . Pacala, S. W. 1986. Neig h borhood models of pla nt po p u lati on dy n ami cs . 4. Sin g le -spe ci es and multis p e cie s models of annuals wi th dormant seeds. Americ a n Natu r ali st 128:8 5 9-878. Pacala, S. W. , a nd J. A. Sil a nder, Jr. 1985. Neig h borhood models of pla nt po p u lati on dy n a mics . 1. Sin g le -spe cie s models of annuals. Americ a n Natu r ali st 1 25:385-411 . Pa rrish , J. A. D., and F. A. Bazzaz. 1982. Respo n ses of pla nts from thr ee successio n al communit ies to a nutr ien t grad ie n t. Jo urnal of Ecolog y 70:233-248. Pasto r , J., J. D. Aber, C. A. McClaug he rt y, and J. M. Melill o. 1982.
Geolog y, soil s , and veg e ta t i on of Blackhawk Island, Wi sc onsin . Americ a n Mi dl and natu r ali st 108:266-277 . Pasto r , J., J. D. Aber, C. A. McClaug h erty , and J. M . Melill o . 1984. Aboveg r ound pr oducti on and N and P cy c li ng along a nit ro g e n mi ne rali za ti on gra die n t on Blackhawk Island, Wi sc onsin . Ecolog y 65:256-268. Pasto r , J., M . A. Sti llw ell, and D. Til ma n. 1987. Ni tro g e n mine rali za ti on and nit rif i ca ti on in fou r Mi nn esota old fiel ds. Oecolog ia 71:481-485. Pig o tt , C. D., a nd K. Tayl o r. 1964. The dis t r ibu ti on of some woodland herbs in relati on to the sup ply of nit rog e n and ph osph orus in the soil . Jo urnal of E colog y 52(sup ple ment) : 175-185. Qu in n , J. F., and A. E. Dunham. 1983. On hyp o th e sis tes ti ng in ecolog y and evoluti on . Americ a n Natu r ali st 122:602-617. Rabin o vit ch -Vi n, A. 1979. Infl ue nce ofp a rent rock on soil pro p e rt ies and comp o sit ion of veg e ta t i on in the Galile e. Ph. D. Thesis . Hebrew Univ e rsit y of Jer usalem. Rabin o vit ch -Vi n, A. 1983 . Infl ue nce of nutr ien ts on the comp o sit ion and dis t r ibu ti on of pla nt communit ies in Med iter ranean- T y pe ecosys te m s of Israel. In F. J. Krug e r, D. T. Mi tch ell, and J.V .M. Jarv is , eds., Medit er ranean-Typ e Ecosys te m s, 74-85. New York: Sp ringe r-Verlag. Rapp o rt , D. J. 1971 . An op timiza ti on model of foo d selecti on . Americ a n Natu r ali st 1 05:575-578. Rapp o rt, D. J., and J. E. Turner. 1975. Feed ing rate s and po p u lati on grow t h. Ecolog y 56:942-949. Raunkia e r, C. 1934. The Li fe F orms of Plants and Sta t i st i ca l Plant Geog ra p hy . London: Oxfo rd Press . Rein e rs , W. A., I. A. Worley, and D . B . Lawrence. 1971 . Plant dive rsit y
in a chronoseq ue nce at G lacie r Bay, Alaska. Ecolog y 52:55-69. Ric e , E. L., W. T. Penfo u nd, and L. M. Rohrbaug h. 1960. Seed dis p e r sal and mi ne ral nutr i t ion in successio n in abandoned fiel ds in centr a l Oklahoma. Ecolog y 41:224-228. Ri ck lefs , R. E. 1987. Communit y div e rsit y: relati ve roles of local and regi on al pro cess. Sc ien ce 235: 167-171. Roberts o n, G. P. 1982. Facto r s reg u lati ng nit rif ica ti on in pr im ary and seconda ry successio n . Ecology 63:1561-1573. Robert so n, G. P., a nd P. M. Vit ou sek. 1981 . Ni trif ica ti on po te n ti al s in prim a ry and seconda ry successio n . Ecolog y 62:376-386. Salisb ury , E. J. 1942. The Rep r oducti ve Cap a c ity of Plants . London: G. Bell and Sons, Ltd . Schaff er , W. M. 1981 . Ecolog ica l abstr a cti on : the conseq u ences of reduced dim ensio n ality in ecolog ica l models. Ecolog ica l Monog r ap h s 51:383-401 . Schoener, T. W. 1983. Fie l d exp e ri m ents on int e r spe c ific comp e ti tion . Americ a n Natu r ali st 122:240-285. Schop f, J. W., J. M. Haye s, and M. R. Walte r . 1983. Evoluti on of ear th' s earlie s t ecosy s te m s: recent pr og re ss and unsolved pro blems. In J. Wi lliam Schop f, ed., Earth ' s Earlie s t Bi os ph ere: Its Orig in and Evoluti on , 361-384. Pri nc eto n : Pri nc eto n Univ e rsit y Press. Shamsi, S. R. A., and F. H. Whit eh ead. 1977. Comp a rati ve eco- ph y s io l og y of Ep ilob iu m hir s utu m L. and Ly thr um sali ca ria L. III. Mi ne ral nutr ition . Jo urnal of E colog y 65:55-70. Shaver, G. R., and F.S. Chap in, III. 1980. Respo nse to fertiliza ti on by vario us pl ant gr owt h for ms in an Alaskan tu ndra: nutr ien t accumulati on and grow t h. Ecolog y 61:662-675.
Snayd on, R. W. 1962. Mi cr o-dis t r i b u ti on of Trifo li um rep e ns L. and its relati on to soil s f ac to r s. Jo urnal of E colog y 50: 133-143. Snayd on, R. W. 1970. Rap id po p u lati on dif fere nti at i on in a mosaic envir o nment. I. The respo nse of Anth o xanth u m odoratu m po p u lati on s to soil s. Evoluti on 24:257-269. Snayd on, R. W. 1976. Geneti c chang e wit hi n spe cie s . In J. M. Thursto n , G. V. Dy k e and E. D. Wi lliam s, The Park Grass Exp e rim ent, 14-20. Ha rpe ndon, U.K. : R oth a mste d Exp e rim enta l Sta t i on . Snayd on, R. W. , a nd M. S. Davie s . 1972. Rap id po p u lati on dif fere nti at i on in a mosaic envir o nment. II . Morp h olog ica l varia t io n in Anth o xanth u m odoratu m . Evoluti on 26:390-405. Sonleit ne r, F. J. 1977. A sto c hasti c comp u te r model for sim ulati ng po p u lati on gro wt h. Researches on Pop u lati on Ecolog y 19:10-32. Sp itter s, C.J .T . 1983. An alt er nati ve app ro ach to the analys is of mixe d crop ping exp e ri m ents . 1. Esti m ati on of comp e ti tion eff ec ts . Neth erlands Jo urnal of A g ri c u ltu r al Sc ien ce 31 : 1-11. Ste b bin s , G. L., and G. J.C . Hi ll. 1 980. Di d multic e llular pla nts inv ade the land? Americ a n Natu r ali st 1 15:3 4 2- 3 53. Ste m , W. R., and C. M. Donald. 1962. Ligh t relati on ship s in gras s-clover swards. Austr al i an Jo urnal of A g ri c u ltu ral Research 13:599-614. Summerhaye s , V. 1941. Ef fec ts of voles(Mi cr otu s ag re stis ) on vege t a tion . Jo urnal of E colog y 29:14-48. Swank, W. J., J. B. Waid e , D. A. Crossley, Jr. , a nd R. L. Todd. 1981 . Insect defo liat i on enh·an ces nit ra te exp o rt from for est ecosys tm es . Oecolog ia( B e rlin ) 51:297-299. Tamm, C. 0. 1985. The Swedis h op tim um nutr ition exp e ri m ents in fore st sta n ds-ai m s, meth o ds and yiel d results. Jo urnal oft he R oy a l Swedis h
Academy of A g r ic u ltu r e and Forestr y , Sup pl. 17:9-29. Tamfomo ,d Cp. la0 n.,t s a . n Pd aAg e. sA r1o8n1ss-o1n 9.6 1. 9i8n2 .O Op pt itmim i zui mn g n uYtire ilt di osn— oTf hseo mReo lneo nof- Fertil iz e rs, Proceedin g s of the 12th Cong r. Int. Pota s h Insti tut e , Bern. Tamm, C. 0., A . Aronsson, and H. Burgt or f. 1974. The op tim um nutr ition exp e rim ent Str as an . A brie f descri ption of an exp e rim ent in a you ng sta n d of Norway spr u ce(Pic e a abie s Karst. ) . Rap po rte r och Up ps ats e r, No. 17, Dep t. of Plant Ecolog y and Forest Soil s . Roy a l College of F orestr y, Sto c kholm, Sweden. 29 pp. Tansley, A. G. 1949. The Briti sh Isles and Their Veg e ta t i on . Cambri dg e: Cambri dg e Press . Thursto n , J. 1969. The eff ec t of lim i ng and fer t iliz e rs on the bota n ic a l comp o sit ion of pe rmanent gra ssland, and on the yiel d of hay. In I. Rori so n, ed., Ecolog ica l Aspe cts oft he Mi ne ral Nutr i t ion of P lants , 3- 10. Oxfo rd : Blackwell Sc ien ti fic P ubli ca ti on s. Ti lm an(Tit m an), D. 1976. Ecolog ica l comp e ti tion betw een alga e: exp e rim enta l confi rm ati on of resource-based comp e ti tion the ory. Sc ien ce 192:463-465. Ti lma n, D. 1980. Resources: a gr ap h ic a l-mechanis t i c ap pr oach to comp e ti tion and pre dati on . Americ a n Natu r ali st 116:362-393. Ti lm an, D. 1982. Resource comp e ti tion and Communit y Str u ctu r e. Pri nc eto n : Prin c eto n Univ e rsit y Press. Til ma n, D. 1983. Plant successio n and gop he r dis t u r bance along an exp e ri m enta l grad ie n t. Oecolog ia 6 0:285-292. Til m an, D. 1984. Plant domi na nce along an exp e rim enta l nutr i e n t grad ie n t. Ecolog y 65:1445-1453. Til m an, D. 1985. The resource rati o hy po th e sis of successio n . Americ a n
Natu r ali st 1 25:827-852. Til m an, D. 1986a. Evoluti on and dif fere nti at i on in ter restr i a l pla nt communit ies : the im p o rt an ce of the soil resource:lig h t grad ie n t. In J. Di am ond and T. Case, eds., Communit y Ecolog y, 359-380. New York: Ha rpe r and Row. Til m an, D. 1986b. Ni tro g e n-li m i ted gr owt h in pla nts from dif fere nt successio n al sta g e s. Ecolog y 67:555-563. Til m an, D. 1987a. Secondary successio n and the pa tt er n of pla nt domi n ance along exp e ri m enta l nit ro g e n gr adie n ts . Ecolog ica l Monog ra p hs 57:189-214. Til m an, D. 1987b. The im p o rt an ce of the mechanis m s of int e r spe cif ic int e r acti on . Americ a n Natu r ali st 129:769-774. Toumey, J. W. and R. Ki en holz, 1931 . Trenched plo ts under for est canop ies . Bull. No. 30, School of Forestr y, Yale Univ e rsity , New Haven , Connectic u t. 31 pp . Vandermeer, J. H. 1980. Indir e ct mutu a li sm : Va riat io n s on a the me by Ste p h e n Levin e . Americ a n Natu r ali st 116:441-448. Vit ou sek, P. 1982. Nutr ien t cyc lin g and nutr ien t use eff ici e n cy. Americ a n Natu r ali st 1 19:553-572. Vit ou sek, P. M., J. R. Gosz, C. C. Gr ier , J. M. Meli llo , W. A. Rein e rs, and R. L. Todd. 1979. Ni tra te losses from dis t u rbe d ecosys te m s. Sc ien ce 204:469-474. Vit ou sek, P. M., J. R. Gosz, C. C. Gr ier , J. M. Meli llo, and W. A. Rein e rs. 1982. A comp a rati ve analys is of po te n ti al nit rifica ti on and nit rat e mobil ity in for est e cosys t e m s. Ecolog ica l Monog r aph s 52:155-177. Vi tou sek, P. M., and P. A. Mats o n. 1985. Di st u rbance, nit ro g e n avali ab il ity, and nit ro g e n losses in an int e n siv e ly manage d loblolly
pine pla nta t i on . Ecolog y 66: 1360-1376. Walker, J., C. H. Thomp so n, I. F. Fergu s , and B. R. Tunsta l l. 1981 . Plant successio n and soil develop m ent in coasta l sand dunes of subtr o p ica l easte r n Austr al ia . In D. C. West, H. H. Shug art, and D. B. Botk i n , eds., Forest Successio n : Concep ts a nd Ap pli c a ti on s, 107-131 . New York: Sp ring e r-Verlag . Waller, D. M. 1986. The dy na mi cs of gro wt h and form . In M. J. Crawley, ed., Plant Ecolog y, 291-320. Oxfo r d: Black-well Scie n t ific Publi ca ti on s . Walte r , H. 1985. Veg e ta t i on of the Earth and Ecolog ica l Sys te m s of the Geo-bio s ph ere, 3rd ed. New York: Sp ri n g er -Verlag. Wa ring , R. H., W. H. Emm ing ton , H. L. Gholz, and C. C. Gr ier . 1 978. Va riat i on in maxim um lea f area of conif ero us fore sts in Orego n and its ecolog ica l sig n if ica nce. Forestr y Sc ien ce 24:131-140. Werner, P. A., and W. J. Platt . 1976. Ecolog ica l relati on ship of co- occurrin g go ldenrods(Soli da g o : Comp o sit ae ) . Americ a n Natu r alis t 110:959-971 . West, D. C., H. H. Shug a rt, and D. B. Botk i n (eds.). 1981 . Forest Successio n : Concep ts a nd Ap plica ti on s. New York: Sp ring e r-Verlag. Westo b y , M. 1984. The self- thi n n in g rule. Advances in Bi ol og ica l Research 14:167-225. W 血 e, A. S. 1983. The eff ec ts of 13 yea rs of annual pre scrib e d burn ing on a Qu ercus ellip s oid alis communit y in Mi n nesota . Ecolog y 64: 1081-1085 . Whitne y, G. 1986. Relati on of Mi ch ig a n' s pre sett le ment pine fore sts to substr at e and distu rb ance histo r y . Ecolog y 67: 1548-1559. Whittaker , R. H. 1953. A consid e rati on of clim a x the ory : the cli ma x as a
po p u lati on and pa tt er n. Ec ol og ica l Monog r ap h s 23:41-78. Whit tak er, R. H. 1975. Communit ies and Ecos ys te m s. New York: Macm illa n Press. Wi en s, J. A. 1984. Resource sys te m s, p o p u lati on s, and communit ies . In P. W. Pric e , W. S. Gaud, and C. N. Slobodchik o ff , e ds., A New Ecolog y, 397-436. New York: Wi ley Inte r scie n ce. Wi llis, A. 1963. Braunto n Burrows: the eff ec ts on the veg e ta t io n of the addit ion of mi ne ral nutr i e n ts to the dune soil s . Jo urnal of Ecolog y 51:353-374. Wi ls on, S. D. , and P. A. Keddy . 1986. Sp e cie s comp e ti tive abil it y and po sit ion along a natu r al stre s s/dis t u r bance gra die n t. Ecolog y 67: 1 236-1242. Woods, K. D. 1979. Recip r ocal rep la cement and the main t e n ance of codom ina nce in a beech- m apl e fore st. Oik o s 33:3 1 -39. Woods, K. D. , a nd R. H. Whit tak er. 1 981 . Canop y- un dersto r y int e r acti on and th e in te r nal dy n ami cs of matu re hardwood and hemlock- hardwood fore sts . In D. C. West, H. H. Shug art, a nd D. B. Botk in, eds., F orest Successio n : Concep ts and Ap plica ti on s, 305-322. New York: Sp ring e r -Verlag. Worley, I. A. 1973 . The black crust ph enomenon in upp e r Glacie r Bay, Alaska. N orth w est S c ien ce 47:20-29. Zak, D. R., K. S. Preg itze r, and G. E. Host. 1986. Landscape va riat i on in nit rog e n mine rali za ti on and nitrifica tio n . Canadia n Jo ur!'al of Forest Research 16:1258-1263. Zedler, J., and P. Zedler. 1969. Assoc iat i on of sp e cie s and the ir relati on ship to micr oto p o g ra p h y wi thi n old field s. Ecolog y 50:432-442.
찾 0 ~.!i!.:71
기 가변성 46 가소성 317- 3 27 가우스 반응 297 개체군 동태 25 경쟁 능력의 통합 개념 286 경쟁 동태 197-214 경쟁 우세종 197 경쟁적 교체 30, 205 경쟁적 평형 11 계절성 141 고유치 30 공간적 구조 327 공간적 이질성 328 공존 54,91 광 구배 85 광합성 산물 68 교란 구배 17 교체 계 열 222, 223 구부러진 등구배곡선 87 기계론적인 모델 63 기계론적인 접근 방법 22,24 L 내음성 179 E: 다년성 140
다영양 단계 157 다종 경쟁 55,203 단일종 지소 29 대응책 13, 27, 58 둥구배 곡선 38, 100 □ 모래평원 255 모의 모델 74 무기화율 178 밀도 독립적 18 밀도 조작 실험 222 닌 반포화율 88 분배 가소성 183-185 분배비 118, 119 분배 삼각형 119 분배 패 턴 29, 64, 109, 110, 183 불안정한 공존 96 불안정한 평형점 95,96 人 산록군계 계열 166 상대 생장률 32 상대수도 233 상호 작용의 필수 자원 41 생 리 적 가변성 43, 47, 195
생리적인 대응 131 생산력 구배 15,61 생장 동태 81 생장 등구배곡선 28,37 생장률 30 생 활사 76, 109, 144 소비억제 42 소비 억제곡선 43, 45 손실률 30, 34 손실률 구배 17, 125 수직적 광구배 85 순환율 70 숲톰 329 습도 구배 166 식물 경쟁의 동태 202 식물 상관 159 식생 패턴 149, 160 。 R* 29, 30 안정한 공존 95 ALLOCATE 72, 73 야초 진화 249-251 에너지 분배 14 영양억제곡선 43 영양염류 공급률 78, 85, 91, 208, 210 2 차 천이 228, 244
일시적 인 동태 215, 219, 246, 303 일시적인 우점 217, 219 1 차 천이 228 大 자연 선택 59 자원 경쟁 27-34, 63 자원 공급률 15,48 자원 공급 벡터 50, 51 자원 공급점 50 자원비 58 자원비 가설 232 자원 소비 30 자원 소비 벡터 49 자원 소비율 48 자원 수준 15,29 자원 이용도 15 자원 획득 이론 318 전체 영양염류 78 정착-경쟁 가설 251 정착-초식 가설 252 제로 순생장 등구배곡선 57 제약 13 제한 자원 29 종간 경쟁 47,88 종내 경쟁 52 종자의 크기 135, 180 질소 구배 268
질소 무기화율 172, 174, 265 天 초강력 경쟁종 58 최대 광합성률 66 최대 상대 생장률 65 최대 생물량 56 최대 생장률 64, 65, 67, 150 최대 영양 생장률 67 최적 획득 41,42 더 크기 구조 20 E 타감 효과 23 톡양극상 177, 179 토양 모재 177
토양 자원 15 토양 자원:광 구배 125, 144 퇴행천이 234 투영행렬 30 五 평형 상태 36 평형점 52 표현형의 가소성 317 피드백 효과 179 필수 자원 38,47 己 형태적 가변성 43,46,47 형태적 가소성 194 호흡률 70 환경제약 13
지은이 소개 데이비드 틸만은 McKnig h t Di st i ng u is h ed Un i vers ity의 생 태 학 교수이 며, Un ive rsity o f M i nneso t a 의 Cedar Creek Natu r al Area 의 연구책 임자 이다. 그의 연구 분야는 종간 경쟁의 기작, 생물 다양성이 유지되 는 과정 생물 다양성이 개체군과 생태계의 안정성 및 기능에 미 치는 영향, 그리고 이러한 주제와 관련된 수학적인 이론들이다. 지난 15 년 동안 그는 Cedar Creek Natu ra l Area 에서 생물 다양성과 생태계의 동태를 연구하였다. 또한 새로 침입하는 종의 생물학적 조절, 그리고 서식처의 분단이 생물 다양성에 미치는 영향에 관해 서도 연구하였다. 1984 년에는 Gug ge nheim Fellow, 1985 년에는 Ameri ca n Associa t i on for the Advancement of Sc i ence 의 특별 회 원을 역임하였고, 1989 년에는 미국생태학회로부터 W. S. Coo p er 상을 수 상하였으며, 1995 년 Ameri ca n Academy of Arts and Sc i ence 에 선정 되 었다. 또한 같은 해에 Conservati on B i olo gy에 Pew Scholar 로 선정되 었으며, 1996 년에는 미국생태학회에서 Mac Arth ur 상을 수상하였다. 그는 미국생태학회에서 발간하는 새로운 학술지의 창립 편집자였 고, Sc i ence 의 편집위원으로 활약하고 있다. 그는 Pri nc eto n Mono- grap h 총서로 두 권의 책을 출판하였고, 그 밖에 두 권의 책을 편 집하였으며, 100 편 이상의 논문을 발표하였다.
문형태 서울대학교 생물교육학과 졸업 서울대학교 대학원 식물학 박사 뉴멕시코 주립대학 박사후 연구원 현재 공주대학교 생물학과 교수 식물의 전략과군집 구조 대우학술총서 번역 114 1 판 1 쇄 펴냄 • 1998 년 3 월 30 일 지은이 • 데이비드 틸만 옮긴이·문형태 펴낸이·朴孟浩 펴낸곳· (주)민음사 출판등록 1966. 5. 19 제 16-490 호 서울특별시 강남구 신사동 506 대표전화 515-2000 팩시밀리 515-2007 값 • 22,500 원 한국어판 © (주)민음사, 1998 식물학. 환경에 대한 식물의 적응 KDC/481.52 Pri nte d in Seoul, Korea ISBN 89-374-4114-4 94470 89-374-3000-2( 세 트)
L 대우학술총서안 역 ) 1
1 유목민족제국사 칸텐 / 송가중 2 수학의 확실성 클라인/박세희 3 중세철학사 와인버그 / 강영계 4 日本語의 起源 밀러 / 검방한 5 古代漠語音韻學槪要 칼 그렌/최영애 6 말과 사물 푸코/이 광래 7 수리철학과 과학철학 와일/김상문 8 기후와 진화 피어슨 / 검준민 9 이성 진리 역사 파트남 /검 효명 10 사회과학에서의 場理論 레빈/박재호 11 영국의 산업혁명 딘/ 나 경수 이정 우 12 현대과학철학논쟁 쿤 外/조승옥 김동식 13 있음에서 됨으로 프리고 진/ 이철 수 14 비교종교학 바하/검종서 15 동물행동학 하인드/장현갑 16 현대우주론 시아마/양종만 17 시베리아의 샤머니즘 디오세지 호괄/최길성 18 조형미술의 형식 힐데 브란트 /조창 섭 19 힐버트 리 드/ 이일해 20 원시국가의 진화 하스 / 최몽룡 21 商文明 張 光直/윤 내현 22 마음으 r생 태학 베히혼尻적봉 23 혼 돈 속의 질서 프리고진 스텐저스/유기풍 24 생명의 기원 밀러 오르겔/박인원 25 일반언어학이론 야콥슨 1 권재일 26 국가권력의 이념사 마아네케/이광주 27 역사학 논고 브로델/이정옥 28 유럽의식의 위기 I 0~ 자르/조한경 29 유럽의식의 위기 l o t자르/조한경 30 일반국가학 켈젠/민준기 31 일반언어학강의 소쉬르/최승언 32 일반체계이론 버틀란피/현승일 33 현대문명의 위기와 기술철학 애거시/이군현 34 언어에 대한 지식 촘스키/ 이 선우 35 음운학원론 트루베쪼코이/한문희 36 우주의 발견 하위트/강용희 37 수학적 발견의 논리 라카토스/우정호 38 텍스트 사회학 지마/허창운 39 현대물리학의 철학적 테두리 보음/전일동 40 과학과 가치관의 우선순위 스페리/이남표
41 신화의 진실 휘브너/이규영 42 대폭발 실크/홍승 수 43 大同書 1 康有爲/이성애 44 표상 포더 / 이영옥 정성호 45 과정과 실재 화이트헤드/오영환 46 그리스 국가 에렌버그/김진경 47 거대한 변환 풀 라니/박현수 48 법인류학 포스피실/이문웅 49 언어철학 울스튼/곽강제 50 중세 이슬람 국가와 정부론 램톤/ 김정 위 51 전통 쉴즈 / 김병서 신현순 52 몽골문어문법 뽀 빼/ 유원수 53 중국신화전설 | 원가 / 전인초 김 선자54 중국신화전설 ll 원가/전인초·검 선자55 사회생물학 1 윌슨/이병훈 박시룡 56 사회생물학 il 윌슨/이병훈 박시룡 57 일반언어학의 제문제 l 밴베니스트/황경자 58 일반언어학의 제문제 ll 밴베니스트/황경자 59 폭력과 성스러움 지라르 /김진 식 박무호 60 갑골학 OO 년 董作합/ 이형구 61 현대수학의 여행자 피터슨 / 김안수 주형관 62 프랑스 혁명의 지적 기원 모르네/주명철 63 해석학과 과학 커널리 코이트너/이유선 64 서양고대경제 핀리/지동식 65 음악사회학 베 버/이건용 66 앙띠 오이디푸스 들뢰즈 가따리/최명관 67 과학커뮤니케이션론 프라이스/남태우 정준민 68 교양교육의 개혁 다니엘 벨/송미섭 69 과학과 가치 래리 라우든/이유선 70 초기제국에 있어서의 교역과 시장칼 폴라니/이종욱 71 폭정론과 저항권 헬라 만트/심재우 72 생명과학철학 데 이 비드 헐/하두봉 구혜영 73 사랑의 역사 쥴리 아 크리스 테바 /김영 74 大地의 노모스 칼 슈미트/최 재 훈 75 일반 공법학 강의 레 옹 뒤 기 / 이 광윤 76 텍스트학 반 다이크/정시호 77 문명의 발생 찰스 레드만/최몽룡 78 근대국가의 발전 G 폿지/박상섭 79 과학적 발견의 패턴 N R 핸슨/송진웅 조숙경 80 아랍 문학사 R A 니콜슨/사회만 81 18 세기 중국의 관료제도와 자연재해 P. E 빌/정철옹
82 역사비교언어학개론 R 안틸라/박기덕 남성우 83 계몽주의 철학 E 카시러 / 박완규 84 토양 미생물학과 생화학 폴 클라크/이도원 ·조병철 85 수학, 과학 그리고 인식론 I 라카토시/이영애 86 봉건제의 이해 러쉬튼 쿨본/검동순 87 그라마톨로지 자크 데리다/검성도 88 殷代貞卜人物通考 1 라오쏭이 / 손예철 89 殷代貞卜人物通考 l 라오쫑이 / 손예철 90 殷代貞卜人物通考 l 라오쫑이 / 손예철 91 순수경제학 레옹 왈라스/심상필 92 문화유물론 마빈 해리스 / 유명기 93 포유동물의 가축화 역사 줄리엣 클루톤 브록 / 김준민 94 르네상스 철학에서의 개채와 우주 E. 카시러/박지형 95 화학적 진화 SF 메이슨/고문주 96 과학의 한계 바이츠제커/송병옥 97 러시아 연해주와 밭해 역사 예 웨 샤브꾸노프 / 송기호 정석배 98 경제정책의 원리 발터 오이겐 / 안병직 황신준 99 비코와 헤르더 이사야 벌린 / 이종홉 강 성호 100 실용주의의 결과 리처드 로티/김동식 101 국제 지층구분 지침서 A 살바도르/백인성 102 국가론 헤르만 헬러/홍성 방 103 경험과 판단 후설/ 이 종훈 104 17 세기 프랑스 정치사상 앙리 쌔/나정원 10518 세기 프랑스 정치사상 〈근간〉 106 里鄕見聞錄 유재건/실시학사 고전문학 연구회 107 수학학습심리학 리처드 스켐프/황우형 108 고생태학 로버트 도드 로버트 스 탠 턴/ 양 승영 109 지리학의 본질 l 리처드 하트손/김덕현 외 110 지리학의 본질 Il 리처드 하트손/ 김 덕 현 오 111 수학과 물리학 유리 마닌 1 명효철 채 동호 112 과학의 합리성 뉴턴 스미스/양형진 조기숙 113 뇌의 진화 존 애클스/박찬웅