
존 에클스 Joh n C. Eccles 오스트레일리아의 생리학자 멜버른 대학을 졸업한 후 옥스퍼드 대학에서 셰링턴의 지도를 받아 신경생리학을 연구하였다, 신경세포의 흥분과 억제 기구를 발견한 공로로 1963 년, 호지킨 헉슬리와 함께 노벨 생리 의학상울 받았으며 이러한 그의 연구는 신경 계통의 질병 치료와 심장 뇌의 기능 연구에 큰 영향을 미쳤다. 저서로 『신 경세포생리학 』 『 뇌에 대한 이해 』 등이 있다. 박1-초꽝 서울대학교 의과대학 졸업 서울대학교 대학원 의학박사 현재 서울대학교 의과대학 교수 서울대학교 의학연구원 원장겸 신경과학연구소 소장
-자뇌의아 의진 화창 조
EVOLUTION OF THE BRAIN: Creati on of the Self by Joh n C. Eccles Cop yright © 1989 Joh n C. Eccles All right reserved. Korean Translati on Cop yright © 1998 Minu msa Publis hing Co., Ltd. Korean transla ti on edition is pub lis h ed by arrang em ent with Routl ed g e. 이 책의 한국어 판 저작권은 Rou tl edg e 와 독점계약한 (주)민음사에 있습니다. 저작권법으로 한국 내에서 보호를 받는 저작물이므로 무단 전재와 무단 복제를 금합니다.
 뇌의 진화
뇌의 진화
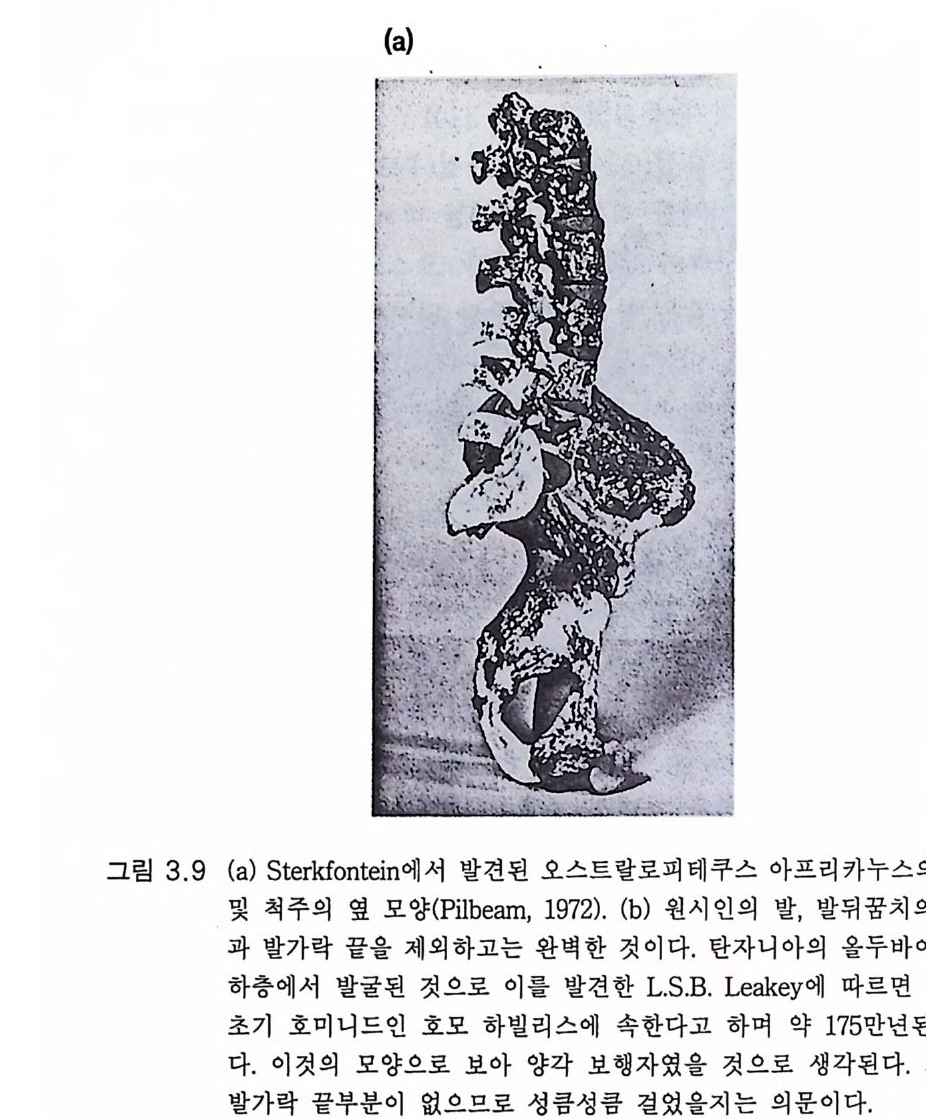
책의 번역에 부쳐 지금으로부터 36 억년 전에 나타난 생명의 기원이 돌연변이와 자연 도태를 거쳐 다시는 반복되지 않을 호모 사피엔스 사피엔스라는 진화 의 최종 산물에 이르는 과정을 살펴보는 일은 실로 마음 설레는 일이 라 아니할 수 없다. 뇌의 진화 과정 특히 현재 지구상에 살고 있는 사람의 뇌가 있기까 지의 과정은 특히 관심의 대상이 아닐 수가 없다. 물론 저자가 지적했 듯이 사람 뇌의 진화에 대한 명쾌한 해답은 아직까지는 없다 . 현대 과 학은 인간의 발자국을 달에 남기게 하였고 생명의 원천이라고 할 수 있는 유전자를 조작하기에 이르렀다. 그럼에도 불구하고 뇌의 기능은 아직도 신비의 커튼이 벗겨지지 않고 있다 . 사람은 의사 소통에 언어를 이용할 수 있는 유일한 동물인가? 그 언 어의 사용은 언제부터 시작되었는가? 사람 이외의 동물에게도 감정이 있는가? 동물의 행동은 학습에 의한 것인가 아니면 타고난 본능적 행 동의 일환인가? 그 발달 과정은 어떠했는가? 영혼이란 무엇인가? 사 람이 자아를 인식하게 되는 과정은 어떠했는가? 온통 궁금한 것투성 이댜 이들 궁금증을 규명하기에는 너무나도 부족한 기록과 근거밖에는 없으나 저자는 이를 제한된 기록과 근거를 가지고 뇌의 진화, 자아의 창조 과정을 더듬어 보고자 하였다. 이 같은 과학적 실증적 시도는 기 록과 실험 기법의 제한으로 결국 철학적, 종교적 논의에 이르게 되기 는 하지만 언젠가는 우리가 해결해야 할 과제인 것이다. 이진숙 교수는
마음의 학문에서의 진화를 논하면서 < 사람은, 특히 삶의 정신은 신이 특별하게 만든 까닭에 동물계를 지배하고 있는 자연의 법칙의 지배를 받지 않고 자연법칙과는 전혀 다른 고귀한 신의 법칙에 지배되고 있다. 이런 사람은 다윈의 진화론 때문에 치명적 타격을 받게 되었다 > 고 하 였다 랙 Lack 은 <영혼이란 영성을 견지해야 하므로 이것의 존재에 대 한 의문은 과학의 범위 밖에 있는 것이다. 따라서 다윈 학설과의 관계 에서 문제가 생기지 않는다 > 고 하였고 저자는 이에 동의하고 있다. 그러나 저자 에클스는 마음의 학문도 과학적으로 접근해야 한다는 입장에서 다윈의 진화론적 접근을 시도하고 있다. 저자의 의도한 바를 충실하게 전달하기에는 역자의 능력이 너무나 도 모자란다고 생각하나 이 방면에 관심을 가진 사람들에게 소개하고 싶은 욕심에서 감히 번역을 시도하여 보았다 . 부분적으로는 대단히 난 해한 곳이 없지 않으리라 생각되나 전체적으로 의미를 파악할 수는 있 으리라 생각된다. 좋은 책을 소개해 준 서울대학교 의과대학 생리학교실 검전 교수에 게 감사드리며 그 바쁜 미국의 인턴 수련 과정중에 시간을 쪼개어 번 역 초고를 꼼꼼히 읽고 다듬어 준 나의 서랑 이경민 군에게 깊이 감사 한다 그리고 책을 번역할 수 있도록 지원하여 주신 대우재단과 출판 올 맡아 애써주신 민음사에 심심한 감사의 말씀 올립니다 .
4
차례
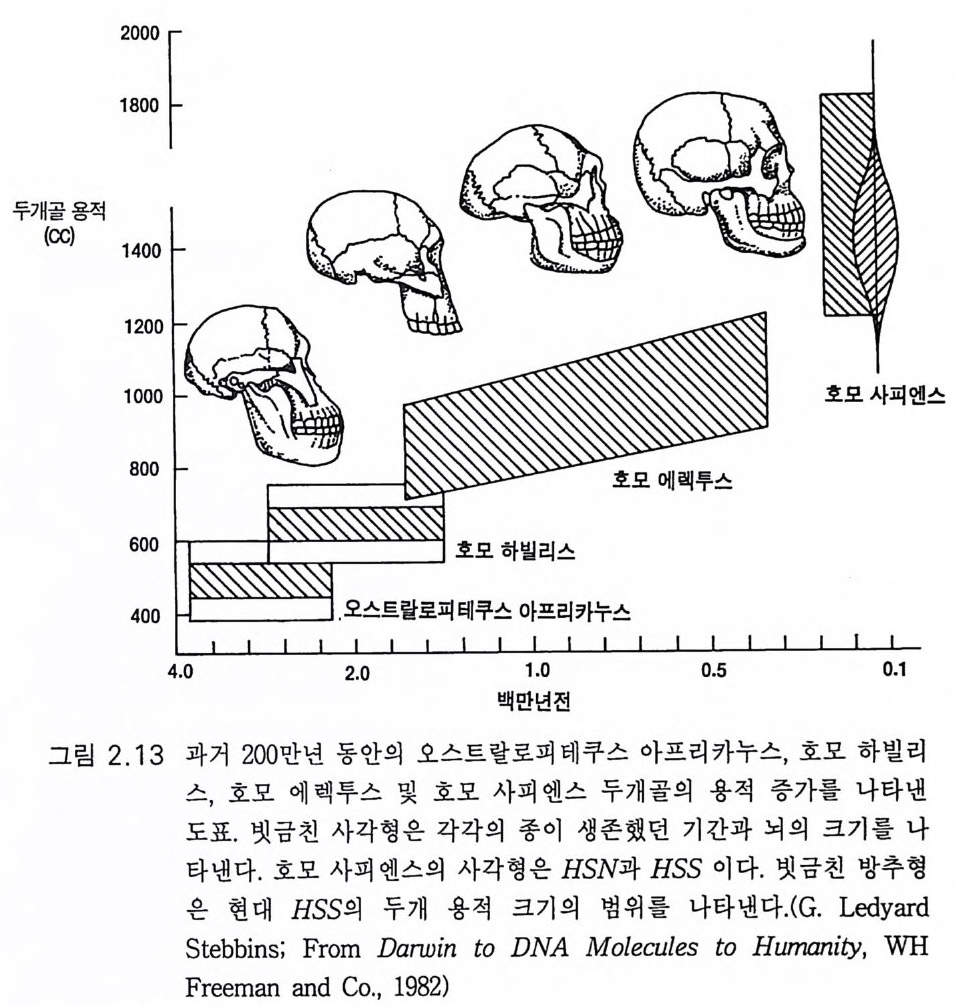
책의 번역에 붙여 3서문 9제 1 장 생물학적 진화 131.1 유전자 부호 • 131.2 현대적인 종합이론 : 점진적 계통발생설 • 201.3 한시적 평형 • 211.4 유인류 진화의 유전 기전 • 231.5 종의 진화적 기원에 관한 일반적 결론 • 25제 2 장 인간 진화의 일반론 272.1 유인류 조상 • 272.2 오스트랄로피테신 • 332.3 호모 하빌리스 • 432.4 호모 에렉투스 • 442.5 네안데르탈 • 472.6 호모 사피엔스 사피엔스 • 512.7 호미니드 진화의 문제점 • 59제 3 장 호미니드 뇌의 진화 633.1 기본 구조 • 633.2 뇌의 기능적 작동 • 70
3.3 직립, 보행 및 반응 • 753.4 운동의 미세 조절을 위한 신경기전의 발달 • 923.5 능숙한 손 움직임 • 101제 4 장 호미니드 진화와 언어 소통 1074.1 언어의 수준 • 1074.2 언어적 표현 • 1094.3 인간 언어의 학습 • 1114.4 유인원의 언어 훈련 • 1144.5 언어 중추와 관계되는 대뇌피질의 해부 • 1214.6 청각 회로 • 1294.7 언어 발달과 관계되는 뇌의 진화 • 1324.8 언어 생성의 진화 • 1374.9 언어와 진화적 생존 • 139제 5 장 대뇌 변연계 감정 및 생식계 진화의 관계 1435.1 해부학적 개관 • 1435.2 변연계와 감정의 표현 • 1485.3 변연계와 시상하부의 약리학 • 1515.4 영장류 진화 과정중의 변연계 구성분의 크기 지수 • 1535.5 호미니드 진화에 있어 뇌 증대의 결과 • 1555.6 호미니드의 인구학적 전략 • 1585.7 애타심의 진화 • 166제 6 장 시각-운동 진화 1716.1 영장류 대뇌피질의 시각 영역 • 1716.2 일차 시각피질에서의 신경 반응 • 173
6.3 입체시 • 1776.4 선조전 시각 영역 • 1796.5 선조 및 선조전 영역의 손상 • 1846.6 시각반응에서 사람 뇌의 연구 • 1866.7 시각 진화에 대한 결론 • 1916.8 석기문화의 진화 • 1946.9 골패의 판화 • 195제 7 장 학습과 기억의 진화 ·2057.1 호미노이드 조상의 모델로서의 유인원 • 2057.2 의사소통을 위한 부호의 학습 • 2107.3 유인원의 학습과 인간 학습의 비교 • 2127.4 기억에 관계되는 뇌의 부위별 크기 • 2137.5 학습과 기억의 신경과학 • 2167.6 사람 기억의 일부 특별한 양상 • 2437.7 결론 • 246제 8 장 진화상의 마음과 뇌 문제 2498.1 역사적 고찰 • 2498.2 비인간 동물의 의식 • 2508.3 의식의 진화 • 2528.4 마음과 뇌 문제의 철학 • 2568.5 마음과 뇌 문제의 실험적 검사법 • 2598.6 마음과 뇌 상호작용에 관계되는 신경 구조 • 2678.7 마음과 뇌 학설들의 도형적 비교 • 2708.8 양자물리에 근거한 마음과 뇌 상호작용의 새로운 가설 • 2718.9 마음과 뇌 문제의 재고 • 278
제 9 장 자아의 창조와 그 뇌 2819.1 해부학적으로 관찰된 비대칭성 • 2819.2 기능적 비대칭성 • 2839.3 대뇌신피질의 모듈적 설계 • 2919.4 진화적 극점 : 자아의식의 발단 • 2939.5 자아의 단일성 • 2969.6 교차연결 절개술 • 2979.7 대뇌 비대칭성의 진화적 의의 • 3049.8 신피질의 진화적 기원과 개체발생이 늦다는 발생학적 및 구조적 증거 • 306제 10 장 인간의 인격 31310.1 인간의 인격 • 31510.2 문화적 진화 • 31710.3 생물학적 및 문화적 진화 • 32110.4 마음의 개념 • 32610.5 특이적 인간 특성 • 33110.6 인간 특성의 진화 • 34110.7 자아 또는 영혼의 창조 • 342후기 : 반성과 상상들 347옮긴이 후기 355참고문헌 372찾아보기 399서문 생물학적 진화 과정 중 가장 중요한 창조 과정인 뇌의 진화, 즉 우 리의 유인 동물 조상으로부터 900 만내지 1000 만년의 세월을 통하여 걸 출한 창조력을 지닌 사람의 뇌에 이르기까지 인류 진화에 관한 서적은 의의로 드물다. 유인류로부터 인류 Homo sapi en s sap i ens 로의 진화에 관한 이야기는 아주 경이로운 것이다. 이는 곧 우리 자신에 관한 이야기이다. 우리 모 두는 과거사를 논함에 있어 유인류의 대단한 성공적 진화야말로 우리를 인간으로 존재할 수 있도록 한 유일한 기회였음을 인식해야 할 것이다. 그런데 왜 이 책에서 다루어진 것과 같이 인간의 뇌가 있기 까지의 근 본적 양상에 관하여 별로 다루어지지 않았을까? 이는 뇌의 진화에 관한 이야기가 근거가 없고 검증되지 못한 추론에 불과한 것처럼 보이기 때 문일 것이다. 많은 부분은 불명인 상태이거나 불충분하게 알려진 것임 은 사실이나 나는 합리적 비판을 엄격하게 적용한 창조적 상상력을 동 원하여 인간 뇌의 진화에 관한 매혹적안 이야기를 풀수 있었다. 다윈설 Darw ini sm, 심지어 그 합리성의 훼손이 유행하는 시기에 이 책은 생물학적 진화의 다윈설을 확인한다. 다만 계통 발생의 점진성은 때때로 한시 적 평 형p unc tu a t ed equilibr i um 같은 수정 (2 절 1.3) 그리고 염색체 재조합 가능성 (1 .4)에 자리를 내주는 일은 예외이다. 이 책의 내용은 단지 마지막 세 장만이 다윈의 실질적 개념을 벗어난 것이다. 거기서는 가장 논쟁의 초점이 되는 진화 현상을 다루었다. 첫째는 고 둥 동물에서 의식의 발현(제 8 장)이고 둘째는 유인류가 자아의식을 경
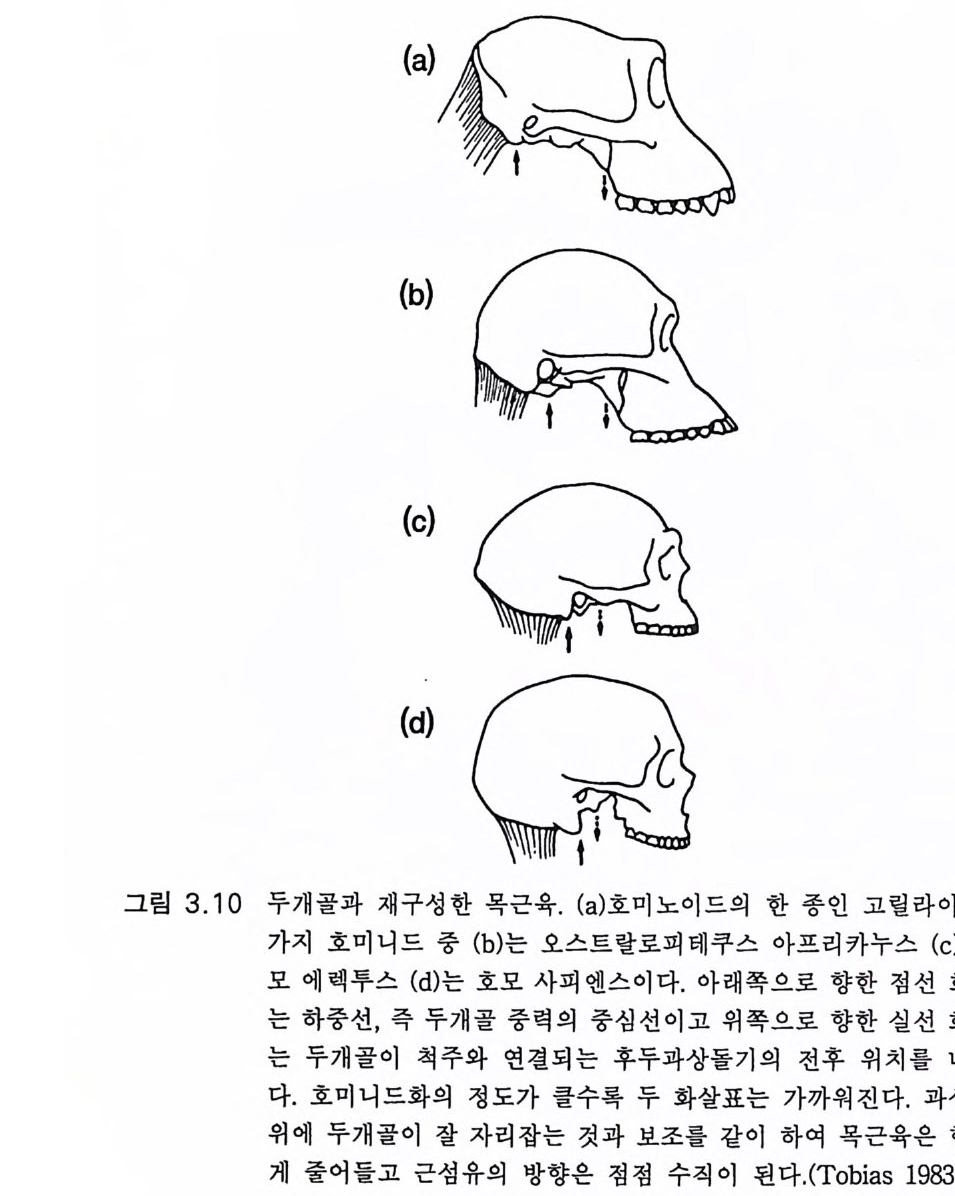
험(제 9, 10 장)했을 때 훨씬 놀라운 초월 현상이 있었다는 것이다. 유인 류 진화는 바로 그 시작부터가 신비이다. 단백연대법으로 밝혀진 바에 의하면 900 만내지 1000 만년 전에 사람상과(유인동물) 계통은 사람과(유 인류)와 폰지드과 계통으로 나누어진다 (2 절 2.1 및 표 2.1 ) 불행히도 사 람과 진화의 가장 결정적 시기인 이 시기 이후의 500 만년 동안은 거의 화석 이 소멸되 었다 (2 절 2.1 및 2.2). 아마도 그 기 간 동안 사람과의 개 체 수효가 대단히 적었을 것이다. 그 500 만년 동안에 3.3 에서 언급한 양각 보행으로의 진화적 변형이 있었다. 수상 서식 유인동물과 육상 서식 오스트랄로피테쿠스 사이에는 일련의 몇 단계가 있었을 것이다 . 커튼 이 걷혀진 400 만년 전, 화석 기록에 따르면 골격과 근육 계통이 완전히 양각 보행으로 변형되었음이 밝혀진다(그림 3.8 , 3.9 , 3.1 0 ). 놀랍게도 그 때 뇌는 조금밖에 커지지 않았다. 사각 보행에서 양각 보행으로 이행 하는 과정에는 양각 보행을 가능하게 하는 신경계의 변화가 있었을 것 임은 가장 훌륭한 화석 라에톨리 Lae t o li 발자국으로 나타난다.(그림 3.1 1 ) 제 2 장과 제 3 장에서 간단하게 언급한 바와 같이 최근 수십년 동안에 400 만년 전부터 근년에 이르기까지의 풍부한 유인류 화석이 발견되었 댜 뇌의 변형까지도 내부 주형 endocas t으로 확인되었다(그림 2.7, 2.9). 유인류 진화로 이루어진 변화를 말하려면 현대 폰지드의 뇌를 유인동 물 조상의 뇌 모델로 이용할 필요가 있다. 유인류 진화에서 뇌의 변화 롤 그려 보려는 시도는 하인츠 스테판 He i nz S t e p han 과 그 공동 연구자 들의 절묘한 연구에 크게 힘 입었다. 그들은 사람을 포함하는 다양한 영장류 뇌에서 핵 같은 해부학적으로 구별이 되는 구조의 크기를 측정 하였댜 계산된 크기의 지표들이 이 책의 여러 표들의 근거가 된다. 고등 영장류에 있어서만이 인간의 교묘한 감각 및 운동계를 가진 생물로의 진화 과정이 있었음을 인식해야 한다. 인간의 진화는 고등 영장류에서 이미 이루어진 진화 위에 더하여 이루어졌다. 특히 유인동
물에 의함이 그렇다 . 가장 좋은 예는 눈이 양안시로 완전하게 적응된 시각 계통이댜(제 6 장) 원시적 시각피질과 전줄무늬피질에 이르는 시 각 경로는 인간 Homo sa pi ens 에 이르는 진화에 있어 크게 변하지 않았 다 . 그중 가장 중요한 것은 대뇌피질로서(제 8, 9 장) 고등 영장류의 대뇌 피질은 사람과 매우 유사하다. 또 중요한 것은 변연계(제 5 장)와 학습 계통(제 7 장)이다 . 이것도 전반적인 구성이 사람과 매우 유사하다. 대뇌피질과 함께 간뇌의 가장 중요한 기능으로서 발생된 새로운 부 위 특히 언어 분야는 폰지드의 뇌에는 혼적에 불과했고 다론 영장류에 는 존재하지 않았다 . 제 9 장에서 기술한 바와 같이 이들 새로운 부위는 기능적으로 비대칭적이다. 이것은 가장 나중에 발생되었을 뿐 아니라 개체 발생 과정에서도 가장 나중에 기능한다. 제 9 장과 제 10 장에서는 특히 인간 대뇌피질의 이점에 관하여 집중적으로 살펴보았다. 이것은 영적 기능을 가지는 신신피질 neoneoco rte x 이라 한다. 영장류 진화에서는 우리가 보수적 지혜라고 하는 것이 있었다. 이는 진화적 잠언으로 표현할 수 있다. 매력적으로 보이는 단기간의 이득을 위해 물려받은 근본 형체와 홍정하지 않는다 . 예를 들면 마음대로 움 직일 수 있는 다섯 손가락을 발이나 발굽 또는 날개와 바꾸지 않는다 . 그래서 유인류 진화는 손가락을 가진 초기 척추동물로부터 시작하여 귀 중한 손과 발로 변할 수 있게 된다 .(3.3-3.5) 특히 손은 유인류 진화 에서의 걸작이고 결국 신경계의 발달과 함께 완성품을 이룬다 .(3.5) 다음과 같은 의문이 종종 제기된다. 우리의 진화 계통만이-오직 생각할 수 있는 한에서-우리들 것에 걸맞는 또는 그것을 능가하는 상상력과 지능을 가진 생물에 이르게 할 수 있는 것이었나? 예를 들어 어떤 초지능 원숭이가 유인류 계통을 능가하는 또는 그에 걸맞는 다른 진화계통을 창시할 수 있지 않았을까? 대답은 <절대 그렇지 않다>이 댜 유인류 진화는 기존의 유전자 정보군에서 분리된 대단히 작은 분 절들에 의한 불연속적인 진보에 의존한다. 더욱이 대단히 오랜 시간
一―수십만년_후에야 각각의 새로운 종이 탄생할 수 있었다. 이같 은 조건들은 이제 통신망과 그 통신망을 통해 연결된 개체들로 뒤덮인 지구 행성 Plane t E art h 에서 다시는 재연될 수 없다. 사실상, 과거에도 유인류 진화는 단 한번 있었을 뿐이고 그 이후에는 완전히 멸종당할 위험이 상존하는 가운데 극소수의 인구에 의해 그 유인류 진화의 결과 가 유지될 수 있었다 . 그래서 내가 이 책에서 묘사하고자 하는 지구 행성의 유인류 진화 에 관한 이야기는 다시는 반복되지 않을 것이다 . 인간 Homo sap ien s sap i ens 은 새로운 경쟁자의 출현을 두려워 할 필요가 없다. 이 책에서는 사람의 뇌 진화 과정에서 의식과 자아 의식의 생성을 중 점적으로 다루었다. 그전까지 의식이란 존재하지도 않던 세계에 의식과 자아 의식이 생성되는 이 신비로운 과정에 대하여 물리과학적인 설명이 불가능함을 인정한다 . 이 문제에 관한 철학적 고찰이 제 8, 9, 10 장에 있 . 고 이는 제 10 장에서는 우리가 각자 경험해 온 자아 인식 출현 과정에서 의 종교적 개념으로 이어졌다. 우리의 의식세계의 중심, 포퍼 Po pp er 의 제 2 세계(그림 9.5, 10 .4),에는 신에 의해 창조된 영혼이 있음을 암시하고 있다. 이 문제에 관하여는 부록 후반부에 좀더 논의하였다 .
제 1 장 생물학적 전화 l.l 유전자 부호 진화 과정의 본질을 이해하기 쉽게 설명하려면 우선 세포의 유전 물질인 데옥시리보핵산 (deox yri bonucle ic acid , DNA) 과 유전자 부호를 통한 이것의 작용 양상에 관하여 간략하게 짚고 넘어갈 필요가 있다. 이 진화의 기본 물질이 세포핵 속으로 분리된 것은 진핵단세포의 진화 초기인 18 억년 전에 이루어졌다. 이는 진화 발전에 대단히 중요한 것 으로 번식을 비롯한 모든 세포 활성의 중심인 복합 기구를 방어하였던 것이댜 세포핵의 DNA 는 조밀하게 꼬인 대단히 긴 이중나선이다 . 그림 1.1 에 보이는 바와 같이 각각의 가닥은 인산p hos pha te과 당ri bose 분자가 교대로 나열되어 있다 각각의 당에는 다음의 네 가지 분자 중 한 가 지가 결합된다. 푸린 염기인 아데닌과 구아닌 그리고 피리미딘 염기인 티민과 시토신이다. 두 나선은 3 .4 A 마다 효과적으로 교차 결합을 이루 고 있다(그림 1.1). 한쪽의 아데닌은 티민과 결합하고 구아닌은 시토신 과 짝을 이룬댜 그래서 그 서열은 다음과 같다.
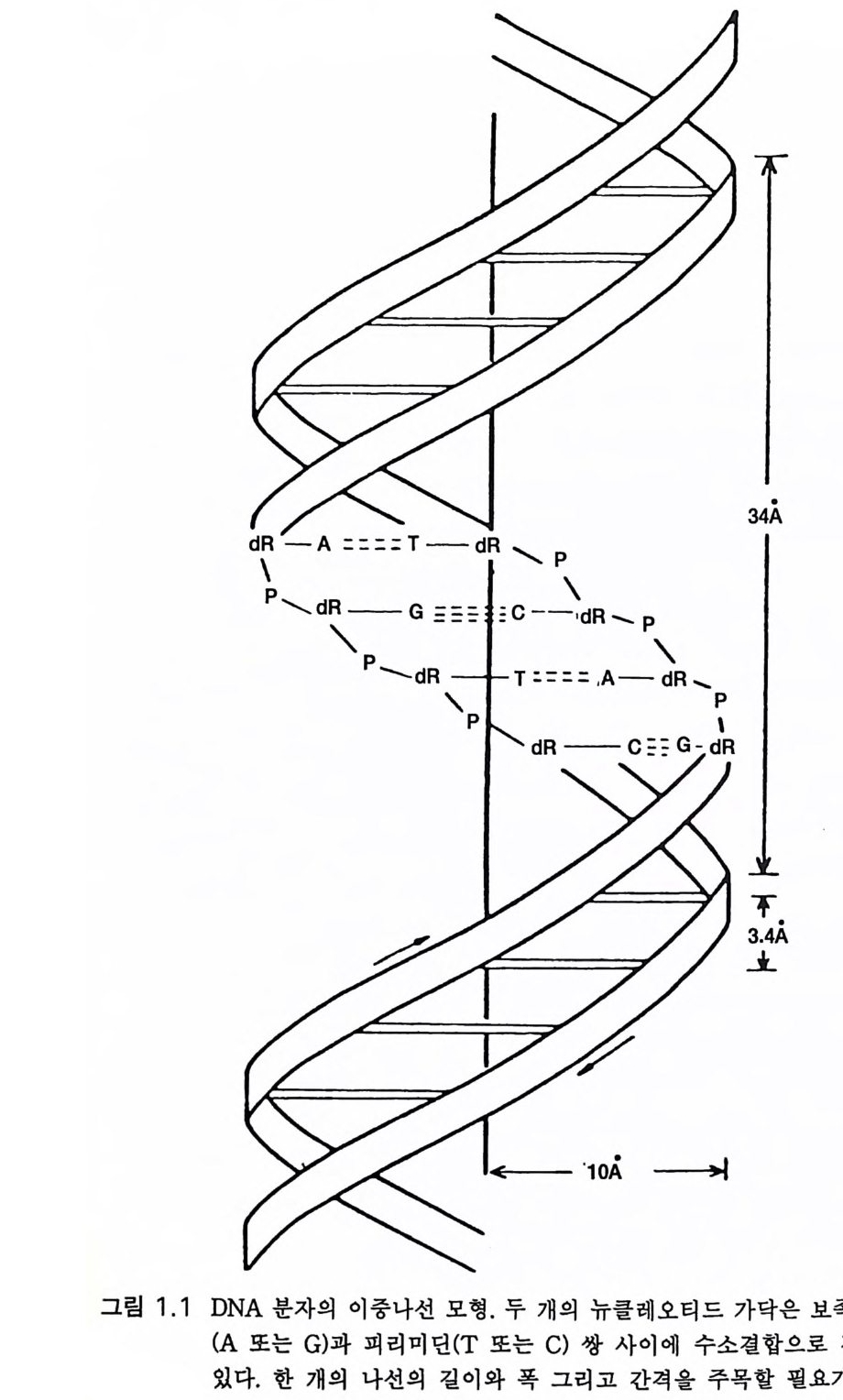 뼈 P\ _A]U d―-一[RP T\- —-G-RidR dC \ -lTP—-\- R 익―u《- d—8P R d\갱 l-P '.dR
뼈 P\ _A]U d―-一[RP T\- —-G-RidR dC \ -lTP—-\- R 익―u《- d—8P R d\갱 l-P '.dR
GTAGCAT CATCGTA 그래서 뉴클레오티드 부호는 각각의 가닥을 따라 일렬로 나열된다. 그 림 1. 2a 는 인산기와 당에 결합된 푸린 염기와 피리미딘 염기가 아데닌과 티민 사이 그리고 구아닌과 시토신 사이의 수소결합으로 이중나선을 이 루는 분자적 구조를 나타낸 것이다 . 그림 1. 2b 는 일렬로 나열된 DNA 부 호가 mRNA( 리보핵산)으로 전사되는 것을 나타내고 있다. mRNA 는 단백 구성 아미노산의 서열을 결정하는 세 자로 이루어진 부호로서 그림에 . 서 다섯 가지 특정 아미노산의 서열을 예로 들었다. 세균에서 각각의 가닥의 부호는 약 150 만 자로 되어 있다. 인간의 DNA 가닥은 약 350 만 자로 되어 있고 이것이 사람의 모든 세포를 구 성하는 기본 정보를 제공한다. 세포가 분열하기 전에 이중나선의 각 가닥은 분리되고 효소 반응에 의하여 각각의 보족 가닥을 만든다. 이 같이 재조성된 두 개의 이중나선은 원래의 것을 꼭 닮은 복사판이다. 세포를 구성하고 조절하는 유전 정보는 DNA 가닥을 따라 ATGC 로 나 타내는 뉴클레오티드 서열 속에 부호로 부여된다 . 단백 구성 아미노산 서열의 DNA 부호를 읽어내어(그림 l.2b) 세포의 구조를 구성하고 세포의 대사를 효소적으로 조절하는 유전자 전사와 복제의 자세한 과정을 논하는 것은 이 책이 목적하는 바가 아니다. 효 소는 단백질이다. 어떤 단백질이든 DNA 가닥에 일렬로 배열된 부호로 구성된다. 이것은 앞서 예로 든 바와 같이 몇 글자로 이루어진 짧은 것이 아니고 수천 자의 서열로 이루어진 유전자g ene 라고 하는 것이다. 유전자는 특정 단백을 구성하는 아미노산 서열의 자세한 명령을 가지 고 있댜 세균 세포가 살아가는 데 필요한 여러 종류의 단백을 구성하 기 위한 DNA 가닥이 300 만 자의 서열로 이루어졌다는 것은 지나친 것 이 아님을 알 수 있을 것이다. 사람의 세포에서는 그 숫자가 1000 배나
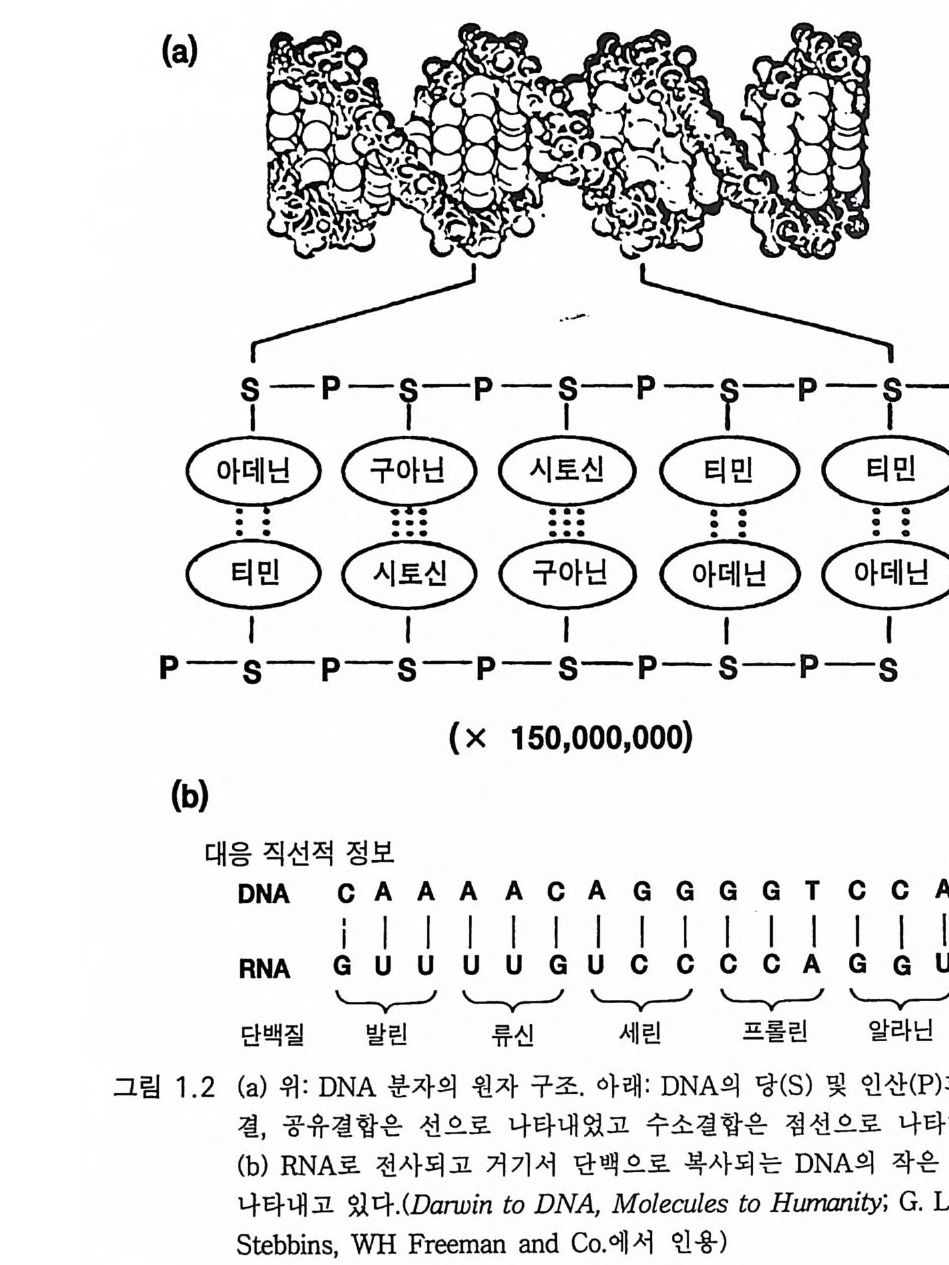 (a)
(a)
많아서 35 억 개에 이른다. 사람의 세포를 구성하는 데 필요한 정보로 는 다소 지 나친 감이 있다 . 도브잔스키 Dobzhansk y의 추산에 의 하면 사 람은 최소한 3 만 개의 유전자를 가졌다고 한다 . 단백이 평균 500 개의 아미노산 서열을 가졌다면 하나의 아미노산은 3 개의 뉴클레오티드 쌍 을 요하므로 1500 개의 뉴클레오티드 쌍을 필요로 하고 결국 3 만 개의 유전자는 4.5 X 10 7 개의 뉴클레오티드 쌍을 필요로 한다. 그러나 여분을 고려하면 숫자는 몇 배로 늘어날 수 있어서 1. 4% 라는 이 낮은 비율은 늘려 잡아야 한다. 더욱이 풀리지 않는 문제는 포유류 유전자계(게놈)의 최소한 30 - 70% 가 무활동이라는 것이다. 활성 DNA 분절을 분리시키는 DNA 스 페이서로 일부 설명이 되고 있다. 정상적으로 번식시에 DNA 위에 쓰인 직선적 부호는 정확하게 복사 된댜 따라서 세대를 거듭하여도 안정이 유지된다. 그러나 DNA 부호 내에 유전자 돌연변이라고 하는 변화가 일어난다 . 이는 복사 과정에서 잘못되어 한 개의 뉴클레오티드가 다른 것으로 치환되는 것으로 생각 한댜 즉 구아닌이 아데닌으로 바뀌든가 때로는 보다 근본적인 변화로 한 개 또는 몇 개의 뉴클레오티드 염기쌍이 절제되거나 전도되기도 하 고 때로는 상당히 큰 DNA 분절의 전도가 일어나기도 하는 것이다. 이 같은 복사 오류는 단백에서 다른 아미노산으로의 치환을 초래한다. 그 러나 이들 변화의 대부분은 개체의 번식이나 생존에 대하여 유해한 것 이므로 결국 자연도태 과정에서 제거된다. 다만 드물게 돌연변이가 번식이나 생존에 유리한 경우가 있다. 이 같은 돌연변이는 세대를 따라 전파될 것이고 이 돌연변이를 공유하는 생물계의 생존율을 향상시킬 것이다. 아직까지 여러 세대에 걸쳐 자연 도태에 의하여 이 같은 유리한 돌연변이가 모든 개체에서 나타나며 결 국 약간의 유전자형의 변화를 반영하는 것이다. 뒤이어 또다론 돌연변 이적 도태가 첨가되고 또 이것이 반복된다.
이것이 다윈의 자연도태설 또는 적자생존설에 대한 최신의 개정 개 념의 기초가 된댜 유리한 돌연변이는 선택되고 불리한 변이는 제거된 댜 고로 순전히 우연한 유전자 돌연변이라는 초기 과정에 의해 놀라 운 창조력과 적응 능력을 가진 생명체의 경이로운 모양과 기능으로의 자연도태가 이루어질 수 있었다. 이처럼 진화이론은 순전한 생물학적 과정으로서 현재는 그 원리가 잘 알려진 운행기전이다. 그리고 어떤 단순한 원시 생명의 형태가 아니고 모든 생명체 발생의 만족스러운 설 명으로서 받아들여진 것이다. 다윈과 월라스에 근거하는 이 이론은 인 류가 이룩한 가장 위대한 개념의 하나이다. 그러나 아직 개선의 여지 는 있 다(1. 2, 1.3) . 최근의 발전으로는 DNA 복사의 많은 오류는 실제에 있어서 중성임 을 알게된 것이다. 예를 들면 돌연변이로 인한 단백 일부의 아미노산 의 변화는 기능적으로는 치명적이지 않은 것이다. 또 DNA 일부의 돌 연변이는 단백 합성에 참여할 수 없으므로 도태 과정에서 중성이다. 어느 시기에 이 같은 중성 돌연변이가 많이 축적되어 원래 DNA 집단 의 상당 부분이 변하게 되면 이들 돌연변이는 그 환경에서 더 이상 중 성일 수는 없게 된다. 세포핵 속에서 DNA 는 2m 정도나 되는 대단히 긴 이중나선으로 존 재하는 것은 아니고 작은 조각으로 나뉘어 염색체를 이루고 있다. 염 색체는 세포가 분열하는 과정에 있을 때 뚜렷하게 나타난다. 사람의 유전자계(게놈)는 각기 특징적인 23 쌍의 염색체에 포장되어 있다(그림 1.3) . 감수 분열시 DNA 를 함유한 염색체는 나누어지고 분리되어 성세 포룰 만든댜 수정이 이루어지면 각각의 성세포로부터 온 반쪽씩으로 완전한 DNA 가 재구성된다. 유인 동물 중 네 가지 생물종은 그 핵구조가 대단히 유사하다(표 2.1 ). 폰지드 중 세 가지 종__침팬지, 고릴라, 오랑우탄一一은 48 개 염색체를 가졌다. 인간 Homo 에 있어 두 쌍의 염색체는 중앙 융합에 의
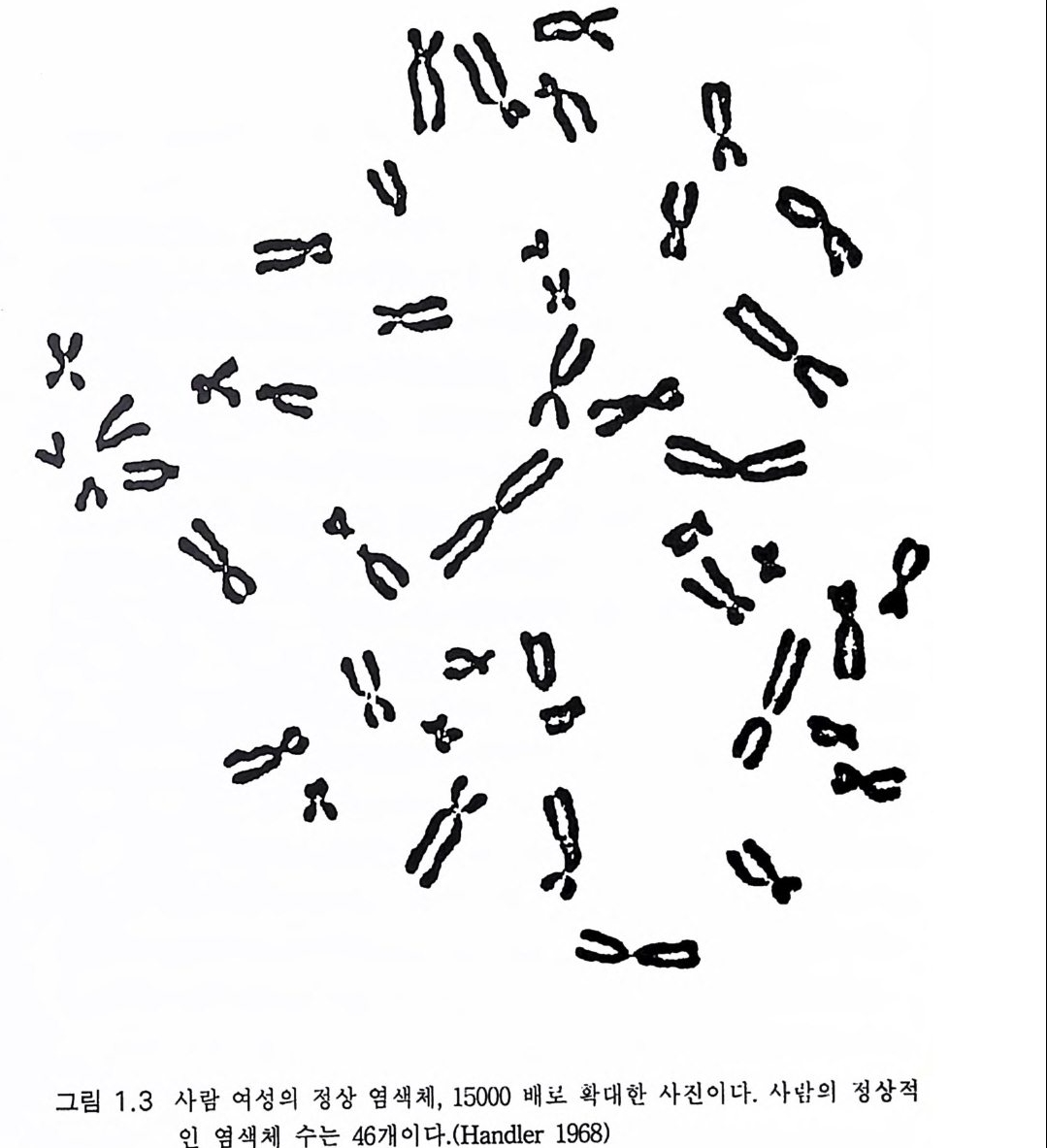 t
t
해 합쳐져 염색체 2 를 만든다. 따라서 인간의 염색체 는 46 개가 된다(그 림 1.3) . 다른 각도에서도 대단한 유사성을 보이는데 각각의 종의 염색 체의 무늬 모양이 예라 하겠다 . 1.2 현대적인 종합 이론: 점진적 계통발생설 (Ma y r 1963) 다윈 이후 생물학적 종이 진화의 단위로 인식되어 왔다. <종> 이라 함은 비상관성 개체들이 아니고 집단을 이루는 것이다. 한 가지 종의 집단은 수정 조건 때문에 다른 종과 생식성이 격리된다. 비교적 비슷 한 종끼리 같은 영역에 살 수는 있다. 그러나 같은 지역에 함께 살더 라도 교잡은 일어나지 않는다. 종이라는 것은 각기 치밀하게 짜여진 유전계로서 주어진 환경에서 분명한 생태적 조건에 맞도록 여러 세대 에 걸쳐 선택된 결과이다 . (Ma y r 1963:109) 다윈적인 관점을 통해 고생물학에서는 주거지역 내 수많은 개체들 이 대단히 느린 과정에 의한 선조집단의 변형으로 새로운 종이 형성된 다고 생각한다. 이 과정을 엘드레지 Eldred g e 와 굴드 Gould 는 1972 년에 계통 발생의 점진성이라고 하였다 . 불행히도 이 점진성은 화석 기록에 서는 찾아볼 수 없었다. 고전적 진화학자들은 이를 화석 기록의 불완 전성 탓으로 돌렸다. 화석 기록에는 진화과정에서 예리한 돌파 또는 돌변이 있었음을 나타낸다. 종의 유전적 다양성은 한 세대에서 다음 세대로 유전적 전파과정에서의 돌연변이, 재조합, 절제 등에 의한다. 그러나 무제한적 집단 번식의 세대교체 과정에서 유전자 흐름의 총체 적 과정으로 조절되는 것이다. 그렇지만 일란성 쌍생아를 제외하고는 성생식에 의한 집단 내 어떤 개체들도 유전적으로 동일하지 않다. 유전자 흐름의 혼합 효과에도 불구하고 계통 발생의 점진론이라는 현대적 종합 이론도 국소적 집단으로부터의 유추라는 관습을 견지하
고 종의 기원에 대한 범례로서 적응성 지역적 변이, 죽 자연도태에 의 한 점진적 대립인자 치환을 합당한 모델로 이용하였다 .(Gould 1982:134, 여기서 대립인자란 유전자를 총칭하는 것이다) 이 같은 총체적 개념을 공 통 조상적 연속성 s y m p a tri c con ti n uity이라고 한다 그러나 마이르(1 963) 는 분화는 작은 격리된 집단에서 보다 효과적이고 빠르게 일어날 수 있음을 인식하였다. 작은 기본 집단은 이주하여 커다란 조상 집단의 유전자 흐름으로부터 격리되었을 것이다. 그러나 이 모델은 주변 격리 집단의 계통 발생 점진성의 원리를 견지하였다. 수많은 세대를 통한 조그마한 적응성 변이의 효과적인 축적에 의하여 성공적인 분화가 이 루어졌다. 격리 집단이란 작은 군집 내에서 유전자 흐름의 혼합 효과 룰 감소시킨다는 데에만 그 이점이 있다. 1.3 한시 적 평 형 (Punctu a te d eq uilibr i um ) 엘드레지와 굴드는 새로운 종이 발생하는 것에 관하여 타조상적 종 화 allo p a tri c spe cia t io n 이론을 개진하였다. 국소적 소집단이 모집단의 지리적 영역 변두리로 격리될 때만 새로운 종의 발현이 일어날 수 있다. 그 같은 국소적 집단을 변두리 격리 집단이 라 한다 변두리 격리 집단은 유전자 흐름의 재창조를 방지하는 격리 기 전에 의하여 그리고 새로운 형질이 장래 어떤 시점에 조상을 다시 만나 면 새로운 종으로 발전한다 . 타조상설 allo p a tri c th eo ry에 따르면 새로운 종의 화석은 그들의 조상이 살았던 장소에서 생기지 않는다 .(1972: 94) 엘드레지와 굴드는 변두리 격리 집단에서 새로운 종의 발생은 종의 지속기간에 비하면 짧은 기간 동안에 생겼다고 주장하고 선조 종의 영
역으로 다시 이주하더라도 지역 공생 형태로 공존하기는 하지만 교잡 은 일어나지 않는다고 주장하였다. 이는 화석 기록에서 볼 수 있다. 따 라서 장시간의 정체기간은 타조상적 종화라는 돌발사건으로 구분지어 지는 것이다. 이것이 <한시적 평형설p unc tu a t ed e quili b ri a> 이다 . 위에서 언급한 바와 같이 계통 발생의 점진성은 오랜 기간에 걸친 도태에 의하여 축적되면서 적응되고 계승된 단위 유전자 돌연변이에 따른다. 보다 신속한 한시효과를 나타내는 대체 모델도 있다. 주요 염 색체의 변화는 변두리 격리 집단이 몇 세대 내에 종을 형성하는 데 필 요한 유전자 변화가 될 수도 있다. 이같이 종화 s p e ci a ti on 는 계통 발생 의 점진성에 있어 새로운 유전자를 만드는 전형적 점돌연변이에 의한 다기보다 유전자의 조절 또는 재배열에 의할 것이다. 또 조절 유전자 변화의 시기에 따라 커다란 표현형의 변화가 초래될 수도 있다. 이 같 은 방법으로 새로운 종이 발생될 것이다 .(Bush 등 1977) 카손 Carson 은 다음과 같이 제안하였다. 일련의 확률적인 유전적 대이변으로 종 분화가 시작되고, 종의 발생을 향한 중요한 유전적 도약이 이루어질 것이다. 이는 폐쇄적 변이계의 상위 초유전자의 비정상적인 강제적 재조직이 일어날 때 시작된다. 나는 이러한 해체와 재조직의 순환이 종화 과정의 핵심이라고 생각한 다 .(1975: 88) 도태에 의한 전형적 계통 발생의 점진론과는 달리 이 종의 기원의 도약설은 무작위적 점돌연변이의 조절 기전이므로 적응성이 약하다. 방대하고 신속한 유전적 변화의 도약으로 생식적 격리가 우선 일어나 나 이는 적옹성이 아니다. 종의 도약적 발생은 도태될 수 있는 원료를 제공하기는 하지만 이들 두 가지 양자택일적인 종화의 가설에는 상반 된 차이가 있다고 굴드 (1982) 는 말하였다. 계통 발생의 점진성에 따르
면 점돌연변이는 국소 집단 내 대립 유전자 치환을 초래하고 이는 도 태에 의하여 순차적으로 느리게 적응한다 . 한시적 평형설에 따르면 새 로운 종의 도약적 기원은 비연속성이고 비적응성이며 단지 이차적인 도태의 대상일 뿐이다. 역 설 적으로 < 바람직한 괴물 > 이라고 하는 극단적 성질의 도약적 발 생이 있을 수 있다. 도태는 < 바람직한 괴물 > 의 생존 여부를 결정할 것 이댜 그러나 이것의 발생과 방향에 대한 일차적인 압력은 유전된 개체 발생에 있는 것이지 도태 모델에 있는 것이 아니다 (Gould 1982: 142). 새로운 종의 번식 제한 조건은 유인류의 진화과정을 이해하는 데 중요하다 예를 들면 변두리 격리 집단에서 새로운 종이 발생했을 때 이는 조상 집단으로의 역이주가 가능하나 유전자 흐름의 확산성에도 불구하고 그 정체를 유지한다. 종간의 불임성 때문에 이 유전자 흐름 은 비효과적이다 . 잘 알려진 예는 말과 당나귀이다. 이들 두 종의 유전 자 구성은 매우 유사하여 당나귀 정자는 말의 난자를 수정하여 노새라 는 잡종을 만든다. 혼합된 유전 성분은 힘센 동물을 만드는 개체발생 적 의미에서 대단히 효과적이다. 그러나 노새는 불임이다. 생식과정에 서 분리 된 DNA 가 유전자 사이 사이 에 존재하는 공간 s p ace DNA 를 비 롯한 DNA 의 차이 때문에 효과적으로 합쳐지지 못한다. 수놈 잡종의 생식세포는 고도의 비정상적 유전자 복합체를 갖는다. 따라서 기능을 할 수 없다 (S t ebb i ns 1982). 1.4 유인류 진화의 유전 기전(Whit e, 1978) 진화에 따른 변화의 유전 기전에 관한 이론적 검토는 다음 장들에 서 화석 기록에 근거한 유인류 진화에 대하여 살펴보게 될 때 대단히 도움이 될 것이다.
표 1.1 몇 동물종의 DNA 뉴클레오티드 서열 차이의 백분율 사람 / 침팬지 2.5 사람/기본 5.1 사람/녹색(구세계)원숭이 9.0 사람/카푸친(신세계)원숭이 15.8 사람/여우원숭이 42.0 유인류 진화에서 일부 핵심적 사건은 화석의 흔적을 남기지 않고 일 어난 것 같댜 예를 들면 유인류 진화에서 대단히 중요한 양각 보행을 가지고 수상 서식에서 지상 서식으로의 이행에는 골반골과 하지골의 큰 적응성 변화를 수반했다. 그러나 이행 과정을 보여주는 화석은 없다 (그림 3.9). 아직도 더 많은 완전한 화석 기록을 필요로 한다. 사람과 원숭이는 생화학적으로 놀랄 만큼 비슷하다. 인간과 영장류 의 유전적 유사성은 이중나선 DNA 를 짧은 길이의 외가닥 DNA 로 쪼 개어 측정할 수 있댜 사람의 외가닥 DNA 를 다른 사람의 외가닥 DNA 와 섞으면 재조합으로 완전한 이중나선을 재생시킨다. 다론 동물의 의 가닥 DNA 와 섞으면 DNA 서열의 근사성의 정도에 따라 정도의 차이 를 가지고 재조합을 이룬댜 표 1.1은 이 같은 방법으로 측정한 결과로 서 사람과 침팬지 사이에는 유전자의 2 .5%만이 차이가 있고 다른 영 장류와는 분류학적인 근거로 예상되는 것보다 더 큰 차이를 보인다. 표 1.1로 짐작할 수 있듯이 유전자의 전사와 복사로 만들어지는 단백 도 사람과 침팬지는 매우 유사하고 유인원과는 좀더 다르고 원숭이나 여우원숭이와는 더 많이 다르댜 사리치 S ari ch 와 크로닌 Cro ni n(1977) 은 진화 역사를 추정하기 위해 단백 차이를 이용하였다. 특히 유인류 진 화에서 사람과 침팬지의 선조 혈통이 분파한 시기를 추정하였다. 이른 바 분자 시계라는 것이 있는데 이에 따르면 유인 동물 계통이 유인류 와 폰지드로 갈라자는 데는 500 만 내지 1000 만 년이 걸렸다 . 유전학자들은 돌연변이의 효과가 평균적 무익이라는 것에 일반적으
로 동의한다. 단지 대단히 적은 부분만이 유익하여 진화의 재료가 된 댜 한 개체가 물려받은 유전자 중 1 만분의 1 이 생식과정에서 미세한 변화 가능성을 갖는다. 그 확률은 1 만분의 1 내지 25 만분의 l 이다. (Do bzhansky, 1960) 돌연변이가 무익한 것이라면 자연도태로 제거된다. 때로는 겸상 적혈구성 빈혈처럼 이중 효과가 있다. 빈혈 그 자체는 무 익하나 말라리아에 저항성을 갖는 이점도 있다. 고로 그 유전은 확고 할지 모르겠다 . 보다 문제가 되는 경우는 어린아이의 눈에 생기는 암 인 망막아세포종에서 생기는 일로서 생식세포 5 만 개 중 약 1 개가 가 지고 있는 유전자에 의한 것이다. 이 병은 치료되지 않으면 거의 전부 사망한다 그러므로 자연도태로 조절된다. 그러나 이제는 이 유전자를 가전 사람의 70% 가 적절한 치료로 어른이 될 때까지 살아 남아서 그 자식의 반에게 이 병을 전파한다. 그래서 윤리적 문제가 제기된다. 그 들에게 자식을 갖게 할 것인가? 이것은 우리가 인간 유전학에서 경험 하는 수많은 복잡한 문제들의 한 가지 예에 불과하다. 1.5 종의 진화적 기원에 관한 일반적 결론 엘드레지와 굴드 (1972) 는 지질학적 시간 속에서 진화의 역할을 정체 와 도약적 단속성으로 설명하였다. 점진적 변화는 종의 정상상태가 아 니댜 어떤 종의 커다란 중심 집단은 근소한 적응성 변이를 나타낼 수 있댜 그러나 이것은 유동적인 성질의 것으로서 골드슈미트 (Goldsch rnidt, 1940) 는 이를 종내의 분파된 막다른 골목이라 하였다. 중심 집단 내 집중적 유전자 흐름의 항상성 영향은 이들 초기 종화를 조절할 것 이다 굴드 (1982) 는 종화는 거대 진화의 기초이고 대단히 빠른 가지치 기 과정 clado g enes i s 이 라고 주장하였다. 이 기 간은 수천년으로 종의 지 속 기간이 수백만 년인 것에 비하여 빠른 것이다. 그는 또 유전적 변
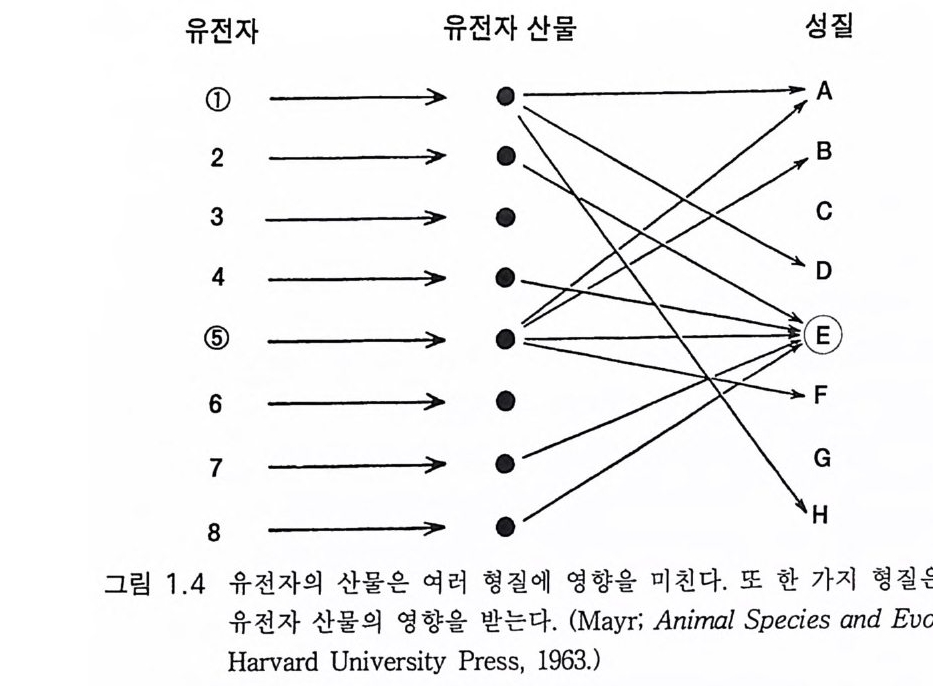 유전자 유전자산물 성 질
유전자 유전자산물 성 질
이는 도태의 재료 역할을 하고 도태가 진화의 방향을 근본적으로 조절 한다는 계통 발생 점진성의 환원법적 개념으로 간과되어 왔던 유기체 의 개념을 한시적 평형설이 생물학적인 것으로 재정립하는 것이라고 주장하였댜 한시적 평형설에서는 어떤 도태 조절 이전에 도약적 변화 가 일어나고 새로운 종이 완전하게 형성된 이후에 도태 조절기전이 작 용한다. 개체 발생이 도태에 선행한다. 유기체들은 자기들의 운명에 홍 미롭고 복잡하며 이해가 가능한 방법으로 영향을 미친다. 우리는 이 유 기체의 개념을 진화생물학으로 되돌려야 한다 .(Gould 1982: 144) 유기체의 개념은 유인류 진화에서 발생하는 여러 가지 문제에서 가 장 중요한 것이 될 것이다. 그림 1. 4 는 유기체를 구성하는 유전자들의 복잡한 관계를 일반화한 것이다. 유전자들은 그림 1. 2b 에서처럼 유전 자 산물인 단백을 구성하고 이들 단백은 대개 효소들인데 대단히 복잡 한 개체 발생 과정을 통하여 독특한 유기체의 모양을 만든다.
제 2 장 인간 전화의 일반론 이 책의 주제는 포유류의 뇌와 인간의 뇌에 관한 것이므로 영장류 뇌의 진화에 관하여 중점적으로 다루고자 한다. 영장류 등급의 최초의 화석 은 80 my B P(rn illion ye ars befo re pre sent 백 만년 전)의 치 아이 다. (Ste b bin s , 1982) 작은 영 장류는 65-40 m y BP 에 수상 서 식 동물로 알려 졌댜 이 원시 영장류의 현대 후손은 나무들쥐tr ee-shrews 일 것이다. 2.1 유인류 조상 (Tob i as 1975a, Sim o ns 1981, Copp en s 1983) 우리의 관심은 현대의 대유인원이 속하는 원인동물초과와 폰지드과 의 우리들 조상의 화석 역사에 초점을 맞출 것이다(표 2.1 ). 30 내지 35 m y BP 에 이집트 사막의 파이움 평원에 수상 서식 유인동물 화석이 있 었댜 주목할 것은 이집토피테쿠스 Aeg ypt op it hecus 의 화석으로서 이는 현대 폰지드와 유인류의 공통의 조상이 될 수 있을 · 것이다. 당시에 파 이움은 우거진 열대 밀림이었고 습지였다.(Pil beam lr.J /2 , Sim o ns 1983, 1985)
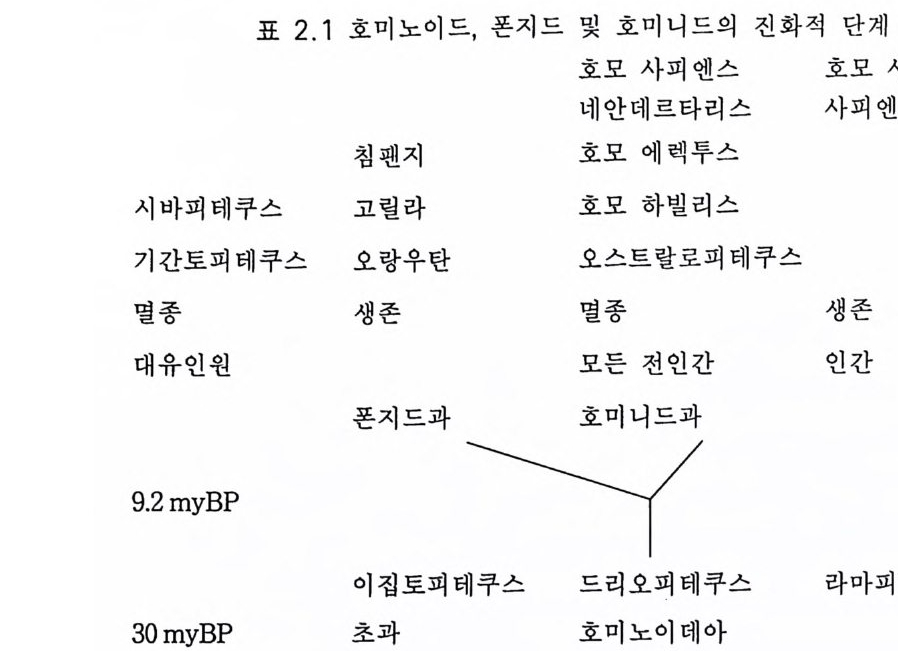 표 2.1 호미노이드, 폰지드 및 호미니드의 진화적 단계
표 2.1 호미노이드, 폰지드 및 호미니드의 진화적 단계
유인동물초과의 드리오피테쿠스 D ry op ith ecus 는 초기 일반적 유인동 물에 붙여진 이름으로서 표 2 .1처럼 폰지드와 유인류 두 과의 · 선조였 을 것 이 다 .(P il beam 1972, 198. 5, Sim o ns 1972, 1977) 드리 오피 테 쿠스는 유 인원이었고 약 30 내지 12m y BP 에 걸쳐 넓은 지역에 서식했을 것이다. 주목할 것은 케냐에서 발견된 프로콘솔 Proconsul 이라고 하는 16my B P 경의 두 개의 두개골로서 이는 케냐피테쿠스 아프리카누스 Ken y ap i the cus afri canus 와 연 계 된 다. (Pic k for d 198. 5) 드리오피테쿠스는 헝가리, 그리스, 터키, 인도, 케냐 등 대단히 광범 한 지역에 분포하고 있음에도 불구하고 유인류 진화의 다음 단계인 오 스트랄로피 테 신 Aus tra lopith e 아 nes 과 호모 하빌 리 스 Homo habil is 양자 모두는 아프리카에 국한하였다(그림 2.1). 따라서 왜 아프리카의 드리오 피테신만이 호모로의 진화에 참여하게 되었는가라는 의문이 제기된다. 엘드레지와 굴드 (1972) 가 한시적 평형설(1. 3) 에서 주장했듯이 오스트랄
 하다
하다
로피테신의 기원이 일관된 진화적 변환을 대표했을 것이라고 믿는다. 따라서 아마도 이는 적은 격리된 집단에서 일관되었을 것이다. 나머지 드리오피테신은 결국 소멸하였다(표 2.1 ). 최근까지 라마피테쿠스속은 그것의 턱뼈와 치아의 구조에 따라 호 미니드과에 두었다(그림 2.2c). 그러나 라마피테쿠스는 보통 12 내지 14 my B P 시대의 것으로 이는 lOmy B P 이내의 유인동물 계통이 분파 하기 이전이며, 최근에 중국 루팽에서 두개골이 발견되었는데 이는 유 인동물인 것으로 밝혀졌다 (Wu 1984). 불행히도 8 내지 4m y BP 의 화석 결손이 있는 것 이 다 (Tob i as 1975a, Sim o ns 1981, Cop pen s 1983). 가장 풍
표 2.2 인간 조상의 화석 기록 종 또는 속 출현시기 멸종시기 평균두개 기타 특징 (년전) (년전) 용적 (cc) 호모 사피엔스 현대 인종 30,000 현존 1330 막달레니아, 오리나시 크로마뇽 아 및 현대 문명 네안데르탈 100,000 30,000 1470 무스테리아 문화, 최초의 의식과 예식 고대 300,000 200,000 1300 후기 아슐 문화 호모 에렉투스 1,300,000 400,000 950 아슐 문화 최초로 불 사용 호모 하빌리스 2,000,000 1,600,000 700 올두바이 문화, 최초로 마제석기 사용 오스트랄로피테쿠스 3,000,000 2,000,000 550 직립보행, 알려진 도구 로부스투스 없음, 채식성(?) 원시 형태 아님 오스트랄로피테쿠스 3,000,000 2,000,000 500 직립보행, 알려진 도구 아프리카누스 없음, 잡식성(?) 원시 형태일 가능성 오스트랄로피테쿠스 3,500,000 2,500,000 425? 직립보행 아파렌시스 원시 형태일 것임 라마피테쿠스 14,000,000 10,000,000 ? 네발로 걸었음, 치아는 호미니드형에 접근 드리오피테신스 22,000,000 12,000,000 ? 뼈마디 걸음(?) 유인원과 사람의 공동 조상일 것임
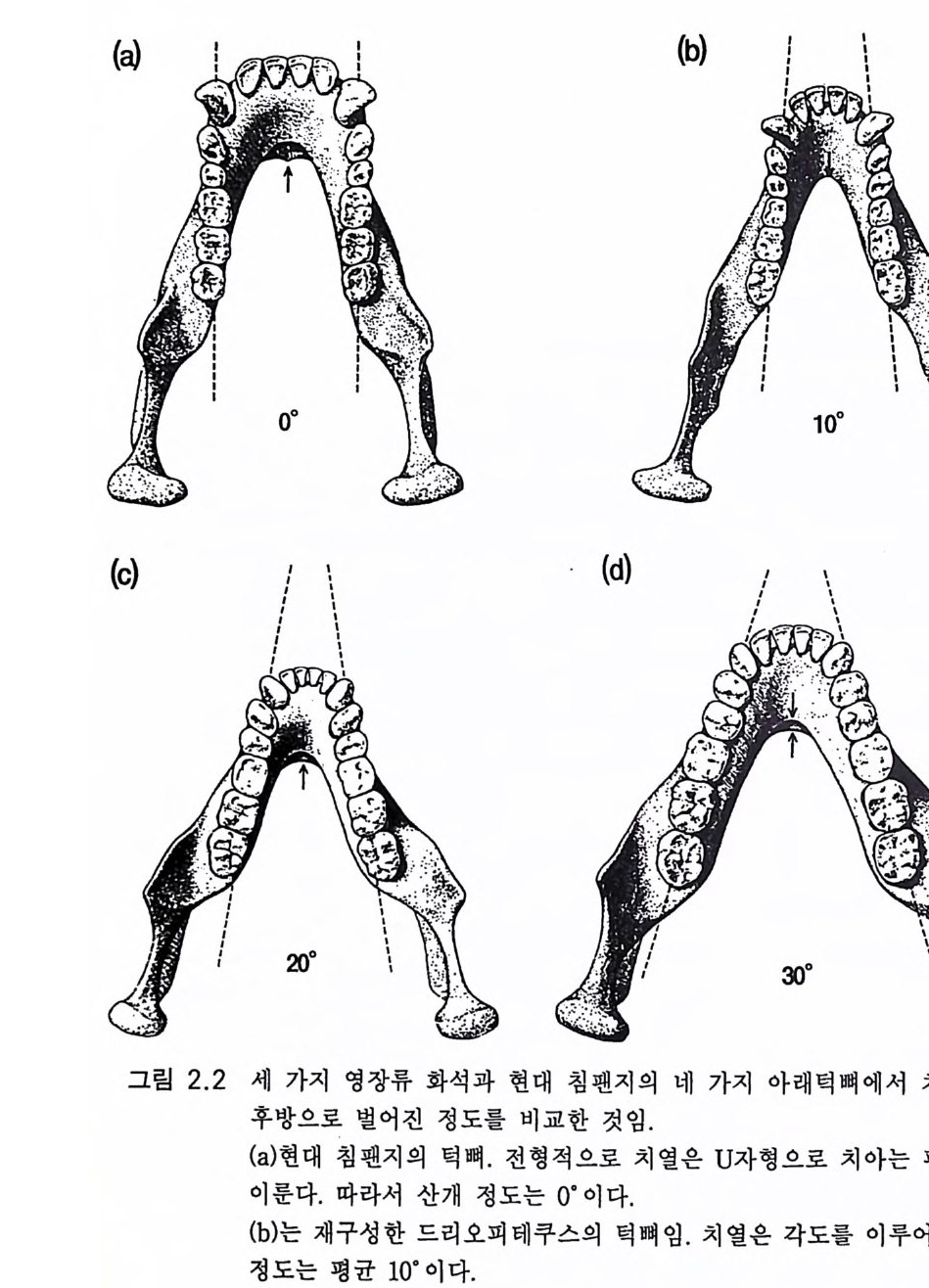
부한 화석 기록은 우리의 진화 역사에 결정적인 시기의 것이 절대적이 댜 즉 호미 니 드와 폰지 드가 분파하는 시 기 이 다(표 2.1 ). 표 2.2 는 호미 니드 진화의 대강을 간추린 것이다. 호미노이드와 호미니드의 턱뼈, 특히 치아는 잘 보존된 화석이다. 치아를 식별하고 기술하는 고도의 복잡한 기술이 있다. 때로는 호미니 드의 남아 있는 유일한 화석은 치아이다. 여기서 턱뼈와 치아의 진화 상의 의의만을 그리는 것이 가능하다. 그림 2.2a 에서 침팬지의 턱뼈는 큰 견치까지의 양측 치열이 평행임을 나타내고 있다. 유인동물 드리오 피테쿠스의 치열은 10· 의 각도를 보이고 견치는 역시 크다(그림 2.2b). 라마피테쿠스의 턱뼈는 20· 의 각도를 이루어(그림 2.2c) 30° 의 각도를 이루는 오스트랄로피테쿠스에 가까워진다(그림 2.2d). 라마피테쿠스의 견치는 작아지고 오스트랄로피테쿠스의 견치는 더 작아져서 호모 사 피엔스와 큰 차이가 없다 . 아름답게 구부러진 사람의 턱뼈 치열은 그림 2.2a 의 침팬지의 치열 에 합치할 정도로 평행을 이루고 있는 오랑우탄의 그것과 대조를 이룬 댜 턱뼈와 치아는 호미니드 진화의 여러 단계를 나타낸다(그림 2.3, 2.4 , 2.8, 2.1 0 ). 여 러 유인동물들은 유인류의 특징 적 인 모양을 각기 다른 정도로 갖추면서 유인류화되었다 . 유인류화는 단일한 공통 진보 방향 울 따른 것이 아니라고 토비아스 (1975a) 는 말하였다. 제 1 장에서 기술한 바와 같이 사리치 S ari ch 와 크로닌 Cro nin (l
 @ (b)
@ (b)
2,2 오스트랄로피테신 화석 기록은 호미니드 진화가 동아프리카(케냐, 탄자니아, 에티오피 아) 그리고 남아프리카(트란스발) 둥 아프리카에 국한함을 나타낸다 . 그곳에는 400 개체 이상의 골격이 남아 있다 (Tob i as 1981a, 1983). 아프 리카에서의 최초의 발견은 1925 년 다트 D art에 의한 것으로 남아프리카 타옹에서 발견한 놀랍도록 잘 · 보존된 두개골이다. 그림 2.3 은 이 소년 의 두개골인데 두개강 용적은 500cc 이하이다 .(Tob i as lITTl) 이 정도의 크기는 작은 체격을 감안한다면 현대의 유인원의 것보다 약간 큰 것이 댜 그리고 오스트랄로피테신 Aus tr alo pit hec i nes 의 골격은 양각 보행이 었음을 나타낸다 (3.3). 토비아스 (1983) 는 작은 성간의 이형성을 없애기 위하여 많은 수의 여성 및 남성 뇌 크기의 평균치를 다음과 같이 구하 였댜 1 ) 침팬지 394cc 고릴라 506cc 오랑우탄 411cc (c) 는 라마피테쿠스의 턱뼈를 재구성한 것이다. 치열의 산개 정도는 평균 20· 이다. (d) 재구성한 오스트랄로피테쿠스의 턱뼈. 전형적 치열의 산개 정도는 30° 이다. 후기 호미니드의 치열은 더 큰 산개 각도를 보이기도 한다. 화 살표는 상하 융기로 알려진 두 개의 턱뼈 융기 버팀벽의 차이룰 가리키 는 것이다. 현대 유인원은 크고 선반 모양의 하융기를 가졌고 드리오피 테쿠스는 상융기가 월등하다 . 라마피테쿠스와 오스트랄로피테쿠스에서 는 양융기 가 모두 발달되 었다.(E. L. Sim o ns; Ramapi the cus, Scie n ti fic Ameri ca n, May 1977)
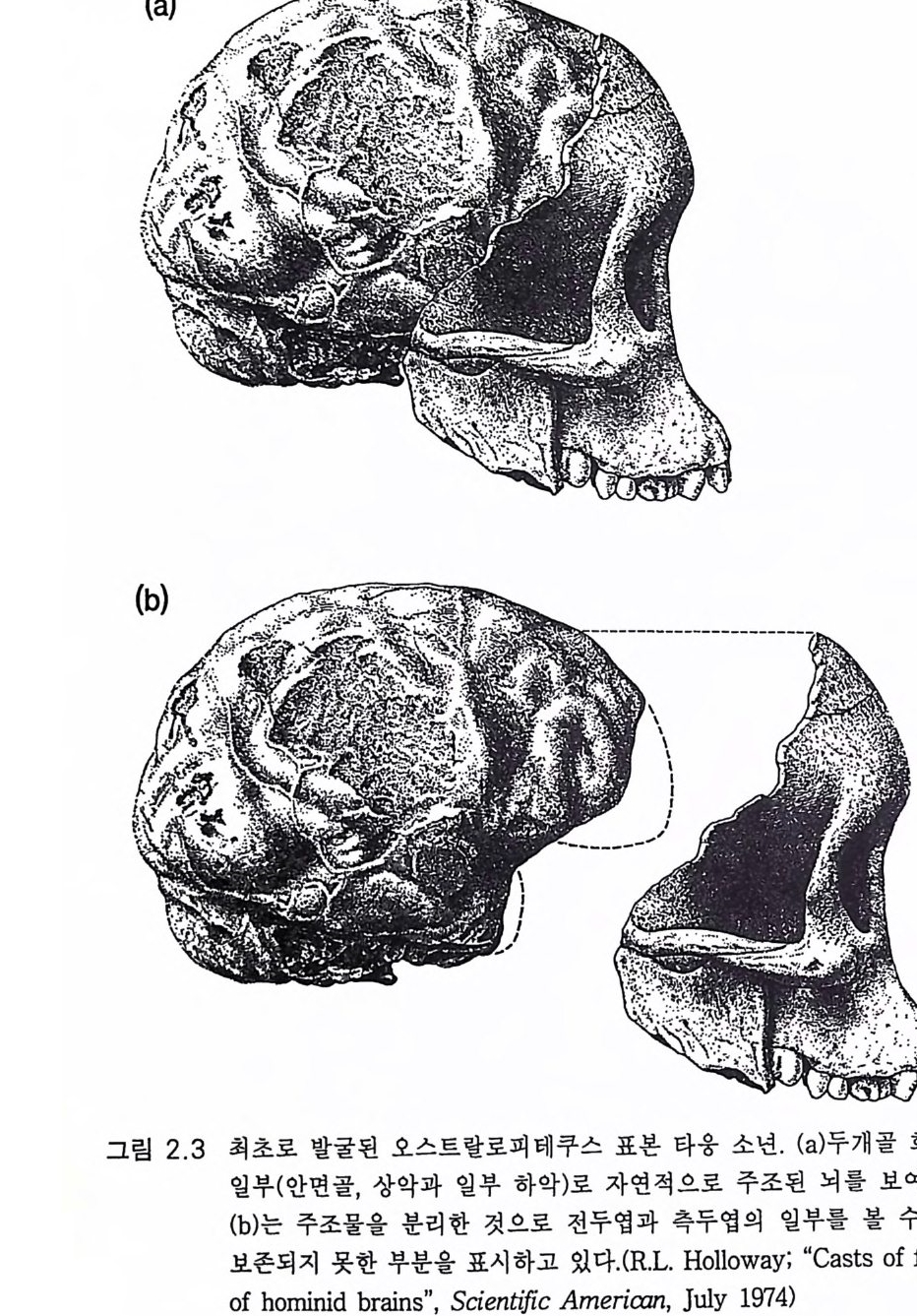 그림 2.3 최초로 발굴된 오스트랄로피테쿠스 표본 타옹 소년 . (a) 두개골 화석의
그림 2.3 최초로 발굴된 오스트랄로피테쿠스 표본 타옹 소년 . (a) 두개골 화석의
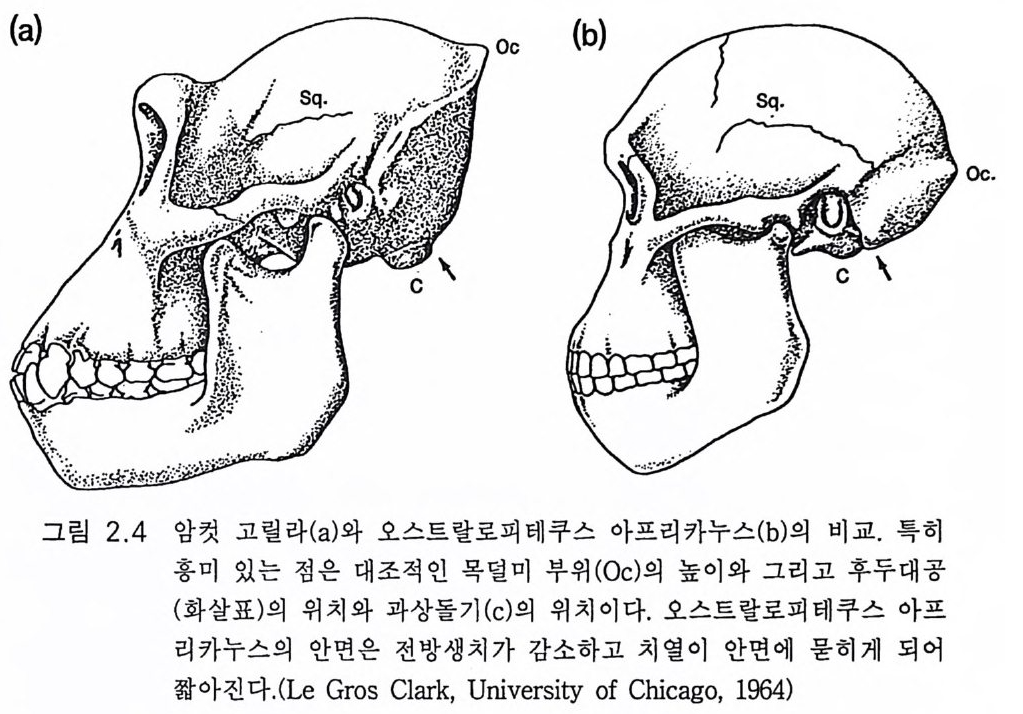 (a) Oc (b)
(a) Oc (b)
이어서 남아프리카(그림 2 .1의 지도)에서 발견된 기타 호미니드는 뇌 용 적이 425-480cc 였고 여섯 개의 호미니드 뇌 용적 평균치는 441cc 였 댜 오스트랄로피 테 쿠스 아프리 카누스 Aus tra lopit hecus afri canus 라는 이 름은 다트 D art가 만든 것 이 다 (Tob i as 1971, 1981a, 1983). 동아프리 카 에서는 오스트랄로피테쿠스 아프리카누스로 분류된 작은뇌 양각 호미 니드가 발견되었다. 홀로웨이(H ollowa y 1983) 는 동아프리카에서 발견 된 여섯 개의 뇌 크기를 잠정적으로 400-582cc 로 보고 평균치는 445cc 라 하였다. 그림 2 .4는 유인동물의 모델로 취한 고릴라로부터의 굉장한 변화를 보이는 것으로 이것은 최초 호미니드 오스트랄로피테쿠스 아프리카누 스로 진화하였다 . 뇌의 크기는 비슷하지만 얼굴과 턱은 크게 다르다.
치아는 크기가 작아지고 견치도 많이 작아졌다. 오스트랄로피테쿠스 아프리카누스에서 이미 호미니드 진화 계통을 따라 변화가 있어 왔던 것이다. 불행히도 동아프리카와 남아프리카에서 발견된 것들간의 관계에 관 한 논쟁이 제기되었댜 탄자니아의 레톨리에서(그림 2.1 ) 1960 년대에 약 3.8m y BP 의 일련의 호미니드 화석이 발견되었댜 후에 1970 년대에는 에티오피아의 하다에서 많은 화석이 발견되었다. 이 두개골들을 재구 성하여 뇌의 크기가 남아프리카에서 발견된 오스트랄로피테쿠스 아프 리카누스의 것과 비슷하다는 증거가 제시되었다. 골격 화석은 모두가 양각 동물이었다. 하다 화석의 연대는 3 .1 -2.6m y BP 였고 이는 실제로 남아프리카의 3.0-2.5m y BP 와 같은 것이다 .(Tob i as 1981a) 요한손J ohanson 과 화이트 W 血 e(1979) 는 교묘한 분류학적 방법으로 하다와 레톨리에 널리 산재 하는 화석들을 연관지어 에티오피아의 아파라는 지명을 따라 오스트 랄로피테쿠스 아파렌시스라는 별개의 종으로 명명하였다. 레톨리 호미 니드는 하다로부터 1600km 에 위치할 뿐 아니라 그들은 약 60 만 년이 나 일찍이 그곳에 살았다. 이 방법으로 레톨리, 오모, 쿠비에서 발견되 었던 오스트랄로피테쿠스 아파렌시스에서 호모 하빌리스가 되었을 것 으로 생각되는 호미니드 진화의 본류로부터 트란스발과 오모에 있는 오스트랄로피테쿠스 아프리카누스를 분리하게 되었다(J ohanson 과 White 1979). 종화의 기준으로 하다와 레톨리롤 합친 화석은 트란스발의 오스트 랄로피테쿠스 아프리카누스와 구별이 안 된다는 점을 들어 토비아스 (1981a) 와 다른 사람들은 오스트랄로피테쿠스 아파렌시스라는 신종의 창조를 크게 비판하고 있다. 요한손 (1985) 은 오스트랄로피테쿠스라는 신종의 창조에 대한 비판에 몇 가지 해답을 주려고 하였다. 그는 이를 지지하는 22 가지 원시 형태적 형질을 열거하였다. 그러나 특정 종이
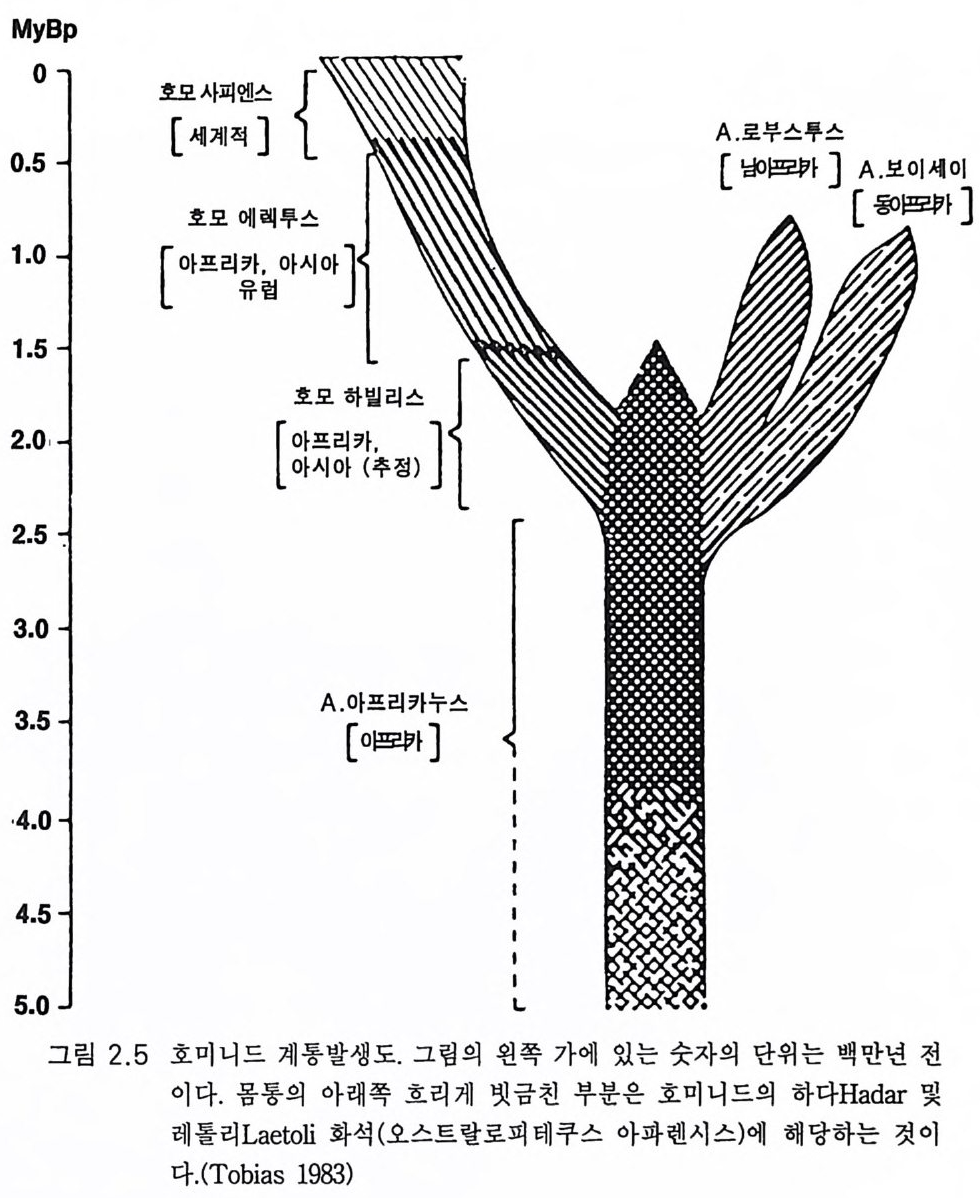 My Bp
My Bp
넓은 지역에 산재하였던 지역성 및 시간성을 고려할 때 이들 형질은 근소한 정량적 또는 확률적 개체 차에 지나지 않는 것으로 보인다 (Tobia s 1981a). 이 책에서는 종의 다양성을 그림 2.5 에서처럼 오스트랄로피테쿠스 아프리카누스를 넓은 지리적 뿌리로서 나타내는 것으로 족하다고 생
각한다 수백만 년에 걸친 대단히 긴 오스트랄로피테쿠스 아프리카누 스의 시대에는 변종, 아종이 있었을 것이다. 토비아스(1 983) 는 이들을 지명에 따라 다음과 같이 제안하였다. 오스트랄로피테쿠스 아프리카누스 트란스발렌시스 (A. africa nus tran svaalensis ) 오스트랄로피테쿠스 아프리카누스 탄자니엔시스 (A. africa nus tanz anie n sis ) 오스트랄로피테쿠스 아프리카누스 에티오피쿠스 (A. africa nus aeth i o p i cu s) 오스트랄로피테쿠스 아프리카누스종은 지리적 재구성 시기에 교잡 과 격리 기간 동안에 아종으로 진화하여 그물 모양의 성질을 갖게 되 었을 것이다 . 그래서 유전자 흐름을 통하여 종의 원상은 실제적인 정 적 상태를 유지하게 하였을 것이다. 그림 2 .5에서 보여주듯이 약 2.5 m y BP 에 분지 발생의 격변이 있었다. 오스트랄로피테신은 직립 자세와 양각 보행에 의해 그 조상 유인동 물과 구별된다(제 3 장) . 둘째 구별점은 작은 견치를 지닌 치열이고 셋째 는 가장 중요한 점으로 추정된 체중에 비하여 뇌가 조금 커진 것이다 (그림 2.6). 직립 자세와 양각 보행은 3.3 에서 자세하게 다룰 것이다 . 치 열에 관하여는 2 .1에서 언급하였다. 발굴 당시에는 조각난 상태인 두개골을 재구성하는 정교한 기술이 있었다(그림 2.7 및 2.9). 그리고 플라스틱 물질을 이용하여 두개골을 채 우고 있었던 뇌의 모형을 만드는 특수한 내부 주조 기술이 개발되었다 (Holloway 1974, 1983). 그러나 그림 2.3b 는 타옹 두개골 앞쪽에 붙어 있 던 자연적 내부 주조물이다. 발굴 당시 두개골 내부는 석회화된 모래 로 채워져 있었다. 오스트랄로피테신의 내부 주조물을 현대 유인원의 두개골 내부 주조물과 비교하면 뇌의 모양이나 크기가 별로 다르지 않
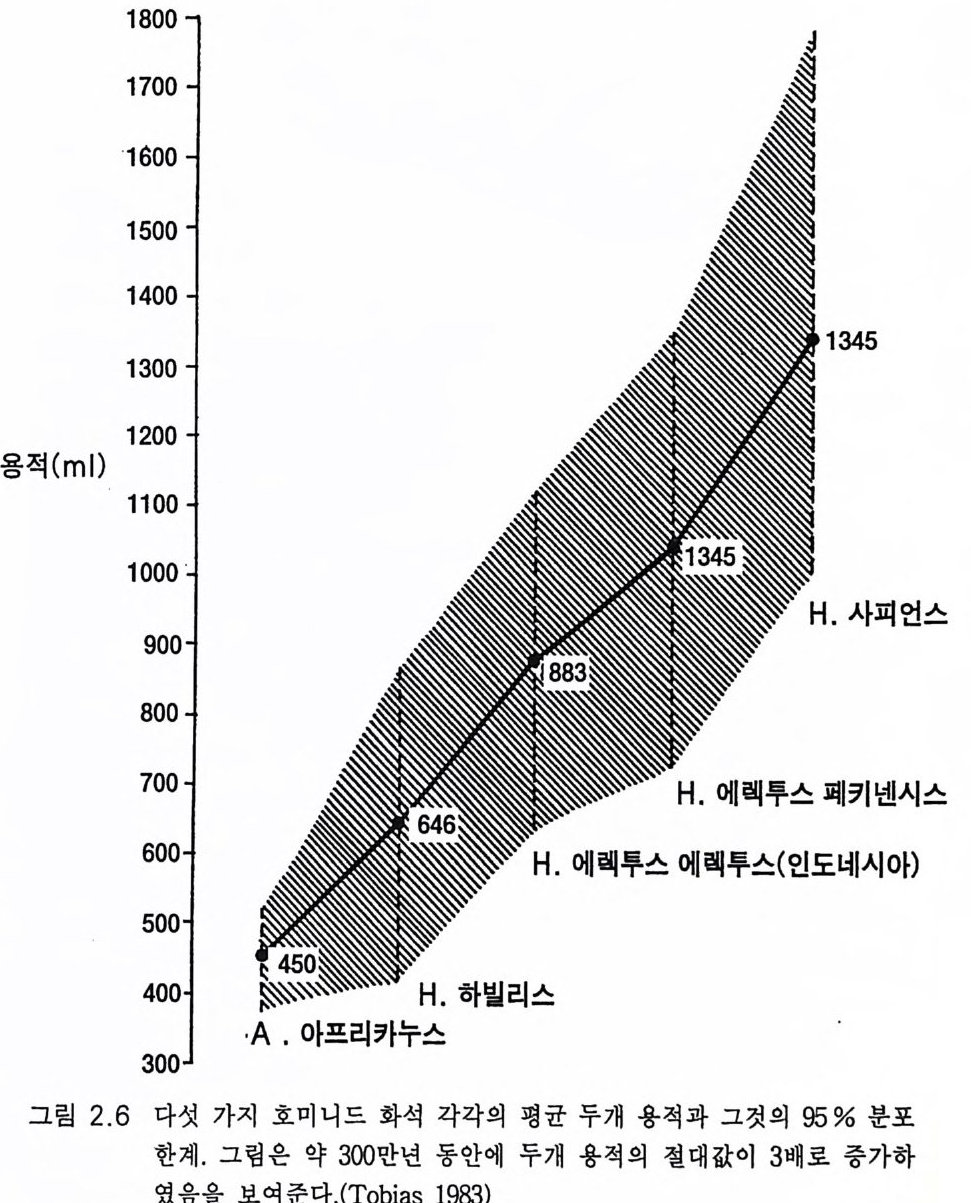 1800
1800
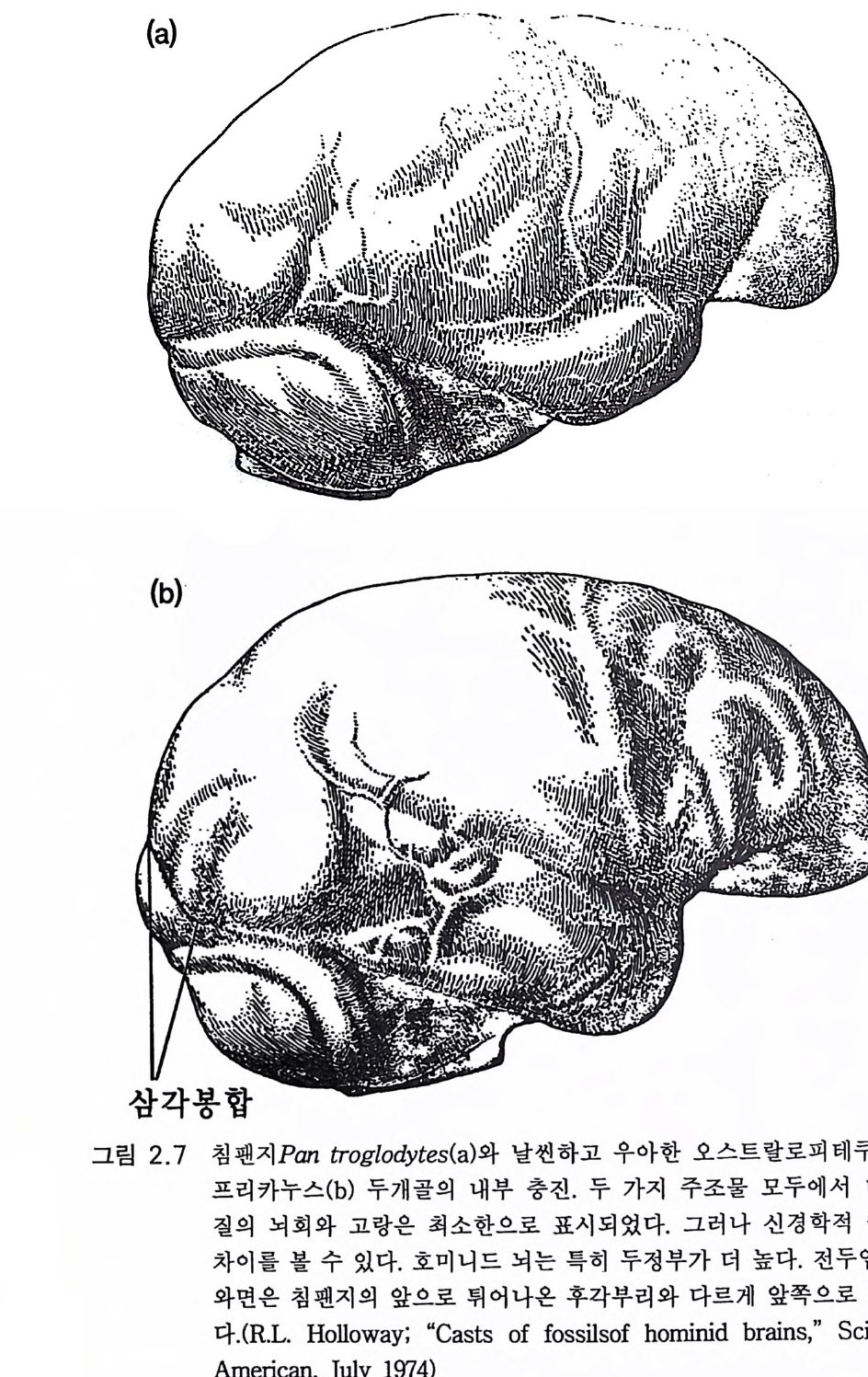 (a) ----’· -. ~. . •., '、. .. `_` • • _ —-··. •.. `` ..`
(a) ----’· -. ~. . •., '、. .. `_` • • _ —-··. •.. `` ..`
다(그림 2.7). 대체로 사람의 언어 영역의 대부분인 하전엽이 약간 발달 되 어 있고 상측두엽 이 약간 커 져 있다 (Tob i as 1983, Holloway 1983). 오스트랄로피테쿠스는 200 만년 이상 존재하면서 대부분 점진적으로 진화하였다(그림 2.5 ). 그러고 나서 2.5m y BP 에 도약적 분지분파가 있 었다 (Tob i as 1983). 한편 오스트랄로피테신속에는 남아프리카의 오스 트랄로피테쿠스 로부스투스와 동아프리카의 오스트랄로피테쿠스 보이 세이의 두 신종이 있었다. 이들 양자는 좀더 건장한 체격을 가졌고 뇌 용적이 조금 컸다. 이 같은 발달은 이들에게 진화적 이점이 되었을 것 으로 추측된다 그럼에도 불구하고 양자는 약 lm y BP 에 소멸된다(그림 2.5). 본래의 오스트랄로피테쿠스 아프리카누스도 분파분지 이후 오래 지속하지 못하고 사실상 폭발적 방사에 의해 새로운 종으로 귀착되는 데 이 새로운 종은 너무도 달라져서 우리가 지금 속하는 새로운 속, 죽 호모속의 첫 구성원의 지위를 부여받았다. 그러면 오스트랄로피테쿠스 아프리카누스의 과거력을 어떻게 말할 것인가? 양각 보행이라는 그리고 지상 생활의 모든 이점과 위험에서 초기의 큰 성공을 이룬 후 진화적 정체로 빠져들었다. 그러나 그 작은 뇌를 가지고도 생존에 충분하였다. 오스트랄로피테쿠스 아프리카누스 만이 아프리카의 생물계 변천 속에서 호미니드 진화계를 이루었다. 이 것이 더 일찍 소멸되었다면 호미니드 진화는 끝장일 수도 있었다. 그 러나 이 종은 지속되어서 큰 뇌의 호모 하빌리스의 시조로서 유전적 진화의 여명기를 맞기에 충분했다. 우리의 상징적 상상력으로도 오스 트랄로피테신은 호미니드의 이어달리기 첫번째 주자로 간주할 수 있 고 그들의 정확한 유전자를 운반하는 일관된 역할을 수행하여 이 탁월 한 호미니드 이어달리기에 참여한 다음 주자 호모 하빌리스에게 배턴 울 넘겨주었다.
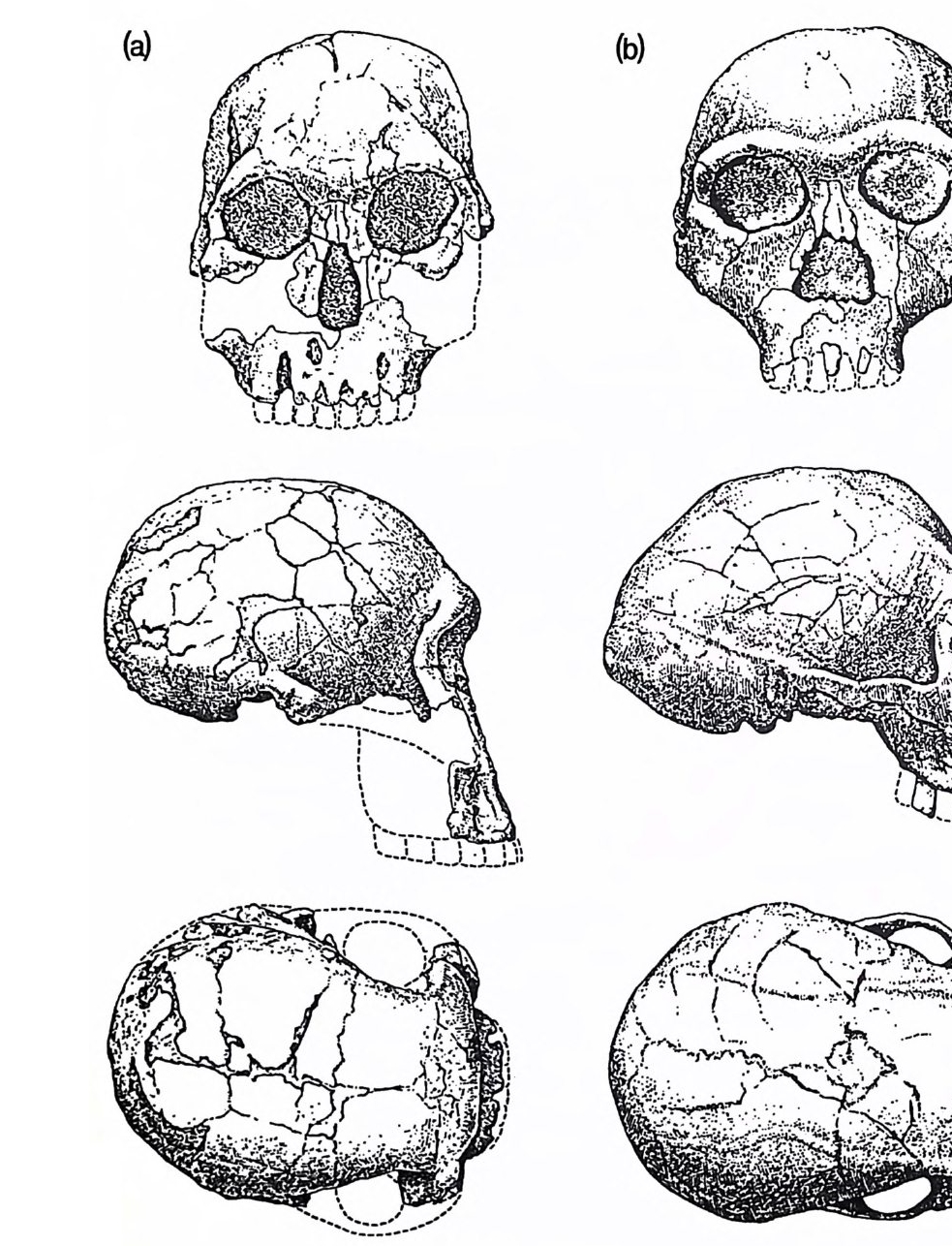 固 (b)
固 (b)
그림 2.8 (a) 호미니드 화석, KNM-ER 1470 이 두개골은 쿠비 포라 형성의 하 위 구성원에서 온 것이다. 160 만년 이상 되었음은 확실하며 아마도 250 만 년 이상 되었을 것이다 . 두개 용적은 약 775cc 이다 . (b) 호모속의 두개골로서 동투르카나 화석충에서 나온 것이며 초기 인종인 호모 에 렉투스를 나타내는 것이다. 자바와 중국에서 처음 발견된 호미니드 화 석 이 다 KNM-ER 3733 은 150 만 년 이 상된 것 이 고 중국에 서 발견된 것 보다 100 만년 이상 오래된 것이다. 대연대는 호모 에렉투스가 아프리 카에서 처음 발생하였음을 강력하게 시사하는 것이다. KNM-ER '373 3 의 두개용적은 약 850cc 이다 .(Walker 와 Leakey l'J7 8 )
2,3 호모 하빌 리 스 (Tob i as, 1987) 남아프리카와 동아프리카에서 1950 년대에 괄목할 만한 일련의 발견 이 있었는데 그곳에서 오스트랄로피테신으로부터 진화과정이 계속 전 진하였음을 보여주었다. 이 진화 발전의 첫번째 증거는 남아프리카에 서 발견되는 치아 화석과 석기이다. 그러고 나서 1960 년대에 리키 Leakey 가족과 그 일행이 탄자니아의 올두바이와 케냐의 투르카나 호 수(그림 2 .1)에서 호미니드 화석을 발견하였는데 이는 뇌의 크기가 진 화적으로 크게 발전하였음을 보여주었고(그림 2.6, 2.8 a ) 그에 상응하는 석기문화의 발달을 나타내어 올두바이 문화라고 불렸다(그림 6.l Oa ).21 오스트랄로피테쿠스 아프리카누스의 뇌 크기 평균값이 450cc 에서 646cc 로 44% 나 크게 증가하였으므로(그림 2.6 ) 이들 호미니드는 호모 속으로 간주되었다 .(Ma yr , 1973) 하빌리스 hab i li s 라는 종명은 석기문화 가 시작되었으므로 솜씨 hand y라는 뜻에서 다트 D art가 명명한 것이다. 체구는 현저한 증가가 없었으므로 뇌 크기의 증가는 괄목할 만한 발전 이댜 호모 하빌리스의 두개골 내부 주조(그림 2 . 8a) 에 따르면 뇌 크기의 증가는 일관된 것은 아니 었다 (Tob i as 1983, Holloway 1983). 부로카 영 역이 있는 하전두엽이 더 발달하였다. 그러나 가장 두드러진 것은 하
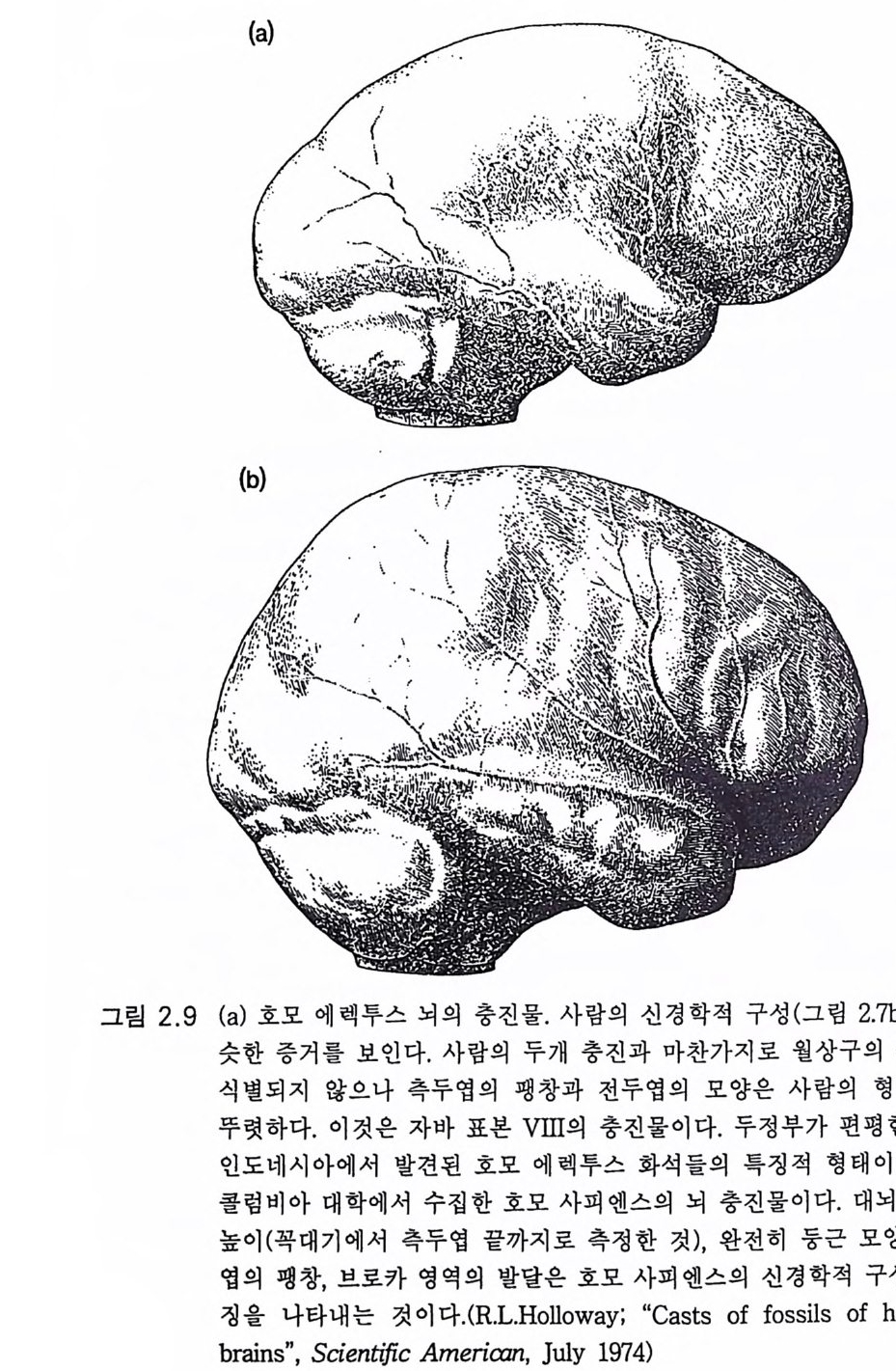
측두엽의 둥근 충만성이댜 이것은 베르니케의 중요한 언어 영역에 해 당하는 부분이 다(제 4 장 그림 4.3 ) . 상측두엽 의 발달도 두드러 졌다. 토비 아스 (1986) 는 다음과 같이 말하였다. 호모 하빌리스의 모양은 인간 진화에 새로운 시기의 시작을 점한다. 죽 큰 뇌, 기구 연마, 문화 의존적 호미니드 발달이다 . 호미니드화의 과정 은 오스트랄로피테쿠스가 현대 인간을 향하여 불연속적인 정량 도약적 전진을 이룩함으로써 이루어졌다. 호모 하빌리스는 동아프리카와 남아프리카에 국한하였던 것 같다. 약 2.5my BP 의 오스트랄로피테쿠스 아프리카누스 뿌리로부터의 분파 분지에 있어 그 시조로부터 약 1. 6m y BP 까지는 큰 변화가 없었던 것 같다(그림 2.5 ). 그래서 90 만 년의 진화적 정체기에 도약적 변환이 있었 다. 이 다음의 커다란 진화는 호모 에렉투스 Homo erec t us 로의 발전이 었다(그림 2.5 및 2.6). 호모 하빌리스의 언어 영역 증가는 그 용도를 말하는 것이다. 호모 하빌리스의 각 개체는 의사 전달을 위한 효과적 언어를 개발하는 큰 창조적 발전을 이룩하였다고 할 수 있다. 제 4 장에서 상세하게 다룰 것 이다. 2.4 호모 에렉투스 두보아 Dubo i s 는 1890 년대에 놀라운 발견을 하였다. 죽 자바에서 비 교적 두껍고 용적이 약 850cc 가 되는 두개골 화석을 발견하였다. 이들 온 70 만년 이전의 것이었다. 후에 자바의 다른 곳에서도 발견되었다. 1920 년대에는 북경 근처에서 50 만년 내지 80 만년 전 시대의 더 큰 뇌
(평균 1040cc) 의 호미니드 화석이 발견되었다. 이는 호미니드 계통이 아시 아로 왔던 것 으로 생 각되 는 것 이 다 (P i lbeam 1972, Howells 1966). 그러나 보다 이론 시기인 1.5 my B P 때의 큰 뇌를 가진 호미니드 화석 이 아프리카에서 발견되면서 상황은 변환된다 (Walker 와 Leakey 1978). 아프리카에서 발견된 장소는 탄자니아의 올두바이와 케냐의 투르카나 호수이댜 이들 아프리카와 아시아에서 발견된 모든 화석은 두개골, 턱뼈 및 치아가 대단히 유사하였다 . 그림 2.8b 의 두개골은 투르카나 호 수에서 발견된 것으로 뇌의 크기는 850cc 이고 시기는 1. 5m y BP 였다. 그 내부 주형(그림 2.9a) 은 사람의 것(그림 2.9b) 과 대조를 이룬다. 발견한 것의 거의 대부분을 발견 장소에 따라 명명하므로 분류학상 의 혼동을 가져왔다. 그래서 뇌의 크기가 800 내지 1200cc 인 것과 0.5 내지 1.5 my B P 시대의 것을 한 군으로 하여 호모 에렉투스로 크게 단 순화하였다(H owalls 1966). 그리고 때에 따라 아종으로 그림 2.6 에서 처 럼 인도네시아의 것을 호모 에렉투스 에렉투스 Homo erect us erect us 그리고 호모 에렉투스 페키넨시스 Homo erectu s p e ki nens i s 로 하였다. 호모 에렉투스로 확인된 화석은 자바의 트리닐과 제티스, 중국의 란티 안, 유럽의 하이델베르크에서도 발견되었다. 그러므로 동아프리카에서 기시하는 약 1. 5m y BP 의 발견장소로 추정 한 시기를 고려하면 호모 에렉투스는 아프리카, 유럽 및 극동지방에 걸쳐 널리 이동하였다고 단순하게 가정할 수 있다 . 이는 호미니드의 첫번째 대이동이었다. 호모 에렉투스는 분명 대단히 성공적인 호미니 드이고 대모험가였다. 하지만 극동아시아로의 이동에는 50 만년이라는 대단히 장구한 세월이 있었음이 확실하다. 동아프리카와 유럽의 호모 에렉투스 화석은 아슐리안 Ache uli an 이라 고 하는 일관된 석기문화를 수반한다. 도구들은 깎아서 만든 손도끼, 깎는 도구, 칼날, 송곳이다(그림 6.l Ob )(Ste b bin s 1982). 이 석기문화의 발달은 루이 Lo ui s 박사와 마리 리키 M ary Leakey 박사의 탄자니아의
 (a)
(a)
올두바이 퇴 적 층의 발굴로 밝혀 졌다 .(Leake y와 Lewi n 1977) 호모 하빌 리스의 올두바이 도구가 최하위에 있었고 10 만년에 걸쳐 점차 개선된 도구가 네 층을 이루고 중첩되어 묻혀 있어 정교한 아슐리안 도구가 제일 위에 있었다(그림 6.1 0 b). 이는 특정 목적에 따르는 도구를 만든 복잡한 문화의 좋은 증거이다. 이 동아프리카 지방에는 10 만년 이상에 걸쳐 좀더 원시적인 호모 하빌리스가 독자적 석기문화를 가지고 호모 에렉투스와 공존하였다. 우리는 우리들 호미니드 조상의 보수성을 경 탄할 따름이 다 (Leake y와 Lewi n 1977). 2.5 네안데르탈 1856 년에 독일 라인랜드 지방의 네안더 골짝에서 사람 뼈의 화석이 발굴되었다. 화석은 호모의 원시형일 것으로 생각되었다. 비슷한 화석이 유럽의 다른 지방에서도 발견되므로 호모 사피엔스 Homo sap i ens 의 중 요한 조상의 증거로 인식되었다. 트린카우스 T rink aus 와 호웰스 Howells (1979) 는 네안데르탈 Neande rtal 진화에 관한 권위 있는 의견을 개진하였 다 . 최근 옐리넥J e li nek(1985) 은 풍부한 새로운 발견들을 통해, 그리고 새 로운 연대 측정 기술로 발전해 나가는 복잡한 이론들을 비판적으로 조 망해 발표하였다. 네안데르탈은 분명하고 대단히 중요한 호미니드 진화 의 실체를 이룬다. 이것을 바르게 표시하면 호모 사피엔스 네안데르탈 렌시스 Homo sap ien s neander ta lens i s 로서 HSN 으로 약해서 쓰고 분명히 HSS 즉 호모 사피엔스 사피엔스 Homo sap ien s sap i ens 와 구별된다. 후 자는 현존하는 인류를 포괄하는 분류학적 용어이다. 그림 2.1 0 c 와 2.10d 의 네안데르탈 두개골은 그림 2 .1 0a 와 2.10b 의 HSS 두개골과 비교하면 몇 가지 특징이 있다. 후자는 대단히 홍미 있 는 HSS 두개골의 예이다. 이돌은 대단히 초기의 HSS 이다. (b) 는 팔레
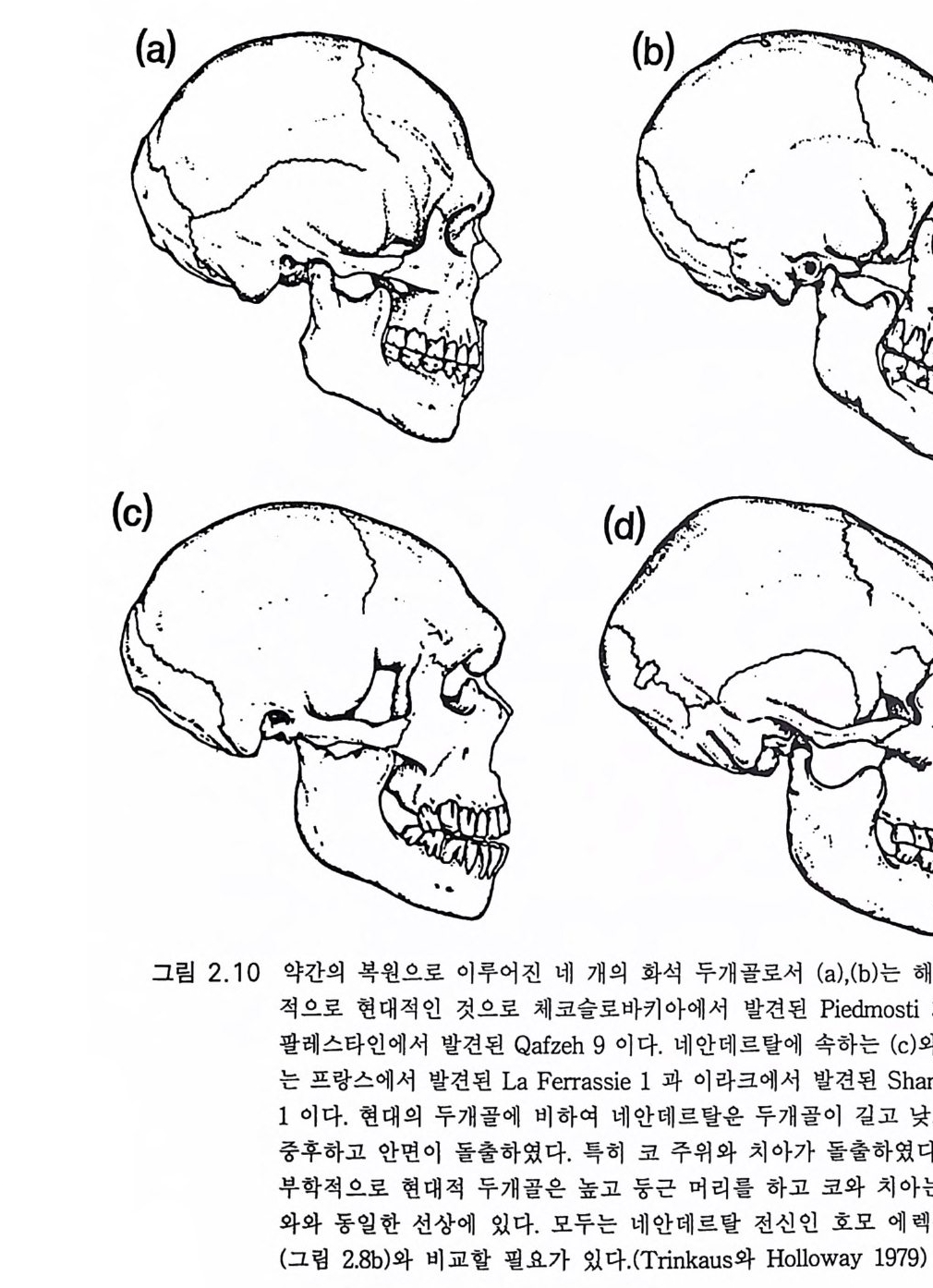 그림 2.10 약간의 복원으로 이루어진 네 개의 화석 두개골로서 (a) , (b) 는 해부학
그림 2.10 약간의 복원으로 이루어진 네 개의 화석 두개골로서 (a) , (b) 는 해부학
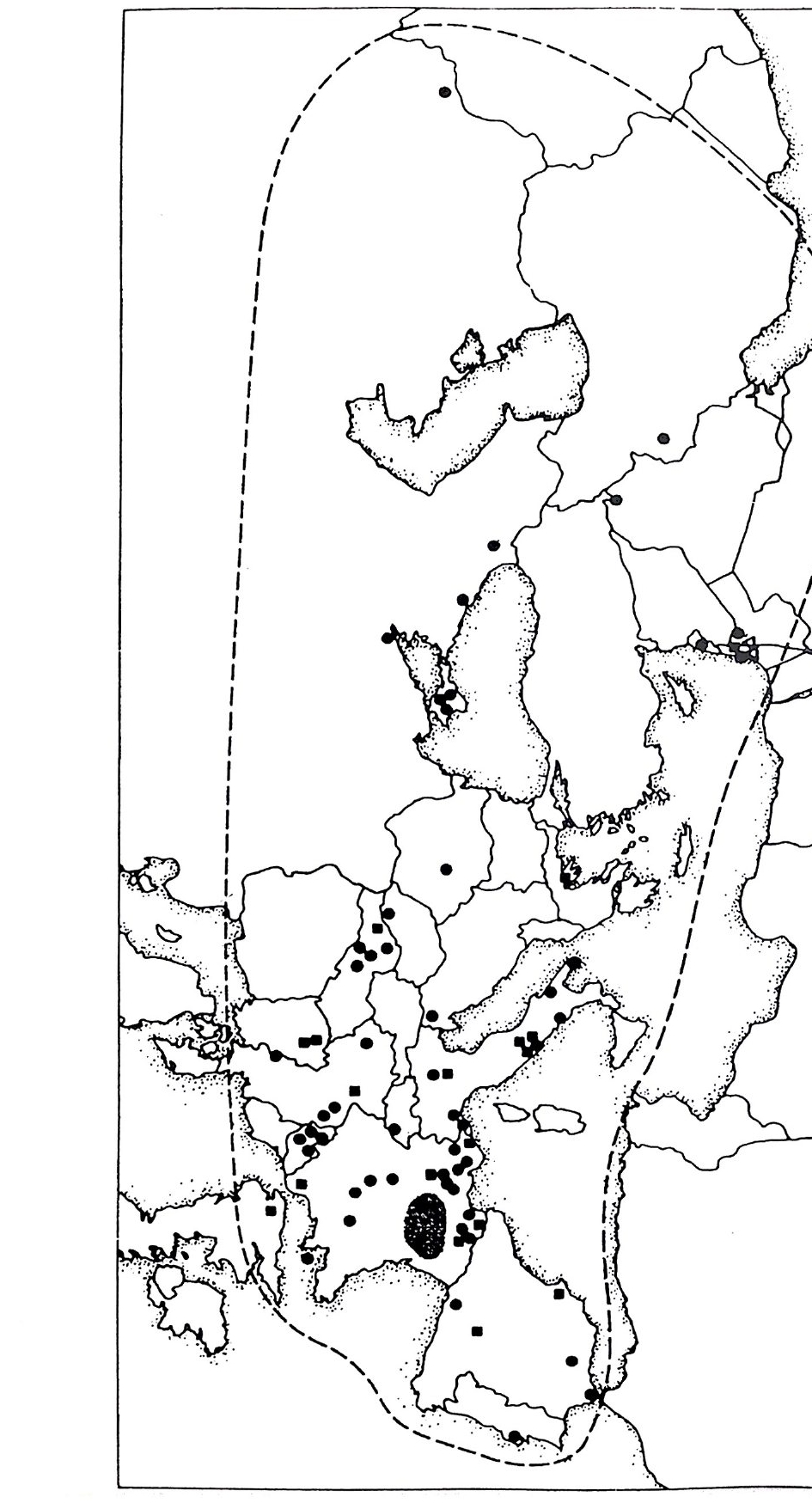
스타인의 나사렛 근처 카프체 동굴에서 발견된 것으로 이상적으로 빠 른 시기인 60000y B P 시대의 것으로 생각된다. 이는 뒤에 다시 설명하 게 될 것이댜 (a) 는 체코슬로바키아의 프레드모스티에서 발견된 것으 로 25000y B P 시대의 것이다. HSN 의 두개골은 두드러진 눈두덩이 길 고 낮은 두정으로 이어지는 이마는 몰입하듯 쑥 들어갔다. 뇌 용량이 HSS 의 평균 1350cc 보다 크다(평균 1450cc). 턱뼈는 앞으로 쭉 뻗어나 와서 최종 어금니와 턱뼈의 위로 구부러지는 끝 사이에 상당한 간격이 있댜 그림 2 .l Oc,d(HSN) 과 그림 2 .l Oa,b(HSS) 에 그 차이가 잘 대조되 어 있댜 코와 볼뼈는 앞으로 불거져나와 얼굴이 눈두덩보다 돌출한 모양을 보인다. HSN 의 치아는 HSS 의 것보다 다소 크다. 그러나 얼굴 과 머리는 강하고 잘 구성된 인간의 아름다운 몸 위에 서있는 특징을 가지고 있다 . 네안데르탈 화석은 유럽과 근동의 넓은 전지역에 걸쳐 놀랄 만큼 유 사하다 (T ri nkaus 및 Howells 1979, Tr ink aus 1984). 호모 에 렉 투스에 서 HSN 으로의 이행은 그림 2 .1 2 에 사각점으로 나타냈고 유럽과 레반트에 국한되어 그림 2.11 에 보여주듯이 5 만년에서 12 만년 사이에 이루어진 것 으로 생각된다. 두개골은 점차로 커져서 호모 에렉투스의 뇌 용량 800-1200cc 는 HSN 의 크기에 가까워지고 이행형 얼굴 모양을 가졌다. 옐리 넥 (19 8.5)은 이들을 모두 합하여 호모 프레네안데르탈리스 Homo p reneander l:a li s 라고 하였다 13 만y BP 후에는 단지 HSN 화석과 도구만 이 발견된댜 HSN 화석은 그림 2 .1 2 에 둥근 점으로 나타냈고 프랑스의 중앙 산괴 Cen tr al Mass if를 중심으로 유럽과 근동에 널리 분포한다. 그 림 2 .1 2 에서 보여주듯 HSN 의 광범위한 분포는 유럽의 호모 프레네안데 르탈리스 터로부터 이동했음을 쉽게 짐작할 수 있다. 채집수렵인들은 버딩 bud ding, 죽 한 세대 동안 다음 삶터를 향해 20lan 씩 무작위 이동을 했음직한데, 이것이 더하여져 1000 년 동안에 lOOOlan 까지 되었을 것이 다. 따라서 HSN 의 전체 영 역에 이르는 데에는 5000 년이 걸렸을 것이다.
 유럽호미니드화석 근동호미니드화석
유럽호미니드화석 근동호미니드화석
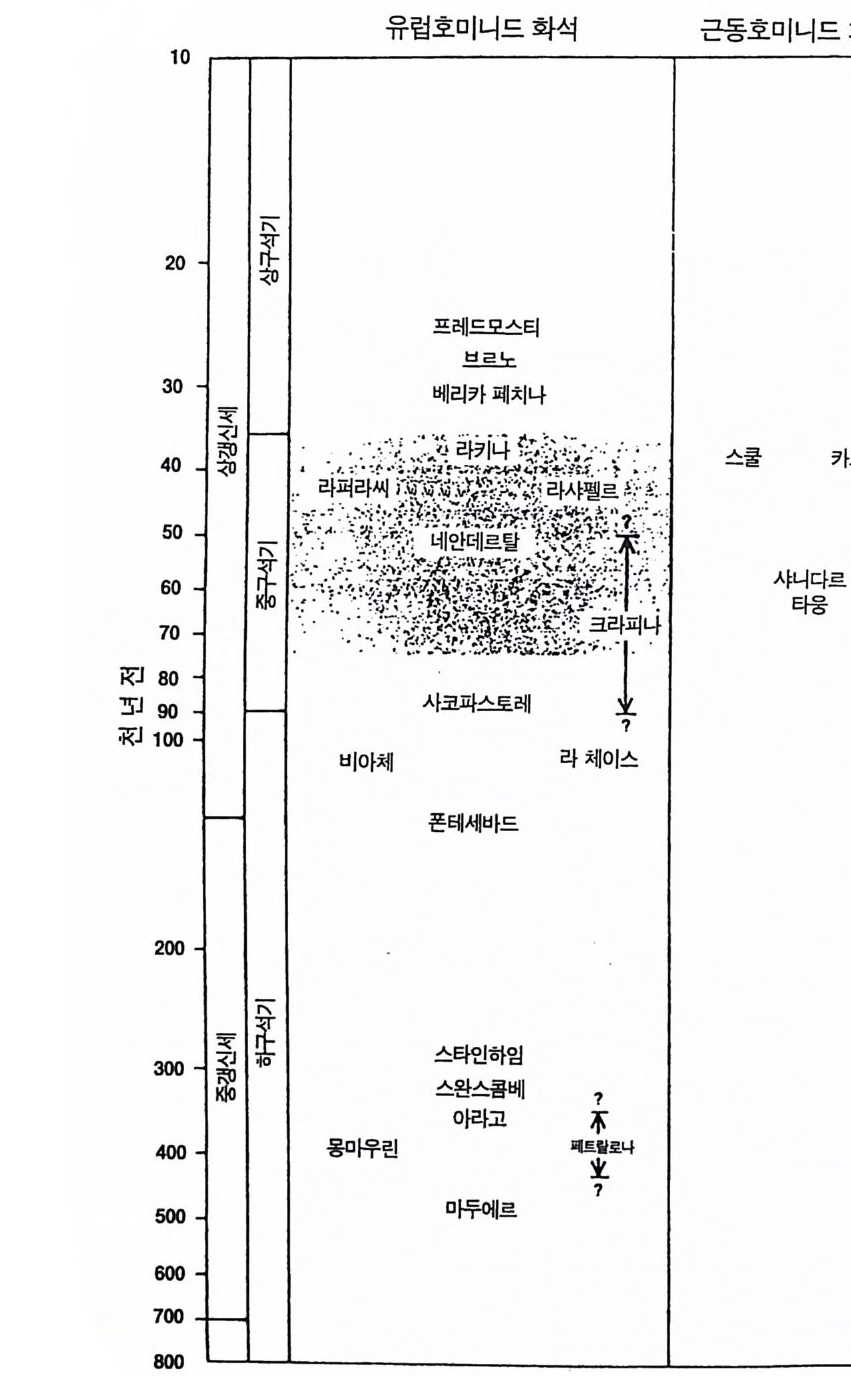
그림 2.11 네안데르탈의 시대적 분포. 이 도표는 80 만년 전부터 1 만년 전까지로 되어 있다 . 중 및 상구석기 시대와 상갱신세를 포함하는 충분한 여유 를 갖 기 위해 시간 간격은 대수눈금으로 되어 있다 . 상갱신세의 최후 빙하기는 8 만년 전에서 1 만년 전까지이고 35000 년 전의 온난기에 의 해 중단되었다 . 유럽의 여러 네안데르탈 발견 지역이 모두 정확하게 연대가 밝혀지지는 않았으나 대부분은 75000 년에서 35000 년 전 사이 이다(빗금친 띠) . 크라피나에서 발견된 가장 오래된 화석은 유럽의 다 른 네안데르탈보다 약간 오래된 것이나 대부분은 동시기의 것이다. 8 만년 전보다 오래된 화석에는 초기 네안데르탈로 분류될 수 있는 것 이 있다. Saccop a sto r e, Bia c he, La Cha ise 둥이다. Fon t echevade 에 서 Mauer 에 걸쳐 발견된 더 오래된 유럽 화석은 정도의 차이는 있으 나 네안데르탈과 호모 에렉투스에 가깝다. Velik a Pecin a , Brno, Predmosti, Skhul 및 Qafze h 둥지의 상구석기 지역에서는 현대인형 의 화석 이 발견된다 . (E . Tr ink aus and W. W. Howells; The Nean- dert ha l Sc ien ti fic America n, Dec. 1979)
2.6 호모 사피 엔스 사피 엔스 (Facch ini 1984, Sm ith 1984t 두개골의 특징적 모양(그림 2 .1 0) 은 두개골 계측치를 후기 두개골 화 석들과 비교하여 표 2.3 에 나타내었다 . 유럽과 동방(주로 샤니다르)에서 온 네안데르탈은 두드러진 돌출 안면과 강한 저작기를 보인다. 일반적 으로 HSN 에서 HSS 에로의 변화는 건장한 HSN 이 비교적 가날픈 HSS 가 된 것으로 생각될 수 있다 (T ri nkaus 1983, 1984). 진화적 변화는 식 품 취득 기술(사냥과 수확) 그리고 먹기 전의 음식 조리 기술의 개선을 수반한다고 생각된다. 결국 건장한 네안데르탈은 불필요하게 근육이 발달하고 기술에 비하여 힘에 의존하므로 HSS 에 비하여 불리하였다 . HSS 의 사지뼈는 가늘고 길었으며 여성과 남성의 키가 평균 12cm 정 도 증가하였다. 팔의 만곡도는 줄어들어 요골의 만곡지수는 5.5 에서 그 반인 2.66 이 되었다 . 견갑골은 더 가늘어져 HSN 의 평균 넓이 11. 4 mm 이 HSS 는 9 . 7mm 이 되었다 (T ri nkaus 1984). 설계의 개선으로 손뼈 특
표 2.3 HSN 과 HSS 화석의 차이점 특징 동네안데르탈 유럽네안데르탈 카프체 - 스쿨 초기 상구석기 현내인 키 평균 (cm) 166 165 177 178 대뇌 용적 (cc) 1545 1577 안면 돌출지수 (mm) 99 97.5 83 85 -81 하악 길이(정규화치) 25.3 26.6 22.8 21. 5 -21 . 0 하악가지 넓이 (mm) 43 44.5 38.3 36 34 얼굴 비근점 치조점 높이 (mm) 94 85 74.6 69 57.5 양악 넓이 (mm) 114-120 109 107 96 98 치골 관골구 치골 결합 길이 (mm) 93-80 59- 83 58- 73 키에 대한 지수 166 167 117 120 견갑골 넓이 (mm) 11. 4 9.7 요골 만곡지수 4.3 3 5.9 6 2.73 2.5 9 (Tri nka us 1983) 히 엄지손가락의 변화는 해부학적으로 가날픈 것에 비하여 힘있게 쥘 때의 정확성을 얻게 되었다 . 치골의 변화는 관골구 치골결합 사이가 줄어든 것이다. 골반이 좁아져 운동 효율이 개선되나 분만이 어려워진 댜 그러나 이는 출산 개시를 빠르게 함으로서 상쇄될 수 있고 (5 . 5) 조 산아에 대한 자녀교육 개선이 있었다면 자연도태로서도 받아들여질 수 있다 (T rink aus 1984). 음식 조리방법의 개선으로 저작근의 감퇴와 씹기에 전적으로 참여하는 구치 및 전구치에 비하여 앞니 크기의 감소 를 가져왔다. 따라서 골격에 있어 HSN 에서 HSS 로의 변화는 모자이크 식 적옹 변화라 하겠다. 수십만 년에 걸친 호모 에렉투스의 분명한 점진적 계통발생 과정에 서 HSN 의 시작이 점진적인 것에 반하여 HSS 로 대치되는 것은 대단 히 빨라서 어느 지역에서건 5000 년을 넘지 않는다. 표 2 .4에 HSN 에서
HSS 에로의 개략적 이행시기를 나타내었다. 이것은 물론 개선된 연대 기법으로 개정한 것이다. 그러나 이 표에서도 HSN 에서 HSS 로의 이행 이 가장 빨랐던 곳은 팔레스타인이고 중앙 유럽은 그보다 5000 년 늦고 서유럽은 그보다 수천년 더 늦었음을 보여주고 있다 . 점진적인 시기의 지연은 팔레스타인의 원점으로부터 네안테르탈이 현대 인간 (HSS) 으 로 대치되면서 서쪽으로 유럽을 통하여 이동함에 의한 유전자 흐름 탓 이라고 할 수 있다. 이는 화석으로도 나타날 뿐 아니라 무스테리아 Mouste r i an 석기문화가 오리나시아 A urig nac i an 석기문화로 바뀌는 것 으로도 알 수 있 다 (6.8). 팔레스타인 나사렛 근처의 카프체 동굴에는 대단히 흥미 있는 화석 이 있었다. 이는 스쿨에서 발견된 것과 비슷했고 해부학적으로 현대 호모 사피엔스라고 유라시아 지방에서 알려진 것 중 가장 초기의 것이 다 . 카프체-스쿨에서 발견된 화석은 대부분 HSS 의 범주에 속하나 일 부 네안데르탈의 특징을 보였다.(표 2.3 ) 예를 들면 얼굴의 나시온-프 로티온 Nas i on-Pro thi on 높이와 위턱 넓이가 이행상이다. 따라서 잡교배 의 가능성이 제기된다 . HSN 과 HSS 는 동일종인 호모 사피엔스의 변이 이다 고로 교잡을 생각할 수 있다 HSN 이 HSS 로 대치되는 이행형의 이에 비견되는 증거는 다른 장소에도 있다. 큰 고유 HSN 집단이 HSS 로 변형되었다는 것은 잡교배 과정에서 특징적 HSS 유전자의 표현형 표현에 편중이 있었음을 뜻한다 . 이 생각은 네안테르탈인이 완전히 소 멸당했다는 이론보다는 더 받아들일 만하다 . 가장 중요한 의문을 마지막으로 제기해 본다. 어떻게 호모 사피엔스 사피 엔스(현대 인간)는 나타나게 되었는가? 언제 어디서? HSN 화석과 도구(무스테리아 문화, 그림 6 .1 0c) 의 실질적 소멸과 오리나시아 문화를 가진 최초 HSS 의 등장을 설명하는 두 가지 대립하는 의견이 있다 (Je lin e k 1985).
 / / / / --` `\` `
/ / / / --` `\` `
그림 2.12 네안데르탈 화석이 발견된 지역 분포도. 가장 밀집된 지역은 프랑스 의 Massif Cen tr al 이다(빗금천 부분). 이 지역에는 적어도 10 개의 초 기 네안데르탈과 25 개의 후기 네안데르탈이 분포되어 있다. 기타 지 역에서 초기 지역은 네모로 지도에 표시하였고 (75 화석) 후기 화석지 역 은 검 은점 으로 나타내 었다 (200 화석 이 상). Levan t의 두개 삼각표 시 는 Nu g hare t -Skh 따과 Jeb el Qaf zeh 의 위 치 이 다 .(E. Tr inka us and W. W. Howells; Neandertlr: zls , Sc ien ti fic Ame rica n, Dec. 1 罪 9).
표 2.4 HSN 에서 HSS 로의 이행시기(년전) 팔레스타인 중앙유럽 서부유럽 타분 스쿨 카프체 최후 HSN 45,000 40,000 35,000 최초 HSS 41,000 70,000 34,000 32,000 T ri nkaus(19 腐 1984) CD HSS 은 다른 곳에서 발생하여 유럽 내로 침입한 사람들이었다. HSS 의 우수한 효율 때문에 네안데르탈은 소멸하였거나 홉수되었다 . 이 가설은 가상적 원격 주거지역의 확인에 의한 것이다. 다음에 설명하는 바 와 같이 이 장소는 아프리카였을 것이다 .41 ® 호모 사피엔스 사피엔스는 아마도 일부 격리된 작은 사회의 어떤 신속한 도약적 발전에 의해 네안데르탈로부터 발생되었다(1. 3). 이것은 호웰의 이른바 노아의 방주 가설이다 (S tri ng er 등 1984). 큰 네안데르탈 사회 의 침 투성 유전자 흐름 때문에 (그림 2.1 1 , 2.1 2 ) 국소적 유전자 도약 온 전체 네안데르탈 사회 속으로 신속하게 전파될 수 없었을 것이다. 대 량 학살이 아니고 잡교배가 있었을 것으로 생각할 수는 있다. 네안데르탈 은 풍부한 동물과 식물이 자라는 비옥한 영토에 살던 자비심을 가진 매 력 있는 사람들이었다(제 5 장). 결과적으로는 HSS 로 대치되기 위한 국소 적 HSS 조상이 점진적으로 형성되었다 . 다만 호모 사피엔스 사피엔스의 발생 장소는 의문으로 남는다.
팔레스타인 내 카프체와 스쿨의 초기 상구석기 시대 발견 에 이미 대 답이 있을지 도 모르겠다 (T ri nkaus 와 Howells 1979, Tr ink aus 1983, Jel in e k 1985). 그 두개골들은 별 의미없는 네안데르탈의 모양(표 2.3) 이 있을 뿐 해부학적으로는 현대 호모 사피엔스 사피엔스로 분류될 수 있 다(그림 2.1 0 b). 그러나 타분 근처의 네안데르탈 화석은 분명히 달라 그 림 2 .l Oc,d 와 비 슷하다 호모 사피엔스 사피엔스가 실제로 팔레스타인에서 유래한 것이라면 종화의 제안된 진화기전의 좋은 예가 된다(1.3; Gould 1982). 호모 사피 엔스 네안데르탈리스는 거의 10 만년간의 정체 또는 평형기가 있었다 (그림 2.11). 그러고 나서 주변 격리장소(카프체)에서 도약적 유전적 변 화가 일어나 이는 유전자 흐름의 희석 영향을 덜 받으면서 작은 국소 집단 내에 유전적으로 파급되었다. 초기 연대 측정에 의하면 카프체가 스쿨보다 앞선 것으로 알려졌으 나 70,000-80,000 y BP 라는 이른 시기가 믿을 만한지에 대하여는 의문 이 제기되어 왔다(J el i nek 1985). 그런데 보다 현대적인 아미노산 라세 미 화법 에 의 한 연 대 측정 으로도(카프체 는) 68,000-78,000 y BP 가 되 어 스 쿨의 41,000 y BP 보다 훨 씬 이 르다(J e li nek 1982, 1985). 카프체 화석 의 조 기 연대에 대한 논쟁의 초점은 25000 년 동안의 지나치게 긴 기간 동안 의 격리라는 심한 제한 때문에 30km 의 거리에 이웃하여 살았던 사람 들 간의 유전적 문화적 교류가 거의 없었을 것이라는 점이다. 그러나 이 논쟁은 호미니드 존재의 놀랄 만한 보수성을 간과하고 있다. 예를 들면 이미 지적했듯이 (2 .4) 동아프리카에서는 호모 하빌리스는 100,000 년 동안이나 새로 발생한 호모 에렉투스와 같은 장소에서 살았다 . 심 지어 이 시기의 격리 부족들의 희귀한 예가 뉴기니 고원지대에서 발견 되었댜 20 세기 중반 오스트레일리아 지배계급이 점령할 때까지 그 부 족들은 완전한 격리상태에서 살았고 각 부족은 서로 다론 언어를 썼 다. 뉴기니에는 대충 700 가지의 언어가 있었다고 추산된다. 심지어 지
리학적으로 특별하게 다를 것이 없는 지역에 서로 이웃한 부족들간에 도 완전히 다른 언어를 사용하였다. 주의 깊은 언어학적 연구에 의하 면 관찰된 것과 같은 언어적 분포를 나타내려면 각 부족은 약 4000 년 이라는 이웃과의 격리 기간이 필요하다는 결론에 도달하게 된다. 기미 부족의 포어족 여자 납치, 그래서 쿠루에 의해 괴롭힘을 당하게 되는 예처럼 여자 사냥을 위한 부족간 싸움으로 유전적 침해가 있었던 예는 있지만 그 같은 격리는 무한정 계속될 수 있었다 (Ga j dusek 1CJ 73 ). 그와 같은 문화적 격리는 70,000y B P 시절부터도 팔레스타인에서 있 었댜 즉 그 시절 아프리카로부터의 파종에 의했거나 도약적 유전자변 화로 기원하는 카프체의 현대인에서 있었다. 20,000-30,000 년의 격리 후에 HSS 의 유전적 변화는 스쿨로 확산되고 표 2 . 3 에 지적된 바와 같 이 제한된 네안데르탈의 특징만을 가진 초기 호모 사피엔스 사피엔스 의 집단에 귀착한다. 그래서 확대된 HSS 집단은 다음 10,000 년 동안에 유럽에 그들의 분명한 유전자를 확산시킬 수 있었다(그림 2.1 1 , 표 2.4 ). 결국 발전된 수렵 수확 기술과 음식 조리 기술 등 우수한 지능과 기술 로 그들은 유럽의 네안데르탈 영토에서 집락을 형성하게 되었다. 현대 인간은 그들 특유의 특징을 해치지 않고 네안데르탈 유전자의 혼입을 어느 정도 수용할 수 있었던 것 같다. HSN 과 HSS 는 단순히 호모 사 피엔스의 아종으로서 교배가 가능했던 것으로 생각되어 왔다. 이제 첫번째 대안 즉 네안데르탈 세계는 HSS 집단에 의해 침범되었 다는 설로 돌아가자. 그림 2.1 0 , 2.1 1 , 2 .1 2 에서처럼 유럽과 근동지방의 화석은 수적으로나 유지 상태가 좋아 호모 사피엔스 사피엔스의 기원 을 밝히려는 장소의 중심이 되었다(두번째 대안은 위에서 논했다). 일부 는 트린카우스와 호웰스 (1979) 의 종설에 근거하고 뒤에는 트린카우스 (1983, 1984) 의 광범 한 저 술에 근거 한다. 그러 나 브라우어 (Br auer, 1984) 나 라이트미르(Rig h tmir e, 1984) 그리고 스트링거 및 동료 (S tring er 둥 1984) 가 호모 사피엔스 사피엔스의 아프리카 기원에 관하여 제기하였
던 반론에도 주목할 필요가 있다 . 호모 에렉투스의 발생까지의 유안류화가 아프리카에서 기원하였다 는 것에 관하여는 이론이 없다 (2 .4). 그러나 호모 에렉투스가 유라시아 로 이주한 후부터는 아프리카의 지역적 중요성이 상실된 것 같다 . 그 러나 지난 15 년 동안에 보다 연대가 확실한 화석들이 많이 발견되어 아프리카 동부와 남부의 호모 에렉투스가 유럽의 전네안데르탈인과 비슷한 특징을 가진 유인류를 거쳐 네안데르탈로이드 시기를 지나 호 모 사피 엔스 사피 엔스로 발전하였다는 가설에 이르게 되 었다 (Brauer 1984). 호모 사피엔스 사피엔스가 처음 출현한 연대는 화석 증거로는 90,000y B P 또는 그보다 빠르댜 이는 유라시아의 HSS 출현보다 훨씬 빠르고 심지어 의심스럽도록 빠른 카프체 HSS 화석보다도 빠르다. 불 행히도 브라우어(1 984) 가 해부학적으로 초기 현대인 호모 사피엔스라 고 제시한 아프리카 화석들(크리지에스, 보더 동굴, 오모 I, 캄제라, 뭄바 의 것들)은 단편적이고 연대 측정이 적절치 못한 것들이다. 하지만 유 라시아 HSS 화석보다 더 중요함은 일반적으로 인정된다. 훨씬 우수한 화석 기록들은 현재도 진행되는 철저한 탐구로 곧 밝혀질 것이다. 이 미 이들 HSS 화석은 동아프리카와 남아프리카에 널리 분포하고 있음 이 밝혀졌다. 더욱이 하사하라 sub-Sahara 아프리카인의 돌다루기는 유 라시아의 중석기 시대와 같은 시대이거나 어쩌면 더 빠를 것이라는 증 거들이 있다 (Brauer 1984). 유라시아 HSS 의 기원에 관한 가장 중요한 의문은 이는 그 지방 고 유의 것이었는가 아니면 나일 계곡 상류에 있던 HSS 집단의 이주에 의하여 유전적으로 촉발된 것인가이다. 이는 아마도 동아프리카의 호 모 에렉투스 집단이 유라시아로 이동하고 그래서 자바와 중국에서 크 게 발전하게 되는 길이었을 것이다 (2 .4). 아프리카 하사하라에 해부학 적으로 현대 호모 사피엔스라 할 수 있는 것이 100,000y B P 이상의 이 른 연대에 있었음은 이동할 시간적 여유가 있었을 가능성울 제시한다 .
팔레스타인의 카프체와 스쿨은 유라시아로 널리 퍼져 나가는 길목에 위치하고 있다 불행히도 단지 한 개의 화석 기록만이 이 북쪽으로의 이동로에서 발견되었다. 이는 수단의 싱가에 있는 것으로 연대는 확실 치 않다. 이것은 초기 호모 사피엔스로 생각되며 해부학적으로는 현대 HSS 가 아니 다(Rig h tmir e 1984). 2.7 호미 니 드 진화의 문제 점 (Facch ini 1984) 사람이 호미니드의 후손이라는 일관된 생각은 지리적으로 넓은 지 역에 거주하는 여러 종족으로부터 채취한 많은 수의 미토콘드리아 DNA 분석 결과에 따른다 (Cann, Sto n eki ng , W ilso n 1987). 미토콘드리 아 DNA 는 정자가 접합체를 이룰 때는 관여하지 못하므로 모계에 의해서 만 유전된다 . 제한지도법 res tri c ti on ma pping이라는 특수한 기법으로 모 계 계보를 더듬어 올라갈 수 있다. 그래서 지구상의 모든 인간은 20 만 년 전에 아프리카에 살았을 것으로 생각되는, 한 어머니의 후손이라는 잠정적인 결론에 이르렀다. 계보의 초기 특정 가지에는 아프리카 흑인 만이 포함되기 때문이다. 칸과 공동연구자 (1987) 들이 지적했듯이 이는 호모 사피엔스 사피엔스가 아프리카에서 기원하여 그곳에서 불어나고 전세계로 퍼져나갔음을 말하는 것이다. 그러나 이 생각은 아직도 확인 되고 비판될 여지가 있다. 또 한가지 의문은 자바와 중국에 있던 진보된 호모 에렉투스의 종 국적 운명에 관한 것이다. 이것의 뇌는 1000cc 를 넘어섰다(그림 2.5, 2.6, 2.1 3 ). 더욱이 약 30 만년 전에 자바에는 솔로인 Solo Man 이 있었고 이는 후에 호모 에렉투스로 발전하여 뇌가 1200cc 에 이른다. 대체로 유럽에서 있었던 것과 마찬가지로 호모 에렉투스는 격리 상태에서 전 네안데르탈 상태로 발전하였다고 생각된다(그림 2.1 1 , 10 만년 이전의 사
항). 스트링거와 공동연구자들 (1984) 은 북아시아에 있어 호모 에렉투스 로부터 호모 사피엔스로의 진화 과정은 네안데르탈의 상사형일 것이 라고 주장하였댜 비슷한 발전과정을 자바에도 적용할 수 있을 것이 다. 호모 사피엔스 사피엔스가 근동지방 근원지 또는 아프리카의 전이 인으로부터 전파되었을 것이라는 제안은 아직도 의문의 여지가 있는 것 같댜 여기서 호모 사피엔스 사피엔스에 의하여 세상이 점령되는 것은 유라시아에서 이루어져야 하기 때문에 아프리카의 전이인설은 아프리카 HSS 집단을 이동상태로 간주하고 있음에 주목해야 한다. 이 같은 호모 사피엔스 사피엔스에 대한 기원의 추정으로부터 약 15,000 년에 걷쳐 아시아에 이어 베링 해협을 거쳐 아메리카로 그리고 순다 협곡을 거쳐 오스트레일리아로 세계의 모든 주거 가능지역을 점 유할 수 있도록 하는 가히 경이적인 성취가 이루어진다. 이들은 모두 가 마지막 빙하기 동안의 걸어다니는 호미니드였다. 지난 4 만년 동안 에는 생물학적인 진화는 없었던 것 같다. 약 70,000 y BP 에 출현하는 HSS 의 뇌는(그림 2.10b) 막대한 문화적 발달 가능성과 엄청난 동기를 가졌다. 이는 인간의 생활과 활동뿐 아니라 온 지구를 변화시켰다. 생 물학적 진화는 문화적 진화로 이어지는데 이는 제 10 장에서 다룰 것이 댜 제 6 장에서는 문화적 소산, 죽 석기, 동굴벽화, 원시적 조각 동을 다 룰 것이다. 제 5 장에는 꽃장식, 장례의식 둥 네안데르탈인의 영적 여명 을 다루었다 (Solec ki 1971, lfJ7 7 ). 세계의 팽창에 관하여 호미니드 진화 후기의 인구를 추정해 볼 필 요가 있다. 오스트랄로피테신, 호모 하빌리스, 호모 에렉투스 둥 수렵 인들은 30 또는 50 명의 작은 부족 집단을 이루었을 것이며 그래서 아 프리카 전체의 호미니드 인구는 모두 합하여 수천 명을 넘지 않았을 것이다 호모 에렉투스가 유라시아로 확산되던 시기인 1,000,000 y BP 에 도 세계 인구는 125,000 으로 추산한다(D eeve y 1960). 네안데르탈 시기 에는 인구 중가가 별로 없었다. 샤니다르 Sh ani dar 부족사회는 100 명을
 2000
2000
넘는 시기가 한번도 없었다 (1971). 인구의 팽창은 가축과 농작물을 기 르는 농업경제가 자리잡는 신석기 시대에 시작되었을 것이다. 디비 (De evey 1960) 는 25,000y B P 때의 세계 인구는 300 만이었을 것으로 추 정하였다.
그림 2 .1 3 은 호미니드 두개골의 크기와 연대에 따라 진화하는 모양 울 그린 것이다. 네안데르탈인은 직사각형 안에 호모 사피엔스로 표시 하였고 오른쪽 검은 방추 모양에 호모 사피엔스 사피엔스가 표시되었 댜 그림 2.6 에는 지난 300 만 년 동안에 있어 95% 인구의 현저한 뇌의 진화적 팽창이 나타나 있다. 가설에 의하면 이 같은 뇌의 팽창은 진화 의 한시적 평형 과정에 있어 도약적 유전자 변화의 결과라는 것이다 (1.3, Eldr edg e 와 Gould 1972). 뇌 기능의 점진적인 중가는 자연도태의 중심적 역할을 하였을 것이다 .
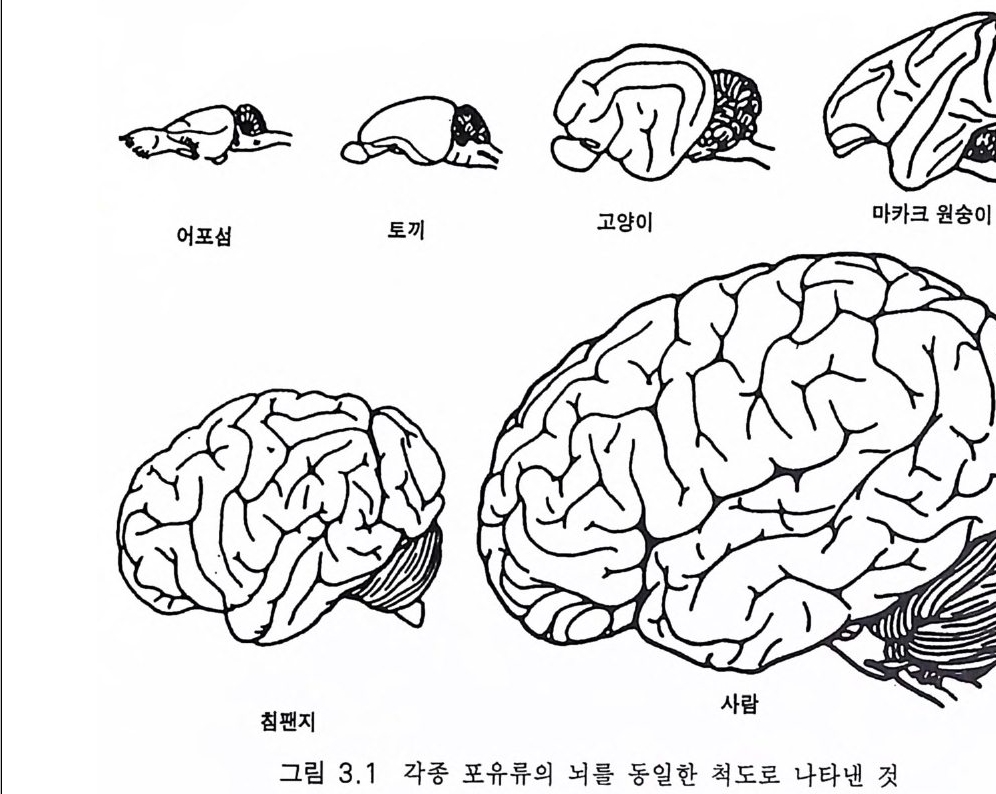
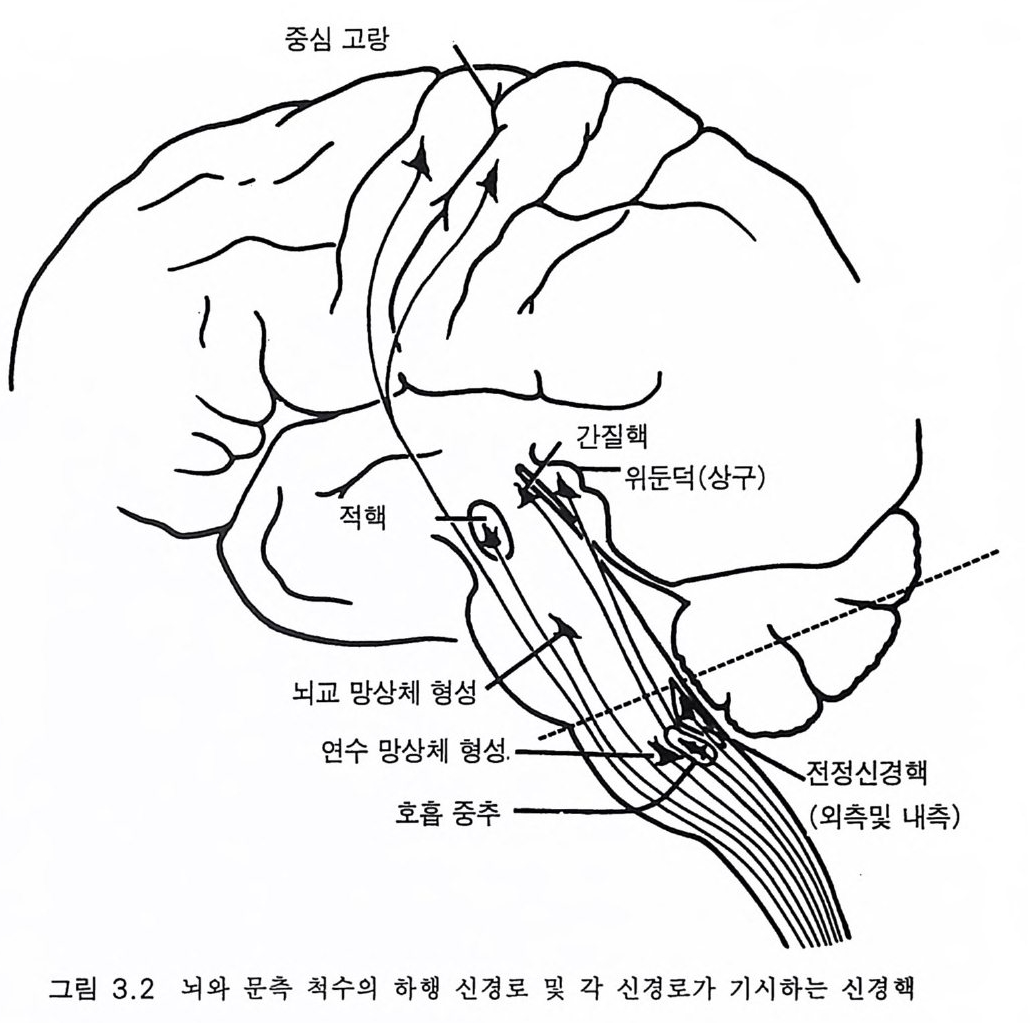
제 3 장 호미니드 뇌의 전화 ―양각 보행, 민활 행동 3.1 기본 구조 그림 3 .1은 세 가지 영장류 포유류의 뇌를 동일한 축척으로 나타낸 것이다 대뇌가 점차적으로 커지는 것이 두드러진 모양이다. 소뇌도 그 정도는 작지만 평행하여 커진다. 뇌의 여러 중요 부분의 측면과 상 부 척수는 그림 3.2 에 나타내었다. 그리고 뇌의 세로 절편은 그림 3.3 에 있다 이들 그림은 광범한 영장류 뇌의 진화 특성을 이해하는 데 적당 한 것으로 스테판과 앤디 (S tep han 과 Andy 1969) 의 연구를 시작으로 뒤 이은 스테판과 공동 연구자들에 의한 일련의 연구를 최근의 종설로 마 무리 한 것 이 다 (S t e p han 1987). 포유류 중추신경계의 일반적 형세는 첫째로 척수와 관련지어 생각 할 수 있는데 경수와 요수는 척수 분절 신경에 의하여 팔과 다리가 신 경 지배되는 정도에 따라 중대한다. 각각의 분절 신경은 전근과 후근 으로 구성되고 전근은 신경섬유를 근육에 보내고 후근은 피부, 근육, 관절 둥 말초 감각 기관으로부터 척수에 보내지는 신경섬유이다. 척수 내에는 많은 신경로가 있는데(그림 3.6, 3.1 4 ) 뇌에서 하강하여 전근과
 ¢3 도
¢3 도
사지 신경을 통하여 운동 신경으로 지배되는 근육에 정보를 전달하고 (그림 3.7, 3.1 5) 상승하는 많은 신경로는 피부, 근육, 관절 둥의 수용체 로부터 정보를 뇌로 전달한다. 끝으로 척수 자체(그림 3.7, 3 .1 5) 는 중추 신경계의 일부로서 여러 반사작용과 상위 중추의 조절과는 독립적인 통합 기능을 한다 이 같은 기본적인 형식은 모든 포유류 진화 변천을 통하여 유지되었다. 제리슨(J e ri son 1973, 19 8.5)은 서로 다른 종들의 대뇌 발달을 단지 체 중 (BoW) 에 대비한 뇌 중량(B rW) 만으로 구하였다. 이는 크게 서로 다 론 체구를 가진 넓은 범위의 종들에 대한 계측치를 경험적으로 구했기 때문이며 그 관계는 아래 식과 같이 대수적 관계를 보인다.
 ------- -
------- -
BrW = k • BoW213 제리슨은 대뇌화 계수 (ence ph aliz a ti on qu oti en t E Q)의 개념을 개발하 였다. 죽 실제 뇌 크기에 대한 위의 식으로 구한 체중을 감안한 살아 있는 평균 포유류 뇌 크기의 비율이다. E Q값의 범위는 프로시미안(원 시 영장류)의 1. 1 에서 폰지드의 1. 9( 침팬지 2.3) HSS 의 8.5 사이이다.
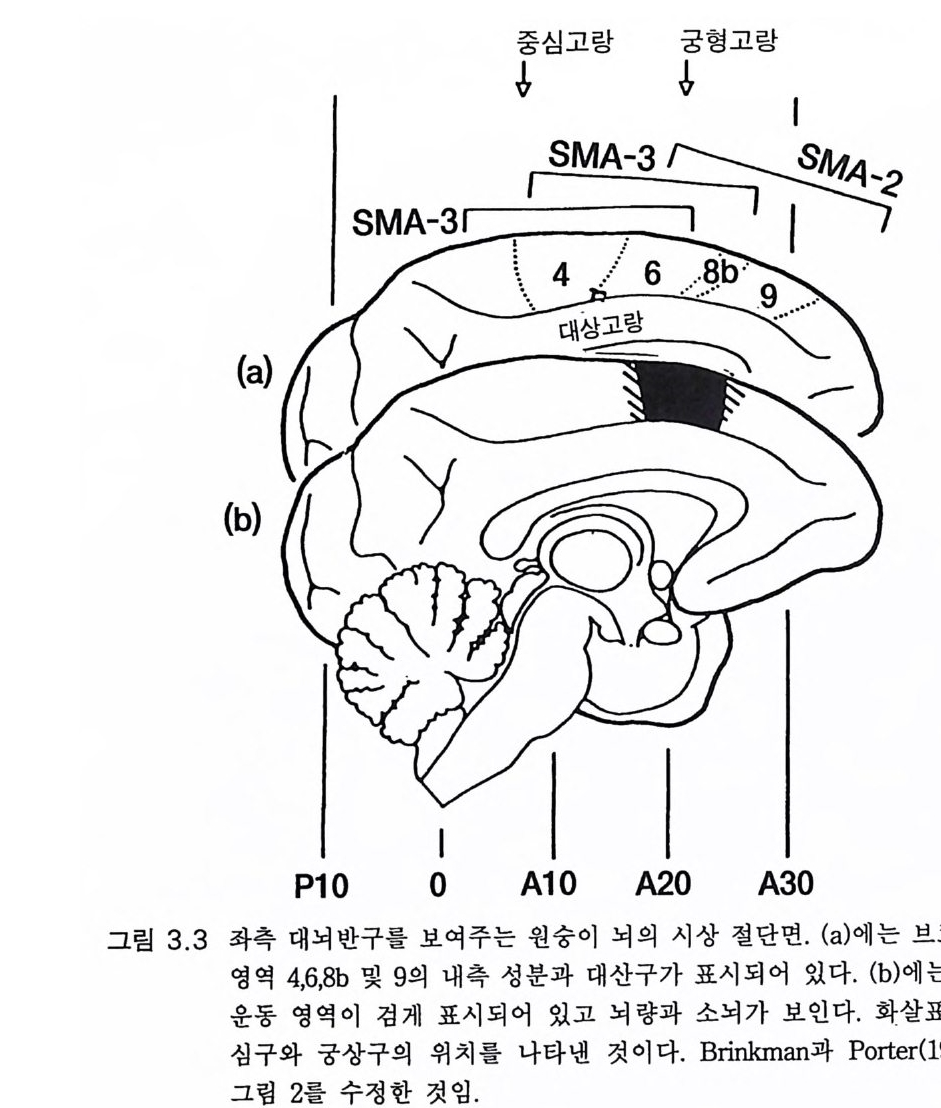 중 심고랑궁형고랑
중 심고랑궁형고랑
한편 스테판과 공동 연구자들 (19 얽)은 여러 종류의 영장류 뇌의 특 정 부위 크기를 비교하는 방법을 찾았다. 그들은 가장 원시적인 살아 있는 영장류인 기초적 인 식충동물 Tenrec in ae 을 기준으로 이용하였다. 이 동물의 뇌는 소멸된 동종의 동물 뇌와 비슷한 것으로 생각되며 영 장류를 포함하여 여러 포유류급의 선구자로 생각되는 것이다. 체중이 크게 변하는 데 대한 BrW:BoW 비의 광범위한 연구로 0.63 이라는 대
표 3.1 주요 뇌 성분 크기 지수의 텐레크류 (Tenrec i nae) 에 대한 비교치 텐레크류 식충류 원원류 신, 구세계 폰지드(기 호모 원숭이 본, 침팬지, 고릴라) (N=4) (N=50) (N=18) (N=23) (N=3) (N=l ) 연수 1.00 1.27 1.56 1.g 1.61 2.09 중뇌 1.00 1.31 2.71 3.4 0 2.86 5.1 6 소뇌 1.00 1.64 4.64 6.20 8.81 21. 75 간뇌 1.00 1.56 5.56 8.00 8.57 14.76 후구 1.00 0.81 0.52 0.08 0.0 6 0.03 후각피질 1.00 0.94 0.65 0.34 0.3 1 <0.3 편도체 1.00 1.10 1.73 2.24 1.8.5 4.48 중격 1.00 1.22 1.91 2.09 2.1 6 5.4 5 해마 1.00 1.75 2.91 2.64 2.99 4. 없 분열피질 1.00 1.68 2.8 0 2.23 2.38 4.43 선조체 1.00 1.80 5.99 10.1 2 11. 78 21. 98 신피질 1.00 2.65 20.37 48.4 1 61. 88 196.4 1 대뇌화지수 1.00 1.43 4.24 8.1 2 11. 19 33.7 3 자료 : Ste p h an 둥, per sonal commu nica ti on , 1988 수 지수를 얻게 되었는데 이는 제리슨의 2/3 에 비해 별 차이가 없었다. 그러나 대뇌화 지수 (ence phliz a ti on ind ex EI) 는 기준치가 변하였으므로 제리슨의 대뇌화 계수와는 다르다. 몇 가지 EI 값이 표 3.1 에 있다. 스 테판과 공동 연구자들 (1987) 은 폰지드와 HSS 를 제외한 18 원원류 p ros irni an 와 23 유인원 s imi an 의 EI 값을 구했는데 원원류의 평균 EI 값 4.24 는 식충 동물에 비하여 현저한 진화적 발달을 보이는 것이고 이는 계속해서 유인원 (EI-8 .1 2), 폰지드 (EI-1 1.1 9) 를 거쳐 HSS 의 33.73 으로 크게 발달한다. 스테판 둥 (1987) 의 연구는 포유류 뇌의 주요 부위의 크기를 주의 깊 게 측정한 것이므로 특히 중요하다. 뇌의 연속 절편법으로 주요 부위
 l. >--_ _— — — _- ...... _ 피질피30질0m간 기등
l. >--_ _— — — _- ...... _ 피질피30질0m간 기등
그림 3.4 피질 피질간 주상 구조 또는 모듈(중앙에 있는 약 300 µm의 수직 원통 형 구역)(그림 9.4 참조)의 신경세포 연결 관계. 모듈은 인접한 두 개의 납작한 원주와 판 IV 에서 구역을 공유하고 있다 . 그곳에서 특이적 상 행섬유 (SPEC. AFF.) 가 가지치게 된다. 피질 피질간 상행섬유(밑바닥 에 표시됨)는 모든 피질 피질간 모듈에 걸쳐 종착한다. 그러나 종말의 밀도는 서로 다르다. 판 I 에서 피질 피질 섬유의 접선 방향 확산은 모 듈을 훨씬 벗어난 곳까지 뻗는다. 추상세포의 출력 선택은 그림의 오 른쪽 반에 있는 흥분성 중간 뉴런(극성 위성세포 SS) 와 탈억제성 중간 뉴런(라몬 이 카할의 Cellules a double bou q ue t (CDB) 로서 억제성 중 간 뉴런이고 억제성 중간 뉴런에 특이적으로 작용하며 검게 표시하였 다)으로 정해진다. 그림의 왼쪽 반은 일부 추상세포(무늬 있는 것)의 작용을 억제하는 일종의 신경섬유로서의 억제성 중간 뉴런의 작용을 설명한다. 상당한 근거로 억제성이라고 규정할 수 있는 중간 뉴런은 심판에 있는 바구니세포 (BC), 판 11 에 있는 작은바구니세포 (SBC), 축 삭타래세포 (ATC) 그리고 추상세포 축삭의 첫 분절에 작용하는 대단 히 특이적인 축삭축삭세포 (AAC) 로서 검게 나타내었다 .(Szentag o thai, 1983)
를 해부학적으로 식별할 수 있었고 용적을 측정할 수 있었다. 각각의 해부학적 부위의 측정치를 대뇌화 지수에서와 동일한 기준으로 계산 하여 크기 지수 (s i ze ind ex SI) 를 구했다. 표 3 .1은 원원류, 유인원, 폰지 드(긴팔원숭이, 침팬지, 고릴라), HSS 의 뇌의 열두 개 주요 부위의 평균 SI 값이다 가장 큰 점진적 증가를 보인 것은 신피질로서 HSS 의 SI 는 196 으로 폰지드 (SI=6 1. 88) 에 비해 세 배나 된다 .5) 기타 주목할 만한 증가 를 보인 것은 소뇌, 간뇌 그리고 선조체였다. 이에 반하여 취구는 점진 적으로 작아졌댜 표 3 .1에 열거된 구조는 다음 장들을 참조하기 바란다. 뇌의 진화적 발달은 정량적인 것이고 정성적인 것은 아닌 것 같다. 대뇌피질도 마찬가지여서 조직학적 구조는 기본적으로 변하지 않은 채로 남아 있다. 예를 들어 대뇌피질의 절편 모식도(그림 3 .4)에서 고양 이나 원숭이, 사람에서 제 II, ill, V 판의 지배적 추상세포들을 볼 수 있 댜 물론 대뇌피질의 기능적으로 특이한 부위의 크기는 뒤에 설명하는
것처럼 크게 변한다 . 이들 부위의 구성 양상도 달라서 대뇌피질은 다 양한 특성을 가진 부위들의 모자이크라고 할 수 있다. 이것도 뒤에 다 시 설명하게 될 것이다. 3.2 뇌 의 기 능적 작동 (Brooks 1986, Eva rts 1981) 사람의 운동을 이해하려면 뇌에서 척수를 거쳐 운동신경을 이루는 추체로의 진로를 이해할 필요가 있다. 이것의 축삭은 전근을 통하여 신경 흥분을 전달하여 근수축을 일으킨다(그림 3.6). 여기서는 인간의 뇌에 의한 운동 조절에 관계되는 개략적인 기본 원리를 설명하는 것으 로 족하다고 생각한다. 그 기본 원리는 유인원의 뇌나 원숭이의 뇌에 서도 크게 다를 것이 없다. 그림 3.5 에 왼쪽 대뇌피질 모식도에 발가락 (상중방)에서 입술(하측방)까지가 띠 모양을 이루고 있는 운동피질을 보여주고 있다. 이룰 제 4 브로드만 영역이라 하는데 중심 고랑(롤란도) 앞쪽에 위치하고 바로 뒤쪽으로 인접하여 감각피질(제 3,1,2 브로드만 영역)이 운동피질과 마찬가지로 띠 모양으로 배열되어 있다 . 제 4 영역 에는 많은 추상신경세포가 있고 축삭은 추체로를 따라 내려가서 근육 을 지배하는 운동신경과 접합을 이룬다(그림 3.15). 추체로는 교차하므 로 왼쪽 운동피질의 엄지손가락 영역에서 기시한 축삭은 반대쪽 경수 의 운동신경으로 가서 오른쪽 엄지손가락을 움직이게 하는 근육을 지 배하는 운동신경을 이룬다. 모든 운동신경의 지배는 이와 같다 (P hillip s 1971). 그림 3.7 은 관절의 신근(E)과 굴근 (F) 의 운동신경 지 배를 나타낸 것 이다 이 간략한 모식도는 수백 개의 신경세포 또는 신경섬유를 마치 한 신경세포인 양 나타내었다. 또 그림 3.7 과 3 .1 5 에서 보여주듯이 근 수축은 수용체를 활성화하여 근육은 수축할 때 수용체를 홍분시켜 근
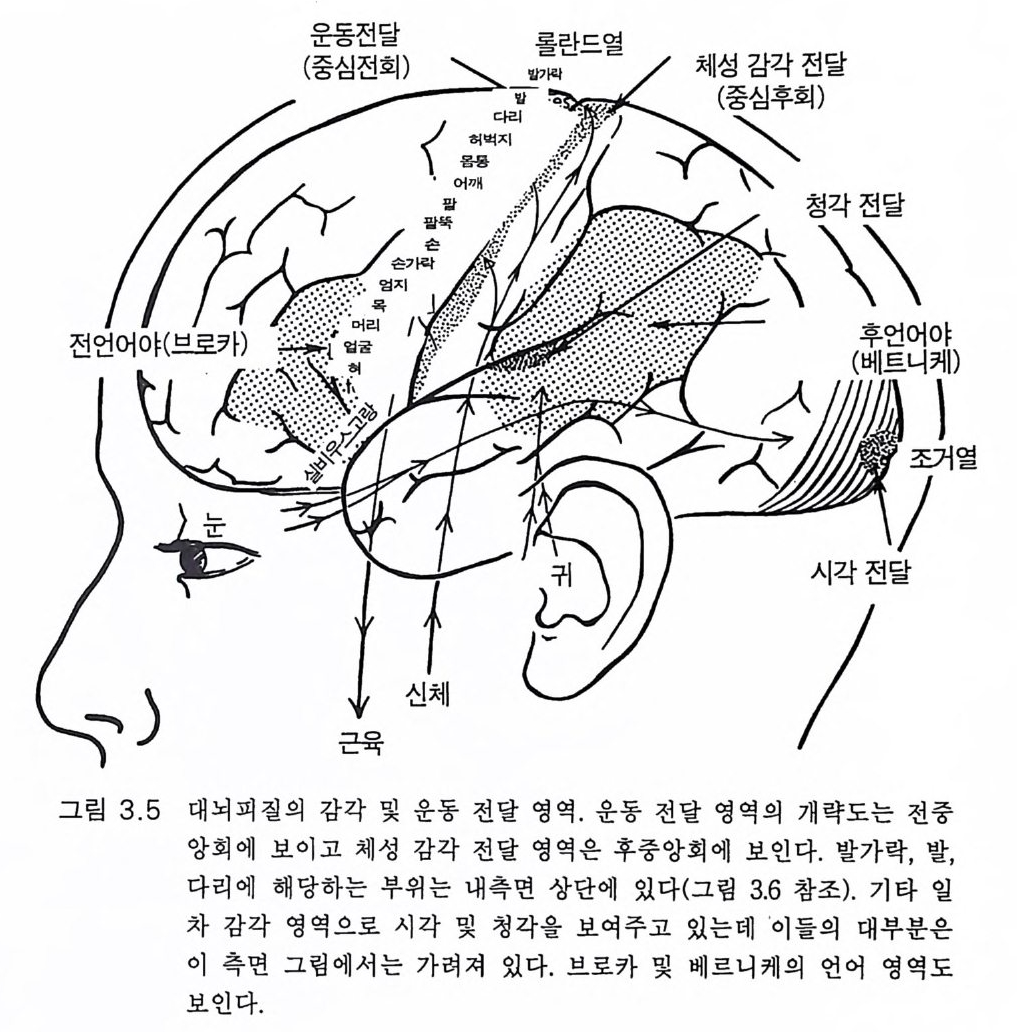 롤란드열 체성감각전달
롤란드열 체성감각전달
수축이 일어나고 있다는 정보를 중추신경계에 보낸다. 그림 3 . 5 와 3.1 5 에서는 이 같은 구심성 섬유를 Ia 로 나타내었다. 간단한 운동을 하고자 할 때 예를 들면 발가락으로 서려고 하면 대 뇌피질의 부수 운동 영역 (SMA) (그림 3.3 , 4.3) 같은 고위 중추가 하고자
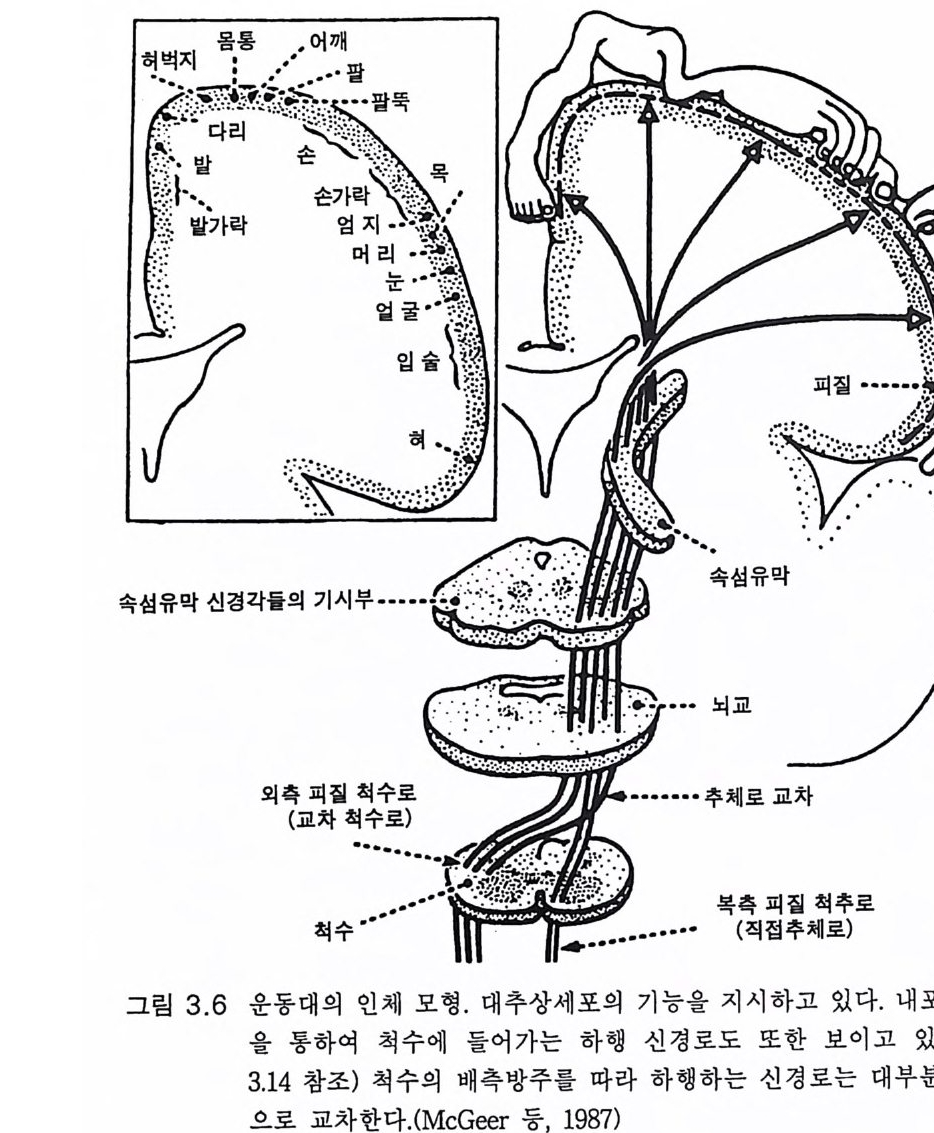 피질 -----
피질 -----
하는 의도에 따라 우선 활성화된다. 이어서 운동피질의 해당 신경세포 가 활성화하여 추체로를 따라 운동신경에 이른다(그림 3.1 5 ). 결국 발목 관절의 신근(장딴지근, 가자미근)이 수축한다. 이는 신경 활동의 기본에
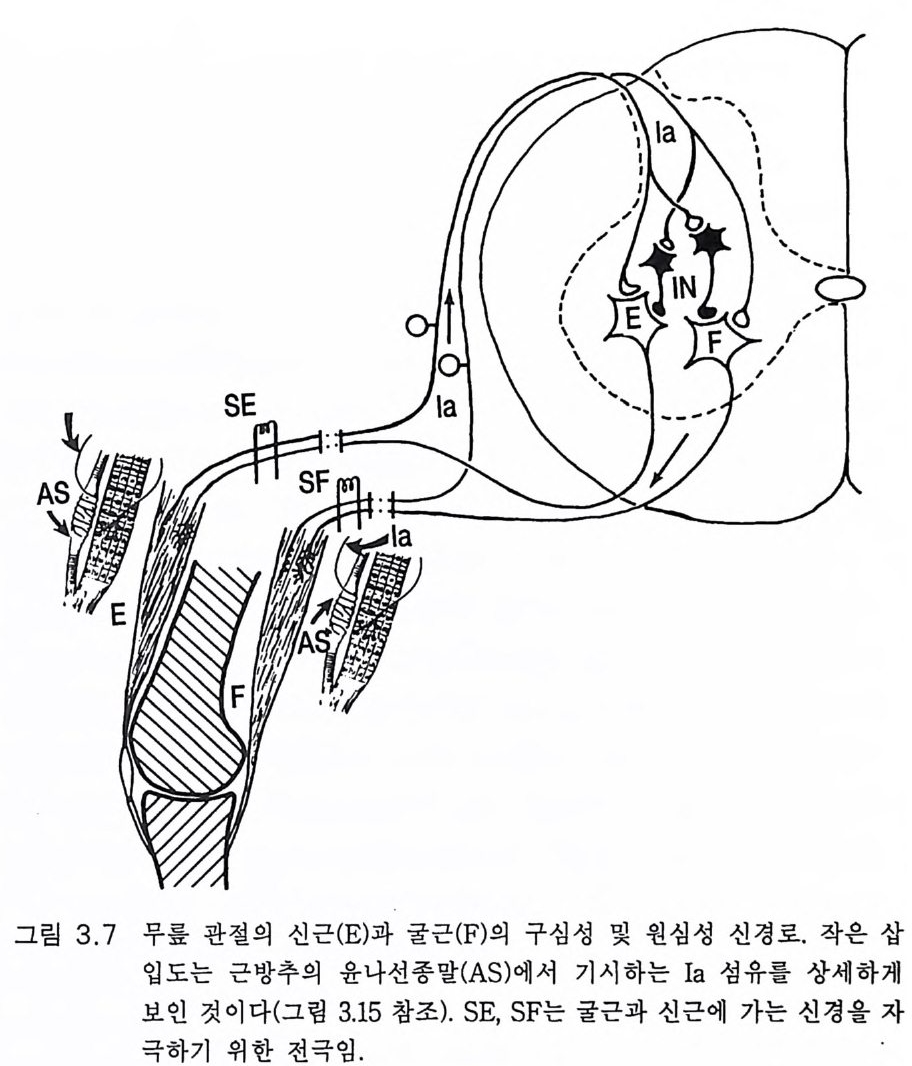 `` ` ` ``
`` ` ` ``
지나지 않고 활동에는 뇌의 기타 여러 계통이 참여하게 된다 . 죽 운동 피질의 추상세포가 추체로를 따라 임펄스를 방출하기 전에 소뇌와 기 저핵은 반응을 조절함으로써 과거의 경험과 현재 진행되고 있는 운동 을 고려하여 서있는 자세가 유지되도록 한다. 추체로 이외에도 척수를 따라 운동신경에 이르는 또 다른 신경로가
있다(그림 3.1 4 ). 그러나 진화과정에서 추체로가 유리하게끔 이 같은 신 경로의 일부는 소멸하였다. 예를 들면 뇌간의 적색핵(그림 3.2) 으로부 터 척수로 하강하는 신경로인 적색척수로는 사람의 척수에서는 대단 히 축소되어 있다 (Na th an 과 Sm ith 1955, Brodal 1981). 전정핵에서 나오 는 전정척수로는 중요한 신경로의 하나이다(그림 3.2 ). 이것은 내이의 전정기구에서 머리의 움직임이나 돌림에 관한 감각 정보를 전달한다 . 또 한가지 사람 뇌의 대뇌척수로는 뇌간의 그물핵에서 기시하는 그물 척수로이다(그림 3.2, 3.1 4 ). 이것은 운동신경에 작용하기 전에 척수내의 한 개 또는 그 이상의 신경계를 통하여 정보를 전달하는 것이다 (Na th an 과 Sm ith 1955, Brodal 1981). 뇌의 고위중추에서 기시하는 이 같은 신경로에 더하여 근수축 및 운 동이 수행되는 동안 순간 순간의 정보를 뇌로 전달하는 신경로가 있다 . 사람의 신경계 활성에 관한 지식들은 혈관 손상이나 총상 등 우연 한 뇌 손상의 효과를 연구하여 얻어진 것들이다. 그러나 이들 손상은 과학적 연구에는 대단히 적당치 못하다. 아직까지는 사람 뇌의 이해는 다른 영장류를 이용한 실험적 연구에 의존한다. 조절된 조건에서 사람 뇌의 기능을 밝히는 모델로 이용되는 것이다. 포유동물에서 사람으로의 뇌의 진화를 고려할 때 좀더 단순한 포유 동물 뇌로부터 호미노이드 조상을 포함한 고등 영장류에 이르기까지 많은 진화가 이미 일어났을 것으로 생각된다. 호미노이드로부터 호미 니드를 거쳐 일어나는 진화적 발전이 특히 우리가 다루고자 하는 홍미 의 대상이 된다. 특히 관심의 대상이 되는 것은 양각 보행, 무기와 도 구를 생산하는 미세한 손놀림, 언어 표현에 관계되는 인두, 혀, 후두, 호홉근의 움직임이다. 이것에 관하여는 제 4 장에서 다룰 것이다.
3.3 직립, 보행 및 반응 3.3.l 골격 진화 (Washburn 1978) 골격 변화의 증거로 직 립자세라는 중요한 진화적 발달을 알아볼 수 있다고 처음 제안되었다. 화석 증거만으로는 호미노이드로부터 양각 직립자세를 취하는 오스트랄로피테신에 이른다고 생각되는 변화 과정 울 알아보는 데 충분치 못하다. 그래서 네 다리로 서고 걷는 현대의 유인원(꼬리 없는 원숭이) 골격을 원시 호미노이드 조상 오스트랄로피 테쿠스 아프리카누스 골격의 모델로 이용한다 . 화석 증거는 사람 골격 에 근사한 골격 발달이 어떻게 이루어지는가를 보여준다. 그림 3 . 8 은 HSS 와 침팬지의 몸통과 골반 관계를 보여주는 것이다 (Schultz 1968). 사람의 척추는 머리와 몸통의 무게를 수직으로 관골절 구로 전하여 대퇴골에 체중이 실리게 된다. 침팬지는 장골이 길어서 네 다리로 서는 데 적합하다. 초기 호미니드의 화석 기록을 꿰어 맞추 어 비교하는 것은 불가능하다. 그러나 그림 3 . &-e 에서 오스트랄로피 테쿠스 아프리카누스의 골반골은 HSS 의 것(그림 3.8e) 과 매우 유사하 고 침 팬지 의 골반골(그림 3.&) 은 그림 3.8a 와 유사함을 보여 준다. 오스 트랄로피테쿠스 아프리카누스의 척추와 골반 관절이 HSS 의 것(그림 3.8b) 과 유사함을 추측게 하는 훌륭한 화석 (그림 3.9a) 이 있다. 화석 기 록에 의하면 호미니드의 뒷다리 긴뼈들(대퇴골, 경골, 비골)은 HSS 와 유사하고 원형이 놀랍도록 잘 유지된 호모 하빌리스의 발 화석(그립 3.9b) 은 사람의 발과 매우 닮았다. 특히 첫째 발바닥뼈가 유사하다. 이 것은 가장 튼튼한 것이고 다른 발바닥뼈들과 가지런히 놓여 있고 비호 미니드 영장류의 발과 같이 벌어지지 않았다. 이들 화석 기록으로 미루어 호미니드 오스트랄로피테쿠스 아프리카 누스의 척추, 골반 및 뒷다리의 골격 구조는 이미 직립자세에 적합하
 (c)
(c)
그림 3.8 (a) 와 (b) 는 성인과 성숙 원숭이의 척주의 만곡도와 골반뼈의 위치와 크기 이 다 (Schul tz , 1968). (c),(d) 및 (e) 는 오른쪽 요골로서 (c) 는 침 팬 지, (d) 는 오스트랄로피테쿠스 아프리카누스, (e) 는 현대인이다 . 각각 온 장골면을 직각으로 보이게 하였고 전상장골극이 오른쪽으로 보이 게 한 것이다. 사람과 오스트랄로피테쿠스의 장골은 원숭이에 비하여 옆으로 퍼졌고 원숭이의 장골은 좁고 아래 위로 커져 있다 .(Tob i as 1983)
도록 발달되었음을 알 수 있다. 그러나 머리와 척추를 연결하는 관절 은 아직 충분히 발달되지 못하였다(그림 3.1 0 ). 직 립자세를 취할 때의 고릴라 머리는 중력의 중심( i )이 척추 관절( f )보다 훨씬 앞쪽에 있 댜 따라서 가끔 직립자세를 취할 때 머리를 바로 지탱하기 위해서는 목뒤 근육의 강력한 수축이 필요할 것이다 . 오스트랄로피테쿠스 아프 리카누스의 경우는 그 같은 불균형이 훨씬 덜해서 목뒤 근육이 강할 필요가 없었을 것이다. 이 같은 경향은 진화 과정에서 호모 에렉투스 (그림 3 .1 0c) 를 거 쳐 호모 사피 엔스 사피 엔스(그림 3 .1 0d) 로 이 어 진다 (Tobia s 1983). 3.3.2 양각 보행과 민첩성 여기서 사람의 양각 보행을 상세하게 논할 필요는 없다고 생각한다 (Napi er 1967 참조). 양 다리는 교대로 움직여 한쪽 다리가 앞으로 나갈 때 다른 쪽 다리는 체중을 지탱한다 . 움직이는 다리는 구부려 지면에 서 떼고 발뒤꿈치로 땅을 차면서 걸음을 내딛는다 . 이때 체중을 앞으 로 옮기고 곧이어 지지하던 다리는 긴엄지발가락굽힘근의 수축으로 엄지발가락으로 땅을 차면서 움직이는 다리가 된다. 걸음을 걸을 때 다리가 움직이는 기간은 60% 이고 지지하는 기간은 40% 이다. 따라서 보행시 25% 는 양 다리가 동시에 지면에 닿아 있다. 빨리 걸을 때는 이 기간이 짧아진다. 그러나 뛸 때와는 달리 걷는다는 것은 최소한의 기간 동안은 양 다리로 지지하는 것을 특징으로 한다 . 양각 보행은 발 뒤꿈치로 내차는 것으로 시작하여 엄지발가락으로 밀어올리는 것으로 끝난다. 물론 더 얌전하게 걸을 수도 있다. 보행시 중간둔부근과 작은 둔부근은 수축하여 체중이 실리는 쪽으로 골반을 꽉 린다. 동시에 골 반을 회전시켜 보폭을 길게 한다. 이 양각 보행 걸음걸이는 사람 특유의 것으로 지면 위룰 효과적으
 '`,1,
'`,1,
로 어동하는 방법인 것이다. 이에 반하여 폰지드는 잠시나마 양각 보 행시 절뚝거린다. 진화적으로는 몇 가지 해부학적 변화가 있다. 팔 길 이에 비한 다리 길이의 증가, 골반이 짧아지고 넓어짐(그림 3.&,d), 고 관절 근육의 조정, 중간둔부근과 작은둔부근 대신 큰둔부근이 고관절
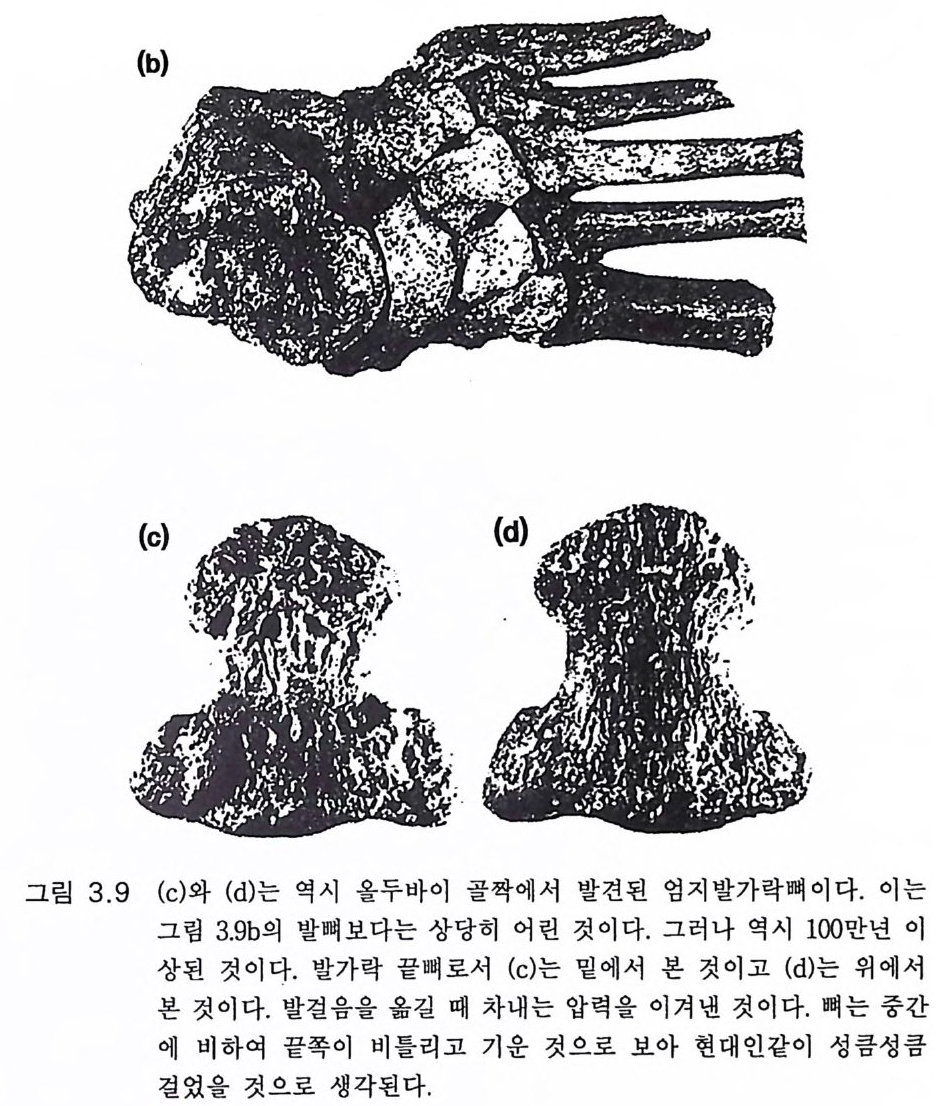 그림 3.9 (c) 와 (d) 는 역시 올두바이 골짝에서 발견된 엄지발가락뼈이다 . 이는
그림 3.9 (c) 와 (d) 는 역시 올두바이 골짝에서 발견된 엄지발가락뼈이다 . 이는
펴 짐 기 능을 담당함, 발 모양의 상당한 새 모습(그립 3.9b), 척 추의 요 추 부위 전방 만곡과 골반의 장골 부위 전방 회전으로 체중이 직접 관 골절구와 대퇴골에 실리 게 함(그림 3.8 b ). 이상과 같은 모든 진화적 변 화의 관찰에도 불구하고 대체로 호미니드는 비틀거리는 양각 보행을 했을 것으로 믿어지고 있다 (Ash t on 1981). 레톨리의 발자국은 오스트랄
 (a)
(a)
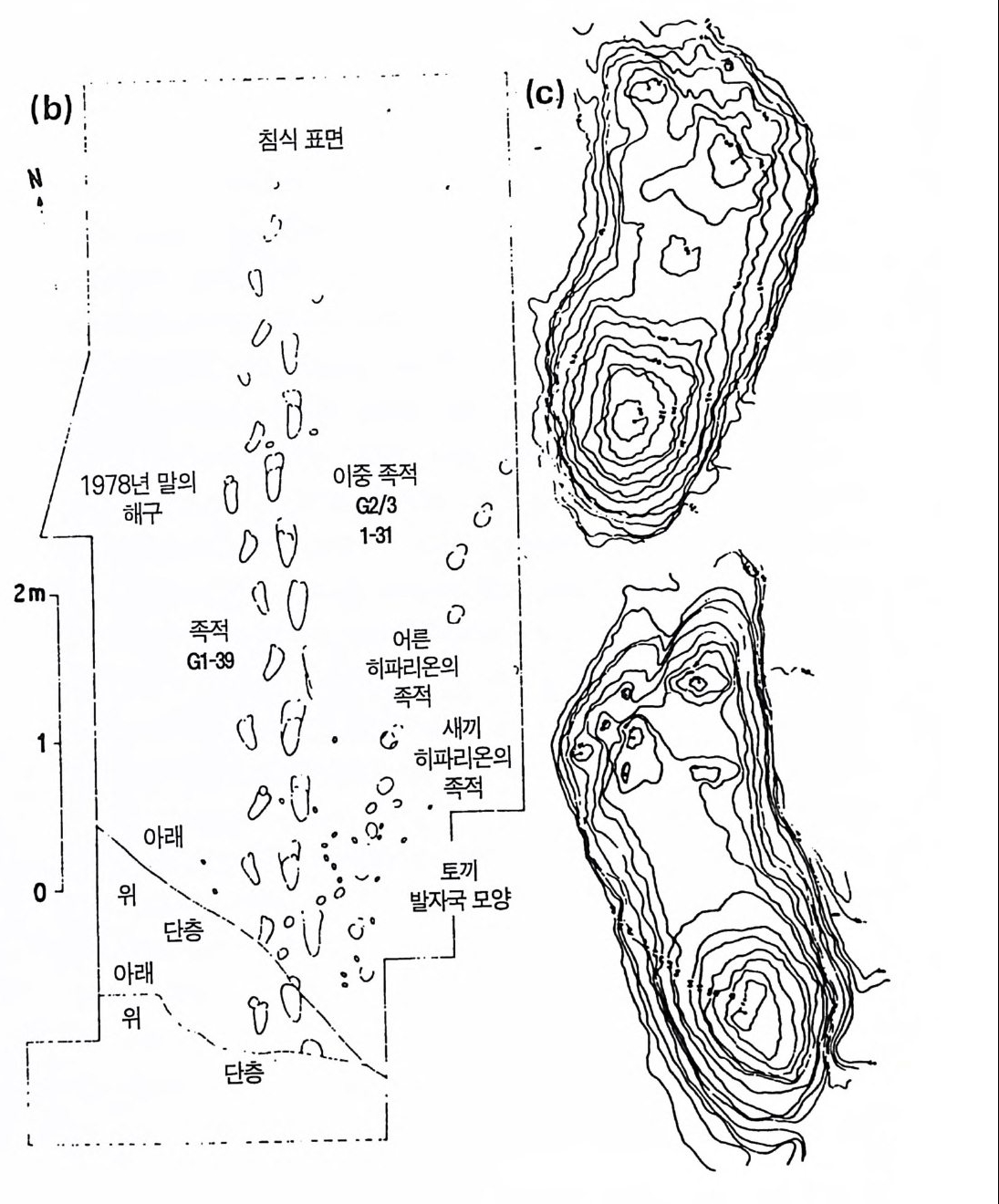
로피테쿠스 아프리카누스와 그 이후의 모든 호미니드가 사람 모양으 로 양각 보행을 했음을 결정적으로 훌륭하게 보여주는 것이다 (Tu ttl e 1985). 오스트랄로피테신이 남긴 가장 훌륭한 화석은 북탄자니아의 레톨리 하상에서 발굴된 독특한 발자국이다(그림 3.1 1 , Leake y와 Hay 1979, Leakey 1981). 이 들 발자국 화석 은 신 경 학자들에 게 3.6-3.7 my B P 때 의 호미니드 조상의 초기 역사를 말하여 준다. 이 연대는 화산 응괴암의 K-Ar 방사성 연 대 측정 법 으로 밝혀 진 것 이 다(D rake 등 1980). 그 같은 화석은 화산 폭발과 함께 대단히 특이한 기상학적인 조건에서만 생길 수 있댜 첫째 나트리오카르보나타이트 na tri ocarbona tit e 와 메릴리타이 트 me lilitit e 용암괴의 성분인 화산재가 수 센티미터 떨어져 쌓이고 그 위에 발자국이 만들어졌다 . 이것은 수용성 탄산염들이 비에 녹아나와 서 뜨거운 햇빛으로 결정을 만들면서 급속하게 굳었다. 그러고 나서 두번째 화산 폭발이 발자국을 남긴 응괴암을 덮어 더 이상의 발자국이 생기지 못하게 하였고 수백만 년 동안 침식과 풍화를 막았다 . 그림 3.lla,b 의 발자국은 잘 훈련된 양각 보행을 나타내고 있으며 정 상적인 좌우 발의 위치와 사람과 유사한 큰 엄지발가락을 보인다. 키 가 130-150cm 되는 두 호미니드의 것일 것이다. 특기할 것은 한 사람 이 앞서간 다른 사람의 발자국을 정확하게 따라밟고 가는 형상이다. 세번째 것은 좀 작은 것으로 약간 삐뚤빼뚤하여 손을 잡고 옆에서 걸 어간 것으로 생각되는 것이다. 그림 3.llc 는 세번째 것의 사진제도법으 로 그린 발자국이 다(D a y 1985). 이 발자국은 의심할 여지없이 오스트랄로피테신의 잘 배운 양각 보 행을 나타낼 뿐 아니라 인간관계를 나타내는 것이다. 화석은 오스트랄 로피테신이 서로 손을 잡고 걸었으며 인도자의 발자국을 정확하게 밟 으며 따라갔음을 보여주는 것이다. 이로서 마치 우리가 파도에 씻긴 부드러운 모래 위를 걷듯이 360 만년 전에 오스트랄로피테신의 한 가족
 그림 3.11 (a),(b) 레톨리 지역 G 에서 발견된 호미니드의 족적 .(Leake y, 1981)
그림 3.11 (a),(b) 레톨리 지역 G 에서 발견된 호미니드의 족적 .(Leake y, 1981)
 (Nb)‘ I’’’’ . . • • --·침 - -拜 면 ·· 1':i': '_ c' -
(Nb)‘ I’’’’ . . • • --·침 - -拜 면 ·· 1':i': '_ c' -
이 새로 형성된 화산재 위를 산책하는 광경을 상상할 수 있다. 그림 3.1 1 의 발자국을 현대 인간이 맨발로 걸은 발자국과 비교하면 모든 면에서 똑같고(D a y 198. 5) 폰지 드의 발자국과는 분명 히 다르다 (Tu ttl e 198. 5). 분명히 오스트랄로피테신은 능숙한 양각 보행을 하였다. 호미니드 화로의 첫번째 커다란 진화적 발달을 완전하게 이룩한 것이다 . 네발 자세와 보행에서 양각 보행으로 기능이 변하는 데 따르는 골격의 진화 적 변화도 있었다(그림 3.8, 3.9 , 3.1 0 ). 그림 3 .1 1 의 확실한 보행은 의 지 하지 않고 똑바로 선 자세를 유지하면서 걸을 수 있었음을 말한다. 발자국 화석이 보여주듯 앞선 사람의 발자국을 정확하게 밟고 따라 가는 행위는 정신 집중을 요하는 행위이다. 능숙한 자동 운동에서 다른 발자국을 정확하게 밟고 가는 능숙한 수의적 조절로 발전한 것이다. 때때로 직립자세로 뒤뚱거리는 양각 보행을 하던 유인원의 둔중한 네발 걸음에서 그림 3 .1 1 에서와 같은 산뜻한 두발걷기로 가는 데는 엄 청 난 진화적 발달이 있었다 (Na pir e 1967). 골격 진화를 수반하여 사지 근육 특히 고관절 주위 근육의 변화가 있었다. 유인원과는 달라서 고 관절 근육, 중간둔부근과 작은둔부근은 보행 운동시 굴신운동에 따르 는 고관절의 동요를 제한함으로써 고관절을 안정시키는 작용을 한다 . 고관절의 펴는 기능은 큰둔부근이 커지면서 떠맡게 된다 . 오스트랄로 피테쿠스 아프리카누스의 골반 위의 근육 부착 부위는 이 같은 근육의 변화가 일부 이루어졌음을 보여준다 . 따라서 그들은 사람과 마찬가지 로 산뜻한 걸음걸이를 하였을 것이다. 엄지발가락의 중요한 역할도 그 림 3.llc 의 족적에 나타나 있다. 3.3.3 양각기립 네발걷기에서 두발걷기로 변하려면 신경계의 재조정이 있어야 한 다. 그러나 불행히도 양각기립에 관계되는 신경기구에 대한 지식은 충
 무릎전략 엉덩이 전략
무릎전략 엉덩이 전략
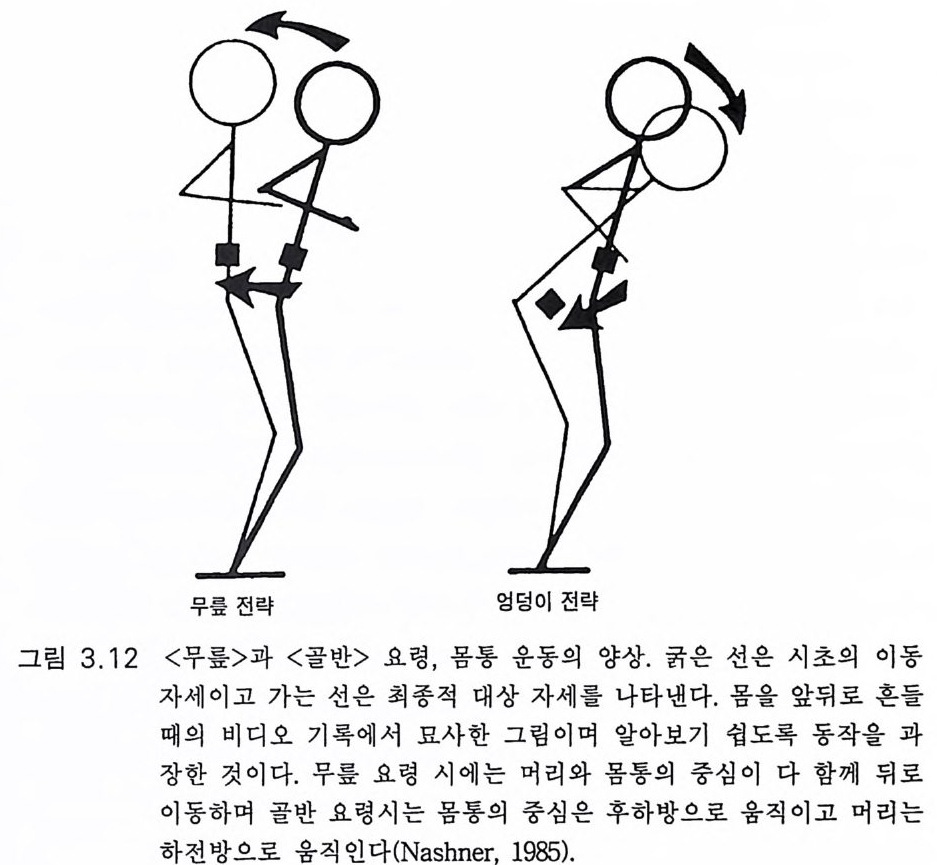
분치 가 못하다. 양각기 립 에 관한 당대 의 권위 자 내 시 너 (Nashner 1985) 는 여러 가지 상황에 대처하여 사람이 직립자세를 유지하게 하는 복잡 한 기전에 대한 이해는 이제 시작에 불과하다고 말하였다. 그림 3 .1 2 에 간단한 실험을 보여준다. 앞뒤로 움직일 수 있는 실험대 상이 평면 위에 서있다. 인위적으로 몸을 앞뒤로 혼들 때 실험대상이 몸을 원 위치로 유지하려는 전략을 알아보기 위하여 몸통과 사지의 여 러 근육에서 활동 전압을 측정하였다. 몸의 중심을 지지하고 있는 발
의 중심보다 뒤쪽으로 움직였을 때의 반응을 목표로 하였다. 실험대상 은 두 가지 전략 중 하나를 선택하게 된다. 무릎 전략 ankle s tr a t e gy에 서는 무릎펴점근들이 먼저 수축하여 그림에서 굵은 실선이 가는 실선 위치가 되었을 때처럼 다리를 뒤쪽으로 움직이게 한다. 그러고 나서 무릎 굽힘근과 하체의 근육들이 그림에서 굵은 화살표로 나타낸 것처 럼 평면 위에서 몸의 중력 중심이 제자리를 찾도록 한다. 이때 머리도 뒤로 움직인다. 고관절 전략hip s tr a t e gy에서는 몸이 많이 흔들리고 바 닥이 고르지 않을 때로서 첫째로 고관절을 많이 구부려 몸을 굽히고 머리를 앞으로 움직여 중력 중심이 제자리를 찾게 한다. 이 같은 간단한 보정에도 수용체와 중추신경 활성에 많은 일이 일 어난댜 대단히 복잡한 일이 특히 머리의 위치와 회전 신호에 관계되 는 시각과 전정감각 기전이 관여한다. 전정은 머리 위치의 신속한 적 옹에 관여하고 시각은 머리 위치의 안정적 유지에 관계된다 (Nashner 1981). 영장류 중에서 사람만이 두 다리로 직립하므로 직립 자세에 영 향을 미치는 여러 가지 뇌 손상을 일으켜 실험한다는 것은 기술적으로 나 윤리적으로 매우 곤란하다. 죽 대뇌피질, 소뇌 또는 시각계, 전정계 에 실험적 손상을 사람에게 일으키는 것은 곤란하다. 양각 보행과 민 첩성에 관계되는 이들 문제는 다음에서 재론한다. 적어도 오스트랄로 피테신이 직립자세를 취하게 되는 데는 대단히 복잡한 중추신경계의 변화가 있었어야 했음은 확실하다. 3.3.4 신경기구 양각 보행 운동(그림 3.11) 은 뇌의 신경조절 기구의 변화가 있었음을 나타내는 것이다. 그러나 사람의 양각 보행의 신경조절에 관하여는 상 세하게 아는 바가 별로 없다고 말할 수밖에 없다. 챗바퀴돌리기나 선 택적 손상을 이용한 사지보행에 관한 과학적인 연구가 거의 전부이다
(Gr illn er 1981 ). 쳇바퀴돌리기 같은 인공적인 실험실 조건이 아닌 실생활에서의 동 물의 이동운동에 관한 기초적인 문제가 무시되어 왔다고 그릴너 (Gr illn er 1981) 는 지적하였댜 예를 들어 도로에서의 통상 보행에는 잘 발달된 감지능력이 있어야 한다. 특히 시각과 순간순간의 의지적 보행 운동 조절 능력이 있어야 한다. 놀랍게도 보행에 관한 신경계 연구는 척수, 뇌간, 기저핵 또는 소뇌에 관심이 집중되어 있고 대뇌 운동피질 이나 추체로의 역할은 무시되고 있다. 운동불능증을 주중상으로 하는 파킨슨씨 병에 있어 기저핵의 병변은 보행의 시작을 대단히 어렵게 한 댜 정상적인 보행시 조절 또는 조정 감각신호는 소뇌와 기저핵으로부 터의 신호와 함께 대뇌피질로 입력되는 것을 그림 3.13 에서 보여준다 (D eLon g과 Georgo p ou los 1983). 어쨌든 보행운동은 대뇌피질의 적절한 추상세포들에서의 율동적 방전의 순차적 폭발에 의한 근수축으로 이 루어진댜 그렇지 않으면 우리가 군중 속이나 낯선 지역을 걸을 때처 럼 한발짝 한발짝을 수의적으로 조절할 수 없을 것이다. 사지와 체간 근육의 조절을 상세하게 설명한 사람 근육의 대뇌피질 조절에 관한 훌륭한 종설이 프로인드 Freund 와 훔멜샤임 Hummelsh eim에 의해 1985 년에 출판되었다. 그림 3 .1 4 에 두 가지 중요한 하향 신경로를 보이는데 피질-망상 척수로와 추체로로서 주로 운동피질(브로드만 제 4 영역)에서 기시한다(그림 3.5 , 3.6). 전자는 대체로 전운동피질에서 기시 하여 뇌간으로 하향하여 그곳에서 큰 망상세포와 연접하고 척수의 양 옆을 달리는 두 개의 하향 신경로를 이룬 것이다(그림 3.1 4 ). 척수에서 체간과 상지근을 지배하는 운동신경을 활성화한다. 후자는 연수에서 교 차하여 추체로를 따라 하강하고 반대편 하지근을 지배한다. 양각 보행 에서의 한걸음 한걸음은 말단 하지근육의 하나인 긴엄지굽힘근의 수축 으로 시작되므로 추체로를 따라 하향하는 운동피질에서의 임펄스 방전 이 양각 보행에서 주역을 담당함에 틀립없다. 기타 하지근도 물론 운동
 'IIIIII_'
'IIIIII_'
피질이 활성화한다. 피질-망상 척수로는 체간과 고관절 근육의 운동 조 정을 담당할 것으로 생각한다. 그러나 보다 철저한 연구가 필요하다 . 비센댕거 (W i esendan g er 1981b) 는 추체로증후군이라고 분명하게 구 분하는 추체로의 임상적 병변 증례를 모았다. 이들 증례에서는 병변이 추체로에 국한되어 있었고 이완성 사지마비가 후에 경직성으로 변하 였다. 비센댕거는 잔류 추체로라는 특별한 예에 대하여 기술하였는데 나머지 부위는 모두 망가졌는데도 환자는 수의운동을 할 수 있었다. 추체로의 부분 병변시에는 운동력의 상실이 예상보다 적었다. 그러나 여러 개의 추체로 신경섬유가 하나의 운동신경을 담당하므로 자동적
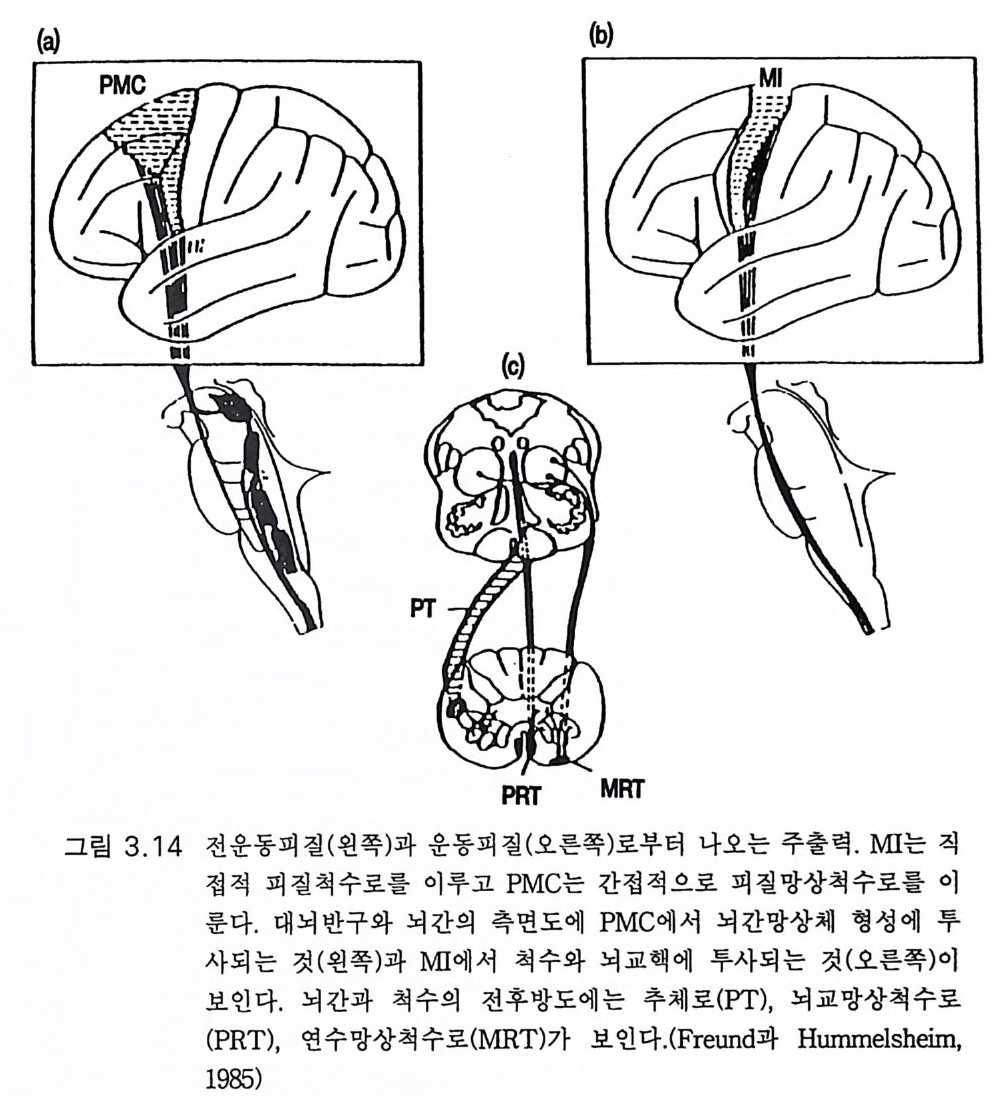 이{b)
이{b)
으로 보상될 수 있다는 사실을 상기 할 필요가 있다 (P hillip s 와 Port er 1977). 사람과 영장류 추체로의 비교 특히 타워 (Tower 1940) 의 고전적 연구로부터 비센댕거 (1981b) 는 대뇌피질 병변에 의한 결손에 대하여 단지 부분적으로만 추체로가 관계된다고 결론지었다. 또 자세와 운동
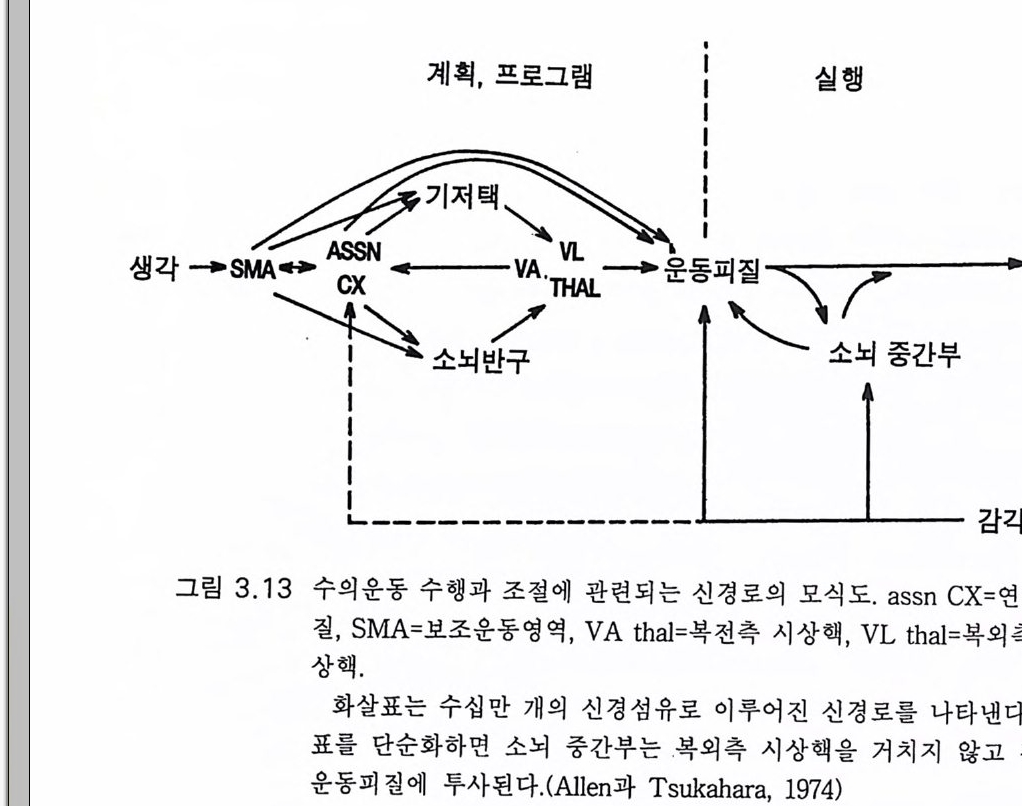
의 양분은 가공된 것이라고 결론지었다. 자세의 조절도 능숙한 운동만 큼이나 정밀성이나 복잡성이 다같이 관계된다. 양자가 모두 추체로 같 은 신경조절계의 빠른 전달을 필요로 한다. 여기서 사람이 걷는 동안의 비복근의 반응에 관한 멜빌 존스 Melv i ll J ones 와 와트 (Wa tt 1971a) 의 특기한 연구를 소개한다 사람이 아래로 걸어내려갈 때 발이 지면에 닿는 순간의 135msec 전에 비복근이 활성 화(근전도 EMG) 되었댜 보폭을 크게 하면 EMG 는 발이 지면에 닿는 것보다 더 일찍 시작되고 더 커졌다. 결국 비복근의 수축은 체중을 지 탱하려는 준비인 것아다 . 발이 지면에 접촉한 후 다리를 뻗기 위한 근 육의 반응이 그림 3.7 의 회로를 따른다면 이는 너무 느리고 적절치 못 하댜 멜빌 존스와 와트 (1971b) 그리고 윌슨 (W i lson 1983) 은 다음과 같 이 제안하였다. 아래로 내려가는 걸음 초기에는 머리의 움직임이 내이 의 평형석 기관을 활성화하여(전정 수용체 기전) 망상척수로와 전정신 경로를 거쳐 비복근을 수축시킬 것이다. 그러나 내려서는 시각 신호로 운동피질도 활성화될 수 있다 . 평형석 의 역할은 뜻하지 않은 자유낙하의 효과로 연구되었다 . 비복근의 EMG 는 74msec 의 잠재기후에 시작되었댜 이것은 15cm 이하의 짧은 거리를 자유낙하할 때의 충격을 완충하기에는 너무 느린 것이다. 결국 뒤꿈치에 괴로운 충격을 받게 된다 . 심지어 2.5cm 의 자유 낙하일지라 도 대단히 괴롭다 . 자유 낙하로 인한 중력 자극의 순간적 소실에 의한 이 같은 평형석의 자극은 연속적 펌뛰기 때의 근육운동을 설명하는 데 도 고려되어야 한다 (M~lv ill J ones 와 Watt 1971a). 원하는 펌뛰기 빈도를 근육수축으로 성취하는 데 다음 세 가지 요인이 고려되어야 한다. 예 상되는 착지시간과 다음 뛰어오르기의 시작이 되는 기능적인 뻗기반 사 (FS R) (Chan 둥 1979a), 그리 고 뛰 어 오르기 가 시 작되 었을 때 의 무중 력 상태의 개시에 대한 전정반옹이다. 이들 요인은 율동적 보행을 설 명할 때에도 고려되어야 하는 것이다 . 이 모든 것은 대뇌중추의 기능
임은 말할 것도 없댜 사람의 율동적 보행운동은 척수기전으로는 설명 이 어렵다 그리고 망상척수로 또는 전정척수로에 전정자극을 전달하 는 하위 대뇌기능만으로는 부분적인 설명만이 가능하다. 그림 3 .1 3 에서 지적했듯이 보행운동은 기저핵, 소뇌, 기타 대뇌 여러 부위가 서로 고리를 이루는 활성의 영향을 받는 운동피질의 율동적 방 전을 기본으로 한다. 이 모든 신경기전은 오스트랄로피테신에 있어 네 발걷기에서 두발걷기로의 운동 기능의 진화로 개조되었다. 어쨌든 네 발걷기의 기전은 다리를 교대로 움직이도록 조절하는 기전을 설명하 는 데 응용된다. 이제 다시 3.6m y BP 에 두 발로 걸었다고 생각되는 세 개의 발자국 (그림 3.11) 에 대하여 살펴보기로 한다. 이중 두 개는 단순한 양각 보행 으로서 계속적인 수의적 조절은 필요치 않았던 것으로 생각된다. 그러 나 세번째 것은 인도자의 발자국을 정확하게 밟고 따라간 것이었다. 이는 잘 조절된 행동으로서 주로 시각에 의한 감각 입력의 조절을 받 은 운동피질의 신호 방출이 추체로를 통하는 수의적 조절을 필요로 하 는 것이다. 이것은 오스트랄로피테신의 보행에 중요한 의미를 부여하 는 것이나 그림 3.11 의 틀에 박힌 보행은 다양한 지면 위를 걷는 양각 보행의 정상 범위에 속한다고 하겠다. 그림 3.11 은 두 다리로 움직였던 사냥, 전투, 공동생활터로 식량을 동점으로 나르는 등 모든 활동에 적 용되었을 것으로 생각할 수 있다. 그림 2.5 에서 지적했듯이 오스트랄로 피테쿠스 아프리카누스는 그림 3.11 의 기록이 만들어지기 전 최소한 100 만년 동안 존재했었다. 이는 뼈, 관절, 근육 동이 양각 보행에 적합 하도록 진화적 변화를 일으키기에 충분한 시간이다(그림 3.8, 3.9, 3.10). 오스트랄로피테쿠스 아프리카누스는 특기할 변화없이 그림 3.11 에 보여주는 것과 같은 방식으로 보행운동을 계속하였다. 이를 제 1 장에서 진화적 정체라고 하였다. 그림 2.5 에는 약 2.5m y BP 에 가지치기에 의한 종말을 보여준다. 오스트랄로피테쿠스 아프리카누스는 그 긴 기간을
통하여 뇌는 약간의 증가만을 보였다. 즉 조상으로 생각되는 호미노이 드의 뇌 평균 용적이 350cc 에서 450cc 로 늘어났다 . 두 다리로 걷게 되 는 데는 틀림없이 상당한 중추신경계 재구성이 따랐을 것이다. 그러나 두 다리로 걷는 데는 네 다리로 걷기보다 단순한 신경기전이 필요했을 것이댜 네 다리로 걷는 데는 네 다리 모두가 시간적 및 공간적 상태 에 적합하도록 조절되어야 한다. 그러나 네 다리의 지지는 기립시의 안정도를 높인다. 전호미니드는 현재의 영장류의 것과 마찬가지로 대 단히 효과적이고 복잡한 운동조절계를 이미 가졌다 . 이는 어느 포유류 급 보다도 다양하고 잘 조절된 운동기구이다. 오스트랄로피테쿠스 아 프리카누스의 조절된 민첩성은 오늘날의 훈련되지 못한 사람 수준이 었을 것으로 생각된다. 오스트랄로피테쿠스 아프리카누스는 석기를 만들어 쓰지는 않았다 고 생각한다. 그러나 뼈나 뿔 또는 나무 둥을 도구로 사용했을 것으로 생각되나 이것들은 화석으로 남아있지 않은 것이다 (Leake y와 Lewi n 1977). 그들은 사냥이나 전투에 사용하는 원시적인 무기, 곤봉, 창 등을 나무를 다듬어 만들었을 것으로 생각한다. 이 같은 손재주의 발달은 생존을 위하여 대단히 중요하였을 것이다. 오스트랄로피테쿠스 아프리 카누스는 동아프리카와 남아프리카에 널리 퍼져있었지만 아직도 유라 시 아에 존재 했다는 화석 의 증거 는 없다 (Tob i as 1983). 3.4 운동의 미세 조절을 위한 신경기전의 발달 우리의 모든 운동의 섬세한 조절은 말초 수용체로부터의 되먹이기 전에 의한다는 사실은 대단히 중요하다. 만일 사지로부터 중추신경계 로의 감각 입력이 차단된다면 그 사지의 움직임은 감퇴하거나 혼란에 빠지게 된다(D enn y Brown 1966). 피부나 관절의 수용체로부터의 감각
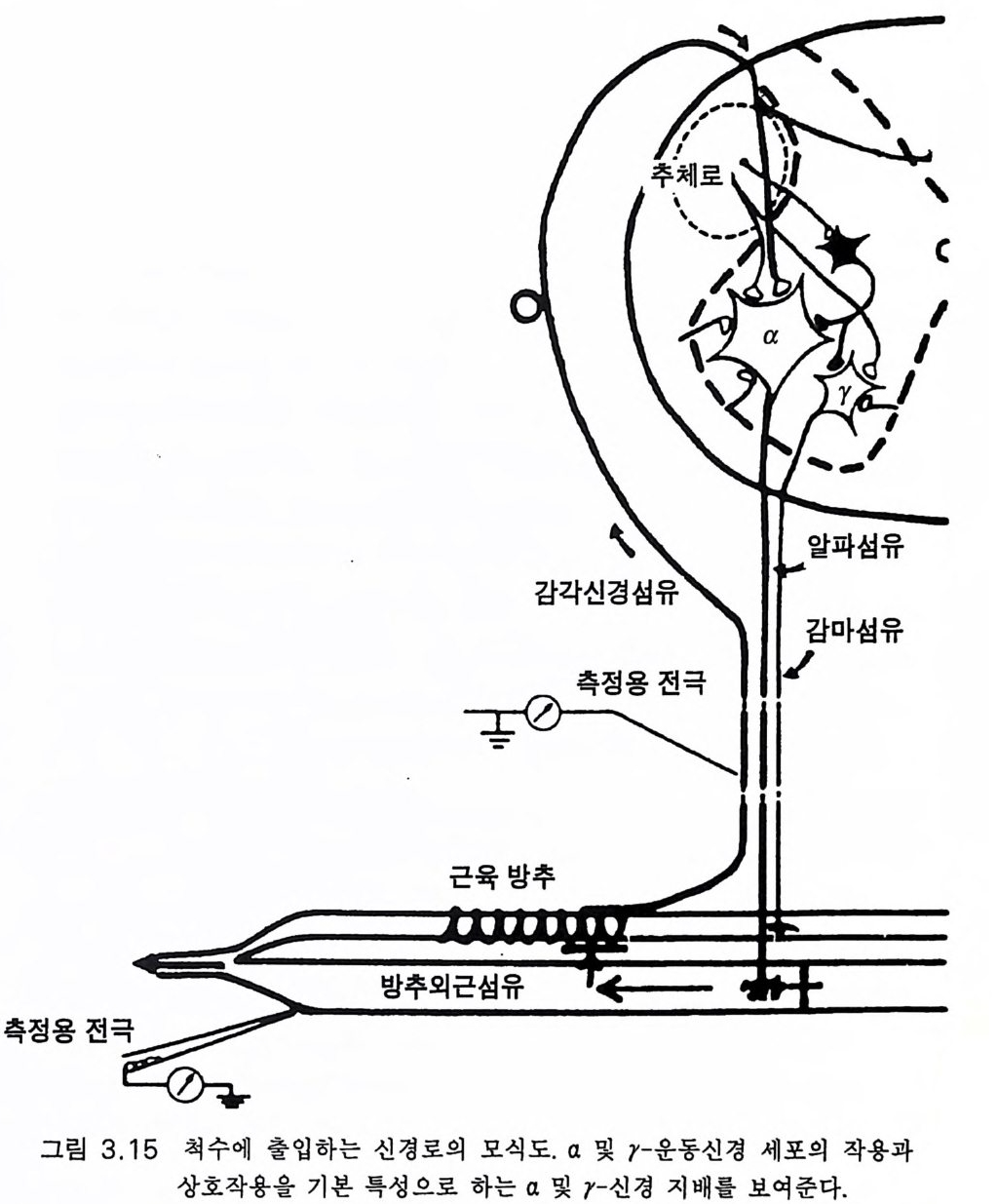
입력은 중요하다. 그러나 더욱 중요한 것은 근육 수용체로부터의 감각 입력이다 (Ma tt ews 1981). 그림 3 .1 5 의 개념도에서 근방추의 근육 섬유 를 둘러싸고 있는 윤나선상종말 annulos pir al en di n g을 볼 수 있고 이것 의 신경방전은 이미 그림 3.7 에서 보였던 Ia 군의 구심성 섬유를 따라 전달된댜 몇 개의 근방추내 섬유는 근방추에서 함께 다발을 이루어 분명한 두 가지 무리를 만든다. 작은 구심성 섬유(제 2 군)로 된 이차 종 말은 윤나선상종말의 큰 Ia 구심성 섬유보다 작다. 그러나 이 책에서 설명하려고 하는 것은 그림 3.15 의 개념도로 충분하다고 생각한다. Ia 섬유는 한 번의 접합으로 운동신경을 지배한다(그림 3.7). 근육의 건을 그림 3 .1 5 에서 왼쪽에 보이는 화살표 방향으로 끌어올 리면 근방추의 흥분으로 발생된 임펄스가 Ia 섬유를 따라 방출된다. 근 방추내 섬유가 홍분하게 되면 r- 운동신경 임펄스에 의해 수축이 일어 나고(그림 3.1 5 ) 윤나선상종말의 강력한 홍분으로 a- 운동신경의 단일접 합 활성이 증강된다. 한편 a- 운동신경의 방전으로 방추의 근섬유가 수 축하게 되면 이와 평행하게 있는 근방추의 장력이 떨어져 윤나선상종 말의 방전은 감소하거나 없어진다. 그러나 r ―운동신경이 a- 운동신경 과 동시에 방전하게 되면 근방추가 수축하여 늦추어지지 않는다. 이같 이 자동제어 기전이 잘 짜여져 있다 . 구심성 섬유 la 에 반응하는 a- 운 동신경의 신호발사 강도가 클수록 방추의 근섬유 수축은 강해지고 소 위 r- 고리 loo p에 의한 a- 운동신경의 la 활성은 작아진다. 그런데 이 고 리의 작용은 넓은 범위의 r- 운동신경 방전 정도에 의해 변동될 수 있 댜 따라서 y-운동신경 자극에 의한 변동을 조정하는 근수축의 자동제 어 고리 조절기전이 있는 것이다 (Gr anit 1970). 최근까지도 근방추로부터의 구심성 Ia 입력은 척수 수준에서 이 같 은 자동제어 고리 조절에 국한한다고 생각하였다. 그런데 필립스 (Ph illips 1969) 는 근육에 서 뇌 로 올라가는 구심 성 입 력 은 고위 자동제 어로서 작용한다고 제안하였다. 원숭이에서 la 군의 신경섬유는 주로
운동피질 바로 뒤에 있는 감각피질의 제 3a 영역으로 뻗어있음이 밝혀 졌 다 (Ph illip s 등 1971, Maendly 등 1981). 그 림 3.16 에 서 척 수라고 표시 한 화살표는 VPLc 시상의 심부 성분으로 가는 신경섬유들이고 거기서 시 상-피질 섬유는 제 3a 및 제 2 체감각 영역으로 가는 신경과 접합을 이 룬다 . 영장류와 기타 고등 포유류에서는 Ia 섬유가 척수로 들어갈 때 두 갈래로 갈라진다(그림 3.15). 가지 하나는 척수의 동일 높이에서 동 측성 운동신경을 포함하는 여러 신경으로 가고(그림 3.7) 또 한 가지는 동쪽 기둥을 따라 올라가서 뇌간의 외측쐐기다발핵에서 접합을 이루 고 내측 섬유띠라고 하는 신경로에 의해 VPLc 의 심부덮개로 간다(그 림 3.1 6 ). 근자에 정상인에 있어 근육의 이들 구심성 신경섬유의 작용기전에 관한 대단히 중요한 연구가 있었다 (Chan 1983). 예를 들면 발목 관절을 갑자기 등쪽으로 구부리면 척수반사로 장딴지근 수축을 일으킬 뿐 아 니라 기능적 신전반사 (FSR) 라고 하는 강한 지연성 수축을 유발한다. 두 가지 반사반응의 지연시간은 척수반사의 경우 평균 37msec 이고 FSR 의 경우 108msec 이다 (Chan 등 1979a). 감지되는 신전에 대한 가장 빠른 수의적 반응 속도는 평균 157msec 이다. 이는 강하고 긴 FSR 의 개시 시간보다 현저하게 느린 것이다 . FSR 의 지연시간은 근육 신전으 로 발생된 구심성 입력이 대뇌, 소뇌 둥 고위 중추로 올라갔다 내려오 는데, 즉 필립스 (1969) 가 제안했던 고위 중추의 자동제어 조절기전이 운영되기에 충분한 시간이다. 찬 동 (Chan 둥 1979a) 은 신경회로의 길이가 짧아지면 지연 시간도 짧아진다는 사실을 들어 이 같은 FSR 에 있어서의 긴고리 신경회로를 증명하였댜 예를 들면 팔의 두갈래근의 지연시간은 평균 50msec 이고 다리의 네갈래근은 67.8msec 이다. 척수반사인 건반사와 FSR 사이의 시간적 차이는 척수의 감지 중추로부터 고위 중추까지의 거리로 설명 될 수 있다. 이 시간적 차이는 두갈래근은 평균 32msec 이고 네갈래근
 c
c
 L심 피L 부부 수모대
L심 피L 부부 수모대
은 47msec, 장딴지근은 73msec 이다 FSR 이 긴고리 반사회로라는 또 한 가지 증거는 척수 손상시 그 손상 부위 이하에서는 FSR 이 나타나 지 않는다는 것이다 . 또 찬 등 (1979b) 은 발이나 발목의 피부와 관절의 완전한 부분 마취로도 FSR 은 변하지 않음을 관찰하였다 . 따라서 긴 고리 반사는 근육으로부터의 구심성 자극에 의함을 알 수 있고 이는 주로 근방추의 구심 성 신경 에 의 한 것 이 다(그림 3.1 5 , 3.1 6 ). 찬 등 (1979a) 은 이 같은 긴 지연시간이 운동중추를 왕복하는 예상되 는 긴고리 신경회로의 자극 전도시간에 의한다고는 생각하지 않았다 . 신경의 Ia 섬유를 전기적으로 직접 자극했을 때의 반응에서 훨씬 짧은 긴고리 지연시간도 관찰되었다. 예를 들면 아일스(Il es 1977) 는 사람의 비골신경울 자극하여 건반사처럼 29msec 의 지연시간을 갖는 H 반사와 55 msec 의 긴고리 반사를 관찰하였다(그림 3.1 7 d). 긴고리 반사는 운동 피질을 통한 신경회로에 의한다고 생각되는데 이는 안정시에는 0 이고 수의적으로 일정하게 수축하는 동안 비골신경이 자극되었을 때 반응 이 상당히 커지기 때문이다. 비슷한 현상은 가자미근에서도 관찰되는 데 그림 3 .1 7e 에 보이는 것처럼 건반사와 긴고리 반사는 각각 31 과 57 msec 의 지연시간을 나타내었댜 이 같은 짧은 긴고리 지연시간은 운 동피질로 올라가는 신경회로의 전도시간으로 간주되는 것으로 Ia 임펄 스가 동쪽 기둥핵으로 올라가는 동쪽 섬유단은 대단히 전도 속도가 빠 른 신경섬유이기 때문이다. 마르스덴 등 (Marsden 등 1W8a,b, 1983) 은 사람의 긴엄지굽힘근에 일 으킨 반사반응에 관한 기발한 연구룰 하였다. 이 근육은 오로지 엄지 손가락의 끝관절을 굽히는 데에만 관여한다. 굽힘근이 수축하고 있는 상태에서 갑자기 힘을 가하여 펴게 하면(그림 3.17a) 근육은 일련의 반 사반옹을 나타낸다(그림 3.1 7 b). 제일 먼저 그림 3.7 의 반사회로에 의한 통상의 건반사가 25msec 의 지연시간을 가지고 나타난다. 연이은 두 가지 반응은 지연시간이 평균 40 과 55msec 인 것으로 역시 자동적인
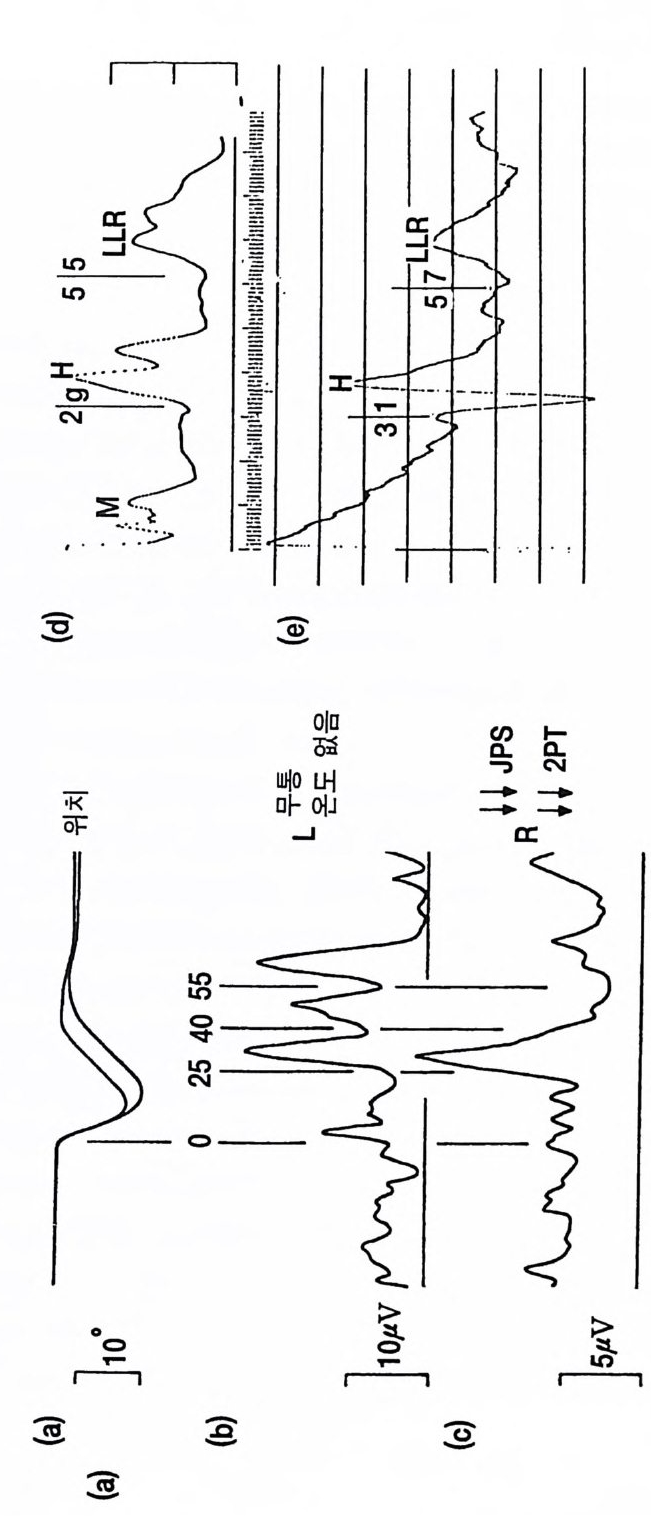
그림 3.1 7 (a)-(c) 우측 뇌간의 병변이 있는 환자에서 장모지굴근의 빠른 단기신 전에 대한 반옹 . 이 환자는 왼쪽 팔의 통각 및 온각의 소실, 오른쪽 부위의 관절 위치감각, 진동감각 및 촉각 판별(JP S 및 2PT) 의 소실 이 있었으나 운동장애는 없었다 . (a) 우측 및 좌 측 엄지의 자세각도의 기록 (b), (c) 좌측 (b) 와 우측 (c) 손의 장모지굴근에서 기록된 정류근 전도의 완전파형. 엄지는 2N 의 고정된 힘을 유지하도록 하였다. 수직 선으로 표시한 시간 0 때에 30N 의 힘을 30msec 동안 가했다. 각각의 기록은 24 번 시행한 것의 평균이다. 좌측 장모지굴근에서 기록한 근 전도에는 척수단 접합 잠복기 (25 msec) 에 분명한 반응을 보인다. 그 리고 후기 반응들은 40 및 55msec 에 있다. 우측 장모지굴근의 기록 에 서 는 척 수단 접 합 반옹은 나타났으나 LLRs 은 분명 치 않았다. (Marsden 둥, 1987b) (d) 사람의 비골신경을 자극하고 경골전근에서 기록한 것이다. 자극은 운동신경 섬유에 대한 역치 이상을 가했으며 이는 초기 M - 반응으로 나타났고 이어서 H- 반사(잠복기 29msec) 가 있고 LLR( 잠복기 55msec) 로 이어진다. LLR 은 수의적 배경 수준에 의존적이고 안정시에는 0 이다.(Il es, 1'57 7 ) (e) 경골신경을 운동신경 섬 유에 대한 역치 정도로 자극한 것이다. H- 반사는 31msec 의 잠복기 롤 가지고 초기 양성파를 보이고 LLR 의 잠복기는 57msec 이다. LLR 은 발목 관절에 대한 운동피질이 자극되도록 발가락으로 서서 몸을 앞으로 기울였을 때 나타났다. 자극 빈도는 3Hz 였고 50 번의 반옹을 평균한 것이다 .
것이며 수의운동에 의한 것이 아니다. 이들 40 과 55msec 의 자동적 반 옹은 고위 중추 특히 운동피질로부터의 긴고리 반사 (LLR) 라는 임상적 증거도 있다 (Marsden 둥 1978a,b). 운동피질로 오가는 긴고리 반사회로 에 병변이 있으면 이 반사는 없어지거나 억제된다(그림 3.1 7 c). 그러나 소뇌핵(개재 및 치아핵)도 또한 55msec 반응에 관계된다. 곧이어 갑자 기 펴는 데 대한 의식적 반응이 있다. 이 반응은 강한 저항이거나 이 완인데 그 지연시간은 평균 126msec 로서 일반적인 반응속도 reac ti on tim e 보다는 짧은 것 이 다 . 이 등 (Lee 등 1983) 은 사람의 손목굽힘 근을 갑자기 펴는 실험으로 비슷한 LLR 을 관찰하였다 . 6)
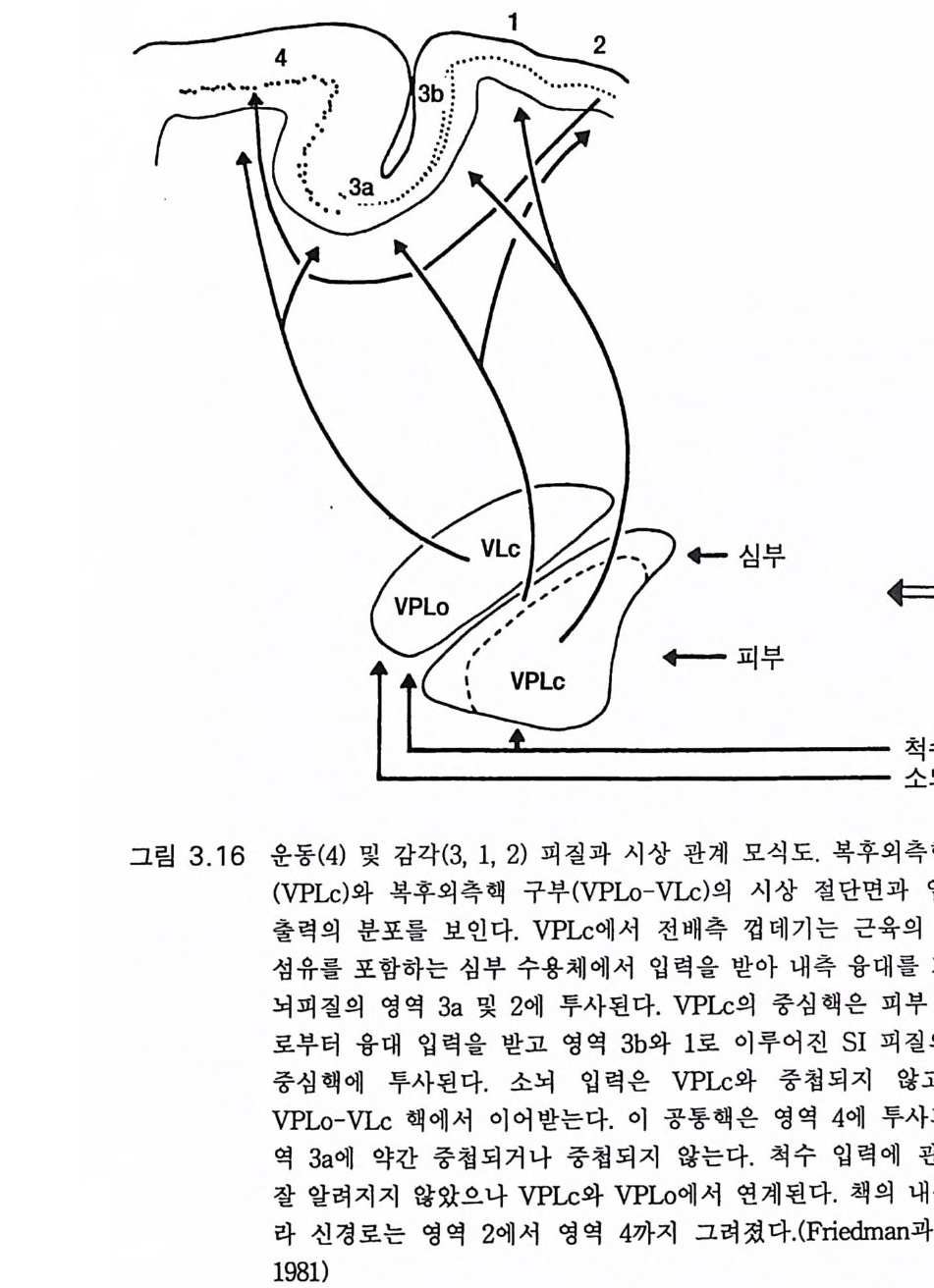
고위 중추 특히 운동피질의 자동제어 기전인 긴고리 반사는 수의적 운동에도 점점 중요하게 취급되는데 이는 실제로 손재주 를 나타내는 효 과적인 운동조절 기전으로 생각되기 때문이다. 이 를 비센댕거 W i esen dan g er 와 마일스Mi les 는 다음과 같이 잘 표현하고 있다. 운동의 자동제어 기전의 원리는 운동의 실제 행위를 계속적으로 재고 또 의도하는 그리고 원하는 행위와 비교하는 것이다. 의도하는 행위와 실 제 행위 간에 어떤 차이가 있게 되면 그에 상응하는 오차 신호가 만들어 지고 이는 자동적으로 오차 를 보상하는 명령신호 를 발하게 한다 . 대뇌피 질에 걸쳐 있다고 생각하는 자동제어 기전에 있어 사지 를 움직이는 동안 에 일어나는 혼란은 자극 강도에 병행하는 강도로 근육의 신전 수용체를 활성화한다. 이 오차 신호는 운동신경으로 전달된다. 결국 M 2 지연시간에 근수축을 변화시켜 애초에 가해진 혼란을 보상하게 된다(1 982:1256). 움직이려고 할 때에는 우선 근수축의 적정한 강도에 대한 판단이 서야 한다. 이 예측에 오차가 있으면 Ia 를 포함한 근방추 수용체가 활 성화하게 된다. 손 움직임에서 예측의 오차가 생기면 이룰 수정하기 위한 LLR 이 일어나서 운동피질에서의 신호 발생이 적절하게 변하여 50msec 이내의 지연시간을 가지고 손 움직임의 수정 반응이 일어난 다. 발목 신전에 대한 건고리 수정반사 지연시간은 약 70msec 일 것으 로 생각된다. 이 같은 수정 보상은 자동적이고 무의식적인 것이다. 만 일 존스(J ones 1983) 가 제의했듯이 피질 수준에서의 Ia 신경로가 제 2 영 역을 거쳐 운동피질로 간다면 잘 짜여진 설계라고 할 수 있다. 포웰과 마운트캐 슬 (Powell 과 Mountc a stle 1959) 은 관절 이 나 근막으로부터 의 모 든 심부 감각 입력은 제 2 영역이 담당함을 관찰하였다(그림 3.16). 이 같 은 입력이 Ia 입력과 어울려 운동피질을 통한 자동제어를 효과적으로 일으키기 위해 제 2 영역에서 제 4 영역(운동피질)으로 필수적인 정보를
전달하는 것이라면 옳은 관찰이었다고 생각된다 . 또 VPLc 에서 제 3a 영역으로 가는 신경로는 근수축을 감지하는 것이고 제 2 영역으로 가서 제 4 영역에 연계되는 것은 자동제어 기전이라고 생각한다. Ia 수용체에서 운동피질로 가서 긴고리 반사에 참여하게 되는 신경 로는 고양이 같은 포유류에서도 알려져 있다 (Land g ren 동 1984). 운동 반옹에 있어 건고리 조절은 호미노이드에서부터 발생하였을 것임은 이미 지적하였다 (P hillip s 등 1971). 여기서 제안하려고 하는 것은 호미 니드의 이 긴고리 조절기전의 점진적 진화는 호미니드의 손재주를 이 루는 데 필수적이었을 것이라는 점이며 이는 석기시대를 거치는 동안 더욱 뚜렷해진다(그림 6.10). 비센댕거 (W i esendan g er 1981a) 는 보닌 Bo nin을 인용하여, 원숭이의 제 4 및 제 6 브로드만 영역은 비슷한 크기이나 사람의 제 6 영역은 제 4 영 역보다 6 배나 크다고 하였다(그림 4.4 ). 이같이 제 6 영역이 상대적으로 크다는 것은 부시 (Buc y 195. 5), 펜필드와 자스퍼 (Pe nfi eld 와 Jas pe r 1954) 등도 지적하였다. 피질의 제 6 영역이 크다는 것은 HSS 의 고급 운동조 절이라는 면에서 중요성이 증대된다 (Ful t on 1949, Wi es endang e r 1981a). 따라서 제 6 영역의 손상은 운동 실조와 시각운동 기능 수행의 장해를 초래한다 (Freund 와 Hurnmelsheirn 1985). 그림 3 . 6 은 또 다른 진화적 발 달을 개념적으로 보여주는 것이다. 손가락들을 담당하는 운동피질이 대단히 커져 있다. 이는 다음에 기술하는 섬세한 손재간의 진화적 발 달과 관계가 있는 것이다. 3.5 능숙한 손 움직임 오스트랄로피테쿠스 아프리카누스가 두 발로 걷게 됨으로써 또 다 른 큰 장점을 얻게 되었다. 앞다리(팔)는 보행의 의무로부터 해방되어
다양하고 능숙한 활동에 이용할 수 있게 되었다. 이는 손의 변화에서 뚜렷하게 나타나는데 엄지는 발달하고 회전하여 엄지와 손가락 끝으 로 정확하게 잡을 수 있게 된다. 그 이전까지는 폰지드나 사람은 손으 로 움켜쥘 때 손바닥을 구부리는 힘으로 잡았다 . 그림 3.1 8 (Nap ier 1962a,b) 에 도시한 화석 증거로 보면 호미니드의 엄지 (e) 는 호미노이드 의 조상으로 간주되는 어린 고릴라 (d) 로부터 거의 사람의 엄지(f)에 가 까워지는 이행 단계라 할 수 있다. 호미니드의 엄지는 그림 3 .1 8a,b,c 에 나타난 것처럼 뼈의 강도와 킬이 그리고 손가락 끝의 넓이가 발달하였 다. 이는 모두가 다른 손가락을 마주 대하기 쉽게 하는 변화이다. 주의 깊은 탐사에도 불구하고 오스트랄로피테쿠스 아프리카누스가 3.6m y BP 에 살았고 거기서 두발걷기가 잘 발달하였다고 생각되는 레 톨리 하상에서는 원시적인 석기조차도 발견되지 않았다(그림 3.11, 3.1 9 )(Leakey 1981). 기 타 오스트랄로피 테 쿠스 아프리 카누스의 거 주지 역에서도 발견되지 않았다 (Tob i as 1971). 그러나 1.9 my B P 시대인 올두 바이 퇴적층 인접 지역에서는 다양한 석기, 죽 도끼, 정, 다면체형 , 원 판형, 편구형 긁기 석기들이 발견되었다(그림 3.1 0 a). 모두가 용암석으 로 만들어졌고 자르는 데 사용하였을 것으로 보이는 석영암으로 만든 예리한 칼날도 발견된다 (Leake y 1981). 이 같은 1. 9m y BP 의 조잡한 석기 제작은 그곳에서 발견되는 좀더 발 전한 호미니드 즉 650-670cc 의 뇌를 가진 호모 하빌리스의 화석과 함 께 발견되 는 수도 있다 (Leake y와 Lewi n 1977). 3.6m y BP 의 오스트랄로피테쿠스 아프리카누스는 다듬은 석기를 만 들 만큼 충분히 발달하지는 못하였던 것 같다. 그들의 뇌는 능숙한 손 재주로 석기를 만들 만큼 발달하지 못하였을 것으로 추측할 수 있다. 중거로는 오스트랄로피테쿠스 아프리카누스의 손이 힘있게 잡고 석기 를 다듬는 데 적합했어야 한다. 폰지드조차도 잘 조절된 악력을 가졌 다. 모자라는 것은 손재주라기보다는 대뇌의 능력이라 생각된다.
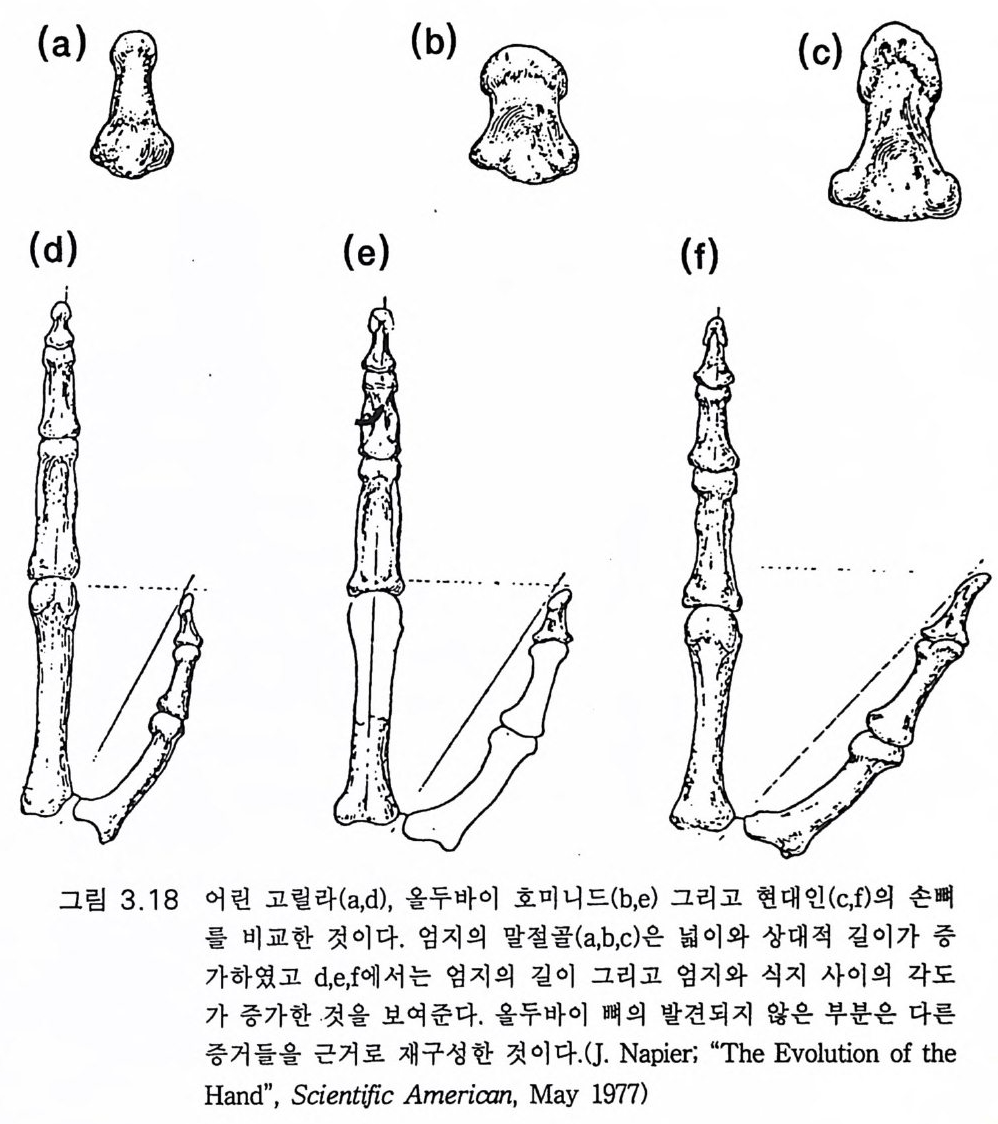 {a) (b)
{a) (b)
300 만년이라는 오랜 동안 존재했음에도 불구하고 오스트랄로피테쿠 스 아프리카누스가 석기제작 기술을 개발하지 못하였다는 것은 놀라 운 일이다 .
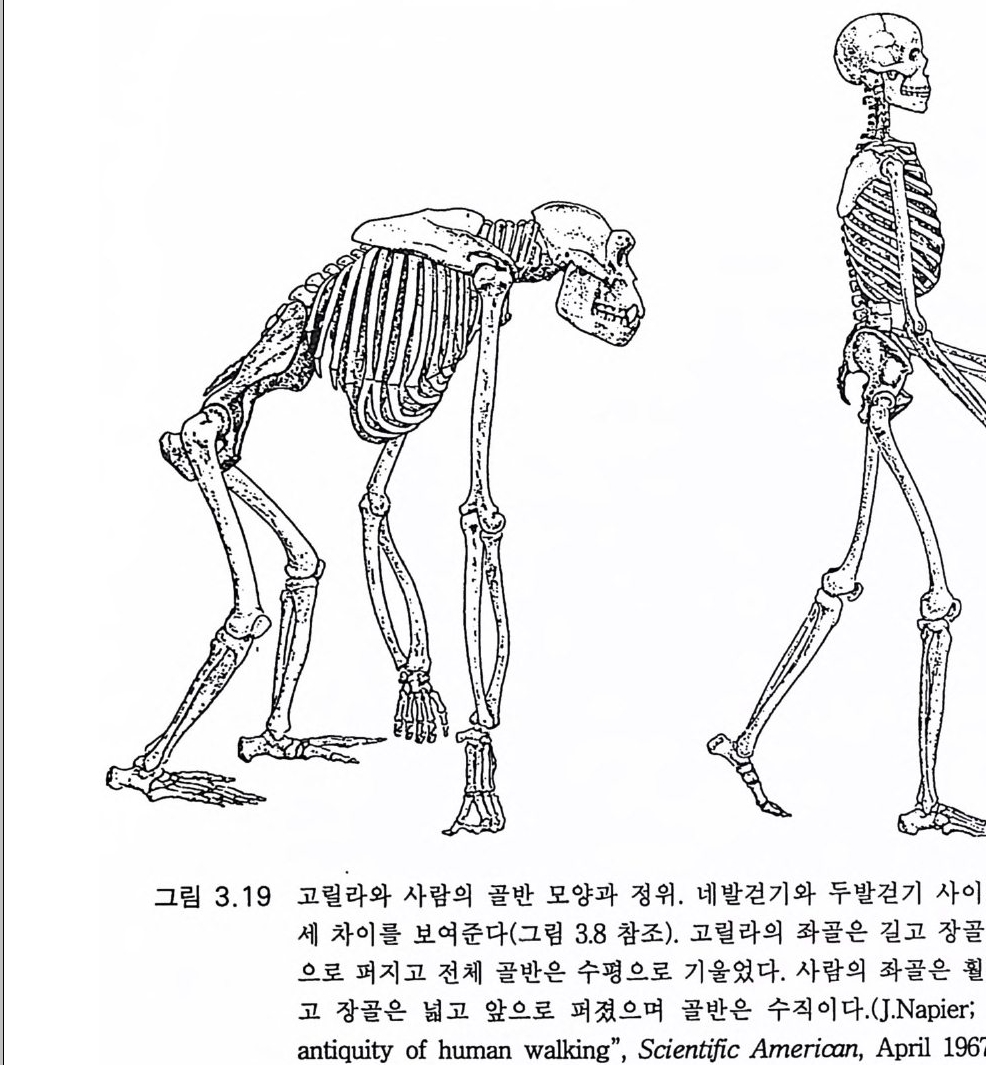 그림 3.19 고릴라와 사람의 골반 모양과 정위. 네발걷기와 두발걷기 사이의 자
그림 3.19 고릴라와 사람의 골반 모양과 정위. 네발걷기와 두발걷기 사이의 자
그러나 그들은 뼈, 뿔, 치아로 만든 도구 (Tob i as 1971) 가 있었으며 현 재의 원시종족이 사용하는 것 같은 원시적인 무기 또는 파내는 데 사 용했을 곤봉 또는 창 모양의 조잡한 목제 도구는 사용했을 것으로 생 각되나 나무는 화석이 되지 못하여 모두 부패하여 없어졌다 (Leake y와
Lewi n 1977). 이 같은 나무 도구도 오스트랄로피 테쿠스 아프리 카누스 의 생존력을 향상시켰을 것이다 . 원시적인 손도끼를 만들 때 이것을 만들기 전에 만들어질 모양을 마음에 그려야 한다 . 호모 하빌리스는 제한된 범위이긴 해도 이 같은 능력을 가졌다고 추측된다. 호모 하빌리스가 호모 에렉투스로 그리고 호모 사피엔스 네안데르탈리스에서 HSS 로 진화하면서 각 단계마다의 석기 종류와 멋이 있었다 (6 . 8) . 이는 대뇌 반구의 진화적 발달에 연유 하는 운동 능력과 창의력이 점차로 향상되었기 때문이라 생각된다. 특 히 전운동피질 및 운동피질의 부속 부위와의 연계가 발달함에 의한다 고 생각한다(그림 3.1 , 3.1 3 , 3.1 4 , 3.16, 4.4 ). 손이 정확하게 잡을 수 있도 록 섬세해지는 것은 도구를 효과적으로 사용하는 데 중요하다(그림 3.1 8 ). 와쉬번 (Washburn 1960) 은 호미니드 진화에서 도구의 역할을 강 조했다. 도구를 사용하는 것은 도구의 개량 또는 제조를 유도한다. 도 구 제조에서 호미니드는 예술성을 터득하여 네안데르탈인의 박편문화 또는 크로마뇽인의 칼날문화로 나타난다. 호미니드의 진화는 HSS 의 창의성에서 절정을 이룬다. 유사 이전의 동굴벽화나 조각에서 볼 수 있는 것으로 제 6 장에서 다룰 것이다.
제 4 장 호미니드 전화와 언어 소통 호미니드 진화에서 언어의 역할을 논하기 전에 사람의 언어와 고등 포유류가 구사하는 언어의 구조와 성질을 철학적으로 검토해 볼 필요 가 있다. 폰지드는 우리의 호미노이드 조상들이 사용했던 의사 소통을 검토하기에 가장 좋은 모델이다. 4.1 언어의 수준 뷸러 (Buehler 1934) 는 언어라고 하는 범주에 속하는 모든 것을 다각 적으로 공식화하였고 포퍼 (Po ppe r 1972) 는 이룰 더욱 발전시켰다 (Pop per 1972 제 6 장, Po pper 와 Eccles 1977 제 3 장). 동물의 언어를 사람의 언어와 함께 생각하는 것은 중요하다. 일반적으로 언어 소통에는 발송 자, 소통 방법 및 수납자가 있다. 이는 특수한 기호체계이다. 뷸러_포퍼 분류(그림 4 .1)에 의하면 두 가지 하급언어 (1 및 2) 가 있는 데 이는 동물과 사람이 공통으로 가지고 있는 것이며 두 가지 고급형 은 유독 사람만이 가지고 있는 것이다. 이것에 관한 시비는 뒤에 다룰
기능 가치 (4) 논쟁적 기능 정당성 / 비정당성 (3) 서술적 기능 거짓 /참 } 사람 (2) 신호 기능 효율성 /비효율성 (1) 표현적 기능 나타냄 /못나타냄 그림 4.1 포퍼가 분류한 인간 언어의 네 가지 수준. 수준과 그에 연관된 가치들 . 수준 3 과 4 는 인간에게만 국한된다 .
것이다 한편 두 가지 하급형의 언어는 다음과 같은 특성을 갖는다고 할 수 있다. ® 표현적 또는 징후적 기능 동물은 내적 감정이나 느낌을 표현한다 . 사람은 소리치거나 울거나 웃는 것으로 표현한다. ® 방출 또는 신호 기능 발송자는 그의 징후적 표현의 소통 방법으로 수납자에게서 어떤 반응이 일어나게 한다. 예를 들면 새의 다급한 지저뀜 은 무리에게 위험 신호가 된다. 생태학적 연구에 따르면 사회적 동물 특 히 영장류에는 이 같은 신호가 대단히 다양한 것으로 알려졌다 . 더욱이 사람들 사이 또는 사람과 동물 사이의 소통에서는 무한한 신호법이 있다. 예를 들어 사람과 양치기 개 사이 또는 사람과 말 사이 등에서 볼 수 있 는 것이다. ® 서술적 기능 이는 사람의 소통법의 대부분을 이룬다. 우리는 우리 의 경험을 다른 사람에게 기술한다. 예를 들면 정원에 대한 기후의 영향, 상점 물건들의 품질과 가격, 최근의 여행 경험, 친지 또는 이웃 사람들의 태도, 최근 과학상의 발견 둥 이루 헤아릴 수 없이 많다. 언어의 두 가지 하급 기능은 발성과 관련된 표현과 신호라는 점이 중요하다. 언어의 서술
적 기능의 특징은 실제적 사실이거나 거짓일 수 있는 진술이라는 것이다. 거짓일 가능성을 함축한다. ® 논쟁적 기능 원래 뷸러의 3 원소는 아니었으나 포퍼가 첨가하였다 (Pop pe r 1972, 제 4 장) . 이는 언어의 최고급 기능이다. 이것은 복잡한 성격 을 지니므로 계통발생학적으로 가장 뒤에 발달하였으며 개체발생학적으 로 반영된다. 날카로운 논쟁 기술은 사람의 논리적 사고 능력과 밀접하게 연관된다. 언어의 네 가지 수준은 유아가 어린아이로 발달하는 과정에서 잘 나타난다. 즉 초기의 순전한 표현 수준에서 신호 수준을 거쳐 서술 수 준을 지나 결국 논쟁 수준으로, 단계적으로 각 수준을 극복해 나간다. 언어의 각 수준에는 하급 수준의 언어가 스며든다는 사실을 알아야 한 다 예를 들어 논쟁시 느낌을 표현하고 반대 의견을 뒤집기 위한 신호 룰 하고 실례를 들어 쟁점을 지지할 수 있도록 설명한다. 또한 언어로 표현하는 동시에 몸짓을 더하는데 다만 수준이 1, 2, 3, 4 로 올라갈수록 이들의 중요성은 덜해진다. 4.2 언어적 표현 우선 존재하는 모든 것과 모든 경험을 포괄하는 포퍼의 3 계 철학을 소개할 필요가 있다. 제 1 계는 사람의 뇌까지를 포함하는 물리적 대상 및 상태계이고 제 2 계는 의식의 주관적 경험 또는 의식 상태들이고 제 3 계는 객관적인 의미에서의 지식 세계이다. 이는 언어를 포함하는, 순 전히 사람이 만들어낸 문화의 세계이다(그림 4.2). 주관적 의미에서 생각이라는 말은 정신적 경험 또는 정신적 과정을 뜻한댜 우리는 이를 <사고과정>이라 하며 이는 제 2 계 상태이다. 반면
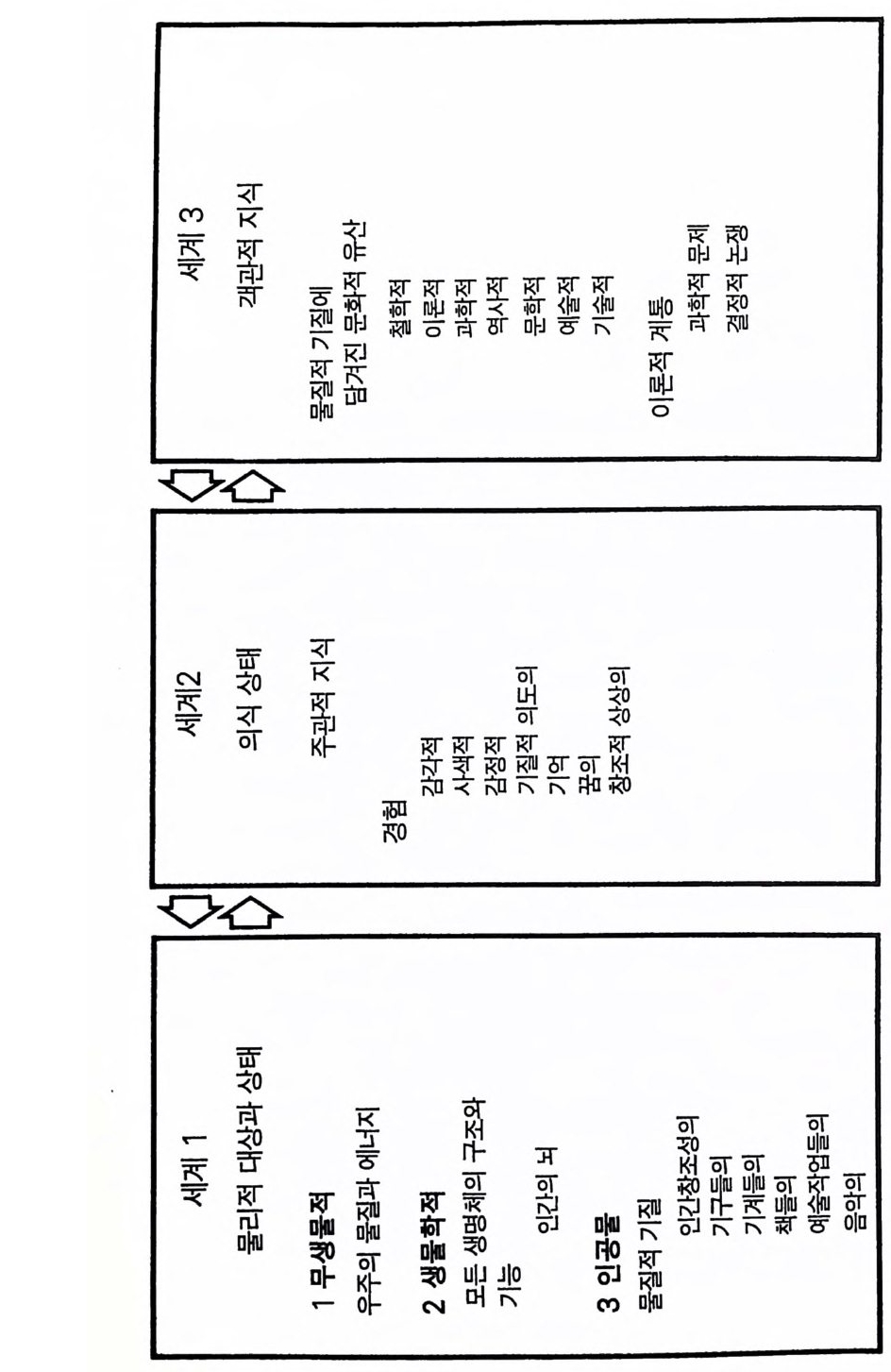
에 사고 과정의 산물로 된 세계가 있다. 즉 사람의 창조성의 세계로서 포퍼의 제 3 계에 속하며 10 .2와 10.3 에서 논할 것이다. 언어적 표현에 의 해 주관적 사고 과정은 객관적 상황을 이룬다(제 3 계 )(Eccles 1981b). 언어에 있어 말로 표현함에는 세 가지 단계를 고려할 수 있다. 첫째, 담화의 모든 여러 부분에서 어휘의 충분한 이해와 정확성의 필요이고 둘째, 문법에 따르는 단어의 정확한 어순 과정으로 구문론법의 요건이 댜 언어의 경험적 구사에 의하여 잘 짜여진 어구를 판단하는 것이 기 준이 된다 셋째, 문장들은 표현하고자 하는 의미를 판단할 수 있어야 하는데 이는 어의학적 요건이다 . 구문론적으로 만족스럽게 구성된 문 장들일지라도 의미론적으로는 무의미할 수도 있다. 여기 촘스키 (Chomsky 1962:15) 의 글에서 예를 들어보겠다 무색의 녹색 생각은 맹렬한 기세로 잠자다. (Colorless gre en ide as sleep furiou sly) 사람의 언어에 있어 이들 기준은 언어의 3 또는 4 수준인 사람의 언 어로 인식될 수 있는 것을 훈련된 유인원이 얼마나 따라 할 수 있는가 를 판정하는 결정적안 기준이 될 수 있다. 또 언어 장애, 대뇌 손상에 의한 실어증을 이해하는 기초가 되며 그리고 대뇌의 여러 언어 기능 위치를 알아보는 기초가 된다 . 43 인간 언어의 학습 렌네버그 (Lenneber g 1969) 는 어린 아이의 발육 과정에서 운동 기능 과 언어능은 동시적으로 발달한다고 제안하였다. 표 4 .1에 0.5 세에서 3 세까지 그리고 4 . 5 세까지의 발달 이정표를 보여주고 있다. 그러나 이
표 4.1 어린 아이의 운동 및 언어 발달 관계 나이(세) 운동 지표 언어 지표 0.5 손으로 짚고 앉는다 . 꾸르 록 거리는 소리는 화음의 도입 한쪽으로 도달하기 으로 재잘거림으로 변한다 1 선다 . 한 손으로 잡고 걷는다 음절 반복 . 일 부 단어 를 이해하는 표시 . 특정 대상이나 사람을 가리키 는 소리 를 규 칙적 으로 낸다. 최 초 의 단어이다 1.5 잡기와 놓기가 완전하게 발달 어 구 를 구성 하지 못하나 3 - 50 단어 한다. 추진성 발걸음 . 계단을 를 구사함. 담화에 유사한 음성 과 뒷걸음으로 기어내려움 음조 연습. 이해력의 좋 은 진전 2 뛰기(넘어지며) . 한쪽 다리로 50 단어 이상. 보통 두 단어 어구. 언 만 계단오르기 어 의사소통에 홍 미 를 가짐. 더 이 상 재잘거리지 않음 2.5 두 다리로 뛰기. 1 초 동안 한 매일 새 단어. 세 단어 이상의 언어 다리로 서기 . 여섯 개 입방체 능력. 자신이 들 은 것 을 거의 모두 로 탑 쌓기 이해하는 것 처럼 보임. 아직 문법적 으로 완전치 못함 3 발끝으로 2.7m 걷기. 양발로 약 1000 단어로 회화. 이해력 80% . 계단오르기. 0.9m 뛰어오르기 문법 구사력 성인에 접근 . 구문상의 오류 다양. 계통적 , 예견적 4.5 줄넘기. 한발로 됩뛰기 . 언어는 잘 성립됨. 문법적 오류는 직선으로 걷기 일상 회화에서 벗어난 경우에 한함 등식은 일반적인 사항일 뿐 언제나 타당한 것은 아니다. 왜냐하면 극 심한 운동능력 장애를 가진 어린이의 예를 들어 소아마비의 후유증을 겪는 아이들일지라도 언어 발달은 정상적일 수 있기 때문이다. 또 침 팬지 새끼는 사람보다 운동 능력이 훨씬 빨리 발달하지만 인간 사회에 들어온다 하더라도 언어 구사 능력의 발달은 신호에 지나지 않는 초보 적인 언어의 범위를 넘지 못한다. 특별하게 훈련된 유인원의 언어 발
달에 관하여는 4 .4에서 논한다 . 우리가 잘 알다시피 생후 1 개월째라 하더라도 유아는 끊임없이 발 성 연습을 하여 가장 복잡한 협동 운동인 언어를 배우기 시작한다. 발 성 학습은 듣는 것으로 유도되며 처음에는 들은 것을 흉내내는 것으로 시작한다. 일년쯤 되면 간단한 단어, 죽 엄마 아빠 등의 형태를 이룬 다. 주목할 것은 말이란 말한 것을 듣는 것에 의한 되먹임에 의한다는 것이다 죽 귀머거리는 벙어리가 된다. 언어 발달에 있어 인식은 표현 울 앞지른다 . 어린아이는 진정으로 말에 굶주려 있다 . 그래서 사물의 명칭을 물어보고 혼자 있을 때도 이를 끊임없이 연습한다 (Pa p ousek 동 1985). 이는 자신이 정한 규칙에 따라 잘못을 일으킬 수도 있다. 예를 들면 사물의 복수형을 불규칙하게 사용하는 것이다. 언어란 단순한 모 방으로만 이룰 수는 없다 . 어린아이는 자신이 들은 것들의 관계와 규 칙성을 정리하여 이를 언어적 표현을 구사하기 위한 문장 구성 원칙에 적용한다 . 힐Hi ll 은 어린아이의 언어 발달에 관하여 할리데이(Halli da y 1975) 의 말을 인용하였다. 초기의 기능적 발달은 거의 전적으로 실용주의적인 것이다. 죽 어린아 이가 사용하는 초기 언어는 자신의 주위에서 요구하는 것을 인지하게 하 고 상호작용이 일어나도록 하는 동 주의를 끌기 위한 것이다 . 어린아이들 이 사물들을 상호작용의 초점으로서 사용하는 이들 초기 기능들은 보다 성숙된 학습적 기능으로 점차 발전해 나간다 (1980 : 348). 어린아이는 인지라는 입장에서 세상을 배우기 위하여 언어를 사용 한다 그러나 물론 이들 두 가지 기능, 죽 실용적 및 학습적 기능은 어 린아이들이 말을 구사하는 매 순간에 피할 수 없이 섞이게 된다. 이와 관련하여 테라스 Terrace 와 베버 Bever 는 다음과 같이 말하였다.
어린아이가 자신을 주목하게 되는 능력, 이것의 욕구와 주위 환경으로 부터의 사회적 압력은 문장 구성능을 별로 요구하지 않는다. 그러나 언어 의 이 기본적 기능은 지대한 효과를 갖는다. 감정을 표현하고 사회적으로 바람직하거나 바람직하지 않은 행동을 식별하는 언어의 신비는 보다 세 련된 언어 구사를 발달시키는 동기를 부여하고 언어 구사는 문장 구성을 요구하게 되는 것이다. 인간 언어의 필요조건은 아마도 자기 자신을 표현 하고자 하는 능력이라고 할 수 있을 것이다(1 980:180). 어린아이의 첫해 동안의 놀라운 언어 발달은 자신을 인식하고 자신 을 표현하기 위한 노력 속에서 어린아이가 자신의 자아의식을 발달시 키는 과정이라 할 수 있다. 어린아이의 지능 발달과 언어 발달은 서로 긍정적으로 상호작용한다. 말하는 것에 별로 접촉한 일이 없다 하더라 도 말할 수 있게 되는 것은 우리의 생물학적 유산의 하나이다. 이러한 잠재 능력과 예민성은 유전적 기초로 보장되는 것이다. 그러나 렌네버 그 Lenneber g는 언어유전자는 없다고 한 도브잔스키 CDobzhansk y 1967) 의 의견에 동의한다. 그러나 유전자에 의해 언어에 관계되는 특정 부 위가 대뇌피질에 마련되고 문장 구사에 관계되는 부속 구조가 대뇌피 질에 마련되는 것이다. 이 구조의 특성에 관하여는 뒤에 기술한다. 44 유인원의 언어 훈련 데카르트 Desc art es 는 사람과 동물의 질적 차이는 언어로 나타낼 수 있다고 하였다. 동물은 사람의 자아의식에 비견할 만한 것이 결여된 자동인형과 같다. 그들은 제한된 발성 신호로 의사를 소통한다. 그러 나 사람의 지능 과정 또는 사고의 성격이나 흐름을 반영하는 언어는 아니 다 (Choms ky 1968). 사람은 이 성 에 의 해 지 배 되 며 동물은 본능에
지배된다. 데카르트에 따르면 인간의 언어는 인간의 영적 활동이다. 유인원이 그림 4 .1에 있는 두 가지 하급언어를 대단히 잘 구사하는 것 은 틀림이 없다 . 죽 행동의 표현과 소속 사회의 구성원과 관계되는 신 호체계이다 (Goodal 1971, 1986). 그러나 그림 4.1 의 3, 4 수준인 고급언 어가 유인원에서 완전하게 결여되어 있으므로 데카르트가 주장했듯이 질적 차이가 있는 것이다. 인간을 영장류로서 그리고 유인원의 근친으로서 계통발생적 체계에 확립시킨 것은 다윈주의적 혁명이다 . 단지 약간의 정량적 차이만을 가 질 뿐, 여러 각도에서 연속성이 있음을 보이는 일은 대단히 홍미 있는 연구 대상이 된다. 이 같은 순조로운 이행 과정을 설명하는 데 한 가지 장애는 인간 언어의 유일성이다. 사람의 언어와 유인원의 언어 사이에 는 분명한 정성적 차이가 있었다 . 따라서 유인원의 언어 능력을 밝히고 언어상의 차이가 단지 정량적 차이일 뿐인가를 밝히는 연구계획이 수 립되었다. 그래서 계통발생상의 간격을 제거하고 그리핀 (G riffin 1976) 이 말한 지능적 경험의 진화적 연속성을 밝히고자 하였다. 초기의 연구는 푸르네스 Furness, 헤이스 Ha y es 씨 가족, 켈로그 Kello gg에 의한 일련의 대담한 시도로서 유인원을 인간 가족과 함께 기르면서 말을 가르쳤다. 수년이 지나서도 아빠, 엄마, 컵, 서 등 네 단 어밖에 익히지 못하였다. 이 같은 실패는 몇 가지 소리의 발음에 국한 하는 발성 기 의 해부학적 결함 때문이 라고 생각하였다(Li eberman 1975). 그러나 사람은 발성기에 현저한 병변으로 예를 들면 후두나 혀가 완전 히 없어져 심한 기능 장해가 있더라도 말을 할 수 있다 (L im ber 1980). 또한 발성기에 여러 실험적 장해를 일으키더라도 대단히 잘 적응할 수 있댜 그래서 림버 (L im ber 1980) 는 다음과 같이 결론지었다. 이들 모든 요인은, 사람 성대의 형태는 인간 언어의 기본이라는 주장을 신중하게 생각하게 하는 것이다 . 사람의 정상적인 성대 자체는 사람의 언
어 능력에 꼭 필요한 것도 아니며 충분한 것도 아니다 ....... 최근의 주요 생물학적 연구 결과는 인간의 언어에 대한 우리의 이해를 별로 진전시키 지 못한 듯하고 단지 데카르트에서처럼 유물론을 미결인 채 남겨 놓은 대신에 하나의 선험적 가정으로 유물론을 받아돌이는 데 기여했을 뿐이 다(1 980:211,214). 유인원의 발성 언어 훈련에 실패하자 여러 다른 훈련 방법을 시도 하게 되었다. 이 같은 시도는 사람과 동물 사이의 장벽을 돌파할 수 있을 것으로 생각되어 대단한 관심을 불러일으켰다. 호미니드 진화에 서 의사소통이라는 관점에서 정성적인 변화는 없었다고 생각한다. 그 러나 단지 점진적 정량적 발달이 있었다고 생각한다. 동시에 유인원의 자연상태의 습성에 관한 계통적 연구가 있었다. 예를 들면 제 인 구달J ane Goodall 은 곰베 Gombe 유역 보호구의 침 팬지 를 연구하 였다 (1971,1986). 세 베옥 Sebeok 과 우미 케르 세 베옥 (Umi ker-Sebeok 1980) 은 유인원 언어에 관한 훌륭한 저술을 출판하였는데 여러 연구자들에 의한 연구에 언어학 전문가들에 의한 결정적 평가가 견들여 있다. 더 욱이 연구자들 자신이 서로를 비평하고 있다 . 이 비평은 대중성으로 골머리를 앓고 있는 이 분야에서 대단히 요구되는 카타르시스라는 점 이 대단히 인상적이다. 유인원의 언어 능력을 대단히 광범하고 훌륭하게 나타내려는 시도 는 미국신호언어 (Ame ri can Sig n Lang u ag e ASL) 를 이용하여 가드너 Gardners 가 1966 년 이래로 수행한 것이다. 침팬지의 자연적 몸짓 의사 소통 방법과 관계되는 손짓 신호체계를 이용하는 이점을 찾았다. 그래 서 유인원은 그들의 부적절한 발성에 의한 장애를 극복하고 언어롤 습 득하는 능력을 나타낼 기회를 가졌다. 젊은 암컷 침팬지 와슈 Washoe 는 수년 동안에 걸친 그들 연구의 대상이었다. 와슈가 깨어 있는 동안 에는 항상 사람과 같이 있었고 ASL 로 사람과 와슈 사이에 의사소통이
이루어졌댜 와슈는 130 개 신호의 어휘를 습득했고 이를 조합하여 네 가지 말을 엮을 수 있었다. 거의 모든 신호로 된 의사 표시는 음식을 요구하거나 사회적 주의에 관한 것이었고 따라서 실용성을 지향하는 도구적 의사 소통이었다. 한편 어린아이의 언어는 말에 대한 탐구와 학습으로 간주되는 것으로 이 학습적 기능은 4.3 을 참조하기 바란다. 가드너 (1980) 는 침팬지와 어린아이의 어휘 학습 속도를 정량적으로 비교하였다 . 처음 1 년반 동안에는 침팬지가 앞선다. 그러나 2 년째는 어 린아이의 어휘가 커진다. 그러나 어휘의 수만이 언어능력의 평가 기준 이 될 수는 없다. 말을 사용하는 방법으로 평가되어야 한다. 유인원이 의지를 전달할 수 있다는 것에는 의심할 바가 없다. 죽 그들이 나타내 는 ASL 에는 어의학적 능력이 있다. 그러나 그들이 손짓으로 엮어내는 신호가 문장구성 법칙에 따를 것이라고 생각되지는 않는다 . 예를 들면 < 너 > < 나 > < 간질이다 > 등의 신호를 여러 순서로 조합하여 <너는 나를 간질인다 > 라고 나타낼 수도 있다. 그러나 어린아이는 3 세만 되어 도 요구, 명령, 물음, 타협을 정확하게 구별하는 문장 구성법을 안다. 이 가드너의 선구자적 연구에 대하여는 여러 비판이 있어 왔다. 예 를 들면 우미케르 세베옥과 세베옥 (1980) 은 비록 가드너가 엄격한 기 준을 적용하였다고는 하나 유인원의 신호가 과대평가될 위험이 있다 고 지적하였다. 더욱 신랄한 비판은 유인원이 나타낸 신호는 훈련자의 신호룰 무의식적으로 따랐을 것이라는 것이다. 죽 곡예효과 clever Hans e ff ec t일 것 이 다. 가드너 (1 980) 는 이 비 판에 답하기 위 하여 이 중맹 검법에 의한 실험을 시도하였다. 그러나 이것조차도 세베옥 (1980) 과 룸 바우 (Rumbau gh 1980) 가 비판하였다 . 와슈 같은 유능한 신호자는 원시적인 행위로 순박한 침팬지에게 신 호언어를 가르칠 수 있으리라고 낙관적으로 생각하였다. 그러나 파우 츠 Fou t s 와 리그비Rig by (1980) 는 종안에서의 신호에 의한 단지 최소한 의 의사소통을 보고하였고 와슈가 그 자손에게 ASL 을 가르치리라는
희망은 실망적인 것이었다. 와슈 자신은 실제로 사람의 딸처럼 자랐다 . 그런데 지금은 죽었지만 그 새끼에게 모성으로서의 관심을 별로 갖지 않았다(Umi ker-Sebeok 과 Sebeok 1980:50 ). 이 같은 종족 내의 교육 실패로 인하여 ASL 로 훈련된 유인원은 실 제로 의사 소통 수단으로서의 가치를 인정하였는지가 의문시된다. 그렇다면 ASL 을 이용한 이 연구로 무엇을 알게 되었는가? 첫째 유 인원의 행동과 사물에 대한 신호의 학습능력이고 둘째 유인원은 음식 을 요구하거나 즐거운 경험 그리고 기분을 표현하기 위해 신호를 도구 적 방법으로 사용할 수 있었다는 것이다. 따라서 ASL 에 의한 의사소 통은 언어의 두 가지 하급수준에 속하는 것이다(그림 4.1 ). 이를 서술적 방법으로 사용했다는 아무런 분명한 증거가 없다. 죽 < 개가 고양이를 물었다>라거나 이룰 변형시켜 < 고양이가 개에게 물렀다 > 라고 하는 간단한 것조차도 안된다. 더욱이 힐(Hi ll1980) 은 침팬지는 간단한 함축 적 표현법도 나타내지 못하나 세 살짜리 어린아이는 이를 이해한다고 지적하였다. 즉 <사라는 초콜릿을 가져온 교관을 좋아한다>라는 말은 <사라는 교관을 좋아한다>라는 말과 <교관은 초콜릿을 가져왔다>는 말에서 파생된 것임을 이해한다. 이는 간단한 상관성의 예이다. 유인원이 언어와 관계가 있는 상징적 의사소통법을 배우는 능력을 보다 분명하고 조직적으로 알아보기 위해서 프레막 (Premack 1976) 은 단어롤 상징하는 플라스틱 조각을 이용하는 대단히 기발한 방법을 개 발하였다(그림 7.1 ). 이 폴라스틱 조각의 모양과 색깔은 특별한 단어와 물건을 의미하게 된다. 침팬지와 훈련자 사이에 있는 언어판에 플라스 틱 조각을 자석으로 부착시켜 상징적 의사소통으로 이용한다. 이 상징 물은 나타내려는 물건과는 닮지 않은 것이다. 예를 들면 삼각형은 사
과를 나타낸다 훈련 방법은 성공을 보상하는 사역적 조건훈련법이다. 젊은 암컷 침팬지 사라 Sarah 는 사물, 색깔, 행위의 단어와 전치사
득하는 놀라운 능력을 보였다고 할 수 있다. 이 소통 방법은 유인원이 음식을 요구하거나 시회적 접촉 등 실용적인 목적으로 사용하였다 . 그 러나 세살배기 어린아아도 효과적으로 구사하듯이 주위 환경에 대하여 배우는 학습적 목적으로는 사용하지 못하였다. 분명히 유인원은 제 2 수 준 언어인 신호로서 상징언어를 택하였고 이를 제 3 수준 언어인 서술적 언어로서 취급했다고 생각하기는 힘들며 제 4 수준 언어는 분명히 아니 었댜 유인원을 대단히 힘들여 훈련시켰더라도 사람의 특징적 언어를 구사하지는 못하였다. 유인원은 언어를 특히 ASL 에서 어의적으로 사 용하였으나 언어 표현에서 구문적 증거를 찾기는 힘들다 (Lenneber g 1980, Brown 1980, Bronowsk y와 Bellug i 1970, 1980, McNeill 1980, Lim b er 1980, Hill 1980, Chomsky 1980). 촘스키는 다음과 같이 결론지 었댜 사람의 지능이 아주 낮은 수준일 때, 그리고 육체적 또는 사회적 장애 에도 불구하고 발달하는 사람의 언어는 그것의 극히 초보적인 성질조차 도 사람 이외의 다른 종에게는 능력 밖의 것이라는 별로 의외가 아닌 전 통적인 가정이 최근의 연구들에 의하여 상당히 광범위하게 재확안되었 댜 이 점은 최근 몇년 동안 에릭 렌네버그, 존 림버 둥 여러 사람이 강조 했다. 이 언어 능력의 차이는 질적 차이인데 양적인 다소의 문제가 아니 라 지적 능력 구성의 전혀 다른 종류의 문제이다 (1980: 439). 유인원의 언어 훈련 계획을 제 3 자의 입장에서 보더라도 유인원이 사람의 수준으로 의사·소통이 가능하게 되리라는 초기의 큰 기대는 실 망적이었다고 생각된다 . 유인원이 의사소통법을 원했다고 보는 것은 홍미에 지나지 않는다. 사람의 생각에 필적하는 아무 것도 갖지 않았 댜 특히 홍미로운 연구는 테라스와 베버 (Terrace 와 Bever 1980) 가 시도 한 것으로 침팬지에게 자아의 개념을 가르치는 것이었다. 이미 갤럽 Gallu p은 1977 년에 침팬지가 거울에서 자신을 인식하는 것을 보았다.
갤럽과 베버는 다음과 같이 말하였다. 우리는 자아의 개념에 의하여 침팬지가 주위 환경의 다른 개체와 관계 되는 자신의 느낌과 의도 등을 개념화할 수 있게 되는 것은 침팬지가 사 람 언어의 문장 구성능적 특징을 알고자 하는 동기부여의 결정적 단계일 것임을 강조하려고 한다. 우리가 아는 한, 사라나 라나는 자아의 언어적 개념을 개발하지 못하였다는 것이다 . 와슈의 경우도 기껏해야 불명확한 증거 가 있을 뿐이 다 (1980: 188). 호미노이드 진화에 있어 침팬지가 우리와 가장 가까운 근친이라는 것에는 별 이론이 없다. 고로 침팬지에게 그림 4 . 1 의 제 1, 2 수준 이상 의 언어를 개발하려던 시도를 장황하게 다룬 것은 타당한 것이었다. 단지 실용적 목적에 사용되는 그림 4 .1의 하급수준에서 세 살짜리 어 린아이도 자신이 속한 세상을 이해하려고 하는 욕구에 의해 부단히 사 용하는 서술, 논의, 그리고 질문 등의 능력을 가진 사람의 고급언어로 진화한 호미니드의 언어 능력에 있어 굉장한 진화적 성취가 있었음을 강조하고자 한 것이다. 이에 반해 유인원은 질문을 하지 않는다. 따라 서 뇌의 언어중추의 특징은 무엇이며 그리고 언제 그것들이 발생하였 는가라는 의문이 제기된다. 4.5 언어 중추와 관계되는 대뇌피질의 해부 지난 세기 이래로 사람의 뇌에는 언어와 관계되는 커다란 두 개의 영역이 좌측 대뇌피질에 있음이 알려져 왔다. 이것이 파괴되면 실어중 을 초래 한다 (Pe nfi eld 와 Robert s 1959, Geschwsin d 1'57 2 , Darna si o 와 Geschw ind 1984). 베 르니 케 We rni cke 의 후방 언 어 중추는 특히 언 어 의
관념화와 관계가 있다. 이 실어증은 문어체 및 구어체 언어 이해의 부 전이 특징이다. 환자는 정상적인 속도와 운율 그리고 정상적인 구문으 로 말한다 하더라도 그 내용은 허황된 것으로 어의가 결여되어 있다. 브로카 Broca 의 전방 언어중추라고 부르는 셋째 전두희의 후방부의 손 상은 브로카의 운동성 실어증을 초래한다. 이 환자는 유창하게 말하지 못하고 간단한 소리를 내는 데도 대단히 힘이 든다. 그러나 말을 이해 할 수는 있다. 브로카 영역은 언어근을 조절하는 대뇌피질 영역 바로 앞에 있다 운동성 실어증은 발성 근육의 마비에 의한 것은 아니지만 그것의 사용 장해에 의한다. 우측 대뇌반구의 병변은 어느 부위에 있 건 심한 언어장애는 초래하지 않는다. 약 5% 의 사람들은 왼손잡이로 이 경우는 반대가 되어 베르니케 브로카 영역이 우측에 있다. 유아나 소아의 뇌 손상의 연구로 초기 발달시에는 대뇌의 양 반구 가 모두 언어에 참여함이 알려졌다(B asser 1962). 정상적으로는 차츰 좌측 대뇌반구가 표현과 해석 등 언어 구사의 우위를 차지하게 된다. 천부적 신경성 우월성 때문이라고 생각한다. 한편 우측 대뇌반구는 언 어 생성이란 면에서는 퇴행하나 언어 이해 능력은 약간 유지하게 된 다. 이 같은 언어 능력의 이행 과정은 대개 4-5 세에 완성된다(Kim ura 1967). 완전하지는 않으나 상당히 좋은 상관관계가 손잡이 상태와 언어 능력의 편재성 사이에 존재한다. 거의 모든 오른손잡이는 좌뇌 언어자 이고 왼손잡이는 좌뇌 또는 우뇌 언어자이다 (Pen fi eld 와 Jas pe r 1954). 우리는 언어영역의 신경 구조의 특징에 관하여 별로 아는 바가 없 다. 다만 임상적 경험으로 그것의 가소성이 알려졌다. 예를 들면 유아 나 소아에서 베르니케 영역이 손상되면 불완전할 때도 있으나 다른 부 위가 언어 기능을 대신한다(Mi lner 1974). 렌네버그 (Lenneber g 1967) 는 언어 영역을 사춘기가 되기까지 사용하지 않으면 학습 능력을 상실하 게 될 것이라고 예상하였다. 13 세가 넘을 때까지 말없이 지냈던 제니 Ge ni e 라고 하는 비극적 예에 따르면 그녀는 말 배우기가 대단히 힘들
었고 그녀는 오른손잡이이므로 좌측 대뇌반구가 언어 영역이어야 함 에도 불구하고 우측 뇌로 언어를 익혔다 (C urti ss 1977). 펜필드와 로버츠 (Pen fi eld 와 Robert s 1959) 는 수술시 때로는 간질환자 의 치료시 노출된 사람의 뇌룰 이용할 수 있어서 언어 영역에 관하여 대단히 만족스럽게 연구할 수 있었다. 자발적이건 반복적이건 다른 부 위는 건드리지 않고 손상 연구로 밝혀진 언어 영역 위에 약한 반복 자 극을 가하여 언어의 장해 또는 정지를 일으킬 수 있음을 발견하였다. 여러 실험 절차로 실어증 손상에 근거한 잘 합치되는 영역 구획이 이 루어졌댜 그림 4.3 은 자극 방법으로 얻어진 언어 영역을 나타낸 것이 댜 우측 대뇌반구의 자극으로는 언어가 변경되지 않았다. 언어중추가 우측 대뇌반구에 한정되었던 약 5% 만이 예의였다. 양측 모두 언어근 의 운동중추를 직접 자극하면 운동성 언어 반응(소리)을 일으켰다(그림 3.5 , 3.6, 9.1 ). 그러나 이는 쉽게 구별할 수 있었다. 그 같은 영 역은 그림 4.3 에는 표시하지 않았다. 이와 유사한 언어 영역 구획은 대규모의 수 술적 절제법으로도 만들어졌다. 실어증 구획법으로 만들어진 언어영역 지도에서 특히 다른 점은 상위 언어중추를 구별할 수 있었던 것이다 (그림 4.3). 이 부위는 예상했던 대로 언어운동의 개시에 특별한 역할을 한댜 이는 부속운동영역 (SMA) 에 속하기 때문이다(그림 8.3 , 8.4 ). 오제 만과 크로이 츠펠트 (O j emann 과 Creutz feld t 1987) 는 언어 영 역 에 관한 임상적 실어증에 의한 또는 생리학적 연구에 의한 광범한 연구 결과를 검토하였다. 대뇌피질의 약한 반복 자극으로 여러 언어기능, 죽 이름 대기, 단문 읽기, 문구의 단기간 기억, 음성 구별 둥을 구분하는 연구도 있었다. 이들 기능은 언어영역에 조각보 모양으로 분포되어 있 고 어떤 기능은 다른 기능과 일부 중첩되어 있다. 표면기록 또는 이식 전극법에 의한 전기생리학적 기록으로 자국법에 의해 추정된 구획에서 전압을 기록할 수 있었다. 장차 이식 전극으로 단일 신경세포의 활동을 세포의적으로 기록하여 더욱 구체화될 것이다. 이 방면의 연구는 이
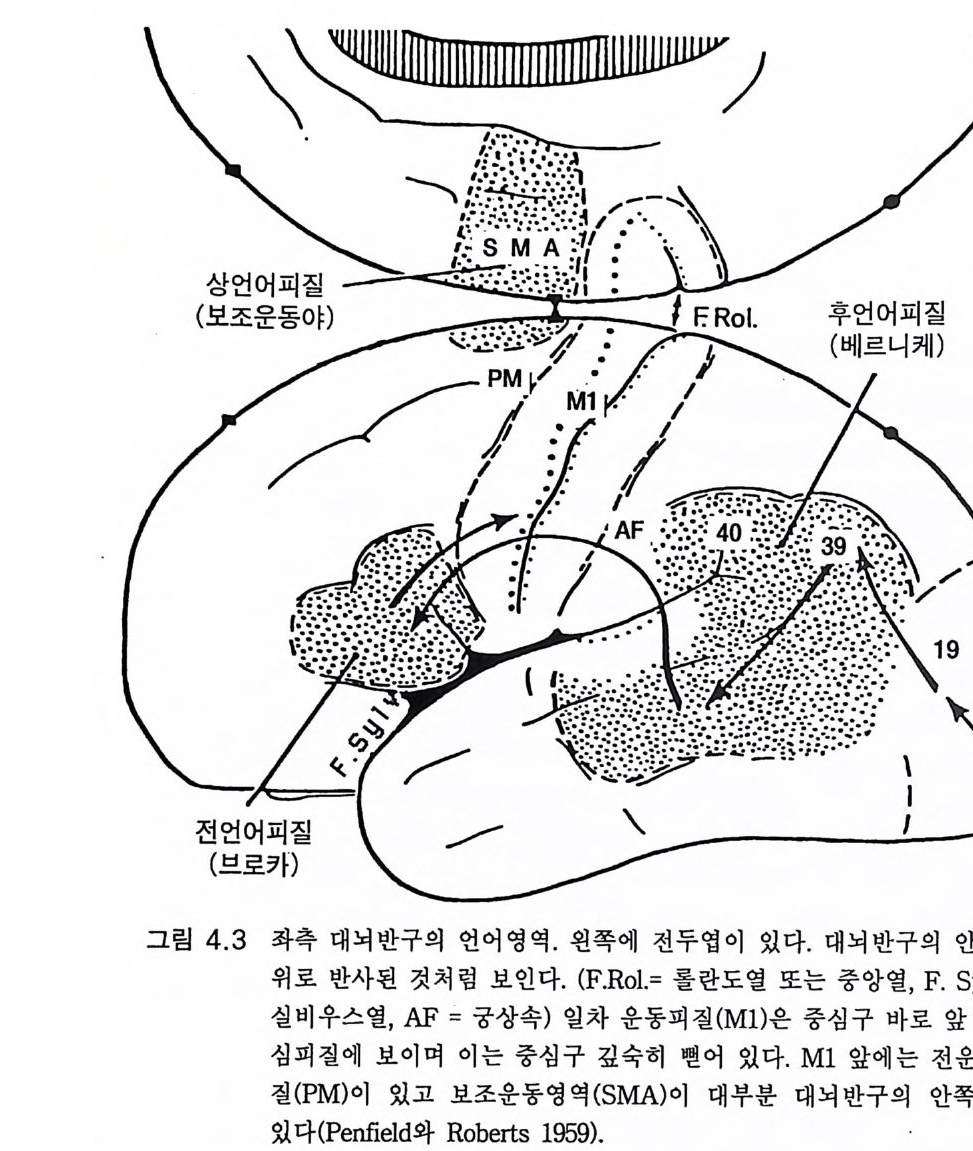 二二죠;.:〈二~ -u베\ 르니\케) 17
二二죠;.:〈二~ -u베\ 르니\케) 17
제 시작에 불과하나 신경세포는 언어의 형태에 따라 방전 형태가 변한 다는 것이 알려졌다. 대뇌에서의 언어기전에 관한 우리의 지식은 아직 유치한 수준이다.
그림 4.3 의 화살표는 소리내어 읽을 때의 신경회로라고 생각되는 것이 다. 시각영역 (17,18,19) 으로부터 신경회로는 영역 39( 각회)로 간다. 영역 39 의 손상은 실독증을 일으킨다. 다음 단계로 베르니케 영역에 의한 의미론적 해석이 있고 궁상속을 거쳐 브로카의 영역으로 가서 언어 생 성을 위한 운동중추의 활성화를 일으키게 하는 복잡한 수속이 이루어 진댜 이는 신경기능의 무한한 복잡성을 단순화한 것으로 우리의 이해를 초월하는 것이다. 그러나 브로드만 Brodmann 에게 있어 언어영역의 구 조는 분명하였다. 그는 1909 년 방대한 조직학적 연구로 사람의 대뇌피 질을 개별적으로 식별되는 영역으로 구획하였다(그림 4.4 ). 펜필드와 로버츠(1 959) 의 규정에 따르면 베르니케 영역은 영역 39,40, 영역 21, 22 의 후방부, 영역 37 의 일부를 포함하는 넓은 영역이다. 한편 브로카 의 영역은 영역 44,45 를 포함하는 것이다. 유인원에서 이에 합당하는 구획은 브로드만의 제자 마우스 (Mauss 1911) 가 만들었다. 그림 4.5 는 오랑우탄 (Pon g o) 의 것이다. 그림 4 .4에 있 는 대부분의 특수한 언어영역이 보이지 않는다. 베르니케 영역은 영역 21,22 가 대부분을 차지하고 영역 40 은 실비우스 구열에서 비쭉 나와 있는 정도다. 그러나 브로카 영역 44,45 는 찾아볼 수 없다. 불행히도 침팬지의 대뇌피질 구획은 이에 합당하는 것이 없다(그림 3.1). 그러나 베 일리 B ail e y, 폰 보닌 von Bon in, 매컬로흐 McCulloch 는 해부학적 증거 들을 수집하였다. 영역 39 는 그들이 말한 영역 PG 로 대신할 수 있었으 나 브로드만 형의 구획이 만들어져야 할 것이다. 만일 침팬지의 뇌가 그림 4.5 와 비슷하다면 사람의 언어수준 3, 4 단계를 학습하지 못하는 것이 쉽게 설명된다(그림 4.1 ). 즉 사람의 언어 수준에 이르는 특수한 대뇌피질 영역이 없거나 충분히 발달되어 있지 못한 것이다. 그림 3 .4에 보이는 바와 같이 대뇌피질 언어영역은 특이적인 성질을 가지나 불행히도 우리는 아직도 자세한 해부학적 구조를 이해하지 못
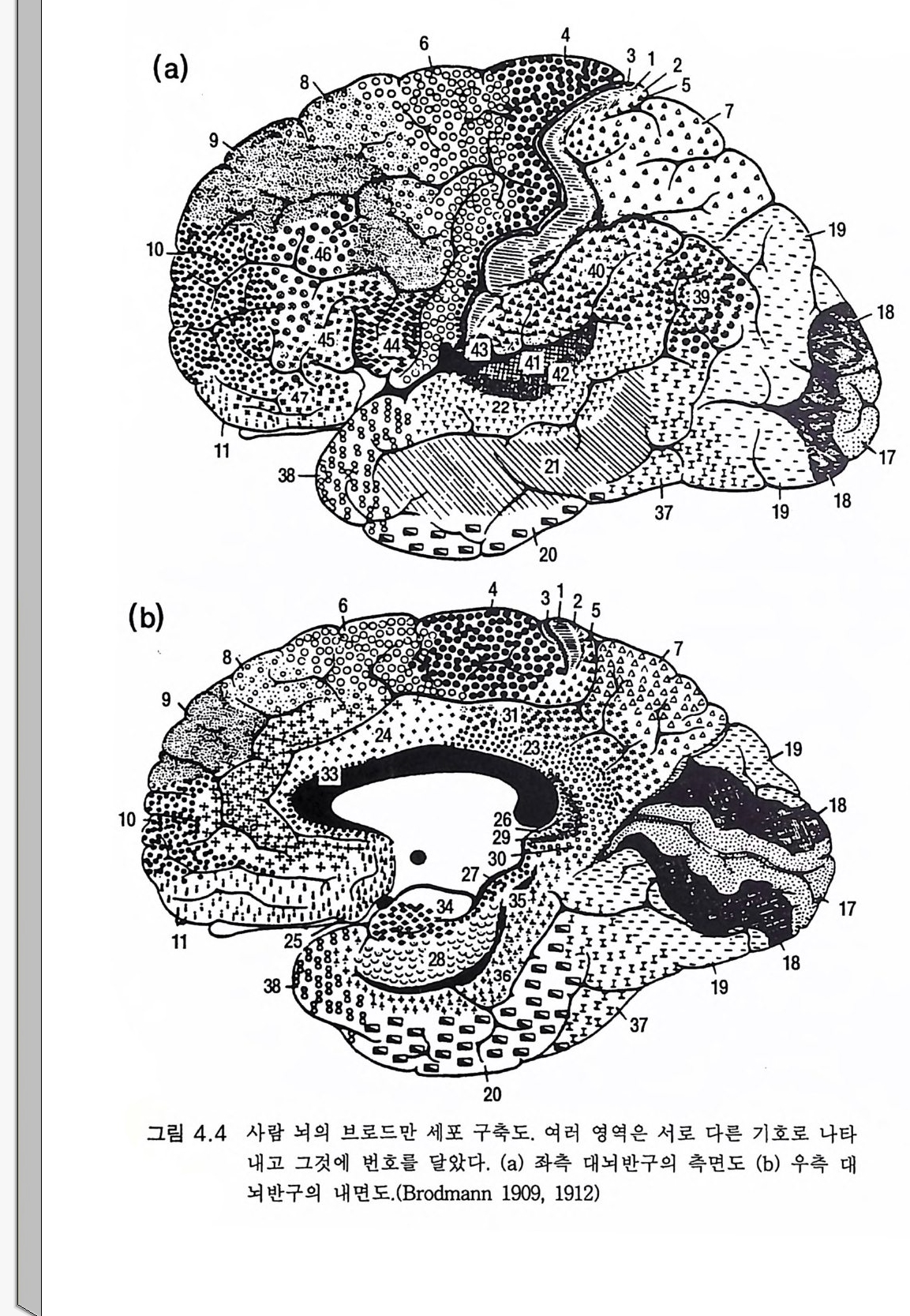 (o
(o
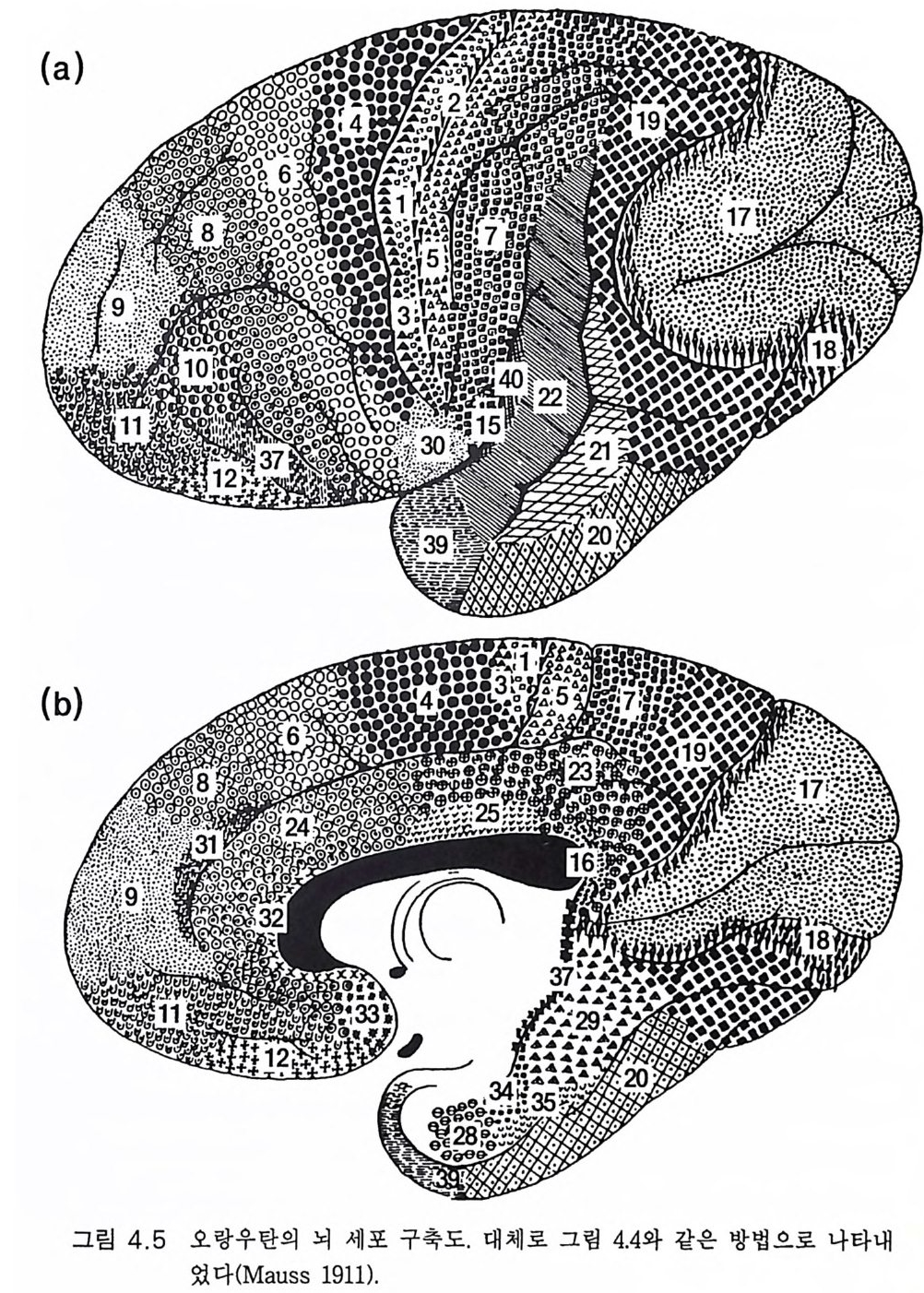 (a)
(a)
하고 있댜 상기한 바와 같이 생후 처음 몇년 동안은 기능적 언어영역 이 우측 대뇌반구에도 있다 . 좌측 대뇌반구의 언어영역을 파괴하면 우 측 대뇌반구의 언어기능이 발달한다. 그러나 어린아이 시기에 이 같은 대치 능력이 소실된다 . 우측 대뇌반구의 잠재적 언어영역의 가소성이 퇴화된다고 할 수 있다. 진화 과정에서 대뇌의 영역은 두 가지 이상의 감각 종류의 입력이 수렴될 때 특이적 기능을 습득하게 되는 것이라고 게슈윈드 (Geschw i nd 1965) 는 제안하였다 . 예를 들면 촉각이나 시각의 감각정보가 예컨대 영 역 39 같은 동일한 신경으로 수렴되면 이들 신경은 느낀 것과 본 것을 인식하는 신호를 발하여 이를 객관화하고 명명하게 된다. 이 같은 감 각 수렴 영 역은 영장류의 상측두구 (STS) 에 있다(J ones 와 Powell 1970). 그리고 이 수렴영역은 진화 과정에서 명명하고 말할 수 있게 발달하였 다고 게슈윈드는 주장하였다. 이와 관련된 토이버 Teuber 의 평을 보면 언어……존재하지 않는 대상을 사람의 마음속에 나타낼 수 있는 도구 를 제공한다. ……언어는 감각의 압제로부터 우리 를 대단히 자유롭게 한 다. ……서로 다른 감각 양식으로부터의 정보 를 통합하는, 따라서 상호감 각적이고 초감각적인 개념들을 생성하게 하는 데 이 과정이 어떻게 이루 어지는가는 수수께끼로 남아 있다 (1967:209). 최 근까지 의 중거 로는 침 팬지 (Da venp ort 1976) 는 보았던 사물을 어 두 운 곳에서 만져보고 식별하거나 그 반대로 사물을 식별하는 교차감각 양식에 의한 인식이 가능하나 원숭이 (E ttring er 와 Blakemore 1969) 는 하 지 못하는 것으로 알려졌다. 그러나 원숭이도 유인원과 마찬가지로 교 차감각 양식으로 인식한다는 것이 밝혀졌다 (Cowa y와 Weis k rantz 1975, J arv i s 와 Ett linge r 1977). 물론 사람은 월등히 잘 하고 말을 하기 전의 유아도 할 수 있다. STS 의 수렴영역은 진화적으로 발달하여 베르니케
의 영역 39, 40 이 되었을 가능성은 충분히 있다. 그러나 불행히도 그림 4.5 에서 영역 39 및 40 은 실비우스 구열에 있는 작은 영역이고 STS 에 있는 것이 아니다. 중요한 것은 사람의 언어영역은 출생 전에 이미 형성된다는 점이고 쉽게 언어를 배울 수 있도록 개체발생학적으로 발달한다는 것이다. 이 는 유전자적으로 결정되는 과정이고 놀랍게도 이 언어영역은 어떤 인 간 언어든 완벽하게 습득할 수 있다 . 어린아이들은 서로 다른 종족간 에도 동등한 언어 습득 능력이 있다는 것은 잘 알려진 사실이다 . 촘스 키 (Chomsk y 1967, 1968, 1978) 는 이 같은 사실을 이용하여 세계 공통 문 법의 일반 원리를 구체화하였다. 나는 문법의 기본적 구조가 뇌의 언 어영역의 미세구조와 상동적 관계에 있다고 생각한다 . 이 같은 관점에 서 어린아이는 언어의 저변 구조의 지식을 가지고 태어난다고 할 수 있다 왜냐하면 이 문법 구조가 출생 전부터 유전자에 의해 구축되는 대뇌피질의 언어영역의 미세구조 속에 새겨져 있기 때문이다 (Wada 등 1975). 4.6 청 각 회 로(Imig와 Morel 1983) 고등 영장류에서 호모 사피엔스 사피엔스까지 와우관 수용체에서 대뇌피질에 이르는 청각 계통에는 별 진화적 변화가 없었다고 알려졌 다(B rodal 1981 ). 와우각에는 고도로 특이적인 변환기전이 있다. 교묘하게 설계된 공 명기전이 있고 복잡한 양상의 음파의 빈도를 분석하며 이룰 뇌로 전달 하는 전기적 신호로 변환시킨다. 몇 단계의 접합부 연계를 거쳐 상측 두회(헤실 회)에 있는 일차 청각영역에 부호화된 정보가 전달된다(그림 4.6). 우측 와우각에서는 주로 좌측 일차 청각영역으로 가고 좌측 와우
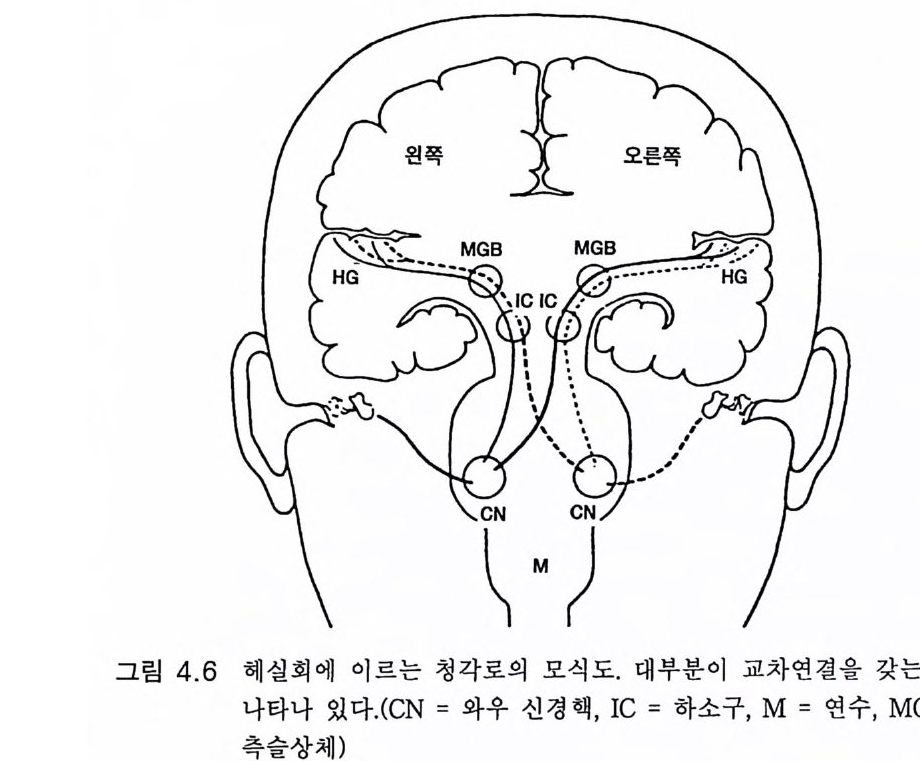 넒-S솔 ,”’ ‘ I
넒-S솔 ,”’ ‘ I
각은 그 반대다. 직선적 체성감각 분포를 하고 있어 가장 높은 음성 주파수는 헤실 회의 가장 내측에 위치하고 가장 낮은 주파수 제일 바 깥쪽에 있다 (Merze ni ch 와 Brug ge 1CJ l3 ). 앞서 언급했듯이 약 95% 의 경우는 좌측 대뇌반구가 언어를 담당한 다. 그리고 이와 더불어 그림 4.7 의 측두평면 (PT), 죽 이차 청각영역은 보통 좌측이 훨씬 크다 (Gesch win d 와 Levi tsk y 1968, Damas i o 와 Gesch- wi nd 1984). 헤실 회와 측두평면은 대충 그림 4 .4의 브로드만 영역 41 및 42 에 해당한다 (Brod a! 1981). 사람 뇌의 이들 영역은 마취된 사람에 서도 소리자극에 대하여 짧은 지연 전압을 보이며 반응한다 (Celes i a 1CJ l6 ).
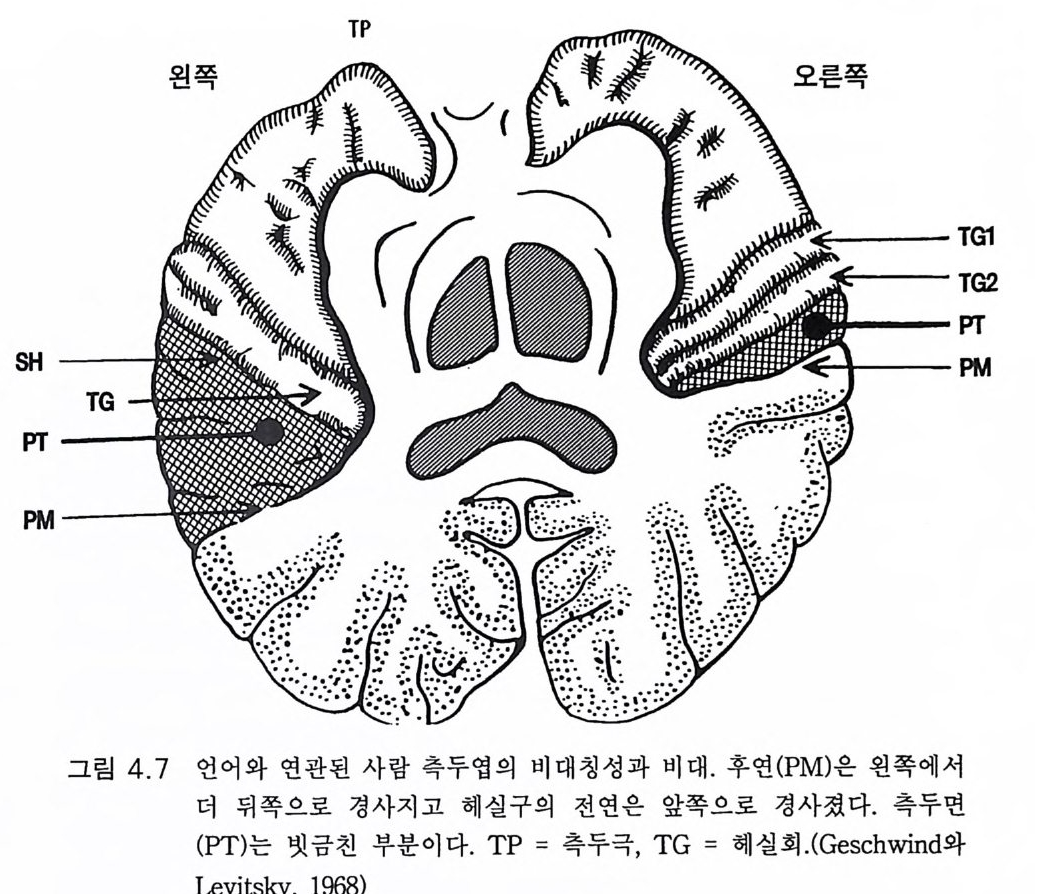 TP
TP
원숭이에서 단계적 퇴행에 의한 해부학적 연구에 의하면(J ones 와 Powell 1970) 영 역 41 과 42 에 서 영 역 22( 상측두희 )와 전두엽 의 여 러 영 역 8a, 9 및 10( 그림 4 .4)으로 가는 회로가 알려졌다. 이에 부합하여 의 식이 있는 사람에서 소리자극은 전두엽과 두정엽의 광범한 부위를 홍 분시 킨다 (Celes i a 둥 1968). 이와 유사한 광범한 청각신호의 분포는 사람의 언어 행위중의 대뇌 피 질 부위 별 대사율 (rCMR) 과 혈류 (rCBF) 의 연구로도 알려 졌다(Ing var 1983). 고전적 브로카-베르니케 영역은 전두전부피질(영역 6,8,9) 을 포
함하는 넓은 부위로 확대되었다. 이는 셀레시아 (Celes i a 1968) 등의 발 견과 합치한다 두정피질은 생각했던 것보다 더 많은 활동이 있었다. 우측 대뇌반구가 상당한 활동을 한다는 사실은 예측하지 못했던 것이 나 뇌량을 가로지르는 활동으로 설명되었다. 그림 4 . 3 에 보여주는 언어 영역은 기본적 언어행위에 필요한 것이며 청각기능에 대한 대뇌 중추 영역이다 . 이들 영역으로부터의 회로는 개념적 사고 등 고급 언어기능 에 필요한 것으로 제 9 장에서 논한다. 4.7 언어 발달과 관계되는 뇌의 진화 대뇌피질의 일차 및 이차 청각영역에 이르는 청각회로(그림 4.6) 의 관점에서는 원숭이에서 유인원을 거쳐 사람에 이르는 진화 과정에서 아무런 변화도 보고된 일이 없다. 그러나 이밖에서는 굉장한 변화가 있었다. 브로카의 전방 언어영역에 해당하는 영역은 유인원에서 알려지지 않았다(그림 4.5). 더욱 두드러진 것은 큰 하두정영역, 각회 그리고 연 상회로서 대개의 경우 오랑우탄(그림 4.5) 이나 기본의 뇌에서는 겨우 식별할 수 있을 정도이고 침팬지의 뇌에서는 존재가 의심스러울 정도 로 작은 영역이다 . . 대단히 홍미로운 것은 영역 39 및 40 은 사람 대뇌피질의 융기 부위 의 모든 영역 중 가장 늦게 유수화된 것임을 플레흐지히 (Flechs ig 1970) 가 발견한 것이다(그림 4.8, 4.9 ). 수초 발생은 출생 후 수상돌기가 발달할 때까지 지연된다. 그리고 세포의 성숙은 소아기까지는 완성되 지 않는다고 생각한다. 이 같은 소견은 영역 39 및 40 이 대뇌피질의 새 로운 영역으로서 계통발생학적으로 발달한 것임울 나타내는 것이고 (Geschw ind 1965, Tobia s 1983) 결국 그림 4.5 에 이것들이 실제로 존재
하지 않는다는 결론에 이른다. 사람의 뇌와 유인원의 뇌를 비교할 때 영역 39 및 40 이 늦게 발달하여 강한 진화적 < 힘 > 이 시각영역을 뒤로 밀어냈음을 의미한다. 이 계통발생학적 과정의 각 단계는 호미니드 진화에서 찾아볼 수 있다. 각 주요 단계를 나타내는 좋은 내부 충전 자료가 있다 . 토비아스 (19 83, 1987) 는 오스트랄로피테신의 뇌는 현대의 유인원 뇌에 비하여 별로 발달되지 않았음을 발견하였다 . 죽 장차 하전두엽의 브로카 영역 이 될 부위에 작은 충만을 보았댜 그러나 훨씬 큰 호모 하빌리스의 뇌에서는 하전두엽의 충만이 뚜렷할 뿐 아니라 하두정엽도 분명하여 영역 39 및 40 의 계통발생학적 발달이 이미 시작되었음을 나타낸다 (Tobia s 1983, 1987, Holloway 1983). 호모 에 렉투스에서는 이들 장래 전 방 및 후방 언어영역의 발달이 진전되었고 네안데르탈인의 큰 뇌는 호 모 사피엔스 사피엔스의 수준까지 완전하게 발달한 것으로 생각된다. 상두정엽의 동시적 발달이 있었는데 이 영역(브로드만 영역 7) 은 특별 히 언어영역과는 관계되지 않고 촉각-시각과 관계되는 것으로 이는 제 6장 에서 다룰 것이다. 대단히 특이한 진화현상을 살펴보았다. 진화적 변화는 이미 존재했 던 구조가 서로 다르지만 연관된 기능이 발달함에 의하여 일어나는 것 이 정상적이다. 그런데 특이하게도 영역 39 및 40 은 꽃 피듯이 상측두 엽의 상부둑으로 돋아나고 진화 시간적으로 대단히 일찍 나타나 시각 영역을 뒤로 밀어붙여 후두엽의 중간 쪽에 위치하게 하였을 것으로 생 각된 다 (Tob i as 1983, Holloway 1983). 영장류급의 진화적 변화·속도를 알아보기 위하여 뇌의 큰 구성 성 분을 측정하였다 (S t e p han 둥 1987). 대뇌피질의 진화적 발달에는 근사 한 상동성이 있을 것으로 가정한 것이다 (Frahm 둥 1982). 예를 들면 그 림 3 .1에서 사람의 신피질 크기 지수 s i ze in dex 는 199 이고 이는 원인류 의 신피질 크기 지수보다 약 3.2 배 큰 것이다. 피질의 팽창은 표면적이
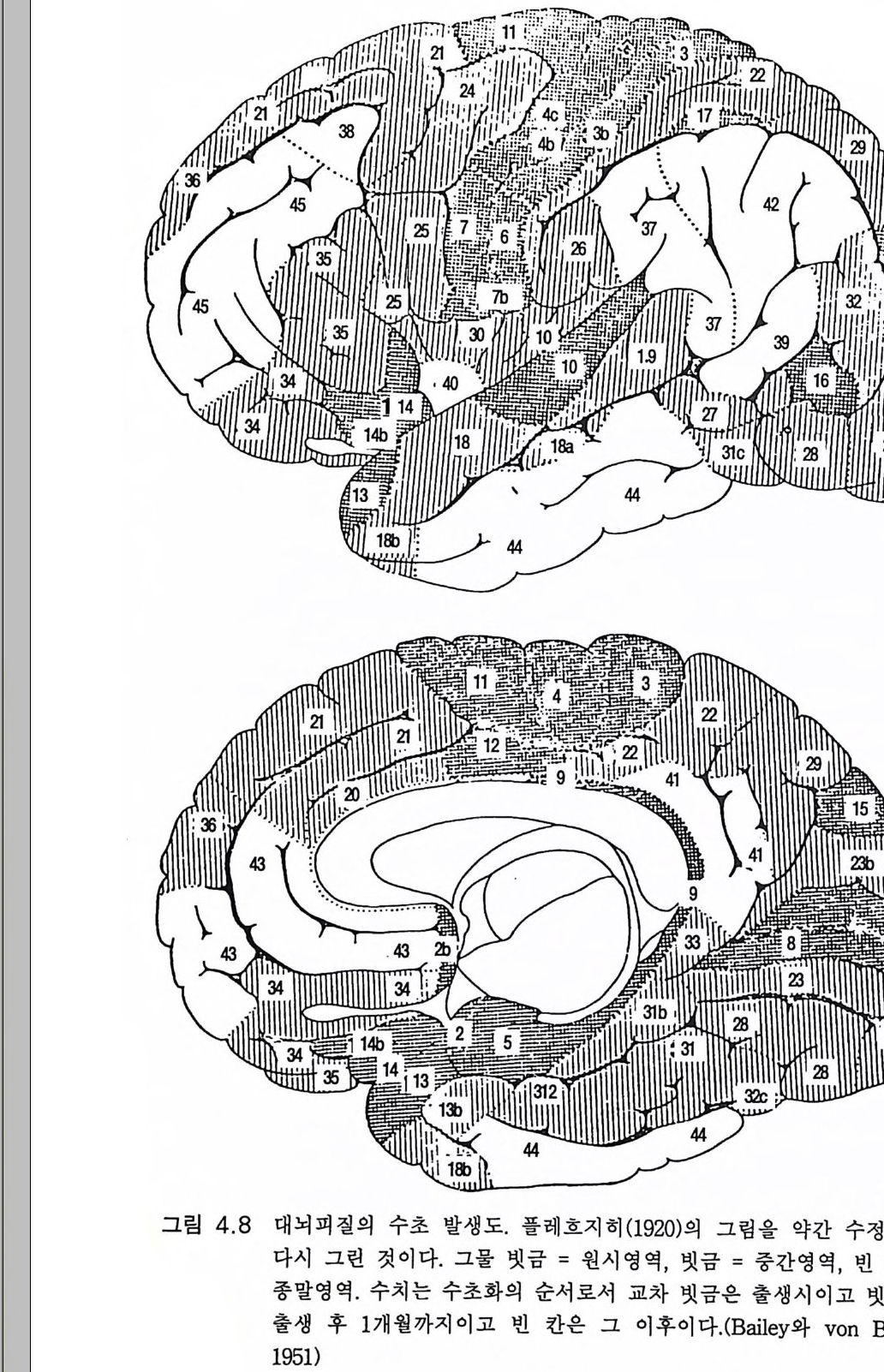 그림 4.8 1출종鬪9말생5?1영 ) 린역후.의 1 것수개치:월는까: 지 수발이;초고생화물 의\빈 : 순러칸서〔은흐로 ;서그\ ?교鬪이차°후) 이빗:다금금그 .은(=B a립출i중l을 e간생 y 영약와시역 간이v, 고o 수n 빈 정빗B 하칸금on여 은 in=
그림 4.8 1출종鬪9말생5?1영 ) 린역후.의 1 것수개치:월는까: 지 수발이;초고생화물 의\빈 : 순러칸서〔은흐로 ;서그\ ?교鬪이차°후) 이빗:다금금그 .은(=B a립출i중l을 e간생 y 영약와시역 간이v, 고o 수n 빈 정빗B 하칸금on여 은 in=
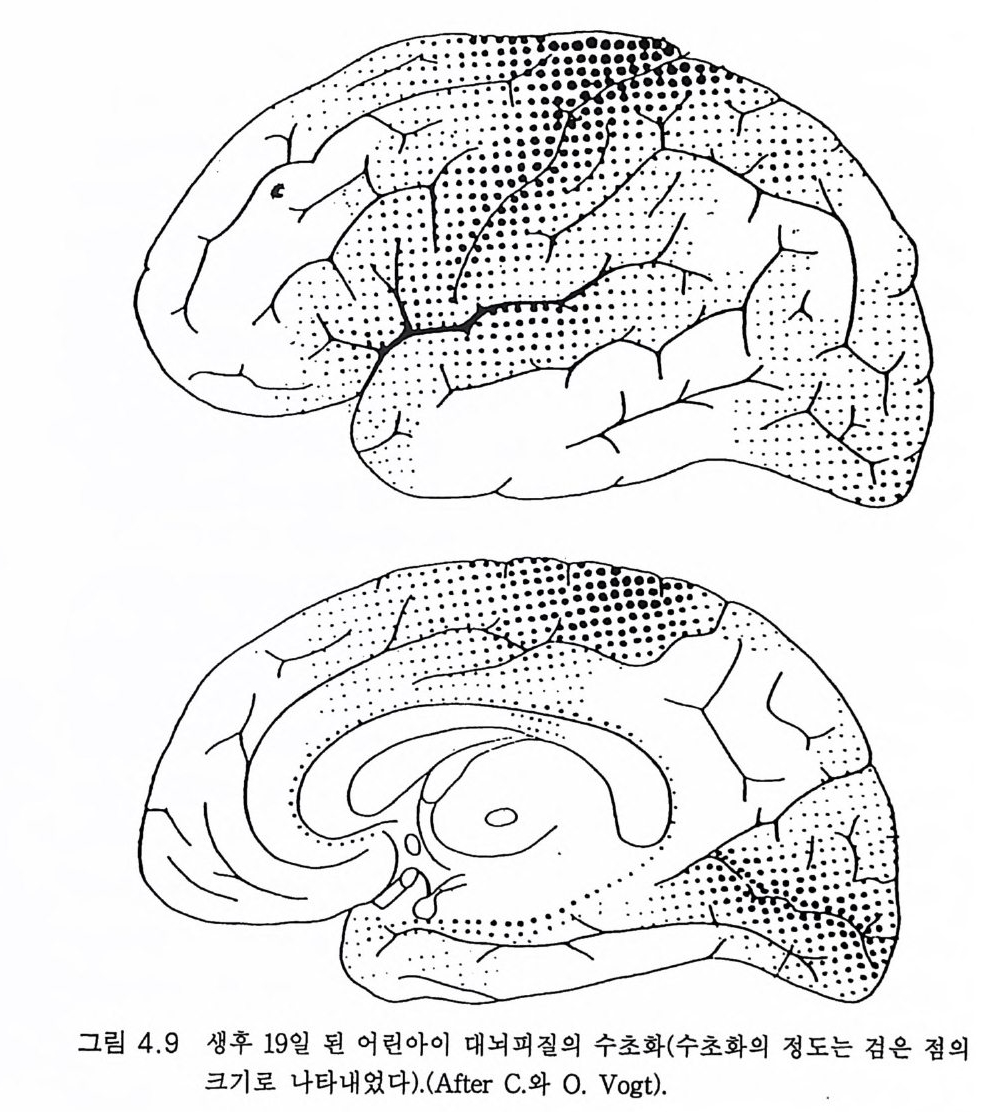 ... : ..
... : ..
커진 것이고 깊이는 별로 커진 것은 아니다 (Frahm 등 1982). 그런데 피 질의 평균 팽창이 3.2 배인 것에 비하면 영역 39 및 40 은 굉장히 팽창한 것이다. 제 3 장에서 또 하나의 특별한 팽창을 보인 부위를 들었는데 운 동피질에 대한 전운동피질의 비는 사람 뇌가 원숭이 뇌보다 수 배 크 댜 죽 브로드만 영역 44 및 45 가 크게 증대하여(그림 4.4 ) 브로카의 전 방 언어영역(그림 4.3) 을 이루었다고 생각되는 것으로 원인류의 뇌에서 는 식별되지 않는다(그림 4.5). 물론 더 철저한 연구가 필요할 것이다. 게슈윈드 (1965) 는 영역 39 및 40 은 사람의 뇌 진화과정에서 유일하 게 새 롭게 나타나는 구조라고 하였다. 또 영 역 39( 각회 )는 시 각과 촉각 정보가 만나는 장소에서 발달하였으므로 언어의 특별한 기능을 가졌 고 이들은 서로 특히 밀접한 관계가 있을 것이라고 생각하였다 . 그래 서 시각과 촉각을 통하여 관찰한 사물을 식별하고 명명하는 데 특히 적합하게 되었을 것이다. 이로써 그림 4 .1에 있는 언어의 제 3 단계를 설 명할 수 있는 것이다. 이것으로 예상할 수 있듯이 영역 39 의 손상은 실 서증을 수반하는 독서불능증 또는 난독증을 초래한다(D amas i o 와 Geschw ind 1984). 대뇌반구의 해부학적 비대칭성에 대하여 많은 사람들이 홍미를 가 졌다(D amas i o 와 Geschw ind 1984, 그리고 제 9 장). 그러나 이를 예룰 들어 호모 하빌리스의 내부 주조물에도 적용하는 데에는 문제가 있다. 실비 우스 구와 후두 극에서 발견되는 비대칭성은 호모 하빌리스에 있어 언 어능력을 나타낼 만큼 뚜렷하지 않다. 하측두엽의 양측성 곡면성 충만 은 훨 씬 뚜 렷 하다 (Tob i as 1987). 소두증 난쟁이의 뇌는 대뇌피질의 영역들에 있어 특이적 성질의 이 상을 나타낸다(H ollowa y 1968, Pass ing ham 과 Ett linge r 1974). 유인원 범 위에 속하는 뇌의 크기에도 불구하고 분명히 사람의 행동을 나타내며 수백 단어 정도의 제한은 있으나 사람의 언어능력을 나타내고 단순한 수준의 사역을 수행할 수 있다. 이들의 뇌는 고릴라의 뇌에 비하여도
작지만 그림 4 .1의 인간 수준의 언어를 나타내도록 특정화된 영역들을 가지고 있음에 틀림없다. 실제로는 체구가 더 작으므로 소두증의 뇌는 고릴라의 뇌에 비하여 대뇌화 지수 ence p ha li za ti on i ndex 가 크다 (3 .1). 4.8 언어 생성의 진화 앞서 유인원은 발성 장치의 구조 때문에 말을 할 수 없다는 리버만 (Lie b erman 1975) 의 주장을 지 적 하였다. 공기 가 입 과 코를 통하여 들어 오고 나가기 때문에 원숭이로부터 유인원을 거쳐 사람에 이르는 동안 후두의 점진적인 하강으로 발성 통로가 열리는 것을 그림 4.10 에 보이 고 있댜 그림 4 .1 0b 에 유인원의 비 인강이 나타나 있고 이 것 이 사람에 서는 더욱 발달했음을 그림 4 .l Oc,d 에 보여주고 있다. 토비아스는 이를 다음과 같이 말하였다. 중요한 점은 유인원이라 하더라도 비인강이 있다는 것이다. 죽 발성의 형태적인 전제 요건은 입을 통한 공기 흐름의 구조상의 여지와 후두의 하강이다 오스트랄로피테쿠스에 있어서 이 같은 형태가 분명치 못하다 는 것은 뜻밖이다. 그러나 호모 하빌리스와 그 이상 발전한 호미니드에서 는 현대인의 조건에 매우 접근해 있었음에 틀림없다 (1983 : 129). 침팬지의 발성 기구가 음소p honeme 를 만들기에 충분할 정도로 발달 한 것이라면 미숙한 수준의 발음도 이루지 못하는 것은 대뇌 수준의 비적합성에 기인한다고 할 수 있는 것이다. 이는 사람의 어린아이와 침팬지 새끼의 극단적 차이로 잘 나타난다. 사람의 소아는 끊임없이 조잘댄다. 그래서 계속해서 차츰 음소의 생성이 개선되어간다는 것을 파푸색 (Pa p ousek 198. 5) 등이 연구했다. 그들은 어린아이가 혼자있을 때
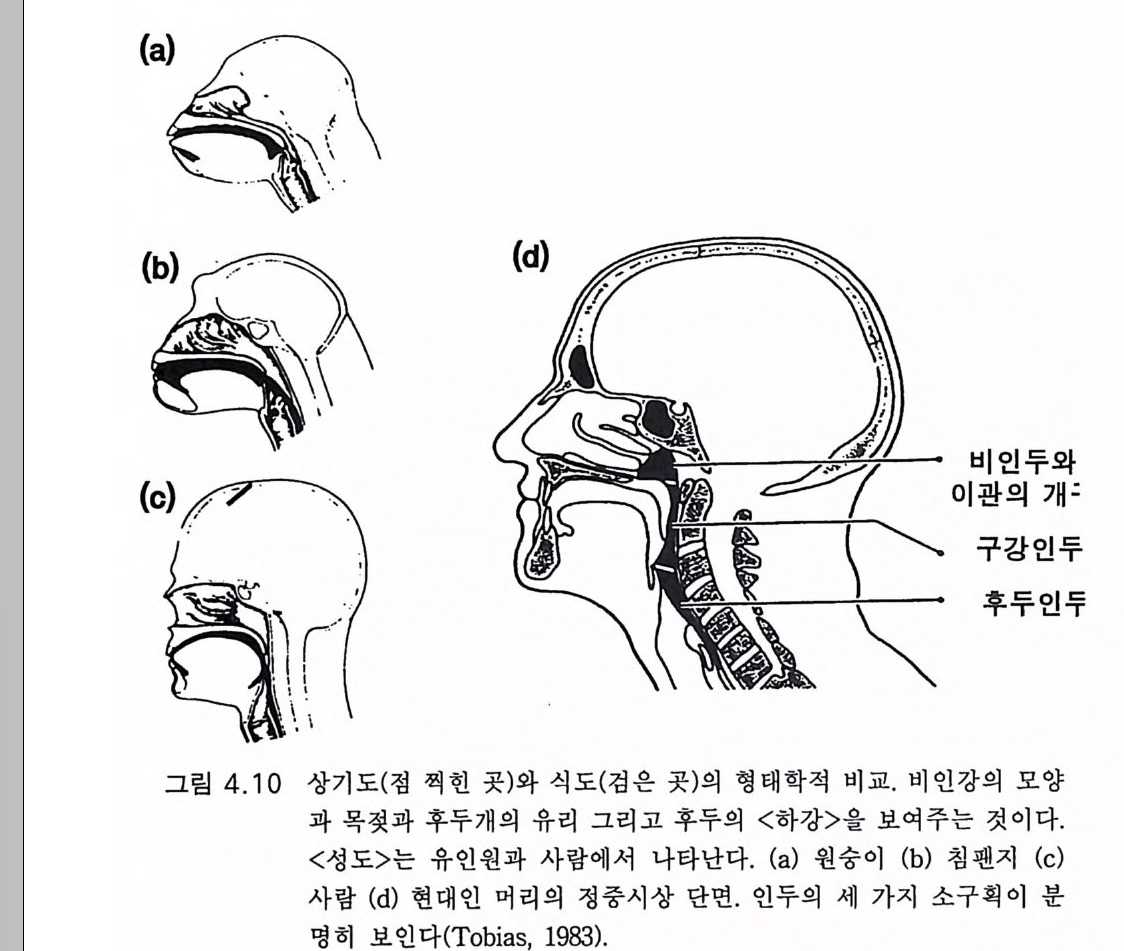 (d}
(d}
의 음성을 조칙적으로 광범하게 녹음하였다. 어린아이들은 자동 학습 과정으로 생각되는 수시간의 연습을 하고 있었다. 그래서 간단한 단어 롤 발음하게 되고 이어서 몇 음절과 어구를 이루게 된다. 이에 반하여 침팬지 새끼는 놀랍도록 조용하다. 그래서 그것들의 대뇌 구조는 실험 적 발음 생성이 시작되지 않는다. 이는 4 .4에서 푸르네스 FUI11ess, 헤이 에스 Hay es 및 켈로그 Kello gg의 실패로 예를 들었다.
렌네버그 (Lenneber g 1969) 는 대단히 홍미로운 관찰을 하였다. 양친 이 모두 선천성 귀머거리인 어린아이의 듣기를 관찰하였다 . 어린아이 는 양친으로부터는 말을 들을 수 없었다. 양친의 발성으로는 그들이 원하는 것을 얻는 데는 소용이 없었다. 그런데 어린아이는 제때에 말 을 시작하고 정상적인 언어발달을 보였다. 이는 집밖에서 맞게 되는 기회가 학습을 유도하기에 충분했다고 생각되는 것이다 . 렌네버그는 이와 관련하여 다음과 같이 결론지었다. 언어 능력은 그 자체의 자연 경과에 따른다. 어 린아이는 환경으로부터 최소한의 자극과 기회만 주어지면 언어 능력을 얻을 수 있다 . 어린아이의 언어 활 동은 환경 조건에 좌우되나 잠재능력은 쉽게 정지되지 않는다 (1969:637). 언어의 진화라는 입장에서 호모 하빌리스의 능력에는 몇 가지 음소 가 있었다고 토비 아스 (1986) 는 말하였다. 이는 오스트랄로피 테신보다 는 크고 호모 에렉투스나 호모 사피엔스보다는 못했다고 결론지었다. 구조적으로나 기능적으로 오늘날의 인간 수준의 진화적 발달에 미 치지는 못하였으나 하빌리스도 언어의 신경적 기초와 말초해부학적 능력을 가졌다고 생각한다 (1986:757) . 4.9 언어와 진화적 생존 오스트랄로피테쿠스 아프리카누스는 200 만년 이상 오랜 동안 존재 했으나(그림 2.5) 그 인구는 크게 중가하지 않았던 것 같다. 이는 화석 이 남아프리카의 다섯 군데 그리고 동아프리카의 열 군데에 국한한다 는 것으로 알 수 있다. 화석은 수백 개체가 발견되었다 (Tob i as 1983).
이 같은 장기간의 오스트랄로피테신의 존재는 10 만 세대 이상에 해당 한댜 만일 화석이 백만 중 하나꼴로 발견되었다 하더라도 (Leake y와 Lewi n 1977) 어느 한 시기에 존재했던 인구수는 수천에 지나지 않는다 . 호미니드 진화계는 대단히 미약하게 유지되었을 것이다 . 어떤 지역에 서는 몇 명으로 압축되었을 것으로 추측된다 . 실제로 오스트랄로피테 신 속의 모든 종은 약 100 만년 전에 사라지게 된다 . 대담한 양각 보행의 경험으로 초기 호미니드는 수상생활의 상대적 인 안전성에 비하여 지상을 근거로 하는 생활의 값을 치루어야 했다 . 이는 특히 어린아이나 유아의 약탈자로부터의 위험을 수반하는 위태 로운 선택이었다. 그러나 생활 터전을 마련하여 음식을 서로 나누고 동굴 또는 나무나 기타 소멸되는 재료로 방어할 수 있는 터전을 마련 하여 야간 생존을 도모할 기회를 갖게 되었다. 여기에 언어라는 소리 에 의한 의사소통의 개발에 직면하게 되었을 것이다. 내부 충진법에 의한 전방 및 후방 언어영역의 존재를 근거로 토비아스 (1983, 1987) 는 호모 하빌리스가 구어체의 언어를 시작한 것으로 간주할 수 있다고 하 였다. 이 같은 획기적인 발달이 있으려면 생존성을 중가시키기 위한 유전적 정보가 뇌에 형성됨으로써 초보적인 소리 신호룰 이용하는 모 종의 전단계가 있었을 것이다 . 이것도 진화의 점진성이라는 범주에 들 어갈 수 있다. 오스트랄로피테신에게는 하전두회, 브로카 덮개 발달의 증거 도 있다 (Tob i as 1983, 1987). 앞 사람의 발자국을 정 확하게 따라 밟 은 훌륭한 보행은(그림 3.11) 오스트랄로피 테신은 유인원보다 우수한 대뇌 활동이 있었다는 증거이고 그림 4 .1의 수준 3 언어의 시작까지도 생각할 수 있다. 이어서 호모 하빌리스의 시초가 되는 큰 유전적 변화 로 싹트기로 가지쳐서 우리가 속한 호모라는 새로운 속의 첫번째룰 이 룬다(그림 2.5). 이때에 뇌의 크기는 40% 중가하였고 더욱 중요한 것은 전 방 및 후방 언 어 영 역 의 존재 중거 이 다 (Tob i as 1983, Holloway 1983). 토비아스는 호모 하빌리스의 효과적인 언어 능력을 확신하였다. 오
스트랄로피테쿠스 아프리카누스는 그 뇌가 작았으므로 위에서 추측한 정도의 초보적인 언어조차도 없었을 것이라고 하였다 . 언어 능력이 추 측될 수 있음에도 불구하고 호모 하빌리스의 인구가 크게 증가하였다 고는 생각되지 않는다. 그리고 타지방으로 이주하였다는 증거도 없다. 수십만 년 후 호모 에렉투스의 진화로 크게 확산된다 . 여기서 뇌(그림 2.5 , 2.6, 2 .1 3) 와 언어영역이 크게 증대하는 큰 유전적 발전으로 새로운 종을 이룬다. 초보적 언어라 하더라도 표현 목적으로 이용될 수 있었을 것이다. 따 라서 그림 4.1 의 수준 3 언어가 될 수 있다. 좋은 식물성 음식 또는 사냥 할 동물의 위치 그리고 확인된 약탈자의 위치와 움직임을 표현할 수 있 었을 것이댜 또 이 수준의 언어는 사회적 단결력을 기르고 집단간의 배 려 그리고 제 2 장과 5 장에서 · 설명한 핵가족 내의 배려에 이용될 수도 있 댜 그래서 한정된 진화적 성취를 확고히 하고 이 성취는 호모 사피엔스 사피엔스에 이르기까지 진화적 발달을 거듭하여 의심할 바 없이 지구상 에서 가장 지배적인 종을 이루게 된다. 문화에 있어 언어의 뛰어난 역할 은 뒤에 사고와 자아의식을 다룰 때에 언급하게 될 것이다(제 8, 9, 10 장).
제 5 장 대뇌 변연계와 감정 및 생식계 전화의 관계 변연계는 신경 중추의 복합체 그리고 중추간의 소통 회로로 이루어 졌댜 대뇌피질의 저변에 위치하고 대뇌를 중심선을 따라 절개하면 내 측에 일부가 보인다(그림 4.4 , 5.1 ). 여기서는 진화적 생존 그리고 결국 에 호미니드의 성취를 이름에 있어 변연계의 중요 역할을 다루게 된 댜 진화를 설명하는 데에 해부학을 상세하게 다룰 필요는 없다. 상세 한 것은 브로달 (Brodal 1981) 이나 아이삭손 (Isaacson 1974) 의 문헌을 참 고하면 될 것이다. 5.1 해부학적 개관 편도체라고 하는 신경핵의 집합체가 작용의 중심이다(그림 5.2a). 이 것은 편도처럼 생겼기 때문에 붙여진 이름이다. 영역 34( 그림 4.4 ) 밑으 로 측두극 내측에 묻혀 있다. 화살표로 보여주듯이 시상하부로 직접 투입하고 분계선조를 통하여 간접적으로도 투입한다. 시상하부는 수직 금으로 표시되어 있다(H). 이곳의 구성핵들은 다루지 않는다. 브로달
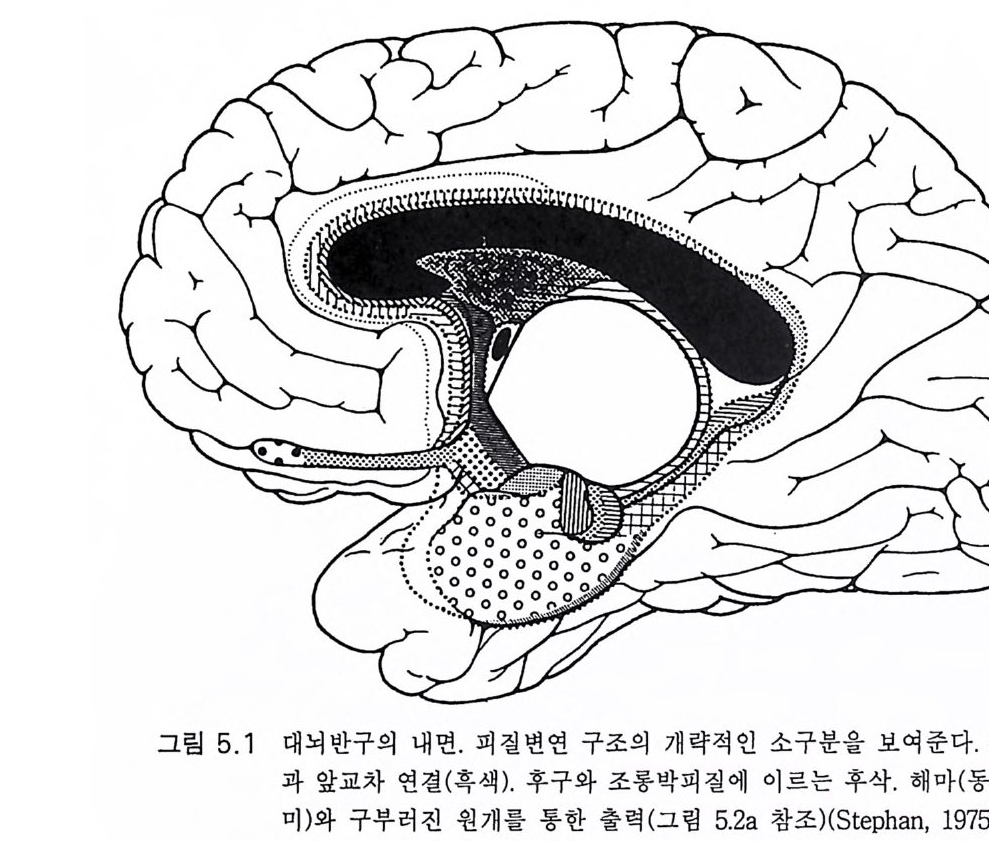 그림 5.1 대뇌반구의 내면 . 피질변연 구조의 개략적인 소구분을 보여준다 . 뇌량
그림 5.1 대뇌반구의 내면 . 피질변연 구조의 개략적인 소구분을 보여준다 . 뇌량
(1981) 은 편도체를 피질내측군과 기저의측군으로 세분했는데 이는 자 극과 제거 효과의 연구에 가치가 있다 . 또 편도체에서 중격핵들로 가는 회로도 보이고 있다. 그림 5.2a 에는 보이지 않으나 후구로부터 직접 또 는 조롱박피질(그림 5.3) 을 거쳐 편도체로 입력되는 회로도 있고 여러 시상핵들로부터 그리고 피질부로부터도 편도체로 입력된다(그림 5.3 ). 그림 5 . 2b 에서는 중격핵들 (S) 이 작용의 중심이다. 특히 중요한 것은 내측전뇌 속으로 이는 피개와 시상하부 간의 쌍방 교통이다 . 수조를 거쳐 고삐핵으로 가서 피개와 각간핵에 이르는 하행회로도 있다. 따라 서 중격핵은 뇌간으로 가는 좋은 교통로이고 전체적으로 신체 반응을 유발하는 참재성 이 있다. 그림 5.2a 및 b 에서 시상하부는 배경으로 나 타냈다. 그러나 이것은 감정이나 성적 반옹을 나타냄에 있어 매우 중
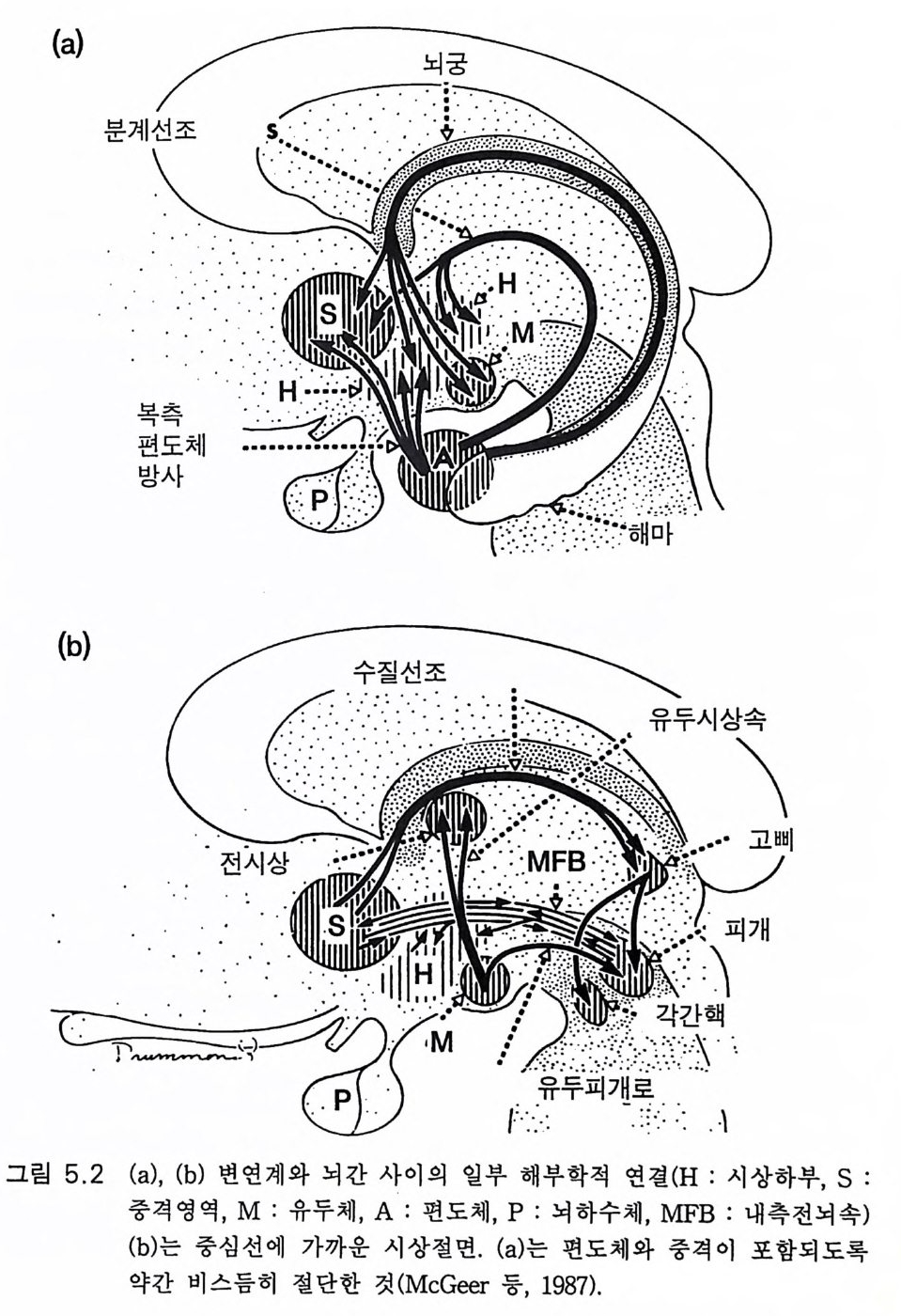 (a)
(a)
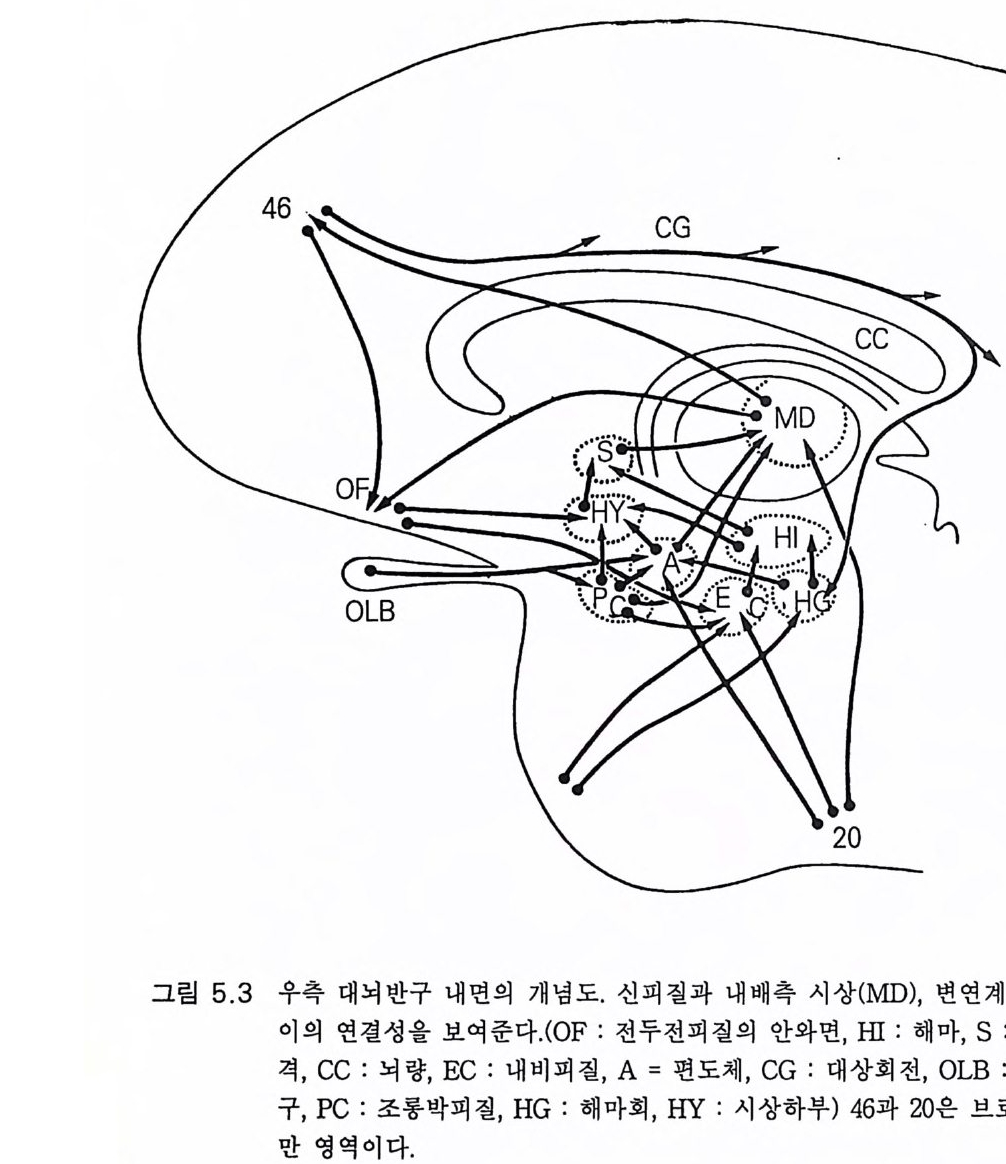 \\° F 二
\\° F 二
요하고 자율 신경계의 중추신경절이라고 하는 것이 옳다. 이는 심혈 관계와 내장을 조절하는 것이다. 시상하부는 또한 뇌하수체 (P) 의 분비 도 조절하여 그 분비물은 신체 전체를 통하여 화학적 전달물질로 작용 하는 것이다. 사람의 중격핵은 퇴화한 것이라고 잘못 생각하여 왔다. 하등 포유 동물에서는 신경 세포가 없는 얇은 막 투명 중격이 되어 측뇌실을 가 르고 있댜 그러나 앤디와 스테판 (And y와 Ste p h an 1968) 은 철저한 비교 연구로 중격핵은 복측으로 이주하여 피막중격 se pt um verum 을 형성하 였음을 증명하였다. 사람의 중격 복합체는 실제로 대단히 잘 발달해 있고 크기 지수가 5 .4 5 이며 이는 유인원의 2.0 9 내지 2.16 에 비하여 배 이상이 된다(표 3.1 ). 크기 지수는 동등한 체중의 기초 식충 동물에서의 크기에 대한 비교치이다. 기타 몇 가지 중요한 회로를 그림 5.2 와 5.3 에 보여주고 있다. 뇌궁 (그림 5 . 2a) 은 백만 개 이상의 신경섬유로 이루어진 큰 신경로이다 . 이 것은 해마에서 시작하여 뇌량 밑을 싸고돌아 중격핵 (S), 시상하부(H), 유두체 (M) 에서 끝나는 주원심성 회로이댜 유두체에서 출발하는 신경 섬유는 전부시상으로 가서 대상회로 힘차게 뻗어 결국 신피질에 이르 게 되는 것이다(그림으로는 보이지 않음). 중격핵과 편도체에서 큰 중배 측 시상핵 (MD) 으로 가서 대부분 전두엽 전방인 신피질에 이르는 회로 가 그림 5.3 에 있다. 그림 5.3 에는 여러 신피질 영역에서 변연계로 가는 몇 가지 회로도 있다. 이들 회로는 신피질과 변연계 간의 상호 정보 교환에 대단히 중요한 의미가 있을 것으로 생각되며 감정의 표현과 발 달에 중요한 것으로 생각된다. 이 같은 간단한 해부학적 개관은 변연계롤 자극하거나 제거하였을 때 나타나는 여러 반응들과 정서와의 관계를 검토함에 필요하다. 여기 서는 비교적 근소한 사람에 대한 실험이 특히 중요하게 다루어질 것 이다.
5.2 변연계와 감정의 표현 (U tt al 1978, 제 5 장 E) 헤스(H ess 1932) 의 선구적 연구로 의식 이 있는 동물의 시상하부와 변연 부위를 자극했을 때의 효과가 처음으로 계통적으로 밝혀졌다. 자 극 전극은 마취 상태에서 미리 수술적으로 심어두었다. 자극 부위를 극소화하기 위하여 세심한 주의를 기울였다 . 이렇게 해서 헤스는 자극 에 의하여 동물의 정서 행동을 유발할 수 있었고 이는 대부분 자율 신 경계를 통한 효과였다. 으르링거림, 위협적 치찰음, 입모 등의 공포나 방어반응을 일으킬 수 있었고 필요 여부에 불구하고 먹기, 마시기, 토 하기, 배변, 배뇨, 성행위 증가 등을 일으킬 수 있었다. 그 이후로 시상 하부나 변연계를 자극하거나 제거했을 때의 효과가 대단히 광범하게 이루어졌다. 때로는 상반된 결과도 보고되었다. 이 부위는 뇌의 대단 히 복잡한 부분으로서 다양한 신경핵들이 근접하게 위치해 있고 이들 은 서로 다르게 때로는 정반대의 작용을 하기 때문이다. 또 상당한 종 간의 차이도 있다. 장기적으로 전극을 심어놓는 기술을 이용하여 올즈와 밀너 (Olds 와 Milne r 1954) 는 정서 연구의 새로운 방법을 시도하였다 . 전극은 중격 핵 또는 내측 전뇌 속을 자극하도록 심어졌고(그림 5.2 b ) 쥐는 자신이 누 름막대를 누름으로써 자극을 받도록 장치하였다. 그래서 한 시간에 7000 번까지도 자기 자극을 일으키는 광적 행동을 유발할 수 있었다 . 행동실험자는 쾌락 중추의 자극에 의한다고 해석되는 행동반응을 경 험할 수도 있었다. 그래서 아이삭손은 행동의 제한적 자극」간옹설 대 신에 <생체의 경험하는 바도 행위 내용을 기술하거나 설명하는 데 중 요 요인으로 간주되어야 한다 . 쾌락과 고통은 행위의 신경기전을 밝히 고자 노력하는 과학자들에게 다시 한번 적절한 용어들로 받아들여졌 다>고 하였다(1CJ7 4:104). 동물 실험의 해석에 관한 이 같은 태도의 변화는 사람을 대상으로
하는 실험의 보고에 의하여 크게 자극받았다 . 이 점에 관하여는 간단 히 기술할 것이다. 쥐가 자기 자극을 얻기 위하여 성행위나 음식을 포 기하고 자극 장치에 들어가기 위하여 발 자극에 의한 동통을 감수한다 는 것은 주목할 일이다. 변연계-시상하부의 다른 부위들은 중격-내측 종축속 부위보다 자극 효과가 강하지 못하였다. 어떤 다른 부위의 자 극으로는 회피반응을 학습하도록 하는 기피반응을 일으킬 수 있었다. 고양이에서는 편도체의 자극은 대부분 기피반응을 일으켰다 . 그러나 자극하면 공격행동을 억제하는 부분도 있었다. 양측 중격핵의 제거는 대체로 쾌락 자극에 반대되는 결과를 보였다. 죽 신경질이나 공격성을 보였다. 양측 편도체를 제거하면 동물은 온순 해지고 성 행위가 증가하며 때로는 이상한 성 행위가 나타난다. 클레 멘테와 체이스 (Clemen t e 와 Chase 1973) 는 동물의 공격성과 변연계-시 상하부의 구조의 관계를 상세하게 연구하였다. 변연계(편도체 및 중격) 늠 시상하부의 조정에 의하여 공격적이거나 공격성이 억제되는 작용 을 하는 것으로 생각된다. 드물게 변연계에 이상이 있는 환자가 자극 또는 제거시의 효과를 연구하는 대상이 되었다 . 사람에서의 실험은 비교적 동물실험의 결과 와 잘 부합되었다. 델가도 Del g ado 는 18 세 때에 뇌 염을 앓았던 편도체 이상이 있는 20 세 여자의 주목할 만한 경우를 보고하였다. 그녀의 주된 사회적 문제는 빈번하고 예측할 수 없는 극도로 난폭한 분노 발작이었다. 환자는 형사상으로 문제가 되는 광기로 입원하였고 신 경과 이상을 조사하기 위하여 해마와 편도체에 전극을 심었다. ……그리 고 해마와 편도체 모두에서 심한 전기적 이상을 보였다. ……환자의 자발 적 분노 발작과 비슷한 난폭한 행동을 우측 편도체의 전파자극으로 일으 킬 수 있었다 . 그녀는 자기의 기타를 내던지고 난폭하게 벽을 들이박고
수분 동안 마루를 뛰어돌고 차츰 조용해져서 원래의 명랑한 상태로 회복 되었댜 이 같은 소견은 국소적 응고법에 의한 단계적 치료를 선정할 때 임상적으로 매우 중요한 것이었다. 이와 비슷한 보고가 더 있다. 마크와 어빈 (Mark 와 Ervin 1970) 은 조 절되지 않는 난폭을 수반하는 측두엽 간질이 있는 두 명의 환자에서 편도체 자극 효과를 관찰하였다. 전극을 내측에 위치하고 자극하면 조 절되지 않는 난폭 행동이 유발되었다. 그러나 전극을 외측에 위치하고 자극하면 상기되고 더워지며 뜨는 듯한 줄거움을 느꼈다. 이 두 환자 는 편도체를 수술적으로 제거하여 치유 효과를 보았다. 히스(H ea th 1954, 1963) 는 다수의 정신분열증 환자를 중격핵에 심어 놓은 전극으로 자극하여 성적 충동이 커지는 느낌을 일으키고 그래서 올즈의 쥐처럼 어떤 환자는 자기 자극을 하게 되는 것을 보고하였다. 델가도 (1969) 는 세 명의 정신운동성 간질환자의 편도체에 전극을 심고 (외측부라고 생각됨) 자극에 의하여 지나치게 친근한 행동을 보이는 즐 거운 기분을 유발할 수 있었다고 보고하였다. 맥레안 (McLean 1966) 은 원숭이의 변연계를 자극하여 성행위가 유발 됨을 발표하였다. 특별히 효과가 있었던 부위는 내측전뇌속 (MFB), 의 측 시상하부(H), 중격 (S) (그림 5.2 ) 그리고 내배측시상 (MD), 대상회 (CG), 전뇌 전부피 질의 안와면 (OF) (그림 5.3) 이 다. 결론적으로 맥레안(1 966, 1970, 1982) 이 주장한 간단한 가설은 지지하 기 힘들고 변연계에는 적어도 두 가지 성분이 있어서 쾌적 효과와 불 쾌 효과에 관계되는 것이다. 중격핵, 내측전뇌속, 그리고 부속 시상하 부(그림 5.2b) 는 쾌적 효과와 관련이 있고 종종 성적 상기를 일으키는 정서와 관계된다. 편도체(내측부)와 일부 이것에 연결된 분계선조(그림 5.2a) 는 불쾌감과 관계된다. 과도한 편도체 활성은 환자에 따라 비참한 결과를 가져올 수도 있어서 범죄 심지어는 살인도 할 수 있는 위험 인
물로 분류될 수 있다. 지금까지 정동에 관계되는 변연계 및 관련 구조에 관하여 검토하였 댜 그러나 정동이라는 것은 의식적 경험임을 상기할 필요가 있다 (Mc Lean 1970). 이 원론적 상호 작용설 du ali s t-i n t erac ti o ni sm 에 따르면(제 8 장) 정동이란 일부 특정한 시공간의 패턴으로 신피질내 기본 단위가 활성화되는 것과 관계되는 것이다. 예를 들면 그림 8.5 와 9.5 에 있는 정 동감은 영적 Inner Sense 인 것으로 되어 있고 적절한 기본 단위(모듈)의 활성화를 통해 감정 표현이 이루어지게 된다. 따라서 변연계와 신피질 간의 연결 회로는 정서와 정동의 의식적 체험에 있어 필수적인 연결선 을 제공한다. 그림 5.3 에 대강의 희로를 나타내었다. 내배측 시상핵은 중심적 구조이다. 이것은 편도체와 중격으로부터 정보를 받고 신피질 에 넓게, 특히 전뇌엽전부의 거의 모든 부위에 발신한다(그림 5 . 3 에서 46, OF). 전방시상(그림 5.2b) 또한 중요한 것으로 대상회(그림 5.3 의 CG) 를 거 쳐 신피 질에 넓 게 발신한다. 5,3 변연계와 시상하부의 약리학 세로토닌, 카테콜아민, 노르아드레날린, 도파민은 변연계와 시상하 부에 관계되는 신경전달물질이다. 따라서 행동에 영향을 미친다. 이들 아민이 행동 및 정서 상태를 매개하는 중요한 역할을 한다는 많은 증 거들이 있다 (Ke ty 1970, 1972). 노르아드레날린과 도파민은 특히 각성, 공격성, 자기 자극 및 느낌과 관계가 있고 세로토닌은 이완, 수면과 관 계가 있다고 한다 (Eccles 1980, 제 6 장). 케티 Ke ty는 다음과 같이 결론지 었다. 한두 가지 생리적 아민의 활성으로 정서 상태를 설명하려고 시도하는
것은 무리한 것이라 생각한다. 이들 아민은 복잡한 신경망의 결정적 접합 부에서 각기 독립적으로 또는 협동적으로 작용하여 정서 상태를 나타내 게 된다고 생각한다. 이 같은 협동작용은 여러 정동의 공통적인 특징과 원초적 성질이지만 이들 각 상태의 특이적인 성질은 아마도 개인의 경험 에 기초한 감지 및 인지 인자를 나타내는 신경 망으로 확장될 때 유도될 것 이 다(1 972:120). 여러 약리학적 물질의 작용이 이들 신경전달물질과 관계될 수 있다. 예를 들면 LSD 의 환각작용은 세로토닌 수용체 부위에서의 작용으로 생각된댜 레세르핀의 진정작용은 항정신병약으로 이용되는데 도파민 을 고갈시키는 작용에 의한 것으로 생각한다. 변연계와 신경전달물질 의 약리 학에 관한 자세 한 것 은 맥 기 어 (McGeer 1987) 등의 책 제 15 장을 참조하기 바란다. 변연계에서는 아민성 전달물질보다도 펩티드 전달물질이 더 중요하 다 (McGeer 동 1987, 제 12 장). 변연계 중에서 펩티드가 가장 많이 포함되 어 있는 부위는 시상하부이다. 편도체에도 많이 포함되어 있는데 여러 펩티드 신경전달물질이 여러 세부에 분포한다(R obe rt s 등 1982). 펩티 드를 함유하는 회로가 편도체에서 분계선조를 거쳐 시상하부로 간다 (그림 5.2a). 중격핵(그림 5.2 ) 그리고 브로카의 대각대핵에도 펩티드 신 경전달물질이 풍부하게 함유되어 있다. 아편류를 주사하면 편도체 및 부속핵 그리고 시상하부에 축적된다. 또 에스트로겐(에스트라디올)은 시상하부와 편도체에 결합한다(그림 5.7). 생리적 아민, 펩티드 둥 신경전달 후보물질들의 변연계의 여러 부위에 있어서의 생리학적 작용에 관하여는 분별적인 연구가 있어야 한다. 변연계핵의 아편류 결합부는 약물 의존성과 관계가 있음을 상기할 필요가 있댜 앞서 말했듯이 중격, 내측 종속, 그리고 외측 편도체를
전기적으로 자극하면 성적 충동을 수반하는 쾌감을 일으킨다. 따라서 심어 놓은 전극을 통한 쥐의 자기자극도 의존성을 일으킨다. 이 같은 자기자극 의존성은 사람에서도 볼 수 있다. 쾌적하고 선정적인 느낌은 정상 생활중의 즐거운 경험의 일부이고 원하는 것이다. 변연계의 오피 움류 수용체는 엔케팔린이 작용하는 부위이고 동통을 자동적으로 해 소하는 신 경 계 이 다(H o kf el t 동 1977, Eccles 1984). 불행히도 바람직한 생리학적 및 정신적 작용에 관계되는 이들 수용 체는 모두 의존성 약물이 결합할 수 있는 부위이다. 이에 따라 유발된 신경반응들은 약물 중독자를 만들 만큼 불가항력적인 쾌락을 경험하 게 한다 진화 연구에서, 현존하는 대부분의 인간들은 쾌감을 맛볼 수 있는 여러 식물들을 발견하였고 그래서 약물 의존성의 위험에 노출되 었음을 볼 수 있다. 반대로 사람 이의의 영장류에서는 약물 의존성을 자연생활에서 찾아볼 수 없다. 변연계 신경핵의 크기 지수로 보아 공 격성 신경핵의 발달보다는 쾌감 신경핵이 점진적으로 더 발달함을 알 수 있는데 이는 다음 절에서 설명한다. 5.4 영장류 진화 과정중의 변연계 구성분의 크기 지수 호모를 포함하여 존재하는 영장류의 변연계 구성분의 크기를 관찰 함으로써 호미니드 진화에 대해 통찰할 수 있다. 그림 3 .1에 스테판과 동료들 (1988) 이 제시한 변연계 구성분의 크기 지수가 있다. 대체로 신 피질의 중가에 따르는 큰 변화는 없다. 그러나 변연계의 일부와 부속 부위들에서는 상당한 변화를 보인다. 예를 들면 사람 해마의 크기 지 수는 4.87 까지도 되는 데 비하여 유인원은 2.99 이고 해마의 가장 발달 한 부위인 CA1 피질(그림 7.4 a )(Ste p h an 1983) 은 더 커서 6.5 이고 이곳이 인지적 기억이 저장되는 중요 구조이다(제 7 장). 해마와의 밀집한 연결
성으로 짐작할 수 있듯이 분열피질(내비 및 바주위피질 그리고 지각)의 크기 지수는 해마와 상관이 깊어 호모에서는 4 .4 3 이고 유인원에서는 2 .3 8 이다. 사람의 중격 크기 지수는 5 .4 5 로 크게 증가하였고 반대로 취구는 위 축되어 호모의 크기 지수는 0.03 인데 유인원은 0.06 이고 원원류는 0.52 이댜 후각피질도 호모는 0.3 이고 유인원은 0.31 이다. 분명히 진화함에 따라 원시 포유류보다 후각은 감퇴하였다. 변연계의 중요 구성분인 편도체는 성격이 크게 다른 두 부위로 나 누어져 있다 (S t e p han 과 Andy 1977). 표 5 .1에 이를 보이고 있다 (S t e p han 둥 1988). 기초 식충동물에서는 두 부위의 크기가 대략 비슷하지만 점 차 내측 중심핵 (CM) 이 차지하는 비율이 줄어들어 24.7% 가 되고 피질 외측기저핵 (CBL) 은 75.3% 를 차지한다. 편도체핵의 의측기저군은 사람 으로 진화함에 따라 점차로 커지고 내측중심군은 작은 채로 남아 있게 된 것이라고 브로달(1 981)( 그림 10.5) 이 말하였다 . 따라서 CM 의 크기 지수는 진화 과정에서 늘 작아서 비인간 원원류는 1. 30 이고 호모는 2.52 인데 CBL 은 상당히 커져서 비인간 원원류는 3.05 이고 호모는 6.0 2 이댜 중격의 진화적 발전으로 호모의 크기 지수가 5 .4 3( 표 5.1)인 것을 편도체의 구성분들의 수치와 관련지어 생각하면 진화는 변연계에 있 어 쾌락과 관계되는 성분(중격 및 CBL) 은 증강되고 공격성에 관계되는 성분 (CM) 은 발달하지 않은 채로 남아 있게 된 것이라고 결론지을 수 있댜 논란의 여지가 많은 편도체 및 중격으로부터의 복잡한 회로 (Brod a! 1981) 를 다루는 것은 이 책이 의도하는 바가 아니다. 몇 가지는 그림 5.2 와 5.3 에 나타나 있는데 이는 굉장히 단순화한 것이다. 대상회 (CG) 를 자극하거나 제거했을 때 위에서 언급한 편도체나 중격에서 있었던 것 같은 정서의 변화를 나타낸다는 것은 홍미있는 일이다. 즉 그림 5.3 에 일부 나타나 있듯이 대상회는 전뇌전부와 안와피질에 대하여 중간
표 5.1 편도체와 중격의 진화 (Tenrec i nae 와의 비교치) 중심내측군 피질 기저 외 측군 중격 전체 백편분도율체 의 크기 지수 전체 백편분도율체 의 크기 지수 크지기수 평균 Tenrecin a e(N=4) 46.8 1.00 53.2 1.00 1.00 평균 식충류 (N=50) 46.8 1.08 53.2 1.13 1.22 평균 원원류 (N = 18) 31. 1 1.16 68.9 2.23 1.91 평균 신구세계 원숭이 (N=23) 26.5 1.30 73.5 3.0 5 2.0 9 기본 23.4 1.20 76.6 3.2 4 2.46 고릴라 26.4 0.8 8 73.6 1.94 2.1 6 침팬지 27.4 1.11 72.6 2.28 l.87 평균 폰지드 (N=3) 25.7 1.06 74.3 2.4 9 2.1 6 호모 (N=l) 24.7 2.5 2 75.3 6.0 2 5.45 N = 종의 숫자 (Ste p h an, 1988) 적 작용을 할 것으로 생각되는 것이다. 5.5 호미니드 진화에 있어 뇌 증대의 결과 굴드 (Gould 1977) 는 뇌의 출생 전 및 출생 후의 성장에 관하여 의문 을 제기하면서 신생아는 특이하게 성인에 비하여 작은 뇌를 가지고 태 어남을 지적하였다. 이는 뇌의 체중에 대한 이중 대수도표로 볼 수 있 다(그림 5.4 ). 그는 <사람의 신생아는 시기적으로 일찍 태어나는 것인
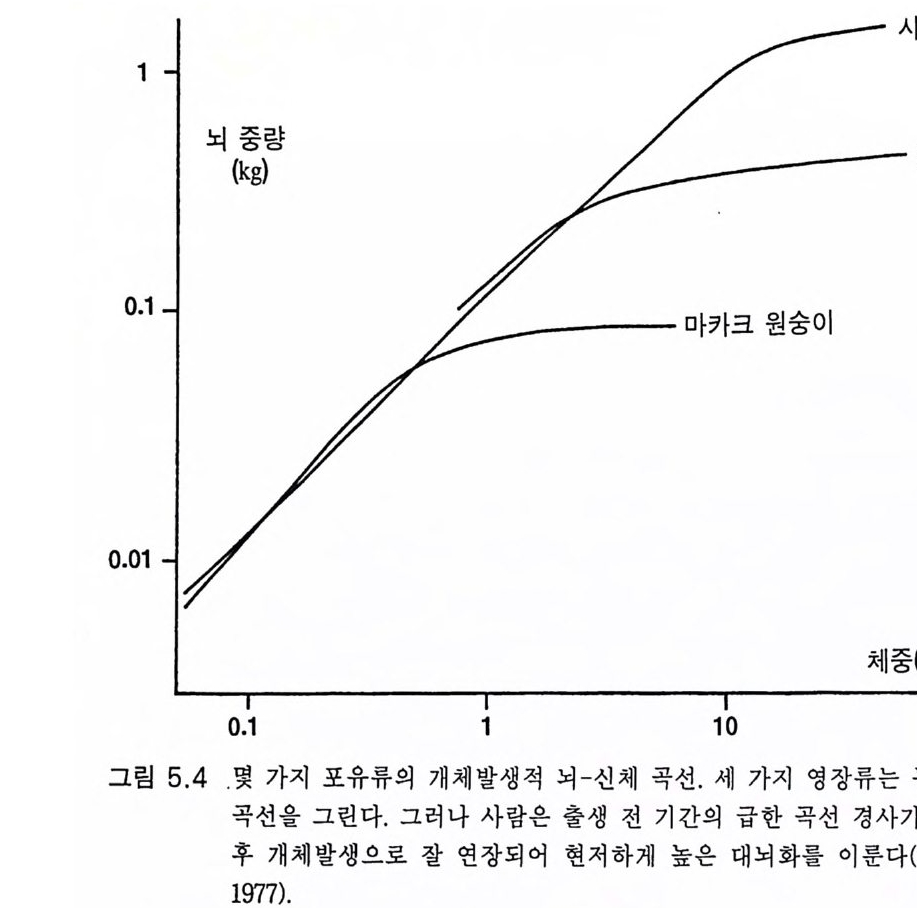 1
1
가?>라는 의문을 제기하였다. 토바아스(1 981b) 는 이같이 출산 시기가 점차 빨라지는 필요성에 대한 산과적 근거를 제시하였다 . 뇌 크기의 진화적 증대를 대비하여 여성의 골반을 통한 출산로의 크기가 태아의 두개골보다 크게 되지 않으려면 조기에 출산해야 한다고 하였다. 그림 5.2 에는 유인원과 호모의 성인 및 출생시의 뇌 크기 평균치를 보여주 고 있다. 현대의 유인원이 호미노이드 조상의 모델이라고 가정하면 놀 랍게도 호미니드 진화에 있어 성인의 뇌는 세 배로 커진 데 비하여 출 생시 뇌는 별로 커지지 않았다 (300-350cc) . 호미니드 진화에서 출생시 뇌의 크기가 성인의 60% 였다면 표 5.2 의 넷째 줄에 보여주듯이 이는 생각하기 힘들 정도로 컸던 것이라 하
표 5.2 유인원 , 호모(실제) 및 호미니드(추측)의 출생시 및 성인의 뇌 크기. 성인 뇌 출생시 성인 크기의 조기에 출산하지 평균 크기 뇌 크기 백분율 않았을 경우 계산된 출생시 (cc) (cc) 뇌 크기 (cc) 유안원 480 300 60 오스트랄로피테신 480 300 60 300 호모 하빌리스 646 300 46 390 호모 에렉투스 890 300 35 530 호모 에렉투스 페키넨시스 1,043 300 29 625 호모 사피엔스 1,3 4 4 350 26 810 (Tobia s , 1981b) 겠댜 그래서 태아의 뇌가 발달하기 이전에 점차로 조기에 출산해야 할 진화적 필요가 있었다. 이 같은 조기 출산의 결과에 관하여는 5.6 에 서 논한다 . 동시에 생물학적 기전에 관하여 검토하게 된다 . 리긴스 (L iggi ns 1967) 둥은 양의 출산시 첫번째 단계는 태아 뇌하수 체의 신경성 활성으로 시작된다는 놀라운 사실을 발견하였다 . 이는 ACTH 를 유리하고 결국 부신에서 코르티솔을 분비하게 한다 (Thorborn 둥 1972). 태아에서 뇌하수체 활성은 뇌하수체의 문맥계로 부신피질 자 국호르몬 분비호르몬을 분비하는 궁상핵의 시상하부 신경에 의하여 촉발된다 (Brod a! 1981, 그림 11 .12 , 11 .13 , 11. 14 ). 태아에서 코르티솔이 증 가하면 에스트로겐의 생성이 중가하고 이는 자궁으로부터 프로스타글 란딘을 유리한다 . 이는 결국 자궁근을 감작하여 자궁 수축을 일으킨다 (Klop pe r 1974). 호모를 포함하여 영장류에서는 이러한 과정이 훨씬 복잡하다 (L iggin s 둥 1977). 그러나 분만의 개시는 역시 태아에 의하여 주도된다고 생각 한다. 태아막(양막 및 융모막)은 호르몬을 생성함으로써 분만의 개시를
돕는다고 생각한다. 그러나 분만의 정확한 개시 시간은 양에서와 마찬 가지로 태아의 부신기전에 의하여 조절된다고 생각한다 . 프로스타글란 딘은 뒤 에 참여 하여 분만이 시 작된 자궁 수축을 조력 한다 (Na t ha ni elsz 1978). 문제는 표 5 . 2 에 나타나듯이 진화 과정에서 분만의 시상하부에 의한 개시가 점차로 빠르게 되었다는 것이다. 여기서 주도적 역할은 편 도체 같은 변연계가 담당한다고 생각한다. 출산시의 위험성 때문에 자 연도태 기전을 통한 유전자 조작으로 출산 시기가 앞당겨지게 되었다 고 생각되지만 이것이 어떻게 초래된 것인지는 아직 알려져 있지 않다 . 5.6 호미니드의 인구학적 전략 영장류의 진화 발달 과정에서 회임기간과 생명의 점진적 증가를 그 림 5 . 5 에 보여주고 있다. 그림 5.6 은 수명과 본질적 인구 증가율을 감소 시키는 네 가지 요인과의 평형관게를 나타낸 것이다 (Love j o y 1981). 오 른쪽 눈금에서 1 은 정적 인구수이고 R=l 이면 인구수는 정적인 상태를 유지하고 R 값이 -1 이면 인구는 줄어들어 결국 소멸하게 된다. 현재 서 독은 R=0.6 이 고 스위 스는 R=0.7 이 라는 점 은 주목할 일 이 다. 침팬지는 호미니드의 기원 이전의 호미노이드 인구 전략상 가장 적 절한 모델이라고 생각된다. 침팬지의 성적 성숙은 10 세에 일어난다(그 림 5.5). 평균 출생 간격은 5 년 반이다. 여성의 불임 기간이 길기 때문 이라고 생각된다. 임신 기간은 34 주이고 유아 기간은 약 6 년이다. 4 년 반 내지 6 년이 수유기간이고 4 년 동안 어미에 매달려 보살핌을 받는 댜 어미가 나무에서 이동하는 동안 떨어져서 다치거나 죽는 일은 드 물다. 어미가 죽으면 어린것은 5 세 이내에 죽는다. 양자는 혼히 있는 일이 아니다. 양육에 있어 양식을 조달하는 일조차도 아비로서 할 일 이 없다 . 순전히 모성 중심사회이다. 침팬지는 보통 40 세를 살지만 일
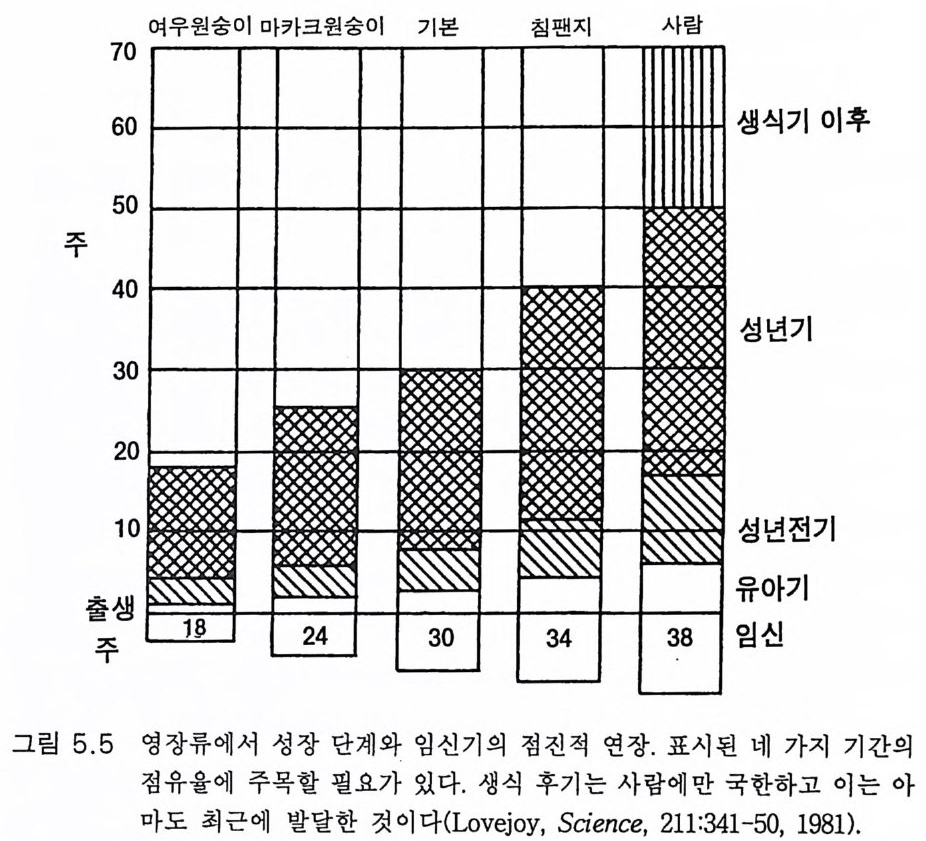 여우원숭이 마카크원숭이 기본 침팬지 사람
여우원숭이 마카크원숭이 기본 침팬지 사람
찍 죽는 수도 있댜 구달 (Goodall1986) 은 곰베 유역 보호구에서의 연구 로 방목군에서 도살이 자주 있음을 보고하였다. 5 년 동안에 50 마리 중 12 마리의 도살이 있었다 . 대부분은 높은 지위의 암컷이 낮은 지위의 새 끼룰 죽이는 것이다. 침팬지가 인구학적인 견지에서 근근히 종족을 유 지하고 있음을 그림 5.6 에서 알 수 있다. 비슷한 일은 오랑우탄에서도 볼 수 있는데 역시 모성 중심사회이고 여성의 불임기간이 길며 출산 간격은 약 7 년이다 보르네오 밀림에 사는 약 7000 마리는 가까스로 종
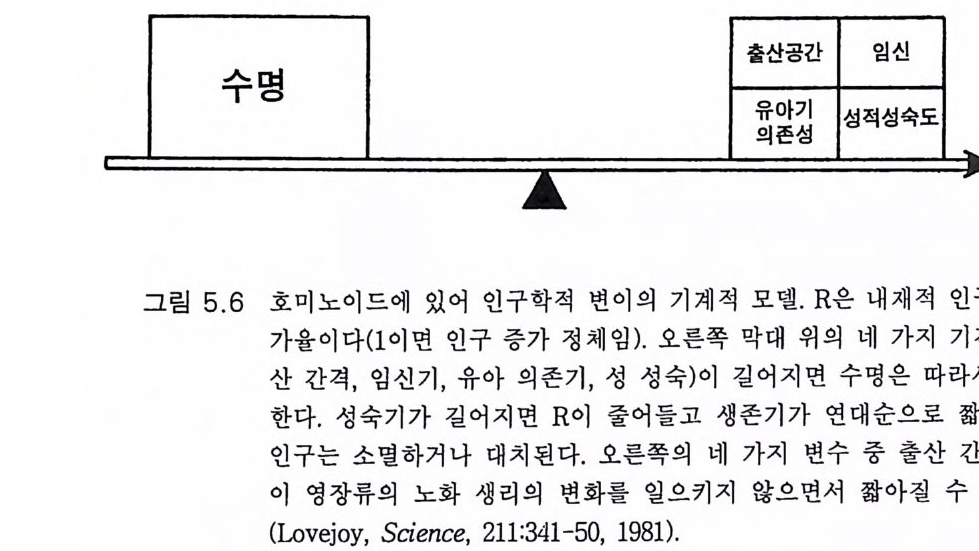 출산공간 l 임신 +
출산공간 l 임신 +
족을 유지하고 있다. 그러나 가장 작은 유인원인 흰손 기본 H y loba tes 은 사정 이 완전히 다르다 (W il son 1975: 528). 짝을 이루는 핵가족이 있고 네 마리까지의 새끼를 갖는다. 가족은 서로 가까이 있고 방어와 성교 전 성행위에 있어 암컷은 수컷과 동동한 역할을 한다. 어미는 어린것 을 돌본다 그러나 아비도 이에 참여하여 특히 큰 새끼를 돌보고 같이 놀아주고 보호해 준다 . 정상적으로 뒤섞여 있는 침팬지도 때로는 초기 짝짓기를 보인다 . 구달 (1986: 464) 은 완전하게 관찰한 짝짓기 행위 여덟 예를 보고하였다. 쌍을 이룬 것은 수일의 구애기간 동안 같이 지내기 위하여 무리로부터 떠난다. 러브조이 (Lovej o y 1981) 가 지적했듯이 인구학적 상황은 호미노이드 가 모험적으로 나무에서 내려와 두 발로 지상생활을 함으로써 개선될 수 있었다 . 두발걷기의 중요한 이점의 하나는 음식을 쉽게 상당히 먼 거리로 운반할 수 있다는 것이다. 이때 나무 또는 나무껍질로 만든 쟁
반을 사용했을 것이다. 이것이 오늘날에도 있는 수렵-농경사회의 기본 생활양식이다. 지상생활을 하려면 나뭇가지 또는 나무로 만든 보호 움 막 같은 생활 터전이 있었을 것이다. 이런 것들은 아무것도 화석으로 남아있지 않으나 보호 구조물의 기초로 생각되는 터 또는 돌들은 발견 된다 (Lumle y 1969). 아이삭(I saac 1978) 은 도구와 동물 뼈 무더기를 가 지고 약 2m y BP 의 공동 생활 터전에 관한 증거를 기술하였다. 음식을 나누어 먹었을 것으로 생각되고 남성은 음식, 때로는 먼 거리의 사냥 으로 얻은 육류를 조달하고 여성은 생활터전 근처에서 소동물, 먹을 수 있는 열매, 곡물, 나무뿌리, 나뭇잎 등을 수확하였을 것이다. 지상생활의 중요한 결과는 여성이 자식을 돌보는 일이 훨씬 수월해 진 것이댜 생활 터전의 둥지에서 어린것을 키우게 되었다. 어린것을 돌보는 일은 어미들끼리 나누어 할 수도 있었을 것이다. 이는 당시와 같이 위험한 시절에는 대단히 중요한 것이다. 그리고 어미가 죽더라도 고아는 길러질 수 있었을 것이다. 그러나 러브조이가 강조했듯이 가장 중요한 인구학적인 이점은 자식을 돌보는 일을 아비와 나눌 수 있었다 는 것이다. 이와 같은 대변혁이 갑자기 일어났다고는 생각되지 않는 댜 러브조이 (1981) 와 여러 사람들도 몇 가지 강화 요인이 있었을 것으 로 생각하였다. 첫째, 어미의 노고가 줄어들고, 음식을 나누어 먹음으로써 영양이 좋아져 차츰 여성은 현대인처럼 언제든지 성행위를 받아들이게 된다. 월경 주기까지도 이에 영향을 미쳤다. 결국 출산 간격은 줄어들었다. 이 성적 용납성은 에스트로겐 혈중 농도에 의해 초래되는 것으로 월경 주기 동안 계속 올라가서 월경 직전에 가장 높고 월경 후 떨어져서 1/3 수준이 된다. 둘째, 직립자세로 암수가 모두 음경, 유방, 음모, 액모 둥 그리고 성 적 유인 냄새의 분비물을 분비하는 외분비 ecc rin e 와 유출분비 a pocrin e 선 둥을 들어낸 성기 노출형이 되었다. 더욱이 몸을 뒤덮고 있던 털은
퇴화하여 매력적인 피부가 드러나게 되었다. 셋째, 음식나누기와 어린것 돌보기에 있어서의 연합은 양친으로서 의 역할을 분담하는 것으로 일부일처 짝짓기를 더욱 공고히 하였을 것 이댜 기본에서 먼저 나타나기는 하였지만 핵가족이 탄생되었다. 결국 생존성이 개선되었다. 그러나 짝짓기에 있어 수컷이 자식에 대한 생물 학적 부성을 인식하였으리라고 생각되지는 않는다. 성교는 그 자체가 보수였다. 출산 간격이 현대의 호모에서처럼 평균 2 년 반으로 줄어들 어 핵가족 내에는 동시에 여러 형제가 살았을 것이다 . 호모에서처럼 어린아이의 의존성은 15 세 이상으로 연장되었을 것이다(그림 5.5 ). 생식능의 이 같은 변화는 변연계에 의하여 좌우되었을 것이다. 예를 들면 어미의 노고가 줄어들어 시상하부에서 난포 자극호르몬 유리 인 자가 생성되어 혈중 에스트로겐이 계속해서 고농도로 유지되었을 것 이댜 난포 자극호르몬 유리 인자는 뇌하수체에서 난포 자극호르몬을 유리시키고 이는 난소에서의 에스트로겐 생성을 자극한다. 에스트로겐 농도가 높아지면 시상하부-변연계에 대한 작용으로 여성의 성욕을 촉 발한다. 에스트로겐을 주사하면 편도체 , 중격핵 그리고 내측 시상하부 (그림 5.7) 에 결합한다는 것이 증명되었다(Pf a ff 1980). 이들 신경핵에서 는 그림 5.3 에 보이듯이 대뇌피질로 신경섬유를 보낸다. 특히 대상회와 전뇌엽전부로 보낸다 . 성기 노출 또한 시각적 촉각적 및 후각적으로 성욕을 자극한다. 따라서 암수의 짝짓기가 강화되고 이는 핵가족 전체 로 퍼져서 결국 사랑의 감정으로 순화되었을 것이다. 그림 5.6 에서 이들 인구학적 전략으로 인구가 크게 중가하였을 것이 라고 생각할 수 있다. 그러나 약탈자로 인해 지상생활은 특히 어린것들 을 위험하게 하였다. 또 여러 가지 심각한 열대성 질환이 호미니드 진화 지역에 유행하였을 것이다. 솔레키 (Solec ki 1971) 는 샤니다르 Sh ani dar 에 는 단지 약 25 명으로 된 한 부족군이 있었음을 제시하였다 . 일부일처 짝짓기 사회에 대한 대안도 있다. 마이르 Ma yr가 가장 생
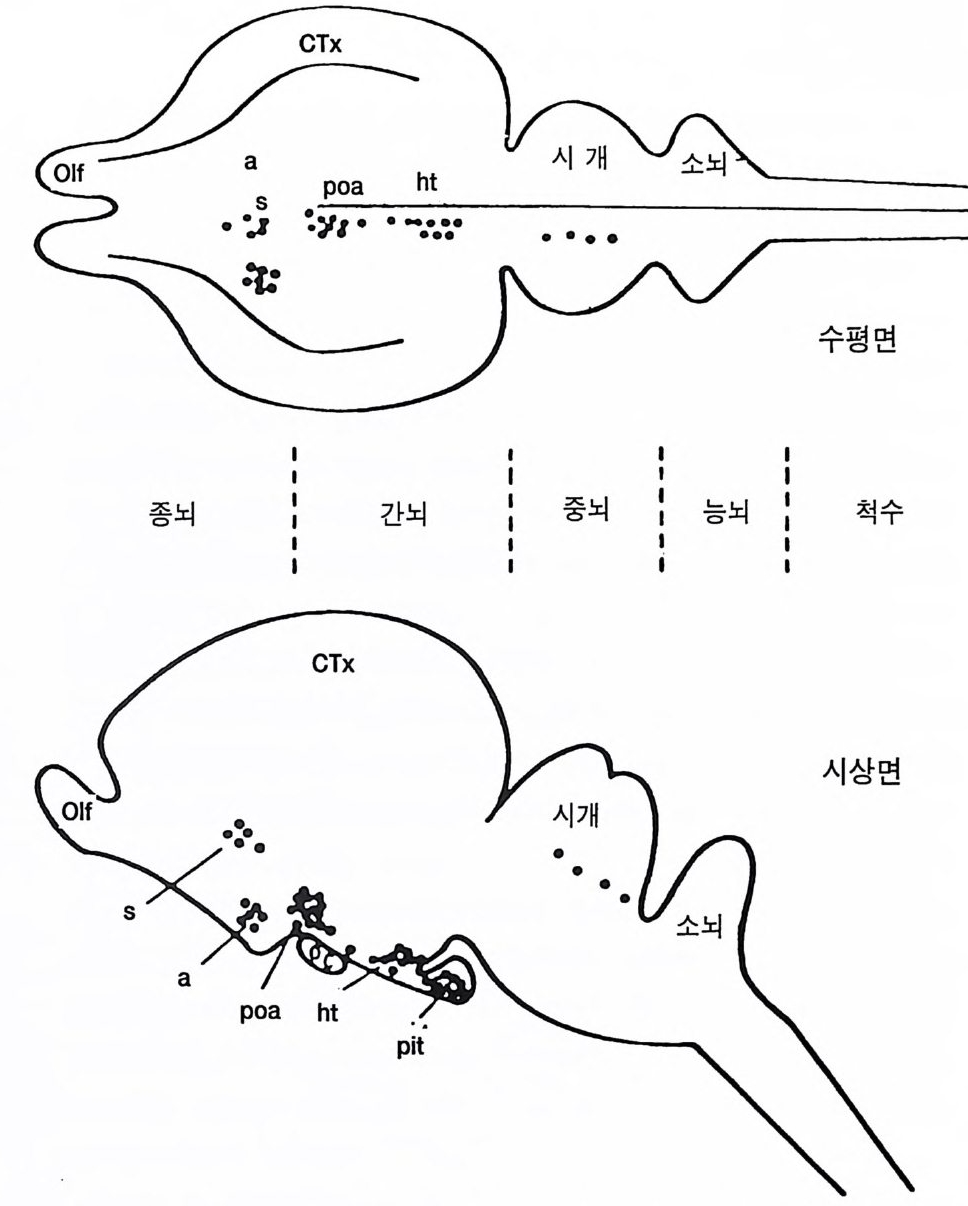 poa ht
poa ht
그림 5.7 일반화한 척추동물 뇌의 개략도. 현재까지 연구된 모든 척추동물에서 공통으로 보이는 에스트라디올 및 테스토스테론 함유 신경세포의 위치. 검은 점은 스테로이드 함유 세포군을 나타냄. 척추동물 공통의 3H- 에 스트라디올과 3H- 테스토스테론의 분포 양상은 변연종뇌(중격, 편도체, 원시선조체 둥), 시신경 교차전 영역, 결절성 시상하부핵, 그리고 중뇌 의 특수 개막하 좌위가 포함된다 .(a: 편도체 또는 원시선조체, cbllm: 소 뇌, cbc: 피질(비포유류는 일반피질, 포유류는 신피질), ht: 시상하부의 융기부에 있는 핵, oc: 시신경교차, OLF: 후구, pit: 뇌하수체, poa : 시신경 교차전 영역, s: 중격, t ec tum(비포유류는 시신경계와 하소구, 포유류에 서 는 상 및 하소구)(Pfaff 1980).
산적인 유전적 발전으로 간주한 일부다처p ol yg am y 의 경우 < 각 무리의 지도자가 굉장한 생식 기능의 우월성을 갖는다. 여기서 지도력을 가진 개인은 여러 처를 갖게 될 기회 를 최고로 누린다 >( 1973: 57). 그러나 이 같은 사회에서는 핵가족에 있어 아버지의 아이보기는 하지 않는다 . 이는 점차로 남자 아이는 도구나 무기를 만들어 사용하는 기술을 배우 고 자라는 것이 더 중요하게 된다. 따라서 어머니는 여자 아이에게 식 품을 수확하고 요리하며 위생 활동과 몸을 덮을 것 만들기 등을 가르 치게 된다 마이르는 생물학적 진화에 있어 유전적 우월성에 주안점을 두었다 그러나 호미니드 진화를 포함하여 생물학적 진화는 문화적 진 화로 대치된다. 그래서 핵가족의 진화적 우월성이 커지게 된다(1 0 . 3, 10.4 ). 핵가족에서 양친이 아이보기 역할을 분담함으로써 자식의 생존성이 증가하였음은 틀림이 없다. 이것이 바로 진화라는 것이다. 러브조이는 <핵가족을 초기 호미니드의 중심적 성취로의 막대한 적응으로 간주할 수 없는가?>라는 의문을 제기하고 다음과 같이 결론지었다 . 오히려 물질문화의 발달과 뇌 발달의 홍적세 가속화는 이미 형성되었 던 호미니드 성격 체계로의 귀결이었다. 여기에는 양친성의 강화, 사회적 유대, 일부일처 짝짓기, 특수한 성적 생식행위, 두발걷기 둥이 포함된다. 핵가족과 인간의 성행위는 홍적세의 태동보다 훨씬 이전에 그 궁극적 기 원이 있었음을 의미하는 것이다 .(1981: 348) 이 같은 일이 일어났던 시기는 오스트랄로피테신과 함께였을 것이 다(그림 2.5 ). 만일 러브조이가 주장했듯이 호미니드의 궁극적인 진화 전략이 핵가족이었다면 상호협조와 이타적인 행위는 이에 도움이 되 는 것이댜 이 같은 생각은 쉐링턴 She rri n gt on 이 생각했던 인간 이하의 생 활infr a-human lif e 과 매 우 대 조적 인 것 이 다.
그 세계는 < 힘이 정의 > 라는 법칙으로 지배되었고 고난 을 통해 건설된 댜 그러나 < 살아야겠다는 절규 > 로 이 모든 것 울 극복했다 . 우리가 이해 하는 한도에서 그 생 활을 재조명하면 우리가 그런 세계에서 벗어나게 된 것은 희열과 경탄할 일이다 .(1940: 377) 우리는 평화로의 진화적 이행을 가정하고 있지만 짚고 넘어가야 할 사실이 있댜 호모 하빌리스를 낳게 한 싹트기 방사 후 오스트랄로피 테신은 약 100 만년 전에 사멸되었다(그림 2.5). 그러나 이것이 반드시 호모속의 공격에 의한 것이라고 생각하지는 않는다. 그림 5.6 의 평형도 에서 R 값이 1 보다 작으므로 공격에 의하지 않고도 소멸하였을 것이다. 한 예로 우리는 질병의 발생률에 관하여는 모른다. 일부일처 짝짓기와 핵가족 가설은 호미니드 진화 초기에 대하여는 지나치게 이상적인 사회로 간주한 것일 수도 있다. 두발걷기로 집단생 활을 영위하고 음식을 나누었을 것이라는 증거가 있을 뿐이다. 그러나 호미니드 진화에 있어 변연계 신경핵의 상당한 증가가 있었고 이는 쾌 락적 및 우호적 행동과 관계가 있다 (5 . 2). 표 5 . 1 에 원원류 중격의 크기 지수 2.09 는 호모의 5.45 로 커졌고 외측 편도체는 3 . 05 에서 호모의 6.02 로 커졌음을 보여주고 있다. 이에 반하여 공격성과 관계되는 신경핵 내측 편도체는 크기 지수가 별로 증가하지 않아서 원원류는 1. 30 이고 호모는 2.52 이다. 진화 과정에서 자연도태로 유전자 부호는 개체발생 학적으로 분노와 공격성에 비하여 쾌감과 우호감에 관계되는 신경핵 이 중강되도록 짜여졌다고 생각된다. 오스트랄로피테신의 행동에 관한 기록은 오직 그림 3 .1 1 에 있는 발 자국뿐이다. 상상력을 동원하여 이룰 해석하면 이는 3.6my B P 시절의 핵가족에 관한 중거이고 짝을 이룬 어른이 주의 깊게 발자국을 겹쳐 걷고 어린것이 그중 하나의 손을 잡고 걸었다고 생각할 수 있다.
5.7 애타심의 진화 <애 타심 altr uism > 이 라는 말은 어 거 스트 콩트 Au g us t e Com t e 가 19 세 기 중엽에 소개한 것으로 정확하게는 어떤 개인적인 소득도 생각하지 않고 남에게 좋게 해주려는 특수한 도덕적 행위를 일컫는다 . 불행히도 사회생물학자들 (W il son 1975, 1978, Dawk ins 1976) 에 의하 여 많은 혼란이 야기되었다 . 그들은 어떤 개체가 일종의 희생적인 그 리고 같은 종족을 위한 것이라고 생각되는 행동 방식을 애타심의 관점 으로 보았다 . 인용한 대부분의 예들은 사회적 곤충에 관한 것으로 그 행동은 순전히 본능적인 것이다 . 그러나 그 같은 사회적 희생행동과 애 초에 정의되었던 애타심은 분명하게 구분할 필요가 있다 . 이 본능적 행 동은 <가성 애타심p seudo-al trui sm > 이라 하자고 제안하였고 (Eccles 1980, 제 8 장) 이는 이 책에서 다루고자 하는 진정한 애타심과는 전혀 관계가 없다 . 애타심에는 두 가지 특징이 있다 . 첫째는 근본적으로 의지가 있어야 한다 그 행위는 대개 자동적이기는 하더라도 계획된 행동인 것이다 . 둘째로 의도적 행동은 타인의 이익을 고려하여 계획된 것이어야 한다. 정상적인 인간 생활이 애타적 행동투성이임은 분명하다. 그러면 호미 니드 진화에 있어 이 같은 도덕적 행동이 언제 처음 있었는가라는 의 문이 제기된다 . 우선 고동동물의 행동을 살펴볼 필요가 있다. 돌고래가 상처 입은 동료에게 보인 사려깊은 행동에 대한 우화적 예가 있다. 그러나 와시 번 (Washburn 1969) 에 의하면 야생 영장류에서는 동정심의 혼적이 없 다. 투쟁에 의하여 먹이차지의 순위와 같은 지위가 결정된다. 침팬지 는 예외일 수 있지만, 강자는 언제나 남아있는 것을 참고 기다려야 하 는 유약자를 생각하지 않고 먼저 먹는다. 곰베 유역 보호구 내의 침팬 지 군에 는 동정 심 이 없는 것 같다 . 구달 (Goodall 1971) 은 소아마비 로 다
리가 마비된 성숙 수컷 맥그레거 McGre g or 가 어떻게 타격을 받는가를 보았댜 그는 무능상태로 하반신이 감염되고 가죽이 벗겨졌다. 그의 큰 고통에 대하여 아무런 감정도 갖지 않은 이전의 모든 동료들로부터 소외되었고 결국 그들이 보이지 않는 곳에서 죽었다 . 이제 호미니드로 돌아가서 애타적 행동의 첫 흔적을 찾아보고자 한 댜 집단 생활 터전에서 음식을 나누었다는 것이 애타심의 태동이라고 하겠다 (Isaac 1978). 그리고 핵가족이 애타적 사회의 시작이라 하겠다. 함께 걸은 360 만년 전의 도형 예가 그림 3 .1 1 에 있다 . 네안데르탈인의 매장 습관에서는 분명히 애타적 행동을 확인할 수 있다. 거기에는 의식적 자아로서의 자신과 타인의 인식이 있었다고 생 각된댜 이 같은 인식으로부터 살아있는 동안 남을 돌보고 죽었을 때 매장 의식으로 돌보는 사고의 흐름이 약 8 만년 전 네안데르탈인에 의 하여 행하여졌을 것이다(H awkes 1965). 시체는 음식 단지, 도구 또는 무기와 함께 묻었고 때로는 죽엄의 창백함을 제거하려는 짓으로 생각 되는 것으로 붉은 황토로 물들였다. 솔레키 (Solec ki 1971) 는 인류의 선사시절 (6 만년 전) 태어날 때부터 또 는 생후의 상해로 심하게 무능력했던 네안데르탈인의 골격으로부터 동정적인 행동의 첫 증거를 발견하였다. 이 무능력한 사람이 40 세까지 살았고(그림 2.10d 의 두개골) 이는 부족의 다른 사람들로부터의 보살핌 으로만 가능한 것이다. 동정심은 샤니다르 동굴시대의 놀라운 발견에 서 찾을 수 있다. 그 매장물은 꽃가루 분석으로 확인된 바로는 꽃으로 장식되어 있었다 (Solec ki 1971, 1977). 따라서 동정심의 가장 빠른 징조 는 인류의 선사시대인 6 만년 전이라고 할 수 있다. 적어도 10 만년 전일 것이라고 생각하는 사람도 있다. 네안데르탈인의 뇌는 현대인의 것과 크기가 거의 같기 때문이다(그림 2.1 0 , 2.1 3 ). 역사 이야기는 애타심의 사건으로 충만해 있다. 첫 기록은 기원전 2200 년에 수메르 Sumer 에서 쓰여진 길가메시의 서사시 E pi c of G ilg amesh 이다 .
이것은 지하세계에 바쳐진 동료 엔키도르 E nki dor 를 찾아서 삶을 되찾 아 주려는 주인공 길가메시의 고뇌를 읊온 것이다 . 위대한 종교적 행동 의 발로와 함께 애타적 행동의 교훈이었고 여러 가지 투쟁이 있기는 하 지만 우리 사회는 근본적으로 애타적임을 가르치고 있다. 불행히도 이 것은 출판계에서 빛을 보지 못하였는데 일반적으로 애타적인 것은 잘 팔리지 않는댜 그러나 주목할 만한 예는 캘커타의 테레사 수녀이다. 이미 5 . 6 에서 논했듯이 호미니드 진화에서 쾌감과 온순성에 관계되 는 신경핵의 크기 지수는 증가하였으나 공격성에 관계되는 내측 편도 체는 커지지 않았다. 그러나 이 신경핵도 인간 생활에서는 중요한 것 으로 용기, 모험심 그리고 조절된 공격성을 필요로 하는 헌신성의 동 기이기 때문이다 . 쉐링턴은 도덕성 그리고 애타심의 신비한 기원을 극적으로 보여주 었댜 생물학적 진화에서 나타났듯이 어머니 자연 Mo th er Na tur e 은 사 람에게 다음과 같이 강조하였다. 너는 내가 도덕적이라고 생각했다. 이제 너는 내가 도덕적이 아님을 알았다. 너는 내가 필요에 무감하고 기계적이어서 어찌 도덕적이라고 할 수 있는가고 말하지만, 나는 장구한 세월 동안 너, 도덕적인 너를 산출했 다 . 그래 너는 너의 세계에서 가장 도덕적인 사람이다 그래서 가장 부도 덕한 것이다. 너는 나를 지적이라고 생각했다. 심지어 슬기롭다고 생각했다 . 이제 너 는 내가 합리적이지도 않고 슬기롭지 못하다는 것을 알았다 . 어떻게 순전 히 기계적인 내가 이성이나 목적의식을 갖겠는가? 그러나 장구한 세월 동안 나는 너를 너의 이성을 가진 너로 만들었다. 네가 만일 조금만 생각 해보면, 이성을 가진 너는 이를 알 것이다. 너, 이 세상에서 유일하게 이 성적인, 따라서 유일하게 광기어린 것 …… 너는 나의 산물이다. 내가, 필 요에 눈먼 내가 어찌 사랑을 하랴? 나는 사랑할 수 없다 . 미워할 줄도 모
른댜 그러나 이제 나는 너와 너의 무리 를 만 들 었으므로 네가 네 자신에 대하여 새로운 세계라는 것 을 기억하라. 그 세계 는 너로 인하여 사랑과 미움, 합리성과 광기, 도덕과 부도덕, 선과 악 을 가진다. 네가 사랑해야만 사랑을 느낄 수 있다 . 죽 너희가 서로 사랑해야만 한다. 너는 또한 생각해 야 한댜 나 를 알고자 할진대 네가 목적 P urp ose 의 도구, 지금의 너의 소 견으로 가늠하기엔 너무 크다는 것을 알 수밖에 없으리라. 그러므로 너의 시야 를 넓히도록 노력해라(1 940: 399-400). 나의 대스승 쉐링턴의 이 심오한 글은 큰 감명을 준다. 생물학적 진 화는 단지 우연과 필연만은 아니라고 믿는다. 그것들만으로는 우리가 지닌 가치를 지닌 우리를 만들 수 없었으리라. 진화란 목적 P urpose 의 도구였을 것이라는 그의 의견에 동의한다. 그 도구는 진화를 최소한의 우연과 필연 수준에서 끌어올려 자아의식을 보유한 인간을 창조하는 초월을 이룩했다. 그후 문화적 진화 (10.2, 10.3) 는 생물학적 진화를 대치 하고 곧 자연도태 과정에 있어 결정적인 것이 되었다. 단지 기술적인 진보뿐 아니라 가치의 창조와 발달 때문이다. 예를 들면 애타심은 사 회 구성원의 복지에 헌신하는 사회의 도덕적 기초를 이루는 것이다.
제 6 장 시각운동 전화 —예술적 창조성 6.1 영장류 대뇌피질의 시각 영역 호미노이드로부터 호모 사피엔스에 이르는 시각운동계의 진화적 변 화를 검토함에 있어 유인원 뇌에서 시각정보 처리 과정에 관한 현재의 지식을 살펴볼 필요가 있다. 마카크 Macaca mula tta의 복잡한 시각계에 관한 연구가 가장 철저하게 이루어졌다. 이것은 다른 고동 영장류의 시각계와 기본적으로 대단히 유사하므로 이 시각계를 호미니드 진화 초기의 호미노이드 시각계의 모델로 간주할 수 있는 것이다. 영장류는 지극히 시각적인 동물이고 그 진화 과정에서 이미 유인원은 포유류 중 어느 것도 능가하였다. 그들의 눈은 앞으로 향하였고 완전한 양안시이 댜 시각 경로가 그림 6 .1에 있는데 시신경 교차에서 부분적으로 교차 하고 있어서 양쪽 눈의 우측 시야는 외측 슬상체 (LGB) 내의 접합연결 을 거쳐 좌측 후두엽의 일차 시각피질(브로드만 영역 17 또는 시각 영역 Vl) 에 투사된다(그림 4.4 ). 왼쪽 시 야는 그 반대가 된다. 일차 시각피질은 원숭이 대뇌피질에서는 대단히 많은 부분(1 6%) 을 차지하는데 사람 뇌에서는 4% 를 차지한다 (Van Essen 198. '5). 이는 사람
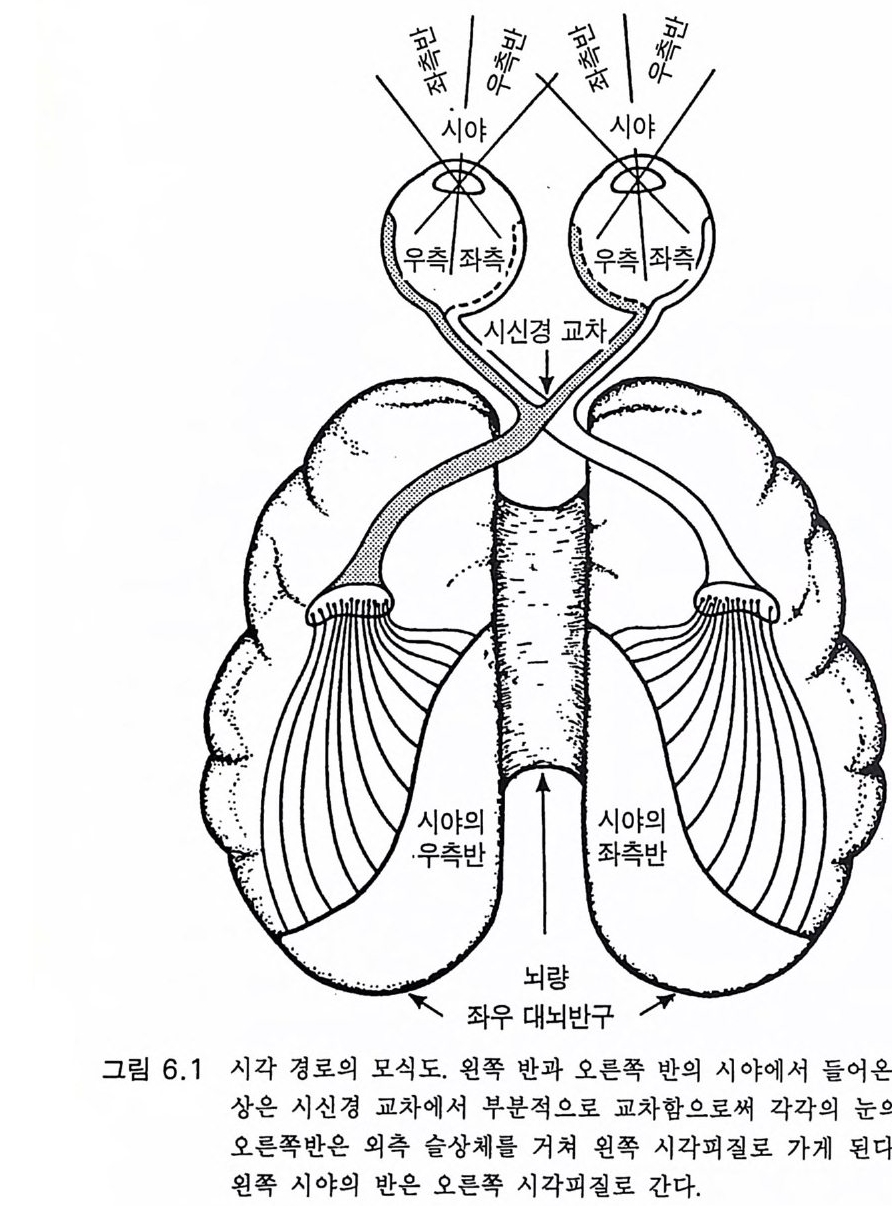 l1
l1
의 영 역 17( 그림 4.4) 을 원숭이(그림 4.5) 와 비교하여 볼 수 있다. 다음에 설명하겠으나 시각 정보의 처리는 영역 17 에서만 이루어지 는 것은 아니고 인접한 시각영역 18 및 19 그리고 측두엽과 두정엽의 인접 영역에서도 이루어진다 . 사람의 대뇌피질에서 시각영역에 관여하 는 부위를 모두 합하면 15% 가 되고 (Van Essen 1985) 원숭이에서는 최 소한 30% 가 된다. 6.2 일차 시각피질에서의 신경 반응 미세전극을 이용하여 단일 신경세포에서 발생하는 신경 충격을 기 록할 수 있다. 예를 들면 휴벨과 비 셀(H ubel 과 Wi es el 1962) 은 일차 시 각피질에서 성공적으로 기록하였다. 그림 6.2a 는 일차 시각영역에 삽 입된 미세전극으로 단일 신경세포에서 발생하는 임펄스를 기록한 것 이다. 그림 6.2b 에서 사선으로 나타낸 삽입 궤도에 횡선으로 표시된 것 이 그 궤도상에 있는 여러 단일세포이다. 미세전극을 주의하여 위치를 정하면 세포 밖에서 단일세포 활동전위를 기록할 수 있다. 세포들은 느린 배경방전(그림 6.2a 의 맨 위)을 보이나 빛의 띠가 망막을 지나가면 빛이 망막의 특정 지역을 지날 때 그림의 왼쪽에 보이듯이 세포의 격 렬한 방전이 나타나고 빛의 띠가 그 지역을 지나자마자 방전은 죽시 정지된다(그림 6.2a 의 밑쪽 기록). 빛띠의 방향을 회전시키면 그림의 가 운데 기록에서처럼 방전의 강도가 감소한다. 끝으로 빛의 각도를 직각 으로 회전시키면 어떤 효과도 나타나지 않는다(맨 위쪽 그림). 이는 특 정 세포는 한쪽 방향의 빛띠의 움직임에 특히 민감하고 이것에 대하여 직각인 빛띠의 움직임에는 감수성이 없다는 것을 말한다. 그림 6.2b 에서 미세전극이 지난 자리를 가로지르는 선들의 방향은 감수성의 방향을 나타낸 것이다. 위쪽의 12 개 세포군은 동일한 감수성
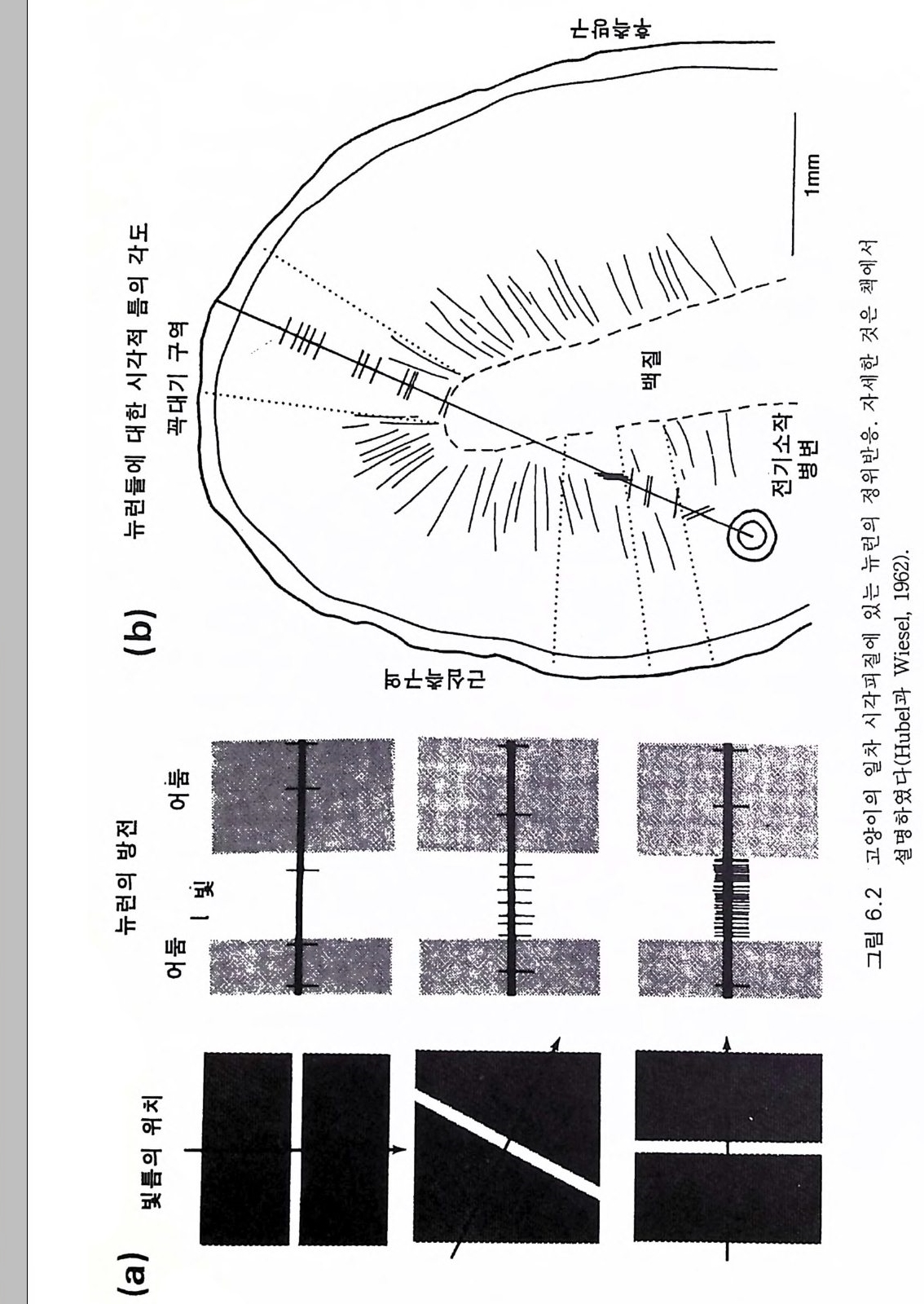 t如째아
t如째아
방향을 보이는데 궤도 를 따라 종대로 달리는 이들 세포는 표면에 대하 여 직각 방향을 이루고 있음을 알 수 있다. 그러나 중심 회백질을 지 나서는 세 개의 세포군이 전혀 틀린 감수성 방향을 나타낸다. 궤도는 분명히 점선으로 표시된 서로 다른 감수성 방향의 세포 종대를 지나고 있다. 시각피질에서 비슷한 감수성 방향을 가진 신경세포는 피질 표면에 대하여 직각으로 종대를 이루는 경향이 있다. 따라서 사람의 광범한 일차 시각피질에는 약 4 억 개의 신경세포가 조각보 모양으로 종대를 이루고 있고 수천 개의 신경세포가 있는 각각의 종대는 동일한 감수성 방향을 갖는 것 임 을 그려 볼 수 있다(H ubel 과 Wi es el 1963). 이 같은 배열은 망막 영상 재구성의 첫번째 단계로 생각된다. 이 같 은 방향성은 물론 망막 시계도와 합치되고 이 시계의 각 구역이 종대 를 이루어 빛띠의 방향과 명암의 경계를 판별하게 된다. 동측 및 반대측 눈은 다같이 LGB 를 거쳐 시각피질에 입력됨은 이미 그림 6.1 에 보여주었다. 그러나 영장류에 있어 이들 입력은 각각 다른 판에 의하여 중계되는데 동측에 대하여 세 개 (2 i, 3i, 5i) 그리고 반대측 눈에 대하여 세 개 (le, 4c, 6c) 가 있다(그림 6.3 ) . 이 그림에는 영역 17 의 종 렬 로 가는 입 력 을 대 단히 도식 화한 것 이 다(H ubel 과 W ies el 1974). LGB 의 동측 및 반대측 판은 선, 종렬 또는 시각적으로 우세한 종렬이 교대하여 입력된다. 이와는 직각 방향으로 또 다른 띠 모양의 배열이 정위 특이성에 의해 결정되는데(그림 6.2) 이는 그림 6 . 3 에서 보이듯이 대뇌피질의 상충 표면에서 정위 특이성이 점차 회전하는 것으로도 알 수 있다. 종대의 구성은 실제 원숭이 피질에서는 이 그림에서처럼 엄 밀하게 배열되어 있지는 않다. 그림 6 . 3 의 판 IV 상부에는 단순세포가 있는데 이는 엄격하게 단안 시 monocular 이 고 그림 6 . 2 에 서 보듯 선 또는 모서 리 에 단순하게 반응 하는 것이다. 영상 재구성의 다음 단계에는 영역 17 그리고 주위의 이
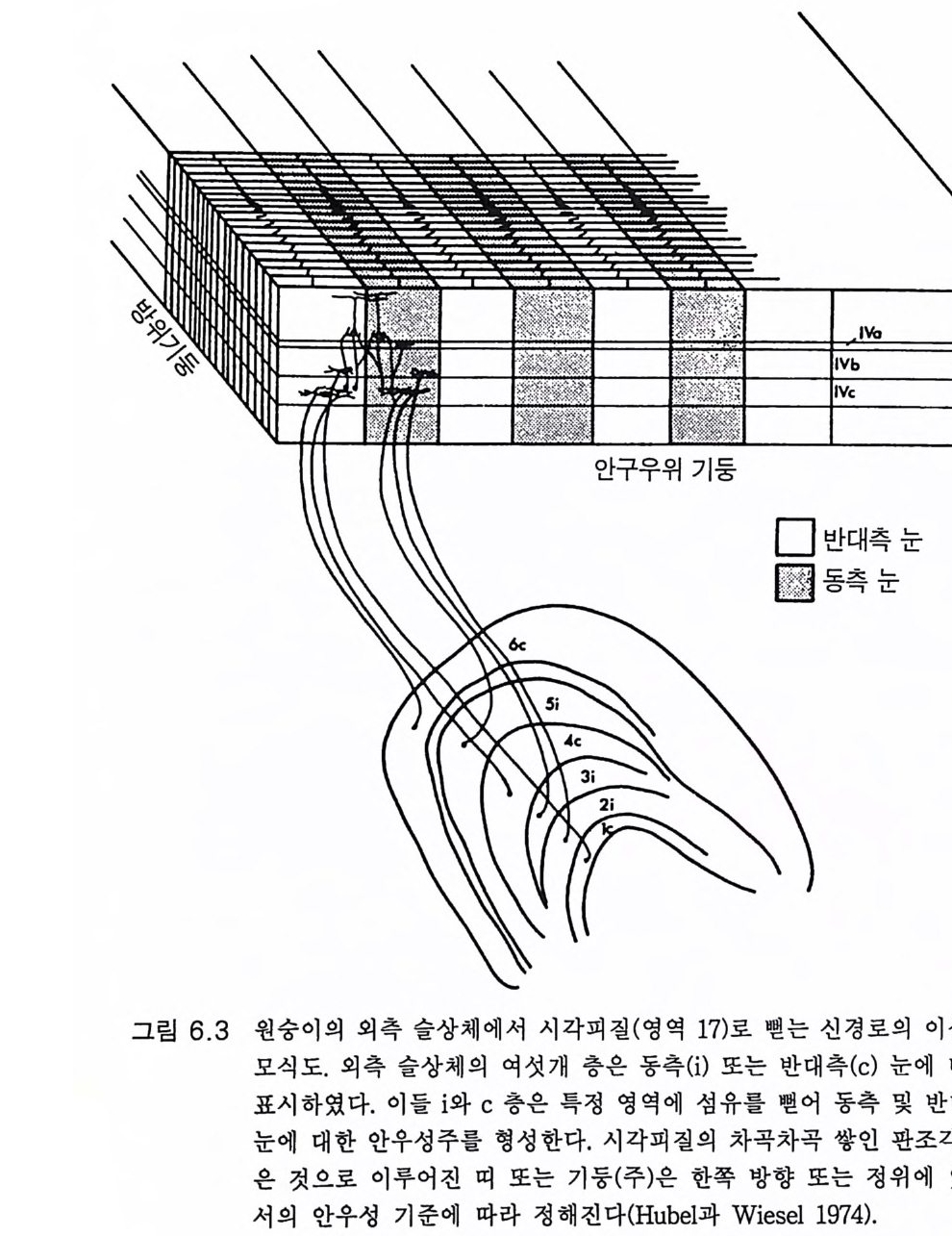 IVo
IVo
차 및 삼차 시각영역(브로드만 영역 18 및 19, 그림 4.4 및 4 . 5) 의 다른 수 준의 신경세포가 있다. 이들 신경세포는 밝거나 어두운 선의 폭과 길 이 그리고 그 방향 심지어 두 선이 만나는 각도에 특히 민감하다 . 이 들 소위 복잡 및 과복잡 신 경 세 포(H ubel 과 Wi es el 1963, 1965, 1977) 는 형체 인식의 다음 단계를 이룬다. 그림 6.3 에 상판에 있는 두 가지 복 잡 세포의 예가 있는데 각각은 서로 다른 시각적 우세띠의 두 개 단순 세포로부터 받는다 . 6.3 입체시 이미 그림 6.3 에서 나타냈듯이 각각의 눈은 LGB 의 6 개관 중 3 개에 입력되고 나아가 일차 시각피질에 입력된다. 이 같은 방법으로 동측 및 반대측이 서로 엇갈려 눈지배띠를 이룬다. 그래서 시신경 교차에서 부분적으로 교차하지만 왼쪽 또는 오른쪽 눈으로부터의 신경섬유는 각각의 시각 경로를 따라 섞이지 않고 LGB 의 특정 판에 분포하고 일 차 시각영역의 눈지배띠에 입력된다. 인접한 눈지배띠의 일차 수용세포는 보다 표충에 있는 공동의 추 상세포에 입력한다(그림 6.3 ). 두 눈은 60-70mm 의 동공간 각격을 갖 고 삼차원 세계를 보게 되므로 두 망막 영상 간에는 차이가 있을 것 이다 . 이 차이는 입체시 S te reo p s i s 의 근거로서 결국 인접 눈지배띠에 도달한다. 입체시의 초기 연구는 두 눈으로부터 입력되는 차이에 특히 민감한 일차 시각피질의 신경세포를 밝히는 데에 경주되었다. 눈을 고정한 마 취 된 고양이 실험 (Barlow 둥 1967, Bis h op 1970, Hubel 과 Wi es el 1970) 으 로 두 눈의 차이룰 감별하는 시각피질(영역 17) 의 단일 신경세포를 발 견하였댜 포지오 Po ggi o 와 그 동료들은 정교한 실험기법을 이용하여
이 연구를 계속하여 훈련된 마카크 원숭이를 마취하지 않고 철저하게 연구할 수 있었다 (Po ggi o 와 Fis c her 1977, Pog gio 1984). 여 러 방향의 한 개의 밝은 막대(그림 6 . 2) 를 시계를 가로질러 움직였다. 고정점을 기준 으로 여러 거리에서 한 방향 또는 다른 방향으로 움직였다. 브로드만 영역 17 과 18 에서 단순 및 복잡 세포의 상당 부분이 수평의 양안차에 대하여 상이한 감수성을 보였다. 양안차에 대한 반응에는 여러 종류가 밝혀졌지만 지금 우리가 논하고자 하는 바를 위해서는 시각 정보의 피 질 처리과정 초기에 신경세포의 거리 인식이 있는데 각 눈의 이차원적 망막 입력은 양안시의 차이로 변형되어 시각영역 Vl 및 V2 에서 반응 을 일으켜 신경세포는 거리를 감지하는 특이적 정보를 지니게 된다는 것을 인식하는 것으로 충분하다 (Poggi o 와 Pog gio 1984). 단지 20 % 의 신 경세포에서 보일 뿐이지만, 이 같은 변형은 산재점으로 된 입체도에서 도 일 어 난다 (Poggi o 1984). 고로 시 각정 보의 피 질 처 리 과정 의 가장 초 기 단계에서 이미 입체시의 뇌 c y clo p ean br ai n 가 출현한다. 그러나 어 떻게 3 차원적 정보가 3 차원 체험으로 변형되는지는 알려져 있지 않고 다른 여러 요인들이 관련되어 있다. 입체시는 학습을 통해 얻어진다 . 양안시는 수상생활을 하는 영장류에서 대단히 발달되었다 . 양안시는 모든 수상생활 유인원 중 말레이시아의 작은 영장류인 기본종들에서 가장 발달하였다. 우리의 가장 뛰어난 체조선수 , 발레 무용수 또는 육 상선수라도 기본의 재주에 비하면 얼마나 될까를 의심케 된다 . 기본의 브로드만 영역 (Mauss 1_ 911) 을 살펴보면 시각국은동세포가 특히 많이 분포하는 두정엽 내에 어떤 특별한 부위가 발달해 있는 것 같지는 않 다(그림 6.5 ).
6.4 선조전 시각영역 원숭이 대뇌피질의 후방 1 /3 은 시각영역들이 조각보처럼 배열되어 있는데 각각의 영역들은 망막 중심적 부위를 나타낸다. 그들은 형태, 차이, 방향, 움직임, 색깔 등 특수한 시각 기능에 대한 반응의 분석에 관 계 된 다 (Ze ki 1969, 1973, 1978, Coway 1982). 마카크 원 숭 이 의 영 역 Vl 에서보다 상위 영역으로 보내는 시각정보룰 분석하는 데는 세 가지 방 법이 주로 이용된다 . 첫째는 손상 후의 퇴행을 관찰하는 고전적 방법 이고 둘째는 신경섬유의 경로를 방사성 물질로 추적하는 방법이다. 예 롤 돌면 방사성 물질을 영역 Vl 에 주사하면 인접한 영역 V2 에서 강한 방사능을 나타낸다. 그리고 V2 에 주사하면 다른 두 가지 시각영역에 분포함을 볼 수 있다. 죽 하측두 영역에 있는 V3 와 V4 그리고 상측두 구에 있는 MT 이다(그림 6.5 , 6.6 ). 셋째는 가장 중요한 것으로 선택된 시각 입력에 대한 단일 신경세포의 반응을 기록하는 방법이다. 그림 6 .4는 마카크 원숭이의 좌측 대뇌피질 외측 및 내측면의 V2, V3, V4 의 위치룰 보여주는 것이다. 이들은 위의 방법들로 밝혀진 것이 다 그림에 월상구는 열려서 깊숙이 V2, V3, V4 가 보이도록 그려졌다. 이 돌기는 시각 자극의 양상과 특별한 관계가 있다. 예를 들면 영역 V4 의 세포는 색깔 인지에 선별적으로 민감하다 (Ze ki 1973, 1980). 그리 고 V2, V3 세포의 70 %는 방향 특이성을 보인다 (Ze ki 1978). 그림 6.5 에는 Vl 에서 상측두구 깊숙이 있는 MT 영역에 주로 투사되 는 것을 보여주고 있다. MT 영역에서 위쪽 네 부위로 투사되는 것이 그림 6.6 에 있다` 두정간구에 있는 두 부위는 그림 6.5 에 열려져 보여주 고 있고 이 두 부위는 브로드만 영역 7 에 있다. 그림 6.5 의 MT 복합영 역은 움직임의 방향과 주변과의 관계를 신호한다. 즉 이것은 공간시각 기능을 갖는다 . 여기서 마운트캐슬 Moun t cas t le 과 그 동료들의 원숭이 브로드만 영역 7 에 있는 신경세포에 관한 수년간에 걸친 연구를 상기
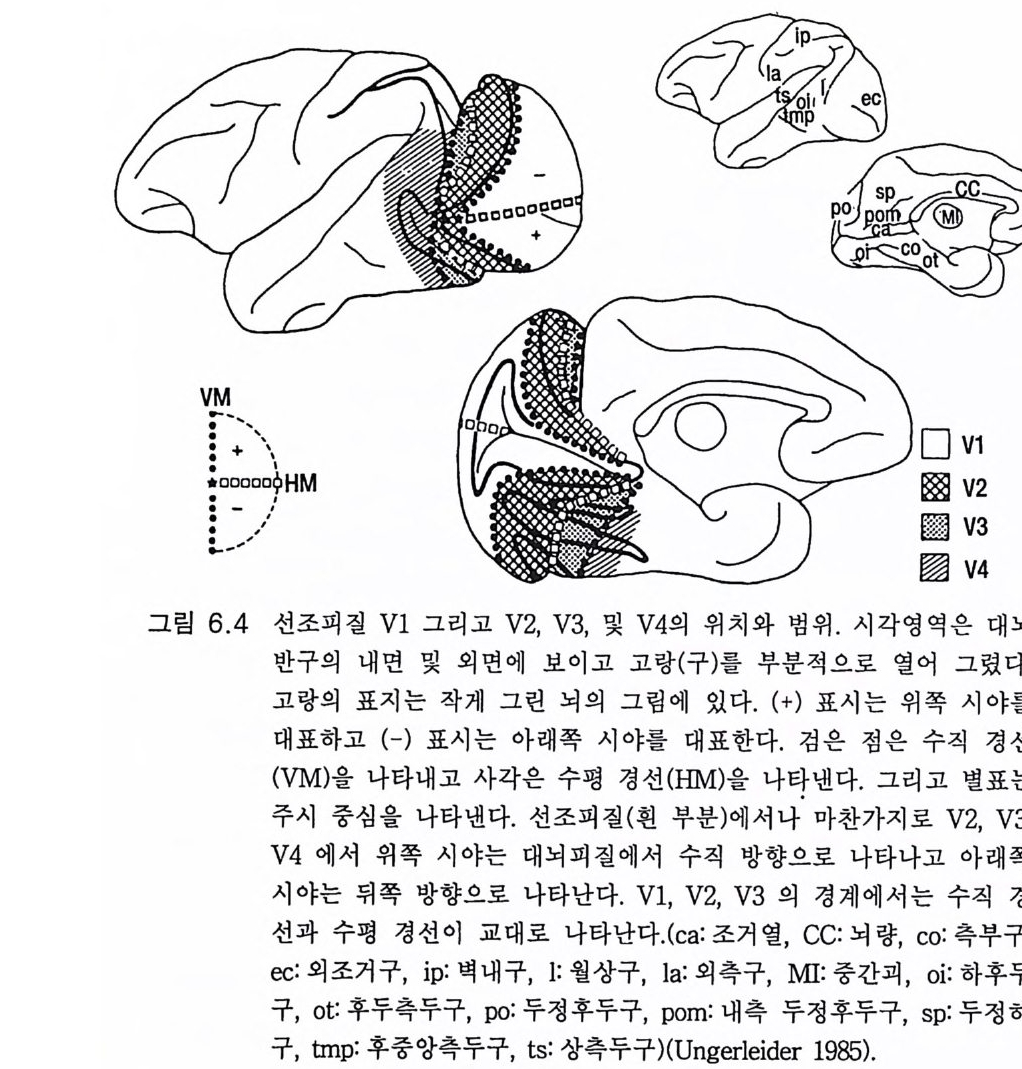 V...•.•-..••••M - o - o +-`,o,` o \, ’o`, ` , ❖I, HM 回霧□臨 wwww
V...•.•-..••••M - o - o +-`,o,` o \, ’o`, ` , ❖I, HM 回霧□臨 wwww
할 필요가 있다 (Moun tc as t le 등 1975, 1984). 이들 신경세포는 시각 입력 에 민감하고 특히 움직임에 관계되고(시각 추적 신경세포) 시각 공간에 관계되며 신체상의 형체를 감지하는 것에 연계된다.
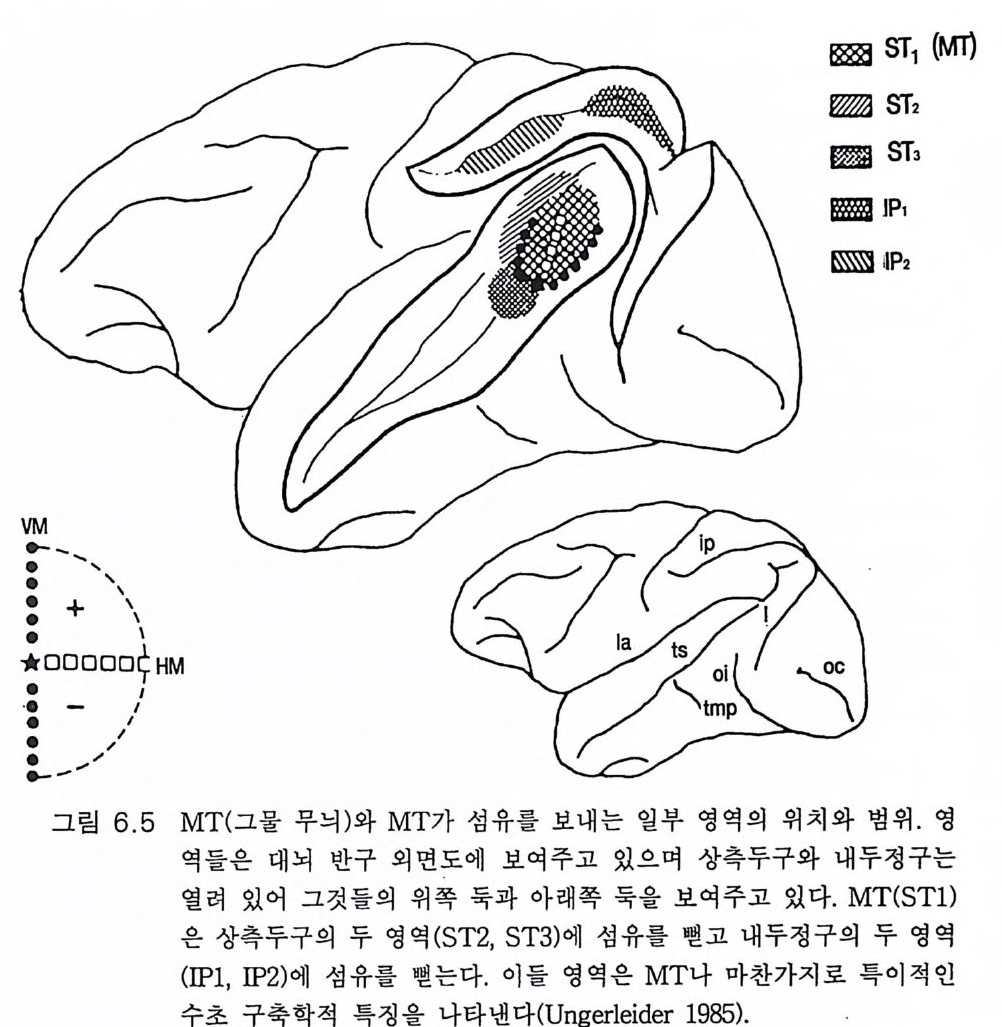 ~ ST1 (MT)
~ ST1 (MT)
반 에센 (Van Essen 1985) 은 Vl 으로부터 여러 선조전 영역으로의 투 사와 이들의 상호연결에 관한 복잡한 연구 결과를 발표하였다. 그러나 이것은 운게르라이더{U n g erle i der 1985) 의 단순한 도형(그림 6 . 6) 과 근본 적으로 다룰 것이 없다 . 마카크 원숭이에게 사용하였던 것과 같은 방법으로 올빼미 원숭이
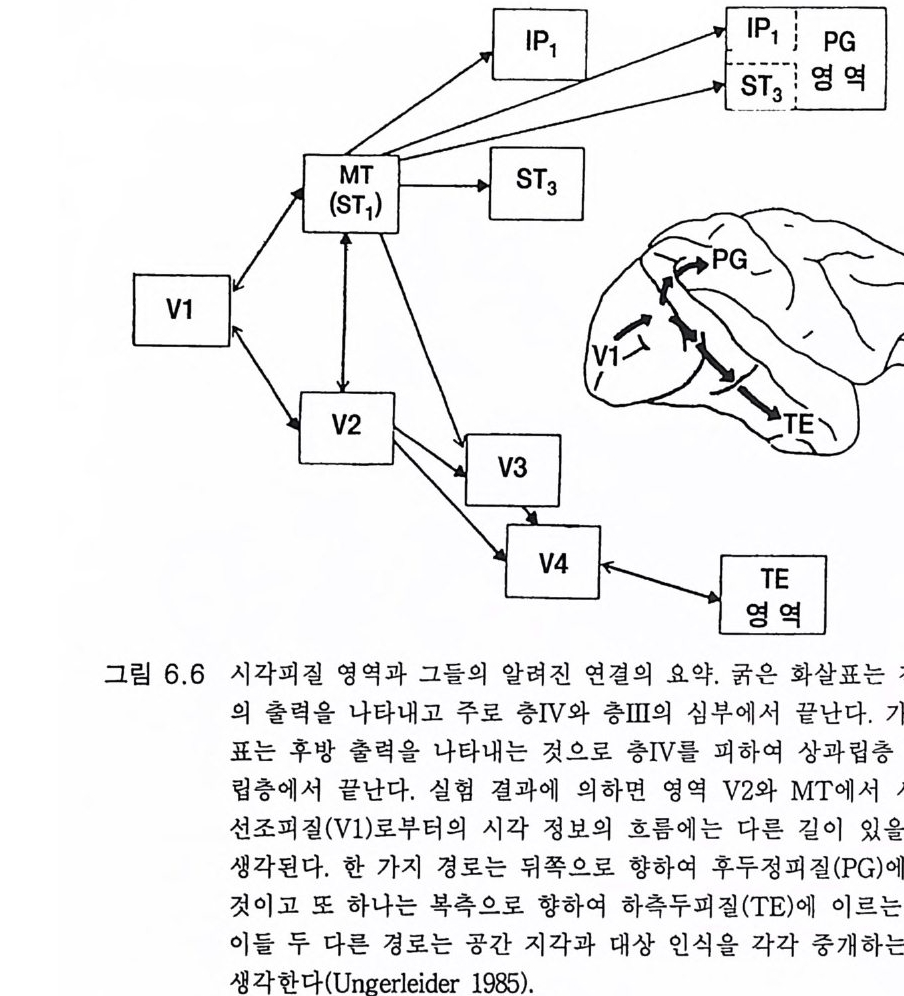 그림 6.6 시각피질 영역과 그들의 알려진 연결의 요약. 굵은 화살표는 전방으로
그림 6.6 시각피질 영역과 그들의 알려진 연결의 요약. 굵은 화살표는 전방으로
의 대뇌 시각계통을 연구하였다 (Baker 둥 1981, Coway 1982, Van Essen 198. 5). 이 동물의 연구상 이점은 대뇌피질이 홈으로 깊이 파여 있지 않 으므로 연구하기 쉽다는 것이다. 여러 선조전 구성분이 확인되었고 일 부는 마카크의 것과(그림 6.6) 일치하였다 (Baker 둥 1981, Van Essen 198. 5). 유인원류에서 비교 연구는 별로 없다. 그러나 구조적 측면에서 (그림 4.5) 선조전 시각영역의 비슷한 복잡 구조를 추정할 수 있고 호
미니드 진화 계통상의 호미노이드 조상에 이를 적용할 수 있다. 선조전 시각영역의 이 연구는 최근에 이루어진 것이므로 시각정보 를 처리하는 복잡한 신경망의 기능적 의미를 아직 완전하게 이해하지 못하고 있다. 그림 6.6 에도 대단히 복잡한 분포 상황을 나타내고 있다. 시각 입력의 이 같은 특수한 양상을 고려함에 있어 시각 자극의 망막 국소적 특이성은 영역 Vl 에서의 정밀성과는 상당한 거리가 있음을 주 목해야 한다. V2, V3, V4 그리고 MT 의 단일 신경세포는 광범한 망막 입력을 수용하는 것이다 . 측두엽의 TE 영역(그림 6.6) 에는 형체 탐지 신경세포가 있다 (Gross 1973, Gross 등 1985). 예를 들면 네모, 직각, 삼각, 별 모양에 특이적으 로 반응하는 신경세포가 있다. 더욱 놀라운 것은 얼굴이나 손 그리고 그 부분들에 특이적으로 반응하는 적은 수의 세포가 있다는 것이다. 따라서 TE 영역에는 일정 형태에 관한 시각정보를 놀라울 정도로 선 택적으로 처리하는 신경세포가 있는 것이다. 어떤 경우에는 TE 영역 이 또 다른 기능 예를 들어 그 동물에게 그 시각자극이 중요한가 아닌 가를 신호하기도 한다. 망막에서 피질영역 20 및 21( 그림 4 .4)에 이르는 시각정보 처리의 각 단계는 순서적 배열의 양상으로 서열이 매겨져 있는 것으로 생각된다. ® 시 계는 점차로 특이성 이 감소된다. 이 같은 일반화의 중가는 영 역 20 및 21( 그림 4.4 , 6.4 , 6.6) 의 모든 신경세포들이 중심와의 시각자극을 반 영하는 것으로 초래된다 . 또한 이 단계에서는 모든 신경세포는 뇌량 팽대 (그림 6 .1)를 거쳐 양쪽 후두엽으로 오는 입력을 통하여 중심와를 포함하 는 양쪽 반시계 모두에서 신호를 받는다. ® 적절한 자극의 특이성은 중가한다. 점에서 밝은 선으로, 또는 특정 한 방향으로 그리고 선의 특정한 폭과 길이 때로는 특정한 방향으로의 움직임 그리고 영역 20 및 21 의 일부 신경세포의 보다 복잡한 형체 탐지
등이댜 ® 영역 20 및 21 의 신경세포는 추가적인 반응 성질을 가졌다는 증거 도 있댜 죽 동물에 대해 그 시각자극의 유의성 여부에 따라 반응한다. 6.5 선조 및 선조전 영역의 손상 원숭이 일차 시각영역의 손상은 시각 국소적으로 시계상에 예상되 는 결함을 일으킨다. V2 영역의 일부를 제거하면 중심 시력 또는 주변 시력의 예상되는 결함을 나타낸다 (Cowa y 1981, 1982). V4 영역을 선택 적으로 제거하면 색상 판별 불능도는 증가하나 방향 판별 불능도는 증 가하지 않는다. TE 영역을 제거하면 시각 학습의 장애가 초래되나 시 각 자체 는 영 향을 받지 않는다 (Cowa y 1981, 1982). 사람의 시각 계통이 원숭이의 것과 유사한지를 알아보려면 선조 또 는 선조전 피질의 국소적 손상으로 인한 환자의 증상을 기록 수집해야 한다 윤리적인 문제로 원숭이에게서와 같은 방법으로 사람의 시각계 통을 실험 할 수는 없다(퇴 행, 방사성 물질 추적 , 단일 신경세포 기록 등). 이 같은 난점에도 불구하고 사람에서의 연구는 관심의 대상이 된다. 인식, 활동, 생각, 감정 둥을 포함하는 시계의 분석은 그것에 대한 우리 의 반옹, 예를 들면 인지, 반응 동작, 개념 형성, 감정 유발 등의 시작점에 지나지 않는다. 동일한 자극이라도 그것의 학습 경험에 따라 다른 반옹을 유발할 수 있다 (Cow~ y 1981: 405). 불행히도 사고에 의한 뇌 손상은 특정한 시각영역에 국한되는 경우 가 거의 없다. 그러나 뇌 손상의 여러 보고를 종합하면 마카크 원숭이 에 실험적으로 손상을 일으켰을 때 나타나는 중상과 합치시킬 수가 있 다 (Cowa y 1981, 1982, Wa ning ton 1985).
양측성 후두엽 손상으로 인한 피질성 실명의 경우 완전한 실명은 드물고 시력, 형태 , 색상, 위치, 입체시 동 시각 기능이 부분적으로 손 상되므로 환자는 여러 상대적인 장애를 나타낸다. 일측성 손상은 일측 성 시각장애를 일으킬 수 있다. 외복측 선조전 영역의 손상은 색맹을 초래하여 색시가 심하게 손상되거나 완전한 색맹이 된다. 그러나 망막 피질섬유의 삼색기전은 온전하다. V4 의 손상으로 추측되는 것이다. 의 배측 두정엽의 손상을 입으면 대상물의 위치는 잘못 인식하나 색깔과 모양은 제대로 인식한다. 우측 측두엽의 손상은 시각 인식의 결함을 초래한다 코웨이 (Cowe y 1982) 는 사람에서 시각 감각의 결함은 없으면 서 복잡한 대상 또는 얼굴의 시각적 인식불능증 v i sual a g nos i a 이 있던 경우를 기술하였다. 예를 들면 얼굴의 인식 장애인 상모실인증 p roso p a g nos i a 은 내 복측 후두측두피 질 의 손상으로 나타난다. 이 부위 는 마카크 원숭이의 일부 신경세포가 얼굴의 시각 입력에 반응하였던 TE 영역에 비하면(그림 6.6) 조금 후방에 있다고 생각된다. 이와 비슷한 증 후에는 대상 인식불능증이라고 하는 것이 있는데 여기에는 여러 아형 이 있다. 시각 입력 또는 움직임에 대한 브로드만 영역 7( 그림 4 .4)의 신경세 포반응 (Moun tc as t le 둥 1975, 1984) 은 주변 공간의 존재를 주체에게 신 호한댜 이 영역이 손상된 환자는 공간의 반대측 반쪽에 있는 대상의 위치룰 인식하는 데 장애가 있다. 그들은 해당 공간에 있는 사건과 사 물, 심지어는 그쪽에 있는 자신의 신체 부위에 주의를 집중시키는 데 결함을 보인다. 이와 비슷한 증상들이 원숭이의 MT 영역 및 관련 영 역(그림 6.6) 에 손상이 있을 때도 나타난다. 그들은 도달동작 reac hing에 서 오류를 나타내고 대상의 공간적 관계 인식에 결함이 있다. 반 에센 (Van Essen 19 8.5)은 사람의 선조전 영역은 유인원에 비하여 대단히 크기 때문에 여러 특수 영역을 더 많이 보유할 수 있다고 하였 다. 그러나 이는 단지 추측일 뿐이다. 필요한 것은 사람의 뇌를 연구하
는 방법의 향상이다. 예를 들면 새로운 조직학적 기술, PET 또는 NMR 같은 주사 영상기기에 의한 국소 해부학적 기술 등이다. 방사성 제논 Xenon 법은 이미 값진 증거들을 제공하고 있다(그림 6.8, 6.9 ). 6.6 시각반응에서 사람 뇌의 연구 특수한 판별적 상징적 시각 과업을 수행할 때 사람 뇌의 부위별 활 동 상황을 연구할 수 있게 되었다. 가장 의미 있는 결과는 롤랜드 Ro land 와 그 동료들의 연구 결과이다(R oland 와 Lassen 1976, Roland 와 Sk inh ~j 1981, Roland 와 Fri be rg 1985, Roland 1985). 뇌 의 한쪽 반구 에 혈 액을 공급하는 주동맥인 내측 경동맥에 방사성 제논 (133Xe) 을 주사하는 방법을 이용하였다. 그림 6.7 에서와 같이 254 개 가이거 측정기를 포진 하고 주사 후 약 40 초 동안 한쪽 대뇌 반구의 254 개소에서 방사능을 측 정하였댜 각 구역은 약 lcm2 였댜 눈을 감고 귀를 막은 완전한 휴식 상태에서 부위별 뇌혈류량 (rCBF) 의 대뇌피질 지도를 만들 수 있었다. 그러고 나서 일종의 시각과업을 수행하는 동안에 측정을 반복하여 그 과업으로 인한 변화를 알아보았다 . 특정 시각 과업에 의해 변화가 유 발되는 부위를 찾아내기 위하여 여러 대조군과 시각 과업군에서 측정 하였다. 이렇게 해서 발견된 모든 변화는 혈류량의 중가를 나타내었는 데 이는 신경 활동의 증가를 뜻하는 것이다. 그림 6.7 은 편심 정도가 서로 다른 두 개의 타원을 구별하는 과업을 준 실험 설계이다. 방사성 제논을 주사한 후 40 초 동안 가이거 측정기 로 측정하면서 실험 대상은 타원 l 또는 2 를 마음대로 자신의 결정에 따라 보게 하였다. 타원을 보는 동안 당연히 눈은 움직인다. 그러나 그 외의 움직임은 전혀 없었고, 1 인가 2 인가를 판별하여 표현하는 것은 실험이 끝난 후에 허용되었다. 통계적으로 유의한 부위별 뇌혈류량의
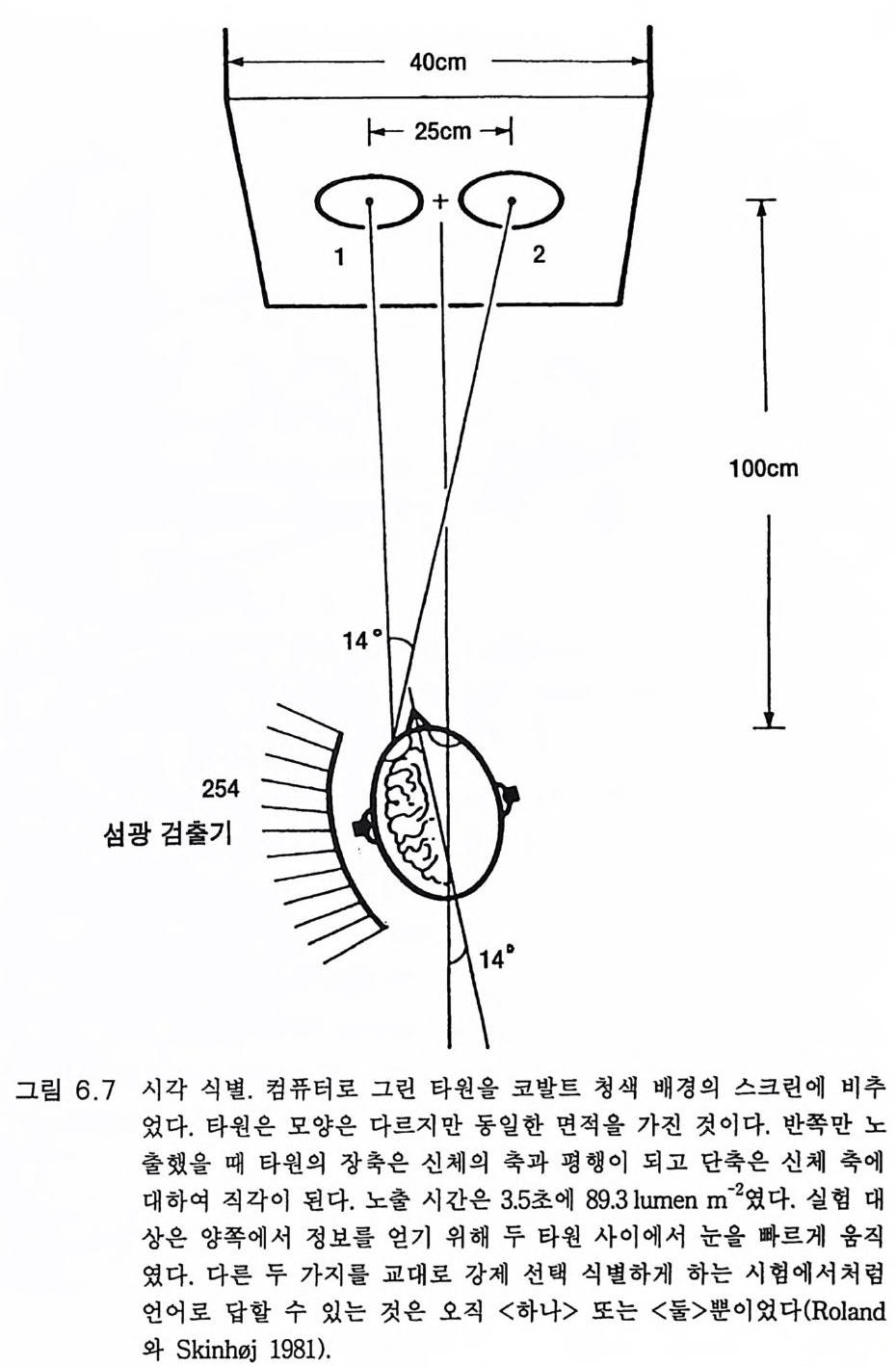 40cm
40cm
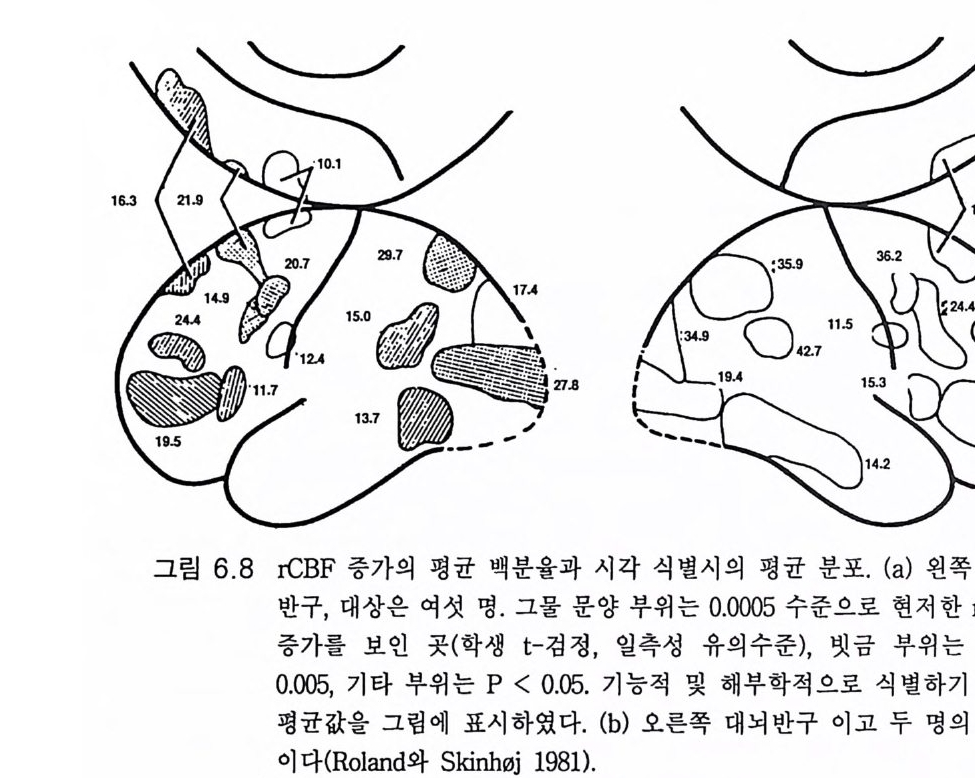 ”.8
”.8
증가를 나타낸 부위는 양쪽 대뇌반구에 광범하게 분포하였다. 그림 6.8 에 %로 표시하였다. 그림 6.6 에 나타낸 마카크의 실험 결과에서 예상 되듯이 양측 후두엽 그리고 인접한 두정엽 및 측두엽 영역에서 rCBF 의 유의한 증가가 있었다. 불행히도 후두극은 내측 경동맥에 주사한 물질이 미치는 범위 밖이었다. 또 대뇌반구의 내측 표면의 rCBF 도 측 정되지 않았다. 대뇌반구 후방에서 증가되었던 대부분의 구역은 마카크에서 발견되 었던 시각영역에 해당하는 것이다(그림 6.4 , 6.5, 6.6 ) . 하측두 영역에 있
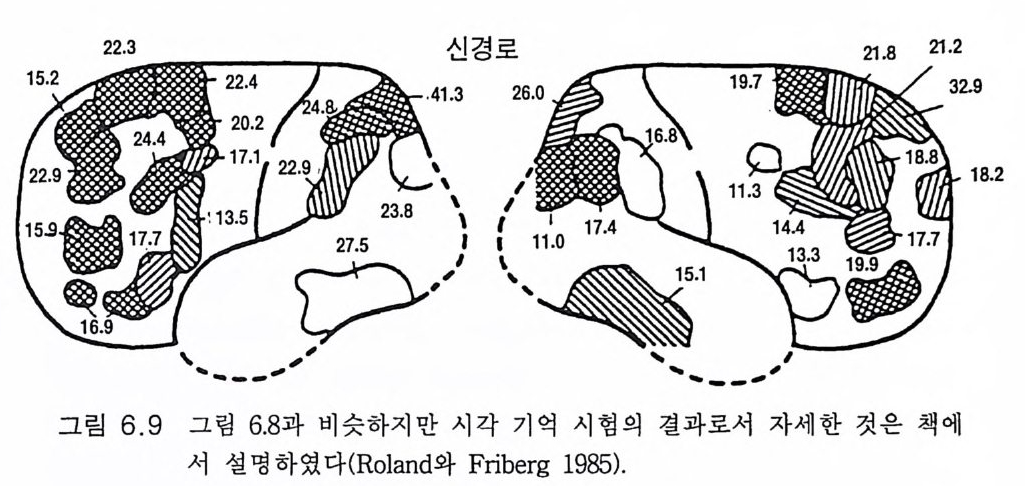 22.3 신경로
22.3 신경로
어서는 상당한 rCBF 의 증가가 있었다(그림 4.4의 브로드만 영역 20, 21). 특히 오른쪽의 증가가 많았다 (14%). 양쪽 대뇌반구에서 그림 6 .4의 V2, V3, V4 에 해당하는 영역에서 증가하였댜 그림 6.6 의 MT 영역에 해당 하는 것으로 생각되는 부위는 좌측에서 15% 의 증가를 보였고 우측에 서는 42% 의 증가를 보였다. 롤랜드(R oland 1985) 는 후상두정부에서 36% 및 30% 의 증가가 있음에 놀랐다. 그러나 이것은 마운트캐슬과 그 동료들 (Moun t cas tl e 둥 1975, 1984) 의 발견으로 예상할 수 있는 것으로 생각한다. 그리고 형체의 판단과 시각 관계는 그림 6.6 의 PG 영역인 브로드만 영역 7 의 신경 활동이 관련되는 것으로 생각한다. 양측의 전두전엽에 높은 rCBF 가 광범하게 분포함은 놀라운 소견이 댜 전두엽 시야는 예상대로 좌측에서 21%, 우측에서 36% 증가하였다. 그러나 전두전엽의 다른 여러 부위에서도 상당한 rCBF 의 중가가 있었 다. 그 부위는 주의와 집중에 관련되는 부위로 생각되며 청각 및 체성 감각의 판별 및 판단에 관련되 는 것 이 다(R oland 와 Sk inh ~j 1981). 그림 6.9 와 같은 양상의 rCBF 를 나타낸 실험 대상에게 또 다른 과업
을 수행하게 하였다(R oland 와 Fri be rg 1985). 실험 대상은 눈을 감고 귀 룰 막은 안정상태에서 잘 알려진 길을 따라 움직이는 연속적 광경을 상상하는 과업이 주어졌다. 불행히도 후두엽 대부분에서 rCBF 를 측정 할 수 없었다. 그러나 양측 하측두엽은 활성화되었다 . 이는 이 부위의 형체 탐지성 때문에 예상되는 것이다. 또한 MT 및 PG 영역은 양측이 모두 홍분되었다 . 이는 공간과 움직임에 관계되는 시각 과업 때문에 예상되는 것이다. 전두전엽에 있어서는 광범한 rCBF 의 증가 를 보였다 . 이는 시각 기억 과업과 관계되는 것으로 생각된다. 그러나 이 모든 활 성이 일차적으로 과업에 의하여 유발되는 것으로 생각 할 필요는 없다 . 이돌 여러 전두전엽 영역은 서로 잘 얽혀 연결되어 있으므로 한 부위 가 일차적으로 활성화되면 여타 부위의 이차적 및 삼차적 활 성을 초래 하게 될 것이다 . 전두전엽의 광범한 홍분은 일부 시각 기억 과업에 관 련된다는 사실은 알아둘 필요가 있다. 불행히도 rCBF 형의 실험을 원숭이에서는 할 수가 없다. 한 가지 생 각에 집중해야 하는 시간이 너무 길다 (40 초) . 그러나 원숭이의 전두전 엽 피질이 기억과 관계된다는 사실은 오래전에 알려졌다. 예를 들면 지연성 반응으로 동물은 기다리는 기간에 먹이통을 치우더라도 먹이 가 들어있는 용기의 시각기억을 일정시간 동안 보유한다 (Fus t er 1980). 관계되는 반응은 우발적 음성 변이 con ti n g en t neg a ti ve va riat i on : CNV 로서 보상에 연계되는 후속 자극을 위한 신호를 제공하는 자극에 의하 여 전두전엽에서 발생하는 것이다 . 이 두 가지 자극은 시각적인 것일 수도 있어서 시각 자극에 대한 반응으로 원숭이의 전두전엽 피질이 광 범하게 활성화된다는 증거이다. 단일 단위 기록으로 이들 전두전엽 반 옹의 위치와 강도를 알아볼 수 있다 (Fus te r 1980). CNV 는 애초에 사람 의 두피 표면 기록으로 관찰된 것이다. CNV 의 전두전엽 활성은 그림 6.8, 6.9 에서 보이는 광범한 rCBF 의 증가와 상관관계가 있다. 따라서 원숭이의 전두전엽 활성은 양적 차이는 있을지라도 질적으로는 호모
사피 엔스 사피 엔스와 유사하다고 할 수 있다 . 원숭이의 전두전엽 손상은 (Fus t er 1980) 학습과 행동 과업의 시행 결 함을 초래한다 . 일반적으로 시각과 시각운동 반응에 관계되는 것에 결 함이 있다. 이는 행위의 선택을 요구하는 과업에서 특히 분명하다. 지 연성 반응이나 지연성 맞추기시험 같은 것에서 뚜렷하다. 동물은 계속 적으로 주의가 산만하여져서 과업에 집중할 수가 없다. 전반적인 정서 적 반옹의 감소도 있다 . 이 증상은 전두전엽을 절제한 환자에서 특징 적인 것으로 의욕 상실이 있고 식물인간처럼 되어버린다. 6.7 시각 진화에 대한 결론 다양한 종류의 시각 활성에 관하여 일반적인 결론을 내린다면 후두, 측두 , 두정 및 전두엽에 있어 유인원, 마카크, 올빼미 원숭이, 사람의 반웅 사이에는 현저한 상동성이 있다고 할 수 있다. 그렇지만 사람 대 뇌피질의 시각 과업에 의하여 활성화되는 영역은 원숭이 대뇌피질보 다 훨씬 크다는 것을 기억해야 한다. 호모 사피엔스의 진화 과정 초기 에 망막과 LGB 에서 일차 시각피질에 이르는 시각 경로는 이미 완전한 발달이 이루어졌다고 생각된다(그림 6.1 ). 마카크의 일차 시각피질 (Vl) 은 전체 대뇌피질에 대하여 사람의 Vl 보다 훨씬 큰 비율 (4 배)을 차지 한댜 유인원에 대한 상세한 비교연구는 없다. 그렇다고 해서 대뇌피 질의 시각 계통이 원숭이와 다를 것이라고 생각할 이유는 없다. 입수할 수 있는 증거들(그림 6.8, 6.9) 에 의하면 사람의 선조전 영역 은 마카크의 것(그림 6.4 , 6.5 , 6.6) 에 해당하는 것으로 보인다 . 그러나 사 람에서는 그 영역이 훨씬 크고 전체 대뇌피질에 대한 비율도 크다. 토 비 아스 Tob i as 와 홀로웨 이 Hollowa y의 관찰 (2.2 및 2 . 3) 을 상기 해 보는 것 도 홍미 있는 일이다. 호모 하빌리스 혹은 심지어 오스트랄로피테신에
게 상두정엽이 대략 일치한다고 하였다. 그것은 마운트캐슬이 관찰한 피질영역(영역 7 또는 PG) 이었을 것으로 생각되고 이는 시각 공간 및 움직임에 관계되는 것이다. 이 같은 발달은 양각 보행에 있어 특히 진 화적 가치가 있는 것이다. 따라서 호미니드 진화 초기에 대단히 효율적인 시각 계통을 물려받 았다. 사람과 비교할 때 전체 시각피질 영역은 유인원으로부터 세 배 이상의 면적 증가가 있었으나 기능이나 설계에 있어서는 근본적인 진 화적 발달은 없었다고 생각한다. 또한 제 3 장에서 말했듯이 중추신경계 에서 근수축까지의 운동 계통도 호미니드 진화 초기에 기본적으로 발 생되었다. 다만 화석 기록에 의하면 엄지가 교묘하게 발달되어 호모 에렉투스 시절에 정확하게 잡을 수 있게 되었다 (3.5 의 그림 3.18). 그러 나 정확한 파지는 솔루트레아 Solu tr ea 의 정교한 도구를 만드는 데에나 필요했던 것 이 다 .(6 . 8) 만일 호미니드 진화 초기에 시각과 운동 계통이 인간 수준까지 완 전하게 발달하였다면 어째서 호미니드에게는 일찍부터 석기문화에 기 술적 발달이 없었는가라는 의문이 생긴다. 해답은 전두전엽의 결핍에 있다고 할 수 있다. 이는 개념적 과업(그림 6.8 , 6.9 , 7.1 4 ) 그리고 맡겨진 일에 대한 집중력과 밀접한 관련이 있는 것이다. 원숭이와 사람에서의 손상에 의한 연구에 의하여 전두전엽의 결핍이 동기 부여의 상실이라 는 비극적 결과를 보였다. 따라서 도구 문화는 호모 하빌리스로부터 호모 사피엔스 사피엔스에 이르는 100 만년 이상의 진화를 통하여 대단 히 서서히 발달했음을 알 수 있다(그림 6.1 0 ). 호미니드는 전두전엽이 충분히 발달하지 못하였으므로 동기와 집 중력의 결함이 있었을 것이다. 호모 하빌리스 그리고 올두바이 문화 에서 나타났듯이 그들은 수백 또는 수천년 동안 기존의 도구 기술 수 준에 만족하였을 것이다.(그림 6.1 0 a) 아마도 진화 정체기로 생각된다. 그러고 나서 호모 에렉투스로의 유전적 발달이 있어서 아슐 Ache uli a
 I
I
문화(그림 6 .1 0b) 로 진입하였을 것이다 . 도구만들기가 개선되고 새로운 도구룰 발명하였다. 이 같은 개선은 젊은이를 훈련시켜 명맥이 이어졌 고 장기간 지속될 수 있는 잠재력을 가지고 새로운 석기문화가 탄생하 였는데, 각각의 석기문화에 따라 그 석기시대가 명명되었다 . 죽 호모 하빌 리 스는 올도완 문화 Oldovan cul tur e 로, 호모 에 렉 투스는 아슐 문화 로, 호모 네안데르탈리스는 무스테리아 문화 Mous t e ri a 로 부른다(그림 6.1 0 c). 각 단계의 석기기술은 매우 미미한 정도의 진보를 보이면서 수 백 또는 수천년 동안 계속되었다. 이런 낮은 수준의 진보는 아마도 변 연계 특히 내측 편도체와의 관련하여 전두전엽이 미미한 수준으로 발 전하고 작동하게 된 것에 따른 것일 수도 있다(제 5 장). 이제 석기문화 를 간략하게 살펴보기로 한다. 6.8 석기문화의 진화 호미니드 진화에 있어 그들이 이룩한 문화적 업적의 대부분은 단지 내구성인 석기문화의 증거뿐이다. 앞서도 언급했듯이 오스트랄로피테 신은 찰 발달한 목기문화를 가졌음에 틀림없다. 그러나 이것은 화석으 로 혼적을 남기지 않았다. 그리고 남아프리카의 오스트랄로피테신은 뼈, 뿔 그리고 이가 있는 턱뼈를 사용했다는 중거도 있다. 첫 석기는 올도완 형에 속한다(그림 6.1 0 a). 이는 호모 하빌리스와 연 관된다. 이 같은 석기들은 작은 돌을 날이 서도록 쳐내어 만들어 잘게 썰기, 잘라내기, 긁기 둥에 사용했다. 훨씬 뒤에 호모 에렉투스는 생산 기술이 훨씬 향상되어 아슐 형(그 림 6.1 0 b) 석기를 만들었다. 손도끼, 깎는 도구, 뾰족한 도구 둥 보다 만 족할 만한 도구로 사용할 수 있는 것을 만들었다. 이것으로 동물을 도 살하거나 육류를 먹기 쉽게 만드는 데 효과적으로 사용했을 것이다.
아슐 형은 호모 에렉투스와 호모 프레네안데르타리스의 확산과 더불 어 널리 분포한다. 가장 잘 알려진 여러 종류의 석기는 무스테리아 형이다. 엷은 조각 내기 기술로 만들어진 것으로 호모 네안데르타리스에서 발달하였다 (그림 6.1 0 c). 잘 고른 돌을 돌망치로 주의하여 때려서 편편한 돌조각으 로 만들어 여러 가지 유용한 도구를 만들었다. 잘 만들어진 도끼 외에 도 긁어내기, 창끝 및 특이한 날이 있는 돌편이 있다. 이들 도구는 동 물을 도살하는 데뿐 아니라 가죽을 다듬는다거나 천을 꿰매는 데 효과 적이었고 빙하기라는 사나운 기후에 대단히 필요한 것이었다. 호모 사피엔스 사피엔스의 가장 정교한 도구는 40,000 y BP 부터이다. 크로마뇽 Croma g non 및 솔루트레아 Solu tr ea 시대의 칼날문화라고 하는 대단히 정교한 돌조각과 돌편이 있다. 그중 가장 정교한 것은 월계수 잎 모양 칼날이다. 그러나 이것은 실제로 사용하기 위한 것이라기보다 는 아름다움을 뽐내기 위한 것이었을 것이다 . 드 럼 레 이 (De Lumley 1969) 는 약 40 만 내 지 30 만 y BP 에 스페 인 과 프 랑스 남부에 있었던 주거지역 구조를 기술하였다. 기초석들이 둥글게 배열되어 있는 것을 발견할 수 있었는데 l0m 와 5m 사방의 공간이 되 는 상부 목조 구조물이 있었을 것으로 생각된다. 이보다 훨씬 전 2 m y BP 에 아프리카의 올두바이 지방에 있었던 주거지역의 초석 증거도 있다 (S t ebb i ns 1982). 6.9 골패의 판화 석기 도구문화의 진화로 설계와 제작기술이 복잡하게 되었다. 채집수렵 사회에서의 실제적인 필요성에 의했을 것으로 생각할 수 있다. 재료의 이 점을 이용하여 점차로 설계와 제작기술을 소중히 개선해 나갔을 것이다.
그렇게 해서 호모 사피엔스 사피엔스의 구석기시대(약 30,000y B P) 초기에 인식체계라는 놀라운 발견을 이룬다. 즉 관찰, 추상 그리고 기 록이 발달하게 되었다. 그림 6.lla 는 프랑스의 볼랑카드에서 발견된 골 패에 눌러 그린 판화이다. 복합적 부호가 새겨져 있다 (Marshark 1985). 그림 6.llb 에 상세하게 보여주듯이 2 개월 보름 동안의 달의 순차적 인 모양이 그려진 것이다 . 이 그림은 날짜별로 달의 변화한 모양을 그린 것으로 이 같은 달의 작은 변화를 새겨넣기 위해서는 대단히 정교한 도구를 사용했을 것이다. 이 그림은 두 달 주기 이상을 추적한 것이다. 이것 이후 만년 동안에 이것에 비교될 만한 것이 발견된 일이 없다 . 이것은 초기 수렵_채집시대의 선구자적인 작품으로 간주될 수 있는 것으로서 천문 관찰자는 달 주기의 규칙성을 도식적으로 작성하였다. 이 같은 기록은 수렵―농경인들에게는 실제적인 의미가 있는 것으로 동물의 행동과 계절의 변화를 주의 깊게 따라갈 수 있게 하였을 것이 댜 또 집단이 정기적인 모임이나 의식의 날잡기에도 이용되었을 것이 댜 이 같은 초기의 인간들이 천체의 질서를 이해하고 그들 사회의 시 간적 질서룰 수립하였음을 알 수 있다. 블랑카드의 판화는 비산술적인 관찰적 달 부호이다 . 휘어지는 것은 달 주기의 이행기를 나타낸다. 좌측에 만월상이 있고 우측에 초생달과 보이지 않는 상이 있고 중간에 반월이 있다. 이 기록은 관찰 당시에 만들어진 것이지 나중에 기억을 더듬어 기록된 것이 아니다. 마르삭 (Marshack 1985) 은 다음과 같이 말하였다 이 부호는 문자언어라고 볼 수 없다. 왜냐하면 각 단위와 조가 언어가 아니기 때문이다. 그리고 또한 수학적 기술도 아니다. 왜냐하면 각 조와 조의 조합은 정량적이기는 하지만 산정되거나 합산된 것이 아니기 때문 이댜 . 그러나 부호의 구성과 배열을 통한 문제 해결 과정은 이후의 문자 적 기록 수학적 기록 유지의 발달 과정에서 볼 수 있는 것과 동일한 차원
 (a)
(a)
이고 그 발단이라고 하겠다. 완전한 불랑카드 판화를 가지고 그들은 달이 차고 기우는 형상을 알게 되었을 뿐 아니라 시간의 주기성과 계속성에 관한 개념을 창출할 수 있 었다 그 당시 문화에 속한 누구든지 그 형상을 보고 그 전통에 익숙한 자는 꾸불꾸불한 형태로부터 시간의 계속성과 주기성을 볼 수 있었을 것 이다. 마르삭(1 985) 은 또 훨씬 후인 약 11,000 y BP 에 만들어진 것이기는 하 지만 그로테 두 타이 Gro tt e du T ai에서 발견된 비슷한 부호 판화를 보 고하였다. 이것도 꾸불꾸불하게 배열되었으나 좀더 복잡하고 불랑카드 판화보다 발달된 부호로서 3 년 반에 걸친 해와 달의 관찰이었다. 마르 삭은 이들 초기 부호에서 발견되는 의향적, 신경학적, 시각지향적 서 열 및 개관 능력 과정은 후에 수학적, 천문학적 및 지리학적 부호체계 에 이용되었을 것이라는 논리를 정립하였다. 6.10 시각-구성 대뇌영역 대뇌피질의 시각-구성vi suo-cons tru c ti ve 영역은 언어영역(그림 4 . 3) 처 럼 분명하게 구분되어 있지는 못하다. 이것은 처음에는 손상에 의한 연 구로 구획도를 만들고 뒤에 전기자극 실험으로 펜필드와 로버츠 (1959) 가 기술하였다. 그러나 운동의 장애(실행중)를 일으키는 손상에 의한 연구 (9.2) 그리고 최근의 교련절개 환자 (9.6) 의 연구를 통해 시각-구성 영역은 주로 우측 대뇌반구, 특히 하두정부(그림 9 .1)에 위치하는 것이 발견되었다. 주로 브로드만 영역 39 와 40 에 위치할 것으로 생각되며 해 부학적으로는 좌측 대뇌반구의 베르니케 언어영역 부분의 거울상 mirror im a g e 이다. 이들 합성 시각-구성 영역은 분석적 언어영역 반
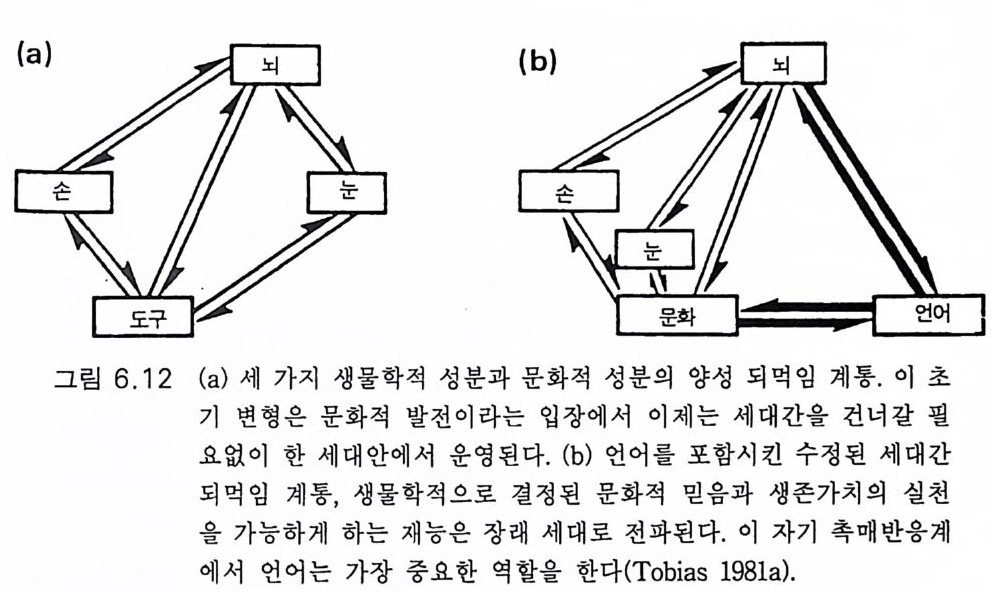 그림 6. 1 2 (a) 세 가지 생물학적 성분과 문화적 성분의 양성 되먹임 계통. 이 초
그림 6. 1 2 (a) 세 가지 생물학적 성분과 문화적 성분의 양성 되먹임 계통. 이 초
대측에서 진화의 보조를 같이 하여 발달한 것으로 생각된다. 이는 대 뇌신피질의 기능 수행에 있어서 비대칭성이 어떻게 반복을 피하고 대 뇌의 기능이 증강하게 되는가를 보여주는 좋은 보기가 된다 (9.7). 토비아스 (1981a) 는 시각-구성의 생리학적 회로를 그렸다. 그림 6.12a 는 도구의 사용과 구성에 관련되는 상호 활동을 나타낸 것이다. 운동 계에 관하여는 제 4 장에서 검토하였고 시각계통은 이 장에서 다루었다. 뇌와 도구 사이의 상호연관적 화살표는 도구의 설계와 제작에 관계되 는 사고과정 을 상징 적 으로 나타낸 것 이 다. 그림 6 .1 2b 는 좀더 복잡한 그림으로 뇌, 문화 그리고 언어를 강조한 것이다. 이는 언어를 포함하 여 고도로 발달한 문화 사회의 복합적 개인 관계를 포괄하므로 단순한 도구문화와는 거리가 상당히 멀다.
6.11 성형적 기술의 창조성 스테빈스 (S t ebb i ns 1982: 363) 는 < 현대 인간사회는 정성적 및 정량적 으로 현존하는 어떤 동물사회와도 다르다 > 라고 말하였다 . 그는 사람 의 특이적인 세 가지 특성은 기공성arti sans hip, 의식적 시간-결합 그리 고 상상력이라고 하였다. 기공성은 경탄할 만한 사람의 시각 _ 구성 수 행 능력의 기초가 된다. 의식적 시간 구 결합성은 호미니드 진화과정에서 달의 판화로서 처음 알려졌다(그림 6.1 1 ). 상상력은 도구문화에서 찾아 볼 수 있댜 예를 들어 조각가는 월계수잎 칼날의 형상을 그것이 조각 되어 나올 돌 재료에 미리 그려 볼 수 있었을 것이다. 우측 두정엽 외 에도 사람에게 주어진 모든 창조성은 변연계와 관계되는 전두전엽에 도 의존하는 것으로 알려졌다. 따라서 그림 6.12b 에 그려진 뇌는 대단 히 복합적 방식으로 운영되는 것이다 . 가장 복잡한 형의 석기는 발달된 호미니드의 시각 - 구성 계통을 대 단히 인상적으로 나타내는 것이다. 이 도구문화는 성형적 기술 창조성 의 시작으로 간주할 수 있는 것이다. 성형적 기술에 있어 사람이 크게 성취한 점들 중 하나는 호미노이 드의 대뇌 영역에서 호미니드 진화 이전에 이미 발달되었던 것을 보다 효과적으로 이용하였다는 것이다. 이 같은 진화적 과정을 예상 진화 anti cipa to ry evolu ti on 라고 할 수 있다 . 다른 몇 가지 예를 뒤에 다시 들 것이댜 전두전엽피질의 진화적 발달과 아마도 측두 및 두정엽 영역들 의 발달은 이미 발생되어 있었던 호미노이드의 시각_운동 계통을 좀 더 효과적으로 이용하게 하였을 것으로 생각한다. 이것이 괴리진화 disp a rate evolu ti on 라고 말하는 것의 예이다 시각-운동 계통의 · 일부는 완전하게 발생되었다. 그러나 그 잠재능력은 이후의 두정 및 측두엽 영역이 진화되기까지는 충분히 발휘될 수 없었다. 제 3 장 그리고 이 장에서 설명한 시각-운동계통은 폰지드의 능력을
능가하는 훌 륭한 계통이라고 생각할 수 있다. 그러나 이것은 도구문화 에서 나타났듯이 대단히 서서히 발생한 호미니드의 발달 과정의 정신 적 요구에 대한 생물학적 도구로서 적절한 것이었다. 이는 뒤에 호모 사피엔스 사피엔스의 창조적 활동으로 꽃피우게 된다. 막달레니아 시 대의 동굴벽화 그리고 중석기 시대를 거쳐 신석기 시대를 통하여 수메 르에 이르러 처음으로 크게 문명을 이룬다. 이는 호크스(H awkes 1'51 6 ) 가 잘 기술하였다. 넓은 의미에서의 성형적 기술인 건축, 조각, 조판, 도예, 그림, 유리, 가구, 벽걸이, 융단 등은 모든 인간 문화의 평가와 인식의 기초인 것이 댜 이는 쉽게 창조적 상상력을 들여다보게 하는 것이다. 일부 성형적 기술은 특히 조각과 그림은 묘사적 주제를 가지나 상징성이 훨씬 중요 하댜 라스코 Lascaux 의 동굴 벽화까지도 동물들은 묘사적으로 그려졌으 나 사람은 아이들의 그림처럼 막대기 모양으로 상징적으로 그려졌다. 성형적 기술을 창조하고 평가함에 있어서의 감각적 경험의 역할을 논할 때 바움가르트너 (Ba umg artne r 1986) 는 가소성 또는 학습에 서 뇌 기능의 절대적 역할을 강조하였다. 이는 제 7 장과 제 10 장에서 살피게 될 것인데 문화적 진화는 우리의 인간성 발생의 기초임을 알게 될 것 이다 그래서 진화의 절정에는 학습이 문화 생활을 점차 풍요하게 하 는 열쇠이다. 미학은 지적 및 정서적 가치를 갖는다. 전자는 특히 전두 전엽피질에 의하고(제 9 장) 후자는 변연계에 의해 좌우된다(제 5 장). 이 들 두 계통은 서로 연계되어 있다. 바움가르트너는 다음과 같이 결론 지었다. 시각 계통의 기능적 조직은 신경 활성의 동적 양상을 통해 주위 환경 의 표상을 창출한다. 물질적 외계와 우리가 상호작용하는 데 적절하도록 현실 세계를 우리를 위해 창출해 주는 것이다. 우리가 아름답다거나 즐겁 다고 느끼는 것은 계통의 처리과정 법칙에 가장 적절하게 맞추어진 시각
입력일 것이다 . 이들 법칙은 이미 결정되어 있는 것이다. 그러나 시각 경 험의 축적과 인습의 테두리 안에서 학습을 통하여 어떤 시각 입력을 선 호하게 되는지는 변화될 수 있다(1 986: 813) 이와 같은 주제에 대하여 포퍼 Po pp er 는 다음과 같이 말하였다. 화가가 하는 일은 때로는 놀랄 만큼 비슷하다 . 그의 캔버스 위에 물감 으로 점을 찍고 이를 그대로 받아들일 것인지 아니면 파기할 것인지를 판단하기 위해 뒤로 물러선다. 그러고 나서 점 찍기를 계속한다. 효과를 그려진 대상과 비교하든 또는 내적 구상과 비교하든 아니면 단순히 효과 를 인정하거나 부정하든 여기에서 중요한 것은 아니다 . 중요한 것은 에른 스트 곰브리치 (Erns t Gombri ch 1960) 가 <창조가 선택을 선행한다 Mak ing Comes befo r e ma t c hi n g.>라고 적절하게 표현한 것이댜 이 어구는 선택의 모든 경우에 유익하게 적용될 수 있고 특히 가설의 정 립과 중명 과정에도 적용될 수 있는데 이런 과정들에는 지각 과정 특히 총체지각 Gesta lt pe rce pti on 이 포함된다. 물론 <창조가 선택을 선행한다>는 어구 는 다윈의 도태설에도 적용될 수 있다. 여러 유전적 변종이 만들어지는 것은 환경에 의해 도태되고 그러고 나서 환경에 맞추어지는 것에 선행한 댜 환경의 작용은 우회적이다. 왜냐하면 도태 또는 선택이 일어날 수 있 는 대상을 생성 또는 창조하는 과정이 이 환경의 작용에 선행하기 때문 이 다 (1987: 146). 진화에 관하여 또는 문화적 진화에 관하여 이 책에서 창조적 기술 의 거대한 역사적 단계를 간결하게 취급하는 것은 적절치 못하다. 우 리가 위대한 예술이라고 하는 것의 문턱에 왔다고 하는 것으로 충분하 댜 그 너머에는 여러 종족의 창조적 예술을 학술적으로 기술해야 하 는 무한한 분야가 있다. 예를 들어 토인비는 인류의 위대한 문화적 신
기원을 여덟 가지로 잡았다 . 수메르, 이집트, 중국, 인도, 이슬람, 그리 스, 유럽, 아메리카 인디안이다 .
제 7 장 학습과 기억의 전화 학습이란 기본적으로 뇌에 저장되는 과정이다. 그리고 기억이란 뇌 의 자료 은행에 저장되어 있는 것을 상기하는 것이다. 많은 경우 혼동 하여 취급되기는 하나, 두 가지 분명한 형태의 학습과 기억이 있다. 첫 째는 운동 학습과 기억이다. 제 3 장에서도 언급한 선다거나 걷기까지를 포함하는 능숙한 움직임의 학습이다. 둘째는 인지적 학습 또는 기억이 라고 하는 것이다 . 여기에는 모든 지각, 관념, 언어적 표현이 포함되고 실제로는 문화를 통틀어 그 안에서 나타나는 모든 일이 포함된다. 그 림 4.2 에서 제 3 세계로 나타낸 것이다 . 인간 기억의 모든 형태가 관련되 는 대뇌 기전을 설명하고 추론하기에 앞서 진화적 기원을 살펴보는 것 이 중요하다. 7.1 호미노이드 조상의 모델로서의 유인원 인간의 종으로서의 경탄할 만한 성취는 뇌의 진화적 발달에 연유한다. 이는 여러 일들 중에서도 도구 사용, 도구 제작, 그리고 합리적 이론, 사
려 깊은 협동 및 언어에 의한 문제 해결 능력을 갖게 하였다. 가장 두드 러진 것의 하나는 뇌의 구조에 있는데 침팬지는 생물학적으로 사람과 비 슷하댜 침팬지는 원시적 추리력으로 현재 살고 있는 어떤 포유동물보다 도 사람에 가까운 지능을 나타낸다 . 현대 침팬지의 뇌는 수백만년 전의 첫번째 유인원인의 행동을 산출해낸 뇌와 크게 다르지 않다고 생각된 다 (Goodal 1971: 243) 구달 (Goodall 1971) 은 그녀의 훌륭한 저서 『인간의 그림자에 관하여 In The Shadow of Man 』에서 탄자니아의 곰베 유역 보호구에 자연 상 태로 자유롭게 살고 있는 50 마리의 침팬지에 관한 독특한 연구를 기술 하였댜 연구는 수년 동안 계속된 것으로 이 무리의 각 개체에 이름을 주고 이들을 식별할 수 있었다 . 이들은 순수한 여족장 사회로서 새끼 는 태어나서 성숙할 때까지 어미의 계속적인 보호로 자란다. 이 자웅 동서 사회에 부성은 알려지지 않았으며 그리고 어떤 경우에도 침팬지 는 교집으로 임신되고 새끼를 출산한다는 사실을 몰랐다 . 호모에서와 마찬가지로 침팬지 새끼는 출산 후 대단히 무기력하다. 그러나 침팬지 새끼의 태어날 때의 뇌는 성숙한 것의 60% 이나 사람은 26% 밖에 안 된다(표 5.2). 침팬지 어미는 새끼를 잡고 나무에 오른다거 나 둥지를 만드는 둥 어려운 과업을 떠맡는다. 조만간 새끼는 어미의 머리털을 잡고 매달리는 것을 배운다. 5 개월이 되어서야 어미 등에 매 달리는 것을 배우게 된다. 한편 2 개월쯤에는 사물을 보고 접근하는 것 을 배운댜 이는 사람의 어린아이보다 조금 빠른 것이다 . 3 개월째에는 조화로운 운동을 배우게 되고 5 개월째에 첫발을 딛게 되어 나무에 기 어오른다. 이는 사람의 어린아이보다 훨씬 빠른 것이다. 초기 학습 과 정은 밤에 쉴 수 있는 둥지를 나무 위에 만드는 것이다. 8 개월째가 되 면 침팬지 새끼는 이 중요한 기술을 습득하려고 노력한다. 그러나 수 년 동안 밤을 보낼 수 있는 둥지를 만들지는 못한다. 이것은 어린아이
들의 놀이 갇은 것이다 . 어미는 이 초기 학습의 스승이다. 그러나 새끼 가 주위를 돌아다닐 때가 되면 모방하는 것으로 배운다 . 8 개월째에는 암내 난 암컷에 교집하는 것까지도 시도한다. 3 년이 지나면 어미는 처 음으로 도구 사용법을 가르쳐 흰개미 둥지를 막대기로 쑤셔 흰개미 유 충을 꺼내 먹는다. 그러나 5 살에 젖을 뗀 후에도 1-2 년 동안 어미의 보 살핌을 받는다. 한편 다른 젊은 것 또는 성숙한 것에게서도 행동을 배 운다. 때로는 적절치 못한 행위에 대하여 늙은 수컷에게 공격을 받는 것 으로도 배운다 . 그러나 어미는 항상 조력할 태세에 있다. 제인 구달의 이야기에서 침팬지 어린것의 원시적 행위는 오늘날의 원시적 사회에도 있듯이 사람의 유년기와 크게 다를 바가 없다. 큰 차 이라면 인간의 원시적 사회에서는 남성 특히 아버지가 젊은 남자의 훈 련에 참여한다는 것이다. 침팬지의 경우 7-8 세가 될 때까지 사춘기가 시작되지 않는다. 이때쯤 어린 수컷은 어미를 떠나 수컷 사회에 끼어 들기를 시도한다. 그 사회는 어린 수컷을 환영하지는 않으나 단지 내 버려둘 뿐이다. 그들은 순종하는 태도를 보여야 한다. 심한 공격성을 보이면 어미가 상관하게 된다 . 극단적인 감정을 가진 복합사회이다. 그러나 심하게 다치는 일은 드물다 . 결국 수컷은 15 세가 되면 완전하 게 성숙하고 우세를 다투게 된다. 한편 암컷은 8-9 세에 성적으로 성숙 하고 대단히 활발한 성생활을 가져서 11-12 세에 첫 새끼를 낳는다. 침팬지의 청년기는 사람처럼 길지는 않지만 그래도 의외로 긴데 이 는 그 복잡한 사회구조를 배우는 긴 학습 과정 때문이다. 개체간의 상 호작용에는 대단히 다양한 몸짓들 그리고 적어도 14 가지로 식별되는 울음과 소리치기가 포함된다. 공동체 내에서 각 개체는 음성만으로도 서로 구별되는 것 같다 (Goodal 1971). 그러나 소리치기와 울음은 두 하 위수준 언어(그림 4.1 ) 이상의 의사 소통은 이루지 못하고 감정 상태를 표현한다거나 음식, 침입자, 다른 동물 등에 대한 정보로서의 신호 정 도이댜 이는 모든 사소한 사건과 행동에 관한 지나치게 자질구레한
대화를 즐기는 오늘날의 가장 원시적인 호모 사피엔스와 비교하면 대 단히 빈약한 의사 소통이다. 쿰메르와 구달 (Kummer 와 Goodall 1985) 의 논문에서 쿰메르는 호미 니드는 호미노이드에서 진화하였다고 추론하였다 . 현대의 침팬지처럼 본능적 행동이 결여되어 있기 때문이라고 하였다. 정신적 융통성과 학 습은 혁신에 대한 보상으로서 무엇보다도 중요하다. 자연도태에 의한 진화에서 좋은 기회가 될 수 있다. 침팬지에 대한 이 연구에서 그들의 자연 습성에 실험적 시험과정을 가미할 필요는 없다. 멘젤 (Menzel 1984) 의 어린 침팬지에 관한 연구는 구달의 연구를 보충한 것이다. 동물들은 식물로 뒤덮인 상당히 넓은 폐쇄 지역에 풀어 두었다. 일부 시험에서는 어린 침팬지를 자연 상태 에서 어미의 등에 업혀 돌아다니는 것을 본 따 상자에 넣어 실험자가 옮겼댜 이전의 방문 이후 위치가 바뀐 물체들을 곧 식별하는 것으로 보아 사물이나 공간에 대한 기억이 대단히 좋다는 것이 밝혀졌다 . 그 동물들은 매일 매일 그 우리 속에 있는 웬만한 사물들 모두의 시각 적 형태와 선택적 위치를 학습하고 기억하는 것으로 생각되는데 각 사물 들이 음식물 둥의 분명한 보상과 연계되어 있는지 아닌지와는 무관하였 다 (Menzel 1984: 513). 학습의 효과는 동물을 18 조각의 음식이 숨겨진 장소로 옮겨 보여줌 으로써 실험하였다 . 정보를 받은 동물은 우리에 갇힌 다른 동물이 있 는 곳으로 되돌려 보냈다. 그리고 잠시 후에 모든 동물을 풀어 놓아 개방된 지역을 마음대로 돌아다니게 하였다. 정보를 받았던 동물은 원 래의 길에서 벗어나 잘 선택된 길을 따라 숨겨진 음식 거의 전부를 효 과적으로 찾아내었다. 이는 단지 한 번의 시행으로 공간적 학습이 효 과적으로 이루어졌음을 뜻한다. 예상대로 정보를 받지 않은 동물은 정
보를 받은 동물과 함께 찾을 때만 성공하였다. 이 실험은 정보를 받은 동물은 지도 위의 숨겨진 음식의 위치로서 일종의 영역지도를 만들었 음을 뜻한다. 사람의 행위와의 관계에서 이들의 상황을 알아보려면 침 팬지의 공간 기억에 관한 연구가 더 필요하다. 침팬지 학습에 관한 이 분야의 실험은 학습의 보다 정밀한 실험성 시험 특히 일종의 부호 계 통에 의한 언어적 의사소통의 학습과의 관계로 이어진다. 그 같은 연구의 시작은 아기 침팬지가 수용된 생활에 적응하는 것 을 배워나가는 과정에 대한 연구였다. 켈로그와 켈로그(1 933) 는 어린 암컷 침팬지 구아 Gua 를 양육하였을 때 주의 깊게 생각해 낸 과정을 상세하게 기술하였다. 그들은 가족과 함께 정상적인 아이들처럼 구아 룰 키우려고 시도하였다. 훈련은 우발적인 것이었다. 구아는 사람의 아이들처럼 배웠다. 대체로 놀랄 만큼 성공적이었다. 가드너와 가드너 (1 983) 도 주의 깊게 그 방법을 따랐다. 태어나서 약 10 개월째인 1965 년 에 아프리카에서 잡혀 실험실에 도착해서부터 시작된 아주 어린 침팬 지 와슈 Washoe 의 생 활은 장기 간의 우발적 학습과정 이 었다. 와슈는 사 람의 어린아이처럼 행동해야 했다. 단지 다른 점은 유일한 의사소통 방법으로 신호언어 (ASL) 를 사용하는 환경에서 살았다는 것이다. ASL 을 선택한 이유는 어린 침팬지에게 초기에 음성언어를 가르치려던 시 도가 실패했기 때문이다 . 와슈는 컵으로 마시는 것을 배웠고 식탁에 포크와 숟가락을 가지고 앉고 옷을 입고 벗기 그리고 변기 사용하는 것을 배웠다. 와슈는 장난감을 가지고 놀며 인형을 돌보았다. 책에 있 는 그림을 보고 ASL 을 이용하여 내용을 이야기하였다. 결국 와슈는 야생 생활로부터 획기적으로 변한 생활방식에 적옹하였다고 제인 구 달은 말하였다. 와슈의 적응은 타고난 본능에 의하여 크게 영향받지 않는 어린 침팬지의 학습능력을 나타내는 것이다. 모든 것이 청년기 동안에는 잘 진행되어 와슈는 130 가지 ASL 신호를 배웠다(제 4 장). 그 러나 사춘기 후에 와슈는 심한 정신적 혼란에 직면하게 되었다. 자신
을 사람의 딸로 생각하고 자신의 두 침팬지 새끼에 실망한 듯 보였으 며 결국 새끼들은 오래 살지 못하였다. 7.2 의사소통을 위한 부호의 학습 제 4 장에서 신호나 부호를 가지고 침팬지의 의사소통을 가르치는 여 러 방법을 이미 살펴보았다. 특수하게 설계한 기술로 막대한 노력이 기울여졌댜 브라운 Brown 은 그 동기를 다음과 같이 생생하게 표현하 였다. 왜 사람들은 관심을 갖는가? 아마도 우리가 우주 여행에 관심을 가지 는 이유와 같을 것이다. 이는 오로지 우리가 우주에서 유일하게 언어를 사용하는 종이기 때문이다. 우리는 침팬지도 말할 수 있기를 바란다 . 그 래서 <여보세요, 저, 침팬지라는 존재는 무엇 같습니까? > 라고 물어 보고 싶은 것이다 (1980: 88). 동일한 학습 계획을 가지고 사람과 침팬지를 비교하는 것은 홍미 있는 일이다. 이것을 프레막 (1976) (제 4 장)이 기술한 방법으로 렌네버그 (1975) 가 수행하였다. 그림 7 .1에 보여주는 것처럼 침팬지를 언어 자석 판 앞에 앉게 하고 그위에 플라스틱 조각을 선택하여 부착하게 하였 댜 어린 암컷 침팬지 사라 Sarah 와 대단히 효과적으로 의사소통이 이 루어지게 되었다(제 4 장). 렌네버그 (1975) 는 정상적인 고등학교 학생을 사라와 똑같이 훈련하였다. 고동학교 학생은 사라보다 훨씬 빨리, 틀 리는 점수가 적어졌다. 그러나 언어판에 있는 단순한 문장을 정확하게 영어로 옮기는 데는 실패했다. 실제로 그들은 플라스틱 부호와 언어 사이의 합치성을 이해하지 못하였고, 오히려 수수께끼 풀기로 알고 있
 5
5
는 것 같았다. 이에 반하여 사라는 이 과정 전체를 단순히 주스를 보 상받기 위한 것이라고 우리는 추측할 수 있다. 침팬지가 보여준 의사소통의 신빙성을 여러 방법으로 알아내려 하 는 학습을 통해 침팬지가 인지적 기억에 있어 잘 발달된 능력을 가졌 다는 것을 밝힐 수 있었음을 인식하는 것은 학습에 관해 다루는 이 장
의 주제에 중요성을 지닌다. 기억은 아마도 주기적인 강화에 의하여 수년 동안 유지될 것이다 . 유아에서부터 계속되는 인지적 기억은 유아 에서 소년, 청년, 성인에 이르는 정상적인 발달에 절대적인 것이다. 그 래서 대단히 보수적인 침팬지 사회의 일원으로 받아들여지게 되는 것 이댜 7.3 유인원의 학습과 인간 학습의 비교 제 4 장 그리고 위에서 말한 유인원의 학습에 관한 연구는 사람의 학 습과 유사한 점이 많다는 것을 보여주었다. 예를 들어 새끼에서 청년 기에 걸친 모든 사회적 상호작용의 복합적 학습이 이루어지며, 가두어 기른 어린 침팬지는 사람의 어린아이처럼 행동하는 것을 대단히 잘 배 웠댜 운동 학습은 잘 이루어졌으며 일부 언어소통도 신호 또는 부호 언어로 가능했다. 침팬지의 사물 식별능도 잘 발달되었다. 그들은 자 신이 하는 일을 알았고 주스의 보상없이도 작업에 홍미를 가졌다. 피 그미 침팬지 칸지Kanzi의 수행 능력은 보통의 침팬지보다 훨씬 우수했 다고 사배지-룸바우 (Sava g e-Rumbau gh 1985) 는 기술하였다. 예를 들어 칸지는 놀라울 만치 영어 회화를 잘 이해하였으나 영어를 발음하는데 는 실패했다. 이 같은 언어학적 연구로 어린아이와 어린 침팬지의 근본적인 차이 는 언어에 있음을 알게 되었다 . 어린아이는 의욕을 가지고 언어의 세 계로 들어가고 문법적으로 바른 문장 구성을 빨리 배운다. 그렇게 해 서 세상을 배우게 된다는 것은 의문의 여지가 없다. 테라스 Terrace 는 인간의 학습과 기억에 있어 언어의 핵심적 역할을 강조하면서 다음과 같이 말하였다.
사람의 기억에 관한 대부분의 연구는 언어 자극을 이용한다 . 비언어적 자극을 이용하는 경우라도 기억은 언어적 가억술과 조절 과정에 의해 증 진된댜 그 같은 기억술과 조절 과정이 없는 경우에는 동물이 자극을 언 어적으로 예행 연습한다거나 동물과 사람 간의 부호화 과정이 대단히 중 첩되었으리라고 추측하는 데는 무리가 있다고 생각된다 . 동물에서의 인 지과정은 사람의 경우에 비하여 그 생물학적 한계에 의하여 훨씬 제한될 것이라는 것은 분명한 것 같다 (1985 : 126). 호미노이드(침팬지 같은)로부터의 진화에서 학습이나 기억에 대한 고도로 발달한 대뇌기전은 이미 발달될 준비가 다 되어 있었다. 이점 에 관하여는 뒤에 논한다. 하지만 사람의 수행 능력은 차원이 다른 정 도의 차이가 있다. 침팬지가 습득한 수행 능력은 세 살짜리 어린아이 보다 못한 수준이다. 사람 성인의 수행 능력과는 비교가 되지 않는다 . 그리고 이는 살아가며 계속 발달하고 문자 기록, 컴퓨터 기억, 또는 수 학적 부호 등의 불변하는 형태로 습득한 학습 내용을 이용할 줄 안다. 7.4 기억에 관계되는 뇌의 부위별 크기 호미니드 진화 과정에 뇌에 새겨진 진화적 변화의 일부는 인지적 및 운동 학습에 관계되는 영역의 크기 지수를 조사함으로써 살필 수 있다 . 이미 그림 3.1 과 5.1 에서도 보여주었듯이 스테판 S t e p han 과 그 동 료들은 광범한 각종 영장류의 여러 대뇌 성분의 크기를 측정하여 발표 하였다. 표 7.1 에는 인지적 기억과 관계된다고 믿어지는 뇌의 부위별 크기 지수를 기초 식충류를 기준으로 나타낸 것이다. 기본은 다른 폰지드보 다 지수가 높다. 이는 아마도 지수 계산에 있어 상대적으로 체중이 작
표 7.1 인지적 기억에 관계되는 뇌 영역의 크기 지수 (Tenrec i nae 에 대한 비교 값) 분열피질 해마 간뇌 신피질 Tenrecin a e 평 균 (N=4) 1.00 1.0 0 1.00 1.00 식충류 평균 (N=50) 1.68 1.75 1.56 2.65 원원류 평균 (N=l8) 2.80 2.9 1 5.56 20.37 신구세계 원숭이 평균 (N=23) 2.23 2.64 8.0 0 48.4 1 기본 3.89 5.07 10.9 0 65.58 고릴라 1.45 1.72 6.4 3 48.3 2 침팬지 1.81 2.1 8 8.3 9 71. 73 폰지드 (N=3) 2.38 2.99 8.5 7 61. 88 호모 (N=l) 4.43 4.8 7 14.76 196.4 1 N : 종수 (S t e p han , 1988)
표 7.2 동작 및 운동학습에 관계되는 뇌 영역의 크기지수 (Tenrec i nae 에 대한 비 교값) 소뇌 간뇌 선조 신피질 Tenrecin a e 평 균 (N=4) 1.00 1.00 1.00 1.00 식충류 평균 (N=50) 1.64 1.56 1.80 2.65 원원류 평균 (N=18) 4.64 5.5 6 5.99 20.37 신구세계 원숭이 평균 (N=23) 6.2 0 8.00 10.1 2 48.4 1 기본 9.76 10.9 0 15.4 1 65.58 고릴라 7.95 6.43 8.4 1 48.3 2 침팬지 8.7 1 8.39 11. 51 71. 73 폰지드 평균 (N=3) 8.81 8.57 11. 78 61. 88 호모 (N=l) 21. 75 14.7 6 21. 98 196.4 1 N : 종수 (S t e p han 둥, 1988)
표 7.3 소뇌에 대해 입 출 력으로 연 결 된 뇌 영역 들 의 상대적 크기지수(원원류에 대한 비 교값 ) INO VPO CER MCN ICN LCN DIE NEO 원원 류 평균 1.00 1.00 1.00 1.00 1.00 1.00 1.00 1.00 신 구 세계 원숭이 평 균 1.66 2.5 5 1.34 1.23 1.61 1.74 1.44 2.3 8 기 본 3.6 9 5.3 7 2.1 0 0.91 1.87 2.8 0 1.96 3.2 2 고 릴 라 1.97 3.5 3 1.71 0.34 0.4 5 1.51 1.16 2.3 7 침팬 지 2.20 4.5 3 1.88 0.3 7 0.6 1 2.23 1.51 3.5 2 폰 지드 평 균 2.6 2 4.4 8 1.90 0.5 4 0.9 8 2.1 8 1.54 3.0 4 호 모 4.1 4 9.4 4 4.6 9 0.79 1.06 4.5 4 2.6 5 9.64 !N O : 하 올 리브 , VPO : 복측 교뇌, CER : 소뇌 , MCN: 내 측 소뇌핵, ICN : 중간소뇌핵, LCN : 의 측소 뇌핵 , DIE: 간뇌 , NEO : 신피 질 (St e p h an 동, 1988)
은 것 (단지 5-8k g)에 기인하는 것으로 생각된다. 호모의 해마는 의의 로 지 수가 낮다 (4 . 87). 그러나 해마의 가장 발달한 부위 (CAl) 는 크기 지수가 6 . 6 이고 다른 부위들은 훨씬 작아서 CA3 은 1.7, 치상막은 2.8, 지각은 3 . 3 이다 (S t e p han 1983). 예상대로 분열 피질의 크기 지수는 해마와 거의 비슷하다. 표 7.1 과 관계되는 더 자세한 것은 7.5.1 의 인지적 기억을 참조하기 바란다. 표 7.2 는 보행과 운동학습에 관계되는 대뇌 부위들의 기초 식충류를 기준으로 한 크기 지수이다 . 힐로바테스 H y loba t es 는 다른 폰지드에 비 하여 크기 지수가 큰데 판 Pan 의 신피질은 예외이다. 표 7.3 에는 소뇌피 질의 입출 회로에 관한 것으로 지수는 유인원의 평균치에 대한 비교값 이댜 진화의 정도는 낮지만 소뇌반구는 대뇌반구와 나란히 발달한다 (Mata n o 등 1985a). VPO 영 역은 대체로 대뇌-소뇌 회로에 관련되고 크 기 지 수가 대 단히 크다 (9 . 44) . 소뇌 반구에 서 나오는 섬 유는 의 측소뇌 핵 (치상핵)을 거친다. 따라서 호모 (4.54) 와 폰지드 특히 힐로바테스 (2.80)
의 크기 지수가 높다 . 이에 반하여 다른 소뇌핵 MCN 과 ICN 은 전유인 원p ros imi an 에 바하여 퇴행하는 경향이 있다. 힐로바테스의 높은 값은 ’ 일부 그 체중이 작은 것으로 설명된다. 그러나 이 월등한 완족류에서 소뇌와 그것에 입출하는 회로가 진화적으로 고도의 발달을 보였다는 것은 생각해 볼만한 일이다 . 표 7.2 와 7.3 에 관하여는 운동학습에서 더 살펴보게 될 것이다. 7.5 학습과 기억의 신경과학 인간의 기억에 있어 진화적 발달을 제대로 평가하려면 포유류의 인 지적 및 운동 학습과 기억에 관한 신경과학적 이해의 현황을 알아볼 필요가 있댜 최근의 발전은 눈부신 바가 있다. 그러나 아직도 갈길이 멀다. 제 3 장과 제 6 장에서 살펴본 바와 같은 기초 신경과학 연구의 발견 들은 영장류에서 이미 완전하게 발달하였고 호미니드를 이어받아 종으 로 계승되었다고 생각한다. 그러나 호미니드 진화에 있어 위대한 호모 사피엔스 사피엔스를 부여받게 되는 데는 한충 높은 완성과정이 있어 야 한다 여기서 다루려고 하는 것은 호미노이드 뇌 기능의 현황을 조 망하고 고등 포유류를 호미니드 진화의 모델로 고려해 보는 것이다. 7.5.l 인지적 학습과 기억 제 3 장과 제 6 장에서 뇌의 기본적 생물학적 단위인 신경세포와 신경 섬유의 임펄스에 의한 신경세포간의 정보 교환에 관하여 살펴보았다. 이제 신경세포간의 정보 교환이 이루어지는 미세구조인 신경접합부에 관하여 살펴봄으로써 신경 기능을 자세하게 알아보고자 한다· 그림 7.2a 는 여러 개의 신경섬유가 한 개의 신경세포에 모여들어 접합마디
(syn ap tic knob, bou t on) 를 이루고 있는 것이다 . 그림 7.2b 에는 대뇌피질 의 추체세포가 있는데 큰 첨단수상돌기의 가지들을 특별히 보여주고 있댜 거기에는 실제로는 약 1 만 개나 되는 작은 돌출부에 붙은 접합 종말들이 몇 개 그려져 있다. 흥분성 접합부룰 확대한 그림 a, b, c, d 에는 접합전마디에 접합부소포가 차있고 그 소포에는 접합 간격을 건 너 전달되는 화학물질을 함유하고 있음을 나타내었다. 그림 7.3b 에는 신경세포의 세 개의 커다란 수상돌기 그루터기를 보여 주는데 그중 긴 그루터기에는 그림 7 . 3a 에 확대하여 나타내었던 것과 같 은 모양의 여러 개의 돌출부 신경접합을 볼 수 있다. 접합 간극 근방에 두어 개의 접합부소포가 그려져 있는데 실제로는 30 개 정도이다. 거 의 100 년전에 스페 인의 신경 해부학자 로만 이 카할 Roman y Caja l 은 학습이라는 것은 접합부의 극심한 활동으로 접합부가 강화되어 이 루어지는 것이라고 제안하였다. 많은 변천(H ebb 1949) 을 거쳐 이 개념 은 가설을 이루게 되었다. 이를 간략하게 살펴보기로 한다. 그림 7 .4는 토끼 해마를 이용한 실험 절차를 나타낸 것으로 현대적 연구의 시작이다 (B li ss 와 Umo 1973). CAI, CA3 및 치상막 신경계 신경 구조의 개 략이 그림 7 .4 a 에 있다. 치 상과 립 세 포의 EPSP(Excit ation Posts yna p tive Po t en ti al, 홍분성 접 합 후 전위) 빈도를 세포 밖에서 기록한 고전적 결과가 그림 7 .4에 있다. 치상과립세포로 가는 관통회로(pp)의 단일 자극으로 유발된 하향음편 향을 (b) 에서 볼 수 있다. 그 회로(pp)에 대한 네 번의 짧은 강직 자극 (15 초 동안에 15/sec) 후 EPSP 빈도는 접 합부가 두 배 강화되 고 10 시 간 이상 지속됨을 (c) 에 보여주고 있다. 이 같은 신경접합의 장기적 강화 의 발견은 곧 기억의 훌륭한 접합부 모델로 인정되었다. 이는 마치 접 합부가 집합부 활동을 기억하는 것이라 생각되는 것이다. 뇌의 하위수 준 접 합부는 이 같은 소위 장기 간 강화 (LTP, lon g term pote n ti a ti on) 를 나타내지 않는다.
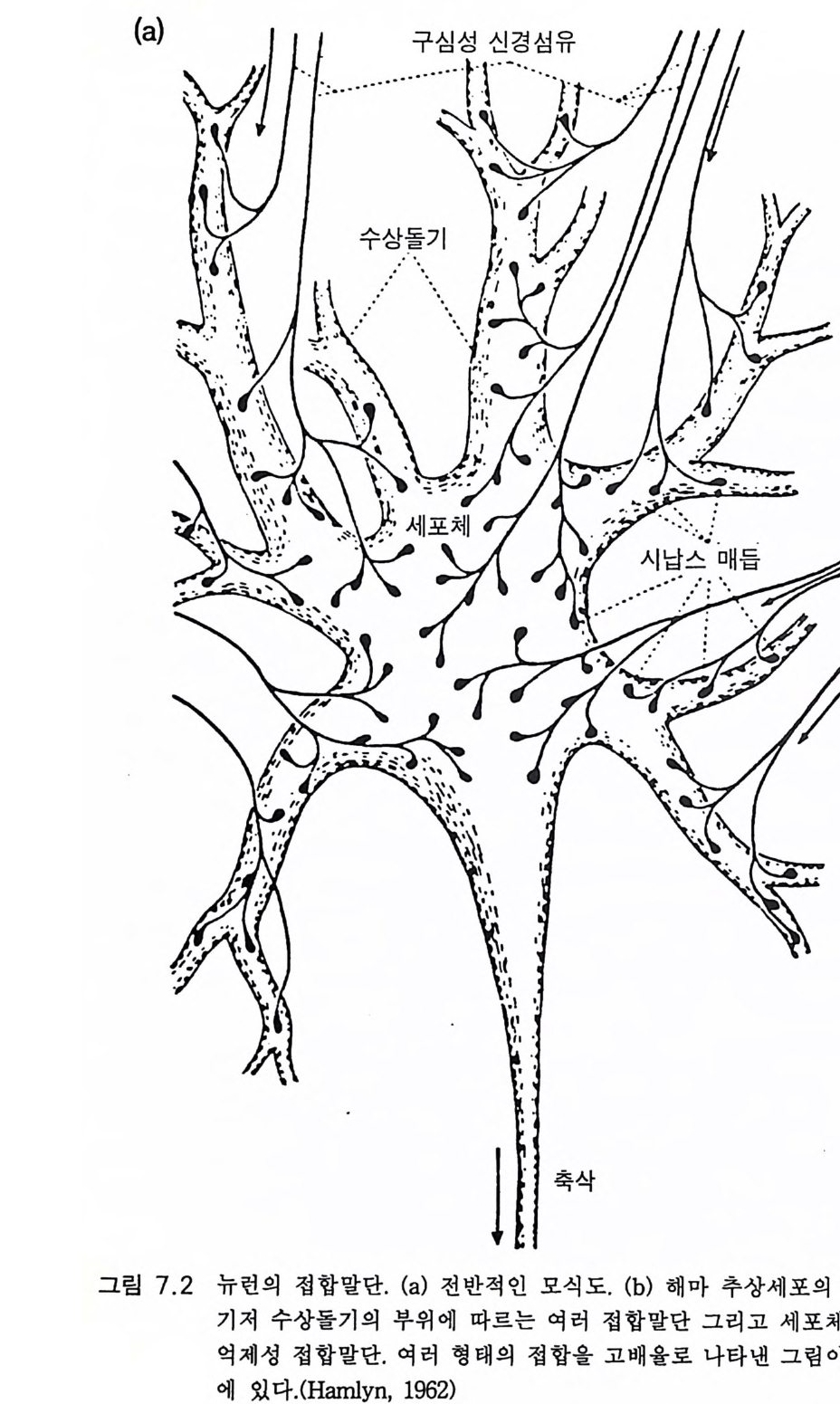 (a) I I I 구심성 신경십유
(a) I I I 구심성 신경십유
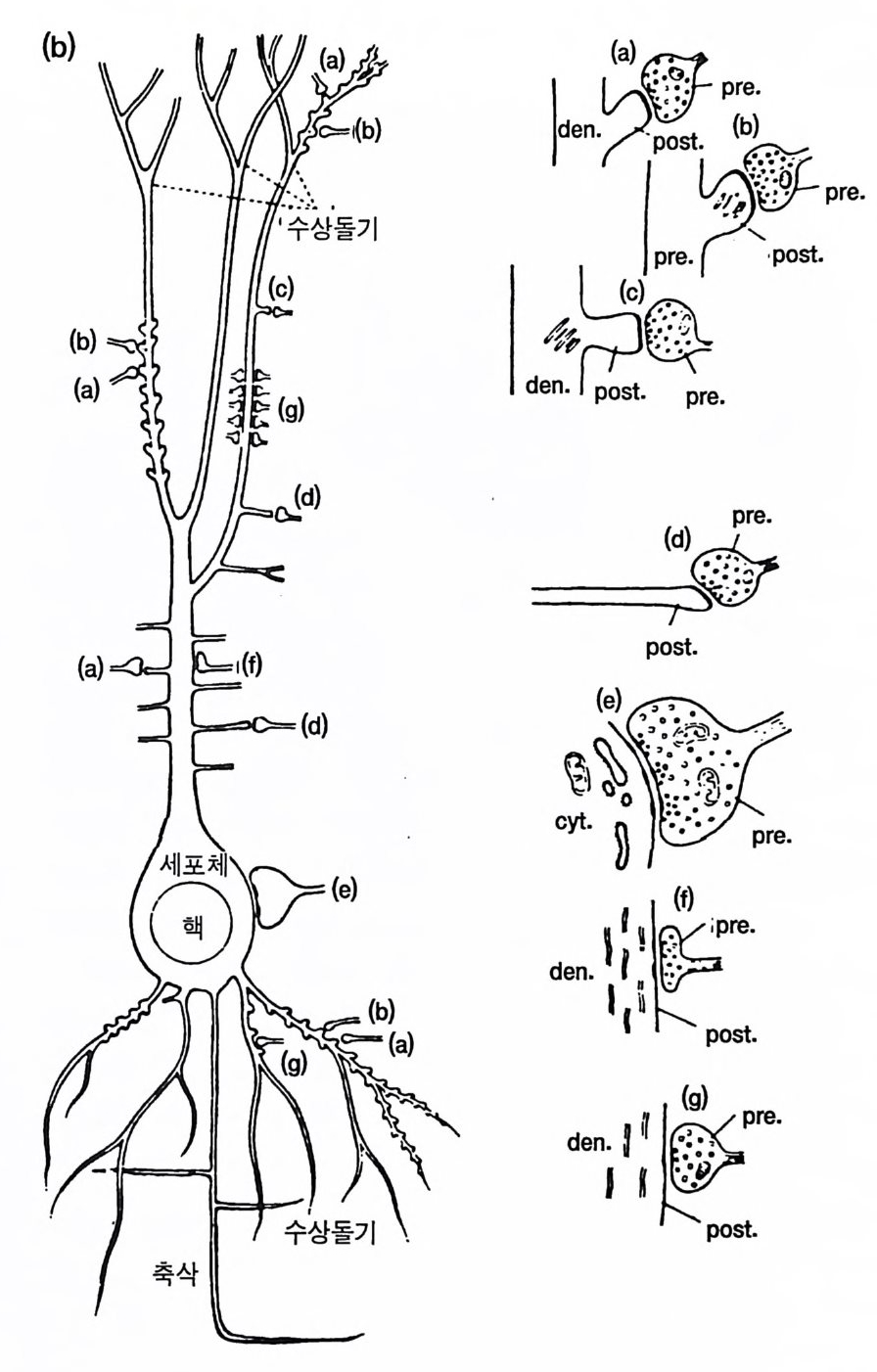 回
回
기억에 대한 모델로서 더욱 의미가 큰 것은 LTP 가 조건자극 강직의 결과에 국한하는 것이 아님을 발견한 것이다. 이같은 동종접합성 LTP 외에 조건자극 강직에 의해서 약간 약한 정도로 활성화된 그 신경세포 의 다른 신경접합들도 강화되었다 . 이를 이종접합성 LTP 라고 명명할 수 있댜 반면 조건강직 동안에 활성화되지 않는 접합부는 강화되지 않는다 . 이 이종접합성 LTP 는 대뇌피질의 학습 및 기억 가설을 수립 함에 있어 특히 중요하다. 학습이란 단지 일부 초기 극열자극의 기억 인 것은 아니다. 서로 연관관계에 있는 경험들을 기억하는 것이 더 중 요할 수도 있다. LTP 는 접합부의 접합후 반응의 우선적인 강화이고 신경세포에 있 는 수백 개의 접합부가 활성화될 때 발생되며 이 접합부들이 신경세포 가 장기간의 강한 탈분극을 일으키는 데 협동적으로 기여한다는 믿을 만한 증거들이 있다 . 이 협동성을 설명하는 가설들은 후에 살펴보기로 한다. 한편 LTP 의 기본 성질은 일차적으로는 접합후성이라는 것이고 가장 중요한 진화적 혁신으로 간주할 수 있는 것이라는 것이며 인지적 기억의 열쇠 역할을 한다는 것이다. 또다른 진화적 발달은 접합전 LTP 로 이루어졌는데 연체동물 아플리시아 A p l y s i a 는 이것으로 대단히 제 한된 기 억 을 수행 한다 (Kandel 과 Schwarz 1982). LTP 에 관련한 최근의 발견들이 그림 7 . 3a 에 있다 (Gus taf sson 과 Wi gs tr om 1986, McGeer 등 제 16 장 1987, Eccles 1988). 이중 수용체 구조 로 되어 있는 접합후막과 접합부 전달물질 글루타민이 접합소포로부터 접합부 간격으로 유리되는 것을 도식적으로 나타내었다(그림 8.3 a ). 글루 타민산염 은 접 합후 막의 키 스카레 이 트 감수성 부위 (quisq u alate sensit ive comp o nent: QQ)에 작용한댜 그래서 특별히 나트륨 이온 통로 룰 열어 강한 전기화학적 경사를 따라 나트륨 이온이 접합가시 s yn a pti c spi ne 내로 쏟아져 들어온다. 그래서 그림 7 .4 b 에서 볼 수 있는 탈분극, 죽 EPSP 를 생성한다 그의의 글루타메이트 수용체는 글루타메이트와
그림 7.4 (a) 해마 절단면 . 내비피질 (en t o) 로부터 관통신경로(pp)를 거쳐 그 축 삭(태상섬유: mf)이 근막의 과립세포에, CA3 추상세포에 그리고 쉐퍼 부축삭 (Sch) 에 의해 CAl 세포에 이르는 신경로 . (b) 과립세포의 세포 체 , 수상돌기 그리고 태상섬유(mf)를 형성하는 축삭의 모식도 . 관통신 경로(pp)는 수상돌기를 가로지르면서 홍분성 시냅스를 만든다 . pp 시 냅스 수준에서의 영역 전압의 기록에는 크고 지연성안 음전위가 유발 된다. < 집단 e . p . s . p . >로 그림에 표시하였다. 기록 전극을 세포체의 수 준으로 전진시키면 세포체에서 생성되는 임펄스인 예리한 음성극파 (집 단극파)가 나타난다. (c) 집 단 EPSP 의 상대 적 진폭을 화살표로 표 시한 네 번의 조건화 자극 후에 10 시간까지 도시한 것이다 . 화살표가 하나인 것은 초당 15 번의 자극이고 화살표가 두 개 포개진 것은 3 초 동안 초당 100 번 자극이다. 100 퍼센트 선은 0 . 5 초 때의 자극 전 반옹을 나타내고 조건화 강축자극 후에도 비슷한 낮은 수준으로 회복된다. (d) 집단극파의 그림 (c) 에서와 같은 자극후 중강의 시간 경과. 단 한 번의 조건화 강축자극을 15 초 동안 초당 15 회 주었다 .(B li ss 와 Lomo, 1973)
유사한 기 질 (N-me t h y I - D-as part a t e; NMDA) 에 대하여 특이 적 인 감수 성을 나타낸다. 그러나 이 반응은 수상돌기에 있는 다수의 접합부 활 성의 누적된 EPSP 에 의하여 접합후막이 강력하게 탈분극되었을 때에 만 일어난다(그림 7.3 b ). 그래서 접합부소포에서 방출된 글루타메이트 는 NMDA 이온 통로를 열 수 있고 대단히 강력한 전기화학적 구배를 따라 칼슘 이온이 접합가시 내로 들어오게 한다(그림 7.3a). 이 NMDA 작용은 정상상태에서 NMDA 수용체를 차단하고 있던 마그네슘 이온 을 EPSP 의 강력한 탈분극으로 밀어냈을 때에만 발생할 수 있다. 그림 7.3a 의 QQ와 NMDA 수용체 는 APV(2- ami no - 5-phosp hono-valera te)라 는 물질에 의하여 선택적으로 구별할 수 있다. 죽 APV 는 NMDA 수용 체는 차단하지만 글루타메이트에 대한 QQ 수용체는 차단하지 못한다. 그래서 큰 EPSP 의 적당한 조건하에서 NMDA 통로는 열리고 칼슘이 온이 7) 접합가시 내로 쏟아져 들어온다는 것을 알 수 있다 (Ma y er 와
Westb r ook 198. 5). 이 작용에 의하여 가시 내로 들어온 칼슘 이온은 칼모 듈린의 도움을 받아 단백활성효소를 자극한다. 그래서 접합부 소포에서 접합부 간격으로 주입된 글루타메이트에 대한 가시 수용체의 수용성이 증가한다. 이 작용의 자세한 신경화학은 아직 완전하게 밝혀지지 않았 지만 (L yn ch 와 Baud ry 1984) LTP 를 이루는 기본 요인이라고 생각할 수 있다. 이같이 간단한 이야기보다는 훨씬 방대한 연구가 가시 내로 들어 온 칼슘 이온의 이온적 작용에 관해서만도 이루어졌다 (McGeer 등 제 16 장 1987, Eccles 1987). 그러나 LTP 에 관한 실험 결과는 여기서 이야기한 것으로 대부분 설명되었다. LTP 는 활성화된 부톤으로부터의 글루타메 이트의 유리를 증가시키고 가시 내로 들어온 칼슘 이온은 그 가시 안에 오랫동안 독립적으로 촌재하지 못한다는 것이 발견됨으로써 문제가 복 잡하게 되었다. 수일 또는 수주일 동안 오래 유지되는 LTP 를 생성하려 면 칼슘 이온이 작용하여 단백, 전달물질 수용체, 여러 가지 거대분자 등을 만들어 그림 7.3b 에서 화살표로 표시했듯이 미세관을 따라 그 세 포의 핵으로 영향을 미치는 작용회로가 존재해야 한다. 문제는 이제 해마에서의 LTP 연구로 발견된 원리를 이용하여 대뇌 피질에 있어서 인지적 기억울 보다 단순한 측면들부터 설명할 수 있는 가설을 구성하는 것이다. 기억에 있어서 해마의 중요성은 처치 곤란한 해마성 전간을 해소하기 위해 양측 해마 절제술을 시행한 비극적 증례 인 환자 HM 과 또 다른 두 명의 환자들의 예에 의해 밝혀졌다(Mil ner 1968. 1972). 그들의 간질병은 치료되었지만 예상 외로 환자는 인지적 기 억능력을 완전히 상실하는 비극적 일이 일어났다(전향적 기억상실). 그러나 수술 이전에 기억했던 대부분의 기억은 유지되었다. 죽 후향적 기억상실은 별로 없었다. 이 증상들은 수술 후 30 년 이상 전혀 회복되 지 않고 있다. HM 환자는 순간순간 단기간 기억만으로 살았고 불과 수초전에 일어났던 사건이나 사실도 기 억하지 못하였다(Miln er 1966). 원숭이에 양측 해마 절제술을 시행하여 이에 비견되는 기억상실을 볼
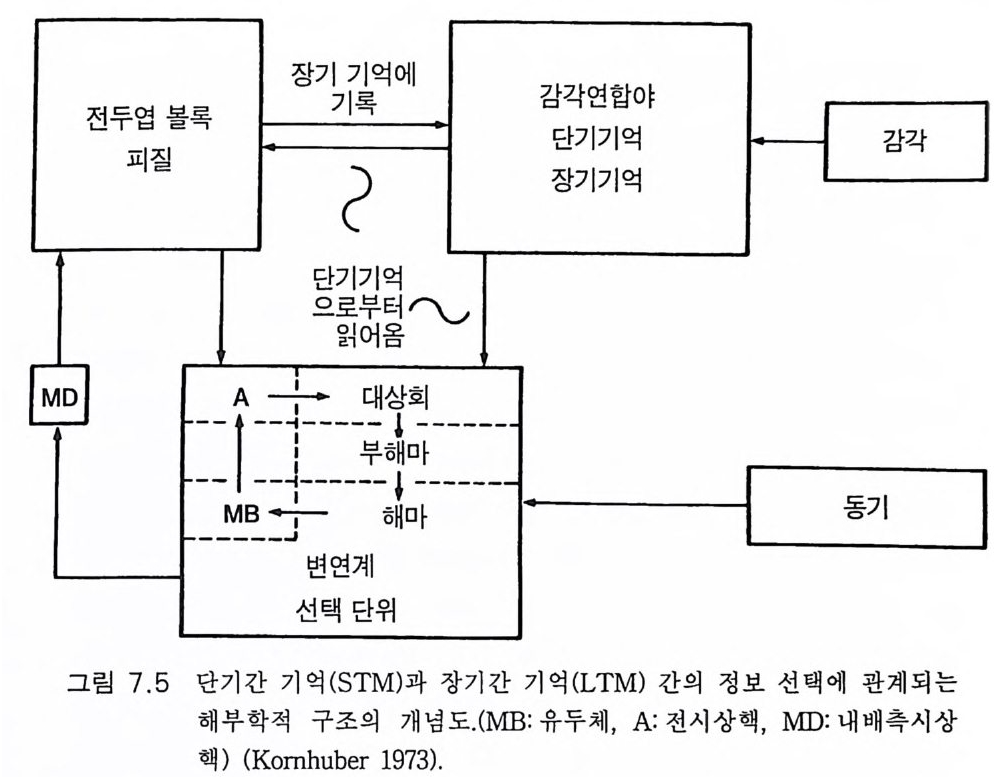 전두피엽질볼 록 장기기기록?억 에 감장단각 7711연7711합 억억야
전두피엽질볼 록 장기기기록?억 에 감장단각 7711연7711합 억억야
수 있었다. 이제 다루고자 하는 가설은 콘후버의 이론 (Kornhuber l c.!7 3) 에서 개 발된 것인데 그림 7 . 5 와 같다. 측두 및 두정엽의 감각연합영역(그림 3.5, 4.3 , 4.4 , 6 . 6) 은 촉각, 청각 및 시각의 입력을 가지고 결정적인 역할을 한다. 첫째
두 가지 입력 회로의 수렴을 통해서 생산되는 것이다 . 그림 7.6 에 이 합치점을 보여주고 있다. 한편으로는 대뇌피질 극성 위 성 세 포 (Ss t)의 축삭 (Ax) 을 거 치 는 MD 시 상으로부터 의 입 력 으로서 이는 상행하여 그림에 표시된 세 개의 추상세포의 첨단수상돌기와 복 합접합을 이루고, 다른 한편으로는 그림 7 . 5 의 감각연합영역에서 기시 한 수평섬유로서 이는 제 1, 2 층의 첨단수상돌기와 접합을 이룬다. 그림 7.6 의 첨단수상돌기의 복합접합이 이들 수상돌기의 큰 접합 탈분극 (E PSP) 을 생성할 수 있다고 추측할 수 있다 . 비슷한 시기에 활성화되 는 이들 수평섬유는 그들의 NMDA 통로를 열어서 글루타메이트에 의 한 접합 활성을 일으키고 결국 칼슘 이온이 유입된다(그림 7.3a). 해마 모델에서처럼 칼슘 이온은 단백 키나제를 활성화하여 가시접합에서 QQ 수용체 부위의 글루타메이트에 대한 감수성을 증가시킴으로써 LTP 효과를 나타낸다. 따라서 LTP 는 장기 기억의 기본이 되는 것이다. 해마 모델의 한 차이점을 주목할 필요가 있다. 그림 7 . 6 에서의 접합 부의 비대칭성은 극성 위성 세포의 QQ 수용체에 의한 강력한 접합종 말은 추상세포의 첨단수상돌기를 탈분국시키는 데만 이용됨을 뜻하는 것이댜 반면 제 1, 2 층에 있는 약하고 서로 교차하는 복수 접합은 이 탈분극으로 NMDA 통로를 열어서 칼슘 이온의 유입에 의해 LTP 를 초 래한댜 모나간 (Mona g han 1985) 둥은 면역조직화학적 방법으로 NMDA 수용 체가 신피질의 제 1, 2 층에 밀집해 있음을 증명하였다. 그림 7.6 에서 1, 2 층이라고 표시한 것이다 . 인지적 기억에 관한 이 가설을 뒷받침하는 또 하나의 증거는 해마에서처럼 LTP 가 신피질 수용체에서도 증명되 었다는 것이다 (Lee 1983). 그러나 연구가 더 필요하다 . LTP 가설의 발 전에서 제기되는 문제는 효과적인 신피질 활성을 위해서는 단일 접합 에서 방대한 공간적 증폭이 형성되어야 한다는 점이다. 이 점에 관한 초보적 시 도가 있었다 (Eccles 1981a, 그림 7 과 8). 장기 기 억 에 관한 보
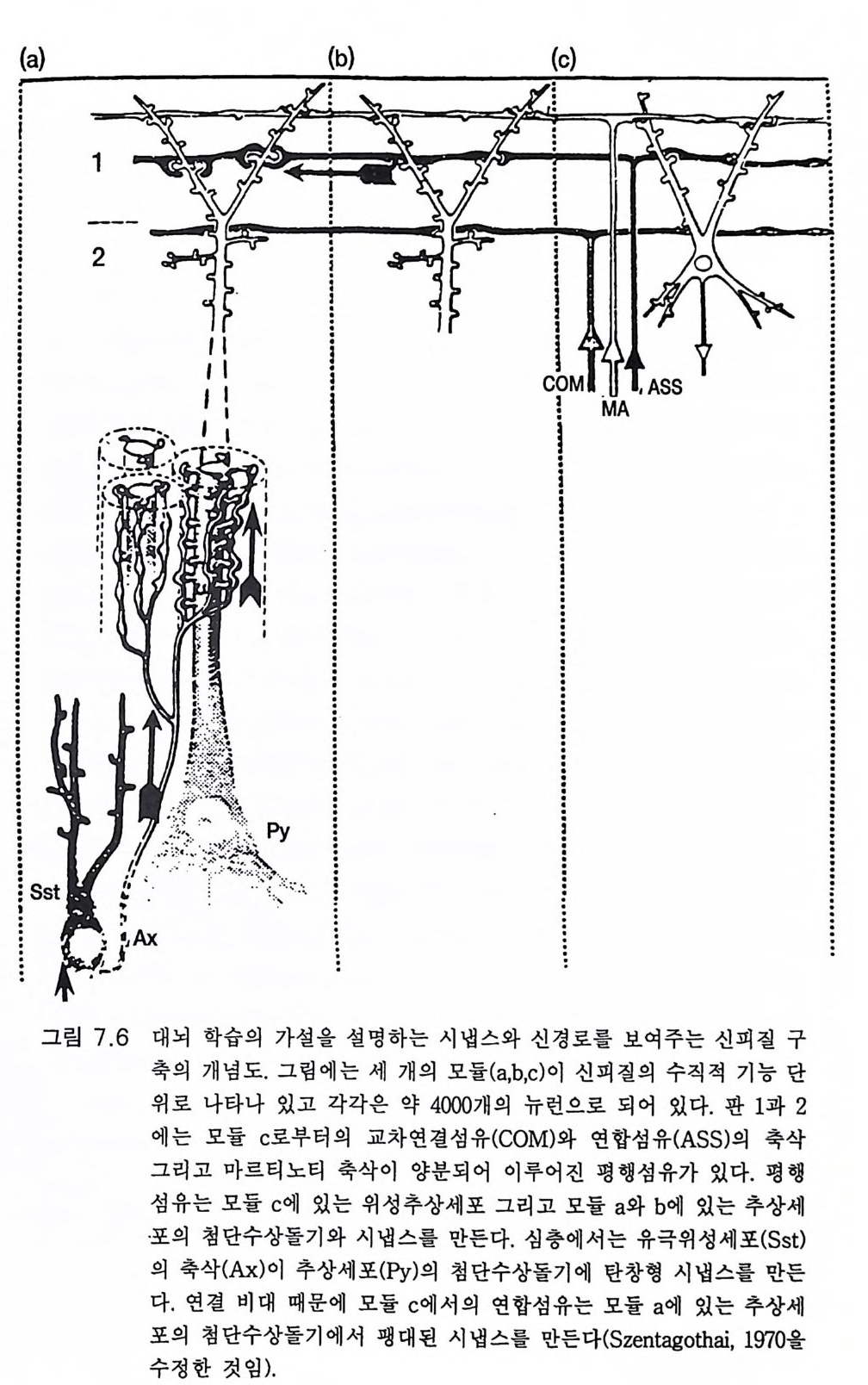 bL.!. @-f
bL.!. @-f
다 결정적 문제는 의식 상태를 경험하게 하고 의식적으로 유발시킬 수 도 있는 장기 기억들에 관한 것이다. 이 마음과 뇌의 문제는 제 8 장에 서 다루게 된다. 이 LTP 의 신경과학은 포유류 진화 초기부터 발달하였고 쥐, 기니피 그, 토끼의 해마에서 완전하게 볼 수 있다 . 영장류에서는 특별하게 연 구되어 있지는 않으나 표 7 .1에서 해마 그리고 간뇌와 분열피질에 관 계되는 구조의 크기 지수의 증가를 나타내고 있는데 이 증가는 모든 폰지드와 호모의 경우 신피질의 크기 증가에 비하면 훨씬 작은 것이 다. 사람에서 인지적 기억능의 엄청난 증가를 고려할 때 두 가지 중요 한 요인이 있다 . 첫째 연합피질 특히 전두전엽이 크게 증가하였다는 것이댜 브로드만(1 912) 은 전두전엽의 전체 신피질에 대한 비율을 호모 의 경우 29% 로, 침팬지는 16.9% 로, 마카크는 1 1. 3% 로 하였다 . 전체 신 피질의 상대적 크기에서도 사람의 전두전엽피질은 유인원의 것보다 5 배나 크다. 둘째 언어의 역할이다. 사람의 인지적 기억은 대부분 언어 로 작성된다. 인지적 기억에 대하여 해마가 상대적으로 크게 기여하지 못한 것(표 7 .1)에 비하여, 인지 능력의 발생에 있어서 이들 두 가지 측 면의 진화상 발달은 훨씬 중요한 것이라 할 수 있다. 호미니드 진화에 있어 호모 하빌리스는 뇌가 중대하고 언어영역이 발달하여 가장 큰 진 화적 도약을 이루었다고 제 2 장에서 제안한 바 있다. 이는 또한 석기 사용에 기초한 문화를 창시하였다. 이 문화는 인지적 기억의 개선에 의존하였을 것임을 짐작할 수 있다 . 7.5.2 운동 학습과 기 억 사람과 원숭이의 신생아는 몇 가지 본능적 동작 외에는 대단히 무 기력한 상태라는 것을 7 .1에서 살펴보았다. 첫해 동안에는 운동 학습에 집중한다. 이 같은 관점에서 침팬지 새끼는 일어서고 붙잡고 기어오르
는 동작을 사람의 어린아이보다 두 배나 빠르게 배운다 . 이는 아마도 사람의 어린아이는 미숙하게 태어나기 때문이라고 생각된다 (5 . 5) . 소뇌는 뇌의 상당 부분을 차지한다(그림 3.1 ). 그리고 동작의 조절에 거의 전적으로 관련된다 . 세 가지 종의 폰지드 소뇌의 크기 지수는 유 인원보다 거의 두 배나 크고 동작과 관계되는 기타 구조(간뇌와 선조一 기저핵)도 비슷한 정도로 크다는 것을 표 7.2 에서 볼 수 있다. 모든 구 조의 크기 지수는 호모가 훨씬 크지만 비율은 신피질에 대한 것보다 훨씬 작다. 소뇌의 진화에 있어 소뇌반구는 층부와 중간부가 비교적 내측에 위 치하여 자랐다는 것은 오래전부터 알려졌다. 이 같은 상대적 변화는 소뇌피질의 세 가지 구성분에 해당하는 세 가지 소뇌핵에 반영되는 것 으로 볼 수 있다(표 7.3 )( Mata no 둥 1985b). 유인원에 비하여 표 7.3 에 있는 모든 종은 내측소뇌핵 (MCN) 이 충부의 감퇴에 수반하여 감퇴하 였댜 이는 몸체와 사지의 자동운동의 감퇴를 반영하는 것이고 이는 척수와 뇌간이 담당하는 것이다(그림 3.2 및 3.7). 중간소뇌핵(I CN) 은 혼 합적 진화 변천을 보인다. 소뇌 및 중간소뇌핵의 중간엽의 이중 역할 에 해당하는 것이다. 한편으로는 뇌간과 척수의 동작조절에 관계되는 것이고 또 한편으로는 반대측 대뇌피질에 관계되는 것이다(그림 3.6, 3.1 4 ). 이에 비하여 원숭이와 호모에서, 특히 호모에서 대뇌와의 관계가 더욱 밀접하게 발전한 외측 소뇌핵 (LCN) 이 발전되었다. 더욱이 완벽 한 상완동물인 기본 Hy loba t es 은 하올리브 (IO), 소뇌핵 그리고 소뇌 전 체가 모든 원숭이 중 가장 높은 지수를 갖는다. 이것의 운동 민첩성은 진화상에서 소뇌에 반영된 것이다. 하올리브 (INO) 에 관한 표 7.3 의 수치는 특히 홍미 있는 것이다. 소뇌 의 학습과정에서의 주요 역할 때문이다 . 이는 다음에서 설명한다. INO 의 크기 지수는 LCN 과 밀접하게 연관된다 . 양자는 모두 동작의 고수 준조절에 관계되는 것이다. 복측교 (VPO) 의 수치가 높은 것은(표 7.3)
놀라운 일이댜 교의 외복측부는 대뇌반구로부터 반대측 소뇌반구를 대부분 이어주는 것이다(그림 7.7 의 PN). 동작의 소뇌 조절이 학습 과정에 관계된다는 많은 증거들이 있다 . 그림 7.7 에 보여주듯이 푸르킨제 세포에서 나오는 섬유는 소뇌 핵(D E) 의 표적 신경세포를 억제한다. 이는 독특한 설계로서 이토(It o 1984) 가 발견하였다 이 독특한 설계와 합치하여 독특한 학습과정이 있다 . 이 학습과정도 이토가 발견하였는데 이에 따르면 평행섬유 (P . F) 에 의한 푸르킨제 세포의 흥분은 정확하게 LTP 에 반대되는 장기간 억제 (LT D) 를 나타낸다는 것이다. 이 같은 비상한 설계는 광범한 연구 (I t o 와 Kano 1982) 로 밝혀졌고 최근에 사쿠라이 (Sakur ai 1987) 에 의해 대단히 정확 하게 연구되었다 . 하올리브 (IO) 에서 소뇌의 푸르킨제 세포로 가는 상승섬유 (C.F. )를 그 림 7.7 에 보이고 있다. 각 푸르킨제 세포마다 단 하나의 상승섬유가 존 재하는데 이 상승 섬유는 수상돌기 주위를 광대하게 감싸고 오르면서 많은 홍분성 접합부를 이룬다(그림 7.1 0 ). 푸르킨제 세포의 수상돌기로 들어가는 기타 주섬유는 평행섬유로서 이는 각각의 푸르킨제 세포와 약 10 만 개의 홍분성 접합을 이룬다 . 이 중의 하나를 그림 7 .1 0 에 보여 준댜 이토 (1986) 는 상승섬유가 푸르킨제 세포의 스승으로 작용할 것이 라는 가설을 세웠다. 학습 과정은 상승섬유와 평행섬유의 활성이 연합 할 때 이루어진다는 것이다. 이 가설을 층명하기 위한 여러 실험이 있 었는데 사쿠라이의 연구에서 절정을 이룬다 . 소뇌피질의 박편을 산소가 공급되는 영양액에서 수시간 동안 유지 할 수 있는 실험 장치롤 그림 7.8 에 보여주고 있다 . 여기서 동일한 푸 르킨제 세포를 지배하는 상승섬유 (STIM 1) 와 평행섬유속 (STIM 2) 에 자극을 가할 수 있도록 장치하였다. 대단히 가는 미세전극으로 푸르킨 제 세포의 수상돌기 (REC) 에서 세포내 전위를 기록하는 대단히 힘든 조작을 적용하였다. 이 실험으로 3 초마다 가한 단일 자극 (STIM 2) 에
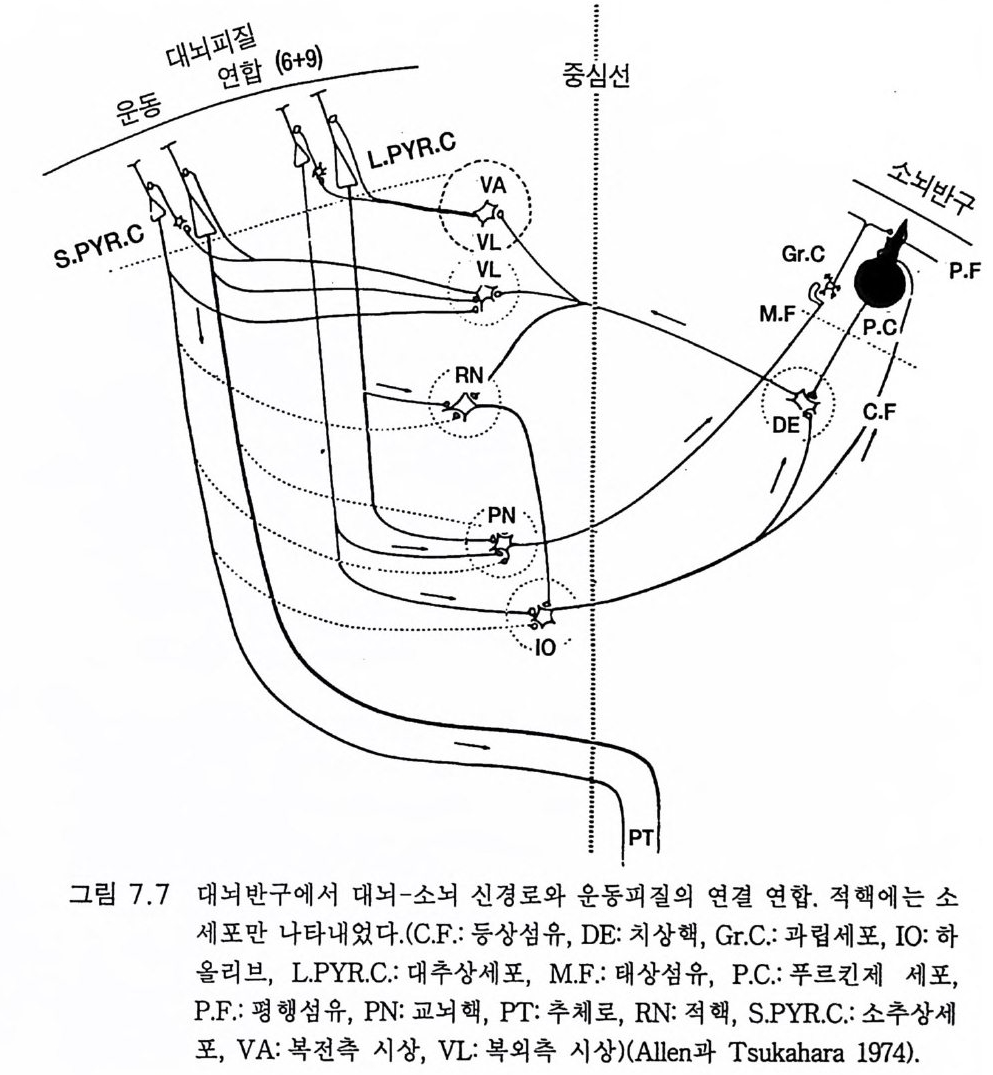 중『선
중『선
의하여 EPSP 반옹을 유발할 수 있었고 40 개의 연속적 반옹의 평균치 를 구하여 그림 7.9b 에서 영보다 작은 시간값 부분에 그 수치를 기록 하고, 또 그림 7.9a(l) 에 예시하였다. 그림 (b) 의 화살표에서 연합자극 (STIM 1 + STIM 2) 4Hz 를 25 초 동안 가하였다. STIM 1 자극은 상승
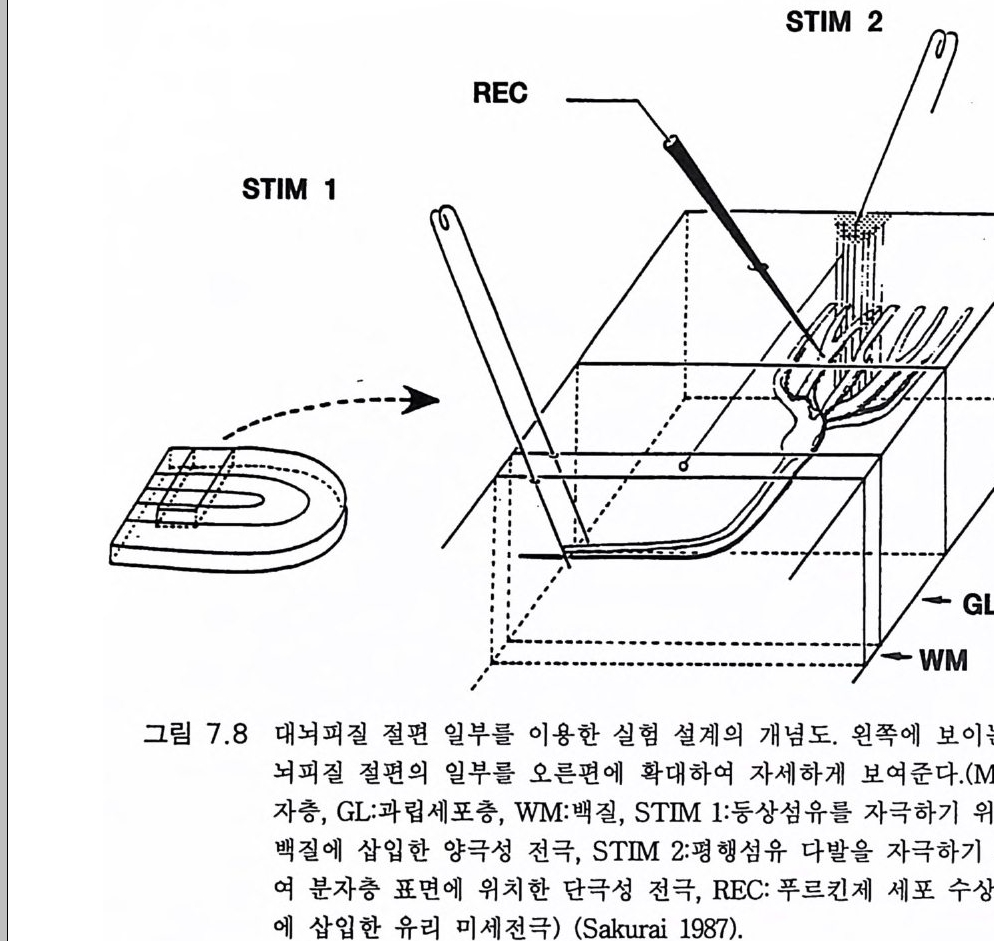 REC
REC
섬유의 역치를 훨씬 상회하는 것이었다. 연합자극으로 평균 EPSP 의 저하를 가져왔고 이는 그림 (b) 에서처럼 연합자극 후 80 분 동안 계속 되었다 . 그림 (b) 에 번호로 표시된 반응을 그림 (a) 의 기록 2 와 3 으로 예시하였는데 이를 보면 평균 EPSP 가 원래의 크기보다 60% 감소한 것을 보여주고 있으나 시간 경과에는 변화가 없었다. 또 그림의 (c) 에 서 안정막전위는 -70mV 에서 일정하게 유지되는 것을 보여주고 수상 돌기의 입력저항은 약 30M Q울 유지하였다 . 이 실험은 수상돌기에서
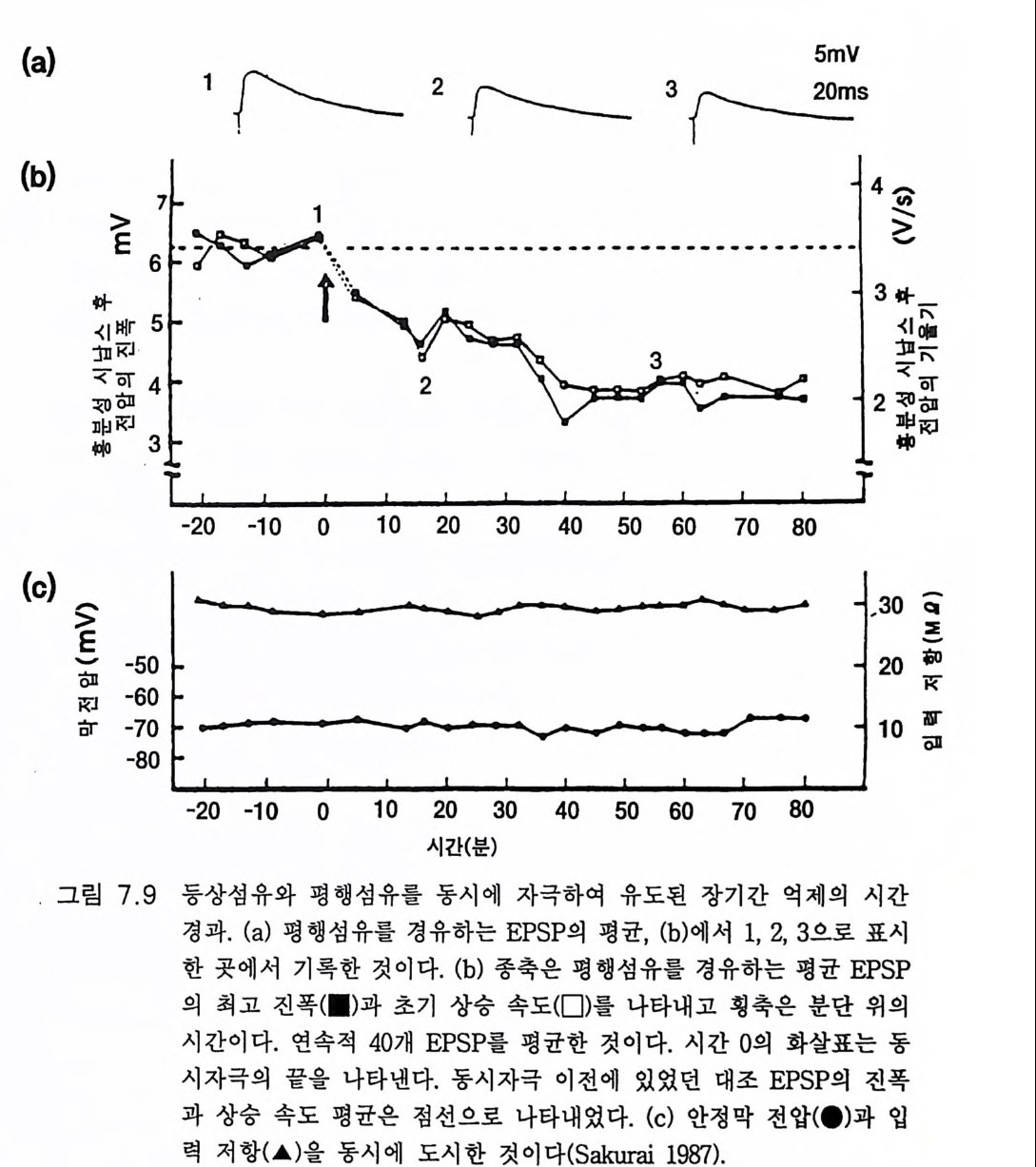 ((ba)) :1 f-t..-...... . .- .- ”-- · · -2· -r -2- -- · - -- · · --· - · - -3- 三-3- · · · · · --5·m -V .s ->)(s/아 lc-~-ck
((ba)) :1 f-t..-...... . .- .- ”-- · · -2· -r -2- -- · - -- · · --· - · - -3- 三-3- · · · · · --5·m -V .s ->)(s/아 lc-~-ck
의 세포내 기록을 2 시간 동안 일정하게 유지해야 하는 어려운 실험이 었다 10 개의 푸르킨제 세포에서 조건에 합당한 기록을 얻었는데 그중 9 개에서 그림 7.9 같은 지속적 연합억제 (LTD) 가 기록되었다. 이 같은 억제는 세포내의i n tr asoma ti c 기록 실험 세 개 모두에서도 관찰되었다. 상승섬유 임펄스에 의한 푸르킨제 세포의 강력한 활성은 수상돌기 내로 칼슘 이온을 유입시키고, 이 칼슘 이온은 아마도 단백 키나제에 작용하여 접합부 전달물질인 글루타메이트에 대한 수상돌기 접합부의 감수성을 저하시킨다는 증거가 있다 (I to 1984, 그림 7.1 0 ). 그래서 칼슘 이온은 인지적 학습의 LTP 와 운동학습의 LTD 모두에서 중심적 역할 을 한다. 평행섬유에 의한 푸르킨제 세포의 활성화를 장기 저하 (LTD) 시키면 핵세포 (D, E) 의 억제를 감퇴시키고, 따라서 억제에 의한 학습은 그 핵 세포로부터의 출력을 증강시키는 것으로 변환된다. 그림 7.9 에서 나타 낸 단순한 접합기전으로 운동 학습을 설명하는 이론을 수립하는 데는 문제가 있다 . 그러나 이와 관련한 관찰들은 이미 있어 왔다. 이에 관하여 원숭이에게 손잡이의 위치를 조절하도록 훈련시키고 손잡이를 잡고 손목의 신근과 굴근에 교대로 같은 강도의 힘을 주되, 작용시간을 1. 5-:-4 초로 다양하게 한 실험 (Gi lbe rt와 Thach, 1977) 이 인상 적이다. 원숭이는 매번 손잡이의 위치가 바뀐 후 0.5 초 이내에 그 손잡 이를 중간 위치로 되돌리는 것을 학습하였다. 그러고 나서 실험법을 바꾸었다. 예를 들면 굴근에 작용하는 부하는 변화시키지 않은 채로 두고 신근에 작용하는 부하를 두 배로 하였다 . 원숭이는 이 변화에 적 옹하는 것을 12-100 회 시행으로 배웠다. 소뇌의 정확한 위치에 미리 심어놓은 전극으로 한 상승섬유 및 평행섬유의 홍분에 의한 푸르킨제 세포의 반응을 기록할 수 있었다. 신근 · 부하의 변경은 즉시 큰 점으로 나타낸 짧은 잠복기를 가진 상승섬유의 반응과 몇 개의 느린 반응(복 합 전위 활동 the comp le x s pik es) 을 유발하였다(그림 7.l lb) . 동시 에 평 행
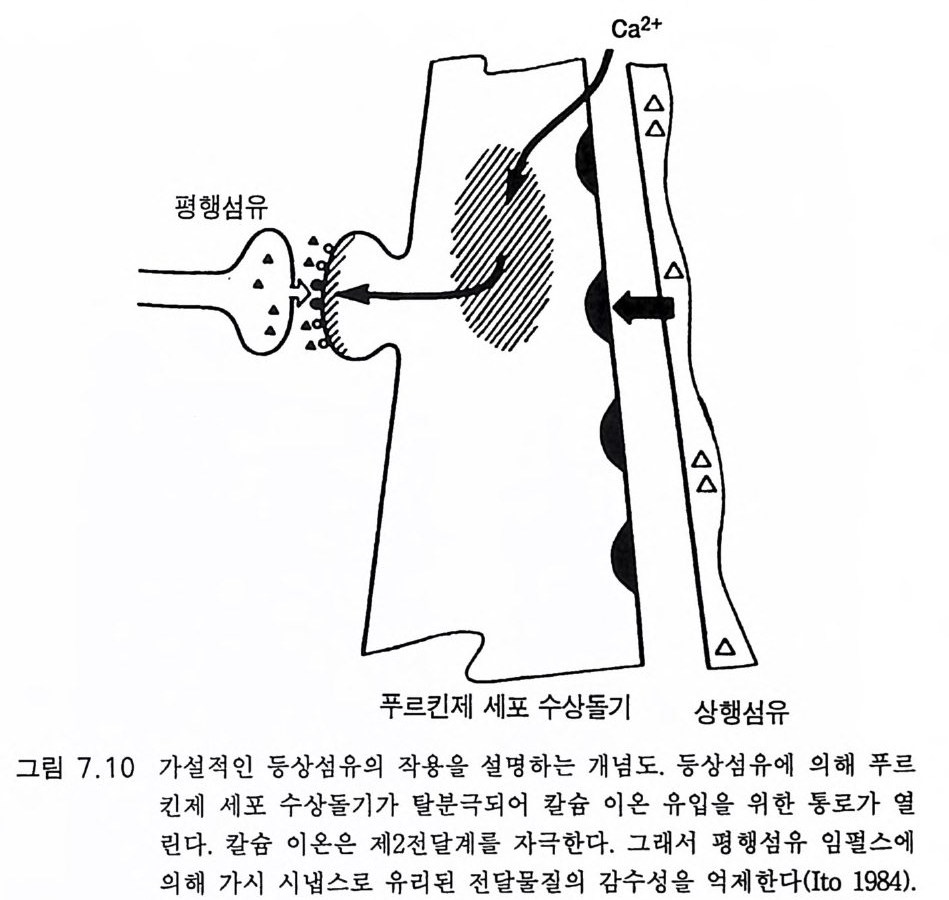 Ca2+
Ca2+
섬유의 활동에 의하여 . 생성되는 단순극파 s imp le s pik es 는 감소하였다 (작은 점). 실험 내용의 변화에 따라서 운동 수행 성적상의 변화가 일 어나고 약 80 회 시행 후 성적이 · 회복되는 것을 7.llc 에 보이고 있다. 복합극파 반도의 시간 경과가 (c) 에 있는 운동 수행 성적의 변화에 관 련되고 있다 . 원숭이가 새로운 실험법에 적옹하게 되면서 초기의 낮은 빈도 수준으로 회복되었다. 상승섬유의 입력 중가와 기존하는 평행섬 유 입력이 협동하면 그림 7.9b 에서처럼 그 입력에 대한 반응의 억제를 초래한다 이는 단순극파 홍분 빈도가 계속되어 있고(그림 7.l ld ), 그림
 (c) 신근운동
(c) 신근운동
근 반응으로 이어진다. (b) 와 (d) 에서 삼각접으로 표시된 곳에서는 알 고 있는 굴근 부하는 310 g으로 유지하면서 알고 있는 신근부하 300g 을 새로운 부하 450 g으로 변경하였다. 부하를 변경하기 전에는(삼각 형 표시 이전) 신근 시험이 시작된 후 약 lOOmsec 에 관계되는 CS 활 성의 빈도가 낮았다. 부하 변경 후(삼각 표시 이후)에는 CS 빈도가 크 게 중가하였고 이는 50 시행까지 지속하였다. 이는 새로이 학습한 적옹 에 익숙해짐에 따라 (b) 와 (d) 의 포개진 삼각 표시가 있는 곳에서 시 초에 학습하였던 수준으로 회복되었다. 이 같은 일시적 CS 빈도 중가 의 수반은 새로운 운동 과업을 학습함에 따라 SS 빈도를 감소시킨다. (c) 운동 수행 성적과 시행 횟수와의 관계. 수행 성적은 부하 변경 시 험 20 번째에는 불안정하나 50 번째 이후에는 개선되었다. (d) 중복 시 행에 따른 CS 및 SS 빈도의 변화. SS 빈도는 새로운 운동 과업을 습 득하면 감소되고 나서 안정된다. CS 빈도는 대충 50 시행 동안에 (20-70 시행) 중가하고 이는 새로운 부하에 대한 좋은 수행 성적을 습 득하는 데 필요한 횟수이 다 (Brooks 와 Thach, 1981).

7.9a 에 있듯이 EPSP 가 감소되는 것으로 잘 알 수 있다 . 그림 7 .1 1 에 운동 학습에 관한 대단히 간단한 실험을 보여주고 있다. 여기서 푸르킨제 세포로 들어가는 상승섬유의 입력은 푸르킨제 세포 에 오류 신호자로 작용하여 단순극파의 발사 속도를 감소시키는 것이 댜 결국 핵세포로부터의 출력이 증가되어 변경된 실험법에 근사하게 반응되도록 적절히 작용한다 . 그것이 그림 7.llc 이다 . 결국 약 80 회 시 행후에 그림 7.llb 와 d 에 나타나 있듯이 오류 신호는 사라지 게 된다 . 길버트와 타취 (G il be rt와 Thach 1977) 는 모든 종류의 변종실험으로 운 동학습은 상승섬유에 의한 오류 신호 발생을 기초로 하여 이루어진다 는 설명을 확인하는 결과를 얻을 수 있었다 . 7.5.3 인지적 학습과 운동 학습의 복합 인지적 학습과 운동 학습을 분명하게 구분하려는 초기 시도는 이들 두 범주의 학습 기전이 상당히 다르다는 것으로 정당화되었다. 그러나 동물이 직면하게 되는 정상적인 복합 상황에 있어 이들 두 학습 과정 을 복합할 필요가 있다. 훌륭한 예를 사사키와 겜바 (Sas aki와 Gemba 1982) 가 제공하였다. 원 숭이에게 그림 7 .1 2 의 삽입도에 나타낸 것처럼 부위 A, B, C, D, E 에 양극성 기록 전극을 심어 놓았다. 각 전극은 삽입된 장소에서 8 ) 대뇌피 그림 7.12 원숭이가 손을 움직일 때 반대쪽 다섯 군데(위쪽 작은 그림의 A-E) 대뇌피질에서 기록한 뇌파 . I-IV : 시각적으로 유도된 손동작 학습과 정의 네 단계, 훈련이 시작된 후 각각 3, 21, 24 및 59 일 . 여기서는 S-D( 표충 빼기 심층)만을 나타내었다. 100 회의 평균이다 . VS-LE 의 막대그림은 100 번 시료의 반옹 속도이다. I-III 에서 일부는 삭제되었 는데 여기서 경자극은 900rnsec 였고 끝은 기록되지 않았기 때문이 다. 모든 전압은 50µV 로 보정하였고 모든 기록은 100rnsec 로 보정하 였다 (Sas aki와 Gemba 1982).
 ?
?
질의 표면과 심부 사이의 전위 차를 기록하였다. 학습 절차는 대단히 단순한 사역 조건 학습이었다. 원숭이에게 작은 녹색 전구에 의한 시 각 자극을 주었는데 전구는 2.5-6 초간의 무작위 적 간격 후에 900msec 동안 켜지게 하였다. 원숭이는 불빛 자극이 있는 동안에 손목을 신전 하여 손잡이를 들어올리면 과실즙의 보상을 받도록 학습시켰다. 초기 에는 원숭이가 무작위적으로 손잡이를 들어올려서 가끔 보상을 받는 댜 그러나 일주에 약 1000 회의 시행으로 수주간 훈련하면 차츰 인지 적 학습과 운동학습이 이루어지고 결국 원숭이는 빛 자극이 있을 때만 손잡이를 들어올리게 된다. 그리고 시행 잠복기는 차츰 감소하여 일정 한 짧은 시간에 이르게 된다. 빛자극 시간을 500msec 로 단축해도 여전 히 잘 해 낸다(축 IV). 대강 500, 3000, 4000, 9000 번의 시행 후 다섯 장소에서 기록한 양극 성 기록이 그림 7 . 12 의 축 I-W 에 있다. 빛 자극에 대한 피질반응이 점 차 증가되는데. 특히 전두전피질 (A) 과 선조주위피질(D)에서 증가되었 댜 이 같은 증가는 인지적 학습에 의한 것으로 간주되는 것인데 그림 7 . 6 과 잘 상응하는 결과이다. 가장 홍미 있는 반응은 운동피질에 있는 것인데 이 피질부위는 손잡이 들어올리기의 운동을 시작하게 하는 부 위이기 때문이다 . 그림 7 .1 2 의 기록 C, 축 IIl 은 4000 회 시행 후의 기록 으로 작은 음전을 보이는데 과일즙 보상을 받는 때를 나타내는 손잡이 조작반응 시간들이 서서히 한 곳에 모여 도표상 작은 언덕을 이루기 시작하는 것과 상응한다 (VS : 반옹시간) 진정한 성공은 9000 회 시행 후에 (CIV) 있었다. 이 후로는 원숭이가 매번 짧은 반옹잠복기 (VS) 를 나타내고 운동피질에서의 기록예 큰 반 옹이 기록된다. 그러나 기록 A 와 D 에서의 큰 음전위보다 긴 잠복기 후에 나타난다 . 사사키와 겜바는 c 에 있는 이 후기 음전위를 소뇌로부 터의 홍분성 입력에 의한 것으로 생각하였다 . 이것은 소뇌를 절제하면 소실되고 반옹에 크게 장애를 일으킨다 . 그리고 미리 소뇌를 절제한
동물에서는 관찰되지 않는다. 고로 빛자극으로 손목을 드는 실험의 효 과적인 학습에는 빛에 의한 대뇌 활성이 인지적 학습과 연계되는데 특 히 전두전부의 대뇌 활성이 소뇌로 투사되고 운동피질로 희귀되어야 만 VS IV 에 서 처 럼 잘 학습된 반옹이 보장된다. 똑똑한 원숭이에서조차도 빛신호에 대한 이같이 간단한 동작을 배 우는 데 일주에 1000 번씩 4000 번을 시행하여야 한다는 것은 놀라운 일 이다 이에 비해 어린아이(취학전)는 빛과 손목들기를 연관시키는 문제 에 관하여 쉬지 않고 말함으로써 몇 번의 시행으로 배운다 . 분명히 원 숭이에게 있어 빛에 대하여 손목들기 과업은 너무나도 낯선 일인 것 같댜 그림 7.11 에 100 회 시행 이내에 학습된 것도 있으나 이때의 실험 동작은 원숭이가 주위를 배회하며 정상적으로 경험하는 바와 유사한 것이었다 . 원숭이와 유인원에서 인지적 및 운동 학습을 위한 신경기구가 잘 발달된 것같이 보이나 문제를 언어적으로 생각할 수 없으므로 생소한 상황에 대하여 대처하기가 대단히 힘들다. 그래서 호미니드 진화에 있 어 진화적 성취에는 언어가 지대한 역할을 한 것이다. 현대의 폰지드 와 유사했을 우리의 호미노이드 조상은 인지적 및 운동 학습을 위한 영 역 이 잘 발달된 훌륭한 두뇌 를 가졌던 것 같지 만(표 7.1 , 7.2, 7.3 ) , 그 러나 제 2 장에서 설명했듯이 그들은 대단히 느리게 발전하였다. 결과적 인 호미니드의 큰 성취는 제 4 장에서 언급한 바와 같이 진화적인 언어 의 발달에 기인하는 것으로 생각된다. 그리고 호모속의 선두 주자인 호모 하빌리스에서 그 언어의 발달이 비로소 영향력을 발휘하기 시작 했다고 생각한다 (Tob i as 1966). 학습에 관련되는 대뇌 기구의 상세한 검토를 마무리함에 있어 그림 7 .1 3 이 유용하다. 대뇌의 인지적 기억과 소뇌의 운동기억의 어떤 측면 들을 연합시키고 있기 때문이다. 그림 오른쪽에 평행섬유 입력(그림 7.7) 을 생성하는 태상섬유 입력 (MF) 과 상승섬유 입력 (CF) 의 상호작용
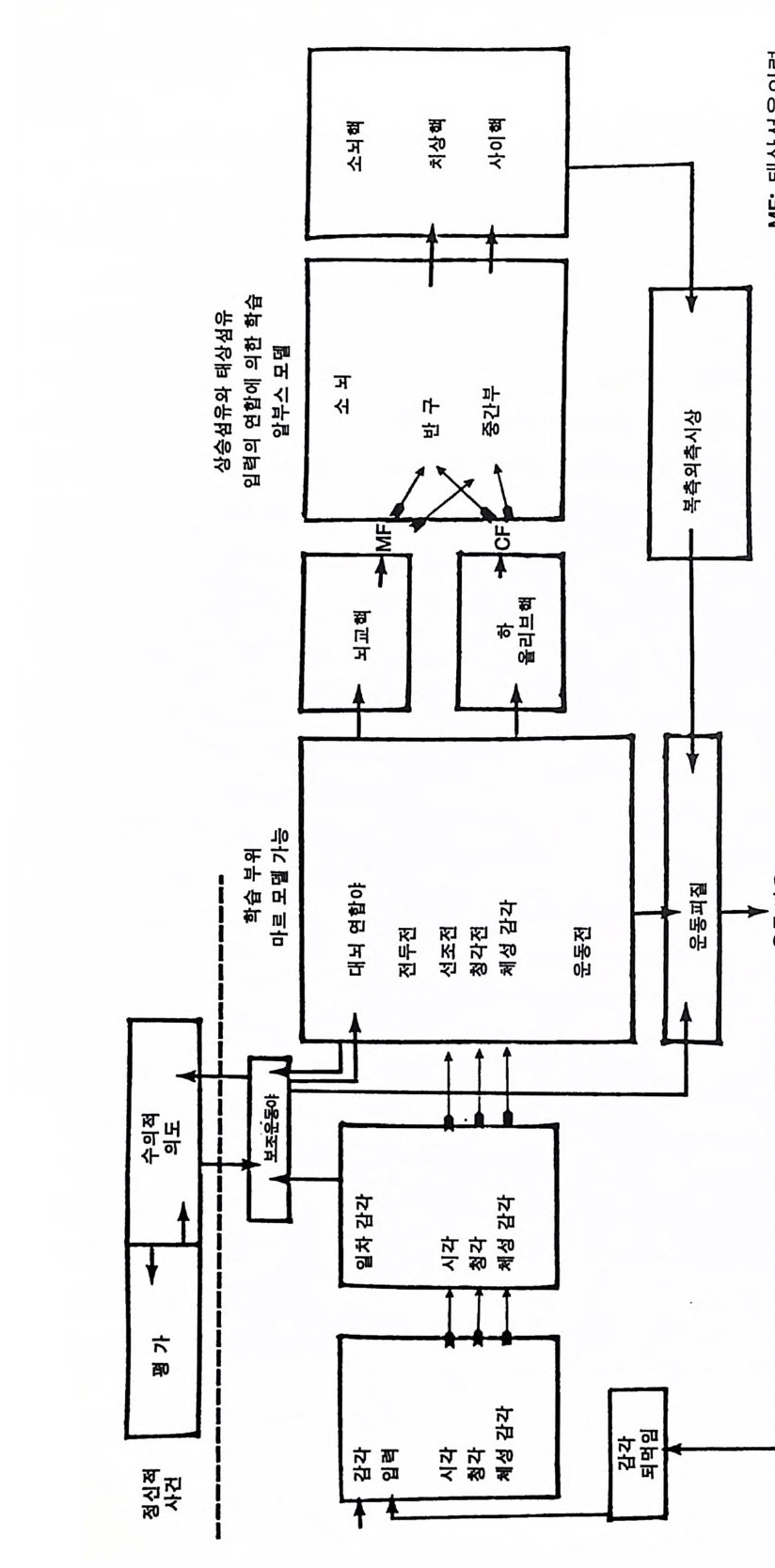
그림 7.13 숙련된 동작의 학습에 관련되는 심적 및 신경적 사건의 가설적 개념 도. 인간의 운동학습에 대한 심적 사건은 점선 위에 보여주는 것으로 신경성 사건과는 분리되어 있다. 동물의 운동 학습에서 심적 사건의 역할은 아직 규명되어 있지 않다. 화살표는 활동의 주경로 방향을 나 타낸다 . 자세한 것은 책에서 설명되어 있다. 모든 경로는 흥분성이고 소뇌핵에서의 소뇌의 억제작용만 예외이다.
을 통하는 학습에 있어 소뇌의 역할을 보여주고 있다 . LTD 에 의한 학 습은 MF 입력에서 일어나고 소뇌핵의 반응 중가로 표현된다. 이는 시 상으로 전달되고 그래서 운동피질로 전달된다(그림 7.1 2 C IV, 그림 7.7).9 ) 그림 7 .1 3 의 왼쪽에 여러 감각 입력이 있는데 이는 대뇌연합야에 투 사될 때 인지적 기억의 기초가 된다(그림 7.5 ). 대뇌피질로 가는 해마로 부터의 입력과 합치되는 것(그림 7.5, 7 . 6) 은 그림 7 .1 3 에는 나타나 있지 않댜 그림 7 .1 3 의 왼쪽 윗부분은 제 8 장을 참조하기 바란다. 7.6 사람 기억의 일부 특별한 양상 인간의 기억의 지속시간을 관찰할 때 우리는 기억 지속을 무한대로 계속시키는 반복적 회상효과롤 검토하여야 한다. 양측 해마 절제 환자 인 HM 이라는 특별한 경우에 수술이 있었던 시점인 1953 년부터 역행 효과를 검사하였다. 1953 년 이 전에 TV 에 흔히 동장했던 인기 인들을 알아볼 수 있는가를 검사하였다. 이 검사로 환자는 해마 절제술을 받 기 1 - 3 년 전의 사건에 대한 역행성 건망증이 있음을 알게 되었다. 양 측성 전기쇼크 치료 후에도 일시적으로 비슷한 기간에 해당하는 역행 성 건망증이 발견되었다 (S quir e 1982, 1983). 장해감도의 기간은 정상적 안 건망증의 시간 경과와 상관관계가 있었다. 고로 1-3 년이라는 기간
은 장기 기억의 공고화와 관계되는 것으로 보이며 이 기간이 지나면 더 이상 정상적 건망증이나 또는 양측성 해마 절제나 전기 쇼크 치료 와 같은 기억장애 과정에 의한 소실을 일으키지 않는다. 기억의 영구 적 공고화가 작용하려 면 1 - 3 년 동안의 소위 회 상기 간 동안 초기 경 험 에 따라 같은 신피질(그림 7.5, 7 . 6) 로 들어가는 해마 입력이 재연되어야 한다는 것을 가상해야 한다. 이 재연이 안 되면 정상적인 기억 소실 과정이 초래된다. 3 년 후에는 대뇌피질의 기억 부호가 훨 씬 안정적으 로 수립되고 더 이상의 해마 입력에 의한 강화를 필요로 하지 않는 것 같댜 따라서 이것들은 양측 해마 절제나 전기 경련 치료로 소실되지 않는다. 포퍼 (Po ppe r 1977) 는 홍미 있는 가설을 제창하였다. 두 가지 종류의 기 억 죽 암시 적 기 억 implicit memory과 명 시 적 기 억 exp lici t memo ry이 있다는 것이다. 암시적 기억은 특별한 기억 사건으로 인식하지 않고도 기억하고 있는 것들이다. 가장 좋은 예는 우리의 언어지식이다 . 흔히 타고난 것으로 생각하고 기억된 지식으로 생각하지 않는다. 마찬가지 로 우리의 모든 동작 즉 걷기, 뛰기 , 자전거타기, 스키타기, 스케이트 타기, 헤엄치기 둥의 수행 능력은 연습으로 개선되기는 하지만 암시적 으로 기억된 것이다. 이들 단어를 글로 쓰는 것도 암시적 기억에 의한 것이다 반대로 명시적 기억이란 특정한 사건, 대상, 얼굴, 이야기, 음 률, 주제 등이댜 명시적 기억은 일반적으로 암시적 기억보다 오래가 지 못하나 반복적으로 재연하면 영구적으로 지속시킬 수 있다.
HM 은 수술 후에 일어나는 모든 사건에 대한 선행성 건망증이 있었 댜 이 환자는 명시적 기억이 없었다• 그러나 암시적 기억으로서의 운 동학습은 계속 발달시킬 수 있었다. 그러나 그 자신은 이를 알지 못했 댜 그 환자는 인지적 학습에 있어 극심한 장애가 있음에도 불구하고 복잡한 운동기법을 습득할 수 있었다. 예를 들면 환자는 정상적인 시 간 내 (3 일)에 거울을 통해 보면서 이중선별표의 평행선 사이에 선그리기를 배웠다 다음날 그 환자는 그가 그 과제를 배웠다는 사실은 회상 하지 못하였으나 그 과제를 수행하는 운동기술은 남아 있었다(Mi lner 1966). 실험 절차의 차이에 따라 밝혀진 기억의 여러 측면들의 다양한 명 칭들을 언급할 필요가 있다. 예를 들어 스콰이어 (S quir e 1983) 는 서술 적 기 억 declarati ve memo ry과 절차적 기 억 pro cedural memory 사 이 의 중 요한 구분을 제시하였다. 건망증 환자에 대한 일반적 기억시험은 과거 사건 또는 형태에 관한 지식의 결함을 알아내도록 설계된 질문에 의해 서술적 기억의 상실을 조사하는 것이다. 한편 절차적 기억은 학습한 기술에 관련되는 것으로 운동 그리고 일부 인지적 기능에 관계된다. 해마가 손상되면 서술적 기억에 대한 기억 상실이 있으나 절차적 기억 에는 장해가 없다 . 이는 아마도 신경회로망의 차이에 기인할 것이다. 여 기 에 는 보조 운동 영 역 the sup plr nenta ry moto r area, 전운동 영 역 the pre - m oto r area, 소뇌 그리고 기저핵이 주로 관여한다 이 분류는 실제 에 있어 여기서 설명한 인지적 및 운동 기억과 동일한 것이다. 기타 용어들도 실험적 검사법에 관계되는 것이다. 즉 사역 및 참조 기억 (Olto n 1983), 연합기억과 인식기억 (G aff an 과 Weis k rantz 1980) 둥이다. 이 장의 시작에서 인지적 기억은 대뇌피질의 자료 은행에 들어있는 것이라고 제안하였다. 기억을 되찾아내는 데 활발하게 관여하는 피질 영역의 지도를 발견할 수 있다 . 그림 6.7, 6.8, 6.9 에 있는 대뇌 활동 지 도는 내경동맥에 133Xe 를 주사하고 뇌에서 방출되는 방사능을 기록하 는 방사성 제논법으로 얻어진 것이다. 그림 7.14 는 특수한 기억과업 수 행중의 대뇌 활동 지도이다 (Roland 와 Fr ibe rg 198. 5). 그림 7 .1 4a 에서 50_3 은 실험대상이 50 에서 3 씩을 순차적으로 빼는 셈을 조용하게 수행하는 동안 통계적으로 유의한 중가를 보인 부위를 표시한다. 죽 기억절차는 50-47-44-41 둥이다 . 2 이하로 계속해서 -1, -4 동으로 이 어 져 약 45 초에 국소 뇌 혈류 (rCBF) 의 측정 이 끝난 후까지
시행하였다. 이 실험의 전체 기간 동안 실험 대상은 주위 환경에 의하 여 방해받지 않도록 하였다. 이후 최초의 방해는 rCBF 기록이 끝난 후 실험자가 실험 대상에게 뺄셈이 어디까지 도달하였는가를 질문한 것 이었댜 두정엽의 각회에서 20.3% 와 14.7% 의 증가를 보인 것을 제외하 고는 양측 전두전엽에서 현저한 rCBF 의 증가를 보안 것은 주목할 만 하댜 이와 더불어 임상적 조건인 계산불능증이 양측 각회를 파괴했을 때 관찰되었다. 그림 7 .1 4b 에
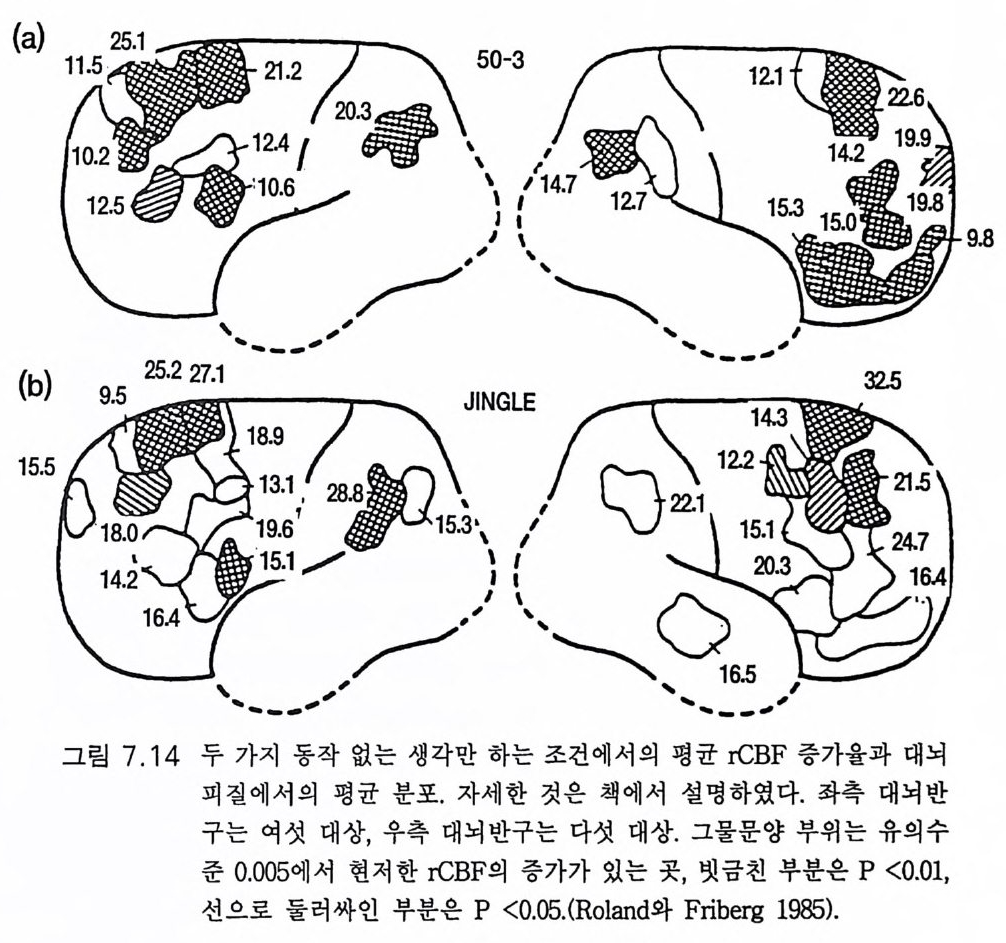 (a) 50-3
(a) 50-3
만 영역 39 와 40( 그림 4.3, 4 .4)이 혼적에 지나지 않는 유인원의 뇌(그림 4.5) 로부터 크게 중대하였다는 것을 살펴보았다. 그리고 이 후기 진화 의 기원은 대단히 늦은 수초화(그림 4.8 , 4.9) 로 미루어 짐작할 수 있다. 현대 유인원을 모델로 한 호미노이드에서부터 전두전엽이 굉장하게 진화적으로 커졌으나 (5 배) 일률적으로 커진 것은 아니라고 말할 수 있
댜 영역 39, 40 처럼 새로운 영역이 특별한 목적을 가지고 발생하여서 전두전엽이 인간의 사고와 기억에 두드러진 역할을 하게 된 것일 수도 있다 이들 영 역 중 일부는 그림 7 .1 4a , b 의 전두전엽 지도에 있는 것일 지도 모른댜 그림 4.8, 4.9 에는 전두전엽의 커다란 후기 수초화 영역을 보여주는 것이다. 이러한 개념들을 마음과 뇌의 상호작용이라는 문제 로 연결하여 제 8 장에서 설명한다.
제 8 장 전화상의 마음과 뇌 문제 8.1 역사적 고찰 그리스 사상에서는 철학의 여명기에서부터 의식이라는 현상에 많은 주의가 기울여져 왔으며 이를 여러 가지로 정의하였다. 애초에 영혼 soul 을 공기, 미립자 또는 불처럼 물질로 생각하였다. 당시에는 물질적 마음과 물질적 신체의 상호작용을 생각하는 데 큰 어려움이 없었다 . 플라톤 Pla t o 과 아리스토텔레스Ari s t o tl e 의 후기 그리스 철학에서는 영 혼을 신체와 상호작용하는 비물질적 본질로 보게 되었지만 초보적인 그리스 과학에서는 아무런 문제도 제기되지 않았다. 당시의 최초의 위 대한 의사 히포크라테스는 이원론자였다. <동작을 할 때 뇌는 의식의 번역사이며 뇌는 사지가 어떻게 움칙일 것인지를 명령하고 또한 뇌는 의식에 대한 전달자이다>라고 그는 말하였다. 그러나 이는 물론 이것 이 어떻게 일어나는가에 대한 초보적 생각에 지나지 않는다. 요약하면 보통 그리스의 철학적 믿음은 이원론적이고 상호작용론적이었다 . 따라서 데카르트가 그 유명한 그의 이원론과 상호작용론을 수립하 기 이전에 많은 선각자들이 있었다. 처음으로 그는 비물질적인 마음
res co git ans 이 물질적 뇌 res ex t ensa 와 어떻게 상호작용할 수 있을까를 설명하려고 시도하였다. 고전적 철학적 표현인 신체-마음 문제에서 신 체를 뇌로 대치한 그의 생각은 옳았다. 그러나 그는 대단히 부적절한 근거에 기초하여 송과체가 인간의 영혼으로 인해 직접 움직여지는 기 관이라고 하는 지나친 추측을 하였다. 동물은 영이 없으므로 자동인형처럼 행동한다고 하였다. 당대의 사 람들은 이 지나친 독선을 받아들이지 않았다 . 이원론자처럼 기회원인 론자 occas i ona li s t들은 마음一신체의 환상적 인 신-조절 병 행론을 채 택하 였다. 스피노자와 라이프니츠도 이 데카르트의 상호작용론을 배척하고 마음-신체 문제를 신-조절 병행론 또는 조화의 다른 운영으로 간주하 려고 하였다. 마음과 뇌의 문제에 관하여 꽤 의미있는 기여가 이루어지기까지는 오랜 휴식기가 있었다. 뇌 과학이 상당히 원시 상태였기 때문일 것이 댜 재미있는 예의 하나는 프랑스의 의사이며 철학자인 라 메트리 La Me ttri e 인데 그는 <뇌는 간이 담즙을 분비하는 것처럼 생각을 분비한 다>고 말하였다. 이 조잡한 뇌 분비설은 세기에 걸쳐 받아들여졌고 후 에 위대한 진화학자인 찰스 다윈은 다음과 같이 썼다. 왜 생각은 뇌의 분비물이어야 하는가? 이는 중력이 물질의 성질인 것 보다 더 신 비 로운 일 이 다 (Gruber 1974, 451). 8.2 비인간 동물의 의식 최근 생물학자와 심리학자들이 행동주의의 오랜 암흑기에서 깨어났 댜 그 암흑기에는 동물의 의식에 대하여 의문을 제기하는 것은 과학 적으로 적당치 못하다고 생각하였다. 다행히도 이 어둠온 예를 들어
도르프 (Tho rp e 1974), 로렌츠 (Lorenz 1977) 그리고 그리핀 (G riffi n 1976, 1984) 의 글에 의해 벗겨지게 되었다. 특히 그리핀(1 976) 은 인성학의 문제에 심적 경험의 개념을 도입함으 로써 중요한 발전을 이룩하였다. 동물 행동의 복잡성은 사람 행동의 경우와 공통점이 대단히 많은 것같이 보이기 때문에 동물에게 있어서 도 연관된 심적 경험이 있다고 추론하는 것이 적절한데 그는 이룰 다 음과 같이 정의하였다 . 정상적인 모든 사람은 감각의 직접적인 흐름에서 시간적, 공간적으로 멀리 떨어진 사물과 사건에 관하여 생각한다. 이것이 바로 내가 심적 경 험이라고 하는 것이다 . 심적 영상의 존재 그리고 동물이 행동을 조절하기 위해서 그것을 사용한다는 것은 의식에 대한 실용적 작업 정의를 제공한 다 (1976: 5). 그러나 그는 <의식이라는 용어를 행동주의자들은 과학적 분석에 유 용한 개념이라고 강력히 주장하고 있다>고 경고하였다. 동물이 가질 수도 있는 어떤 심적 경험도 고려하지 않고 동물 행동 울 연구하는 심 리 학자 또는 생물학자들을 총괄적 행동주의자in clus i ve behav i o ri s t라고 부르자고 그리 핀 (1984) 은 제 안하였다. 단지 동물이 무 엇을 행동하였는가만이 동물의 생존과 번식에 대하여 효과를 가질 수 있고 따라서 어떤 경험도 진화적 결과를 가질 수 없을 것이라고 그돌 은 암시적으로 생각한다. 그러나 그리핀은 이에 대하여 반론을 제기한 다. 만일 동물의 심적 경험이 동물의 안녕과 생식에 영향을 미친다면 자연도태에도 효과가 있다는 것이다. 호미니드 진화에 대한 이것의 많 은 예는 제 4, 5, 7 장에서 기술하였다. 동물이 표면상 느낌이나 감정에 의하여 동작했을 때 그리고 동물이 과거의 경험에 비추어 현재의 상황을 평가할 능력이 있을 때 그래서
판에 박힌 본능적 반응이 아닌 행동의 적절한 과정에 도달할 수 있을 때 동물은 의식이 있다고 말할 수 있다. 이 같은 방식으로 원래의 행 동양식이 표현될 수 있다. 행동양식은 학습될 수 있는 것이며 정서적 반응을 풍요롭게 하는 것이다 . 좋은 예를 들면 유인원을 폐쇄된 방에 두고 한쪽 구석에 움직일 수 있는 상자를 놓고 다른 쪽 구석에 바나나 다발을 놓아두는데 손이 닿지 못하는 높이에 매어단다. 유인원은 장시 간의 생각 끝에 상자를 바나나 밑에 옮겨놓고 따먹는 데 성공한다. 의 식적 경험을 증명하는 또 다른 방법은 포유류 특히 어린 동물들의 자 발적 유희 이 다(제 7 장). 8.3 의식의 진화 이 책의 목적이 동물 진화에 있어서 의식의 기원에 관하여 자세하 게 연구하고자 하는 것은 아니다. 그러나 이 문제에 관하여 언급하기 위해서는 최소한의 몇 가지 의문이 제기될 수 있다. 어떻게 종래에는 생각이 없던 세계에 마음 또는 동물의 의식이라는 것이 진화 과정에서 존재를 나타내게 되었는가? 무의식으로 충만된 이제까지의 어둠으로 부터 처음으로 나타난 심적 경험의 섬광인 심적 사건의 경험은 어떻게 동물의 진화적 발달 초기에 나타나게 되었을까? 그 같은 심리적 사건 들은 틀림없이 진행중인 뇌 신경 활동과 관계가 있는 것이다. 그러나 어떻게 뇌의 기능이 이들 심적 사건들에 의하여 향상되었는가? 그리 고 어떻게 이들 심리적 사건들이 진화과정에서의 생존에 진가를 발휘 하게 되는가? 끝으로 이제까지 일원론적으로 물리적 세계, 죽 물질과 에너지의 세계라고 간주하였던 세계에 갑자기 둥장한 심리적 사건들 을 어떤 존재론적 위상으로 평가할 수 있는가? 만일 신경심리학자가 단순한 유기체-꿀벌이라도 좋다-~의 행
동을 연구한다면 가장 복잡한 행동일지라도 학습으로 가미된 타고난 본능의 개념으로 그럴싸하게 설명할 수 있다. 동물의 본능적 행위는 신경계의 개체발생적 구조에 기초하고 유전적 정보에 의한 구조에 관 계된다. 그리고 학습이란 사용합으로써 증가된 접합부 효과일 수 있다 (제 7 장). 고로 우리는 유물론적 질서 내에 머무르면서 이들 동물 행동 들을 설명할 수 있다. 지금까지 새나 포유류 이하의 동물 행동에 관하 여 가장 많이 연구된 것은 꿀벌에 관한 것이다 . 그러나 나는 시간과 공간의 양식으로 정교하게 작성된 부호를 나타내는 꿀벌의 무도양상 에 근거한 그리핀의 유심론적 가정을 인정할 수 없다. 꿀벌이 자신이 하는 일을 알고 있다고 가정할 아무런 이유도 없다. 그 뇌는 너무나 작고 또 구조상 판에 박힌 것 이 다 (S tr aus f el t 1976). 양서류의 수준에서조차 개구리의 효과적인 파리 잡기를 시각적 인 식(벌레 탐지)과 그에 대한 반응이라는 간단한 말로 설명할 수 있다. 로 렌츠 (Lorenz 1971, 1977) 는 조류에 관한 그의 방대한 연구에서 정신 상 태를 포함하는 행동양상을 기술하였다 . 도르프 (Tho rpe 1974: 299) 는 조 류의 숫자 인식 실험으로 결론짓기를 <사람의 어린아이가 의식적 뇌 작용에 의해서만 수행할 수 있는 숫자 개념의 추상적 사고를 동물들이 수행할 수 있다는 대단히 강력한 증거를 얻었다>고 하였다. 의식의 계통발생학적 문제의 범위는 조류와 포유류인 것같이 생각 된다. 가장 간단한 전략은 비인간 고동동물인 유인원의 행위에 있어서 의 의식을 연구하는 것이다. 이는 제 4, 5, 6, 7 장에서 다루었다. 가축_―개, 고양이, 말――의 심적 경험에 대하여는 의심할 바가 없댜 어린 동물의 놀이는 의식의 믿을 만한 기준이 된다고 생각한다. 또한 호기심 그리고 감정의 표현, 특히 깊은 애정의 증거도 기준이 된 댜 그러나 이들 가상적인 정신 상태를 인간들이 경험하는 것과 동일 시하는 것은 주의해야 한다. 사람들 사이에서 이루어질 수 있는 정도 로 사람과 동물 사이에 부호에 의한 의사 소통이 이루어지지 못하기
때문이다. 다음 질문은 대뇌 작용에 관련되는 이 심적 경험의 출현으로 어떤 긍정적 발전이 있었는가 하는 것이다. 예를 들어 윌리엄 제임스 Wi lliam J ames 는 마음이 란 그 자신을 주체하지 못할 만큼 너무 복잡하 게 발달한 두뇌가 획득한 특성이라고 제안하였다. 의식은 동물에게 일 종의 전체론적 경험을 주는 데에 가치가 있다고 주장되어 왔다. 이 주 장은 시각적 경험이라는 관점에서 더욱 발전하였다. 지난 20 년 동안 고양이와 원숭이를 이용하여 뇌에서의 시각 정보 처 리에 관한 방대한 과학적 연구가 있었다 (6 .4 , 그림 6.2-6.6 ). 이 단계적 처리 과정에는 망막 위에 영상으로 존재하는 원래 그림 모양으로부터 점진적인 추상과정이 있다(그림 6.1 ). 신경성 정보 처리의 어떤 단계에 서도 그림을 종국적으로 재구성함에 있어 도움이 될 것으로 생각되는 신경세포가 발견되지 않는다. 예를 들어 할머니를 보고 있다고 말해주 는 신비한 <할머니 세포>와 같이 각각의 세포가 그 자신 안에 일부 특수한 그림을 지닌다고 생각되는 것이 발견되지 않는다. 그래도 우리 는 영상을 감지할 수 있다. 이 같은 설명에 대한 부분적 예외가 있는 데 그림 6.6 의 TE 영역에 있는 희귀한 모양을 탐지하는 신경세포가 그 것이댜 신경세포의 틀에 박힌 활성의 방대한 다양성은 그림의 재구성 에 이용될 수 있을 것으로 생각되는 부호화된 정보를 가지고 있다. 그 러나 그같은 전체론적 운영은 대뇌피질의 기구에 의하여 이루어질 수 있는 것으로 생각되지 않는다 (We i skran tz 1974). 그러나 이는 의식적 경험으로 이루어지고 우리가 눈을 떴을 때 불가사의한 방법으로 나타 난다 그리고 그것은 시각 입력과 명백히 동시발생적으로 순간순간 변 한다. 시각피질과 그 이후 신경영역에서 일어나는 복잡한 신경 처리 기구의 운영 과정은 대뇌피질 내에 시공적 양상으로 표현되는 신경 활 성인 부호적 정보를 다룬다 (Crue tzf eld t 1087). 진화 과정에서 의식적 심 적 경험의 등장은 시각 처리 기전의 진화와 발맞추어 발생했고 이것은
동물의 행동을 조절하는 데에 필수적인 것이었다고 추측할 수 있다. 동물의 간단한 행동을 유도하는 간단한 시각 입력은 전체적인 시각 영상에 통합될 필요는 없을 것이다. 예를 들어 위에서 언급했던 개구 리의 시각 계통은 통합적 운영이 아니고도 기능할 것이다. 그러나 조 류, 포유류 등 고등동물의 대단히 개선된 시각 계통에서는 통합적 영 상을 얻는다는 것이 자연도태에 있어 큰 장점이 되었을 것이다 . 또한 이 통합은 청각 , 후각, 촉각 등의 감각 입력도 포함할 수 있어서 우리 가 즐기는 것 같은 통일된 심적 경험을 이루게 한다(제 9, 10 장) . 따라서 진화상의 심적 경험의 출현은 고도로 발달한 동물의 뇌로 들어오는 광범하고 다양한 입력의 통합을 이루기 위한 것이라고 이해 될 수 있다는 가설이 수립되었다. 간단한 신경계통을 가지고 보다 제 한된 감각 입력과 행동적 출력을 가진 동물은 중추신경계에 의하여 이 루어지는 것 같은 통합을 필요로 하지 않는다. 물론 아 가설은 이전까 지는 순전히 물리적이었던 세계에서 그것에 속한 것의 심적 경험의 불 가사의한 진화적 출현을 설명하지는 못한다. 단지 이 같은 출현이 어 떤 진화적 장점이 되었는가를 암시할 뿐이다. 오늘날 다윈적 진화론은 이전까지는 총체적으로 포괄적이었던 물질 _에너지 세계와는 또 다른 세계에 있는 비물질류의 심적 경험을 생명 체가 얻게 됨으로써 제기되는 범상치 않은 문제를 인정하지 않았다는 점에서 결함이 있다 . 데카르트의 해답도 일반적으로 타당한 것은 아니 다. 즉 그는 인간은 의식적 경험을 갖는데 그 경험은 영의 신성한 창 조에 기인하고 고등동물은 심적 경험이 결여된 기계 같은 자동인형일 뿐이다라고 주장했다. 마찬가지로 전심론자의 문제 얼버무리기도 타당 한 것 은 아니 다 (Po pper와 Eccles l
과 진화』 또는 모노 (Monad 1971) 의 『우연과 필연』 또는 윌슨 (W il son 1975) 의 『사회생물학: 새로운 합성』에서 심성의 진화에 대한 언급이 일체 없다 . 그리핀 (G riffi n 1976) 이 그의 저술 『동물 의식성의 문제』에 서 기술한 것처럼 생물학자들의 의견의 분위기는 행동주의자들의 독 단에 의해 지배되어 왔다고 설명할 수 있을 것이다. 그러나 최소한 고 동동물에 있어 동물의 의식성은 이제는 인정되어야 하고 따라서 진화 론자들에게 큰 도전이 제기되는 것이다 . 문제를 무시하는 것이 문제를 해소하는 길이 아니라고 말할 수 있는 단계에 우리는 이미 와 있는 것 이댜 포퍼 (Po pp er 1982: 150) 는 다음과 같이 말하였다. < 동물계에 의식의 출현은 아마도 생명 그 자체의 기원이나 마찬가지로 커다란 불가사의 일 것이댜 그러나 도저히 이해할 수 없다 할지라도 우리는 이것 또한 진화, 즉 자연도태의 산물이라고 생각해야 할 것이다> 의식의 출현은 정통파 진화론의 꺼림직한 집안의 비밀이라고 믿는 다. 동시에 총체적 개념이 의식의 출현에 의미를 부여하기는 하지만 이 출현을 설명할 수 없음을 인정해야 한다. 의식의 출현은 정통파 진 화론자가 이를 극단적인 유물론적 세계에 있는 극단적 자연현상으로 간주하는 한 오직 수수께끼로 남아있을 수밖에 없다. 8.4 마음과 뇌 문제의 철학 앞서 제 4 장과 그림 4.2 에서 설명했듯이 포퍼 (Po ppe r 1972) 는 제 3 세 계, 문명과 문화 세계의 추가를 제안하였다. 전통적 두 개의 세계는 우 주의 물질-에너지 세계(세계 1) 와 주관적 경험과 의식상태의 세계(세계 2) 이다. 세계 l 과 세계 2 의 기초 위에서는 현재 통용되고 있는 뇌와 마 음에 관한 여러 학설을 일반적인 용어로 설명할 수 있다(그림 8.1 ).
급진적 유물론 : 세계 1 = 세계 lp; lM = 0; 세계 2 = 0 전심론 : 모든 것은 세계 1-2 . 세계 2 는 독립적으로 존재하지 않음 . 부가현상: 세계 1 = 세계 1p + 세계 lM 세계 lN I 一 세계 2 동일론: 세계 1 = 세계 1P + 세계 lM 세계 lM = 세계 2( 자동 自同) 이중 상호작용설: 세계 1 = 세계 1P + 세계 lM 세계 lM ;=t 세계 2; 이 상호작용은 뇌의 중개로 일어난 다. LB = 세계 lM 고로 세 계 1 = 세 계 1P + 세 계 1LB, 세 계 lLB =2 세 계 2 그림 8.1 뇌와 마음의 관계에 관한 여러가지 학설들의 개념도. 자세한 설명은 책에 있다 . 세계 l 은 물질 또는 뇌를 포함하는 물리적 세계의 모든 것 세계 2 는 모든 주관적 또는 심적 경험 세계 l p는 심적 상태를 가지지 않는 모든 물질세계이고 세계 lM 는 심적 상태가 관련되는 물질세계의 작은 분획이다. 위의 8.2 와 8.3 에서 언급했듯이 고집불통의 급진적 유물론자를 제외 하고는 사고과정과 같은 심적 사건에 있어서 의식의 존재에 일반적으 로 동의한다. 물론 생각한다는 것은 주관적인 경험이고 우리를 둘러싸 고 있는 세계에서 감각을 통하여 감지하는 것 같은 방식으로 객관적인 식별이 되지는 않는다. 다른 사람에게 생각하고 있는 것을 이야기하여 확인시킴으로써 경험의 객관성을 부여한다 . 이 기준으로 환각 또는 망 상 둥의 경험을 교차 검사함으로써 조사하였다. 이는 그것을 느끼는 사람에게는 세계 l 의 것이며 객관적인 것으로 해석되지만 실제로는 전적으로 세계 2 에 속한다는 것이 확인된다. 예를 들면 꿈 경험의 표 면상 객관적 세계는 실제에 있어 제 1 세계 상태를 포함하지 않고 전적 으로 세계 2 에 속한다. 물론 꿈과 동시에 객관적 제 1 세계 사건이 뇌에
서 감지될 수 있고 꿈꾸는 사람의 신체로도 감지될 수 있다. 그러나 각성시에 이야기하는 꿈의 내용과 아무 연관 증거가 없다. 마음에 관한 여러 유물론적 학설이 있다 . 그림 8 .1의 처음 네 가지 항목에 정리되어 있다. 급진적 유물론은 이 자체를 제외시킨다. 세 가 지 다른 유물론적 학설은 심적 사건 또는 마음의 존재를 인정한다. 그 러나 그것에 독립적 상태를 부여하지는 않는다 . 마음에 관한 위의 세 가지 유물론적 학설에 따르면 심적 상태는 물질 또는 물리적 세계, 죽 모든 물질(범심론p anp s y c hi sm 의 경우) 혹은 고동 동물과 사람의 고도로 조직화된 신경조직의 형태로 특별히 존재하는 특정한 상태의 물질의 소산이댜 이것의 한 가지 변이는 부가현상설 e pip henomen ali sm 로서 더 자세하게 설명할 필요는 없는데 이는 파이글 (Fe ig! 1967) 이 개발하였던 동일론i den tity th eo ry에 의하여 대치되었다. 포퍼는 다음과 같이 주장 하였다. 네 가지 주장은 모두 물리적 세계(세계 l) 는 폐쇄적 또는 자체 수용적 이라고 주장한다. 물리적 세계 1 의 폐쇄성의 이 물리주의적 원칙은……물 리주의 또는 유물론의 특징적 원칙으로서 결정적 중요성을 가진다…… (1977: 51) 심적 사건과 동일하다고 생각할 수 있는 정확한 신경성 사건과 관 계되는 철학적 논증을 발견하기는 힘들다. 그러나 파이글 Fe ig l 은 다음 과 같이 분명하게 표현하였다. 내가 분명히 하고 변호하고자 하는 동일론에 의하면 의식상태에 있는 인간이 <살아가면서 느끼는> 직접 경험의 상태나 우리가 고등 동물도 가진다고 분명히 주장할 수 있는 직접 경험의 상태들은 이들 유기체들의 신경조직의 특정한(아마도 구조적인) 특성들과…… 특히 대뇌피질의 작
용과정과 동일한 것이다. 신경생리학적 개념으로는 신경 방출의 복잡하 고 고도로 분지 된 양상을 뜻한다 (1967: 79). 이들 유물론 학설 또는 병행론 학설에 대조되는 것이 이중론적-상호 작용론적 학설이고 그림 8.1 의 하단에 표시되어 있다. 이들 학설의 근 본적인 특성은 마음과 뇌는 속성이 별개라는 것이다. 뇌는 세계 1 에 속 하고 마음은 세계 2 에 속하여 이들은 모종의 방법으로 상호작용한다는 것이댜 이는 뒤에 설명한댜 고로 세계 1 과 2 사이에는 선두가 있고 이 선두를 교차하여 양쪽 방향으로 상호작용한다(그림 4.2 , 8.5 ). 이는 정보 의 흐름이지 에너지의 흐름은 아니라고 생각할 수 있는 것이다 . 따라서 고전물리학의 기본 원리인 에너지 불변의 법칙인 물질-에너지의 세계 (세계 l) 가 완전하게 인정된 것이 아니라고 하는 상궤에서 벗어난 생각 울 갖게 된다. 반면 이미 살펴보았듯이 마음에 관한 모든 유물론적 가 설들은 세계 1 의 폐쇄성을 대단히 교묘하게 고수하고 있다. 8.5 마음과 뇌 문제의 실험적 검사법 고동동물 특히 영장류는 의식을 가졌다고 생각할 수 있으므로 이 책에서 특별히 관심을 가지는 문제는 호미니드 진화에서 뇌가 크게 발 달하는 동안에 마음-뇌의 상호작용에는 어떤 변화가 일어났는가에 있 다(제 2-7 장). 유인원에 관한 과학적 연구는 결여되어 있으나 원숭이나 바분은 호미노이드 조상의 모델로 받아들여지고 있다. 시각, 청각, 촉각 등 조절된 감각 입력의 실험 조건에서 고동 포유류 의 의식하 경험을 철저하게 연구하는 것은 가능하지 않다. 그러나 몇 가지 일화적 중거는 있어 이는 제 4-7 장에서 설명하였다. 그들은 자신 의 경험을 표현하는 언어 능력이 결여되어 있다. 그러나 물론 언어의
두 가지 하위수준을 이용하여 이들 경험을 신호로 나타낸다(그림 4.1 ). 이에 반해 고등 영장류의 뇌는 촉발 감각 입력과 독립적으로 동물에 의해 수행되는 수의적 결정이 있을 때 연구될 수 있고 이는 뇌에 대한 마음의 작용에 관한 것이다 . 그림 8.2 는 마카크 원숭이룰 훈련시키는 실험장치로서 (B ri nkman 과 Port er 1979) 음식의 보상을 원하면 손잡이 (L) 를 표적 지 역까지 잡아당 기게 훈련하는 것이다 . 한 가지 행동 제한은 머리를 고정한 것이다. 아
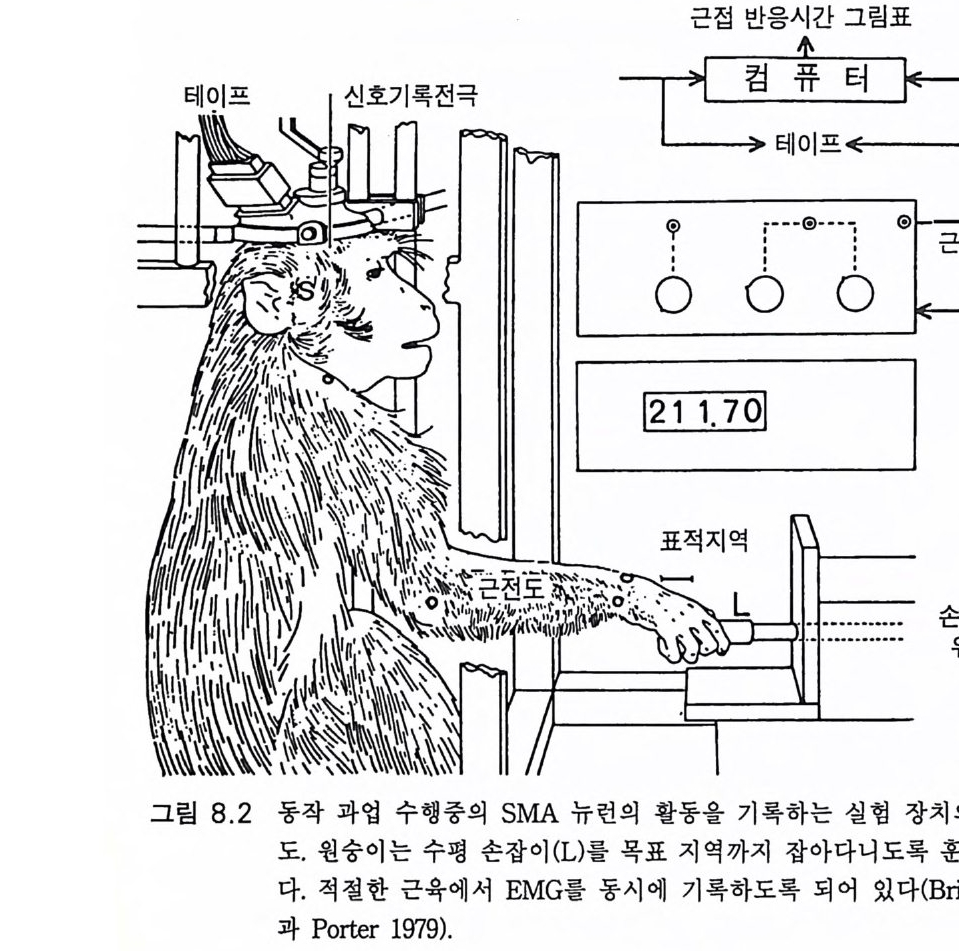 근접 반응시간그림표
근접 반응시간그림표
무 때나 원하면 양쪽 손 어느 것으로나 손잡이를 자유롭게 잡아당길 수 있게 하였다. 촉발신호는 주지 않았다. 동작의 수의적 개시에 있어 서 뇌의 보조 운동 영역 (SMA) 이 일차적으로 관여한다는 많은 증거가 있었다(그림 3.3, 4.3 ). 실험 전에 마취하에서 뇌의 SMA 영역을 노출시 키고 부착물을 두개골에 고정하였다. 부착물을 통하여 실험중에 미세 전극을 SMA 내에 삽입하여 단일신경세포의 반응을 기록할 수 있도록 하였댜 만일 SMA 가 동작의 수의적 개시에 관계된다면 (Eccles 1982b) 결과 적으로 동작을 일으키게 하는 운동피질신경에 앞서서 여러 신경세포 들의 임펄스 발사가 예상된다. 그림 8 . 3a 에 SMA 신경세포가 동작을 일으키는 근육의 활동전압(그림 8.3b) 보다 약 250msec 전에 임펄스 발 사가 시작되는 것을 볼 수 있다. 이 같은 조기 개시는 다른 쪽 손을 사 용할 때도 볼 수 있다(그림 8.3 c ). SMA 뉴런의 94% 가 편측성 과업 에 대하여 양측성 반응을 나타내었다 . 동작은 자체적으로 조율되었고 대 개 성공적으로 손잡이를 잡아다녀 음식 보상을 받았다. 앞팔의 동작을 일으키는 데에 있어 운동 추상뉴런의 임펄스 발사는 추체로(그림 3.6 ) 를 따라 내 려 가고 대 개 근육 반응을 신호하는 근전도(E MG) 보다 40-60 msec 앞서서 시작되었다 . 가장 빠른 것은 140-120msec 앞선 것이었다 (Ev a rts 1972). 따라서 그림 8. 3 의 SMA 뉴런은 운동추상세포의 가장 빠 론 발사보다 어림잡아 약 lOOmsec 앞서서 임펄스를 발사한 것이다. 이 같은 SMA 뉴런의 조기 방출은 흔히 발견되는 소견이다. 따라서 그림 8. 3 은 어떻게 원숭이가 심적 의도에 의하여 양측에 있는 SMA 의 많은 뉴런들을 활성화할 수 있는가를 보여주는 것이다. 그림 3 .1 3 에서 볼 수 있듯이 다른 측부 작용도 효과적 심적 의도(또는 아이디어)에서 일어난 댜 그러나 직접적 계열은 SMA 뉴런의 활성화이고 이는 결국 운동피 질의 추상세포(운동 EX) 를 홍분시키고 아마도 운동전피질(연합 EX) 의 중개에 의하여 신경 방출이 추체로를 따라 내려가서(그림 3.6)
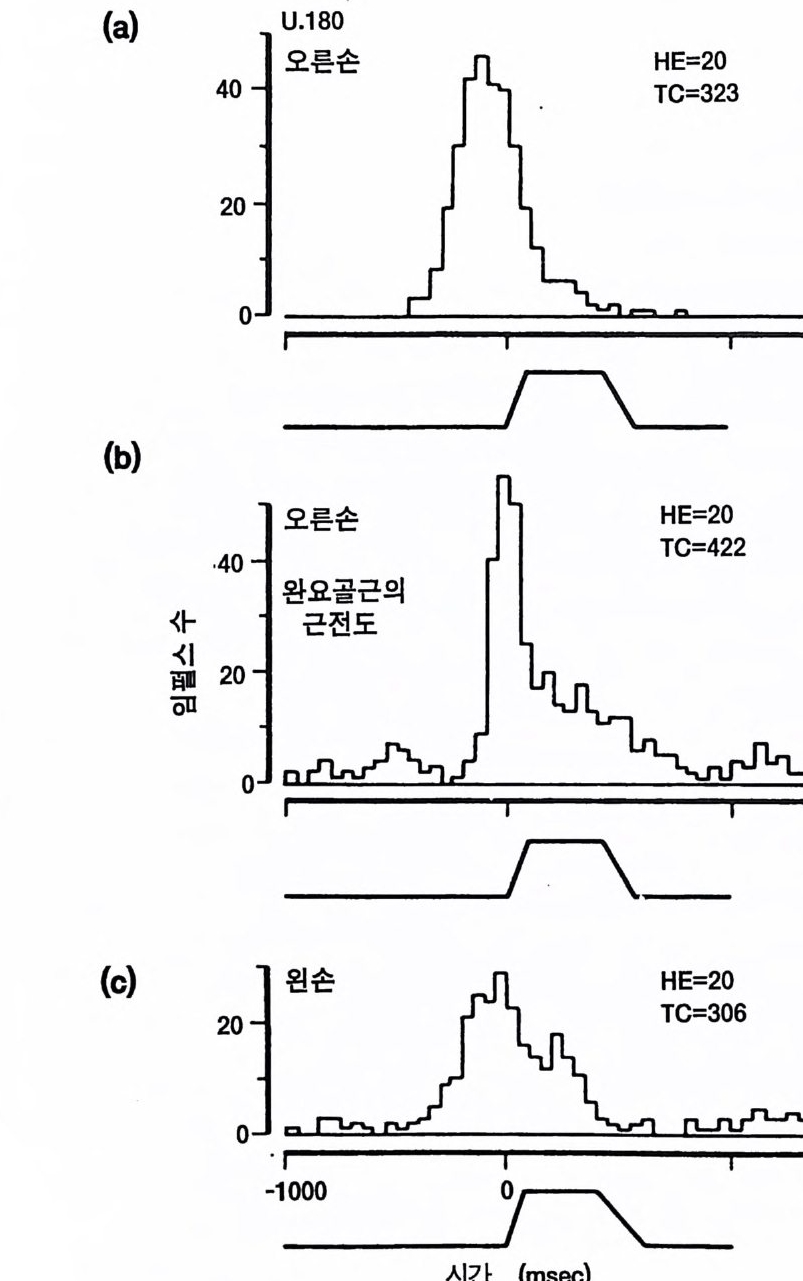 (a) U.180
(a) U.180
그림 8.3 오른손 (a) 과 왼손 (c) 으로 손잡이룰 잡아다닐 때 팔꿈치 굽힘에 관계되 는 뉴런에서의 방전 양상. (b) 는 대표적 팔꿈치 굴근 상완요골근의 EMG 를 나타내는 주위 반옹시간의 막대그립. (a) 에서와 같이 오른팔 로 20 번 잡아다닐 때의 것으로 EMG 활동의 중가 전에 뉴런의 방전 증가가 잘 나타나 있다. 이것은 대부분의 뉴런에서 방전 양상과 EMG 변화를 비교할 수 있게 하는 것이다 .(HE 잡아다닌 횟수, TC:20 번 잡 아다닐 때 의 임 펄스 수 )(B rinkm an 과 Port er 1'57 9 ).
손잡이 를 잡아다니 는 근육이 수축하게 된다 . 브링 크만과 포터 (Br in- kman 과 Pott er 1983) 는 수의적 의도가 어떻게 광범한 SMA 뉴런의 반 옹을 개시하고 손잡이를 잡아다니는 복잡한 동작을 이루도록 운동전 피질이 반응하는가를 설명하였다 .10) 윤리적인 문제 때문에 그림 8 . 3 에서와 같은 실험을 사람의 SMA 단 일 뉴런에 관하여 연구하는 것은 불가능하다 . 그러나 SMA 를 포함하 여 사람의 대뇌피질 여러 부위의 활성은 그림 6.7, 6.8, 6.9, 및 7 .1 4 에 보여준 방사성 물질 추적법으로 연구할 수 있다. 수의적 동작은 엄지 로 손가락을 차례로 촉지하는 것을 학습시킨 것이고 검사하는 40 초 동 안 주의를 집중하도록 하였다. 이렇게 해서 자동적이 아닌 수의적 동 작을 정성화하였다(R oland 둥 1980). 그림 8 .4 a 는 운동-서열 시험을 시행하는 동안 국소 대뇌혈류 (rCBF) 가 현저하게 증가한 부위와 평균 증가 백분율을 보여주는 지도이다. 그림 8 .4 a 의 좌측은 운동一서열 시험을 오른손으로 시행했을 때의 좌측 대뇌반구의 rCBF 를 보여주는 지도이다. 우측에 있는 것은 왼손 동작 때의 우측 대뇌반구 지도이다. 예상대로 각 대뇌반구에 있어 활동하는 손의 반대측 운동피질 영역에 현저한 rCBF 의 증가가 있었다. 뒤쪽으 로 감각운동 영역과 손감각 영역에는 rCBF 의 증가가 적었다. 특히 홍 미로운 것은 한쪽 손으로만 시험을 시행했을 때 양측 대뇌반구의 SMA 전체에 rCBF 의 현저한 증가를 보인 것이다. 예상한 대로 동측의
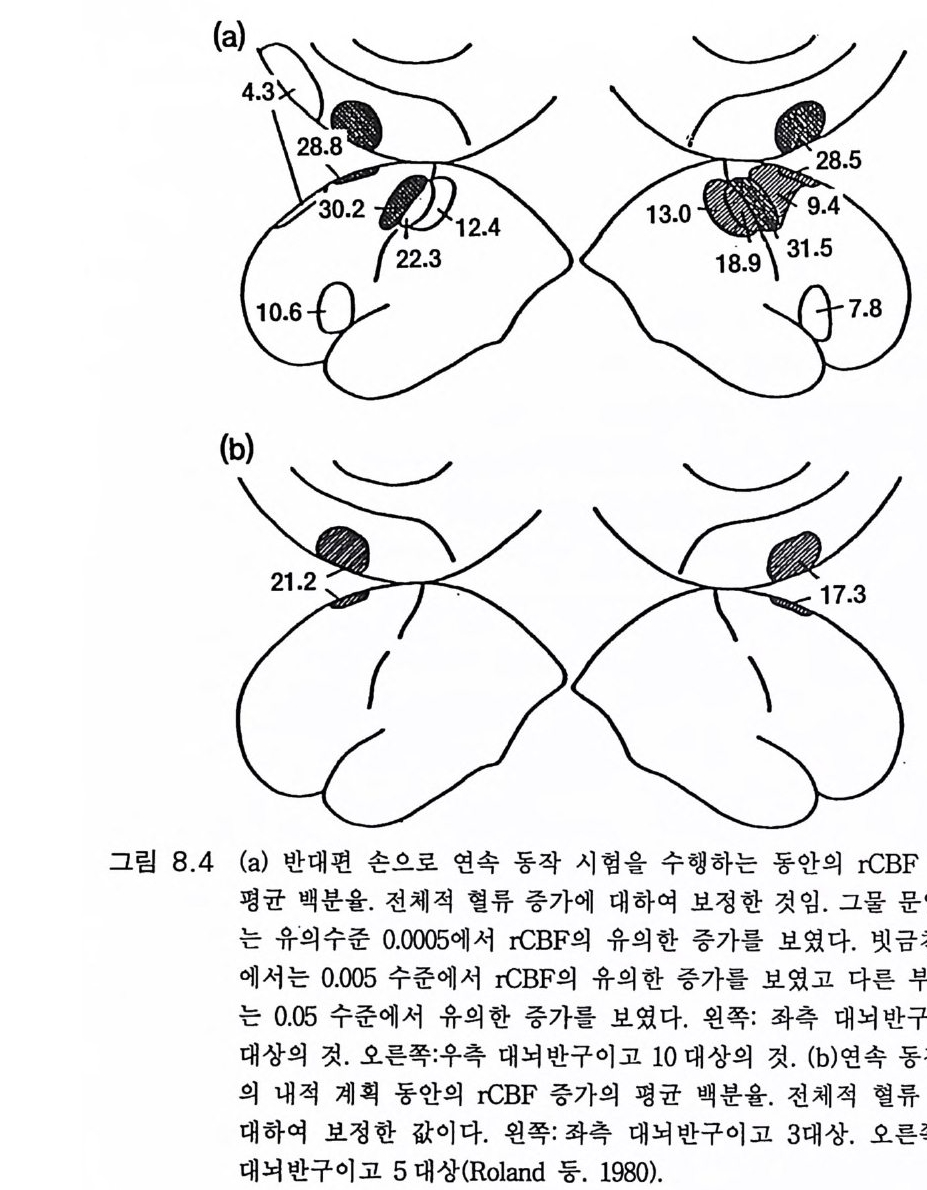 (a)
(a)
운동 및 감각운동 영역에는 rCBF 의 중가가 없었다. SMA 의 양측성 활 성은 원숭이에서도 보였다(그림 8.3a,c). 가장 홍미 있는 것은 그림 8 .4 b 로서 실험 대상은 운동-서열 시험의
< 내부 계획i n t ernal p ro g r ammi n g>이라고 하는 것에 집중하였다. 죽 동 작을 수행하기 전에 마음속으로 동작의 순서를 연습하는 것이다. 놀랍 게도 동작이 있을 때보다는 적었지만 SMA 에 걸쳐 rCBF 의 상당한 증 가를 보였다. 대뇌피질의 다른 어떤 부위에서도 현저한 증가를 보이지 않았댜 조용히 생각만 했는데도 rCBF 의 현저한 증가를 보였다는 점에 서 그림 8 .4 b 는 그림 6.9 및 7 .1 4 와 비 교가 된다 . 그러 나 아마도 심 적 과 업은 대단히 복잡한 것이기 때문에 많은 영역에 걸친 증가도 있었다. 그림 6.9, 7.1 4 , 8.4 및 10.3 에 추가하여 집중 사고의 대뇌피질 활성 유 발 효과는 데스메트와 로버트슨(D esmed t와 Robert so n l
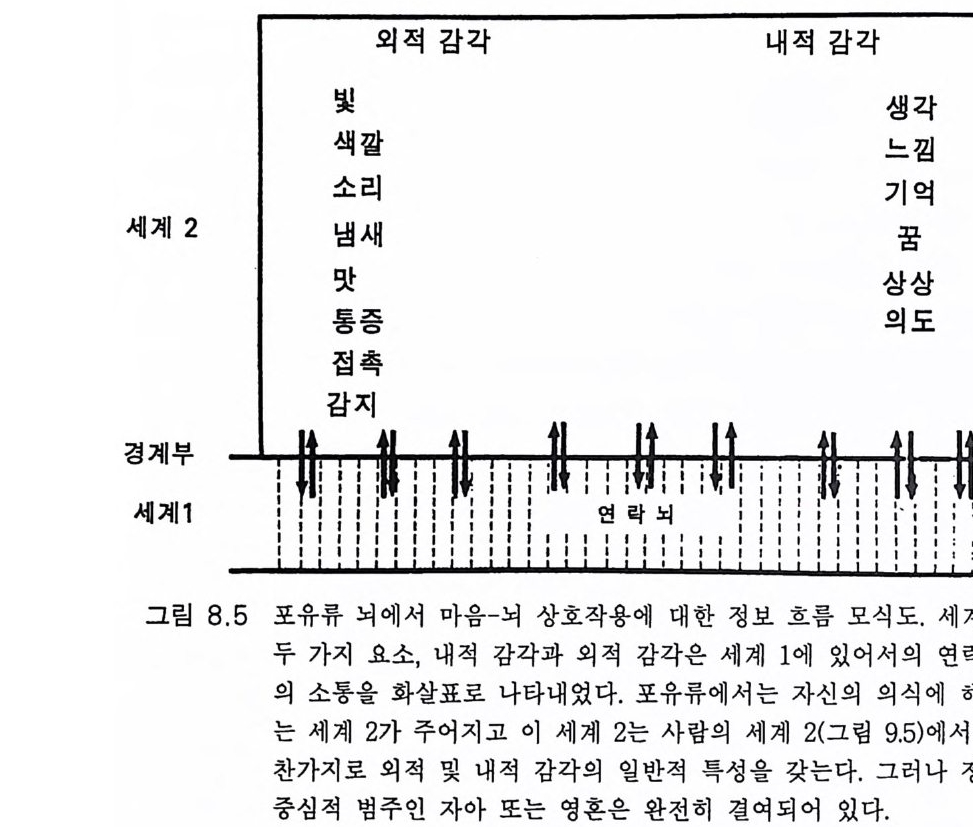 외적 감각 내적 감각
외적 감각 내적 감각
야 한다. 이것은 호미니드 진화에서 더욱 발달했을 것이다(그림 6.1 0 , 6.11). 고등 동물은 심적 경험을 가졌고 이들 세계 2 의 사건은 효과적으로 대뇌피질의 세계 1 사건에 작용한다는 것에 관하여는 8.2, 8.3, 그리고 8. 5 에서 논의했으므로 이 주목할 만한 상황의 도식화를 시도해 볼 수 있다. 그림 8.5 는 이원론-상호작용론에 의하여 필연적으로 짜여진 것 이다. 여기서 위에 있는 세계 2 는 여러 외적 감각과 내적 감각의 관여 룰 나타내고 있고 아래쪽의 대뇌피질 분절(세계 l) 은 수직 점선으로 나
타낸 연락뇌 L iai son Brai n 모듈이다(그림 3.4 및 7.6 참조). 일차적 운동 및 감각 영역은 마음과 직접적인 연락이 없다는 실험적인 증거가 있다 (R oland 와 Fri be rg 1985, 그림 12). 화살표는 경계면을 가로지르는 상호 작용을 나타낸다. 세계 2 를 연락뇌 위에 그려넣은 도식적인 편리성은 있으나 이것의 위치가 주어져야 한다면 이것은 피질 내에 있을 것이고 양 방향 화살표의 시작과 끝에 의하여 보여주는 것이다. 그림 8 . 5 로서 예상되듯이 고등동물의 모든 대뇌피질을 제거하면 이 동물들은 마음 이 없는 자동인형이 될 것이다 . 이는 바로 바드 (Bard 1968) 가 피질 제 거 고양이에서 증명한 바 있다 . 그림 8. 5 의 사람에 대한 경우는 제 9 장에서 설명하게 된다. 8.6 마음과 뇌 상호작용에 관련되는 신경 구조 마음과 뇌의 문제를 좀더 살펴보려면 대뇌피질의 뉴런(신경세포)과 함께 뉴런끼리 밀접하게 접촉하고 있는 부위로서 그것을 통하여 한 뉴 런이 다른 뉴런을 홍분시킬 수 있는 접합부(시냅스)에 대하여 상세하 게 알아보아야 한다(그림 7.2 a ). 이 같은 방식 으로 충분하게 홍분되 면 뉴런은 임펄스를 방출하고 임펄스는 축삭을 따라 내려가 다른 뉴런의 시냅스로 이동하여 그 뉴런을 홍분시킴으로써 그 뉴런이 임펄스를 방 출할 수 있게 한다. 기본적인 과정은 충분한 임펄스가 한 뉴런의 시냅 스(수백 개)에 모이면 임펄스를 방출하고 이는 분지된 축삭을 거쳐 다 른 뉴런의 수백 개 시냅스를 활성화하는 것이다. 그림 7.2b 는 대뇌피질 신경세포를 도형화한 것으로서 세포체 soma 에 는 긴 첨단수상돌기가 있고 각 수상돌기에는 가시 s pin e 가 돋아나 있어 그것은 다른 신경세포의 신경말단과 시냅스를 이룬다 . 각 추상세포에 는 약 1 만 개의 가시 시냅스가 있고 그 일부를 그림 7.2b 에 보여주고
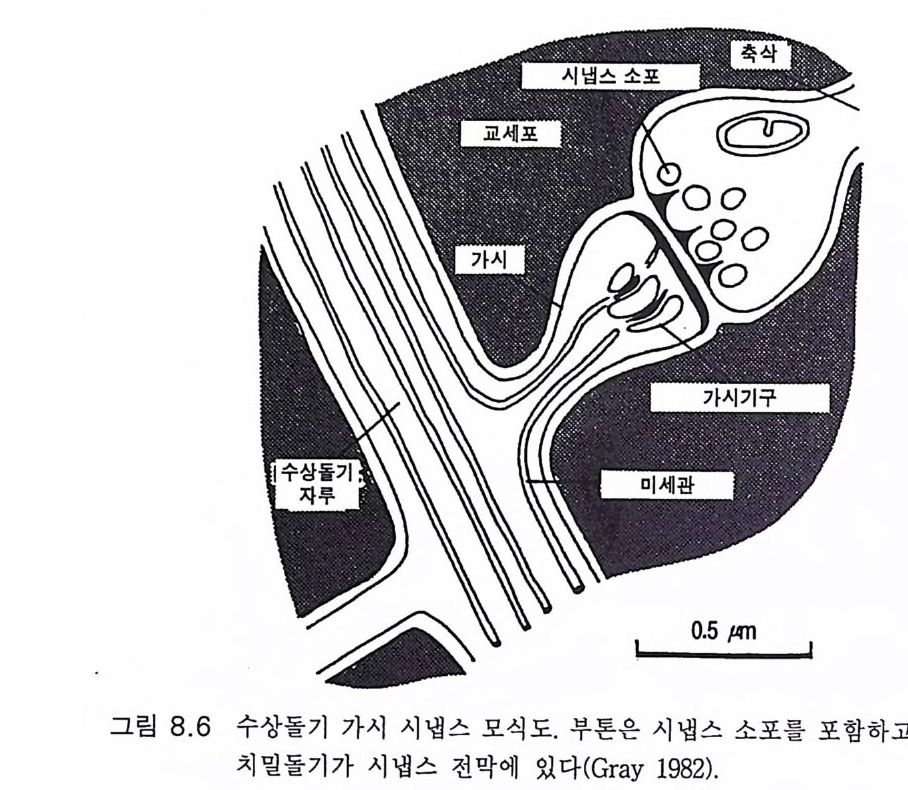 그림 8.6 수상돌기 가시 시냅스 모식도. 부톤은 시냅스 소포 를 포함하고 있으며
그림 8.6 수상돌기 가시 시냅스 모식도. 부톤은 시냅스 소포 를 포함하고 있으며
있댜 그림 8 . 6 은 그 같은 시냅스의 도형인데 신경섬유는 늘어나서 말 단 신경분지 팽대부 bou t on 가 되고 이것은 가시의 특수하게 두터워진 세포막과 밀접하게 접촉하고 있다. 신경분지 팽대부 속에는 수많은 소 포가 있는데 각 소포에는 특이한 시냅스 전달물질 5000-10000 분자가 포함되어 있다. 대뇌피질의 홍분성 신경분지 팽대부에 있는 것은 대부 분 글루타메이트와 아스파르테이트이다. 일부 · 시냅스 소포는 접합 전 막과 밀접하게 접촉하고 있고 극히 좁은 시냅스 간격을 사이에 두고 접합후막과 마주하고 있다. 이들 시넵스 소포는 치밀돌기 dense pro je c ti on 사이 에 배 열된 것처 럼 보인다. 더 자세한 구조 분석은 특히 취리히의 콘라드 애커트 Konrad Akert
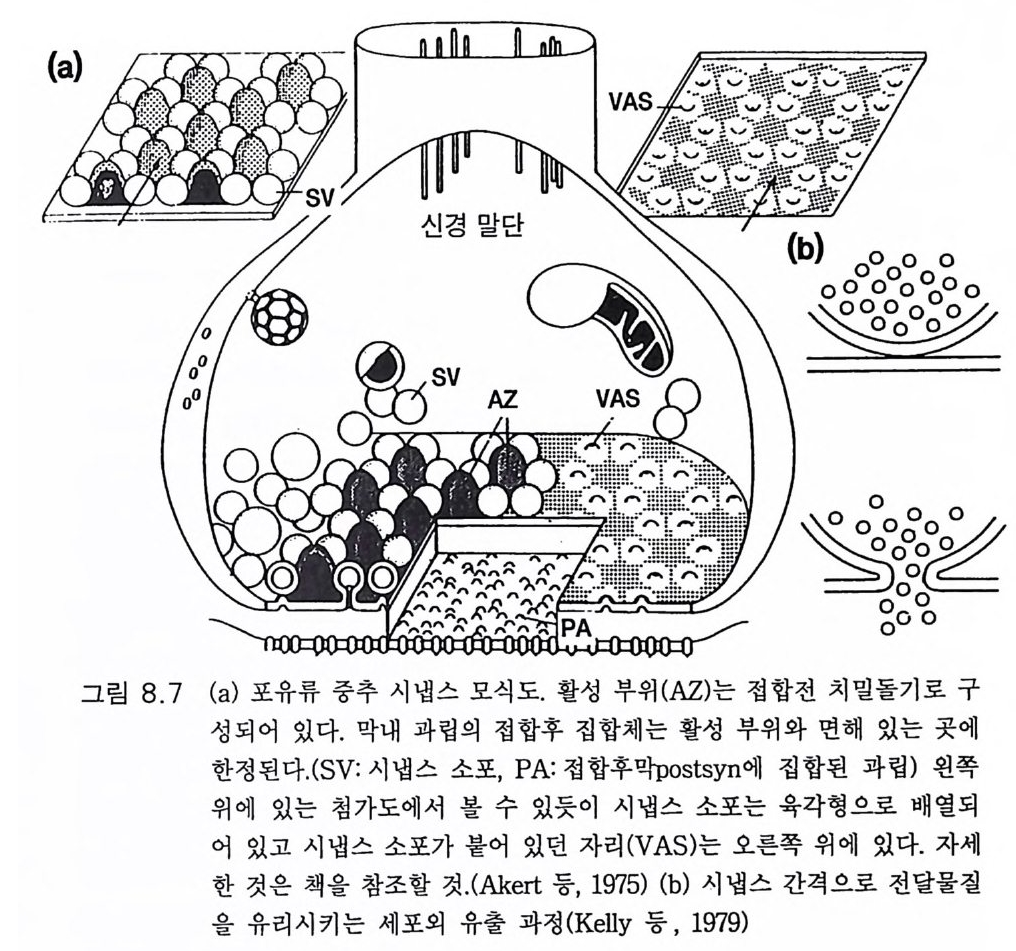 (a)
(a)
와 공동연구자들 (1975) 의 동결-절단 방법에 의한 것으로서 이상적인 가시 시냅스를 구성할 수 있었다(그림 8.7a). 내부 구조를 나타내기 위 하여 일부를 절제한 투시도로 보여주고 있다 . 시냅스 소포와 치밀돌기 (그림 8 . 6) 는 비교적 치밀하지 않게 배열되어 있고 그림 8.7a 의 왼쪽 위 에 정확한 배치룰 보여주고 있다. 삼각형을 이루고 배열된 접합전 치 밀돌기 사이에 시냅스 소포가 육각형으로 배치되어 있다. 이 같은 복 합 구조를 접합전 소포망이라 하며 불완전 결정의 성질을 가졌다고 할 수 있댜 뇌 시냅스의 신경분지 팽대부는 단일 접합전 소포망을 가졌
고 그림 8 . 6 과 8.7a 에 있는 것이다. 하나의 접합전소포망에 함유되는 시냅스 소포의 수는 단지 어림짐 작일 뿐이다. 보통 30 내지 50 이라고 한다. 따라서 신경분지팽대부(약 2000) 의 시냅스 소포는 단지 적은 분획(약 2000) 만이 접합전소포망의 발사 위치에 놓여있다. 나머지는 신경분지팽대부 속에 성글게 배치되 어 있다 그 일부를 그림 8 . 6 과 8.7a 에 보여주고 있다 . 그림 8.7b 는 시냅스 소포의 일부를 확대한 것으로 접합전막과 접촉 하고 있고 그 안에 함유된 전달물질 분자를 보여주고 있다 . 아래쪽 것 은 접합 간격으로 전달물질 분자를 유리하는 세포의 유출 exoc yt os i s 과 정이댜 그림 8.7a 의 왼쪽에도 이와 같은 소포를 볼 수 있다 . 그림 8.7 a 의 오른쪽에는 소포와 치밀돌기를 밀어낸 후의 그림으로 소포부착 부 위 (VAS) 가 육각형을 이루고 있는 것을 볼 수 있다 . 이를 확대한 그림 이 오른쪽 위에 있다. 접합전 임펄스가 단일 신경분지 괭대부를 활성화했을 때 발생되는 접합후막 전위의 변동이 정확하게 분석되었고 세포내 미세전극을 이 용하여 기록되었다 . 임펄스에 반응하여 신경분지 팽대부로부터 소포가 방출(세포외 유출)되는 확률은 일반적으로 0.5 또는 그 이하이다(J ack 등 1981a,b; Korn 과 Faber 19 얽). 이 확률은 상황에 따라 변동될 수 있 다. 접합전 소포망은 함몰된 소포의 세포의 유출을 조절하는 일종의 기능조직을 가졌을 것이라고 생각된다. 8.7 마음-뇌 학설들의 도형 적 비 교 한편으로 이 원론적 상호작용론 가설(그림 8 .1)과 동일론의 추론적 예 측둘을 각각 실험적으로 검증할 수 있을까 하는 의문을 제기할 수 있 댜 그림 8. 8a 는 자동가설의 기본 특징을 간단하게 나타낸 도형이다.
파이 글 (Fe ig! 1967) 에 따르면 마음-신 경 의 자동은 뇌 의 고위 수준 특히 대뇌피질에서 뉴런들 또는 뉴런 계통에 대해서만 일어난다. 이들 뉴런 을 마음_신경 사건 (MNE) 뉴런이라고 할 수 있고 뇌내의 다른 뉴런 특 히 입력과 출력 신경로에 있는 뉴런들은 단순한 신경사건 (NE) 뉴런일 뿐이라고 하겠다 . 자동 가설로 예측할 수 있는 것은 MNE 뉴런은 특수 한 상황에서 그들의 자극 발사는 심적 사건과 일치(자동)할 것이기 때 문에 구분이 뚜렷할 것이라는 것이다. 그러나 이 자극 발사는 물론 다 른 뉴런들 MNE 또는 NE 로부터의 입력에 반응하는 것일 것이며 심적 사건에 의하여 결정되거나 조정되는 것은 결코 아니다. 이것이 위에서 인용했던 물리적 세계의 폐쇄성이다 (Po pp er 와 Eccles 1977). 그러나 그림 8.8a 의 도형은 내적으로 생성된 생각이 대뇌피질의 특 정 부위에 있는 뉴런을 강력하게 흥분시킨다는 증거(그림 6.9, 7.14, 8.3 a ,c, 8.4 b , 9.3 , 10 . 3) 에 의해 도전받게 된다. 이들 소견으로 심적 사건 (ME) 자체로부터의 입력을 도형에 추가하게 되었다. 그림 8.8b 에 추가 된 화살표로 보여 주고 있다. MNE 뉴런의 자극 발사는 반응을 나타낼 것이고 이는 주의 집중, 조용히 생각하기, 혹은 의지적 사고, 심적 사 건이 없을 때의 반응과는 다른 것이다(그림 6.10, 7.14, 8.3, 8.4 , 9.3, 10.3 ). 그림 8. 8b 는 이원론적 상호작용론을 나타내는 도형이다. 8,8 양—자물미 리세 에국소 근설거 (E한c c le마s 음19과86 ) 뇌 상호작용의 새로운 가설 생각과 같은 비물질적 심적 사건이 어떤 방식으로든 대뇌피질의 뉴 런과 같은 물질적 구조에 작용할 수 있다는 가설(그림 8.5, 9.5) 은 극복 하기 어려운 곤란에 직면하게 된다고 유물론자들은 비판한다. 그 같은 추상적 작용은 물리학의 에너지 보존의 법칙 특히 열역학 제 1 법칙에
위배되는 것이라고 단언한다. 이 반론은 19 세기 물리학자들에 의하여 고수되었고 19 세기 물리학의 관념에 빠져 있어 20 세기의 양자물리학 에 의한 혁신을 인식하지 못한 신경과학자 및 철학자들에 의하여 고수 되었댜 불행히도 마음-뇌 문제에 도전해 보려는 양자물리학자가 드물 었다 그러나 양자물리학자 마르게노 (Mar g enau 1984) 의 최근의 저서는 근본적 기여를 하였다 . <양자역학의 확률계와 같은 어떤 장(場)들은 에 너지도 물질도 지니지 않는다>고 한 그의 주장은 19 세기 물리학으로부 터의 주목할 만한 변환이다. 그는 계속해서 다음과 같이 말했다. 뇌와 같은 대단히 복합적 물리 계통에 있어서 뉴런과 감각기관 그리고 그 구성분은 확률적 양자 법칙에 의하여 지배되기에 충분할 정도로 작다 . 이런 경우에, 물리적 기관은 항상 각각 분명한 확률을 가지고 다양한 변 화 가능한 상태에 있다. 만일 에너지, 다론 것에 비해 다소의 에너지를 필 요로 하는 어떤 변화가 일어나면 복잡한 유기체는 자동적으로 그 에너지 를 공급한다. 마음 그 자체가 에너지롤 공급할 필요는 없다(p .96). 결론적으로 마르게노는 다음과 같이 말했다. 마음이란 적합한 물리적 반응계라고 할 수 있다. 그러나 비물질계로서 가장 근접 한 상사형 은 아마도 확률계 일 것 이 다(p . CJ?). 마음-뇌 상호작용의 이원론적 가설을 좀더 정확하게 규정하는 데 있어 초기 개념은 심적 사건의 세계 전체(세계 2) 는 물질-에너지의 세 계(세계 l) 나 마찬가지로 자율적인 존재를 갖는다는 것이다(그림 4.2). 현재의 상호작용론적 가설은 이들 존재론적 문제와 관계가 없고 다만 심적 사건의 신경 활동에 대한 작용 양상에 관계되는 것이며 그것은 그림 8.5 와 9.5 의 접경을 가로지르는 하향 화살표의 성질에 관계되는 제 8 장 진화상의 마음과 뇌 문제 273
것이다 마르게노에 따르는 가설은 마음-뇌 상호작용은 양자역학의 확 률계가 상사형이라는 것이다. 이것은 질량도 없고 에너지도 가지지 않 으나 미세국소에서 효과적인 작용을 유발할 수 있다는 것이다. 더 특 이적으로는 의도나 계획된 생각에 관련되는 마음의 정신 집중은 양자 역학의 확률계와 상사형인 과정으로 신경 활동(사건)을 유발할 수 있 다고 제안하였다. 어떤 신경 활동(사건)이 양자론적 확률계와 상사형인 심적 사건에 대한 적절한 수령자가 될 수 있는가? 한 신경세포가 다른 신경세포(그 림 8.6 , 8.7) 와 특히 활동의 미세국소에 소통하는 접합부 기전의 성질에 관한 최근의 발견에서 이미 해답을 얻었다고 할 수 있다. 제기될 수 있는 첫번째 질문은 양자역학의 확률계에 의하여 발생될 수 있는 효과의 크기에 관한 것이다 . 시냅스 소포의 질량은 하이젠베 르크 He i senber g의 불확실성 원리의 범위 밖에 놓일 정도로 큰 것인가? 그림 8. 7b 에서 세포의 유출이 일어나려면 이중막의 작은 부분이 치환 되어야 하는 것을 볼 수 있다. 이중막의 두께는 10nm 이하이며 만일 치 환되는 부분이 lOnm X lOnm 라면 이 입자의 질량은 단지 10-1 8 g에 지나 지 않을 것이며 이는 쉽게 양자역학이나 하이젠베르크의 불확실성 원 리의 범위에서 취급할 수가 있다 (Eccles 1986). 특히 소포는 이미 접합 전 소포망 위에 위치해 있는 것이므로 세포외 유출은 점성 매체를 통 한 이동과는 상관이 없다. 정해진 심적 영향은 세포외 유출을 위해 이 미 자리잡혀진 소포를 선택하는 일 이상은 아닐 것이다. 접합전 소포 망에 있는 전체 소포의 세포의 유출 확률은 l 이하이다(J ack 둥 1981a, Korn 과 Faber 1987). 결론적으로 하이젠베르크의 불확실성 원리에 따른 계산으로 접합전 소포망(그림 8.7a) 의 소포가 양자론적 확률계와 상사 형으로 작용하는 심적 의도에 의하여 세포의 유출을 위해 선택될 수 있다고 할 수 있다. 그림 8.7b 에서 지적했듯이 세포외 유출을 개시하는 데 필요한 일은 고농도에서 저농도로 전달물질 분자가 탈출함에 의하
여 동시에 그 자리에서 보상된다. 양자물리학에서 에너지는 미세국소 에서 빌려올 수 있고 즉시 되갚을 수 있는 것이다. 그래서 세포의 유출 의 수지는 물리학의 에너지 보존의 법칙을 어기지 않아도 되는 것이다. 두번째로 제기되는 의문은 단지 단일 소포(그림 8.7a,b) 로부터 방출 된 효과의 크기이다. 이것은 뇌의 작은 부분에서조차도 신경 활동의 양상을 변경시키기에는 너무 작은 차원의 크기이다 . 그러나 대뇌피질 의 추상세포에는 수천 개의 비슷한 신경분지 팽대부가 있다(그림 7.2 b , 7.3 b , 7.6). 미세국소설에 따르면 접합전 소포망은 심적 의도에 대해 신 경분지 팽대부로부터 소포의 세포외 유출을 골라 선택하는 기회를 마 련한댜 이는 그 시간에 활성화되는 가시 시냅스 전체에 걸쳐 일어날 수 있고 단일 피질추상세포에는 약 1 만 개가 있으므로 활성화되는 시 넵 스는 수천 개 가 될 것 이 다 (Szen tag o thai 1978, 1983). 이제 가설적인 미세신경 사건의 구조를 살펴보기로 하겠다 . 포유류 대뇌피질의 주목할 구조적 특징은 제 v, m 및 II 판의 추상세포 첨단 수상돌기의 배열이다. 이것들은 다발 또는 무리가 되어 제 1 판으로 올 려보낸다 (Fle i schhauer 1974, Feldman 1984, Pe t ers 와 Kara 1987). 세 개의 첨단수상돌기의 이 같은 밀접한 연관의 일부를 그림 7.6 에 나타내었다. 수상돌기의 다발짓기는 대뇌피질 모든 영역의 특징이고 제 W 판에 있 는 대뇌피질의 접절편에서 볼 수 있다 . 평균적으로 수상돌기 다발은 제 V 판 추상세포에는 약 30 개의 수상돌기를 함유하고 직경은 약 50 陣 이고 간격은 비슷하다 . 그 수는 제 田 및 II 판에서 더 증가하여 피질 표면 쪽으로는 100 개까지 된다. 피터스와 카라 (1987) 에 따르면 수상돌 기 다발(속)은 대뇌피질의 미세단위 구조이며 정확하게는 덴드론 dendron 이라고 할 수 있다는 것이다. 또한 이것은 단위 심적 사건과 상 호작용하는 미세신경 사건의 피질구조라는 것이다. 사람의 대뇌피질에 서 이 같은 덴드론의 수는 엄청나서 약 4000 만 개나 된다. 하나의 통합적인 가설이 미세국소 가설에서 발전하였고 그림 8.9 에
도시되었댜 세 개의 덴드론(신경단위)이 있는데 각각은 검은 사각형, 빈 사각형 및 검은 점으로 표시된 심적 단위 또는 사이콘p s y chon 에 의 하여 관통되어 있다. 이 같은 통합적인 관계에 대한 많은 증거가 있으 며 (Eccles 1989) 대단히 단순하면서 설득력이 있는 것이다. 이 가설은 세계 2 가 완전한 자율성을 가진다는 점에서 동일설(그림 8 .1)과 다르다. 심적 의도는 사이콘을 통하여 작용하고 검은 사각형의 경우처럼 그 덴 드론에서 선택을 기다리고 있는 소포를 가진 수만 개의 활 성화된 접합 전 소포망을 갖는다. 뇌에서 마음으로의 반대방향 작용은 다음과 같은 가설의 전개를 요한 다. 매번 사이콘이 세포의 유출(그림 8.7) 에 필요한 소포를 성공적으로 선택할 때마다(양자론적 확률계에 따라) < 미세 - 성공mi cro-success > 은 심 적 세계(세계 2, 그림 9.5) 를 통한 소통을 위하여 사이콘에 등록된다. 지각 은 특수한 피질 영역을 활성화하는 주의 집중 방향dir ec t ed a tt en ti on 에 의존하는데 그림 10.3 에 손가락 감각 예상 실험의 경우에서 나타낸바와 같댜 물론 비슷한 시간에 사이콘이 그 덴드론의 접합전 소포망에 있는 수만 개의 소포로부터 상당히 많은 소포를 성공적으로 선택하면 크게 중폭된다 . 사이콘의 <성공> 신호는 그 단위의 특수한 경험적 특징을 세계 2 로 보낼 것이다. 어떤 감각 입력이 한 덴드론의 접합전 소포망의 강력한 홍분을 유 발하면 연관된 사이콘(그림 8. 9) 은 양자론적 확률계에 따른 세포의 유 출을 위한 소포 선택 기회가 증가하게 된다. 이 <성공 > 은 사이콘에 의하여 등록되고 그것의 경험적 특징이 세계 2( 그림 9.5) 를 통하여 전 달될 수 있다. 마음-뇌 경계면은 교차될 수 있는 것으로 생각된다 ; 감각신경계로 들어오는 입력은 감각 경험을 이룬다는 관찰에 대한 잠정적 설명이 제시될 수 있다. 예를 들면 영역 V 4( 그림 6.4 , 6 . 6) 에 있 는 통합 덴드론은 경계면(그림 9.5) 을 교차하여 이것에 연결된 사이콘 (그림 8.9 의 점 모양)에 작용하여 고정된 단위 연결에 따라 적색 감각을
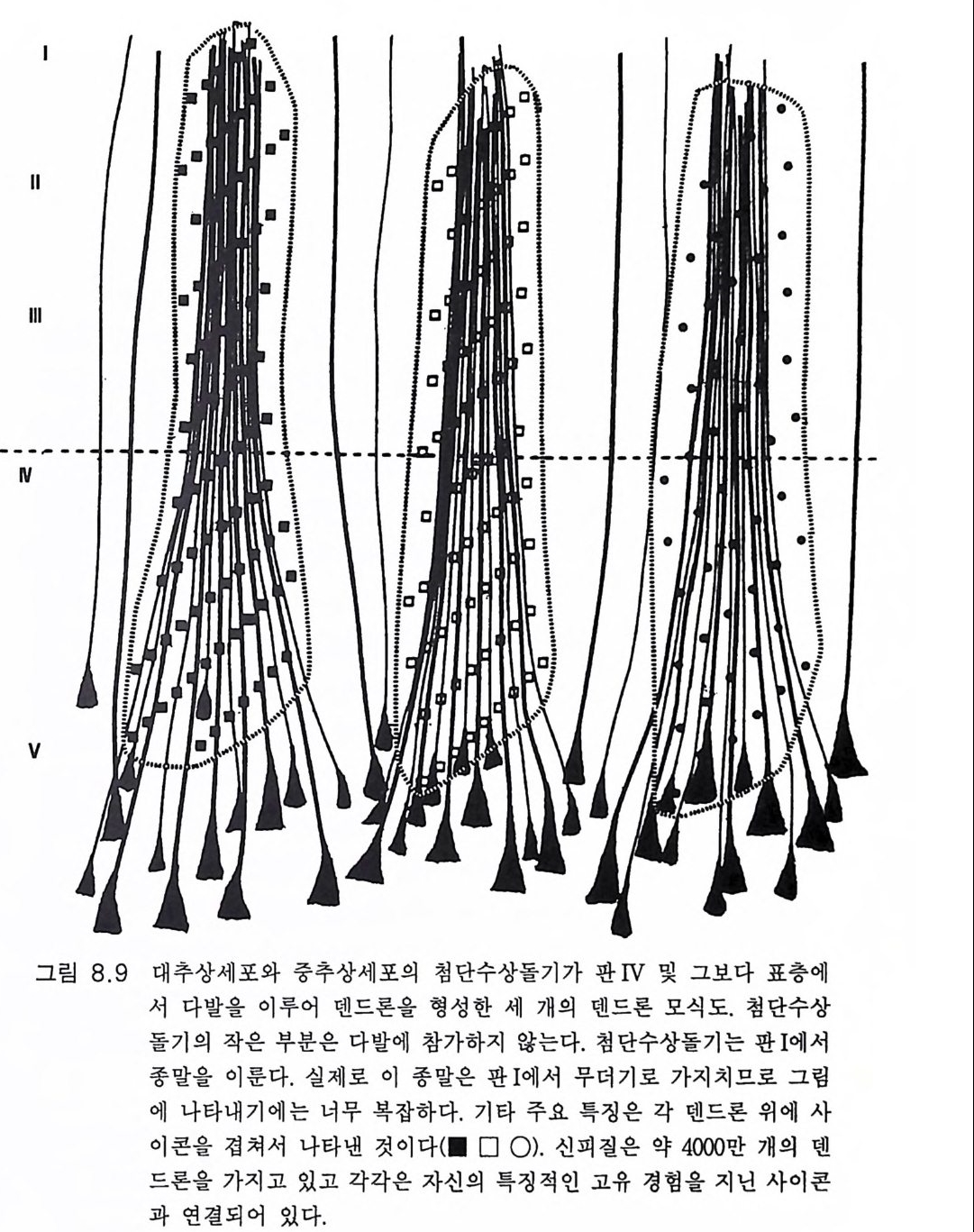 ”lll 2•g•: ::~•.틀-•• • • ―墓..….z,.`...l .,. ·.. .,m ' --:---I,-_' -I-'-,.', -』E··s'2,· -.'. I|,-투l’ii_' ._” s%?,.‘ w..,,• 1 ••.•• w·•• ·•••1•w•• •1 ••·3•W••u•.•2• •·•2 ·*•2 . •.••
”lll 2•g•: ::~•.틀-•• • • ―墓..….z,.`...l .,. ·.. .,m ' --:---I,-_' -I-'-,.', -』E··s'2,· -.'. I|,-투l’ii_' ._” s%?,.‘ w..,,• 1 ••.•• w·•• ·•••1•w•• •1 ••·3•W••u•.•2• •·•2 ·*•2 . •.••
나타내게 될 것이다. 우리가 주의를 집중하면 수백만의 단위 지각(사 이콘)이 우리가 맛보는 전체적인 경험으로 순간순간 통합된다. 8.9 마음_뇌 문제의 재고 각기 기초적 우위를 차지하는 세계 l 과 세계 2 가 나란히 놓여있는 그림 4.2 , 8.5, 및 9 . 5 를 되돌아 볼 수 있다. 이것은 두 가지 구별되는 차 원의 존재들로 받아들여야 할 어떤 것이다. 이런 문맥에서 이 장에서 는 어떻게 뇌의 미세국소가 이들 두 완전하게 구분되는 개체 사이를 소통하는 통로의 탁월한 성질을 갖게 되었는가를 설명하고자 하였다. 그림 8.3, 8.4 및 10.3 에서 지적했듯이 심적 사건이 효과적으로 뇌에 작 용한다는 것이 인정된다면 철학적으로 큰 의미를 가질 것이다. 사람들 은 모두 우리의 행동 특히 언어적 표현에 대하여 최소한 어떤 조절과 책임이 있는 것처럼 생각하고 행동한다. 그러나 환원설학자들은 이것 이 물리학의 에너지 보존의 법칙에 위배되기 때문에 이는 환상에 지나 지 않는다고 비판한다. 그러나 이 비판은 이제 쉽게 파기할 수 있다. 미세국소설에 따르면 마음-뇌 상호작용은 뇌의 흥분성 시냅스의 두 가지 주목할 만한 특징과 밀접한 관계가 있다. 첫째 접합전 소포망(그 림 8.6, 8 . 7a) 의 구조와 배치는 보통 하나의 신경분지 팽대부에 단지 하 나의 접 합전 소포망만을 갖도록 되 어 있다 (Ake rt 둥 1975). 둘째 접 합 전 소포망으로부터 시냅스 소포의 방출은 확률적 함수로 결정되는데 이는 1 이하이고 때로는 훨씬 낮으며 상하로 조절될 수 있는 것이다 (Ja ck 둥 1981b, Korn 과 Faber 1987). 마음一뇌 상호작용에 있어 아무런 기능도 할 수 없을 것이라는 상황 에서 이 같은 성질을 가진 시냅스가 인정되었다는 것은 놀라운 일이 댜 예를 들면 금붕어의 척수에 있는 마우트너 Mau t hner 세포의 억제성
시냅스는 소포 방출의 확률 연구에 주로 이용되는 것의 하나이다. 이 것에 관하여는 콘 및 파버 (Korn 과 Faber 1987) 에 철저하게 기술하였다. 조직 학적 으로 (T ri ller 와 Korn 19 82) 접 합전 소포망은 애 커 트 등(1'57 5) 이 기술한 것과 유사하였다 . 소포 방출의 확률 연구에 주로 이용되는 또 다른 부위는 운동신경의 Ia 구심성 임펄스를 만드는 시냅스이다(그림 3.7 Jac k 등 1981a). 여기서도 역시 심적 사건의 전달과 신경적 사건과 는 기능적 관계를 볼 수 없다. 더욱 놀라운 일은 연체동물 시냅스에서 접합전 소포망을 발견한 것이다 (Kandel 등 1987). 분명히 여기 선행적 진화의 또 다른 예가 있다. 그 개념은 제 6 장에 서 소개하였다. 화학적 접합부 전달의 진화 훨씬 초기에 소포 방출을 조절하는 기전이 발달하였다 . 접합전 소포망의 기능적 설계는 매 임펄 스에 대하여 1 이하의 제한적 세포외 방출로 시냅스 소포를 보존하도 록 하였을 것이다. 그렇지 않았다면 고빈도의 시냅스 홍분으로 신경분 지 팽대부의 소포가 고갈되어 버릴 위험이 있는 것이다. 이 같은 관계 로 볼 때 거대한 신경근 시냅스에서는 임펄스마다 100 이상의 시냅스 소포가 세포의 유출이 되도록(H ubbard 1973) 설계되었다는 것 (Ak ert 등 1975) 은 흥미 있는 일이다. 그같은 다량의 시냅스 전달물질(아세틸콜린) 유출은 운동 종판의 효과적인 탈분극에 필요한 것이다. 이제 고등동물이 의식을 갖게 되는 진화적 사건 (8 . 2) 으로 돌아가기 로 한댜 포퍼 Po pp er 는 다음과 같이 말하였다. 완전한 의식의 출현……은 물론 가장 위대한 기적의 하나이다 (l
진화의 예라 규정할 수 있다. 이는 화학적 시냅스에서 자극 전달이 효 과적으로 이루어지도록 발생하였고 , 굉장히 오랜 진화 기간 후에 미세 국소론에 따라 마음-뇌 상호작용에서 이용되어 동물은 의식을 갖게 되었다. 호미니드가 자아의식을 갖게 되는 더 이상의 호미니드 진화적 발달 에 관하여는 제 9 장, 제 10 장에서 다루게 된다. 심적 사건에 의한 신경적 사건의 조절 방식에 근본적인 변화는 없었다고 제안할 수 있다 . 이 장 에서 논의한 미세국소 가설은 호미니드 진화에서 발달하는 마음의 자 아의식에도 응용될 것이다 . 따라서 제 8 장은 호미니드 진화에서 일어난 일, 자아의 의식과 관련된 뇌의 특수 부위의 발달을 설명하는 기준이 된 다 (Creu tzfe ld t 1979, 1987).
제 9 장 자아의 창조와 그 뇌 이 장에서는 사람 뇌의 비대칭성이라는 고도로 의미 있는 특징 그 리고 자아의식과 밀접하게 관계되는 비대칭성 소엽에 관하여 알아보 려고 한다. 따라서 우선 대뇌의 일부 비대칭성은 호미니드 진화로 이 루어진 것이라고 제안한다. 9.1 해부학적으로 관찰된 비대칭성 대뇌의 비대칭성에 관한 상반된 관찰의 역사는 길다. 그러나 의미 있는 관찰들만 열거하자면 대단히 짧다. 크기, 무게, 뇌회의 분지 둥 대뇌피질의 특징에 있어서는 유의한 비대칭성을 볼 수 없으나 좌우 대 뇌반구의 기능적 특성에 있어서의 비대칭성은 놀라운 것이다• 원숭이와 바분의 뇌에서는 비대칭성이 발견되지 않았다 (Wada 둥 l
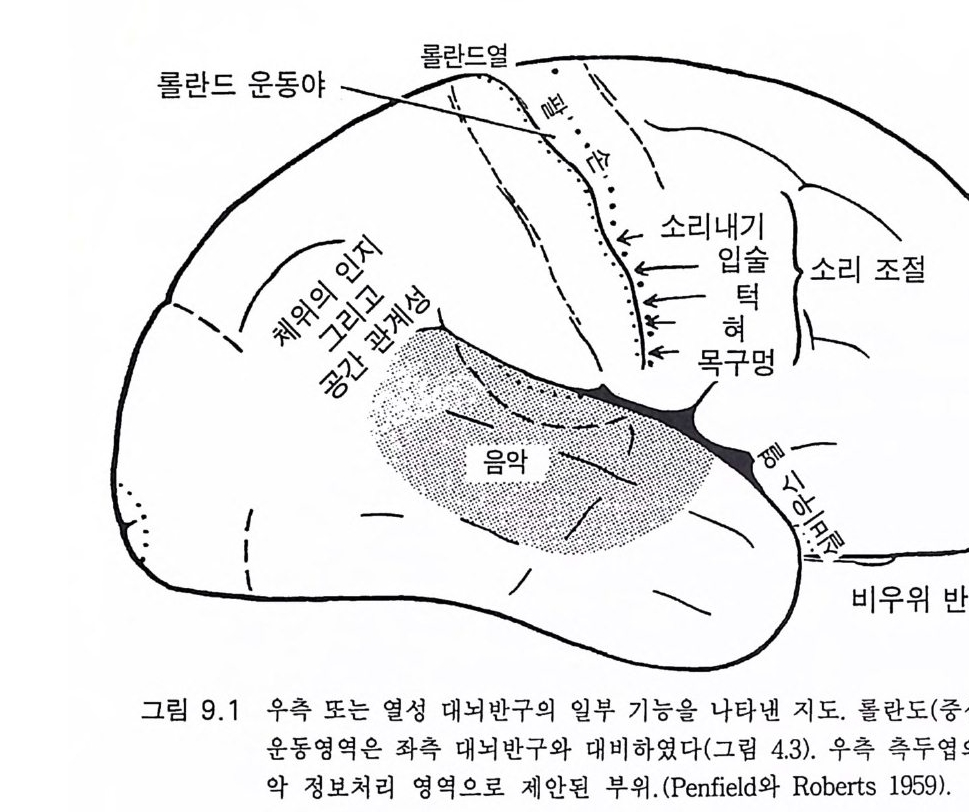 大
大
길 다 (Ye ni -Koms hi an 과 Benson 1976). 사람의 뇌에 관하여는 루벤스 (Rubens 1977) 가 대단히 주의 깊게 실 비우스열을 연구하여 오른쪽이 보통 (36 번 중 25 번) 왼쪽보다 앞서서 위 쪽으로 각을 이룬다고 보고하였다. 왼쪽은 각을 이루기 전에 평균 1.6 mm 가 더 계속된다고 하였다. 이 비대칭성은 측두평면의 비대칭성과 관계있는 것으로(그림 7 .4 )(Gesch wind 와 Lev itsk y 1968) 실비우스열 믿 에 있댜 이것은 65% 의 경우 왼쪽이 더 큰데 언어의 극단적인 비대칭 성에 비하면 훨씬 적은 것이다. 언어의 경우 95% 에서 좌측 대뇌반구 가 대표한다 (4.5) . 와다 둥 (Wada 둥, 1975) 은 성인의 보통 비대칭성을 확인하였고 나아
가 100 사람의, 아주 어리게는 29 주짜리 태아에서도 이를 확인하였다. 피질 언어영역의 비대칭성은 증명된 바가 없다(그림 4.3). 브로드만 영역 39 와 40( 각회 및 연상회)는 하두정엽에서 쉽게 식별되나 대칭성인 것으로 보인다. 좌측은 언어의 읽기와 듣기의 어의학적 기능을 갖는다 (4.5). 그러나 오른쪽에는 그 같은 분명한 기능은 있는 것 같지 않다. 이 수수께끼는 9.2 에서 검토하게 될 것이다. 9.2 기능적 비대칭성 원숭이에서 대뇌 비대칭성의 한 가지 간단한 증거는 손 우선성에서 찾아볼 수 있을 것이다. 이에 대한 많은 연구가 있었다. 해밀턴 (Hamilton 1977a,b) 은 원숭이의 (왼)손잡이에 관하여 대단히 주의 깊게 연구하여 평균적으로 대칭성이었음을 보고하였다. 여러 가지 대뇌 교 련 절단술 후의 학습에 관한 연구에서 어떤 비대칭성도 증명하지 못하 였다. 해밀턴 (1977a:60) 은 다음과 같이 요약하였다. <분리된 대뇌반구 에서 편측성 기억 형성이나 학습 및 수행 능력의 차이를 발견할 수 없 었다.> 해밀턴 (1977b:230) 은 끝으로 다음과 같이 결론지었다. <수집된 모든 증거로 레수스 원숭이에는 대뇌반구의 특성화가 있지 않다는 믿 음을 재확인하였다.> 불행히도 침팬지의 왼손잡이에 관하여는 철저한 연구가 이루어지지 않은 것 같다. 오랜 기간의 몸치장 행동 관찰에 따르면 침팬지는 보통 양손잡이라고 추정되나 (Goodall1986: 387-408) 왼손잡이에 관한 언급은 없고 확실하게 사진으로 나타난 것도 없다. 비인간 영장류에 반하여 호미니드 진화과정에서 좌우 대뇌반구의 해부학적으로 대칭인 부위의 기능적 비대칭성이 크게 발달하였다. 이 들 차이는 치료적으로 뇌량 절단술을 받은 환자에서 분명하게 나타난
댜 이는 9.6 절에서 논의하게 될 것이다 . 그러나 우선 국소 병변에서 증거를 찾아보는 것도 적당하다 . 초기의 관찰들은 임상적인 것이었고 뒤이어 흔히는 뇌의 수술적 절제에 의한 손상을 관찰한 것이었다. 두드러진 비대칭성은 4.5 에서 언급한 언어영역에서 볼 수 있다. 좌 측 두정엽과 두정엽의 대부분은 언어의 인지 및 생성에 관한 어의적 특성화가 이루어진 것으로 그림 3.5 와 4.3 에 보여주고 있다(베르니케 영 역). 그러나 우측 반구의 대칭 부위(그림 9 .1)는 언어와의 기능적 관련 성이 거의 없다. 마찬가지로 우측 하전두엽에 있는 브로카 영역의 대 칭에 해당하는 부위는 언어 생성에 쓰이지 않는 것으로 생각된다. 문 제는 <우측반구 대칭 부위의 기능은 무엇인가? > 이다 . 기능적 비대칭 성의 평형이 이루어지는가? 이것의 해답은 운측 또는 좌측 대뇌반구 의 국소 절제시의 임상적 증거에서 찾아볼 수 있다. 두정엽이 특히 홍 미의 대상이 된다. 브로드만 영 역 39 와 40( 그림 4.3, 4 .4)에 관심 의 초점 을 맞추어 보고자 한댜 좌측의 이 영역들은 특이적 언어 기능을 갖는 것으로 생각한다. 영역 39 는 시각 입력(쓰기 또는 전사)를 의미 부여로 바꾸는 곳으로 생 각하며 영역 40 은 청각 입력과 관련되는 것으로 생각한다. 우측에 병 변이 있는 환자의 임상적 관찰로 무동작증, 체성인식 등 몇 가지 기능 적 장애 그리고 공간적 자료의 인식 장애를 볼 수 있다 . 우측 두정엽이 손상되면 환자는 대단히 이상한 행동 양상을 나타낸 댜 이를 헤켄 (Hecaen 1967) 은 <광범한 무시의 무언극>이라고 적절하 게 표현하였다. 환자는 대측성 사지의 존재를 무시하거나 부정하고 그 것에 옷 입히는 것까지도 무시한다. 시계의 반대쪽 반구영역에 있는 대상을 관찰할 수 없을 때도 있다. 대측성 사지는 거의 움직이지 않는 다. 그러나 마비된 것은 아니다. 환자는 대측성 공간의 1/2 에서 피하려 는 경향이 있다. 이 같은 무시의 무언극에도 불구하고 환자는 그가 병 들었다는 사실을 전적으로 부인한다. 좌측을 무시하는 대표적인 경우
를 융(J ung 1974) 이 기술한 바 있다. 화가는 손상을 받은 후 자화상에 거의 오른쪽 얼굴만 그렸는데 점차로 회복되었다 . 우측 반구에 큰 손상이 있으면 환자는 주위 환경에 대한 방향감각 의 장애를 일으켜 지도 보기나 길 찾기가 힘들어진다. 이 과업은 체성 감각과 특히 부차적 반구 기능인 시각 신호와의 협동이 관련되는 것이 다. 환자는 그림을 묘사하지 못하고 조각을 맞추어 삼차원적 구조를 만들지 못한다. 우측 두정엽의 손상은 또 옷 입기에 장애를 초래한다. 요컨대 환자는 공간적 조직 결함이 있는 것이다. 우측 두정엽은 특별히 공간 정보의 취급과 공간과 신체 사이의 관 계에 있어 비언어적 형태와 관련되는 것이다. 특히 공간적 기능과 관 련된다 이것이 손상되면 무동작증, 측 섬세한 조직적 동작에 관계되 는 기능의 상실을 초래한다. 공간 정보의 취급에 있어서의 장애는 쓰 기에서 나타난다. 죽 줄은 파도치게 되고 단어의 간격이 둘쭉날쭉해지 며 지속증 때문에 변형되기도 한다. 죽 겹쳐진 두 글자는 종종 세 글 자로 겹쳐 쓴다. 예를 들면
반구 손상 때보다 약하다 . 그러나 물론 주된 장애는 실어증, 실독증 둥 언어에 관련되는 것이다. 좌측 두정엽에서는 감각 정보가 언어와 통합 된댜 좌측 두정엽의 손상으로 몸짓, 쓰기, 계산 그리고 양쪽 신체에 대한 언어적 지식의 장애를 초래한다는 사실로 이를 알 수 있다. 결국 운동능의 장애(실행중), 구성능의 장애, 계산능의 장애가 있는 것이다. 헤켄 Hecaen 은 양측 두정엽의 차이를 다음과 같이 요약하였다. 오른쪽 뇌의 손상온 여러 활동의 공간적 관계 인식에 지장을 가져 오고 왼쪽의 손상은 신호, 부호 및 분류 계통적 활동의 장애를 초래한 댜 따라서 주(우세)대뇌 반구의 활동에 언어적 중개의 조직적 역할이 필수적이라고 생각할 수 있다. 좀더 일반적인 우세 및 열세 대뇌반구의 역할을 고려한 양 뇌엽의 기능적 차이는 9.6 에서 더 다루게 된다. 밀너Miln er 와 그 동료들 (1972, 1974) 은 좌우 측두엽의 대조적 기능에 대한 대단히 홍미 있는 연구를 하였다. 약 95% 의 대상에서 베르니케 영역(그림 4 . 3) 에 중심을 두었다고 생각되는 특수한 언어적 기능이 있 었댜 이점에 관하여는 제 4 장에서 언급한 바 있다. 보충적으로, 우측 측두엽은 특히 음악 감상(그림 9 .1)과 공간적 양상의 인식에 관계가 있 다고 한다. 광범한 뇌 영역의 손상으로 초래된 전간을 치료하기 위해 한쪽 측두엽을 제거한 환자를 이용하였다. 음악 감상에 관한 측두엽 손상의 효과 연구에서 우측 측두엽에 손 상이 있는 환자는 단순한 음도나 음률을 구별함에 정상인과 다르지 않 았다. 그러나 이들 환자는 두 가지 시쇼어 시험 Seashore tes t 죽 음색과 음조 기억시험에서 차이를 나타내었다(Mil ner 1972). 예를 들면 짧은 서열의 음조시험으로 4 또는 5 음을 연속하여 두 번 빠르게 연주하고 두번째 연주에서 음도의 변화가 있을 때 그 음을 찾아내게 하는 것이 다. 우측 측두엽을 절제하면 이 시험에서 선율 식별에 대한 오류가 수 술 전보다 많아진다. 그러나 좌측 측두엽 절제시에는 점수의 차이가
별로 없댜 우측 측두엽에 관계되는 음악 감상에 대한 증거는 생크바일러 (Shankweil er 1966) 가 전통가락의 기억울 이용하여 더 제시하였다. 몇 소절을 듣게 한 다음 콧노래로 가락을 계속하게 하거나 그 이름을 대 게 하는 것이었다 . 오른쪽 측두엽 절제 환자는 이 두 가지를 모두 잘 해내지 못하였다. 그러나 예상할 수 있듯이 좌측 측두엽 절제시에는 콧 노래로 따라부르기는 잘해냈으나 언어기능의 결함이 있으므로 그 가락의 이름을 대는 언어적 과업은 잘 해내지 못하였다. 뇌의 음악적 수행 능력에 대한 우측 측두엽의 중요성은 음악적 재 능이 있던 여성의 우측 측두엽 절제에 의한 비극적 결과로 확실하게 보여 주었다 (Go tt 1973). 그녀 는 음악 전공으로 피 아노 연주자로 입 신하 였다. 수술 후에 그녀의 음악적 재능은 소실되어 가락을 연주할 수는 없었으나 잘 알고있는 노래말은 정확하게 반복할 수 있었다. 밀너Mil ner 와 공동 연구자들(1 972 , 1974) 또한 우측 측두엽의 손상이 불규칙적 자극 특히 음성적으로 식별되지 않는 양상의 자극의 감지에 있어 지속적 결함을 나타내는 경우를 제시하였다. 사용한 세 가지 시 험 중에서 얼굴 식별시험은 환자에게 특히 중요한 것이었고 둘째로 중 요한 시험은 비언어적 시각기억으로 기무라(Kim ura 1 '!7 3) 가 고안한 이 른바 반복적 무의미한 그림 시험 Rec urring Nonsense Fig ures te s t이다 . 이 시험은 낯선 기하학적 또는 불규칙적 곡선 설계에 대한 환자의 기 억을 시험하는 것이다. 또 한 가지 시험(Miln er 1 '!7 4) 은 직선상의 특정 위치에 있는 작은 원을 가리키고 나서 다시 첫번째 직선을 시야에서 가리고 같은 길이와 위치를 가진 직선에서 원의 위치를 재현하게 하는 것이다. 이 손상시험으로 우측 측두엽온 시각적 특징의 식별에 특히 관계됨을 알 수 있다. 요약하면 이들 실험으로 우측 측두엽은 사람 뇌의 음악, 공간적 식 별 그리고 기억재생 능력과 중요하게 관계됨을 알 수 있는 것이다. 우
측 측두엽 단독으로 이들이 수행된다고 주장할 수는 없다. 다만 그 영 역이 주로 관계된다는 것이다. 대뇌반구에 넓게 분포하는 이들 기능의 대뇌 위치에 관하여 살펴보는 것이 이 장의 주제이다 . 사람의 우측 측 두엽의 이들 기능은 언어적 수행능에 있어 좌측 측두엽 및 두정엽의 광범 한 관련으로 균형 잡히 게 된다 (4.5 참조). 열성 대뇌반구가 음악 감상을 관장하는 손상 연구에 의한 이 증거 들은 정 상 인 에 서 의 이 분 청 취 시 험 Dich oti c Lis t e n ing tes t( Kim ura 1973) 으로 얻어진 증거에 의해 확실하게 되었다. 이 시험에서는 헤드폰을 이용하여 동시에 연주되는 두 가지 곡조를 각각의 귀로 듣게 하는 것 이댜 실험 대상은 정상 상태에서 순서적으로 청취한 네 개의 곡조 중 에서 두 가지 곡조를 알아맞추는 것이다. 왼쪽 귀로 청취한 곡조에 대 하여는 현저하게 점수가 높았다. 이는 오른쪽 대뇌반구가 왼쪽에 비하 여 음의 인식이 우세하다는 것을 말하는 것이다. 각각의 귀에서의 신 호는 반대편 측두엽의 청각영역(그림 4.6) 으로 주로 들어가기 때문이 댜 이 같은 이분 청취법으로 단어나 숫자를 순서적으로 듣게 하면 예 상한 대로 오른쪽 귀로 입력된 것을 더 잘 인식하였다. 죽 오른쪽 귀 의 입력은 주로 왼쪽 측두엽으로 가서 처리되고 언어적으로 인식되는 것이다 이 같은 현상은 무의미한 어구 또는 뒤집어 말하기에서도 관 찰되었다. 따라서 좌측 대뇌반구는 어의적 인식에 선행하는 청각 정보 룰 처리하는 단계를 담당한다고 생각된다. 그림 9.2 의 좌우측에 있는 숫자는 시험 점수를 비교하기 위하여 비율로 나타낸 것이다. 죽 구어 의 인식은 좌측 대뇌반구가 우세하여 1. 88 에서 1. 0 이고 우측 대뇌반구 는 곡조에 대하여 우세하게 1.88 에서 1. 0 을 나타내었다. 그림 9.2 는 대 뇌의 비대칭성을 잘 나타낸 것으로 좌측 대뇌반구는 언어에 우세하고 우측 대뇌반구는 곡조에 우세하다. 베버와 키아렐로(B ever 와 Ch iar ello 1974) 는 음악에 대한 대뇌반구의 역할 분담에 관한 중요한 업적을 이룩하였다. 음악을 오른쪽과 왼쪽귀로
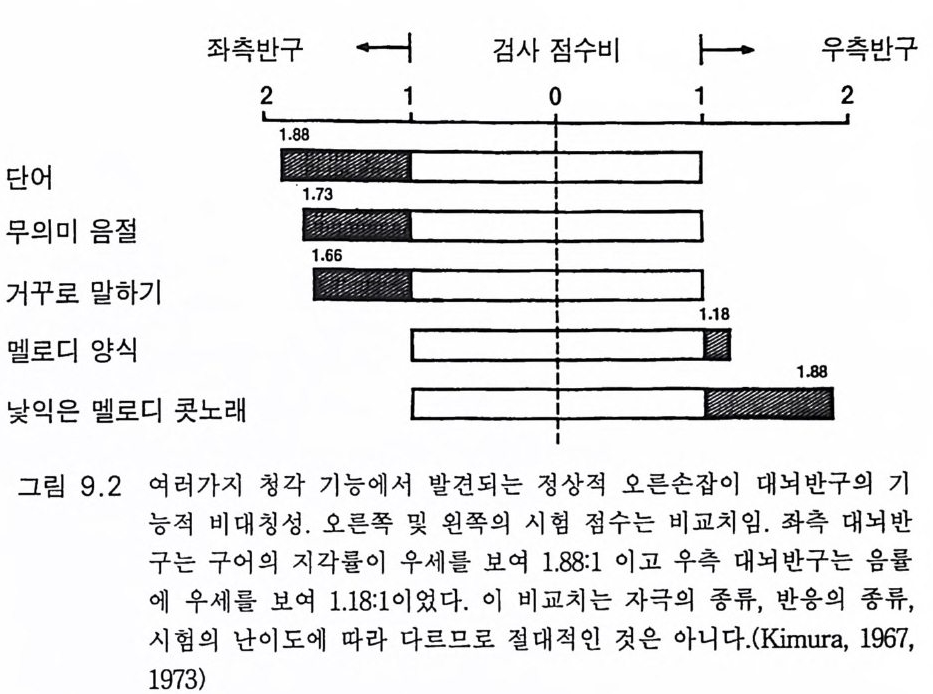 좌측반구 __1 검사점수비 I-- 우측반구
좌측반구 __1 검사점수비 I-- 우측반구
입력시켜 오른쪽과 왼쪽 대뇌반구에 우세하게 입력되도록 하여 음악 에 경험이 있는 사람은 곡조를 오른쪽 귀가 더 잘 인식하고 경험이 없 는 사람은 반대였음을 발견하였다. 그러나 경험의 총체로서의 음악 감 상은 양자 모두에서 왼쪽 귀를 통한 우측 대뇌반구로의 입력이 우세하 였다 고로 좌측 대뇌반구는 분석적 처리에 우세하고 오른쪽은 전체적 처리에 우세하다는 가설을 지지하게 되는 것이다. 음악적 해석에 있어서의 대뇌 위치에 관한 가장 직접적인 시험은 방사성 제논을 이용한 대뇌 국소혈류 (rCBF) (그림 6.7, 6.8, 6.9, 7.1 4 , 8.4 ) 에 의하여 시행되었다. 시험 절차는 한쪽 이어폰을 이용하여 짧은 간 격으로 두 가지 짧은 리듬을 듣게 하는 것이다. 그리고 비슷한지 아니 면 틀린 것인지롤 알아맞추게 하였다. 질문은 rCBF 측정 후에 시행하
 >
>
였고 그전에는 완전한 안정 상태에 있도록 하였다. 이어폰 입력은 항 상 측정하려는 대뇌피질의 반대편에 두었다. 오른쪽 대뇌반구 특히 측 두엽과 하 및 중두정엽이 훨씬 활성화되었음이 그림 9.3 에 나타나 있 댜 이 모든 영역은 좌측 대뇌반구의 베르니케의 후 언어영역(그림 4.3) 에 잘 일치하는 부위이다. 좌측 대뇌반구는 훨씬 활성화되지 않은 것 이 그림 9.3 에 나타나 있다 (Roland, S kinh싸와 Lassen 1981). 이들 관찰은 그림 9 .1에서 <음악>이라고 표지된 넓은 영역의 중요 한 실험적 근거가 되는 것이다. 사람 대뇌피질의 기능적 비대칭성은 호미니드 진화에 가장 중요한 발달이다. 유인원이나 원숭이에서의 중 거는 우리의 호미노이드 조상온 대칭성 뇌를 가졌었음을 시사한다. 비
대칭성은 호미니드 특유의 것이다. 9.3 대뇌 신피질의 모듈적 설계 대뇌피질을 보다 전체적으로 살펴보기 전에 대뇌의 연합적 및 교차 성 연결성에 대하여 일부 소개할 필요가 있다 . 그림 9 .4 (Szen 炫g o thai 1978) 는 골드만과 나우타 (Goldman 과 Nauta 1977) 의 비 범 한 발견을 그 림으로 나타낸 것이다. 그들은 방사능 측정법으로 사람 신피질의 방대 한 절편 (2000 cm2 에 약 1010 개의 뉴런)을 준불연속적 구역 단위 의 조각 보로 소분하였다. 이 같은 한 단위 내에서의 신경의 연결성은 그림 3.4 에 나타나 있다 . 각 구역 단위는 모듈이라고 생각하는데 신피질의 기 능적 설계에 있어서 기본적 전달 요소를 형성하는 것이다. 빽빽하게 들어찬 12 개의 추상세포를 그림 9 .4 a 의 왼쪽에 있는 한 개의 모듈 또는 기둥에서 볼 수 있는데 그 폭은 약 300 µm이다. 이돌 추상세포로부터 나오는 축삭은 같은 쪽 대뇌반구의 세 개의 다른 모듈로 뻗어 있고 뇌 량을 가로질러 다른 쪽 대뇌반구의 두 개의 모듈로 뻗어 있다. 이렇게 해서 한 개의 모듈의 추상세포들의 연합과 뇌량의 신경 투사를 간단하 게 나타내었다 . 그림 9 .4 a 에서 몇 가지 중요한 특징을 주목해야 한다. 첫째 모듈의 몇 개 추상세포는 완전히 중첩된 방식으로 다른 모듈에 신경섬유가 뻗 어 있다. 다른 모듈들도 마찬가지이고 폭도 역시 300µ m 정도이다. 그림 9 .4 a 에서 이것의 규모는 두 개의 모듈로 가는 단일 연합섬유의 분지로 표현되고 있고 중첩 분포는 두 개의 연합섬유와 4 또는 5 개의 뇌량섭 유들로 보여주고 있다. 둘째 뇌량섬유는 전부는 아니지만 대체로 반대 편 반구의 대칭되는 모듈로 뻗는다. 셋째 대칭되는 모듈 사이의 연결은 상호 왕래한다. 이에 반해 덴드론 (dendron, 8 . 8) 은 입력 접수에 관여
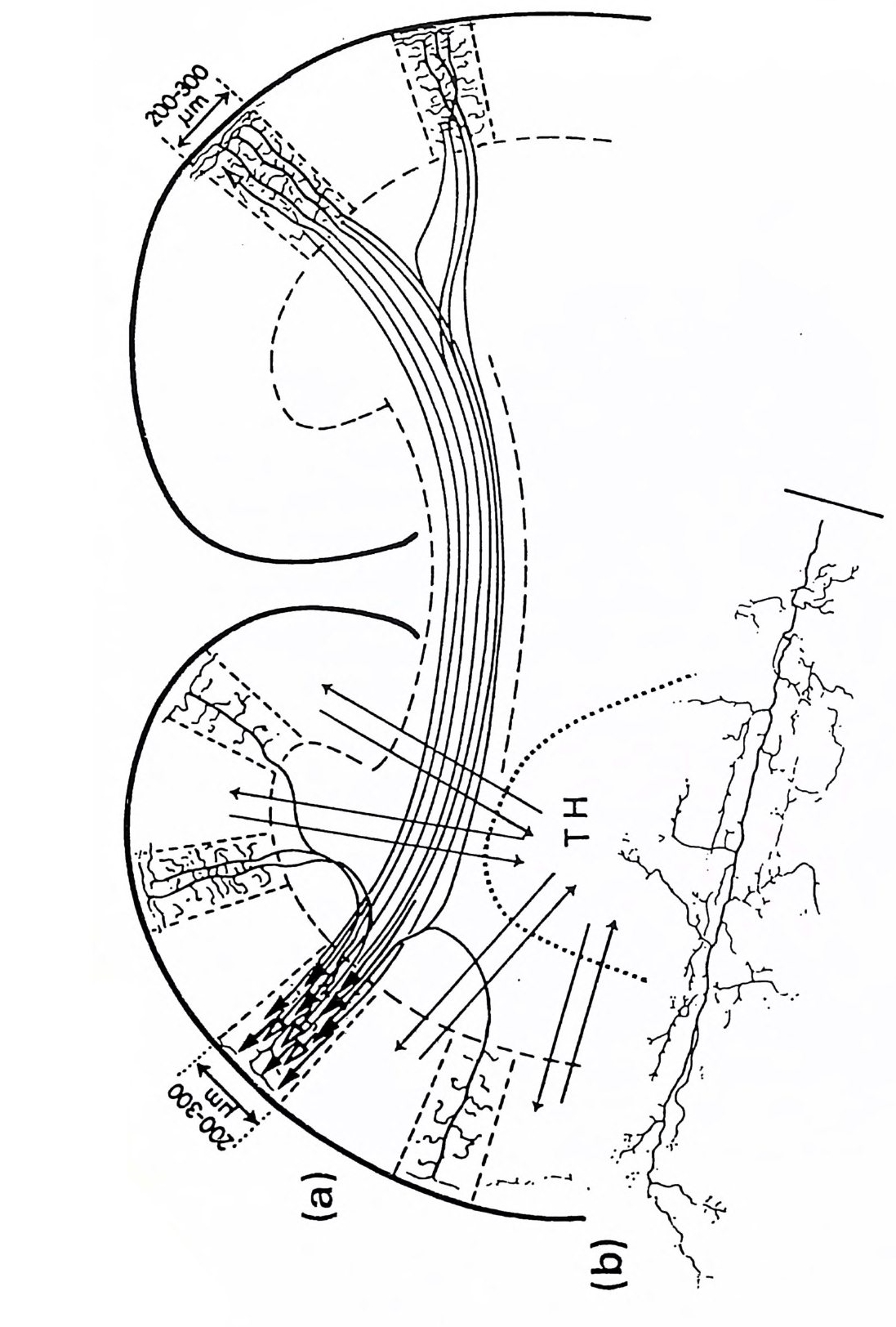 쭙#^ `:` / \~~J'. -- --
쭙#^ `:` / \~~J'. -- --
그림 9.4 (a) 피질피질 연결성의 일반 원라 연결은 양측 대뇌반구에 있는 200- 300 JaD의 직경을 가진 수직 기둥 사이에 고도로 특이적인 양상으로 이 루어져 있다 . 동측 연결은 주로 rn 충에 위치한 세포들(왼쪽에 스으로 나타낸 것)로부터 유래하고 반대측 연결은 II-VI 충에 위치한 세포들 (. ..으 로 나타낸 것)로부터 유래한다 . (TH: 시상) 이 그림에는 피질의 다 른 부위에서 기시하여 동일한 기둥으로 모이는 구심성 섬유는 나타내 지 않았다 (b) 골지 염색된 단일 피질피질 구심성 섭유. (a) 에 있는 단 일 구심성 섬유롤 고배율로 모듈과 관련지어 나타낸 것. 수많은 가지 치기를 보여준다.
하는 훨씬 더 작은 단위체이다.11) 9.4 진화적 극점: 자아의식의 발단 최고의 심적 경험으로서 자아의식적 마음이라는 용어를 쓰려고 한 다. 이는 스스로 안다는 것을 알게 되는 것이라고 말할 수 있는데 물 론 이 규정은 처음에는 주관적이고 내성적인 것이다. 그러나 언어적 소통에 의하여 다른 인간들도 자신을 인지하는 경험을 공유한다는 것 을 확증할 수 있다 . 도브잔스키(D obzhans ky 1967) 는 자아의식 또는 그 의 표현으로는 말로 자아 인식 sel f -awareness 의 비범한 출현을 다음과 같이 잘 표현하였다. 자아인식이라는 것은 인류의 특징 중에서 기본적인 것의 하나이다 . 아 마도 가장 기본적인 것이라 하겠다. 이 특징은 진화상 대단히 새로운 것 인데 인류가 발전해 나온 조상 동물들은 자아 인식의 혼적만을 가지거나 혹은 아예 전혀 갖지 못했다. 그러나 자아의식은 그후 여러 가지 암울한 동반자-공포, 근심 그리고 죽음의 인식 둥-울 가져왔다. 사람은 죽 음의 인식으로 괴로움을 당한다. 자신이 죽을 것이라는 것을 아는 존재가
그것을 모르던 조상으로부터 발생하였다(1 967: 68). 자아인식이 발생하는 결정적 단계는 약 8 만년 전 네안데르탈인이 치루었던 매장 의식에서 처음으로 찾아볼 수 있는 것이다 (2.5, 5.7). 로 렌츠 (Lorenz 1977) 는 다음과 같이 말하였다. 인간의 이해를 전혀 불허하는 장벽들 중 가장 신비스러운 장벽, 그것은 우리 인격의 부정할 수 없는 하나됨을 가르는 장벽이다 . 그 장벽은 우 리 몸안에서 일어나는 객관적이고 확인할 수 있는 생리학적 사건을 우리 의 주관적 경험으로부터 분리시킨다. 유아의 의식으로부터 소아에서의 자아 의식으로의 점진적 발달은 호미니드에 있어 자아의식의 진화적 출현의 좋은 모델이 된다. 침팬지 의 자신에 대한 원시적인 지식(다른 하동 영장류에는 없음)의 증거도 있 댜 침팬지는 거울 속의 자신을 인식하고 거울을 통하여 자신의 얼굴 에 묻어 있는 붉은 표딱지를 떼어 낸다 (Gallu p 1977). 이와 같은 거울에 서의 인식은 약 한 살반짜리 어린아이가 수행하는 것이다 (Ams t erdam 1972). 진화 과정에서 죽음의 인식이 고통스럽게 경험되기 훨씬 이전에 원시적 자아인식이 있었다고 생각한다. 이 죽음의 인식은 각종의 종교 적 의식들로 나타났고 매장의식으로 표현되었다. 마찬가지로 어린아이 의 자아에 대한 지식은 죽음의 인식에 대한 최초의 고통스런 경험에 앞선댜 자아의식을 도식적으로 나타내는 것은 이해에 도움이 된다. 그림 8. 5 를 발전시킨 것이 될 것이다. 뇌-마음 관계의 공식적 정보흐름 도표에(그림 9.5) 의식적 경험의 세계인 세계 2 의 세 가지 주요소가 있 다. 의식적 경험의 세계이다 외적 감각과 내적 감각 구획은 중앙구획 과 관계된다. 중앙 구획은 심리학적, 철학적 또는 종교적 논의의 종류 에 따라 정신, 자아 또는 영혼이라고 이름붙여질 수 있는 것이다.
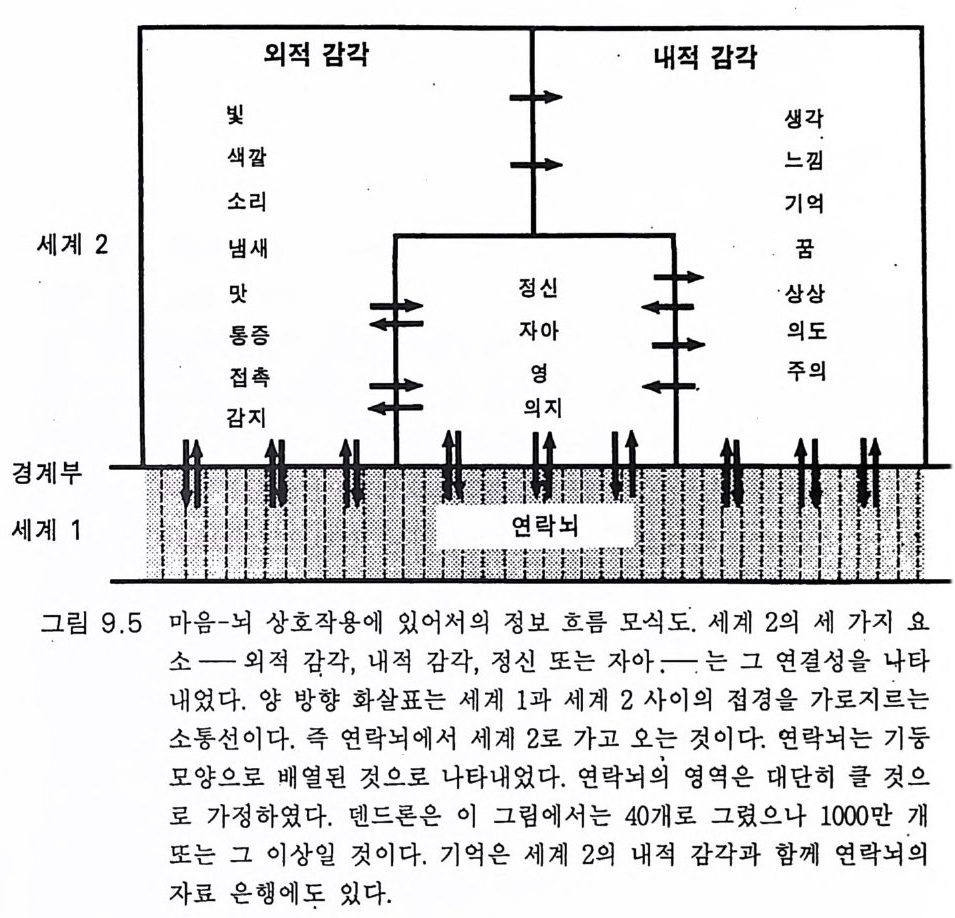 외적 감각 내적 감각
외적 감각 내적 감각
래크 (Lack 1961, p l28) 와 로렌츠 (Lorenz lr;r /7, p l70) 는 둘 다 자아의식 의 진화적 출현에 있어 영과 신체 사이에는 건너갈 수 없는 간격 또늠 심연이 있다고 말하였다. 하지만 종국적으로는 그림 9.5 에 나타냈듯이 정신 또는 영혼의 완전한 . 출현을 이루는 중심적 핵심의 창조와 발달을 상정해야만 한다. 호미니드 진화의 계통발생학적 과정에 있어 그림 8.5 와 9 . 5 에 나타나 있는 상황 사이에는 사람의 유아에서 소아를 거쳐 성인 에 이르는 개체발생학적 과정에서와 마찬가지호모든 종류의 변이가 있
었다고 생각된다 . 그러나 그것은 아직 우리의 이해를 초월하는 것이다. 9.5 자아의 단일성 개인의 최초의 기억으로부터 기억의 계속성이 유지된다는 것에 의 해 인식할 수 있듯이 심적 단일성이 주관적으로 존재한다는 것은 사람 의 보편적인 경험이다. 이것이 자아라는 개념의 기초가 되는 것이다 (Pen field 1975). 가장 간단히 설명하자면 단일성의 경험은 개체의 유일 한 뇌에서 그 단일성이 유래되는 사실에 의하여 완전하게 설명된다. 그러나 그 뇌에는 거의 무한대의 신경 활성 양상이 있는 것이다. 어떻 게 이 극단적인 다양성을 순간순간 통일된 경험으로 통합하는가? 마 음의 성질에 관한 의문 그리고 뇌의 신경 활동과의 관계를 조사한 후 우탈 (U ttal 1978, p 208) 은 <작용 방식 이 란 관점 에 서 보자면 마음은 선 천 적으로 총체적이고, 부분으로 나눌 수 없는 것으로 생각된다 . 심리학 은 분명히 아직 이 문제룰 파악하지 못하고 있다>고 결론지었다. 1977 년에 포퍼 Po ppe r 와 에클스 Eccles 는 자아의식적 마음은 신경활동 을 수동적으로 읽어내기만 하는것이 아니고 융(J un g 1978) 이 말한 탐 조등 비유처럼 능동적 탐색 활동이 있다는 가설을 수립하였다 . 자아의 식 앞에 죽시죽시 복합적 신경 과정 전체를 잠재적으로 나타내거나 묘 사한다. 그리고 주의 집중과 선택 그리고 홍미 또는 의도에 따라 이룰 연락뇌Liai son B rain에서 일어나는 방대한 활동의 합주로부터 선택할 수 있다. 그래서 이것 저것을 탐색하게 되고 연락뇌의 여러 영역의 판 독 결과를 종합한다. 이 같은 방식으로 자아의식적 마음은 경험을 하 나로 통합한다• 이 상호작용은 8. 8 에서 제시한 방식으로 쌍방 통행 과 정이다. 더욱이 이들이 빈번히 활성화되는 신경 양상은 시냅스의 장기간 강
화를 초래할 수 있고 (7.5 .1 참조) 이는 신경회로를 안정화시킨다. 그래서 대뇌피질에 기억 저장을 구축하게 될 것이고 자아의 단일성을 보장하 는 기초가 된다 . 그 같은 기억이 없으면 단일성의 경험은 있을 수 없다 . 9.6 교차연결 절개술 유명한 심리학자 페크너 Fechner 는 만일 뇌의 두 쪽을 중앙에서 길이 로 반씩 나눈다면 인간이 중복되었을 때와 비슷한 일이 일어날 것이라 고 믿었다 (Zan g w i ll1974). 한편 맥도걸 (McDou g all1911) 은 의식의 단일 성은 해부학적 연결에 의해 조건지어졌다는 것을 믿지 않았다 . 장윌 Zan g w ill의 인용에 의하면 그는 뒤에 그가 만일 불치의 병으로 죽게 된 다면, 자기 자신의 뇌량을 갈라서 이 문제에 대한 해답을 얻자고 제안 하기까지 하였다. 만일 페크너와 생리학자들이 옳다면 그는 <분리된 성격 > 을 경험하게 될 것이다. 만일 그가 옳았다면 그의 의식은 통일된 채로 있었을 것이고 이는 맥도걸이 영혼이라고 주장한 것을 확증하게 될 것 이 다. 이 들 이 론적 추론은 스페 리 (Sp e rr y 1969) 와 그 동료들의 철 저한 연구로 인하여 재고할 수 있게 되었다. 그들은 조절되지 않는 심 한 전간을 치료하기 위해 완전한 교차연결 절개술을 받은 환자를 대상 으로 하였다. 사람에게 이와 같은 수술을 시행하기 이전에 고양이에서 교차연결 절제술의 효과에 관한 주의 깊은 연구가 있었음을 주목할 필 요가 있다 . 이 수술로 심한 기능장애는 없었다 (M y ers 1961). 이 수술은 뇌량을 절개하는 것으로 뇌량은 양쪽 대뇌반구를 연결하 는 신경섬유(약 2 억 개)의 대로이다(그림 5.1, 6.1 , 9.6). 뇌의 하부에 있어 대뇌반구 연결은 정상임을 알아둘 필요가 있다. 또 두 대뇌반구는 수 술 전에는 실험 대상의 모든 대뇌 활동이 밀접하게 연관되어 있었다는 것을 알아두어야 한다. 그리고 각각의 대뇌반구는 연대적 행위의 수년
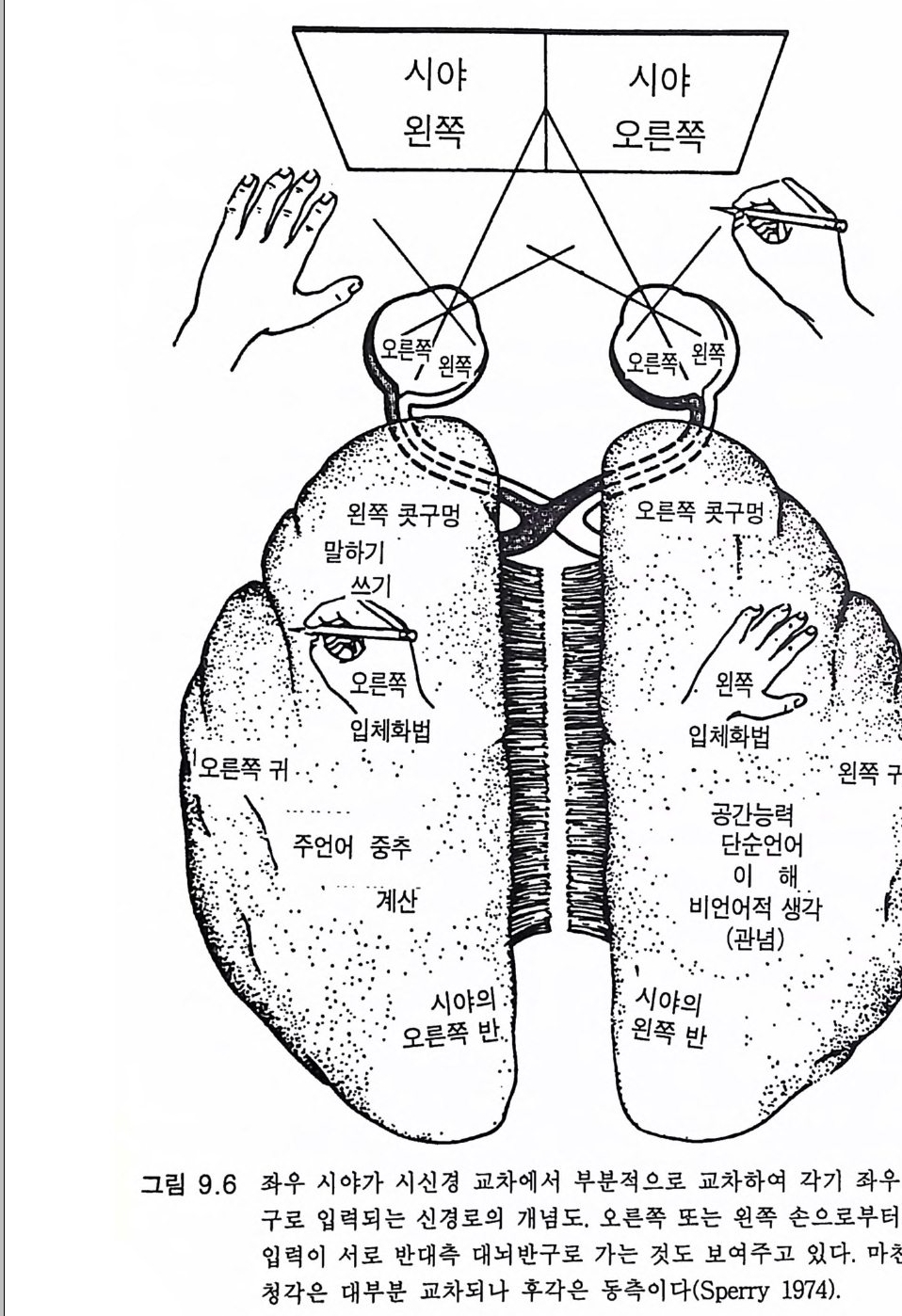 시야 시야
시야 시야
간의 기억을 가지고 있다는 것을 염두에 두어야 한다 . 다행히도 뇌량을 절개하여도 환자에게는 장애가 크지 않다. 따라서 스페리가 대단히 특이한 연구를 수행하기 전까지는 별로 뚜렷한 것이 발견되지 않았었다 (S p e rry 등 1969). 교차연결 절제술에 의해 손상받지 않은 뇌의 하위에서의 교차연결은 정상이기 때문에 서기, 걷기, 다이 빙, 수영 그리고 수면과 각성 둥 신체의 일상적인 행위는 정상적으로 연관된다 스페리와 그 동료들이 개발한 시험법에서는 이 <분리된 뇌 s plit br ai n > 환자의 한쪽 또는 다른 쪽 대뇌반구에 정보를 주입할 수 있고 각각의 대뇌반구 반응을 독립적으로 관찰할 수 있다. 기본적으로 좌 또는 우측 시야룰 통하여 뇌에 정보를 주입한다(그림 9.6). 실험 대상은 중심 점 에 고정 하고 단어 등의 신호를 0 .1초 동안 반짝(섬 광) 비 추어 줌 으로써 눈의 움직임에 의한 시야의 변화를 배제하였다. 7 명을 대상으 로 좌측 대뇌반구의 언어영역(그림 4.3) 을 연구하였다. 그것은 항상 우 세 대뇌반구였다. 이 실험에서 발견한 가장 주목할 결과는 우측 대뇌 반구(이른바 열세 대뇌반구)에서의 모든 신경 활성은 언어 표현을 하는 실험 대상과 연결되어 있지 않다는 것이다. 이 말하는 실험 대상은 우 세 대뇌반구인 좌측 대뇌반구에서의 신경 활성과 연락이 되어 있다 . 단지 우세 대뇌반구를 통해서만 실험 대상은 언어로 의사 소통될 수 있댜 더욱이, 수술 전과 꼭 같은 것으로 인식되는 의식적 존재 또는 자아는 오직 이 우세 반구와 연락되어 있다. 스페리는 다음과 같이 말 하였다. 좌측뿐만 아니라 단절된 대뇌반구들은 각각 자신의 사건을 의식하지 않는 것처럼 행동하였다. 바꾸어 말하면 각 반쪽 뇌는 대부분 분리된, 자 신의 개인적 감지, 학습 및 기억 경험을 가진 인지적 영역을 가지는데 이 모든 것은 분명히 다른 쪽 대뇌반구에 있어 대옹하는 사건들을 알지 못
하는 것으로 보인다(1 982: 1224). 그림 9 . 6 은 스페리가 수년전에 그린 것이다. 이는 분리된 뇌 이야기 전체를 논하는 기초로서 아직도 가치가 있다. 이 도형에는 좌우측 시 야를 교차되는 시각피질에 고도의 선택적 투사로 나타내었고 R 또는 L 로 표시하였다(그림 6.1 참조). 도형에는 취각의 엄밀한 일측성 투사도 보이고 청각이 주로 교차투사됨(그림 4.6 참조)도 보인다. 손의 운동 및 감각 신경의 교차지배도 나타내고 있고 수학적 계산에 대해 좌측 대뇌 반구가 우세함도 보여주고 있다. 우측 대뇌반구는 단지 대단히 간단한 덧셈만을 수행할 수 있다. 우측 대뇌반구는 고도로 발달한 뇌라고 말할 수 있지만 자신을 언 어로 표현하지 못하고 우리가 인지할 수 있는 의식의 어떤 경험도 나 타낼 수 없댜 스페리(1 982 : 1224) 는 <좌측뿐만 아니라 단절된 대뇌반 구들은 각각 자신의 각기 다른 감지, 심적 영상, 연합 및 아이디어를 이용하는 것 같다>고 추론하였다. 우측 대뇌반구에 있어서의 의식은 언어적 표현의 결여로 인해 잘 알 수 없다. 한편 좌측 대뇌반구는 정상적인 언어능을 갖고 그래서 뇌 량 절개 이전의 모든 과거 기억을 가지고 자아의 이전의 존재와의 연 합으로 인식할 수 있다. 이 관점에 따르면 말하는 마음으로부터 언어 로 소통될 수 없는 말 못하는 마음이 분리되어 나와서 있기는 있으되 벙어리 또는 실어증이 된다. 일반적으로 우세 대뇌반구는 모든 묘사와 반응에서 미세한 표상적 구체화라는 점에 전문화되었다. 즉 이것은 분석적이고 순서적이다. 또 이것은 더하기, 빼기, 곱하기 둥 전자계산기 같은 기능을 할 수 있다. 그러나 물론 그 우세성은 언어적 및 관념적 능력과 자아의식과의 연락 으로 인한 것이다. 열세 대뇌반구는 이 같은 관점에서의 결여로 인하 여 열세라고 말하나 이는 여러 중요한 성질에 있어 뛰어나다. 특히 강
 우위 반구 열세 반구
우위 반구 열세 반구
력하게 발달한 회화적 및 구조 감각을 가진공간적 능력의 관점에서는 뛰어났다(그림 9.7 ). 예를 들면 왼손의 계획을 작성하는 열세 대뇌반구 는 모든 종류의 기하학적 그리고 투시화법에 있어 대단히 우수하다. 이 우월성은 색조각 맞추기, 죽 모자이크 그림맞추기로도 증명된다. 우세 대뇌반구는 이런 종류의 단순한 과업도 수행하지 못한다. 그리고 복사장애로 나타나듯이 회화적 그리고 구조적 감각에 있어 문맹이나 마찬가지이다 . 우세 대뇌반구는 수학적 반구이지 기하학적 반구는 아 니다(그림 9.7). 섬광시험법의 모든 미세한 방법들은 다른 기법으로 개선되었다 (Sp erry 둥 1979). 즉 오른쪽 눈에 안과적 장치를 장착한 콘택트 렌즈를 끼워 눈을 어떤 방향으로 움직이더라도 왼쪽 시야로부터는 입력이 되 지 않도록 하는 것이다. 이 같은 방법으로 두 시간까지도 시험할 수가
있다 그리고 섬광시험법보다 훨씬 복잡한 검사도 가능하다. 이 검사 는 오른쪽(열세) 대뇌반구가 복잡한 시각영상을 이해하는 능력을 고려 한 것으로 왼손의 적당한 반옹으로 나타내게 하였다. 시험 대상의 왼 쪽 눈은 가렸고 오른쪽 눈의 콘택트 렌즈에 장착한 기구는 왼쪽 시야 의 것만 망막에 입력되도록 하였다. 그래서 오른쪽 시야로부터의 입력 은 완전히 제거하여 의식적 시각 경험이라는 관점에서 장님이라 할 수 있댜 우측 대뇌반구의 회화 이해는 예를 들면 고양이의 그림과 그 밑에 고양이와 개라는 단어가 있는 구림으로 검사 절차를 꾸민 것이다. 검 사대상은 왼손으로 적절한 단어를 가리키게 된다 . 마찬가지로 두 개의 그림이 있을 때 즉 컵과 칼 그리고 그 밑에 해당하는 단어가 < 컵 > 하 나만 있을 때 왼손으로 컵을 가리키게 한다. 더 복잡한 그림 식별 검 사로는 풍경화 밑에 맞는 이름과 톨린 이름을 주는 것이다. 예를 들면 그림 밑에 <여름>과 <겨울>이라는 이름을 주면 풍경화를 옳게 식별 하고 <여름> 대신 <겨울>이라는 단어를 가리키게 된다. 열세 대뇌반 구에 대한 회화적 및·언어적 과업의 이 같은 지적 수행능에도 불구하 고 우측 대뇌반구는 간단한 것일지라도 문장을 완성시킬 수 없다. 침 팬지의 언어 검사로서 (4 .4 참조) 얻어진 대부분의 증거도 이와 유사한 데, 침팬지는 애매한 요구는 할지라도 문장 완성 능력은 없다. 이는 물 론 열세 대뇌반구나 침팬지의 뇌에는 어의학적 능력을 나타내는 베르 니케 영역이 없다는 것에 기인한다. 이들 대단히 철저한 검사 절차 (S p e rry 동 1979) 로 교차연결 절제술 후 우측대뇌반구가 상당한 단어 청취능력이 있음을 알았고 명령을 인 식하는 능력과 시각과 청각으로 주어진 단어와 시각적으로 주어진 그 림을 관계짓는 능력이 있음을 알았다. 특히 일반적인 실험 조건에서 혼 히 주어지는 그림 과제의 식별에 특히 효과적이었다. 또 놀라운 것은 우측 대뇌반구가 행위 명칭에서와 마찬가지로 동사에도 매우 효과적으
로 반응한다는 점이다. 동사에 대한 반응은 섬광검사법으로는 감별되 지 않았다 이들 모든 언어 이해 능력에도 불구하고 우측 대뇌반구는 문어 및 구어적 표현능은 극도로 결여되어 있어 실질적으로는 없는 것 과 같댜 또 정확하게 순서대로 기억해야 하는 여러 사항이 포함된 지 시 문장을 이해하지 못한다. 고도로 의미 있는 소견은 우측 대뇌반구의 언어 이해 능력과 표현 능력 사이에 큰 차이가 있다는 점이다. 스페리 등(1 979) 은 두 교차연결 절제 환자에서 우측 대뇌반구에 자 아의식 및 일반적 사회적 인식의 관점을 검사하도록 설계한 연구 결과 를 보고하였다. 이 검사에서는 여러 종류의 인물 사진과 잘 알려진 사 물 및 풍경의 그림을 조합하여 환자의 왼쪽 시야에 제시하였다. 따라 서 우측 대뇌반구에 한정된다(그림 9.6 ) . 실험 대상은 자신의 사진을 포 함하여 낯익은 사진을 항상 식별하게 된다 . 그러나 그것이 무엇이었는 지를 구체적으로 알지 못한다. 그리고 우측 대뇌반구의 식별을 언어로 표현하려면 상당한 일련의 교시적인 암시를 주어야만 했다 . 이런 언어 적 표현은 아마도 좌측 대뇌반구에 의한 것이다. 식별능에 있어서의 양 대뇌반구의 동질성에 관한 이들의 극적인 결론은 비판의 여지가 있 댜 실험 설계에서 나타났듯이 실험 대상의 반응을 낙관적으로 과대 평가한 결과일 것이라는 점이다 . 어찌되었건 우측 대뇌반구에도 제한 적이나마 자아의식이 존재한다는 좋은 증거가 되는 것이다. 마음 그리고 자아의식적 마음의 존재에 관한 이들 검사는 비교적 단순한 회화적 및 감정적 수준에 있다 . 아직도 우측 대뇌반구가 완전 한 자아의식적 존재인가는 의문이다. 예를 들면 우측 뇌는 장래를 걱 정하고 계획하는가? 어떤 가치 체계를 근거로 토의하고 결정을 내리 는가? 이것들은 보통 이해되는 인간성의 . 갖추어야 할 기본이다 (S tr a wson 1959; Po pp er 와 Eccles 1977, p 31, 33). 이 제 의 식 적 으로 경 험 한 자 아의 유일성에 관한 이들 발견들의 중요성을 살펴보려 한다. 우측 대뇌반구에 자아의식이 존재한다는 것은 스페리 둥의 이들 연
구에서 실마리를 찾을 수 있다 . 그러나 이는 제한적인 것이고 위에서 언급한 기준에 비추어 우측 대뇌반구가 인간성을 갖는다고 규정하지 못한댜 따라서 교차연결 절제술은 자아의식적 마음으로부터 한 조각 을 떼어내었지만 그 사람은 표면상 홈이 없는 채로 남아 전적으로 좌 측 대뇌반구만의 연합으로 정신적 단일성이 온전하게 보존된다. 이는 아마도 맥도걸을 즐겁게 한 결과이다. 그러나 우측 대뇌반구에서 나오 는 감정적 반응이 분리되지 않은 변연계를 통하여 좌측 대뇌반구에 관 여하게 될 것이라는 점은 인정해야 할 것이다. 따라서 그 사람은 감정 적으로 아직 우측 대뇌반구와 연결된 채로 있는 것임을 인정해야 할 것이다. 9.7 대뇌 비대칭성의 진화적 의의 대뇌의 비대칭성을 지나치게 강조한 감이 없지 않다 . 이제 진화상의 의의를 살펴보려 한다. 앞서 호미니드 진화 과정에서의 광범한 대뇌 변 화를 언급했댜 언어가 대뇌 비대칭성의 발달에 의존한다는 것은 제 4 장 에서 분명히 했다. 그러나 제 5-8 장에서는 비대칭성이 별로 중요한 것 같지 않다고 하였다. 호미니드계의 진화적 성취에 기여한 뇌에서의 핵심 적 변화를 정의하기는 힘들다. 그러나 대뇌 비대칭성의 진화가 호미니드 진화의 기본 과정이었을 것이라고 하는 홍미로운 가능성이 존재한다. 이 같은 전망은 제르 레비J erre Le vy가 두 대뇌반구의 상호보완적인 기능을 연관시켜 잘 표현하였다. 사람의 대뇌반구는 공생적 관계로 존재하며 그 안에서 양 반구의 행위 의 능력이나 동기는 상호보완적인 것이다. 양측 뇌의 각각은 다른 쪽 뇌 가 어렵다거나 마땅치 못하다고 여기는 인지적 과업을 수행할 능력이 있
고 그 수행을 선택한다. 이 기능들의 두 벌의 성질을 고려하면 논리적으 로 양립할 수 없는 것처럼 보인다 . 우측 대뇌반구는 공간상에서 합성하고 좌측 대뇌반구는 시간상에서 분석한다. 우측 대뇌반구는 개념적 유사성 은 제외하고 시각적 유사성을 감지한다. 좌측 대뇌반구는 그 반대이다. 우측 대뇌반구는 형체를 감지하고 좌측은 그 상세함을 감지한다. 우측 대 뇌반구는 감각 입력을 영상의 형태로 부호를 만들고 좌측은 언어적 기술 의 형태로 만든다. 우측 대뇌반구는 음성학적 분석기가 없고 좌측은 형체 합성 기가 없다 (1974: 167). 스페리 (1982) 는 이제까지는 비교적 발달이 부진하다고 생각했던 우 측 대뇌반구의 우월한 수행능을 강조하여 다음과 같이 말하였다. 우측 대뇌반구의 특기는 모두가 성질상 비언어적이고 비수학적이며 비 순서적이다 . 다분히 영상적이고 공간적이다. 예를 들면 얼굴의 판독, 설계 룰 대체적 모체의 형태로 맞추기, 작은 반원에서 전체 원의 크기를 판단하 기, 비서술적인 모양의 재생 및 감별, 심적 공간적 변환, 음악적 음계의 감 벌 조각의 크기와 모양을 계통별로 가려내기, 부분적 집합에서 전체를 파 악하기 그리고 기하학적 원칙의 직관적 지각과 이해 둥이다 (1982: 1225). 그림 9.7 은 대뇌반구의 이들 특성화를 대단히 간단하게 요약하여 나 타낸 것이다. 상단선에 우측 대뇌반구는 의식과 연락이 이루어져야 되 나 좌측 대뇌반구에서와 같은 자아의식과의 연락은 없다고 되어 있다. 이는 자아라는 것은 좌측 대뇌반구 특유의 것임을 말하며 9.6 에서 설 명한 바 있다.
9.8 신피질의 진화적 기원과 조직 발생이 늦다는 발생학적 및 구조적 증거:신신피질 9.8 . l 신신피질 신피질의 수초화(그림 4.8, 4.9 ) 시기는 플레흐지히 (Flechs ig 1920) 에 따르면 진화 연대를 나타낸다고 한다 . 또 수초화의 지연은 뉴런과 수 상돌기 발달의 지연과 상관관계가 있고 접합부 입력 발달의 지연과도 상관될 가능성 이 있다 (Gesch wi nd 1965, p.2 73). 영역 39 와 40 이 특히 홍미 있는 것이다. 유인원의 뇌(그림 4.5) 와 사 람의 뇌(그림 4 .4)에 대한 브로드만 지도를 비교하면 유인원의 영 역 39 와 40 은 혼적에 지나지 않는다 . 늦은 진화적 발달은 수초화의 지연과 좋은 상관관계를 보인다(그림 4.8, 4.9 ). 그림 4 . 8 에서 수초화가 늦은 전두전엽과 하측두엽의 넓은 영역을 볼 수 있댜 만일 이들 영역이 모두 진화적 발달이 늦은 것이라면 피질증 대의 상당 비율이 이들 새로운 영역에 할당되었을 것이다 . 이를 신신 피질이라고 할 수 있다. 이 사실은 또 다른 호미니드 진화에 있어 일 차 감각 및 운동 영역의 크기가 거의 변하지 않았다는 것이다. 예를 들면 그림 4 .4와 4.5 에 있는 영 역 17 이 다. 9.8.2 인지 기능 신신피질의 영역들은 호미니드 진화의 특이적 성질인 특별한 인지 적 기능들을 위하여 진화상 발달되었다고 제안되었다 (S p e rry 1982). 이 제까지 사람의 뇌의 우수한 인지적 능력은 그 크기에 의한다고 믿었 댜 이는 창조적 사고가 결여된 조잡한 생각이다. 한편 지금은 사람의 뇌의 우수한 기능은 신피질에서 유래되는 것이라고 주장되고 있다. 신
신피질은 유인원을 모델로 살펴본 바와 (그림 4.5 ) 마찬가지로 가장 발 달한 호미노이드에서조차도 무시할 정도였을 것이다. 영역 39 와 40 은 신신피질에서 가장 분명하게 정의되는 영역이다. 그러나 중전두전엽과 하측두엽(그림 4.8, 4.9) 도 신신피질이라 할 수 있다 . 이들 신신피질 영 역들의 소모듈에는 틀림없이 모종의 알 수 없는 미세구조적 특징이 있 울 것이라고 생각된다. 이것은 신경해부학자들이 도전해 볼 큰 과제이 댜 신신피질 영역들은 틀림없이 여러 비대칭성의 구조적 기초가 될 것이다. 예를 들어 좌측의 언어(그림 4.3) 그리고 우측의 공간 구성 및 음악 영역(그림 9 .1 ,9 . 7) 이다. 9.8.3 신신피질의 개체 발생 인지적 피질 영역의 개체발생학적 발달의 특성은 그림 9 . 8 과 관련하 여 생각해 볼 수 있다. 이는 97 일된 원숭이 태아의 신경관의 그림으로 서(Raki c 1972) 신피질 생성 과정에 있는 것이다 . 사람 태아의 신경관 도 비슷하리라고 생각되나 다만 중요한 차이는 신피질에 더하여 신신 피질의 신경 발생에 관계되는 특별한 부분이 있을 것이라는 점이다. 그 같은 부분에서는 신경상피세포의 미숙 신경세포를 생성하기 위한 세포 분열이 늦올 것으로 생각한다. 이들 신경세포는 특정 신신피질 영역을 구성하는 과업에 적합한 넓은 범위의 특별한 유전적 기질을 가 지고 있을 것이라 생각한다 . 이는 아마도 고도의 특이적 기능을 나타 내기 위한 신경세포의 단세포군을 생성할 것이다 (Meller 와 Tatz le ff 1975). 이들 신경세포는 마운트캐슬 (Moun tc as tl e 1978) 이 제안한 바와 같이 작은 수직의 단세포 기둥으로 배열된다. 따라서 호미니드 진화에 있어 호미노이드의 신경관은 그것의 신피 질 생성과 함께 신신피질의 생성을 위한 조각들을 보유하게 되었을 것 이다. 이들 조각들은 오스트랄로피테신에서는 작았으리라고 생각되고
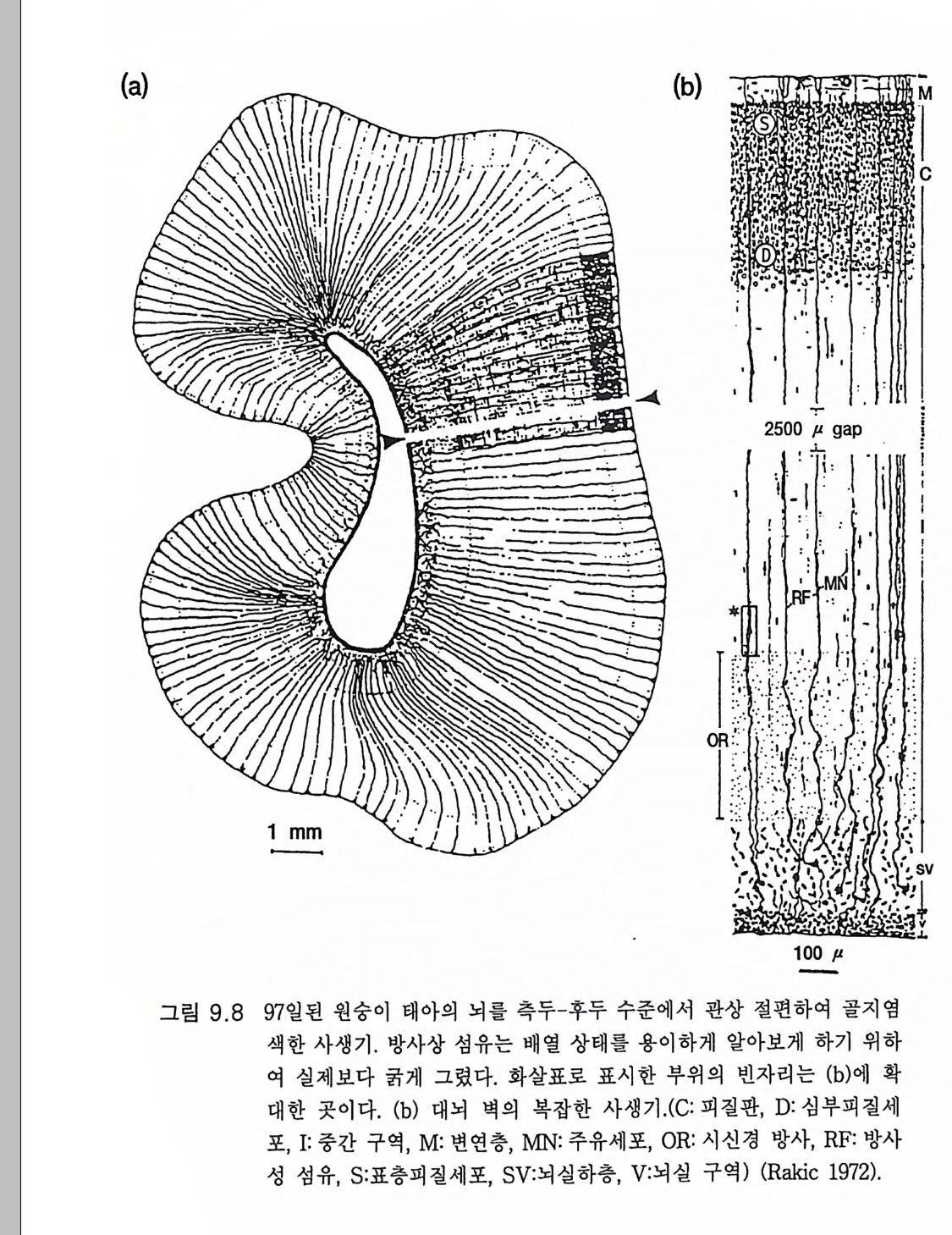 (a) (b)
(a) (b)
호모 하빌리스의 크게 증대한 뇌는 커다란 신신피질 생성 부위에 의하 여 이루어졌다고 생각된다. 호미니드 진화의 이후의 단계에서 이들 부 위는 신피질 생성 부위보다 더 큰것으로 자랐을 것이다. 결국 호모 사 피엔스 사피엔스에 와서는 신경관은 대부분 신신피질 생성 부위로 대 치되었을 것이다. 그러나 이러한 상대적 크기가 신신피질 생성 부위의 유일한 성질은 아니다. 오히려 대단히 다양한 종류의 인지적 기능에 따라 광범위한 전문화가 된 점이 매우 독특한 특질이다. 신신피질의 후기 발달설은 사람의 신경관의 신경상피세포의 커다란 조각들에서 세포 분열이 지연됨에 의하여 증명되어야 한다. 이는 아직 연구된 바 없댜 먼 장래일지라도 신경 발생의 핵심 문제를 해결할 수 있을 것이 다. 문제란 특유의 인지적 기능 그리고 이들 기능의 비대칭성 분포의 특성을 나타내는 특수한 내적 연결성과 관계되는 것이다. 9.8.4 신신피질의 영양적 발달 인지적 기능의 제한적 변환이 어린 나이에 일어날 수 있다는 것을 상기할 필요가 있다. 예를 들어 유아 시절에 베르니케 영역을 파괴한 후에 언어가 우측 대뇌반구로 변환되는 것이다(Mil ner 1974). 제 8 장의 8. 8 에서 단위 덴드론과 단위 사이콘 간의 고정된 관계에 관한 가설을 세웠다. 사춘기 이전에는 융통성이 있을 것으로 생각하며 나이가 어릴 수록 그 융통성이 커진다고 생각한다. 오그덴 (Og den 1988) 은 이 영양적 보상에 관한 훌륭한 예를 기술하였 댜 결정적 사춘기 동안에 거의 쓸모가 없게 되고 하루에도 수 차례씩 의 심한 전간 발작의 원인이 되었던 좌측 대뇌반구의 심한 손상을 유 아 시절에 받았던 두 명의 환자에게 17 세와 18 세 때에 거의 완전한 좌 측 대뇌반구 절제술을 시행하였다. 놀랍게도 환자 상태는 개선되고 이 제 나이가 46 세와 34 세가 되어서도 언어적 I Q와 공간 시각적 과업에
있어 80% 를 IQ 수준으로 우측 대뇌반구가 계속 수행하고 있었다. 그 두 환자는 모두 정상적인 직장 생활을 하여 잘 적응하고 있다. 유아 시절의 심한 좌측 대뇌반구 손상 때문에 우측 대뇌반구가 언어반구가 되었다는 것은 놀라운 일이다. 그러나 정상적 우측 대뇌반구 기능에 있어서나 습득했던 언어기능에 있어 모두 결함이 있었다. 이 같은 사 실은 밀너(Mi lner 1974) 도 기술하였다. 9,8,5 신신피질 기능의 바대칭성 호미노이드의 기능적 대칭성에 있어 신피질 수지의 특징은 모든 신 피질 기능의 중복에 있다고 하였다. 호미니드 진화에 있어서는 새로운 진화적 발달에 따른 많은 요구를 수행하는 데 필요한 정교한 설계의 더 많은 신경회로에 대한 압도적 요구가 있었다고 생각한다. 특히 고 위 수준의 언어에 있어서이다(그림 4.1 ). 따라서 중복되는 기능의 신피 질 구성은 더 이상 되지 않게 진화적 작전이 수립되었다. 대신에 게슈 윈드 (1965) 와 렌네버그 (1967) 가 기술하였듯이 출생시에 지연 성숙에 따 라 한 가지 또는 다른 인지적 기능에 대한 왼쪽 또는 오른쪽 편파성이 있었을 것이다 . 생애 초반 동안의 인지적 기능의 변환으로 표현되는 영양적 조절과 제 7 장 7.5 .1과 7 . 6 에서 설명한 학습의 기초가 되는 전 생 애를 통한 가소성 변화는 분명하게 구별해야만 한다 . 모차르트는 사춘기 전에 있어서 신신피질의 영양적 발달에 관한 최 상의 예가 된다. 그의 생애를 보면 생의 초기부터 그의 뇌는 음악에 대한 극단적 편파성을 나타내었다. 그의 아버지도 이를 인식하였다. 부단한 음악교육으로 음악의 인지적 기능에 대한 신신피질의 사춘기 전 성숙에 따른 철저한 자체 편성이 이루어졌을 것이다. 그래서 전 생 애를 통한 가장 훌륭한 음악적 뇌가 창조되었다. 물론 평범한 신신피 질을 가진 뇌를 어버이가 지나치게 교육적 과도 훈련을 시킬 위험이
상존한다. 신신피질에 있어 개체발생학적으로 발달한 편파성을 실감하 는 사춘기 전 시절의 결정적 시기를 인식함에 있어 부모와 선생은 슬 기롭게 노력해야 한다는 것을 여기서 지적하고자 하는 것이다. 이것은 언어, 음악, 수학, 시, 조각, 철학, 종교 그 어느 경우도 될 수 있다. 9.8.6 영양적 신신피질 발달 가설 신신피질의 덴드론(제 8 장 8.8) 은 점진적으로 변형된다는 가설을 세울 수 있다 이들 덴드론은 여러 인지적 기능에 따라 비대칭적으로 편파 적인 지연성 개체 발생에 의해 서서히 구축된다. 정상적으로 사춘기 전 시기예는 이들 편파성은 보다 미묘하고 정선된 반응을 일으키는 사 용으로 점점 발달한다. 자아 창조와 자체 편성의 영양적 과정에 의해 신신피질과 상호작용 하는 경험 세계의 관계 아래 인지적 기능들은 극히 다양하게 발달된 댜 제 8 장 8. 8 에서 대뇌 소모듈에 있어 단위 사이콘과 이에 연계되는 단위 덴드론 사이의 고정관계에 관한 가설을 들었다. 이 영구적 관계 는 발육의 사춘기 전 시기 동안의 영양적 상호작용에 의해 형성된다. 정상적으로 발견되는 비대칭성은 한쪽 또는 다른 쪽에 개체발생학적 으로 구축된 편파성의 영양적 발달에 의해 초래된다 (Lenneber g 1967). 이 가설은 어린 나이 때 한쪽 대뇌반구에 심한 손상을 입었을 때 성 인의 비대칭 양상에서 이탈해 나가는 과정을 설명할 수 있다. 사춘기 가 되면(약 14 세) 기능적 비대칭성은 고정된다. 9.8.7 신신피 질의 요약 호미니드 진화의 지대한 성취는 피질의 잠재적 능력을 거의 배가하 는 이 비대칭성 수지에 의하여 확고하게 되었다 (Le vy 1977). 피질의 비
대칭성은 그 성취의 핵심점이다. 감각과 운동 기능을 가진 구 old 신피 질은 그 대칭적 기능을 유지한 채로 남아 있게 된다. 신피질 비대칭성 의 최고 진화적 유익성을 계산할 수 있다. 호모의 신피질 = 3.2X 침팬 지 신피질이다. 그래서 모든 새로운 피질이 두 배로 복제되면 대뇌 기 능은 3.2 배로 증가한다. 만일 새로이 추가된 것이 단순 복제가 아니고 비대칭성이라면 대뇌의 능력은 5 .4배 (1+2.2+2.2) 로 증가한다. 따라서 비대칭성이라는 작전으로 산과적 장해를 일으키지 않고도 신신피질은 능력의 큰 증대를 이룰 수 있는 것이다(표 5.2 ) . 신신피질에는 다섯 가지 특수한 특징이 있다. ® 계통발생학적으로 최후로 출현한 것이고 특별한 호미니드에서의 발 달이다 ® 개체발생학적으로 지연성 수초화(그림 4.8, 4.9) 와 지연성 수상돌기 및 시냅스의 발달에 의해 최후로 성숙된다. ® 언어 및 공간 시각 그리고 음악 둥에 있어서 기능적 비대칭성(그림 9.7) 이 있다. @ 어릴 때에는 손상을 보상하는 등의 성숙상의 가소성이 있다. ® 신신피질의 활성화는 의식, 자아의식, 생각, 기억, 느낌, 상상, 창조 둥 넓은 범위의 인지적 기능을 수반한다.
제 10 장 인간의 인격 이제까지의 아홉 장에서, 특히 뇌라는 관점에서 호미니드 진화의 기 본적 특성을 살펴보았다. 한편으로는 우연한 유전적 돌연변이가 중심 역할을 하고 다른 한편으로는 자연도태의 <체 s i eve > 를 이용해 걸러진 다 s ifti n g고 표현되는데 핵심적 역할을 하는 철저하게 다윈적인 설명이 었다 이 같은 방식으로 인간 뇌의 발달을 호모 사피엔스 사피엔스라 는 그 진화적 절정에 이르기까지 그럴싸하게 설명할 수 있었다. 우리 의 과학적 지식이 아직도 상당히 초보 단계이므로 상세한 것을 모르는 것은 당연하다. 그러나 원칙에 있어 뇌의 해부학적 및 기능적 변화를 초래한 진화의 방식 즉 경탄할 또는 초월적이라고까지 할 만한 결과를 가져온 호미니드 진화 과정을 설명할 수 있었다 . 장차 언젠가 호미니드 진화에 비견할 만한 또 다른 생물학적 진화 가 있을 수 있을 것인가라는 질문은 고려할 가치가 없다. 이 책에서 전하려고 하는 것은 호미니드 진화는 영장류 조상의 비할 데 없는 신 경계의 발달에 전적으로 의존한다는 점이다. 호미니드 진화는 결코 반 복되지 않을 세계의 개화기이다. 자연도태는 근본적으로 기회주의적이다. 단지 특정 세대의 생존과
번식을 위한 것이고 기회를 타서 다음 또 다음 세대로 이어진다고 생 각한다 통상적으로 자연도태는 위험한 환경에서 생존하고 번식하는 유기체의 능력에 의존한다고 이해된다. 포퍼 (Po pp er 1981, 1989) 는 이를 수동적 다윈설이라고 하였다 . 이에 비해 생명체가 보다 쾌적한 생태 조건과 생존 양식을 찾아 환경을 능동적으로 탐사하는 것을 탐사적 또 는 능동적 다윈설이라고 한다 . 죽 < 능동적으로 선택하고 선호하는 더 좋은 새로운 주거지와 새로운 생활 방식에 적합하도록 이를 형성하는 도태압력>이다 (Po pp er 1981: 42). 약 35 억년 전의 생명의 기원으로부터 의 유구한 시간을 통하여 생물학적 진화는 전적으로 수동적 다윈 법칙 에 따랐다고 할 수 있다. 능동적 다윈 법칙은 극히 최근에 아마도 2 억 년 전쯤부터 가능했을 것이다 . 왜냐하면 그들의 발견을 효과적으로 개 발하도록 하는 도태 압력에 의해 환경을 탐사할 수 있었던 고도로 발 달한 유기체가 나타났기 때문이다. 포퍼는 이 마음 비슷한 행동을 포 유류, 조류 등 고동동물에서의 의식적 행동의 선두 주자라고 제안하였 댜 이 같은 의식적 행동이 자연도태에서 중요하다는 것은 의심할 여 지가 없다 . 호미니드 진화에 전혀 새로운 현상이 나타났다는 것을 9.2 , 9.4 , 9.5 에 서 설명하였다. 포유류와 조류 등 고등동물의 행동 양상 (8 . 2, 8.3) 으로부 터 알 수 있는 의식 상태가 있었을 뿐 아니라 적어도 호모 사피엔스 네안데르탈리스에서부터 인정되는 사람의 자아의식 상태가 있었다 (9.4 ). 엄밀한 다윈설의 유물론적 학설에 충실한다면 동물 의식의 존재는 8.3 에서 설명했듯이 이미 불가해한 변태였다. 세계 2 의 주관적 경험이 인간 사이의 의사 소통에 의해 객관성을 성취하였음이 그림 9.5 에 있 댜 이는 그림 9.5 의 내적 감각과 외적 감각 밑에 나열한 모든 항목들 의 유의성을 검사하는 데 사용될 수 있다. 또 선택적 전기적 기록 방 법 또는 rCBF 와 PET 주사법을 이용한 연합뇌의 연구법에 의한 연합
된 뇌 상태에 관한 방대한 과학적 연구가 있다. 이들 어떤 검사법도 세계 2( 그림 9 . 5) 의 핵심에 직접적으로 접근하지는 못하며 이는 다분히 철학적 논쟁의 불씨이다. 10.l 인간의 인격 포퍼는 다음과 같이 추론하였다. 자아 sel f란 < 순수한 절대아p ure eg o > 즉 단순히 주체에 불과한 것은 아니댜 이것은 오히려 믿을 수 없을 만큼 풍요하다. 조종사처럼 관찰과 동시에 행동을 취한다 . 괴로워하며 행동하고 과거를 회상한다. 그리고 장 래 를 계획하고 계획표를 작성한다. 예측하고 처리한다 . 이것은 신속하고 연속적으로 또는 거의 동시에 행동의 청원, 계획, 희망 그리고 행동의 중 심인 행동하는 자아의 존재에 대한 생생한 의식을 함유한다. 그리고 이 자아는 대부분 다른 사람, 다른 자아들 그리고 세계 3 과의 상호작용의 덕 분에 그 자아됨을 성취한다. 그리고 이 모든 것은 그의 뇌에서 일어나는 방대한 <활동>과 밀접히 상호작용한다 (1977 : 120). 우리들 모두는 계속적으로 단지 의식만이 아닌 자아의식으로 인격체 임을 경험한다 . 그리고 안다는 것을 안다. <인격p erson> 을 정의함에 있 어 임마누엘 칸트의 찬양할 만한 두 가지 진술을 인용하고자 한다. <인격은 그의 행동에 책임을 지는 주체이다.> 그리고 <인격은 시 간이 다른 때에 자신의 숫자적 동일성을 의식하는 그 무엇이다 . > 이 진술들은 최소한의 것이고 기본적인 것이다. 그리고 엄청나게 확 대될 수 있는 것이다. 인격을 그 뇌와 관계지어 정의함에 있어 칸트에서 더 전진할 수 있
댜 인격을, 우리들을 구성하고 있는 얼굴, 신체, 사지 등의 집합체로 간주하려는 경향이 있다 . 이것이 틀렸다는 것은 쉽게 증명할 수 있다. 예를 들어 사지를 절단하거나 눈을 잃으면 장애자가 될지는 모르지만 자신의 기본적 정체로서 인간 인격은 유지된다 . 내장을 제거하였을 때 도 마찬가지이다. 여러 장기를 부분적 또는 전체를 들어낼 수 있다 . 신 장 이식 심지어 심장 이식 후에도 인간 인격은 변하지 않은 채로 존재 한다 . < 뇌를 이식하면 어떤 일이 벌어질까? > 라고 물을 수 있다. 다행 히도 이런 수술은 현재 불가능하다 . 그러나 지금이라도 머리를 성공적 으로 이식할 수도 있다. 이식된 머리를 소유하고 있는 인격은 지금 취 득한 신체를 소유할 수 있으나 그 반대는 아님을 누가 의심할 수 있는 가! 인간 인격을 가지고 이것이 상상의 실험으로 남아 있기를 우리는 희망할 수 있다. 그러나 이미 이것은 포유류에서 성공적으로 시행되었 댜 뇌와 관계없는 머리의 모든 구조는 이 이식된 소유성에 관련되지 않는다는 것을 알 수 있다 . 예를 들면 눈, 코, 턱, 두개골 등은 신체의 다른 부분과 다를 것이 없는 것이다. 따라서 뇌, 그리고 뇌만이 우리 인성pe rsonhood 의 물질적 기초가 되는 것이라고 결론지울 수 있다 . 뇌를 의식적 인성의 자리로 생각한다면 뇌의 대부분은 필수적인 것 이 아님을 알 수 있다. 예를 들어 소뇌를 제거하면 동작은 중대한 장 애가 있으나 인격은 달리 영향을 받지 않는다. 뇌 주요 부분 죽 대뇌 반구의 경우는 전혀 다르다. 대뇌반구는 동일하지는 않지만 사람의 의 식과 대단히 밀접하게 관계된다(그림 9.6, 9.7). 사람의 95% 는 좌측 대 뇌반구가 우세하다. 이것은 언어 s peeking 반구로서 그 안에서 언어학 적 소통과 특이적으로 연합한다(제 4장). 유아를 제외하고 이것을 제거 하면 인격의 심한 파괴를 초래하나 전멸하지는 않는다. 한편 열세 반 구(통상 우측 반구)를 제거하면 좌측 신체의 동작이 소실(반신불수)되거 나 왼쪽 눈이 실명(반맹)되나 인격은 별로 크게 손상받지 않는다. 뇌의 다른 부위에 손상이 있으면 인격의 장애를 크게 가져올 수도 있다. 이
것은 아마도 대뇌반구의 필요한 기본 활동을 정상적으로 생성하는 신 경 입력의 제거에 의해서인 듯하다. 10.2 문화적 진화 문화의 세계, 죽 포퍼 (Po pp er 1972) 의 세계 3 을 이미 제 4 장에서 그림 4.2 로 소개하였다 . 이는 우리 인간 문명의 모든 측면을 포함한다. 그리 고 몇 가지 의심스러운 예외를 제외하고는 이것은 전적으로 호미니드 의 창조이다. 더구나 호모 사피엔스 사피엔스의 출현 이전에는 도구문 화(그림 6 .1 0) 의 고고학적 기록에 단지 드물게 세계 3 의 희미한 깜박임 이 있었을 뿐이다. 그림 4.2 에서 세계 3 에 나열된 것들은 개요일 뿐이 댜 각각의 항목은 훨씬 확대되어야 하는 것이다. 예를 들어 <예술적> 이라는 항은 조소미술과 공예 그리고 건축과 음악을 모두 포함하는 것 이댜 포퍼는 세계 3 을 자기 자신들의 창작의 문제를 해결하고자 했던 창 조적 인간 개체들에 의해 만들어진 것으로 생각했다. 수학과 관련하여 그는 다음과 같이 기술하였다. 우리는 이들 문제를 파악하고 이해하거나 발견할 수 있다 . 그리고 그중 일부는 해결할 수 있다 . 따라서 세계 2 에 속하는 우리들의 생각은 부분적 으로는 자율적인 문제와 세계 3 에 속하는 일반 원리의 객관적 진리에 좌 우된댜 세계 2 가 세계 3 을 창조하기만 하는 것은 아니다. 부분적으로는 일종의 되먹임 과정으로 세계 3 에 의해서 창조되기도 한다 (1982: 128). 호모 사피엔스 사피엔스의 초기 수만년 동안의 세계 3 의 성장이 얼 마나 느린 것이었는가는 놀라운 일이다. 오늘날까지도 문화적 창조성
에 대한 인류의 경쟁은 무시할 정도이다. 단지 사회는 주거 , 식량, 의 복 그리고 안전에 대한 일차적인 요구만 충족시켜줄 수 있으면 그 구 성원들은 효과적으로 문화적 창조성을 발휘할 수 있고 그래서 세계 3 을 풍요롭게 한다. 오늘날에도 볼 수 있듯이 아마도 공동체 내에 결정 적 수효 이상의 집단이 있어 창조적인 사람들 사이에 상호작용할 수 있어야 했을 것이다 . 시초에 문헌과 역사의 세계 3 은 사람의 기억 속 에 있었댜 예를 들면 약 5000 y BP 의 수메르인들에 의해 문자 기록이 발명되기 전에 음영시인들의 시암송 같은 것이다. 어찌되었건 거의 오 늘날까지도 문맹사회에는 시인들의 시암송에 의한 세계 3 의 전파가 있었다. 이를 로드 (Lord 1960) 는 < 이야기를 노래하는 사람 > 에서 생생 하게 기술하였다. 세계 3 이 책의 잉크와 종이 같은 물질적 기질에 기록되었다는 것 은 중요하다 그러나 세계 3 자체는 비물질이고 근본적으로는 예술가, 사 색가 , 이야기꾼 그리고 결국 철학가, 과학자들의 창조적 관념인 것 이 댜 언어를 보존할 수 있는 물질 위에 기록할 수 있다는 수메르인들에 의한 발견 죽 진흙 조각에 새겨진 설형문자는 현대 세계의 태생기적 시발이라고 생각할 수 있고 이는 세계 3 의 모든 종류의 아이디어의 기록이라는 커다란 기술적 발전에 의해 창조되고 좌우되는 세계이다 . 거기에는 우리들 사회가 매체에 의해 압도될 한 가지 커다란 위험이 있댜 인간은 태어날 때부터 인간의 뇌를 갖는다. 그러나 이것의 세계 2 의 경험은 대단히 미미하고 세계 3 은 미지상태이다. 태어날 때의 뇌는 태 생기부터도 인간으로 간주하여야 한다. 그러나 인간 인격은 아니다. 세계 3, 죽 문화의 세계와의 계속적인 상호작용에 의한 자아의식의 출 현과 발달(세계 2) 은 신비의 과정이다. 이는 효과적인 교차연결에 의해 성장하고 상행하는 이중 구조(그림 10 .l)에 비유할 수 있다. 수직 화살 표는 어린아이의 최초의 경험으로부터 완전히 발달한 인간까지의 시
간 경과를 나타낸다. 각 세계 2 의 위치로부터 한 화살표는 점점 더 높 고 큰 세계 3 으로 유도된다. 이는 그 개체의 문화적 성장을 상징적으 로 나타낸 것이다. 역으로 자아의 세계 3 자원은 그 자아(세계 2) 의 의 식의 더 높고 확대된 수준을 이끌도록 작용한다. 그림 10 .1은 인간성의 사다리롤 상징한 것으로 볼 수 있다. 그리고 우리들 각각은 자아 창조 가 점진적으로 발달하였고 이는 우리의 전 생애를 통하여 계속될 수 있댜 인간 인격의 세계 3 자원이 크면 클수록 상호 풍요화에 의해 세 계 2 의 자아의식을 더 획득하게 된다. 우리가 무엇이 될 것인가는 우 리가 담고 있는 세계 3 에 의해 좌우되며 우리 뇌의 잠재력을 최대한으 로 하는 우리의 기회를 어떻게 효과적으로 이용하는가에 달렸다. 최근의 한 비극적 경우를 그림 10 . 1 이 나타낸다. 지니 Ge ni e 라는 어린 아이는 정신병자인 아버지에 의해 모든 세계 3 이 결핍되었다. 그녀는 로스앤젤레스에 있는 그녀의 집 작은 다락방에 격리되어 감금되었다. 20 개월째부터 13 년 8 개월이 될 때까지 말을 해본 일이 없었고 최소한 의 보살핌을 받았다. 이 끔찍한 폐쇄에서 풀려났을 때 물론 그녀는 인 간이었으나 인간 인격은 아니었다. 그녀는 그림 10 . 1 의 사다리 맨 밑바 닥에 있었다. 그후 의사 수잔 커티스 (Susan Cu rtiss 1977) 의 정성스런 도움으로 그녀는 서서히 인성의 사다리를 기어오를 수 있었다. 언어의 박탈은 그녀의 좌측 대뇌반구를 심하게 손상시켰다. 그러나 우측 대뇌 반구가 상당히 결핍된 언어수행능을 대신할 수 있었다 . 세계 3 에의 접 촉이 이같이 끔찍스럽게 지연되었으나 지니는 인간 인격이 되어 자아 의식과 감정을 가지고 손재주와 시각적 인식을 훌륭하게 가지게 되었 댜 인간 인격 발달에 있어서의 세계 3 의 필요성을 알 수 있는 것이다. 그림 10 .1에 있듯이 뇌는 유전적 지시, 죽 생물학적 조건 Na tur e 에 의해 형성되나 인간 인격성의 발달은 세계 3 환경(죽, 환경요소 N urtur e) 에 좌 우된다. 지니에게 있어서 생물학적 조건 Na tur e 과 환경요소 N urture의 개시 사이에는 13 년이라는 간격이 있었다.
 세계 2 세계 3
세계 2 세계 3
인간 특유의 특성인 기공성, 의식적 경험의 전수, 그리고 상상적 생 각 둥의 스테빈 (S te bb ins 1982) 의 개념에 관하여는 6 .1 1 에서 이미 간략 하게 살펴보았다. 이들 특징은 인간 고유의 세계 3 의 기초를 형성하며 유인원과 유전적 유사성이 있음에도 불구하고 호모 사피엔스 사피엔 스를 정성적으로 다론 모든 동물과 구별하는 것이다. 세계 3 은 호모의 진화적 발달을 변형시켰다. 이제 생물학적 진화와 문화적 진화를 중점 적으로 비교하여 보려 한다.
10.3 생물학적 및 문화적 진화 스테빈은 문화적 진화가 생물학적 진화보다 얼마나 훨 씬 빠르고 효 과적인가를 보여주었다. 문화적 소인은 10 년 이내에 대집단을 통해 전 파된다 그러나 도태 압력에 의한 인간 유전자의 적응에는 수백 년이 걸린댜 문화적 진화라는 관 점 에서 인간은 적응력이 있든 없든 간에 주로 관습 , 모방 및 학습에 기초한 차이가 매우 심하다는 것이 독특하다 (1982: 392). 스테빈 (1982) 은 < 형판t em p la t es > 이라는 개념을 도입하였다 . 죽 복제 기구이다. 생물학적 진화에 있어 DNA 는 유전적인 생물학적 능력을 전 도하는 유전적 형판이다. 문화적 진화에 있어서 문화적 형판은 도처에 산재한댜 예를 들면 모든 종류의 구조물에 대한 설계도, 직물, 의복, 도자기 , 선전물 등의 설계에 있는 것이다. 항목을 나열하자면 끝이 없 댜 우리는 상상을 초월하는 방식으로 문화적 형판에 의해 조건지어지 는 것이다 . 이 모든 문화적 행위는 포퍼(1 972) 의 세계 3 안에 있는 것이다. 도킨 스(D aw ki ns 1976) 는 그가 문화적 요소에 대한 훨씬 덜 포괄적인
유전형의 생물학적 진화 문화적 진화, 인간제한적 유전형에 의한 표현형 형성 가성이타주의적 행동의 동물뇌 언어적 성질의 인간뇌 특이적 인간언어의 학습 이타적 행동 성질의 인간뇌 이타적 행동의 학습 모든 문화적 성취성질의 인간뇌 문화의학습 그림 10.2 진화의 일반적 개념도. 수직선은 생물학적 진화와 문화적 진화 를 분 명하게 분리한다. 왼쪽에는 순전히 세계 l 의 대상 죽 유전형, 표현 형, 동물 뇌, 사람 뇌가 있다 오른쪽에는 이에 반해 세계 3 의 대상 죽 언어 , 가치관 그리고 그림 4.2 에서 지적했던 모든 문화에 관한 것 이 있다 .
다양성으로부터의 선택에 좌우된다. 생물학적 진화에 있어 다양성이란 유전자 풀g ene po ol 이 다 문화적 진화에 있어 서 는 저 장이 란 문화적 다 양성 을 가진 세 계 3, 즉 문화적 풀 cul tur al po ol 이 다 (S t ebb i ns 1982). 사회는 불합리한 또는 본능적인 호미니드의 집합체라는 원시적 기 초로부터 가치관으로 다스려지는 고도의 조직적인 구조로 문화적 진 화를 통해 발달하였다. 생물학적 진화가 언어 학습의 가능성, 애타적 행동의 가능성, 그리고 다른 모든 문화 활동, 죽 사회를 다스리고 형체 룰 만드는 가치체계를 포함하는 활동에 대한 가능성을 가진 인간의 뇌 를 형성하는 인간의 유전자형을 창조하였다는 것이 그림 10.2 에 나타 나 있댜 그림 10.2 에서는 생물학적 및 문화적 진화 사이의 절대적 차 이점을 강조하고 있다. 생물학적 진화는 구조(유전자)를 창조하였고 유 전자적 유전이라는 유전자의 역할에 좌우된다. 문화적 진화는 호미니 드의 문화적 성취에 좌우되며 전혀 유전자적으로 기록된 것이 아니며 라마르크의 용불용설에 해당할 것이다. 이는 전적으로 지도와 학습에
의하여 전달된다 . 언어에 대한 유전자는 없다(D obzhansk y 1967). 단지 인간의 언어를 학습할 수 있도록 하는 언어영역(제 4 장)에 대한 유전자 가 있을 뿐이다 (Chomsk y 1978). 그림 10.2 에는 세계 2 의 근본적 역할이 빠져 있다. 이는 세계 l 의 뇌 구조와 문화적 세계 3 의 학습 사이를 매 개하는 것이다 . 생각, 느낌, 기억, 상상, 주의 등 세계 2 의 내적 감각의 그 같은 특징은 필요한 것이다 . 가치란 세계 3 의 중요한 구성 요소이다. 이는 그림 4 .2에 있는 철학 에 포함되는 것이다. 문화적 풀은 우리의 ·가치 체계를 반영한다. 가치 란 간단하게는 우리가 가치있게 여기는 것이라 생각할 수 있다. 그것 온 우리의 판단과 이것이냐 저것이냐를 선택하는 기준이 된다. 누구나 가치 척도와 가치 체계를 가지고 있다. 대개는 이를 의식하지 못하지 만 판단의 기틀이 되는 것이며 물론 이것의 역할은 제반 조건을 제공 하는 것일 뿐 결정 짓는 것은 아니다. 행위의 이 같은 법칙은 언어에 서와 마찬가지로 학습된 것이다. 제 4 장에서 언급했듯이 뇌는 언어에 대해 가능성을 가지고 형성되었다. 신피질의 큰 언어 영역은 생물학적 진화에서 발달하였고 출생시 들었던 어떤 형태로의 언어로든 사용되 기 이전에 미리 형성되는 것이다. 따라서 또한 뇌는 가치 체계에 따르 는 행위의 가능성을 가지고 형성된다. 그리고 처음으로 배우게 되는 가치 체계는 그를 둘러싸고 있는 문화의 가치 체계이다. 동물의 의사소통이 발달하지 못한 것과 마찬가지로 사회구조도 발 달하지 못하였고 그 안에서 행동 양식은 본능 또는 학습을 기초로 이 루어진다. 아마도 우리들 영장류 선조의 이 사회는 초기 호미니드의 문화를 발달시켰던 기초를 이룬다. 동기는 조칙적 사냥 또는 전투에 있어 그리고 음식나누기와 도구 제조의 발달에 있어 협동의 필요였을 것이라고 생각할 수 있다. 한 가지 예로는 수백 년, 수천 년에 걸친 대 단히 점진적인 손도끼의 발달에 관한 기록이다(그림 6.1 0 ). 그러나 가장 중요한 것은 사회적 협동과 이해를 가능하게 했던 언어적 소통의 점진
적 개 선 이 다 (Tob i as 1981b). 불행히도 전통적 가치 체계를 쓸모없는 것으로 신용하지 않는 진취 적 생각을 가진 사람들에 의한 고정관념이 있다 . 아마도 종교적 기초를 가졌기 때문일 것이다. 그래서 전통적 가치를 과학적 합리성 (Monod 1971, Sp e rr y 198. 3, Chang e ux 1985) 또는 보다 진화적 인 기 초 (Pu g h 1978, Wi lso n 1975, 1978) 에 근거한 윤리로 대치하려고 시도되었댜 이들 모든 가치 체계의 특징은 숨겨져 있을 수 있는 유물론적 기초이다. 전형적 전통적 가치에 대한 옹호자로는 포퍼 (1972), 랭거 (195 1), 도르프 (1974), 하이 에크(1 978) 및 에클스(1 980) 들을 인용할 수 있다. 불행히도 유물론 적 선입관은 우리의 전통 가치 체계를 혼동시키고 모노(1 971) 의 경우 처럼 이룰 독단적 유물론적 억설로 대치하였다. 호미니드의 생물학적 진화는 절정에 도달했다 . 물론 인간 게놈 g enome 의 돌연변이는 계속될 것이다 그리고 이는 방대한 인구로 인 하여 그 빈도가 대단히 커질 것이다. 예를 들어 호모 사피엔스 사피엔 스의 기원이 되는 가장 중요한 의미를 갖는 유전적 변형은 호미니드 세계의 인구가 2 백만 정도일 때 일어났을 것이다(D eeve y 1960). 이제 인구는 2000 배 이상 크다. 그러나 돌연변이가 방대한 유전자 풀에서의 희석 효과로 인해 유지되기는 힘들다(제 1 장). 이 현세계의 모든 곳에 교통 통신이 가능하므로 작은 변두리 격리집단이 수백 수천 년에 걸친 호미니드 진화의 경우처럼 수 세대에 걸쳐 계속 격리되어 번식할 수 있는 기회가 주어지지 않는다. 더욱이 모든 사회의 목표는 사회 복지와 건강 관리에 의하여 모든 개인의 생존을 보장하는 것이다 . 자연도태의 방침인 적자생존의 원리 로 되돌리기는 생각할 수 없다 . 사실상 현재의 상황은 사회의 최하위 구성원에 있어서 번식률이 더 높은 것으로 보아 이에 정반대되는 경향 이 있다. 필사적인 방법으로서 히틀러가 아리안족 사회를 위해 획책했 던 것처럼 선별적 번식 계획의 개시를 제안할 수 있다. 이 같은 계획
은 가축의 번식에 있어서는 대단히 성공적이다 . 그러나 인간의 수백 년에 걸친 번식이 이루어지려면 독재 정치에 의해 강제되지 않으면 안 될 것이다. 따라서 호미니드의 생물학적 진화는 더 이상 가능하지 않 다고 생각한다. 이는 진화론자 그리고 그들의 저술에 의하여 인정되어 야 한다 그렇지 않으면 과학소설가에 의한 공상이 될 것이다. 한편 문화적 진화에 있어서는 그 같은 생물학적 제한은 없다. 이것 의 장래는 인간의 창조성에 달려 있다 . 역사를 검토해 보면 세계 3 의 모든 구성분이 전반적인 진화가 계속된 것은 아니다(그림 4.2 참조). 이 문제에 관해서는 많은 논란이 있을 수 있다. 새로운 색다른 분야에 있 어서의 창조성에 대한 인간의 뇌의 잠재력에 대해서는 의혹이 있을 수 있다 그러나 가장 놀라운 일은 호미니드 진화의 세속적 과정이 여러 가지 종류의 능력과 질서를 가진 인간의 뇌를 창조하였다는 것인데 이 능력과 질서는 호모 사피엔스 사피엔스의 초기 구성원의 생존을 위해 필요했던 것들과는 전혀 다른 것들이다. 수학과 음악의 경이를 생각해 보라! 이것이 월라스 Wallace 의 문제였다 (10.6). 현세는 문화적 산물의 보존에 있어 탁월하다. 박물관과 도서관은 이 전에 비할 데 없을 만큼 발달되어 있고 비판적 평가를 통한 걸작들의 집합체가 되고 있다. 더욱이 시각적 및 청각적 기록과 재생 기술에 힘 입어 예술적 산물의 보존에 있어 제한이 없어졌다. 그러나 상상하기 힘든 지구의 장래를 상상할 때 보존된 물질의 단순한 집단으로는 불안 하다 그 지구는 다음 15 억년 동안 태양을 돌고 오늘날과 마찬가지로 많은 방사선을 받게 될 것이다 . 예를 들어 과학적 문헌들이 장래의 몇 백 년이 지난 후에 처할 상황을 상상하는 것은 우리의 상상을 초월하 는 것이다.
10.4 마음의 개념 의식의 창조와 진화라는 주제로 들어가기 전에 현대의 철학자들의 의견들을 간략하게 살펴보려 한다(좀 아이로니컬하게 이 절 의 제목을 택 했다). 철학자들의 마음에 대한 일원론적 유물론적 철학에 는 놀라운 의견 의 일치가 있는 것 같다 . 그러나 자세히 살펴보면 그 믿음의 범위는 넓다는 것을 알 수 있다. 그리고 철학자들의 믿음은 행동주의의 긴 암 흑기로부터의 점진적 탈출이 일어났던 지난 수십년 동안에 변하여 왔 다(B elo ff 1965). 언어 의 사용은 간단한 예 이 다. 라일 R y le 의 『마음의 개 념』 (1949) 이라는 도전적 저서의 영향의 절정(또는 천저)에서 보면 마음, 의식, 생각, 목적, 믿음 같은 단어들은 용납되지 않았다. 뇌 - 신체 기구 로부터 데카르트 학파의 유령을 내몰려는 노력에 있어서는 데카르트 학파의 이원론 맛이 나는 모든 단어는 지저분한&rty 단어가 되고 교양 있는 철학 강의에서는 용납될 수 없다. 아이러니컬하게도 가장 심하게 철학적으로 추악한 것은 네 글자로 된 새로운 부류의 단어이다 . 죽 마 음min d, 자아 sel f, 영 soul, 의지 w ill이다. 자신을 일원론적 유물론자 또는 물리주의자로 생각하는 철학자들의 신념들도 굉장히 넓은 변이를 보인다 . 그러나 이 변이는 시간의 함수 죽 일종의 문화적 진화라 할 수 있다. 암스트롱 Arms tr on g이 1968 년과 1981 년의 저서 사이에서 그의 관점을 현저하게 바꾸었다는 것은 홍미 있다 이 런 일은 덴네트(D enne tt 1969, 1981) 와 헤브(H ebb, 1949, 1980) 에 게서도 발견된다. 라일의 영향은 최근 십여년 동안에 이들 철학자들의 용어 사용의 변화에 의해 기울어감을 느낄 수 있다. 데카르트의 이원론의 망령으로 남아 있는 민속 철학인 <머릿속의 소인> 죽 정자미인 homunculus 을 극복하려는 철학자들의 고집이 있다 (De nnett 1969 p.1 90). 덴네토는 우리는 데카르트의 정신으로 유도되는
마음이나 아무 소용이 없는 신체가 아니고 사람이 무엇을 할 수 있는 가만을 고려해야 한다고 생각했다. 그러나 이 내용에서 사람이란 무었 인가? 마음-뇌의 문제를 과학적으로 분석하려고 노력하려면 뇌내의 궁극적 미소 부위에서의 신경 활동에 대한 총체적 행위의 주체로서의 사람 밑바닥을 살펴야 한다(제 8 장) . 의식의 경험에 관한 만족할 만한 평가는 암스트롱이 『마음의 성질 』 (1 981) 이라는 그의 책 제 4 장에 기술하였다. 완전한 무의식 상태인 사 람에게 지식과 믿음을 가진 마음이 있다고 말하는 것은 이상하다. 지 식과 믿음은 소실되는 것이 아니고 단지 정지될 뿐이라는 것이다 . 그 러나 그와 같은 조건에서 때로는 문제를 해결하는 것처럼 보이는 또는 어떤 생각을 적절하게 표현하는 방법을 발견한 것처럼 보이는 일이 일 어난다 이는 우리의 각성시 의식 상태에서 나타난다. 암스트롱은 이 룰 최소한의 의식적 행위라고 간주하였다. 심적이라고 할 수 있는 어 떤 일이 실제로 일어났기 때문이다 . 지각과 연합된 특수한 부류의 심적 활동이 수면이나 꿈에서 깨어날 때 그리고 주위에 있는 것을 알아차리게 될 때 인식된다. 최소 의식은 지각적 의식으로 변화되는 것이다. 이 지각적 의식 상태는 우리의 부 분적 각성 상태 또는, 더 좋게 표현하면, 주의 집중을 하지 않은 의식 상태를 나타내는 것인 듯하다 . 어찌되었든 사람은 정신의 지각인 의식이 아직도 완전하지는 못하 댜 (그 같은) 내 적 지 각은 전통적 으로는 내 관i n tr os p ec ti on 또는 내 관적 의 식성i n tr os p ec ti ve awareness 이라고 불린다. 따라서 우리는 이 제 3 의 의 식을 내관적 의식이라고 말할 수 있다. 이는 우리 자신의 마음속의 현재 상태와 활동에 대한 지각과 비슷한 의식성이다…… 내관 자체가 심적 활 동이므로 이것 또한 내관적 의식성의 대상이 될 수 있다 . (Arms trong
1981: 61) 내가 암스트롱의 이 글을 직접 인용한 이유는 그의 생각이 너무나 도 나의 생각과 일치하므로 내가 그의 글을 잘못 오해하고 있지는 않 을까 하는 우려 때문이었다. 내관적 의식은 자아 의식이라고 할 수 있는 것이다 (Po pp er 와 Eccles 1977). 지각적 경험을 주의 깊게 살펴보면 내관적 의식은 높아진 의식 성 안에서 활동을 시작한다. 이는 또한 그림 9.5 에서 내적 감각에 나열 한 느낌 , 기억, 생각, 의도 따위의 다른 심적 상태에 관련해서도 일어 난댜 우리가 내관적으로 알고 있는 이들 모든 상태와 활동들은 유일 한 지속적 주체인 자아 안에서 통합적인 기반을 가진다. 자아에 있어 다양한 내관적 의식의 이 통일은 대단히 중요한 것으로 이는 우리의 가치 체계가 어떻게 우리의 판단에 연합될 수 있는가를 함께 살펴볼 기회가 되기 때문이다. 따라서 암스트롱의 말처럼 < 완전한 각성 상태 의 내관적 의식은 특징적으로 표준 관행으로는 수행할 수 없는 문제 상황에 닥칠 때 일어난다. > 암스트롱이 말한 이들 정신주의적 논의와 결론 뒤에 <물리주의자는 내관적 상태와 활동을 뇌의……모든 물리적 상태로 간주할 것이다>라고 한 것은 놀람다. 내관적 의식은 개관하의 심적 활동에 대한 강한 주의 집중을 포함 한다 따라서 암스트롱은 이것이 사건을 기억하는 데에 중요한 역할을 한다고 제안하였다. 주목하지 않았던 사건은 기억되지 않는다는 것에 는 일반적으로 동의한다. 특히 사건의 기억은 자아의 과거력에 관한 기초가 되는 것이다. 이에 관해 암스트롱은 다음과 같이 강력하게 결 론짓고 있다. 내관적 의식이 없이는 우리는 우리가 존재한다는 사실을 알지 못할 것 이다. 우리의 자아는 자기 자신에게 자아가 되지 못할 것이다. 그래서 우
리는 내관적 의식이 어떻게 심적으로 존재하는 것 심지어 존재하는 모든 것의 조건으로 보이게 되는가를 이해할 수 있다. 의식적 인식이 존재한다는 강력한 증거가 있는 고등동물들에서는 어 떠 한가 하고 묻게 된다 .(8.2, Gri ffin 1976, 1984, Eccles 1982a). 최 소 의 식 과 감지 의식으로 충분한가? 내관적 의식 또는 자아의식이 이들 동물 에서 존재한다는 증거는 거의 없다. 그러나 감지, 느낌, 기억, 의도 그 리고 심지어 생각 같은 심적 속성이 천부적으로 부여되는 것으로 생각 된다(그림 8.5). 그리고 많은 종들은 사건 기억능력이 잘 발달되어 있 댜 그들에게는 암스트롱이 기술했던 것보다는 더 발달된 감지 의식이 있다고 생각되며 이는 내관적 의식으로의 변천 과정상에 있는 것이다. 덴네트는 라일의 철학을 고수하고 있으나 그의 진술은 조금 변하였다. 마음의 가장 중심적인 특성, 무엇보다도 가장 전형적으로 심적이고 비 물질적인 < 현상 > 은 의식이다 (1969: 99). 그러나 그는 이것이 마음의 유일한 특성이 아니고 조화되지 않은 특성들의 집적이라는 것을 증명하기 위한 시도로 의식을 분석하였다. 덴네트의 목표는 이원론적 상호작용론적 논의상 자아(그림 9.5) 의 독립 적 주체이며 기계에서 유령을 쫓아내려는 라일의 주문 이후 다시 부활 한 듯이 보이는 <뇌속의 소인>을 제거하려는 것이다. 그는 무오성의 보고자라고 한 데카르트적 관점 때문에 문제를 일으키는 내관적 접근 을 바판하였다. 그러나 그는 의식을 두 부분으로 구분한다. 한 가지는 특히 감지에 대한 의도적 감각의 의식성이고 다른 하나는 사고와 관계 조차 되지 않는 일종의 애매한 잔재에 대한 의식이다(D enne tt 1969). 홍미로운 것은 헤브가 마음에 관하여 기술한 것이다. 그는 세포-집 합설, 죽 매우 생리학적인 가설을 수립하려고 생각하였다. 그는 생각
울 여러 차원의 추상을 거친 표현적 과정이라고 말하였다. 종국적으로는 언어적 수행에 관련한다 해도 비언어적 생각은 높은 수 준에 존재할 수 있다 . 그러나 글을 쓰는 사람은 누구나 자신의 아이디어 를 단어로 표현해야 하는 것이 생각을 예민하게 할 수 있음을 안다 . 언어 는 인간 행위의 걸출한 고유의 특성이며 아마도 인간 사고의 이러한 특 성 이 기 도 한 듯하다 (1980: 116). 이들은 유물론적 설명이 아니다. 설 Searle 또한 자신을 물질주의적이 아니고 정신주의적으로 표현하 였댜 그의 저서 끝에 다음과 같이 말하였다. 나는 의식이 있다고 생각되기 때문에 나는 의식이 있다. 우리는 우리 들 자신과 우리들의 행동에 관한 모든 종류의 우리를 놀라게 하는 일들 을 발견할 수 있다. 그러나 우리에게 마음이 없다는 것, 우리의 마음이 의 식적, 주관적 및 의도적 심적 상태를 포함하지 않는다는 것은 발견할 수 없다. 또한 우리는 우리가 최소한 자발적, 자주적 및 의도적으로 행동하 려고 시도하지 않는다는 것도 발견할 수 없다.……나의 일반적 주제는 특 정한 중요한 예의들이 있지만 우리들 자신의 우리 자신에 관한 상식적 정신주의적 개념들이 물리적 체계로서의 자연에 대한 우리들의 개념과 완전하게 조화된 것이라는 것이다(1 984 : 99). 이 모두는 그 진술들 자체로서 훌륭하다. 그리고 이는 그림 9.5 에 있 는 이원론적 상호작용론의 도식적 표현과 잘 맞아 떨어진다. 암스트 롱, 덴네트, 헤브 및 설 같은 철학자들이 가진 어려움은 비물질적 심적 사건이 뇌의 시냅스에 작용할 수 있다는 명백한 수수께끼에 있다. 그 래서 그들은 비록 스스로를 일원론적 유물론자로 자처하면서도 이원
론적으로 썼는데 스페리(1 982) 도 마찬가지였다 . 그러나 8.8 에서 그것들 에 접근하는 방법이 제시되었다 . < 기계 속의 유령 > 이라고 라일이 비웃는 것을 나는 그가 모르는 사 이에 귀중한 식견을 표현했다고 비꼬며 결론짓고자 한다. 우리가 가질 수 있는 마음-뇌의 이원론은 비유하건대, 소위 전통적으로 유령이라 불린 것은 뇌라는 기계의 계획을 짜는 자로서 작용하는 비물질적 마음 이라는 것이다. 이것은 또한 소프트웨어나 하드웨어의 면에서 컴퓨터 에 대옹하는 것이다. 이 비유는 10.7 끝에 더 전개할 것이다. 10.5 특이적 인간 특성 이 제목 밑에 단순히 정량적인 것이 아니고 의심할 여지없이 정성 적인 진화적 발달의 예들을 들게 될 것이다 10.2). 10.5.l 의식적 시간의식 스테빈스는 다음과 같이 말했다. 의식적 시간 의식의 개념은 과거 경험의 기억을 이용하면서 장래를 계 획하는 비상한 인간의 능력을 나타낸다. 유인원과 기타 동물에 있어 이것 의 선주자는 최소한으로 혼적 정도이다 . 그들의 행동을 자세히 분석하면 그들은 자동적이고 판에 박은 것이고 비교적 융통성이 없음을 알 수 있 댜 이돌 동물들은 비정상적 또는 예측하지 못했던 상황에 봉착했을 때 지능적 인간이 자신이 계획한 행동 양식을 수행할 때와 같은 그들의 행 동을 조정하는 능력이 적다(1 982: 364).
잉그바르 ( In g var 1985) 는 뛰어난 통찰력으로 인간의 의식적 의식성 의 시간적 구조에 관하여 기술하였다. 우리는 과거 一 현재-미래의 시간 구조 속에 살고 있다 . 사람이 의식적으로 < 현재의 > 시간을 의식 할 때 이 경험은 과거 사건에 관한 기억뿐 아니라 미래 사건의 예측까지도 포함한다 . 이 예측은 미래에 대하여 과거에 예측했던 기억까지 포함하 는 심적 과정의 복합이다. 우리는 상상 속에서 우리가 계속해서 연습 했던 여러 가지로 다르게 계획한 활동에서 일어날 수 있는 모든 가능 성을 생각해 본다 . 잉그바르는 조금 역설적으로 이 심적 과정을 미래 의 기억이라고 하였다. 이것은 이미 일어났던 일에 관한 기억이 아니 고 모든 가능한 경우를 감안한 미래의 계획된 활동에 대하여 우리가 생각할 때 경험하는 예측의 복합을 기억하는 것이다. 체스놀이가 뛰어 난 예가 될 것이다. 달리 표현하면 < 기억된 예측 > 이라 할 수 있다. 이 개념이 제안되고 검토되자마자 우리의 자아의식적 경험의 흐름 . 의 대부분은 정밀하게 평가되어 계획된 활동의 변화를 유도하는 기억 된 예측 속에 들어있다는 것이 분명해진다. 그래서 우리의 현재 경험 에서 <지금>이라는 것의 상당 부분은 미래로 투사되는 것이다. 잉그 바르 (1985) 는 연속적 정보를 취급하는 전두-전두전 신피질의 능력 때 문 (Fus t er 1980) 에 이것이 주로 관계된다는 임상적 증거룰 제시하였다. 그림 10.3 은 롤란드(R oland 1981) 의 주목할 만한 발견을 나타낸 것 이 댜 사람을 대상으로 하여 손가락에 겨우 감지할 수 있는 자극을 가했 을 때 대뇌피질의 후중심회의 손가락一접촉 영역과 중앙전두전 영역에 걸쳐 국소 대뇌혈류 (rCBF) 의 증가가 있었다. 손가락의 접촉이 있기 전 에 기록된 것이므로 이 같은 중가는 정신적 주의 집중의 결과이다. 따 라서 그림 10.3 은 주의 집중이라는 정신적 활동이 대뇌피질의 적절한 국소를 활성화시켰다는 것을 분명하게 보여주는 것이다. 비슷한 결과 는 입술에 접촉되는 것이 예상될 때에도 일어난다. 그러나 물론 입술 에 대한 체감각 영역이 활성화된다. 예측하는 동안에는 전두전 영역의
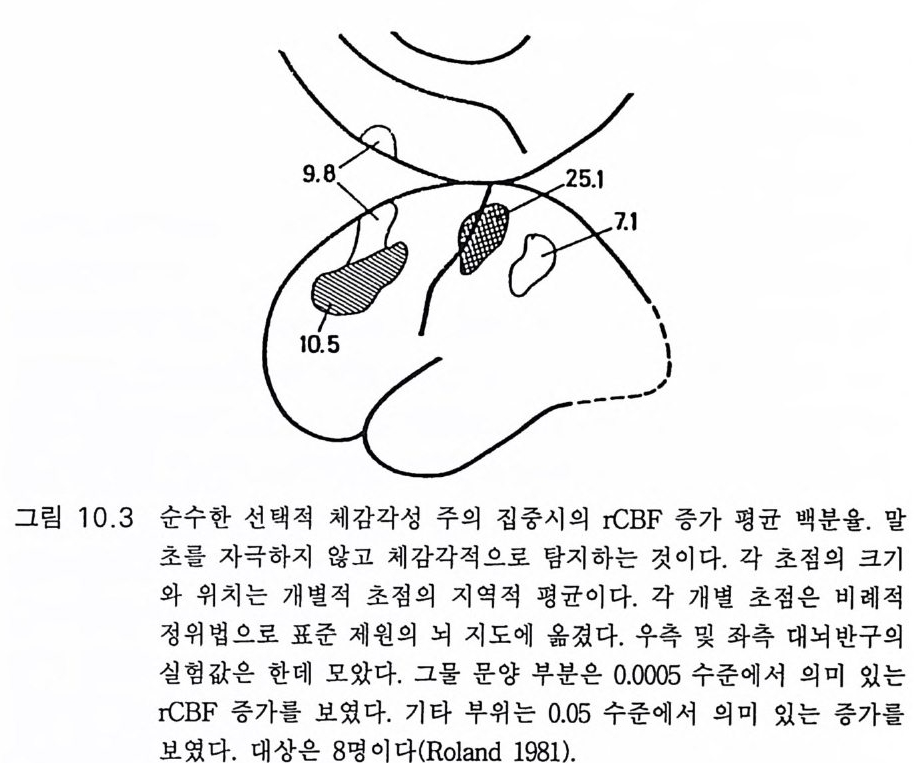 `\
`\
넓은 부위가 활성화된다. 안정상태에서 몽상을 할 때도 전두전피질은 신경 활성을 보여 rCBF 검사상 신피질의 다른 부위의 안정시 수준에 비하여 10-30% 높은 수 준의 뇌활성을 보인다 (In g var 1983). 전두전피질의 양측성 병변은 무관 심, 의욕상실, 선견의 소실을 특징으로 하는 <미래의 상실> 중후를 초 래한댜 이와 관련하여 잉그바르는 다음과 같이 덧붙였다. 단어 발음과 문장 생성을 위한 일련의 순서적 활동 계획에 접근하는 수단이 없이는 우리는 일련의 순서적 단어 전문과 연설의 뜻을 감지할
수 없다 이 같은 개념은 일반적 타당성을 가질 수 있다. 즉 미래의 행동 과 인지에 대한 일련의 계획 , 죽 우리의 < 기억미래 >를 이용해야만 우리 의 뇌가 지속적으로 노출되어 있는 막대한 감각 포화 속에서 우리는 의 미 있는 전문을 선택하고 감지할 수 있다(1 985: 134). 우리의 의식적 생활에 있어 시간 의식의 역할은 과소 평가할 수 없 댜 이는 반성 re fl ec ti on 에 대해서도 중요한데 반성은 그 자체가 과거에 경험한 사건과 문제, 그리고 그들의 예측 결과로서 미래에 투사되는 것을 생각하는 것이이다 . 이는 인생극을 형성하고 많은 위대한 문헌, 시, 연극, 소설의 기초이며 세계 3 의 모든 구성 요소이다. 셰익스피어 의 연극 또는 탐정소설의 기초에서 철저한 시간 의식을 생각해 보라. 시간 의식은 인간 인격이 그의 호미노이드 조상을 얼마나 많이 초월한 것인가를 분명하게 하는 것이다. 10.5.2 창조적 상상 우리의 마음이 감각 경험으로 넘쳐나지 않을 때면 우리의 상상 속 에 어떤 영상이 자주 떠오를 것이다 . 더욱이 이 영상은 몽상의 통속적 인 과정에서 우리가 경험하는 것처럼 다른 영상을 환기시킬 수도 있고 더욱 많이 불러일으킬 수 있다. 우리가 풍요로운 마음을 가지게 되면 이돌 영상은 조화를 이루어 아름답고 미묘해질 것이다. 우리가 이 영 상을 일종의 언어-문어, 음악, 회화-로 표현하면 예술적 창조가 되는 것이다. 이 같은 창조적 상상은 통찰력을 제공하고 남을 이해하 게 만든다 . 그리고 사람의 활동 중에서 가장 뛰어난 것 중의 하나이다. 한편 훨씬 낮은 수준의 상상은 어린아이들의 놀이나 성인의 환상에 서 나타날 수 있고 여러 종의 어린 포유류의 놀이에서까지도 나타날 수 있을 것으로 생각한다.
뇌에서 상상의 위치는 어디인가라는 의문도 홍미 있는 것이다. 방사 능 추적 방법으로 여러 형태의 상상에 관련된 대뇌피질 부위는 대부분 전전엽에 위치함을 알 수 있고 그림 6.9 와 7 .1 4 에 세 가지 형태의 상상 즉 3 씩을 감하는 계산, 의미없는 단어의 열거, 거리 풍경의 열거 등에 대한 결과를 나타낸 것이다. 대뇌피질의 다른 영역들 죽 언어영역과 시각영역도 상상 과업에 특이적으로 관계된다. 특히 과학적 상상에 관 계되는 추상적 상상에 대한 이 같은 연구는 없었다 . 그러나 전두전피 질이 훨 씬 더 우세할 것이라는 것은 추측할 수 있다. 모노 (Monod 1975) 는 과학탐구놀 이 에 있 어 서 의 상상은 극도로 엄 격 한 규칙이 요구된다는 점에서 어린아이들의 놀이의 경우와 대단히 다 르다고 하였다. 이는 대단히 특이한 종류의 창조성이다 . 이는 기본 전 제에 대한 분명한 개념이 없이는 수행할 수 없고 이를 존중하지 않으 면 과학적 창조에 참여할 수 없는 것이다. 내가 받아본 뇌와 뇌-마음 의 문제를 다루는 많은 문헌들은 사실 이점에서 실패작들이었다. 그것 들은 단지 가짜 과학일 뿐이 다. 창조적 상상력을 특이적으로 통찰하면 과학적 창조성과 관계지울 수 있다 . 포퍼 (19 72, 1975) 의 말에 따르면 모든 과학적 발견은 이 미 존 재하는 과학적 믿음이나 새로운 통찰에서 야기되는 문제 상황에서 발 전되는 것이다 . 현존하는 문제 상황에 만족하지 못할 때 괴로움을 느 낀다 그래서 특정한 문제를 파악하고 연구하게 되고 과학자들에게는 그들이 잘 이해하는 분야에서 야기되기 때문에 큰 도전이 되는 문제에 부닥친다 나 자신이 잘 알고 있듯이 그런 문제들은 우리를 완전히 압 도하게 될 수도 있다. 포퍼는 사람들은 당면한 자신의 문제에 큰 관심 을 가져야 하고 그 문제를 동정적으로 칙관해야 한다고 하였다. 사람 들은 자신의 문제로 들어가서 그 일부가 된다. 이는 매우 홍미로운 모 험이 된다. 그러나 한편으로는 잠에서까지도 문제가 붙어다니게 된다. 정상적으로는 문제를 인식하고 관련되는 모든 문헌을 고찰하여 문
제의 모든 내용을 알 수 있게 되고 시도된 해결 방법을 알 수 있게 된 다. 가끔 사람들은 주관적 모방 sub j ec ti ve s i mula ti on 이 라는 것을 구사한 다. 자신이 전자, 이온, 원자, 분자, 세포 성분, 접합 가시, 세포 등 문제 의 대상인 것처럼 인식한다 . 이는 아이디어를 유발하는 생생한 상상적 직관을 가져오고 과학적 합리성이라는 과정에 의하여 비판적 평가를 받게 된댜 창조적 아이디어의 많은 단계들은 생각 실험에 의해 배제 된댜 이것이 과학놀이의 재미이댜 이런 가운데서 문제에 대하여 더 많은 것을 배우게 된다 . 이 경험을 통하여 우리는 자신 또는 다른 사 람의 잠정적 아이디어를 더 효과적으로 비평할 수 있게 된다. 과학적 비판을 극복하고 검정 가능한 예측을 유도하는 종국적인 창조적 아이 디어에 도달하는 데는 행운도 있는 것 같다 . 예상하지 못했던 이 같은 행운에 대하여 < seren dipity > 라는 훌륭한 말이 있다. 이 단어는 1754 년에 호라스 월폴 Horace Wal p ole 이 쓴 「세렌 디 프(스리 랑카의 옛 이 름)의 세 왕자 Three Pri nc es of Seren dip」 라는 이 야기에 근거하여 만들어진 것이다 . 월폴은 다음과 같이 썼다. 왕자님들은 여 행을 하면서 늘 그들이 바라지 않았던 일들을 우연에 의 하여 또는 약삭빠르게 발견하였다. 중요한 의문은 과학에 있어 좋은 아이디어는 어떻게 얻어지는가 하 는 것이다. 포퍼 (1975) 는 마음에 떠오르는 모든 아이디어를 취하고 그 것들에서 비평에 견디어내지 못하는 것들을 제거하라고 제안하였다. 이는 생물학적 진화에 있어서의 자연도태와 비슷한 것이다. 살아 남는 좋은 아이디어는 대단히 드물다. 나 자신의 경험으로는 나는 새로운 좋은 아이디어를 찾으려고 할 때 문제에 관한 지식과 그 문제에 시도되었던 해결책들에 관한 철저한 평가로 머리를 채운다. 그러고 나서 그러한 마음의 긴장에서 결과가
만들어질 때까지 기다린다 . 음악을 듣거나 아인슈타인이 했듯이 산보 룰 하였다. 이 기간을 소위 잠복기라고 한다. 나는 긴장 상태에 있는 마음과 애써 싸우려 하지 않았다. 다만 좋은 창조적 아이디어가 솟아 나기를 희망했고 가끔 그렇게 되었다. 이는 아이디어나 문제를 글로서 표현하는 것도 가끔 도움이 되었다 . 분명한 것은 대부분의 창조적 과 정은 무의식적으로 이루어진다는 것이다. 그러나 좋은 아이디어가 갑 자기 솟아나면 강렬한 심적 정신 집중에 빠지게 되고 이는 상당히 지 속될 수도 있다. 아이삭 뉴턴에 관한 이야기에도 트리니티 대학의 그 의 방에 무아경상태에 있었다는 이야기가 있다. 지능과 상상력이라는 두 가지 기본적 심적 속성을 분명하게 구별하 는 것이 중요하다. 우리는 모두 파악의 신속성, 표현의 선명성, 이해의 깊이, 지적 홍미 그리고 특히 통찰력 등의 속성에서 우리가 남들의 지 능을 판단하는 데 익숙해 있다. 이룰 측정하는 검사법이 있어 근거는 박약하지만 이를 I Q라는 일차원적 함수로 나타낸다. 상상이라는 것은 훨씬 형언하기 힘든 심적 현상이고 이를 평가할 수 있는 방법은 없다. 그러나 창조적 상상이란 지대한 중요성을 갖는 뇌의 속성이다. 상상이 란 배울 수 있는 것이 아니다. 이는 무엇보다도 감사해야 할 유산이다. 내가 이루었던 모든 일을 가능하게 했던 나 자신의 유산에 감사하는 것을 나 개인의 태도로 하고 있다. 포퍼는 라플라스의 결정론을 배척하면서 다음과 같이 말하였다. 그것(라플라스의 결정론)은 인간의 창조성은 순전히 우연의 문제로 만 든다. 의심할 바 없이 그 안에는 우연이라는 요소가 들어 있다. 그러나 음 악이나 예술의 걸작품 창조를 결국 물리학이나 화학의 용어로 설명할 수 도 있다는 이론은 내겐 우스꽝스럽게 보인다. 아직까지 음악의 창조를 설 명하는 한, 적어도 부분적으로는 다른 음악(이것은 음악가의 창조성을 자 극한다)의 영향으로 설명하여야 하고 더욱 중요한 것은 내적 구조라는 입
장에서 음악과 기타 세계 3 의 모든 현상에서 역할하는 내재적 법칙 또는 제한, 죽 그들이 흡수하고 때로는 도전하는 법칙과 제한이 음악가의 창조 성에 있어서는 무한히 중요하다는 것이다. 따라서 우리의 자유 특히 창조에 있어서의 자유는 분명히 세 가지 세 계 모두의 제한 하에 있는 것이다. 불행하게도 베토벤이 귀머거리로 태어 났더라면 그는 작곡가가 되기 힘들었을 것이다. 작곡가로서 그는 세계 3 의 구조적 제한에 대해 자유로이 그의 자유를 복종시켰다. 자율적 세계 3 은 그가 위대하고 천재적인 발견을 이룬 세계였고 히말라야에서 탐험가 들이 길을 찾듯이 자유로이 선택한 것이었다. 그러나 선택한 노정의 범위 에 제한은 있었고 그가 발견하였던 세계의 규정에 제한되었다 .(1982:128) . 10.5.3 의지의 자유와 도덕적 책임 제 8 장에서 사람(그림 8.4 ) 또는 원숭이(그림 8.3) 의 수의적 동작을 일 으키는 보조운동영역 (SMA) 의 초기 활성화에 대하여 설명하였다 , 수의 적 심적 의도가 연락뇌, 죽 SMA에 작용하는 양식을 그림 8 . 5 에서 설명 하였다(E ccles 1982b). 디 크와 콘후버 (D eecke 와 Kornhuber 1978) 는 간단 한 의도적 행위에 반응하여 SMA 위의 머리에서 기록되는 준비성 전 압을 증명하였다. 그림 10 .4는 사람을 대상으로 한 도형이다. 이것은 중앙에 자아와 의 지가 있다는 점에서 그림 8.5 와 다르다. 의지는 수의적 동작을 운영하 는 것으로 간주하는 것이 관례이다. 그 동작의 도덕적 경과는 자아에 의하여 평가된다. 따라서 의지적 행위는 도덕적 책임감을 수반한다. 이것을 도식적으로 나타내기 위하여 그림 10 .4에 <의도>로부터의 화 살표는 의지롤 거쳐 연락뇌로 그려졌고 비인간 동물의 경우에 있듯이 도덕성이 고려되지 않는 단순한 수의적 행위에서처럼 직접 가지는 않 는다
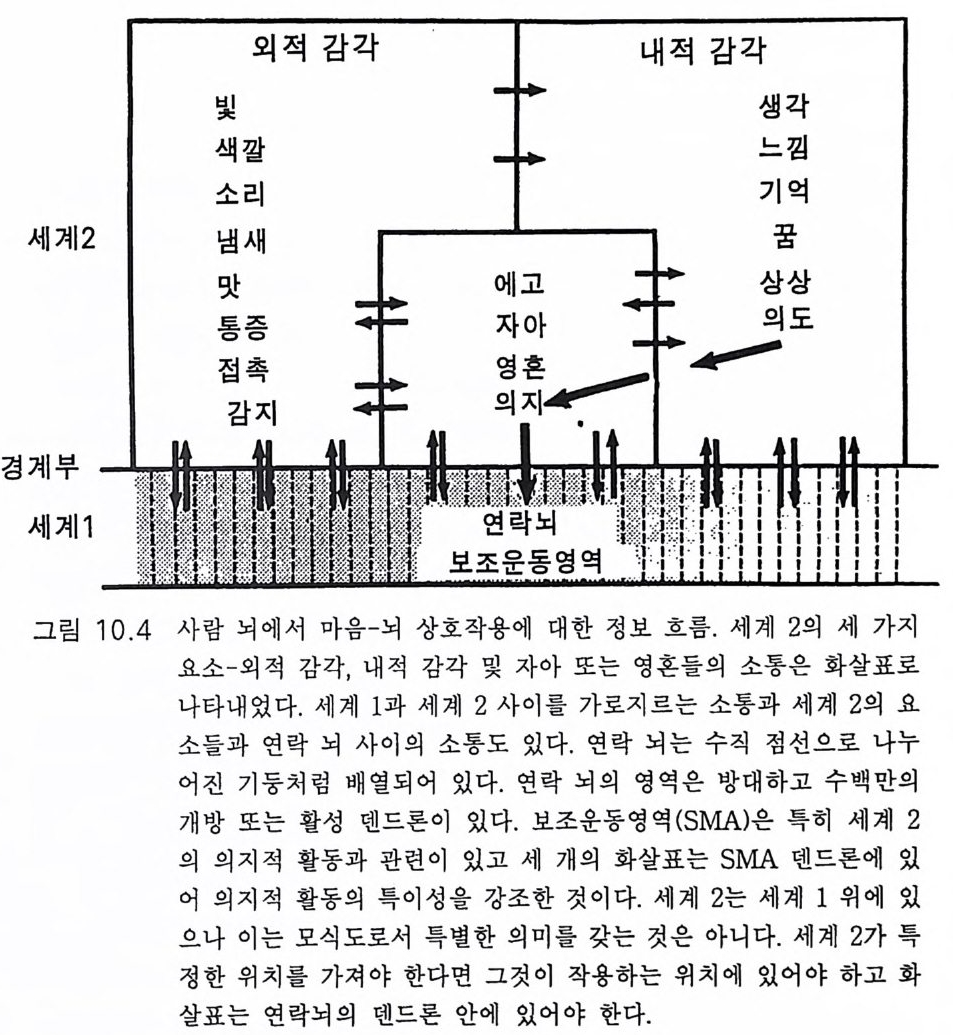 외적 감각 내적 감각
외적 감각 내적 감각
자유의지에 관하여 설 (Searle 1984:92) 은 다음과 같이 말하였다. <우 리가 행하였던 방식과는 달리 할 수도 있으리라는 믿음은 실제로 그 일을 한다. 그리고 이 믿음은 결국 도덕적 책임감에 대한 믿음과 인격 으로서의 우리의 성격과 연결된다.> 그러나 이어서 그는 말하기를
<우리가 근본적인 자유를 갖기 위해서는 우리는 마치 자연의 인과적 질서를 거스를 수 있는 자아가 우리 각자의 내부에 존재하는 양 가정 해 야만 할 것 이 다. > (1 984:92) 그래서 설은 물리학에 위·배되기 때문에 < 근본적 자유 >를 배척하였 댜 그러나 8.8 에서 자아는 뇌의 접합부 미세 부위에서 양자역학의 확 률 분야에 해당하는 작용으로 실제로 물리학을 위배한다고 제안하였 댜 따라서 물리학에 위배된다고 해서 의지의 자주성에 관한 믿음을 포기할 필요는 없다. 설은 결론적으로 다음과 같이 말하였다. 내가 정말로 이해하지 못하는 이유로 인해 진화는 우리에게 자주성의 경험을 부여하였는데 자유의 경험은……의식적 의도적 수의 행동의 기본 구조의 내부에 존재한다 . 이런 이유로 해서 우리의 행동이 자유롭지 못하 다는 것을 우리는 논의 를 통해서는 결코 납득하지 못한다(1 984 : 98) 우리는 유아기로부터 계속해서 우리들 행동의 자주성을 시험한다는 것을 알아두어야 한다. 만일 우리가 우리의 자주성이 침해되었다는 것 을 발견하게 되면 뇌의 병변을 의심해야 하고 신경과의 진찰을 받아야 한다. 도덕적 책임감이란 비인간 동물에게는 없다 . 그들이 수의적 동작을 하기는 하지만(그림 8.3) 그것은 도덕성과 비도덕성에 근거하여 행동하 기 이전에 그 행동의 결과에 대한 선악을 평가하는 것을 의미하는 <의지>로 행동한 것이 아니다.
10.6 인간 특성의 진화 유명한 인성학자인 데이비드 래크 Dav i d Lack 는 1961 년에 『 진화법칙 과 그리스도교의 믿음 』 이라는 책을 출판하였다 . 여기서 그는 이 책에 서 시도했던 것처럼 정 통 다윈학설을 지지하였다. 그리고 그는 눈 같 은 복잡한 구조의 진화를 < 생명력 > 이라든가 < 전체론적 주장>의 개 념으로 설명하려는 시도 ( 기독교도와 불가지론자)를 비판하였다 . 그는 다 음과 같이 결론지었다 . 자연도태에 의하여 진화가 이루어졌다 함은 < 무작위적 > 이 아니었고 자연법칙의 결과임을 의미한다. 이들 법칙이 진화과정을 엄격하게 결정 하였는가 아닌가 그리고 이 과정이 신성으로 계획되었는가 아닌가 하는 형이상학적 질문은 생물학의 범주를 벗어난다(1 961:71). 그는 계속해서 세 가지를 지적했다. 세속의 인본주의자들은 기독교적 신앙은 배척되어야 한다는 주장을 논 박할 것이다 . 그러나 고도의 도덕적 기준을 정당화한다거나 진화과정예 서 진실을 추출하는 것 이 불가능하기 때문에 사실상 그들 자신도 진퇴양 난에 빠져 있다 (196 1:1 09) . 만일 도덕성, 진실 그리고 개인적 책임감이 과학 밖에 있으나 값진 것 이라면 다윈학설로는 사람의 특성을 결코 적절하게 설명할 수 없을 것이 다(1 961:109). 도덕성, 진실, 아름다움, 개인적 책임감 또는 자아인식은 과학으로 설 명되지 않는다……따라서 인간의 경험 중에서 값진 중심 부분은 과학 밖
에 있다 (1961:1l4). 실제로 과학의 윤리와 과학의 철학은 과학 밖에 있고 철학의 범주 에 속한다. 인간 특성의 진화를 논함에 있어 자연도태의 법칙을 다윈과 함께 발견한 월라스의 의견을 참조할 필요가 있다. 다윈과는 달리 월라스는 안간의 지성은 우주적 지성 Cos mi c In t e llig ence 의 직접적 간섭에 의해서 만 설명될 수 있을 것으로 생각하였다. 월라스는 < 계간총설Q uar t erly Rev i ew }> 0869) 에 원시 인(그의 생의 대부분을 함께 살았다)에 관한 다 음과 같은 글을 써서 다윈을 분노시켰다. 자연도태는 원숭이보다 약간 우수한 뇌를 야만인에게 부여할 수 있 었을 뿐이다. 그러나 실제로 그들이 가졌던 것은 우리들 교육받은 사 회의 평균적 사람의 것보다는 약간 뒤떨어진 것이다. 이는 그가 일생동안 반복해서 추구한 명제였다. 자연도태에 의한 인간의 진화에 찬성하는 기독교도는 인간이 선과 악 이라는 영적 속성을 가졌다는 것을 추가해야 한다. 이는 진화의 결과가 아니고 초자연적인 것이다 .(Lack 1961 :11 5) 10.7 자아 또는 영혼의 창조 내가 견지하는 두 가지 전제는 우리들 자신과 타아롤 포함하는 세 계에 대한 우리들과의 관계를 이해하려는 시도에 대해 기초가 되는 것 이댜 그들은 일차적 확실성으로 분류할 수 있는 것이다. 첫째는 사람은 일관된 자아의식적 존재로서 존재한다는 확실성이다. 둘째는 신체와 뇌를 포함하는 물질적 세계가 존재한다는 확실성이다.
각자 자아의 경험적 일관성에 관계되는 문제는 현대 철학에서는 무 시되었다. 아마도 이것은 유물론의 만연에 기인한 것이다. 이는 영적 경험에서 야기되는 근본적인 문제에 어둡기 때문이다. 이제 우리들 경 험 세계의 가장 비상한 사건들에 관심을 집중하려 한다. 죽 특유의 자 아의식적 존재로서 우리가 존재하게 되었다는 사건이다. 이는 과학을 훨씬 초월한 기적이다. 인격이 자신의 독특성을 인식한다는 것은 의심할 바 없다. 그리고 이것은 사회생활과 규범의 기초로 받아들여지고 있다. 이 믿음에 대한 기초를 탐구해 나갈 때 현대 신경과학자는 이것이 신체에 의한 것이라 는 설명을 배척한다. 두 가지 대안이 남는다. 죽 뇌와 정신이다. 유물 론자는 전자에 찬성할 것이댜 그러나 이원론적 상호작용론자는 세계 2 의 자아(그림 9.5) 를 경험적 일관성을 가진 존재로서 간주할 것이다. 자아의 일관성을 유아론적으로 해결하려는 것을 부인하는 것은 중요 하댜 우리들의 직접적 경험은 물론 주관적인 것이고 전적으로 우리의 뇌와 자아에서 유도되는 것이다 . 타아의 존재는 주체들간의 소통으로 형성된다. 경험적 단일성이 뇌의 단일성에서 직접적으로 유도된 것이라면 인 간 뇌의 단일성의 수준을 조사해야 한다. 인간의 대뇌피질에 있는 100 억 개의 세포의 무한정한 자세한 연결 모두를 포함한 단일성일 수는 없댜 이들 연결은 가소성과 퇴행으로 끊임없이 변하는 것이다. 가장 보편적 유물론적 설명은 경험적 단일성이 유전적 단일성에서 유도된 다는 것이다. 이 설명이 의미하는 바를 결정적으로 검사하려는 시도는 없었댜 우선 제닝스(J en ni n g s 1930), 에클스 (Eccles 1970, 1'57 9 ), 도르프 (Thorp e 1966, 1978) 의 논의를 따르자면 이른바 경험적 단일성의 기초 를 이룬다는 단일 게놈g enome 은 적게 잡아도 5 만 개로 추산되는 사람 의 유전자 중에서 무한대로 확률이 낮은 유전자 제비뽑기(실패율 1015,000) 의 결과로 이루어진 것이다. 더욱이 뇌의 표현형적 발달은 워딩
턴 (Wad di n gt on 1969) 이 < 발달상의 잡음 develo p men t al no i se > 이라고 말 하였던 현상 때문에 유전형의 지령과는 상당한 거리가 있다고 스텐트 (Ste n t 1981) 는 지적하였다. 예를 들면 유전형은 뇌의 구축에 관련되나 이는 표현형적 구축과정을 현격하게 조절되도록 작용하는 환경 내에 서 일어난다. 일란성 쌍생아에서 동일한 게놈은 발달상의 잡음의 다양 성 때문에 서로 다른 뇌를 구축하게 된다. 워딩턴(1 969) 은 초유전적 설 계 e pig ene ti c landsca p e 와 양립되도록 크레오드 chreods 라는 개념을 개발 하였댜 초유전적 설계란 그가 만든 용어로서 초기 유전형적 지령이 뇌를 포함하는 표현형을 구축한다는 것이다 . 여기서 중요한 사실은 사 람 뇌의 기본적 연결성은 출생 전에 구축되며 접합부의 연결성은 상당 한 변화가 쉽게 일어날 수 있는 상태에 있어서 생애를 통한 학습과정 에 의해 발달한다는 것이다 (7.5.1). 따라서 접합체 z yg o t e 에 의하여 제공 되는 유전적 지령과 신생아의 뇌 사이에는 무한한 발달상의 간격이 있 댜 발달상의 잡음 때문에 우리의 유전적 단일성으로부터 우리의 경험 적 단일성을 유도하려는 어떤 시도도 혼돈되고 무의미하게 된다 . 그리 고 이미 언급하였듯이 이 시도는 개체의 일관된 게놈의 존재를 지배하 는 무한히 낮은 확률의 제비뽑기에도 직면하게 되는 것이다. 이 수수께끼에 대한 빈번하고 피상적으로 그럴 듯한 해답은 결정 요인이 생애를 통한 자아의 축적된 경험의 일관성이라는 주장이다. 우 리의 행동과 기억 그리고 실제적인 우리의 의식적 내면 생활의 전체 내용은 우리 생에 있어서의 경험 축적에 의한다는 것에는 이의가 없 댜 그러나 절박한 사정에 의해 발생할 수 있는 어느 특정한 쟁점에 있어서 변화의 폭이 얼마나 극단적이든 간에 한 살 또는 그 비슷한 연 령의 초기 기억까지의 기억의 계속성을 추적할 수 있는 동일한 자아, 상당히 다른 모습의 동일한 자아가 유지된다. 새로운 자아가 창조되지 도 않고 자아가 제거되지도 않는다. 유물론은 우리들 경험적 단일성을 해결하지 못하였으므로 자아 또
는 영혼의 일관성을 초자연적 영적 창조에 의한다고 나는 생각한다. 신학적으로 설명한다면 영혼은 신의 새로운 창조물이고 수정과 출산 사이의 어느 시기에 성장하는 태아에게 이식된 것이다 . 이는 < 신성 창 조 > 를 필요로 하는 단일한 개성(그림 9.5) 의 내적 핵심의 확실성이다 . 다른 어떤 설명도 있을 수 없다고 생각한다 . 환상적이고 불가능한 제 비뽑기에 의한 유전적 단일성도 아니고 개체의 단일성을 결정하지는 못하고 이를 변조시킬 수는 있는 환경적 개체회도 아니다. 이 결론은 측량할 수 없는 정도의 신학적 중요성을 가진다. 이는 사 람의 영혼 그리고 그것의 신성 창조에 의한 그 기적적 기원에 대한 우 리의 믿음을 강화하는 것이다 . 우리의 존재를 의존하는 초월적 신, 우 주의 창조자, 아인슈타인이 믿었던 신에 대한 인식뿐 아니라 우리의 존재 자체를 그에게 의존하는 사랑의 신을 인정하게 되는 것이다. 기독교적 다윈론자로서 래크는 자신의 믿음을 간결하게 표현하였 댜 이는 나의 믿음과도 가까운 것이나 다른 점은 나는 이룰 모든 고 위 종교에까지 확대하려는 것이다. 영혼이란 기독교에서는 사람의 영적 부분으로 간주하는 것이다.… ... 이 는 각 개인의 기본적 인간성에 묶여있는 것이고 각 영혼은 고유하다 . 이 는 또한 도덕적 결정과 합리적 결론을 내리는 책임을 지고 불멸한다. 영 혼이란 영성을 견지해야 하므로 이것의 존재에 대한 의문은 과학의 범위 밖에 있는 것이고 따라서 다윈학설과의 관계에서 문제가 생기지 않는다 …… 사람에게 동물처럼 그의 본질상 영적인 부분이 없거나, 아니면 기독 교직 관점에서처럼 인간이 최초로 출현하였을 때 초자연적 현상이 일어 났거나 둘 중의 하나이댜 그 최초의 출현 이전에 우리의 조상은 원시 인 간 포유류였고 그 이후에는 영혼이라는 신의 선물을 통하여 진정한 인간 이 되 었다 (1961:88-9).
호소력 있는 비유 그러나 비유 이상의 것이 될 수 없는 이론은 신체 와 뇌를 유전자 부호에 의하여 생물학적 진화의 훌륭한 과정을 통하여 구축된 비할 바 없는 컴퓨터로 간주하는 것이다. 이 비유에 있어 영과 자아는 컴퓨터의 프로그래머이다. 프로그래머로서의 우리들 각각은 태 생기 초기에 컴퓨터와 함께 태어난다 . 우리는 생애를 통하여 이를 발 전시킨댜 이는 생애롤 통하여 일어나는 모든 일을 함께하는 동반자 다. 이를 통해 다른 자아들을 포함하는 세계와 교류한다 . 가장 큰 미스 터리는 프로그래머로서 또는 경험적 자아로서의 우리들의 창조에 있 고 각 개안이 자신의 컴퓨터를 가지고 생애를 통해 우리들이 연합하는 데 있댜 이를 세계 1 과 세계 2 사이를 가로지르는 첩경으로 그림 9.5 에 도식화하였다.
후기 회고와 상상들 제 10 장에서 빼어난 다윈 학설적 진화론을 크게 혼동시켰는지도 모 르겠다 고식적 진화론들이 호미니드 진화의 마지막 단계에서 나타나 게 되는 자아의식에 대한 고려를 간과하여 왔다는 것으로 나 자신을 변호할 수 있으리라. 나는 여기서 이 심각한 간과를 보충하려고 시도 했을 뿐이다. 생물학적 진화에 있어 최고의 홍미의 대상은 인간의 기원에 있다. 다윈은 『종의 기 원』을 쓴 다음 『사람의 혈통』을 썼다. 나의 생 은 거 의 70 년 동안 이 책을 쓰기 위한 준비 과정이었다. 내가 진화적 기원에 관 한 문제에 매달리게 된 것은 내 나이 17 세 때부터이다. 호미니드 진화 는 바로 내가 어떻게 존재하게 되었는가의 이야기라고 생각하였다. 나 는 나의 상상 속에서 수백만 년의 호미니드 진화의 멀리 떨어진 조상 들이 밟아온 위험하고 희미한 진화 경로롤 느낀다. 되돌이켜 생각하면 이 경로는 나의 존재에 대한 유일한 기회가 아니겠는가. 이 경로는 진 정 우연과 필연의 자연적 과정이었는가? 아니면 어떤 신비한 초자연 적 유도가 있지 않았을까? 래크 (Lack 1962) 가 말했듯이 (10.6) 인간의 특 성에서 모든 영적 관점은 다윈 학설의 과학적 설명을 영원히 뛰어넘는 것이다. 이런 이유 때문에 나는 처음의 9 장에 걸쳐 신뢰를 가지고 견지했던 다윈 학설의 유물론적 설명에 목적론적 개념을 덧붙였다. 생물학적 진 화의 모든 변천에 최후의 목적이 있었음이 가정되어야 한다. <생물학 적 진화의 유물론적 사건 위에 영위된 신의 섭리가 있었음을 나는 믿
는다 > (Eccles 1979:235) 이는 결국 인간의 유전형을 창조하였다(그림 10.2). 이는 최종점을 가지는 때이야르적 Te i lhar di sm 궁극원인론과 관계되는 특수한 종류의 인류학적 원리이다 (Te i lhard 1959). 이는 도브잔스키의 『 궁극적 관심의 생물학 B i olog y of Ultim ate Concern 』 이라는 책에서 공감적으로 논의되 었다 (1967). 더욱이 기회주의적으로 생물학적 진화의 대설계를 깊이 생각하는 것은 적절하다 . 이 책에서 다루었던 호미니드 진화라는 최후의 대창조 기의 끝에 왔다고 결론지을 좋은 사유가 있기 때문이다 . 호모 사피엔 스 사피엔스의 진화적 첨탑까지의 가파른 등정을 이제까지 기술하였 댜 이는 진정 첨탑으로 보인다 우리는 적자생존에 의한 자연도태의 운영 종말에 있기 때문이다 (10 . 3). 결국 생물학적 진화 원칙에 따라 첨 탑으로부터 서서히 하강할 것임을 예견할 수 있다 (10.3). 나는 나 자신을 물리학과 무기화학적 법칙에 의하여 지배되는 생물 이 시작되기 전의 지구를 관찰하는 육체에서 분리된 영혼으로 상상하 기를 즐긴다 (Eccles 1979). 그러면 신비하고 무리없이 약 3.6b y BP 의 생 명의 기원이 나타나고 돌연변이와 자연도태에 의한 유전자 부호의 생 화학적 발달에 의한 상상할 수 없을 정도로 느린 단백과 핵산이 창조 된 생물학적 과정이 나타난다. 결국에는 해방되는 생물학적 창조성은 모든 상상력을 초월하는 것이 댜 밝혀지지 않은 수백만의 생물학적 종이 발생하고 번영하다가 종국 에 는 소멸 했다 .(Ma yr 1963, 1982, Sim ps on 1964). 누구든 모든 이 해 와 지 식을 동원하여 호미니드 진화에 대한 장차의 과정을 예측할 수 있었겠 는가라고 물을 수 있다 . 도브잔스키 (1967) 는 단호히 아니 라고 대 답했다. 어쨌든 수억 년이 지난 후에 생물학적 진화의 초자연적 드라마의 육체 에서 분리된 관찰자는 비교적 최근 (1000 만년 전)에 폰지드 혈통에서 호 미니드 혈통이 분리되어 나오는 것을 목격할 것이다. 이는 이 책의 시
나리오가 되었던 것이다 .(2.2) 생물학적 진화론자로서의 우리는 이 초 자연적 현상은 막연하게나마도 예측된 일이 없었음을 믿어야 한다. 생물학적 진화의 짧은 기간에 미래를 향한 창문이 열려 호미니드 계통이 떨어져 나와 오스트랄로피테쿠스를 거쳐 호모에 이르렀음을 짐작할 수 있다.(제 2 장) 의심할 바 없이 이 개방 시기 동안에 호미노이 드들인 이집토피테쿠스, 라마피테쿠스 그리고 드리오피테쿠스 사이에 는 대단히 복합적인 교섭이 있었을 것이다. 우리 모두가 짐작할 수 있 는 것 은 그 < 창문 > 은 오직 한 번만 열렸다는 것이다. 지금으로부터 장래에 이르러서도 어떤 폰지드 계통으로부터 호미니드 계통을 능가 하거나 이에 필적할 만한 새로운 진화적 계통이 시작될 가능성은 없 댜 그러나 호미니드로의 엄청난 귀결이 없었다면 지구는 소위 생물학 적 <서식 i n fe s t a ti on > 이 무한히 계속되었을 것이다. 이는 그 자체로서 최고로 훌륭한 것이나 개념적으로는 영원히 사멸한 것으로 간주되어 왔을 것이다. 호모 사피엔스 사피엔스를 창조한 자아의식의 문화적 진 화에 의해 주어진 초월적 계시나 의미에는 어떤 희미한 불빛도 없는 계속적인 암흑일 뿐이기 때문이다. 계통발생적 점진성의 점돌연변이를 넘어서서 돌연변이 과정 내에 전 지구적 대설계룰 포함한 가능성이 있지 않았을까 질문할 수 있다. 그리고 그것이 자연도태에 의해 검증된 생존을 위한 실제적 필요성을 넘어서는 사람의 뇌의 진화적 발달을 가져오지 않았을까? 이룰 왈라 스 Wallace 의 문제라고 할 수 있다 (10.6) . 호미니드 뇌가 자연도태를 초 월하여 필연적으로 점점 더 발달하게 되는 소위 유전적 역동g ene ti c dy n ami sm 이라고 할 수 있는 어떤 과정이 있지 않을까? 예를 들면 (2.6) 이는 겉보기에 독립적인 여러 지역에서 동시에 일어났던 것처럼 보인 다 크기가 1200cc 인 50 만년 전 북경인 Pe king man 의 뇌, 크기가 1200cc 까지 되는 30 만년 전 자바인J ava man 의 뇌, 전 네안데르탈인을 거쳐 네안데르탈인 유사한 것에서 호모 사피엔스 사피엔스에 이르는 9 만년
전 아프리카에 있어 호미니드 진화, 유라시아에서의 호모 프레네안데 르탈리스(그림 2 .1 2) 가 네안데르탈인으로 진화, 7 만년전 카프체Q a fz eh 에서의 호모 사피엔스로의 진화 가능성 등이다. 중국과 자바에서는 호 미니드가 멸종되고 후에 유라시아로부터의 호모 사피엔스 사피엔스가 이주하여 군락을 이룬 것으로 생각된다. 그러나 고고학적으로 좀더 탐 사해 보면 다론 설명이 가능할지도 모른다. 호모 사피엔스 사피엔스 종의 현존하는 세계 인구 분포에는 여러 가지 서로 다른 진화적 기원 이 있었을지도 모르겠다 (2.6, 2.7). 제 2 장에서 제 9 장까지 언급한 호미니드 진화에 관한 이야기는 지구 행성 아니 진정 우주 전체에 관한 어떤 이야기보다도 놀라운 것이다. 이는 휠러 (1974) 의 원저에서 다루었던 주제인 인간 중심적 원리의 기 초가 되는 것이고 폴킹혼 (Po lki n g horne 1986) 에 의해 간추려졌던 것이 댜 바로우와 티플러 (Barrow 와 Tip le r 1986) 의 최근 서적에서는 이를 지 나치게 발전시켰다고 나는 생각한다. 그러나 제 9 장과 제 10 장에서 언급 한 바와 같이 인간 중심적 윈리는 우리 각자가 단일한 자아의식의 존 재로 되는 사건을 통해서 새로운 의미를 갖게 된다. 이 초월성이 바로 마음과 뇌의 문제를 과학적으로 설명하기 위하여 뇌를 이해하려는 노 력의 절정에 달하는 이 책을 나로 하여금 생을 바쳐 만들게 했다. 인 간의 신비가 과학적 환원설에 따른다면 결국 신경 활동의 양상으로 간 주하게 되는 약정적 유물론의 주장을 통해 모든 영적 세계를 믿을 수 없을 만큼 격하하는 것이라고 나는 생각한다. 이런 주장은 미신으로 분류되어야 한다. 그림 9.5 와 10 .4에서처럼 우리는 영적 세계에 존재하 는 영혼을 가진 영적 존재인 동시에 물질 세계에 존재하는 신체와 뇌 룰 가진 물질적 존재임을 인식해야 한다. 인간 인성 (10 .1, 10.5, 10.6, 10. 7) 의 철학에 기초하는 인류를 위한 전언 올 우리는 어떻게 만들 수 있을까? 대단한 겸손을 필요로 한다. 우리 는 진화 기원의 초기부터 얼마 되지도 않은 지금 우리가 모든 지혜의
총본산이라고 상상해서는 안된다. 쉐링턴 (She rri n gt on 1940) 은 우리를 < 알에서 이제 막 깨어난 > 존재라고 비유해서 말했다. 현실론자적 입장은 우리들 각자는 달을 동반하고 인간 생활에 극도 로 해로운 대기와 기후 조건을 가진 다른 여덟 개의 행성과 함께 태양 주위의 궤도 안에 있는 지구 행성이라는 이 경이로운 공간 위에 자신 울 발견한다는 것이다. 그러나 우주복을 입은 인간이 이미 달을 정복 하였고 틀림없이 비슷한 방법으로 화성도 정복하게 될 것이다. 그 같 은 방문은 단지 아주 짧은 기간에 불과할 · 것이고 대단히 비싼 비용을 필요로 한다. 인간 생리학의 기초적 지식으로도 태양계를 벗어나는 또 는 우주의 어떤 집락에로의 우주 여행의 가능성은 영원히 배제할 수 있댜 인간은 영원히 지구에 귀속한다고 강조하여 말할 수 있다. 사후 에 영혼이 살아남아서 물리학과 생리학의 굴레에서 벗어나게 된다면 사정은 달라질 것이다. 그러나 우리는 사후에 어떤 일이 일어나는가에 관한 확고한 지식을 결코 가질 수 없을 것이다. 이는 인간의 한계라 할 수 있다. 여기에 공상과학 소설가들 그리고 심령 실험의 사기꾼들 에게 기회가 있다. 그들의 몽환경과 결과표의 조작을 허용하는 여지가 된댜 유명한 공상과학자가 과학소설가의 끊임없는 공격으로 그들의 예리한 판단력을 잃게 되는 것은 심각한 일이다 . 현실적으로 호모 사피엔스 사피엔스는 장래에 있어서도 이 축복받 은 주거인 지구 행성에서 무한한 세대 동안 존재할 것임은 인정해야 한다 . 할로우 샤프리 Harlow Sha p le y가 즐겨 표현했듯이 지구는 물질계 에 있어 보잘것없는 비교적 작은 바위 행성이다 . 지구의 궤도는 태양 계에서 평범한 것이고 태양은 1000 억 개의 별들로 이루어진 은하계의 한쪽 구석 가장자리에 있는 중간 정도의 크기를 가진 별이다. 이 온하 계는 또 1000 억 개의 은하계 중의 하나일 뿐인데 이들은 모두 170 억년 전의 대폭발 B ig Ban g로 생겨났다 . 하지만 인류학적 원리로 보면 지구 행성상의 우리의 주거는 대설계의 중심이다. 우리들 각자는 인간의 신
체와 뇌를 가지고 이 작은 천상의 별 지구 행성 위에 길어야 10 만년 전에 호모 사피엔스 사피엔스의 진화적 조상으로부터 깨어났다 . 우리 가 자아의식적 존재로서 촌재한다는 자체가 기적이다(제 10 장). 우주의 다론 곳에 생명이 존재하고 그것이 지능적 존재로 진화했을 가능성은 너무도 작기 때문에 생물학자들은 지구상의 인간 생명체는 더 논의할 것도 없이 고유한 것이라고 가정하게 된다. 생물학을 잘 모르는 천문학자가 환경이 적당하면 우주의 다른 곳에 서도 생명이 돋아나게 될 것이라고 제안하는 것은 그리 놀라울 일이 아니다. 더욱이 설혹 생명이 존재하고 진화한다 해도 이 책에서 언급했듯이 지능적 존재로 나타나게 되는 호미니드 진화상으로 귀결되지 않으면 그다지 깊은 의미를 가질 수 없다. 태양계를 벗어난 우주 여행은 영원 히 불가능하다. 그래서 세이건 Sa g an 과 드레이크 Drake 는 수소 방사선 의 파장에 부호 전언을 만드는 청취 장치를 개발하였다 . 아직까지의 기 록으로는 우주는 조용하다 . 나의 주제는 지구 행성을 소중히 여기는 데에 있다. 영겁을 통하여 계속되는 세대의 인간 인성으로서의 존재인 우리의 주거는 오직 지구 뿐이기 때문이다. 우리는 육체를 가진 자아의 상태로서 영원히 지구 굴레에 남아 있게 될 것이다 . 이런 의미에서 위에서 언급했듯이 우리 는 신체와 뇌의 죽음을 우리의 이원론적 존재의 해리로 간주할 수 있 다(그림 9.5). 전통적 기독교적 교리를 따라 폴킹혼 (Po lking home 1986) 이 제안했던 것처럼 해리된 영혼이 어떤 새로운 육체적 존재 안에 더 깊은 존재 의미와 더 황홀한 경험의 또 다른 미래를 찾게 되기를 희망 할 뿐이다. 종말론적 시각은 대종교들의 주제의 하나이다. 두 가지 근본적인 종교적 개념이 있다. 하나는 소위 대폭발과 그 결과 들이라고 하는 절묘한 정량적 설계로부터 시작된 (Lovell1978, Polk ing - home 1986) 근본적 자연 법 칙 들을 가진 우주의 창조자로서 의 신 이 다.
이 초월적 신 Tanscenden t God 을 아인슈타인은 믿었다. 또 하나는 우리 가 우리 자신의 모든 존재를 의존하는 내재하는 신 Immanen t God 이다. 어떤 신비한 방법으로 신은 진화 과정의 모든 생명체를 창조한다. 특 히 호미니드 진화에 있어 불멸의 영혼의 자아의식을 가진 인간 인성의 창조자이다. 희망을 찾고, 사랑, 진리, 그리고 아름다움을 추구하는 것을 본성으 로 하는 자아의식적 존재를 위해 그 물질적 기초인 뇌를 제공함으로써 생물학적 진화 과정은 그 자체를 초월한다는 것으로 우리는 결론지을 수 있다.
옮긴이 후기 사람은 누구나 머리뼈 안에 한 개씩 뇌를 가지고있다 . 사람에 따라 발휘되는 능력에는 차이가 있으나 그 사고 능력은 상상할 수 없을 정 도로 무한하다. 이처럼 생물학적 진화 과정 중에서 가장 중요한 창조 적 과정 인 뇌 의 진화 죽 900 만 내지 1000 만 년의 세월을 통하여 우리 의 유인동물 조상으로부터 걸출한 창조력을 지닌 인간의 뇌에 이르기 까지의 인간의 뇌의 진화에 관한 서적은 의외로 많지 않다. 이 책의 저자 존 에클스 박사는 저자 자신이 책의 후기에서 밝혔듯이 자신의 70 년 생애는 이 책을 쓰기 위한 준비 과정이었다고 말하고 있다 . 뇌의 진화 과정을 다루기에는 너무나 많은 사실의 공백이 있다. 그러나 에 클스 박사는 알려진 사실들에 대하여 철저하게 합리적으로 비판하고 그의 창조적인 상상력을 동원하여 뇌의 진화 과정을 전개하고 있다. 존 에클스 박사는 1903 년 오스트레일리아의 멜버른에서 출생하여 멜버른 대학에서 의학을 공부한 후 옥스퍼드 대학에서, 인간의 본성에 관한 연구의 권위자이며 뉴런의 기능에 관련한 발견의 업적으로 1932 년에 노벨상을 수상한 찰스 스코트 쉐링턴 교수에게서 사사하여, 1929 년에 박사학위를 취득하였다. 이후 존 에클스 박사는 인간의 정신에 관한 신경생물학적 연구 분야에 수많은 저서와 업적을 남겼고 1963 년 에는 신경세포의 홍분과 억제에 관련한 이온의 활동 기전을 밝혀 호지 킨 및 헉슬리와 공동으로 노벨상을 수상하였다. 존 에클스 박사는 금 세기 신경과학의 큰 획을 이룩한 학자의 한 사람이다. 에클스 박사가 옥스퍼드에서 연구할 당시에는 신경의 신호전달에 있어서의 전기적
기전에 대하여 강력하게 반대하는 입장이었다. 그러나 미세한 유리전 극을 이용하여 세포내 및 시냅스 전압을 기록할 수 있게 되면서 신경 계 시냅스에서의 화학적 전달기전에 확신을 갖게 되었다. 그리하여 홍 분성 또는 억제성 시냅스후 전위를 측정함으로써 신경전달물질에 반 응하여 발생되는 신경세포막에서의 이온기전을 밝히게 되었다. 에클스 박사는 이 미세전극기법을 소뇌의 시냅스 전달기전에도 적용하였고 이어서 시상, 대뇌피질 그리고 해마에도 적용하여 그의 저서이기도 한 『마음의 신경생리학적 기초』(1 953) 를 이해하기 위한 연구에 이어져서 15 편의 저서로 연구결과를 정리하고 있다. 죽 『신경세포의 생리학』 (1957), 『시 냅스의 생 리 학』(1 964), 『신경 기 구로서 의 소뇌 』 ( 1967), 『뇌 의 이해』(1 973), 『자아와 그 뇌 』 (1 977), 『포유동물 뇌의 분자신경생물학』 (1978), 『쉐링턴, 그의 생애와 생각』 (1979), 『인간의 신비』 (1979), 『인간 의 정신』(1 980), 『경이적인 인간됨』(1 984), 그리고 이 책 『뇌의 진화』 (1989) 로 에클스 박사는 일생의 연구를 정리하고있다. 에클스 박사는 뇌의 진화를 다윈적 진화론의 입장에서 설명하고 있 댜 다만 계통발생의 점진성만으로 해석하기에는 무리한 부분이 적지 않으므로 경우에 따라 한시적 평형설 또는 유전자 재조합의 가능성으 로 설명하고 있다 . 종의 발생 우선 종의 발생부터 살펴보았다 . 유전자는 세포가 살아가는 데 필요 한 여러 단백을 구성하는 아미노산 서열의 자세한 명령을 지니고 있 댜 사람의 경우 유전 정보를 구성하는 DNA 는 35 억 개의 핵산으로 이 루어졌다. 정상적으로 DNA 의 부호는 정확하게 복사되어 세대가 거듭 되어도 안정을 유지한다. 그러나 때로는 유전자 부호 내의 돌연변이가 일어날 수 있다. 이 같은 돌연변이는 대체로 개체의 생존이나 번식에
해로운 것이어서 자연도태 과정에서 제거된다. 다만 드물게 돌연변이 가 번식이나 생존에 유리할 수가 있어서 세대를 따라 전파될 수 있고 이 같은 돌연변이를 공유하는 생물계의 생존을 향상시킬 것이다 . 이것 이 다윈의 자연도태설 또는 적자생존론의 기초 이론이다 . 이 같은 점 진적인 돌연변이와 자연도태 과정에 의한 선조 집단의 변형으로 주거 지역 내에 비상관성 집단을 이루는 새로운 종이 형성된다고 한다. 이 를 계통발생의 점진성 또는 공통 조상적 연속성이라고 한다 . 그런데 화석 기록에 의하면 이 같은 점진성은 찾아볼 수 없고 예리한 돌변이 있었음을 나타낸다. 그래서 한시적 평형설에 의한 타조상적 종화라는 이론이 개진되었다. 죽 국소적 소집단이 모집단의 지리적 영역으로부 터 변두리로 격리될 때 새로운 종의 발현이 일어날 수 있다는 것이다. 작은 집단은 커다란 모집단의 유전자 흐름에서 격리되었을 때 분화가 효과적이고 빠르게 일어날 수 있다는 것이다. 그리고 격리집단은 유전 자 흐름의 혼합 효과를 감소시키는 효과도 있다는 것이다. 이 한시적 평형설에 의하면 어떤 도태 조절에 앞서 도약적 변화가 일어나고 새로 운 종이 완전하게 형성된 후에 도태 조절기전이 작용할 것이라는 것이 댜 즉 개체발생은 도태에 선행한다는 것이다. 여기서 인간 진화를 화석 기록을 중심으로 살펴보았다. 이집트 사막 의 화이움 평원에서 지금으로부터 3000 만 내지 3500 만년 전의 것으로 추산되는 이집토피테쿠스의 화석이 발견되었다 . 이것은 수상서식 유인 동물의 화석으로서 현대의 폰지드와 유인류의 공통의 조상으로 생각 하는 것이다. 분자시계 연구방법에 의하면 호미니드와 폰지드의 분파 시기를 지금으로부터 500 만 내지 1000 만년 전으로 잡고 있다. 화석 기 록에 따르면 호미니드의 진화는 아프리카에 국한하여 나타난다. 여기 서 발견되는 오스트랄로피테신의 골격은 두발걷기였음을 나타낸다. 오 스트랄로피테신은 호미니드 진화의 이어달리기 첫번째 주자로 간주될 수 있고 일관되게 정확한 유전자의 운반 역할을 수행하여 탁월한 호미
니드 진화 이어달리기의 다음 주자인 호모 하빌리스에게 배턴을 넘긴 댜 호모 하빌리스는 석기문화가 시작되어 올도완 문화를 만들었고 체 구는 현저하게 커지지 않았으나 뇌는 현저하게 커 졌 다 . 호모 하빌리스 는 인간 진화에 있어 새로운 시기의 시작을 점하고 오스트랄로피테신 으로부터 불연속적인 정량적 도약 전진을 이룩함으로써 이루어졌다고 한댜 다음의 커다란 진화는 호모 에렉투스로의 발전이라고 한다. 이 들의 화석 발견 장소로 미루어 호미니드의 첫번째 대이동이 있었다고 보는 것이다. 그리고 좀더 발달한 아슐리안 석기문화를 이루었다. 동 아프리카에서는 10 만년 이상에 걸쳐 호모 하빌리스가 독자적 석기문 화를 가지고 호모 에렉투스와 공존하였다고 생각한다 . 1856 년에 호모 사피엔스에 속한다고 생각되는 화석이 독일의 네안데르 지방에서 발 굴되었다. 이를 네안데르탈 또는 호모 사피엔스 네안데르탈렌시스라고 하는데 유럽과 근동 여러 지역에서 대단히 유시한 화석들이 발견되었 댜 이는 호모 사피엔스의 중요한 조상일 것으로 인정되는 것이다. 이 어서 현대인을 통칭하는 호모 사피엔스 사피엔스로 대치되는데 그 발 생 장소는 아직도 의문으로 남아 있다. 사람이 호미니드의 후손이라는 생각은 넓은 지역에 걸친 여러 종족의 미토콘드리아 DNA 분석 결과 로 뒷받침되고 있다고 하나 아직도 비판의 여지는 있다 . 뇌의 진화 지난 300 만년 동안에 있었던 뇌의 현저한 진화적 팽창은 진화의 한 시적 평형 과정에서의 도약적 유전자 변화의 결과라고 생각한다. 그리 고 뇌 기능의 점진적 중가는 자연도태 과정에서 중심적 역할을 했다고 생각한다. 이 같은 뇌의 진화를 뇌의 각 부위별 크기 지수를 식충동물을 기준 으로 영장류의 뇌 부위별 크기 지수와 비교하여 살펴보았다. 대체적으
로 뇌의 진화적 발달은 정량적인 것이고 정성적인 것은 아닌 것 같다 고 생각하는 것이다. 뇌에서 척수를 거쳐 운동신경을 이루는 척수로의 기본 원리는 유인원이나 원숭이의 뇌에서 크게 다룰 것이 없는 것이 댜 추체로 이외에도 운동신경을 이루는 또 다른 신경로가 있다(예 적 색척수로) . 그러나 진화 과정에서 추체로가 유리하게 작동할 수 있도록 그 같은 신경로의 일부는 소멸하는 것이다. 골격 변화를 가져온 중요 한 진화적 발달로는 직립자세를 들 수 있다, 화석 기록으로 미루어 호 미니드인 오스트랄로피테쿠스 아프리카누스의 척추, 골반 및 뒷다리의 골격 구조는 이미 직립자세에 적합하도록 발달되었다고 생각한다. 한 편 두발걷기는 사람 특유의 것으로 지면 위를 효과적으로 이동하는 방 법이다 북탄자니아 레톨리 하상의 발자국 화석은 360 만년 전 오스트 랄로피테신의 것으로 호미니드 조상의 초기 역사를 말해주는 것이다. 이 발자국은 잘 훈련된 두발걷기를 나타내고 있으며 사람과 유사한 큰 엄지발가락을 보여주고 있다. 이것은 사람이 맨발로 모래 위를 걸은 발자국과 매우 유사하고 폰지드의 것과는 분명히 다르다. 오스트랄로피테신이 직립자세를 취하기 위해서는 대단히 복잡한 중 추신경계의 변화가 있었을 것이라 생각한다. 불행히도 양각 기립에 관 계되는 신경기전에 대한 지식은 충분치가 못하다. 한편 두발걷기는 기 저핵, 소뇌를 비롯한 뇌의 여러 부위가 고리를 이루어 영향을 미치는 대뇌피질의 율동적 방전이 기본 기전이다. 오스트랄로피테신은 두발걷 기로의 운동 기능의 진화로 이 같은 신경기전이 개조되었다고 생각되 는 것이다. 모든 운동의 섬세한 조절에는 긴-고리 반사 따위 운동의 자동제어 기전이 중요한데 이것의 점진적 진화는 호미니드의 손재주 롤 이루는 데 필수적이었음은 석기문화로 미루어 알 수 있다. 오스트랄로피테신이 두발걷기를 함으로써 앞다리(팔)가 보행의 의 무로부터 해방되어 다양한 활동에 이용할 수 있게 된다. 이는 손의 변 화에서 뚜렷하게 알 수 있는데 엄지가 발달하고 회전하여 정확하게 잡
을 수 있게 된댜 그러나 오스트랄로피테쿠스 아프리카누스는 석기를 만들어 쓰지 않았다고 생각된다. 그들의 뇌는 능숙한 손재주로 석기를 다듬어 만들 만큼 발달하지 못하였을 것으로 추측된다. 폰지드도 잘 조절된 악력을 가지고 있으나 모자라는 것은 손재주라기보다 대뇌의 능력이라 생각된다 . 이것이 호모 하빌리스, 호모 에렉투스, 호모 사피 엔스로 진화하면서 각 단계마다 특유의 석기 종류와 멋을 가진 석기문 화가 발달한다. 이는 대뇌반구의 진화적 발달에 연유하는 운동 능력과 창의력이 점차로 향상되었기 때문으로 생각하는 것이다. 언어의 발달 호미니드 진화에 있어 언어의 발달 과정은 대단히 중요한 의미를 갖는댜 뷸러와 포퍼는 언어를 네 가지 수준으로 분류하였는데 표현적 또는 징후적 기능과 방출 또는 신호적 기능인 두 가지 하급언어는 사 람과 동물이 공통적으로 가지고 있으나 기술적 기능과 논쟁적 기능의 두 가지 고급언어는 유독 사람만이 구사할 수 있다고 하였다. 언어의 네 가지 수준은 어린아이의 발육 과정에서 잘 나타난다. 즉 초기의 표 현적 수준에서 신호 수준을 거쳐 서술 수준을 지나 결국 논쟁 수준으 로 단계적으로 각 수준을 극복해 나간다. 호미노이드 진화에 있어 침 팬지가 사람과 가장 가까운 근친이라고 보는, 죽 인간을 영장류로서 그리고 유인원의 근친으로서 계통발생적 체계에 확립시킨 것은 다윈 주의적 혁명이라고 하였다. 단지 약간의 축 }량적 차이는 있으나 여러 각도에서 연속성을 보이는데 이 같은 순조로운 이행 과정을 설명하는 데 한 가지 장애는 인간 언어의 유일성이라는 것이다. 사람의 지능이 아주 낮은 수준일 때 그리고 육체적 또는 사회적 장애에도 불구하고 사람의 언어 발달은 사람 이외의 다른 종에게는 능력 밖의 것이라는 것은 여러 연구자들에 의해 거듭 확인되었다.
사람의 뇌에는 좌측 대뇌피질에 언어에 관계되는 두 개의 커다란 영역이 있다. 베르니케의 후방 언어중추는 언어의 관념화에 특히 관계 가 있고 브로카의 전방 언어중추는 운동성 언어중추라고 할 수 있다. 오랑우탄이나 침팬지의 뇌에는 사람의 언어 수준에 이르는 특수한 대 뇌피질 영역이 없거나 충분히 발달되어 있지 않다. 중요한 것은 사람 의 언어영역은 출생 전에 이미 형성되고 쉽게 언어를 배울 수 있도록 개체발생학적으로 발달한다는 것이다. 고등 영장류에서 호모 사피엔스에 이르는 진화과정에서 대뇌피질에 서 일차 및 이차 청각영역에 이르는 청각 계통에는 진화적 변화가 별 로 없었다고 생각한다 . 그러나 브로카의 전방 언어영역에 해당하는 영 역은 유인원에는 없다. 이 같은 계통발생학적 과정의 각 단계는 두개 골 화석의 내부 충전 자료에서 찾아볼 수 있다. 오스트랄로피테신의 뇌는 장차 하전두엽의 브로카 영역이 될 부위의 작은 충만이 있을 뿐 이고 호모 하빌리스에서는 하전두엽의 충만이 뚜렷하여 언어영역의 계통발생학적 발달이 시작되었음을 나타내고 호모 에렉투스에서는 전 방 및 후방 언어영역의 발달이 진전되어 네안데르탈인에 이르러서는 호모 사피엔스 사피엔스의 수준에 이름을 나타낸다. 베르니케의 언어 영역에서 영역 39, 40 은 사람의 뇌 진화 과정에서 유일하게 새롭게 나 타나는 구조라고 생각한다. 앞 사람의 발자국을 정확하게 따라 밟은 레톨리 하상의 발자국 화석은 오스트랄로피테신에게 우수한 대뇌 활 동이 있었다는 증거이고 수준 3 언어의 시작까지도 생각할 수 있는 것 이댜 호모 하빌리스의 뇌는 40% 나 커지고 전방 및 후방 언어영역의 존재는 큰 유전적 변화로 호모라는 새로운 속의 시작이라 할 수 있다. 호모 하빌리스는 언어 능력이 있었다고 생각되는 데도 불구하고 그 인 구는 크게 증가하지 못하였다고 생각된다. 수십만년 후 호모 에렉투스 에 이르러 언어영역이 크게 증대하는 유전자적 변화로 새로운 종을 이 루면서 넓은 지역으로 확산되었다고 생각한다.
시각계의 진화 호미노이드로부터 호모 사피엔스에 이르는 시각-운동계의 진화적 발달을 검토하기 위해 유인원의 뇌를 대상으로 시각정보 처리과정에 관한 대단히 많은 연구가 이루어졌다. 마카크 원숭이의 시각계에 관한 연구가 가장 철저하게 이루어졌다 . 이는 호미니드 진화 초기의 시각계 모델로 간주할 수 있는 것이다. 이것의 눈은 앞으로 향하였고 완전한 양안시이며 그 시각경로는 시신경 교차에서 부문적으로 교차하여 양 쪽 눈의 우측 시야는 외측 술상체 내의 접합 연결을 거쳐 좌측 후두엽 의 일차 시각피질에 투사된다. 왼쪽 시야는 그 반대가 된다. 미세전극 을 이용하여 단일 신경세포에서 발생하는 임펄스를 기록할 수 있어서 일차 시각영역에서의 조사에 의하면 비슷한 감수성 방향을 가진 세포 들이 조각보 모양으로 피질 표면에 대하여 수직으로 종대를 이루는 경 향이 있다는 것이다 . 그리고 영상의 재구성에는 일차 시각영역 주위의 이차 및 삼차 시각영역의 신경세포들이 참여하여 입체 영상과 그 형 태, 방향, 움직임과 색깔 둥 특수한 시각 기능에 대한 반응을 분석할 수 있었다 . 다양한 종류의 시각 활성에 관한 연구의 일반적인 결론은 유인원 마카크 원숭이와 올빼미 원숭이 그리고 사람의 후두, 측두, 두 정 및 전두엽에 있어서의 반응에는 현저한 상동성이 있다고 할 수 있 다는 것이다. 망막과 의측술상핵을 거쳐 일차 시각피질에 이르는 시각 경로는 호모 사피엔스 진화 과정 초기에 이미 완전한 발달이 이루어졌 음을 뜻하는 것이다. 즉 호미니드 진화 초기에 대단히 효율적인 시각 계통을 물려받았다. 다만 사람의 대뇌피질에서 시각 활동에 의하여 활 성화되는 영역은 원숭이에 비하여 훨씬 크다는 것이다 . 다시 말해 전 체 시각피질영역은 유인원에 비해 사람이 세 배 이상 크지만 실제 기 능이나 설계에 있어서는 근본적인 진화적 발달은 없었다고 생각하는 것이다.
인식체계의 발전 호미니드는 전두전엽이 충분히 발달하지 못하여 동기와 집중력의 결함이 있었을 것이라고 한다 . 호미니드 진화의 문화적 발달은 석기문 화 같은 내구성 도구로 추정되는 것이다. 오스트랄로피테신은 잘 발달 된 목기문화가 있었을 것으로 추측된다. 그러나 그것들은 화석으로 혼 적을 남기지 않았다. 호모 하빌리스의 올도완 석기문화는 수천년 동안 지속되었다 . 진화 정체기로 보는 것이다. 그러고 나서 호모 에렉투스 의 유전적 발달로 아슐리안 문화로 진입하여 도구만들기에 개선이 있 었다 이어서 호모 네안데르탈리스의 무스테리아 문화로 이어지는 각 단계의 석기 기술은 매우 미미한 진보를 보이면서 수백 또는 수천년 동안 계속된다. 이 같은 낮은 수준의 진보는 변연계 특히 내측편도체 와 관계되는 전두전엽의 발달이 미미한 수준이었기 때문으로 생각하 는 것이다. 호모 사피엔스 사피엔스의 구석기 시대(약 3 만년전) 초기에 이르러 인식체계라는 놀라운 발전을 이루었다고 생각한다. 한 가지 중거는 프 랑스의 브랑카드에서 발견된 판화이다. 두달 이상에 걸친 달 모양의 변화를 골패에 눌러 그린 판화이다. 이것은 여러 의미가 있다. 판화를 그리기 위한 정교한 도구의 발달, 주기성의 개념, 나아가 부호화된 문 자언어까지도 생각할 수 있는 것이다. 스테빈스는 사람의 특이적인 세 가지 특성은 기공성, 의식적 시간결 합 그리고 상상력이라고 하였다. 석기문화로 대표되는 도구문화를 성 형적 기술 창조성의 시작으로 보는 것이다. 성형적 기술에 있어서 사 람은 호미니드 진화 이전에 이미 발달되었던 호미노이드의 대뇌 영역 들을 효과적으로 이용하였다고 본다. 이 같은 진화를 선행 진화라고 한다. 전두전엽 그리고 측두 및 두정엽 영역들의 진화적 발달은 호미 노이드에서 이미 발달되어 있었던 시각 - 운동 계통을 효과적으로 이용
할 수 있게 하였을 것이라는 것이다. 이는 괴리 진화라고 하는 것의 한 가지 예이다 . 성형적 기술인 건축, 조각, 도예 , 그림 , 융단 둥은 모든 인간 문화의 평가와 인식의 기초라 할 수 있고 창조적 상상력을 들여 다보게 하는 것이다. 변연계는 신경중추의 복합체이고 중추간의 소통회로이다. 이는 진 화적 생존 그리고 그 결과로 이루어진 호미니드 진화에 중요한 역할을 한다 변연계를 구성하는 편도체, 시상하부, 중격핵 및 수조 따위의 동 물 실험을 이용한 자극 실험 그리고 이들 구성분의 크기 지수로 미루 어 보면 변연계의 진화는 쾌락과 관계되는 성분은 증강되고 공격성에 관계되는 성분은 발달하지 않은 채로 남아 있는 것이라고 생각된다. 한편 애타심이라는 것은 의지적인 것으로 그 행동은 자동적이기는 하 지만 계획적이며 타인의 이익을 고려하여 계획된 것이라야 한다는 것 이댜 호미니드 진화상에서 애타심의 흔적으로는 360 만년 전 레톨리 하상의 발자국 화석을 들 수 있다. 네안데르탈인의 매장 습관에서도 찾아볼 수 있다. 여기서 에클스 박사는 그의 스승 쉐링턴의 의견에 동 의하면서 생물학적 진화는 단지 우연과 필연만은 아니라고 하였다 . 기 술적 진보뿐 아니라 가치의 창조와 발달 때문에 사회 구성원의 복지에 헌신하는 애타심이라는 도덕적 기초를 이루는 것이라고 하였다. 학습과 기억 학습이란 기본적으로 뇌에 저장되는 과정이다. 그리고 기억이란 뇌 의 자료 은행에 저장되어 있는 것을 상기하는 것이다. 많은 경우 혼동 하여 취급되기는 하나 두 가지 분명한 형태의 학습과 기억이 있다. 첫 째는 운동 학습과 기억이다. 선다거나 걷기 따위의 능숙한 움직임의 학습이다. 둘째는 인지적 학습 또는 기억이라고 하는 것이다. 여기에 는 모든 지각, 관념, 언어적 표현이 포함되고 실제로는 문화를 통틀어
그 안에 나타낼 수 있는 것이다 . 사람에서 인지적 기억능의 엄청난 증가를 고려할 때 두 가지 중요 한 요인이 있다. 첫째 연합피질 특히 전두피질엽이 크게 증가하였다는 것이다 . 브로드만 (1912) 은 전두전엽의 전체 신피질에 대한 비율을 호모 의 경우 29% 로, 침팬지는 16.9% 로, 마카크는 1 1. 3% 로 하였다. 전체 신 피질의 상대적 크기에서도 사람의 전두전엽피질은 유인원의 것보다 5 배나 크다. 둘째 언어의 역할이다. 사람의 인지적 기억은 대부분 언어 로 작성된다. 인지적 기억에 대하여 해마가 상대적으로 크게 기여하지 못한 것(표 7.1)에 비하여, 인지 능력의 발생에 있어서 이들 두 가지 측 면의 진화상 발달은 훨씬 중요한 것이라 할 수 있다. 호미니드 진화에 있어 호모 하빌리스는 뇌가 증대하고 언어 영역이 발달하여 가장 큰 진화적 도약을 이루었다고 생각하는 것이다 . 이는 또한 석기 사용에 기초한 문화를 창시하였다. 이 문화는 인지적 기억의 개선에 의존하였 을 것임을 짐작할 수 있다는 것이다. 신피질의 호미니드 진화에 있어 언어영역 특히 브로드만 영역 39 와 40 이 흔적에 지나지 않는 유인원의 뇌로부터 크게 증대하였다는 것은 이미 언급하였다 . 그리고 이 후기 진화의 기원은 대단히 늦은 수초화 로 미루어 짐작할 수 있다. 현대 유인원을 모델로 한 호미노이드에서 부터 전두엽이 굉장하게 진화적으로 커졌으나 (5 배) 일률적으로 커진 것은 아니라고 말할 수 있다. 영역 39, 40 처럼 새로운 영역이 특별한 목적을 가지고 발생하여서 전두전엽이 인간의 사고와 기억에 두드러 진 역할을 하게 된 것일 수도 있다. 이들 영역 중 일부는 전두전엽 지 도에 있는 것일지도 모른다. 언어학적 연구로 어린아이와 어린 침팬지의 근본적인 차이는 언어 에 있음을 알게 되었다. 어린아이는 의욕을 가지고 언어의 세계로 들 어가고 문법적으로 바른 문장 구성을 탈리 배운다. 그렇게 해서 세상 을 배우게 된다는 것은 의문의 여지가 없다. 테라스는 인간의 학습과
기억에 있어 언어의 핵심적 역할을 강조하면서 다음과 같이 말하였다 . 호미노이드(침팬지 같은)로부터의 진화에 있어 학습이나 기억에 대 한 고도로 발달한 대뇌기전은 이미 발달될 준비가 다 되어 있었다 . 하 지만 사람의 수행 능력은 차원이 다른 정도의 차이가 있다 . 침팬지가 습득한 수행 능력은 세 살짜리 어린아이보다 못한 수준이다 . 사람 성 인의 수행 능력과는 비교가 되지않는다. 그리고 이는 살아가며 계속 발달되고 문자기록, 컴퓨터 기억, 또는 수학적 부호 등의 불변하는 형 태로 습득한 학습 내용을 이용할 줄 안다. 마음과 뇌 고동동물 특히 영장류는 의식을 가졌다고 생각할 수 있으므로 이 책에서 특별히 관심을 가지는 문제는 호미니드 진화에서 뇌가 크게 발 달하는 동안에 마음-뇌의 상호작용에 있어 어떤 변화가 일어났는가에 있다. 유인원에 관한 과학적 연구는 결여되어 있으나 원숭이나 바분에 관 한 발전된 연구는 호미노이드 조상의 모델로서 받아들여지고 있다. 시 각, 청각, 촉각 둥 조절된 감각 입력의 실험 조건에서 고등 포유류의 의식하 경험을 철저하게 연구하는 것은 가능하지 않다. 그러나 몇 가 지 일화적 증거는 있다. 그들은 자신의 경험을 표현하는 언어 능력이 결여되어 있다. 그러나 물론 언어의 두 가지 하위수준을 이용하여 이 들 경험을 신호로 나타낸다. 이에 반해 고등 영장류의 뇌는 촉발 감각 입력과 독립적으로 동물에 의해 수행되는 수의적 결정이 있을 때 연구 될 수 있고 이는 뇌에 대한 마음의 작용에 관한 것이다. 양자 물리학자 마르게노의 최근의 저서는 근본적 기여를 하였다. <양자역학의 확률계와 같은 어떤 장(場)들은 에너지도 물질도 지니지 않는다>고 한 그의 주장은 19 세기 물리학으로부터의 주목할 만한 변
환이다 그는 계속해서 다음과 같이 말했다 . 뇌와 같은 대단히 복합적 물리 계통에 있어서 뉴런과 감각기관 그 리고 그 구성분은 확률적 양자법칙에 의하여 지배되기에 충분할 정도 로 작댜 이런 경우에, 물리적 기관은 항상 각각 분명한 확률을 가지고 다양한 변화가 가능한 상태에 있다. 만일 다른 것에 비해 다소의 에너 지를 필요로 하는 어떤 변화가 일어나면 복잡한 유기체는 자동적으로 그 에너지를 공급한다. 마음 그 자체가 에너지를 공급할 필요는 없다 . 결론적으로 마르게노는 다음과 같이 말했다. 마음이란 적합한 물리적 반응계라고 할 수 있다. 그러나 비물질계로 서 가장 근접한 상사형은 아마도 확률계일 것이다. 마음-뇌 상호작용의 이원론적 가설을 좀더 정확하게 규정하는 데 있어 초기 개념은 심적 사건의 세계 전체(세계 2) 는 물질 구 에너지의 세 계(세계 l) 나 마찬가지로 자율적인 존재를 갖는다는 것이다. 현재의 상 호작용론적 가설은 이들 존재론적 문제와 관계가 없고 다만 심적 사건 의 신경 활동에 대한 작용 양상에 관계되는 것이며 마르게노의 가설에 따르면 마음소니 상호작용은 양자역학의 확률계가 상사형이라는 것이 다 . 이것은 질량도 없고 에너지도 가지지 않으나 미세 국소에서 효과 적인 작용을 유발할 수 있다는 것이다. 더 특이적으로는 의도나 계획 된 생각에 관련되는 마음의 정신 집중은 양자역학의 확률계와 상人}형 인 과정으로 신경 활동(사건)을 유발할 수 있다고 제안하였다. 미세 국소설에 따르면 마음소니 상호작용은 뇌의 흥분성 시냅스의 두 가지 주목할 만한 특징과 밀접한 관계가 있다. 첫째 접합전 소포망 의 구조와 배치는 보통 하나의 부톤에 단지 하나의 접합전 소포망만을 갖도록 되어 있다. 둘째 접합전 소포망으로부터 시냅스 소포의 방출은 확률적 함수로 결정되는데 이는 l 이하이며 때로는 훨씬 낮으며 상하 로 조절될 수 있는 것이다. 호미니드가 자아의식을 갖게 되는 호미니드의 진화적 발달에 있어
심적 사건에 의한 신경적 사건의 조절 방식에 있어 근본적인 변화는 없었다고 제안할 수 있다. 미세국소 가설은 호미니드 진화에서 발달하 는 마음의 자아의식에도 옹용될 수 있는 것이다. 래크 Lack 와 로렌츠는 둘 다 자아의식의 진화적 출현에 있어 영과 신 체 사이에는 건너갈 수 없는 간격 또는 심연이 있다고 말하였다. 하지 만 종국적으로는 정신 또는 영혼의 완전한 출현을 이루는 중심적 핵심 의 창조와 발달을 상정해야만 한다 . 호미니드 진화의 계통발생학적 과 정에 있어 상황 사이에는 사람의 유아에서 소아를 거쳐 성인에 이르는 개체발생학적 과정에서와 마찬가지로 모든 종류의 변이가 있었다고 생각된다. 그러나 그것은 아직 우리의 이해를 초월하는 것이다 . 개인의 최초의 기억으로부터 기억의 계속성이 유지된다는 것에 의 해 인식할 수 있듯이 심적 단일성이 주관적으로 존재한다는 것은 사람 의 보편적인 경험이다 . 이것이 자아라는 개념의 기초가 되는 것이다 . 1977 년에 포퍼와 에클스는 자아의식적 마음은 신경 활동을 수동적 으로 읽어내기만 하는 것이 아니고 융이 말한 탐조등 바유처럼 능동적 탐색 활동이 있다는 가설을 수립하였다. 자아의식 앞에 즉시 즉시 복 합적 신경 과정 전체를 잠재적으로 나타내거나 묘사한다. 그리고 주의 집중과 선택 그리고 홍미 또는 의도에 따라 이를 연락뇌에서 일어나는 방대한 활동의 합주로부터 선택할 수 있다 . 그래서 이것 저것을 탐색 하게 되고 연락뇌의 여러 영역의 판독 결과를 종합한다. 이 같은 방식 으로 자아의식적 마음은 경험을 하나로 통합한다. 이 상호작용은 쌍방 통행 과정이다. 신신피질 인지적 피질영역의 개체발생학적 발달의 특성은 신피질 생성 과정 에 있는 97 일된 원숭이 태아의 신경관에서 알아볼 수 있다. 사람 태아
의 신경관도 비슷하리라고 생각되나 다만 중요한 차이는 신피질에 더 하여 신신피질의 신경 발생에 관련되는 특별한 부분이 있을 것이라는 점이다. 그 같은 부분에서는 신경상피세포의 미숙신경세포를 생성하기 위한 세포 분열이 늦울 것으로 생각한다 . 이들 신경세포는 특정 신신 피질영역을 구성하는 과업에 적합한 넓은 범위의 특별한 유전적 기질 울 가지고 있을 것이라 생각한다. 이는 아마도 고도의 특이적 기능을 나타내기 위한 신경세포의 단세포군을 생성할 것이다. 신신피질에는 다섯 가지 특수한 특징이 있다. ® 계통발생학적으로 최후로 출현한 것이고 호미니드에서의 특별한 발 달이다 @ 개체발생학적으로 지연성 수초화와 지연성 수상돌기 및 시냅스의 발달에 의해 최후로 성숙된다. ® 언어 및 공간 시각 그리고 음악 둥에 있어서 기능적 비대칭성이 있다. ® 어릴 때에는 손상을 보상하는 둥의 성숙상의 가소성이 있다. ® 신신피질의 활성화는 의식, 자아의식, 생각, 기억, 느낌, 상상, 창조 등 넓은 범위의 인지적 기능을 수반한다. 켜;三: 로L.... 이제까지 뇌라는 관점에서 호미니드 진화의 기본적 특성을 살펴보았 다. 한편으로는 우연적 유전적 돌연변이가 중심 역할을 하고 다른 한편 으로는 자연도태의 <체 s i eve> 를 이용한 표현형의 <걸러짐 s ifting>이 핵심적 역할을 한다는 철저하게 다윈적인 설명이었다. 약 35 억년 전 호미니드 진화에 있어 전혀 새로운 현상이 나타나 포 유류와 조류 동 고등동물의 행동 양상으로부터 알 수 있는 의식 상태 가 있었을 뿐 아니라 적어도 호모 사피엔스 네안데르탈리스에서부터
인정되는 사람의 자아의식 상태가 있었다. 인간은 태어날 때부터 인간의 뇌를 갖는다 . 그러나 이것의 세계 2 의 경험은 대단히 미미하고 세계 3 은 미지상태이다 . 태어날 때의 뇌는 인 간 태생기부터도 인간으로 간주하여야 한다. 그러나 인간 인격은 아니 다 . 세계 3, 즉 문화 세계와의 계속적인 상호작용에 의한 자아의식의 출현과 발달(세계 2) 은 전혀 신비의 과정이다 . 결론적으로 첫째, 사람은 일관된 자아의식적 존재로서 존재한다는 확실성이다 . 둘째는 신체와 뇌를 포함하는 물질적 세계가 존재한다는 확실성이다. 특유의 자아의식적 존재로서 우리가 존재하게 되었다는 사건이다 이는 과학을 훨씬 초월한 기적이다 . 두 가지 대안이 남는다. 죽 뇌와 정신이다. 유물론자는 전자에 찬성 할 것이댜 그러나 이원론적 상호작용론자는 세계 2 의 자아를 경험적 일관성을 가진 존재로서 간주할 것이다. 우리들의 직접적 경험은 물론 주관적인 것이고 전적으로 우리의 뇌와 자아에서 유도되는 것이다 . 타 아의 존재는 주체들간의 소통으로 형성된다 . 경험적 단일성의 기초를 이룬다는 단일 게놈g enome 은 적게 잡아도 5 만 개로 추산되는 사람의 유전자 중에서 무한대로 확률이 낮은 유전 자 제비뽑기(실패율 1015,000) 의 결과로 이루어진 것이다. 유물론은 우리 들 경험적 단일성을 해결하지 못하였으므로 자아 또는 영혼의 일관성 을 초자연적 영적 창조에 의한다고 에클스는 생각한다 . 신학적으로 설 명한다면 영혼은 신의 새로운 창조물이고 수정과 출산 사이의 어느 시 기에 성장하는 태아에게 이식된 것이라고 생각하는 것이다 . 이는 <신 성 창조>룰 필요로 하는 단일한 개성의 내적 핵심의 확실성이고 다른 어떤 설명도 있을 수 없다고 생각한다. 환상적이고 불가능한 제비뽑기 에 의한 유전적 단일성도 아니고 개체의 단일성을 결정하지는 못하고 이룰 변조시킬 수는 있는 환경적 개체화도 아니라는 것이다. 영혼이란 영성을 견지해야 하므로 존재에 대한 의문은 과학의 범위
밖에 있는 것이고 따라서 다윈 학설과의 관계에서 문제가 생기지 않는 다고 생각하는 것이다. 에클스 박사는 < 생물학적 진화의 유물론적 사 건 위에 영위된 신의 섭리가 있었음을 나는 믿는다 > 고 하였다. 그리고 약 3.6by BP 의 생명의 기원이 나타나고 돌연변이와 자연도태에 의한 유전자 부호의 생화학적 발달에 우리는 영적 세계에 존재하는 영혼을 가진 영적 존재인 동시에 물질세계에 존재하는 신체와 뇌를 가진 물질 적 존재임을 인식해야 한다고 주장하고 있다. 끝으로 자신의 주장은 지구 행성을 소중히 여기는 데에 있다고 하였다. 영겁을 통하여 계속 되는 세대의 인간 인성으로서의 존재인 우리의 주거는 오직 지구뿐이 기 때문이라는 것이다.
참고문헌에 사용된 약어표
Acta Anat Adv Neurol Arner J Obst Gy ne col Amer J Phy s Anth r op Amer Nat Amer Phy si o l Soc Handbook Amer Psyc h ol Amer Scie n ti st Anat Anz Ann NY Acad Sc i Annu Rev Neurosci Annu Rev Phy si o l Arch Neural Arch Psyc h i at r Nervenkr Bio l Repr o d Bio s ci Rep Brai n Behav Evol Brain Res Bull Acad Roy Med Belg VII
Acta Anato m ica Advances in Neurolog y Ameri ca n Jou mal of Obste t r ics & Gy n ecolog y Ameri ca n Jou rnal of Phy s ic a l Anth r o- po log y Ameri ca n Natu ralist Ameri ca n Phy si o l og y Socie t y Hand-book Ameri ca n Psyc h olog ist Ameri ca n Scie n ti st Anato m isc her Anzeig e r Annals of the New York Academy of Scie n ce Annual Revie w of Neurosci en ce Annual Revie w of Phy si o l og y Arch ive s of Neurolog y Arch ive fur Psyc h i at r ie und Nerven- krankheite n Bio lo g y of Repr o ducti on Bio s cie n ce Repo r t s Brai n Behavio r and Evoluti on Brai n Research Bulletin of the Academy of Roy al Me dicin e , Belgi um VII
Bull Joh ns Hop kins Hosp Cell Tis s ue Res Cib a Found Sy m p Cli n Neurosurg Cour Forsch Inst Senckenberg Dev Psyc h obio l Electr o enceph alog r Cli n Neuro- phy si ol Exp Brai n Res Foli a Pri ma to lo g Hum Neurobio l Int Rev Neurobio l J Anat J Anat Entw ic k l-Gesch J Comp and Phy si o l Psyc h J Comp Neural J Hirnfors ch J Hum Evol J Neurop hy si o l J Neurosci J Phy s io l J Psyc h Neural J Repr o d Fert MTP Intl Rev Sc i Phy si o l Neurosci Biob ehav
Bulleti n of The Joh ns Hop kins Hos- pital Cell Ti ss ue Research Cib a Foundati on Sy m p os iu m Cli nica l Neurosurge ry Couri er Forsch ins ti tut Senckenberg Develop m enta l Psyc h obio l og y Electr oe nceph alog rap h y & Cli nica l Neurop hy si o l og y Exp e rim e nta l Brai n Research Foti~ Pri ma to l og ica Human Neurobio lo g y Inte r natio n al Rev iew of Neurobio l og y Jou rnal of Anato m y Jou rnal Anato m isc her Entr wick lung - Gesch ich te Jou rnal of Comp ar ati ve & Phy si o l og - ica l Psyc h i at r y Jou rnal of Comp ar ati ve Neurolog y Jou rnal fur Hirnfor schun g Jou rnal of Human Evoluti on Jou rnal of Neurop hy si o l og y Jou rnal of Neurosci en ce Jou rnal of Phy si o l og y Jou rnal Psyc h olog ie und Neurolog ie Jou rnal of Repr o ducti ve Fe rtility MTP Inte r nati on al Rev iew of Sc ien ce Phy si o l og y Seri es One, Volume 8 Repr o ducti ve Phy si o l og y Neurosci en ce and Bio b ehav ior al Re- vie w s
Neurosci Lett Neurosci Res Phil Trans R Soc Lond Phy si o l Rev Proc Natl Acad Sc i(US A) Proc R Soc Lond Proc 3rd Intl Cong r Prim ate s Prog Cli n Neurop hy si o l Rev Phy si o l Bioc hem Pharmacol Schweiz Mschr Zahruned Soc Neurosci Abstr TINS
Neuroscie n ce Lett er s Neuroscie n ce Research Ph ilo sop hica l Transacti on s of the Roy al Soci et y of London Phy si o lo g ica l Revie w s Proceed ing s of the Nati on al Academy of Sc ien ce(U S A) Proceed ing s of the Roy al Socie t y of London Proceed ing s 3rd Inte r nati on al Con- gre ss of Pri ma te s Prog re ss in Cli nica l Neurop h y si o lo g y Rev iew s of Phy s io lo g y, Bio c hem ist r y & Pharmacolog y Schweiz e ri sc he Monats c h rift filr Zahruned izn Socie t y of Neurosci en ce Abstr ac ts Trends in Neuroscie n ce
참고문헌 Akert , K., Pepe r , K. and Sand ri, C . (1975) 'Str uc tu r al orga niza ti on of moto r end pla te and centr a l syn ap se s', in P.G. Waser (ed.) Cholin e rgi c Mechanis m s, New York : Raven Press, pp. 43-57. Allen, G. and Tsukahara, N. (1974) 'Cerebrocerebellar commu nica ti on sys t e m s', Plr ;sio l Rev 54 : 957-1006. Amste r dam, B. (1972) 'Mirror self ima ge reacti on s befo r e the age of two ', Dev Psyc h obio l 5 : 297-305. Andy , O.J , and Ste p h an, H. (1968) 'The sept um in the human bra in', J Comp Neural 133 : 383- 40 9. Armstr o ng , D.M . (1968) A Mate r ia l ist Theory of the M ind , London : —R o(1u9t8l e1 d) g Te h&e NKaegtua rn e Poaf u Ml. ind , Ith a ca, NY : Cornell Un ive rsit y Press. Ashto n , E.H. (1981) 'Pri ma te locomoti on : some pro blems in analys i s and int e r p re ta t i on ', Ph il T rans R Soc Lond [Bio l ] 292 : 77-ITT. Ba ile y, P. and von Bon in, G. (19 51) The Isocort ex of Ma n, Urbana, 파. : Un ive rsit y of Illino is Press. Ba ile y, P., von Bon in, G. and McCulloch, W. S.( 1950) The Isocort ex of the Chim p a nzee, Urbana : The Un ive rsi ty of Illino is Press. . Baker, J ,F. , Pete r sen, S.E., Newsome , W.T. and Albnan, J.M . (19 81) 'Vi su al respo nse pro p erties of neurons in fou r extr as tr iate visu al areas of the owl monkey (A otu s trivirga tu s ) : a qu anti tat i ve comp ariso n of me dial, dorsome dial, dorsolate ra l and midd le tempora l areas', ] Neurop lr;sio l 45 : 397 一 416. Bard, P. (1968) 'Postu ral coordi na ti on and locomoti on and their centr al contr ol', in V.B . Mountc a stl e( ed.) Medic a l Plr ;sio l og y, Sa int Lou is,
Mo, Mosby CV, pp. 1750-70. Barlow, H.B., Blakemore, C. and Pett igre w, JD . (1967) 'The neural mecha nism of bin o cular dept h disc rim ina ti on ', J Ph ysi o l (London) 193 : 327-42. Barrow, JD . and Tip le r, F .J (19 86) The Anth r op ic Cosmolog im l Princ i ple , Oxfo rd : Clarendon Press. Basser, L.S. (19 62) 'Hem iple g ia of early onset and the fac ulty of spe e ch with spe c i al refe r ence to the eff ec ts of hem isp h erecto m y ', Brain 85 : 427-60. Baumg artne r, G. 0986) 'Psyc h op h y si c s and centr a l pro cessin g ', in A. Asbury , G.M. McKhann and W. I. McDonald (eds) Di se ases of the Nervous Sy st e m : Clin i m l Neurobio lo g y, Vol. 2 , ·Lo ndon : Hein e mann Me dica l Books, pp. 804-15. Beloff , J .(19 65) 'The ide nti ty hyp o th e sis : a cri tiqu e', i n J.R. Smy thi es (ed.) Brain and Mi nd , New York : Human ities Press, pp. 35-61. Bever T.G . and Ch iare llo, R.J. (1974) 'Cerebral dom ina nce in music ia n s and non- m usic ian s', Scie n ce 185 : 537- 9. Bis h op, P.0. 0 970) 'Cort ica l beg inning of vis u al form and bin o cular dep th per cept ion ', in F.O. Sch mitt (ed.) Neuroscie n ces Research Stu d y Prog ram , New York : Rockefe l ler Un ive rsity Press, pp. 471-85. Bli ss, T.V.P ., and Lom< P, T. (1973) 'Long -l astin g po te n ti at i on of syn apt ic tran sm iss io n in the denta t e area of the anaesth e ti ze d rabbit foll owin g stim u lati on of the per f or ant pa th ', J Ph ys io l (London) 232 : 331-56. Bordes, F. (19 68) The Old Sto n e Ag e , London : Weid e nfe ld & Nic o lson. Brauer, G. (1984) 'A cia niolo g ica l app ro ach to the orig in of anato m ica lly modem Homo sapi en s in Africa and implica ti on s for the app e arance of modem Europe a ns', in F.H . Sm ith and F. Sp e ncer (eds) The Ori gi.ns of Modem Humans, New York : Alan R. Lis s , pp. 3'27 - 410. Bri nkma n, C. and Port er, R. (1979) 'Supp le menta ry moto r area in the monkey : acti vi t y of neurons dU iing per f or mance of leruned moto r task ', J Neuroph ysi o l 42 : 681-709.
(1983) 'Supp le menta ry moto r and pre moto r areas of the cerebral cort ex in the monkey : acti vi t y of neurons du ring perfor mance of a learned movement tas k', in J.E . Desmedt( e d.) Moto r Contr o l Mechanis m s in Man, New York : Raven Press, pp. 393-420. Brodal, A (19 73) 'Self- o bservati on s and neuro anato m ica l consid e rati on s after a str ok e', Brain 96 : 675-94. - (1981) Neurologi a:zl Anato m y , 3rd edn, New York/ Ox fo rd : Oxfo rd Un ive rsit y Press. Brodmann, K. (19 09) Vergl e ic h ende Lokalisa ti on slehre der Grosshi m r ind e in der Prinz ip ien darge ste l lt a u f Grund des Ze/l en baues, Leip z ig : J. —A (1B9a 1 r2th) . 'Neue Erge b n iss e tibe r die vergl e ic h ende histo l og isc he Loka lisa ti on der Grosshi rnrind e', Anat Anz 41 : 157-216. Bronowski , J. and Bellug i, U. (lITTO) 'Lang ua ge , name and concept ', Sdenct 168 : 669 구 3. (19 80) 'Lang u ag e , name and concept ', in T.A. Sebeok and D,J . Um 恥 r-Sebeok(eds) Sp e akin g of Ap e s, New York : Plenum Press, pp. 10 3-14. Brooks, V.B. (1986) The Neural Basis of Moto r Con trol, New York Oxfor d : Oxfo rd Un ive rsity Press. Brooks, V.B. and Thach, W.T. (1981 ) 'Cerebellar contr ol of pos tu re and movement', in Handbook of Ph ysi o l ogy , The Nervous Sy s te m , vol.2, Beth e sda, MD : Ameri ca n Phys i o l og ica l Soci et y , Chapt er 18, pp. 얽 7- 946. Brown, R. (1980) 'The first sente n ces of chi ld and chi mpa nzee', in T.A Sebeok and D.J, Um iker -Sebeok (eds) Sp eaking of Ap e s, New York : Plenum Press, pp. 8.5- 101. Bucy, P.C. (1935) 'A comp a rati ve cyt oa rchi tectonic stu d y of the moto r and pre moto r areas of the prima te cort ex ', J Comp Neural 62 : 293-311. Bil hler, K. (1934) Sp r acht he ori e : die Darste l / un g sfunktion der Sp ra che, Jen a : Gusta v Fis c her.
Bush, G.L ., Case, S.M. , Wi ls on, AC. and Patt on , J.L. (19 77) 'Ra p id spe c ia t i on and chromosomal evoluti on in mammals', P roc Natl Acad Sc i (USA) 72 : 5061-5. Cann, R.L., Sto n eki ng , M. and Wi ls on AC. (19 87) 'Mi toc hond rial DNA and human evoluti on ', N atu r e 325 : 31-6. Carson, H.L. (1975) 'Th e ge neti cs of spe c ia t i on at the diplo id level', Amer Nat 109 : 83-92. Celesia , G.G . (1976) 'Orga n iza ti on of aud itor y cort ica l areas in man', Brain 99 : 403-14. Celesia , G.G., Brou gh to n , R.J., Rasmussen, T. and Branch, C. ·(19 68) 'Aud itor y evoked po te n ti als from the exp o sed human cort ex ', Electr oe nceph alog r Clin Neuroph ysi o l 24 : 458-66. Chan, C.W.Y. (1983) 'Segm enta l versus sup ra seg m enta l contr ibu ti on s to long - late n cy str et c h respo nses in man', Adv Neural 39 : 467-87. Chan, C.W.Y. , Melvil l Jon es, G., K earney, RE. and Watt , D.G.D . (1979a) 'Th e late electr om y o g ra p hic respo nse to limb disp l a cement in man. I. Ev ide nce for sup ra spi na l contr ibu ti on ', Electr oe nceph alog r Cli n Neuroph ys io l 46 : 173-81. Chan, C.W .Y., Melvi ll Jon es, G. and Catc h love, R.F. H. (1979b) 'Th e late electr om y og r a p hic respo nse to limb disp l a cement in man. II. Sensory orig in', Electr oe nceph alog r Clin Neuroph ys io l 46 : 182-8. Cha nge ux, J.-P . (1985) Neuronal Man. The Bi ol og y of M ind , New York : Panth eo n Books. Ch iar elli , B. (1985) 'Chromosomes and the ori gin of man', in P.V. Tobia s (ed.) Homi ni d Evoluti on . Past, Present and Futu re , New York : Alan R. Lis s , pp. 397-400. C—homsk y( 1, 96N7. ) (1'9T6h2e) Sfoy rnm ta aclt icn aSttur urec t uo rfe sl aSn 'Ggur aagve e' n. haAgp e p, e Mn do iuxt o An &o fC oE. .H. —Le(n1 n9 6e8b)e rgL an(egd u.)a, gBe i oalno dg ictha le FMou inndd , atNi oen ws oYf o Lrka n: gH uaargceo, u Nrte wB raYcoe rk& : Wi ley, pp. 3 얽 -412.
World. (1978) 'On the bio l og ica l basis of lang u ag e capa cit ies ', in G.A . Mille r and E. Lenneberg (eds) Psyc h olog y and Bio l og y of Lang u ag e and —T h(o1u9g80 h) t , 'NHeuwm aYno rkla n: gA ucaagdee ma inc d P roetsh se , r pps.e 1m0i9 o-t2 i 2c0 .s ys te m s', in T.A. Sebeok and D.J. Um ike r-Sebeok (eds) Sp e akin g of Ap e s, New York : Plenum Press, pp. 429-40. Clemente , D.C. and Chase, M.H. OW3) 'Neurolog ica l substr at e s of agg r essiv e behavio u r', Annu Rev Ph ysi o l 35 : 329-56. Cop pe ns, Y. (1983) 'Les plu s anc ien s fos sile s d'hom inide s', in C. Chag a s (ed.) Recent Advances in the Evoluti on of Prim ate s , Vati ca n Cit y : Ponti fici a e Academ iae Scie n ti ar um Scri pta Va ria 50, 1-9. Cowey, A. (1981) 'Why are the re so many vis u al areas?', in F.O. Sch mitt, F.G . Worden, G. Adelman and S.G. Denn is (eds) The Orga niz a ti on of the Cerebral Corte x , Cambri dg e , Mass. : MIT Press, pp. 395-413. (19 82) 'Sensory and non- se nsory vis u al diso rders in man and monkey' , Phil Trans R Soc Lond [Bio l ] 298 : 3-13. Cowey, A.L. and Weis k rantz , L. (1975) 'Demonstr at i on of cross-modal matc hing in Rhesus monkeys , Macaca mulatt a', N europs ych olog ia 13 : 117-20. Creutz feld t, O.D. (1979) 'Neurop h y s io l og ica l mecha nism s and conscio u s-—n e(s1s9' ,8 7C) ib 'Ia n eFvoi tuanbd le Sdy ema pd . lo6c9 k :s 2 1o7f- 3 t3h. e brai n-mind disc ussio n ', in B. Guy la s (ed.) The Brain - Mi nd Problem, Assen /Ma astr ich t : Leuven Un ive rsit y Press, van Gorcum, pp. 3 갑 7. Cu rtiss , S. (1977) Genie : A Ps ych oling uist i c S tu d y of a Modem-da y 'Wi ld Chil a, New York : Academ ic Press. Damasio , A.R. and Geschwi nd , N. (1984) 'The neural basis of lan guage ', Annu Rev Neurosci 7 : l'Z l-4 7. Da rt, RA . 0925) 'Austr alop i the cus Africa nus, the ~an-ape of South Africa ', Natu r e 115 : 195.
Davenp or t, R.K . (1 罪 6) 'Cross- m odal perc ep tion in ape s', in Confe r ence 'On Or igins and Evoluti on of Lang u g a e and Sp e ech', Ann NY Aazd Sci 280 : 143-9. Dawk ins , R. (1976) The Self ish Gene, Oxfo rd : Oxfo rd Univ e rsit y Press. Day, M. H. (1 98. 5) 'Hom inid locomoti on -fr om Taung to the Laeto l i foo tp rint s ', in P.V. Tobia s (ed.) Homi ni d Evoluti on . Past, Present and Futu re , New York : Alan R. Lis s , pp. 115-27. Day, M.H. and Nap ier , JR. (1964) 'Fossil foo t bones', Natu r e 201 : 969-70. Deecke, L. and Komhuber, H.H. (l<;r78 ) 'An electr ica l sig n of pa rtic ipa ti on of the mesia l sup ple menta ry moto r corte x in human volunta ry finge r movement', Brain Res 159 : 473-6. Deevey, E.S. (1960) 'The human popu lati on ', Sc ien ti fic Americ a n 203(3) : 194-206. Delg a do, JM . R. (19 69) Ph ys ic a l Contr ol of the M ind , New York : Ha rper & Row. DeLong , M. R. and Georgo po ulos, AP. (1983) 'M oto r func ti on s of the basal ga ng lia', Amer Ph ys io l Soc Handbook 21 : 1017-61. Dennett, D.C. (1969) Conte n t and Conscio u sness, London : Routl ed g e & —K e(g1 a9 n81 )P aBulr.a in s to r ms, Cambri dg e , Mass. : MIT Press. Denny - Brown, D. 0966) The Cerebral Contr ol of Movement, Liv e rp oo l : Liv e rp oo l Un ive rsit y Press. Desmedt, J.E . and Robert so n, D. {l<;r77 ) 'Di ffere nti al enhancement of early and late comp o nents of the cerebral somato s enory evoked po te n ti al s dw ing forc ed-pa ced cog nitive tas ks in m 패, J Ph ysi o l (London) 刃 1 : 761-82. Dobzhansky , T. (1960) 'The pre sent evoluti on of man', Sc ien ti fic Americ a n —20 (31(936)7 :) 20T6h-1e 7.B i ol og y of Ultim a te Concern, New York : The New Ameri ca n Libr ar y. Drake, R.E ., Cu rtis, G.H., Cerlin g, T.E. , Cerlin g, B.W. and Hamp el, J. (1980)
'Tu ff dati ng and ge ochronolog y of tuf f ac eous sedim ents in the Koobi- f o r a and Shung u ra for mati on s, East Af rica ', Natu r e 283 : 368-72. Eccles, JC . 0970) Facin g Reality : Phil o sop h ic a l Adventu r es by a Brain ——S c((1 1ie99 n78 9t0i) )s tT, T hhNeee wHH uumYmoaarnkn :P Ssyp cr i hneg , eB r-eVrleinr l/aHge. id e lberg/ Ne w York : Sp ring e r. My s te r y , Berlin /He id e lberg/ Ne w York : Sp ring e r. 一 (1981a) 'The modular ope rati on of the cerebral neocort ex consid e red as the mate r i al basis of menta l events ', Neurosci en ce 6 : 1839-56. 一 (19 81b) 'Lang u ag e, tho ug h t and brai n', Ep ist e m olog ia [ Sp ec ia l Issue] —4 : (9179-8122a6). 'An ima l conscio u sness and human self -c onsci ou sness', Exp e rien ti a 38 : 1384-91. (1982b) 'The initiat i on of volunta ry movements by the sup - ple menta ry moto r area', Arch Ps ych ia t r Nervenkr 231 : 423-41. (1984) 'Phy si o l og ica l and ph armacolog ica l inv estig a ti on s on pain contr o l', Schwei2 Mschr Zahnmed 94 : 1004-13. 一 (1986) 'Do menta l events cause neural events analog o usly to the pro babil ity fiel ds of qu antu m mecha nics ?' Proc R Soc Lond [Biol ] —22 079 :8 471)1 -'M28a.m ma lian sys te m s for sto r i ng and retr iev in g inform ati on ', in C.D. Woody (ed.) Cellular Mechanis m s of Condit ion in g and —B (e1h9a8v9i)o r' Talh eP lmasintid c i-t by ,r aNi ne wpr o Ybloermk :r ePvleisn iu t emd :P trhees sm, picpr. o 2si8t9 e- 3 h0y2 p. othe sis ', in J.C . Eccles and O.D. Creutz feld t (eds) The Pr inc ip le s of Desig n and Op e rati on of the Brain , Vati ca n Cit y : Ponti ficiae Acade miae Scie n ti ar um Scri pta Va ria. Eldredg e, N. and Gould, S.J. (1972) 'Punctu a te d equilibr i a : an alte r nati ve to ph y le ti c grad ua lism ', in T.J.M . Schop f(e d.) Models of Paleobio l og y, San Franci sc o : Freeman, Coope r, pp. 82-115. Ett linge r, G. and Blakemore, C.B . 0969) 'Cross-modal trans fe r set in the
monkey' , N europ s yc h olog ia 7 : 41-7. Eva rts, E.V.(1 9 72) 'Contr as ts betw een acti vi t y of pre centr a l and po stc e ntr al neurons of cerebral cort ex duri ng movement in the monkey' , Brain —R( e1s9 8410 ) :' R25o-le 4 1 o. f moto r cort ex in volunta ry movements in prima te s ', i n Handbook of Ph ysi o l og y. The Nervous Sy s te m , vol. 2. Beth e sda, MD : Ameri ca n Phy si o lo g ica l Socie t y , pp.1 083-120. Facchi ni, F. (19 84) C,amm i no dell' Evoluzio n e Umana, Milan : Jac a Book. Feig l, H . (19 67) The 'Menta l ' and the 'Phy s im l ', M inn eapo li s : Un ive rsit y of Minn esota Press. Feldman, M.L. (19 84) 'M orp h olog y of the neocort ica l pyra m ida l neuron', i n A. Pete r s and E.G. Jon es (eds) Cerebral Corte x , Vol. !, New York : Plenum Press, pp. 123- 20 0. Flechsig , P. (1920) Anato m i e des Menschl .ich en Gehir n s und Ri ick enmarks au f My e log e neti sc hen Grundlag e , Leip z ig : Th iem e. Fleis c hhauer, K. (1974) 'On differ ent pa tt er ns of dend ritic bund ling in the cerebral cort ex of the cat', J Anat Entw ick l-G esch 143 : 115- 26 . Fouts , R.S . and Rigb y, R.L. (19 80) 'Man- ch i mpa nzee commun ica ti on ', in T.A . Sebeok and D.J. U m iker -Sebeok (eds) Sp e akin g of Ap e s, New York : Plenum Press, pp. 261-85. Frahm, H.D., Ste ph an, H. and Ste p h an, M. (19 82) 'Comp ariso n of brai n stru c tu re volumes in ins ecti vo ra and prima te s . I. Neocort ex ', J Hi mfors ch 23 : 375-89. Freund, H.J. and Hummelsheirn , H. (19 85) 'Lesio n s of pre moto r cort ex in man', Brain 108 : 6r;r f-7 33. Fri ed man, D.P. and Jon es, E.G . (1981) 'Th alam ic inp u t to areas 3a and 2 in monkey s' , J Neuroph ysi o l 45 : 59-85. Fulto n , J.F . (19 49) The Ph ys io lo g y of the Nervous Sy s te m , New York : Ox for d Un ive rsit y Press. Fuste r, J.M . (19 80) The Prefr on ta l Corte x , New York : Raven Press. Ga ffan , D. and Weis k rantz , L. (1980) 'Recency eff ec ts and lesio n eff ec ts in
delaye d non-m atc h i ng to randomly bait ed samp le s by monkey s ', Brain Res 196 : 373 -86. Gajd u sek, D.C. (1973) 'Kuru in the New Guin e a Hig h lands', in JD . Sp illan e (ed.) Trop ica l Neurology , New York : Oxfor d Un ive rsit y Press, pp. 376-83. Gallup, G.G. (1977) 'Self- r ecog nition in prim ate s ', Amer Psyc h o/ 32 : 329- 38. Gardner, R.A. and Gardner, B.T . (1980) 'Comp a rati ve ps yc h olog y and lang u age acq u is i t ion ', i n T.A. Sebeok and D.J Um iker -Sebeok (eds) Sp e akin g of Ap e s, New York : Plenum Press, pp. 287-330. Gardner, B.T . and Gardner, R.A. (1985) 'Sig n s of int e l lig e nce in cross fos te r ed chi mpa nzees', P hil Trans R Soc Land [Bi ol ] 308 : 159-76. Geschwi nd , N. (19 65) 'Di sc onnecti on syn dromes in anima l and man. Pa rt 1', Brain 88 : 237-94. 一 (1972) 'Lang u age and the brai n' , Scie n ti fic Ameri(X Jn 226(4) : 76-83. Geschwin d , N. and Levit s k y, W. (19 68) 'Human brai n : left -righ t asym - metr ies in tem p o ral spe ech reg ion ', Scie n ce 161 : 186- 7. Gil b ert , P.F . C. and Thach, W. T. (19 77) 'Purki nje cell acti vi t y du ring moto r lea rning ', Brain Res 128 : 309- 28 . Gin g e ri ch , P.D . (19 85) 'Nonli ne ar molecular clocks and ape - human dive rge nce time s', in P.V . Tobia s (ed.) Homi ni d Evoluti on . Past, Present and Futu r e, New York : Alan R. Liss , pp. 411-16. Goldman, P.S . and Nauta , W.J H . (1977) 'Columnar dist r ibu ti on of cort ico -cort ica l fibr es in the fron ta l associa t i on , limb ic and moto r cort ex of the develop ing rhesus monkey' , Brain Res 122 : 393-413. Goldschm idt, R. (1940) The Mate r i al Basis of Evoluti on , New Haven, Conn : Yale Un ive rsity Press. —Gombri c(1h 9, 8E6.) H .T h(1e9 6C0h) imA pr at naznede sI llouf s iGo no ,m Lboe.n dPona t:t e Prh nas idoof n BPer!er rsosi:o r, Cam-Goodall, J. (19 71) In the Shadow of M an, Bosto n , Mass. : Hou gh to n Mifflin. bri dge, Mass. : Belknap Press, Harvard Un ive rsit y Press.
Gott , P.S . 0973) 'Cog n it ive abil it i es foll owi ng righ t and left hem isp h er- ecto m y ', Corte x 9 : 266-74. Gould, S.J . (1977) Onto g e ny and Phyl o g e ny, Cambri dg e, Mass. : Belknap Press, Harvard Un ive rsit y Press. 0982) 'Is a new and ge neral the ory of evoluti on emergi ng ? ' in J.M . Sm ith(e d.) Evoluti on Now, A Centu ry Aft er Dannin , San Francis c o : W.H. Freeman, pp. 129-45. Gran it, R. 0970) The Basis of Moto r Contr ol , New York : Academ ic Press. Gray, E.G. (19 82) 'Rehabil itat i ng the dend ritic spi ne ', TINS 5 : 5-6. Gr iffin, D.R . 0976) The Qu esti on of Anim al Awareness, New York : —R o(1ck9 8ef4e) l leAr nUim n aivl e rTshit i yn kPinrge s, s.C ambri dg e, Mass. : Harvard Un ive rsit y Press. Gr illne r, S. (1981) 'Contr ol of locomoti on in bip e ds, tet r ap o ds and fish ', in V.B. Brooks (ed.) Handbook of Ph ys io l ogy , Secti on 1, The Nervous Sy s te m . Moto r Contr ol, Pa rt 2, Beth e sda, MD : Ameri ca n Phy s i- olog ica l Socie t y . Gross, C.G. 0973) 'Vi su al func ti on s of inf e r ote m p o ral cort ex ', in R. Jun g (ed.) Handbook of Sensory Ph ysi o l ogy , Vol. VII./3B, Berlin / Heid e lberg/ Ne w York : Sp ringe r-Verlag, pp. 451-82. Gross, C.G ., Desim o ne, R., Albri gh t, T.D . and Schwa rtz, E.L . (1985) 'Infer i or tem p ora l cort ex and pa tt er n recog nition ', in C. Chag a s, R. Gatta s s and C. Gross (eds) Pate m Recog nition Mechanis m s, Vati ca n Cit y : Ponti ficia e Academ iae Sc ien ti ar um Scrip ta Va ria 54, pp. 179- 201. Gruber, H.E. (1974) Dannin on Man, London : Wi ldw ood House. Gusta fss on, B. and Wi gs tr om , H. (1986) 'Hippo camp al long -l astin g poten ti at i on pro duced by pairing sin gle volleys and bri ef cond ition ing tetani evoked in sepa rate affer ents ', J Neurosci 6 : 1575-82. Ha llida y, M. A.K (1975) Leami ng How to Mean, New York : Elsevie r . Ha milton , C.R. (19 Tia) 'Investig a ti on s of perc ept ua l and mnemon ic
late r aliz a t i on in monkey s ', in S. Hamad, R.W . Doty , L. Goldste i n , J. Jay n es and G. Krauth a mer (ed s ) Late r aliz a ti on in the Nervous Sy s te m , Ne w York : Academ ic Press , pp. 45- 62 . (19 77b) 'An assessment of hem isp h eric spe c ia l iz a ti on in monkey s' , Ann NY Acad Sci 299 : 222- 32 . Hamlyn , L.H . (1962) 'An electr o n micr oscop e stu d y of pyra m ida l neurons in the Ammon's horn of the rabbit ', J Anat 97 : 189- 20 1 . Handler, P. (ed.) (19 68) Bi ol og y and the Futu r e of Man, New York : Oxfo rd Univ e rsi t y Press, pp. 134-6. Hawkes, J. (19 65) Prehis t o r y in Hi st o r y of Mankin d . Cultu r al and Scie n ti fic Develop m ent, Vol. 1 , Pa rt 1, London : UNESCO, New —E n(1g9 l7is6h) LTihb er aAr yt. l a s of Early Man, London : Macm illan London. Haye k, F.A . (19 78) The Three Sources of Human Values, London : London School of Econom ics and Poli tica l Scie n ce. Heath , R.G . (19 54) Stu d ie s in Schiz o p h renia . A Multid is c i plina ry Ap pro ach to M ind -Brain Relati on ship , Cambri dg e, Mass. : Harvard Un ive rsit y Press. 一 (19 63) 'Electr ica l self- s tim ulati on of the brai n in man', Ameri ca n jo urnal of Psyc h ia t r y 120.: 5 71-7. H—ebb, D (1.O98. 0 ()1 9E4s9s)a yT hoen OMrgi na d n , izH a itlil osnd aloef , BNeJh a: vLioa rw , rNenecwe EYrolbrka u: mW. i ley . Hecaen, H. (19 67) 'Br ai n mecha nism s sug ge ste d by stu dies of parieta l lobes', in C.H . Millikan and F.L. Darley (eds) Brain Meclrrnis m s Underlyi ng Sp e ech and Lang uage, New York/ Lo ndon : Grune & Str at t on , pp. 146-66. Hess, W.R. (1932) Beit rag e zur P hy s i olo 硏 des Hi m sta mme s. I. Di e Meth o dik der lokalis ie r te n Reiz u ng und Ausscmltu ng subkort ika ler Hi m abschnit te, Leip z ig : Georg Th iem e. Hill, UJm.H i. ker (1-S98e0b)e ok'A (p eedss ) aSnpd e akLian ng g uo fa g A ep ', e si,n NeTw.A Y. orSke :b ePolke nwannd P rDes.sJ ,.
pp. 331-52. Hokfe l t, T., Lju n g d ahl, A., Tereniu s , L., Eid e , R. and Ni ls son, G. 0977) 'Immunoh ist o c hem ica l analys i s of pe p tide pa th w ay s po ssib l y relate d to pain and analge sia : enkeph ali n and substa n ce P', Proc Natl A( XId Sci (US A) 74 : 3081-5. Holloway, R.L. (1968) 'Th e evoluti on of the prim ate brain : some aspe cts of qu anti tative relati on s', Brain Res 7 : 121-72. (19 74) 'The casts of fos sil hom inid brai ns ', Sc ien ti fic Ameriro n —23 (11 (918) 3 :) 10'H6-u1m5.a n pa leonto l o gica l evid e nce relevant to lang u ag e be- hav ior ', Hum Neurobio l 2 : 105-14. Howells, W.W. (1966) 'Homo erectu s ', Scie n ti fic Ameri(X In 215(5) : 46- 53 . Hubbard, J. I. 0973) 'Micr oph y s io lo g y of vert eb rate neµromuscular tra ns- miss io n ', Ph ys io l Rev 53 : 674 구 23 . Hubel, D.H. (1982) 'Evoluti on of ide as on the primary vis u al cort ex , 1955-1978; A bia s sed hist o r i ca l account' (N obel Lectu re ), Bi os ci Rep 2(7) : 435-69. Hubel, D.H. and Wi es el, T.N . (1962) 'Recept ive field s, bin o cular int e r acti on and func ti on al arch itec rure in the cat' s vis u al cort ex ', J Ph ys io l ——(L ((1o1 9n96d65o3)n) )'R 'S1e6ch0ea pp: te1 iv 0e6 a -nf5di4e .l da sr raanndg ef mune cntti ono af l caorlcuhm itnecs ruirne itnh e t wc oa tn' so n ssttrr iiaatt ee cort ex ', j Ph ys io l (London) 165 : 559-68. —v i0su9 a7l0 )a r'eSatse r 0eo8s caonpd ic 1 v9)i sioo f n thine cmaat'c, aJq u Ne emuoronk-pehy y :s ico el ll2s8 s: e2n2s9i-t8 iv9e. to bin o cular dep th in area 18 of the amcaq u e monkey cort ex ', Natu r e ——22 (510 99:7 47417)- )2 'S. 'eF quun ecntci oen arel g ua lar ity and ge ometr y of ori en ta t i on columns in the monkey str iat e cort ex ', J Comp Neurol 158 : 267-94. rch itec rure of the macaqu e monkey vis u al cort ex ', Proc R Soc Lond [Biol] 198 : 1-59.
Iles, JF . (1977) 'Respo nses in human pre ti bi a l muscles to sudden str e ch and to nerve stim ulati on ', Exp Brain Res 30 : 541-70. Im ig, T.J and Morel, A. ( 1983) 'Orga niz a ti on of the tha lamocort ica l aud itor y sys te m in the cat', Annu Rev Neurosci 6 : 90-120. lng v ar, D.H . (1983) 'Seri al aspe cts of lang u ag e and spe ech relate d to pre fr on ta l cort ica l acti vi t y. A selecti ve revie w ', H um Neurobio l 2 : —1 7(71-98 895.) 'Memory of the fut u r e : an essay on the tem p or al orga - niza ti on of conscio u s awareness', Hum Neurobio l 4 : 1 짜 36. Isaac, C. (19 78) 'Food-sha ring behavio r of pro to h uman hom inids ', Sc ien ti fic Americ a n 238(4) : 90-108. Isaacson, R.L. (19 74) The Lim b ic Sy s te m , New York : Plenum Press. Ito , M. and Kano, M. (1982) 'Long - lastin g depr e ssio n of pa rallel fibr e-Purki nj e cell tra nsm iss io n ind uced by conju n cti ve stim u lati on of pa rallel fibr es and clim b in g fibr es in the cerebellar cort ex ', N eurosci Lett 33 : 253-8. Ito, M. (19 84) The Cerebellum and Neural Contr ol, New York : Raven Press. 一 (19 86) 'Long -ter m depr e ssio n as a memory pro cess in the cere-bellum', Neurosci Res 3 : 531-9. Jac k, ].J.B ., Redman, S.J . and Won g, K. (1981a) 'The comp on ents of syn apt ic po te n ti al s evoked in cat spi na l moto n eurones by impu lses in sin g le gro up Ia aff er ents ', J Ph ys io l (London) 321 : 65-96. (1981b) 'Mod ifica ti on s to syn a p tic tran sm iss io n at Group Ia syn - aps es on cat spi na l moto n eurones by 4-am ino p yridine ', J Ph ysi o l (London) 321 : 111-26. Jar vis , M.J. and Ett ling e r, G. (1977) 'Cross-modal recog nition in chi mpa nzees and monkeys ' , Neurops ych olog ia 15 : 499- 50 6. Jel in e k, A.J. (19 82) 'The Tabun cave and Paleolit hic man in the Levant', Scie n ce 216 : 1369-75. Jeli n e k, J. (1985) 'Th e Europe a n, Near East and Nort h Africa n find s after
Austr alo p ithe cus and the princ ip a l consequ ences for the pict u r e of human evoluti on ', in P.V. Tobia s (ed.) Homi ni d Evoluti on . Past, Present and Futu re , New York : Alan R. Lis s , pp. 341-54. Jen nin g s , H.S. (1930) The Bi ol og im l Basis of Human Natu r e, New York : W.W. Nort on . Jer i so n, H.J . (19 73) Evoluti on of the Brain and Inte l lig en ce, New York : —A (c1a9d8e. 5m) ic'A Pnrimes asl. i nt e l li ge nce and enceph a liza ti on ', Phil Trans R Soc Lond [Bi ol] 308 : 21-35. Joh anson, D.C. (19 8. 5) 'Th e most primitive Austr al op ithe cus', in P.V . Tobia s (ed.) Homi ni d Evoluti on . Past, Present and Futu re , New York : Alan E. Lis s , pp. 203- 12 . Joh anson, D.C. and W 血 e, T.D . (1979) 'A sys t e m ati c assessment of early Africa n hom inids ', Scie n ce 202 : 321-30. Jon es, E.G. (19 83) 'The natu re of the aff er ent pa th w ays conveyi ng short -la te n cy inp u ts to prima te moto r cort ex ', Adv Neurol 39 : 263- 8.5. Jon es, E.G. and Powell, T.P. S. (1970) 'An anato m ica l stu d y of convergi ng sensory pa th w ay s within the cerebral cort ex of the monkey' , Brain 93 : 793-820. Jun g, R. (19 74) 'Neurop s yc h olog ie und neurop h y si o l og ie des Kontu r-u nd Formschen in Zeic h ung und Malerei', in H.H. Wi ec k (ed.) Ps yc hopa tho log ie Mi m i sc her Gesta ls tu n g e n, Stu ttga t : F.K . Schat- —ta u( 1e9r,7 8p)p . 'P29er-c8e8p. t ion , consci ou sness and vis u al att en ti on ', in P. Buser and A. Roug eu l-Buser (eds) Cerebral Correlate s of Consci ou s Ex- pe ri en ce, Amste rd am : Nort h Holland, pp. 15-36. K교 el, E.K and Schwa rtz, ].H. (1982) 'M olecular bio l og y of lea rning : modulatio n of tran sm itter release', Scie n ce 218 : 433-43. Kandel, E.R., Klein , M., Hochner, B., Shuste r , M. , Sie g e lbaum, S.A ., Hawk ins , RD., Glanzum, D.L. , Caste ll ucci , V.F. and Abrams, T.W .
0987) 'Sy n ap tic modulati on and learnin g : new ins ig h ts int o syn ap tic tra nsm iss io n from the stu d y of behavio r ', in G.M. Edelman, W. E. Gall and W.M. Cowan (eds) Sy n ap tic Functi on , New York : Wi ley , A Neurosci en ce Instit ute Public a ti on , pp. 471-518. Kellog g, W.N . and Kellog g, L.A . (1933) The Ap e and the Ould, New York : McGraw-H ill. Kelly, R.B ., Deuts c h, JW ., Carlson, S.S. and Wag n er, J.A. (lCJ /9 ) 'Bio - chem ist r y of neurotr an sm itter release', Annu Rev Neurosci 2 : 399- 446. Kety , S.S. (1970) 'The bio g e n ic amine s in the centr al nervous sys te m : the ir po ssib l e roles in arousal, emoti on and lea rning ' , in F.O. Schm itt (ed.) The Neuroscie n ces, New York : Rockefe l ler Un ive rsity Press, —pp .0 392742-)3 6.'N orepi ne ph rine in the centr al nervous sys te m and its correlati on s wi th vehavio r ', in A.G . Karczmar and JC . Eccles (eds) Brain and Behavio r , New York : Sp ringe r-Verlag, pp. 115-28. Kimu ra, D. (1967) 'Functio n al asym me tr y of the brai n in dich oti c liste n i ng', —C o(1r9te7 x3 ) 3 ':T 1h6e7 -7a8sy. m me tr y of the human brai n', Sc ien ti fi c Ame ri따 228(3) : 70- 8. Klopp e r, A. (lCJ/ 4 ) 'Th e hormones of the pla centa and their role in the onset of labour', in R.O . Greep (ed.} MTP Intl Rev Sc i Ph ysi o l (Serie s one), Vol. 8, Repr oducti ve Ph ysi o l og y, London : Butt er - wort hs, pp. 180-201. Korn, H. and Faber, D.S. (19 'ifl) 'Regu lat i on and sig nifica nce of pro babil isti c release mecha nism s at centr al syn a ps es', in G.M. Edelman, W.E. Gall and W.M. Cowan (eds) Sy n apt ic Functi on , New York : Wi ley , A Neurosci en ces Instit ute Publi ca ti on . pp. 57-108. Komhuber, H.H. (lCJ /3 ) 'Neural contr ol of inpu t into lon g-term memory . Lim b ic sys te m and amnestic syn d rome in man', in H.P. Zip pel (ed.) Memory and Tran sfer of Inf orm ati on , New York : Pleum Press,
pp. 1-22. Kummer, H. and Goodall, J. (1985) 'Cond ition of im movati ve behavio r in prima te s ', P hil Trans R Soc Land [Bi ol ] 308 : 203-14. Lack, D. 0961) Evoluti on ary Theory and Chris t i an Belief The Unresolved Con flict, London : Meth u en. Land gre n, S., Sil fv e n ius , H. and Olsson, KA. (1984) 'The sensori m oto r int e g r a ti on in area 3a of the cat', Exp Brain Res [Sup pl] 9 : 359- 75 . Lang e r, S.K . (1951) Phil o sop h y in a New Key, Cambri dg e , Mass. : Harvard Un ive rsit y Press. Leakey, M.D. 0981) 'Tra cks and too ls', Phil Trans R Soc Land [Bi ol ] 292 : 95-102. Leakey, M. D. and Hay, R.L. (19 79) 'Plio c ene foo tp rint s in the Laeto l il b eds of Laeto l i, Nort he rn Tanzan ia' , N atu r e 278 : 317-23. Leakey, R. E. and Lewi n, R. (1977) Orig ins , New York : E.P. Dutt on . Lee, KS. (1983) 'Susta ine d mod ifica ti on of neuronal act ivi t y in the hippoc amp u s and cerebral cort ex ', in W. Seif er t( e d.) Molecular, Cellular and Behavio r al Neurobio l og y of the Hi ppo camp u s, New York : Academ ic Press, pp. 265-72. Lee, R. G., Mu rph y, J.T . and Tatt on , W .G. (1983) 'Lo ng - late n cy my o ta t i c refl ex es in man : mecha nism s, fun cti on al sig nifica nce and chang e s in pa tie n ts wi th Parki ns on's dise ase or hem iple g ia', Adv Neurol 39 : 489-508. Le Gros Clark, F. (1964) The Fossil Evid e nce for Human Evoluti on , Ch ica g o : Un ive rsit y of Ch ica g o Press. LeMay, M. and Geschwi nd , N. (1975) 'He m isp h ere differ ences in the brai n of grea t ape s ', Brain Behav Evol 11 : 48-52. Lenneberg, E.H. (19 67) Bi ol og ica l Foundati on s of Lang u ag e , New York : ——W ((11i l9e9 76y 5.9 )) ''OAn n eexurpol ap i sn yinc gh o lloang gic ua al gc eo ',m S p ca ireins oc ne 1b6e4tw : e6e3n5 -4m3a. n, chi mpa nzee and monkey' , Neurops yc holog ia 13 : 125.
( 1980) 'A word betw een us', in T.A . Sebeok and D.J. Um ike r-Sebeok (eds) Sp e akin g of A p e s, New York : Plenum Press, pp. 71-81. Levy, J. (19 74) 'Psyc h olog ica l im p lica ti on s of bil a te r al asym metr y', in S.J. Dim o nd and J,G . Beaumont (eds) Hemi sp h ere Functi on in the —H u(m19a7n7 ) Br'Taihn e, Nmewam mYoarlika n: Wbi rlae yi n, papn. d1 21th-8e 3 . adap tive advanta ge of cerebral asym metr y', Ann NY Aa id Sci 299 : 264-72. 一 (19 78) 'Late r al differ ences in the human brai n in cog nition and behavio u ral contr ol' , in P. Buser and A. Roug e ul-Buser (eds) Cerebral Correlate s of Consci ou s Exp e ri en ce, Amste r dam : Noth Holland, pp. 285-98. Levy - A g r esti, J. and Sp e rr y, R.W .(1 9 68) 'Differ enti al pe rcept ua l capa cit ies in majo r and mino r hem isp h eres', Proc Natl Acad Sc i (US A) 61 : 1151. Lie b erman, P. (19 75) On the Or igins of Lang uage , New York : Macm illan . Lig gins , G.C., Forste r , C. S., Gr iev es, S.A . and Schwa rtz, A. L. (1977) 'Contr ol of parturition in man', Bi ol Rep r od 16 : 39-56. Liggins , G. C., Kennedy, P.C. and Holm, L.W . (1967) 'Fa ilur e of initiat i on of parturition aft er electr oc oag u lati on of the pituitary of the fetal lamb', Amer J Obst Gyn ecol 98 : 1080-6. Lim b er, J. (1980) 'Lang u ag e in chi ld and chi mp', in T.A . Sebeok and D.]. Um恥 r-Sebeok (eds) Sp e akin g of Ap e s, New York : Plenum Press, pp. 197-220. Lord, AB. (19 60) The Sin g e r of Tales, Cambri dge , Mass. : Harvard Un ive rsit y Press. Lorenz, K. (1971) Stu d ie s in Anim al and Human Bemvio r , vol. 2, London : Meth u en. 一 (1977) Behin d the Mi rr or, London : Meth u en. Lovejo y , C.O. (1981) 'The orig in of man', Sc ien ce 211 : 341-50. Lovell, B. (1978) In the Cente r of Immensit ies, New York : Ha rper & Row. Lumley, H. de (19 69) 'A pa leoli thic camp at Nic e' , Sc ien ti fic Ameri ca n
220(5) : 42-50. Ly n ch, G. and Baud ry, M. (19 84) 'Th e bio c hem ica l inter medi at e s in me- mory form atio n : a new and spe cific hyp othe sis ', Scie n ce 224 : 1057-63. McDoug a ll, W.(1911) Body and M ind , London : Meth u en. McGeer, P.L. Eccles, J.C . and McGeer, E. (1987) The Molecular Neurobio l og y of the Mammalia n Brain , 2nd edn, New York : Plenum Press. MacLean, P.D . (19 66) 'Stu dies on the cort ica l repr e senta tion of cert ain basic sexual fun cti on s', in R. A. Gorski and R.E . Whalen (eds) Brain and Behavio r , vol. 3, Berkeley, CA : Univ e rsity of Ca lifornia Press, —pp .( 3159-7709) . 'The triun e brain , emoti on , and sci en ti fic bia s ', in F.O. Sch mitt (ed.) The Neuroscie n ces, Second Stu d y Prog ram , New York : —T h(e1 9 R82o)c k'Eefve olll eurt i oUn n iovfe rtshiet y p Ps yrec sh so,e npcpe. p 3h3 a6l-o4n9',. Zy go n 17 : 187-211. McNeill , D. (19 80) 'Sente n ce str uc tu re in chi m p a nzee commun ica ti on ', in T.A . Sebeok and D.J. Um ike r-Sebeok (eds) Sp e akin g of Ap e s, New York : Plenum Press, pp. 145-60. Maendly, R., Ruegg , D. Wi es endang e r, M. , W i es endang e r, R., Lago uska,J. and Hess, B. (1981 ) 'Thalam ic relay for gro up I muscle affer ents of fore lim b nerves in the monkey' , ] Neuroph ysi o l 46 : 901-17. Marge nau, H. (1984) The Mi ra cle of Exis t e n ce, Woodbri dge , Conn. : Ox Bow Press. Mark, V.H. and Erv in, F.R . (19 70) Viol ence and the Brain , New York : Ha rpe r & Row. Marr, D. (1970) 'A theo ry for cerebral neocort ex ', Proc R Soc Lond [Bio l ] 176 : 161-234. Marsden, C.D., Mert on , P.A ., Mort on , H.B., Adam, J.E .R . and Hallett, M. —(1(91 7987a8) b) 'A'uTtho em aetfifc e c at n odf vleosliuon n tsa ryo f r ethsep o cn esnetsr alt o nemrvuosculse ssyts r etet cm h oinn man', Prag Cli n Neuroph ysi o l 4 : 167-77.
long - la te n cy str et c h refl ex es in the human thu mb', Prag Clin Neuroph ysi o l 4 : 344- 41 . Marsd e n, C.D ., R oth w ell, J C . and Day, B.L . (1983) 'Long - la te n cy auto m ati c respo nses to muscle str e tc h in man : ori gin and fun cti on ', A dv Neural 39 : 509-39. Marshack A. (19 8. 5) Hi er a rchic a l Evoluti on of the Human Capa city : The pa leolith ic evid e nce. 54th Jam es Arth u r Lectu r e on The Evoluti on of the Human Brain , 1984, New York : Ameri ca n Museum of Natu r al Histo r y . Mata n o, S., • Baron, G., Ste p h an, H. and Frahm, H.D . (1985a) 'Vo lume comp ariso ns in the cerebellar comp le x of prim ate s . II. Cerebellar nuclei', Folia Prim ato l og 44 : 182-203. Mata n o, S., Ste p h an, H. and Baron, G. (1985b) 'Volume comp ariso ns in the cerebellar comp le x of prim ate s . I. Ventr al pon s', Folia Pr ima to l og 44 : 171- 81 . Matt he ws, P.B.C. (19 81) 'Ev olvin g vie w s on the int e r nal ope rati on and fun cti on al role of the muscle spi nd le', J Ph ys io l (Lond) 320 : 1-30. Mauss, T. (1911) 'Di e Faserarch itek to n i sc he Gli ed erung des Cort ex cerebri der anth r opo morp h en Affen ', J Psyc h Neurol 18 : 410-67. Maye r, M. L. and Westb r ook, G.L . (1985) 'Diva lent catio n pe rmeabil ity of N- m eth yl- D-aspa rtate channels', S oc Neurosci Abstr 11 : 785. Mayr , E. (1963) Anim al Sp e ci es and Evoluti on , Bosto n , Mass. : Bellrnap Press, Harvard Un ive rsit y Press. - (19 73) 'Descent of man and sexual selecti on ', in L'Or igine del( —U (o1m98o2, ) RTohmee G: rAowccta hd eofm Biai o Nl oagzi i(Xo 1n la lTe hdoeuig L hti,n cD eii, v ep rps. i3ty 3, -6E1v. oluti on and Inte l l ige nce, Cambri dg e , Mass. : Bellrnap Press, Harvard Un ive rsit y Press. Meller, K. and Tetz l aff W. (1975) 'Neuronal migr ati on du ring the early develop m ent of the cerebral corte x : A scan ning electr on micr oscop ic stu d y' , Cell Tis s ue Res 163 : 313-25.
Melvill Jon es, G. and Watt , D.G.D . 0971a) 'Observati on s on the contr o l of ste p p ing and hop ping movements in man', J Phys io l (London) 219 : —7 0(199-2771b. ) 'Muscular contr o l of land ing from unexp e cte d fal ls in man', J Ph ysi o l (London) 219 : 729- 37 . Menzel, W.W . (1984) 'Sp a tia l orga n iza ti on and memory in cap tive chim p a nzees', in P. Marler and H.S. Terrace (eds) The Bio l og y of Leami ng , Berlin /He id e lberg/ Ne w York : Sp ring e r-Verlag , pp. 509-31. Merzen ich , M.M. and Brug ge , J,F . 0973) 'Re p re senta t i on of the cochlear partition on the sup erior tem p o ral pla ne of the macaqu e monkey ', Brain Res 50 : 275-96. M 恥 r, B. (1966) 'Amnesia foll owi ng ope rati on on the tem p o ral lobes', in C.W.M. Whitty and O.L . Zang will( eds) Amnesia , London : —B u(t1t 9e6r8 w)o rt'V hsi s, u papl . 1r0e9co-3g3 n. ition and recall aft er righ t tem p o ral-lobe exc isi o n in man', Neurop s yc holog ia 6 : 191-209. (1972) 'Diso rders of lea rning and memory after tem p o ral-lobe —le s(1io9 n7 s4 ) in' H me amn i's, p Ch elir ni c N sepwe - coi sa ulirzga t1i o9n : 4: 2i1t-s 4 6 s.c op e and limits', in F.O. schm itt and Worden, F.G . (eds) The Neuroscie n ces. 3rd Stu d y Prog ram , Cambri dg e , Mass. : MIT Press, pp. 75— 89 . M恥 r, B. and Tayl o r, L. (1 頭) ‘Righ t hem isp h ere sup e ri or i ty in tac ti le pa tt er n recog nition after cerebral com miss uroto m y : evid e nce for nonverbal memory ', Neurops ych olog ia 10 : 1-1 5 . Monagh an, D.T . and Cotm a n, C.W. (19 8. 5) 'Di st r ibu ti on of N-meth y l - D-aspa rtate- sensit ive L[1 !] glu ta ma te bin d ing sit es in rat brai n', J Neurosci 5 : 2909-19. M—onod, (1J9. 7(l5C)J ?'lT )h eC IYanlnacley s a isn do fN secciee ns stii tfyi c, mNeewth oY d oarnk d: Kthneo plof g. ic of scie n ti fic disc overy ', in H.A . Krebs and J.H . Shelley ( eds) The Creati ve Process in Sc ien ce and Medic ine , Amste rd am : Excerp ta Med ica , pp. 3-7, 52.
Mountc a stle , V.B. (1978) 'An orga n izi n g princ ip le for cerebral fun cti on . The unit module and the dis t r ibu te d sy s te m ', in F.O . S c hm itt(e d.) The M ind fu l Brain , Cambrid g e, Mass. : MIT Press, pp. 7-50. Mountc a stle , V.B., L y n ch, J.C ., Georgo po ulos, A., Sakata , H. and Acuna, C. (1975) 'Poste r i or pariet a l associa t i on cort ex of the monkey : command fun cti on s for ope rati on wi thin extr a pe r sonal spa c e', J Neuroph ys io l 38 : 871-908. Mountc a stl e, V.B . , Mott er , B.C., Ste i n m etz , M. A. and Duff y, C.]. (19 84) 'Look ing and seein g : the vis u al fun cti on s of the pariet a l lobe', in G.M . Edelman, W .E. Gall and W. M . Cowan (eds) Dy n ami c Aspe ct s of Neocortir o l Functi on , New York : Wi ley , pp. 159-93. My er s, R.E. (19 61 ) 'Co rp u s callosum and visu al gn osis ', in ].F. Delafr es naye ( ed.) Brain Mechanis m s and Leami ng , Oxfor d : Blackwell Sc ien ti fic Publi ca ti on s, pp. 481-505. Nap ier , ]. (1962a) 'Fossil hand bones from Olduva i Gorge ', Natu r e 196 : 409- 11 . - (19 62b) 'Th e evoluti on of the hand', Sc ien ti fic Ameri ca n 207(6) : —56 (1-69267. ) 'The anti quity of human wa lking', Sc ien ti fic Ameri ca n 216(4) : 56- 66 . Nashner, L.M . (1981) 'Analys is of sta nc e pos tu re in humans', in A.L. Towe and E.S . Luschei( e ds) Handbook of Bemvio r al Neurobio l og y, vol. 5, New York : Plenum Press, pp. 5'2: 7- 65. 一 (1985) 'Str at e g ies for orga niza ti on of human pos tu re', in M. Iga rash i and F.0. Black(eds) Vesti bu lar and Visu al Contr ol on Postu r e and Locomoto r Eq u il ib r i um , New York and Basel : Karge r, pp.1 -8. Nath a n, P.W. and Sm ith, M.C. (19 55) 'Lon g descend ing trac ts in man. Revie w of pre sent knowled ge ', Brain 78 : 248-303. Nath a nie l sz, P.W. (1978) 'Endocri ne mecha nism s of parturition ', Annu Rev Ph ys io l 46 : 411-45. Og d en, J.A. (1988) 'Lang u age and memory func ti on s after lon g recovery
period s in left -h e m isp h erecto m ize d subje c ts ', Neurop s yc h olog ia 26 : 645-59. Oje m ann, G.E. and Creutz feld t, O.D . (1987) 'La ng u age in humans and anim als : contr ibu ti on of brai n stim u lati on and recordi ng ', in V.B. Mountc a stle , F. Blum and S.R. Geig e r(eds) Handbook of Ph ys io l og y, l. The Nervous Sys te m , Vol. 5, Part 2 , The Hi gh er Functi on s of the Brain , Beth e sda, MD : Ameri ca n Phy si o l og ica l Socie t y , pp. 675- 99 . Olds, J. and Milne r, P. (19 54) 'Posit ive rein forc ement pro duced by electr ica l sti m ulati on of sept al area and oth e r reg ion s of rat brai n' , J ou rnal of Comp ar ati ve and Phys io l og im l Psyc h olog y 47 : 419-27. (101] Olto n , D.S (1983) 'Memory fun cti on s and the hippo carnp u s', in W. Seif er t (ed.) Molecular, Cellular and Behavio r al Neurobio l og y of the Hi ppo - camp u s, New York : Academ ic Press, pp. 335-73. Papo usek, M., Papo usek, H. and Bornste i n , M. (19 85) 'The natu ralist i c vocal envir o runent of you ng infan ts : on the sig nifica nce of homog e neit y and va riab il ity in pa renta l spe ech', in T.M. Fie l d and N. Fox (eds) Socia l Percep tion in In fan ts , Norwood, NJ : Allen Publi sh i ng Corp , pp. 269-fJ' l. Passin g h arn, RE . and E 떠i n g er, G. (1974) 'A comp ariso n of cort ica l func ti on s in man and oth e r prima te s , Int Rev Neurobio l 16 : 233-99. Penfi eld , W. (1975) The My st e r y of the M ind , Pri nc eto n NJ : Pri nc eto n Un ive rsit y Press. Penfi eld , W. and Jas pe r, H. (1954) Ep ilep s y and the Functi on al Anato m y of the Human Brain , Bosto n , Mass. : Lit tle, Brown. Pen field , W. and Robert s, L. (1959) Sp e ech and Brain - Mechanis m s, Prince to n , NJ : Prince to n Un ive rsit y Press. Pete r s, A. and Kara, D.A . (19'6 1) 'The neuronal comp o sit ion of area 17 of rat visu al cort ex , iv. The Orga nizat i on of pyramida l cells. J Comp Neurol 260 : 573-90. Pfaff, D. W. (19 80) Estr og e ns and Brain Functi on , New York : Sp ringe r Verlag.
Ph illips , C.G 0969) 'Moto r app a ratu s of the baboon hand', Proc R Soc —L a0n9d7 1[ B) i' oElv ] o1lu73ti o: n1 41o-f 7 t4 h.e cort ico spi na l tra ct in prima te s wi th spe c ia l refe r ence to the hand', Proc 3rd Intl Cong r Prim ate s , Zuri ch 1970 vol. 2, Basel : Karge r, pp. 2-23. Ph illips , C.G . and Port er, R. (1977) Cortic o spi na l Neurones. Their Role in Movement, London /Ne w York : Academ ic Press. Ph illips , C.G., Powell, T.P.S. and Wi es endang e r, M. (1<:Jll) 'Pr oje c ti on from low thr eshold muscle aff er ents of hand and for earm to area 3a of baboon's cort ex ', J Ph ysi o l (London) 217 : 419-46. Pic k f or d, M. (19 85) 'Keny a pi the cus : a revie w of its sta tus based on newly disc overed fos sils from Keny a ', in P.V. Tobia s (ed.) Homi ni d Evo- luti on . Past, Present and Futu r e, New York : Alan R. Lis s , pp. 107- 12. Pil be am, D. (1<: J72 ) The Ascent of Man, New York : Macm illan . (19 83) 'Hom ino id evoluti on and hom inid ori gins ', in C. Chag a s (ed.) Recent Advances in the Evoluti on of Prim ate s , Vatic a n Cit y : Ponti ficiae Academ iae Sc ien ti ar um Scri pta Va ria 50, pp. 43-61. Pog gio, G.F. (1984) 'Processin g of ste re oscop ic infor mati on in prima te vi- sual cort ex ', in G.M. Edelman, W. E. Gall and W.M. Cowan (eds) Dy- nami c Aspe cts of Neocort ica l Functio n , New York : Wi ley , pp. 613- 35. Pog gio, G.F . and Fis c her, B. (1<: J77 ) 'Bin o cular inter acti on and dep th sensiti vi t y of str iat e and pre str iat e cort ica l neurons of the behavi ng rhesus monkey' , J Neuroph ysi o l 40 : 1392-405. Pog gio, G.F. and Pog gio, T. (1984) 'The analys is of ste r eop s is ', Annu Rev Neurosci 7 : 379-412. Polk ingh orne, J. (1986) One World : The Inte r acti on of Sc ien ce and Theology , London : SPCK . Pop per, K.R. (1<: J72 ) Obje c ti ve Knowledg e . An Evoluti on ary Ap pr oach, Ox for d : Clarendon Press.
— (1975) 'Th e analys is of scie n ti fic meth o d and the log ic of scie n ti fic disc overy ', in H.A . Krebs, and J.H . Shelley (eds) The Creati ve Proc- ess in Sc ien ce and Medic ine , Amste r dam : Excerp ta Med ica , pp. 17- —19 (.1 9 77) Part I of K.R. Pop pe r and JC . Eccles, The Self and its B rain , —B e(1rl9i 8n1 /)H e' Tid he elb ePrlgac/ Le o onfd o Mn /iNnde win Y Noarktu :r eS', p rNinog b ee rl -VCeonrlfae gr e. nce, St Pete r , —M i(n1n9 8. 2 : )G uTshtae fuOs pA e ndo lpUh n uisv e Crsoell e: gA e. n Argu ment for Indete r mi ni s m , London : Hutc hins on. (19 87) 'Natu ra l selecti on and the emerge nce of mind ', in : G. Rad nitzk y and W.W. Ba rtley (eds) Evoluti on ary Ep iste m olog y. Theory of Rati on ality and the Soc iol og y of Knowledg e , La Salle : —O p( 1e9n 8 8C)o u'A rt , npepw. 1 3i9n-t 5e 5r p. re ta t i on of Da rwinism . The Fir st Medawar Lectu re', Proc R Soc Lond [Bi ol ] (in pre ss). Popp er K.R. and Eccles, JC . (1977) The Self and Its Brain Berlin / Heid e lberg/ Lo ndon /Ne w York : Sp ring e r-Verlag Inte r nati on al. Powell, T.P. S. Mountc a stl e, V.B. (1959) 'Some aspe c ts of the fun cti on al orga niza ti on of the cort ex of the po stc e ntr al gyros of the monkey : a correlati on of finding s obta ine d in a sin g le un it analys is wi th cyt o- arch itecture', Bull Joh ns Hopk in s Hosp 105 : 133-62. Premack, A.J and Premack, D. (1972) 'Teach ing lang u ag e to an ape ', Scie n ti fic Ameri ca n 2'27 ( 4) : 92-9. Premack, D. (1976) Inte l lig en ce in Ap e and Man, Hillsd ale, NJ : Lawrence Erlbaum. Pug h , G. E. (1978) The Bio lo g ica l Orig in of Human values, London : Routle dge & Keg a n Paul. Rakic, P. (1972) 'Mode of cell migr ati on to the sup e rf icial laye rs of fet a l monkey neocort ex ', J Comp Neurol 145 : 61~84. Righ tm ire, G.P. (19 84) 'Homo sapi en s in sub-Sahara, Africa ', in F. H.
Sm ith and F. Sp e ncer (eds) The Orig ins of Modem Humans, New York : Alan R. Lis s , pp. 295-325. Roberts , G.W., Woodhams, P. L., Polak, J. M. and Crow, T.J . (19 82) 'Di s t r i b u ti on of neurop e pi tide s in the limb ic sys ye m of the rat : the amy gd aloid comp le x', Neuroscie n ce 7 : 99-131. Roland, P. E. (1981) 'So mato t o p i ca l tun ing of po stc e ntr a l gyru s du ring foc al —at (t 1e9n 8t.i 5o)n 'Cino rmt icaan l' , aJc tNi vei tu yr oipnh yms aion l 4d6u r:i 7n4g 4 -d5i4s.c ri mina ti on of extr ins ic pa tt er ns and retr iev al of int r ins ic pa tt er ns', in C. Chag a s, R. Gatt as s and C. Gross (eds) Patt er n Recog nition Mechanis m s, Vati ca n Cit y : Ponti fici a e Academ iae Scie n ti ar um Scri pta Va ria 54, pp. 215-46. Roland, P. E. and Fri be rg, L. (19 8. 5) 'Loca liza ti on of cort ica l areas by thinking ', J Neuroph ys io l 53 : 1219-43. Roland, P. E. and Larsen, B. (1976) 'Fo cal inc rease of cerebral blood flow duri ng ste r eog n osti c tes ti ng in man', Arch Neural 33 : 551-8. Roland, P. E. and Sk inh ~j , E. (19 81 ) 'Ex tr as tr iat e cort ica l areas acti va te d duri ng vis u al stim u lati on in man', Brain Res 222 : 166-71. Roland, P. E., Larsen, B., L assen, N.A . and Sk inh ~j, E. (1980) 'Su pp le -menta ry moto r area and oth e r cort ica l areas in orga niza ti on of volun- tary movements in man', J Neuroph ys io l 43 : 118-36. Roland, P. E., Sk inh ~j, E. and Lassen, N.A . (19 81 ) 'Focal acti va ti on of human cerebral cort ex du ring aud itor y disc ri mina tio n ', J Neuro- phys io l 45 : 1139-51. Rubens, A. B. (l
Lib r ar y. Sakurai, M. (19 87) 'Sy n ap tic mod ifiab il it y of pa rallel fibr e-Purki nj e cell tran sm iss io n in in vit ro gu in e a-p i g cerebellar slic e s', ] Phys io l (London) 349 : 463-80. Sa rich , V. M. and Cron in, J. E (1977) 'Molecular sys te m ati cs of the prima te s ', in M. Goodman and R. E. Tashan (eds) Molecular Anth r op o logy , New York : Plenum Press, pp. 141 -70. Sasak i, K. and Gemba, H. 0982) 'Developm ent and chang e of cort ica l field poten ti al s du ring lea rning pro cesses of vis u ally initiat e d hand movements in the monkey' , Exp Brain Res 48 : 429-37. Savage -Rumbaug h , E. S., Sevcik , R. A., Rumbaug h , D.M. and Rubert , E. 098.5 ) 'The capa ci ty of anima ls to acq uire lang u ag e : do spe c ie s differ ences have any thing to say to us?' Phil Trans R Soc Lond [Biol ] 308 : 177-8.5 . Schultz, A. H. (1968) 'Th e recent hom ino id prima te s ', in S. L. Washburn and P. C. Jay (eds) Perspe cti ve s on Human Evoluti on , vol. 1, New York : Holt, Rine ha rt & Wi ns to n . Searle, J. 0984) Mi nd s, Brain s and Sc ien ce, London : Bri tish Broadcastin g Corp ora ti on . Sebeok, T. A and Um ike r-Sebeok, D. J. (1980) Sp e akin g of Ap e s, New York : Plenum Press. Shankweile r, D. P. 0966) 'Eff ec ts of tem p ora l-lobe damag e on per cep tion of dich oti ca lly pre sente d melod ies ', J Comp and Ph ys io l Ps ych o! 2 : 115-19. Sherr ington , C. S. (1940) Man on Hi s Natu re , Cambri dg e : Cambri dg e Un ive rsity Press. ——S—im o ns (,(( 111 9E99887. . 713L))) . '''RMR(1eaa9cm en7n2'as)tp ii aPmthdrmev icmaeun d ascit'ae,e ts e S E icnpv ie roken l nurtuiot fniiw ocnnl e e A, drsNmg', e ee Pwroifh c Y a it lnh o Te r2 kr3ea 6:an (rMs5li )e R as:c t2m S8 o oi-fl3 lc a5t hn.L e.o Endg y[pBtiio aln ] 292 : 21-41.
olig o cene, inc lud ing the most recent known pre sumed ancesto r s of man', in C. Chag a s(ed.) Recent Advances in the Evoluti on of Prim ate s , Vati ca n Cit y : Ponti fici a e Academi ae Scie n ti ar urn Scri pta —V (a1 r9ia8 5 5) 0,' Apfp r.i c1a 1n- 없ori gin, characte r i st ic s and conte x t of earlie s t high er prima te s ', in P.V. Tobia s (ed.) Homi ni d Evoluti on . Past, Present and Futu r e, New York : Alan R. Lis s , pp.1 01-6. Sim ps on, G.G . (1964) Th is View of Lif e : the World of an Evoluti on is t ', New York : Harcou rt, Brace & World. Sm ith, F. H. (19 84) 'Fossil hom inids from the Up pe r Pleis t o c ene of centr al Europ e and the ori gin of modern Europe a ns', in F. H. Sm ith and F. Sp e ncer (eds) The Or i.gi n of Modem Humans, New York : Alan R. Lis s , pp. 137-209. —Solecki ,( 1R 9.7 7S). '(T1h97e1 )i mSphlaicna itd ia o rn ,s Noef w t hYe oSrkh a: nKidna orp fC. ave Neandert ha l flow er buri al' , Ann NY Acad Sc i 293 : 141-24. Sp e rr y, R.W . 0974) 'Late r al spe cializa ti on in the surgi ca lly sepe r ate d hem isp h eres', in F.O. Schm itt and F.G. Worden (eds) The Neuroscie n ces. 3rd Stu d y Prog ram , Cambri dg e , Mass. : MIT Press, pp. 5- 19 . Sp e rr y, R. (19 82) 'Some eff ec ts of disc onnectin g the cerebral hem isp h eres', Sc ien ce 217 : 1223-6. - (1983) Scie n ce and Moral Pr ior i ty, New York : Columbia Un ive rsity Press. Sp erry, R. W., Gazza niga , M.S. and Bog e n, J.E . (1969) 'Inte rh e misp h eri c relati on shi ps : the neocort ica l com miss ures; syn d romes of hem isp h ere disc onnecti on ', in P.J. Vin ken and G.W. Bruyn (eds) Handbook of Clin i c a l Neurolog y, New York : Wi ley, pp. 'n3 -90. Sp e rr y, R. W., Za ide l, E. and Za ide l, D.(1 '579 ) 'Self-r ecog nition and socia l awareness in the deconnecte d mino r hem isp h ere', Neurops ych olog ia 17 : 153-66.
Sq u ir e, L.R. (19 82) 'Th e neurop s yc h olog y of human me m o ry ', A nnu Re v —N e(1u9r8o3s) ci' T 5h :e 2 4h1 ip- 7p3o .c amp u s and the neurop sy c h olog y of memory ·, i n W. Seif ert ( ed.) Molecular, Cellular, and Behavio r al Neurobio l og y of the Hi ppo camp u s, New York : Academ ic Press, pp. 491-50 7 . Ste b bin s , G.L. (19 82) Dannin to DNA, Molecules to Humanit y, New York : Freeman. Ste n t, G.S . (19 81) 'Str en g th and weakness of the ge neti c app ro ach to the develop m ent of the nervous sys te m ', Annu Rev Neurosci 4 : 163-94. Ste p h an, H. 0975) 'Allocort ex ', in Handbook der mi kr oscop isc he Anato m i e des Menschen. 4 : 119. Heid e lberg : Berga ms, Sp ring er . (19 83) 'Evoluti on a ry tre nds in limb ic str uc tu r es', Neurosci Bi ob ehav Rev 7 : 367-74. Ste p h an, H. and Andy, O.J (1969) 'Qu anti tative comp a rati ve neuroanato m y of prima te s : an att em p t at ph y lo g e neti c int e r p r eta t i on ', A nn NY Acad —Sc 0 i9 71767) :' Q37u 0a-n8t7i t.a tive comp ariso n of the amy gd ala in ins ecti vo res and prima te s ', Acta Anat 98 : 130- 53 . Ste p h a n, H., Baron, G. and Frahm, H.D. (1987, 1988) per sonal cornmuni— cati on s. St rau sfe ld , N.J. 0976) At la s of an Insect Brain , Berlin /He id e lberg/ Ne w York : Sp ringe r-Verlag. Str aw son, P. 0959) Indiv i d u als, London : Meth u en. Str ing e r, C.B., Hablow, JS . and Vandermeersch, B. 0984) 'Or igin of anato m ica lly modern humans in Weste rn Europ e' , in F.H . Sm ith and F. Sp en cer (eds) The Orig ins of Modem Humans, New York : Alan R. Liss , pp. 57-135. Szenta go th ai, J. 0'5 70 ) 'Les cir cu its neuronaux de l'ec orce cerebrale', B ull —A c(1a9d7 8R) o y'T Mhee dne Bureolng nVeItIw, oXr k: 4o75f - t9h2 e. cerebral
cort ex : A fun cti on al interpre ta tion ', Proc R Sol Lond [Bio l ] 201 : 219-48.一 (198 3) 'Th e modular archit ec t o n ic pri n c ip le of neural cente r s', Rev Phys i o l Bio c hem Pharmacol 98 : 11- 61 . Te ilh ard de Chard.i n , P. (19 59) The Phenomenon of Man, New York : Ha rpe r. Terrace, H.S . (1985) 'Anim al cog nition : think ing wi tho ut lang u ag e ', Ph il Trans R Soc Lond [Bi ol ] 308 : 113- 28 . Terrace, H.S. and Bever, T .G . (19 80) 'W hat migh t be learned from stu d y ing lang u age in the chi mpa nzee? The impo rt an ce of sym boli zi n g oneself', in T.A . Sebeok and D.]. Um ike r-Sebeok (eds) Sp e akin g of Ap e s, Ne w York : Plenum Press, pp. 179-89. Teuber, H.L . (1967) 'La cunae and research app ro aches to the m', in C.H. M 沮i kan and F.L . Darley (eds) Brain Mechanis m s Underlyi ng Sp e ech and Lang u age , New York/ Lo ndon : Grune & Str at t on , pp. 204-16. Thorburn, G.D. , N ic o l, D.H., B assett , J .M . Schutt , J .M . and Cox, R.I.(1'572 ) 'Pa rturition in the go at and sheep. Chang e s in cort ico ste r oid s , pro g e ste r one, oestr og e ns and pro sta gla nd ins ', J Rep r od Fert [Sup pl] 16 : 61- 84 . Thorp e , H.W . (1966) 'Eth o log y and Conscio u sness', i n JC . Eccles (ed.) Brain and Consci ou s Exp e rien ce, New York : Sp ringe r-Verlag, —pp (1. 947740 )- 5A05n.i m al Natu r e and Human Natu re , London : Meth u en. (19 78) Purpo se in a World of Chance, Ox for d : Ox for d Un ive rsit y Press. Tobia s , P.V. (19 71) The Brain in Homi ni d Evoluti on , New York : Columbia —U n( 1iv9e7 r5sai)t y ' BPr raeis ns . evoluti on in the Hom ino id e a', in R.H . Tutt le (ed.) —Pr(i1 m9 7 a5tbe ) Fu'Lnocntgi o no arl Mshoorrpt h ohloomg yin iadn dp hE yv lo luti on , The Ha gue : Mouto n , pp. 353- 92 . og e ni es ? Paleonto l og ica l and molecular evid e nces', in F.M. Salzano (ed.) The Role of Natu ro l Selecti on in Human Evoluti on , Amste r dam : Nort h Holland, pp. 89-
—11 (189. 8 1a) 'Th e emerge nce of man in Africa and bey o nd', P hil Trans R Soc Lond [Bi ol ] 292 : 43-56. (19 81b) 'Ev oluti on of Human Brai n, Inte l lect and Sp irit', Andrew Abbie Memoria l Lectu r e, Adelai de Univ e rsit y Press, Adelai de . 一 (1983) 'Recent advances in the evoluti on of the hom inids wit h spe c ia l refe r ence to brai n and spe e ch', in C. Chag a s (ed.) Recent Advances in the Evoluti on of Pr ima te s , Vati ca n Cit y : Ponti fici a e Academ iae Scie n ti ar um Scri pta Va ria 50, pp. 85- 14 0. (19 86) The Revoluti on in Human Evoluti on , Geneva : Dudley —W (r1 i9g 8h7 t) CTohlloeq ub iraa .i n of Homo habil is : a new level of orga niza ti on in cerebral evoluti on ', J Hum Evol 16 : 741-61. Tower, S.S. (1940) 'Pyramida l lesio n in the monkey' , Brain 63 : 36-90. Tr ille r, A. and Korn, H. (1982) 'Transm iss io n at a centr al inhibi t or y syn a p se . III. Ultr as tr uc tu re of ph y si o l og ical ly ide nti fied and sta ine d ter m ina ls', J Neuroph ysi o l 48 : 708-36. Tr inka us, E. (19 83) The Shanid a r Neanderto o ls, New York : Academ ic —P r(e1s9s 8.4 ) 'Weste r n Asia ', in F. H. Sm ith a
nd F. Sp e ncer (eds) The Orig ins of Humans, New York : Alan R. Liss , pp. 251-93. Tr inkau s, E. and Howells, W. W . (1 罪 9) 'The Neandert ha ls', Scie n ti fic Ameri ca n 241 (6) : 95-105. Tutt le, R. H. (1985) 'Ap e foo tp rints and Laeto l i impre ssio n s : A respo nse to SUNY clai ms ', in P.V. Tobia s (ed.) Homi ni d Evoluti on . Past, Present and Futu r e, New York : Alan R. Lis s , pp. 129-33. Ung e rleid e r, L.G . (1985) 'Th e cort ico cort ica l pa th w ays for obje c t recog nition and spa ti al per cep tion ', i n C. Chag a s, R. Gatt as s, and C. Gross (eds) Patte r n Recog nition Mechanis m s, Vati ca n Cit y : Ponti - ficiae Academ iae Scie n ti ar um Scrip ta Va ria 54, pp. 21-37. Utt al, W .R. (lfl1 8 ) The Ps ych obio l og y of M ind , Hillsd ale, NJ : LawrenceErlbaum . Van Essen, D.C . 0985) 'Functi on al orga niz a ti on of prim ate vis u al cort ex ', in A. Pete r s and E.G . Jon es(eds) Cerebral Corte x , Vol. 3, Visu al Corte x , New York : Plenum Press, pp. 259-329. Wada, J.A., Clarke, R. and Hamm, A. (1975) 'Cerebral hem isp h eri c asym metr y in humans', Arch Neurol 32 : 239-46. Wadd ing ton , C.H . (1969) 'The the ory of evoluti on tod ay' , in A. Koestl er and JR. Smy thies (eds) Beyo nd Reducti on is m , London : Hutc h i ns on, pp. 357-95. Walker, A. and Leakey, R.E .F . 0978) 'The hom inids of East Turkana', Scie n ti fic A meric a n 239(2) : 54-66. Wa rring ton , E.K. 0985) 'Vi su al defi ci t s associa t e d wit h occip ital lobe lesio n s in man',. in C. Chag a s, R. Gatt as s, and C. Gross (eds) Patt er n Recog nition Mechanis m s, Vati ca n Cit y : Ponti fici a e Academ iae Scie n ti ar um Scri pta Va ria 54, pp. 247-61. Washburn, S.L. (1960) 'To ols and human evoluti on ', Sc ien ti fic Ame ri따 203(3) : 63-75. (1969) 'Th e evoluti on of human behavio u r', in The Uniq u eness of —M 0a9n7, 8A) m'Tshtee r deavmol uLtoi onn d oonf :m Naonr',t hS-H c ioenl ltai nfidc, Apmp.e 1r67i-函89 .2 39(3) : 194-204. Weis k rantz , L. 0974) 'The intera cti on betw e en occi pital and tem p ora l cort ex in vis i o n : an overvie w ', in F.O. Sch mitt and F.G. Worden (eds) The Neuroscie n ces. 3rd Stu d y Prog ram , Cambri dg e , Mass. : MIT Press, pp. 189-204. (1985) 'Intr od ucti on : cate g o ri za ti on , cleverness and consci ou sness', Phil Trans R Soc Lond [Bi ol ] 308 : 3-1 9 . Wheeler, J.A. (1昭) ‘The Un ive rse as a home for man', Amer Sc ien t ist 62 : 683-91. W 血 e, M. JD . 0978) Modes of Sp e ci at i on , San Franc isc o : Freeman. Wi es endan ge r, M. (1981a) 'Orga niza ti on of seconda ry moto r areas of the cerebral cort ex ', in V.B. Brooks(ed.) Handbook of Ph ysi o l og y, 7
Cerebral Cont rol Mechanis m s; Neurop h ys i o l og y, Beth e sda, MD : —A m(1e9r8i 1cba )n 'PThhye s ipo ly orga imc a ild a Sl octriae ct yt ., pItps. 1s1t2r u1 c-4tu7 r. e and fun cti on ', in AL, Towe and E.S. Luschei( e ds) Handbook of Behavio r al Neurolog y, vol 5. Moto r Coordin a ti on , New York : Plenum Press, pp. 401- 9 1. Wi es endang e r, M. and JS . Mi le s (1982) 'Ascendin g pa th w ay s of low thr eshold muscle aff er ents to the cerebral corte x and its po ssib l e role in muscle contr ol' , Ph ysi o l Rev 62 : 1234-70. Wi ls on, E.O. (1975) Soc iob io l og y. The New Sy n th e sis , Cambri dg e , Mass. : —T h(1e9 7B8e) lkOnna pH Purmesasn, HNaartvu ar red, CUanmiv be rris digt y e , PMreassss.. : Harvard Univ e rsit y Press. Wi ls on, V.J (198. 3) 'Oboli th- spi na l refl ex es', in M. lga rash i and F. Owen Black (eds) Vesti bu lar and Visu al Contr ol on Postu r e and Locomoto r Eq u il ib r iu m , Basel : Karge r, pp. 177- 8. 5. Wu, R. (19 84) 'The cran ia of Ramap ithe cus and Sil v ap ithe cus from Lufe n g , Ch ina ', Cour Forsch Inst Senckenberg 67 : 41-8. Yeni -K omshi an , G. and D. Benson, (19 76) 'Anato m ica l stu d y of cerebral asym me tr y in the tem p o ral lobes of humans, chi m p a nzees and rhesus monkey s' , Scie n ce 192 : 387-9. Zang w i ll, O.L. (19 74) 'Consci ou sness and the cerebral hem isp h eres', in S.J Dim o nd and JG . Beaumont (eds) Hemi sp h ere Functi on in the Human Brain , New York : Halste d Press, Wi ley , pp. 264-78. Zeki , S.M . (1969) 'Repr e senta tion of centr al vis u al fiel ds in pr estr iat e cort ex —of ( 1m97o3n)k e'yC' ,o lBourra inc o Rd iensg 1i4n : 2rh71e-s9u1s. monkey pr estr iat e cort ex ', Brain —R (e1s9 7583) : '4U2n2 i-f7o.r mity and dive rsit y of str uc t ure and fun cti on in rhesus —m (o1n9k8e0y) 'pTrhe es trr ieaptr e e sveins ut a atli ocno rot ef x c ',o lJo uPh ys io l( London) 277 : 273-90. rs in the cerebral cort ex ', Natu r e 284 : 412-18.
기 가드너 116, 209 가성 애타심 166 가소성 122 가지치기 과정 25 가치 체계 323 감각 입력 93 감각피질 70 감정의 표현 148 게슈윈드 128, 136 겜바 238 고관절 전략 86 고급형 107 곡예효과 117 골격 진화 75 골드슈미트 25 골반골 75 골패의 판화 195 공통 조상적 연속성 21 광범한 무시의 무언극 284 괴리진화 200 교차감각양식 128 교차연결 절개술 297 구달 206 구문론법의 요건 111 구심성 섬유 93
구아닌 13 굴드 20, 25, 155 그리핀 251, 256 그릴너 87 그물척수로 74 그물핵 74 근방추 93 근전도 90 글루타메 이 트 수용체 221 기계 속의 유령 331 기공성 200 기능적 비대칭성 283 기능적 신전반사 94 기무라 287 기억 205 기억의 공고화 244 기저핵 73 기피반응 149 기회원인론 250 긴게리치 31 긴고리 반사 97 긴고리 신경회로 94 긴고리 지연시간 97 길가메시의 서사시 167
L 내관 327 내관적 의식성 327 내부 계획 265 내시너 85 내재하는 신 353 네안데르탈 47, 133 네안데르탈인 167 노르아드레날린 151 논쟁적 기능 109 뇌간 74 뇌궁 147 뇌량 147 뇌 분비설 250 뇌의 음악적 수행 능력 287 뇌하수체 147 C 다윈 18 다윈주의적 혁명 115 다트 33 단안시 175 대뇌반구 136 대뇌의 비대칭성 281 대뇌피질 69
대뇌피질 부위별 대사율 131 대뇌피질의 진화적 발달 133 대뇌화 계수 65 대뇌화 지수 67 대뇌 활동 지도 245 데스메트 265 데옥시리보핵산 13 데이비드 래크 341 데카르트 114, 249, 326 덴드론 275 델가도 149 도구 92 도구 문화 192 도르프 253 도브잔스키 293 도파민 151 동물 의식성의 문제 256 동물의 종과 진화 255 동일론 258 동종집 합성 LTP 221 두보아 44 드리오피테쿠스 28 2 라마피테쿠스 29 라 메트리 250
러브조이 160 레세르핀 152 레톨리 36 레톨리의 발자국 79 렌네버그 111, 210 로렌츠 294 로만 이 카할 217 로버츠 123, 125 로버트슨 265 롤랜드 186, 265 루이 45 룸바우 119 리그비 117 리긴스 157 리키 43 림버 115 □ 마르게노 273 마르스덴 97 마리 미키 45 마우스 125 마우트너 Mau th ner 세포 '2:18 마음-신경 사건 (MNE) 뉴런 업 1 마음의 성질 3'2: 1 마이르 21, 255
마카크 171 마크 150 망상척수로 90 매장 습관 167 매컬로흐 125 멜빌 존스 90 명시적 기억 244 모나간 226 모노 256 모듈 291 무릎 전략 86 무스테리아 53 무스테리아 문화 194 문화적 진화 317 문화적 창조성 318 문화적 풀 322 문화적 형판 321 물리적 세계의 폐쇄성 없 1 미국신호언어 116 미래의 기억 332 미세국소 가설 업 5 미토콘드리아 DNA 분석 59 민첩성 77 밀 너 148, 286, 287
ki 바움가르트너 201 반복적 무의미한 그림 시험 287 반응잠복기 240 발달상의 잡음 344 발성 기구 137 방사성 연대측정법 81 방사성 제 논 Xenon 법 186 방사성 제논법 245 방출 또는 신호 기능 108 버딩 49 베르니 케 121, 125 베버 113, 120 베일리 125 변두리 격리 집단 21 변연계 143, 153 변연계-시상하부 149 변연계와 시상하부의 약리학 151 보닌 125 보조 운동 영 역 (SMA) 261 부로카 영역 43 부속운동영역 123 부수 운동 영역 71 부위별 뇌혈류량 186 부족사회 60 부톤 268
부호의 학습 210 분계선조 143 분리된 뇌 299 분자 시계 24 불확실성 원리 274 뷸러 107 브라운 210 브로달 285 브로드만 125 브로카 122, 125 블랑카드의 판화 196 비대칭성 136 비복근 90 비센댕거 88 비셀 173 人 사고과정 109 사라 210 사리치 24, 31 사사키 238 사역 적 조건훈련법 119 사역 조건 학습 240 사이콘 276 사회생물학 256 상모실인중 185
상상력 200 상징적 의사소통법 118 상측두엽 44 상호작용론 249 생물학적 조건 319 생크바일러 287 샤니다르 60 서술적 기능 108 서술적 기억 245 석기문화 45, 53, 194 선조전 시각영역 179 설 Searle 330 섬광시험법 301 성형적 기술 200 세계 1 256 세계 2 256 세로토닌 151 세로토닌 수용체 152 세베옥 116 세포외 유출 270 소뇌 73 소뇌의 진화 229 소두중 난쟁이 136 손 우선성 283 손재주 92 솔로인 59 수동적 다윈설 314
수렴영역 128 수상 서식 24 수용체 92 쉐링턴 168 스콰이어 245 스쿨 53 스테빈스 331 스테판 63, 153 스트링거 60 스페리 305 시각-구성 영역 198 시각영역 125 시각 영역 171 시각운동계의 진화 171 시각적 인식불능증 185 시각정보를 분석 179 시각 정보 처리 254 시각 진화 191 시간의식 331 시냅스 소포 268 시몬스 31 시상-피질 섬유 94 시 상하부 143, 147 시쇼어 시험 286 시토신 13 식충동물 66 신경 반옹 173
신경분지 팽대부 268 신경사건 (NE) 뉴런 271 신경전달물질 151 신신피질 306 신-조절 병행론 250 신피질 147 신피질 크기 지수 133 실독증 125 실어증 121 실어중 구획법 123 실용적 113 。 아데닌 13 아미노산 라세미화법 56 아슐리안 45 아슐 문화 192 아이삭 161 아일스 97 아편류 152 암스트롱 3 업 암시적 기억 244 애타심 166 앤디 63 양각기립 84 양각 보행 77, 91
양안시 171 양자역학의 확률계 274 어머니 자연 168 어빈 150 어의학적 요건 111 언어기전 124 언어 생성의 진화 137 언어 영역 44 언어영역 129 언어의 수준 107 언어의 학습 111 언어적 표현 109 언어 중추 121 에른스트 곰브리치 202 에스트로겐 152 엔케팔린 153 엘드레지 20, 25 역행성 건망중 243 연락뇌 296 연 락뇌 Lia iso n Brai n 모듈 267 염색체 18 영 역 39 125 예상 진화 200 예술적 창조성 171 엘리넥 47 오리나시아 53 오스트랄로피테신 28, 33, 91, 133,
165 오스트랄로피 테 쿠스 로부스투스 41 오스트랄로피테쿠스 보이세이 41 오스트랄로피테쿠스 아파렌시스 36 오스트랄로피테쿠스 아프리카누스 35 오스트랄로피테쿠스 아프리카누스 에티오피쿠스 38 오스트랄로피테쿠스 아프리카누스 탄자니엔시스 38 오스트랄로피테쿠스 아프리카누스 트란스 발렌시스 38 오제만 123 올도완 문화 194 올두바이 문화 43, 192 올즈 148 와슈 116, 209 와시번 166 와우관 수용체 129 와트 90 외측 술상체 171 의측쐐기다발핵 94 요한손 36 우미케르 세베옥 116 우발적 음성 변이 190 우연과 필연 256 우주적 지성 342
운동-서열 시험 263 운동성 실어증 122 운동신경 70 운동피질 70 운동 학습과 기억 205 워딩턴 344 원인동물초과 27 윌리엄 제임스 254 윌슨 90, 256 유두체 147 유인 동물 18 유인원의 언어 훈련 114 유전자계 17 유전자 돌연변이 17 유전자 풀 322 유전적 형판 321 윤나선상종말 93 의식의 진화 252 의식적 시간-결합성 200 의존성 약물 153. 이분 청취시험 288 이식전극법 123 이원론 249, 326 이원론적 상호작용론 'l:tl 이 원론적 상호 작용설 151 이 종접 합성 LTP 221 이집토피테쿠스 切
이차 청각영역 130 이토 230 인간의 기억 243 인격 315 인구학적 전략 158 인성 316 인성의 사다리 319 인지적 기능 306 인지적 학습 또는 기억 205 일원론적 유물론적 철학 326 일차 시각영역의 손상 184 일차 시각피질 171 일차 청각영역 129 입체시 177 잉그바르 332 x: 자동가설 270 자동제어 고리 93 자동 학습 과정 138 자아 315 자아의 개념 120 자아의 단일성 296 자아의식 293 자아 인식 293 자연도태 17
자연도태설 18 자유의지 339 잔류 추체로 88 장기간 강화 LTP 217 장기간 억제 230 적색척수로 74 적색핵 74 적자생존설 18 전네안데르탈인 58 전두전엽 192 전이인설 60 전정신경로 90 전정척수로 74 전정핵 74 절차적 기억 245 점돌연변이 22 점진적 계통발생설 20 접합부(시냅스) 267 정서 행동 148 정신분열증 150 정신운동성 간질환자 150 정자미인 326 제 4 브로드만 영역 70 제니 122 제르 레비 304 제리슨 64 제인 구달 116
제한지도법 59 종화 22 주관적 모방 336 중격 핵 144, 147 중성 돌연변이 18 중심 고랑(롤란도) 70 중추신경절 147 지상생활 161 지상 서식 24 직립자세 75 天 : 찬 94 찰스 다윈 250 창조성 111 창조적 상상 334 척수 63 척수반사 94 청각 계통 129 청각 회로 129 체감각 영역 94 초월적 신 353 초유전적 설계 344 총괄적 행동주의 자 251 추상신경세포 70 추체로 70, 87
추체로증후군 88 축삭 70 측두엽 간질 150 치밀돌기 268 침팬지 116 = 카손 22 카프체 53 칸 59 칼날문화 195 케냐피테쿠스 아프리카누스 28 켈로그 115, 209 콘후버의 이론 225 쾌락 중추 148 크기 지수 69, 153 크레오드 344 크로닌 24, 31 크로이츠펠트 123 E 타옹 38 타조상설 21 타조상적 종화 21 탐사적 또는 능동적 다윈설 314
테라스 113, 120 토비아스 31, 156 트린카우스 47 티민 13 고 파우츠 117 파이글 258, 270 파이움 27 파킨슨씨 병 87 파푸섹 137 펜필드 123, 125 펩티드 전달물질 152 편도체 143, 154 평형석 기관 90 포유류 중추신경계 63 포 퍼 107, 202, 244, 256, 315 폰지드과 27 표현적 또는 징후적 기능 108 푸르네스 115 푸르킨제 세포 234 푸린 13 프레막 118 프로인드 87 프로콘솔 28 피리미딘 13
피막중격 147 피질-망상 척수로 87 필립스 94 근 하급언어 107 하이젠베르크 274 하전두엽 43 하측두엽 43 학습 205 학습적 기능 113 한시적 평형 21 할리 데이 113 해마 147, 217 해마 절제술 224 해밀턴 283 헤스 148 해이스 115 헤켄 284, 286 혈류 131 형체 탐지 신경세포 183 형판 321 호모 사피엔스 네안데르탈렌시스 47 호모 사피 엔스 사피 엔스 47, 51 호모 에 렉투스 44, 133 호모 에렉투스 에렉투스 45
호모 에렉투스 페키넨시스 45 호모 프레네안데르탈리스 49 호모 하빌리스 28, 41, 43, 133 호미노이드 31 호미니드 31 호웰스 47 홀로 웨이 35 화이트 36 환경요소 319 훔멜샤임 87 휴벨 173 흥분성 접합후 전위 217 히포크라테스 249 힐 118
3 계 철학 109 EPSP 217 HSN 47 HSS 47 H 반사 97 LSD 152 NMDA 이온 통로 223 QQ 수용체 223 rnRNA 15 a- 운동신경 93 r- 운동신경 93
박찬웅 서울대학교 의과대학 졸업 서울대학교 대학원 의학박사 현재 서울대학교 의과대학 교수 서울대학교 의학연구원 원장겸 신경과학연구소 소장 뇌의 진화 一자아의 창조 대우학술총서 번역 113 1 판 1 쇄 펴냄 _1998 년 3 월 30 일 지은이―一-촌 에클스 옮긴이_박찬옹 펴낸이 * ~朴孟浩 펴낸곳-(주)민음사 출판등록 1966. 5. 19. 제 16-490 호 서울특별시 강남구 신사동 506 대표전화 515-2000, 팩시밀리 515-2007 값 25,000 원 한국어판 ® (주)민음사, 1998 뇌 KDC/511.4 8 13 Printed in Seoul, Korea ISBN 89-374-4113-6 94470 89-374-3000-2 (세트)
1 대우학술총서번역) 1
1 유목민족제국사 란덴 / 송가중 2 수학의 확실성 클 라인 / 박세희 3 중세철학사 와인버그/강영계 4 日本語의 起原 밀러 / 김방한 5 古代漢語音韻學槪要 칼그 렌 /최 영애 6 말과 사물 푸코/이 광래 7 수리철학과 과학철학 와일/김상 문 8 기후와 진화 피어슨/김준민 9 이성 진리 역사 파트남 /김 효명 10 사회과학에서의 場理論 레빈/박재호 11 영국 의 산업혁명 딘 / 나경수 이정우 12 현대과학철학논쟁 쿤 外/조승옥 김동식 13 있음에서 됨으로 프리고진 / 이철수 14 비교종교학 바하 / 김종서 15 동물행동학 하인드/장현갑 16 현대우주론 시아마/양종만 17 시베 리아의 샤머니즘 디오 세지 호괄/최길 성 1 8 조형미술의 형식 힐데브란트/조창 섭 19 힐 버트 리드/ 이일해 20 원시국가의 진화 하스 / 최몽 룡 21 商文明 張光直/윤 내현 22 마음의 생태학계 1 이츤/서석봉 23 혼돈 속의 질서 프리고진 스텐저스/유기풍 24 생명의 기원 밀러 오르켈/박안원 25 일반언어학이론 야콥슨1 권재일 26 국가권력의 이념사 마이네케/이광주 27 역사학 논고 브로델/이정옥 28 유럽의식의 위기 I 아자르/조한경 29 유럽의식의 위기 Il 아자르/조한경 30 일반국가학 켈젠/민준기 31 일반언어학강의 소쉬르/최승언 32 일반체계이론 버틀란피/현승일 33 현대문명의 위기와 기술철학 애거시/이군현 34 언어에 대한 지식 촘스키/이선우 35 음운학원론 트루베쯔코이/한문희 36 우주의 발견 하위 트/강용희 37 수학적 발견의 논리 라카토스/우정호 38 텍스트 사회학 지마/허창운 39 현대물리학의 철학적 테두리 보음/전일동 40 과학과 가치관의 우선순위 스페리/이남표
41 신화의 진실 휘브너 / 이규영 42 대폭발 실크/홍승수 43 大同 警 / ;ft有 爲/ 이성애 44 표상 포더 / 이영옥 정성호 45 과정과 실재 화이트헤드 / 오영환 46 그리 스 국가 에렌버그/김진경 47 거대한 변환 폴 라니/박현수 48 법인류 학 포스피실/이문웅 49 언어철학 울스톤/ 곽강제 50 중세 이슬람 국가와 정부론 램톤 /김정 위 51 전통 쉴즈/감병서 신현순 52 몽골문어문법 뽀 떼/ 유원수 53 중국신화전설 l 원가/ 전인 초김선자 54 중국신화전설 l 원가/ 전인초 김선자 55 사회생물학 l 윌슨 / 이병훈 박시 룡 56 사회생물학 ll 윌슨 / 이병훈 박 시룡 57 일 반 언어학의 제문제 l 밴 베니 스트 /황경자 58 일 반언어학의 제문제 Il 밴베니 스트/ 황경자 59 폭력과 성스러움 지 라 르/김진식 박무호 60 갑골학 60 년 董 作賓 / 이형구 61 현 대수 학의 여행자 피터 슨/김인수 주형관 62 프랑스 혁명의 지적 기원 모르네/주명철 63 해석학과 과학 커널리 코이트 너 /이유선 64 서양고대경제 핀리/지동식 65 음악사회학 베버/이건용 66 앙띠 오이디푸스 들뢰즈·가따리/최명관 67 과학커뮤니케이션론 프라이스/남태우 정준민 68 교양교육의 개혁 다니엘 벨/송미섭 69 과학과 가치 래리 라우든/이유선 70 초기제국에 있어서의 교역과 시장 칼 푼라니/이종욱 71 폭정론과 저항권 헬라 만트/심재우 72 생명과학철학 데이비드 헐/하두봉 구혜영 73 사랑의 역사 쥴리아 크리스테바/김영 74 大地의 노모스 칼 슈미트/최재훈 75 일반 공법학 강의 레옹 뒤기/이광윤 76 텍스트학 반 다아크/정시호 77 문명의 발생 찰스 레드만/최몽룡 78 근대국가의 발전 G 폿지/ 박상섭 79 과학적 발견의 패턴 N R 핸슨/송진옹 조숙경 80 아랍 문학사 R A 니콜슨/사회만 81 1 8 세기 중국의 관료제도와 자연재해 P. E. 빌/정철웅
82 역사비교언어학개론 R. 안틸 라 / 박가 덕 남 성우 83 계몽주의 철학 E. 카 시라/ 박완규 84 토양 미생물학과 생화학 폴 클라크 /이 도원·조병철 85 수학, 과학 그리고 인식론 I 라카토시 / 이영애 86 봉건제의 이해 러쉬튼 쿨본/김 동순 87 그라마 톨로지 자크 데리다/김성도 88 殷代貞 卜人物通 考 1 라오쫑이 / 손예철 89 殷代貞 卜 人物 通考 ] 라오쫑이 / 손예철 90 殷代貞 卜人物 通考 빼 라 오쫑이 / 손예철 91 순수경제학 레 옹 왈라스 / 십상필 92 문화유물론 마빈 해리스 / 유명기 93 포유동물의 가축화 역사 줄리엣 클루톤브록/김 준민 94 르네상스 철학에서의 개채와 우주 E 카시러/박지형 95 화학적 진화 SF 메이슨/고문주 96 과학의 한계 바이츠제커/송병옥 97 러시아 연해주와 발해 역사 예 퀘 샤브꾸노프 / 송기호 • 정석배 98 경제정책의 원리 발터 오이켄/ 안병직 황신준 99 비코와 헤르더 이 사 야 벌린/ 이종홉 • 강성호 100 실용주 의의 결과 리처드 로티/김동식 101 국제 지층구분 지침서 A 살바도르/백인성 102 국가론 헤르만 헬러/홍성방 103 경험과 관단 후설/이종훈 104 17 세기 프랑스 정치사상 앙리 쎄/나정원 105 18 세기 프랑스 정치사상 〈근간〉 106 里鄕見聞錄 유재건/실시학사 고전문학 연구회 107 수학학습십리학 리처드 스캠프/황우형 108 고생태학 로버트 도드 로버트 스탠턴/양승영 109 지리학의 본질 l 리처드 하트손/김덕현 외 110 지리학의 본질 rr 리처드 하트손/김덕현 외 111 수학과 물리학 유리 마닌 1 명효철 채동호 112 과학의 합리성 뉴턴 스미스/양형진 조기숙