
데이비드 헐 (Dav i d L. Hull ) 미국 노스웨스턴 대학 철학과 교수 미국 과학철학회 회장, Socie ty of Sys t e m ati c Zoolo g y 회 장 역 임 저서 Darwi n and His Cris t i cs ( l97 3), Scie n ce as a pro cess (19 88) 古片 서울대학교 생물학과 졸업 일본 도오호구(東北) 대학원 이학박사 서 울 대학교 자연과학대학장. 부 총장 역임 현재 서울대학교 분자생물학과 교수 . 국가과학기술자문희의 위원. 광주과학기술원장 구혜영 이화여대 생물교육과 졸업 서 울 대학교 대학원 이학석사 미국 플로 리다 주립 대학교 이학박사 현재 상지대학교 생물학과 교수
생명과학철학
 Phil os op hy of Bio l og ica l Sc ien ce
Phil os op hy of Bio l og ica l Sc ien ce
 생명과학철학
생명과학철학
• 이 책은 Prenti ce -Hall Inc 에 의해 기획 • 출판된 〈 기초철학총서 Foundati on s of Phil oso p h y > 중 의 한 권인 Phil o sop h y of Bi ol og ic a l Sc i enc 례 번역한 것이다.
옮긴이의 말 나는 원래 동물생리학이라고 하는 실험생물학에 종사해 온 사 람으로서 철학에 관해서는 공부를 해 본 적이 없는 사람이다. 다 만 철학이 모든 학문의 기본이 된다는 등의 막연한 개념을 지니 고 있었고, 또 철학이라는 학문의 이름도 좋아서 그리고 철학은 어려운 학문이라고 듣고 있어서 그 학문에 대하여 일종의 존경심 비슷한 감정을 가지고 있을 뿐이었다. 그리고 역자가 하고 있는 것과 같은 실험생물학에도 철학이라는 것이 있는가, 또 나아가서 과학에 철학이라는 것이 있는가, 있다면 왜 그런 것이 필요한가, 자연과학은 자연현상을 있는 그대로 관찰하고 분석하고 해석하고 체계화하면 되는 것이지 철학이란 것이 왜 필요한가 등으로만 생 각하고 있었다. 미리 말하지만 역자의 이런 무식은 지금도 본질적 으로는 같다. 부끄럽지만 역자는 아직도 과학철학, 그 중에서도 생명과학의 철학에 대하여 그 존재 의의나 역할에 대하여 잘 이 해하고 있지 못한 것이다. 이러한 나에게 高麗大學校 金容駿 교수(현재는 정년 퇴임하셨 음)께서 하루는 이 데이비드 헐 David Hull 의 『생명과학철학 Ph i losop h y of Bio lo g ica l Sc i ence 』을 번역해 보라고 권유하셨다. 大宇
財團의 무슨 회의 끝에 역자에게 이렇게 권하시면서, 그분의 말씀 이 자연과학자는 모름지기 철학을 가져야 한다, 특히 생물학자에 게는 더 절실하다, 그러나 우리나라의 과학자들에게는 과학철학에 대한 관심이 없다, 몇몇 중견 과학철학자들이 우리나라에서 고군 분투하고 있으나 대학인들의 인식은 너무나 부족하다, 특히 생물 학 분야에서 이런 현상이 현저하다는 것이었다. 그리고 이 헐 Hull 의 책은 작아도 재미있어 보이니 꼭 번역해 보라고 하시면서 대우재단의 연구번역지원 과제로 선정하여 나에게 맡기신 것이다. 1984 년 초였다고 기억한다. 모르면 겁이 없다고 하는데, 철학에 대해서는 한줌만큼의 소양 도 없는 나는, 처음에는 물론 어려워서 못한다고 사양하였으나 고 작 15~ 구 정도의 작은 책, 그것도 영어로 된 책, 어려우면 얼마나 어려우랴 싶어 떠맡고 밀았다. 그러나 한 1 년쯤 지나서 일에 착수 해 보고는 곧 후회하기 시작하였다. 구구절절이 무슨 뜻인지 알 수가 없었다. 疾!이 가고 疾!이 가도 전도는 지지부전하였다. 그 렇게 하는 사이 몇 년이 또 지났으니 이제 와서 대우재단에 반납 할 수도 없게 되었다. 그러던 중 미국에서 具惠映 박사가 학위를 받고 귀국하여 나의 실험실에서 당분간 일을 같이 하게 되었다. 具 博士는 나와는 달리 철학에 대한 교양이 상당한 분이었다. 그 래서 공동으로 번역하기로 하고 원문의 1 차 번역을 具 博士가 사 실상 다하게 되었다. 그러나 나에게는 우리나라 말로 번역된 것도 이해하기 어려웠다. 그래서 좀더 쉬운 말로 고쳐보느라고 또 1 년 이상이 걸렸다. 이렇게 하여 겨우 이 번역본을 내놓게 되었으나 서두에 말한 바와 같이 나는 아직도 생명과학의 철학에 대하여 이해하고 있다고는 말할 수가 없다. 아직도 아쉬운 감이 적지 않으나 워낙 시일을 오래 끌었기에
6
이 책을 내놓기는 하지만 미비한 점들이 너무도 많을 줄 안다. 그러나 앞에서 언급한 바, 김용준 박사의 말씀처럼 생명과학의 철 학이 황무지인 우리나라 생물학계에 조그마한 철학적 관심을 불 러 일으키는 계기가 되었으면 하고 감히 인쇄에 부치기로 하였다. 번역상의 오류나 내용 표현 등에 관하여 관심 있는 분들의 지적 과 비판을 바라마지 않는다. 끝으로 이런 기회를 마련하여 주신 김용준 박사님과 대우재단의 관계자 여러분께 심심한 감사의 뜻 을 표하고, 또 學長이라는 補職울 맡아 무척 바쁘신 가운데도 이 원고를 끝까지 읽고 좋은 의견을 많이 보내주신 大邱大學校 分子
生物學科梁在涉 교수에게도 사의를 표한다. 그리고 草稿 단계에 서 試案을 만드는 데 많은 도움을 주신, 지금은 불란서에서 內分 認、 生理學울 전공중인 姜光一 博士에게도 감사를 드린다. 1994. 9. 관악산록에서 河斗鳳머리말 나는 이 책을 입문서로 하기 위하여 가능한 한 내용을 이해하 기 쉽고 또 독자 스스로 이해할 수 있도록 서술하였다. 그러나 내용이 너무 압축되어 있기 때문에 누구나 쉽게 접근할 수 있으 리라고는 생각하지 않는다. 대개의 독자는 생물학과 철학의 양쪽 기초를 다 지니고 있지는 않을 것이다. 사실 생물 철학을 강의해 본 사람이면 생물학 전공 학생에게 철학을 강의하거나 또는 철학 전공 학생에게 생물학을 가르친다는 것이 얼마나 어려운 일인가 롤 잘 알고 있을 것이다. 그래서 나는 이 책에서는 먼저 생물학 의 필수적 기초를 간단히 설명하고 이어서 점차 철학적 내용으로 들어가려고 한다. 또한 생물학과 철학의 술어들은 처음 나왔을 때 만 한 번 설명하고 그 이후는 생략하고 있다. 이 책을 이해하기 쉽게 그리고 유익하게 만들기 위하여 많은 생물학자와 철학자들이 원고를 읽어 주셨는데, 그 가운데서도 다 음의 분들에게는 특별히 감사를 드리고 싶다 : Morto n Beckner, Doroth y Grover, Helen Heis e , Ri ch ard Levin s , Ernst Mayr , R. E. Monro, Rosario Morales, Mi ch ael Ruse, Kenneth Scha f- fne r, Leig h Van Valen, Mary Wi lliam s, Wi lliam s Wi m satt , 그
리고 나의 철학과 생물학 강좌의 학생들, 또 본서의 편집과 출판 의 기회를 주신 E li zabe t h 와 Monroe Beardsley, 두 분께도 감사 를 드린다. 그리고 Dawn Klemme, Doroth y Wi ls on, Lin d a Eri- ch 는 타자와 원고 교정에 도움이 대단히 컸었다. 끝으로 이 책은 Nati on al Scie n ce Foundati on Grants GS-1971 과 GS-31 02, 그리 고 Un ive rsit y of W i scons i n-M i lwaukee 의 연구비 보조로 이루어 졌음을 명기하고자 한다. 데이비드 헐
생명과학철학
차례옮긴이의 말 • 5머리말 • 9서론과학철학은 하나뿐인가? • 15이론과 이론의 환원 • 18환원과 생물학 • 23제1장 멘델 유전학의 분자유전학으로의 환원환원과 과학혁명 • 27과학에 있어서의 역사적 재구성과 합리적 재구성 • 34고전적 멘델 유전학 • 36유전자형과 표현형의 관계 • 40구조, 기능, 위치 및 유전자의 개념 • 44우성과 열성 • 48멘델 유전학에 대한 도전 • 51멘델 유전학과 분자유전학의 구분 • 55
유전물질의 분자 구조 • 58생합성 경로 • 66유전학에 있어서의 이론의 환원 • 69환원함수-유전자 • 77환원함수와 서술용어 • 82멘델적 서술용어와 분자 기작 • 86유전학에 있어서의 환원-환원인가? 대체인가? • 92제2장 진화론의 구조과학적 이론, 법칙 및 정의 • 97다윈의 주장과 진화론의 구조 • 105도약 진화와 본질주의 • 108판게네시스 이론과 혼합유전 • 113단일 변이와 개체간의 차이 • 116진화의 유전 이론 • 120진화론의 통계적 특성과 그 근원 • 123창시자원리 • 130진화론의 연역적 성질 • 133자연선택과 적자생존 • 139제3장 생물학적 이론과 생물학적 법칙과학법칙과 서술상의 일반화 • 145인과법칙 • 149개체발생에 관련된 발생법칙 • 154계통과 교차연계법칙 • 157계통학에 있어서의 발생법칙 • 165역사법칙 • 169과학적 설명과 예측에 대한 포괄법칙 모델 • 178설명과 예측 사이의 대칭 • 183대칭명제에 대한 반론 • 187
고유성과 이야기식 설명 • 196제4장 목적론목적론의 문제점 • 205목적론의 본질 • 209목적론적 시스템을 위한 물리적 모델 • 213선호상태와 음성 피드백 • 217선호상태와 음성 피드백-반론들 • 221프로그램과 선택 과정 • 225목적론과 선택이론 • 229목적론적 서술과 포괄법칙 모델에 의한 설명 • 231목적론적 설명과 설명의 포괄법칙 모델 • 236요약 및 결론 • 244제5장 생체론과 환원론논쟁의 성격 • 249살아 있는 액체로서의 생명 • 253생기로서의 생명 • 255생명에 대한 기계론적 견해 • 257환원론과 구성론 • 261환원과 양자물리학 • 266복잡성과 고유성 • 270과학에서의 한계 조건의 역할 • 274결론 . 279참고문헌 • 281역자 해제 • 285찾아보기 • 295서론 과학철학은 하나뿐인가? 전통적으로 형식과학(수학, 논리학 등)과 경험과학(물리학, 생물 학, 심리학 등)은 구별되고 있다. 이 총서에서 논리학과 수학은 바커 S. F . Barker, 살몬 W. C. Salmon 그리고 과인 W. V. Qui ne 에 의해 세 권의 책에서 디루어지고 있으며, 경험과학 전반과 사회과 학 그리고 역사는 각각 행펠 C. G. Hemp el , 러드너 Ri ch ard Ru- dner 그리고 드레이 Wi lliam Dra y에 의해 역시 세 권의 책으로 다루어지고 있다. 후자의 책들에서 다루고 있는 주된 문제는 전통 적으로 경험과학을 지질학, 천문학, 사회학 등 독립된 분야로 나 누는 것이 그 연구 대상의 차이 때문인가 아니면 방법론적 차이 때문인가 하는 것이다 . 단적으로 밀하면, 하나의 과학철학은 모든 자연과학에 공통적으로 적용될 수 있는 것인가 아니면 각 분야마 다 제각기 독특한 과학철학이 존재하는가 하는· 문제이다. 헴펠은 그의 명쾌한 저서 속에서 과학철학은 하나뿐이며 이것 이 자연과학의 모든 분야에 공통적으로 적용될 수 있는 것이라고
하였는데, 이 주장은 자연과학의 철학적 원리로 현재 널리 받아들 여지고 있다. 과학적 탐구는 가설-연역적 방식 hy po th e ti co -de-ducti ve model 에 의해 이루어지는 것이 특칭이다. 행펠에 의하면, 수집한 여러 자료로부터 과학적 법칙들이 자동적으로 도출되는 경우에는 과학적 발견에 도달하기 위한 발견의 논리 log ic of dis- cover y라는 것이 존재하지 않는다. 과학자들은 가설을 여러 가지 방법으로 설정하는데, 한 방법아 다른 방법보다 더 생산적인 경우 가 물론 있기는 하나 대체로 방법들이 가지각색이고, 그 성공 여 부도 일정치 않기 때문에 어떤 방법이 특히 효과적이라고 일반적 으로 이야기할 수는 없다. 그러나 한 가지 분명한 사실은 어떤 과학적 주장의 전실 여부는 그 주장이 어떤 방법에 의거했는가 하는 문제와는 무관하다는 것이다. 어떤 특정한 방법도 전리를 보 증할 수 없는 동시에, 과학적 주장은 그것이 어떤 방법에 의해 도출되었건간에 전리일 가능성을 지니고 있는 것이다. 그러나 행펠은, 정당화의 논리 log ic of j us tifi ca ti on 리는 측면에 서는 상황이 매우 다르다고 주장한다. 과학자들이 그들의 가설을 검증하는 방법은 여러 가지이지만 그 과정에는 어떤 본질적인 공 통점이 있다. 이 본질을 연구하는 것이야말로 과학철학의 전정한 주제라고 할 수 있다. 가설-연역적 방식에 의한다면, 과학적 가설 은 무엇보다도 먼저, 반드시 검증 가능한 것이어야 한다. 그리고 이상적으로는 검증을 거칠 가설들이 보편성을 지니고 있어야 한 다(예를 들면 모든 기체는 열을 가하면 팽창한다 등등). 그래서 어 떤 현상 하나가 이 가설에 어긋나는 경우가 있더라도 많은 다른 현상들이, 비록 완벽하지는 않다 하더라도, 점진적으로 이 가설을 입증해 나갈 수 있어야 한다. 그러나 실제에 있어서는 가설들 자 체가 언제나 보편성을 지니고 있지도 않고, 또 가설들은 하나씩
따로 존재하는 것이 아니라 과학적 이론이라는- 큰 덩어리 속에 묻힌 상태로 존재하고 있다. 그래서 확증과 반증이라는 것 모두가 대단히 복잡한 문제이다. 과학을 이와 같이 볼 때, 가장 충실한 과학적 설명이라는 것은 〈 포괄법칙모델 cover i n g -law model 〉 을 따르는 것이다. 이 법칙에 의 하면 과학적 설명은 추론이다. 그래서 결국 어떤 현상의 상태를 설명한다는 것은 몇 가지 법칙과 주어전 조건들로부터 추론을 하 는 것이다• 이 추론은 연역적일 수도 있고 또 귀납적일 수도 있 다. 연역적 추론일 때 그 결론은 필연적으로 어떤 전제로부터 도 출된다. 반면에 귀납적 혹은 확률적 추론일 때는 그 결론이, 어떤 범위의 확률을 가전 형태로 전제로부터 도출된다. 따라서 포괄법 칙법에 의한 설명에는 두 가지가 있게 된다. 그 하나인 연역적 법 칙 적 설명 deducti ve -nomolog ica l ex p lana ti on 에서 의 추론은 연 역적이고, 또 다른 하나인 통계적-확률적 설명 sta t i sti ca l-p ro ba-bil ist i c ex p lana ti on 에서의 추론은 귀납적이고 확률론적이다. 따라 서 어떤 설명의 설득력은 귀납적인 방법에 의한 추론이 어느 정 도로 연역적 이상(理想)에 접근하고 있는가 하는 것에 달려 있다. 연역적-법칙적 방식에 관해서는 이미 많이 논술되어 왔다. 그런 데 모든 비연역적인 추론이 다 그렇지만 귀납적이라는 말의 불확 실한 정의에서도 알 수 있는 바와 같이 통계적-확률적 설명이라 는 것은 마치 제비뽑기와 같아서 반드시 들어맞는 것이 아니다. 그래서 여러 가지 비연역적 설명들 전부는 고사하고 그 가운데 어느 하나라도 제대로 설명할 수 있는 모델조차도 아직 없는 실 정이다.
이론과 이론의 환원 과학에 대한 논리적 경험주의자들의 개념에서 중심을 이루는 것은 과학적 이론을, 서로 관련된 여러 서술들로 이루어전 한 개 의 종합체로서 분석하는 일이다. 이 서술들 가운데 어떤 것은 기 본적 공리 basic ax i oms 라고도 할 수 있어서 그로부터 어떤 과학 적 법칙을 유도해 낼 수가 있다. 이렇게 유도된 법칙들은 우리 인간이 직접 관찰할 수 있는 자연 현상과 다시 복잡하게 연관되 어 있다. 과학 이론에 대한 증명은 이런 식으로 이루어진다. 또 과학 이론을 실존적으로 해석한다면(이 책이 그러하듯) 위의 공리 가운데 어떤 것은 우리가 관찰하지 못하는 존재, 죽 이론적 존재 를 의미하게 된다. 과학 이론의 실존적 해석에 의하면 이러한 이 론적 존재는 우리들의 일상적인 경험의 대상이 존재하는- 것과 같 은 의미의 존재로서 간주되며 과학법칙은 자연의 실제적인 규칙 성을 반영하는 것으로 생각되어야 한다(과학 이론의 실존적 해석 에 대한 옹호, Sim on, 1971 참조). 전통적으로 경험과학은 그 영역을 좁혀가면서 몇 개의 분야로 나누어전다. 물리학은 가장 넓은 영역을 가전 학문으로 간주되는 데, 그것은 이 학문이 이 세상에 존재하는 모든 물체의 물리적 성질을 다루는 것아며 모든 물체는 물리적 성질을 갖고 있기 때 문이다. 화학은 약간은 덜 기초적인 학문으로 간주되는데 이는 모 든 물질이 화학적 성질을 가지고 있기는 하지만 화학적 성질을 설명하려면 물리적 성질을 인용해야만 하기 때문이다. 그래서 가 장 기본적인 원리에 이르면 물리학과 화학은 결국 하나가 된다. 생물학은 물리학이나 화학보다는 한정된 영역을 갖는다고 생각되 는데 이는 물리적인 실체 중에서 단지 살아 있는 존재만을 디루·
기 때문이다. 다시 말하면, 모든 생명체는 물리적인 실체이지만, 모든 물리적 실체들이 다 살아 있는 존재는 아니기 때문이다. 심 리학은 더욱더 제한된 영역을 가지고 있다. 왜냐하면 생명체 중에 서도 감각을 지닌 존재만을 다루기 때문이다. 사회학은 더욱 좁은 영역의 학문으로서 감각을 지닌 생물 중에서도 사회라는 조직을 구성하는 존재만을 다루고 있다 . 철학자들과 과학자들은 환원(還元, reduc ti on) 이라는 단어의 뜻 을 다양하게 사용해 왔다. 과학 이론에 대해 분석하고 과학의 주 체를 체계적으로 나누어보면 환원에 대한 세 가지 의미가 분명히 드러나는데 그것은 인식론적 환원, 물리적 환원 그리고 이론적 환 원이다. 인식론적 환원은 과학 이론과 우리가 가진 지식의 대상 사이의 관계를 다루고 있다. 물리적 환원은 여러 과학적 이론들이 입각하고 있는 기본적인 존재들 사이의 관계를 디루·고 있다. 이론 적 환원은 과학 이론 그 자체들 사이의 관계를 다루고 있다. 인식론적 환원주의에 있어서는 과학 이론에서 이론적 - 존재를 대상으로 삼는 것을 배제하기 때문에 과학 이론은 우리 지식의 객관적 대상만을 디루도록 다시 만들어져야 한다고 한다. 물론 인 식론적 환원론자들 사이에서도 이 대상에 대해 의견이 서로 일치 되지 않는 면이 있다. 그래서 어떤 사람은 모든 과학적인 서술들 은 물리적 대상의 용어로 다시 만들어져야 한다는 것이다. 자〔尺〕 나 검류계 같은 기구들을 사용함으로써 말이다 . 그 반면 또 다른 사람은 모든 인식론적 지식은 감각의 경험에서 온다는· 것이고 따 라서 경험적 지식은 〈지금 빨간색 헝깊조각 red pa tc h now 〉과 같 이 감각적인 자료를 사용해서 모든 서술들을 새로 고쳐야 한다고 한다. 경험적 환원의 매력적인 힘은 모든 경험적 지식이 감각적인 경험에 의한다는 경험주의자들의 주장으로부터 나오는 것이다. 따
라서 경험적 지식은 감각적인 것으로 환원되어야 한다는· 것이다. 그러나 실제로는 이들 인식론적 환원에 대한 어느 설명도 별로 성공하지 못했을 뿐 아니라 인식론적 환원에 의해 제시된 문제들 어느 것도 생물학과 별다른 연관성을 갖지 못했다. 그래서 이러한 의미의 환원은 앞으로 전개될 내용에서는 일단 무시하기로 한다. 물리적 환원에서는, 일정 수준에 있는 어떤 계를 그 구성 성분 으로 분석하고, 또 그 계의 행동을 구성 성분들의 성질, 행동, 배 열 등으로 설명한다. 철학자들의 저서에서 쉽게 찾아볼 수 있는 환원의 한 전형적인 예는 기체의 성질(온도처럼)을 기체를 구성 하고 있는 분자들의 브라운 운동이라는 말로 설명하는 것이다. 마 찬가지로, 분자유전학자들은 유전자의 행동을 분자라는 용어로 설 명하려고 한다· 이론을 환원하는 데 있어서, 한 이론의 원리는 다 른 이론의 원리로부터 도출된 일반적인 법칙에서 유래하기 때문 에 새로 창출된 이론은 원래의 이론으로 환원될 수 있다는 것이 다. 그러한 예로, 전통적인 열역학 법칙이, 기체를 구성하고 있 는 분자들의 평균에너지와 기체의 온도를 같다고 간주함으로써 통계 역학으로부터 유도된 것을 둘 수 있다. 과학의 주체가 되는 물질들에 위와 같이 높고 낮은 순서가 있 다면, 물리적 환원과 이론적 환원의 결과는 일치하게 된다. 관련 된 이론의 범위뿐만 아니라 물리적인 분석이 이루어지는 단계들 모두에 있어서 물리학은 그 기본이 된다. 물리학은 가장 고도로 조직화된 존재로부터 가장 단순한 소립자에 이르기까지 모든 체 제의 물리적인 특성을 다루는 반면에, 예를 들면, 생물학은 고도 로 조직화된 존재의 속성만을 다루는 것이다. 과학적 이론은 우주 에서부터 생물에 이르기까지 또 소립자에 이르기까지 모든 단계 에서 만들어진다. 두 이론이 같은 단계에 있는 현상을 다루고 과
학의 같은 분야에 속해 있다면 그 두 이론의 환원은 단계내 i n t ralevel 의 문제라 할 수 있고, 그렇지 않은 경우는 단계간 i n t erlevel 의 문제라고 할 수 있다. 따라서 열역학 법칙을 통계 역 학으로 환원시키는 것은 양쪽 이론이 모두 물리적 아론이라는 의 미에서 단계내의 문제이지만, 환원하는- 이론이 환원되는 이론보다 낮은 단계의 현상을 다루고 있다는 면에서 보면 단계간의 문제가 된다. 그러나 환원에 대한 이러한 분석은 도전을 받게 되었다. 예를 들어 심프슨 (G. G. Sim p so n, 1964) 은 모든 과학의 중심이 물리학 이 아니라 생물학이라고 주장하였다. 왜냐하면 모든 중요한 가정 과 그 설명 원리들은 모두 생명체에 적용되는 것이고 무생물에 적용되는 것은 극히 적기 때문이라고 하였다. 과학의 여러 분야들 은 모든 현상에 적용되는 원리를 통해서가 아니라 모든 원리가 적용되는 현상을 통해서 훨씬 더 잘 조직될 수 있다는 것이다. 그래서 하나가 다른 하나로 환원되어 간다면 그것은 물리학이 생 물학으로 환원되는 것이라고 한다. 만일 심프슨이 그의 주장을 논 리적 인 결론으로 잘 이끌어갔다면, 마이클 사이몬 (M i chael Si- mon, 1971) 이 말했듯이 생물학이 아니라 사회과학이 모든 과학의 중심에 있어야 할 것이다. 왜냐하면, 생물학적인 현상과 그 현상 울 설명하는 원리들이 모두 인간에게 적용될 것이고 아주 작은 숫자만이 인간이 아닌 다른 생명체들에 적용된다는 똑같은 논리 롤 주장할 수 있기 때문이다. 따라서 만일 어느 하나가 다른 하 나로 환원된다면 그것은 물리학과 생물학이 사회과학으로 환원되 어야할 것이다. 그러나 심프슨의 주장과 사이몬의 확대 해석이 가전 설득력은 그들이 세운 전제들이 애초부터 잘못되어 있었기 때문에 손싱을`
입을 수밖에 없다. 물질의 모든 현상들이 생명체를 포함한 모든 물리적 존재에 적용된다는 것은 사실이 아니다. 예를 들면, 어떤 물리적 존재들은 전류를 방출할 수 있고 어떤 것들은 방출할 수 없다. 그리고 어떤 것은 자기력을 가지고 있고 어떤 것은 그렇지 않다. 그러나 어떤 경우에는 생명체들도 전류를- 방출할 수 있지만 필자가 아는 바로는 생명체가 나타내는 자기의 효과는 전자석과 는 다르다. 심프슨의 주장에서 옳은 것은 인간이 어떤 생물학적인 법칙들에도 모순되지 않고 잘 부합되며 어떤 생명체이건 간에 물 리적인 법칙에 어긋나는 경우가 없다는 것뿐이다. 그러나 심프슨 과 사이몬의 결론 어느 것도 이러한 전제로부터 자연스럽게 도출 되지는 않는다. 어떤 것이 과연 과학의 중심이 되는가롤 결정하기 위해서는, 자연 현상을 무작위적으로 뽑아보고 어떤 과학 분야가 여러 현상들과 그 현성을 설명하는 원리들을 더 많이 소유하고 있는가를 살펴보아야 한다. 심프슨의 잘못은 자연 현상에 대한 종 래의 학문체계에 도전하는 데 있어 충분히 깊이 둘어가 보지도 않고 그 나름대로 체계를 세웠다는 데 있다. 그 중심에다 인간을 놓고 이렇게 정한 체계는 체계 그 자체가 의심스럽지 않을 수 없 는 것이다. 자석도 어떤 특별한 성질을 중심으로 한다면 단순한 자석이 아닐 수도 있다. 인간 중심적인 체계로 과학을 다루는 데 대한 끊임없는 회의에도 불구하고, 이 책에서는 목표하는 바를 설 명하기 위해 이런 인간 중심적인 체계를 일단 수용하기로 한다. 모든 것을 한꺼번에 문제시할 수는 없기 때문이다.
환원과 생물학 리차드 러드너 Ri ch ard Rudner 는 사회과학에 대한 그의 저서 에서 자연과학에 대한 견해를 사회과학으로 확대하는 일에 대해 논하고 있다. 그는 사회과학은, 어떤 사실을 발견하기 위해 혹은 정당화하기 위해 사용하는 기술은 자연과학과 다르지만 그 둘은 다 같이 정당화의 논리를 사용하고, 과학철학에 관한 한 정당화 가 가장 중요한 문제라고 한다. 이와 같은 러드너의 설이 헴펠의 것과 가장 다른 점은, 사회 이론의 성질을 밝히기 위해서 전체적 으로 또는 부분적으로 형식화된 이론을 사용하는 점이다. 그러나 불행히도 사회과학에는 명백하게 형식화된 과학 이론이 별로 없 기 때문에 이 기술의 사용이 물리학에서보다 훨씬 덜 효과적일 수밖에 없다. 한편, 윌리엄 드레이 Wi lli a m Dra y는 행펠-러드너 식의 과학 분석을 역사의 문제로 확대하는 것에 대해 반대한다. 역사는 그들 이 말하는 의미의 〈과학〉이 아니기 때문이다. 역사학자들은 사회 적 의미를 지닌 인간의 어떤 행동뿐 아니라 그 행동 자체를 이해 하는 일에도 관심을 둔다. 역사학자들의 설명과 이해는 다론 분 야, 죽 물리학과 같이 오차가 없고 보다 엄격한 과학 분야의 것 과는 다르다. 다른 철학자나 과학자들도 사회과학에 대한 비슷한 견해를 피력하였다. 물론 이런 견해가 지닌 설득력은 이러한 특별 한 의미의 〈설명〉과 〈이해〉를 얼마나 적철히 분석하는가에 달려 있다. 그런데 나는 아직 이 개념들을 분명하게 이해하지 못하고 있다는 것을 인정해야만 하겠다. 이 책이 목표하는 바는 물리학이 사회과학으로 급속히 확산되 는 과정에서 간과된 분야들을 좀더 자세히 보고자하는 것이다. 과
학이 처음 발달하기 시작했던 시기에는, 일반적인 생물학적 현상 둘-특히 인간의 행동-울 설명하는 데 가장 적합한 양식이 모든 자연 현상에도 적용될 수 있을 것으로 생각되었다. 인간이 고결해지려고 노력하는 것처럼, 생물의 종 s p ec i es 들이 자신의 종 을 번식시키려고 노력하는 것처럼, 바위가 땅으로 떨어지는 것도 바위가 자연 내의 자신의 위치를 찾으려고 노력하는· 것으로 해석 된 것이었다. 그러나 갈릴레오와 뉴턴 이후에는 점차적으로 정반 대의 견해가 풍미하기 시작하였다. 그리하여 순전히 물리적인 여 러 현상을 연구하는 데서 비롯된 설명 양식이 생물학 그리고 사 회학적인 현상에까지 확대되었다. 그래서 모든 현상은 사슬처럼 또 그물처럼 서로 인과 관계로 연결된 과거의 결과이고, 다가올· 미래나 또는 한차원 높은 단계의 현상은 이 인과론으로는 예측하 지 못하는 것이라고 해석되고 있다. 우리는 이 책에서, 생물학적인 이론과 설명 양식이 과학의 본 질에 대한, 그리고 생물학과 나머지 다른 과학 분소戶陸] 관계에 대한 여러 논쟁에 어떤 해결점을 줄 수 있는가를 살펴보게 될 것 이다. 전통적인 방식으로 과학을 분류할 때나 이론적인 발달 과정 으로 볼 때나, 생물학은 물리학과 사회과학 사이에 놓여 있기 때 문에 이러한 목적에 아주 적합한 분야이다. 과거에는 너무 자주 과학철학의 여러 문제들이 순전히 과학적인 추상이나 아니면 순 전히 물리적 이론의 관점에서 다루어져 왔다. 앞으로 전개될 토론 은 종래의 이런 두 방법과는 전혀 다를 것이다. 우리는 특정한 이론들을 다루게 되는데 이 이론들은 물리학이 아닌 생물학으로 부터 유도될 것이다. 생물학에는 극히 분명한 이론들이 많이 있기 때문에 우리의 설명이 단지 무엇이 원칙적으로 가능할 것인가 아 닌가를 따지는 데 머물러 있을 필요는 없고, 실질적 인 문제들을
다루게 될 것이다. 제 1 장에서 우리는 유전학에서 일어난 변화, 즉 멘델 유전학이 분자유전학으로 환원된 분명한 사실을 논의하게 될 것인데, 이러 한 사실이 입증될 수 있다면 한 단계의 과학에서 다른 단계의 것 으로 환원되는 일은 가능한 것일 뿐 아니라 실제적인 일이 될 것 이다. 순수한 생물학적 이론이 물리 화학적인 이론으로 환원되는 것이기 때문이다. 단계간 환원의 이러한 분명한 예를 증명하거나 반증하는 좋은 예를 찾기 위해서 양쪽 이론의 윤곽을 대충 살펴 볼 필요가 있다. 이들 이론은 그 자체만으로도 홍미로운 것이지만 이 장의 목표는 환원론자들의 의도가 성공하려면 얼마나 많은 일 들이 다루어져야 하는가를 보여 주는 것이다. 제 2 장에서는 우리의 관심을 전화의 이론으로 돌리게 할 것이다. 이것은 환원의 경우처럼 문제가 간단하지 않다. 지금의 상황이 그 러한 질문이 제기될 수 있을 만큼 충분히 발달되어 있다고 볼 수 없기 때문이다. 대신 진화적 이론 그 자체의 타당성을 다루게 될 것이다. 또 제 2 장에서 우리는 전화론의 여러 형식들을 자세히 설명하고 그들의 구조를 서로 비교해 볼 것이다. 전화론이 아 직도 계속 변화하는 유동적인 상태에 있기 때문에 제 2 장에서 도 달되는 결론은 필연적으로 참정적인 것일 수밖에 없다. 실제로 어 떤 사람들은 전화론뿐 아니라 모든 생물학적 이론들이 의심스 럽다고도 하고, 생물학에는 과학적 이론이나 과학적 법칙이 존재 하지 않는다고까지 주장하고 있는 것이다. 제寫써서, 우리는 이러한 반대 의견들을 분명히 명시하고 그 근거를 발견하고자 노력하게 될 것이다. 제炫授든 목적론이 지니고 있는 해묵은 문제에 할애된다. 순수한 형이상학적 이론으로서의 목적론은 오랜 세월 동안 과학을 위해
아무런 역할도 하지 않았다. 하지만 아직도 생물학자들은 목적론 적인 언어를 계속 사용하고 있다. 문제는 아무런 손실 없이 생물 학으로부터 이들 언어들을 제거할 수 있겠는가 하는 것이다. 그것 이 불가능하다면, 이는 환원론자들의 의도가 성공할 수 없는 또 하나의 이유가 될 것이다. 이 책을 쓰는 데 있어 나의 근본적인 관심은 모든 과학에 부합 되는 한 개의 철학이 있을 것인가 아니면 그 각각에 알맞는 여러 개의 철학이 존재하는가를 연구하는 데 있다. 하지만 나 자신이 이 물음 자체를 이 책에서 손대지는 못하였다는 것을 인정해야만 하겠다. 대신 나는 이 관심사에 대한 결론을 내리기 전에, 먼저 결정되어야 할 구체적인 문제들을 다루고 있다. 그러나 이 커다란 질문에 대해 명확한 결론을 내릴 수 있을 만큼 이런 구체적인 문 제들이 충분히 해결되고 있다고는 생각하지 않는다. 이런 문제들 에는 나 자신의 편견도 섞여 있을 것이다. 이 책 전체를 통해서 나는, 있을 법한 반대 의견들에 지나치게 연연하지 않고 다양한 문제들에 대한 나의 견해를 피력할 것이지만, 지면이 부족한 것을 또한 변명해야만 하겠다. 설명은 지나치리만큼 압축되어 있는데도 말이다. 나의 이런 입장을 환원론-생체론 reduc ti o ni sm-or g a ni c i sm 의 연속선 상에 두고서 나 자신을 환원주의자나 생체론자로 호칭하고자 하는 것은 아니다. 이 책에서 내가 과연 환원주의나 생체론을 옹호하기 위해 논쟁하고 있는가? 대답은 둘 다 아닐 수 도 있고 둘 다 맞을 수도 있다. 내가 환원론-생체론의 논쟁에 약 간의 기여를 했다면 그것은 제沼}에서 찾을 수 있을 것이다.
잭 멘델 유전학의 분자유전학으로의 환원 환원과 과학혁명 과학의 발달과정이 항상 순조로운 것은 아니다. 때로는 극적인 변화가 일어나기도 하고 때로는 어떤 특정 분야와 관련해서 극히 점진적인 변화가 일어나기도 한다. 토마스 쿤 Thomas Kuhn1) 은 전자를 혁명적 과학 revolu ti onar y sc i ence 이라 했고 후자를 정상 적 과학 normal sc i ence 이라고 하였다. 이러한 분류 방법과 밀접 하게 관련된 또 하나의 분류 방법이 있는데 그것은 대체 (代替, re placemen t)와 환원(還元, reduc ti on) 이다. 일반적으로 과학혁명 기에는 그때까지 보편화되어 있던 하나의 과학 이론이 다른 것에 의해 대체된다. 이러한 변화는 과학이 발달하기 시작한 초기에, 창조론과 같은 초자연적인 이론이 다윈의 전화론과 같은 자연주 의적인 것으로 대체됨으로써 특히 두드러졌다. 정상적 과학의 시
1 ) T. Kuhn, The Str u ctu re of Scie n ti fic Reuoluti on , 2nd ed. (Ch ica go : Univ e rsit y of Chic a g o Press, 1970). * 이 책에서는 원주를 1), 2), 3) 등으로, 역주를 ®, ®, ® 등으로 표 시하였다.
기에는 변화의 폭이 대단히 작다. 이 경우 과학자들은 이론을 가 능한 한 이해하기 쉽게 만들기 위해, 어떤 때는 종전의 독립된 법칙을 더 일반적인 법칙 속에 포함시키기도 하고 어떤 때는 덜 포괄적인 것을 디론 것에다 갖다 붙이기도 한다. 과거에는, 이와 같은 〈 환원 〉 이 항상 전통적으로 정의된 한 과 학 분야 내에서만 일어났다. 그러나 지난 이십 년 사이에 꼭 환 원처럼 보이는 어떤 것이 유전학에서 일어났는데, 이 경우에는 환 원과 관련된 두 개의 이론이 각각 아주 상이한 과학 분야에 속하 는 것이었다. 죽 생물학자들은 생물학적인 이론인 멘델 유전학을 물리-화학적인 용어로 된 이론인 분자유전학으로 환원하고 있었 다. 이 변화는 대체가 아니라 단지 환원일 뿐인데도 불구하고 과 학적인 혁명이라고 할 만큼 많은 사람들에게 중요한 변화로 인식 되고 있었다. 혁명적 또는 정상적 과학에 있어서나 대체와 환원에 있어서나, 구별이란 것은 그리 명확하지가 않다. 과학혁명의 기간이 정상적 기간에 비하여 대체로 짧기도 하지만, 과학의 역사 속에 일어났던 실제 사건들을 분류하고자 하면 결국에는 모두가 연속적아었던 일로 귀착이 되고 마는 것이다. 물론, 과학 역사상의 몇몇 사건들 은 분명히 혁명적인 것이었다. 그 예로 다윈의 전화론의 등장이나 멘델 법칙의 발견 등을 들 수 있다. 반면, 전화에 있어서 의태(擬 態 mimi cr y)의 역할이나 반성 유전(伴性遺傳)의 발견 같은 것은 분명히 정상적 과학의 범주에 속하는 부분이다. 그러나 세포설과 염색체설의 체계화 또는 종간 교배의 발견 같은 것은 어떤가? 혁 명적인 것으로부터 정상적인 것에 이르는 연속선 상에서 어느 한 쪽 끝에 놓이는 것들도 더러 있겠지만, 과학사에서 일어난 대부분 의 사건들은 그 중간 어디쯤에 속하는 것이었다.
대체와 환원이라는 분류에서도 비슷한 양상을 볼 수 있다. 뉴 턴 물리학이 아리스토텔레스의 물리학을 대체한 것이나, 라부아지 에의 산화 이론이 산소 발견 전의 연소(燃~燒) 이론을- 대체한 것 이나, 전화론이 창조론지들의 교리를 대체한 것 등은 의심의 여 지가 없는 대체의 예라고 할 수 있다. 마찬가지로 어떤 시기에는 그야말로 분명한 환원의 예들도 과학철학에서 쉽게 발견되는데, 그 예로 케플러의 행성의 운동 법칙과 갈릴레오의 자유 낙하 법 칙이 뉴턴의 역학과 만유 인력의 법칙으로 환원되었던 것을 둘 수 있다. 이 경우의 환원은 동질적 이론들의 환원으로 간주되었는 데, 그것은 그 이론들이 대체로 같은 종류의 현상을 다루코. 있고 또 같은 용어를 사용했기 때문이다. 그러나 분명히 다론 현상을 디루는, 각기 서로 다른 서술 용어들을 사용하는 두 이론 사이의 환원도 혼히 볼 수 있다. 그러한 이질적 환원의 고전적인 예로서 열역학이 통계역학으로 환원된 것을 들 수 있다. 애초에 이 두 이론은 각각 독자적으로 발달되어 왔고, 또 그 대상도 서로 전혀 다른 것이었다. 열역학은 기체와 같이 거시적인 대상의 행동이라 는 용어로 설명되었던 반면, 통계역학은 미시적인 대상의 용어로 설명되었다. 그러나 이상기체(理想氣體)의 온도라는 개념이 기체 분자의 평균 운동 에너지라는 역학적 개념과 동일시되면서, 열역 학의 법칙을 비롯하여 그와 비슷한 여러 이론들이 통계역학의 기 본적인 원리로부터 도출될 수 있었다. 하지만 대체와 환원 사이의 위와 같은 차이점을 받아들인다 하 더라도, 우리는 대체 또는 환원 어느 쪽이라고 딱 잘라 말할 수 없는 경우, 즉 그 경계선 상에 놓인 수많은 경우들이 있다는 것 울 알아야 한다. 더욱이 최근에는 여러 과학철학자들이 구별 그 자체를 의문시하고 있다. 그들은 〈환원〉이라는 말을 매우 엄격하
게 정의하고, 과학의 역사에서 실제로 이러한 정의를 충족시킬 만한 사건은 없었다고 주장한다. 대체-환원의 연속선 위에서 환 원의 끝쪽에 위치한 것, 죽 국단적인 환원의 경우는 없었다는 것 이다. 이러한 결론은, 때로는 또 다른 구별, 죽 이론적 용어와 관 찰적 용어라는 구분에 그 바탕을 두고 있는 경우도 있다. 이론적 인 용어란 과학 이론 속에 있는 서술용어이고, 그 과학 이론의 의미 대부분은 이들 서술용어로부터 생겨나는 것이다. 이러한 이 론적 용어들을 이론들로부터 떼어내면 아무 의미도 없어지거나 거의 무의미한 용어가 된다. 또한 이론을 변화시키면 그 용어의 의미도 크게 변화된다. 〈전자〉, 〈질량〉, 〈 유전자 〉 , 〈 종(種, spe - c i es) 〉과 같은 단어들은 이론적인 용어의 몇몇 예라고 할 수 있 다. 관찰에 의한 용어란, 인류의 공유재산이라고 할 수 있는 서술 적인 단어, 죽 사물을 보고 묘사하는 단어들이다. 과학이 독립된 학문 분야로 등장하기 훨씬 전부터 사람들은 〈빨간색〉이나 〈 임 〉 이라는 단어가 무엇을 의미하는지 이미 알고 있었다. 이러한 용 어 사이의 차이점을 설명하기 위해 사용되는 말은 어휘 그 자체 가 잘못된 것이다. 중요한 차이점은 관찰에 의한 것과 관찰에 의 하지 않은 용어들 사이에 있는 것이 아니라, 이론적으로 기울어 져 있는 용어와 이론적으로 중립적인 용어 사이에 있는 것이다. 애초에 이론적 용어는 보통 관찰될 수 없는 존재나 과정을 지칭 했지만 경우에 따라서는 관찰이 얼마간 가능한 경우도 있었다. 그러나 관찰 가능한 것을 지칭했다고 해서 그것이 덜 이론적인 것으로 변하는 것은 아니다• 쿤 T.K uhn, 핸슨 N. R. Hanson, 그리고 파이어아벤트 P. K. Fe y erabend 는, 위에서 언급한 두 가지 경우와 마찬가지로, 이론적인 것과 비이론적인 것의 구분은 그다지 명백한 것이 아니라고 주장
하고 있다 ? 과학의 서술용어들은, 어떤 특정 과학 이론과 밀접
2) T. Ku hn, The Str ucture of &ient i fic Reuolu tion , 2nd ed. (Ch ica go : Univ e rsit y of Ch ica go Press, 1970) ; N. R. Hanson, Patt er ns of Di sc ouery ( Cambrid g e : Cambrid g e Un ive rsit y Press, 1958) ; P. K. Fey e rabend, Ex pla nati on , Reducti on , and Emp iri c i s m , in Mi nn esota Stu d ie s in the Phil o sop h y of Scie n ce, Vol. III (Mi nn eap o li s : Un ive rsit y of Mi nn esot a Press, 1962).
하게 연관된 용어로부터 이론적으로 중립적인 용어에 이르는 한 개의 연속된 직선 위에 하나하나씩 배열될 수 있는 것이다. 그러 나 모든 이론에 관해서 완전히 중립적인 용어란 있을 수 없다고 그들은 주장한다. 서술용어들은 모두 어느 정도의 이론을 내포하 고 있다는 것이다. 오늘날에는, 남자와 여자는 같은 종에 속한다 는 주장은 하나의 상식으로서 전혀 문제가 될 수 없다고 여겨질 테지만, 그 주장은, 〈 남자 〉 와 〈 여자 〉 라는 말은 말할 것도 없고, 〈 종 〉 이라는 이론적으로 정의된 개념을 전제로 하고 있다. 이 경 우는, 관련된 이론이란 것이 언급하기에 너무 평범한 것일 뿐이 다 . 그러나 평범하다고 해서 덜 중요한 이론은 아니다. 과학의 서술적 용어에 대한 이러한 견해는 엄밀한 의미에서 환원이란 불가능한 것이라는 결론을 가져 온다. 두 이론이 서로 비교될 수 없다는 단순한 이유 때문에 갈릴레오의 법칙은 뉴턴의 법칙에 환원될 수 없으며, 뉴턴의 법칙에서 도출될 수 있는 것은 갈릴레 오의 법칙과 아주 근접한 공식일 뿐 갈릴레오의 법칙 그 자체는 아니다. 뉴턴과 아인슈타인은 그들의 이론에서 모두 〈질량〉이라 는 단어를 사용했지만 이 단어가 같은 것을 의미하고 있지는 않 다. 과학 용어에 대한 이러한 견해는 결과적으로 동질적 환원과 이질적 환원 사이의 구별을 사라지게 했으며, 따라서 모든 것은 이질적인 것이 되었다. 두 이론이 같은 용어를 사용하고 있을 때
도 마찬가지이다. 이제 다음 단계는 대체와 환원의 구별을 없애는 것이다. 실제 로 과학에 있어서의 모든 변화란 대체에 해당하며, 엄격한 의미에 서 환원이란 불가능한 일이기 때문이다. 이런 식의 추론이 매력적인 듯이 보이지만, 실제로 〈 대체 〉 그 자체의 개념에 있어서는 극복할 수 없는 어려움이 따른다. 만약 원래부터 비합리적인 것이 아니었다면, 한 과학 이론을 버리고 다 론 과학 이론을 선택하기 위해서는 적어도 현재 공식화되어 있는 이론을 훨씬 능가하는 추론적 도약을 필요로 할 것이다. 한 이론 이 경쟁 관계에 있는 이론보다 더 포괄적이고, 더 정확하게 예측 할 수 있고, 더 간단할 때 그 이론이 선택된다는 것은 당연한 일 이다. 예를 들어, 천문학에 있어 코페르니쿠스의 체계는 아마 세 가지 점 모두에 있어 프톨레마이오스후] 것보다 우수하다고 보아 야 할 것이다. 그러나 이것은 우리가 바라는 것처럼 그렇게 간단 하지가 않다. 양쪽 이론이 다같이 행성 운동을 상상적인 어떤 원 의 주변을 돌고 있는 또 다른 상상적 인 원, 다시 말해서 주전원 (周轉圓 e pi c y cles) 으로 설명한다. 다만 두 이론의 가장 큰 차이 점은 프톨레마이오스는 지구를 중심에 놓은 것이고 코페르니쿠스 는 태양을 그 중심에 놓은 것이다. 태양을 중심에 놓음으로써, 코 페르니쿠스는 더 적은 수의 주전원을 사용하면서도 프톨레마이오 스와 똑같은 정확성을 가지고 예측을 할 수가 있었다. 그러나 이 경우에 단순성의 증대는 그리 대단한 것이 아니었고, 코페르니쿠 스의 가장 뛰어난 점은 역시 태양이 중심에 있고 지구는 그저 또 하나의 행성일 뿐이리는 일관되고 설득력 있는 물리 체계에 있었 다. 예를 들어, 의행성 (外行星) 운동과 내행성 운동의 차이점은 코페르니쿠스 천문학으로부터는 자연스럽게 유도될 수 있었던 데
비하여, 프톨레마이오스주의자들은 약간의 억지 이론을 사용해서 만 설명이 가능했던 것이다. 그러나 만일 두 이론의 모든 서술용어들, 즉 가장 이론적인 것 으로부터 가장 관찰적 용어에 이르기까지 모두가 이론을 지니고 있다면, 어떻게 그들을 서로 비교할 수 있고 어떻게 하나가 다른 하나에 비해 낫다고 할 수 있을까? 코페르니쿠스 천문학에 따르 면 수성과 금성이 지구 바깥쪽에 있는 행성들과 다르게 행동하는 것이 너무나 당연하지만, 프톨레마이오스 천문학에 의하면 그러한 차이점을 예측할 아무런 근거가 없다 . 일단 그러한 차이점이 발견 되었다면 거기에 맞춰 이론에 수정을 가할 수 있을지는 모르지만. 그러나 이 토론에서 논리적인 결론을 얻어야 한다면, 그것은 우리 가 〈 같은 〉 현상을 비교하고 있는 것이 아니라는 것이다. 프톨레 마이오스의 금성은 코페르니쿠스의 금성과 갇은 것이 아니다. 그 러므로 하나의 이론을 버리고 다른 하나를 선택해야 할 아무런 이유가 없다 ; 과학적인 발전이라고 일컬어져 왔던 것들도 실제로 는 그렇게 완벽하게 합리적인 것만은 아니었다. 이론 속의 모든 서술용어들이 이론을- 담고 있고, 환원이 정말 엄격하게 해석된다 면, 과학에 있어서의 모든 변화는, 정도의 차이는 있겠지만, 〈대 체〉에 해당되며, 하나의 이론을 택하고 다른· 하나를 버리는 것은 현재 우리가 이해하고 있는 개념으로서의 증거나 이유가 있어서 가 아니다 . 증거와 이유에 대한 더욱 정교한 개념이 정립되어 실 제로 과학 내에서 통용될는지 모르지만 아직은 그 개념이 명백하 지 않다.
과학에 있어서의 역사적 재구성과 합리적 재구성 앞에서 언급한 논쟁은 사람들이 환원에 대해 서로 다른 견해를 가졌기 때문이라기보다는 과학철학의 속성에 대해 서로 다론 견 해를 가졌기 때문에 생긴 것이라고 볼 수 있다. 한쪽 견해는, 과 학 이론이라는 것이 어떤 특정한 시대에 만들어진 것인 만큼 환 원을 과학 이론과 관련된 역사적 과정으로 보고 있다. 이러한 견 해에 의하면, 환원이란 서로 일치도 되지 않고 특별한 것들로 가 득찬 복잡하고 불확실한 작업이다. 이야기하는 내용은 자세하고 도움이 될 만한 것이 많지만 문제는 각각의 경우가 늘 유일무이 하다는 데 있다. 또 한쪽 견해는 환원을, 역사적으로 정립된 다음 추출된, 그리고 재구성된 두 과학 이론들 사이의 연관성을 연구하 는 것이라고 보고 있다. 이때 두 이론들이 처음과 똑같은 상태여 야 할 필요는 없으며 오히려 교과서적인 설명 방법에 의해 더 이 상적인 것이 될 수가 있다. 그러나 그렇게 하면 물론 논리적으로 는 더 단순해 질 수 있겠지만 동시에 역사적인 정확성이 희생될 것이다. 과학 이론 속에 있는 모든 서술적 용어들이 이론을· 담고 있겠지만 어떤 용어는 분명히 다른 용어에 비하여 덜 이론적일 수가 있다. 그러한 용어들을 가려냄으로써, 잘못될 가능성을 최소 화하면서 동시에 서로 다른 과학 이론들을 비교해 볼 수 있을 것 이다. 엄격한 의미에서는 아마 과학에 있어서의 어떤 변화도 환원 으로 간주될 수는 없을 것이다. 하지만 어떤 변화들은 다른 경우 에 비해 환원적 이상에 더 가까이 접근할 수도 있을 것이다. 널 리 알려진 한 이론이 다른 이론으로 바뀔 때, 이 변화라는 것이 그 주창자들이 혼쾌히 승복할 수 없는 것들도 있고, 또 그러한 변화들이 모두 똑같이 근본적 인 변화라고 할 수도 없다. 보다 근
본적인 변화일 수도 있고 그렇지 않을 수도 있다. 만일 다 똑같 다면 혁명적 과학과 정상적 과학 사이에는 아무런 차이점이 없게 될 것이다. 이 장에서는 위에서 대략 살펴본 두 방법을 사용해서 고전적 멘델 유전학이 현대 분자유전학으로 변하는 과정을 살펴보게 될 것이다. 우리의 고찰은, 논리적 경험주의지들이 사용하는 최신 분석 방법과 그것에 반대하는 견해에 의거하여 표현될 것이다. 왜냐하 면 현재로서는 분석 방법에 있어 선택의 여지가 없기 때문이다. 이 장의 내용이 변질될지 모르지만 어쨌든 나는 환원에 대한 논 리적 경험주의자들의 분석이 나쁘게 말하면 판단 착오이고 좋게 말 해봐야 부적절하다고밖에 할 수 없음을 먼저 밝혀두고 싶다. 또한 앞으로 전개될 내용은 간추려진 유전학사가 아니며 그럴 의도도 전혀 없디는 것을 독자들에게 말해 둔다. 나는 유전학에 있어서 부분적으로 독립된 두 분야가 거쳐온 발전 과정을 합리적으로 재 구성해 보고자 할 뿐이다. 이러한 두 전통 사이에 선을 긋자면 우선 역사적 배경을 전제로 해야 할 것이나, 그렇게 해서 이상적 인 형식상의 환원 같은 것이 얻어진다 하더라도 결국에는 또다시 시대착오적이라는 판단에 따라 새로운 수정을 가해야만 할 것이 다. 그래서 가능할 때마다, 나는 역사적 사실에서 벗어나 보고자 하였다. 물론 이러한 이원론적인 방법에는 위험이 따른다. 가장 위험한 일은 재구성된 후에 단순하고 명백한 모습으로 나타날 두 과학 이론 사이의 관계와, 과학 이론들이 발달하는 과정과 과정 사이에 있었던 복잡한 관계와의 구별이 없어져서 똑같은 것으로 취급되는 일이다. 어떤 사람들은 교과서를 내던지고 보다 근본적 인 자료로 눈을 돌리려 할 것이고 그렇게 되면 이들은 수많은 역 사적인 사건들에 당면하게 될 것이다. 또 다른 문제는, 이러한 수
많은 사건들이 만들어낼 일련의 철학적인 안건들을 재구성하는 과정에서 놓치게 될지도 모른다는 것이다. 고전적 멘델 유전학 다윈이 『종의 기원』 (1859) 을 출판하고 멘델이 완두콩을 가지고 한창 실험하고 있던 시기에는 원형질이 생명의 본질이라고 간주 되었다. 그 복잡한 구조가 이해됨에 따라 그 속에 생명의 비밀이 있다는 것이 알려졌던 것이다. 세기가 바뀌고 멘델 법칙이 재발견 되면서 관심은 원형질에서 유전자로 옮겨졌다. 1930 \'!tjJ 초부터는 유전자의 구조가 생명 현상의 열쇠를 쥐고 있을 것이라고 생각되 었다. 교과서들은 멘델 유전학에 대해 상당히 직선적으로 설명하고 있다. 멘델 유전학은 유성생식을 하는 다세포 생물에는 잘 적용되 지만 다른 형태의 생물에 대해서는 좀 이야기가 달라전다. 이 유 전 법칙에 따르면 유전자는 염색체의 한 부분이며, 염색체 위에 염주알처럼 배열되어 있고, 약간의 예의를 제의하면 염색체들은 쌍으로 존재한다. 쌍을 이루는 염색체들을 상동염색체라고 한다. 한 쌍의 상동염색체 0 에서 하나는 부계로부터 온 것이고 다른 하나
® 생물의 모든 세포 내에는 염색체가 그 생물 특유의 숫자만큼 들 어 있다. 사람의 경우는 그 숫자가 46 개이다. 이 염색체는 유전자 가 자리잡고 있는 물체로서 현미경으로 관찰하면 길고 짧은 여러 가지 형태의 막대기 모양을 하고 있다. 특정 물감에 염색이 잘 되 기 때문에 염색체라는 이름이 붙어 있다• 한 개의 세포 속에 있는 염색체를 관찰해 보면 똑같은 모양의 염색체가 두 개씩 쌍을 이루 고 있는 것을 알 수 있는데, 여러 가지 세포학적 연구 결과에 의하
여 이 각 쌍의 한 짝은 부계에서, 다른 한 짝은 모계에서 온다는 것이 밝혀졌다. 그래서 사람의 경우에 전체 46 개의 염색체 중 23 개 는 아버지로부터, 나머지 23 개는 어머니로부터 온 것이다. 이 한 쌍씩의 염색체에서 한쪽 염색체를 그 짝이 되는 상대방 염색체에 대하여 상동이라고 한다.
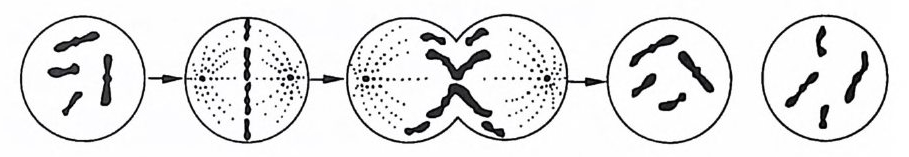
는 모계로부터 온 것이다. 따라서 한 생명체가 가진 염색체의 절 반은 아버지로부터 온 것이고 디론 절반은 어머니로부터 온 것이 다. 유전자도 역시 상동염색체 위에서 한 쌍씩으로 존재한다. 염 색체 위에서 유전자의 상대적 위치를 좌위(座位)라고 하며(때때 로 좌위는 유전자 그 자체를 의미하기도 한다), 같은 좌위를 점하 고 있는 한 쌍의 유전자에서 한쪽을 다른 쪽의 대립유전자(對立 遺 傳子)라고 한다. 유사분열시에 염색체는 길이로 쪼개져서 원래 의 염색체와 똑같은 두 개의 새로운 염색체를 만든다. 세포가 분 열할 때, 새로 만들어전 두 쌍의 염색체는 한 벌씩 니누어져서 새로 형성된 세포로 가게 되며 그 결과로 원래의 세포와 똑같은 두 개의 세포가 만들어진다(그림 1-1 참조).
 효〈〉 \5 습 .A ::.
효〈〉 \5 습 .A ::.
그림 1-1 원래의 세포와 동일한 두 개의 세포가 유사분열에서 만들어지고 있다.
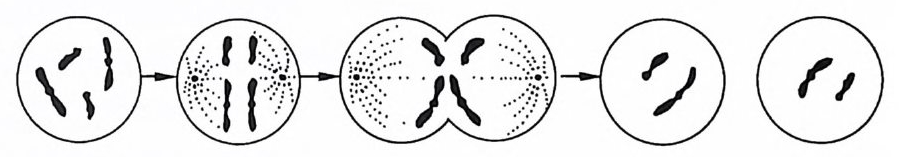
그림 1-2 염색체의 수가 절반인 생식 세포가 감수분열을 통해 만들어지고 있다.
이러한 방식으로, 다세포 생물이 생장하고 또 단세포 생물들이 증식해 나간다. 유사분열로 생긴 결과 중의 하나는 다세포 생물의 모든 세포는 그것이 신경세포이든 근육세포이든 골세포이든 상관 없이 동일한 유전물질을 갖게 된다는 것이다. 한편 수정이 되고 배우자가 형성되는 과정의 어느 시점에서 생식소 내에서는 감수 분열이 일어난다. 감수분열에서도 상동염색체가 쌍으로 배열되지 만 유사분열에서와 같은 식으로 갈라지지 않고 한 쌍의 염색체가 각기 헤어져서 서로 다른 세포로 가기 때문에 새로 형성된 두 개 의 생식세포는 염색체의 수가 절반으로 줄어들게 된다(그림 1-2 참조). 그리고 수정이 일어날 때, 남성의 생식세포와 여성의 생식 세포가 결합하면서 다시 염색체의 수가 원래의 수로 된다. 이 두 생식세포의 결합체를 접합체라고 한다. 이 접합체가 유사분열을 거듭함으로써 새로운 개체로 발달되어 가는 것이다. 감수분열에 있어서, 한 가지 중요한 특칭은 상동염색체가 두 생식세포로 각각 나누어져 들어가는 것은 그 염색체의 출처와는 무관하다는 것이다. 죽, 각 쌍이 따로따로 헤어져 각각의 생식세 포로 가는데 어느 쪽으로 가느냐 하는 것은 그 염색체가 부계에 서 온 것이냐 또는 모계에서 온 것이냐에 관계없이 순전히 우연 의 문제라는 것이다. 따라서 우리 염색체의 절반은 한쪽 부모로부 터, 나머지 철반은 다른 한쪽 부모로부터 받은 것이 틀림없지만, 그 중 얼마만큼이 예컨대 할머니로부터 온 것인가 하는 문제는 알 수가 없다. 예를 들어, 한 아이가 가진 염색체가 모두 의할머 니에게서 온 것이고 의할아버지로부터는 아무 것도 받지 않은 경 우도 이론적으로는 있을 수 있다는 것이다. 이제까지 우리는, 염색체는 변하지 않는 일정한 구조를 지니고 있는 것처럼 얘기했지만 사실은 그렇지가 않다. 염색체 내에 있어

그림 1-3 감수분열의 과정에서 한 쌍의 염색체가 교차 재조합되는 모습
서의 유전자의 순서와 조합을 변화시키는 몇 가지 기작이 있는데, 그 중 교차(交又)와 재조합이 가장 잘 알려진 경우이다. 흔히 상 동염색체는 서로 꼬이고 갈라지고 다시 재조합되면서 잡종영색체 룰 형성한다. 그래서 각 염색체가 어느 한쪽 부모로부터 온 것이 기는 하지만 그런 순수한 상태로 오래 머무는 것이 아니라(그림 1-3 참조) 모계와 부계로부터 온 여러 유전지들 ·로 혼합이 된다. 한 개의 염색체 위에서, 서로 멀리 떨어져 있는 유전자들이 근접 한 유전자들보다 더 자주 재조합을 일으킨다고 가정한다면, 재조 합의 빈도로부터 염색체 지도를 만들 수 있다. 예를 들어, 염색체 위에서 양쪽 끝에 있는 유전자들은 어느 위치에서 교차가 일어나 든간에 한 번 교차가 일어날 때마다 서로 교환이 될 것이고 반면 에 서로 가까이 위치한 유전자들은 교차가 그 두 유전자 중간에 서 일어나야만 분리가 될 것이다. 만일 중간이 아니라 두 유전자 중 어느 한쪽의 바깥에서 교차가 일어난다면 두 유전자는 함께 행동하게 되는것이다. 그러나 재조합이 항상 완벽하게 전행되는 것은 아니다. 감수분 열을 할 때 상동염색체가 제대로 배열되지 않는 경우가 종종 있 는데, 그런 경우에는 재조합중에 교환된 부분이 동일하지 않기 때
문에 한 염색체는 두 개의 대립유전자를 받는 반면 다른 한 염색 체는 대립유전자를 하나도 받지 못하게 된다. 이러한 현상을 중복 (重複)이라고 하는데 단순한 염색체에서 복잡한 염색체가 만들어 지는 한 방법이라 할 수 있다. 염색체 위에서의 유전자의 순서를 변화시키는 또 다른 기작들도 있다. 예를 들어, 한 염색체가 자체 내에서 꼬이고 갈라지고 재결합되면서 유전자의 순서가 역전되기 도 한다. 그 의에도 수많은 다른 종류의 염색체 이상(異常)이 일 어날 수 있는데 어떤 경우에는 염색체의 한 조각이 떨어져 나가 기도 하고 그것이 같은 염색체 위의 다론 부분에 삽입되거나 다 른 염색체의 어느 부분에 삽입되기도 한다. 어떤 경우에는 세포분 열시에 염색체 전체가 소실되어 한쪽 세포에는 염색체가 너무 많 고 다른 한쪽에는 염색체가 너무 적은 결과를 가져오기도 한다. 극단적인 경우에는, 염색체가 중복된 후 제대로 분리가 되지 못해 서 염색체의 수가 두 배로 늘어나는 경우도 있다. 한마디로, 일어 날 수 있는 모든 염색체의 이상이 실제로 거의 다 일어나고 있다 는것이다. 유전자형과 표현형의 관계 앞의 논의에서는 생물의 유전적인 구성을 디루었을 뿐이고 이 러한 유전적인 구성(유전자형)과 그에 의하여 나타나는 표현형의 관계에 대해서는 전혀 언급하지 않았다. 일반적으로 멘델 유전학 의 역사에서는, 유전자형이 생물의 변이(變異)에 대한 한계를 결 정한다는 이론이 지배적이었다 . 한계라 함은 유전자가 발현(發現) 되는 과정에 있어서의 어떤 경계라고 할 수 있다. 그러나 여러
가능한 표현형 중에서 어떤 것이 실제로 나타나게 될 것인가를 최종 결정하는 것은 유전자가 아니라 환경이다. 유전자형과 표현 형과의 관계에 대한 논의는 주로 일단의 유전학자들이 표현 방 식-특히 인간의 지성이나 광기와 같은 형질에 대하여 논의할 때-을 잘못 사용했기 때문에 야기되었다고 볼 수 있는데, 예를 들어 어떤 유전학 교과서에는 〈 인간의 신체적, 정신적 구성을 포 함하여 인간의 모든 것은 인간의 유전적 구성에 의해서 결정된 다 〉 라고 하고 있다. 그러나 이 말은 유전적 구성이 어떤 특성을 결정하는 충분조건이라는 뜻은 아니다. 동시에 어떤 특정한 유전 적 구성이 특정 형질을 만들기 위해 필수적이라는 뜻도 아니다. 서로 다른 유전적 구성이 동일한 표현형을 만들어 낼 수도 있어 서, 유전자형과 표현형과의 관계는 마치 흡연과 폐암의 관계처럼 복잡하고 다양한 것이다. 표현형을 만들어 내는 데 있어서의 유전자형과 환경과의 관계 에 대해서는 멘델 유전학의 초기에 이미 정착이 되었지만(드물게 는 신라마르크주의자들 사이에서도) 특정 유전자와 표현형과의 관 계에 대해서는 별로 알려진 것이 없었다. 사실, 특정 유전자형에 대한 우리의 지식과 그에 의하여 최종적으로 나타나는 표현형에 대한 우리의 지식 사이에 단절이 있다는 사실 그 자체가 바로 고 전 멘델 유전학의 한 특칭이기도 하다. 표현형으로 나타나는 수많 은 특성들이 교배를 통해서 연구가 되고 그러한 표현형을 만들어 낸다고 생각되는 유전자들이 추정되었지만, 과연 어떤 기직을· 통 해서 그 유전자들이 표현형 상의 형질을 만들어내는지에 대해서 는 거의 아는 바가 없었고, 유전학지들은 지금도 아직 그 문제에 골몰하고 있는 것이다. 193% !에 와딩톤 C.D. Wad di n gt o 퍼본 다 음과 같이 불평을 토로하였다. 〈유전학은 그 정확한 법칙성에 있
어서는 논의의 여지가 없으면서도 여러 생물학적인 현상들과의 관련성이 전혀 밝혀지지 않은 유전자들의 행동을 쫓느라고 바빴 고 그래서 유전학은 여러 생물학자들에 의해 유전학 그 자체만의 세계라고 혹평받는 위험한 상황에 빠지게 되었다. 〉 3 )
3) An Int ro ducti on to Modern Geneti cs (London : George Allen & Unw in, 1939), p. 7.
유전자와 형질의 관계에 대한 여러 가설 중, 가장 단순하면서 초기의 멘델 유전학을 완전히 지배했던 것은 1 유전자 /1 형질이었 다. 이 견해에 따르면, 각 염색체는 일정한 숫자의 유전자로 나누 어지고, 표현형은 또 같은 숫자만큼의 형질로 나눌 수 있기 때문 에 유전자와 형질 · 사이에는 1 대 1 의 관계가 성립된다는 것이다. 이 가설은 그 단순성으로 말미암아 그럴 듯하게 보였지만 수많은 개념상의 혼돈을 일으켰고 끝내 사라지고 말았다. 이 혼돈이라는 것은 형질과 유전자 사이의 관계였다. 한 형질에 관련된 유전자와 그 형질 자체를 혼동하기 쉬웠던 것은 어떤 유전자의 존재, 부재 또는 변화 등을 표현형질의 존재, 부재 또는 변화로부터 추정하려 했기 때문이다. 사태를 더욱 악화시킨 것은 초기의 많은 유전학자 들이 물리학의 브리지만 P.W. Br i d g man 과 심리학의 왓슨J.B. Wa t son 에 의해 제기된 실험에 의한 정의 op e rati on al defi ni t ion 라는 개념을 사용했기 때문이다. 실험주의자들의 주장에 따르면, 과학 용어들은 그것이 적용될 수 있는가를 연구할 때 사용하는 방법을 통해서 정의되어야 한디는· 것이다. 예를 들어, 길이나 온 도와 같은 물리적 개념들은 자나 온도계와 같은 것에 의해서 정 의되어야 하고, 분노나 지성 같은 심리적 개념들은 행동과 IQ 검 사 같은 용어로 정의되어야 한다는 것이다. 유전지는- 그 유전자를 추정할 수 있게 한 교배 실험 상의 용어로 정의되는 것으로 해석
되었다. 그러나 실험주의자들의 입장을 엄격한 의미로 따져 본다 면, 1 유전자 /1 형질의 가설은 실속 없는 말장난에 불과하게 된다. 유전학의 역사는 극단적으로 실험주의를 지향한 유전학자들조차 도 그들의 주장을 모두 증명해 보이지 못했음을 분명히 보여주고 있다. 가능한 모든 실험 방법을 동원해 밝혀낸 것보다 실제로는 훨씬 더 많은 것이 유전자 속에 있었던 것이다 .4 )
4) David Hull, The Op er ati on al Imp e rati ve -Sense and Non-sense in Op e rati on is m , Sy st e m ati c Zoology , 16(1968), pp.4 38 - 457 을 참조할 것.
유전학 발달의 초기에 유전학자들은 생물의 표현형이 무수히 많은 특칭으로 니누어질 수 있다는 사실에 부딪쳤다. 그렇게 표현 형을 여러 가지로 나누는 방법 중에서 어느 하나가 다른 것보다 낫다고 한다면 거기에는 어느 정도의 정당성이 있어야 하는데, 문 제는 이러한 정당성의 부여가 유전자형에 대해 갖고 있던 종전의 지식을 그대로 전제했다는 것이고, 또 그 유전자형은 표현형으로 부터 추정될 수밖에 없었다는 대 있다. 표현형은 여러 개의 형질 로 분명하게 나누어질 수 없을는지 모르지만 유전자형은 여러 개 의 유전자로 나누어질 수가 있다. 일단 특정한 유전자가 분리되 면, 그 분리된 유전자를 가지고 실험하여 그 결과로 나타나는 표 현형은 모두 다 그 유전자에 의한 형질로 간주될 수 있다. 그러 나 결과는 실험주의자들이 세운 이러한 명제들이 물리학이나 심 리학에서와 마찬가지로 유전학에서도 결코 아무런 득이 되지 못 했음을증명하고 있다. 게놈g enome ® 상에서 서로 멀리 떨어져 있는 유전자들이 종종
® 게놈g enom~ 한 생물체의 세포 속의 유전자 한 벌을 말한다. 보통 세포는 부계와 모계로부터 한 벌씩의 유전자를 받으므로 두 벌의 게놈울 가지고 있게 된다.
똑같은 형질에 그 영향력울 행사하는데 이러한 유전자들을 · 동의 유전자(同義遺傳子)라고 한다. 예를 들면, 서로 디른 - 유전자 좌위 에 있는 90 개의 유전자들이 모두 초파리의 눈의 색깔에 관여한다 고 한다. 1 유전자 /1 형질의 가설에 의한다면, 이들 유전자 하나하 나는 한 개의 독립된 유전자이고 그래서 초파리의 눈색은 90 개의 다른 형질로 생각되어야 할 것이다 . 마찬가지로 한 개의 유전자가 종종 여러 다른 형질에 관여하기도 하는데 이러한 현상을 다면현 상(多面現象)이라고 한다. 예를 들어, 초파리의 백색 눈 자리에 있는 대립유전자들은 눈의 색깔뿐만 아니라 암컷에 있는 정자 저 장 기관의 모양과 그의 다양한 여러 형질에도 관여한다. 1 유전자/ 1 형질의 가설에 의하면, 이러한 여러 형질은 동일한 형질로 간주 되어야 하거나 또는 그 유전자가 사실은 하나가 아니고 여러 개 의 유전자로 간주되어야 할 것이다. 그러나 사실은 그렇지 않다. 이것은 논리적으로도 맞지 않고 상식에도 어긋나는 것처럼 보였 던것이다. 구조, 기능, 위치 및 유전자의 개념 지금까지 언급한 딜레마는 과학 이론과 그 이론적 실체들에 대 한 연구를 회피했기 때문에 생긴 결과라고 할 수 있다. 이론적 실체를 실존론적으로 해석한다면 이러한 논리적인 혼란들이 해결 될 수 있을 것이다. 〈유전자〉 ®롤 정의하는 데 가장 많이 사용되는
® 유전자는 DNA 라고 하는 길다란 나선형의 물질인데 염색체 위에 길게 한 줄로 놓여 있다. 염색체의 수는 생물의 종류에 따라 다르 지만 대개 100 개 미만이다. 그러나 유전자의 수는 이루 헤아릴 수
없이 많다• 따라서 염색체 하나에는 수많은 유전자가 한 줄로 자리 잡고 있게 된다. 염색체 위의 이 유전자의 자리를 좌위(座位)라고 하는데, 이 위치를 결정하여 지도처럼 표시한 것이 염색체 지도이 다. 유전자의 좌위가 여기서 말하는 위치이다. 유전자의 기능은 말 할 것도 없이 그 유전자에 의하여 나타나는 형질을 보고 알 수 있 는 것이지만, 분자생물학적으로는 단백질이라는 물질 합성에 관여 하는 것이 유전자, 죽 DNA 의 기능이라고 할 수 있다. 이 단백질 이 수많은 화학반응둘을 거쳐 궁극적으로 가시적인 어떤 형질, 예 컨대 피부색이라든지 코의 모양 등으로 나타나게 된다. 그러나 이 러한 형질은 일차적으로 유전자에 의해 결정된다고는 하나 환경에 의한 후천적 영향을 대단히 많이 받는다는 것이 밝혀져 있다. 유전자의 구조라는 것은 DNA 라는 물질의 분자구조를 말하는데, DNA 는 4 종의 단위물질이 일정한 순서로 수천 개 또는 수만 개가 연결되어 있는 물질이며, 이 단위물질의 연결 순서가 바로 유전자 의 구조가 된다• 이 연결 순서에 단 하나의 변화가 일어나도 그 유 전자의 기능은 전혀 다른 것이 된다. 유전자의 구조는 이와 같이 DNA 의 분자구조이기 때문에 구조가 같으면 위치가 달라도 같은 유전자이다• 그러나 위치에 따라 기능에 약간의 차이가 나기도 하 는데, 그것은 한 유전자의 기능, 죽 단백질 합성 기능은 인접한 또 는 관련되어 있는 다론 유전자의 영향을 받는 경우가 많기 때문 Ol 다.
세 가지 기준은 위치, 기능 그리고 구조이다(진화론이 필연적으로 수반하게 될 유전자 개념의 진화적인 차원은 나중에 논의하기로 한 다). 그러면 이 세 기준들 중 어느 하나라도 과연 어떤 두 개의 유전자가 사실은 같은 유전자라고 단정할 수 있게 하는가? 이 세 가지 중에 꼭 필수적이라고 할 만한 것이 있는가? 유전자에 대한 일반적인 분석 방법에 의하면, 위치와 기능 그 어느 것도 어떤 두 유전자가 사실은 동일한 유전자임을 밝히는 데에 충분하지가 못하다. 그러나 두 기준을 동시에 생각해 보면,
위치와 기능이 모두 필수적이면서 그 두 개를 합하면 위와 같은 경우를 충족시키는 일이 종종 있다(하지만 구조에 대한 언급은 하 지 않기로 한다). 염색체 위에서 같은 자리에 위치하고 있고 갇은 형질에 관련하는 유전자라면 그들은 동일한 유전자라고 할 수 있 다. 만일 같은 형질에 관여하지만 그 영향력이 서로 다를 경우 이러한 유전자들은 대립유전자라고 한다. 서로 다른 수많은 유전 자들이 같은 자리에 존재할 수도 있는데 이런 경우는 복대립유전 자(複對立遺傳子)라고 한다(앞에서 언급한 동의유전자와 혼동하지 말 것). 예를 들어, 초파리의 백색 눈에 관련된 유전자 자리에는 14 개의 대립유전자가 있고 이들에 의해서 빨간색으로부터 순백색 에 이르는 여러 가지 눈의 색깔이 만들어 진다. 이렇게 볼 때, 서 로 다른 자리에 위치하고 서로 다른 형질에 관여하는 유전자는· 서로 다른 유전자라고 하는 데에 아무런 문제가 없을 것이다. 이렇게 보면, 〈 유전자 〉 라는 것은 위치와 기능에 의해서만 정의 되는 것처럼 보인다. 그러나 유전학자들은 유전자의 이러한 개념 에 대해 서로 일치된 견해를 보이지 않고 있는데, 이러한 견해의 차이는 위치와 기능 이의의 또 다른 요인도 관련되어 있기 때문 이다. 192 珉텨 말에 이르러 유전학자들은 같은 유전자도 게놈 상 의 서로 다른 위치에서는 다른 기능에 관여하고 있을지도 모른다 고 생각하기 시작하였는데, 이러한 현상을 위치 효과라고 한다. 그런데 사실은, 유전자의 위치가 변하면 그 유전자 자체의 기능에 만 영향을 주는 것이 아니라 다론 유전자들의 기능에도 영향을 주게 된다. 예를 들어, 초파리에 있는 어떤 유전자 하나는 전하게 염색되지 않는 염색체 부위에 있을 경우에는 눈색깔이 고르게 분 포된 상태로 나타나는데, 이 유전자가 염색시에 전하게 염색되는 부위에 위치하게 되면 눈의 색소가 반점 형태로 나타난다. 위치나
기능의 기준으로 보면, 위치와 기능이 바뀔 때마다 이 유전자들은 다른 유전자로 변한다고 보아야 할 것이다. 그러나 유전학자들은 이들 유전자들이 동일한 유전자임을 고집 한다. 일단의 유전학자들이 위치 효과의 존재를 그렇게까지 부인 하려 했다는 사실은 위치와 기능의 기준이라는· 것이 바로 그 유 전자 위에 존재하고 있음을 증명하는 것이 틀림없지만, 위치 효과 가 그러한 문제를 제기했다는 점과 위치 효과라는 것이 시종일관 멘델 유전학의 문맥 내에서 서술 가능했다는 사실은, 위치나 기능 이상의 더욱 근본적인 어떤 것이 멘델 유전자의 개념 밀에 깔려 있음을- 시사하고 있다. 그 어떤 것이란 바로 구조였다. 멘델 유전 학에 있어서 유전자는 이론적 존재이다. 유전자들이 그들의 특별 한 방식대로 각기 행동할 수 있는 것은 바로 이 구조 때문이다. 구조적으로 보아서 동일한 유전자가 게놈 상에서 서로 다른 자리 에 위치할 수도 있고, 특정한 자리에 위치한 한 유전자는 다른 유전자로 변하지 않고도 그 기능을 바꿀 수가 있다. 또한 이러한 유전자는 불활성 상태로 존재할 수도 있고 전혀 기능을 갖고 있 지 않을 수도 있다. 이런 모든 현상이 실제로 일어나고 있고, 이 러한 현상들은 유전학자들이 실험적으로 정의된 기능이나 위치에 의해서가 아니라 특정한 구조로서 유전자를 파악하고 있음을- 제 시하고 있으며, 유전학자들에 의해 그런 식으로 기술되어 왔다. 다음 논의에서는, 유전자를 구조적으로 정의된 실체로서 다루게 되는데, 왜냐하면 바로 그것이 그 시대의 유전학 이론을 - 형성했던 근본적인 가정이었기 때문이며 또 이 의에는 달리 구별할 방법이 없기 때문이다.
우성과 열성 상동염색체 위의 동일한 자리에 같은 구조를 가진 유전자가 놓 여 있다면(이 경우의 염색체는 동형접합(同型接合)이다), 멘델 유 전학자는 그 결과로 나타날 표현형질에 대해 아무런 의문도 갖지 않을 것이다. 그러나 만일 구조가 서로 다른 유전자가(이 경우의 염색체는 이형집합(異型接合)이다) 이 자리를 점하고 있다면 그 표현형은 여러 가지로 달라질 수 있다. 이런 경우, 한쪽 대립유전 자는 상대 쪽에 대해 지배적(우성유전자)이고 다른 한쪽 대립유 전자는 종속적(열성유전자)일 수가 있다. 예를 들어, 사람 눈의 색깔의 경우 갈색 눈을 나타내는 유전자는 파란색 눈을 나타내는 유전자에 대해 우성이다. 여기에서의 형질이란, 한 사람의 눈이 다른 사람의 눈에 어떻게 보이느냐는 것이지 색소 그 자체의 유 무를 얘기하는 것은 아니다. 만일, 파란색 눈을 나타내는 동형접 합체를 가전 사람과 갈색 눈을 나타내는 동형접합체를 가전 사람 이 결혼한다면, 자녀들은 모두가 눈의 색깔에 대해 이형접합체를 지니게 된다. 다시 말해서 한사람 한사람 모두가 파란색 눈과 갈 색 눈을 나타내는 유전자 하나씩을 갖게 되는 것이다 . 이런 경우 우리는 자녀들이 모두 갈색 눈이라고 말할 수 있을 것이다 . 왜냐 하면 갈색 눈의 유전자가 파란색 눈의 유전자보다 우성이기 때문 이다. 유전자를 특정한 구조를 가전 이론적 실체라고 본다면, 이 러한 해석은 매우 타당한 것이다. 그 기작은 모른다 하더라도 말 01 다. 우성과 열성의 관계는 대립유전자에 대해서나 형질에 대해서나 상대적이다. 한 대립유전자는 대립유전자들 중 어떤 것에 대해서 는 우성이고 다른 어떤 것에 대해서는 열성일 수가 있다. 예를
들어, 토끼의 어떤 종에서 정상적인 갈색털을 나타내는 대립유전 지는 흰색을 나타내는 대립유전자에 대해 우성이다(코, 귀, 꼬리, 발 주변을 제의하고). 그런데 흰색을 나타내는 대립유전자는 백화 증( 白化症, alb i n i sm) 을 보이는 대립유전자에 대해서는 우성이다. 앞에서 언급하였듯이, 단일 유전자 자리에 있는 대립유전자들이 여러 개의 다른 형질에 영향을 줄 수가 있는데(다면현상), 이 경 우에도 한 대립유전자가 어떤 형질에 대해서는 열성으로 어떤 형 질에 대해서는 우성으로 행동할 수가 있다. 그래서 단순히 우성 또는 열성이라고 규정할 수 있는 대립유전자란 존재하지 않는다. 어떤 특정한 형질에 한해서만 한 대립유전자가 다른 ” 대립유전자 에 대해 우성이라고 말할 수가 있다. 그러나 어떤 경우에는 특정한 형질만을 얘기한다 하더라도, 대 립유전자가 우성이다 혹은 열성이다 라고 말할 수 없을 때가 있 다. 예를 들어, 동형접합체인 안달루시안 Andalus i an 흰 닭울 동 형접합체인 검은 닭과 교배시키면 자손들은 모두 이형집합체가 될 것이나 겉으로 나타나는 표현형은 푸른색이 도는 회색이 된다. 이것은 불완전 우성의 한 예이다. 따라서 완전한 우성과 완전한 열성 사이에는 연속성이 존재한다는 것이다. 더욱이 형질이 동형 접합체에서보다 이형집합체에서 더 뚜렷이 나타나는 경우도 있는 데 이런 현상을 초우성 (超優性)이라고 한다. 또 어떤 때는 양쪽의 형질이 동시에 이형접합체에서 나타나기도 하는데 이런 현상을 공우성 (共優性)이라고 한다. 그러나 멘델 유전학에서 소개된 가장 복집하면서도 홍미로운 것은 상위(上位) 현상이다. 동의유전자의 경우에는 두 개 혹은 그 이상의 유전자가 동일한 형질에 관여하 는 것으로 생각되는 반면, 상위의 경우는 한 유전자가 다른 유전 자에 영향을 주기 때문에 나타나는 현상처럼 보인다. 예를 들어,
쥐 두 마리를 교배하는데 한 마리는 우성인 회색을 나타내는 동 형집합체이고 다른 한 마리는 열성인 검정색을 나타내는 동형접 합체라면 새끼둘은 모두 회색이어야만 할 것이다. 그러나 이런 경 우, 간혹 백화현상이 나오는데 그 빈도가 돌연변이로 설명할 수 있는 것보다 훨씬 높다. 이러한 명백한 비멘델식 비율의 출현은, 원래의 유전자 좌위에 있는 대립유전자뿐 아니라 또 디른- 제臨] 유전자 좌위에 있는 한 쌍의 대립유전자가 역시 색깔에 관여했기 때문이라고 설명할 수 있을 것이다. 이 제庭除위에 있는 우성 대립 유전자는 정상적인 털 색깔을 나타내고 열성 대립유전자는 백화 증을 나타낸다. 다시 말해서, 제疾持위에 우성 대립유전자가 있을 때는 제 1 유전자 좌위에 있는 대립유전자에 의해서 털의 색깔이 결정되지만, 이 제塗核위에 두 개의 열성 대립유전자를 갖고 있는 쥐는 제 1 좌위에 있는 유전자에 상관없이 모두 알비노가 되는 것 이다. 양쪽 유전자에 대해 이형집합체인 쥐끼리 교배된 경우, 새 끼의 1/4 이 알비노일 것이고, 3/1 얹븐 검정색, 9/1 鈴근 회색이 될 것이다(교배시킨 쥐는 두 마리 모두 알비노를 나타내는 동형집합체 였을 리가 없다. 그랬다면 그것은 회색이 아니라 알비노였을 테니 까). 이러한 경우에 제鈴] 유전자를 제 1 유전자에 대해 상위적이라 고 하며 이 제 2 유전자는 어떤 식으로든 제 1 유전자를 지배하고 있 는 것으로 생각된다. 결론적으로 말하자면, 유전자와 표현형질 사 이의 관계란 매우 복잡해서 1 유전자 /1 형질의 가설을 주장하는 사람들이 상상하는 것보다 훨씬 더 복잡하다는 것이다 .
멘델 유전학에 대한 도전 멘델 유전학은 초기에 많은 변화가 있었다. 독립의 법칙은 서 로 다른 쌍의 상동염색체 위에 있는 유전자들에게만 적용되는 것 이었고, 단순한 우성-열성의 관계는 멘델 자신이 생각했던 것처 럼 그렇게 중요한 문제는 아니라고 생각되었다. 이러한 변화는 멘 델 유전학이 잘못되어서라기보다는 일종의 수정으로 간주되었으 나 어쨌든 캐슬 W. E. Castl e 3! }- 골트슈미트 R i chard Goldsch midt 에 의해 멘델 유전학은 중대한 도전을 받게 되었다. 멘델 유전학의 가장 기본이 되는 것 중 하나는, 이형접합체의 각 대립유전자가 서로의 영향을 받지 않고 원래의 상태로 계속 존재한다는 것이다. 두 개의 서로 다른 동형접합체를 교배시키면 자손들은 모두 이형접합체가 된다. 이 자손들끼리 다시 교배를 시 키면 그 자손의 1/4은 한 개의 동형집합체일 것이고 또 다른 1/4은 다론 동형접합체일 것이고 나머지 반은 이형집합체가 될 것이다. 멘델 유전학의 가장 중요한 현상은 이와 같이 동형접합체 가 이형집합체 상태를 거쳐서 나오는데도 유전자 자체는 전혀 변 하지 않는 것이다. 그렇다면 순수한 상태로 남아 있는 것은 유전 자인가, 아니면 형질인가? 멘델은 거의 모든 것을 형질로써 설명 했고 그가 말한 유전적 〈요소〉에 대해서는 거의 언급을 하지 않 았다. 그러나 초기의 유전학자들이 곧 알게 되었듯이 표현형에만 의거해서 해석을 한다면 멘델 법칙이 적용될 수 있는 형질은 아 주극소수에 불과하다. 캐슬 (1905) 은, 이형접합체가 순수한 상태로 존재하는 것이 아 니라 이형접합체 내의 두 대립유전자가 서로를 오염시킨다고 주 장하면서 문헌 속에 유전자와 형질에 관해 애매한 점이 많음을
지적하였다. 알비노 쥐와 검정색 쥐를 교배시킨 실험을 통해서, 캐슬은, 교배를 거듭할수록 알비노 내에 검은 색소가 증가한다는 사실을 발견하였다. 관련된 유전자들이 이형접합체 속에서 서로를 오영시키기 때문에 이 두 형질이 섞인 것처럼 보인디는- 것이다. 멘델 유전학자들은 섞이지 않는 상위유전지를- 도입함으로써 이런 경우에 대한 해답을 내리고자 하였다. 그래서 알비노 내의 색소가 증가한 것은 오염 때문이 아니라 색소를 만들고 분배하는 데 관 련된 수많은 동의유전자들이 재조합되었기 때문이라고 설명하였 다. 하지만 캐슬은 임의적인 존재를 너무 많이 끌어들이고 있는 것 같다고 생각하였고 따라서 그에게는 멘델주의자들이 이미 천 문학에서 포기되었던 일, 죽 주전원 위에 다시 주전원을 쌓는 일 울 계속하고 있는 것처럼 보였다. 유전자와 형질을 구별하고자 하 는 노력이 대부분 실패했기 때문에 캐슬과 멘델주의자들 사이의 논쟁은 그리 오래가지 못했지만, 유전자와 형질이 명백히 구분되 고 초기의 수많은 유전학자들이 가졌던 실험주의적 사고가 극복 되었더라면, 캐슬 자신도 수많은 경험주의적 증거들이 그의 견해 를 부정하고 있음을 알게 되었을 것이고 그래서 스스로 포기했을 것 0] 다. 1925 년에 스터티반트 A.H. S t ur t evan t는 그가 위치 효과라고 이름붙인 현상을 발견하였다. 멘델 유전학에 대한 캐슬의 반대가 유전자와 형질 사이의 혼란으로 인해 생간 것처럼, 위치 효과를 둘러 싼 논쟁도 역시 여러 다론 혼란들로 인해서 몹시 복잡하게 되어버렸다. 말하자면 서로 다른 수많은 현상들이 위치 효과라는 제목 아래 한 데 묶여졌고, 앞에서 논의된 것과 같은 유전자의 정의와 관련된 많은 주제들은 그러한 혼란을 더 한충 가중시켰다. 이때 가장 중요한 문제는 동일한 혹은 다른 염색체 위의 다른 위
치에서 동일한 유전자(구조적으로 정의된)가 과연 똑갇은 표현형 울 나타낼 것인가 아닌가 하는 것이었다. 어떤 자료들은 유전자들 의 기능이 위치에 의해 좌우된다는 것을 암시하기도 했는데, 그렇 다면 유전자의 기능은 그 구조와 위치 두 가지 모두에 의해 결정 된다고 할 수 있을 것이다. 골트슈미트R. Goldsc hmi d t 5) 는 위치 효과가 멘델 유전학의 원 리에 직접적으로 모순된다고 생각하고 아무리 수정을 가한다 해 도 이런 그릇된 자료를 고칠 수 없을 것이라고 주장하였다. 그리
5) R. Goldschmi dt , The Mate r ial , Basis of Euoluti on (New Haven: Yale Univ e rsit y Press, 1940).
고 특정 유전자에 대한 개념 전체를 포기해야 한다고 했다. 위치 효과는 멘델 유전학에 있어 두 가지 문제점을 제시했는데, 그 첫 째는 동일한 유전자인지 다른 유전자인지를 결정하는 데 있어서 위치와 기능의 관계에 대한 것인데 이는 앞에서 이미 논의된 바 있다. 둘째는 위치 효과의 존재를 설명하기 위한 기작에 대한 것 이다. 멘델주의자들은 위치 효괴를- 인정하지 않을 수 없었지만 그 것을 설명할 수가 없었다. 그러나 이것은 내행성과 외행성의 행동 양식의 차이를 다루던 천문학과는 그 상황이 또 달랐다. 모든 천 문학지들이 그러한 차이점이 존재한다는 것을 알고 있었지만 프 톨레마이오스 체계만으로 그러한 차이점을 설명하는 것은 불가능 했던 것이다. 그럼에도 불구하고 그러한 차이점을 프톨레마이오스 체계 내에서 설명해 보려고 그들은 국단적으로 의도적인(따 hoc) 장치들을 도입했다. 그러나 만일 태양이 태양계의 중심에 있다고 가정했었다면, 지구의 궤도는 당연히 의행성과 내행성 사이에 위 치하게 되었을 것이고, 의행성과 내행성의 차이점은 이 체계의 중 요한 부분을 이루면서 저절로 드러났을 것이다.
점차적으로, 유전학의 발달은 유전자의 구조와 유전자의 작용에 대한 지식에 의존하지 않으면 안된다는 것이 명백하게 되었다. 고 전적인 멘델 유전학에서는 유전자가 돌연변이를 일으킬 수 있다 는 사실 때문에 유전지를 한 개의 변하지 않는 일관된 구조를 지 닌 존재로 생각하지 않았다. 그러나 실제로는 유전자 하나하나가 독립된 존재일까? 유전자들은 - 과연 하나하나씩 나누어질 수 있는 것인가? 만일 유전자들이 염색체 위에 염주알처럼 배열되어 있다 면 구조적으로 쉽게 구별이 될 것이고 재조합은 유전자와 유전자 사이에서만 일어날 수 있을 것이다. 반면에 만일 유전물질이 연속 적으로 연결되어 있는 것이라면 구조 한 가지만으로는 유전지를· 구별할 수 없을 것이며(기능적인 기준도 고려되어야 할 것이다), 재조합은 유전자와 유전자 사이에서 뿐 아니라 유전자 자체 내에 서도 일어날 것이다. 유전자는 돌연변이를 일으킨다. 그러나 어떤 방법을 통해서 일어나는가? 또 어떤 변화들이 일어나는가? 여러 방법이 있겠지만 과연 그 기작이란 어떤 것들인가? 왜 어떤 유전 자는 우성이고 어떤 것은 열성이며 또 어떤 것은 불완전 우성 인가? 어떻게 한 유전자가 디론· 유전자를 억제하거나 조절할 수 있는가? 분자유전학적인 연구는 멘델 유전학의 이러한 여러 의문 울 해결해 주었을 뿐 아니라 멘델 유전학이 대답할 수 없었던 몇 몇 문제에 대한 해답도 주었는데, 195~ 빈에 왓슨J .D. Wa t son 과 크릭 F.H. Cr i ck 에 의해 발표된 DNA 모델은 분자유전학이 발전 하게 된 전환점이 되었다.
멘델 유전학과 분자유전학의 구분 멘델 유전학이 분자유전학으로 환원되었는지의 여부를 결정하 는 데 있어 가장 중요한 문제는, 우선 어떻게 이 두 원리들을 구 분해 내는가 하는 것이다. 유전학의 역사를 자세히 살펴보면서 이 러한 결정을 하는 데 어떤 도움을 얻어낼 수도 있겠으나, 누구나 예상할 수 있듯이, 문제는 멘델 유전학이 끝나고 분자유전학이 시 작되는 일정한 시점이 있을 수 없다는 데 있다. 멘델 유전학이 분자유전학에 비해 훨씬 먼저 발달하기 시작했지만, 이 두 학문은 동시에 그리고 상호 의존하면서 발달되었다고 보는 것이 타당하 다. 이 책 에서 우리는 멘델 유전학에 대한 도브잔스키 Theodosiu s Dobzhansk y의 정의를 따르려고 하는데, 그에 따르면, 〈멘델 유전 학이란 유전자의 차이를 다루는 학문이며, 유전자를 알아내기 위 해 교배라는 방법을 사용한다. 죽 형질이 서로 다른 배우자끼리 교배하고, 잡종인 자손에게서 각 형질이 어떻게 분리되어 나타나 는가를 관찰하는 것이 곧 멘델 유전학이다〉. 유전자의 구조가 어 떻고 생합성 경로가 어떻고 효소의 생성이 감소하고 하는 따위의 지식은, 그것이 예기치 못했던 유전 양상을 발견하는 데 큰 도움 이 되지 않는 한, 멘델 유전학자들에게는 아무런 관심의 대상이 되지 못한다. 그들에게 있어서는 멘델 비(比)에 영향을 주지 않는 차이점이란 차이점이 없는 것과 같은 것이다. 반면에 분자유전학은 생합성에 관한 이론이라고 할 수 있다. 분자유전학자들은 유전물질의 분자 구조나 유전물질이 세대에서 세대로 전달되는 사실에만 관심을 갖는 것이 아니다. 설령 그렇게 하고 싶었다 하더라도 그렇게 할 수가 없었을 것이다. 그들은 분 자 상태가 규명된 유전자들이 어떻게 단백질을 만들고, 그 단백질
은 또 어떻게 표현형질을 만드는가를 밝히는 데 몰두하였다. 와딩 톤 C.D. Waddin g ton 같은 초기의 유전학자는 유전자와 표현형 사이에 복잡한 일련의 화학반응이 존재할 것이라고 이미 예언하 였다. 죽 〈 우성 〉 이나 〈 상위 〉 같은 용어는 유전자들 사이의 반응 울 얘기하는 것이 아니라 유전자가 만들어 놓은 물질 사이의 반 응을 얘기하고 있다는 것이다. 예를 들어, 서로 다른 대립유전자 둘이 이렇게 기능의 차이를 나타내는 것은, 그 유전자들이 생산하 는 효소의 양이 서로 다르기 때문일 수도 있디는· 것이다. 멘델 유전학과 분자유전학의 차이가, 멘델 유전학자들은 전체적 인 표현형질을 다룬 데 반해 분자유전학자들은 분자적으로 규명 된 형질을 다루었다는 데 있는 것은 아니다. 형질전달 유전학에서 사용하는 방법들은 전체적인 표현형질의 유전을 연구하는 데 있 어서 뿐 아니라 예컨대 색소의 전달을 연구하는 데도 사용될 수 가 있다. 사실상, 초기에는 유전자의 미세구조에 대한 연구가 대 부분 전통적인 방법을 통해 이루어졌고(나중에 논의될 시스트론 cis t r o n, 레콘 recon 등을 참고할 것 ), 분자 연구 결괴는- 또 거꾸로 멘델 유전학 속에서 더욱 쉽게 이해가 되었다 . 예를 들어, 앞에서 언급한 안달루시안 닭에 있어서의 불완전 우성 현상은, 양쪽 유전 자가 모두 기능을 갖고 있는 경우이기 때문에 멘델 유전학에서 말하는 공우성 현상으로- 더 잘 설명될 수가 있다. 이때, 실제로는 검정색과 흰색의 색소 양쪽이 모두 생성되며, 그것이 회색으로 보 이는 것은 단지 그것을 보는 사람의 시각이 얼마나 정확한가에 달려 있는 문제이다. 인간에게 있어 갈색 눈은 파란색 눈에 대해 우성인데, 파란색 눈이란 무슨 푸른 색소를 만들기 때문이 아니라 하늘이나 맑은 물이 파란 것처럼 그저 빛의 굴절로 생긴 결과일 뿐.o l 다.
이제 되돌아보면 멘델 유전학에서는 이형접합체에 있어서 유전 자가 순수하게 보존되는 현상을 설명하지 못했고, 상위나 다면현 상 또는 위치 효과 같은 현상들에 대해서도 만족할 만한 설명을 하지 못했다는 사실을 느끼게 된다. 이러한 현상들은 멘델 유전학 의 전통적인 방법에 의해 발견이 되었는데 이것을 설명하기 위해 서는 그러한 현상들에 들어맞는 유전자가 필요했으나, 이러한 현 상이 실제로 존재한다는 사실 그리고 그에 관련된 유전자의 작용 은, 우성과 열성의 경우처럼 멘델 유전학으로부터 그렇게 자연스 럽게 유도되지는 않았던 것이다. 분자유전학의 가장 두드러진 특 칭은, 여러 단백질-구조 단백질, 촉매 작용을 하는 단백질효소, 조절 단백질 등-울 만드는 데 있어서의 DNA 와 RNA 분자의 중추적인 역할과 생합성 경로에 대한 개념이라고 할 수 있다 . 분 자유전학은 고전적인 멘델 유전학이나 현대의 멘델 유전학보다 훨씬 더 강력한 이론이라고 볼 수 있는데, 왜냐하면 고전적인 멘 델 유전학과 현대의 멘델 유전학 그리고 그 의의 여러 다른· 방법 에 의해 설명되었던 모든 형태의 현상들을 분자유전학 혼자서 모 두 설명해 낼 수 있기 때문이다. 예를 들어, 멘델 유전학의 상위 유전자들 사이에는 우선 순위가 있는데 이러한 현상은 분자적으 로 설명될 수가 있다. 한 가지 가능한 설명은, 동일한 생합성 경 로에서 이들 유전자가 먼저 나타난다는 것이다. 마찬가지로, 어떤 경우에는 위치 효과도 오페론 o p eron, 죽 한 개의 작은 DNA 토 막의 통제 하에 있는 복합유전자 단위 때문에 생기는 것이라고 추정해 볼 수 있다(이 문제는 뒤에서 다시 논의될 것이다). 따라서 앞에서 정의한 바와 같이, 멘델 유전학과 분자유전학에 대한 비교는 비대칭적이다. 멘델 유전학이 형질유전이라는 방법을 사용해 유전물질을 유전자로 분석하고 그 결과로 나타나는 멘델
의 비로서 이러한 유전자들의 특성을 설명하는 일로 국한되는 반 면, 분자유전학은 DNA와 그에 의해 합성된 단백질 사이를 연결 하는 분자적 기작까지 포함하는 것으로 확대된다. 이 이상 다르 게, 이 두 영역을 설명할 방법은 없다. 혹자는, 이 두 영역을 형 질유전에 관한 두 학문으로서 혹은 발생에 관한 두 이론으로서 비교해야 한다고 생각할지 모르지만 그것은 불가능한 일이다. 멘 델주의자들이 말하는 유전자와 분자적인 유전지를- 비교해 볼 수 도 있겠지만, 그렇게 비교하다 보면 아마 멘델 유전학과 분자유전 학의 중심적인 내용 대부분을 빠뜨리게 될 것이다. 분자 수준에서 멘델의 우성 유전자와 상관 관계에 있는 유일한 것을 찾는다면, 그것은 아주 세분화된 특정한 분자 기작이지 따로 분리된 DNA 가닥이 아니다. 물론 다른 방식으로 멘델 유전학과 분자유전학을 재조합할 수 있을지도 모른다. 그러나 문제는, 이러한 재조합이 논리적 경험주의자들의 설명 방식을 통해서 멘델 유전학을 분자 유전학으로 환원시킬 수 있느냐 또는 없느냐 하는 것이다. 유전물질의 분자 구조 멘델 유전학자들은 유전자가 어떤 일정한 구조를 지니고 있어 서 유전자의 기능 전부 또는 일부분이 그 구조에 의해 결정된다 고 생각했으나, 이러한 구조의 성격을 알지는 못했다. 여러 물리 학자들이 유전자의 구조를 규명해 보려고 노력했는데, 슈뢰딩거 E. Schro di n g er(1945) 는 유전자를 주기가 일정하지 않은 결정체 라고 생각했고, 델브뤽 M. Delbr ii ck(1949) 은 안정된 상태에 있는 준현미경적 체계일 것으로 생각했다. 그러나 보어 N. Bohr(l932)
는 유전자가 화학적으로 분석 불가능한 것으로 판명될지도 모른 다고까지 했다 .6) 그 분자 구조가 아무리 복잡하고 정교한 것이라 해도 어떤 분자 구조를 가지고 설명하기에는 유전 현상이란 것이 너무 복잡했다. 사실상 1940 년 이전까지는, 단백질이 가장 강력한 유전물질 후보였는데 그 이유는 단백질이 유전 현상을 설명하기 에 충분할 만큼 복잡한 구조를 가지고 있기 때문이었다. 또 다른 중요한 후보였던 핵산은 그 구조가 너무 단순했기 때문에 묵살되 었다. 핵산 ® 은 인(燦), 당(糖) 및 47} 지 영기(鹽基)로 구성되어 있으므로 핵산의 일종인 DNA 의 전반적인 구조는 여러 단백질들 과 비교해 볼 때 사실 매우 단순하다. 그러나 모든 이의 상상을 초월하여 바로 이 DNA7} 유전 현상을 설명하는 데 가장 적합한 물질임이 니중에 증명되었다.
6) 슈뢰딩거, 델브뤽 그리고 보어의 논문에 관해서는 블랙번 Black burn(1966) 을 참조할 것. ® 핵산(核酸)은 처음 발견될 당시 (166 8\'l)에는 세포의 핵 속에 존 재하는 산성물질이라는 의미에서 핵산이라는 이름이 붙었는데, 그 후 핵 속뿐만 아니고 핵 밖에도 존재한다는 것이 알려졌다. 이것은 4 종의 뉴클레오티드 nucleo ti de 라는 구성물질(단위물질)이 수없이 많이 연결되어 있는 물질인데, 이 단위물질의 종류에 따라 DNA 와 RNA 의 두 종류로 나누어진다. 이 중 DN A;가 바로 유전자이다.
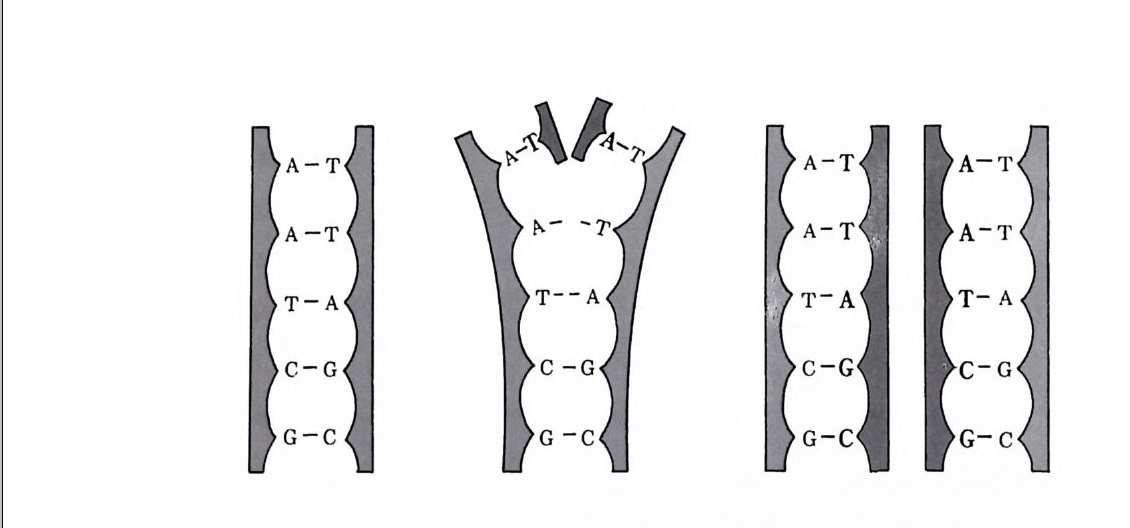
분자적으로 따진다면, 멘델의 유전자란 DNA 의 한 분절(分節) 이라고 할 수 있다. DNA는 보통 나선형으로 서로 꼬인 채 사다 리 모양을 하고 있는 분자, 죽 이중나선(二重螺旋, double heli x) 구조를 하고 있다. 사다리의 양쪽에는 당과 인의 분자가 교대로 배열되어 있다. 이와 같이 핵산에는 인이 있는데 단백질에는 인이 거의 없다는 사실이 핵산이 유전물질임을 밝히는 데 큰 도움이 되었다. 왜냐하면 어떤 세포가 바이러스에 감염되었을 때나 혹은
형질 전환을 일으켰을 때, 바이러스의 단백질은 그냥 세포 밖에 남아 있고 핵산만이 세포 속으로 들어가기 때문이다. 이중나선의 두 가닥을 연결하는 것은 4 개의 염기인 구아닌 (Gua ni ne, G), 아 데닌 (Aden i ne, A), 시토신 (C yt os i ne, C), 티민 (Th ymi ne, T) 이 다 . ” DNA 의 기본 단위는 이 4 개의 영기 중의 하나에 인과 당이 결합하여 이루어전 뉴클레오티드 nucleo ti de 라는 물질아다. 사다리
7) 바이러스와 세균 가운데는 시토신이 메틸시토신 또는 히드록시메 틸시토신으로, 그리고 아데닌이 메틸아데닌으로 바뀌어 있는 것이 있다.
의 가로 부분을 형성할 때 각 염기는 다론 염기 중 어느 하나하 고만 결합할 수 있는데, 즉 구아닌은 시토신하고만, 아데닌은 티 민하고만 결합할 수 있다. 이렇게 각 염기가 언제나 한 종류의 짝만을 택할 수밖에 없다는 사실로 인해 DNA 분자의 구조는 더 욱 단순해진다. 그러나 유전물질이 되려면 어느 정도는 복잡해야 할 필요가 있다. 이러한 복잡성은, 모르스식 전신부호에서 알파벳 이 단순한 점과 선으로 번역될 수 있는 것처럼 염기쌍둘이 순서 롤 지어 한 줄로 배열함으로써 가능하게 된다. DNA 분자의 한 부분은 그립 1- 伊} 같이 도식적으로 표현될 수 있다. DNA 분자의 기능은 두 개의 과정으로 나누어질 수가 있는데, 그 중 하나는 복제(複製)이고 다른 하나는 전사(轉寫)이다. 복제 란 분자 수준의 유사분열이라고 할 수 있다 . 복제 과정에서는 DNA 의 가로 부분을 이루는 염기 사이의 결합이 끊어지면서 분 자의 가운데가 갈라지게 되는데, 이렇게 떨어져나간 반쪽 부분은 거기에 맞는 뉴클레오티드로 다시 채워진다. 사다리의 양쪽은 서 로 상보적(相補的)이기 때문에, 이렇게 해서 생긴 DNA 분자는 갈라지기 전의 구조와 똑같게 된다. 이렇게 하여 새롭게 만들어진
 \/_\/-_\/—_\//_P\ _/ P_\/_P\/ P\
\/_\/-_\/—_\//_P\ _/ P_\/_P\/ P\
그림 1-4 한 가닥의 DNA 를 이루고 있는 4 개의 뉴클레오티드
분자의 반쪽은 원래의 DNA 분자를 이루던 원자들이고 나머지 반쪽은 환경으로부터 새로 얻은 원자들이다(그림 1-5 참조). 따라 서 복제 과정을 통해 생기는 최종 물질은 두 개의 분자라고 할 수 있고, 이 두 분자는 각각 원래의 DNA 분자와 동일한 구조를 갖게 된다(그림 1-5 의 a 와 c 를 비교해 볼 것 ). 만일 DNA 가 이러 한 복제 기능 하나만 갖고 있다면 DNA 분자의 유기적 구성에 대해 더 이상 말할 것이 없겠으나, DNA 는 리보핵산 (RNA) 이라 는 또 한 종류의 핵산을 만드는 기능, 죽 전사의 기능도 지니고 있다. RNA 는 여러 가지 면에서 DNA 와 다르다고 할 수 있다. RNA는 대략 DNA 분자의 한쪽만으로 되어 있어서 말하자면 사 다리의 반쪽에 해당된다. 그 기본 골격은 당과 인이 교대로 배열 되면서 이루어져 있는데, 당은 디옥시리보오스 deox y r i bose7} 아 니라 리보오스 r i bose 이다. 염기의 경우에는 티민이 우라실 uracil 이러는 것으로 바뀌어 있을 뿐 다른 염기들은 DNA 와 동일하다. 그 다음에는 번역이라는 과정을 통해서 단백질이 합성되고 이 과
 AA--TT A-T IA TTACGGC
AA--TT A-T IA TTACGGC
그림 1-5 복제를 설명하는 그림. 연은 부분은 원래의 DNA 분자를 나타내고 진한 부분은 새로 형성된 뉴클레오티드를 나타낸다.
정은 이러한 RNA 분자를 매개체로 해서 이루어전다. 단백질은 분자 구조가 대단히 복잡하지만, 따져보면 204 중의 아 미노산으로 이루어져 있는 물질일 뿐이다. 아미노산들은 단백질 내에서 선상(線狀)으로 배열되어 폴리펩티드 사슬을 이루고 있는 데, 이 아미노산의 배열 순서를 단백질의 1 차 구조라고 한다. 단 백질의 구조가 복잡한 것은 이렇게 선상으로 배열된 분자들이 서 로 꼬이기 때문이며, 이렇게 꼬인 분자들이 다시 또 다른 분자들 과 결합하면서 더욱 복잡한 3 차원적 구조를 이루게 된다. 그러나 이런 복잡한 구조도 기본적으로는 1 차 구조에 의해서 결정된다. 물 론 환경은 여기서도 유전자형과 표현형의 관계에 영향을 미친다. 유전암호(遺傳暗號)를 발견하는 데 있어서의 문제점은, RNA 의 영기쌍을 통해서 DNA 의 염기쌍(유전자 암호 속의 문자)과 단백 질을 구성하고 있는 아미노산의 관계를 밝히는 일이었다. 어떤 염 기들이, 또 몇 개의 염기들이 아미노산을 결정하는 것인가? 유전
암호의 문자는 코돈 codon 이라는 단어로 배열되는데, 문제는 몇 개의 문자가 한 개의 코돈을 이루는 것인지, 어떤 코돈이 어떤 아미노산을 만드는지 하는 것 등이었다. 그러나 해답은 예상했던 것보다 훨씬 간단했다. 만일 코돈의 길이가 모두 똑같다면, 한 코 돈이 가질 수 있는 최소한의 길이는 양 n 의 문자로 되어 있다는 것이다. 만일 코돈이 단 한 개의 문자로 되어 있다면 단지 4 종의 아미노산만이 4 개의 염기 A, U, G, C 에 의해 결정될 수 있을 것 이고, 만일 코돈이 업 n 의 문자로 되어 있다면 16 종의 코돈만이 만 들어질 수 있을 것이다. 코돈의 길이가 沙 n 의 문자로 늘어난다면 64 개의 코돈이 가능하게 되는데 이것은 2 眞은의 아미노산을 충분 히 암호화할 수 있는 수에 해당된다. 이 시점에서, 어떤 이는 모 든 코돈의 길이가 똑같다는 가정을 재검토해 보려하거나(이것은 지극히 매력적이지 못한 대안이지만), 아니면 3 개의 문자로 된 코 돈과 2~ 은의 아미노산의 관계를 정립해 보려고 할 것이다. 그러나 아미노산과 염기와의 관계에는 1 : 1 의 대응 관계가 성립될 수 없 다. 왜냐하면 아무 아미노산과도 관계없는 코돈도 있을 수 있고, 어떤 코돈은 단백질 합성 과정의 시작 또는 종료에 관여하고 있 울 수도 있으며, 혹은 한 개 이상의 코돈이 동일한 아미노산을 암호화하고 있을 수도 있기 때문이다. 또 만일 신이 정말 심술궂 으신 분이라면, 같은 코돈이 한 개 이상의 아미노산울 암호화하고 있을수도 있을 것이다. 암호는 이와 같이 沃문자t r ip le t로 되어 있으며, 위에서 말한 여 러 가지의 가능성 중에서 한 개를 제의하고는 모두가 실제로 일 어나고 있는 것으로 밝혀졌다. 죽 아미노산은 2 종 내지 연돈의 코 돈에 의해 지배되는데, 예를 들면, 아미노산 페닐알라닌은 2 종의 코돈 (UUU 와 uuc) 에 의해, 아미노산 류신은 연은의 코돈 (UU A,
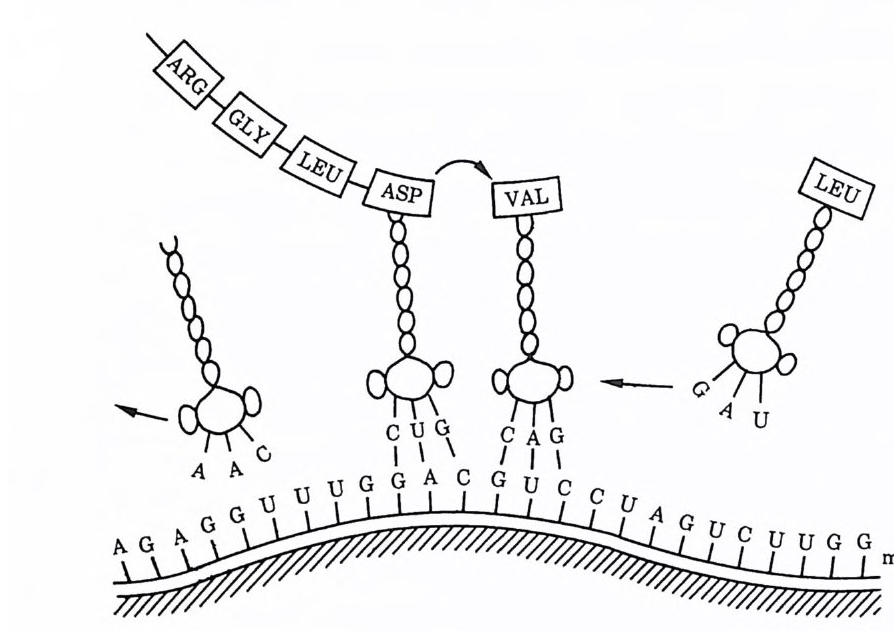
UUG, CUU, CUA, CUC, CUG) 에 의해 지배된다. 어떤 코돈들 은 단백질 합성을 종결하는 마침표와 같은 역할을 하는데 (UAG, UAA, UGA), 이들은 어떤 아미노산도 암호화하지 않기 때문에 〈 넌센스 nonsense 코돈 〉 이라는 오해받기 쉬운 이름을 갖게 되었 다. 한때는 암호란 것이 불확실하고 모호한 것처럼 보이기도 했 다. 예를 들면, GGA7} 글리신과 글루탐산이라는 두 종의 아미노 산 모두를 암호화하는 것으로 생각되었던 것이다 . 그러나 지금은 그러한 모호함은 모두 사라졌고, 유전암호라는 것이 우리가 원하 는 만큼 질서정연하지는 않을지 모르지만 어쨌든 놀라우리만치 단순하다는 사실을 알게 되었다. 이미 언급한 바와 같이 DNA 는 두 개의 기능을 지니고 있는 데, 그 하나인 복제 과정에서는 한 개의 DNA 가 복제되어 두 개 로 늘어나며, 또 하나의 과정인 전사 과정에서는 상보적인 RNA 분자를 만들어낸다. 이때 원래의 분자가 지녔던 염기의 순서는 새 로 만들어지는 분자 속에서도 계속 그대로 지켜진다. 새로 만들어 진 RNA 분자는, 어떤 단백질의 도움을 받아서 자신에게 알맞는 단백질을 만들어내는데 이 과정을 번역이라고 한다 . 이 번역의 과 정을 통해서, DNA 속에 암호화되어 있던 염기 서열은 아미노산 서열로 바뀌게 되고 결국 단백질이 합성되는 것이다. RNA 에는 세 가지 종류가 있는데, 이 중 하나는 단백질과 함께 세포질 내 에서 리보좀 r i bosome 이라고 하는 세포내 소기관을 만든다. 리보 좀 속에 들어 있는 RNA 를 리보좀 RNA(rRNA) 라고 한다. 두번 째는 전령 RNA( 傳令 RNA, mRNA) 로서, 단백질 합성을 준비하 기 위해서 한 개 또는 여러 개의 리보좀에 일렬로 배열한다. 그 러면 세번째의 RNA 인 운반 RNA( t RNA) 가 거기에 맞는 아미노 산을 전령 RNA 상의 적당한 위치를 찾아 운반해 오게 된다.
운반 RNA 의 구조에 대해서 약간의 설명이 필요한 까닭은 그 구조가 단백질 합성에 있어서 중심적인 역할을 하기 때문이다. 운 반 RNA 는 클로버의 잎과 같은 모양을 하고 있으며, 중간 잎에는 세개의 염기가 나와 있어서 전령 RNA 상의 적합한 코돈과 짝을 이루게 되어 있다. 이렇게 운반 RNA 의 중간 잎에 있는 양脂의 염 기로 된 3 문자를 역코돈 (an ti codon) 이라고 한다. 이것은 전령 RNA 의 코돈과 상보적이기 때문이다. 단백질이 합성될 때, ATP (adenosin e tr ip hos pha t e) 와 여러 효소의 도움을 받아 적 합한 아 미노산이 운반 RNA 의 줄기에 와서 붙게 되는데 이 부분은 중간 잎의 반대 쪽에 위치한다. 이처럼 아미노산을 싣고 있는 분자를 아미노아실-운반 RNA(am ino acy l t RNA) 라 한다. 이와 같은 아 미노산을 실은 분자가 전령 RNA 상의 적합한 코돈과 결합하고 나면, 마치 컴퓨터용 테이프가 컴퓨터의 헤드를 향해 상대적으로 움직이는 것과 갇이 리보좀은 전령 RNA 에 대해 상대적으로 움 직이면서 전령 RNA 상의 코돈을 계속 읽어 내려간다. 각 코돈이 지나가면 그에 적합한 아미노아실-운반 RNA 가 들어와서 가져온 아미노산을 폴리펩티드 사슬에 새롭게 부착시키며, 그러고 나면 폴리펩티드 시술은 다음 아미노아실 - 운반 RNA 로 옮겨 붙는다 (그림 1-6 참조). 이 과정들은 모두가 아주 정확할 뿐만 아니라 전령 RNA, 리보좋 아미노아실-운반 RNA, 여러 가지 효소들, 활성화된 GTP(gu a nosin e tri p h osp h ate ) 같은 여러 가지의 분자 둘이 함께 관여하고 있는 대단히 복잡한 과정이다.
생합성 경로 DNA 의 복제, RNA 로의 전사, 단백질로의 번역 및 유전암호는 다 함께 분자유전학의 가장 중요한 구성 요소라 할 수 있다• 이 에 덧붙여 한 가지 또 다른 중요한 요소를 든다면 그것은 생합성 경로라고 할 수 있을 것이다. 분자유전학이 출현하기 이전의 생물 학에는 유전자와 형질 사이에 커다란 틈이 있었다. 후에 다소 수
 cI u1 \ --o 4 u
cI u1 \ --o 4 u
그림 1-6 이 그립은 4 개의 운반 RNA 분자가 단백질 합성에 관여하고 있는 모습을 보여주고 있다. 오른쪽에는 류신을 실은 분자가 전령 RNA 의 적 철한 코돈과 짝을 이루려하고 있다. 발린을 실은 두번째 운반 RNA 분자 는 그것에 맞는 코돈과 짝을 이루고, 길어지고 있는 폴리펩티드 사슬이 막 이 두번째 운반 RNA 가 실어온 발린과 결합하려 하고 있다. 세번째와 네번째 운반 RNA 에 의해 실려온 아미노산은 이미 폴리펩티드 속으로 결 합되었고, 네번째 운반 RNA 는 아미노산을 내려 놓고 떠나가고 있다. 메 시지 message 전체가 다 번역될 때까지 이러한 과정은 계속된다.
정이 되긴 했어도 멘델 유전학에 있어서, 유전자와 형질은 여전히 하나의 가정에 의해서 연결될 수 있었을 뿐이었다. 분자생물학이 발전함에 따라 초파리의 눈색과 같은 하나의 형질이 발현되는 과 정이 실제로는 얼마나 복잡한 것인지롤 비로소 깨닫게 되었다. 생 합성 경로에서 일어나는 여러 일들을 도식적으로 표현한다면, 특 정한 유전자가 RNA 를 통해서 효소를 생산하고 이 효소는 기질 (基質)에 작용하여 필요한 산물을 만들며, 이렇게 만들어전 산물 은 다시 다른 유전자에 의해서 만들어전 효소의 기질로 사용되어 두번째의 산물을 만들게 된다. 이런 식으로 과정은 계속된다(그림 1-7 참조).
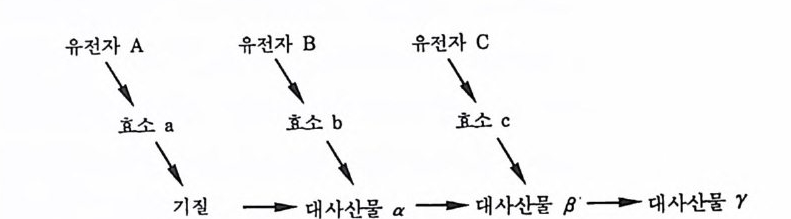 유전자\ A \기 질 유--전-\자-- B 대 사\산 물 a 유--전\-자-- C대 \사산 물 B' - ----대 사산물 Y
유전자\ A \기 질 유--전-\자-- B 대 사\산 물 a 유--전\-자-- C대 \사산 물 B' - ----대 사산물 Y
그림 1-7 생합성 경로를 요약하여 설명한 그립. 세 개의 유전자가 벌이는 연 속적인 작용을 통하여 기질이 최종 산물로 전환된다.
단편적으로 보면, 생합성 경로란 것이 연쇄적으로 일어나는 단 순한 사건처럼 보이지만, 일반적으로 생합성 경로는 수없이 가지 를 치고 순환을 하는 복잡한 그물망과 같은 과정이다. 하나의 대 사 산물(代謝産物)이 한 개 또는 여러 다른 반응에서 기질로 사 용되기도 하고, 또 반대로 동일한 대사 산물이 여러 가지 경로를 통해서 만들어지기도 한다 .. 그 러한 중복성은 분자적인 수준에서나 거시적 수준에서나 전화에 있어서 아주 중요한 의미를 지닌다. 어 느 요소가 연쇄과정 상의 어떤 특정한 부분에 영향을 주는가를 알아보기 위해서 사용하는 가장 일반적인 방법 중의 하나는, 한
번 실험할 때마다 요소를 하나씩 변화시키면서 그에 따라 결과가 어떻게 변했는가를 알아보는 것이다. 만일 A 를 제거시킨 결과로 B 가 사라진다면, A 는 B 를 위한 필수 조건임에 틀림없다. 그러나 생물학적으로 중요한 기능이 다른 방법을 통해서도 생성되는 경 우가 종종 있기 때문에 이러한 방법이 실패하는 경우가 자주 생 긴다. 예를 들어, 낙엽송이 살아남기 의해서는 봄철에 수액이 상 승되어야 하며, 암토끼가 종족을 보존하려면 배란이 일어나야 한 다. 그러나 이러한 과정을 자세히 조사해 보면, 수액이 상승하고 배란이 일어나기 위해서는 이것이 절대적으로 필요하다고 말할 수 있는 딱 한 가지의 현상을 집어낼 수가 없다. 즉 낙엽송의 뿌 리를 찰라내고 나무를 수액이 담겨진 그릇에 옮겨 놓더라도 수액 은 여전히 상승한다는 것이다. 따라서 뿌리의 압력이 수액을 상승 시키는 데 필수 조건이라고 할 수가 없다. 또 잎을 떼어버린다 해도 수액은 역시 상승할 것이다. 그래서 잎도 역시 필수적인 기 능을 수행한다고 할 수 없다. 또 나무 둥지의 체관부p hloem 세포 롤 죽인다 하더라도 수액은 역시 싱승할 것이고, 이런 식으로 실 험을 계속해 볼 수 있을 것이다. 토끼의 배란에 있어서도 같은 식의 얘기를 할 수 있다. 때로 중요한 기능들 하나하나가 너무 과장되는 경우가 많이 있지만, 사실은 다른 여러 기작들과 또 그 것을 보충하는 기작들 하나하나 역시 동일한 기능을 수행할 수 있는 경우가 많아 있다. 그러한 중복성이야말로 생명체로 하여금 전화적으로 대단히 잘 적응해 나갈 수 있게 하는 요인이다. 어떤 과정이 단 한 가지 방법에 의해서만 이루어질 수 있다면, 그러한 방법에 어떤 장애가 생길 경우 그 개체는 당장 죽게 될 것이고 결국 그 종은 멸종이 될 것이다. 예를 들어 진화 과정중 어느 시 기엔가 인류는 비타민 C 를 생성하는 능력을 잃었는데, 만일 인류
가 음식으로 비타민 C 를 섭취하지 않았다면 그러한 돌연변이체 인간은 이미 멸종이 되었을 것이다. 물론 그러한 중복성 자체도 진화적으로 그 대가를 치루기는 했다. 즉 이러한 중복의 존재는 또 다론 중복을 희생시키면서 그 균형을 유지시킬 수 있었던 것 이다 (4 장의 설명을 참고할 것). 유전학에 있어서의 이론의 환원 앞에서 우리는 멘델 유전학과 분자유전학의 윤곽을 살펴보았는 데, 지금부터는 환원이라는 문제에 대해 좀더 자세히 살펴보고자 한다 . 몇몇 철학자들은, 어떤 하나가 다른 하나로 〈환원〉될 때 그 두 이론 사이의 관계를 반형식적 se mifo rmal 으로 분석하였다. 예 를 들어 , 케메니 J. G. Kemen y와 오펜하임 P. O pp enhe i m8) 은 그들 이 〈 간접 환원 〉 이라고 이름을 붙인 것에 대해 설명했으나, 그들
8) J. G. Kemeny and P. Op pe nheim , On Reducti on , Phil o sop h i- cal Stu d ie s , 7(1956), pp. 6-17.
의 분석은 실제로 환원이라기보다는 우리가 〈대체〉라고 부르는 것과 동일한 것이었다. 예를 들어, 연소 이론과 산화 이론은 다같 이 열 현상을 설명하고자 하는 것이었는데, 결국에는 산화 이론에 의해 연소 이론이 사장되었다. 이 경우, 어떤 의미에서든 연소 이 론이 산화 이론으로 〈환원〉되었다고 말할 수는 없다. 그러나 케 메니-오펜하임의 방법은 이론을 대체하기 위한 분석 방법으로써 유용하게 쓰일 수가 있다. 이들에 의하면, 어떤 이론이 디론· 이론 보다 더 우수하려면 그 이론은 더 체계적이어야 하고 그 영역도 훨씬 더 넓어야 하며 더 정확한 예측을 할 수 있어야 한다는 것
이다. 또 다른 두 조건은, 확고히 정립된 다른 이론들과 잘 조화 될 수 있는 양립성과 단순성이라고 할 수 있는데, 이 둘은 가장 중요하면서도 문제의 소지가 많은 항목이다. 대체의 고전적인 예로서 프톨레마이오스와 코페르니쿠스의 천 문학을 흔히 얘기하지만, 그 둘 사이의 관계는 흔히들 말하는 것 처럼 그렇게 일방적인 것은 아니었다. 계산법으로는 두 이론 모두 훌륭하게 체계화되어 있었으며, 완성된 상태에서는 두 이론 모두 똑같이 정확하게 천체의 위치를 예측해 낼 수 있었다. 다만 유일 한 차이점은, 똑같아 정확한 예측을 하기 위해서 프톨레마이오스 의 이론이 코페르니쿠스의 것보다 몇 개의 주전원을 더 필요로 했다는 것뿐이다. 그러나 물리학에서와 마찬가지로 어쨌든 코페르 니쿠스 천문학이 프톨레마이오스 천문학에 비해 우수하였던 것은 사실이다. 프톨레마이오스 천문학의 경우에는 억지로 설정한 가정 들에 의존해야만 설명 가능했던 여러 현상들, 즉 내행성(수성, 금 성)과 의행성 운동의 기본적인 차이점 같은 것을 코페르니쿠스 천문학은 쉽게 설명해 낼 수가 있었다. 하지만 코페르니쿠스 천문 학도 나름대로 문제가 있었다. 이론적으로는 항성 운동 시차(恒星 運動 視差 ste l lar p arallax) 를 관찰할 수 있어야 했는데도 실제로 는 관찰할 수가 없었던 것이다. 그러나 코페르니쿠스 천문학의 가 장 큰 장애 요소가 되었던 것은 무엇보다도, 자연에 대한 물리학 적 이론에서부터 질병에 대한 4 기질(四氣質) 이론에 이르기까지 아리스토텔레스 과학의 이론 그 어느 것과도 조화될 수가 없었다 는것이다. 환원에 대한 고전적 인 분석은 우드거 J. H. Woodg er , 네겔 E. Nage l , 퀸 W. V. Qu in e 등의 방법 9) 이라 할 수 있는데, 그것은 직
9) J. H. Woodg er , Bi ol og y and Lang u ag e (Cambrid g e : Cambrid g e
Un ive rsit y Press, 1952) ; E. Nage l , The Str uc tu r e of Scie n ce (New York : Harcourt, Brace, & World, 1961) ; W. V. Quine , Onto l og ica l Reducti on and the World of Numbers, Jo urnal of Phil o sop h y, 61( 19 64), pp. 97-102.
접 환원이라고 이름 붙여진 분석 방법이다. 이러한 견해에 따르 면, 환원되는 이론이나 환원하는 이론이나 그 각각의 영역 안에서 는 본질적으로 모두 옳은 이론이지만, 환원하는 이론은 환원되는 이론보다 더 넓은 범위의 영역을 지니며, 환원되는 이론의 고유한 영역 내에서조차도 환원하는 이론이 환원되는 이론보다 더 정확 하게 어떤 사실을 예측할 수 있게 한다는 것이다. 이 두 이론이 같은 서술적 용어를 사용한다면 이러한 환원을 동질적이라 하고, 같은 용어를 사용하지 않을 경우에는 이질적이라고 한다 . 이질적 인 환원의 경우에, 환원되는 이론의 보다 원시적인 모든 용어들 (pr im itive te rm , 각 개체 내의 여러 부분을 분류하고 서술하는 모 든 용어들)은 환원하는 이론 내의 적절한 용어와 연관이 되어야 만 하는데 이러한 연관은 환원 함수에 의해 이루어질 수 있다. 하지만 환원되는 이론 내의 어떤 용어와도 연관지울 수 없는 새 로운 서술적 용어들이 환원하는 이론에 더 첨가될 수도 있다. 환 원 함수란, 두 이론의 상호 관계를 표현하고 있는 물리적 가설이 나 또는 그에 상당하는 원리라고 할 수 있다. 어느 경우이전간에, 이것은 두 이론을 종합하여 내린 주장이어야 하며 경험에 의한 지지롤 얻을 수 있어야 한다. 결국, 환원하는 이론과 환원 함수를 종합하면, 그것으로부터 환원되는 이론이 추론될 수 있어야 한다 는것이다. 포퍼 K. Pop pe r, 파이어아벤트 P. K. Fey er abend, 쿤 T. Kuhn 등 10) 은 과학에 있어서의 직접 환원의 가능성을 설득력 있게 반박
10) Karl Pop per, Conje c tu re s and Refu ta t i on s(New York : Basic
Books, 1962) : P. K. Fey e rabend, Ex pl anati on , Reducti on , and Emp iri c ism , Mi nn esota Stu d ie s in the Phil o sop h y of Scie n ce, Vol. III(Mi nn eap o lis : Un ive rsit y of Mi nn esot a Press, 1962) ; Thomas Kuhn, 'Ihe Str u ctu re of Scie n ti fic Revolut ion s(Ch ica g o : Un ive rsit y of Chic a g o Press, 1970).
하였다. 동질적인 환원의 경우 두 개의 이론이 동일한 언어를 사 용하고 있다고 해서, 그 단어가 같은 의미로 사용되었다고 할 수 는 없다는 것이다. 이질적인 환원에 있어서는 용어 자체가 다르므 로 더 말할 필요도 없다. 만일 관찰에 의한 용어도 이론을 담고 있다는 것을 인정한다면, 서로 다른 두 개의 이론이 〈 동일한 〉 현 상을 설명하고 있다고 말하는 것은 더욱 의문인 것이다. 아리스토 텔레스 물리학에서는 천체의 궤도가 완벽한 원형에서 벗어날 때 그에 대한 설명을 요구하는 반면, 뉴턴 물리학에서는 그 궤도가 완벽한 직선을 벗어날 때 그 설명을 요구하고 있는 것이다. 그러 나 다른 관점에서 보면, 포퍼, 파이어아벤트, 쿤 등이 환원이라는 작업을 현재로서는 가장 잘 분석했다고 할 수 있다. 덜 타당하다 고 할 수 있는 이론이 더 타당한 이론으로 환원되기 위해서는 환 원되는 이론에 반드시 수정이 가해져야만 한다. 환원되는 이론들 은 역사적 실체가 아니라 논리적으로 재구성된 것들이기 때문에, 환원하기 위해서 역사적인 정확성과 논리적인 정확성, 둘 다 충족 되어야 한다면 형식 for mal 환원이란 아예 불가능한 일이 될 것이 다. 그러나 샤프너 Kenneth Scha ffner11) 의 주장처럼, 만일 환원되 는 이론을 보다 정확하게 하기 위해서 역사적인 정확성이 어느
11) Ap pro aches to Reducti on , Phil o sop h y of Scie n ce, 34(1 9 67), pp. 137-147 ; Schaff ne r' The Wats o n-Cric k Model and Re-ducti on ism , Briti sh Jo urnal for the Phil o sop h y of Scie n ce, 20 (19 69), pp. 325-348.
정도 희생될 수 있다면, 환원을 부정하는 포퍼, 파이어아벤트, 쿤 등이 제시한 명제도 일반적인 환원의 좋은 예가 될 수 있을 것이 다. 이것이 바로 형식 환원 이론이 의미하는 바 전부이며, 따라서 만일 그 이상을 더 요구한다면 그 요구 자체가 잘못된 일이다. 샤프너의 일반적 환원 패러다임에 따르면 다음의 5 가지 조건을 충족시켜야만 환원으로 간주될 수 있다. (1) 환원되는 이론이 정확하게 수정된 후 그 이론 내에 있던 모든 용어가 환원하는 이론 내에서 나타나거나, 아니면 환원하는· 이론 속에 있는 한 개 또는 그 이상의 용어와 연관이 되어야 한 다. 이러한 연관성에 의해 두 용어는 서로 관련되는 존재 refe r en- tial i den tity가 될 것이다. 즉 그 두 용어의 의미가 똑같지는 않 더라도 그 두 용어가 같은 실체를 표현하게 되는 것이다. 각 용 어의 일차적인 의미는 그 용어가 들어 있는 원래의 이론 내에 있 는 것이지만, 환원 함수를 통하여 이차적인 의미를 새롭게 부여 받을수있다. (2) 환원하는 이론과 환원 함수로부터 환원되는 이론이 정확한 모습으로 유도될 수 있어야 한다. (3) 수정된 상태의 환원된 이론은 원래의 이론을 수정한 것이 어야 할 뿐 아니라 왜 그것이 잘못된 것이었는가를 지적할 수 있 어야한다. (4) 환원하는 이론은 엉성하고 비형식적인 방법으로나마 환원 되는 원래의 이론을 설명할 수 있어야 한다. (5) 원래의 이론과 수정된 이론 사이에는 커다란 유사점이 있 어야한다.
샤프너의 분석법으로 환원을 설명하려면, 세 개의 이론, 즉 원 래의 이론, 수정된 이론 그리고 환원하는 이론을 항상 명시하여야 한다. 유전학의 경우에, 원래의 이론이란 이 장의 앞부분에서 논 의된 고전적인 멘델 유전학의 교과서적 이론이며, 수정된 이론은 초기의 현대적 형질전달유전학이고, 환원하는 이론이란 현대의 분 자유전학이다. 우리가 이미 보아온 바와 같이 고전적인 멘델 유전 학은 관찰이 가능한 표현형질이 한 세대에서 다음 세대로 전달되 면서 나타나는 멘델 비 (比)를 찾는 일 , 또 어떤 유전자에 대한 상위유전자의 수 등에 관심을 둔다. 이때의 형질이란 〈 파랗게 보 이는 눈 〉 처럼 매우 다양한 것이다. 유전지는- 영주알의 하나하나 처럼 독립된 실체이며 구조적으로 정의된 존재, 기능적으로 이미 결정된 존재로 간주되고, 이 유전자들은 직접 또는 간접으로 표현 형질을 지배한다. 교차는 유전지들- 사이에서만 일어나고, 돌연변 이는 유전자 구조에 변화가 생긴 것이기 때문에, 기능 단위, 교차 와 돌연변이는 동시 공존할 수 있다 . 물론 멘델 유전학에 대한 이러한 견해는 잘못된 것들이다. 우리는 소위 〈 허구적인 유전자 str a w g ene 〉를 설정했던 것이며, 초기의 유전학자들은, 물리학자 들이 보어 Bohr 의 축소판 태양계를 가지고 원자를 설명하려 했던 것보다도 더 가볍게, 〈선상에 배열된 구슬 〉 의 비유를 가지고 유 전자를 설명하려 했던 것이다. 두 모델 모두 각각 유전자와 원자 에 대해 그럴 듯하게 있을 법한 가상의 속성을 부여했을 뿐, 실 제로 발견된 이론의 한 부분이 될 수 있는 실질적인 속성을 부여 하지는 못했던 것이다. 한 모델의 성공 여부는 그 모델과 그 모 델이 표현하고자 하는 자연 현상이 얼마나 유사한가에 달려 있다. 원자가 항성계와 어떻게 다르고 염색체와 실에 꿰인 구슬이 어떻 게 다른가 하는 문제는, 원자론과 유전자 이론이 발달하는 데 있
어 매우 중요한 문제 였 음이 증명되었다 (4 장의 유사에 대한 논의를 참조할 것). 형질전달유전학은 고전적인 멘델 유전학의 현대판으로 간주된 다. 왜냐하면 멘델 유전학의 기술(技術)과 설명 방식을 그대로 사 용하기 때문이다. 그러나 동시에 이 두 이론 사이에는 분명한 차 이가 있는데, 그 차이점은 형질전달유전학자들이 추적하고 있는 형질의 형태가 멘델 유전학의 그것과 다르기 때문에 생긴 것이다. 유전학자들은 더 이상 〈 파랗게 보이는 눈 〉 식의 표현형질에 국한 되지 않고, 겉으로는 똑같게 보이는 형질일지라도 생화학적인 차 이를 근거로 하여 선별해 낼 수가 있게 되었다. 고전적인 멘델 유전학이 현대의 형질전달유전학으로 전화하게 된 것과 분자유전 학의 출현은 깊은 관계가 있다고 할 수 있는데, 다시 말해서 형 질전달유전학의 향상은 생화학적 연구를 더욱 용이하게 해 주었 고, 형질전달유전학지들은 표현형질의 생화학적인 성질이 발견됨 으로써 그들의 분석 방법을 개선할 수가 있었다• 손본 (Sonnebor n. 1963 : Ap ple to n , 1970 의 22 - 26 쪽)은 다음과 같이 적고 있다. 출발할 때부터 유전학은 어느 정도 생화학적이고 분자적인 것 이었다. 약 60 년 전에 가로드 Garrod 는, 인간에게는 유전자에 의 해 조철되는 생화학적 형질이 있다는 것을 지적하였으며, 그 이 후로 그에 필적하는 연구와 이론을 구성하려는 노력이 없었던 시 대란 없었다. 그러나 한편으로, 새로운 유전학이 달성한 놀라운 승리 중의 어떤 것, 예를 들어 벤저 Benzer 가 유전자의 아단위 subun it의 숫자와 배열을 발견한 것과 같은 일들은 고전 유전학 의 방법을 통하여 이루어질 수 있었다.
고전적인 멘델 유전학과 현대의 형질전달유전학은 그들이 취급 하는 형질의 형태가 다를 뿐 공통적인 방법론과 설명 양식을 지 니고 있다. 분지유전학의 전보로 인해 이제 형질전달유전학자들은 화학적으로 이미 밝혀진 표현형질이 어떻게 전달되는가를 추적할 수 있게 되었다. 똑같이 파랗게 보이는 두 개의 물건이 있다고 하자. 그 중 하나는 파란 색소를 가지고 있어서 파랗게 보이는 것이고, 다른 하나는 단지 빛의 굴절 때문에 파랗게 보이는 것일 지도 모른다. 이와 같이 유전물질의 분자 구조와 분자적으로 규명 된 표현형질에서 일어나는 생합성 경로에 관심을 둔다는 점에서 분자유전학은 고전적인 멘델 유전학이나 형질전달유전학과 다르 다고 할 수 있다. 따라서 이론의 환원을 위한 조건을 충족시키려 면 형질전달유전학의 원시적인 용어들은 적당한 환원 함수를 통 해 분자유전학의 용어와 연결될 수 있어야 한다. 샤프너는 환원 함수에 사용되는 용어를, 분류하는 용어 class t erm 와 서술하는 용어 pre dic a te t erm 의 두 가지 형태로 구분하 였다. 형질전달유전학에 있어 개체에 적합한 분류 용어는 유전자 이며, 이러한 유전자는 하나 또는 여러 개의 DNA 나 RNA 의 분 절을 지칭하는 것으로 간주되어야 한다. 마찬가지로 형질전달유전 학의 모든 서술 용어, 죽 동형집합체, 우성, 열성, 공우성, 상위 등의 용어들도 분자유전학의 내용과 효과적으로 연결지워 질 수 있어야한다. 처음에는 환원주의자들의 프로그램이 유전학과 잘 맞아 떨어지는 것처럼 보였다. 유전자는 DNA의 한 부분으로 간 주될 수 있었고, 상위는 각 단계가 수령적으로 계속 연결되어 나 가는 생합성 경로를 가지고 설명할 수 있었으며, 다면 현상은 생 합성 경로가 계속 분기되어 나가는 경우를 통해서, 위치 효과는 게놈 상에서 일어나는 한층 높은 수준의 조직화 or gani za ti on 된
집단, 죽 오페론 o p eron 갇은 용어로 설명될 수 있었다. 그러나 특정한 환원 함수를 사용하여 이렇게 인상적인 첫번째의 접근에 좀더 충실을 가하려고 하면, 이야기는 이상하게 점점 장황해지기 만 하고 환원주의자들의 프로그램이 도대체 이치에 맞는 것인지 에 대한 의문이 생기게 된다. 환원 함수__유전자 앞에서 고전적인 멘델 유전학을 살펴보는 동안, 우리는 유전자 롤 따로따로 분리되어 있고 변하지 않는 하나의 일관된 구조를 갖는 것으로 가정하였다. 또한 교차, 돌연변이 및 기능의 단위를 모두 동일한 것으로 간주하였으나, 실제로는 그렇지가 않다 (Pe t er, 1957 속의 Muller, 1927 참조). 정도의 차이는 있겠지만, 유전물질 은 분리되어 있는 것이 아니라 계속 연속되어 있는 존재이며, 일 련의 뉴클레오티드로 되어 있다. 교차와 돌연변이의 최소 단위는 한 개의 뉴클레오티드임이 판명되었고, 점 돌연변이(點 突然變異) 는 한 개의 영기쌍이 변해서 생간 결과임이 밝혀졌다. 예를 들어, 전령 RNA 의 특정 부분을 전사하는 DNA 의 코돈이 CAT 에서 CTT로 바뀔 수가 있는데, 그렇게 되면 전령 RNA는 GUA 에서 G AJ넘_ 바뀌게 될 것이며 결과적으로 아미노산은 발린에서 아르 기닌으로 바뀌게 될 것이다. 이러한 변화는 단백질의 기능에 심각 한 영향을 미칠 수도 있고 그렇지 않을 수도 있다. 다시 말해서, 유전암호에 중복현싱이 있기 때문에, 어떤 돌연변이는 그 결과로 나타나는 단백질에 거의 또는 전혀 영향을 주지 않을 수도 있다 는 것이다. 예를 들어, UUU가 UUC 로 변한다 하더라도 UUC 역
시 UUU 와 갇은 페닐알라닌을 만들게 된다. 그러나 어떤 경우는, 유전 암호가 바뀌는 데 따라서 결과적으로 약간의 변화가 일어날 수도 있는데, 그것은 다른 종류의 운반 RNA 가 필요하기 때문에 생기는 결과이며, 종의 종류나 조직의 종류에 따라 또 운반 RNA 의 종류에 따라 운반 RNA 의 양이 다르기 때문이다. 표현형에 변 화가 일어나든 일어나지 않든, 또 표현형으로 나타나는 정도에는 상관없이, 이러한 변화는 모두 돌연변이로 간주된다. 어떤 경우든 그 DNA의 구조가 변했기 때문이다. 돌연변이와 마찬가지로 교차 도 그것만으로는 기능 단위를 묘사하는- 데 사용될 수 없다. 교 차는 두 뉴클레오티드 사이에서 언제든 일어날 수 있는 일이며, 기능 단위 내에서의 당과 인의 결합이나 기능 단위 사이의 결합 이나 서로 다를 바가 없는 것이다. 만일 현대 형질유전학이 분자유전학으로 환원되었다면, 멘델유 전자는 DNA 나 RNA 의 어떤 부분과 동일한 것이어야 한다. 벤저 (Benzer, 1955 : Pete r , 1959 에서) 이후의 대부분의 생물학자들과 마찬가지로 샤프너도 멘델 유전자와 시스트론 ® 이 동일한 것이라고 주장했는데, 시스트론은 시스-트랜스 c i s- t rans 데스트에 의해 설 명된 기능 단위이다. 돌연변이나 교차는 그 최소 단위가 한 개 의 뉴클레오티드이기 때문에 그 어느 하나만으로는 DNA 의 기능 단위를 구별해 내는 데 사용될 수가 없다. 그러나 그 둘의 효 과를 동시에 분석한다면 더러는 기능 단위가 결정될 수 있을 것이다. 이것이 바로 시스-트랜스 데스트의 핵심이다. 만일 두 개
® 시스트론은 유전자의 최소 단위를 말한다. 분자생물학적으로는 하 나의 폴리펩티드p ol yp e pti de 분자를 합성할 수 있는 DNA 의 토막 울 말한다. 유전자의 정의에는 여러 가지가 있는데, 일반적으로 하 나의 유전자 단위를 시스트론이라고 하기도 한다.
의 돌연변이가 동일한 염색체 위에서 일어난다면 그들은 시스 위 치에 있다고 하고, 대립되는 상동염색체에서 각각 일어난다면 그 것은 트랜스 위치에 있다고 한다. 만일 교차가 상동염색체에서 일 어난다면, 시스 위치에 있는 두 돌연변이는 트랜스 위치로, 트랜 스 위치의 돌연변이는 시스 위치로 바뀔 수도 있게 된다(그림 1- 8 참조). 만일 두 돌연변이가 염색체 위에서 서로 다른 기능 단 위에서 일어난다면 그것이 시스에 있든 트랜스 위치에 있든 아무 런 차이가 없을 것이며, 따라서 이 두 단위는 그것에 상관없이 각자의 기능을 나타내게 될 것이다. 그러나 돌연변이가 같은 기능 단위 내에서 일어난다면, 시스 형과 트랜스 형은 서로 다르게 행 동할 것이다. 시스 위치에서는, 한 염색체 위에서 적어도 한 분절 은 영향을 받지 않고 정상적으로 작용을 할 것이고, 트랜스 위치 에서는, 양 염색체가 모두 같은 기능 단위 내에 돌연변이 유전 자를 갖게 되므로 DNA 의 그 어느 분철도 정상적인 기능을 할 수가 없을 것이다. 그래서 시스-트랜스 효과를 보이는 두 돌연변 이는 같은 시스트론에 속한다고 정의할 수 있다. 시스-트랜스 효 과를 보이지 않는다면 아마 그들은 같은 시스트론에 속하고 있지 않을 것이다 .12)
12) 본문 중의 시스-트랜스 데스트는 상동영색체를 가진 생물에게만 적용되는 것이지만 그렇지 않은 생물에서도 비슷한 적용을 할 수 있다. 시스-트랜스 테스트에서 필요한 것은 돌연변이룰 일으킨 두 유전자가 같은 세포 내에 존재함으로써 기능의 상호 보완 여부를 알 수 있게 해주는 것이다.
멘델 유전자와 시스트론을 동일시하는 것은 타당하지 못한 일 이라고 할 수 있다. 여러 가지 이유가 있을 수 있겠지만 가장 중 요한 이유는, 메이어 Ernst Ma yr와 루스 M i chael Ruse7t 지적한 것처럼, 〈시스트론〉이 분자유전학으로부터 나온 용어가 아니라는
 (a) cis po sit ion (b) cro ss - o ve r (c) tra ns po sit ion
(a) cis po sit ion (b) cro ss - o ve r (c) tra ns po sit ion
그림 1-8 시스 형의 돌연변이가 일어난 상동영색체가 교차에 의하여 트랜스 형의 돌연변이로 바뀌는 것을 보여주는 그림. +는 정상적인 대립유전자 를 나타내고, A 와 B 는 돌연변이를 나타낸다 .
점이다. 그것은 교차와 재조합의 연구에 의존하는· 현대 형질유전 학에 속하는 개념이며, 거기에는 DNA의 분자 구조나 생합성 경 로에 대한 지식이 전혀 포함되어 있지 않다. 〈 시스트론 〉 은 분자 유전학에 의해 유도된 용어에 가깝고 새로운 이론을 발전시키는 데도 유용한 도구로써 사용되긴 했으나, 결코 멘델 유전자나 DNA 와 동일시될 수는 없다. 시스-트랜스 데스트가 문헌에 사용 되는 횟수가 점차 줄어들고 있음이 시사하는 것처럼, 시스-트랜 스 테스트나 그것과 관련된 개념이 유전학에서 점점 경시되어 가 고 있다는 사실은 그러한 주장을 간접적으로 증명하는 것이다. 그 러나 시스-트랜스 테스트가 DNA 분자 내에서 여러 수준으로 조 직화되어 있는 유전자 집단들을 식별해 내지 못하고, 또 유전물질 내의 기능 단위를 조사하는 다른 데스트들의 결과와 항상 일치되 는 것도 아니기 때문에, 설령 시스트론이 분자유전학에서 나온 용 어로 간주된다 하더라도, 샤프너의 동일하다는 주장을 받아들이기 에는 역시 미흡한 데가 있다. 멘델 유전자와 한 분자의 RNA(DNA 의 분자 산물이라고도 할 수 있다)를 생산해 내는 DNA 분절을 동일시하는 것은 바람직한
So
일로 보인다. 1 유전자 /1 형질이라는 가설은 생명체에 나타나는 표 현형을 하나하나씩 분리된 형질 단위로 나누어서 분석할 수 없다 는 사실 때문에 좌절되었다. 반면에 DNA 의 여러 분절이 만들어내 는 산물은 하나하나가 독립된 분자이기 때문에, 1 유전자 /1 분자 산 물이라는 가설은 확고한 기반 위에 서 있다고 볼 수 있다. 따라서 이 하나하나의 독립된 분자들로부터 DNA 분자를 더 작은 단위로 나눌 수가 있다. 또한 유전자를 각 분자로서 경계를 짓는 일에는 종말 코돈 ter m ina to r codon 과 같은 구조를 사용할 수가 있다. 그러나 이러한 동일화도 너무 단순한 것이다. 어떤 DNA 분절 은 아무 아미노산도 암호화하고 있지는 않지만 전체적인 유전 체 제에서는 하나의 중요한 기능을 담당하기도 한다. 어떤 때는 한 개 혹은 여러 개의 시스트론의 기능이 조절(調節)유전자나 작동 (作動)유전자와 같은 특별한 DNA 분절에 의해 조절된다. 조절유 전자는 RNA 분자를 거쳐 단백질을 생산하며, 이렇게 생산된 단 백질은 작동유전자라 불리는 DNA 분절에 가서 결합한다.단백질이 작동유전자에 결합되어 있으면 인접한 시스트론의 기능을 방해하 게 된다. 이렇게 해서 인접 시스트론이 생성하는 물질의 농도가 너무 낮아지게 되면 다시 단백질이 작동유전자에서 분리되고, 이 들 시스트론은 다시 그 물질을 생산하다가 충분한 농도에 이르면 그 단백질이 다시 작동유전자에 결합하게 된다. 작동유전자와 그 것을 조절하는 인접 시스트론을 오페론 o p eron이 라고 한다. 〈유전 자〉에 대한 환원 함수는 적어도 아미노산을 암호화하지는 않지만 다른 분절의 DNA 의 기능에 관여하고 있는 이런 작동유전자리는· 존재를 포함할 수 있어야만 한다. 유전자의 기능에 대한 아주 단 순한 환원 함수가 만들어질 수도 있겠지만, 위에서 언급한 바와 같이 유전작용이 갖고 있는 특징들 때문에 이러한 환원 함수는
아마도 1 대 1 의 관계가 아니라 1 대 다수 one-man y의 관계로 하 나씩 분리하여 표현하는 것이 되기 쉽다. 다시 말해서, 한 개의 멘델 유전자가 여러 개의 서로 다른 분자 구조와 연관되어 있을 것이라는 것이다. 그러나 분자 쪽에서 멘델 유전자 쪽을 향해 나 가는 방향으로 환원을 위한 추론이 이루어져야지 그 반대 방향으 로 되어서는 안 되는 것이기 때문에, 그러한 동일화는 환원의 목 적에 잘 부합되는 것 같다. 만일 환원을 위한 추론이 멘델 유전 자에 대한 설명으로부터 특정한 분자 구조 쪽을 향해 진행되어야 했다면, 그런 식의 동일화는 적절치 못했을 것이다. 유전 암호를 논하기 위해 사용된 용어를 빌려 표현한다면, 중복이라는 말은 환 원에 수용될 수 있겠으나 한 개의 코돈이 여러 종의 아미노산울 지배한다는 뜻의 불명확 amb iguity이라는 말은 수용될 수가 없을 것이다. 환원 함수와 서술용어 멘델 유전학에서는 수많은 형질과 행동들이 멘델 유전자를 가 지고 서술되었는데, 이러한 서술 중의 일부는 멘델 유전학이 분자 유전학으로 환원되는 데 아무런 문제도 야기시키지 않는다. 아마 이 일부라는 것은 〈동형집합체〉나 〈이형집합체〉와 같은 순전히 구조적인 용어들일 것이다. 예를 들면, 두 생명체가 어떤 한 형질 에 관하여 동형집합체라고 한다면 그것은 이 두 생명체가 상동영 색체 위에서 서로 마주 보고 있는 같은 분자 구조의 유전자를 가 지고 있다는 말과 같고, 만일 그 형질에 대해 이형접합체라면 그 유전자들은 서로 디론 구조를 가지고 있다는 말과 같다 .• 이 러한
동일화 과정에서 혼란이 생길 수 있다면 그것은 유전암호의 중 복 때문일 것이다. 그래서 한발짝 더 나아가서, 한 아미노산이 바 뀜으로써 어떤 단백질의 기능이 현저히 변화되는 경우와 그렇지 않는 경우를 구별하려 한다면 불필요한 혼란을 몰고 오게 될 것 이다. 우리가 여기서 주의해야 할 점은, 이 과정에서 상동염색체 리는 개념이 쓰이고 있디는 것이다. 세포학적 측면에서 본다면, 상동염색체란 감수분열시 짝을 이루는 염색체를 말한다. 유전자에 대한 우리의 관심을, 그러한 구조적인 특칭에서 표현 형질로 돌리게 되면 상황은 갑자기 대단히 복잡해진다. 멘델 유전 학에서 하나의 대립유전자가 어떤 형질에 대해 〈우성〉이라고 할 때, 이것은 이 대립유전자가 조절하는 형질이 어떻게 한 세대에서 다음 세대로 전달되는가에 관해 설명하고 있는 것이다. 이들 형질 이란 보통 전체적인 형질을 가리키는 것이고, 대개 인간의 지각 (知 登 )으로 인지되는 것들이다. 서술용어를 위한 환원 함수에 관 심을 가졌던 유일한 과학철학자는 샤프너였다. 그가 시스트론과 멘델 유전자를 동일시했다는 것은 이미 앞에서 논의한 바 있거니 와, 그의 견해가 옳지 못하다고 판명되긴 했어도 어쨌든 그의 시 도는 훌륭한 출발점이 되었다. 하지만 그가 〈우성〉이라는 용어를 위해 제시했던 환원 함수는 전혀 만족스럽지 못한 것이었다. 그는 〈 우성 〉 이라는 서술용어롤 〈 x 는 활성 효소의 합성을 지시할 수 있 다 〉 는 식의 문장으로 표현하기 위하여, 〈 DNA 분절 1 이 활성효소 의 합성을 지시하는 부분〉일 경우에만 한하여 〈유전자 1 은 우성 이다〉라고 할 수 있다는 것이다. 이 경우, 샤프너는 분자 구조와 단백질 합성에 관련된 기능을 논하고 있는 것이다. 이러한 환원 함수가 아주 그릇된 것이라고 단정할 수는 없겠지만, 문제는 그가 우성에 대한 멘델의 개념을 찰못 표현했다는 데 있다. 첫째, 다른
대립유전자들에 대해 우성인 유전자의 대부분은 효소를 생산하는 일과 무관하다는 것이다. 그 예로 구조 단백질의 전령 RNA 를 암 호화하는 유전자들을 들 수 있다. 둘째, 멘델 유전학에 있어서 〈 우성 〉 이라는 것은 상대적인 개념이어서 , 아무런 조건없이 이 유 전자는 우성이다라고 할 수 있는 유전자란 없다는 것이다. 이러이 러한 조건 하에서 특정 형질을 나타내는 대립유전자들 중 어느 것이 어느 것에 대하여 우성이다라고 말할 수 있을 뿐이다. 똑같 은 유전자라 할지라도 주어전 조건이 달라지면, 디~ 형질이나 다 른 대립유전자에 대하여 열성이 될 수도 있다. 효소를 암호화하고 있는 유전자의 대부분은 활성(최소한 어느 정도는)을 지닌 효소를 생산한다. 따라서 샤프너의 분석에 따른다면, 효소를 암호화하고 있는 이 유전자들은 모두 〈 우성 〉 이다라고 정의되어야 할 것이다. 그러나 사실은 이들 유전자 가운데 많은 것들이 디~ 유전자들에 대하여 열성인 것이다. 멘델 유전학이 분자유전학으로 환원되기 위해서는 이보디는- 더 나은 환원 함수가 있어야만 할 것이다. 그러나 그러한 환원 함수 롤 만든다는 것이 처음 예상했던 것보다 훨씬 힘든 일이라는 것 을 사람들은 곧 알게 되었다. 거기에는 여러 가지 문제가 있는데, 첫째 어떤 가시적인 표현형질이 분자적으로 규정된 형질로 바꾸 어져야만 하는 것이다. 멘델 유전학의 방법론으로는 가시적인 표 현형질이나 분자론적으로 규명된 표현형질이나 다 같이 분석해 볼 수 있으나, 가시적 표현형은 아무래도 너무 복잡해서 분자유전 학적인 분석에는 적합하지 않기 때문이다. 둘째, 여러 가지 멘델 적인 서술용어들(예를 들어, 〈 우성〉)이 하나 혹은 여러 형태의 분 자론적 기작(예를 들어, 활성 효소의 생산)과 연관지어져야 한다 는 것이다. 유전자의 환원 함수에서 언급했던 것처럼, 분자적인
용어와 멘델적인 용어 사이의 이러한 관계는 일 대 일 혹은 다수 대 일은 될 수 있으나 일 대 다수는 될 수 없다. 다시 말해서 한 개의 분자 기작이 여러 개의 멘델적 서술용어와 관련되어 있을 수는 없다는 것이다. 셋째, 환원을 위해 필요한 모든 조치가 다 취해진 후에도, 이렇게 수정된 멘델 유전학은 수정되기 전의 원래 의 멘델 유전학과 극히 유사한 모습으로 남아 있어야 한다는 것 이다. 예를 들어 고전적인 멘델 유전학에 있어서, 우성과 열성 사 이에 어떤 대칭성이 존재하는 것처럼 일련의 상위 유전자들 사이 에도 일정한 순위가 존재한다. 이 두 가지가 합쳐져서 우성과 열 성 상위 현상이 생기게 되는데, 이 관계는 수정된 멘델 유전학에 서도 그리고 분자론적으로도 어느 정도 유지되어야 할 것이다. 예 를 들면, 우성과 열성을 만들어내는 분자 기작이 상위를 만들어내 는 분자 기작과 결합하여 이러한 우성과 열성 상위 현상이 일어 난다고 가정해 볼 수 있을 것이다. 그러나 멘델 유전학과 분자유전학 사이의 관계를 그리 깊이 관 찰해 보지 않더라도 그러한 가정이 얼마나 순전한 발상인가를 곧 깨닫게 될 것이다. 가시적인 표현형질을 모두 분자적으로 규명된 형질로 바꿀 수는 있을지 모르지만, 멘델 유전학과 분자유전학의 서술용어들 사이의 관계란 여전히 말도 못하게 복잡한 것이고, 다 수대 다수의 관계로 표현될 것이기 때문이다. 한 개의 멘델적 서 술용어로 설명되는 현상도 실은 여러 개의 다른 분자 기작에 의 해 일어나는 현상일 수가 있다. 따라서 환원이 가능하다 하더라도 그것은 매우 복잡한 것이 될 것이다. 역으로 말하면, 같은 형태의 분자 기작이 만들어내는 현상도 여러 개의 멘델적 서술용어들을 사용해야만 설명될 수 있는 경우도 있다는 것이다. 결론적으로 말 해 환원이란 불가능하다. 후자의 경우, 죽 한 개의 분자 기작이
어떤 멘델적 서술용어와 연관된 것인지 분명치 않은 경우, 그 모 호함은 멘델 유전학을 계속 수정해 나가면서 제거가 될 수 있을 지도 모르지만, 그렇게 된다 해도 결국에는 또 다른 새로운 문제 에 부딪치게 된다. 그것은 이렇게 수정을 거듭해 나타난 최종 결 과가, 대체가 아니라 환원이라는 사실을 정당화시킬 수 있어야만 한다는 것이다 . 일단 멘델 유전학을 이런 식으로 변화시켜 나가면 무언가가 분자유전학으로 환원되긴 하겠지만, 분명 그것이 멘델 유전학은 아니다. 죽 멘델 유전학이 분자유전학으로 환원되는 것 이 아니라는말이다. 멘델적 서술용어와 분자 기작 앞의 논의에서는 가시적인 표현형질이란 것이 분자유전학적 분 석에 적합치 않다는 것을 보여주었다. 멘델 유전학에 있어서, 어 떤 이형집합체가 두 동형집합체의 중간 형질을 나타내는 경우, 그 이형집합체의 두 대립유전자는 불완전우성이라고 한다. 그러나 분 자 수준에서 본다면 이것만으로는 충분하지가 않기 때문에 또 다 른 종류의 구분 방식이 첨가되어야 한다. 앞에서 우리는 검은 안 달루시안 닭과 흰 안달루시안 닭을 교배하여 청회색의 새끼를 얻 는 것을 불완전우성의 예로 들었다. 이러한 불완전우성의 형질은 여러 가지 있을 수 있는 분자적인 상황 중의 어느 하나로 인해 생긴 결과라고 할 수 있을 것이다. 청회색의 색소가 만들어지는 것일 수도 있고, 검정색소만이 만들어지는데 그 양이 줄었기 때문 일 수도 있고, 검정색과 흰색 색소가 똑갇은 양으로 만들어지기 때문일 수도 있다. 또한 이러한 색소가 깃털 전체에 고르게 분포
될 수도 있고, 같은 색끼리 작은 덩어리를 이뤄 여기저기 모여 있을 수도 있다. 후자의 경우, 그것이 회색으로 보이는 것은 보는 사람의 시각이 얼마나 정확한가에 달린 문제이다. 위에서 제시된 여러 상황 중에서 어느 것이 과연 맞는 것인가에 따라 형질은 여 러 개의 멘델적 범주 중 어느 하나로 분류될 수 있을 것이다. 만 일 검정색소만이 만들어지는데 그 양이 줄어든 것이라면 형질전 달의 양식은 불완전우성이 맞을 것이며, 만일 검정색과 흰색 색소 가 모두 만들어전다면 그것은 불완전우성이 아니라 공우성의 한 예가 될 것이다 . 그런데 실제로 안달루시안 닭의 색깔이라는 하나 의 형질은 공우성이리는- 양식을 통해 유전되는 것으로 밝혀졌다. 따라서 두 대립유전자가 모두 완전히 그 기능을 발휘하여 검정과 흰색 색소 모두가 만들어지며 이 두 색소가 각각 작은 덩어리를 이루어 깃털의 여기저기에 분포되어 있는 것이다. 위에서 언급한 예의 요점은(지면이 허락된다면 여러 가지 다른 예도 둘 수 있을 것이다), 멘델 유전학이 분자유전학으로 환원되 려면 멘델 유전학의 전형적인 형질들이 분자적으로 규명된 형질 로 바뀌어야 한다는 것이며, 그러한 변화는 전반적인 재편성을 필 요로 한다는 것이다. 수많은 현상들이 멘델 유전학에서 열성 상위 리는 용어로 설명되고 있는데, 예를 들자면 쥐의 털색깔, 플리마 우스 록 Pl ym ou t h Rock 이나 레그혼 Le g horn 갇은 닭의 깃털 색 깔, 어떤 돌연변이로 일어나는 귀머거리 현상(인간에게 있어) 등 울 들 수 있다. 이 다양한 현싱들이 모두 한 가지의 분자 기작을 통해서 일어난다고 보기는 어렵고, 최소한 몇 종류의 기작이 관련 되어 있다고 보아야 할 것이다. 역으로, 이들 여러 가지 분자 기 작들이 항상 〈열성 상위〉라는 멘델적 현상을 만들어낸다고 보기 도어렵다.
물론 분자유전학이 〈 파랗게 보이는 눈 〉 과 같은 가시적인 표현 형질을 다룰 수 없디는 의견에 반대하는 사람도 있을 수 있다. 그러한 표현형질들은 모두 〈 파란색 색소가 있음 〉 또는 그 비슷한 분자적 용어로 바뀌면 된다는 것이다. 그러나 분자 수준에도 역시 여러 어려운 문제들이 놓여 있다. 예를 들어, 여러 가지 분자 기 작들을 통해 〈불완전우성〉 같은 분자적 현상이 만들어질 수 있으 나, 문제는 이러한 기작들이 불완전우성뿐 아니라 다른 현상도 만 들어낼 수 있다는 데 있다. 예를 들어, 안달루시안 닭의 색깔이 청회색인 이유가 검정색소의 양이 줄었기 때문이라고 가정한다면, 이 말은 청회색 닭이 갖고 있는 검정색소와 검정색 닭의 검정색 소는 똑같은 것인데 다만 청회색의 경우에는 그 양이 줄어들었다 는 뜻아 된다. 그렇게 양이 감소된 데에는 여러 가지 원인이 있 울 수 있을 것이다. 반응에 필요한 효소의 농도가 절반으로 감소 되었기 때문일 수도 있고, 검정색소에 대한 전령 RNA 분자가 절 반만 만들어졌기 때문일 수도 있고, 효소의 농도와 전령 RNA 의 농도에는 변화가 없으나 그 효소의 기능아 저하되었기 때문일 수 도 있다. 마지막의 경우는 효소나 전령 RNA 의 구조에 약간의 변 화가 생겼기 때문인지 모른다. 우리는 이런 식으로 이야기를 계속 전개해 나갈 수 있는 것이다. 반면에 한 가지의 분자 기작이 여러 가지의 디론· 표현형적 효 과를 일으킬 수도 있다. 예를 들어, 이형접합체의 한 대립유전자 는 완전한 기능을 하는 반면 다른 하나는 기능을 전혀 하지 못하 는 경우도 있을 수 있다. 이 경우, 대립유전자의 성질과 생합성 경로에 따라 유전자가 나타내는 효과가 달라전다고 볼 수 있는데, 정상적인 반응을 유지하는 데 필요한 산물 모두가 한 개의 대립 유전자에 의해 생산되는 경우가 있다면, 우리는 이런 현상을 〈우
성 〉 이라고 부를 수 있을 것이다. 한 개의 대립유전자만 작동하는 것은 변함없으나, 만일 그 반응 속도가 약간 또는 절반으로 감소 된다면, 그런 현상을 〈 불완전우성 〉 이라고 이름 붙일 수 있을 것 이다. 따라서 분자적인 수준으로 내려온다 하더라도 멘델적인 서 술용어와 분자적 기작 사이의 관계는 역시 다수· 대 다수가 될 수 밖에 없는 것 같다. 위에서는 한 단계의 반응만을 다루었으나, 앞에서 언급한 바와 같이 대부분의 구조 단백질은 건 생합성 경로를 통해 생산된다. 이러한 반응 과정 중 초기 단계에서 어떤 변화가 생기면 그것은 그 이후의 모든 과정에 영향을 미치게 된다. 뿐만 아니라 생합성 의 초기 경로는 여러 경로가 얽혀 있는 경우가 많아서 어떤 변화 로 말미암아 사용되지 못한 기질은 다른 경로에 사용될 수도 있 는데, 그렇게 되면 동일한 산물이 다른 경로를 통해 만들어지게 된다. 상위를 설명하는 한 가지 기작으로서, 상위유전자는 제 1 유 전자보다 생합성 경로의 초기 단계에 작용하는 것이라고 생각되 었다. 하나의 유전자가 다른 유전자를 억제하는 것이 아니고, 상 위 유전자가 생합성 경로에 필요한 선구 물질을 생산하여 그것을 제 1 유전자에 의해 조절되는 단계로 넘겨준다고 보아야 한다는 것 이다. 그러면 백화증 alb ini sm 돌연변이가 왜 그리 혼하고, 왜 단 한 개의 유전자를 가려내어 이것이 바로 백화증을 일으키는 유전 자다라고 말할 수 없는지 쉽게 이해할 수 있게 된다. 일반적으로 색소가 생산되려면 생합성 경로의 여러 과정을 거치게 되는데, 그 많은 단계 중 어느 하나만 억제되어도 색소는 생산되지 못하게 되며 그래서 그 개체는 알비노가 된다. 따라서 생합성 경로의 초 기 단계에 관여하는 유전자 좌위 loc i는 어느 것이나 알비노를 만 들 수 있는 가능성을 지니고 있는 것이다.
이제까지 얘기한 복잡한 문재들을 원래의 멘델적 서술용어와 결합시키려고 한다면, 유전학에서의 환원을 지지하는 사람들이 부 딪치는 그 많은 문제들울 인식해야 할 것이다. 열성 상위나 우성 상위를 나타내는 기작들을 모두 열거해야 하는 것은 물론이고, 우 성과 열성의 형질들이 서로 대칭적인 것처럼 열거된 두 기작들도 서로 대칭이 되어야 한다 . 논리적 경험주의자나 그 계승자들이 제 시하는 최신의 환원 분석법에 따르-면 환원되는 이론의 현싱들을 다시 고쳐서 재분류를 해야 하는데, 이 경우 멘델 유전학의 기본 적인 사항들이 아예 무시되거나 그 기본을 완전히 분해하고 다시 결합시켜서 전혀 다른 형태의 새 것으로 만들어야 하는 것처럼 보인다. 스트릭버거 M. W. S t r i ckber g er 는 다음과 갇이 말한 바가 있다 . 초기 문헌에서는 유전자의 활동과 상호작용에 대해 알려진 바 가 거의 없었다. 대부분의 유전학자들은 하나의 특정 유전자는 다른 유전자와 상관없이 독립적으로 작용하여 그 표현형적 효과 롤 나타내는 것으로 생각했었다. 따라서 한 유전자가 다른 유전 자의 효과를 변화시키고 수정하는 경우를 발견하는 일은 언제나 대단한 관심거리가 되었다. 유전자간의 상호작용이 일어나는 여 러 예들이 발견됨에 따라, 그러한 상호작용들을 묘사하기 위해 수많은 용어가 생겨났는데, 그 중에서 보족유전자(補足 遺 傳子)와 상위 및 앞에서 언급했던 그 의의 몇 가지 용어들은 아직도 사용 되고 있다. 이것은 특정 유전자의 표현형적 행동을 눈으로 볼 수 있게 하는 데 도움이 되기 때문이다. 물론 우리가 눈으로 관찰하 는 표현형이란 실제로는 우리가 상상하는 것보다 훨씬 더 복잡한 과정을 거쳐 만들어지는 결과이다. 예를 들어 상위라는 현상은
상위라는 단어의 뜻처럼 한 유전자가 다른 유전자를 가리기 때문 에 생기는 것이 아니라, 효소 활동이 증가하거나 감소해서 생기 는 것일 수도 있고, 색깔에 영향을 주는 산도(酸度)의 변화 혹은 그의의 여러 가지 다른 복잡한 반응들 때문에 생기는 것일 수도 있다. 전체적인 표현형질 수준에서 유전자에 관해 우리가 어떠한 용어를 사용하든 간에, 우선 그 유전자의 작용에 대한 발생상의 기초적인 사항들이 먼저 연구되고 밝혀져야 할 것이다 .13 ) 하나의 분자 기작이 여러 가지 표현형적 효과를 일으킬 수 있 다는 의견에 대해 반박하는 사람이 있을지 모르지만, 그러한 일 대 다수의 효과는 다만 관련된 여러 가지 요소들, 죽 온도나 수 소이온농도 등을 생각하지 않았기 때문이다. 일단 이러한 요소들 이 다 고려된다면 분자유전학과 표현형 사이의 일 대 다수의 관 계는 모두 일 대 일의 관계로 바뀔 것이다. 이러한 일들이 가능 하다고 믿는다는 것은 다시 말하면 결정론적 인과율의 기본 원리 를 고친다는 것과 같다. 결정론적 인과율에 따르면, 원인이 같으 면 늘 같은 결과가 나타나야 하고 결과가 달라졌으면 그 원인이 반드시 달라야 한다. 그러나 우리는 분자 기작의 개념이 이제 얼 마나 팽창되었는가를 한번 생각해 보아야 한다. 우리는 이제 멘델 적 서술용어와 분자 기작을 일치시키려고 하는 것이 아니라 멘델 적 서술용어와 분자론적 상황 전체를 일치시키려 하는 것이다. 자 신을 환원주의자라고 생각했던 생물학자들과 자신을 생체론자(生 體論者)탸고 생각했던 생물학자들의 차이점은 바로 이러한 점에
13) Geneti cs (New York: Macm illa n and Co., 1968), p. 202. ® 생체론(生體論, orga n ici s m ). 생명현상이라는 것은 생명체의 구성 단위 또는 구성 물질의 물리적 특성이나 화학적 성질만으로는- 설
명될 수 없고, 이들 단위 물질 또는 단위 구조의 총체적 조직에서 나타나는 한 단계 높은 차원의 종합적 성질이라는 견해이다. 따라 서 생명물질론 또는 기계론적 입장도 아니고 생기론적 입장도 아 니다. 생명체의 기계론적 해석은 물론 철대 필요하지만 그 지식으 로는 전체를 조화시키고 전체로서의 통일된 기능을 나타내는 힘은 이해할 수 없다는 것이다 . 따라서 생명현상 연구에서는 물리화학적 법칙성 이외의 또 다른 생명 특유의 법칙성을 발견해야 한다고 한 다. 그러나 이 생명 특유의 법칙 또는 힘은 생기론에서와 갇은 초 인간적인 그리고 초자연적인 어떤 절대주의 힘이 아니고 물질론적 바탕에 입각한 합리적 힘이라고 생각하고 있다.
있을 것이다. 환원주의자들은- 분자 구조와 기작을 더욱 구체화시키 는 것이 모든 유전 현상을 설명하는 길이라고 생각한다. 그렇다면 다소 복잡하간 해도 전체적으로 보면 환원이란 어쨌든 상당히 단순 한 작업이 될 것이다. 한편 생체론자들은- 분지론적 상황 전체를 참 조해야 한다고 강조하는데, 정말 그렇게 해야 한다면 환원주의자들 이 세운 프로그램이 과연 실현 가능할 것인지 의문시된다.
유전학에 있어서의 환원-환원인가? 대체인가? 우리는 논리적 경험주의자들에 의해 일반화되었던 환원론과 이 에 반대하는 의견 때문에 취해전 변화를 이야기함으로써 이 장을 시작했었다. 그리고 그러한 환원주의자들의 프로그램에 대항하기 위하여 생긴, 보다 근본적이고 〈 철학적인 〉 반대 견해들도 알아보 았다. 이러한 반대 의견에 대한 논의는 맨 마지막 장으로 미루고, 여지껏 우리는 논리적 경험주의자들이 사용하는 전통적 방식을 사용해서 바로 그들의 환원에 대한 분석을 비판해 보았다. 논리적
경험주의자들에 따르면, 환원이란 재구성된 이론들 사이에 존재하 는 형식적인fo rmal 관계이댜 적당한 환원 함수가 만들어지고 수 정이 가해지면, 환원하는 이론으로부터 환원된 이론이 유도될 수 있다. 무엇을 설명하고자 할 때와 마찬가지로 이렇게 유도된 환원 도 추론에 의하여 분석된다. 이 장에서 우리는 유전학의 경우, 그 러한 유도 과정이 실제로 얼마나 어려운 일인가를 살펴보았다. 여 기에 멘델 유전학을 끌어들인다면 분자적 용어와 멘델적 용어 사 이의 관계는 다수 대 디수가 되는데, 이러한 다수 대 다수의 관 계를 일 대 일 혹은 다수 대 일의 관계로 바꾸기 위해서는 상당 한 변화가 멘델 유전학에 가해져야 한다. 이때 두 가지의 문제가 새롭게 대두되는데, 그 중 하나는 이러한 변화를 〈수정〉이라고 부를 수 있느냐는 것이고, 다른 하나는 멘델 유전학이 분자유전학 으로 변하는 것이 〈대체〉가 아니라 정말 〈환원〉에 해당하는가 하는것이다. 그 다양한 여러 생화학적 기작들이 멘델 비에 영향을 주지 않 는 한, 형질전달유전학자들은 형질의 근원이 되는 생화학적 기작 그 자체에는 관심을 두지 않는다. 생화학적 경로에 대한 지식이란 그들에게 아무런 쓸모가 없는 것이다. 그러나 환원 함수가 형질전 달유전학의 용어와 분자유전학의 용어를 서로 연관시킬 수만 있 다면, 중요한 생화학적 차이점들은 형질전달유전학에서도 그 모습 을 드러내게 될 것이다. 만일 동일한 분자 구조가 두 개의 서로 다른 경로를 통해 만들어진다면, 그러한 차이점은 적절한 환원 함 수 속에 반영될 수 있을 것이고, 그에 따라 멘델 유전학에도 적 절한 변화가 일어나게 될 것이다. 그러나 이러한 변화를 〈수정〉 이라고 부르는 것이 다소 이상하게 느껴지는 것은 사실인데, 그 이유는 수정이라는 단어가 지니고 있는 일반적인 의미와는 달리,
형질전달유전학이 그 전통적인 작업을 수행하는- 데 있어 부정확 했다거나 적철치 못했던 것이 결코 아니기 때문이다. 형질전달유 전학의 원리나 기술이 다른 분야에 적용될 수도 있고 형질전달유 전학의 해결 능력이 부족하다고 볼 수도 없다면, 형질전달유전학 은 아주 훌륭하고 타당성 있는 이론이라고 할 수밖에 없다. 실은, 단지 멘델 유전학을 분자유전학으로부터 연역해 보고자 할 때에 한해서만 약간의 변화가 요구될 뿐이다. 논리적 경험주의자들의 환원 분석법에서, 환원되는 원래의 이론 을 수정해야 하는 일은 아주 필수적인 작업이다. 그러나 얼마나 많은 수정이 가능한 것인지, 또 수정이 되고 나서도 어떻게 〈 동 일한〉 이론이라고 부를 수 있는 것인지 의문이 생긴다. 극히 정 교한 착상을 통해 열에 대한 연소 이론이 수정될 수 있었고, 그렇게 수정이 되었기 때문에 그 이론이 현대 통계역학으로부 터 유도될 수 있었던 것은 사실이지만, 과연 누가 그러한 이른바 〈환원〉이라는(대체가 아니라) 작업에 감명을 받겠는가? 이에 대 해 논리적 경험주의자들이 제시한 답은, 환원의 경우에는 원래의 이론과 수정된 이론 사이에 상당한 유사점이 있어야 한다는 것이 었다. 하지만 유전학의 경우에는 그 두 이론이 서로 비교될 수조 차 없는 것처럼 보인다. 먼저 형질전달유전학을 분자유전학으로부 터 연역하기 위해서는 어느 정도의 재구성이 필요한가를 생각해 보자. 이 경우, 연역을 위해 필요한 재구성의 정도란 것은 실로 엄청나서, 마치 전통적인 멘델 유전학과 재구성된 형질전달유전학 사이의 유사성을 완전히 말살시키는 것처럼 보인다. 원래 유사성 이라는 것이 환원과 대체를 구별하기 위해 도입된 개념이며, 또 그 개념 자체가 잘못 분석되었기 때문에 이러한 갈등은 결코 해 결될 수가 없었다(비슷한 예로서 정신mi nd 이라는 것을 뇌의 어떤
상태라는 말로 환원하는 것을 둘 수 있을 것이다. 그러나 이 경우 에도 심리학적 이론이나 생리학적 이론, 그 어느 것도 정신과 뇌를 정확하게 비교할 수 있을 만큼 충분하지 못하다). 이러한 논의가 설득력이 있다면, 논리적 경험주의자들은 딜레마 에 빠지게 될 것이다. 논리적 경험주의자들이 주장하는 환원에 대 한 분석이 옳다면, 멘델 유전학은 결코 분자유전학으로 환원될 수 가 없다. 생물학의 이론을 물리학이나 화학적인 이론으로 환원시 킨다고 할 때의 그 환원은, 실제로 〈 환원〉이 아니라 〈대체〉의 한 예로 판명되었기 때문이다. 그러나 분석하기에 앞서 우리의 직관 적인 통찰에 의한다면 그것은 분명 환원의 한 예가 될 수 있는 것 같다. 그러나 만일 생물학자들이 멘델 유전학을 분지유전학으 로 환원하고 있는 중인데도 논리적 경험주의자들의 분석 방법이 여기에 적용될 수 없다면, 논리적 경험주의자들의 분석 방법을 옳 다고 볼 수는 없다. 여기서 중요한 문제는 과연 환원이라는 것이 과학자들이 실제로 하고 있는 어떤 〈일〉이냐, 아니면 이러한 노 력으로 생겨난 산물에다가 어떤 사후의 행동을 가하고 있는 것인 가 하는 일이다. 관찰해 본 바에 의해 내가 아는 한은, 형질전달 유전학의 원리들을 분자유전학의 원리로부터 유도해 내려고 하는 생물학자들은 없다 . 논리적 경험주의자들의 분석에 따르면, 그것 이야말로 바로 생물학자들이 해야 할 일인데도 말이다. 이 장에서 필자가 분석해 온 것이 옳았다면 결론은 자명하다. 즉 유전학의 경우에는, 논리적 경험주의자들의 분석이 크게 도움 이 되지 못했다는 것이다. 오히려 그것은 멘델 유전학과 분자유전 학 사이의 관계를 이해하는 데 방해를 했다고 생각한다. 아무리 극소화시킨다고 해도 두 이론간의 관계는 너무 복잡하고, 정밀한 유도 작용을 위해 필요한 재구성도 엄청난 것이어서 멘델 유전학
을 분자유전학으로 환원시키려는 노력은 그러한 노력을 기울일 만큼의 가치가 없는 것 같다. 따라서 현재 그러한 유도를 해내기 위해 애쓰는 사람은 아무도 없고 그래야 할 이유도 찾을 수 없 다. 이 장에서 제시했던 구체적인 분석들은 결코 쉬운 일이 아니 다. 왜냐하면 그러한 분석이 실현 불가능한 일들을 다루고 있기 때문이다. 그러나 그러한 분석에 주어졌던 명제들이 옳지 못하다 는 것을 증명하는 일은, 비록 어렵건하지만 의심의 여지없이 확고 한 결론을 이끌어낼 수가 있다. 그러기 위해서 해야 할 일이란 멘델 유전학을 분자유전학으로부터 도출해 내는 데 필요한 환원 함수를 제시하는 것, 환원 함수를 통해 그러한 도출 작업을 실제 로 해보고, 수정된 멘델 유전학과 수정 전의 원래의 유전학이 상 당한 유사점을 갖고 있는지 분석하는 일이 전부이다 .14)
14) S더im o상n(세19한71 )내, 용an에d 관D.해 H서u는ll, MR. eRduuscet,i on R eind u gctein oent ,i cRs —e p l-a Bcie om l oegn yt and Molecular Bi ol og y, Di ale cti ca , 25(1 9 71), pp. 39-72 ; M. or Ph ilo sop h y ? Phil o sop h y of Scie n ce, 39(1 9 72), pp. 491-499 를 참고할것.
잭 진화론의 구조 과학적 이론, 법칙 및 정의 앞 장에서 우리는 멘델 유전학이 분자유전학으로 〈환원〉되는 현상에 대해 살펴보았다. 이 장에서는 생물학적 이론들 중에 두번 째로 중요한 것이라 할 수 있는 전화론으로 우리의 관심을 돌려 보려고 한다. 고전적안 멘델 유전학, 현대의 형질전달유전학 그리 고 분자유전학 사이에 논란의 여지가 없이 확실한 경계선을 긋는 것이 쉽지 않은 것처럼, 전화론의 여러 단계들과 여러 해석들 사 이에 경계선을 긋는 일 또한 쉽지 않다. 그러나 필자는 이 책이 목표하는 바에 따라 진화론의 발달 과정을 고전전화론, 유전학적 진화론 및 종합 전화론의 세 단계로 구분하고자 한다. 고전전화 론은 대충 다윈 Charles Dar wi n 이 평생 동안 이룩해 놓은 바로 그것이라고 말할 수 있고, 유전학적 전화론은 금세기 초 약 30\ :i 간에 걸쳐 유전학자들과 수학자들에 의해 만들어전 이론이라 할 수 있다. 이 유전학적 진화론은 수학적인 표기법을 사용하며 전화
를 유전자와 유전자형의 빈도라는 관점에서 다루고 있다. 유전학 적 진화론자들이 유전자에 대해서만 관심을 기울이고 있다는 사 실을 특별히 강조하고자 하는데(다소의 과장이 있을지도 모르겠으 나), 이는 현대의 진화론자들이 직면하고 있는 문제를 유도해내기 위해서이다. 죽 다윈의 표현형적인 해석을 유전학적인 해석과 접 목하여 종합하기 위해서라고 할 수 있고, 따라서 이러한 시도를 종합 진화론이라고 이름 붙일 수 있을 것이다 . 필자의 이러한 구분 방식 의의 또 다른 방식들도 있을 수 있겠으나 이 책의 성 격상 위의 구분 방식이 그래도 가장 도움이 될 수 있으리라고 믿 는다. 처음부터 철학자들과 과학자들은· 진화론을 과학 이론의 하나로 간주하는 일을 탐탁치 않게 여겼다. 그래서 전화론에 반대하는 사 람들이 무수히 많았는데, 그들에 따르면 진화론은 잘못된 것이며 반증도 할 수 없다는 것이었다. 전화론의 가장 기본적인 원리 중 하나인 잘 적응된 생물체가 덜 적응된 생물체보다 더 성공적으로 번식하고 생존한다는 주장은 같은 말울 되풀이하고 있는 듯한 인 상을 주었고, 논리적인 형태를 판단하기에는 전화론의 구조가 너 무 엉성하다는 것이었다 . 전화론에 의하면 진보가 필연적으로 일 어나는 것으로 되어 있으나, 고등한가 그렇지 않은가를 판단하는 일은 배제되고 있다. 또한 전화론은 어떤 일련의 사건에 대한 미 래를 예측할 수 있도록 그 근거를 제공하는 것이 아니라, 일단 일어난 사건을 설명하는 데 사용될 수 있을 뿐이라는 것이다. 전 화론에 대한 이러한 반대 견해는 대부분이 전화론의 간단한 특칭 에 대한 무지 또는 특칭을 무시하는 데서 연유한다고 할 수 있으 나, 그러한 반대 견해 중의 몇몇은 과학철학의 근본적안 문제와 관련이 깊은것도 있다.
이 장에서는 전화론의 논리적 구조를 살펴보고 이러한 논리적 구조를 철학 문헌 내의 과학 이론에 대한 전통적인 분석과 비교 해 보려고 한다. 진화론의 형태에 어떤 특별한 점이 있는가? 그러 한 증거는 아마 찾을 수 없을 것이다. 그러나, 만일 있다면 과연 전화론의 구조가 예측하고 검증하는 등의 일을 배제하고 있는가? 이러한 질문이 단순히 학문적인 것이라고만 단정할 수는 없다. 왜 냐하면 궁극적으로 인류의 미래가 이러한 질문을 해결할 수 있는 가의 여부에 달려 있기 때문이다. 생태학은 단기적 전화를 연구하 는 학문이다. 만일 인간을 포함하여 모든 전화적 변화가 원래부터 예측 불가능한 것이었다면 생태학, 인구 폭발 그리고 우생학 같은 것도 모두가 공허한 논쟁이 될 것이다. 만일 어떤 종의 미래를 예측할 수 있는 방법이 전혀 없다면, 인류의 미래도 예측할 수 없는 것이다. 우리가 갑작스런 종분화 과정에 있는지 점전적인 진 화의 과정에 있는지, 갑작스럽게 절멸하는 과정에 있는지 누가 알 수 있을 것인가? 이 장의 상당히 많은 부분이 이러한 문제에 할 애되겠지만, 우리의 일차적인 관심사는 과학 이론의 전통적 분석 법과 전화론의 차이점 및 전화론의 형태라는 것임을 잊지 말아야 할것이다• 현재 보편적인 과학 이론의 패러다임은 대개 연역적으로 연관 된 일련의 법칙들의 구조인데, 이 법칙들은 경험적 실체와 진행 과정에 대한 보편적인 진술이어야 한다. 그래서 〈모든 백조는 희 다〉 또는 〈알비노 쥐는 항상 알비노 쥐를 낳는다〉와 같은 식의 전술이어야 한다. 과학법칙들이 보편적인 형태이어야 하는 것처 럼, 이러한 법칙 내의 각 집단들은 일련의 성질에 의해 정의되어 야 하는데 그 성질들은 각각이 그 분류군의 한 구성원이 되는 데 필수적인 것이면서 그것들이 합해질 때는 그러한 구성원이 되기
에 충분한 것이어야 한다. 어떤 특정 집단에 대한 정의가 주어졌 울 때, 그 집단의 각 구성원은 그 집단에 대한 정의 속에 나열되 어 있는 성질을 모두 갖추고 있어야 하고 그렇게 모든 성질을 지 니고 있는 각 개체는 필수적으로 그 집단의 구성원이어야 한다. 예를 들면, 모든 삼각형은 세 변으로 되어 있고 그 세 변이 다 닫혀 있는 도형이며 따라서 이러한 도형은 어느 것이나 다 삼각 형인 것이다. 그러나 어떤 학자들은 과학 이론의 구조가 연역적인 이상(理 想)에 미치지 못하는 경우에도 때로는 그것을 용납하여야 한다고 한다. 그러한 경우 그 이론 속의 법칙들 또한 보편적인 형태를 지니고 있지 못하는 경우도 있을 것이다. 그들에 따르면 통계적인 법칙들, 심지어는 추정하는 일이나 어떤 경향, 그리고 어떤 경향 울 띤 전술들조차도 하나의 과학법칙으로 간주될는지도 모른다. 또 어떤 학자들은 각각은 필수적이면서 결합될 경우 충분조건이 될 수 있는 일련의 조건이라는 전통적인 이상형에서 크게 벗어난 정의조차도 묵인한다. 전통적인 정의(定義)의 개념이 요구하는 그 러한 보편성은 없더라도 자주 함께 취급되는 속성으로 이루어진 정의라면 충분하다고 하는 듯하다. 예를 들어, 시간적으로 연속되 어 온 계보로서의 종의 이름은 한 집단이라는 개념으로만 정의될 수 있을 뿐이다. 즉 한 종을 다른 모든 종과 구별하는 특칭이 그 종의 구성원들 사이에 보편적으로 분포되어 있는 경우란 없으며, 그 역도 마찬가지이다. 기껏해야 한 종의 구성원이 되느냐 못되느 냐 하는 것은 그 종에 대한 정의 속에 나열되어 있는 특성들 중 에서 가장 중요한 것들을 충분히 지니고 있는 개체에 의해 결정 되는 것일 뿐이다 (Beckner, 1959 참조). 따라서 스펙트럼의 한쪽 끝에는 연역적으로 구성된 이론이 있
다고 할 수 있는데, 이러한 이론들은 전통적인 방법으로 정의된 형식과 용어로 이루어전 보편적인 법칙만을 포함하고 있다. 다른 한쪽은 귀납적으로 구성된 이론으로 되어 있는데 이 이론들은 통 계적인 차이를 보이는 다양한 특성들로 이루어전, 말하자면 덜 보 편적인 전술과 용어로 되어 있다고 할 수 있다. 이 중 전자의 이 론 및 정의가 지닌 특칭은 논리학자들이나 철학자들에 의해 상세 히 연구되어 왔다고 볼 수 있는 데 비하여 후자의 정확한 속성에 대해서는 아직도 논란이 많이 일고 있다. 우리가 이 장에서 다루 려고 하는 문제는 법칙과 정의의 형태 모두에 있어서 전화론이 연역적 이상에 근접할 수 있겠느냐는 것이다. 만일 그것이 불가능 하다면 그것은 전화론이나 현재 받아들여지고 있는 과학 이론과 정의가 모두 옳지 못하다는 사실을 반영하는 것이라 할 수 있을 것 0] 다. 과학 이론으로서 필요한 또 하나의 전통적인 조건은 그 이론을 구성하고 있는 법칙들이 시간적으로나 공간적으로 제한이 되어서 는 안 된다는 것이다. 그렇다고 시간적 또는 공간적 다양성이 과 학법칙에서 제의되어야 한다는 뜻이 아니라(그의는· 반대로 그러한 다양성은 과학 전반을 통하여 아주 중요한 역할을 하고 있다고 보 아야 할 것이다), 오히려 과학법칙이 필연적인 어떤 특정한 시간 이나 우주 내의 어떤 특정한 공간에 국한되어서는 안 된다는 뜻 이다. 철저히 폐쇄된 계(系) 속에 존재하는 행성이 있다면, 그 행 성들에만 국한된 과학법칙이 있을 수 있고 또 어쩌면 단 한 개의 별을 중심으로 회전하는 유일한 행성이 실제로 존재할 수도 있다. 그러나 화성, 그것도 예컨대 1935 년 6 월 15 일의 화성만을 다루는 과학법칙이란 있을 수 없는 일이다. 과학법칙이 하나의 사건만을 다平큰 경우는 있을 수 있으나 이 경우 그 사건이란 것도 실제로
는 같은 부류의 사건 전체를 뜻한다. 죽 과학법칙은 절대로 유일 무이한 사건을 다룰 수는 없는 것이다. 보통 유일무이한 사건이라 는 것도 실제로는 같은 부류에 속하는 일련의 사건 전체를 지칭 하는 것이다. 전실로 유일무이한 사건이란 바로 그 사건 자체를 제의하고는 이러한 사건에 속하는 또 디론 경우가 전혀 없다는 뜻인데, 이런 사건은 특정 공간 및 특정 시간에 일어난 사건이라 고 명시되어야 한다. 왜냐하면 두 개의 대상이 동시에 동일한 공 간에 존재할 수는 없기 때문이다. 대부분의 철학자들은 시간과 공간의 연장 ex t ens i on 과 연속성 con ti nu it쩌 가지고 〈 개체 〉 의 개념을 정의하는데, 그러한 정의는 개체라는 단어 자체의 일상적인 의미와 아주 잘 일치하고 있다. 보통 우리가 개체라고 부르는 것들은 공간적으로 그리고 시간적 으로 연속되고 있는 존재들이면서 동시에 상당히 명백한 경계선 울 가지고 있다. 그러므로 개체라는 것을 공간적 시간적 실체라고 정의한 이상, 과학법칙은 특정 개체를 다루는 것이 아니라 그 개 체들의 집단을 다루는 것이라고 해야 할 것이다. 그러나 독자는 여기서 〈개체〉라는 것이 어떤 절대적인 의미로 사용되고 있지 않 다는 것에 유의해 주길 바란다. 인류라는 종으로서의 우리는 보통 하나의 개체를 어떤 복합적인 존재로 생각하는 데 익숙해져 있다. 다세포 생물은 우리의 감각으로 인지할 수 있을 만한 크기이고 또 충분히 긴 시간을 변형되지 않고 존재하기 때문에 우리는 그 것을 개체 pa radig m i n di v i dua 토 간주하지만, 최소한 과학자들은 그래서는 안 될 것이다. 과학자는 생물 개체를 하나의 집단, 경우 에 따라서는 세포의 집단으로 간주하여 이론을 구성할 수도 있고, 또 각 생물 개체들을 생물학적 종의 한 부분이라고 간주하여 이 론을 구성할 수도 있다. 진화론의 역사를 통해 가장 지배적이었던
문제는 전화 법칙에 잘 들어맞는 전화 단위를 발견하는 일이었 다. 그러나 생물학자들은, 개체라는 것은 어떤 단계에서 분석을 하든간에一―유전자, 염색체의 한 부분, 염색체 전체, 유전자형, 개체, 멘델 집단, 생물학적 종 등, 어떤 수준에서 분석하여도 진화에 있어서 하나의 선택 단위가 된다는 것을 잘 알고 있다 .I)
1 ) Mary B. Wi lliam s, The Log ica l Sta t u s of Natu r al Selecti on and Ot he r Evoluti on ary Contr o versie s : Resoluti on by an Axiom ati zat i on , The Meth o dolog ica l Unit y of &ienc e(Dordrecht : D. Reid e l, 1 ':17 3) 을 참조.
그러나 여기서 내가 말하고자 하는 요지는 과학자들이 그들의 이 론 속에서 말하고 있는 개체라는 것이 어떤 실체이든간에 그러한 개체 하나하나가 그 이론 법칙 속에서 기능을 발휘하고 있는 것 도 아니고, 단지 그러한 개체들로 구성된 집단이 그렇게 할 수 있을 뿐이라는 것이다. 예를 들어, 만일 어느 한 전화론이 지칭하 고 있는 〈 개체 〉 란 것이 특정 염색체일 경우, 그 이론 내의 법칙 은 예컨대 어떤 성염색체 전체를 얘기하고 있는 것이지 어떤 특 정 세포 내의 특정 성염색체만을 지칭하는 것이 아니라는 것이다. 그렇게 과학지들이 서로 의견의 차이를 보이는 것을 보면서 혹 자는 감각적 인 자료나 있는 그대로의 적나라한 모습의 사건과 갇 은, 그러면서도 동시에 여느 과학 이론과도 무관한 어떤 실체적 개체에 의지하고 싶을런지도 모른다. 그러나 그러한 개체들의 역 사를 살펴보면 그러한 충동은 결코 매력적인 것이 못된다는 것을 알게 된다. 오히려 그 개체와 연관이 있는 과학법칙들에 의존하면 서 여러 단계에 걸쳐 그 개체성을 인식해 볼 필요가 있는 것 같 다. 일련의 법칙에 의하면 개체란 것이 어떤 복합적인 존재로 간 주될 수도 있고, 또 다론 일련의 법칙에 의하면 하나의 집단으로
간주될 수도 있다. 이러한 〈 개체 〉 는 모두 동일한 실체의 한부분 이기 때문에 각 단계를 분석하여 얻을 수 있는 어떤 시각과 견해 가 하나의 체계로 종합될 수 있어야 하고 그것을 디를 수 있도록 형식화된 그리고 전체를 포괄할 수 있는 하나의 이론이 존재해야 한다고 생각된다. 이것이 바로 과학의 통합이라는 개념인데, 이 개념은 그것이 과학의 유일한 목적이라는 독선적인 주장과 어떻 게든 통합을 해보려는 성급한 시도로 인해 불필요하게 그 명성에 손상을 입어 왔다. 과학 이론은 분석의 여러 단계마다 다 존재할 수 있다. 그리고 과학 이론은 그 이론 자체의 가치를 가지고 평 가되어야지, 그 이론이 환원시킬 수 있는 영역의 크기로서 평가되 어서는 안된다 . 끝으로, 전화론을 분석하기에 앞서 과학법칙의 속성에 대해 부 언할 것이 하나 더 있다. 현재 받아들여지고 있는 과학법칙들은 과정의 법칙 pro cess law 이라고 할 수 있는데, 다시 말해서, 어떤 한 특정한 시간에 있어서의 변수의 상태를 알면 그러한 지식으로 부터 시스템의 일반적인 상태, 죽 어떤 시간이 주어지든간에 그 시간에 해당하는 변수의 값을 추론할 수 있도록 하는 일반화의 과정이라고 할 수 있다. 과정의 법칙은 태양계와 같이 상관 변수 에 대하여 닫혀 있는 시스템에서만 가능하다. 어떤 특정 시간에 있어서의 태양계 내에 있는 각 천체의 운동량과 위치를 알고 천 체역학의 법칙을 알며 동시에 시스템이 닫힌 상태로 유지될 수 있다면, 다른 시간 죽 그 특정 시간의 전이나 후에 있어서의 각 천체의 위치와 운동량도 정확히 추론할 수 있을 것이다 . 하지만 천체와 관련된 문제들이란 너무나 복잡해서 동시에 두 개 이상의 천체를 다룰 수가 없고 또 다룬다고 해도 그 정확성은 엄청나게 떨어질 것이다. 더군다나 아주 거대한 혜성이 태양계를 지나가면
서 태양계 전체를 교란시키는 일이라도 일어난다면 미래를 예측 하여 해 놓은 계산은 모두 쓸모없게 될 것이다. 그것은 천체 법 칙이 잘못되었기 때문이 아니라 시스템의 폐쇄성이 붕괴되었기 때문 o] 다. 생물계에서는 충분할 만큼 오랜 시간 동안 유지되고 있는 페쇄 계란 거의 없다고 보아야 한다. 생합성 경로, 개체, 교배집단, 생 태계 및 종들 모두가 부분적으로 또는 때로는 필연적으로 열려 있을 수밖에 없는 시스템들이다. 그러나 이러한 어려움을 피할 수 있는 방법이 있는데, 그것은 사건과 사건이 일어나는 중간 단계에 서 생물계가 어떻게 움직이는가를 보는 과정의 법칙이 제시될 수 있다는 것이다. 태양계가 철저히 폐쇄되어 있을 때에 한하여 물리 적인 법칙이 태양계 내에 있는 천체의 위치를 추론해 낼 수 있었 던 것처럼, 생물 법칙도 시스템이 충분히 폐쇄되어 있는 경우에만 생물계의 상관 변수의 값을 구하는 데 사용될 수가 있다. 그렇다 고 하여 어떤 종이 존재하는 모든 기간 동안 그 법칙이 적용되어 야 한다는 것은 아니며 특별한 상황, 예를 들면 구성원 중의 어 떤 것들이 어떤 비어 있는 생태계에 들어갔다든가 혹은 그 집단 내의 한 개체가 그 종으로부터 생식적으로 고립되었다든가 하는 상황에 적용될 수 있으면 된다. 마찬가지로 생리적 법칙도 생명체 가 존재하는 전 기간 동안 반드시 적용되는 것이 아니라 발달 과 정중의 어느 특정 단계에 한해서만 적용될 수도 있는 것이다. 다윈의 주장과 진화론의 구조 전화론을 보다 적철하게 설명하기 위하여 진화 과정과 계통발
생 사이에, 또 전화론과 계통발생학적 기술(記述) 사이에 존재하 는 명백한 차이점을 강조해야 할 필요가 있을 것 같다. 전화 과 정은 종이 전화해 나가는 기작이라고 할 수 있고 계통발생은 그 기작에 의하여 생기는 산물이다• 헴펠 (Hem p el, 1965 : 370) 이 경 고한 바와 같이 우리는 진화론을 돌연변이나 자연선택의 기작에 관한 이론과 구분할 수 있어야 한다. 포유동물이 쥐라기(紀)에 원시 파충류로부터 생겨났다는 주장은 진화 또는 계통발생학적 이야기에 속한다고 할 수 있으나 진회론의 한 부분이라고는 할 수는 없다. 한편 종분화는 적절한 격리 기작이 있어야만 일어날 수 있디는 주장은 전화론에 대한 한 가지 해석에 해당하는 부분 이라고 할 수 있다. 계통발생학적 기술(記述)은 역사에 관한 이야 기라고 할 수 있지만 전화론은 역사의 이야기는 아니다. 또한 우리는 전화의 존재에 대한 다윈의 주장과 완전히 정립이 된 전화론을 구별할 수 있어야 한다. 진화의 존재에 대한 다윈의 주장은 이해하기가 쉽다. 생물은 모두 양적으로 많은 양의 자손을 낳는 경향이 있고, 만일 그 자손들도 모두 그런 식으로 번식을 한다면 지구는 단 한 가지 종만으로도 금새 넘치게 될 것이다. 그러나 이런 일은 분명히 일어나지 않는다. 어떤 종은 특정 계절 에 그 수가 증가하지만 곧 감소하게 되고, 또 어떤 종은 어떤 지 질학적 시대에 번성했으나 훗날 절멸되었다. 지구의 역사상 어느 한 시점에 존재했던 생물체들은 그 숫자에 크고 작은 변동이 있 었지만 생식적인 잠재력에 의한 급속한 팽창 같은 것은 일어난 적이 없다. 따라서 다윈은 생물체는 대부분 생식할 수 있는 시기 에 이르기 전에 죽는다는 결론에 도달하게 되었다. 여기에는 두 가지 가능성이 있다. 생물둘의 이러한 죽음은 무 작위적으로 일어나는 것이거나 아니면 최소한 부분적으로나마 그
러한 죽음이 생물 각 개체와 환경 사이의 관계로 인해 생긴 결과 일 것이다. 여기서 환경이란 물론 같은 종 내의 다른· 개체들이나 다른 종 모두를 포함하는 것이다. 많은 생물들은 다른 생물들과의 차이점과는 상관없이 그저 우연히 죽어간다. 예를 들어, 어떤 생 물은 어떤 특정한 특성을 지니고 있든 없든 간에 유충 때에 죽어 버린다. 그러나 어떤 경우에는 생물과 환경 사이의 관계로 인해 그 생존 기간에 차이를 보이는 것 같다. 오래 생존하는 것은 그 들이 다른 생물보다 환경에 더 잘 적응했기 때문이다. 생존 경쟁 울 하는 데 있어서 환경에 잘 들어맞는 생물들은 잘 적응하지 못 하는 것보다 더 높은 빈도로 생존하는 경향이 있다. 한 종내의 모든 개체가 동일하고 같은 환경에 처했을 때, 어떤 것은 살아 남아 자손을 만들고 다론 어떤 것은 그렇지 못했다면 그것은 우연의 문제일 것이다. 적응성은 왜 한 종은 존속하는 반 면 다른 종은 소멸되는가 하는 문제만을 설명하기 위해서 사용될 수 있다. 종 내부에 변이가 존재한다는 것은 종분화가 일어나기 위해서는 필수적인 일이다 . 한 종이 지니고 있는 특칭 중 어떤 것은 유전적이어서 한 세대로부터 다음 세대로 전달이 된다. 만일 한 종내에 어떤 변이가 있고 그것이 유전되고 갈은 방향으로 무 한히 변화를 계속하면서 여러 세대를 거친다면 그 생물은 접점 더 환경에 잘 적응하는 생명체가 될 것이다. 만일 그 종이 부딪 치는 환경에 변화가 없다면 그 종은 최적의 적응점에 도달하게 될 것이고 그 다음부터는 이 점을 중심으로 큰 변화를 보이지 않 게 될 것이다. 그러나 만일 환경적인 변이가 일어난다거나 시간이 흐름에 따라 환경도 변한다면 신종이 전화되어 나울 수 있을 것 01 다. 예를 들어, 어떤 한 종의 분포 지역이 건조지역에서 중간지역
(비가 잘 내리지 않는 건조한 지역과 수분이 적절하게 공급되는 지 역 사이)으로 확장이 된다고 하자. 만일 이때 어떤 장벽으로 인해 서 이 종이 두 개의 집단으로 분리된다면, 결국 이들은 두 개의 새로운 종으로 전화될 것이다. 왜냐하면 건조지역에 있는 개체들 은 건조지역이라는 환경에 알맞는 형질을 통해 생존해 나가려 할 것이고, 중간지역의 생물은 또 나름대로 그 상황에 알맞는 형질을 가지고 생존해 나갈 것이기 때문이다. 말하자면 이 생물들은 두 개의 서로 다른 선택 압력에 놓이게 된다는 것이다. 그러나 그러 한 급격한 변화에 처하는 일 없이 원래의 종 자체가 서서히 변하 는 환경에 적응해 가면서 디른 종으로 진화되어 갈 수도 있다. 예를 들어, 건조지역으로부터 중간지역으로 환경이 서서히 바뀌게 되는 경우, 생물들은 중간지역에 맞도록 서서히 적응해 나갈 것이 다 . 우리는 전자를 〈종분화(種分化)〉라 하고 후자를 〈 계통적 전 화〉라한다. 도약 진화와 본질주의 우리는 위대한 과학혁명을 일으킨 사람들을 가리켜 과거에는 상상할 수도 없었던 어떤 것, 즉 과거와의 완전한 단절을 가져온 사람들이라고 얘기한다. 그러나 코페르니쿠스는 프톨레마이오스와 똑같은 주전원을 사용했고, 다윈 역시 초기의 생물학 이론의 명맥 을 그대로 유지했다고 볼 수 있다. 자연선택이 전화를 일으키는 가장 중요하고 직접적인 힘이라고 믿었던 것은 틀림없지만, 그는 환경이 주는 직접적인 영향이나 기관(器官)의 용불용(用不用) ®의
® 용불용설. 전화의 한 요인으로 라마르크 Je an Bap tist e Pie r re La-
marck (1 744-1829) 가 제창한 설로서, 동물의 기관은 많이 쓰면 쓸 수록 발달하고 쓰지 않으면 위축 퇴화하는데 이 후천적인 용불용 아 디옴- 세대로 전승되고 그것이 대를 거듭함에 따라 축적되어 동 물의 종분화, 죽 진화의 요인이 된다는 이론이다. 그러나 이 설은 그 후 유전학에 의하여 소위 획득형질, 죽 후천적으로 획득된 개체 의 변이는 유전되지 않는다는 것이 밝혀침에 따라 과학적 근거를 갖지 못하게 되었다.
영향둘 역시 인정하였다. 실제로 그의 판게네시스p an genes i s 이 론에는 이 두 가지 형태의 영향을 받아 생겨날 수 있는 기작도 포함되어 있다. 그는 수 년에 걸쳐서 그의 이론에 계속 수정을 가해 나갔지만 그가 끝내 수정을 거부했던 부분이 있는데, 그것은 전화는 점진적으로 일어난다는 것이고 그러나 어떤 목표를 향해 진행되는 것은 아니라는 점이다. 다윈 이후, 생물학자들은- 종이 전화한다는 사실을 순식간에 모두 받아들이게 되었지만, 다윈이 제시한 몇 개의 기작들과 특히 위에서 언급한 두 원리, 즉 전화 란 것이 점진적이며 목표를 가지고 있지 않다는 점에 대해서는 강한 반대를 보였다. 그 중에서도 미바르트 S t. George Jac kson M i var t (l827-1900) 의 견해는 상당한 영향력을 발휘했고 여러 의 미로 볼 때 가장 전형적인 반대 견해라고 할 수 있었다. 그에 의 하면, 대부분의 종은 어떤 내적인 고유의 힘에 의하여 갑작스럽게 단계를 뛰어넘으면서 도약적으로 전화한다는 것이다. 종은 마치 자체 추전력을 지닌 다면체 (多面體)가 한 면에서 다른 면으로 자 체의 힘에 의해 굴러가는 것처럼 전화해 간다는 것이다. 다윈 시 대의 수많은 학자들이 왜 전화를 방향성이 있는 것으로 생각했는 지에 대해서는 이 책의 끝부분에 있는 목적론 부분에서 다시 논 의가 될 것이다. 많은 생물학자들이 도약(挑躍) 전화에 큰 매력을 가졌던 것은 그 나름대로 형이상학적인 근거가 있었는데, 이는
도약 진화가 본질주의자들이 주장하는 자연 종 natu ral ki nds 의 개념을 수용할 수 있었기 때문이다. 소크라데스 이래로 철학자들은 거의 모두, 집단이라는 것은 그 집단이 집단이 되기 위한 필요조건이면서 그 집단의 구성원이 되 기 위해 충분한 몇 개의 특성들로서 정의되어야 한다고 생각해 왔다 . 다시 말해서, 그 특성들 각각은 구성원이 되는 데 필수적이 며, 그 각각의 특성을 합한 것은 집단의 구성원이 되기 위한 충 분조건이 되어야 한다는 것이다. 어떤 자연 종을 정의할 때 그 정의 속에는 그 종이 속하고 있는 더 높은 단계의 명사가 포함되 어 있게 마련이다. 예를 들어, 총각은 결혼하지 않은 남자라고 정 의할 수 있는데, 이러한 정의 속에는 더 높은 단계인 〈 남자 〉 라는 단어가 등장한다. 또, 〈 남자 〉 란 어론이며 수컷으로서의 기능을 하 는 인간이라고 정의할 수 있고, 〈 인간 〉 은 엄지손가락을 다른 손 가락과 마주보게 할 수 있고 발로 걸울 수 있으며, 고도로 발달 된 뇌를 가지고 있는 동물이라는 식으로 정의할 수 있다. 이러한 전통적인 방식은 각각의 자연 종들이 본연의 체계질서를 항상 그 대로 유지하고 있는 세계에서는 대단히 유용한 것이라 할 수 있 겠지만, 서서히 진화해나가는 종을 묘사하기에는 적합하다고 볼 수 없다. 다윈의 이론에 따르면 종 자체나 그러한 종을 정의하기 위해 사용되는 특성이나 그 어떤 것도 개별적인 실체일 수가 없 다. 허파를 예로 들면, 어떤 부류의 동물에서는 허파가 점점 발달 되어 나가는 반면에 다른 부류의 동물에서는 접차 퇴화가 된다. 그러나 이 경우, 하나의 구조가 점점 허파의 모습으로 변해간다거 나 허파가 되는 것을 중단하는 분기점이라 할 수 있는 어떤 일정 한 시점을 집어낼 수는 없다. 이러한 하나하나의 구조들이 점진적 으로 변화함으로써 종도 전화되는 것이므로, 하나의 종이 다른 종
으로 변하는 일정한 시점 역시 있을 수 없게 된다. 실제로는 이 렇게 점진적인 전화를 거치면서 한 계통의 생물들이 여러 종으로 계속 분리되어 나가는 것이지만, 그렇게 분리된 종들의 이름은 전 통적인 방식, 죽 아주 인위적인 방법으로, 보통 종을 구분짓는다 고 생각되는 단 몇 개의 특징에 의해서 만들어지게 된다. 위에서 언급한 것처럼 〈 인간 〉 을 엄지손가락, 손, 뇌 등 단 몇 개의 중요 한 특칭을 가지고 정의하는 것도 그 한 예이다 . 그러므로 이러한 전통적인 방법은 그러한 방법을 통해 만들어진 개념의 실질적이 고 전정한 의미를 희생시키게 된다는 것이다. 만일 현대의 분류학 자들이 의견의 일치를 보이는 것이 있다면, 단 하나의 특칭만으로 동식물의 종을 구분한다면 생물학적 분류의 전정한 목적은 사라 진다는 점일 것이다. 그런데 도약 전화의 경우에는 수많은 형질들 이 한 세대에 갑자기 한꺼번에 변화하게 된다. 따라서 다윈 시대 의 많은 사람들은 도약적인 경우에 한해서만 진화의 개념을 용납 할 수가 있었는데, 이는 본질주의자들의 정의 방식에 대한 그들의 선호도를 반영하는 것이라 할 수 있다. 그들은 종들이 영원불변한 존재까지는 못되더라도 최소한 뚜렷이 불연속적인 존재일 것이라 고 믿었던 것이다 (Hull, 1973 참조). 멘델 유전학이 재발견되기 이전에도 또 그 이후에도 생물학자 들은 전화에 작용하는 변이(變異)란 것에 대해 논란을 거듭해 왔 다. 과연 변이는 드브리스 Hu g o de Vr i es 나 골드슈미트 R i chard Goldsc hmi d t가 주장한 것처럼 크고 급작스러운 것인가? 그렇다 면, 전화는 분명히 도약적 인 것이다. 아니면 갈돈 Francis Galto n , 베이트슨 W illi am Bate s on, 모건 T . H. Morga n 등이 주장한 바와 같이 작으면서 급작스러운 것인가? 그렇다면 진화란 도대체 점진 적이라고 해야 하는가, 도약적이라고 해야 히는가? 근대 유전학의
출현 이후 진화론을 가장 괴롭혀 왔던 혼란 중의 하나는 돌연변 이의 크기와 그것의 돌연성 정도 abru pt ness 를 갇다고 보는 생각 이었다. 크고 갑작스러운 변이를 주장했던 드브리스가 왜 다윈의 견해에 반대를 했는지는 쉽게 이해가 가지만, 갈톤, 베이트슨, 모 건 들도 역시 그들의 견해가 다윈의 그것과 다르다고 주장했던 것이다. 사실 다윈은 진화의 기본이 되는 변이란 것이 〈 연속적 〉 이라고 주장한 적이 없었고, 연속적이라는 말은 베이트슨이 사용 한 용어였던 것이다. 다윈은 변이둘이 〈 뚜렷하다 〉 는 것과는 대조 되는 표현인 〈약간〉 또는 〈 감지할 수 없는 〉 것이라 말했을 뿐이 다. 다만 이러한 일련의 변이는 한꺼번에 일어나는 것이 아니라고 덧붙였는데, 한꺼번에 일어나지 않기 때문에 결국 진화는 점진적 인 것이라는 의미가 된다. 로버트 울비 (Rober t Olby , 1966 : 82) 는 순수유전과 혼합유전 에 대한 그의 저서에서, 왜 다윈이 전화란 여러 개의 아주 작은 그러나 명확하게 구분이 되는 단계들로 이루어져 있다는 사설을 받아들이지 않았는지에 대해서 의문을 제기하였다. 그러나 사실은 다윈은 그것을 무시한 것은 아니었다. 그는 다만 크고 갑작스러운 표현형적 변화를 전화에서 배제하려고 노력하였던 것뿐이다. 그러 한 표현형적 변화들이, 유전자형의 수준에서 볼 때 단 한번의 커 다란 변화에 의한 것인지 아니면 수많은 작은 변화들이 동시에 일어났기 때문인지 하는 것은 문제가 아니었다. 그러나 어쨌든 그 문제에 대한 다윈의 직관은 근본적으로 옳은 것이었다고 볼 수 있다. 왜냐하면 도약 진화를 일으키는 요인으로 발견된 . 유전 기작 은 염색체의 배수성(倍數性)뿐이며, 실제로 대부분의 경우 전화란 점진적이지 도약적인 것이 아니기 때문이다 (Vorz i mmer, 1971 과 Provin e , 1971 참조). 그러므로 전통적인 의미에서의 자연 종이
과학법칙에 필수적인 것이라 한다면, 종의 이름은 자연 법칙 속에 서 결코 아무런 기능도 의미도 가질 수 없게 될 것이다. 〈모든 백조는 희다 〉 라는 문장은 자연 법칙이 될 후보 명단에도 낄 수가 없을 것이다. 실제로 그것이 진리일 경우에도 말이다 . 따라서 정 의라는 개념이 상호 변화하는 특징들까지도 포함한다고 확대 해 석될 수 있는 경우에 한하여, 종이나 혹은 더 높은 단계의 분류 군의 이름이 자연 법칙에 의미있게 쓰일 수 있을 것이다. 판게네시스 이론과 혼합유전 여러 유전현상들을 모두 수용하고 또 전화론이 요구하는 바를 충족시킬 수 있는 유전법칙을 마련하기 위하여 다윈은 판게네시 스라는 이론을 만들었다. 이 이론에 따르면 신체의 각 부분은 그 부분의 특성을 지닌 유아(幼茅, g emmule) 를 만들고, 이러한 유아 들은 각자 다른 비율로 생식소에 모였다가 생식 과정을 통하여 다음 세대로 전달된다. 그렇게 생긴 자손이 어떤 특성을 갖게 되 느냐는 것은 부모로부터 받은 유아의 양과 질에 달려 있다. 따라 서 한 개체가 파란색보다 갈색의 유아를 더 많이 받았다면 그는 갈색눈을 지니게 될 것이다. 간혹 유아가 참재적인 상태로 존재하 는 경우가 있어서, 어떤 알 수 없는 요인으로 인하여 다시 증식 하기 시작할 때까지 동면상태로 세대에서 세대로 전달이 되기도 한다. 유아는 몸 전체에서 만들어지기 때문에 신체의 어느 한 부 분에 변화가 생기면 그 부분과 관련된 유아가 영향을 받게 되므 로, 신체의 어느 부분을 많이 사용하거나 사용하지 않는 것, 혹은 그 의의 다른 여러 환경적인 요인들도 신체에 변화를 초래함으로
써 유전에 영향을 미칠 수 있다는 것이다. 다윈 이론을 해설하는 학자들은 종종 다윈이 혼합유전을 버리 지 않으려는 일념으로 인해 입자론적 유전의 개념 (1 857) 을 만들 어 내게 되었음을 애석해 한다. 최근 나는 예전에 없이 나를 매혹시킨 주제인 유전에 대해 생 각해 보면서, 수정에 의한 증식은 실은 진정한 의미의 융합이 아 니라 일종의 혼합이며, 그 혼합도 두 개체의 혼합이 아니라 결국 수많은 개체의 혼합-부모는 또 각각 그의 부모와 조상을 가지 고 있으므로一_이라는 사실을 어령풋이 깨닫게 되었다• 그런 식 으로 계속 거슬러 올라가서 드디어 이러한 혼합 형태가 최초의 조상에 이르는 것 이의에 나는 달리 유전을 이해할 방법이 없다. 그러나 물론 이러한 나의 견해는 아직 제대로 다듬어지지 않은 상태이다 .2)
2) Francis Darw in, ed., More Lett er s of Charles Darwi n, I (London : Jo hn Murray, 1903), p. 103.
여기서 다윈이 멘델의 입자 유전 같은 것을 예상했던 것은 아 니고, 단지 그는 판게네시스p an genes i s 라는 자신의 이론을 설명 하고 있는 것이다. 멘델 이론 속에 나오는 입자로서의 개념이 아 니라, 그저 단순히 그가 제시한 유아의 개념이 입자라는 의미에서 다윈의 이론은 〈입자론적〉이라고 불릴 수 있으며, 때로 유아 자 체가 융합하기도 하고, 한 형질을 만들어 내기 위해 여러 유아들 이 합해지기도 하는데 그렇게 합해지는 유아의 숫자가 각양각색 이고 또 엄청나게 많아지기도 하기 때문에 그의 이론은 〈 혼합 blen di n g〉에 관한 이론이라고 불리기도 한다. 그에 의하면 잡종인
한 자녀가 부모의 중간쯤 되는 특성을 갖거나 한쪽 부모의 형질 이 다른 쪽보다 우세하게 나타나거나 하는· 것은 그 형질에 관련 된 유아의 숫자와 그 강도, 또는 친화도 a ffi n ity 등에 달려 있는 것이다. 이 경우, 각 세포는 질이 서로 디론· 수많은 유아를 만들 고, 또 수정될 때 접합체로 전달되는 유아의 비율이 서로 다르기 때문에 그 가능한 조합의 수는 엄청나게 클 것이다. 그러나 평균 적으로 볼 때 어느 한 가지 유아는 한 번 교배될 때마다 그 숫자 가 절반으로 줄어들게 된다. 다윈의 유전 개념에 대한 피셔 (Fis h er, 1930) 와 보르짐머 (Vorzim mer, 1971) 의 분석과는 상당히 대조적이지만, 바로 이것이 판게네시스 이론이 〈 혼합유전〉의 이 론이라고생각된다. 다윈의 판게네시스 이론은 188~ 빈에 드브리스에 의하여 수정되 어 되살아나기도 했으나 인기도 없었고 평판도 좋지 못했다. 이 이론의 가장 큰 문제점은 그것이 무한정 확장될 수 있다는 데 있 었다. 또한 무슨 일이든간에 그것을 과거로 소급해 설명할 수는 있었으나 미래에 대한 정확한 예측은 전혀 할 수가 없었다. 죽, 파란색 눈을 가진 사람과 갈색눈을 가전 사람이 결혼한다고 했을 때, 파란색과 갈색이 어떤 바율로 자녀들 사이에 나타나게 될지는 고사하고 그 자녀들이 과연 파란색과 갈색 중 대체 어떤 색깔의 눈을 갖고 태어날지조차 전혀 예상할 수가 없는 것이다. 유아가 몸 전체에서 생성되는 것이라면 획득형질도 유전이 될 수 있어야 한다고 생각할지도 모론다. 그러나 획득형질이 유전되지 않는 것 이 확실하다 하더라도 그러한 사실이 반드시 판게네시스 이론을 잘못이라고 단정할 수는 없다. 유아는 환경에 의해서 변화될 수가 있기 때문이다. 따라서 그러한 변화로 인해 획득형질의 유전이 불 가능하게 되었다고 설명하면 그만이다. 또 유아라는 것이 어떤 고
정된 비율로 생성되어야 할 필요도 없지만 만일 그렇다 하더라도 이러한 사실 역시 판게네시스 이론을 반증하는 설명이라고 할 수 는 없다. 판게네시스 이론을 반증할 수 있었던 유일한 것은 체세 포 속에 있는 유전물질은 생식을 통해 다음 세대로 전달될 수가 없다는 사실이었는데, 이는 바이스만 Au g us t We i ssman 에 의해서 생식세포와 체세포가 명백히 구분됨으로써 결정적인 증거를 갖게 되었다. 단일 변이와 개체간의 차이 다원에게는 단일 변이 sin g le var i a ti on 와 개체간의 차이 ind i- vidu al diffe renc 촐 구분하는 것이 대단히 중요한 일이었다. 단일 변이란 특이한 개체가 하나 출현하는 것이라고 할 수 있는데, 다 윈은 그러한 단일 변이를 흔히 말하는 〈 변종 〉 과 같은 것으로 생 각하였다. 반면 개체간의 차이는 한 종 내의 구성원들 사이에서 흔히 볼 수 있는 아주 미세한 차이라고 할 수 있는데, 다윈은 바 로 이러한 개체간의 차이가 전화의 원인이라고 믿었다. 젠킨 (Fleem ing Jen k in, 1867 ; Hull, 1973 속에서)의 논문을 읽으면서 다윈은 이러한 자신의 견해가 옳다는 것을 다시 확신하게 되었으 나, 이 논문은 혼합유전이라는 가설 하에서 자연선택에 의한 전화 가 과연 새로운 종을 만들어 낼 수 있겠는가 하는 심각한 의문을 불러일으키게 되었다. 젠킨은 다윈의 유전 원리에 대해 이의를 제기하면서 아무리 극 적인 자연선택이 일어난다 하더라도 자연선택에 의해서 어떤 새 로운 형질이 한 집단 내에서 고정이 될 수는 없다고 주장하였다.
예를 들어, 백인 하나가 100 명의 흑인으로 이루어진 집단으로 들 어와서는 흑인 남자들을 모두 다 살해하고, 흑인 여성 모두와 잠 자리를 같이 했다고 하면 그렇게 해서 태어난 다음 세대는 모두 갈색이 될 것이다. 다시 그 백인이 아들과 아내둘을 모두 살해하 고서 딸들과 잠자리를 같이 했다고 하면, 그렇게 해서 생긴 다음 세대는 75% 의 흰색과 25% 의 검은색으로 혼합된 피부색을 갖게 될 것이다. 이런 식의 일이 그가 죽을 때까지 거듭된다고 하자. 그러나 일단 그가 죽고 나면, 피부색이 다른 새로운 방문객이 등 장하지 않는 한, 그 백인이 죽을 당시 그 집단이 갖고 있던 피부 색은 그 상태 그대로 유지가 될 것이다. 그러므로 혼합유전의 가 설 하에서는 한 사람의 백인이 흑인 국가를 완전한 백인 국가로 바꿀 수는 없다는 결론에 이르게 된다. 그러나 젠킨의 결론은 한 종의 구성원들 사이에서 나타나는 무 수한 작은 변이들이 전화의 근원이라는 다윈의 신념을 재확인해 주는 결과를 가져왔다. 예를 들어 구성원의 피부색이 조금씩 다른 어떤 종족이 있다고 하자. 가장 검은 피부를 가전 사람을 106] 라 하고 가장 밝은 색의 피부를 가전 사람을 8 이라 하면 그 종족의 평균 피부색은 9 가 될 것이다. 여기서 참시 밝은 색의 피부가 생 존에 더 유리하다고 가정을 하면 밝은 색 피부를 가전 개체들은 검은 피부를 가전 개체보다 더 많이 살아 남아 자손을 낳게 될 것이다. 따라서 다음 세대의 평균 피부색은 8. 또료 변화될 수 있을 것이다. 원래의 집단은 평균치로부터 양쪽으로 1 점씩의 변이를 보 였었는데, 만일 이러한 상황이 그대로 지속된 채로 여러 세대를 지나게 되면 그 종족 내에서 가장 검은 피부를 가진 개체는 9.5 가 될 것이고 가장 밝은 피부를 지닌 개체는 7.57} 될 것이다. 따 라서 세대를 거치면서 형질이 계속 〈모든 방향으로〉 변해나가기
만 한다면 혼합유전이라는 가설 하에서도 전화는 얼마든지 일어 날수가 있게 된다. 한편으로 젠킨은 다윈의 가장 큰 약점을 붙들고 늘어졌다고 할 수 있는데, 그에 의하면 평균치는 자연선택에 의해서 변동될 수 있을지 모르지만 평균치가 변한다고 해서 변이의 한계도 함께 변 동되는 것은 아니라는 것이다. 각각의 종에는 그 종만의 고유한 변이의 영역이 존재한다고 그는 주장하였다. 처음에는 선택에 의 해서 형질의 평균치가 크게 변동될 수 있겠지만 평균치가 변이의 한계점에 가까워질수록 그러한 변동은 점차 어려워지게 될 것이 다. 더군다나 이러한 특수한 극단 상태에서 그 집단에 대한 선택 압력(選擇壓力)이 제거된다면, 집단은 원래의 상태로 저절로 돌아 가게 된다는 것이다. 젠킨은 자신의 주장을 보강하기 위하여 원예 학자와 동물사육가들의 실험을 그 예로 들었다. 다윈은 젠킨의 주 장이 매우 설득력 있다는 사실을 인정하지 않을 수 없었지만, 그 러한 가설적인 변이의 한계점을 뛰어넘을 수 있다는 확신도 버리 지 못하였다. 모두가 그릇된 것이라 치부해 버리기에는 전화론에 의해 설명될 수 있는 현상들이 너무 방대하고 다양했던 것이다. 물론 다윈의 실수는 유전 이론, 죽 모든 형질은 전달되는 과정 에서 섞이게 된다는 판게네시스 이론에 있었다 . 아형접합의 경우, 어떤 형질들이 완전히 사라졌다가도 훗날 어떤 세대에서 그대로 변형되지 않고 다시 나타난다는 것을 그도 잘 알고 있었다. 그러 나 그는 그러한 현상을 격세유전(~젊世遺傳)으로 간주하여 전화와 는 무관한 일로 무시해 버렸다. 다윈이 도피할 수 있는 어떠한 여지도 남겨주고 싶지 않았던 젠킨은 생물체가 형질을 희석시키지 않고도 그러한 형질을 후손 에게 무한정으로 전달해 나갈 수 있는 가능성에 대해 생각해 보
았다. 그러한 가설 하에서는 전화가 아주 빠른 속도로 일어날 수 있을 것이었다. 그는 〈 종의 기원에 관한 나의 이론은 다윈의 것 과는 분명히 다르다. 나의 이론은 동물이 종종 그 조상과 아주 다른 모습을 지닌 채 태어나고 또 그러한 차이점을 후손에게 전 달할 수도 있다고 하는 가설일 뿐이다. 그러나 이것은 곧 새로운 종이 창조될 수 있음울 의미하는 것이 아니겠는가? 〉 라고 하였다. 그러나 위의 글에서 젠킨은 그의 무지와 편견을 여실히 나타내 고 있는데, 이 점에 있어서는 훗날 멘델 유전학의 기초를 다진 사람들도 마찬가지였다. 다윈 시대에 있어서나 세기가 바뀐 뒤에 있어서나 한 가지 형질에 있어서의 변화는 그것이 아무리 급작스 럽게 일어난 것이라도 신종을 만들어내기에는 충분하지 못하다는 것이 분명하고, 또 초자연적인 의미에서도 이러한 조그마한 변화 를 〈 창조 〉 라고 부를 수는 없는 것이다. 멘델 유전학의 초기 연구 가 모두 진화를 염두에 두고 이루어졌다는 사실만으로도 전화론 의 중요성은 높이 평가될 수 있을텐데, 그러한 학자들의 상딩수가 오히려 전화론을 반박했다는 사실은 그들이 얼마나 전화론에 대 해 무지하였고 얼마나 많은 편견을 가지고 있었는가를 보여주는 것 0] 다. 전화의 유전적 요소에 대한 논쟁에 있어서는, 그 논쟁에 가담 한 모두가 부분적으로는 옳고 부분적으로는 그르다고 할 수 있다. 전화의 가장 중요한 요인이라고 할 수 있는 변이는 아주 조금씩 천천히 일어나는 것이라는 다윈의 견해는 옳다고 볼 수 있지만, 한 종의 구성원들 사이에서 예사로 관찰되는 변이들 모두가 똑같 이 진화의 재료가 된다는 그의 생각은 잘못된 것이다. 개체간의 차이는 대부분 재조합과 환경의 영향 때문이라는 것을 그는 몰랐 던 것이다. 유전물질 자체에 어떤 변화가 일어나서 생긴 이러한
미세한 차이들 중 극히 일부만이 실질적인 〈 돌연변이(변이) 〉 인 것이다. 돌연변이(변이)는 이와 같이 희소하기는 하지만 그대로 혼합되지 않고 순수한 상태로 바로 유전된다. 그래서 아~느 적은 숫자의 변이이지만 그러한 변이는 섞여서 소멸되는 것이 아니라 세대에서 세대로 계속 전해짐으로써 진화의 잠재적인 요인으로서 의 역할을 하는 것이다. 겉으로 나타나는 형질은 섞인 것처럼 보 일는지 모르지만 유전자는 결코 섞이지 않는다. 진화의 유전 이론 유전학과 전화론이 양립될 수는 없다고 그렇게 법석을 떨었음 에도 불구하고 생물학자들은 금새 이들 두 이론이 서로 완전히 일치한다는 것을 깨닫게 되었다 (Prov i ne, 1971 참조). 다윈은 전 화에 필요한 변이를 생성하고 유전시키는 어떤 기작이 있다고 가 정하고 그의 전화론을 만들었는데 멘델 유전학이 바로 그 기작에 해당된다고 할 수 있다. 사람들은 멘델 유전학과 다윈의 전화론을 결합하면 더욱 훌륭한 진화론이 탄생되리라고 기대했었고, 이러한 목적으로 피셔 R. A. Fis h er, 할데 인 J. B. S. Haldane, 라이트 Sewall Wrig h t 등은 수학적 진화론, 혹은 신다윈주의나 유전학적 전화론 등 여러 가지 이름으로 알려진 이론을 만들게 되었다. 어 떤 관점에서는 이 세 사람의 연구가 서로 차이점을 보이긴 했지 만, 그들의 작업에는 두 가지 기본적인 공통점이 있었는데 그것은 표현 방식이 수학적이었다는 점과 세대가 바뀌면서 유전자의 분 포가 어떻게 변해가는지에 관심을 두었다는 점이다. 전화론의 발달 과정을 통해서 보면, 수학적인 것에 관심을 두
었던 생물학자들은 유전형질의 전달과 자연선택의 힘에 대한 개 념들이 얼마나 잘못되었는가를 증명하는 데 중대한 역할을 했다 고 볼 수 있다. 예를 들어, 어떤 생물학자들은 우성과 열성 형질 들이 왜 모든 자연 집단 내에서 3 : 1 의 비율로 나타나지 않는가 에 대해 의문을 가졌었다. 한 특정 형질에 대해 이형접합체인 생 물을 같은 숫자의 동일한 이형접합체로 교배한다면 그 결과 생기 는 자녀는 3 : 1 의 비율을 보일 것이고 교배가 무작위적이고 선택 압력이 없는 상태라면 세대가 계속 바뀌더라도 그러한 3 : 1 의 비 율은 그대로 유지되어야 할 것이라고 생각했던 것이다 . 그러나 190 8\1에 하디 G. H .• H ard y와 와인버그 C. We i nber g는 그러한 수 학적 판단이 얼마나 잘못된 것인가를 지적하였다. 예를 들어, 돌 연변이의 경우에는 같은 숫자의 이형접합체가 아니라 새로운 대 립유전자를 가전 개체 하나가 집단에 유입되는 것이라고 생각할 수 있는데, 이러한 돌연변이는 그것이 우성이건 열성이건 간에 선 랙압력에 대해 중립이라면 그 집단 내에서 아주 낮은 비율로 유 지가 될 것이다. 하디와 와인버그는 크기가 충분히 큰 집단에 있 어서, 특정 유전자에 관해서 볼 때 암수가 무작위적으로 교배되고 그러한 각각의 교배로부터 생기는 자손은 그 수와 생식 능력이 동일하며 그 의의 모든 조건 또한 동일하게 유지될 수 있다면, 그러한 집단에서는 동형집합체와 이형집합체가 대략 같은 비율로 유지될 것이라고 주장하였다. 어떤 집단 내에서 한 대립유전자를· 지닌 배우자(생식세포)의 비율을 院} 하고 또 다른 대립유전자를 지닌 배우자의 비율을 虐} 한다면, 하디-와인버그 법칙은 다음과 같이 간단히 표현할 수가 있다. p2 + Zpq + q2 = 1
여기서 p은 한 동형접합체의 빈도를 나타내며, 2 凶는 이형접합 체의 빈도를, 검은 다른 또 하나의 동형접합체의 빈도를 나타낸 다. 그러나 전화의 유전 이론이 이렇게 수학적으로 표현이 될 수 는 있으나 거기에는 한계가 있다. 하디-와인버그 법칙의 중요성은 그것이 수학적으로 표현되었다 는 데 있는 것이 아니라, 실제로 존재하는 자연 집단들을 서로 비교할 수 있는 어떤 기준을 마련했다는 데 있다. 하디-와인버그 법칙에 따라 우리는, 훗날 세대가 여러 번 바뀌었을 때 유전자의 빈도는 과연 어떻게 변할 것인가를 예측할 수가 있다. 그것이 불 가능하다면 무언가 조건이 달라졌음이 틀림없다. 교배가 어떤 유 전자에 대해 무작위적으로 일어나지 않았거나, 또 다른 새로운 유 전자가 집단 내에 유입되었거나 선택압력이 작용했거나 그랬을 것이다. 어떤 경우이건 중요한 어떤 요인이 작용했음이 틀림없다. 그러나 하디-와인버그 법칙은 이렇게 단순하게 표현될 수 있는 대신 다른 대가를 치루어야만 하는데, 그것은 전화의 유전 이론을 수학적으로 표현하고 있는 이 법칙이 단지 유전자와 유전자형의 빈도만을 다루고 있지 유전자 각각과 그것으로 인해 나타나는- 표 현형의 관계 혹은 생물체 각각과 그들이 부딪치는 환경 사이의 복잡한 관계는 완전히 무시하고 있다는 것이다. 따라서 하디-와 인버그 법칙을 통해 우리가 예측할 수 있는 것은 알고자 하는- 유 전자의 비율이 일정하게 유지될 것이라는- 사실과, 만일 그렇지 않 다면 무언가가 그러한 변화를 초래했을 것이라는 사실뿐이다. 예 롤 들어 伊 1 빈도가 증가되는 경우, 이러한 증가가 계속될 것인 지, 어느 시점에서 멈출 것인지, 아니면 다시 줄어들 것인지 예측 할 아무런 방도가 없다. 그러한 예측을 하려면 이렇게 수학적으로 표현된 유전 이론 의의 다른 것에서 그 방도를 찾을 수밖에 없을
것이다. 그러므로 공식으로서의 하디-와인버그 법칙은 두 개의 요소를 가지고 만들 수 있는 조합의 비율이 어떻게 서로 연관될 수 있는가를 보이는 것일 뿐이다. 요소 그 자체는 일원짜리 동전 두 개와 같은 하찮은 것일 수도 있다. 만일 그 법칙에다 경험적 내용을 부여한다면 그것은 그 법칙이 갖는 한계 조전이 될 것이 다. 즉 경우에 따라 그 내용이 변하면서도 항상 그 법칙에 들어 맞아야할것이다. 진화론의 통계적 특성과 그 근원 과학 이론이 되기 위한 필요조건 중의 하나는 자연 현상에 대 해 추론을 할 수 있어야 한다는 것이다. 그러한 추론은 연역적일 수도 있고 귀납적일 수도 있다. 또 과거나 현재에 대한 것일 수 도 있고 미래에 일어날 사건에 대한 것일 수도 있다 . 그 중에서 가장 전형적인 것이라면 미래에 관해 연역적인 방법으로 추론하 는 것을 들 수 있다. 말하자면, 연역적 예측이라고 할 수 있을 것 이다. 전화론에서의 예측은 몇 가지 형태로 나누어볼 수가 있는데 특정 한 생 물 ( Gar g an t ua 처 럼 ) 이나 종 (Ph t h i r i us p ubus 처 럼 ) 에 대한 예측, 현재로서는 잘 모르고 있는 생물들을 어떤 식으로 그룹을 짓게 될까 하는 데 대한 예측(앞으로 몇 개의 예를 언급하게 될 것이다), 혹은 여러 가지 있을 법한 전화 양상이나 생길 법한 종들의 종류(예를 들어 방사선에 적응해 나가고 있는 종과 갇이) 에 대한 예측이 그것이다. 과거에는 생물학자들이 특정한 종이나 집단에 대한 예측에만 관심을 가졌었으나 이 장과 다음 장에서 우리는 그러한 관심이 잘못된 것이라는 사실에 대해 논의하게 될
것이다. 종이란 것이 현재 정의되어 있는 것처럼 그렇게 뚜렷한 진화 단위가 아닐지도 모르며, 그보다 더 중요한 것은 얘기하고자 하는 분류의 단계가 무엇이든간에 진화 법칙이 어떤 하나의 특 정 생물에 직접적으로 적용될 수는 없다는 것이다. 그러한 법칙들 은 일련의 어떤 것, 죽 일련의 종들, 일련의 집단들과 같은 식으 로 관련지을 수 있을 뿐이다. 다윈 시대에는 뉴턴의 역학이 과학 이론의 본보기였다. 그것은 연역적으로 구성되어 있었고 정확한 예측을 가능하게 했는데, 예 를 들어 적절한 데이터가 역학의 법칙과 함께 주어지면 누구나 아주 정확하게 화성과 같은 특정 혹성의 위치를 추론해 낼 수가 있었던 것이다. 이처럼 디윈 시대나 현재의 생물학자둘도 간혹 각 각의 생물에 대해 정확히 예측을 해야만 하는 입장에 처하게 되 는데 전화론에 있어서는 사실상 그것이 불가능하다고 보아야 한 다. 일찌기 철학자 퍼스 C.S. Pe i rce 가 말한 바와 같이 전화론은 이러한 접에서 뉴턴의 역학과는 크게 다르다고 할 수 있다 . 적철 한 데이터와 진화론이 주어진다 하더라도 어느 특정 생물의 운명 에 관한 합리적인 추론을 한다는 것은 불가능한 일이다. 그러나 퍼스는 뉴턴 이의의 다른 이론들에 비하면 전화론이 그렇게 형편 없는 것은 아니며, 그 당시에 받아들여지기 시작했던 기체 이론처 럼 통계적인 형식도 갖추고 있으므로 예측하고자 하는 것이 수많 은 개체로 이루어전 대단히 커다란 집단이라면 앞으로의 일에 대 한 추론이 가능하다고 주장하였다. 다윈주의자둘의 논쟁은 대부분 논리에 대한 문제이다. 다윈은 생물학에 통계적인 방법을 적용할 것을 제안했는데, 그러한 통계 적 방법은 과학의 여러 다론 분야, 예를 들어 기체 이론 같은 데
서도 이미 사용되고 있다. 기체의 구성에 관한 가설 그 자체에 의해서는 어떤 특정 기체 분자의 운동을 예측할 수가 없었지만, 클러시우스 Claus i us 와 맥스웰 Maxwell 은 확률이라는 원리를 적 용하여 어떤 주어진 조건 하에서 이러이러한 분자들이 이러이러 한 속도를 지니게 될 것이며, 매초 이러이러한 숫자의 충돌이 일 어날 것이라는 예측을 할 수 있게 되었다. 또한 그러한 전제로부 터 기체의 어떤 성질 특히 분자와 열의 관계에 대한 연역을 할 수가 있었던 것이다. 마찬가지로 다윈도 개개의 개체에 대해서는 변이와 자연선택이 어떻게 되리라는 예측을 할 수 없었지만, 동 물들은 결국 주어진 환경에 잘 적응하게 될 것이러는· 사실을 증 명할 수 있었다. 하지만 현존하는 동물들이 그러한 작용에 의해 생긴 것이든 아니든, 이론이 어떤 입장을 취해야 하든, 사실과 논 리라는 논쟁의 주제는 기묘하게 얽히고 설킨 복잡한 문제임에 틀 립없다 •3)
3) Peir c e, The Fix a ti on of Beli ef, Pop u lar Scie n ce Mon thl y, 12 (18 77), pp. 1-15.
생물학자들이 시도해 보고자 했던 것이 전화가 일어날 것인가 아닌가를 예측하는 일이었다면 전화론은 통계적인 것이 되었을 것이다. 이 경우 통계적이라는 것은 과학철학의 관점에서 볼 때 어떤 특별한 하자가 없다는 의미가 될 것이다. 기체 법칙의 경우 에는, 각 기체 분자의 행동은 전혀 예측할 수가 없다 할지라도 그러한 분자들로 이루어전 커다란 집단의 행동은 아주 정확하게 예측할 수가 있었다. 그러나 생물계에서는 다윈 자신이 인정했던 바와 같이 생물 개체가 모인 커다란 집단의 경우에도 합리적인 예측이 불가능했던 것이다. 〈자세히 살펴보면 이것이 변화된 종
이다라고 증명해 보일 수도 없으며, 전화론의 토대라고 할 수 있 는 변화란 것이 과연 유익한 것인지 아닌지도 증명할 수가 없음 울 깨닫게 된다. 또한, 왜 어떤 종은 변하고 어떤 종은 변하지 않 는지에 대해서도 설명할 수가 없다 〉 고 다윈은 기록했다 4). 그러나
4) Franci s Darwi n, 'Ihe Lif e and Lett er s of Charles Dan. uin, II (New York : Basic Books, 1887, reis s ued 1959), p. 210.
집단이나 종과 같은 분류 단계에서는 전화론이 분명히 통계적인 이론처럼 보인다. 물론 전화론이 지니고 있는 이러한 불확실성의 원인은 아주 조심스럽게 분석되어야 할 것이고 우리가 다물 수 있는 종류의 불확실성과 다룰 수 없는 종류의 불확실성도 구분되 어야만 할 것이다. 앞으로 우리는 전화론을 근거로 하여 특정 종 이나 집단에 대한 합리적인 추론을 할 수는 없고, 기껏해야 이 집단은 여태까지 알려진 전화 양상 중 어떤 양식을 나타내 보 이게 될 것이다라는 정도의 예측을 할 수 있을 뿐임을 논의하게 될 것이다. 또한 우리는, 시험적인 것이기는 하지만 종과 같은 전 통적인 생물학적 단위가 전화론에 적합치 못할 수도 있다는 사실 도제시하게 될 것이다. 생물종의 전화가 지니고 있는 통계적 성질은 종합전화론의 유 전적인 요소와 자연선택의 특성에서 기인하는 것이다. 멘델의 유 전 법칙에 의하면 감수분열 때에 일어나는 교차와 분리는 모두 무작위적으로 일어나며, 수정될 때 어떤 정자가 어떤 난자와 결합 할 것인가도 우연의 문제라고 할 수 있다. 그러나 문제의 집단이 충분히 클 경우에는, 기체 분자의 무작위적인 운동이 기체운동에 서 무시될 수 있는 것과 마찬가지로 그러한 우연적인 요소들이 모두 무시될 수 있을 것이다. 여러 세대에 걸쳐 유전자가 통계적 으로 어떻게 분포될 것인지를 예측할 수 있을 만큼 충분히 큰 자
연 집단들도 분명히 있으며, 집단이 이렇게 충분히 클 경우에는 돌연변이의 무작위성조차도 종종 무시될 수가 있다. 돌연변이율이 보통 매우 낮긴 하지만, 보통의 경우에는 집단의 크기가 크다고 해서 그 존재가 상쇄되어도 좋을 만큼 낮다고는 볼 수 없을 것이 다. 그러나 만일 유전적으로 동일한 두 개의 집단이 어떤 제한없 이 무한히 팽창해서 새로운 두 개의 집단이 형성된다면 위에서 언급한 모든 무작위적인 요소에도 불구하고 그 두 집단은 통계적 관점에서 거의 동일하다고 볼 수 있을 것이다. 그러나 환경이 전화에 미치는 여러 가지 영향까지 우리의 연구 에 포함시키고자 한다면, 전화라는 개념은 다루기 힘든 무작위적 다양성까지 포함하게 된다. 이 경우, 그 한계를 어떻게 설정하는 가가 문제가 되는데, 생물 개체와 그 환경 사이의 구분이라는 것 이 그리 완벽한 것은 아니지만 대부분의 경우에 큰 어려움 없이 구분할 수가 있다. 개체라고 하는 것은 표현형적 특성을 통해서 나타나는 게놈과 환경 사이의 끊임없는 상호 작용의 결과라고 정 의할 수도 있겠지만, 어떤 표현형이 출현되어 나올 것인지의 범위 를 정하는 것은 한 생물이 가지고 있는 유전적 구조이기 때문이 다. 그러나 우리가 종이나 집단 같은 전통적인 전화 단위에까지 관심을 둔다면 상황은 사뭇 달라지게 된다. 이러한 시스템들은 그 시스템에 영향을 주는 여러 요소에 대해 개방되어 있기 때문이다. 만일 특정한 종의 발전 과정을 예측하려 한다면 그 종의 구성원 들이 집하게 될 환경 모두를 예측할 수 있어야만 하는데, 그렇게 하기 위해서는 지구의 지질적인 변화와 기상의 변화뿐 아니라 단 기간, 장기간에 걸친 기후 변화까지 아주 정확하게 예측할 수 있 어야 할 것이다. 예를 들어, 갑자기 강이 그 경로를 바꾸는 바람 에 한 개의 집단이 두 개로 나누어지고, 새로운 약탈자가 다른
종의 생존 구역을 침범하는 등의 상황이 벌어지고, 이때 새로운 돌연변이가 하나 나타난다고 하자. 이 새로운 돌연변이체는 환경 에 적응하거나 도태되거나 두 가지 중 하나의 길을 가야만 하는 데, 보통 처음 등장한 돌연변이체는 생존에 매우 불리한 처지에 있게 된다. 그러나 훗날 상황이 많이 달라전 후에 그러한 돌연변 이체가 나타나게 된다면 그 개체는 살아 남아 자리를 잡을 수 있 울지도 모른다. 환경이 변하게 되면, 그 돌연변이체가 자리를 잡 을 수 있도록 어떤 훌륭한 생태적 적소ni che7} 그 돌연변이룰 위 해 열리게 될지도 모르기 때문이다. 그리고 생태적 적소는 그것이 어떤 생물에 의해 점유된 적이 있었느냐에 상관없이 열리고 닫히 고 하는 것이기 때문이다. 현재로서는 특정한 집단의 미래에 대한 장기적 예측을 할 수 있을 만큼 환경 요소들의 값을 정확히 측정할 수 있는 처지가 못 된다. 뿐만 아니라, 설혹 전화에 영향을 주는 그러한 환경 요소에 대한 자료를 가지고 있다 할지라도, 자세하고 정확한 예측을 할 수 있도록 이러한 자료들을 종합할 수 있는 방법조차 없는 실정 이다. 레원틴 Ri ch ard Lewon ti n은 이러한 상황을 다음과 같이 기 술하였다 (Mendelsohn~ 1969 중에서 인용). 대부분의 전화론은 다음과 같은 구조를 가지고 있다. 죽 전화 론에는 일반적인 틀이 있다고 볼 수 있다. 자연을 관찰하면서 우 리는 어떤 매개 변수에 대한 지식을 갖게 되고 그것을 계산에 넣 어봄으로써 납득할 만한 일련의 결과를 얻을 수 있다. 예를 들면, 관찰을 통해 우리는 돌연변이율을 알 수 있고, 생물의 생식률에 대해서도 알 수 있는데, 그러한 조건들이 주어지면 진화론자들은 그것을 토대로 하여 어떤 결과를 예측할 수 있게 된다. 또 찰하
면 결과 자체뿐 아니라 그러한 결과의 확률분포함수까지도 작성 할 수 있을 것이다. 얼마 후에 우리는 결과가 실제로 어떻게 나 타나는지를 관찰할 수 있게 되는데 그때, (1) 전화론자들의 예측 이 과연 용납될 수 있는 결과의 범주 속에 놓였는지, (2) 그렇다 면 그러한 결과에 대한 그들의 예상 확률은 얼마였는지롤 알게 될 것이다. 만일 상당히 높은 확률을 보인다면 전화학자들에게는 줄거운 일이 될 것이고, 만일 그 확률이 변변치 못하다면 확률을 높일 수 있는 적절한 방법을 찾으려 할 것이다. 그러한 방법을 찾을 수 없거나 혹은 그들의 예측이 용납될 수 없는 결과에 속하게 된 다면 법칙이나 자료의 수치 혹은 그러한 자료들을 사용하는 방법 에 문제가 있었을 것이다. 전화론자들은 어떤 구체적인 결과~ 예 측하려고 고집하기보다는 전반적으로 볼 때 가능성이 높고 용납 될 수 있는 결과에 대개 만족해 한다. 왜냐하면 그러한 결과들은 대개 비슷비슷해서 용납될 수 있는 결과의 범주에 속하기만 한다 면 그 범주 안에서 일어나는 차이점들은 그리 대단한 것이 아니 라고보기 때문이다. 그러나 만일 우리가 태양계 전체의 미래(혹은 과거)의 상황을 추론하고자 한다면 천체역학의 경우도 전화론에 비해 그리 나울 게 없다고 반박할 사람이 있을지도 모른다. 태양계 내의 모든 천 체를 동시에 다루는 것이 어떤 특정 종의 전화에 영향을 주는 요 소들을 모두 함께 다루는 것보다 결코 더 쉬운 일은 아닌 것이 다. 또한, 전화의 시스템이 부분적으로는 폐쇄될 수 있어도 완전 한 폐쇄는 불가능한 것처럼, 태양계의 폐쇄성도 거대한 혜성이라 든지 그와 비슷한 것에 의해 언제든지 파괴될 수가 있다. 그러나
실제로 이 두 상황의 차이점은 따로 있으며, 그것을 지적해 냄으 로써 그러한 반대 의견에 대한 답을 줄 수 있을 것이다. 천체역 학에서는 단지 태양과 알고자 하는 어떤 특정한 혹성 하나만을 고려하여도 그 혹성의 위치를 아주 정확하게 예측할 수가 있다. 왜냐하면 다른 혹성이 그 혹성에 미치는 영향은 태양의 그것에 비하면 무시될 수 있을 만큼 작기 때문이다. 만일 디른- 혹성의 영향까지도 알아보고 싶다면 그 혹성들을 하나하나씩 따로따로 고려해 나가면 된다. 그러나 전화의 경우에는 그 수많은 요소 중 에서 어느 하나가 나머지보다 훨씬 중요해서 그의의 다른 요소들 울 모두 무시할 수 있는 경우란 거의 없다고 보아야 할 것이다. 그러므로 천체역학에서는 단지 두 천체 사이의 문제이고 따라서 그것은 쉽게 해결될 수 있지만, 전화의 경우에는 결코 어떤 종과 어떤 하나의 요소 그 둘만의 문제가 아닌 것이다. 창시자원리 환경이 전화 단위에 주는 영향이 다양하고 불규칙적이기 때문 에 야기되는 여러 문제들, 또 어떤 진화 단위에 대해서 예측을 하기 위해 우리가 가지고 있는 자료를 과연 어떻게 종합할 수 있 울 것인가 하는 어려운 문제들은 접어두더라도, 자연 집단이라는 것은 크기가 너무 작아서 우리가 앞에서 언급한 수많은 무작위적 요소들 중의 하나 혹은 몇몇이 아주 중요한 문제점으로 등장하는 경우가 많다. 그러나 메이어 (Erns t May r, 1963) 는 그러한 소집단 들이야말로 진화에 결정적인 역할을 하는 존재들이며 특히 유성 생식을 하는 생물의 경우에는 더더욱 중요하다고 하였다 .• 그 에 의
하면, 생태적인 장벽이나 지리적인 장벽이 생겨서 유전자의 교환 이 방해를 받게 되는 경우에는 종이 두 개 혹은 여러 개로 나누 어지게 된다. 그런데 종종 그렇게 분리된 집단의 크기가 너무 작 아서 유전적 구성이 다양하지 못한 경우가 생기며, 그런 경우에는 그 종이 지니고 있는 유전자형의 상당 부분이 자연선택될 처지에 놓이게 된다. 따라서 크기가 큰 집단에서는 전혀 문제가 되지 않 던 여러 요소들이 이렇게 작고 격리되어 있는 집단에서는 갑자기 대단히 중요한 존재가 되는 것이다. 그러한 집단의 유전적 구성은 유전적 부동(浮動)에 의하여 더욱 축소가 되기도 하는데 이는 유 전적 부동으로 인하여 대립유전자들 중의 일부가 제거될 수 있거 때문이다. 동계교배(同系交配)의 기회가 늘어남에 따라 동형집합체 의 수가 함께 증가하는 것은 하디-와인버그의 법칙을 통해 이미 우리가 알고 있는 바이며, 원래의 모집단에서는 상위 유전자로서 서로 작용하던 유전자들 중 몇몇이 없어지는 데 따라 남아 있는 유전자의 선택값도 변하게 되리라는 것은 너무도 당연한 일이다. 이러한 집단이 존속되려면 그렇게 크기가 작은 채로 남아 있어 도 안 되고, 또 오랜 시간 그렇게 격리된 상태로 있어서도 안 되 며, 아주 빠른 속도로 팽창해야만 한다. 만일 그렇게 격리된 채 시간이 흐르고 생식적으로도 격리가 된다면 그 종은 결국 새로운 종으로 진화하게 될 것이다. 아니면 원래의 집단에 다시 합쳐져 더욱 큰 집단이 되면서 대단히 많은 유전자들이 출현하게 될 수 도 있다. 어떤 경우이건 이러한 현상은 대단히 중요한 것이어서 메이어는 그렇게 빈약해전 유전적 구성이 집단의 발달에 미치게 될 영향을 창시자원리 the fou nder p r i nc ip le 라고 이름하였다. 만 일 우리의 목적이 어떤 특정 계보의 전화, 죽 어떠한 방해도 받 지 않고 계속 진행되는 진화를 예측하고자 하는 것이라면, 이 창
시자원리는 여러 제거할 수 없는 무작위적 요소들을 전화에 끌어 둘이게 된다는 데에 그 중요성이 있다. 따라서 우리는 어떤 생물 이 살아 남을 것인지 아닌지에 대해 전혀 아무런 예측을 할 수 없는 것이다. 그러나 한 가지 분명한 것은 설혹 어떠어떠한 생물 체가 새로운 집단의 기초가 되고 또 그들의 유전적 구성이 이러 이러하다는 것까지 다 알고 있다 하더라도, 어떤 형태의 집단이 어떤 식으로 발전해 나갈 것인가는 전혀 예측할 수가 없다는 것 이다. 왜냐하면 거기에 관여된 개체의 숫자가 너무 소규모이기 때 문에 바늘 구멍을 통해 전체를 다 파악하고자 하는· 격이기 때문 이다. 메틀러 L. E. Me ttl er 와 그레그 T. G. Gre gg는 그러한 상황 울 다음과 같이 요약하였다. 유전자형의 종류는 이루 헤아릴 수 없이 많지만 실제로 나타나 는 것은 아주 적은 숫자에 불과하다. 집단은 유한한데, 유전자형 의 종류는 상상할 수 없을 정도로 많기 때문에 이론적으로 가능 한 유전자형의 조합의 대부분은 현실화되지 않는다. 그런데 어떤 유전자형이 나타나느냐 않느냐 하는 것은 순전히 우연의 문제이 다. 심지어 어떤 것은 일단 만들어졌다가도 훗날 뜻하지 않게 우 연히 없어지기도 한다 .5)
5) Pop u lati on Geneti cs and Euoluti on (Eng el wood Cl iffs; N. J. : Prenti ce -Hall, Inc., 1969), p. 89.
이 장에서 우리는 전화론이 지닌 통계적인 특성의 요인이라고 할 수 있는 세 가지 점에 대해 살펴보았다. 가장 중요한 문제는 진회론에 관한 우리의 지식이 부족하고 또 전화론이라는 것이 언 제 부정될지 모를 원리 속에 존재한다는 데 있다. 그러면서도 집
단과 같은 특정 전화 단위에 전화론을 적용하기 위해서는 끊임없 이 일어나고 있는 변화를 실황중계라도 해야할 것처럼 보인다는 것이다. 그러나 그러한 변화와 관련된 특정 상황이나 한계 조건이 라는 것이 너무 광범위하여 역사 이야기의 한 부분이 될 지경이 다(제 3 장 참조). 또한 우리가 설혹 좋은 자료를 가지고 있다 하더 라도 현재로서는 그것을 제대로 요리할 수도 없는 형편이다. 그것 은 아마도 생물학자들이 적절한 전화 단위와 전화에 있어 가장 중요한 요소들을 가려내는 데 실패했기 때문일 것이고, 한편으로 는 적절한 수학적 도구가 없기 때문이기도 할 것이다. 그러나 이 런 것들이 다 해결된다고 하더라도 또다시 우리는 창시자원리라 는 문제에 봉착하게 된다. 수많은 분자로 이루어전 폐쇄계에서는 극히 정밀한 기체 법칙도 분자의 수가 감소할수록 점점 부정확해 지고 결국에는 아예 적용할 수 없게 된다. 그런데 창시자원리는, 어떤 특정 집단 또는 종의 진화에 관련된 법칙들이 아주 작은 집 단, 죽 그 불확실성의 정도와 다양성에 비추어 크기가 대단히 작 은 집단에도 반드시 적용될 수 있어야 한다는 데에 문제가 있는 것이다. 이렇게 보면 특히 창시자원리만이 전화론은 반드시 통계 적이어야 한다고 주장하고 있는 것 같다. 진화론의 연역적 성질 앞에서 우리는 전화의 법칙 속에서 다루어져야 할 분류군은 집 단이나 종과 같은 전화 단위들의 시간적 단편이라고 생각한 생물 학자들과 의견을 같이 했었다. 생물학자들은 만일 그러한 분류군 들이 적절한 것이고 예측하고자 하는 것이 이들이 앞으로 어떻게
발전할 것인가 하는 것이라면, 우리가 할 수 있는 것이라고는 기 껏해야 확률적인 예측뿐이라는 것을 인정하지 않을 수 없었다. 그 러나 만일 진화론의 기능 단위를 바꿔보고, 또한 하고자 하는 예 측의 형태를 바꿔본다면, 진화론을 이 책의 앞부분에서 설명했던 연역적 과학 이론의 개념과 모순되지 않는 형태로 구성할 수 있 을지도 모른다. 윌리 엄스 Mar y B. W illi ams6 ) 는 진화론의 공리화 (公理化)라는 논문 속에서 바로 이러한 일을 시도하였다.
6) Deducin g th e Conseq u ences of Evoluti on : A Math emati ca l Model, Jo urnal of Theoreti ca l Bi ol og y, 29(1 9 70), pp. 343-385.
여러 철학자들과 생물학자들은 전화론 속에도 연역의 핵심이 되는 요소들이 들어 있다는 문제를 가지고 논란을 거듭해 왔으나, 사실상 그들이 주장하는 연역이란 몇 개의 기본 원리로부터 전화 가 일어날 것이라는 사실을 연역할 수 있다는 것뿐이다. 진화론이 연역적이라는 주장이 의미 있는 것으로 받아들여지기 위해서는 이보다는 더 낫고 더 상세한 연역이 필요할 것이다. 또한 전화론 의 여러 공리를 포함하고 있는 연역적 예측둘을 만들어야 한다. 예를 들면, 진화는 점진적인가, 도약적인가, 아니면 양자가 조금씩 혼합된 것인가? 두 집단 사이에 유전자의 교환이 있을 때에도 종 분화는 일어나는가? 집단의 크기를 환경의 수용 능력을 넘지 않 도록 조절하는 그러한 유전적 형질에 대해서도 선택이 일어나는 가? 종분화에는 여러 단계가 있는데, 현존하는 종 가운데 그러한 각각의 단계에 속한다고 볼 수 있는 것은 몇 퍼센트나 되는가? 하는 것 등이다. 전화의 기능 단위에는 여러 단계가 있다는 사실 을 인식하고 그것들이 서로 어떻게 연관되어 있는가를 조심스럽 게 구체화함으로써, 윌리엄스는 전화론을 그녀의 방식대로 연역적 으로 구성하고 그러한 구성으로부터 여러 가지 다양한 예측을 유
도해 낼 수 있었다• 공리화 과정에서 그녀가 목표했던 것은 〈 다 윈의 진화론을 연역적 시스뎀으로 표현하는 것인데 진화론의 기 본 원리들 중 몇 개는 그러한 시스템 안에서 공리로써 사용되고 전화론의 나머지 원리들은 그러한 공리로부터 연역적으로 도출해 낼 수 있음 〉 을 보이는 것이었다. 그렇게 해서 〈 다윈의 진화론을 정확 간결하며 검증 가능한 형태로 기술할 수 있도록 그 기초를 제공할 수 있으며, 그러한 전화론을 바탕으로 하여 완벽한 진화론 울 정확 간결하고 검증 가능하게 기술할 수 있을 것〉이라고 하였 다 .7 )
7) 앞의 책, pp. 343 전 44.
윌리엄스의 공리화 내용 중에서 가장 중요한 요소는 기본적인 단위, 즉 그녀가 클랜 clan, 다윈 서브클랜 Dar wini an subclan, 그 리고 서브클랜 subclarr 이라고 명명한 단위에 대한 정의이다. 〈클 랜이란 몇 개의 ‘창시자’ 생물의 후손들을 모두 포함하는 하나의 집단으로 정의되는데, 말하자면 분류군을 일컬을 때 그것은 현존 하는 생물들뿐 아니라 창시자 생물과 현존하는 생물 사이에 존재 했던 세대들 모두를 의미한다.〉 다윈의 서브클렌은 클랜의 한 부 분으로 〈 어떤 결속력에 의해 결합되어 있기 때문에 이것은 선택 이 작용하는 하나의 단위로서의 역할을 하게 된다〉. 그러므로 윌 리엄스의 법칙은 클랜이나 다윈 서브클렌을 전체, 즉 그러한 전체 를 시간적으로 쪼갠 단편적인 것으로서가 아니라 시공간적인 전 체로서 다루고 있다. 또한 윌리엄스는 이러한 법칙들이 단지 이미 확립되어 있는 그룹에만 적용된다고 그 한계를 정하고 있다. 따라 서 확립되지 못한 〈서브클랜〉은 서브클랜으로 간주되지 않는다. 이러한 두 가지 이유로 인해 윌리엄스는 메이어의 창시자원리에 수반되었던 우연성을 제거할 수 있었다.
그러나 역시 전화론은 진화되고 있다고 여겨지는 모든 현상들 울 다뤄야만 한다고 불평하는 사람이 있을지도 모른다. 뉴턴의 법 칙이 모든 물체를 다루고 있는 것처럼, 다윈의 전화론도 모든 생 명체를 다 다루어야 하며, 살아 있는 생명체는 어떤 진화 단위에 든지 포함될 수 있어야 한다는 것이다. 그러나 과학 이론은 종종 그 범위가 제한되기도 한다. 예를 들어, 전화의 유전 아론은 멘델 집단% 士 을 디루는데, 그러한 집단은 생식을 통해 유전자를 상호교 환하는 개체들로 이루어전 집단이라고 할 수 있다. 그러나 최소한 윌리엄스의 공리화는 진화론의 주체라고 여겨졌던 현성들- 중 어 떤 것이, 또 얼마나 많은 것이 연역적으로 구성된 전화론의 일부 가 될 수 있는가를 보여주고 있다. 또한 윌리엄스의 공리화는 각 단계가 진화에 미치는 중요성과 어떻게 그들이 서로 연관되어 있는지를 강조하고 있다. 유전자, 염색체, 배우자, 개체, 집단, 종 등 여러 단계는 모두 전화에 있어
® 멘델 집단. 전화요인의 하나로서 유전자 풀p ool 의 변동이라는 개 념이 있는데, 여기서의 유전자 풀은 한 생물 종의 집단 속에 있는 모든 개체의 모든 유전자 전부를· 가리키는 말이다. 그리고 이 집단 이라는 것은 하나의 가상적인 집단으로서, 집단이 충분히 크고, 그 집단 속의 유전자가 집단 밖으로 유출되거나 의부로부터 유입되는 일이 없이 항상 일정한 완전 격리시스템이고, 그 집단의 구성원 속 에서는 돌연변이가 일체 없고, 구성원 사이의 교배는 완전히 우연 에 의하여 일어나며, 모든 구성원의 생식 능력이 똑같은 그러한 집 단을 말한다. 이러한 집단을 멘델 집단이라고 하는데 이 집단은 이 론상의 집단이고 현실적으로는 존재하지 않는다. 한 집단이 멘델 집단일 경우 그 집단에서는 전화라는 것이 전혀 일어나지 않겠지 만, 이 집단의 요소 가운데 어떤 하나 또는 둘 이상이 파괴되면 진 화가 일어날 수 있을 것이다. 따라서 · 진화는 한 집단 속의 유전자 풀의 변동이 일어났을 때 생긴다고 설명할 수 있다.
서로 다른 역할을 하고 있으나 한편으로는 서로 밀접하게 관련되 어 있다. 많은 사람들이 적자생존의 원리가 동의반복(同義反復)이 라고 주장하는 주된 원인 중의 하나는 그들이 이러한 사실을 제 대로 깨닫지 못하고 있기 때문이다. 생식을 통하여 전달되는 것은 유전자를 지닌 염색체뿐이며, 유전될 수 있는 돌연변이는 전화의 필요조전이다. 그러나 환경과 상호 작용을 하면서 생식기까지 살 아남거나 살아 남지 못하는 것은 개체이다. 생존은 생식의 필요조 건이며 생식은 유전자를 전달하기 위한 필요조건이다. 그러나 이 러한 과정에 의해 변화되는 것은 집단이다. 윌리엄스가 설명한 바 와 같이 자연선택은 〈 이상한 힘이다. 왜냐하면 우리가 다루는 힘 은 보통 그 힘이 작용하는 대상의 특성을 변화시키는 것이다. 예 를 들어, 당구공에 힘을 가하면 그 힘으로 인하여 당구공의 운동 상태가 변화된다. 선택도 유전적 특성을 변화시키는 힘이다. 그러 나 이러한 변화는 그러한 힘의 대상인 개체 수준에서 일어나는 것이 아니다. 말하자면 나쁜 형질을 지닌 개체를 벌주려고 할 때, 그 개체로 하여금 자손을 갖지 못하게 하는 식으로 벌을 줄 뿐이 고 그 개체가 이미 거기에 있는 이상 힘의 대상인 그 개체 자체 의 형질을 변화시킬 수는 없다〉 .8)
8) Wi lliam s, The Log ica l Sta t u s of Natu ral Selecti on and Ot he r Evoluti on ary Contr o versie s , The Meth o dolog ica l Unit y of Scie n ce(Dordrecht : D. Reid e l, 1973).
윌리엄스의 업적 중 두번째로 중요한 것은, 진화론을 기초로 하여 만들어낼 수 있는 예측 형태는 어떤 특정 개체나 개체의 성 질, 또는 분류단위 등에 관한 것이 아니라 전화 양상에 관한 것임을 명쾌하게 기록하였다는 것이다. 전화론은 예를 들어 백조 (Cy gn us olor) 라는 특정 종의 진화를 다루는 것이 아니라 종 전반
의 진화를 다루는 것이다. 과거에도 전화론자둘은 수많은 전화 양 상에 관심을 기울여 왔는데, 그들의 관심거리가 되었던 전화 양상 의 한 예로 아직 아무 종에도 점령되지 않은 어떤 생태계에 어떤 종이 들어가 그곳에 맞도록 특수화되는 현상 같은 것을 둘 수 있 다. 진화론을 근거로 하여 그들은 백조라는 종은 이들 진화 양상 중의 어느 것에 해당한다리는· 정도까지 자세하게 백조에 대하여 예 측을 할 수가 있었던 반면에, 모든 백조는 희다리~ 예측을 할 수 는 없다(모든 백조들이 희지는 않기 때문에)는 사실 때문에 너무 오랫동안 시달려 왔다• 그러한 형질을 나타내는 특정한 종에 연연 해 할 필요없이, 우리는 다만 의태(擬態)나 보호색과 같은 전화적 형질 그 자체에 한해서만 예측할 수 있을 뿐이고 또 그래야 한다 는 것을 그들은 깨닫지 못했던 것이다. 전화론을 기초로 한 예측의 전통적인 예로 이형집합체(異型接 合)의 우위성(優位性)을 들 수 있다. 열대성 말라리아가 만연하 는 지역 주민들이 갖고 있는 헤모글로빈 S 유전자의 이형접합체 가 그 한 예이다. 이러한 지역 주민 중, 정상적인 헤모글로빈 유 전자의 동형접합체를 가전 사람은 이형집합체인 사람보다 말라리 아에 걸리기 쉽고, 헤모글로빈 S 대립유전자가 동형접합체인 사 람들은 낫세포 빈혈증(鎌狀赤血球性貧血症)이라는 병에 걸려 죽는 확률이 높다. 우리는 하디-와인버그의 법칙으로 무장한 멘델 유 전학의 원리만을 가지고도 어떤 특정 집단에 있어서의 여러 유전 자 빈도와 유전자형의 빈도를 알기 위해 필요한 정보를 쉽게 얻 어낼 수 있다. 그러나 전화적 예측은 특정한 상황으로부터 선택압 력이 제거되었을 때 어떤 일이 일어나겠는가 하는 일에만 관심을 둔다. 예를 들면, 어떤 개체군들이 열대성 말라리아가 만연하는 지역에서 그렇지 않은 지역으로 이동되었을 경우이다. 그렇게 되
면 다음 세대에서는 헤모글로빈 S 대립유전자의 빈도가 급격히 줄어들게 될 것이다. 그러나 인간에게서 일어나는 이러한 변화 그 자체는 전화론에 기초한 진화적 예측의 요소로는 부적당하며, 말 라리아와 낫세포 빈혈증이라는 두 질병이라는 요소 역시도 부적 합하다. 전화적 예측에 적합한 요소는 이 지역에 있어서의 이형집 합체의 우위성과 그에 따른 두 대립유전자의 상대적 빈도일 뿐이 다. 그리고 이 경우 상대적 빈도란 개체의 특성이 아니라 집단의 특성 01 다. 자연선택과 적자생존 전화의 역사를 통하여 가장 심각하고 지속적이었다고 할 수 있 는 비판은 자연선택이나 적자생존의 원리가 동의반복이라는 것이 다. 일반적으로 과학 이론 속의 어떤 특정한 전술이 동의반복인가 아닌가를 결정하는 것은 쉽지 않은 작업이며, 그 예로서 뉴턴의 제 2운 동법칙을 둘러싼 논쟁을 둘 수 있다. 많은 사람들이 적자생 존의 원리가 동의반복이기 때문에 전화론 그 자체가 모두 잘못된 것일 수 있다고 느끼면서도, 어떤 이유에서인지 뉴턴의 이론에 대 해서는 그러한 주장을 하지 않는다. 그러나 어쨌든 적자생존의 원 리를 동의반복으로 해석하느냐 아니냐는 것은 각자의 자유이지만, 그렇게 된다면 그것은 전화론을 검증할 수 있는 중요한 방법을 제거해 버리는 결과가 된다. 많은 생물학자들은 자연선택이란 곧 유전자들이 서로 다른 정 도의 영속성을 지니는 것이라고 주장한다. 그렇다면 유전자를 환 경에 더욱 찰 적응하도록 만드는 것은 무엇인가? 그것은 다음 세
대의 유전자 풀p ool 에서 그 유전자의 상대적 빈도를 증가시키는 일이다. 유전자들은 각각이 분리되어 전달되는 것이 아니라 유전 자형으로 전달되는 것이기 때문에 신중한 생물학자들은- 자연선택 이란 유전자가 아니라 유전자형들이 서로 디론· 정도의 영속성을 지니는 것으로 해석해야 한다고 말한다• 그렇다면 무엇이 유전자 형을 환경에 더 잘 적응하도록 만드는가? 그것은 다음 세대에 있 어서 그 유전자형의 영속성을 증가시키는 일이다. 생체론자인 생 물학자라면 자연선택은 바로 생식능력의 정도라고 말할 것이다. 죽 무엇이 생물체를 환경에 더욱 잘 적응하도록 만드는가? 더 많 은 자손을 남기는 일이다. 그러나 자연선택의 원리에 관한 이러한 제반 해석은 모두 몇 가지의 공통점을 가지고 있는데, 즉 그러한 해석들은 모두 전화의 필요조건을 자연선택에 의해 구체화하고 있다는 점과 필요조건을 마치 충분조건처럼 다루는- 오류를 범하 고 있다는 점, 그리고 차별적 영속성의 원인인 인과론적 기작을 전혀 언급하지 않았다는 점 등이다. 위에서 언급한 세 가지 해석 울 하나하나씩 따져 보면 그 결과는 동의반복으로 끝나고 말 것 임이 틀림없다• 유전자와 유전자형이 지닌 영속성이 서로 그 정도가 다르다는 사실은 분명히 필요한 일이다. 유전자형 없이 표현형이 있을 수 없고, 물론 표현형이 없다면 유전자형 역시 있을 수 없다. 또한, 이 모든 차별적 영속성은 자연선택의 결과지만, 자연선택은 생물 개체와 환경 사이의 관계이다. 만일 유전자, 유전자형, 표현형, 개 체, 환경 사이의 이러한 상호 관계를 무시한다면 경험적 내용들 중 많은 부분이 진화론에서 삭제되게 된다. 예를 들어 환경과의 관계를 무시한다면, 앞에서 예로 든 열대성 말라리아에 관련된 유 전자의 상대적 빈도나 낫세포 빈혈증의 증가 경향 따위는 무의미
하게 될 것이다. 이제 독자들은 개체와 환경 사이의 관계가 갖는 인과적 역할의 중요성을 인정하면 어떻겠느냐고 할지도 모르겠다. 그러나 그런 마음을 갖고 적자생존의 원리를 해석한다 해도 동의반복적이기는 마찬가지이다. 어떤 개체는 한 형질 혹은 일단의 형질 때문에 특 정 환경 속에서 살아 남을 수 있고 다론 개체는 또 다른 환경 속 에서 또 다른 형질 혹은 또 다른 한 세트의 형질 때문에 살아 남 을 수 있다. 그러나 실제로 살아 남은 개체들은 살아 남았기 때 문에 잘 적응할 수 있는 기회를 갖는 것이고 그렇게 적응된 개체 들은 또 적응되었기 때문에 살아 남는 것이며, 이 경우, 적응성의 정도는 생식 가능한 자손을 얼마나 낳았는가에 의해서 판단된다 는것이다. 하지만 이에 대해 몇 가지 반대 의견이 생길 수 있다 . 그런 식 으로 해석을 하자면 예를 들어, 일벌은 모두 적응도계수가
분으로 인식하는 것이다 . 다른 하나는 이러한 여러 개체로 이루어 진 집단인 개체군 전체를 하나의 〈 개체 〉 로 간주하는- 것으로, 예 룰 들면 벌집이 개체가 되고 여왕벌, 수필, 일벌 등은 그 개체의 한부분으로 생각하는 것이다. 그러나 〈 적응도(適應度) 〉 와 〈 생촌(生存) 〉 을 구별하는 데 있어 서의 이러한 경험적 어려움을 모두 무시한다 하더라도, 애초에 그 러한 구별을 하게끔 우리를 유도해 갔던 형이상학적 집착은 그대 로 남아 있으며, 이러한 사실 자체가 바로 형이상학적인 집착이라 고 볼 수 있다. 적응도에 대한 주장은 어떤 특정한 형질이나 일 련의 형질에 관한 것이었지 그 개체의 〈 모든 〉 형질은 아니었던 것이다. 특정한 환경, 즉 건조기가 닥쳐왔을 때 한 개체가 다른 개체보다 더 잘 적응한디는 것은 그 개체가 다른 개체보다 건조 한 환경을 더 찰 견디기 때문이며, 전자가 후자보다 이러한 하나 의 형질을 더 발전시켰기 때문에 생존할 수 있는 보다 나은 기회 롤 갖게 되는 것이다. 만일 환경의 변화와 함께 유전적 구성, 발 생학적 발달 과정 , 관련된 생물들의 생리 모두를 충분히 알 수 있다면 생물 각각의 적응도를 알 수 있게 될 것이고 따라서 그들 의 생존확률에 대한 어느 정도 정확한 예측을 할 수 있을 것이 다. 그리고 이러한 정보를 토대로 하여 앞으로 그 집단이 어떻게 변화할지도 예측할 수 있을 것이다. 다음으로 전회론자들이 해결해야만 했던 문제는 전화적 발전의 단계 level들 을 찾아내는 것이었는데, 여기서 전화적 발전이란 것 은 시공간적으로 다른 상황에서도 동일한 사건들이 반복되어 일 어날 수 있고 또 전화법칙이 형식화될 수 있도록 충분히 넓은 의 미로 쓰여져야 하는 동시에 그러한 법칙으로부터 합리적인 추론 을 유도해 낼 수 있도록 아주 정확해야 하는 것이다. 만일 생물
과 그 생물의 주변 환경이 너무 좁은 의미로 기술된다면, 사건들 이 너무 특수화되어서 우주의 역사상 단 한 번밖에 일어나지 않 는 사건이 될 것이며, 또 만일 기술이 너무 광범위해지면 법칙이 너무 엉성하고 산만해져서 아무런 역할도 아무런 의미도 없게 될 것이다. 따라서 그 의미가 아주 적절한 범위를 지켜야 하는데, 집 단과 같은 전화 단위에 있어서는 전화적 발전에 관련된 법칙들이 보편적이고 일반적인 형태를 갖추고 있지 못한 형편이다. 또 하나 는 정확성의 문제인데, 우리가 확률적인 예측에만 관심을 둔다 하 더라도 그 예측은 항상 정확하게 기술되어야 한다. 그러나 너무 자세하게 기술되다 못해 사건들이 모두 유일무이한 것이 되어버 린다면 기술의 정확성이 좀더 정확한 예측을 하는 데 아무런 도 움도 줄 수 없는 상황에 이르게 된다. 그러므로 어떤 확실성을 가지고 특정한 종이나 집단의 전화적 발전을 예측한다는 것은 불 가능하며, 다만 어떤 집단이나 종이 현재까지 알려전 전화 양상 중의 하나에 들어 맞는다 하는 정도의 예측만이 가능할 뿐이다. 그러나 그렇게 너무 광범위한 문제들에 매달리게 되면, 전화에 관련된 변수들 모두를 알지는 못하게 될 것이고 설혹 안다 하더 라도 그러한 변수를 의미있게 연결시킬 수가 없게 될 것이다. 그 러나 다행히도 자연이 우리를 위하여 그 모든 것에 대해 개괄적 인 요약을 해주고 있다. 원리상으로는 문젯거리를 아무 것도 남기 지 않고 죽어버리는 생물은 적응도계수가 〈 O 〉이다. 따라서 두 개 체가 같은 유전자형을 가전 일란성 쌍동이라 할지라도 얼마나 자 손을 많이 남겼는가에 따라 하나는 대단히 높은 적응도계수를 갖 고 다른 하나는 아주 낮은 적응도계수를 가질 수 있다. 이러한 소급적 결정론적인 편견에 호감이 가는 것을 피하기는 어렵지만, 만일 한 개체가 자손을 남기지 않고 죽은 반면 다론 개체는 수
많은 자손을 남기는 데 성공했다면 분명히 그러한 차이점이 생기 게 된 이유가 있을 것이다. 우리가 〈 완벽한 〉 지식과 모든 것을 포괄할 수 있는 이론을 가지고 있을 때에 한해서 우리는 절대적 인 정확도로써 생물체 각각의 운명을 예측할 수 있을 것이고 그 러면 적자생존은 동의반복에 지나지 않게 될 것이다. 이제 완벽한 기술(記述), 총체적 지식, 보편적 인과성의 원리와 같은 개념들로 관심을 돌려 조금 더 얘기를 해야 할 필요가 있을 것 같다. 우리가 아는 한 완벽한 기술이라는 개념은 하나로 된, 모든 것을 포괄하는 과학 이론을 전제로 하며 이러한 이론에 의 해 이루어전 기술은 완벽한 것으로 간주될 수 있다. 총체적인 지 식의 경우도 같은 식의 이야기를 할 수 있을 것이다. 완벽한 기 술이나 총체적인 지식에 대한 개념 중에는 이보다 더 강력한 개 념들도 있지만 그들은 모두 절대주의자들의 형이상학에 근거하고 있으며 그러한 형이상학은 많은 문제점을 지니고 있다. 예를 들 어, 우리가 총체적인 지식을 지니고 있다면 과학은 존재 의미가 없게 될 것이고, 이미 모든 것을 다 알고 있다면 무엇인가를 추 론할 필요도 없을 것이다. 어떤 경우이든 간에 전화론은 그렇게 모든 것을 포괄하는 이론은 아니다. 그 해석이 상당히 인위적인 방향으로 치우천 전화론에서는 적자생존의 원리가 동의반복이 될 것이며, 따라서 이러한 해석은 곤란한 입장에 놓이게 될 것이다. 그러나 생물과 환경의 관계가 전화에 결정적으로 중요한 역할을 한다는 식으로 전화론을 해석하는 경우에는 결코 동의반복이 아 닐것이다.
잭 생물학적 이론과 생물학적 법칙 과학법칙과 서술상의 일반화 앞에서 우리는 유전이론과 진화론에 대해 이미 살펴보았는데 여기서 새삼스럽게 생물학적 이론이나 법칙이 존재하는가라는 질 문을 던진다는 것은 불필요한 일처럼 보일지 모른다. 그러나 여태 껏 논의해 온 이론이나 법칙들이 지닌 몇 가지 특칭들로 인하여 스마트(J.J .C. Smart, 1963 : 50) 와 갇은 철학자들은 〈물리학에 서와 같이 세련된 법칙들이 생물학에는 없다〉, 심지어는 〈생물학 에는 법칙이 존재하지 않는다〉고까지 주장하고 있다. 그렇다면 멘델 유전학, 분자유전학, 전화의 유전이론, 종합 전화론, 다윈 이론에 대한 윌리엄스의 공리화, 그리고 레빈스 Le vi ns, 레원틴 Lewonti n, 맥아더 MacArth ur, 윌슨 W il son 등의 집단생물학 이론 은 어떤가? 그것들은 자연과학의 이론이 아니라는 말인가? 그것 둘이 생물학적인 법칙을 포함하고 있지 않다는 말인가? 스마트 같은 철학자들은 위에서 얘기한 것들이 생물학적 이론이 될 수 없는 이유로, 전정한 의미의 과학 이론으로서 필요한 것을 갖추고
있는 경우에는 그것들이 정말로 생물학적이라 보기 어렵고, 또 그 것이 진정으로 생물학적인 경우에는 전정한 의미의 과학 이론이 아니기 때문이라고 밀하고 있다• 전형적인 생물학적 이론이나 법칙에 대한 대부분의 반대 의견 에는 두 가지 문제가 내포되어 있다 . 첫째는, 생물학 문헌 속에 나오는 법칙들은 과정의 법칙이 아니라는 것이다. 다시 말해서, 어떤 특정한 시간에 대한 변수의 값이 주어졌을 때에도 그 시스 템의 과거나 미래에 관해 추론을 할 수가 없다는 것이다. 그러나 이것은 생물학적 시스템이 그러한 법칙을 공식화할 수 있을 만큼 충분한 폐쇄계가 아니기 때문이다. 생물학 문헌에는 다른 형태의 일반화가 그러한 과정의 법칙을 대신하고 있는데, 인과법칙 casual law, 발생법칙 develop m enta l law, 역사법칙 hist o r ic a l law 등이 그것이다. 어떤 사람들은 그러한 일반화를 쉽게 잊는 경우가 있는 데, 이 장에서 우리는 잠정적이지만 일단은 생물학적이라고 생각 할 수 있는 이러한 법칙들이 과연 진정한 의미에서의 과학법칙으 로 간주될 수 있겠는지 살펴보게 될 것이다. 과정의 법칙이야말로 우리가 도전해야 할 이상적인 법칙이라고 생긱하는 · 사람도 있울 지 모르나, 이상적 상태에 미치지 못하는 법칙일지라도 잘 살펴보 고 지나가야 하는 데는 충분한 이유가 있다. 특히 그것이 예측 가능한 미래를 다루는 과학 분야이거나 그 한 부분일 경우에는 더욱 그러하다. 생물학적 이론이나 법칙들이 비록 완벽하지 못한 점이 있다 하더라도 그 중에는 다른 것보다 훨씬 나은 것도 있는 것이다 . 그래서 이러한 〈불완전〉한 법칙이나 이론을 서로 비교하 고 분석하는 것도 충분한 이유가 있다고 생각된다. 과정의 법칙 이의의 법칙들까지도 과학법칙으로 간주할 수는 있겠지만 그래도 여전히 두번째의 문제, 죽 서술상의 일반화와 과
학법칙을 구별하는 데 있어서의 어려움은 그대로 남아 있다. 초대 부터 7 대까지의 미국 대통령 중 기독교인이 한 명도 없었다는 것 은(그들은 이신론자(理神論者, de i s t)였다) 사실이긴 하지만 그렇 다고 그것이 자연법칙이 될 수는 없다. 과학철학에 있어 이러한 구별은 대단히 중요하지만, 그 사이에 적당한 경계선을 긋는 일 또한 쉽지 않다. 이를 위해 수많은 기준이 제시되었으나 가능성을 보여준 유일한 것이라면, 서술상의 일반화가 고립된 전술의 형태 를 갖춘 채 종국적으로 자연법칙 이론으로 통합되어 들어갈 수 있는가 없는가를 알아보는 것이다. 예를 들어, 여러 혹성과 태양 사이의 거리를 다루는 보드 Bod#! 법칙과 그 혹성들의 궤도와 속도를 다루는 케플러의 법칙도 한때는 한 개의 태양계에만 국한 된 고립된 일반화였다. 그 중에서 보드의 법칙은 결국 일반화가 될 수 없었고 따라서 더 이상 전정한 의미에서의 법칙으로 간주 될 수가 없었다. 태양과 여러 혹성 사이의 상대적인 거리는 태양 계에서 일어난 우연적인 일 내지는 역사적인 현상으로 간주되었 다. 즉 태양계가 지닌 이러한 특칭은 그러한 특칭을 만들어내게 된 특수한 상황에 의한 것일 뿐, 이러한 상황이 행성계의 형성 전반에 걸친 전형적인 형태라고 예측할 아무런 이유가 없었던 것 이다. 이와는 반대로 케플러의 법칙은 실제로 뉴턴 이론에 병합될 수가있었다. 그러나 과학은 계속 발전해가고 있기 때문에, 어느 일정한 시 점에서 볼 때는 모두 진리라고 확신할 수 있는 보편적 일반화라 하더라도 실제로는 단순히 서술적이거나 우연적인 것에 불과한 것도 있을 수 있고, 또 새로이 대두되고 있는 이론에 병합될 수 있는 것도 있을 수 있다. 예를 들어, 우리의 태양계 내에 있는 주 요 행성은 모두 태양을 중심으로 하여 갇은 방향으로 회전하고
있다. 동시에 천왕성과 금성을 제의한 모두가 그 행성 자체의 회 전과 동일한 방향으로 회전하고 있다. 하지만 현재로서는 이것이 모든 행성계에 해당하는 진리라고 믿을 수 있는 근거가 없다. 마 찬가지로 육상동물을 구성하고 있는 단백질들은 모두 L 형의 아미 노산 ® 으로 되어 있다(아주 드물게 D 형 아미노산이 어떤 박테리아 속에 있는 경우도 있기는 하지만). 한 생물체 내에 있는 단백질들 이 모두 같은 형태여야 하는 데는 화학적 이유가 있지만, 그것이 D 형이 아니라 L 형이라고 예측할 수 있게 하는 화학적, 생물학적 이론은 현재로서는 없다. 따라서 현존하는 과학 이론의 관점에서 보면 이러한 사실은 순전히 우연적인 일에 불과할 뿐이다.
® L 형 아미노산. 아미노기(基)와 카르복실기의 두 기를 한 분자 속 에 지니고 있는 물질을 총칭하여 아미노산이라고 하는데, 이 아미 노산은 서로 분자구조가 다른 204 용이 있다. 이 20 종의 아미노산이 어떤 배열순서를 이루면서 수십 개 또는 수백 개 길게 한 줄로 연 결된 것이 단백질이라는 물질이다. 서로 다른 20 종이 어떤 순서로 배열되느냐 하는 조합의 종류는 무한대라고 할 수 있고, 따라서 그 결과 만들어지는 단백질 분자의 종류 수도 무한대라고 할 수 있다. 이 지구상에 무한히 많은 종류의 단백질이 존재하고 있는 것은 이 것 때문이다. 그런데 이 20 종의 아미노산 가운데 한 가지만 제의하 고 1~ 적 각각에는 분자식은 갇으나 원자의 공간배치에 차이가 있는 두 종류가 있을 수 있다. 이 두 종류의 한쪽을 D 형이라 하고 다른 쪽을 L 형이라 하는데, 자연에서는 모든 단백질이 L 형 아미노 산만으로 구성되어 있다. 실험실에서 아미노산을 합성하면 같은 양 의 D 형과 L 형이 섞여 나오는데 자연에서 L 형만 있는 것은 L 형으 로 된 단백질이 D 형으로 된 것보다 열역학적으로 더 안정하기 때 문인 것으로 알려져 있다.
마지막으로 우리가 앞으로 사용하게 될 〈 뜻밖 〉 혹은 〈 우연 〉 과 같은 단어들에 대한 설명을 덧붙여야 할 것 같다. 〈 개개의〉라는 단어와 마찬가지로 이러한 단어들은 상대적인 의미로 사용될 것
이다. 과학 발달의 어느 시점에서나 한 과학 이론의 영역 밖에 존재하는 어떤 규칙 같은 것들이 항상 있는데, 이들 규칙들은 우 주가 지니고 있는 〈 우연적인 〉 특징들이라 할 수 있다. 따라서 하 나의 사건은 어떤 일련의 법칙들에 대해서는 우연일 수 있으나, 그보다 더 포괄적이고 강력한 법칙들에 대해서는 확정 가능한 것 이 될 수도 있다. 물론 천계결정론자(天啓決定論者, ap o calyp tic de t er mi n i s t)들은 모든 사건들은 원인이 있어 생긴 것이고 따라 서 우연한 사건이란 존재하지 않는다고 확신하겠지만 말이다. 그 러나 우연한 일치에 해당되는 사건들의 존재도 역사적인 추론을 위해서는 매우 중요한 것이다. 예를 들어, 육상동물을 구성하고 있는 단백질이 모두 L 형이라는 것은 모든 육상동물이 한 개의 공 통되는 근원점으로부터 전화되어 왔다는 훌륭한 증거가 될 수 있 다. 그러나 만일 L 형으로 되어 있다는 사실을 현존하는 과학 이 론으로부터 추적해 올라간다면 그러한 추론은 도출될 수가 없을 것이다. 그러한 일치를 우연이라고 함으로써만 그 사건들은 우리 가 과거를 재구성하는 데 도움을 줄 수가 있을 것이다. 인과법칙 이 장에서 논의될 제반 법칙들은 인과법칙에 해당된다. 이것은 원인, 결과 그리고 이와 유사한 용어에 대한 일반화를 의미하는 것이며, 흔히 사용되는 것처럼 결정론적인 법칙들을 지칭하는 것 은 아니다. 가장 완벽한 형태로서의 인과법칙과 과정의 법칙의 차 이점은 단지 시간에 관한 문제에서만 나타난다. 인과법칙은 원인 이 결코 그 결과를 추구하지 않고, 과정의 법칙은 시간적 비대칭
성을 전혀 구체화하고 있지 않거 때문이다• 이제 우리는 인과법칙 의 용어에 관한 논의를 하게 되는데 이는 그런 용어가 특별히 과 학의 기초가 된다고 믿고 있기 때문이 아니라, 완벽한 지식으로부 터 이탈된 사항들도 이들 어휘 속에서는 보다 쉽게 논의될 수 있 기 때문이다. 과정의 법칙은 폐쇄된 시스템을 다루는 반면, 인과 법칙은 인과관계로 이루어져 있는 개방된 그물에서 선택된 몇 개 의 용어들로 구성되어 있다• 인과법칙이 되기 위한 필요조건은 자 연 현상을 인과적으로 연결된 몇 개의 그룹으로 나누는 것이며, 이러한 그룹 중의 하나에서 생긴 예는 다론 경우를 추론하는 데 사용될 수 있다. 그러나 이러한 연계성이 지닌 특징이 너무나 다 양하고 그러한 연계성을 통해 유도되는 추론도 마찬가지로 너무 다양하다. 예를 들어, 원인 (C) 이 그 결과 (E) 를 위해 필요조건이 면서 동시에 충분조건일 경우, C 의 예를 통해 E 를 연역적으로 추 론할 수도 있고 그 반대로 E 에서 C 를 추론할 수도 있는 것이다. 원인과 결과라는 용어가 지닌 시간적인 의미를 무시한다면 인과 법칙은 과정의 법칙으로 대치될 수도 있지만, 앞에서도 얘기한 것 처럼 생물학에서 그러한 보편적인 연계성의 예를 찾아낸다는 것 은 결코 쉽지가 않다. 그러나 출생이 필연적으로 맞게 될 죽음을 위한 필수적이고 충분한 일이라는 우스꽝스러운 이야기를 하는 사람도 있을수 있다. 철학자들은 보통 인과적인 연계성을 보편적 상호 관계라는 말 로 정의하지만 원인과 결과러는 용어는 상호 관계가 보편적이지 않은 경우에도 사용될 수가 있다. 어떤 경우에는 그 원인이 결과 를 위한 필요조건이 아니라 충분조건에만 해당되는 경우도 있는 데, 이런 경우에는 C 의 경우로부터 E 를 추론할 수는 있지만 그 역은 불가능하다. 예를 들면, 어느 별의 폭발은 그 별에 존재하는
생물 전체를 파괴시키는 충분조건이 될 수 있을 것이다. 그러나 생태학자들이 우리에게 끊임없이 상기시키고 있는 바와 같이 그 것은 결코 필요조건은 아니다 . 역으로 원인이 결과를 위해 필요적 이기는 하나 충분조건은 아닌 경우도 있다. 이때에는 E 의 예에서 C 를 추론할 수는 있지만 그 역은 불가능하다. 대부분의 질병은 그러한 예에 해당된다. 예를 들어, 폐결핵균은 폐결핵에 걸리는 데 필요조건이지만 충분조건은 아닌 것이다. 그러나 보통의 경우 에는 원인이 그 결과에 대한 필요조건도 아니고 충분조건도 아니 다. 이러한 경우에는 C 로부터 E 를 연역적으로 추론할 수도 없고 E 에서 C 를 추론할 수도 없지만, 그렇다고해서 C 와 E71- 서로 무 관한 것은 아니다. 그러한 형식화 또한 인과적인 경우의 한 예인 데도 불구하고 이를 깨닫지 못했기 때문에 과학자들은 흡연이 폐 암의 원인이 아니라고 주장할 수 있었던 것이다. 많은 사람들이 흡연을 줄기면서도 폐암에 걸리지 않았고 폐암에 걸린 사람이라 고 다 흡연 경험이 있는 것은 아니다. 그러나 저명한 과학자들이 한결같이 지적하는 바와 같이 흡연과 폐암은 분명히 서로 관련이 있다. 만일 〈원인〉이 충분조건 아니면 필요충분조건이라는 용어 로 정의되어야만 한다면 우리는 결코 폐암의 원인을 발견할 수가 없을것이다. 위에서 열거한 일반화의 유형들을 〈법칙〉이라고 부르는 것에 대해 많은 반대 의견이 대두되어 왔다. 만일 보편적인 상호 관계 를 찾아내지 못했다면 그것은 자연 현상에 대한 분석이 적절하지 못했다는 것을 의미한다는 것이다. 이럴 때 우리는 전체적인 세트 가 충분한 것이 되도록 만들기 위해서 부수적인 요소들을 필요조 건에다 첨가시킬 수 있을 것이다. 예를 들어, 폐결핵균에다 약한 저항력 등을 덧붙이면 폐결핵에 걸리는 충분조건이 될 수 있을
것이다. 아니면 결과를 더 세분화된 상태로 분석할 수도 있다. 어 떤 질병에 대한 단 한 개의 원인만을 찾아낼 수 없는 경우, 그것 울 여러 개의 질병으로 다시 나눈 후 그 각각에 대한 한 개씩의 원인을 찾아낼 수 있는지 본다. 어떤 때는 이러한 방법들 중 어 느 한쪽이 성공할 수 있을 것이고 또 어떤 때는 성공적이지 못할 수도 있을것이다. 또 다른 가능성은 인과성을 더욱 정교하게 분석하는 것이다. 한 예로서 스크리본 Mi ch ael Scr i ven1) 은 보통 하나의 원인이라는 것은 어떤 결과를 일으키기 위한 충분조건의 세트 내에 있는 중 복되지 않은 구성원이라고 하였다. 똑같은 결과를 가져올 수 있는
1) Ex pla nati on s, Pred ict i on s, and Laws, Mi nn esota Stu d ie s in the Phil o sop h y of Scie n ce, 冊 ( Mi nn eap ol i s : Univ e rsit y of Mi n- nesota Press, 1962), p. 215.
또 다른 충분조건 세트들도 있을 수 있겠지만 어쨌든 이러한 충 분조건 세트가 주어지면 그 세트 내에 있는 특정한 조건 각각은 그 사건을 위한 필요조건이 된다. 필요조건에 해당하는 구성원이 하나 이상일 수도 있다. 그 중 어떤 것이 그 사건의 원인으로 선 랙이 되느냐는 것은 전후 문맥에 달려 있다. 그러나 한 사건에 대한 완벽한 충분조건 세트롤 만들 수 있다고 생각해서는 안 된 다. 실제로 그것은 불가능한 일이다. 예를 들어, 누군가가 케네디 대통령의 사망 원인이 무엇이냐고 묻는다면 거기에는 수많은 종 류의 대답이 가능할 것이며, 그것은 질문자의 의도가 무엇이냐에 따라 달라질 수 있을 것이다. 케네디의 죽음은 총상으로 인한 사 망, 국가 원수의 암살 기도 등 수많은 범주 속에 집어넣을 수가 있는것이다. 그렇게 전후 문맥에 따라 달라질 수 있고 또 부분적으로만 관
련이 있는 진술들을 〈 법칙 〉 이라고 부르는 데 반대하는 사람들은 그런 것들이 일상적인 이야기나 수준이 낮은 과학적 연구들이 지 닌 특징이라고 믿는다 . 자연 현상이 적절하게 연구되기만 한다면 보편적인 연계성은 저절로 생기게 된다는 것이다. 이러한 견해에 대해 두 가지 답변을 할 수 있을 것 같다. 첫째, 유전자형과 표현 형 사이의 관계는 상당히 철저하게 연구되었는데도 그러한 연계 성이 나타나지 않는다는 것이다. 왜냐하면 그러한 연계성이 아예 존재하지도 않기 때문이다. 특정 유전자와 표현형질 사이의 관계 는 마치 흡연과 폐암 사이의 관계와 같아서 어떤 특정 유전자가 특정 표현형질에 필요조건인 경우는 거의 없으며 충분한 조건이 되는 경우는 아예 있을 수도 없다. 따라서 이것은 유전이냐 환경 이냐의 논쟁과 같은 것이다. 이렇게 본다면, 어떤 유전자가 정신 분열증에 대한 필요조건도 혹은 충분조건도 아닐 경우, 그 유전자 를 정신분열중의 원인이라고 할 수 없게 될 것이다. 둘째, 더욱 철저히 연구를 하여 보편성과의 연계성을 결국 밝혀낸다고 하더 라도, 그렇게 되기 전까지는 부분적 상호 관련에 대한 지식이 여 전히 효용 가치가 있다는 것이다. 예를 들어, 흡연이 폐암과 관련 이 있다는 사실만으로도 큰 도움이 된다. 보편적 인 상호 관계가 발견된 뒤라 할지라도, 덜 보편적이지만 쉽게 얻을 수 있는 전체 적인 상호 관계도 여러 가지 목적상 여전히 유용 가치가 있는 것 이다. 많은 유전학자들이 아직도 형질전달유전학에 관심을 기울이 고 있는 것을 그 예로 둘 수 있을 것이다.
개체발생에 관련된 발생법칙 생물학 문헌에서 압도적으로 많이 쓰이고 있는 인과법칙은 바 로 발생에 관한 법칙이다. 인과론에서는 한 사건이 다음 사건의 원인이 된다는 것인데, 발생법칙에서는 연속적으로 일어나는 일련 의 사건들이 구체적으로 명시될 수가 있기 때문이다. 따라서 A 가 일어나고 그 다음에 B, 그 다음에 C, 그 다음에 D 가 일어날 때, A 는 B 를 일으키고 B 는 C 를, C 는 D 를 유발한다고 할 수 있다. 이러한 사건들 하나하나가 다른 사건의 필요충분조건이 될 때 발 생법칙은 과정의 법칙으로 대체가 될 수 있고, 과정의 법칙에 있 어서 A, B, C, D 는 연이어 일어나는 시스템의 상태라고 할 수 있다. 원인과 결과가 보편적 상호 관계를 지닌 두 구성원으로 이 루어져 있는 인과적 형태는 생물학에서 찾아보기가 힘들기 때문 에, 발생법칙의 구성원들이 서로 보편적으로 관련되어 있으리라고 기대하지 않는 것은 당연한 일이다. 그러나 생물학에 있어서 발생 이 차례차례로 일어나는 예는 발생학, 생리학 그리고 고생물학에 서 자주 찾아볼 수 있다. 발생학은 생물체가 수정란에서 성체로 발달하기까지의 초기 발생단계를 다루며, 생리학은 개체가 계속 생명체를 유지시켜 나가는 과정을, 고생물학은 그 종의 계통발생 울 다루는데, 이들 각 분야에 있어 발생 순서는 이미 구체적으로 정해져 있는 것이다. 문제는 이러한 일반화 중의 어느 하나라도 과학법칙으로 간주될 수 있겠느냐는 것이다. 하다 못 해 통계적인 방법을 통해서라도 말이다. 생물학 문헌에서 논의된 많은 순서들은 단순히 서술뿐인 피상 적 모습을 보이고 있다. 예를 들면, 초파리에서 붉은 반점을 나타 내는 크산토마틴 xan t homma ti n 을 생성하는 데 있어서, 트립토판
은 키누레닌으로, 키누레닌은 3- 히드로크산토마틴으로, 이것은 다 시 크산토마틴으로 전환된다. 한 반응 속에 포함된 이러한 단계들 각각에 대한 설명을 과학법칙으로 간주하는 데는 아무 이의가 없 을 것이다. 그러나 이러한 구체적인 반응들 각각이 어느 한 특정 한 초파리 종의 세포 속에서 일어난다고 말하는 것은 옳지 않다. 적절한 pH , 온도, 촉매 등이 주어지면 언제나 트립토판은 키누레 닌으로 전환이 될 것이며 더욱이 그러한 화학법칙은 원칙적으로 양자역학에서 유도될 수가 있기 때문이다. 그러나 극히 단순한 복 합물과 반응, 그 중에서도 가장 단순한 특징들조차도 실은 어마어 마하게 복잡한 것이라는 사실을 간과해서는 안 된다. 하나의 물 분자가 지닌 특성 중의 많은 부분이 양자역학적 계산에 의해 설 명될 수 있겠지만 , 물 분자들로 이루어진 커다란 총체적 집단이 지닌 전체적인 특성을 양자역학으로부터 유도해 내는 것은 아직 도 요원한 상태라고 물리학자들은 말하고 있다. 생합성 경로의 각 단계에 대한 전술은 과학법칙으로 간주될 수 있으나, 그러한 각각의 반응으로 이루어전 발생 과정 전체를 볼 때에는 어떠한가? 문제는 반복되어 일어나는 경우가 부족하다는 데 있는 것이 아니다. 실제로 위에서 얘기한 발생 과정은 되풀이 하여 일어난다. 경험적 일반화와 마찬가지로 그것 역시 기초가 튼 튼하다. 또한 이 생합성 경로가 특정한 종에만 국한되어 있는 데 서 문제가 생기는 것도 아니다. 여러 가지 생합성 경로에 대한 예들이 식물, 동물계 전반에 걸쳐 산재해 있고, 비육상동물계에서 도 쉽게 볼 수가 있다. 그러한 발생 과정에 대한 전술을 법칙으 로 간주하기 위해서는 그 법칙들이 유도될 수 있는 시스템을 제 대로 인식해야 한다. 그러기 위해서 우리는 생물체의 유전자형과 이 유전자형이 그러한 특정 경로를 조절하기 위해 어떠한 작용을
하는가 하는 문제로까지 우리의 관심 분야를 확대시켜야 한다. 유 전적 프로그램을 포함하여 그 생물체의 상태에 대한 지식과 주변 환경의 몇 가지 일반적 특성들에 대한 지식이 주어진다면 생합성 경로의 과정을 예측하는 것은 원리상 가능한 일일 것이다. 하지만 이 경우, 실제 그 유도 과정은 훨씬 더 복잡할 것이다• 마찬가지 로 이러한 반응이 어떤 특정 초파리 종의 세포에서 일어난다고 말하는 것은 옳지 않으며, 생화학 반응은 적절한 유전적 구성과 환경을 가전 세포들에서라면 항상 프로그램된 그대로 진행될 것 이다. 그러나 이러한 유전적 프로그램은 설명 과정에서 사용되는 법칙 속에 포함되어 있는 것이 아니라, 단지 한계조건(限界條件) 속에 포함되어 있을 뿐이다. 비슷한 이야기를 고전적 발생생물학에서도 포배, 낭배, 신경판 이 될 부분, 중배영 등의 용어를 가지고 반복해 볼 수가 있다. 과 거에는 그러한 일반회들이 단순히 서술적인 것에 불과한 것처럼 보았다. 왜냐하면 그들 대부분이 관찰에 근거하여 만들어전 것일 뿐, 어떤 잘 짜여전 법칙으로부터 유도되는 것은 아니었기 때문이 다. 게다가 발생학 전체로 보았을 때는, 발생 과정들이 그 분포에 있어 생화학적 반응에 비해 더 한정된 분류군으로 국한되는 경향 이 있다. 그러나 접합체의 유전적 구성과 그 생물체의 발생을 지 배하는 환경이 모두 알려진다면, 원리상 발생 과정들 모두가 구체 적으로 명시될 수 있을 것이다. 발생학에서의 기작에 관한 연구에 많은 발전이 이루어졌지만 그 전보는 분명히 더디고 고통스러운 것임에 틀림없는 것 같다.
계통(系統)과 교차연계법칙(交又連緊法則) 계통발생麟] 있어서의 발생법칙은 개체발생의 그것과는 매우 다르다. 왜냐하면 계통발생학에는 표현형을 지배하는 유전자형에 비교할 만한 것이 없기 때문이다. 또 〈전화법칙〉은 생물 철학에 있어 가장 혼동되는 문제 중의 하나이다. 계통발생과 전화론과의 혼동은 이미 논의된 바 있거니와 무악류의 어떤 종이 판피어류로 부터 기원한다는 이야기나 판피어류 중의 어떤 종은 경골어류로 발달되었다는 등의 이야기는 전화론의 일부라고 할 수가 없다. 그 것은 계통발생 과정에서 일어난 일련의 특정한 사건들을 묘사한 것일 뿐이다. 이러한 특정 사건들에 대한 추론은 전화론을 포함한 여러 과학 이론의 도움을 받아 가능하겠지만 이러한 사건을 기술 하는 것은 그저 기술일 뿐이지 전화법칙은 아닌 것이다.
® 계통발생. 헤켈 (E. H. Haeckel, 1866) 에 의해 제창된 개념으로서, 한 종의 생물이 그 선조 때부터 현생 종에 이르기까지 거쳐온 형 태적 변화를 말한다. 말하자면 한 종의 진화 역사라고 할 수 있다. 이에 대하여 개체발생은 하나의 개체가 수정란에서부터 성체에 이르기까지 거치는 형태적 변화를 말한다. 헤켈은, 개체발생은 계 통발생을 압축하여 되풀이한다는 설을 제창하였고 한때 이 설은 대단히 설득력 있게 받아들여지기도 했으나 과학적 근거가 희박하 다는 비판이 대단히 많아서 현재는 하나의 가능성 정도로만 취급 되고있다.
이 장에서 우리가 관심을 두게 될 문제는 과학법칙 내의 용어 들이 공간적으로 그리고 시간적으로 제한이 되어서는 안 된다는 요구와 〈종〉에 대한 전화론적 정의 사이에서 생기는 갈등이다. 아리스토텔레스나 데오프라스투스로부터 심프슨 G.G. S i m p son 이 나 하이만 L i bb i e H ym an 에 이르기까지 생물학자들은 생물을 표
현형질(대개 형태적인)의 다양성을 근거로 해서 여러 분류군으로 나누어 왔다. 예를 들어, 〈 척색동물 〉 은 내부에 새열(아가미 톰)이 있고, 등에 신경색(神經索)이 있으며, 발생 과정중 특정 시기에 척색을 지닌다는 점 등의 형질로 정의(定義)된다. 고전적 정의의 개념에 의하면 정의 속에 나열된 일련의 형질들은 특정한 종을 같은 속에 속하는 다른 종들과 구분지울 수 있어야 하며, 그 특 정 종에 속하는 모든 구성원들은 이러한 일련의 형질을 지니고 있어야 한다. 이것이 바로 어떤 용어를 정의하기 위하여 사용되는 특성들은 그 구성원이 되는 필요조건이며, 그 각각이 모이면 구성 원이 되기 위한 충분조건이 된다는 말이 의미하는 바이다. 다론 종들도 이 각각의 형질 중 어느 하나씩을 지니고 있을 수는 있으 나, 이러한 특정한 형질 전체로 된 세트는 그 종에 속하는 모든 구성원들 사이에서만 보편적으로 발견될 수 있어야 한다. 정의에 관한 전통적인 개념에 따르면, 일단 정의에 필요한 톡 정 형질들의 세트가 선택이 되면 거기에 부가되는 상호 관계에 대한 전술은 모두 과학법칙으로 간주되는데, 이러한 것을 교차연 계법칙이라고 부르고 있다. 과정의 법칙이 순간적인 변화나 시간 적인 차이룰 두고 계속되는 변천을 다루는 반면, 교차연계법칙은 현재의 상호 관계에 중점을 둔다고 할 수 있다. 한때는 〈금 〉 을 노란색이라는 성질이 포함된 일련의 성질들로 정의했었는데, 이런 경우 〈금은 노랗다〉라는 것은 분석에 근거한 주장이다. 또 금이 라는 것은 전성 (展性), 죽 펴서 늘이거나 모양을 바꿀 수 있는 성 질을 가지고 있는데, 이 전성은 노란색이나 마찬가지로 금의 성질 을 나타내는 것이지만 〈금은 전성을 가지고 있다〉라는 문장은 분 석적인 것이 아니라 종합적인 전술이며 하나의 교차연계법칙에 해당되는 것이다. 물론 오늘날에는 〈금〉이 원자번호에 의하여 정
의되며, 원자론에 의하면 〈 금은 원자번호가 79 이다〉라고 정의할 수 있다. 그리고 그 원자번호에 해당되는 원소가 지니고 있는 일 련의 물리적 특성들에 대한 전술은 모두 교차연계법칙으로 간주 된다. 이제 문제는 이와 비슷한 관찰이 〈모든 백조는 희다〉와 같 은 전술에서도 만들어질 수 있겠는가 하는 것이다. 앞에서 우리는 계속 진화하고 있는 계보로서의 종은 전통적 방 식으로 그 한계가 정해질 수 없다는 것에 대해 논의했다. 특정한 종의 이름은, 그 구성원이 되기에 필수적이고 또 충분한 형질들의 세트로서 정의될 수 없다. 그렇게 되면 그러한 범주에서 제의되는 개체가 너무 많기 때문이다. 대신 그 종이 지닌 특성으로 명명되 어야 한다. 그러나 그러한 특성은 그 구성원들 사이에 보편적으로 분포되어 있지가 않다. 다시 말하면 〈정의〉를 하는 데 필요한 일 련의 특칭들이 주어지지만, 한 개체가 그 형질들을 모두다 지니고 있을 필요는 없고 단지 그 중 가장 중요한 것들을 충분할 만큼 지니고 있으면 되는 것이다. 어떤 형질이 다른 형질보다 특정 분 류군의 특성을 더 잘 나타내고 있는 것은 사실이지만, 그렇다고 해서 아주 필수적이라고 할 수 있는 형질이 따로 있는 것은 아니 다. 이러한 관점에서 보면 정의를 하는 데 사용되는 특성과 경험 에 의해 연관시킬 수 있는 부가적 특성들 사이의 명확한 구분은 사라지게 된다. 그러나 우리가 살펴본 이러한 어려움들을 여기서 는 일단 무시할 수가 있다. 왜냐하면 문제는 특정 종의 이름이 어떻게 정의되는가가 아니라 〈종〉이라는 단어 그 자체를 어떻게 적합하게 정의하는가 하는 것이기 때문이다. 이는 〈물리적 원소〉 와 〈금〉을 정의하는 문제와 마찬가지이다. 원자론에 따르면 원소 는 원자번호, 죽 그 원소가 지니고 있는 양성자의 수로 구별되므 로 금은 79 번이다. 〈원소〉 그 자체는 79 개의 양성자를 갖는다고
정의되어 있지 않지만, 〈 금 〉 이라는 원소는 79 개의 양성자를- 갖는 것이라고 정의될 수 있는 것이다. 심프슨에 따르면러 진화적 종은 〈 다론 것으로부터 분리되어 나와 그 자신만의 고유한 진화적 역할과 경향을 가지고 전화하는 하나의 계통(한 집단에 있어서 선조와 후손을 잇는 계보)이다 〉 . 전 화론적으로 〈 종 〉 이라는 것은 유성생식이나 무성생식을 하는 모든 종을 말하는데, 이러한 정의는 자연 내에서 종 사이에 어떤 경계 가 존재한다는 것을 암시하고 있다. 종이란 것은 바다의 해류처럼 뒤섞여 전화하는 것이 아니라 삼각주에서의 강의 지류처럼 분리 되어 진화한다는 것이다. 이러한 견지에서 볼 때 종에 대한 전화 적 정의는 〈 조작적 o p era ti onal 〉 인 것은 아니다. 이러한 정의를 쉽게 적용할 수 있기 위해서는 어떤 기준이 필요한데, 〈 종 〉 에 대 한 생물학적 정의는 유성생식을 하는 생물에 대해 그러한 기준을 하나 제공해 준다. 메이어에 따르면라 생물학적 종은 〈 실제로 혹 은 잠재적으로 상호 교배가 가능한 자연집단 〉 이며 그러한 집단은 다른 집단들과 생식적으로 격리되어 있다고 한다. 따라서 교배라 는 것은 유전적 균일성을 높이는 기작이며 진화적 집결성 cohesi- veness 을 중전시 킨다.
2) Prin c ip le s of Anim al 'lhxo nomy ( New York : Colombia Univ e r-sit y Press, 1961), p. 153. 3) Sys t e m ati cs and the Orig in of Sp ec ie s (New York : Columbia Un ive rsit y Press, 1942), p. 121 .
진화론자들이 오랫동안 주장해 온 바와 같이 전화하는 계보로 서의 종은 시공간적인 총체이며 따라서 조상과 후손이라는 관계 에 의해 시간적으로 연결되어 있고 생식을 통해 유전자를 교환함 으로써 공간적으로.(유성생식의 경우) 서로 연결되어 있다. 대부분
의 분석에 따르면, 어떤 복합체가 개체인지 아닌지를 판단하는 가 장 중요한 기준이 시공간적인 연속성이기 때문에 전화론의 관점 에서 볼 때 종이라는 것은 개체들이 모인 것이다 . 이렇게 보면 시조새 (始祖 鳥 ) ®는 쥐라기 후반에 절멸됐으므로 만일 한 무리의 개체들이 고대의 시조새와 똑같이 전화해 왔다 하더라도 그들은· 시조새가 아닌 새롭고 독자적인 종으로 간주되어야 할 것이다. 그 러나 스마트(J. J. C. Smart, 1963, 1968) 가 지적한 바와 같이 만 일 종이 시공간적인 개체이고 그 이름이 적절한 것이라면, 그것은 과학법칙이 시공간적인 제한을 받아서는 안 된디는 조건을 어기 지 않고서는 과학법칙 내에서 아무런 기능을 할 수가 없다. 따라 서 〈 모든 백조는 회다 〉 와 같은 말은 그것이 전실일지라도 과학법 칙이 될 수가 없게 된다. 특정한 종의 이름에 대한 정의와 〈종〉에 대한 정의를 혼동함으 로써 스마트의 논의에는 불필요하게 복잡한 면도 있긴 하나 그의 주장은 좀더 설득력 있게 수정이 될 수 있을 것이다. 스마트와 루스 M ich ael Ruse4> 두 사람은 모두, 우리가 〈종〉에 대한 전화론
4) Are There Laws in Bi ol og y? Austr al asia n Jo urnal of Phil o - sop h y, 48(1970), pp. 234-246. 이 논문에서 루스는 생물학에는 법 칙성이 없다고 하는 스마트의 견해를 반박하고 있다. 그러나 그 는 〈종〉의 정의를 예컨대
덮여 있는 동 현생 조류의 특칭을 대부분 갖추고 있으나, 턱에 이 가 있고 용골돌기가 거의 없는 등 파충류의 특칭도 많이 지니고 있어서, 전화의 과정에서 원시 파충류와 조류가 갈라지는 초기 단 계의 중간 생물로 생각되고 있다.
적 정의를 포기한다면, 그리고 특정 종의 이름을 그 개체가 거쳐 온 진화적 경로와는 무관하게 표현형질만을 가지고 정의한다면 그 이름은 과학법칙 속에서 어떤 기능을 하는 데 필요한 일반성 을 지닐 수 있다고 생각한다. 남은 문제는 이러한 일반화가 과학 법칙인가 아니면 이론이 결핍된 경험적 일반화에 불과한가 하 는 것이다. 루스는 이것을 과학법칙이라고 간주했고 스마트는 그 렇게 생각하지 않았다. 스마트 (1963 : 235) 도 우리가 관찰 대상으로부터 일련의 특성 들, 예를 든다면 〈육상동물에 한한다면, 쥐만이 그러한 특성을 가 지고 있다〉라고 묘사될 수 있는 어떤 특성들을 찾아낼 수 있다는 것은 인정한다. 다만, 〈문제는 우리가 만든 법칙이 전리라고 주장 할 만한 근거가 없다는 데 있다〉는 것이다. 만일 종에 대한 전화 적 정의와 그 생물학적인 관련성을 함께 수용한다면, 〈 종 〉 은 종 합전화론, 죽 멘델 유전학과 고전 전화론이 병합되어 이루어전 하나의 이론적 용어가 된다. 이러한 이론에 따라 전화적 종을 시 간에 따라 나누어보면, 표현형질에 관한 한 나누어전 조각 하나하 나는 여러 개체들로 이루어전 하나의 묶음을 형성하고 있다. 그렇 게 해서 특정한 종의 특성이 일련의 형질을 가지고 통계적으로 설명되면, 이미 조사된 형질을 통해 조사되지 않은 형질에 대해서 도 어느 정도까지 예측을 할 수가 있는데 이는 그러한 형질들이 함께 변화해 나가리라고 예상되기 때문이다. ' 그러나 〈종〉에 대한 전화적 정의와 그 생물학적 관련성을 포기한다면, 우주는 고사하
고 이 지구상에서조차도 그러한 변화를 기대할 수 없게 된다. 혹자는 이렇게 반박할 수도 있을 것이다. 〈 그러나 온도가 급강 하된 환경 속에서 온혈동물이 살아가게 된다면, 그 동물은 털을 갖게 되지 않겠는가? 〉 그러나 반드시 그럴까? 깃털이든 지방총이 든 그의의 다른 것이든 무엇이 필요한지 하늘만이 알 것이다. 한 때 큐비에 Cuvie r 같은 이상적 형태학자들은· 특정한 형질들은 서 로 반드시 연관되어 있어야 하고 그렇지 않으면 조화가 이루어지 지 않는다고 생각했었다 . 예를 들어, 어떤 생물이 깃털을 가지고 있다면 그 생물은 이 (齒)를 가질 수가 없다는 것이다. 그러나 여 러 가지 표현형질이 불가능하다고 생각된 상태로 혼합되어 있는 시조새, 오리너구리 등의 생물들이 발견됨으로써 생물학자들은 표 현형질의 수준에서는 환경에 적응해서 살아가는 수단에 여러 가 지 방법들이 있다는 것을 인정하지 않을 수 없었다. 모든 삼각형 은 세 개의 각을 가져야 하지만 모든 파충류가 세 개의 심방 심 실로 된 심장을 가질 필요는 없는 것이다. 실제로는 그렇다 할지 라도 말이다. 담수에 사는 척추동물이 바닷물로 가게 되었다면 그 둘은 질소 노폐물을 배설하는 방법 의에 체내의 수분량을 일정하 게 유지하는 방법도 찾아야 할 것이다 . 그들이 어떤 방법을 택하 느냐 하는 것은 대개 그들이 처한 전화적 상황의 특성에 달려 있 다. 만일 종에 대한 전화적 정의를 버린다면 특정한 종에 대해 설명할 때에는 전화론 이의의 다른 곳에서 이론적 뒷받침을 받아 야할것이다. 이 장의 앞부분에서, 순수한 과학법칙은 실제적으로 그리고 궁 극적으로 이론 속에 병합될 수 있음으로써 경험적 일반화와 구분 이 된다고 했었다. 금이 변형될 수 있다는 것은 사실이나 이러한 경험적 일반화는 태양계의 모든 행성들이 태양을 중십으로 하여
같은 방향으로 회전하고 있다는 사실이나 마찬가지로 우주의 한 우연적 특성에 불과한 것으로 생각될 수도 있다. 그러나 실제로는 그렇지 않다. 이제 우리는 물리적 원소의 특성을 그 원소의 원자 구조와 양자론으로부터 유도할 수 있게 된 것이다. 〈 금은 변형될 수 있다〉와 같은 것은 양자론으로부터 너무 쉽게 유도되는 결과 이긴 하나, 그래도 결과는 결과이다. 이와 유사한 교차연계법칙을 생물학에도 적용시킬 수 있을까? 만일 진화론이 그러한 상호 관 계에 대한 추론을 허용하지 않는다면 과연 다른 무엇이 그럴 수 있을까? 이제 남아 있는 유일한 답은 생물의 유전 구성에 대한 지식까 지도 포함하는 넓은 의미에서의 발생학이다. E1 에서 E 계 이르는 환경 속에서 유전적 구성 G 를 가전 생물체는 R 에서 R 에 이르 는 생화학적 반응을 통해 CI, C 료-부터 Cn 에 이르는 표현형질을 지니게 될 것이다. 따라서 한 생물집단의 유전적 구성과 이 집단 의 구성원들이 처하게 될 환경의 범위와 생화학적 반웅들을- 안다 면, 그 결과로 어떤 형질들이 나타나게 될는지를 추론할 수 있을 것이다. 그러나 그러한 주장이 정당화되려면, 반드시 알고 있어야 할 사실들에 대한 세심한 주의가 필요하며, 이러한 추론의 극히 가설적인 성질에 대한 주의도 필요할 것이다. 이것은 현재로서는 양자론의 경우보다 훨씬 더 어려운 일이다. 본질주의자들은 전화하는 계보로서의 종의 이름이 과학법칙에 서 아무런 기능을 할 수 없다고 주장하였다. 왜냐하면 그러한 이 름은 계속 변화되는 특성을 통계적인 용어로 정의할 수밖에 없기 때문이라는 것이다. 한편 스마트와 루스는 그러한 종이란 결국 개 체들인 데 비하여 과학법칙은 어떤 집단에 대한 것이기 · 때문에, 진화하는 계보로서의 종의 이름은 과학법칙 내에서 아무런 기능
울 할 수 없다고 주장했다. 스마트는 표현형질들의 상호 변화에 관한 전술에 〈 종 〉 에 대한 전화적 정의가 결여되어 있다면, 그것 울 진리라고 가정할 아무런 근거가 없기 때문에 생물학적 법칙이 아니라고 주장한다. 만일 그것이 진리라고 하더라도 그것은 우연 적인 상호 관계에 대한 경험적 일반화에 지나지 않는다는 것이다. 이 장에서 필자는 극히 약한, 루스의 주장보다도 더 약한 명제를 놓고 논의를 해 왔다. 표현형질의 상호 관계에 대한 진술들은 낮 은 수준의 생물학적 법칙으로 간주될 수도 있을 것이다. 그러나 그것은 생물의 유전적 구성과 그것이 만들어내는 생화학적 반응 울 포함시키는 경우에만 가능하다. 사실상, 생물학에는 그러한 배 경을 갖고 있는 교차연계법칙이 거의 없고, 그들 대부분이 〈모든 백조는 희다 〉 라는 진술의 변형과 같다. 계통학에 있어서의 발생법칙 종을 전화하는 계보로 보든 개방된 존재로 보든 간에 특정한 종의 계통발생에 대해서는 경험적 일반화가 불가능하다. 종을 전 화하는 계보로 간주한다면 그러한 일반화에 필요한 반복성이 필 연적으로 사라져 버린다. 동일한 전화적 종은 오직 한 번만 전화 하기 때문이다. 종을 단계통 mono p h y le ti~로- 취급하지 않는다 해도 진화적 발전에 대한 경험적 일반화를 공식화할 수 있을 만 큼 충분한 빈도로 〈같은〉 종이 진화한다는 것은 거의 불가능한 일이다. 따라서 고생물학에도 발생법칙이 존재한다면 그 일반화는 특정한 종이라는 단계보다는 더 높은 단계에서 만들어져야 할 것 이다. 죽 집단, 세포군(細胞群), 다윈의 서브클랜, 적응도 집합 등
과 같이 집단에 대한 용어로 되어야 한다는 것이다. 사실상 고생 물학에 있어서의 발생법칙이란 주머니쥐, 개, 까마귀 같은 특정 종에 대한 것이 아니고 원시종, 다형적 종, 그리고 보편종(普 退 種) 같은 것을 다루는 것이다. 따라서 전화 법칙은 〈 모든 백조는 X 이다 〉 와 갇은 형태가 될 수 없고, 대신 〈 Y 형에 속하는 모든 종 은 X 다 〉 라는 식이 될 것이다. 우리는 고생물학 문헌에서 진화적 발전 양식에 관한 수많은 일 반화를 찾아볼 수 있다. 렌쉬 Rensch 는 무려 백 가지나 되는 것을 열거하고 있으며 5)' 코프 E.D. Co p~근 그러한 전화 법칙들 중에서 두 가지 법칙을 높이 평가하고 있는데, 첫째는 분화되지 않은 종
5) Bio ph ilo sop h y( N ew York : Columbia Un ive rsit y Press, 1971).
은 분화된 종보다 더 오랫동안 멸종되지 않은 채 존속된디는- 것 이고, 둘째는 한 그룹이 전화하는 동안 그 그룹 내의 생물체의 크기가 계속 중가한디는 것이다. 또한 닥케 Dacq u eS ] 법칙에 따 르면, 같은 지역에 살고 있는 현존 종은 모두 비슷한 방식으로 변화되는 경향이 있다는 것이다. 윌리스톤 W illi s t on 의 법칙은 다 소 복잡한데, 어떤 생물체에서 본질적으로 같은 기능을 하는 조직 이나 기관이 여럿인 경우, 전화가 계속됨에 따라 이들 구조들의 종류는 줄어들면서 남은 부분들이 더욱 특수하게 분화되어 간다 는 것이다. 마지막으로 루이스 돌로 Lo ui s Doll~ 즌 진화의 비가역 성이라는 발표로 높히 평가받고 있는데, 이는 전화하고 있는 그룹 들이 그들이 전화 과정에서 거쳐온 여러 단계를 다시 그대로 거 슬러 울라가더라도 원래의 상태로 되돌아갈 수는 없다는 것이다. 물을 떠나서 육상에서 살게 된 생물이 수중생활로 다시 돌아갈 수 있을지도 모르나 이 경우 처음 수중에서 살았을 때와 다시 되 돌아갔을 때의 형태에는 아주 큰 차이가 있으리라는 것이다.
고생물학자들은 계통발생적 순서를 재구성하기 위해 주먹구구 식 경험적 방법들을 사용한다. 이들 방법들은 마치 미스테리 살인 사건의 단서와도 갇이 어떤 것도 특별히 결정적인 것일 수가 없 다. 왜냐하면 돌로의 법칙을 제의하고는 모든 경우에 수많은 예의 가 존재하기 때문이다. 그러나 만일 그 중 몇 개의 법칙이 화석 에 정확하게 반영되어 나타난다면, 그러한 일치는 재구성에 대한 어떤 신뢰감을 줄 수 있을 것이다. 이제껏 이러한 〈전화법칙〉둘 을 열거한 것은 그것이 극히 정확하기 때문이거나 그것이 진짜 전화법칙이냐 아니냐를 논의하기 위해서가 아니라 그러한 법칙들 이 어떤 특정한 종을 지칭하는 것이 아님을 보이기 위해서였다. 그 법칙들은 단지 종의 형태를 이야기하고 있을 뿐이다. 따라서 전화하는 계보로서의 종에 대한 스마트의 반대 견해는 옳지 않다 고 생각된다. 그러나 이러한 법칙들이 옳다고 믿을 만한 어떤 근 거가 있을까? 헤켈 Ernst Haeckel 과 쉰트볼프 0. H. Sch ind ewolf 는 종도 한 생물 개체와 마찬가지로 일정한 생활사life c y cl 혀를 거친다고 했다. 우리는 이러한 법칙이 그릇된 것임을 분명히 알고 있다. 왜냐하면 그러한 주장을 화석 기록에서 전혀 찾아볼 수 없 기 때문이다. 동시에 우리는 그러한 생활사를 만들어내는 기작도 전혀 모른다. 그러나 우리가 어떻게 위에서 언급한 법칙들이 전정 한 과학법칙인지 아니면 우연한 일치일 뿐인지를 구별할 수 있겠 는가? 그것들은 모두 경험적인 일반화에서 출발했지만 이제 그들 의 이론적 정당화에 관심을 기울여야 할 때가 왔다. 위에서 언급한 법칙들 중에서 돌로의 법칙이 가장 진리에 가깝 다고 볼 수 있고 따라서 가장 이론적인 지지를 많이 받아왔다. 유전적 변화의 특성을 알고 또 이러한 변화가 게놈 상에 들어 있 다는 것을 안다면, 이러한 변화가 게놈 상에서 모두 지워지고 게
놈이 원래의 상태로 정확하게 되돌아가게 되는 경우도 가능한 일 이라고 생각될지도 모른다. 그러나 어떤 육상동물이 바다로 다시 되돌아갔다 하더라도, 그것은 원래의 수중 조상에는 없었던 바로 자신의 선조 즉 육상동물의 흔적을 지니게 될 것이다. 한편 닥케 의 법칙은 별로 논리적이지가 못하다. 환경이 변화할 때마다 그 곳에 사는 생물 모두가 그 변화에 적응하게 되리라는 것은 분명 하지만 그들 모두가 비슷한 방법으로 적응한다고는 할 수 없다. 어떤 지역이 건조해지면 거기에 사는 모든 식물은 습기를 보존하 는 방법을 찾게 될 것이다. 그러나 그 방법은 식물마다 서로 다 를것이다. 그러나 대체적으로 볼 때, 이러한 참정적인 법칙들에 대한 정 당화는 그리 훌륭한 편은 못 된다는 것을 인정해야만 한다 . 이 법칙들은 대략 앞에서 논의한 교차연계법칙과 동일한 상황에 놓 여 있으며, 그렇게 된 이유도 동일하다. 전체적인 표현형질 사이 의 상호 관계에 대한 교차연계법칙을 정당화시키는 일은, 생물체 의 유전적 구성과 환경 변화에 따른 생물의 반응에 대해서 우리 가 얼마나 알고 있느냐에 달려 있는데, 이 경우 우리는 환경에 대해서 거의 아무 것도 모르고 있다는 것이다. 마찬가지로 위에서 열거한 일반화는 진화적 발전에 대한 것을 좀더 커다란 범주에서 추론해 보기 위해 만들어진 것이다. 그 모두가 다 별로 정확하지 못하다는 것이 밝혀진다고 해도 놀랄 사람은 아무도 없을 것이며, 또 어떤 진짜 전화법칙이 만들어전다고 해도 그것은 현대의 집단 생물학자들이 제시하는 형태의 것이 되기 쉬울 것이다. 이들 잠정 적인 법칙들은 다음과 같은 변수들간의 관계를 다룬다. 예를 들면 볼록형 적응도집합 convex fitne ss se ts 과 오목형 적응도집합 con cave fitne ss se t s( 환경의 범위가 그룹의 내성(兩性)보다 작은가 큰
가에 따라 구분됨)의 관계, 소환경(小環境)과 대환경 사이의 관계 그리고 다양한 표현 양상 사이의 관계 등이다 (Lev i ns, 1968 : MacArth ur, 1972 참조). 현재 이러한 전화이론들은 모두 그 영역 에 한계가 있고 각각이 고립된 채로 그리고 부분적으로 서로 일 치가 되지 않는 몇 개의 덩어리로 존재하고 있을 뿐이다. 이들 모두 하나의 시스템으로 종합이 되어야 할 것이다. 역사법칙 생물학에는 역사법칙 hist o r ic a l laws 이 많기 때문에 생물학이 특별하다는 이야기를 종종 접하게 된다. 사실상 이미 논의한 여러 형태의 일반화 가운데 많은 것이 무분별하게 〈 역사법칙〉으로 불 려왔다. 언뜻 보기에도 무엇이 하나의 법칙을 역사적으로 만드는 가를 결정하는 일은 쉽지 않다. 단지 과거에 대한 추론이 가능하 다는 이유만으로 〈 역사적인〉 법칙이 될 수는 없다. 과정의 법칙 도 그러한 추론이 가능하기 때문이다. 현재나 미래를 추론하는 데 과거에 대한 지식이 사용되었다고 해서 그 법칙이 〈역사적인〉 것 이 될 수도 없다. 과정의 법칙도 마찬가지이기 때문이다. 그러나 역사법칙으로 불려도 괜찮을 생물학적 논술에서 두 가지 형태의 일반화를 찾아볼 수 있는데, 그 중 첫번째는 단지 과거에 대한 추론만을 허락하는 법칙이다. 예를 들어, 전화론은 예측은 불가능 하지만 과거의 사건에 대해서는 설명을 할 수 있는데, 이러한 추 론은 과학적 설명과 과학적 예측이 대칭이라는 명제와 함께 조금 후에 다시 논의될 것이다. 베르그만 Gus t av Ber gm ann6) 은 심리학
6) Phil os op hy of Scie n ce(Madi so n : Un ive rsit y of Wi sc onsin Press, 1957).
에서, 〈 역사법칙 〉 이라고 불리는 두번째 형태의 일반화를 신중하 게 분석했다. 그러나 그가 설명한 것 중의 많은 부분이 생물학에 도 똑같이 적용될 수 있는 것이었다. 베르그만에 따르면 역사법칙의 일반성은 과거에 대한 지식이 미래를 예측하는 데 필수적이라는 데에 있다. 현재에 대한 지식만 가지고 미래를 예측할 수는 없디는 것이다. 예를 들어, 앉아 있는 고양이를 쳐다본다고 해서 그 고양이가 앞에 놓인 먹이를 먹을 것인지 아닌지를 정확하게 예측할 수는 없다. 배가 고프지 않을지 도 모르기 때문이다. 그러나 고양이가 아주 건강한 상태이며, 여 러 날 동안 아무 것도 먹지 못했다는 것을 안다면 우리는 그 고 양이가 앞에 놓인 먹이를 먹을 것이라는 확실한 예측을 할 수 있 울 것이다. 이 경우에, 〈 배고픈 〉 이라는 것은 역사적인 개념이며 생물의 과거 역사에 대한 어떤 것을 담고 있는 것이고, 따라서 〈배가 고픈 고양이는 먹이를 보면 먹는다 〉 라는 진술은 역사법칙 이다. 베크너 (Beckner, 1959) 의 정의에 따르면, 역사법칙은 〈 배고 픈 〉 이라는 말과 같은 역사적 개념을 지니고 있는 법칙을 말한다. 그러나 〈배고픈〉이라는 말은 현재 그 생물의 생리적 그리고 심리 적 상태를 말하는 것으로 해석할 수도 있으며 그러한 해석에 따 른다면 개념과 그에 따른 법칙은 결코 역사적인 것일 수가 없게 된다(행동심리학자, 생리심리학자 그리고 정신심리학자들 사이의 논쟁을 찰 아는 사람은 지금 이 논쟁의 일반적인 과정에도 익숙해 질 것이다). 많은 역사적 개념과 역사법칙들이 역사적인 특징을 지니고 있 지 않은 개념이나 법칙으로 대치될 수 있음은 의심의 여지가 없 다. 중요한 문제는 그러한 대체가 항상 가능하냐는 것이다. 역사 적 개념 또는 법칙과 비역사적 개념이나 법칙 사이의 차이는 유전
학에서 가장 분명히 나타난다. 형질전달유전학에서 보면, 〈 순계 (純系) 〉 , 〈 이형접합체 〉 , 〈 잡종 〉 과 같은 용어들은 역사적 개념이라 고 볼 수 있다. 〈 순계 〉 는 수많은 세대에 걸쳐 동계교배가 이루어 진 한 가계의 생물을 말하는데, 이 경우에는 어떤 특정 형질에서 특이한 형태가 나타나는 일이 거의 없다. 역사적 개념으로서의 〈 잡종 〉 은 양粹의 아주 디론 집단을 교배시키는 것을 말한다. 〈이 형접합체 〉 와 〈 동형접합체 〉 도 역사적인 정의를 내릴 수 있다. 그 러나 이들 개념은 아주 분명하여야 한다. 왜냐하면 여러 개의 서 로 다론 과거 역사가 하나의 동형집합체나 이형집합체를 만들어 낼 수도 있기 때문이다. 그러나 표현형의 단계가 아니라 유전자의 미세구조를 논의한다면 〈 동형집합체〉와 〈이형접합체〉는 역사적인 특성을 잃게 된다. 두 대립유전자의 구조가 서로 다르기 때문에 이 두 대립유전자는 이형접합이고 따라서 그 유전자에 대한 형질 이 이형집합이므로 결국 한 개체는 특정한 형질에 대해 이형집합 이 되는 것이다. 따라서 두 대립유전자의 조상이나 가계를 전혀 모른다 하더라도 그들이 이형집합이다라고 유전학자들이 단정할 수 있는 것이다. 유전학에 대한 논의 전반을 통해서 우리는 유전자 개념에 관한 역사적 차원을 무시해 왔다. 이제 그 문제를 이야기해 보자. 상동 염색체는 감수분열시 짝을 이루는 염색체이다. 대립유전자는 상동 염색체 위에서 서로 마주하는 자리에 위치한 유전자인데, 상동(相 同)이라는 말을 좀더 확대시킨다면 두 대립유전자는 상동적이라 고 바꾸어 말할 수도 있을 것이다. 그러나 〈상동〉이라는 말은 전 화적 상동성을 언급할 때도 사용되는데, 전화적 상동의 개념은 표 현형으로 나타난 형질을 말한다.@ 예를 들면, 포유류의 유스타키
@ 상동(相同). 여기서 이야기하는 상동은 상동영색체에서의 상~과
는 개념이 다른 말이다 . 상동염색체의 상동은 부계에서 온 염색체 와 모계에서 온 염색체가 감수분열시에 짝을 지을 때 그 짝 각각 울 지칭하는 것이고, 여기서의 상동은 두 종류의 생물에서 의견상 형태적으로나 기능적으로 전혀 다른 기관처럼 보이더라도 그 발생 기원이나 체제상 동일한 기관으로 간주되는 것을 말한다. 이 반대 어는 상사(相似)이다.
오관은 상어의 숨구멍과 전화적으로 상동이다• 그 둘은 다 초기 유악 척추동물의 아가미 툼에서 발달되어 나온 것이다. 대부분의 현존 포유류는 유스타키오관이 인두에서 중이(中耳)까지 연결되 어 있어서 고막 내의의 공기 압력을 같게 해주는 역할을 한다. 대부분의 현존 상어류에 있는 숨구멍은 바깥에서 아가미로 물을 나르는 통로로 사용된다. 포유류의 유스타키오관과 상어의 숨구멍 은 구조나 기능이 이렇게 다르지만 서로 진화적 상동이다. 왜냐하 면 일련의 조상들을 계속 거슬러 올라가면 같은 조상 그룹에 있 었던 같은 구조에 이를 수 있기 때문이다. 어떤 생물학자들은 그 중간에 얼마나 많은 구조적 변화가 일어났는가에 상관없이 계통 발생적으로 같은 조상 구조에서 파생된 두 구조는 서로 진화적으 로 상동이라고 말한다. 또 어떤 생물학자들은 얼마나 많은 변화가 일어났고 얼마나 오래 전에 두 개가 서로 다른 계통으로 갈라지 게 되었는가에 따라 진화적 상동 정도를 따지기도 한다. 유전자와 염색체에 대해서도 같은 이야기를 해볼 수 있다. 용 어의 의미를 확장해서 두 유전자는 〈 전화적으로 상동이다 〉 라고 말할 수 있다. 일련의 자기 복제 과정을 거슬러 울라가서 공통 조상의 유전자까지 도달할 수 있다면 말이다. 마찬가지로 일련의 세포분열을 거쳐 공통의 조상염색체로 거슬러 올라갈 수 있다면 두 염색체도 전화적으로 상동이다. 그러나 교차가 일어나기 때문 에 유전자와 염색체 단계에서의 진화적 상동성을 그렇게 간단히
알아낼 수는 없다` 생물학지들은 서로 디른 두 개의 구조가 전화적 의미에서 서로 상동인지 아닌지에 대해 큰 관심을 보여 왔다. 계통을 재구성한다 는 견지에서는, 수령이나 평행적 발달로 인해 생긴 유사성과 공통 조상을 가졌기 때문에 생기는 유사성을 구별하는 것이 극히 중요 한 일이다• 우리가 목적하는 바에 있어 결정적인 문제는 이러한 구분이 생물학적 이론 특히 진화론의 목적으로 만들어져야 할 필 요가 있는가 하는 것이다. 두 개의 시스뎀이 한 법칙의 영역 내 에 있을 만큼 아주 비슷하면서 그러나 과거 역사가 다르기 때문 에 서로 다르게 행동하는 것이 가능한 일일까? 생물학자들은· 정 교하게 체계화된 시스템을 제시하였는데, 이러한 체계화를 무시할 수가 없었던 것이다. 또한 이러한 체계화는 기나긴 진화 과정의 결과이며 그 중의 일부는 시스템의 구성 속에 그 흔적을 남기고 있다. 이러한 흔적이 과거 역사를 추론하는 데 사용될 수 있다는 것은 틀림없으나, 진화론을 적용시키기 위해 전화적 상동성과 비 전화적 상동성을 꼭 그렇게 구분해야만 할 필요가 있을까? 저명한 생물학자들의 견해도 양쪽으로 갈라져 있다. 양자의 차 이라는 것은 생물학적 이론에 있어서의 역사의 역할에 관한 것인 데, 이 문제와 관련해서 두 가지 연구가 계속되어 왔었다. 그 첫 째 연구에서 도브잔스키 Dobzhansk y는 한 집단의 초파리를 여러 개의 소집단으로 나누고 그 각각을 환경 변화가 비슷하게 일어나 는 곳에 놓았다. 이 경우, 그 각각의 소집단은 모두 비슷하게 반 응하였다. 다음에 그는 지리적으로. 다른 몇 개의 장소에서 같은 종의 초파리들을 잡은 후 그들을 서로 섞어서 혼합된 집단을 만 들고 그들을 역시 환경 변화가 비슷하게 일어나는 곳에 놓았다. 하지만 이 경우에는 초파리들이 서로 다르게 반응하였다. 따라서
도브잔스키는 〈 결과는 실험동물의 조상이 어떤 지리적 장소에 있 었는가에 달려 있다 〉 라고 결론지었다 (Munson, 1971 : 194). 좀더 정확히 말하자면 실험 결과는- 실험동물의 조상들이 어떤 유전적 구성을 가졌는가에 달려 있다고 할 수 있을 것이다. 지리적으로 다른 여러 장소에서 취한 집단둘은 같은 종이면서도 그 유전적 구성이 조금씩 다르기 때문에 그들을 섞어 놓지 않는다 하더라도 같은 환경 변화에 대해서 서로 다르게 반응하게 될 것이다. 결국 도브잔스키의 실험은 생물학적 시스템이 반드시 역사적이어야 할 필요는 없지만, 특정한 종의 진화에 대하여 예측하려면, 그 종을 구성하는 각 집단간의 유전적 변이에 대해 알고 있어야만 한다는 것을보여주고 있다. 레원틴 Ri ch ard Lewon ti n7 ' 은 진화에 있어서의 역사성의 원리 에 대해 더욱 정교한 논증을 제시하였다. 유전적으로 동일한 집단 을, 선택압력은 같지만 그 순서가 거꾸로인 상황에 놓게 되면 〈 그 두 집단은 서로 다른 종착지에 도달하게 될 뿐 아니라 매우 다론 역사를 지니게 된다. 그러나 더욱 중요한 것은 그들의 보편적 행 동이 완전히 다르다는 것이다 〉. 따라서 그는 〈 한 집단의 보편적 인 유전구조는 환경의 정적 확률분포(靜的 確率分布)에 의존할 뿐 아니라 역사적인 순서에도 의존한다. 만일 환경에서 나타나는 역사적 순서가 그 집단이 적응하는 데 있어 중요한 변수로 작용 한다면, 그러한 역사적 순서 또한 그 집단의 고유한 특성을 만드 는 하나의 유입 요소로 보아야 한다〉고 결론지었다. 레원틴의 실 험은 뛰어난 것이기도 하고, 또 법칙이 지배하는 이러한 현상들과 양립될 수도 있을 것이며, 나아가서는 그러한 현상을 비역사적인
7) Is Natu re Proq ab le or Cap ri c i o u s? Bi oS cie n ce, 16(1 9 66), pp. 25-26.
법칙 심지어는 과정의 법칙으로 대치시킨다 해도 이들과 양립 가 능하게 될 것이다. 동일한 유전구성으로 된 집단이라 해도 처음부 터 그들을 서로 다른 선택압력에 놓았다면 그들이 서로 다르게 반 응하리라는 것은 누구나 쉽게 예상할 수 있다. 그러나 같은 선택 압력이지만 그 순서가 거꾸로인 상황이라는 것은 전혀 다론 이야기 인 것이다. 여기서 우리가 말하고자 하는 것은, 서로 다른 역사를 지녔으나 대단히 바슷한 유전적 구성으로 전화해 온 두 집단의 경우, 유전구성이 비슷하다고 해도 이 두 집단은 같은 환경 변화 에 대해서 서로 다르게 반응한다는 것이다. 왜냐하면 그들은 서로 다른 역사를 거쳤기 때문이다. 주의할 점은 만일 생물학적인 현상이 위에서 논의한 것과 같은 의미에서 〈 역사적 〉 이라면, 그것은 애초부터 비인과적인 것임을 스스로 드러내 보였을 것이라는 점이다. 현재의 상태에 관한 〈완 벽한 〉 지식만으로는 미래를 예측하기에 불충분하고 시스뎀의 과 거 역사에 대한 지식도 함께 요구된다. 그러나 〈완벽한〉 지식에 대한 요구를 버리고 우리 스스로를 현존하는 이론으로 국한시킨 다면, 역사법칙이 의미하는 바가 그렇게 엄청난 것만은 아니다. 현존 이론에 따르면, 두 시스템이 동일한 법칙의 영역 내에 속할 만큼 충분히 유사하면서도 아주 다르게 행동하는 것은 가능한 일 이다. 하지만 그것은 그들의 과거 역사가 다르기 때문이 아니라 현재의 구성에 차이가 있기 때문이다. 앞에서 설명한 도브잔스키 의 실험은 만일 예상 밖의 결과를 피하려면 두 집단이 얼마나 비 슷해야 충분히 비슷하다고 간주할 수 있는가를 보여준다고 볼 수 있다. 델브뤽 (Max Delbriic k , in Blackburn, 1966 : 119) 은 〈수십억 년에 걸친 진화의 기록을 지니고 있는 세포는 물리적인 사건보다
는 역사적인 것을 더 많이 보여주고 있다. 역사가 오랜 새 한 마 리를 물리적인 몇 마디 말로 설명할 수 있을 만큼 우리가 현명하 다고 생각해서는 안 된다〉고 주장하였다. 심프슨 (S i m p son, 1964) 은 〈중력은 역사가 없는 〉 반면, 전화하는 종은 역사가 있기 때문 에 전화론과 중력이론은 다르다고 했고, 메이어도 생명체는 역사 의 산물이며 무생물과는 다르게 다루어야 한다는 것을 수차 강조 했다. 이 말들에 담긴 메시지는 부분적으로는 분명히 쉽게 받아들 여 질 수가 있다. 생명체는 전화하고 있다. 따라서 생물학에서 전 행되고 있는 훌륭한 작업 중의 하나는 계통적 발생순서를 재구성 하는 일이다. 그러기 위해서는 전화적 상동성이 유전자 수준에서 일어나는 것인지, 염색체 수준에서 일어나는 것인지 아니면 표현 형적 구조에서 일어나는 것인지를 구별해야 한다. 또 전화론에 따 르면, 생물은 경쟁자보다 에너지원을 더 효율적으로 활용할 수 있 을 때만 생존할 수가 있다. 따라서 생물은 모두 대단히 효율적인 기직을 가지고 있을 것이라고 예상되고 또 실제로도 대부분이 그 렇다. 인체의 경우 콩팥의 헨레 고리, 인두에서 볼 수 있는 호 흡기와 소화기의 교차, 망막에서 상이 거꾸로 맺히는 것 등은 그 들이 기능을 수행하는 데 있어서 최선의 방법이라고 할 수는 없 다. 그러나 우리의 조상들이 처했던 그 당시의 상황으로는 그것이 최선의 것이었고, 또 그것은 충분히 괜찮은 것이었다(이것은 제 4 장에서 좀더 논의될 것이다). 그러나 이 모든 것을 듣고 나서도, 우리는 앞에서 언급한 인용 문동기 암시하고 있는 것 중, 몇몇에 대해서는 아직도 반대를 할 수밖에 없다. 계통을 재구성하는 일은 매우 타당한 일이다. 그러 나 과학법칙과 이론을 구성하는 일 또한 그와 마찬가지로 타당하 다는 것이 그러한 반대 중의 하나가 될 것이다• 또 중력에는 역
사가 없지만 자연선택에도 역사가 없기는 마찬가지이며, 종이 역 사를 지니고 있는 것은 사실이지만 은하계도 마찬가지로 역사를 지니고 있다. 특정 종이 지닌 특성 중 어떤 것은 그들이 거쳐온 역사의 특수성을 가지고서만 설명이 될 수 있는 것처럼, 특정 은 하계의 성질 중 어떤 것은 그들의 과거 역사를 가지고서만 설명 될 수 있다. 실제로 우주론자들도 은하계의 과거 역사를 재구성하 는 데 있어, 전화론자들이 종의 과거 역사를- 재구성하는 데 사용 하는 기술과 비슷한 것을 사용한다. 또한 은하계에는 종보다 과거 역사에 대한 흔적이 적겠지만, 그러한 흔적이 아예 없다 하더라도 역사적 재구성은 가능한 것이다. 이제까지 생물학지들은· 유전자나 총체적인 표현형적 특성, 혹은 종들이 현재 서로 다르게 기능하고 있는 이유가 단지 그들의 과거 역사가 다르기 때문이라는 것을 증명하지는 못했다. 왜냐하면, 어떤 시스템이 현재 지니고 있는 구조란 과거 역사로 인한 것이기는 하지만, 문제가 되는 것은 그 들의 과거 역사 모두가 아니라 그 중 중요한 부분뿐이며, 중요한 부분이란 현재 그 시스템의 구성 속에 남아 있는 부분이기 때문 이다 . 여기서 오해의 소지를 줄이기 위해 어떤 단서를 첨가하는 것이 좋을 것 같다. 대부분의 철학자들처럼 필자도 어떤 것이 원리적으 로 가능한 것이며 어떤 것이 불가능한 것인지가 일차적 관심사이 다. 반면에 과학자들은 무엇을 할 수 있고 무엇을 할 수 없는가 에도 이와 똑같은 관심을 가지고 있다. 왜냐하면 그들은 바로 과 학을 실행해야 할 사람들이기 때문이다. 필자는 원리상 역사법칙 은 생물의 현상태(유전적 구성에 대한 방대한 지식까지 포함하여) 를 다루는 법칙으로 대치될 수 있다고 주장해 왔다. 그러나 실제 로 생물학자들은 역사법칙 없이는 그렇게 할 수 있을 만큼 충분
한 지식을 갖고 있지 못하다. 그둘은 그들 앞에 놓여진 표본의 유전적 구성도 모르며, 단지 그들이 아는 것은 흰 눈을 가진 암 컷과 붉은 눈을 가전 수컷의 자손이라는 것뿐이다. 실제로 현대 형질전달유전학의 대부분은 유전물질의 분자구-조나 생합성 경로 에 대한 지식이나 관심없이 이루어지고 있는 것이다• 역사 법칙 이 비역사적인 법칙으로 대치될 수 있다 하더라도, 그렇게 해야 할 의미가 전혀 없는 경우도 많다. 그런 경우, 역사 법칙이면 충분하다. 역사 법칙이 쓸모가 있다면 그것을 사용하는 것이 현 명한 일일 것이며, 그것이 사용 가능한 유일한 것이라면 그것을 사용할 수밖에 없을 것이다. 필자가 주장하고자 하는 것은, 역사 법칙을 비역사적인 법칙으로 대체시키는 과정에서 우리가 현재 가진 것이라고는 불완전한 자료뿐이라는 것이다. 과학적 설명과 예측에 대한 포괄법칙 모델 거의 모든 백조가 흰색인 이유는 무엇인가? 질문자가 알고자 하는 것이 무엇인가에 따라 답이 달라질 수 있을 것이다. 질문을 던전 사람은 백조의 계통발생적 역사에만 관심이 있는지도 모론 다. 그러나 그렇다 하더라도 백조의 전화 과정 중 어느 단계에서 인가 하얀색으로 적응할 수밖에 없었던 이유에 대한 설명이 함께 이루어지지 않는 한 그러한 대답만으로는 충분하지가 않다. 아니 면 질문자는 현재도 계속 흰색으로 적응되어 가고 있는 이유를 알고 싶은 것인지도 모른다. 아니면 이 세상의 백조는 모두 흰색 으로 적응되었는데 유독 호주의 백조만이 검은색으로 적응되게끔 (혹은 적어도 잘못 적응된 것은 아니게끔) 만든 요인이 무엇인가
를 알고 싶은지도 모른다. 아니면 그는 흰 백조의 게놈이 어떻게 흰색의 결과를 낳게 되었는가를- 알고자 하는지도 모론다. 표면적으로 볼 때는 여러 과학자들의 설명은 서로 대단한 차이 룰 보이고 있다. 행펠과 오펜하임 (Hem p el 과 Op pe nheim , 1948) 의 고전적 논문에 동조했던 여러 과학자들은, 단 하나의 분석만으로 여러 가지 형태의 예측과 여러 형태의 과학적 설명을 할 수 있다 고 주장했다. 이는 바로 포괄법칙 모델 coverin g -l aw model 을 뜻 한다 . 우리가 목적하는 바에 비추어 볼 때, 이 모델에는 중요한 점이 두 가지 있다. 첫째 이 모델은 과학에서의 철학적 분석의 예를 보여주고 있다는 것이며, 둘째 포괄법칙 모델의 타당성에 대 해서 논의할 때마다, 그 중에서도 특히 과학에서의 설명과 예측은 어떤 의미에서 〈 대칭적 〉 이러는 명제(앞으로는 대칭명제라 칭함) 에 대해 논쟁할 때마다 언제나 진화론이 거론되어 왔다는 것이다. 과학철학자들은 과학을 하는 것이 아니라 과학 그 자체를 연구 한다. 이 말은 아주 간결하기는 하지만 상당히 애매모호하게 들린다. 과학을 연구하는 데 있어서, 철학자들은 발견된 사실을 그저 단순히 기술하는 것뿐인가, 아니면 과학이 어떠어떠해야만 한다는 법칙도 만드는가? 또한 그들이 연구하는 〈과학〉이란, 역 사적 사건과 같이 실제적인 것, 즉 실질적인 과학 이론인가 아니 면 이상적인 재구성일 뿐인가? 그들은 과학적 탐구의 산물만을 연구하는 것인가, 아니면 과정 자체를 연구하는 것인가? 어떤 역 사가들은 자신들은 사건을 수동적으로 기록하는 사람일 뿐이라고 주장하는 반면, 또 다론 이들은 단순히 기술만 하는 것이 아니라 고 이야기한다. 선택을 하고 해석하고 체계화시키고 재구성까지 한다는 것이다. 과학철학자들은 거기에서 한걸음 더 나아가 재구 성을 한 후, 수정과 재해석까지 제시한다. 때때로 과학철학과 이
론 과학이 구별되지 않는 경우도 있지만, 과학에서는 확실하게 법 칙화된 것이 과학철학에서는 시대에 뒤떨어전 것인 경우도 많다. 과학철학자들은 자신들에 의해 이루어지는 재구성과 제시들에 대 해 자신들이 주장을 해야만 하고, 그러한 재구성과 제시들이 과학 이 가전 〈 최상의 것 〉 과 잘 일치된다는 것을 보여주어야 한다고 느끼고 있다. 물론 무엇을 최상의 것으로 간주하느냐는 그것이 그 들이 재구성한 것과 얼마나 잘 일치되는가에 의해 부분적으로 결 정된다. 따라서 포괄법칙 모델을 반대하는 사람들은 전화론에 있 어서의 설명과 예측이 포괄법칙의 패러다임에 맞지 않기 때문에 그러한 모델은 그릇된 것이라고 주장하며, 또 모델을 옹호하는 사 람들은 전화론은 이 모델에 잘 들어맞는다고 주장하거나 혹은 전 화론이 이 모델에 찰 들어맞지 않는다면 그것은 전화론 자체가 이 모델에 부적합한 것이기 때문이라고 주장한다. 이런 식으로 과 학철학을 하는 것을 종종 논리적 해석 ex p li ca ti on 이라 일컫는다. 이 경우, 논리적으로 해석하는 데 있어 주제는 실질적인 과학이지 만, 어떤 철학자들은 반드시 그래야 할 필요는 없다고 생각한다. 또한 다른 철학자들이나 대다수의 과학자들은 아마도 이에 대해 어떤 특정한 구분을 하지 않는 것 같다. 하지만 그러한 구분은 반드시 필요하다. 왜냐하면 이 책의 대부분은 이러한 논리적 해석 에 해당되기 때문이다. 과학철학에 적합한 주제에 대해서는 이미 앞에서 간단히 언급 을 했지만, 어떤 과학철학자들은 그들 스스로가 과학 이론을 재구 성했던 당사자인 만큼 과학 이론의 논리적인 형태에만 관심을 두 논 경향이 있다. 크게 놀랄 것도 없이 그들은 재구성된 이론 속 에서 그들 스스로가 재구성 과정에 삽입해 넣었던 것을 정확히 찾아낼 수가 있다. 또 다른 철학자들은 과학 이론의 역사적 발전
I8o
을 그대로 따라야 한다고 생각한다. 따라서 그들은 기껏해야 부분 적이고 일시적인 재구성만을 제시할 뿐이다. 그들은 또한 실용성 에 관심을 두며, 방법론적이고 인식론적인 질문 또한 그것들이 얼 마나 논리적인가 하는 것만큼 중요한 것이라고 생각한다. 포괄법 칙 모델을 둘러싼 논쟁들은 과학적 설명과 예측에 대한 견해의 차이만큼이나 큰, 과학철학에 있어서의 철학의 차이에 의해 야기 된 것이라고 생각된다. 포괄법칙 모델에는 37} 지 요소가 있는데 그것은 하나 혹은 여 러 개의 일반 법칙 (Li, L2, … , L.), 특정한 상황 (C i, C2, …, ck) 에 대한 전술로 된 세트, 그리고 두 세트의 진술에서 추론된 사건 전술 (E) 이다. 일반 법칙은 특정한 상황 전술과 함께 엑스플래난 스 ex p lanans 를 형성하고 추론된 사건에 대한 전술은 엑스플래난 둠 ex p lanandum 을 형성한다. 설명과 예측을 형성하는 논증의 도식 들을 모두 합하면 아래와 같이 표현될 수 있을 것이다 (Hem pe l, 1965 ; Rudner, 1966 참조) . LcI1,, Lc22,, ……,, LCkr } 엑스터난스 l설 명 E } 엑스플래난둠 헴펠과 오펜하임은 적어도 한 개 이상의 통계법칙을 담고 있는 확률적 설명들도 있다는 것을 인식했지만, 그들의 원논문은 보편 적 법칙만을 담고 있는 연역적 설명과 예측에만 국한시켰으며, 확 률적 설명은 나중에 발표된 논문에서 다루었다 (Hem pe l, 1965). 포괄법칙 모델의 옹호자들 중 일부는 형태에 상관없이-보편적 이든 통계적이든―-연역적 연관성만 있으면 과학적 설명이나 예
측을 하는 데 충분하다고 주장했고, 이 모델에 대한 초기의 비판 중 많은 부분이 바로 이 점에 초점을 맞추고 있다. 하지만 이러 한 논쟁은 우리가 하려는 논의에서는 무시하기로 한다. 또 위에서 언급한 특정한 상황이란, 원래 〈 선행(先行, ante c edent) > 조건이 라고 불리다가 논쟁이 진행됨에 따라 이러한 표기가 부적당하다 고 판명되었던 것인데, 이것은 저자들의 원래 의도를 파악하는 하 나의 단서가 될 수 있을 것이다. 마지막으로, 포괄법칙 모델은 유 일무이하게 특이한 진술이나 일반적 전술 양쪽에 대한 추론에 모 두 적용되는 것이지만, 문제를 보다 단순히 하기 위해서 우리는 전자에만 국한시키기로 한다(통계적 일반화는 통계적 일반화 그 자체에서 연역될 수 있다. 그러나 통계법칙에서 유일무이하게 특이 한 전술로 추론을 하게 되면 필연적으로 확률적이 된다). 대칭명제에 대한 논의로 들어가기 전에 몇 개의 단어에 대한 〈일상적인 용법〉에 대해 이야기해야만 하겠다• 행펠과 오펜하임 이 처음 포괄법칙 대칭명제를 만들었을 때 독자들은- 그들의 분석 이 해석스펙트럼 상에서 서술 쪽에 더 치우쳐 있다고 생각히는 것이 당연했으나, 논쟁이 진행됨에 따라 옹호자들은 스펙트럼의 법칙쪽 끝을 향해 계속 조금씩 조금씩 이동해 갔다. 이 책의 일 반적 취지에 따르면 필자 자신의 입장이 어떤가를 밝히는 것이 마땅하겠지만, 이 스펙트럼 상의 어디쯤에 필자의 견해가 놓여 있 는지에 대해 전혀 제시하지 않으려 한다. 이미 소개한 것처럼 대 칭명제는 과학에 있어서 설명과 예측 사이의 중요하고 또 분명한 관계이다. 만일 그것이 전부라면, 대칭명제에 대한 비판은 둘 사 이의 관계가 중요할 때는 그 관계가 불분명하고 또 그것이 분명 한 관계일 때는 둘의 관계가 중요하지 않다는 것을 보여주었다고 볼수있다.
설명과 예측 사이의 대칭 일반적인 법칙을 담고 있는 하나의 사건에 대한 설명에만 국한 한다면, 〈 설명 〉 은 혼히 사용되는 것처럼 엑스플래난스만을 지칭 한다. 한 사건이 일어났다는 것과 그 사건이 법칙 세트(일련의 법 칙으로 된)와 특정 상황에 대한 전술을 향해 전행되고 있다는· 것 울 안다면, 우리는 그러한 법칙 세트와 상황에 대한 전술로부터 사건의 발생을 추론할 수 있다. 그러므로 법칙과 특정 상황과 사 건에 대한 지식이 있어야만 한다. 가장 논란이 적고 엄격한 의미 로서의 〈 설명 〉 속에서, 알려지지 않은 상황에 관련된 알 수 없는 법칙을 가지고 아직 일어나지 않은 사건을 설명할 수는 없다. 혼 히 사용되는 것처럼 〈 설명 〉 은 인식론적인 공약(公約, com mit- men t)이다. 그러나 또 다른 식으로 사용할 수 있는 선택의 여지 는 그대로 남아 있다 . 설명된 사건은 특정한 상황의 이전, 이후, 혹은 그 중간 언제나 일어날 수 있지만, 그 사건이나 특정한 상 황은 모두 설명이 주어전 시간 이전에 일어나야만 한다. 이것이 바로 특정 사건이 설명에 〈선행한다〉는 뜻이다. 한편 〈예측〉은 그 사용이 더욱 제한되어 있다. 예측이란 미래에 일어날 사건의 결과에 대한 전술이다. 〈예측〉이라는 단어의 기본적인 의미에 의 하면 이미 일어난 것을 예측할 수는 없다. 그러나 예측은 그러한 예측을 하는 데 대한 이유를 필요로 하지 않고 보통 그러한 이유 를 포함하고 있지도 않다. 좀더 큰 의미로 본다면 예측은 어떠한 근거를 가지고 있을 필요도 없고 추론의 결과일 필요도 없다(예 롤 들면 예언들). 그러나 예측이 그 근거를 가지고 있고 추론의 결과일 때는 특정한 상황이 일어날 상대적 시간은 설명의 경우보 다 그 범위가 더욱 좁고 정확하게 된다. 사건이 일어난 후에 그
사건을 예측할 수도 없고 알려지지 않은 상황으로부터 추론할 수 도 없는 일이기 때문에 특정한 상황은 예측을 하기 전에 그리고 추론된 사건이 일어나기 전에 일어나야만 한다(과학자들은- 또 다 론 세트의 특정한 상황을 근거로 하여 어느 특정한 상황에 대하여 예측을 할 수도 있을 것이다. 그러나 그 경우도 결국은 현재나 과 거의 상황에 의존해야 한다). 원논문에서 행펠과 오펜하임 (Hemp el, 1965 년 재판)은 다음과 같이 주장했다. 죽 동일한 분석이 〈 과학적 설명뿐 아니라 과학적 예측에도 똑같이 적용된다. 다만 둘 사이의 차이점은 실제적인 사 용상의 문제에 있다. 만일 E 가 주어지면, 죽 우리가 E 에 의해 기 술되는 현상이 먼저 일어나고 적합한 전술세트 CI, c2, …, ck, L i. Li, …, Lr 이 나중에 주어졌다는 것을 알고 있다면, 우리는 문제의 현상에 대해 설명을 하고 있는 것이다. 그러나 만일 진술 세트기· 먼저 주어지고, 기술하는 현상이 일어나기 전에 E 가 유도되었다 면, 우리는 예측을 하고 있는 것이다. 따라서 시간상으로 볼 때는 맞는다 하더라도 그 엑스플래난스가 알고자 하는 현상을 예측하 는 근거가 될 수 없는 한 그 설명을 완벽한 것으로 볼 수는 없다 고 할 수 있을 것이다〉. 주석에 보면 또 이렇게 덧붙여 있다. 〈하 나는 과거를 향하고 다론 하나는 미래를 향하고 있다는 사실과, 설명과 예측의 논리적 유사성은 라이헨바흐 Re i chenbach7} 사용 했던 ‘후기술가능성 (後記述可能性, p os tdi c t ab ility)'과 ‘선기술가 능성 p re di c t ab ility’이라는 말 속에 잘 나타나 있다(Q uan t um Mee- hanic s , 12 쪽) .> 과학철학자들에 의해 그렇게 무의식적으로 사용되면서 또 그렇 게 면밀히 검토된 단어도 아마 없을 것이다. 만일 행펠과 오펜하 임이 이러한 견해가 불러올 대소동을 예상했더라면, 분명히 표현
에 좀더 신중을 기했을 것이다. 설명과 예측이 어떤 의미에서건 확실히 대칭이라면 우리는 여러 가지 의미로 쓰는 일상적인 사용 에서 탈피해야만 하며, 상당한 해석이 필요할 것이다. 훗날 밝혀 진 바와 같이 대칭명제를 유지할 수 있는 유일한 방법은 사람들 이 그렇게 흔히 쓰는 것이 아닌 새로운 방법으로 이들을 다시 정 의하는 것이다. 그렇다고 해서 대칭명제가 과학적 추론 자체가 지 니고 있는 중요한 관계를 보여 줄 수 있다는 가능성이 배제되는 것은 아니다. 포괄법칙 모델에는 세 가지 요소가 있는데, 법칙, 특정 상황, 그리고 추론된 사건이 그것이다. 지질학에서 균일설(均一說, uni- fo r mit ar i an i sm) 이 채택된 이래 과학법칙은 비시간적인 것으로 되어 왔고, 따라서 법칙은 어떤 특정한 시간에만 일어나는 것이 아니다. 그러나 법칙 이의의 다른 요소들은 특정 시간에 일어나는 것이다. 예를 들면, E는 CI, …, ck 이전에, 이후에 혹은 중간에 일어날 수 있는 것이다. 또한 예증하기 위한 논쟁에 포괄법칙 모 델을 사용하려면, 과학자들은 각 요소들에 대해 상당한 지식을 지 니고 있어야 한다. 그러나 법칙을 알고 CI, …, C-, 7} 일어났다는 것도 알지만 E 가 일어났다는 것을 모를 수가 있다. 어쩌면 E 와 C1, …, G 가 일어났다는 것은 알지만 법칙을 모를 수도 있으며, 이런 식의 조합을 6 개나 더 만들 수가 있다. 그런데도 설명과 예 측 사이의 관계에 대한 논의를 하는 데 있어 저자들이 그들이 얘 기하고 있는 사건의 상태를 항상 주의깊게 구체화시키고 있었던 것 같지는 않다. 예컨대, 현재는 후기술p os tdi c ti on 이나 선기술 pre d ict i o ~ 설명이나 예측과 구별되고 있다. 또한 역기술(逆記 述, retr o -dic t i on ), 전기술p re di c ti on 이나 역설명 re t roduc ti on5 斗 의 구별은 말할 것도 없고 역기술이나 선기술과도 구분되고 있다.
그륀바움 Gr ii nbaum8 ) 은 단언가능성 ( 斷言可能性, asserti bi l it y ) 울 추론가능성 inf erab i l ity에 대조시키면서 대칭명제에 대한 비판 울 줄여보려고 했다. 어느 특정 과학자가 특정한 설명-논증이나 예측-논증을 단언하는 데 있어 그것이 정당화될 수 있는가 없는 가 하는 것은 가능한 모든 종류의 인식론적인 고찰에 달려 있다.
8) Temp o rally Asy m metr i c Prin c i ple s, Parit y betw een Exp la - nati on and Predic t i on , and Mecha nism versus Teleolog y, In- ducti on : Some Current lssues(Mi dd leto w n, Conn. : Wesley a n Un ive rsit y Pr~ss, 1963).
그러나 대칭명제는 추론가능성을 다루는 것이지 단언가능성을 다 루는 것은 아니라는 것이다. 설명-논증과 예측-논증을 구별하는 기준은 그러한 논증이 제기된 시간과 E 의 시간적 관계에 있다. 따라서 포괄법칙 모델의 요구를 충족시키면서 추론되는 사전보다 시간적으로 앞서서 제시되는 논증이면 그것은 예측이며 사건 이 후에 제시되면 설명이 되는 것이다. 그륀바움의 해석에 따르면 E 가 일어났다는 사실을 안다는 것과 같은 그러한 인식론적인 고찰 은 대칭명제에 적합하지 못하다. 연역된 논증이 연역되는 사건보 다 앞서 제시된다 하더라도 그러한 논증 역시 사건 이후에 제시 되는 논증처럼 연역적인 것은 마찬가지이기 때문이다• 귀납적 논 증에서도 마찬가지이다. 러드너(1 966 : 63) 는 다음과 같이 이러한 의견에 대해 동조를 표명하고 있다. 〈과학자가 사용하는 논증이 설명에 해당하는가 예측에 해당하는가 하는 것은 그러한 논증이 엑스플래난둠에 의해 서술된 사건보다 먼저 사용되었는가(이 경 우에는 사건을 예측하기 위해 논증을 사용하게 된다) 아니면 엑스 플래난둡에 뒤이어 사용되었는가(이 경우에는 사건을 설명하기 위 해 그러한 논증을 사용하게 된다)에 달려 있다.〉
하지만 논증이 지닌 비시간적 본질 ate m p o ral na t ure 에 대한 주장처럼 대칭명제도 예의일 수는 없다. 다른 사람들이 모두 예의 로 취급하는 것을 반대하는데 어느 한 사람만이 특별히 그에 매 달린다고 상상하기는 어렵지 않겠는가? 포괄법칙 모델과 대칭명 제에 대한 반대는 설명력 exp la nato r y p owe 려t 추론의 강도와 동 일시하는 데 대한 근본적인 불만에서 기인한다. 설명에는 그 이상 의 무엇이 있다고 생각되며, 특히 포괄법칙 모델이 확률적 설명을 포함하는 것으로까지 확대될 경우는 더욱 그렇다는 것이다. 행펠 울 비판하는 사람들은 과학적인 이야기에서는 연역이 일상적인 이야기에서처럼 〈 설명 〉 을 많이 내포하고 있지 않으며 귀납도 현 재 해석되고 있는 것만큼 많은 설명을 내포하고 있지 않다고 주 장해 왔다. 오히려 설명력은 전달된 정보나 통계적으로 그것이 얼 마나 적합한가에 의해 평가될 수 있으며, 가장 통계적인 설명이란 엑스플래난둠에 대한 가장 정확한 가능성 correct li ke li hood울 지 정해 주는 것이다. 그러므로 실제로 그 사건이 일어날 확률은 낮 다 하더라도 그것이 가장 정확한 가능성이기만 하면 그것은 곧 정확한 설명이라는 것이다 .9)
9) Wesley C. Salmon , Sta t i sti c a l Exp la natio n a 뼈 Sta t i sti c a l Rele- vance(Pit tsb urgh : Univ e rsit y of Pittsb urgh Press, 1971).
대칭명제에 대한 반론 포괄법칙 모델에 대한 많은 반증 사례들은 설득력과 추론의 강 도는 서로 별개의 것임을 증명하는 데에 목적을 두고 있다. 그것 들은 포괄법칙 모델이 어떤 것을 빠뜨리고 있다는 것을 보여주려
하고 있는데, 그 〈 어떤 것 〉 이란 법칙과 우연적인 일반화와의 구 분을 말한다 . 우리는 한 사건이 일어날 것이라거나 일어났다는 것 을 과학법칙에 의해 추론하는 것과 마찬가지로 우연적인 일반화 에 의해서도 추론할 수가 있다. 하지만 이 경우에는 그 추론이 설명적이 아니다. 순수한 과학법칙이 보증하고 있는 것은 그것이 무엇이든간에 우리가 과학적 설명 속에서 얻어내고자 하는 바로 그것에 대한 보증이다. 따라서 과학법칙과 이 책에서 말하는- 우연 적 일반화를 구분짓는 기준은, 과학법칙이 궁극적으로 과학 이론 속으로 포함될 수 있는 데 반해 우연적 일반화는 그렇지 못하다 는 것이라고 할 수 있다. 실제로 포괄법칙 모델을 옹호하거나 반 대하는 수많은 예들이 지니고 있는 공통적인 문제점은 그것들이 서로 연관성이 없이 제 각각이고 또 가끔은 이러한 고립성이 설 명을 만족스럽지 못하게 만들고 있는 것이라 생각된다. 그러나 포 괄법칙 모델을 반대하는 사람들은 과학법칙과 우연적인 일반화를 구분하는 우리의 기준을 별로 만족스럽게 생각하지 않는다. 그것 도 역시 무언가를 빠뜨리고 있다는 것이다. 예를 들어, 〈 헨리 앙1 ] 는 왜 죽었는가? 그것은 그가 태어났기 때문이다 〉 라고 한다면, 사람이 태어나고 죽는 일 사이에 놓인 이러한 상호관계는 보편적 인 것이기는 하지만 무엇인가가 빠져 있다는 것이다. 빠뜨린 그 무엇인가를 서술하는 가장 좋은 방법은 인과적 기작이라 불리는 것이다 .. 이 철에서는 포괄법칙 모델 그 자체보다는 이 모델의 대칭명제 에 대한 반론을 디루게 된다. 이 명제에 대한 반론들 대부분이 인식론적 비대칭성을 거론하고 있고, 또한 과학자들은 논리적 고 찰보다는 인식론적인 고찰에 더 민감한 경향이 있기 때문에 생물 학자들은 대체로 포괄법칙 모델을 반대하는 사람들 편에 서 왔다.
여기서는 대칭명제에 대한 여러 가지 반론 중에서 두 가지만 논 의하고자 하는데, 그것은 보통 원인과 그 결과 사이에 존재하는 비대칭성이며, 또 한 가지는 과거에 대한 기록과 미래에 대한 〈 기록 〉 사이에 존재하는 거의 보편적인 비대칭성이다. 대칭명제에 대한 반대 견해 중 이 두 가지가 특별히 선택된 것은 전화론이 그들 견해를 지지하는 데 인용되었기 때문이다. 여기서 우리는 이 두 가지가 포괄법칙 모델에 대한 중요한 반론이긴 하지만 진화론· 이 이들 견해에 특별히 적절한 것은 아니라는 것을 논의하게 될 것 0] 다. 갈리 Gallie , 스크리본 Scr i ven, 툴민 To ulmi n10) 은 때때로 결과를 위해 충분하다고 볼 수는 없지만 필요조건인 것들을 규정하는 것 이 어떤 의미에서는 곧 설명에 해당된다고 주장하였다. 앞에서 언
10) W. B. Gall ie, uExp la nati on in Hist o ry and the Geneti c Sc ien -ces, repr in t e d in Theor ies of Hi stor y (G lencoe, ill. : The Free Press, 1959) : M. Scriv e n, in Munson(l<:!1 1 ) : S. Tou lmin. Fore-sig ht and Understa n din g(B loom ington : Ind ian a Un ive rsit y Press, 1961).
급했던 것처럼 원인이 결과를 위해서 필수적이기는 하지만 충분 하지는 않은 경우가 종종 있고, 그 반대 역시 마찬가지여서 충분 하지만 필수적이지는 않은 경우도 종종 있다. 그러므로 양자의 경 우 모두에 있어 원인과 그 결과 사이에서 만들어질 수 있는 추론 들 사이에는 비대칭성이 존재하게 되는데, 대칭명제를 옹호하는 사람들은 충분조건에 대한 진술은 항상 설명적인 반면, 필요조건 에 대한 전술은 이러한 필요조건이 충분조건에 가까운 경우에 한 해서만 설명적이라고 주장하였다. 이를 위해 흔히 매독(일차 매 독)과 매독성 진행마비 (삼차 매독) 사이의 관계가 그 예로 인용 된다. 이 경우, 매독에 걸리는 것은 마비 상태로 전전되는 필요조
건이기는 하지만 충분조건은 아니다. 사실상 매독에 걸린 사람이 그러한 마비 상태로 진전되는 일은 아주 드물며, 치료가 되거나 아니면 마비 상태가 나타나기 전에 다른 원인으로 인해 죽게 되 는 것이 보통이다. 이제 어떤 사람이 〈 헨리 앙1 ] 는 왜 마비 상태 가 되었는가? 〉 라고 묻는다면 〈 그는 매독에 걸렸기 때문이다 〉 라는 대답을 얻을 수 있을 것이다. 그러나 이 답은 그럴 듯하지만, 어 떤 사람이 매독에 걸렸디는 사실을 알았다고 해서 그 사람이 마 비 상태로 가게 될 것이라고 예측할 수는 없으며, 오히려 귀납논 리에 대한 전통적인 재구성에 의한다면 우리는 그가 마비 상태로 전전되지 않을 것이라는 예측을 해야 될 것이다. 물론 그 사람에 대해 전혀 아는 바가 없다면 그가 마바 상태에 이르지 않을 것이 라고 우리가 예측하는 것은 당연한 일이다. 그러나 그가 매독에 걸렸다는 정보가 그 위에 추가되더라도 우리는 여전히 그가 결코 마비 상태로 되지 않을 것이라고 예측하게 될 것이다. 대칭명제의 옹호자들은 필요조건이기는 하지만 충분하지는 못 한 조건들에 의한 설명은 〈 왜 헨리 앙1 ] 가 죽었는가? 그가 태어났 기 때문이다〉라는 전술과 똑같은 것이라고 반격해 왔다. 그것들 이 모두 설명적이 아니라는 점에서 충분조건이 아닌 필요조건은 모두 동일하다는 것이다. 만일 포괄법칙 모델이 확률적인 설명이 나 예측을 포함하는 것으로까지 확장된다면 대칭명제에 대한 이 러한 방어는 대단히 강력해져서, 최상의 필요조건들이란 기껏해봐 야 다른 필요조건들에 비해 충분조건에 좀더 가까운 것일 뿐, 그 이상 아무 것도 아니다. 만일 설명력이라는 것을 추론이 얼마나 확실히 보장되는가 하는 정도와 같은 것으로 간주한다면, 거의 충 분조건에 가까운 필요조건들은 그렇지 못한 것들보다 좀더 설명 적이어야만 하는데도 말이다. 그러나 대칭명제를 반대하는 이들은
설명에는 이 이상의 무엇이 더 있다고 반박한다. 어떤 필요조건은 다른 것들보다 훨씬 설명적이며, 따라서 이러한 설명력은 필요조 건으로부터 그 결과를 얼마나 확실히 추론할 수 있는가 하는 정 도의는 최소한 부분적으로라도 무관한 것으로 보아야 한다는 것 이다. 그러나 대칭명제에 대한 이러한 특정한 반론은 실제적으로 귀납논리의 재구성, 즉 그 확률이 0.5 수준을 넘지 않으면 그 추 론은 확실하지 않은 것이라는 데 대한 불만에서 기인하는 것이다. 실제로, 조건들로 구성된 세트가 더욱 완벽해지면서(세트 내의 조 건들 수가 많아지면서 ) 그것은 0.5 라는 확률 수준에 도달하기 이 전부터 더욱 설명적으로 변하는 것처럼 보인다. 특히 그러한 조건 둘 중 일부가 필요조건인 경우에는 더욱 그렇다. 포괄법칙 모델의 대칭명제에 대한 이러한 반론이 지닌 매력은 반대의 상황, 즉 명시된 원인이 충분조건이지만 필요조건이 아닌 상황에서 더욱 명백히 나타난다. 예를 들면, 지구의 자전 방향이 역전되어도 조석파를 만들기에 충분한 여건이 된다 (Ve li kovsk y에 의함)고 했울 때 그것이 필요조건은 아니다. 조석파가 실제로 이 런 식으로 만들어지는 경우가 있다 하더라도 그것은 아주 희귀한 일일 것이다. 이러한 논증은 포괄법칙 모델의 요구 조건을 훌륭히 만족시키고는 있지만 역시 추론의 비대칭성을 보이고 있다. 왜냐 하면 원인으로부터 그 결과를 추론할 수는 있으나, 그 결과로부터 원인을 추론할 수는 없기 때문이다. 앞에서 말한 수용 가능한 인 과적 토론에 대한 분석을 우리가 받아들인다면, 그리고 이것이 바 로 행펠과 오펜하임이 의도했던 대칭성이라면, 대칭명제는 분명히 잘못된 것이다. 원인이 그 결과에 대한 필수조건이건 하지만 그 결과가 원인과 함께 일어나지 않는 인과법칙에서는, 미래에 대한 추론은 보장되나 과거에 대한 추론은 보장되지 않는다. 따라서 필
수적이지만 절대로 충분할 수가 없는 조건에 의해 이루어지는 설 명 그 자체가 오류이거나 아니면 대칭명제가 잘못된 것이라는 데 딜레마가 있다. 대칭명제를 옹호하는 사람들은 앞의 것을 선택할 수밖에 없고, 반대자들은 뒤의 것을 선택할 수밖에 없을 것이다. 필수적이지만 충분조건이 아닌 법칙들이 다른 과학 이론에 비 해 전화론에 더 많다는 것을 증명할 수 있는 경우에 한해 전화론 은 이러한 논쟁에 특히 적합하다고 할 수 있을 것이다 . 예를 들 면, 메이어는 양성 생식을 하는- 종에서 종분화가 일어나려면 유전 자 흐름을 막는 어떤 장벽이 필히 형성되어야 한다고 주장한다. 따라서 종분화가 일어났다는 사실로부터 우리는 과거에 유전자 흐름을 막는 어떤 장벽이 분명히 있었을 것이라는· 추론을 할 수 있다 . 그러나 그러한 장벽의 존재가 충분조건은 아니기 때문에 반 대의 추론은 그렇게 보장될 수가 없다. 마찬가지로 어떤 종에 의 해서도 점령되지 않고 비어 있으면서 어떤 종이건 적응해 나갈 수 있는 지역에 미분화된 종이 침입해 온다면, 그것은 빠른 속도 로 널리 확산되는, 즉 적응방산(適應放散)에 대해 거의 충분조건 이 된다 . 그러나 그것은 필요조건은 아니다. 미분화된 종이 아무 에게도 점령되지 않은 그리고 자신이 적응해 갈 수 있는 지역에 들어오게 되었다면, 우리는 상당히 높은 확실성을 가지고 미래의 어느 땐가 적응방산이 일어나리라는 것을 추론할 수가 있다. 그러 나 그 반대의 경우는 이보다는 훨씬 불확실하다. 현재 전화에 있 어서는 거의 필수적이라고 할 수 있는 조건들을 찾아내는 것이 거의 충분하다고 생각되는 조건을 얻는 것보다 더 쉬운 일이라는 것이 전리로 되어 있다. 그러나 그것은 과학의 영역 전반에 걸친 전리이지 특별히 전화에 있어서만 그런 것은 아니다. 따라서 전화 와 전화론이 대칭명제를 해석하는 데 특별히 적합한 것으로 보이
지는않는다. 우리는 원인과 결과 사이에는 비대칭성이 존재한다고 주장해 왔다. 러드너나 그륀바움에 의해 정립된 대칭명제의 해석에 따르 면, 원인과 결과 사이에 존재하는 비대칭성은 포괄법칙 모델의 대 칭명제에 조금도 어긋나지 않는다. 그들의 견해에 따르면 대칭명 제는 엑스플래난스와 엑스플래난둠 사이의 논리적인 관계를 디루 는 것이므로, 엑스플래난둠 속의 사건이 일어나기 전에 논의가 제 기되었거나 사건이 일어난 후에 제기되었거나 추론의 강도는 전 혀 변하지 않는다는 것이다. 예롤 들면, 헨리 앙1 1 의 매독 감염에 대해 알고 있다고 해서, 그가 매독에 감염되기 전에 이미 마비 상태에 이르렀다고 추론할 수 있는 것도 아니고, 또 매독에 걸린 후에 마비 상태에 이르게 되었다고 추론할 수 있는 것도 아니다. 마찬가지로, 만일 지구의 자전 방향의 역전을 안다는 것이 조석파 가 일어나기 전에 조석파에 대해 추론을 할 수 있도록 해준다면 일어난 후에도 마찬가지로 조석파에 대해 똑같은 추론을 할 수 있게 해줄 것이다. 따라서 러드너나 그륀바움의 해석에 대한 반증 들이 실제로는 얼마나 받아들이기 힘든 것인지 알 수가 있을 것 이다(행펠은 러드너나 그륀바움의 해석에 동조하지 않았다는 것을 언급하고 지나가야 하겠다. 물론 동의하지 않은 것은 그의 실수였 는지도모른다). 설명과 예측 사이에 존재한다고 생각되는 두번째의 비대칭성은 인식론적인 것이고 이미 앞에서 암시된 바 있다. 과거 혹은 미래 에 대해 추론하기 위해서는 먼저 그에 적합한 특정한 상황을 알 0 向 F 하는데, 이 특정 상황들이 지닌 특성 때문에 우리는 미래에 대한 예측보다는 과거에 대해서 더 확실하고 자세하게 재구성을 할 수가 있다. 어떤 자연 현상은 그들의 과거에 대한 흔적을 지
니고 있어서 하나의 기록으로 사용될 수가 있는 데 반해, 태풍이 오기 전에 기압계가 급강하하는 경우처럼 미래를 알리는 기계가 있기는 하지만 보통 자연 현상이 미래에 대한 〈 흔적 〉 을 지니고 있는 경우는 거의 없다. 태풍과 기압계의 경우, 둘의 일치는 우연 적인 것이 아니라 보편적인 원인의 결과이다. 다시 말해서 태풍울 일으키는 어떤 변화는 역시 기압계의 수은도 내려가게 한다. 그러 나 수은이 내려갔다는 것이 폭풍을 일으키는 원인은 아니다. 그러 므로 기압계의 변화를 통해서 태풍을 예측할 수는 있으나, 태풍을 설명할 수는 없다. 일반적으로 미래에 대한 〈 기록 〉 이란 존재하지 않는다. 이러한 주장에 대한 예의가 있을 수 있다면 그것은 예찰 (豫察 p reco niti on) 일 것이나 여기서는 그것에 대해서는 언급하 지 않는다. 현재의 전화론이 그대로 사실이라고 한다면, 그리고 현재의 사 건 속에 어떤 기록이 존재한다는 것과는 무관하게 사건의 현상황 에 대한 지식이 주어진다면, 생물학자들은 시스템의 과거와 미래 상태 모두를 추론할 수 있을 것이다. 죽 현재의 상태 중 어떤 것 이 미래의 특정한 상황을 야기시킬 것인가를 추론할 수 있을 뿐 아니라, 과거의 상태 중 어떤 것이 현재의 상황을 야기시켰을까를 추론할 수도 있다. 현재의 자료 중의 일부는 곧 기록이라는 사실 울 무시한다면, 과거에 대한 가능성의 종류는 미래에 대한 가능성 의 종류보다 결코 적지 않을 것이다. 양자의 경우 모두 추론력은 상당히 약하겠지만. 우리는 미래에 일어날 날씨 변화에 대해 찰 추론할 수가 없으며, 특히 장기간에 걸친 변화의 경우에는 더욱더 그렇다. 그러나 과거에 대해서도 이는 마찬가지이다. 다시 말해서, 기록의 역할을 하는 지질학적인 기록들을 무시한다면 같은 규칙 과 원리를 가지고 과거에 일어난 기후의 변화를 높은 정확도를
가지고 추론할 수는 없는 것이다. 또 기록이라는 지식 없이는 우 리가 미래보다 과거에 일어난 진화적 사건들을 더 많이 추론할 수도 없을 것이다. 이는 실험실 내에서 통제된 실험을 하는 경우 에도 마찬가지이다. 어떤 생물집단의 현상태와 과거 환경이 어떻 게 변화되어 왔는지를 알고 있다면 그 집단의 과거의 상태를 상 당히 정확하게 추론할 수가 있을 것이다 . 그러나 우리는 미래에 대해서도 동일한 추론을 할 수가 있을 것이다. 즉 집단의 현상태 와 앞으로 환경이 어떻게 변화해 갈 것인지를 알고 있다면 과거 의 경우와 똑같은 만큼의 정확도를 가지고 미래의 상태를 추론할 수 있을 것이다. 그러나 우리가 사건의 현상황 중 어떤 것은 기록에 해당한다는 것을 인식한다면 상황은 대단히 달라지게 된다. 죽 과거에 대한 가능성의 종류는 상당히 감소될 것이다. 지질충의 순서와 그 특 성, 화석, 현재의 DNA 염기서열 등 과거의 사건에 대한 수많은 기록들이 있다면 전화론은 과거의 계통발생적 순서를 더욱더 상 세히 재구성하는 데 사용될 수 있을 것이다. 그리고 같은 만큼의 자료가 주어전다면 미래에 대해서도 과거에 견줄 만한 재구성을 해볼 수가 있을 것이다. 그러나 실제로는 그렇지가 않은데, 전화 생물학자들이 미래롤 예측하기보다는 과거에 대해서 훨씬 더 잘 설명할 수 있는 것도 바로 이러한 사실 때문이다. 그러나 이러한 사실이 전화론이 특별히 〈역사적〉임을 뜻하는 것은 아니다. 계통 의 재구성은 전화론과 조화가 되어야만 하고, 전화론은 또 계통을 재구성하는 데 사용될 수도 있다. 그러나 뉴턴 이론과 우주발생론 에 대해서도 이와 똑갇은 말을 할 수가 있다. 계통을 재구성하는 것은 역사적인 작업이며 전화분류학 역시 역사적이다. 왜냐하면 전화분류학은 분류군을 가지고 계동발생적인 종분화를 보여주도록
되어 있기 때문이다. 그러나 전화론은 역사적인 이론이 아니다. 러드너 (1966 : 60) 는 〈 설명의 경우, 우리는 말하자면 사건 E 를 갖는다 〉 고 말했다. 그륀바움과 러드너에 의해 정립된 설명-논증 과 예측-논증의 관점에서 보면 이러한 차이는 차이가 없는 것과 마찬가지이나 인식론적인 관점에서 보면 큰 차이가 있다. 스크리 본이 강조한 것처럼, 과거에 어떤 사건이 일어났다면 그에 필요한 모든 필요조건들도 함께 일어났을 것임에 틀림없으며, 이러한 필 요조건에 대해 언급하는 것은 곧 설명에 해당된다. 어떤 상황에 있어서는, 또 어느 정도까지는 말이다 . 또한 설명되는 사건이 그 사건 자신의 역사적 흔적을 포함하고 있을 수도 있고, 나아가서는 그 사건보다 앞서 일어난 사건들을 더욱 잘 추론할 수 있도록 우 리를 도와줄 수도 있게 된다. 고유성 u niq ueness 과 이야기식 설명 과학적 설명을 하는 데 있어서 포괄법칙 모델은 여러 면에서 도전을 받아 왔다. 굿지 (T. A. Goudg e, 1961) 는 진화 연구에서 발 견되는 특이한 형태의 설명, 죽 그가 이야기식 설명 narrati ve ex- p lana ti on 이라 이름 붙인 것을 설명하기에는 포괄법칙 모델이 부 적합하다고 주장하였다. 이야기식 설명에서 포유류의 전화나 공룡 의 절멸 같은 사건들은 그러한 결과에 도달하기까지 일어난 일련 의 사건들을 자세히 나열함으로써 설명된다. 물론 설명될 사전보 다 먼저 일어난 사건들을 모두 언급한다는 것이 아니라, 선행조건 중 가장 중요한 것들만을 골라서 언급한다는 것이다. 때때로 이 조건들은 필수적이거나 거의 필수적인 것이기도 하며, 또 때로는
예상 밖의 상황이기도 한다. 거의 희박하긴 하지만 때에 따라서는 충분조건일 수도 있다. 굿지가 필요조건이 때로 설명적이다라고 했을 때, 그것이 단순히 분석 이전의 〈 설명 〉 에만 국한된다면 우 리가 그와 논쟁을 벌일 아무런 이유가 없겠으나 그가 의미하는 바는 그렇게 단순하지가 않다. 이야기식 설명을 할 때 노골적이든 은연중이든 절대로 법칙에 의지해서는 안 된다는 것이 그의 주장 이므로, 포괄법칙 모델을 옹호하는 이들의 계속되는 지루한 주장, 죽 최소한 은연중으로는 모든 설명들이 법칙을 언급하고 있다는 것(심지어는 그러한 법칙이 만들어질 수 없는 경우에도)에 대해 그가 분노를 보이는 것은 당연한 일일 것이다. 그러나 문제는 그 가 여기에서 그치지 않고 전화 속의 사건들은 모두가 고유한 것 이라는 입장을 견지하는 데 있다. 그렇다면, 설명이 과학법칙이라 는 이름 아래 포함된다는 것은 완전히 불가능한 일이다. 그에 의 하면, 〈 전화 속의 한 사건에 대해서 이야기식 설명을 할 때, 언제 나 그것은 일련의 사건으로 이루어전 어떤 부류의 사건들 가운데 속하는 한 예가 아니라 오직 단 한 번만 일어날 뿐이며 다시는 반복되지 않는 고유한 사건이다. 따라서 전화적 사건은 어떤 일반 화나 법칙을 만드는 데 적합한 재료가 될 수 없다〉. 굿지의 주장에서 어떤 의미를 찾고자 한다면 우리가 제 2 장에서 본 바와 같이 고유 사건과 필수적 고유 사건을 구분해야만 할 것 같다. 고유 사건이란 한 부류의 사건에 속하는 것이다. 예롤 들어, 카톨릭교회는 언젠가 아일랜드인 교황을 선출할 수도 있을 것이 다. 그렇게 된다면 그것은 고유한 사건이 될 것이다• 그러나 필수 적으로 고유한 사건은 아니다. 왜냐하면 그것은 다시 반복될 수도 있기 때문이다. 그러나 첫번째의 아일랜드인 교황을 탄생시키는 선거는 필수적으로 고유한 사건에 해당된다. 왜냐하면 정의상 그
것이 다시 반복되어 일어날 수가 없기 때문이다• 이러한 예가 우 리에게 주는 의미는, 한 사건은 그 자체가 고유한 것도 아니고 스스로 고유해질 수도 없으며 단지 어떤 특정한 서술 아래서만 고유할 수 있다는 것이다. 따라서 필수적 고유 사건은 그에 적합 한 이름과 어떤 명확한 서술을 지니고 있다 . 그러나 그것이 전부 다. 따라서 우리는 굿지의 견해를 전화에는 중요한 부류의 사건이 없다는 말로 해석해야 할 것이다. 굿지의 견해를 이와 같이 해석한다면, 우리는 두 가지 질문에 부딪치게 된다. 전화나 전화론에 대한 그 무엇이 굿지로 하여금 진화적 사건들은 필수적 고유 사건이라고 결론맺도록 하고 있는 가? 역사적인 이야기에 의해서 고유한 사건이 설명된다는 것은 어떤 의미인가? 굿지가 전화적 사건들이 고유하다고 주장했을 때 그는 단계통성 (單系統性)의 원리를 염두에 둔 것인지도 모른다. 모든 전화적 사건들이 분류군을 포함하고 있고 그 분류군이 필수 적으로 단계통적이라면 진화적 사건들이 필수적으로 고유하다고 주장해 보고 싶을 수도 있을 것이기 때문이다. 정의상, 단계통적 단위로서의 포유류는 단지 한 번만 전화될 수밖에 없다 . 따라서 포유류의 전화는 필수적 고유 사건이다. 그러나 다른 기술상에서 보면, 여러 가지 분류군에 있어서의 전화는 고유한 사건이 아니 다. 예를 들어, 포유류의 전화는 점령되지 않은 상태에다 적응도 가능한 지역으로 한 분류군이 침입하는 경우의 한 예로서 간주될 수도 있기 때문이다. 그런 사건은 얼마든지 한 번 이상 일어날 수 있고 실제로도 일어났었다. 요약해서, 분류군이 단계통적이어 야만 한다면 그것은 개체 하나하나가 될 것이고 따라서 과학법칙 울 그것에 직접 연관지울 수는 없게 될 것이다. 그러나 그렇다고 해서 모든 서술 하에서 모든 전화적 사건들이 고유하다는 뜻은
。四다. 보통 어떤 것이 고유하다고 말할 때 위에서 말한 것처럼 그렇 게 엄격한 의미로 〈고유하다〉는 말을 사용하지는 않는다. 그저 어떤 사건이 우주의 역사상 다시는 일어날 것 같지 않은 상태임 울 의미하는 것이다. 예를 들어, 아래의 인용문에서 심프슨 (G. G. Si m p s on, 1964 : 186) 은 물리학에서의 설명과 역사 과정에서의 설명을 대비시켰다. 물리법칙이나 기계론적 법칙들은 보통, 직접적으로 관련된 조 건들의 조합으로 이루어진 조건 세트의 존재에 의존하는데, 이런 조전들은 의도적으로 얼마든지 반복될 수가 있고 또 그들 자체가 반응이나 결과를 결정하는 데 적합한 것이다. 그러나 역사적인 과정에 있어서의 결정 조건들은 단순한 것과는 거리가 멀며 직접 적이지도 않고 반복 가능하지도 않다. 역사적 원인이란 앞서 일 어난 사건 전체를 포함하는 것으로, 그러한 원인은 결코 반복될 수 없고 경우에 따라 항상 변할 수밖에 없다. 그 중에 어떤 요소 가 반복된다 해도 그것은 역사적 원인의 반복은 아니다. 비슷한 조건들이 한 번이 아니라 두 번 일어난다는 것은 단순한 사실이 지만 그 둘 사이에는 본질적인 차이가 있을 것이며, 그 재료나 시료(전화 과정 상에 존재하는 생물 갇은 것들)에는 어떤 면에서 든 분명한 차이점이 있을 것이다. 좀더 직선적으로 해석한다면 심프슨의 이러한 견해는 종의 전 화뿐 아니라 모든 사건에 적용될 수 있을 것이다. 어떤 사건이건 다시 반복되는 일이 없을 만큼 그 특성을 아주 세분화시켜 볼 수 가 있을 것이고, 또 만일 〈원인〉이란 것을 특정한 사건 이전에
일어난 사건들 전체라고 정의한다면 일반 법칙 아래에 놓고 설명 할 수 있는 사건이란 아무 것도 없을 것이다. 예를 들어, 오래전 파충류가 차지했던 것과 똑같은 육상의 생태적 장소를 지금 어떤 새로운 생물 집단이 차지하고자 한다면 다른 모든 조건들이 동일 하다고 할지라도 그 상황이 완전히 동일한 것일 수는 없다. 왜냐 하면 이둘 장소는 현재 다른 생물에 의해 점령되어 있는 상태이 기 때문이다. 그러나 물리적 시스템에서도 이와 비슷한 관찰을 할 수가 있다. 태양계가 완벽하게 고립되어 있는 시스템이라 할지라 도 태양계의 역사를 통틀어 태양계 내의 천체 모두가 서로서로에 대해 상대적으로 동일한 위치에 한 번 이상 놓여 있었다는 것은 거의 상상 불가능한 일이다. 우리는 심프슨의 견해롤 통해 물리학과 생물학을 구분지을 수 있도록 해석하는 데 실패하였다. 그러나 그와 굿지는 아마도 그보 다 더 미묘한 점을 염두에 두고 있었던 것 같다. 경사전 면에다 공을 놓으면 당연히 굴러가는 것처럼, 사실상 자세한 설명이 없어 도 우리는 행성에 대해 정확한 예측을 할 수가 있다. 그 경우, 서 술은 짧고 단순하며 〈같은〉 사건을 수없이 일어날 수 있게 하면 서 한편으로 법칙은 보편적인 형태로 되어 있고 극히 정확하다. 사실상 과학자들의 주된 작업 중의 하나는 그들이 연구하는 현상 모두를 그렇게 보편적이고 정확한 법칙을 형성하도록 서술하는 것이다. 그 둘은 함께 존재한다. 그런데 도브잔스키, 메이어, 레원 틴 같은 유전학자들은 우리가 어느 정도 정확하게 집단의 미래를 예측하기 위해서는 집단의 유전적 구성을 얼마나 정확하게 구체 화시켜야 하는가를 우리에게 보여주었다. 심프슨과 굿지가 주장하 는 바는 그 구체화에 대한 요구 정도가 너무 커서 그러한 사건을 지배하는 어떠한 법칙도 있을 수 없다는 것인지도 모른다.
아니면 그들은 전화적인 설명에서 특정한 상황과 전화법칙으로 간주될 수 있는 어떤 것 사이에 존재하는 독특한 불균형을 이야 기하고 있는지도 모른다. 포괄법칙 모델에 따르면, 설명에는 법칙 과 특정한 상황 모두에 대한 논의가 이루어져야 한다. 포괄법칙 설명에 따르는 일반적인 예의 경우 법칙은 가장 핵심이 되는 설 명적 요소이다. 그에 비해 특정한 상황에 대한 구체화는 필수적이 긴 하지만 그렇게 중요한 것은 아니다. 그러나 진화적 설명에 있 어서는 그 반대이다. 설명에 대한 의무를 지니고 있는 것은 법칙 이 아니라 역사적 이야기의 형태로 나타나는 특정 상황이다. 어딘 가에 법칙이 숨어 있을지도 모르지만 하여튼 법칙이 언급되는 경 우는 거의 없다. 설혹 언급이 된다 하더라도 설명에 그리 큰 보 뎀이 되지 못한다. 아마도 〈 동물은 다리가 길수록 빨리 달린다〉 와 같은 형태가 되겠지만, 그것은 지극히 당연한 말인 동시에 툴 린 말이다. 포괄법칙 모델을 지지하는 사람들은 이에 대해 〈전화 적 설명의 경우에는 더욱 심하다〉고 말할 것이며, 비판가들은 〈 포괄법칙 모델에 의한 과학적 설명의 경우에는 더욱 심하다〉고 말할 것이다. 어떤 의미에서는 역사적 이야기가 상당히 설명적이 라는 우리의 느낌과 포괄법칙 모델이 요구하는 바 사이에는 일치 되지 않는 면이 있는 것이 사실이다. 그러나 어떤 경우든간에, 심 프슨이나 굿지가 이야기하는 것이 진화적 사건이 필수적으로 고 유하다는 것이라면, 역사적 이야기가 어떻게 그 필수적으로 고유 한 것을 설명할 수 있는 것인지 (어떤 의미에서든) 이해하기 어렵 다 .11)
11) 굿지의 입장에 대한 비판에 관해서는 루스 M. Ruse 의 Narra- tive Ex pl anati on and the Theory of Evoluti on , Canadia n Jo urnal of Phil o sop h y, 1(1971), pp. 59-74 를 참고할 것.
이 장에서는 〈 전화법칙 〉 을 여러 가지 의미로 구분하였다. 때로 는 특정한 계통발생적 순서에 대한 묘사를 진화법칙으로 간주했 는데, 분류군이 단계통적이라면 그러한 모든 순서는 필수적으로 고유한 것이며, 현재의 문헌에 의하면 어떠한 과학법칙에 대한 분 석에 의해서도 사건의 고유한 발생 순서에 대한 묘사는 과학법칙 으로 간주될 수가 없을 것이다. 그러나 우리가 단계통성의 원리를 버린다면 계통발생적 순서는 고유할 필요가 없게 된다. 그러면서 도 그것이 특정한 분류군을 잘 나타낼 수 있을 만큼 충분히 자세 하게 서술된다면, · 반복될 가능성이 거의 없는 계통발생적 순서는 그러한 서술에서 배제될 수가 있을 것이다. 따라서 만일 그러한 전술이 과학법칙으로 간주된다면 우리는 전회론으로부터 그러한 전술, 죽 법칙을 추론할 수 있어야만 하는데, 그러한 추론 과정에 서 부딪치는 어려움은 제 2 장에서 이미 논의한 바 있다. 돌로의 법칙처럼 눈대중으로 하는 경험적 방법도 계통발생적 순서를 재구성하는 데 사용되었는데 이들도 역시 종종 전화법칙 이라고 불렸다. 그러한 법칙들은 특정한 분류군을 다루는 것이 아 니기 때문에 단계통성의 원리는 여기서 전혀 문제가 되지 않는다. 그러나 수백 개에 이르는 그러한 잠정적 법칙에 대한 이론적 뒷 받침은 그리 대단하지 않다. 기껏해야 극히 제한된 영역 내에서의 경험적 일반화일 뿐이며, 언제나 수많은 예의를 지니게 된다. 원 인이 그 결과를 위해 필수적이거나 충분치가 않은 인과법칙은 과 학법칙과 과학적 설명 모두에 있어 개념 분석상 여러 문제를 야 기시킨다. 그런 경우, 나타난 결과로부터 원인들을 추론할 수는 있겠지만 그 반대는 불가능하다. 그러나 어쨌든 일단 계통발생적 재구성을 전화론으로부터의 추론과 구분할 수 있다면, 이런 특정 논쟁이 유독히 전화론에 부적합한 것이라거나 아니면 전화론이
그러한 논쟁에 부적합한 것이라거나 하는 것이 확실히 밝혀질 것 이다. 그렇게 된다면, 〈 전화법칙 〉 은 제 2 장에서 살펴본 전화론에 대한 여러 해석 중 하나 혹은 그 이상을 구성하는 전술을 언급하 게 될 것이다 . 이들 법칙은 전화 과정을 언급하고 있다는 점에서 〈 전화적 〉 이지만 그렇다고 해서 특별히 〈 역사적 〉 이라고 할 수는 없다.
제 4 장 목적론 목적론의 문제점 앞 장에서 논의된 주제들과 초기 그리이스인이나 후기 기독교 도들의 형이상학적 원리 사이에는 메울 수 없는 큰 틈이 있다. 돌이켜 보면 이들 목적론적 형이상학은 유추보다 조금 나을까 말 까 하는 정도에 불과하다. 아리스토텔레스의 내적 목적론에서는 유추란 생명체에 의해 나타나는 구조적인 체제와 거대한 우주 사 이의 것이었고, 기능적인 것이었다. 기독교인들과 신풀라톤주의자 들이 주장하는 의적 목적론의 경우에 있어서는, 유추란 현명하게 행동하는 선한 사람과 우주를 지배하는 전지전능한 신 사이에 존 재하는 목적적인 것이었다. 이러한 목적론들과 전화론은 분명히 거리가 멀며 결코 그게 그것이라고 말할 수 없을 것이다. 혹자는 반대 견해를 가지고 있을지도 모르지만 말이다. 그러나 아직도 목 적론적 개념을 보여주는 생물학적 현상들은 우리 주위에 많이 있 으며 생물학자들 역시 여전히 목적론적인 방식으로 이야기를 하 고 있다. 생물학적 현상을 설명하기 위해서 완전히 다른 용어가
별도로 필요할 만큼 생물학적 현성들이 물리학적 현상이나 화학 적 현상들과 그렇게 다른가? 생명계의 행동과 체제를 기술할 때 목적론적 표현 양식이 일상적인 인과론적 표현보다 편리한 경우도 있긴 하나 과연 그것 없이는 안 될 만큼 필수불가결한 것일까? 이 장의 주제가 앞 장의 주제들과 비교해 볼 때 명백한 차이점 이 있기는 하지만 그렇다고 완전히 다른 것은 아니다. 제 1 장에서 의 주요 논제는 유전자를 적절하게 정의하는 것이었다. 따라서 유 전자는 무엇인가? 유전자는 기능적으로, 구조적으로, 혹은 역사적 으로 정의되어야 하는가? 아니면 염색체 위에서의 상대적 위치로 서 정의되어야 하는가? 이런 것이 주요 관심사였다. 이 장에서 논 의될 우리의 주된 관심사 중의 하나는 목적론적 체계를 적절하게 정의하는 일이다. 목적론적 체계란 무엇인가? 이것은 기능적으로, 구조적으로, 또는 역사적으로 정의되어야 하는가? 유전자와는 달 리 어느 누구도 목적론적 체계를 〈 위치 〉 로서 정의할 것을 제안한 사람은 없었다. 그 위치가 무엇이 될는지는 모르지만 말이다. 이 름 제의하고는 공통성이 거의 없는 여러 형태의 목적론적 체계 가 존재하는가? 뚜렷이 구별이 되는 여러 가지의 개념들을 모두 포함할 수 있는 한 개의 종합적인 개념이 있겠는가? 아니면 목적 론적 체계라는 개념은 모든 것이 뒤범벅된 잡동사니일 뿐인가? 목적론의 문제점들을 경험적으로 접근해 볼 수가 있다• 죽 몇 개의 특성이나 그 집합이 그 체계 전체의 특성이 될 수 있는지 알아보고자 할 때, 전통적으로 목적론적인 것으로 간주되어 온 시 스템들을 우선 연구해 보는 것이다. 보통 두 개의 쌍으로 나눠질 수 있는 47} 지의 특성이 제시되어 왔는데, 그 첫번째 쌍은 기능적 인 정의 하나와 구조적인 정의 하나로 되어 있다. 기능적 정의는 빈도(顔度)로 정의될 수 있으며 이 경우 빈도란 시스템이 큰 폭
으로 변할 때, 혹은 시스템과 환경이 함께 큰 폭으로 변할 때 그 시스템이 어떤 선호상태(選好狀態) 또는 어떤 목표상태에 도달하 게 되는 빈도를 말한다. 기능적 정의에 동반되는 구조적 정의는 선호상태나 인과적 피드백 fe edback 의 고리 (특히 억제 피드백 ) 등을 일으키는 인과적 기작에 의해 정의된다. 두번째 쌍은 역사적 정의 하나와 구조적 정의 하나로 되어 있는데, 역사적 정의는 선 랙 과정을 통해서 본 목적론적 시스템의 기원에 관한 것이고 구 조적 정의는 프로그램이라는 말로 표현된다. 목적론의 문제는 언어학적으로도 접근해 볼 수가 있다. 물리학 자들이나 생물학자들은 자연 현상을 기술하고 설명할 때 흔히 일 상적인 인과적 용어들을 사용한다 . 물리학자들이 기체에 열을 가 하면 기체가 팽창한다고 말하는 것처럼, 생물학자들은 포유동물에 열을 가하면 땀울 흘린다고 이야기할 것이다. 그러나 생물학자들 은 열이 가해지면 포유동물이 체온을 일정하게 유지시키기 위해 땀을 홀리게 된다고 설명할 수도 있을 것이다. 하지만 어느 물리 학자도 열이 가해졌을 때 기체가 팽창하는 것이 • 기체의 온도를 유지하기 위한 것이라고 얘기하지는 않는다. 그것이 혹 사실이라 할지라도 말이다. 따라서 우리는 제 1 장에서 논의했던 멘델 유전학 의 분자유전학으로의 환원과 유사하지만 그보다는 좀더 어려운 문제에 이제 부딪치고 있다 . 표현을 목적론적인 양식에서 일상적 인 어법으로 환원하는 것이 바로 그것이다. 이 문제는 제 1 장에서 논의된 것과 비슷하다고 할 수 있는데, 그것은 두 개념 사이에 존재하는 언어적 그리고 논리적 관계를 해결해야 하기 때문이다. 그러나 이 경우는 훨씬 더 어려운 일이다. 왜냐하면 유전학의 경 우에는 비교적 합리적으로 잘 정리된 두 개의 과학 이론이 있었 던 데 반해, 목적론적인 양식으로 된 논법을 일상적인 인과적 방
식으로 환원하기 위해서는 비형식적인 두 개의 표현 양식을 놓고 해결해야 하기 때문이다. 때로 생물학자들은 전문적인 문헌에서 목적론적인 용어롤 사용하기도 하지만 목적론적인 논법 그 자체 는 전문적인 것이 아니다. 또한 과학자들은 목적론적 언어를 사용 하지만 그것을 분석하는 일은 거의 없다. 따라서 목적론적인 양식 울 인과적 양식으로 환원하는 일에 대한 결론은 멘델 유전학을 분자유전학으로 환원함으로써 도달한 결론보다 훨씬 참정적일 수 밖에 없는 일이다. 앞으로의 논의에서 〈목적론적〉이라는 말은 목표, 목적, 기능이 라는 말로 표현된 모든 것을 지칭하는 일반적이고 형이상학적으 로 중립적인 의미로 사용될 것이다. 〈합목적적(合目的的)〉이라는 말도 많이 사용될 텐데 이는 우리의 분석이 결국 목적법칙을 지 지하는 사람들의 것과 같은 분석에 이르게 되기 때문이다 .I) 〈기 능적〉이라는 말은 생명체가 패러다임의 성격을 띠는 경우에만 사 용되며 인간의 의식적인 행위가 패러다임이 되는 경우에는 대신 〈목적 p ur p os i ve. 〉이라는 말을 사용한다. 때로 사람들은 목표에 도 달하기 위해 정밀하게 계산된 행동을 한다. 그러나 때로는 다른 생명체나 심지어는 인공물들도 이런 식으로 행동한다. 아래에 계 속되는 논의는 목적론적인 체제에 관한 것인데 이것은 목적론적 과정, 행동, 실체라는 용어들을 가지고 쉽게 논의될 수 있다.
1) C. S. Pit ten drig h , in Behavio r and Evoluti on , A. Roe and G. G. Sim p so n , eds.(New Haven : Yale Un ive rsit y Press, 1958) ; and Mayr in Blackburn(1966) and Munson(1 9 71).
목적론의 본질 여기서 우리는 목적론적인 체제의 일반적 특징이라고 할 수 있 는 기준둘을 살펴보게 될 것이다. 이에는 선호상태, 닫힌 피드백 고리, 프로그램, 그리고 선택 과정에 의해 성립된 이둘의 기원 등 을 들 수 있다. 이들을 함께 고려해 볼 때, 이러한 기준들은 우리 로 하여금 체제의 단계를 구분할 수 있게 하고, 그러한 구분은 목적론적 방법에 대한 우리의 직관적인 관념 및 분석 이전에 지 니게 되는 관념들과도 잘 부합되는 것처럼 보인다. 하지만 이러한 기준들에 의해 결정되는 단계간의 경계선이 위의 관념들과 항상 일치하는 것은 아니다. 그렇다고 목적론적 시스템에 적합한 기준 들 중에서 각각은 필수적이면서 전체로는 충분조건이 되는 것들 만을 나열해서 하나의 목록을 만들어낼 수도 없다. 놀랄 일도 아 니지만, 우리가 목적론적 시스템의 본질을 아칙 발견하지 못했다 는 것을 필자는 확신하고 있다. 하나도 다듬어지지 않은 귀납적 근거만을 가지고 목적론적 시스템의 본질에 대한 연구가 사회, 예 술, 우주, 말(馬), 인류 같은 단어들의 본질에 대한 연구보다 더 많은 결과를 얻을 수 있으리라고 기대할 수는 없다. 많은 학자들 은 이러한 개념들 각각이 본질을 지니고 있으며 그러한 본질이 무엇인가를 알고 있다고 주장하고 있다. 그러나 불행하게도 실제 로 그러한 본질이 무엇인가 하는 데 있어서는 어느 학파도 의견 의 일치를 보이지 못하고 있다. 필요충분조건으로만 이루어전 단 한 개의 세트를 가지고 고도로 발달되고 견고하게 체제화된 과학 이론 속의 용어둘을 정의한다는 것은 대단히 어려운 일이다. 목적 론적 시스템의 개념에 대한 것만큼 뒤틀린 역사를 가지고 있는 그 어떤 것을 이러한 방법에 의해 정의할 수 있다면 그것은 실로
놀라운 일이다. 물론 다음 단계는 〈 목적론적 시스템 〉 을 개념의 집합체라고 생 각하는 주장이다. 조건들의 세트 가운데, 한 구성체에 대하여 각 각의 세트는 필요조건이면서 총체적으로는 충분조건이 되는 그러 한 세트는 있을 수 없다. 다만 서로 밀접하게 관련이 되어 있는 몇몇 세트들을 가지고 그 집합의 구성원이 지니는 특칭을 설명할 수는 있을 것이다. 이러한 주장은 사실일지도 모르며 사실 그렇게 거리가 먼 얘기처럼 둘리지는 않는다. 그러나 어느 특정한 용어를 집합적 개념이라고 주장하는 것은 쉬운 일이지만 그러한 개념을 둘러싸고 있는 모호함을 줄이기 위하여 그 특성들을 쭉 열거하고 또 그들이 어떻게 연관되어 있는지를 정확하게 구체화시킨다는 것은 쉬운 일이 아니다. 그러한 구체화를 위해서 우리는 우선 목 적론적 시스템으로 제시되어 왔던 두 개의 물리적 모델들, 즉 기 능적 시스템에 적합한 온도조절 시스템과 의도적 시스템에 적합 한 어뢰 모델을 논의하게 될 것이다. 이러한 모델들을 사용하고자 하는 이유는, 적어도 목적론적 시스템의 여러 특징들이 이러한 물 리적 시스템, 즉 일반적인 물리법칙들을 통해 설명될 수 있기 때 문이다. 그리고 나아가서 이렇게 단순한 시스템을 연구함으로써 얻어지는 원리들을 생명체가 지니고 있는 의식적 요소(意識的 要 素)의 구조나 기능의 문제로 확대해 나갈 수 있기 때문이다. 다음에는 목적론적 시스템의 개념을 정의하기 위해 제안된 두 쌍의 조건에 주의를 기울여 보자. 이들 조건들이 얼마나 목적론적 시스템에 잘 들어맞는가? 그 한계가 불분명한, 경계선상에 놓여 있는 여러 경우에도 잘 들어맞을 것인가? 만일 어떤 것이 목적론 적 시스템으로 간주될 수 있다면, 생명체 또한 당연히 그렇게 될 수 있어야 한다. 생명체를 배제하게 된다면 그러한 분석은 잘못된
것이다. 반대로 태양계가 포함되는 경우 그것도 잘못된 것인데 이 는 앞의 경우와 정반대의 이유 때문이다. 경계선상에 놓여 있는 경우는 또 다른 문제를 제기하고 있는데, 이들이 목적론적 시스템 의 특징 중의 일부를 가지고 있는 것은 분명하나 또 다른 일부가 결여되어 있다는 것이다. 어떤 특정한 분석 하나가 과연 나침판 자침의 움직임, 감수분열시 염색체의 배열, 컴퓨터, 매시간 일정한 간격을 두고 분출하는 올드 패이트풀 Old Fait hfu l 온천 등과 갇 이 다양한 경우를 모두 잘 설명할 수 있을까? 설령 그렇다고 할 지라도 그러한 분석이 얼마나 훌륭한 것인가를 우리가 제대로 판 단할 수 있을까? 따라서 중요한 문제는 우리가 내리는 결정에 대 한 정당성을 찾아내는 일이다. 예를 들어, 어떤 사람이 걸어다니는 투구풍뎅이라는 아이들 장 난감을 관찰하고 그 움직이는 모양을 목적론적으로 서술한다고 하자. 태영을 감아 책상 위에 올려 놓으면 그것은 책상 위에서 이리저리 돌아다니다가 모서리에 이르러 떨어지려고 할 때마다 방향을 툴게 된다. 이러한 현상은 다른 바퀴들과 수직 방향으로 놓여 있는 풀라이휠flyw heel 이 회전함으로써 나타나는 것이다. 장 난감의 앞바퀴가 책상 모서리에 이르러 떨어지려고 할 때 플라이 휠이 책상의 표면과 접촉하게 되므로 자연히 장난감은 그 모서리 에서 멀어지게 되는 것이다. 우리가 이러한 인공 물들을 목적론적 으로 다루려고 할 때 단순히 풍뎅이 같은 모양이나 장식 등에만 관심을 둘 것인가? 그 장난감이 어떻게 작동하는지 그 기작을 알 게 되었다면 그래도 대상을 여전히 목적론적으로 다루려 하게 될 것인가? 되풀이되는 선호상태, 인과적 피드백 고리들의 연쇄망, 프로그램 그리고 그것들을 유도하는 선택 과정은 어디에 있는가? 이러한 질문에 대한 답은 점차 분명해질 것이다. 문제는 그것
들을 평가하는 몇 가지 근거를 찾는 일인데, 이것은 분석을 하기 이전에 갖게 되는 직관을 제의한 것을 말한다. 목적론적 개념에 대한 모호함을 줄이고자 시도할 때 우리는 곧 어떤 하나의 점에 도달하게 되는데, 이 점은 어떤 결정을 내리게 된 특정한 이유가 없는 점, 그 이유란 단지 동물적인 선임관일 뿐인 점이다. 철학의 역사를 통해 볼 때 한 사람에게 있어서는 자명한 전리가 다른 사 람에게는 동물적인 선임관에 불과한 경우가 많았다. 전자, 유전자 혹은 종이란 무엇인가 하는 문제를 결정하는 것은 쉬운 일은 아 니지만 그래도 결정은 내려질 수 있고 또 여러 훌륭한 논증들이 그러한 결정을 뒷받침하고 있다. 왜냐하면 그러한 용어들이 과학 이론 속에 들어 있기 때문이다. 하지만 목적론적 시스템의 개념들 은 그렇지가 않다. 이것이 바로 우리가 현존하는 과학 이론 특히 전화론과 학습설에 기초를 둔 기준들을 강조하는 이유이다. 이 두 가지 이론 어느 것도 우리가 원하는 정도로까지 발달하지는 못했 겠지만 적어도 이 이론들은 분석을 전후한 직관보다는 훨씬 실제 적인 어떤 것을 우리에게 제공해 준다. 마지막으로 이 장에서 우리는 생물철학에 있어서의 세 가지 문 제들을 다룰 것인데 그 세 가지의 문제들은 서로 연관이 되어 있 다. 죽 〈포유동물에 있어서 심장박동의 기능은 혈액을 순환시키 는 것이다〉와 같은 목적론적 서술과 〈포유류의 심장박동은 혈액 순환이라는 작용을 일으킨다〉와 같은 인과론적 서술은 어떻게 다 론가? 목적론적 설명과 일상적인 인과론적 설명은 어떻게 다른 가? 그리고 이러한 각각의 설명은 과학적 설명에 있어서의 포괄 법칙 모델과 어떻게 다른가? 의 세 가지가 바로 그 문제들이라고 할 수 있다. 그러나 무엇보다도 우리의 최우선적인 관심사는 우리 가 어떠한 결정을 내리든지간에 그에 대한 어떠한 정당성을 찾아
내는 일이다. 목적론적 시스템을 위한 물리적 모델 과학자들은 유추리는 말을 빈번히 사용하면서도 그것을 믿지는 않는 경향이 있다. 아리스토텔레스의 목적론적 형이상학으로부터 사회과학의 기능주의에 이르기까지 과학에 있어서의 기능적 유추 가 가져온 커다란 실패들로 인해, 기능적 유추는 아주 나쁜 평판 울 얻게 된 것이다. 그러나 유추에 의한 추론은 발견 과정에 있어 서 가장 성공적인 방법 중의 하나이며 기능적 유추에 대한 대부분 의 비판들은 생물학 때문이 아니라 사회과학에서 그것을 제대로 사용하지 못해서 생긴 것들이다 (Hem pe l, 1965 ; Rudner, 1966). 유추에 의한 추론을 할 때, 제대로 이해가 되지 않는 시스템의 운동을 충분히 이해가 되어 있는 패러다임 시스템의 운동에 비유 시킨다. 패러다임 시스템의 운동을 지배하는 원리들을 잘 알려져 있지 않은 시스템에 적용할 수도 있기 때문이다. 때로는 패러다임 시스템과 비유된 시스템 사이에 상호적인 관계가 성립되기도 한 다. 처음에는 패러다임에 대한 지식을 비유된 시스템에 확대 적용 함으로써 그러한 시스템을 이해하는 데 도움을 얻울 수 있지만, 비유된 시스템에 관해 얻어진 새로운 지식이 거꾸로 패러다임을 이해하는 데 도움이 되기도 한다. 때로는 유추로 말미암아 비유될 시스템으로 선택된 시스템이 패러다임보다도 더 이해가 찰 되기 도 한다. 사실상 이러한 현상은 현대 생물학과 심리학에서 실제로 일어났던 일이다. 과학자들은 현재 생명체, 특히 인간의 뇌의 구 조나 기능에 대해서보다는 거대한 우주의 움직임에 대하여 더 많
이 알고 있다고 생각한다. 과학자와 철학자들은 생명체나 자각을 지닌 행위자의 운동으로부터 순수한 물리시스템으로 추론해 가는 대신에 물리적 모델을 가지고 생명체를 이해하고자 노력하고 있 댜 이제 그러한 물리적 모델을 분석하는 일로 관심을 돌려보자. 기능적 시스템, 특히 항상성 (恒常性, homeos t as i s) 0을 지닌 시 스템에 적합한 모델로 온도조절 장치가 지주 거론되어 왔다. 이
® 항상성은 생물 개체 내에서 어떤 상태가 일정한 범위 내에서 항 상 유지되는 현상을 말한다. 사람의 체온이 37°C 안팎울 항상 유지 하고 있는 것도 하나의 항상성이다.
장치로 실내 온도는 일정하게 유지된다. 예를 들어 온도조절 장치 롤 72°F 에 맞추어 놓으면, 실내온도가 그 이상으로 울라갈 때는 온도가 다시 내려갈 때까지 냉방장치가 가동되다가 72 ° F 에 이르 면 저절로 꺼지게 된다. 온도가 72°F 이하로 내려가면 이번에는 반대로 난방장치가 가동될 것이다. 따라서 온도는 정확히 72 ° F 는 아니더라도 그 근방을 오르락내리락 한다. 이에 비해 어뢰는 목적 행위에 적합한 모델로 제시되어 왔다. 어뢰는 동력 프로펠러와 소 리가 나는 근원지를 찾아가는 장치를 갖추고 있다. 어뢰는 표적 선박이 있는 방향 쪽을 대충 겨냥하고 있다가 선박의 엔진에서 나오는 소리를 정확히 포착하게 되면 조종장치에 의해 그곳으로 유도되므로 표적 선박이 아무리 이리저리 움직인다 하더라도 어 뢰는 계속해서 그 표적을 찾아갈 수가 있다 . 어떤 면에 있어서는 이러한 모델들이 두 시스템의 전형적인 모 델이라 할 수도 있지만 다론 측면에서 보면 그렇지도 않다. 온도 조절 모델에서는 하나의 변수가 시스템 내의 여러 다른 부분의 변화들을 희생시키면서 어떤 일정한 한계 내에서 유지가 되지만, 어떤 항상성 시스템의 경우에는 그처럼 계속 유지되어야 할 것이
하나의 변수가 아니라 시스뎀 전체의 상태인 경우도 있기 때문이 다. 예를 들면, 유전자형과 표현형의 관계는 발생적 (發生的) 항상 성의 한 예로 간주될 수가 있는데, 발생적 항상성에 있어서 어떤 특정 유전자형은 환경 조건이 달라져도 항상 동일한 표현형으로 나타나게 된다. 마찬가지로 한 집단은 환경의 변화에 의해서 혹은 새로운 유전자의 유입으로 인해서 집단의 평형 상태가 끊임없이 방해를 받는다 하더라도 그렇게 변화되는 환경에 대해서 늘 적응 된 상태를 그대로 유지하여야 한다. 그러한 조절 능력을 우리는 유전적 항상성이라 부른다. 항상성 시스템에서는 하나의 변수 또는 시스템 전체의 상태가 환경이나 시스템 그 자체의 여러 변화에 상관없이 (그렇다고 무한 정한 것은 아니지만) 일정한 범위 내에서 유지된다. 그러나 기능 적 시스템의 또 다른 형태, 죽 시간이 경과함에 따라 규칙적이고 순차적으로 일어나는 일련의 변화들까지도 목적론적으로 기술되 는 경향이 있다. 생명체의 생활사의 어느 한 시점에서 보면 수많 은 항상적 기작들이 그 체제를 일정하게 유지시키고 있지만, 한편 으로 그 생물체는 발생학적 발달을 계속하고 있다. 이러한 안정된 흐름을 카날화 cana li za ti on 혹은 호메오레시스 homeorhes i s 라 한 다. 호메오레시스에서는 어떤 하나의 선택된 상태만이 유지되는 것이 아니라, 구체적으로 명시된 통로를 따라 일련의 선택된 상태 들이 차례대로 유지된다. 말의 목에 씌운 굴레는 목이 커짐에 따 라 차차 늘어나지만 한 번 늘어난 굴레는 다시 줄지는 않게 되는 데, 이것은 호메오레시스 시스템에 대한 하나의 간단한 물리적 유 추라고할수있다. 호메오레시스 시스템은 여러 면에 있어서 목적론적 시스템의 두번째 형태, 죽 목적 시스템과 비슷하다. 많은 철학자들과 과학
자들이 주장하는 것처럼 목적행위(목적이 있는 행동)는 미래에 일어날 사건이 인과론을 도입하지 않아도 설명이 가능하다. 어뢰 의 경우, 보통 인과론적 고리는 표적 선박의 엔진에서 나는 소리 로부터 그것을 감지하는 장치로, 그리고 다시 어뢰의 조종장치로 이어지는데, 어뢰의 목표는 어떤 미래의 사건, 즉 선박의 파괴라 는 것이지만, 이 미래의 사건은 어뢰의 현재의 움직임을 결정하지 는 않는다 . 따라서 어뢰가 쫓는 것은 목표 대상인 선박일 뿐이지 미래의 사건이 아닌 것이다. 온도조철 장치와 어뢰 사이의 일차적인 차이점은 전자의 경우 에는 목표물이 없다는 것이다. 그러나 이 점에 있어서 어뢰를 목 적론적 행위의 전형적인 모델로 볼 수는 없다. 목적 행위는 어떤 목표물이 없는 상태에서도 빈번히 나타나기 때문이다. 예를 들어, 프로 권투선수의 목표는 다가오는 시합에서 이기는 것일 데지만 링에 오르기 전까지는 목표의 대상이 없다. 어떤 개가 토끼를 뒤 쫓는 경우도 있겠지만 , 어뢰가 적절한 신호를 포착할 때까지 이리 저리 움직이는 것과 마찬가지로 개 또한 어떤 특정한 대상을 쫓 는 것이 아니라 뒤쫓아갈 어떤 것을 찾기 위해 그냥 쿵쿵거리고 있는 경우도 있다. 호메오레시스 시스템을 하나의 주기를 통해 전 행하는 항상적 시스템으로 보거나, 혹은 항상적 시스템을 연속적 인 (success i ve) 주기를 통하여 진행하는 호메오레시스 시스템으로 본다면, 항상적 시스템이나 호메오레시스 시스템은 동일한 패러다 임에 비유될 수가 있을 것이다. 이러한 방법을 목적행위에도 똑같 이 적용해 보는 데 방해되는 것이 있다면 그것은 몸과 마음의 문 제일 뿐일 것이다.
선호상태와 음성 피드백 많은 철학자들과 과학자들은 목적론적 시스템을 비목적론적 시 스템과 구분할 때 사용하는 두 개의 기준, 죽 선호상태 pre fe r red st a t es 와 음성 피드백 neg a ti ve fe edback 엇을 분석해 보려고 노력 해 왔다. 그러나 그러한 분석들은 대단히 복잡하고 고도의 전문적 인 개념 및 용어들을 사용하고 있기 때문에 분석 형식의 정교함 울 그대로 반영하면서 좀 쉽게 표현한다는 것은 문외한에게는 대 단히 어려운 일이다. 어려울 뿐만 아니라 그와 같은 시도는 사실 상 성공할 수 없을 것이다. 그러므로 여기서 필자가 하고자 하는 일은 목적론적 시스뎀울 분석하는 데 사용되는 두 개의 대표적인 시도들을 선호상태와 인과적 피드백 고리라는 측면에서 서술해 보고 그 각각이 어디서 그리고 왜 성공하지 못하는가를 살펴보는 것이다. 좀더 자세히 알고 싶은 독지들은 인용 문헌들을 참조하기 바란다 .2)
2) Beckner(1 9 59) : Can fiel d(1 9 66) : Hemp el ( l9 65) : Munson (1971) ; Rudner(l966) : Hug h Lehman, Functi on al Ex pla -nati on in Bi ol og y, Phil o sop h y of Scie n ce, 32(1965), pp. 1-20, and Wi lliam Wi m satt , Teleolog y and the Log ica l St ru ctu re of Functi on al St at e m ents , Stu d ie s in the Hi sto r y and Phil o - sop h y of Scie n ce, 3(1972), pp. 1~8 峰 참조할 것. @ 피드백 fee dback. 어떤 일련의 반웅 또는 과정에서 결과가 원인 에 작용하여 그 반응 또는 과정 전체의 양이나 속도 등을 변화시 키는 현상을 말한다. 되돌려먹임이라고도 말한다. 그리고 결과가 원인의 크기를 감소시키는 방향으로 작용하는 것을 음(陰, nega - ti ve) 의 피드백이라 하고, 그 반대로 결과가 원인의 크기를 중가시 키는 것을 양(陽 p os iti ve) 의 피드백이라고 한다. 생물체 내에서의 거의 모든 반응은 음의 피드백에 의하여 항상성이 유지되고 있다
고 할 수 있다. 예를 들어, 혈액 속의 포도당의 양(혈당량)이 높아 지면(원인) 인슐린의 분비가 많아지고(결과), 이 인슐린이 혈당량 을 줄이게 된다. 이에 따라 혈당량이 낮아지면 이번에는 인슐린의 분비가 줄어들어 혈당량이 다시 상승하게 된다. 이와 같은 음의 피 드백을 통하여 혈당량은 항상 어떤 일정량(대개 혈액 100cm3 당 100mg 정도)을 유지하게 된다.
목적론과 비목적론을 구분하는 방법 중의 하나는 목적론적 시 스템이 추구하고 도달하고 혹은 유지하고자 하는 어떤 선호상태 룰 빈도로 표시하는 것이다. 이러한 분석에서 목표지향적인 시스 뎀으로서의 필수 조건은 그 시스템의 어떤 상태, 죽 목표상태가 다른 상태에 비하여 충분히 높은 빈도로 일어니는· 것이다. 이런 식으로 비교를 하기 위해서는 시스템의 요소들을 모두 파악해야 하고, 그러한 요소들의 특성을 규정해야 하며, 그리고 이들로부터 또한 일단의 법칙이 마련되어야 한다. 그렇게 되면, 그러한 법칙 들로부터 요소들 사이의 상호 관계나 시스템의 연속상태를 추론 할 수 있을 것이다. 예를 들면, 태양계를 구성하고 있는 요소는 태양과 9 개의 행성들인데, 이들 각각의 천체는 시스템의 상태변수 로 작용할 수 있는 무수한 종류의 특성들에 의해 서술될 수 있 다. 이들 중에서 천체역학에 적합한 두 개의 특성을 든다면 위치 와 운동량이라고 할 수 있다. 어느 특정한 시간의 태양계의 상태 를 서술하려면 태양계에 속하는 천체 각각의 위치와 운동량이 그 주어전 시간에 어떤 상태에 있는가를 구체적으로 설명하는 1~J - 의 문장을 만들면 된다. 그렇게 만들어전 법칙은 천체역학 법칙의 한부분이 될 것이다. 천체역학의 법칙은 시스템의 상태, 죽 가능한 상태들을 명시하 는 것이다. 문제는 태양계의 목표상태라고 이름 붙일 수 있을 만 큼 충분한 빈도로 일어나는 어떤 한 상태 혹은 일련의 상태를 발
견할 수 있겠는가 하는 것이다. 가령 모든 천체가 일직선 상에 배열되는 경우를 목표상태로 선택했다면 그러한 목표상태가 일어 나는 경우는 대단히 드물 것이고, 따라서 태양계는 목적론적이라 고 할 수 없을 것이다. 어느 두 행성이 태양과 일직선 상에 놓이 는 상태를 목표상태로 택한다면 그러한 목표상태는 전자보다는 빈번히 일어나긴 하겠지만 태양계를 목적론적인 시스템이라고 할 수 있을 만큼 충분한 빈도로 일어나지는 않을 것이다. 태양계가 목적론적인 시스템이라고 할 수 있을 만큼 충분한 빈도로 반복될 수 있는 목표상태가 되는 유일한 방법은 태양계의 목표상태를 천 체역학 법칙에 어긋나지 않는 어떤 천체배치(天體配置)라고 묘사 하는 길뿐이며, 이러한 정의에 의한다면 태양계는 항상 목표상태 에 있게 될 것이다. 목적론적 시스템과 비목적론적 시스템을 구분하는 또 다른 방 법은 두 시스템의 구조, 즉 인과적 피드백 고리를 촉진시킬 수 있는 장치가 있는가를 고려해 보는 것이다. 앞에서 살펴본 바와 같이 인과적 순서들은 사슬의 형태보다는 나무 모양 혹은 그물의 형태로 서술되는 것이 더 적절하다. 이러한 인과적 그물망 가운데 는 폐쇄된 고리를 포함하고 있는 것도 있다. 죽 A 라는 사건이 B 형을 포함한 여러 가지 결과를 초래하고, 이 B 형의 사건은 C 를 포함한 여러 결과를 초래하며, 다시 C 는 또 다른 여러 결과들을 초래하는데 이 속에는 A 도 약간 변형된 형태로 포함되는 경우키 다 . 다시 말하면 어떤 과정에 의해 나타나는 결과 중의 일부분이 다시 원인으로 재순환하는 것이다. 이런 경우 양성 피드백에서는 A7} 증가되고 반대로 음성 피드백에서는 A7} 감소되는 것인데, 일반적으로 사람들은 그 결과가 만족스럽지 못할 경우 양성 피드 백을 〈악순환〉이라고 부른다. 예를 들면, 어떤 처녀가 너무 뚱뚱
해서 그 나이 또래의 남자들에게 매력적이지 못한 존재로 여겨진 다. 이러한 일이 그녀를 불행한 기분으로 만들고 그래서 그녀는 더 많아 먹게 되고 그래서 더욱 뚱뚱해지고 남자들은 더욱 그녀 를 싫어하게 되는 따위이다. 음성 피드백의 예는 대단히 혼하다. 어떤 교사가 수줍읍 을 많이 터는 학생에게 발표를 해보도록 권유 하는데 그렇게 해서 그 학생이 토론을 독점하는· 정도에까지 이르 게 되면 다시 그가 토론에 지나치게 참여히는· 것을 막게 되는 것 과 같다. 제 1 장에서 논의했던 오페론 o p eron 모델도 음성 피드백 의 아주 좋은 예라고 할 수 있다. 표면적으로 볼 때 음성 피드백의 개념은 놀랄 만큼 분명한 것 같지만 이 음성 피드백을 일상의 인과적 그물망과 구분이 되도록 정의하기란 극히 어려운 일이다• 예를 들어, 음성 피드백에 의해 목적론적 시스뎀을 정의하는 네겔 Na g el 의 방법은 무엇을 목표상 태로 간주할 것인지에 대한 사전 지식에 달려 있는데 (Munson, 1971), 이것은 대략 다음과 같이 표현될 수 있다. 만일 시간 T 동안에, 일정한 범주 내에서 시스템 S 또는 환경 E 의 일차적 변이가 적응적 변이 adap tive var i a ti on 를 동반하게 된다면 그런 경우에 한하여, 그러한 시스템 없큰 목표 G 에 대하 여 목표지향적이라고 할 수 있다. 이 경우, 필요한 용어는 〈일차적 변이〉와 〈적응적 변이〉이다. 네겔은 목표상태의 정의부터 시작하는데, 그에 따르면 어떤 시스 뎀이 G 라는 성질을 가지고 있을 때에 한하여, 혹은 일련의 변화 가 S 내에서 순차적으로 일어나고 그 결과 일정한 시간이 흐른 후 시스템 S7t 성질 G 를 소유하게 되는 경우에 한해서 시스뎀은
어떤 특정한 시간에 G 상태에 있게 된다. 예를 들어, 온도가 72°F 이거나 , 지금은 그렇지 않더라도 난방시설이 작동을- 하고 있다면 온도는 조만간 72 ° F 에 도달하게 될 것이다. 일차적 변이라는 것은 시스템 내에 존재하는 하나 혹은 그 이상의 상태변수들 사이의 변화인데, 만일 이 변화가 시스템 내의 다른 상태변수의 변화를 수반하지 않는다면 그 시스템은 G 상태에서 멀어지게 된다. 예를 들어, 만일 부엌의 오븐이 켜졌는데도 자동 온도조절 장치가 냉각 장치를 가동시키지 않는다면 실내 온도는 아주 높아지게 될 것이 다. 그러나 이 일차적인 변화에 뒤이어 일어나는 또 다론 변화들 에 의해 결국 일차적 변화는 수정되고 따라서 그 시스템은 G 상 태로 복귀하게 되는데, 이때의 변화가 적응적 변이인 것이다. 어 뢰 모델에도 이런 식의 이야기를 적용해 볼 수 있다 . 선호상태와 음성 피드백-반론들 앞에서 우리는 목적론적인 시스템을 비목적론적인 것과 구분하 기 위해 제시된 두 개의 기준을 살펴보았다. 그 둘 모두 적절한 것이기는 하지만 그 중 어느 것도 필수적이거나 충분하지는 않다. 어떤 선호상태에 도달하거나 그것을 유지하는 일은 목적론적인 시스템이 되는 데 필수적인 것은 아니며, 때로는 분명히 목적론적 인 시스템인데도 불구하고 선호상태를 유지하지 않는 경우도 있 다. 예를 들면, 온도조철 모델에서 시스템은 선호상태를 유지하는 것이 아니라 선호상태 주변에서 오르락내리락 하며 맴돌 뿐인 것 이다. 또 어떤 경우에는 결코 선호상태에 도달하지도 못하고 유지 되지도 못하며, 대신 주기적으로 혹은 점차적으로 그러한 선호상
태를 향해 접근해 갇 뿐인 경우도 있다. 예를 들어, 어떤 종들은 다른 종들에 비해 좀더 완벽하게 환경에 적응하기도 하고 또 그 러한 완벽한 상태에 더욱 빈번히 접근해가기도 하지만 어떤 종이 든 결코 완벽하게 적응할 수는 없다 . 때로는 목적론적인 시스템이 그 목표하는 바에 도달하는 경우가 있기도 하지만, 하나의 시스템 에서 그것은 단지 한 번 있는 일일 뿐이다. 또 목적론적인 시스 템은 그 상태가 완벽하지 않아도 목적론적이 될 수도 있다. 애초 에 목표에 도달하도록 계획된 시스템일지라도 그 체제 내의 작은 결점, 죽 목표오류g oal fai lure 라는 것 때문에 목표에 도달하지 못 하고 계속 실패하는 경우도 있다. 따라서 설혹 어뢰가 유도 장치 의 결함으로 인해 목표물인 선박을 그냥 지나쳐 버리는 일이 생 기더라도 어뢰는 여전히 목적론적인 시스템인 것이다. 목적론적 시스템의 선호상태에 관한 반론들 가운데 이 마지막 것은 그리 만만치가 않다. 이것은 목적론적 시스템에 대한 분석을 목표상태의 유지나 도달뿐만이 아니라 목표상태 부근을 맴돌거나 그에 가까이 접근하는 것까지 다 포함하도록 확장시킬 수가 있고, 또한 한 시스뎀울 목적론적이라고 칭하는 데 필요한 빈도의 문제 는 각각의 어뢰를 하나의 집합으로 취급함으로써 구제될 수가 있 다는 것이다. 어뢰 하나하나는 단 한 번만 그 목표에 도달할 수 가 있다. 그러나 어뢰를 하나의 집합으로 본다면 무작위적으로 발 사하거나 옛날 방식으로 목표물을 겨냥했을 때보다 지금은 훨씬 더 높은 빈도로 목표물을 맞추게 될 것이 분명하다. 그러나 그렇 다고 하더라도 이러한 목표오류의 예들은 선호상태에 무엇인가가 빠져 있다는 것을 분명히 지적해 주고 있다. 청색아 (blue baby ) 의 경우, 비록 생존력이 없다 할지라도 그 아이는 여전히 목적론 적인 시스템이다. 이렇게 하나의 시스템이 그 목표상태에 도달할
수도 유지할 수도 없고 심지어 목표상태 주변을 맴돌 수도 접근 할 수도 없는 경우에도 우리가 그 시스템을 목적론적인 것으로 규정할 수 있는 것을 보면, 목적론적인 시스템에는 선호상태에 의 한 정의가 시사하는 것 이상의 무엇이 있음을 알 수 있다. 목표상태를 가지고 목적론적인 시스템을 정의하는 데 있어서의 가장 큰 결점은 시스템을 목표상태에 도달하도록 만드는 기작에 대한 언급이 없다는 점이다. 인과적 피드백 고리에 의한 정의는 바로 이러한 기작들의 일반적인 성질을 구체화하려는 시도라고 할 수 있다. 음성 피드백 고리에 의해 분석을 하는· 데 있어 가장 적응이 안 되는 목적론적 시스템의 형태를 두 가지 든다면, 그 중 하나는 목적지향적(目的指向的)이 아닌 인공물이 어떤 행위자 에 의하여 사용되는 경우이며, 다른 한 가지는 목적지향적이긴 하 지만 음성 피드백에 의한 것이 아닌 경우를 들 수 있을 것이다. 망치는 못을 박거나 못을 빼는 등의 특정한 기능을 수행하도록 만들어진 것이지만 때로는 그 의의 다른 일들을 위해 쓰이기도 한다(창문이 닫히지 않도록 받쳐 놓거나, 살인을 할 때 등). 그러 나 전혀 사용이 불가능한 경우도 있다(피임 기구로서 혹은 해부 도구로서 등). 어떤 일을 하는 데 그것을 쓸 수 있느냐 없느냐를 결정하는 것은 그 물체의 물리적 특성이지만, 사용 가능한 범위 내에서는 사용자의 의지에 달려 있다. 비목적지향적 인공물을 사 용하는 경우를 음성 피드백 모델에 이용하려면 행위자가 못을 박 기 위해 망치를 사용하는 일 같은 일시적 상황은 목적지향적 시 스뎀으로 간주해야 한다. 이러한 시스템에서 만일 우리가 정신적 실체에 대한 언급을 하지 않는다면, 이 시스템이 가지고 있는 목 적지향적 특칭은 (행위자의) 신경계에 의해서 이루어지는 것이라 고 바꾸어 말할 수 있을 것이다.
어떤 선호상태들은 음성 피드백이 아닌 것처럼 보이는(적어도 처음에는) 기작에 의해서 유도되기도 한다. 다른 종류의 경로를 사용하는 것도 그 한 예가 될 수 있다. 콩팥이 과다한 수분을 체 내에서 제거하지 못하면 땀으로 그것을 해결할 수 있는데, 이러한 시스템에서 나타나는 효과는 특정한 선호상태 또는 일련의 선호 상태들을 유지하는 것이다. 그러나 이러한 시스템은 음성 피드백 의 일반적 정의나 그것이 담고 있는 정신에 들어맞지가 않는다. 그러므로 그러한 정의는 너무 경직된 것이라 볼 수 있다. 그러면 서 또 너무 포괄적이다. 왜냐하면 적어도 처음에는 목적론적인 것 처럼 보이지 않는 여러 형태의 시스템에도 적용되기 때문이다. 그 러한 예로, 정지 상태에 있는 복합전자(複合振子, comp o und pe n-dulum), 탄성을 지닌 고체, 전도체를 흐르는 일정한 전류, 그리고 열역학적으로 평형상태에 있는 화학계 등을 둘 수 있다. 음성 피 드백 모델은 어떤 시스템이 그 선호상태로 되돌아가는· 경우뿐 아 니라 사전에 규정된 일련의 선호상태를 순차적으로 거치면서 앞 으로 진행되는 경우에도 적용될 수 있도록 고안되어 있다. 그러나 후자의 경우 프로그램, 죽 그러한 규칙적인 순서를 거치게 하는 원인에 관한 언급은 없다. 그렇기 때문에 이러한 방식을 통해 목적론적 시스템을 정의하 고자 시도할 때에는 반대되는 사례들까지도 수용할 수 있도록 본 래의 정의에다 몇 개의 조항을 첨가하고 제약을 가하는 일이 타 당하고 일반적인 철차이다. 이렇게 수정된 몇몇 정의들은 목적론 적 시스템의 직관적 개념을 상당히 비슷하게 그려낼 수가 있다. 예를 들면, 선호상태에 대한 기준이나 음성 피드백에 의한 분석 그 어느 것도 그 하나만 가지고서는 충분할 만큼 정확하고 명확 하게 희망하는 바를 만들어낼 수가 없지만, 그 둘을 함께 사용하
면 요술처럼 성공할 수가 있는데, 하나의 기준으로 볼 때는 그 한계를 정하기 어려운 경우도 또 다른 기준으로 보면 상대적으로 쉽게 처리될 수가 있기 때문이다. 만일 시스템의 어떤 특정한 상 태가 분명히 목표상태에 해당한다면 어떤 인과적 기작이건 그 시 스템을 유도하는 것은 모두 음성 피드백의 예로 간주될 수 있다 . 이러한 방법이 성공한다고 한들 그 시도가 지닌 중요성은 그럼 무엇인가? 우리는 아직도 이러한 분석이 과학철학에 무언가 기여 하고 있으며 결코 언어적 분석을 위한 연습이 아님을 증명해야 하는 것이다. 위에서 이야기한 분석이 생물철학이 되기 위해서는 이러한 논리적 분석과 현대 생물학 이론 사이의 어떤 연관성을 보여주어야만 한다. 목적론적 시스템의 개념을 정의하기 위해 제 시되었던 기준 중에서 두번째 쌍이 지닌 강점이 바로 그것이 현 대 생물학 이론에 기초하고 있고 동시에 현대 생물학 이론에 기 여하고 있다는점이다. 프로그램과 선택 과정 앞에서 논의한 분석 중 그 어느 것도 일반적인 의미에서전 제 한된 의미에서건 목적론적인 시스템의 개념을 올바르게 나타내지 못하고 있다. 두 가지 기준 모두 너무 엄격해서 목적론적으로 보 이는 것들까지도 제의시키고 있는 한편, 또 너무 허술해서 목적론 적 시스템이 아닌 것까지 포함시키기도 한다. 목적론적 시스템과 비목적론적 시스템에 대한 경계선이 모호하기는 하지만 그러한 모호함이 태양계나 정지 상태에 있는 복합전자에까지 확장되는 것은 아니며, 무엇보다 중요한 것은 기준 그 자체에 대한 모호함
이다. 어떻게 목표상태를 시스템 내의 다른 상태와 구별할 수 있 는가? 어떻게 음성 피드백 고리를 일상적인 인과적 그물망과 구 별할 수 있는가? 이 두 가지 분석법이 지닌 결점은 더 깊은 분석 울 위한 어떤 기반도 제공하지 못한다는 점이다. 왜 신축성이 있 는 고체를 목적론적인 시스템이라는 집합에서 제의시키며, 왜 날 아가는 나비는 그에 포함시키는 것인가? 목적론적인 시스템을 정의하기 위해 제시되었던 두번째의 기준 쌍이 지닌 중요한 장점은 이들이 과학 이론 특히 전화론, 학습이 론, 그리고 정보이론에 근거를 두고 있다는 점이다. 전화론이 목 적론에 대한 새로운 분석에 있어 중심 역할을 해왔다는- 것은 그 리 놀랄 만한 일은 아니다. 왜냐하면 1 양1 l 기를 지배했던 목적론에 치명타를 가한 것도 역시 전화론이었기 때문이다. 이에 비해 학습 이론과 정보이론의 역할은 다소 모호한 점이 없지 않다. 생물이 어떻게 학습을 하는가 하는 근본적인 문제에 대한 연구는 아직은 대단히 초보적인 단계에 있는 터라 이들 이론에 바탕을 둔 분석 들이 모호한 것은 당연한 일일 것이다. 또한 정보이론은 경험적인 이론이 아니며 수학의 한 부분인 형식이론이다. 따라서 정보이론 은 기껏해야 학습과 전화의 기작을 연결시키는 형식적인 유추로 사용될 수 있을뿐이다. 메이어 (Ernst Mayr ; Blackburn, 1966 및 Munson, 1971 에서 ) 는 목적론적 시스템의 구조적 정의로서 프로그램의 존재를 주장 하였다. 그는 〈컴퓨터 언어를 쓴다면 소위 ‘프로그램'된 개체는 목적하는 바에 따라 행동할 수 있다〉고 주장하면서, 〈합목적적 t eleono mi c 이라는 용어는 정보를 암호화한 것, 죽 프로그램을 바 탕으로 하는 시스템에만 엄격하게 국한시켜야 한다〉고 결론지었 다. 생명체가 생명체로서 기능을 하는 데 필수적인 정보는 DNA
또는 RNA 에 암호화되어 있고 이러한 정보는 세대에서 세대로 전달될 수 있다. 그러나 목적이 있는 행동의 경우 프로그램된 것 은 신경계(혹은 이와 비슷한 시스템)이며, 신경계에 축적된 정보 는 세대에서 세대로 직접 전달될 수가 없기 때문에 매 세대는 이 러한 정보를 새로이 획득해야만 한다. 인공물의 기능도 그 내부에 들어 있는 프로그램(예를 들어 자동 온도조철 장치에 들어 있는 바이메탈) 자체에 의해서건 또는 그것을 만든 사람이나 사용하는 사람에 의해서건 이러한 패러다임에 속한다고 볼 수 있다. 물론 다음으로 메이어가 부딪친 문제는 프로그램의 개념을 정 의하는 일이다. 인용부호가 있음으로 해서 그 문구가 인용문임을 알 수 있듯이, 메이어는 정보이론 언어의 폭넓은 성공을 통해서 자신이 어느 정도 정당화되어 있는 그러한 유추를 다루고 있다는 것을 알고 있었다. 그러나 그의 분석을 만족스럽게 만들기 위해서 는 이러한 유추 이상의 어떤 것이 필요하기 때문에 메이어는 현 재도 〈 프로그램〉에 대한 좀더 적절한 정의를 제시하기 위해 노력 하고 있다. 그는 목적론의 개념이 시스템보다는 과정이라는 용어 로 훨씬 잘 설명될 수 있다고 생각하고 있다. 그러나 그의 분석 이 우리에게 주는 가장 큰 중요성은 그가 목적론적인 시스템과 비목적론적인 시스템(혹은 과정)을 구분하기 위해 프로그램에 의 존했다는 사실이 아니라 그의 분석이 선택이론에 의존했다는 데 있다고생각된다. 윔사트W illi am W i msa tt는 목적론적인 시스템에 대한 역사적 정의를 논하면서, 시스템이 목적론의 범주에 속하기 위해 필요한 사항들은 직접적이건 간접적이건 선택의 과정을 통해 형성되어야 한다고 주장하고 있다. 선택 과정에는 어떤 기작이 있어서 이것에 의해 항목it em 이 서로 구분되고: 또 구분된 항목들 중에서도 선
택 여부가 결정되고, 그렇게 선택된 항목들이 시스템의 구성원으 로 통합된다. 생물체가 복제, 돌연변이, 자연선택 등을 통하여 진 화하게 되는 것처럼 목적을 가전 행위지는 - 학습과정이나 선택을 받아들이는 과정, 자극을 계속 보유하는 과정, 그리고 그렇게 행 위자 내에 보유하게 된 자극을 필요에 따라 다시 새로이 유도하 는 과정 등을 통해서 의도적인 행동을- 나타내게 된다(한 가설에 의하면, DNA 는 학습에 대한 중요한 화학적 기초가 된다. 예를 들 어 기억이란 것은 DNA, RNA 그리고 단백질로 이루어전 복잡한 상관 관계의 결과라는 것이다). 윔사트의 분석은 두 가지 문제에 부딪치게 되는데, 첫째 선택 과정에 대한 적절한 분석을 제시하는 일이며, 둘째 〈 목적론적인 시스뎀〉을 역사적 개념을 가지고 해석한 그의 방법에 정당성을 부여하는 일이다. 아무리 보아도 그의 분석을 수용하려면 최소한 목적론적 시스템을 지금과 같이 사용해서는 안 될 것 같고 거기 에 어떤 수정이 가해져야 한다고 생각된다 . 왜냐하면 현재 우리가 사용하고 있는 목적론적인 시스템이란 그 기원에 관계없이 인정 되고 있는 것이기 때문이다. 사실 우리가 목적론적이라 여겨왔던 모든 시스템들은 직접적으로 혹은 간접적으로 결국 선택의 과정 을 동해 생긴 것이다. 생명체는 자연선택을 통하여 형성되었고, 행위자는 자국을 선택적으로 보유하고 체계화함으로써 의식을 지 닌 존재가 되었다. 선택 과정에 의해 직접 만들어전 인공물은 거 의 없다고 하지만, 결국 그들은 그러한 선택 과정을 거친 존재에 의해서 만들어지는 것이다. 따라서 인공물들은 〈간접적으로 〉 선 택 과정을 통해서 생겨난 것이라고 볼 수 있는 것이다 .3 )
3) 나는 윔사트 W illi am W i msa tt와 윌리엄스 Mar y B. W illiam s 두 사람과의 대화를 통하여 목적론적 체계에 대한 이해를 깊이 할 수 가있었다.
목적론과 선택이론 목적론적 시스템을 정의하기 위해 제시된 4 개의 기준들은 이런 저런 면에서 모두 적합하다고 볼 수 있다. 각 기준 하나하나에는 다소 모호한 면이 있지만 이 4 개의 기준들을 함께 사용하면 그러 한 모순점들이 해결된다. 그러나 우리의 목적하는 바에 있어서는, 목적론적 시스뎀의 특칭을 정의하는 이러한 기준들이 궁극적으로 선택설에 의존한다는 데 그 중요성이 있다. 다론 시스템과는 달리 목적론적 시스템에는 일련의 법칙들이 있어서 이 법칙들에 의해 선호상태가 점점 더 좁은 범위로 국한되어 간다. 메이어가 도입 한 용어를 사용하면 (Blackburn, 196~ Munson, 1971) 목적론적 시스템에는 일련의 근접 원인p rox i ma t e cause 과 궁극 원인 ul ti mate cause 이 존재한다. 가장 근접적인 원인과 관련된 법칙들은 시스템의 상태를 극히 조금밖에 제한하지 않기 때문에 이러한 법 칙들만 주어졌을 때는 시스템은 넓은 범위의 상태에 존재할 수 있게 된다. 그리고 궁극 원인에 대한 법칙이 첨가될수록 이 시스 템의 상태는 점점 좁은 범위로 국한된다. 생물에 있어서 가장 궁 극적인 법칙은 전화론이어서, 이것이 시스템의 가능한 상태를 크 게 제한하고 있다. 다른 말로 하자면 이 법칙들이 결국 시스템의 목표상태를 결정하는 것이라고 할 수 있다. 예를 들면 생리학은 생물체가 필요로 하는 아주 넓은 범위를 다룬다. 그러나 현재보다 훨씬 많은 종류의 생물이 과거에 존재했 울 수도 있고 또 현존하는 생물의 어떤 특칭둘은 생리학의 법칙 만으로는 설명이 불가능한 것도 있다. 하지만 보다 궁극적인 전화 론의 법칙들은 현존 생물이 선택되어 온 과정을 설명할 수 있을 뿐 아니라 그들의 생리적 특성까지도 설명할 수 있다. 사실 전화
론을 수용하기 이전에는 구조와 행동의 주요 목적은 개체의 유지 였고, 종의 유지는 자동적으로 그에 따라오는 것이라고 생각하였 다. 그러나 전화론을 수용하게 된 이후, 개체는 부분적으로 서로 상충되는 목표, 즉 개체의 이익과 종의 이익 사이의 타협의 결과 로 인식하게 되었다. 언뜻 보기에는 개체를 희생하여 종의 생존을 증진시킨다는 속 성은 이상한 일처럼 보인다. 그러한 속성은 단 한 세대 동안에 방대한 양의 게놈 재구성을 할 수는 없다는 것과 각 개체는 생식 가능했던 그 조상들의 산물이라는 서실에 기인한다. 개체는 생존 되어야 하겠지만 각 개체는 또한 과거에 이루어전 타협의 결과이 기도 하다는 것이다. 특정 포유류가 가지고 있는 유선(乳脈)은 생 존에 필수적인 것이 아니며 오히려 해가 되기도 한다. 그러나 이 포유동물이 이렇게 생존할 수 있었던 것은 그 어미가 유선을 지 니고 있었기 때문이다. 유선을 만드는 유전자가 없다면 그 특정 개체에게는 이득이 될지 모르지만, 동시에 그 가계는 멸망될 수밖 에 없다는 것, 그리고 그와 함께 그런 유전형질도 소실될 것임은 분명하다. 유전자 구성 속에는 각 개체 자신의 생존에 필요한 유 전자뿐만 아니라 대의 계승에 필요한 유전자들도 들어 있는 것이 다. 개체가 이러한 속성들을 가지고 있기 때문에 그 자손들의 생 존 능력이 다소 감소되는 한이 있어도 이러한 속성들이 그대로 유전되어 대를 이어 가는 것이다 . 생식의 대가는 크고 때로 죽음 울 초래하기도 하지만 그것은 모든 생명체들이 지불해야만 하는 대가이다. 또한 아러한 유전적 속성은 흔적기관(지금은 아무 기능 도 하지 않지만 그 선조대에서는 기능했던 기관)이 계속 유전되는 이유이기도하다. 이 철에서 우리는 프로그램에 의해 직 • 간접으로 지배되는 인
과적 피드백 고리, 특히 음성 피드백 고리에 의해 야기된, 그리고 매우 중요하면서 좁은 의미로 정의된 목표상태가 목적론적 시스 템의 중요한 특징이라고 하였다. 그러나 보다 중요한 것은 이러한 시스템을 지배하는 법칙들 사이에 위계질서가 존재한다는 것이다. 목적론적 시스템은 직접 혹은 간접적으로 선택 과정을 통하여 일 어나지만 우리의 분석에 있어 선택 과정이 특히 중요한 이유는 가장 좁은 의미에서의 목표상태를 결정하는 가장 궁극적인 이론 이 바로 선택이론이라는 점이다. 비목적론적인 시스템의 경우에는 그러한 법칙의 위계가 존재하지 않는다. 예를 들면, 태양계의 위 치를 지배하는 것은 한 세트에 속하는 일련의 법칙들, 죽 천체역 학 법칙뿐이며 단순한 기하학적 고려는 태양계의 선호상태를 정 의하는 데 불필요하다. 아마도 한 천체의 위치를 알면 다른 천체 의 위치까지도 저절로 알 수 있을 것이다. 그러나 목적론적 시스 렘에서는 선택이론이 시스템의 선호상태에 대한 한계를 결정하며, 이러한 선호상태를 아주 좁은 범위로 국한시키는 법칙들은 그 시 스템의 상태를 보다 폭넓게 규정하는 하위의 법칙들과는 최소한 부분적으로나마 독립되어 있다. 목적론적 서술과 포괄법칙 모델에 의한 설명 앞에서의 논의는 주로 목적론적 시스텐에 관한 것이었지만, 이 제 목적론적 서술과 목적론적 설명을 분석해야 할 때가 되었다. 목적론적 언어와 비목적론적 언어 사이의 관계를 연구하는 데 좋 은 방법 중 하나는 목적론적 서술을 일상적인 인과적 서술과 비 교해 보는 것이다. 여러 과학 논문 속에서 우리는 양쪽 서술 모
두를 쉽게 찾아볼 수가 있는데, 양쪽 서술에서 나타나는 차이는 그 서술의 분석 부분에서 자세히 나타내고자 하는 느낌상의 차이 와 대략 일치될 것이다. 그리고 계속해서 목적론적 서술과 비목적 론적 서술의 사용을 비형식적인 설명 가운데서 바교해 볼 수 있 을 것이고, 마지막으로 목적론적 서술과 설명들을 보다 형식적으 로 분석하고, 인과적 서술과 설명들도 이와 같은 방법으로 분석하 여 서로 비교해 볼 수 있을 것이다. 그러나 보통은 목적론적 서 술과 목적론적 설명을 동일시하고 그런 다음 포괄법칙 모델의 설 명 기준에 따라 이러한 설명의 개요를 판단하도록 해 왔었다. 예 롤 들자면 네겔 (Na g el, in Munson, 1971 참조)과 행펠 (1965) 은 다같이 목적론적 서술을 생략논증 el lipti cal ar g umen ts 으로 간주 하였는데, 네겔은 기능과 목표를 필요조건들과 동등하게 놓는 단 순한 방법에 의해 목적론적 서술이 포괄법칙 모델의 요구 조건을 충족시킬 수 있는 논증으로 전환될 수 있다고 생각했으나, 행펠은 그러한 방법은 실패할 것이라고 하였다. 그는 기능적 설명 속에 나타나는 전형적인 정보들이 연역적으로든 귀납적으로든 문제시 되는 구조의 존재를 설명하는 데 필요한 근거를 제공하지 못한다 고결론지었다. 이 철학자들이 범한 오류는 목적론적인 서술이나 설명들이 내 포하고 있는 내용을 밝히려 했다는 데 있는 것도 아니고, 이러한 형식화가 포괄법칙 모델을 따라야 한다는 무언의 가정에 있는 것 도 아니다. 잘못은 목적론적 설명들과 포괄법칙 모델 사이의 모든 불일치가 목적론적 형식화가 목적론적이라는 것에서 기인한다고 생각히는 데 있다. 목적론적 설명과 포괄법칙 모델 사이에 너무나 큰 차이가 있는 것은 사실이지만 그러한 차이는 설명이 목적론적 이라는 사실과는 아무 상관이 없다. 많은 경우에 있어 일상적인
인과적 설명들은 포괄법칙 모델과 다른 점이 있는데, 예를 들 어, 〈 포유류가 땀을 홀리는 까닭은 무엇인가? 〉 라고 묻는다면, 그 포유류 주변 환경의 온도가 높기 때문이라는 것이 한 가지 대답 이 될 수 있다. 또 〈 포유류가 땀을 홀립으로써 어떤 기능을 하는 가? 〉 라고 물을 수도 있으며, 체온을 일정하게 유지시킨다는 것이 한 가지 답이 될 수 있을 것이다. 두 질문 다 적절하고 그 답도 정확하며, 여러 전후 관계로 볼 때 더 이상의 답을 제시해야 할 필요도 없다. 그러나 포괄법칙 모델의 입장에서 보면 이들 답은 설명의 개략에 지나지 않으며 따라서 무언가를 더 채워넣어야 할 필요가 있다. 그러나 채워넣는 과정은 양쪽 질문 모두의 경우에 있어 상당히 어려운 일이며 종종 핵심에서 벗어나기가 쉽다. 목적론적 서술을 배제하는 가장 쉬운 방법은 기능, 목표, 목적 둘을 결과와 동일시하는 것이고, 그리고 나서 그 결과를 필요조건 과, 원인을 충분조건과 동일시하는 것이다. 만일 우리가 이렇게 동일시하는 것을 받아들인다면 다음 세 가지 전술은 모두 같은 것이 된다. (1) A 의 기능은 B 이다. (2) A 의 결과는 B 이다 . (3) B 의 원인은 A 이다. 그러나 이들 세 가지를 동일시하는 것에 대한 반론도 제기되고 있다. 이제 결과를 필요조건과 그리고 원인을 충분조건과 동일시 하는 것에 대한 반론부터 살펴보기로 한다. 먼저 이러한 반론들이 목적론적 논술 그 자체와는 아무런 상관이 없다는 것에 유의해야 한다. 오히려 이런 반대 의견들은 목적론적 시스템에만 국한된 특
수 상황이라고 할 수 없는 여러 원인과 결과들이 있다는 데서 비 롯된다. 다음으로 우리는 기능과 결과를 동일시하는 것에 대한 반 론들을 다루게 되는데 이들은 양성적 기능p os iti ve fu nc ti on 에 그 근거를두고 있다. 기능적 등가성 (機能的 等價性, fun cti on al equ iv a lents ) , 죽 같은 기능을 수행할 수 있는 방법에 여러 가지가 있다는 것을 고려하 지 않고는 어느 누구도 목적론적 서술을 논의할 수 없다. 수액이 나무의 윗부분으로 빨려 울라가고 토끼들이 배란을 하고 또 동일 한 표현형적 특칭들이 생성되는 데는 수많은 방법들이 있다. 그러 므로 기능들이 단순히 필요조건만으로 설명될 수는 없다. 또한 이 러한 기능은 여러 가지 원인이 존재하기 때문에 생겨나는 것이지 만 그것이 목적론적 시스템에만 국한된 특별한 것은 아니다. 또한 여러 학자들은 목표실패, 죽 목적론적 시스템이 그 목표에 도달하 지 못하고 실패하는 일이 있다는 것에도 주목해 왔다. 포유류에 있어서 심장의 기능은 혈액순환이다. 갓 태어난 아기의 몸에서는 심장이 제대로 그 기능을 하지 못한다 하더라도 말이다. 프로 권 두선수의 경우 시합에 지게 된다 하더라도 열심히 연습을 한다. 다시 말해서 목적론적 시스템은 완벽하지 않아도 목적론적이 될 수 있다는 것이다. 그러나 이러한 상황은 인과론적 서술에서도 비 일비재하다. 어느 특정한 구조나 행동이 특정한 기능을 수행하는 데 있어 기능적 등가성이 필요조건이 아닌 것처럼 목표실패의 존 재는 그 어느 것도 충분조건이 될 수 없다는 것을 의미하고 있 다. 죽음이란 것이 수없이 많은 여러 원인들에 의해서 비롯되는 것처럼 주식 시장에 투자하는 일도 무수히 많은 여러 가지 결과 를 가져올 수 있다. 이러한 이유로 인과론적 전술과 목적론적 전 술은 결국 같은 배를 타고 있는 셈이다.
네겔 (Munson 에서, 1971) 은 목적론적 전술과 비목적론적 전술 사이의 차이는 〈 Y 는 X 의 결과라는 이야기와 X 는 Y 의 원인 또 는 조건이라는 이야기와의 차이에 해당하는 것이고, 요약하면, 차 이라는 것은 선택상의 문제이다〉라고 말하였다. 만일 우리가 다 양한 원인들과 결과들이 있다는 것을 명심한다면 네겔의 이야기 에는 분명히 어떤 진리가 담겨 있다고 생각된다. 목적론적 진술은 인과적 순서의 어떤 일시적인 역전을 요구하지 않는다. 그런데도 일시적인 역전이 겉으로 나타나는 것은 단지 선택상의 고려에서 비롯되는 것이다. 일상적인 인과적 서술에서는 문제시되는 사건의 원인이 된 초기의 사건에 관심을 두는 반면 목적론적 서술에서는 문제시되는 사건이 무엇을 일으켰는지 알기 위해 나중에 일어난 사건에 관심을 둔다. 포유류가 일정하게 체온을 유지할 수 있는 것이 땀울 홀리기 때문에 생기는 결과라는 것과 땀을 홀리는 것 이 체온을 일정하게 유지시키는 원인이라고 말하는 것에는 아무 차이가 없다. 그러나 이 둘 중 어느 것도 포유류에 있어서 땀이 하는 기능은 체온을 일정하게 유지시키는 것이라는 목적론적 주 장을 대신할 수 있을 것 같지 않다. 따라서 기능과 결과를 동일 시하는 것은 반드시 옳은 일은 아닐 것이다. 모든 기능은 결과임 에 틀림없지만 모든 결과가 기능에 해당하는 것은 아니다. 탈수현 상은 땀을 흘린 결과로 생기는 것이지만 탈수현성이 땀울 홀리게 하는 기능은 아니기 때문이다. 마찬가지로 포유류의 경우, 심장의 박동은 혈액을 순환시키는 일을 하지만 동시에 박동소리를 만들 어내는데, 그렇다고 해서 박동소리의 생성이 십장박동의 기능은 아닌 것이다. 기능과 결과 사이에 존재하는 이 분명한 차이를 해결하는 일은 의의로 상당히 간단하다. 목적론적 시스템에서 우리는 그 가치를
평가할 수 없는 일반적인 의미로 사용된 〈 기능 〉 과 긍정적인 기능 의 의미로 사용된 〈 기능 〉 을 구별해야 한다. 일반적 의미의 경우 에는, 〈 결과 〉 가 〈 필요조건 〉 과 동일시되지 않는다면 〈 기능 〉 과 결 과는 동일한 것이다. 아렇게 그 가치에 대한 평가가 불가능한 일 반적 의미에서 본다면, 탈수는 땀을 홀리는 일의 기능(결과)이며 박동소리의 생성은 심장박동의 기능(결과)이다. 그러나 일반적으 로 〈 기능 〉 은 긍정적인 의미를 내포하고 있으며, 이는 곧 구조나 행동이 시스템의 선호상태로의 접근, 도달 또는 그 유지에 기여한 다는 것을 의미한다. 이러한 맥락에서 본다면, 구조나 행동이 역 기능적 혹은 기능이 없는 것일 수도 있는데, 만일 구조가 시스템 이 선호상태로 접근, 도달 혹은 유지하는 것을 방해한다면 그 구 조는 역기능적인 것이며, 구조나 행동이 도움도 방해도 하지 않는 다면 그것은 기능이 없는 것이다. 기능이 없는 구조나 행동이란 것은 단지 우리가 긍정적인 기능을 얼마나 세밀하게 구분하는가 에 달려 있다. 평가 내용이 세분화되면 될수록 기능이 없는 구조 나 행동은 감소될 것이다. 목적론적 설명과 설명의 포괄법칙 모델 목적론적 설명은 적어도 하나 이상의 조작적인 목적론적 전술 울 포함하고 있다. 인과론적 설명에서는 원인으로 기술되지만, 목 적론적 설명에서는 기능이나 목표로 기술되는 것이다 . 목적론적 설명을 포괄법칙 모델과 비교해 보면 세 가지 질문이 생기는데, 목적론적 설명을 그에 적합한 연역적 논증으로 바꿀 수 있는가, 아니면 정당화된 귀납적 논증으로 바꿀 수 있는가, 양쪽 어느 경
우이건 조작적인 목적론적 전술을 순수한 과학법칙으로 여길 수 있겠는가? 하는 것이 그것이다. 어떤 사람이 식물은 왜 엽록소를 가지고 있는지를 알고 싶어 한다고 하자. 어떤 이는 〈 광합성의 촉매로 쓰기 위해서〉라고 답 할 것이다. 그렇다면 광합성의 기능은 무엇인가? 식물이 살아가는 데 필요한 양분을 만드는 것이라고 답할 수 있을 것이다. 만일 이러한 기능적 전술이 필요한 연결 관계를 제대로 표현하고 있는 것이라면 이 목적론적 설명은 다음과 같이 재구성될 수 있을 것 01 다. (1) 식물에 있어서 영록소는 광합성에 필수적이다. (2) 광합성은 식물의 생존에 필수적이다. (3) 식물은 생존한다. 그러므로 식물에서 광합성이 일어나고 있다. 그러므로 식물에는 영록소가 있다. 철학문헌에 나오는 수많은 예들과 마찬가지로 이 예도 약간의 수정을 요하는데, 이는 처음의 두 전제가 톨리기 때문이다. 엽록 소가 광합성에 필수적인 것은 아니다. 죽 이와 아주 유사한 물질 울 대신 촉매로 사용할 수도 있는 것이다. 또 모든 식물이 엽록 소나 이와 유사한 물질을 가지고 있는 것도 아니다. 어떤 식물은 숙주의 살아 있는 조직에서 필요한 양분을 섭취하기도 하고(기생 식물), 또 어떤 것은 죽은 생물의 유기물질로 살아가기도 한다(부 생식물). 더욱이 엽록소를 가지고 있는 식물에서조차도 엽록소는 생존에 필수적인 것은 아니다. 왜냐하면 이들 중 일부는 빛이 없 는 곳에서는 기생성이나 부생성 식물로 변할 수가 있기 때문이다.
문제는 더욱 복잡해서 동물 가운데도 엽록소를 가지고 있는 것이 있다. 물론 문제는 같은 기능을 할 수 있는 것이 여러 개이고, 같은 결과를 가져올 수 있는 행동들이 여러 개인 기능적 등가성이 목 적론적 시스템에 산재하고 있다는 점이다. 생물을 현재와 같이 분 류하고 또 기능 위주로 분석하는 체제 아래서는 어떤 특정 그룹 에 속하는 생명체의 경우 한 가지 기능은 한 가지 이상의 여러 방법에 의해 수행될 수가 있다. 기능적 등가성을 배제하기 위하 여, 죽 목적론적 설명의 연역적 특성들을 구제하기 위하여 두 가 지 서로 엇갈리는 방법이 제안되고 있는데, 그 하나는 목적론적 시스템의 여러 부 분 늘 을 기능적으로 정의하는 것이다. 예를 들면, 베크너 Beckner4) 는 혈액의 펌평은 어떤 구조가 심장이 되도록 하 는 필요조건이라고 하였다. 앞에서 든 예에서는 엽록소를 가지고 있다는 것이 식물이 되기 위한 필요조건이 될 것이다. 따라서 겨 우살이는 식물로 간주될 수가 없다.
4) Meta p h y si c a l Presup po sit ion s and the Descrip tion of Bi o- log ica l Sy st e m s, in J. R. Gregg and F. T. C. Harris , Form and Str at e g y in Scie n ce(Dordrecht : D. Reid e l Publi sh in g Co., 1964).
베크너의 그러한 견해에 대해 두 가지 반론이 제기될 수 있는 데, 첫째는 그것이 현행 과학적 사고와 반대가 된다는 것이다. 망 치로 문을 괴어 놓았다고 해서 망치가 문을 괴는 기구인가? 단추 를 장기의 졸로 사용했다고 해서 단추가 졸이 되는가? 이러한 질 문에 대해 아무런 답을 할 수 없을 것 같지만, 과학에서는 하나 의 답을 다른 답보다 선호하게 되는 경우 그 답이 어떤 것이든간 에 그것을 택한 데는 타당한 이유들이 있다. 요즈음에는 화학반응
울 특정 촉매를 가지고 정의하지는 않는다. 그러나 실제로는 그렇 게 정의되는 반응도 많다. 마찬가지로 엽록소를 가지고 있다는 단 하나의 특성을 가지고 어떤 분류군을 정의하지는 않는다. 엽록소 뿐 아니라 어떤 특성도 그 분류군에 속하기 위한 엄격한 필요조 건이 되지는 않는다. 하지만 때때로 조건으로 요구되는 정의를 과 학에 도입해야 하는 경우가 있으며 거기에는 그럴 만한 이유들이 있다. 그러나 실제로 기능적 정의에 〈 광합성〉, 〈식물〉, 〈심장〉과 같은 용어를 사용하는 단 하나의 이유란 기능적 설명의 연역적 특성을 살리기 위한 것이다. 그러나 이 시도는 결국 실패할 것이 다. 왜냐하면 목적론적 시스템이 이런 식으로 해석되는 한 그것은 과학법칙으로 인정될 수 없을 것이기 때문이다. 대신 이러한 목적 론적 진술은 논리적으로는 전리가 된다. 만일 심장이란 혈액을 펌 프질하는 것이다라고 정의한다면 심장이 혈액을 펌프질한다는 주 장은 더 이상 경험적 서술이 될 수 없고, 따라서 목적론적 설명 은 과학적인 설명이 될 수 없을 것이다. 목적론적 설명을 연역적으로 만둘기 위한 두번째 방법은 목적 론적 시스템을 재분석하여 기능적 등가성을 제거시키는 것이다. 이렇게 한다면 특정 구조나 행동이 특정 기능에 필수적이라고 하 는 것은 우연적인 사실이 될 것이다. 이것은 앞에서 논의했던 원 인에 대한 상황과 비슷하다. 만일 인과법칙이 과정의 법칙이 된다 면 원인과 결과 사이에는 보편적 인 공동변화 covaria t i on 7} 있음 이 틀림없다. 그러나 현재 분자생물학에서 행해지는 것처럼 인과 적 나무 causal t ree 나 인과적 그물망 causal ne t work 으로 분석된 다면 이 보편적인 공동변화란 불가능하다. 항상 다수의 원인과 다 수의 결과들이 존재하기 때문이다. 하지만 인과적 나무나 그물망 의 경우, 그에 요구되는 보편적 공동변화를 얻어내기 위하여 상황
을 재분석하는 것은 가능할지 모른다. 때로 이러한 시도는 성공을 거두었다. 그러나 생물, 생물학적 종, 그리고 이와 유사한 것들이 적절한 시스템이라고 생각되는 한 이러한 방법이 생물학에서 먹 혀들지 않으리라고 믿는 데는 타당한 이유둘이 있다. 앞에서 지적 한 바와 같이 적응성을 증진시키기 위한 가장 일반적인 기작이 중복성 redundanc y이다. 죽 같은 기능을 수행하는 여러 가지의 서 로 다론 방법이 있다는 것이다. 따라서 생물계는 혼히 중복성 혹 은 우리가 기능적 등가성이라 불렀던 것들을 가지고 있는데, 만일 기능적 등가성을 제거하기 위하여 연구하고자 하는 현상들을 재 분석한다면, 이러한 분석 아래서는 생명체를 시스템으로 취급할 수가 없게 될 것이기 때문이다. 목적론적 시스템은 그 자체를 우리에게 강요하고 있는 것처럼 보인다. 만일 어떤 것인가를 개체로 간주해야 한다면 분명히 생물 체가 그러한 개체가 되어야 한다. 특별한 교육을 받지 못한 원주 민들이 훈련된 박물학자들처럼 쉽게 생물학적 종을 구별하는 것 을 보면 생물학적 종은 거의 명백한 존재로 생각된다. 목적론적 시스템은 보통 열려 있는 상태, 죽 개방계여서 끊임없이 조금씩은 변하고 있으나 항상 같은 상태로 있는 것처럼 보인다. 따라서 목 적론적 시스템에는 불변성이 있다고 말할 수 있다. 그러나 이러한 불변성은 실제 변화가 없음으로써 같은 상태를 유지하는 피라미 드의 불변성과는 다른 것이다. 목적론적 시스템은 변화에 의해서 지속되는 것이다. 과학자들은 태양계와 같이 철저히 폐쇄되어 있 는 시스템을 다루는 데는 성공했다. 이러한 시스템은 잘 폐쇄되어 있기 때문에 시스템의 행동을 지배할 수 있는 과정의 법칙을 만 들어 낼 수가 있다. 언뜻 보면 목적론적 시스템도 실제보다 완벽 하게 폐쇄되어 있는 것처럼 보인다. 왜냐하면 폐쇄계와 마찬가지
로 이 시스템도 그 본래의 일정한 모습을 유지하고 있는 것처럼 보이기 때문이다. 따라서 과학자들은 시스템의 행동을 지배하고 있는 보편적인 과정의 법칙을 발견할 수 있다고 생각히게 되었다. 그러나 이렇게 본래 모습을 일정하게 유지할 수 있는 것은 변화 가 없기 때문이 아니라 여러 가지 방법을 통해 에너지가 일정하 게 방출되고 일정하게 유입될 수 있기 때문이다. 만일 이러한 양 상이 지속되지 않는다면 과학자들이 그토록 복잡한 것을 하나의 단위로 취급할 수 있으리라는 생각을 하지 못했을 것이다. 그러나 문제는 상대적으로 몸집이 크고 또 감각기관이라는 것 울 가지고 있는 우리 인간에게도 일반적인 생물학적 단위라는 것 이 명백한가 하는 데 있는 것은 아니다. 인간에게도 생물과 생물 학적 종이란 것이 아주 명확한 존재처럼 보인다. 그러나 그것이 중력장처럼 모호한 것이라 해도 중요한 생물학적 개념으로서의 그들의 위치는 변화되지 않을 것이다. 문제는 이러한 개념이 생물 학적 이론의 향상에 기여할 수 있겠느냐는 것인데, 생물학적 종이 그렇게 중요한 전화 단위는 되지 못한다는 것이 맞는 답일 것 이다. 따라서 과거에 그러한 단위가 강조되었던 것은 잘못된 일이 다. 미래의 생물학자들은 생물체 혹은 생식관계와는 전혀 관계없 는 새로운 생물학적 이론, 아마 새로운 전화론을 만들어낼지도 모 른다. 그러나 이러한 변화에 필수적인 개념적 혁명은 너무 거대한 일이기 때문에 기능적 등가성을 배제하여 목적론적 설명을 연역 적으로 만드는 일을 제쳐놓고 이 일을 해야 할 이유가 없다는 것 을명심해야할 것이다. 목적론적 설명을 연역적으로 만들기 위한 세번째 방법은 어느 특정한 기능에 관련된 기능적 등가성을 모두 열거하고, 적어도 그 열거된 항목들 중의 하나에 의해서 그 기능이 수행된다는 극히
빈약한 결론을 연역하는 것이다. 예를 들면, 식물은 기생을 하거 나, 썩은 생물에 붙어서 생활하거나, 또는 광합성을 통하여 자신 에게 필요한 양분을 얻을 수 있다. 그러므로 식물이 생존한다면 우리는 그 식물이 적어도 이러한 방법 중 하나에 의존한다고 연 역할 수 있다. 이렇게 함으로써 논증의 연역적 성격은 구제가 되 었지만 그 대신 결론이 상당히 빈약해졌다. 결론을 강화시키는 한 가지 방법은 영국에서 엘리자베스 시대의 베이컨 Francis Bacon 과 다윈 시대의 밀J ohn Stu art M ill에 의해 제안된 것으로 한 번에 하나씩 기능적 등가성을 제거해 나가는 것이다. 이러한 방법 을 이용하려면 각 기능마다 서로 확연히 구별이 되는 몇 개의 항 목들이 있어야 하는데, 그러한 항목들이 있다 하더라도 실제의 예 에서는 위의 두 사람의 제안처럼 등가성을 하나씩 제거해 나가는 것은 거의 불가능한 일이다. 기능적 등가성의 존재는 자연의 전리 이고 따라서 아무리 많은 지식을 거기다 보탠다 하더라도 결코 그것을 제거할 수가 없을 것이기 때문이다. 아직 마지막 방법이 하나 더 남아 있는데, 그것은 목적론적 설 명을 통계확률적 설명의 예로 보는 것이다. 이는 행펠 (1965 : 312) 에 의해 제시된 것으로, 그는 목적론적 설명은 〈전제되어 있는 상황 중에서 대단히 가능성이 높은 ‘어떤 항목’을 나타내 보이는 귀납적 논증으로 더 적절히 설명될 수 있을 것이다〉라고 하였다. 그는 계속해서 〈그러나 이 과정은 대부분의 경우 어떤 확고함을 약속하지는 못한다. 왜냐하면 대부분의 경우 대안이라 할 수 있는 행동양식, 제도, 관습 등, 주어전 기능에 대한 전제 조건이나 필요 룰 충족시킬 수 있는 것들의 범위를 정확하게 구체화시킬 수가 없을 것이기 때문이다. 그리고 그 범위가 정해진다 해도 그 범위 를 몇 개의 경우로 나누고 그 각각에 확률을 부여할 수 있는 만
족할 만한 방법이 없다 〉 고 말하고 있다. 행펠의 논문에서 연구된 예들은 사회학에서의 기능적 분석으로 부터 인용한 것이어서, 생물학에서는 어떤 필요에 대한 기능적 등 가성을 모두 열거하고 그 각각에 확률을 부여할 수는 없을 것이 다. 그러나 가능성이 아주 높은 대안들이 숫자적으로 볼 때는 소 수이지만 또 잘 알려져 있는 경우도 많이 있다. 진화론에 의하면 생물체 내의 구조는 늘 높은 효율성을 가지고 긍정적 기능을 수 행하는 것으로 생각된다. 물론 이에는 예의도 있지만 그러한 예의 는 상대적으로 아주 드물다. 여태까지 예로 든 생물학의 목적론적 전술을 보면, 거의 모든 경우에 있어서, 어떤 항목이나 행위는 극 단적으로 비정상적인 상황에서도 거의 항상 특정 기능을 수행하 고 있다. 포유류에 있어서 보통 심장박동의 긍정적 기능은 혈액을 순환시키는 것이고 또 심장은 분명히 그런 일을 하고 있다. 목적 론적 시스템에는 대개 기능적 등가성이 존재하지만 그들 모두가 똑같이 작용하고 있는 것은 아니고, 그 중 가장 작용가능성이 높 은 것들만이 잘 알려져 있다. 물론 모든 목적론적 설명이 다 그 런 것은 아니고, 때로는 기능적 등가성이 너무 많은데다 그 중 어느 것도 다른 것들에 비해 우세하다고 볼 수 없는 경우도 있 다. 그러한 경우에는 기능적 등가성 중 어떤 것이 어느 한 경우 에 있어서 높은 확률을 가지고 그 기능을 수행할 것인지를 추론 하는 것은 불가능하다. 그러나 이러한 상황은 예의에 해당하며 하 나의 규칙으로 볼 수는 없을 것 같다. 그러나 목적론적 설명들을 통계-확률적 설명으로 해석한다고 해도 주요한 문제점들은 그대로 남아 있다. 동계-확률적 설명에 있어서의 일반화는 과학법칙이어야 하는데, 목적론적 설명에 있어 서의 일반화는 목적론적 전술인 것이다. 따라서 목적론적 진술도
과학법칙으로 간주되어야 할 것이다. 그러나 불행하게도 여태까지 들었던 모든 예들은 그에 부합된다고 볼 수가 없다. 예를 들어, 엽록소가 광합성에 필수적이라는 것이 전리라 하더라도 그것을 자연의 법칙으로 볼 수는 없다. 보통 촉매라는 것은 반응에 필수 적인 것이 아니라 그 반응의 효율을 높이는 일을 하는 것뿐이고 대개의 경우 여러 가지 종류의 촉매들이 똑같은 기능을 수행할 수가 있는 것이므로, 어느모로 보나 엽록소(혹은 이와 비슷한 복 합물)가 광합성에 필수적이라는 진술은 우연적인 일반화이지 자 연의 법칙은 아니라고 생각된다. 많은 철학자와 과학자들이 과거 에 목적론적 설명에 만족하지 못했던 것은 사실이지만 그들의 불 만은 사실 엉뚱한 데 있었다. 정말로 중요한 것은 목적론적 진술 의 법칙성에 관한 문제이다. 그들 전부가 자연법칙이 될 수는 없 겠지만 목적론적 설명을 포괄법칙 모델에 의한 설명으로 간주한 다면 적어도 그들 중의 일부는 자연법칙이어야 하기 때문이다(제 3 장에서 논의되었던 법칙들과 우연적 일반화를 참조할 것). 요약및 결론 철학지들은 종종 인과적 진술과 설명이 이론적 배경에 역행하 여 만들어전다고 이야기해 왔다. 어떤 항목들이 인과적 그물망의 요소로 고려되어야 하는지 그리고 그들이 서로 어떻게 연관되어 있는지를 결정하는 것은 이론인데도 말이다. 이는 목적론적 진술 과 설명에서도 마찬가지여서 이둘 역시 배경에 역행하여 만들어 지고 있다. 그러나 시스템으로 혹은 시스템의 한 부분으로 그리고 시스템의 선호상태로 간주되어야 하는 것이 무엇인지를 결정하는
것은 이론이다. 그러나 목적론적 시스템이나 설명의 경우, 이러한 이론들이란 선택이론 selec ti on t heor y울 말한다. 철학자들 편에서 는 인과론적 이야기가 목적론적 이야기보다 더욱 많은 정보를 담 고 있다고 생각하는 경향이 있다. 따라서 재구성되어야 하는 것은 언제나 목적론적 이야기 쪽이다. 죽 목적론적 이야기를 순수한 인 과론적 용어로 재구성해야 한다는 것이다. 목적론적인 주장이 상 당히 인과적인 경우도 있고, 또 그것이 이론이라고 할 수도 없을 만큼 극히 초보적인 이론을 근거로 해서 만들어진다는 것도 맞는 말이다. 일상적인 이야기 속에 나오는 목적론적 주장의 경우에는 특히 그렇다. 일상적인 이야기 가운데서 목적론적인 전술이나 설 명을 정확하게 말할 수 없는 이유도 바로 이것이다. 예를 들면, 대개의 목적론은 〈사람〉의 상식 이론에 불과한 것이다. 그러나 일상적인 인과론적 이야기에서도 이와 비슷한 일이 관찰되기는 마찬가지이다. 때로는 설명을 요구하는 대상 자체가 상당히 인과적이고 또 인 과적 설명을 요구하고 있는데, 과학은 인과적 사건이 아니다. 인 과적 설명이나 목적론적 설명 모두 그에 필요한 상당한 양의 기 본 지식을 전제하고 있으며, 그것이 필요에 따라 만들어질 수 있 다고 가정하고 있다. 하지만 배경 지식도 전혀 없고 형식화의 설 명력은 그저 허울 좋은 것에 지나지 않는 경우가 너무나 많다. 인 과론적 설명과 목적론적 설명 모두에 있어서, 위험한 일 중의 하 나는 가끔 그들의 설명이 실제보다도 더 설명적인 것처럼 들리고 따라서 그 이상의 질문을 막아버린다는 데 있다. 그러나 목적론적 설명의 경우가 인과론적 설명의 경우보다 더 위험이 큰 것 같다. 왜냐하면 목적론적 시스템은 놀랄 만큼 분명한 것처럼 우리에게 보이기 때문이다. 예롤 들면, 윌리엄스 (G.C. Wi lliam s, 1966 :
260-261) 는 합목적론t eleonom y에 대한 그의 연구에서 〈 관례인데, 기능적 설계 fun cti on al des ig n란 연구자가 직관적으로 이해할 수 있고, 그러한 이해롤 다른 사람에게 납득이 가도록 전달, 교환할 수 있는 어떤 것을 말한다. 이것이 전실이라 하더라도 니는- 합목 적론이 발달하면 곧 적응성을 증명하는 표준화된 기준과 그것을 기술하는 공식적인 용어를 요구하게 되리라는 데는 의문을 가지 고 있다 〉 고 밝히고 있다. 생물학지들이 일찍이 그렇게 하지 않은 가장 큰 이유는 기능적 관계를 결정하는 데서 발생하는 문제 가 운데 〈 직관적으로 쉽게 해결될 수 있는 〉 것이 너무나도 많았기 때문 01 다. 목적론적 시스템이 실제보다 훨씬 폐쇄되어 있는 것처럼 보이 기 때문에 우리는 그 체계나 작용이 실제보다 훨씬 문제가 적다 고 믿게 되었다. 과연 생물을 목적론적 시스템으로 인식하는 데 필요한 이론들은 얼마나 정교해야 하는가? 대답은 그렇게 정교할 필요가 없다는 것이며, 바로 이 점이 왜 목적론적 시스템이 가끔 불만족스럽게 되어버리는가 하는 이유이다. 이러한 특별한 상태는 우리 자신이 바로 목적론적 시스템이며, 다른 목적론적 시스템과 동일한 선택 과정을 통하여 나타난다는 시실로 설명될 수 있을 것이다. 그리고 부분적으로는, 그것들을 인식하고 그에 대해 반응 하는 일관성 있는 실체로서의 우리의 능력 때문이다. 목적론적 이야기와 인과론적 이야기의 차이는 그 둘이 부분적 으로 개방되어 있는 시스템의 서로 다른 형태에 대해 서술하고 있기 때문이라는 데 있지, 어느 한쪽이 다른 쪽보다 애초부터 더 많은 정보를 담고 있다는 데 있는 것은 아니다. 때로는 인과론적 형식이 더 많은 정보를 담고 있고 목적론적 전술은 더 확장되어 야 할 필요가 있기도 하며, 어떤 때는 목적론적 전술이 더 많은
정보를 지니고 있고 인과론적 진술이 더 확장될 필요가 있기도 하다. 인과론적 전술과 목적론적 전술이 상대방의 용어로 서로 바 꾸어 표현될 수 있게 하려면 양쪽 모두에 있어서 상당히 많은 재 구성이 이루어져야 한다. 그러나 주목해야 할 점은 교류상의 어려 움이 목적론적 이야기가 지니고 있는 특칭들에 의해서만 야기되 는 것이 아니라 인과론적 이야기의 특칭에 의해서도 야기된다는 것 01 다. 인과론적 진술과 목적론적 전술은 모두 부분적으로 개방되어 있는 시스템을 묘사하기 위해 사용된다. 그러나 그 개방의 형태는 두 경우에 있어 각기 다르다. 인과론적 묘사를 가능하게 하는 상 황이 개방되어 있는 것은 인과적 그물이 모든 방향으로 뻗어 있 기 때문이며, 이때 그물을 이루고 있는 모든 울과 매듭이 똑같이 중요한 것은 아니다. 보통 인과적 원인은 그물망의 올 중 하나 그리고 아마도 하나의 매듭만을 가지고 분명하게 언급될 수 있으 며, 나머지는 그것을 뒷받침하는 지식으로 간주된다. 필요조건을 구체화하는 과정이 설명적이다 못해 가능성의 증가라는 개념보다 도 더 설명적으로 보이는 하나의 이유가 바로 이것이다. 알고자 하는 사건에 도달할 수 있는 길이 여러 개 있다 할지라도, 필수 조건을 구체화할 때는 이들 대체 가능한 모든 울들이 반드시 거 쳐 지나가야만 하는 하나의 매듭울 지칭하게 된다. 목적론적 시스템으로 귀착되는 상황이 개방되어 있는 것은 목 적론적 시스템의 체계가 끊임없는 에너지의 교환과 부분적인 변 형에 의해 유지되기 때문이다. 순수하게 인과적인 용어만을 가지 고 목적론적 시스템을 다시 묘사하고자 한다면, 우리는 폐쇄된 피 드백 고리를 통해 그런 시도를 하지 않으면 안될 것이다. 물론 과학이 발달하는 데 따라 이러한 제반 상황들은 태양계 모델에서
와 갇이 절대적으로 과정의 법칙에 의해서만 지배되는 폐쇄 시스 템으로 성공리에 재분석될 수도 있을 것이다. 그렇게 된다면 과학 적 논술에서 인과론적인 언어와 목적론적 언어 모두가 제거될 수 있을 것이다. 그러나 그렇게 될 때까지는 그 두 가지 방법에 의 한 묘사와 설명은 과학의 한 부분으로 계속 남아 있을 것이며 과 학자들에 의해 정중하게까지는 못되더라도 그 역할에 맞는 적절 한 대우를 받아야만 될 것이다.
잭 생체론과 환원론 논쟁의 성격 헉슬리 Ju li an Huxle y는 최근에 발간된 그의 자서전에서 할데인 J. S. Haldane 과 나누었던 대화를 다음과 같이 적고 있다. 할데 인 박사는 자신이 생체론(生體論, or gani c i sm) 의 신봉자라 고 했는데, 그것은 반기계론자(反機械論者)임을 뜻하지만 그렇다 고 신화적 생기론자도 아니라는 것을 뜻한다. 나는 그가 의미하 는 바를 잘 이해하지 못했고, 그래서 약간의 논쟁이 벌어졌다. 내 가 기계론적 설명을 필요로 하는 몇 가지 실험 결과에 대해 이야 기하니까 그는 〈여보게, 하지만 그건 노르가니즘 nor gani sm 이야 노르가니즘〉하고 말했던 것이다.l)
1) Memorie s (New York : Harpe r and Row, 1971), p. 138. 줄리앙 헉슬리 Ju li an Huxle y는 T. H. 헉슬리의 손자이고, 알도스 헉슬리 Aldous Huxle y와 형제간이다. J. S. 할데 인 J. S. Haldan e{는 진화의 유전학 이론의 창시자인 J.B .S. 할데인의 아버지이다.
하지만 언어의 이러한 모호함은 모두가 소위 기계론자와 생체 론자, 유물론자와 생기론자, 혹은 환원주의자와 전체주의자 hol i s t 사이에서 생기는 논쟁들의 중요한 특성이어서 이러한 오랜 논쟁 의 양쪽 면을 다 설명할 수 있는 용어는 몇 개되지 않는다. 게다 가 이러한 소수의 용어마저도 아직 그 정의가 정확하지 않고 사 용하는 사람마다 그 뜻이 제각각인 것 같다. 이러한 용어상의 혼 란은 반대편 견해를 우스꽝스러운 이야기로 치부해 버리고 자신 의 견해가 잘못 이해되고 있다고 상대편을 격렬히 비난하게 만 든다. 필자는 가능하다면 이러한 논쟁을 회피하고 싶지만 이것을 완 전히 무시해 버리기에는 그들이 과학철학에서 너무나 중심적인 역할을 해왔다고 생각한다. 그러나 솔직히 말해서 필자는 대단한 열정을 가지고 이 논쟁에 참여하고 있지는 않았다는 것을 인정해 야 하겠다. 이러한 논쟁 속에서 선택할 것이라고는 아무런 비판없 이 수용해 버리거나 철저히 반대하는 것인데, 나는 그 어느 것도 택하고 싶지가 않다. 사태를 더욱 악화시키는 것은 어떤 이가 자 신이 옳다고 생각하는 주장을 표명한다거나 혹은 자신을 생체론 자나 환원주의자라고 자처한다면 그 죽시 반대 편에서는 위협적 인 경고가 나울 것이며 감은 편의 사람들에게서는 정중한 침묵이 나올 것이 분명하다는 것이다. 이들 논쟁은 과학이라기보다는 정 치적인 논쟁이나 성경의 주석에 대한 회고록에 가깝다고 할 것이 다. 과학자 집단의 전도사(傳道師)들이 우리로 하여금 믿게 하려 는 과학적 사상은 정치적 혹은 종교적 사상과 크게 다를 바 없는 것이다. 특히 이론적인 공약이나 근본적인 형이상학적 신념에 관 한 것일 때는 더욱 그렇다. 그러나 최소한 몇 가지 문제에 있어 서는 과학에서의 논쟁은 정치나 종교 분야에 비해 훨씬 많은 증
거, 증명, 추론 등에 바탕을 두고 있다고 볼 수 있다. 이제 우리 앞에 놓인 문제는 생체론-환원론의 논쟁이 얼마만큼 경험적 사 실, 이론적 공약, 철학, 형이상학 등에 있어서의 차이에 근거하고 있는가 하는 것이다. 만일 이러한 질문에 대한 답이 가능하다면, 그러한 문제를 과연 어떻게 해결할 수 있을까? 역사상 어떤 시기에는 경험적 사실에 의해 과학적 논쟁이 해결 될 것 같이 보이기도 했다. 그래서 실험을 하고 다시 검증을 하 고, 유기물질이 합성되기도 했으나 결국 그 이상의 아무런 의미를 찾을 수는 없었다. 또 어떤 시기에는 주제가 거의 이론적인 것이 어서, 예를 들면 기계론은 모든 과학이 궁극적으로는 물리학의 한 부분인 역학으로 환원될 것이라는 식으로 해석하였다. 그러나 역 학은 모든 과학은 고사하고 물리학의 기초로도 부적합하다는 사 실을 알게 되었다. 그러나 그러한 사실이 기계론에 심각한 타격을 주었을 것이라고 생각하는 사람이 있을지 모르지만 사실은 그렇 지가 않았다. 또 l 양1 l 기에 전화론이 도입되고 2 伊1 ] 기에 유전의 분 자적인 기초가 밝혀짐으로써 기계론자와 생체론자 사이의 논쟁에 큰 영향을 주었다고 생각할지도 모르나 실제로는 그 반대였다. 한 편 그렌 Marjo r ie Grene 이나 폴라니 Mi ch ael Polany i 같은 반환 원론자들은, 생물을 마치 수많은 분자들로 이루어전 분자더미로 취급하는 환원론자들이 종합적 전화론을 성경처럼 삼고 있는 것 을 비난하였다. 심프슨이나 메이어 같은 생체론자들은 전화론이야 말로 기계론적 환원론에 대한 가장 큰 장애라고 생각하였다. 둘 중 어느 한쪽 견해만이라도 확신하고 싶다고 해서 이러한 논쟁의 역사를 자세히 공부할 필요는 없다. 경험 세계에 대한 그 어느 것 도 이쪽 혹은 저쪽 방식으로 확고히 정착될 수가 없기 때문이다. 하인(Hi lde Hein , Hi nt i kk a , 1969 : 239 에서)은 이러한 논쟁이
지속되는 것, 논쟁이 계속되는 동안의 언짢은 기분, 과학이 발전 됨에 따라 그것이 어느 한쪽을 지지하게 되는 것 등은 모두 메타 과학적이거나 메타이론적 생각에 달려 있는 것이라고 하였다. 사이 몬 (M i chael Sim on, 1971 : 237) 은 대체로 거기에 동의하면서도 〈 서로 반대되는 견해 사이의 충돌은 진정한 과학적 논쟁의 표시 이다 〉 라고 말하고 있다. 현재 널리 받아들여지고 있는 견해의 하 나에 의하면 과학이라는 것도 추론 혹은 논증, 증거의 정도 등에 서 정치적인 혁명보다 더 나을 것이 없다는 것이다. 아마 그 말 이 맞을 것이다. 그러나 과학적 논쟁과 정치적 논쟁 사이, 그리고 생기론과 기계론의 논쟁 사이에는 분명히 다른 점아 있다. 과학의 세계에 있어서 때로 한 과학 이론이 다른 이론보다 우세한 경우 가 있다. 예를 들어 과학자들은 지구 중심적인 체계보다는 태양 중심적인 체계를 택할 것이다. 마찬가지로 정치적 논쟁도 때로는 극적인 타결을 보기도 하고 아니면 한쪽이 승리하고 다른 쪽은 패하기도 한다. 그러나 생체론과 환원론 사이의 논쟁은 영원히 계 속될 것이다. 생기론-기계론 사이의 논쟁에 대한 수많은 문헌에서 나타나는 한 가지 경향이 있다면 초기에는 그것이 상당히 단순했던 데 비 해 후기에는 훨씬 모호해졌다는 것이다 . 처음에 생기론은 생명체 와 비생명체 사이에는 독특한 차이가 있다는 견해였다. 생명체는 독특한 형태이거나 아니면 비생명체와는 다른 물질들로 만들어진 것이며, 생명은 절대로 물질론으로 환원될 수가 없다는 것이었다. 기계론자들은 역학, 좀더 좁게 정의하자면 운동하는 물체에 관한 과학은 생물학을 포함하여 모든 과학의 기초가 된다고 주장하였 다. 이러한 견해들은 수세기에 걸쳐 그 오류가 드러날 때마다 과 학자들과 철학자들에 의해 정교하게 수정되어 왔다. 다음 쪽부터
는 생명의 특성, 생물학과 다른 과학 분야 사이의 관계 등에 대 한 논쟁이 초기의 단순한 설명에서부터 현재 유행하고 있는 좀더 정교한 설명에 이르기까지 계속 변천되어 온 과정을 살펴보게 될 것이다. 독자들은 〈 생체론 〉 이나 〈 환원론 〉 과 같은 메티이론적인 용어도 〈 힘 〉 이나 〈 종 〉 같은 이론적 용어나 마찬가지로 때로는 급격하게 혹은 우연히 그 의미가 계속 변화되어 왔음에 주의해야 한다. 따라서 이 용어들은 〈 유전자 〉 라는 단어가 제 1 장의 앞부분 에서 정의된 후 멘델 유전학과 분자유전학에 대한 논의에서 줄곧 사용된 것처럼 이 장에서 더 이상 정의되지 않고 그대로 사용될 것이다 . 살아 있는 액체로서의 생명 생기론을 극히 단순하게 설명하자면, 생명체와 비생명체는 각기 다론 재료로 만들어졌기 때문에 서로 다르다는 것이다. 죽 생명체 와 비생명체는 서로 다른 물질로 구성되어 있고, 이들 재료는 다 론 쪽의 재료로부터 얻을 수도 없고 다른 재료로 환원될 수도 없 다는 것이다. 그리고 생명체는 물질적인 재료로 구성되어 있는 것 이 아니고, 비생명체는 생명을 가전 재료로 되어 있지 않다고 한 다. 이를 더 자세히 설명한다면 모든 것은 다 동일한 기본재료로 되어 있으나 생명체에는 그러한 기본재료 이의에 생명을 지닌 재 료가 더 들어 있다는 것이다. 뉴턴 이후, 생명을 지닌 재료라는 것은 칼로리나 풀로지스톤 또는 양을 측정할 수 없는 어떤 액체 등과 유사한 특수한 액체, 죽 생명액체 vit al flu i d 라고 설명되었 다. 열이 따뜻한 물체에서 차가운 물체로 흐르는 액체라고 생각되
었던 것처럼 생명은 생식을 통해 전달되며 죽으면서 몸으로부터 이탈되어 나가는 액체로 생각되었던 것이다. 생명을 생명액체라고 생각한다면 필연적으로 두 가지 결론에 도달하게 되는데, 그것은 생명체는 비생명체로부터 전화될 수 없 다는 것과 생명은 실험실에 앉아서 창조해 낼 수 없다는 것이다. 이 두 가지는 같은 말이기도 하다. 죽 생명은 곧 생명액체이고 지구가 이 액체를 가지고 있지 않다면 생명체가 이 지구 위에서 자연적으로 생겨난디는· 것은 불가능하다는 이야기가 되는 것이다. 생화학자들이 모든 필요한 물리적 원소들을 적당한 순서로 결합 시킬 수 있을지는 모르나 거기에다 생명액체를 첨가하지 않는 한 결코 시험관에서 생명이 창조될 수는 없을 것이다. 생화학자들이 생명액체를 어떻게 얻을 수 있는지 알지 못히는· 한 결코 생명체 롤 창조할 수는 없다는 것이다. 사람들은 이와 유사한 논증을 통 해 컴퓨터의 사고능력에 대해서도 반대를 하고 있다. 마음이라는 것이 인간이 소유하고 있는 특별한 종류의 물질이고 기계에는 그 런 것이 없다면, 사람이 하는 정신적인 일 중의 일부(예를 들어 장기를 두고기 수학 문제를 풀고, 귀납적 추론을 하고, 시를 쓰고, 사 기를 치는 일 등)를 수행하는 컴퓨터를 생산할 수 있을지는 모르 지만 그 컴퓨터는 인간의 마음과 같은 정신적인 물질을 결여하고 있기 때문에 결코 의식을 지닌 것이 될 수는 없다는 것이다. 이것은 극히 단순한 견해이다. 오늘날 이렇게 단순한 생각을 갖고 있는 과학자는 거의 없을 것이다. 사람들은 18281d 에 요소 (辰素)가 합성되었고® 최근에는 DNA 의 분절들을 만드는 데 성
® 요소 합성. 19 세기 초까지 생명체의 구성물질인 유기화합물은 생 명체에 의해서만 만들어지고, 생명체가 아닌 자연에서는 합성되지 않는 것이라고 생각되었다. 유기화합물의 유기(有機, or ga ni c) 라는
말은 그래서 붙은 것이다. 그러나 이러한 개념은 182 8'-i빈에 뵐러 Frie d lic h Wohler 가 시험관 속에서 요소를 무기물로부터 합성하는 데 성공함으로써 깨어지게 되었다. 이 요소(辰素)는 O=C(NH2h 라는 분자식을 가전 물질로서 사람을 비롯한 거의 모든 동물 오줌 의 주성분이다. 이 요소의 시험관내 합성에 이어 수많은 유기물들 이 속속 인공 합성되어 생명기계론의 기초를 구축하게 되었다.
공하였으며 그 의에도 여러 가지 실험들이 성공을 거듭하고 있으 니 이러한 발전이 생기론을 사라지게 하는 데 결정적인 역할을 했다고 생각할지도 모른다. 그러나 사실은 그렇지가 않다. 183~ 빈 에 별의 시차(視差) 현상을 관찰하기 훨씬 이전부터 이미 천문학 자들이 프톨레마이오스 시스템을 포기했던 것처럼 대부분의 생물 학자들은 생명액체의 존재가 결정적으로 거부되기 훨씬 이전부터 생명액체의 개념에 대한 유혹에서 벗어나고 있었다. 대체로 생물 학자들은 생명액체에 대한 연구가 큰 진전을 가져오지 못하고 또 그것과 모순되는 다른 이론들, 특히 전화론이 나타나기 시작했기 때문에 그러한 개념을 버리게 되었다. 생명체가 오래전 비생명물 질로부터 저절로 생간 것이라면 생명은 생명액체가 될 수 없을 것이며, 따라서 시험관 안에서 생명을 창조한다는 것은 최소한 원 리적으로는 가능한 일이다. 생기로서의 생명 생기론을 옹호히는 사람들 중 일부는 생명은 생명액체가 아니 라 생기(生氣 vit al fo rce) 라고 주장하였다. 이 문제는 그렇게 간 단하지가 않았다. 왜냐하면 물리학에 있어서의 힘의 존재론적 상 태에 대한 대혼란 때문이다. 어떤 물리학자들은 중력과 같은 어떤
힘이 어느 상황에나 늘 작용하고 있다고 주장하고 있다. 중력이 물질을 움직이는 것처럼 말이다. 그러나 또 다른 물리학자들은 중 력은 어떤 물체가 아니고 물체의 성질이라고 주장한다(이 문제에 대 한 훼 웰 Wi lliam Whewell 과 밀 Jo hn St u art Mi ll 사이 의 논쟁 을 참조할 것). 생기의 개념에 있어서도 이와 비슷한 혼란이 생기고 있다. 생기를 물질적으로 해석한다면 그것은 생명액체와 마찬가지 로 후세의 생물학자들에 의해 배척당하게 될 것이다. 그러나 만일 생기를 물체가 지닌 하나의 특성으로 본다면 생기론은- 상당히 그 럴듯한 위치에 설 수 있을 것이며 그에 대한 큰 반대도 피할 수 있을 것이다. 자력이란 것이 쇠막대에 첨가된 단순한 물질이 아닌 것처럼 생명도 생명체가 잉태될 때 그 생명체에 첨가된 물질은 아닌 것이다. 자성과 생명 모두 그들을 구성하고 있는 물질이 체 계화됨으로써 생겨나는 것이다. 철분자는 자력을 나타낸다. 그러 나 보통의 쇠막대는 분자가 불규칙하게 아무렇게나 배열되어 있 기 때문에 이들의 자기효과는 서로 상쇄되어 버린다. 그렇지만 분 자들이 점차적으로 극과 같은 방향을 향해 배열되면 막대는 서서 히 자기를 띠게 된다. 상황은 물리학자들이 시간을 강물처럼 흐르 는 것으로 간주하지 않게 된, 그리고 공간을 모든 방향으로 무한 정 확장될 수 있는 그릇 같은 것으로 간주하지 않게 된 당시의 물리학적 상황과 유사하게 된다. 현재 공간과 시간은 모두 물체의 체계화에서 생기는 특성 orga niza ti on al p ro p er ti es 에서 비롯되는 것으로 생각되고 있다. 이 모든 특성을 허구라고 하지 않는 한 시간과 공간은 허구가 아니다. 눈에 띄게 드러나는 경향성이란 것은 과학에 그리 많지가 않지 만 중요한 과학적 개념들이 사물과 물질이라는 범주에서 특성이 라는 범주, 특히 연관성과 체계화라는 범주로 옮겨가고 있는 것은
이러한 경향성 중의 하나가 될 것이다. 생명이라는 것은 시간, 공 간, 중력 혹은 자성과 마찬가지로 하나의 사물에 불과한 것이다. 이 사물에 〈마음〉이라는 것을 더 첨가하면 족한 것이다. 만일 생 명을 이와 갈이 어떤 물질계의 체계적 특성으로 간주한다면, 우리 는 최초의 생명체가 어디에서 왔는지 혹은 생명체가 죽으면 어디 로 가는 것인지 궁금해 하지 않아도 된다. 쇠막대가 자성을 떨 때 그 자성이 어디에서 오는 것인지, 자성을 잃을 때는 그 자성 은 어디로 가는 것인지에 대한 관심 정도나 마찬가지인 것이다. 말하자면 좋은 자장은 죽어서 자성의 천국에 간다는 등의 말을 할 필요가 없다는 것이다. 생명에 대한 기계론적 견해 초기의 기계론은 두 개의 기본 개념으로 구성되어 있었다. 모 든 과학이 역학으로부터 유도된다는 믿음과, 그 결과로 생명체도 단순한 기계로 취급될 수 있다는 결론이 그것이다. 세기가 바뀌면 서 많은 물리적 현상들이 물체의 운동만으로는 설명될 수 없다는 것을 알게 되었을 때 물리학자들은 첫번째 개념을 포기하였다. 그 러나 기계론적 생물학에서는 여전히 생명체를 기계에 비유하고, 생명체를 활차(滑車)나 호흡튜브 같은 것으로 이루어전 기계로 해석하고 있었으며, 또 생물이 액체를 포함하고 있다고 한다면 그 액체도 순수한 역학법칙을 따를 것이라고 생각하고 있었다. 불행히도 생기론과 기계론 양쪽에 대한 현재의 반론 중 많은 것들이 양자의 초기의 입장에 대한 것이다. 폴라니 Mi ch ael Pol- an yi는 〈환원이 불가능한 여러 원리들이 생명 현상을 설명하는
논리 단계들을 변화시키고 있다. 예를 들면, 갈릴레오나 가상디 Gassend i로부터 우리에게 이르고 있는 개념, 즉 사물의 모든 방 식은 궁극적으로 운동하는 물체로 이해되어야 한디는- 개념은 이 미 거부되었다 〉 는 것을 깨달았다고 주장하고 있다 .2 ) 그러나 이러 한 개념은 오래전에 이보다도 훨씬 더 결정적인 견해들에 의해 이미 부정되었다. 기계론지들이 과거에 생물을 분자더미라고 생각 했든 안했든 간에 현재 그들이 믿고 있는 것은 현재의 생체론자 둘이 생명을 생명액체라고 믿고 있는 정도 이상은 아닌 것이다.
2) .. Life's Irreducib l e Str u ctu r e, Scie n ce 160(1968), pp. 1308- 1312.
기계론자와 생체론자들은 생명체가 지닌 체계가 비생명체의 그 것과는 다르기 때문에 생명체와 비생명체는 서로 다르다는 데는 동의하고 있다(제 4 장 참조). 그러나 이 두 편을 가르고 있는 문제 가 두 가지 있는데, 하나는 존재론적인 것이고 다른 하나는 방법 론적인 것이다. 존재론적인 문제는 체계 자체의 특성에 관한 것, 죽 한 시스템의 체계라는 것은 반환원주의자들이 주장하는 것처 럼 그 시스템의 구성 요소들의 단순한 집합이 아닌 그 이상의 어 떤 것일까? 어떤 한 단계 더 높은 존재론적 단계 onto l og ica l level 가 생명과 마음에 더 첨가되어야만 하는· 것일까? 등의 문제이다. 다윈 시대와 마찬가지로, 기계론적 환원주의에 반대하는 사람들이 가장 큰 관심을 가지고 있는 이 존재론적 단계는 인간을 나머지 생물들로부터 분리시키고 있다. 얼마나 많은 존재론적 단계가 어 디에 도입되어야 하는가를 결정하는 것은 어려운 일이다. 왜 그러 한 존재론적 단계를 종의 진화, 행성계의 출현, 도시국가의 출현 등에는 도입하지 않는 것일까? 그렌 Marjo r ie Grene3) 은 일차원적
3) Bi ol og y and the Problem of Levels of Reality , The New
Scholastic , 41(1967), pp. 427-449.
존재론은 과학 내에서나 그 밖에서나 인간 경험의 주된 영역을 설명하기에는 부적합하다고 주장한다. 다차원적 존재론이 필요하 다는 것이다. 그러나 그녀는 어떻게 그러한 단계의 수를 결정하며 어디에다 그것을 도입해야 하는지에 대해서는 언급이 없다. 포르 트만 Ado lf Por t man 은 〈 원심성 cen t r i c ity 〉 이라고 부르는 것이 식 물에 있다고 믿었으나, 그렌은 (식물에서 발견된) 그러한 개념을 이해할 수 없다고 말했다. 그러나 비판자들은 원심성이라는 것은 동물에서건 식물에서건 이해할 수 없는 개념이라고 말한다. 이들 문제는 대단히 모호하지만 또 대단히 중요한 것이다. 현재 행해지 고 있는 시도들은 지나치게 모호하거나 지나치게 인상적이어서 별로 도움이 되지 못하지만, 우리가 생물을 완전히 이해하고자 한 다면 이것은 언젠가는 반드시 해결되어야 할 문제들이다. 자연을 연구하는 데 있어서 자연은 복잡하다는 것과 체계의 단 계성이 존재한다는 것보다 더 분명한 것은 없을 것이다. 그 중에 서도 생물계보다 더 체계의 단계가 여러 충이고 더 복잡한 것은 없을 것이다. 그러나 존재론적 차원, 죽 개체, 부분, 전체 등의 개 념이 경험에 의해 저절로 〈주어지는〉 것은 아니다. 오히려 이런 개념은 현상을 연구하면서 나타나는 것이며, 이것은 상식적 개념 과 일치되어야 할 필요가 없다. 예를 들면, 보통 사람에게도 숲과 나무의 차이는 극히 명백한 것처럼 보이지만 이렇게 분명해 보이 는 차이도 현대 식물학에서는 별로 대수롭지 않은 것이다 . 마찬가 지로 한때는 지상(달 아래)의 현상과 하늘(달 위)의 현상 사이의 차이는 결정적인 것이라고 생각되었다. 그래서 한쪽에 적용되는 법칙이 또 다른 쪽에도 적용될 수는 없다고 생각되었다. 그러나 현재는 이러한 차이란 대수롭지 않은 것으로 생각된다. 식물과 동
물 사이의 차이나 인간과 다른 종의 차이 혹은 인종간의 차이는 아직 분명하지 않다. 인간은 분명히 다른 종과 질적으로 다르다. 그러나 자석도 다른 쇠막대와 질적으로 다르기는 마찬가지이다. 생물학에서 더욱 중요한 것은 현상을 분석하고 법칙을 만들고 그 결과를 이론화하는 방법의 문제이다. 어떤 생물학자는 생태계 룰 그의 연구대상으로 하고 그 구성원들 사이의 상호 관계를 연 구한다. 이때 구성원들이란 생물개체, 지층의 여러 특성들, 계절에 따른 기후의 변화 등이 될 것이다. 그들은 분류학에서처럼 전화적 계보에 따라 생물을 분류하는 것이 아니라 생태계에서의 역할에 따라 나누게 될 것이다. 분류학에서는 두 종이 같은 속에 속하는 구성원일지 모르지만 그 중 하나가 야행성 육식동물이고 다른 하 나는 주행성 초식동물이라면, 생태학자들에게 있어서 이 두 종은 서로 다른 문에 속하는 것이나 마찬가지다. 어떤 생물학자는 특정 종에 속하는 생물의 해부에 관십이 있을 수도 있다. 이 경우에는 근육, 동맥, 기관 등이 그의 시스템의 부분이 될 것이다. 그러나 생리학자들은 동일한 시스템을 또 다른 측면에서 보게 될 것이다. 또 다른 차원에서 볼 때는 세포가 연구대상이 되는 시스뎀일 수 도 있을 것이다. 그러한 경우에는 이들 세포의 기원은 중요하지 않은 것인지도 모른다. 따라서 두 세포가 서로 다른 종에서 왔다 할지라도 그 두 세포를 같은 것으로 취급할 수도 있을 것이다. 이는 한 탄소 원자는 석탄에서 얻은 것이고 또 하나는 다이아몬 드에서 얻은 것일지라도 물리학자들이 그 두 탄소 원자를 동일한 것으로 생각하는 것과 마찬가지이다. 이들 현상 모두가 동일한 실체의 한부분을 이야기하는 것이라 는 사실에는 의심의 여지가 없으나, 이런 사실을 인식하는 것과 그것을 반영하는 과학 이론을 구성하는 것은 별개의 일이다. 여러
가지의 분석 단계에서 일어나는 수많은 현상을 설명하는 수많은 이론들이 있지만 현재로선 그것들이 하나의 통합된 이론으로 조 직되지는 못하고 있다. 상당한 우연성이 여러 분석 단계에서 일어 나는 현상들 사이에 존재하며, 어떤 경우에는 그러한 분석들이 서 로 양립되지 않는 경우도 있다. 하지만 기계적 환원론에 대한 논 쟁 중의 하니는 조만간 해결될 것이고 과학자들은 하나의 통합 이론을 만들 수 있을 것이다. 많은 과학자들 특히 생물학자들과 사회과학자들은- 과학의 통합이 실제로 물리학으로의 통합임을 두 려워하면서 그러한 작업이 가능하지도 않고 바람직하지도 않다고 주장한다• 그러나 그러한 초이론이 가능하다면 그것은 분명히 현 대 생물학의 이론과는 다를 것이며, 또 현대 물리학의 이론과도 다를 것이다. 그렇다면 그것을 뭐라고 불러야 좋을까? 우리가 궁 금해하는 그 모든 것을 위한 마작(麻雀)이라고 부르자. 환원론과 구성론 앞에서 이야기한 것들 가운데 마지막 것만이 현재 환원에 대한 과학자들간의 논의에 중요한 역할을 하고 있다. 논쟁의 방향이 방 법론적인 것으로 바뀌었기 때문이다. 심프슨 (G.G. Sim p so n, 1961) 은 과학에서 환원론과 구성론(構成論, com p os iti o ni sm) 이라 부르는 두 가지 접근방법을 대조시켰다. 이제 문제는 생물이 부분 으로 나누어질 수 있느냐 또는 무기물질로부터 합성될 수 있느냐 하는 것이 아니라, 과학적인 연구와 설명에 대한 여러 방법들 중 어느 것이 더 큰 결실을 맺을 수 있겠느냐 하는 것이다. 심프슨 에 의하면, 환원은 〈체계화의 어느 특정 단계에서의 구조나 현상
과 그보다 낮은 단계에서의 구조나 현상과의 〉 연관성이다. 그러나 순수한 환원에서는 시스템을 구성하는 요소들의 작용은 그들이 속하는 더 높은 단계에 있는 시스템과 완전히 분리시켜 설명되므 로, 심프슨은 〈 순수 환원론은 적절한 설명을 주지도 못하고 확실 한 예측을 가능케 하지도 않는다 〉 는 결론에 도달하였다. 또한 〈 원소, DNA 분자, 효소 등의 작용은 그러한 작용이 일어나는 시 스템 밖에서는 설명될 수도 예측될 수도 없기 〉 때문에, 〈 두번째 형태의 설명이 첫번째 형태의 설명, 죽 환원주의자들의 설명, 다 시 말해서 물리적, 화학적, 기계적 원리에 의한 설명에 첨가되어 야만 한다. 이 두번째 형태의 설명은 환원론에 대비시켜 구성론이 라고 부를 수 있으며, 이것은 생물개체나 그것이 속한 종의 구조 와 과정의 적응유용성 adap tive usefu ln ess, 나아가서는 그 종이 속한 군집 내에서의 생태적 기능에 의해 이루어지는 설명이다 〉 라 고 그는 주장했다. 이들 환원론자와 구성론자들의 설명에는 전화 적 발전에 기초를 둔 역사적 설명도 첨가되어야 할 것이다. 심프슨은 환원론자와 구성론자들의 설명 양식이 경쟁관계에 있 는 것이 아니라 보완관계에 있다고 생각하고 있다. 순수한 구성론 적인 설명도 순수한 환원론적 설명과 마찬가지로 불완전하다는 것이다. 분자생물학자들은 생물학적 시스템의 한쪽 면만을 강조하 려는 경향이 있지만 이 두 형태의 설명이 역사적 설명과 합쳐질 때만이 생물학적 현상에 대한 적절한 설명이 될 수 있다는 것이 다. 시스템 전반에 고루 적용될 수 있는 이러한 심프슨의 견해가 탁월한 것임에는 틀림없지만, 그는 이러한 구분을 통해 생물학과 물리학이 분명히 구분될 수 있다고 믿고 있는 것이 분명하다. 생 물학에서는 그 두 형태의 설명이 모두 주어질 수 있고 주어져야 하는 데 비해 물리적 현상에서는 환원론적 설명만이 의미가 있고
또 가능한 형태가 되는 경우가 많다. 그러나 심프슨의 주장 속에 들어 있는 이러한 암시에 대해 필자는 동의할 수가 없다(앞에서 논의한 역사적 설명과 목적론적 설명을 참조할 것). 가장 간단한 생명체에서도 그 체제의 복잡성은 물리학자들이 다루는 순수한 물리적 시스템에 비할 바가 아니며, 또 생명체의 체제 속에 남아 있는 과거의 역사적 기록은 비생명체의 그것에 비할 바가 아니다. 특히 개체나 게놈 수준에서는 더욱 그렇다. 또한 생물학적 현상에 대한 설명 속에는 어떤 종류의 물리적 현상보다도 더 많은 우연 성이 들어 있을 것이다. 그러나 그것은 정도의 차이일 뿐이다. 만 일 교과서적 예가 아닌 실제적인 물리 이론을- 연구한다면, 생물학 적 현상과 순수한 물리적 현상과의 차이가 그렇게 크지 않다는 것을 알게 될 것이다. 흑연과 디이아몬드는 겉으로 보기에는 아~느 다른 물질이다. 그러나 그들 모두 탄소라는 동일한 원소로 되어 있다. 이러한 의관상의 차이는 그 구성분자의 체제의 차이에서 기 인하는것뿐이다. 만일 심프슨의 의도가 물리학과 생물학 사이에 명확한 방법론 상의 차이가 있다는 것을 보이는 것이었다면 그는 실패했다고 볼 수 있다. 기껏해야 그는 양쪽의 강조하는 바가 다르다는 것을 보 여줄 수 있었을 뿐이다. 하지만 여기에 예의가 있는데, 그것은 목 적론적 시스렘에서 찾아볼 수 있는 특별한 형태의 체제에 의거한 설명이다. 만일 순수한 목적론적 시스템은 자연선택이나 어떤 생 물의 개입 또는 간섭으로 나타날 수 있는 시스템뿐이고 또 그러 한 조건 중의 하나를 충족시킬 수 있는 어떤 시스템이 생물학의 영역 내에 반드시 있다고 주장할 수 있다면 생물학의 독특함과 자율성에 대한 근거가 마련된 것이라 볼 수 있다. 하지만 이러한 조건전술을 위한 선행요소들 그 어느 것도 아직 확고히 정립되지
못하고 있다. 메이어 (in Mendelsohn, 1969 : 128) 는 심프슨보다 더 강력한 주장을 하였다. 그러나 그는 설명에 대한 방법이 아니라 발견의 논리에 관심을 두고 있다. 결국 환원론은 말하자면 위로 향하는 방향으로는 결코 성공하 지 못하고 있다. 생물학적 기능의 대부분이 물리학적 요소로 분 해될 수는 있겠지만 나는 낮은 단계의 성분을 모두 끌어모아 한 충 높은 단계에서의 새로운 통합을 시도하는 과정에서 어떤 새로 운 생물학적 발견이 이루어졌다는 것을 둘은 적이 없다. 생물학 적 발견, 특히 기본 성분을 연구하는 데 도움이 되는 생물학적 발견을 한 분자생물학자는 아무도 없다. 다론 말로 한다면 환원론이 그르냐 옳으냐를 주장하는 것은 무 의미하지만, 환원론이 대단히 불충분한 접근 방법이라고 말할 수 는 있다는 것이다. 옹호자들의 주장과는 달리 그것이 상위 단계 에서의 새로운 통합으로 유도되는 경우는 거의 없으며, 복잡한 시스템을 이해하기 위한 여러 접근 방법 중 가장 형편없는 것에 지나지 않는다. 그것은 알맹이 없이 텅 빈 설명 방법일 뿐이다. 끝 부분을 제의하면 메이어의 주장은 발견 방법으로서의 환원 에 가치를 두고 있다고 볼 수 있다. 그는 〈하강환원 downward reduc ti on 〉과 〈상승환원 u p ward reduc ti on 〉을 대비시켰는데, 하 강환원에서는 한 단계의 시스템들이 물리적으로 그롤 구성하는 부분시스뎀 subs y st em 으로 분해될 수 있으며 그러한 방법에 의해 더 낮은 단계에서의 통찰이 가능하게 된다. 상승환원에서는 한 시 스템을 구성하는 요소들이 물리적으로 결합되어 더 높은 단계의
시스템을 형성하게 된다. 그러나 메이어에 의하면, 이 높은 단계 에서의 통합으로 얻을 수 있는 새로운 통찰은 거의 없다는 것이 다. 따라서 어느 방법도 어떤 시스템이 존재하고 있는 바로 그 단계에서는 그 시스템을 이해하는 데 별 도움을 주지 못한다는 것 0] 다. 메이어의 이러한 주장을 생물학의 실제 예에 적용하기란 대단 히 어렵다. 메이어 자신은 한 생물집단을 소집단으로 나누고 이 소집단을 여러 가지 방식으로 다시 합천 후 그 결과를 연구하였 다. 그는 전화 과정과 유전 과정의 특성에 대한 그의 결론을 그 의 연구대상이었던 소집단뿐 아니라 종에도 적용하였다. 이와 마 찬가지 시도로 세포학에 있어서 가장 성공적인 방법 중 하나인 세포 배양을 둘 수 있다. 하지만 메이어가 염두에 두고 있었다고 생각되는 환원의 형태에 가장 가까운 예는 오초아 S. Ochoa 와 그 의 동료들의 연구에서 발견될 수 있을 것이다. 그들은 수년 간에 걸쳐 뉴클레오티드와 뉴클레오티드를 연결하여 DNA 분절을 만 들어내는 데 성공하였고, 최근에는 드디어 복제가 가능할 만큼 큰 조각의 분절을 만들어내게 되었다. 이런 방법이 그다지 생산적이 지 못한 방법이었는지는 모르지만, 어쨌든 그러한 연구는 분명히 유전 암호를 푸는 데 큰 도움을 주었고 이러한 지식은 통합의 상 위 단계를 이해하는 분기점이 되었다. 예를 들어 생물의 수많은 질병이 코돈 하나가 변화됨으로써 생기는 것임을 알게 되었던 것 01 다. 분석의 어느 단계에서든지 풀지 못하는 문제는 항상 존재한다. 과학(생물학뿐만 아니라)에서 가장 훌륭한 방법 중의 하나는 이 러한 문제들을 다른 단계로 옮겨서 분석해 보는 것이다. 죽 더 낮은 단계로 혹은 더 높은 단계로 옮겨 놓고 문제의 해결 가능성
울 타진해 보는 것이다. 포유류의 콩팥의 미세구조를 연구하는 사 람이 종의 진화라는 단계로 그의 시야를 넓히지 않는다면 신소체 의 특칭들은 영원히 수수께끼로 남게 될 것이며, 역으로 비교해부 학자가 만일 세포생리학을 전혀 무시한다면 콩팥의 헨레 고리가 얼마나 중요한지를 인식하지 못할 것이다. 만일 기계론적 환원론 을, 모든 문제와 모든 해결이 분석의 가장 낮은 단계에서(즉 양자 론의 수준) 이루어져야 한다는 것으로 해석한다면 그것은 분명히 잘못된 것이다. 과학자들은(생물학자뿐만 아니라) 어떤 현상을 - 조 사하는 데 분석이라는 방법과 종합이라는 방법을 다 사용한다. 다 만 생물학에는 고도로 체계화된 시스템의 존재 때문에 다른 과학 에서보다도 종합적 방법이 더욱 필요한 것이다. 그리고 대상이 되 논 시스템이 워낙 복잡하기 때문에 종합적 방법을 사용하여 체계 의 상위 단계로 올라가는 것이 다른 과학에서보다 훨씬 더 어렵 다. 그러나 생물학적 현상과 순수한 물리적 현상 사이의 이러한 차이는 본질적인 것이 아니고 그 정도의 차이일 뿐이다. 환원과 양자물리학 환원에 대한 앞의 반론들- 대부분은 생물학자들에 의해 주장된 것이며 생물학과 생물학 이론에 대한 그들의 지식에서 비롯된 것 이다. 그러나 수많은 물리학자둘 역시 환원주의의 개념에 반론을 제기하고 있다• 양자물리학은 자연 현상을 미시적 수준에서 다루기 때문에 이들 반론은 대개 양자물리학 주변을 맴돌고 있다. 슈뢰 딩거 Erw in Schrod ing e r , 보어 Nie l s Bohr, 델브뤽 Max Delbrii ck , 하이젠베르크 Werner He i senber g로부터 위그너 Eug en e Wi gn er,
폴라니 Mi ch ael Polany i, 엘자서 Walte r Elsasser 에 이르기까지 물 리학자들은 양자물리학이 생물학에 대해 아주 중요한 암시를 지 니고 있다고 생각해 왔다. 언뜻 보기에는 이러한 주장을 받아들이 기가 극히 힘들 것 같이 생각된다. 왜냐하면 원자 수준 이하의 현상의 특수성이 거시적 대상으로 귀결되는 경우를 찾아보기 힘 들기 때문이다. 이상하게도 이 물리학자들은 환원론에 대한 반대 에 있어 생물학자들보다도 더 극단적이다. 물리학과 생물학의 관계에 대한 최초의 그리고 가장 분별없는 주장 중의 하니는 고전 열역학에서 비롯된다. 열역학 제 2 법칙에 따르면 우주의 모든 과정들은 엔트로피가 증가하는 방향, 즉 질서 를 감소시키는 방향으로 향한다. 그러므로 고도로 체계화된 생명 체가 계속 존재할 수 있다는 것은 이 원리에 모순되는 것처럼 보 인다. 그러나 질서의 정도가 증가된 일시적이고 국부적인 현상으 로서의 생명 현상은 열역학 제 2 법칙에 반드시 어긋나는 것은 아 니다. 왜냐하면 생물은 환경으로부터 에너지를 끊임없이 취함으로 써 그들의 체제를 유지시키고 있기 때문이다. 생물학에 있어서 중요하다고 생각되는 양자물리학의 두 가지 원리는 하이젠베르크의 불확정성 원리 (pr in c ip le of uncerta int y 혹은 pri n c ip le of i nde t er mi nac y)와 보어의 상보성 원리 pri n c ip le of com p lemen t ar ity이다. 모든 과학법칙이 형식상 보편적이어야 한다는 의미에서 보면 수 세기 동안 물리학자들은 결정론자였다 고 볼 수 있다. 그들은 엄격한 보편성에서 어긋난다는 것은 증거 가 불충분하거나 분석하는 데 적합한 단계에 대한 조사를 제대로 하지 못했기 때문이라고 생각했다. 분석의 한 단계에 있는 현상에 서 결정론적 법칙들을 발견하지 못한다면 좀더 낮은 단계에서 발 견할 수 있을 것이며, 절대적 정확성이란 것은 불가능한 일이겠지
만 관찰의 정확도를 무한정하게 높이는 것은 가능한 일이라고 그 들은 믿었다. 그러나 적어도 원자수준 이하에서는 그러한 두 개의 가정이 잘못된 것이었음이 증명되었다. 하이젠베르크에 의하면 소 립자의 위치와 운동량을 동시에 결정한다는 것은 불가능한 일이 며, 둘 중 하나가 확정된다면 다른 하나에 대해선 확률적 계산만 이 가능하게 된다. 따라서 소립자의 경우, 커다란 집단 ensemble 에 대해서만 확정적일 수가 있을 뿐, 개개의 입자에 대해선 반드 시 통계적일 수밖에 없다는 것이다. 상보성 원리는 . 불확정성 원리와 그 치원이 다르다. 그것은 양 자역학의 한 부분이라기보다는 그것에 대한 주석에 해당된다고 볼 수 있다 . 빛의 특성 중 어떤 것은 빛에 대한 입자적 해석을 필요로 하는 반면, 또 다른 어떤 특성은 빛이 파동과 같디~ 것 울 암시하고 있다. 그러나 입자론이나 파동설 그 어느 것도 빛의 특성 모두를 설명할 수는 없으며, 보어의 상보성 원리에 따르면 이 두 이론은 서로 환원될 수는 없으나 상호 보완의 관계에 있 다. 이 두 해석이 양립되지는 않지만 빛의 성질을 완전히 이해하 기 위해서는 이 두 가지롤 모두 사용해야만 한다는 것이다. 양자물리학은 극히 간접적이기는 하지만 생물학에 대단히 중요 한 영향을 끼치고 있다. 이것은 결정론적인 법칙의 절대적 우위성 울 마음에 지니고 있는 철학자들과 과학자들의 신념을 다~ 혼들 어 놓고 있다. 양자물리학의 기본 법칙이 통계적일 수밖에 없다면 아마 생물학에서의 통계법칙의 만연도 그리 나쁜 것만은 아니라 는 것이다. 그러나 양자물리학의 통계적 성격은 양자론의 이론 형 성에 있어서 불가피하게 필요했던 것임을 염두에 둘 필요가 있다. 통계적 성격을 배제하려면 양자론은 전혀 다론 형태로 이론화되 어야 했을 것이다. 제筑려서 우리는 전화론의 형성에서 거의 모
든 것이 통계적 성질을 띠고 있는 이유에 대해 이야기했다. 메이 어의 창시자 원리를 제의하면, 그것들은 모두 전통적인 연역 방식 에 의해 수립되어, 전화론과 양립 가능한 일종의 우연성을 포함하 고 있기 때문이다. 그러나 물리학자들은 앞에서 말한 (생물학에 대한) 양자물리학 의 영향에 만족하지 않는다. 오히려 그들은 유추를 더 확대시키려 하고 있다. 보어는 생물학의 연구대상이 반드시 살아 있는 것이어 야 하기 때문에 양자 수준에서의 불확정성과 갇은 상황이 나타나 는 것이라고 생각하였다 (Blackburn, 1966). 〈따라서 우리가 어떤 기관의 기능을 연구하여 생명현상에서 각 원자들이 갖는 역할을 이해하기 위해서는 동물을 죽일 수밖에 없게 된다. 살아 있는 생 명체에 대한 모든 실험에서는 생명체에 관련된 물리적 조건에 관 해서 불확정성이 존재할 수밖에 없다• 그러므로 이 개념 자체가 생명의 비밀 혹은 신비성을 불가피하게 하고 있는 것이다.〉 유추 롤 더 확대시켜서 델브뤽은 아마도 살아 있는 세포의 특성 중 어 떤 것은 그것을 구성하는 원자들의 구조와 상보적 관계에 있을 것이고, 세포에 관한 생물학적 지식이나 분자론적 지식만으로는 생명의 본질을 이해할 수 없고, 그 두 단계의 보완적인 지식이 필요하다고 하였다 (Blackburn, 1966). 이러한 견해는 상당히 홍미로운 것이어서, 우리는 그들을 좀더 깊히 음미해 볼 수 있을 것이다. 생물학자들은 생체밖i n v it ro 이나 생체내 in viv o 양쪽에서 그들의 연구를 계속하고 있다. 동물을 죽 이지 않고 살아 있는 상태에서 실험할 때도 그들은 그 체제에 대 해 상당히 폭넓게 연구할 수가 있다. 하지만 동물 하나를 사용해 서 알고자 하는 모든 것을 다 발견할 수는 없기 때문에 그들은 쉽게 구할 수 있는 수많은 실험동물을 사용해야만 한다. 여기서
상보성 원리를 생물학적 현상에 적용할 수 있는 가능성은 더욱 희박해진다. 상보성 원리는 형식상 양자물리학의 한부분이 아니 며, 이미 많은 물리학자들에 의해 심각하게 의문이 제기되어 있 고, 빛의 경우와 같은 수준에서 분석을 할 때에도 두 개의 다른 해석을 할 수밖에 없는 것이다. 이것을 생물학적으로 보면, 세포 수준 또는 그 이상의 연구에 의한 지식과 생화학적 연구에 의한 지식 사이의 관계와 같다. 어떤 의미에서는 그러한 지식이 서 로 〈상보적〉이라고 말할 수도 있겠지만 그것은 양자물리학에서의 의미와는다르다. 복잡성과 고유성 앞에서 우리는 물리학적 이론들, 그 중 무엇보다도 양자물리학 과 생물학 사이의 유추 가능성에 대한 물리학자들의 극히 피상적 인 견해에 대해 이야기했다. 20 여 년 이상에 걸쳐 출간된 여러 간행물 속에서 엘자서 Walte r Elsasser4) 는, 생물학의 양자물리학 으로의 환원에 반대하는 결정적 논증이라 생각하는 것을 발표하 면서 이들 유추를 더욱 확대시켜 나갔다. 그는 양자역학 내에 있 는 실체들의 집합이 균질하다고 주장했는데, 그것은 한 집합 내의 요소는 모두 그 구분이 불가능하다는 것이다. 예를 들어, 시공간
4) Ato m and Orga n is m (Prin c eto n : Prin c eto n Univ e rsit y Press, 1966) : see crit icism by Stu a rt Kauff m an, What Can We Know about a Meta z oan's Enti re Contr o l Sy st e m ? : on Elsa-sser's and Ot he r Ep ist e m olog ica l Problems in Cell Sc ien ce, Towards a Theoreti ca l Bi ol og y, Vol IV(Ch ica go : Aldi ne Publi - shin g Co., 1972) .
적인 위치를 제의하고는 동일한 양자 상태에 있는 원소들은 모두 똑같기 때문에 이러한 집합은 실제로는 유한하지만 무한한 것처 럼 취급될 수 있디는 것이다. 이렇게 균일한 집합 내의 한 개체 는 또 다론 개체의 역할을 할 수 있을 것이다. 그러나 엘자서는 생물학에서는 그러한 집합이 존재하지 않는다고 주장한다. 생물학 적 실체의 집합은 극도로 불균질하며 각각 고유성을 지니고 있다 는 것이다. 그는 물리학에는 균질한 집합이 있는 데 반해 생물학 에는 그러한 것이 없다는 사실에 의해 물리학과 생물학을 구분하 기까지 했다. 그는 〈물리학은 본질적으로 균질한 시스템과 집합 울 다루는 과학이며, 생물학은 불균질한 시스템과 집합을 다루는 과학이다〉라고 생각하였던 것이다. 만일 그가 옳다면 생물학자들 은 딜레마에 빠지게 된다. 알고자 하는 것을 찾기 위해서는 엄청 난 양의 실험을 해야 하고 무수한 생물을 죽여야 되기 때문이다. 또 같은 집합 내의 구성원일지라도 그들이 지닌 고유성 때문에 한 개체에 대한 연구를 통해 얻은 지식은 그 개체에 국한된 것일 뿐이기 때문이다. 만일 엘자서의 주장이 옳은 것이라면 생물학적 이론을 물리학 이나 화학으로 환원하는 일은 불가능하다. 왜냐하면 생물학적 이 론이란 것은 아예 존재할 수 없기 때문이다. 생물학뿐 아니라 물 리학에서도 양자론을 제의하고는 어떤 이론도 있을 수 없을 것이 다. 그 구성원들이 아주 동일하여 구분이 불가능한 경우란 원자 이하의 수준에서만 가능한 일이기 때문이다. 원자수준에서조차도 문제가 발생하기 시작할 것이다. 엘자서는 〈원자나 특정 분자의 가장 두드러진 특성 중의 하나는 그들이 아주 똑같다는 것이다〉 라고 주장했지만 곧 그는 동위원소의 존재를 인식하지 않을 수 없게 되었다(예를 들면 중수는 더 많은 중성자를 지니고 있는 물
분자의 비율이 높은 물이다). 물리학에 대한 엘자서의 주장은 이 성질체, 죽 그 구조는 동일하나 서로에 대해 거울상인 분자의 존 재에 의해 더욱 곤란하게 되었을 뿐 아니라, 거시적 수준에서 본 다면 물리학에 대한 엘자서의 명제는 완전히 와해되어 버리고 만 다. 보통 사람들에게도 (두 개의 양성자가 구분 가능하다는 의미에 서처럼) 두 개의 폭스바겐 자동차가 구분 불가능할 만큼 동일하 게 보이는 일은 없을 것이다. 균질한 집합이 과학에 필수적이라면 과학이라고 할 수 있는 것은 양자물리학뿐이다 . 확실히 엘자서는 과장을 했고 어떤 곳에서는 그도 그것을 깨닫 고 〈집합의 본질적 특성에 의해서 그 구성원들은 공통되는 특성 을 지니고 있으며(일반적으로 생물 집합의 구성원들은 수많은 공 통적 특성을 지니고 있다), 따라서 집합의 구성원들이 지닌 공통 적인 특성에 근거한 부분적인 예측이 가능하다〉는 것을 인정하였 다. 하지만 여기에서도 그는 여러 개의 분명치 않은 가정을 하고 있다. 예를 들면, 그는 생물집합이라는 것은 각각이 필수적이면서 다 합하면 그 구성원이 되기에 충분한 특성들로 이루어전 세트라 고 정의될 수 있다고 하는데, 앞에서 논의한 바대로 이 가정은 어떤 생물집단을 시간 차원을 떠난 어떤 시점에서만 간주할 때 가능한 것이다. 만일 시간적인 차원을 인정한다면 도약적으로 전 화하는 종들만이 엘자서가 이야기하는 방법에 의해 정의될 수 있 을것이다 . 덧붙이면 엘자서는 생물 집단의 요소를 생물체라고 가정하고 있다. 하지만 생물체는 매우 복잡한 시스템이다. 복잡성과 동일성 간에는 반비례의 관계가 존재한다. 따라서 시스템이 복잡해질수록 모든 점에서 동일하다고 생각되는 시스템을 찾기가 어려워진다. 생물학에서 엘자서의 균질 집합에 가장 가까운 것을 찾는다면 클
론 clone 이 될 것이다. 클론이러는 것은 모두 동일한 한 짝의 염색 체에서 온 것들로 이루어진 것이다. 그러나 이러한 가장 극단적인 경우에서도 그 클론의 구성원들은 유전의 불완전성으로 인해 유 전적으로도 또 환경의 다양한 영향으로 인해 표현형적으로도 조 금씩 변이를나타낸다. 하지만 엘자서의 주장과는 달리 엄격한 균질 집합이 없이도 과 학은 가능하다. 물리학지들이 천체를 연구할 때 그들의 복잡성을 모두 고려하면서 연구하는 것은 아니다. 그들은 다만 이들 천체 사이의 어떤 중요한 관계를 보여주는 공통적인 특성들을 찾으려 노력하고 있을 뿐이다. 예를 들어 뉴턴의 법칙은 물체의 질량과 속도만을 다루며 색깔, 모양, 화학적 구성 따위는 상관하지 않는 다. 마찬가지로 생물학이 각 생물의 복잡성 모두를 다루어야 한다 고 생각하는 것은 잘못이다. 모든 과학자들과 마찬가지로 생물학 자들도 과학법칙과 이론으로 통합될 수 있는 중요한 특성을 생물 체에서 찾아내려고 노력하고 있다. 적절한 특성을 찾아내고 그것 울 적절한 방식으로 결합시키는 것이 이론과학자들의 주된 임무 이다. 예를 들어 전화에서 말한다면 중요한 것은 무엇인가? 한 종 에 속하는 생물 개체의 숫자인가, 집단의 밀도인가, 아니면 동일 지역 내에 있는 여러 종들(연구하고 있는 종을 포함하여)의 숫자 인가 등과 같은 것이 될 것이다. 제 2 장에서 살펴본 것처럼 생물학 에서 가장 중요한 문제 중의 하나는, 집단의 유전적 구성을 구명 하는 데 필요한 정확성의 정도란 것이 전화법칙을 만둘 필요도 없을 만큼 커야 되는가 하는 것이다.
과학에서의 한계 조건의 역할 폴라니는 〈 기계와 생명은 둘 다 물리학이나 화학적 법칙으로 환원될 수 없다〉고 하였다 .5) 기계는 〈 그 전체가 고등과 하등- 두 개의 서로 다른 원리 아래서 움직이는데, 고등한 원리는 기계의 디자인이고 하등한 원리는 물리-화학적 과정을 수행하는· 장치들 이다. 마찬가지로 생물도 두 개의 원리에 따라 작동하는 시스템이 라고 볼 수 있다. 이 경우, 생물의 구조는 각 기관들로 하여금 그
5) Life's Irreduci bl e St ru ctu re, Scie n ce 160(1 9 68), pp. 1308- 1312. Scie n ce 162(1 9 68), p. 410ol] 실린 로널드 기어 Ronald Gi er e 의 회답과 사이먼 S i mon(1971) 도 함께 참조할 것.
기능을 수행하여 물리-화학적 과정을 전행시키도록 하는 한계 조 건이다〉라는 것이다. 폴라니가 상위단계 원리에 의하여 하위단계 원리나 현상이 진행된다고 한 것은 한 수준에서의 구조가 그보다 더 하위단계에 작용하는 법칙에 대한 한계 조건을 부여한다는 뜻 이라고 생각된다. 그는 또한 〈한계 조건은 하위과정의 총체를 포 괄하는 것이기〉 때문에 기계의 구조나 생물의 형태와 같은 구조 룰 지닌 시스템은 〈물리, 화학적 법칙을 초월하고 있다 〉 고 주장 한다. 더군다나 잘못 이름 붙여전 용어를 너무 막연한 의미로 사용했 기 때문에, 그는 인과적인 결과를 모두 디자인, 어휘, 문법, 정보 등의 탓으로 돌리면서 그의 원리라는 것을 구체화하려 했던 것 같다. 때로 그의 원리는 하위단계의 현상이나 법칙을 이용한다 . 예를 들면 어떤 곳에서 그는 기계를 만들 때 〈우리논 자연의 법 칙을 이용한다〉고 말하고 있다. 또 다른 곳에서는 〈기계의 구조〉 가 〈자연의 힘〉을 이용하고 있다고 말하고, 또 다른 곳에서는
〈 기계의 디자인의 원리 〉 가 그러한 자연의 힘을 이용한다고 쓰고 있다. 그는 한계 조건을 설명하기 위해 갈릴레오의 실험에 있어서 의 경사면의 기울기를 그 예로 들고, 기울기의 선택은 역학법칙과 관계가 없다고 하였다. 또 특정한 경우에 있어서의 천체의 위치와 운동량이 태양계에 대한 천체역학의 한계 조건이 된다고 하였다. 그러나 불행하게도 그가 든 예 중에서 설득력 있게 한계 조건이 라 불릴 수 있는 것은 별로 없다. 만일 경사면의 기울기가 한계 조건이라면 이러한 유추를 아무리 확대시킨다 해도 마음이라는 것까지 포함시킬 수는 없을 것이다. 그는 그의 유추를 기계에서 시작해서 생물의 형태로 전전시켜가긴 했지만 그의 주된 관심은 정신의 차원 s t a t us of mind 에 있었던 것 같다. 앞에서 언급했던 다른 것들과 마찬가지로 〈 정신은 신경생리학적 기작을 이용하는 것이지 그러한 기작에 의해 결정되는 것은 아니다〉라고 그는 말 했다. 그러나 마음이 어떻게 신경생리학적 기작에 대한 한계 조건 으로 작용할 수 있는지 이해하기란 대단히 어려운 일이다. 폴라니는 기계와 생물 사이의 유추는 그들의 기원이 다르기 때 문에 다소 문제가 된다는 것을 인식하였다. 기계를 만들고 그 구 조를 결정하는 것은 인간이다. 그러나 생물의 구조는 전화과정을 통해 생긴다 . 그도 현재 유행하는 전화론들이 많은 결점을 지니고 있다는 것을 알고 있고, 따라서 구조가 우연적인 변이나 자연선택 울 통해 생기는 것이 아니고 〈그렇게 극도로 잘 유지되고 있는 질서 모두가 원자들의 우연적 배열에 의해 생긴다고 믿기는 어렵 다〉고 하였다. 또 그는 종합 전화론은 〈질서원리 orderin g pr in - c ip le 라는 것으로〉 보완되어야 하며, 〈생명을 탄생시킨 질서원리 라는 것은 개방되고 안정된 시스뎀이 지니고 있는 참재력이다〉라 고덧붙였다 .6)
6) Polany i, Personal Knowledge ( Ch ica go : Un ive rsit y of Ch ica go Press, 1958), p. 35.
전화론을 풍자화시키는 경우가 종종 있는데 가장 일반적인 것 중 하나는 마치 많은 공기돌들이 빈 공간 여기저기서 튀어오르듯 이, 원자가 우연히 배열되어 생명이 기원되었다는 것이다. 그러나 원자는 결합하는 성질을 지니고 있고 또 종류에 따라 특정한 방 법을 통해 결합된다. 이것은 지극히 평범한 상식에 속한다. 따라 서 이러한 사실을 무시한 생명의 기원론으로는 전화의 종합이론 이나 기계론적 세계관에 맞설 수가 없다. 폴라니가 말하는, 생명 을 탄생시킨 질서원리라는 것은 아마도 화학원소들의 결합성질일 것이다. 폴라니의 주장에는 부적철한 데도 많지만 그것이 형식화되기만 한다면 그가 내린 결론은 대단히 중요한 것이라 생각된다. 생명계 가 유전자형과 표현형이 서로 구분될 수 있을 만큼 복잡한 수준 으로 진화되어 온 이상, 미래는 유전자형의 구성, 표현형의 구성 그리고 그 둘 사이의 복잡한 관계 등에 의해 결정될 것이다. 돌 연변이의 발생은 그러한 것들과 무관할지도 모르지만 돌연변이의 정착은 무관하지가 않을 것이다. 그러나 폴라니가 새로운 어떤 것 을 말하고 있는 것은 아니다. 다윈에서 메이어에 이르기까지 생물 학자들은 생식을 위해서는 게놈g enome 과 페놈p henome 모두에 있어 어떤 일정한 체제가 유지되어야만 한다는 것을 강조해 왔다. 진화에 대한 연구를 처음 시작했을 때 피셔 F i sher 와 할데인 Hal dan~ 한 번에 한 개의 유전자 좌위만을 고려할 수밖에 없었으 면서도, 〈전화모델에서 각 유전자 좌위를 하나의 독립된 현상으 로 다룰 수는 없다. 하나의 유전자가 모든 형질에 영향을 준다고 주장한다면 그것은 너무 과장이 되겠지만 그래도 그것이 초기
멘델주의지들의 신념보다는 더 전리에 가까울 것이라는〉 사실을 잘 인식하고 있었다 .7)
7) Ernst May r, in Math e mati ca l Challeng e s to the Neo-Darwi ni a n Inte r p re ta tion of Euoluti on (Ph ila delph ia : The Wi st a r Insti tut e Press, 1967), p. 53.
마지막으로 폴라니는 과학자들의 관심은 항상 과학법칙에만 국 한되어 있는 것이 아니라는 중요한 사실을 알게 되었으며, 때로 그 자신도 한계 조건 그 자체에 더 관심이 있었다. 〈우리가 냄비 에 수프롤 끓일 때 우리가 관심을 두는 것은 냄비가 아니라 수프 이며, 마찬가지로 시험관에서 어떤 실험을 할 때 우리의 연구대상 은 시험관에서 일어나는 반응이지 시험관은 아니다. 그러나 장기 는 그 반대이다. 전술은 장기의 규칙에 의한 몇 가지 방법에 의 해 그 한계가 정해전다. 그러나 우리의 관심은 장기 규칙의 정석 에 해당하는 몇 가지 방법이 아니라 그 한계, 즉 전체로서의 전 술 자체에 있다 . 〉 진화학자들도 법칙보다는 진화 과정의 한계 조 건에 더 자주 관심을 두고 있다. 제筑}에서 이미 살펴본 것처럼 생물학자들이 사용하는 법칙, 그 중에서도 계통발생적 발달을 재 구성하는 데 도움이 되는 법칙들은 그 자체가 너무 빈약해서 오 히려 자료에 의해 대부분의 설명이 이루어진다. 태양계처럼 폐쇄 된 시스템의 경우에는 천체역학 법칙과 어떤 특정 시간에 있어서 여러 천체들의 위치와 운동량만 알면 시스템의 미래 상태를 예측 할 수 있다. 따라서 우리의 관심은 법칙에 그 초점이 맞춰진다. 그러나 생물이나 종 s p ec i es 처럼 부분적으로 개방되어 있는 시스 템의 경우에는 어떤 돌연변이가 일어날지, 그 돌연변이가 언제 어 떤 순서로 일어날지, 이 돌연변이들의 표현형에 영향을 주는 요인 들이 어떻게 변화될지 등을 모두 알아야 한다. 그러나 현존하는
생물학 이론, 지질학 이론, 기상학 이론 등이 총동원된다고 해도 어떤 특정 시간에 있어서의 이 변수들을 구체화시킬 수는 없을 것이다. 그렇기 때문에 부가적인 자료가 철대적으로 필요하게 된 다. 그래서 전화론에 있어서의 한계상황을 구체화함으로써 전화에 역사적 이야기로서의 특성을 부여할 수 있어야 하는 것이다(제 3 장을 참조할 것). 그렇게 한계 조건에 많이 의지하는 것이 전화적 설명을 약하게 만든다고 느끼는 사람은, 대부분의 물리학자들도 물리학에서 가장 기본적인 개념 중의 하나인 시간의 방향을 현재 와 같은 우주 팽창을 일으키게 된 한계 조건으로 거슬러 올라가 서 추적하고 있다는 것을 상기해 볼 필요가 있다.
켜근 1 로'-- 지금쯤은 이미 생물철학의 문제점들이 어렵고, 서로 아주 밀집 하게 연결되어 있다는 것을 따로 언급해야 할 필요가 없게 되었 울 것이다. 생물학자들은 생물학적 시스템의 복잡성과 그에 따른 고유성, 여러 단계로 분리된 그리고 목적론적인 체제, 그리고 생 물학에 있어서 역사적인 고찰의 중요성 등을 강조해 왔다. 한 시 스템이 목적론적으로 행동하기 위해서는 일정한 수준의 복잡성을 획득해야만 한다• 인공물을 제의한 모든 시스템은 선택과정을 통 해서만 이러한 수준의 복잡성을 지니게 된다. 앞에서 나는 생물학 자들의 그러한 주장이 무엇을 의미하는지를 설명하였다. 논의된 문제둘이 이해하기 쉽다거나 내가 그것들을 명료하게 설명했다고 생각하지는 않는다. 다만 생물학의 가장 중요한 철학적 문제들을 집약하고, 최소한 다른 사람들이 내가 이 짧은 책에서 제시할 수 있었던 것보다 더 나은 해결점을 찾을 수 있도록 충분히 명료하 게 그 문제들의 관계를 보여주기를 바랄 뿐이다.
참고문헌 Ap ple runan, Ph ilip, Darwi n, A Nort on Cri tica l Edit ion . New York : W. W. Norto n & Co., 1970. Beck, Wi lliam S., Modern Scie n ce and the Natu re of Lif e. Garden Cit y, N. J. : Anchor Books, 1961 . Beckner, Morto n , The Bi ol og ica l Way of Thoug ht. Berkely : Un ive rsit y of Calif or n ia Press, 1959. Blackburn, Robert T., ed., Inte r relati on s : The Bio l og ica l and and Phys ic a l Scie n ces. Ch ica go : Scott , Foresman and Co., 1966. Bossert, W. H., and E.O. Wi lso n, A Pri m er of Pop u lat ion Bio l og y. Sta mfor d, Conn. : Sin a uer Assoc., 1972. Ca nfiel d, Jo hn V., Purp o se in Na ture . Engl e wood Cli ffs, N. J. : Prenti ce -Hall, Inc., 1966. Carlson, Elof Axel, The Cene : A Cr itica l Hi sto r y . Ph ila delph ia : W. B. Saunders. Comp n ay, 1966 , ed., Modern Bi ol og y: its Concept ua l Foundati on . New York : Brazil ler, 1967. Dobzhansky , Theodosiu s , Geneti cs of the Evolut ion ary Process. New York : Columbia Un ive rsit y Press, 1970. Ellega rd, Alvar, Darwi n and the General Reader: The Recep tion of Dar-win ' s Theory of Evoluti on in the Briti sh Peri od ic a l Press, 1859-1 872.
Goth e nburg : Got eb orgs Univ e rsit et s Arskrif t, 1958. Gabrie l, Mordecai L., and Sey m our Fog el , eds., Great Exp e rim ents in Bio l ogy . Eng le wood Clif fs, N.J. : Prenti ce -Hall, Inc., 1955. Ghis e lin , Mi ch ael T., The 'Irium p h of the Darwi ni a n Meth o d. Berkeley : Univ e rsit y of Cali for nia Press, 1969. Glass, Bentl e y, Owsei Temkin , and Wi lliam L. St ra us, Jr. , eds., Fore-runners of Darwi n: 1945-1859. Balti mo re : The Jo hns Hop k in s Press, 1959. Goudge , T.A., The Ascent of Lif e. Toronto : Un ive rsit y of Toronto Press, 1961. Hemp el , Carl. G., Aspe cts of Scie n ti fic Exp la nati on . New York : The Free Press, 1965. Hi nt i kk a, Ja akko, ed., Sy n th e se(vol. 20, no. 2, devote d to philo sop h y of bio l og y), Aug u st 1969. Hull, David L., ed., Darwi n and Hi s Crit ics : The Recept ion of Darwi n' s Theory of Evoluti on by the Scie n ti fic Communit y. Cambrid g e : Har -vard Un ive rsit y Press, 1973. Levin s , Ri ch ard, Evoluti on in Chang ing Envir o nments . Prin c eti on : Prin - ceto n Un ive rsit y Press, 1968. Lovejo y , Arth ur 0., The Great Chain of Bein g . New York : a Harpe r Torchbook, 1936. MacArth u r, Robert H., Geog ra ph ic a l Ecology . New York : Harpe r and Row, 1972. Mayr , Ernst, Anim al Sp e cie s and Evoluti on . Cambrid g e : Harvard Un ive rsit y Press, 1963. Mendelsohn, Everett , ed., Jo urnal of the Hi st o r y of Bi ol og y( vol. 2, no. 1, devote d to hist o ry and philo sop h y of bio l og y), Sp r in g 1969. Munson, Ronald, ed., Man and Natu re . New York : Delta Books,
1971 . Olby , Robert, Orig ins of Mendelism . New York : Schocken Books, 1966. Pete r s, Jam es A., ed., Classic Pape rs in Geneti cs . Eng le wood Clif fs, N. J. : Prenti ce -Hall, Inc., 1959. Provin e , Wi lliam B., The Orig ins of Theoreti ca l Pop u lati on Geneti cs . Chic a g o : The Univ e rsit y of Chic a go Press, 1971 . Ruse, Mi ch ael, Phil o sop hy of Bi ol ogy . London : Hutc h in s on's Un ive r-sit y Lib r ary, 1973. Sim on, Mi ch ael A., The Matt er of Lif e. New Haven : Yale Un ive rsit y Press, 1971 . Sim p so n, George Gay lo rd, This Vi ew of Li fe. New York : Harcourt, Brace & World, Inc., 1964. Smart, J.J. C., Phil o sop h y and Scie n ti fic Realism . London : Routl e dg e & Keg an Paul, 1963. , Betw e en Scie n ce and Phil o sop h y. New York : Random House, 1968. Vorzim mer, Pete r , Charles Darwi n. The Years of Contr ov ersy. The Or i- gin of Sp e cie s and its Criti cs. Phil a delph ia : Temp le Un ive rsit y Press, 1970. Wats o n, Jam es D., Molecular Bi ol og y of the Gene. New York : W.A. Benj a min, Inc., 1970. Wi lliam s, G.C. Adapt at i on and Natu r al Selecti on . Prin c eto n : Prin c eto n Un ive rsit y Press, 1966.
역자해제 현대 자연과학은 무서운 속도로 발전하고 있다. 그 발달의 속 도는 최근 삼사십 년 동안에 쌓인 자연과학적 지식의 양이 그때 까지 인류가 전 역사에 걸쳐 쌓아 놓은 양과 맞먹을 정도일 것이 라고 하는 표현으로도 짐작할 수 있을 만큼 실로 정산을 차릴 수 없을 정도이다. 자연과학자들은 그 탐욕스러운 지식욕과 경쟁심으 로 자연의 신비에 도전하여 이와 같은 오늘날의 과학 체계를 구 축하고 방대한 지식을 문헌에 담아놓았으나, 한편으로는 생계를 위한 직업화로 전락(?)한 이 자연 탐구에 지나치게 몰두한 나머 지 마치 커다란 숲 속에서 나뭇잎 하나하나를 분석하는 데만 온 정열을 기울이고 숲 전체를 보는 것을 잊고 있으며 또 볼 줄도 모르고 볼 필요성도 느끼고 있지 못하다는 비판을 받기도 한다. 더욱이 국제경쟁이 치열해진 최근에 이르러 과학은 더 이상 종래 와 같은 인류 공동 소유의 개방적 지식이 아니고 자국의 국제경 쟁력 제고를 위한 수단으로 인식되어 기술과 함께 한 국가와 민 족의 생존을 위한 무기로 변하고 있다. 이에 따라 과학자들은 자 신의 국가와 민족의 소명에 따라 거의 맹목적이라고까지 할 수 있을 정도로 과학의 연구에 더욱 박차를 가하고 있어서 과학 발
달의 속도는 날이 갈수록 빨라지고 있다. 가위 제동장치가 풀린 수레가 비탈길을 곤두박질치고 있는 것과 흡사하다고 할 수 있다. 이런 이유로 과학에 철학이 필요하다는 말들이 오래전부터 회자 되어 왔다. 이와 같은 현상은 자연과학의 한 분야인 생물학에 있어서 더욱 현저하다. 2 안1 ] 기 후반은 생명과학의 시대라고 할 만큼, 195 0\반대 이후 분자생물학의 대두로 확장된 생명에 관한 지식 영역은 생명 기계론과 생명물질론을 일반화시키고 있고 인공 생명의 합성까지 바라볼 수 있게 하고 있다. 그리고 생명과학의 이러한 발달은 필 연적으로 그 응용 분야인 의학, 농학, 약학 등에도 파급되었고, 나 아가 생물공학 혹은 생물산업의 발전으로 이어지면서 인류의 3 대 난제라고 하는 식량난, 암, 공해롤 단번에 해결할 것 같은 보랏빛 미래상을 우리 앞에 펼쳐 주고 있다. 그러나 이러한 낙관론에 대 하여 비판의 소리 또한 높다. 물질과학화된 생물학으로 말미암아 만연된 생명물질론은 생명 경시 사상을 낳고, 전통 가치관의 붕괴 와 물질만능주의의 팽배를 가져온다고 한다. 또한 내세(來世)에 대한 부정은 현세(現世)에서의 향락주의를 부채질할 뿐이라고 한 다. 무엇보다도 과연 현대 분자생물학이 인류의 3 대 난제를 해결 할 수 있느냐라는 의문과 함께 해결은 커녕 해결하는 과정에서 더 해결하기 어려운 난제만 만들어낼 것이라는 우려도 대단히 높 은 것이 사실이다. 이러한 부정론의 측면에서 현대 생물학에는 철 학이 없다, 생명의 물질론적 탐구에 앞서 모든 지식인이 공감하는 철학이 필요하다는 주장이 적지 않다. 그러나 문제는 현대의 대부 분의 생물학자는 철학에 대한 관심이 많지 않은 것 같고 반면에 대부분의 철학자는 이제 극도로 전문화된 현대 생물학에 대한 지 식을 그들에게 필요한 양만큼 가지고 있지 않다는 데 있다. 그래
서 다론 과학 분야에서도 사정은 비슷하겠지만 생물학에서는 생 명과학철학이라고 할 만한 철학의 형성이 어려운 것이 아닌가 생 각된다. 뿐만 아니라 생명과학의 철학이라고 하여도 그 철학을 하 는 사람의 입장에 따라 방향 혹은 목표가 여러 갈래일 수가 있을 것이다. 따라서 한마디로 생명과학철학이라고 하여도 그 종류가 여러 가지 있을 수밖에 없다. 여기에 소개하는 이 책은 위와 같이 여러 갈래로 있을 수 있는 생명과학철학의 하나이다. 이 책은 현대 생물학의 발전에 대하여 공과론을 펴는 것을 목적으로 하고 있는 것이 아니고, 또 현대 분자생물학적 생명관을 철학적으로 분석 비판하는 것을 목적으로 하고 있는 것도 아니며, 생물학적 이론과 설명 양식이 과학의 본 질에 대한, 그리고 생물학과 나머지 다론 과학 분야와의 관계에 대한 여러 논쟁에 어떤 해결점을 줄 수 있는가를 중점적으로 다 루고 있다. 전통적인 방식으로 과학을 분류할 때나 이론적인 발달 과정으로 볼 때나 생물학은 물리학과 사회과학 사이에 놓여 있기 때문에 이러한 목적에 아주 적합한 분야라고 원저자는 말하고 있 다 . 이 책의 곳곳에서 논급되고 있지만 현대 생물학을 철저히 환 원론적으로 다룬다면 물리학과 다른 접이 없어전다. 따라서 저자 가 생물학을 물리학과 사회과학 사이의 중간적 성격의 학문으로 디두는 것은 아직은 생물학을 고전적인 의미로 보고 있기 때문이 라고 생각된다. 저지는 과거 과학철학의 여러 문제들이 너무 자주 순전히 과학적인 추상이나 아니면 순전히 물리적 이론의 관점에 서 다루어져 왔다고 보고, 이 책에서는 이러한 방법을 떠나 순전 히 생물학에서 유도된 몇 가지 이론을 생물학적 관접에서 다루고 있다. 그러나 이 책의 전부를 통하여 역자가 느끼는 것은 대체로 고전 생물학적 관점에서 중요시된 사항들을 중심으로 문제를 제
기하고 있고 그에 대한 해답은 시도하고 있지 않다는 것이다. 원 래가 이런 문제는 말하자면 주관적인 문제라고도 할 수 있어서 명쾌한 객관적 해석 또는 해답이란 기대하기가 어렵기 때문일 것 01 다. 제 1 장에서는 현대 생물학 최초의 분석적 이론 중 하나라고 할 수 있는 멘델 유전학이 최근의 분자유전학으로 환원되어 물리화 학적 언어로 표현될 수 있으려면 어떤 전제가 필요하며 또 어떤 논리가 필요한가 그리고 그것이 과연 가능한가에 관하여 논술하 고 있다. 또한 유전자를 적절하게 정의하는 문제, 죽 유전지는- 무 엇인가, 유전자는 기능적으로, 구조적으로 혹은 역사적으로 정의 되어야 하는가 하는 문제들이 논의된다. 저자는 이 장에서 위와 같은 환원은 전통적으로 정의된 한 과학 분야 내에서만 일어났는 데 지난 이삼십 년 사이에 서로 다른 두 분어: 사이에서 이러한 환원이 일어났다고 지적하고, 그것은 생물학자들이 전형적인 생물 학적 이론인 멘델 유전학을 물리-화학적인 용어로 된 이론인 분 자유전학으로 환원한 것이라고 하였다. 이 변화는 과학에 있어서 의 대체라기보다는 차라리 혁명이라고 평가하고 있다. 그러나 이 책이 출판된 20\ '! 전과 현재의 생물학은 또 다르다. 197 떠대 초만 하더라도 아직 인간의 손으로 유전자를 직접 조작 하여, 자연 상태에서는 일어날 가능성이 없는 재조합 DNA를 만 든다는 것은 이론적으로는 충분히 가능하다고 인식되고 있기는 했어도 현실적으로.는 그리고 일반 대중에게는 상상의 세계에 불 과하였다. 그러나 바로 그 무렵부터 시작하여 급격히 전전된 분자 유전학적 기술은 재조합 DNA 를 수없이 만들어내게 되었는데, 이 재조합 DNA 는 말하자면 부분적이긴 하지만 인공적으로 만들어 전 인조생명이라고 할 수 있는 것이다. 이것은 비단 분자유전학
분야에서 일어난 현상만은 아니고 생물학의 여러 다른 분이에서 도 그 변혁은 마찬가지여서, 생명현상의 핵심적 원리들이 속속 분 자론적으로 해명이 되고 있고 물리-화학적인 용어로 서술되어 가 고 있다. 저자가 이 책을 집필할 당시는 이러한 변혁이 막 막을 올리기 시작한 무렵이었다고 할 수 있는 것이다. 제 2 장은 멘델 유전학과 더불어 현대 분자생물학의 초석적 이론 이라고 할 수 있는 전화론에 대한 고찰이다. 진화의 요인 혹은 원동력이 무엇인가에 관해서는 현재도 어떤 뚜렷한 해답이 있는 것은 아니다. 아직도 전화라는 현상 자체가 사실인가, 전리인가에 대해서도 논쟁이 있는 형편이고, 또 전화라는 개념을 실험으로 입 증하기도 어려운 것이 현실이다. 물론 거의 모든 자연과학자들은 전화를 믿고 있다. 사실 생물은 진화하였고 또 전화하고 있다고 가정함으로써만 오늘날의 이 자연의 생물의 다양성을 가장 잘 설 명할 수가 있는 것이다. 이것은 중력의 존재를 직접 입증하지 않 더라도 질량을 가전 모든 물체에는 중력이 있다고 가정함으로써 물체의 닉하 운동을 가장 잘 설명할 수 있는 것과 같다. 그러나 그렇다 하더라도 무엇이 전화를 일으키는가에 관해서는 아직도 가설의 단계를 벗어나지 못하고 있다. 적자생존을 통한 자연선택 이 그 1 차적 요인이라는 데는 거의 이의가 없으나 적자생존의 가 장 큰 조건은 무엇인가, 개체변이인가 또는 돌연변이인가 등에 관 해서는 논란이 많다. 다윈은 진화의 가장 중요한 요인으로 개체변 이를 들고, 이 변이는 아주 조금씩 그리고 서서히 일어나는 것이 라고 하였는데, 저자는 이러한 견해에는 물론 동의하고 있으나 다 만 한 종의 구성원들 사이에서 예사로 관찰되는 변이들 모두가 똑같이 전화의 재료가 된다는 그의 생각은 잘못이라고 주장하고 있다• 그 이유로 그는 개체간의 차이는 대부분 재조합과 환경의
영향 때문이라는 것을 다윈은 몰랐기 때문이라고 한다. 이 점에 관해서는 획득형질은 유전되지 않는디는- 현대유전학적 개념으로 보아 옳은 해석이라고 할 수 있다. 진화론의 해석에 있어서 이 책이 제기하고 있는 논란의 하니는· 전화론이 과연 과학 이론이 될 수 있느냐 하는 점이다. 일반적인 정의에 따른다면 과학 이론이란 과거와 현재의 상태를 설명할 수 있어야 할 뿐만 아니라 그것으로 미래를 추론할 수 있어야 한다 는 것이다. 현재의 현상만 설명할 수 있고 미래에 대하여 어떤 추론도 할 수 없다면 그것은 과학 이론이 될 수 없고 또 아무런 쓸모도 없다는것이다. 마찬가지 논리로 종(種)이라는 개념에 대해서도 그 과학적 법 칙성에 의문이 제기된다고 저자는 말하고 있다. 종은 반복되지 않 는다. 전화론적으로 보면 종은 시간적으로 변화되어 온 계보이다. 종은 항상 유동적으로 변하고 있으며 이 변화는 설혹 환경이 원 상태로 환원되었다 하더라도 원래의 종으로 되돌아간다고는 결코 생각할 수 없다는 것이다. 전화는 단 한 번의 현상이라는 것이다. 그런데 과학법칙에는 반복성이 있어야 한다. 따라서 진화론적인 종은 과학법칙의 범주에 들어갈 수 있는 것이 아니라는 것이다. 물론 이러한 논리 전개가 현대 생물학의 이해와 발전에 얼마만큼 도움이 될 것인가에 대해서는 의문이 적지 않을 것이다. 전화의 비반복성, 또 종의 비반복성은 그것이 사실이든 아니든, 그래서 과학적 이론으로 취급받든 아니든, 그것은 어떻게 보면 언어의 장 난에 지나지 않을런지 모른다 . 그렇지 않으면 과학적 용어들의 정 의부터 다시 다듬어야 할런지 모를 일이다. 제筑텨서는 주로 전화론을 대상으로 하여 생물학적 이론과 생 물학적 법칙에 관하여 논술하고 있는데, 생물학에서의 인과법칙,
계통발생과 발생법칙, 역사법칙, 과학적 설명과 포괄법칙 모델 등 을 해설하고 있다. 그리고 생물학에는 과학적 이론이나 과학적 법 칙이 존재하지 않는다고 하는 일부의 주장에 대하여 그 근거를 찾고 그에 대한 반론을 시도하고 있다. 제緖授든 목적론에 관한 서술인데, 여기서는 주로 목적론적 체계 를 적절하게 정의하는 문제를 다루고 있다. 목적론적 체계란 무엇 인가, 이것은 기능적으로, 구조적으로 또는 역사적으로 정의되어 야 하는가 등등이다. 저자의 견해에 의하면, 목적론적 서술보다는 인과론적 서술이 항상 많은 정보를 담고 있기 때문에 재구성되어 야 하는 것은 언제나 목적론 쪽이라고 한다. 죽 목적론적 이야기 를 순수한 인과론적 용어로 재구성해야 한다는 것이다. 이 점에 관해서는 대부분의 생물학자도 이의가 있을 수 없을 것이기 때문 이다. 지금 이 시점에서 생기론(生氣論)을 신봉하는 생물학자는 없을 것이기 때문이다. 저자는 이 장의 말미에서, 인과론적 전술 과 목적론적 전술은 모두 부분적으로 개방되어 있는 시스템을 묘 사하는 데 사용되고 있으나 그 개방의 형태는 서로 다르다고 지 적하고, 인과론적 전술에서는 그 계가 수많은 요인에 의하여 모든 방향으로 얽혀 있는 데 비하여 목적론적 계에서는 그 계가 끊임 없는 에너지의 교환과 부분적인 변형에 의해 유지되고 있다고 한 다. 그리고 순수한 형이상학적 이론으로서의 목적론은 오랜 세월 동안 과학을 위해 아무런 역할도 하지 않았다고 서술하고, 그럼에 도 불구하고 아직도 많은 생물학자들이 목적론적 언어를 사용하 고 있다고 비판하고 있다. 그러나 저자의 이러한 비판은 다소 과 장된 점이 없지 않다고 생각된다. 물론 아직도 생물학적 문헌에는 〈무엇은 무엇을 하기 위하여 이런 기능을 지니고 있다〉는 식의 목적론적 서술이 많은 것은 사실이다. 그러나 이것은 반드시 목적
론적 견해에서 그렇게 서술했다기보다는 서술의 표현상의 편이에 서 생긴 문제일 뿐인 경우가 대부분이 아닌가 생각되는 것이다. 제 5 장은 생체론(生體論)과 환원론에 관한 논설이다. 생체론은 요즈음은 별로 거론되지 않는 생명관이지만, 오스트리아 태생의 버트란피 Ludwi g von Ber t ala ffy (l901- )에 의하여 1950 년경 제창 되었던 학설로서, 생기론을 배제하고 있으나 그렇다고 철저한 기 계론적 입장도 아닌 다소 모호한 생명관이라고 할 수 있다. 이 설에 의하면 생물체의 구성성분이나 반응 또 시간적 요인 등을 분석하는 것은 철대적으로 필요한 일이지만, 그러한 분석만으로는 생명을 이해할 수 없다, 생명이라는 것은 하나의 계 (系)이고 이 계라는 것은 수많은 구성요소들의 복합적인 기능의 총체적 산물 이어서 구성요소들의 물리화학적 법칙성만으로는 설명이 되지 않 는 한 단계 높은 차원의 어떤 힘에 의하여 성립되는 것이다, 따 라서 전체를 파악하지 못하고 부분만의 분석으로는 생명을 이해 할 수 없다는 것이다. 우리는 현재 생명의 물질적 본질을 상당히 알고 있는 것 같으면서도 일면 전혀 알지 못하고 있다고도 할 수 있다• 그것은 우리가 알고자 하는 것이 도대체 무엇인가, 무엇을 알고자 하고 있는 것인가 하는 것 자체가 애매하기 때문이기도 할 것이다. 따라서 우리에게는, 생기론은 배제된다 하여도 생체론 과 기계론은 여전히 양립된 논쟁거리로 남게 된다. 물론 기계론적 인 〈입장〉이란 점에서는 양자가 본질적으로 같다고 할 것이다. 따라서 이 책의 저자가 말하고 있는 것처럼 이들 모두가 동일한 실체의 한 부분을 이야기하는 것인지도 모른다. 그래서 저자는 다 음과 같이 말하고 있다. 〈여러 가지 분석단계에서 나타나는 수많 은 현상을 설명하는 수많은 이론들이 있지만 현대로서는 그것들 이 하나의 통합된 이론으로 조직화되지는 못하고 있다. 상당한 우
연성이 여러 분석단계에서 일어나는· 현상들 사이에 존재하며, 어 떤 경우에는 그러한 분석들이 서로 양립되지 않는 경우도 있다. 하지만 기계론적 환원론에 대한 논쟁 중의 하나는 조만간 해결될 것이고 과학자들은 하나의 통합이론을 만들 수 있을 것이다 . 그러 한 초이론이 가능하다면 그것은 분명히 현대 생물학의 이론과는 다를 것이며 또 현대 물리학과도 다를 것이다.〉 생체론에 대한 저자의 이러한 전망이 현실화될 것인지는 아직 아무도 모른다고 밖에 할 수 없다. 가령 그렇게 된다면 그것은 현대 과학에 있어 서의 일대 혁명적인 사건이 될 것이다. 물론 그렇다고 해서 현대 과학의 모든 법칙성에 변동이 생기는 것은 아니다. 다만 전화론의 수십 배의 영향력을 가전 새로운 이론이 현재의 과학에 보태지는 것 01 다. 이 해제의 서두에서도 썼지만, 이 책은 문제의 제기에 주목적 이 있고 그것의 해결 또는 종합 통일에 목적이 있는 것 같지는 않아 보인다. 생물학적 이론에 대한 철학적 해석 또는 견해라는 것이 원래가 주관적인 문제이기 때문일 것이다. 따라서 독자들~ 이 책을 읽고 이 책이 제기하고 있는 문제들을 각자의 주관대로 해석하고 논리구성을 해볼 수 있을 것이다.
기 가로드 75 가상디 258 갈리 I89 갈릴레오 객, 29-3 1, 258, 275 갈톤 III, II2 감수분열 37-3 9, 83, I26, 2II 개체 roo, 102-IOS, II7, I27, I36, I37, I40- I4 3, I 紅 I62, 230, 273 개체군 I42 개체발생 I54-I 56, I57( 주) 게놈 43, 47, 76, I27, I67, I79, 230, 2 야, 276 격세유전 n8 결정론적 인과율 9I 경험적 일반화 I55, I62, .I63 계통 I57-I 65, I72, I95 계통발생 106, I54, I57, I651 I67, I7 헉 I95, 202, 203, 277 고유성 I96-2 03, 270-2 73 골트슈미트 SI, 53, III 공동변화 239 공우성 49, 56, 76, 87 과정 의 법 칙 104, 105, I46, I49, I50, 158, I69, I75, 239, 24I, 248 관찰적 용어 30, 31 교배 42, 55, II5, I2I, 122, I60 교차 39, 77-8 0, 126, I72
교차연계법칙 I57-I65, I68 구성론 261-266 굿지 196-l 셈, 200, 201( 주) 궁극 원인 229 그레그 132 그렌 251, 258, 259 그륀바움 186, 193, 196 근접 원인 229 기계론 250-2 52, 257, 258, 266 기능 단위 77-7 9, 134 기능적 등가성 234, 238-243 기어 274( 주) L 낫세포빈혈증 138-1 40 네겔 70, 220, 232, 235 논리적 경험주의 18, 35, 58, 90, 92-95 논리적 해석 180 뉴클레오티드 60, 77, 78, 265 뉴턴 24, 29-3 1, 72, 124, 136, 139, 195, 253, 273 C 다면현상 44, 49, 57, 77 다윈 27, 28, 36, 97, 98, 106, 108, 109, 110, 112-116, 118, 119, 124-
126, 135, 136, 145, 242, 276 다윈 서브클랜 135, 165 닥케 166 단계통 165, 198, 202 대 체 27-3 0, 32, 69, 92-9 6 대칭명제 182, 185-196 델브뤽 58, 59, 175, 266, 269 도브잔스키 55, 173, 175, 200 독립의 법칙 51 돌로 166, 167, 202 돌연변이 50, 54, 74, 77-79, 89, ro6, 112, 120, 121, 127, I28, 137, 228, 276, 277 접 돌연변이 77 동계교배 IJI , 171 드 브리스 III, 112, Il5 드레이 15,23 DNA 44, 45, 54, 57-6 4, 66, 76 - 81, 83, 195, 226, 228, 254, 262, 265 2 라마르크 Io8( 주) 라부아지에 29 라이트 120 라이헨바흐 184 러드너 15, 23, I86 ’ 193, 1¢, 213, 217( 주)
레빈스 I45 레원틴 128, 145, 174, 200 레흐만 2I7( 주) 렌쉬 166 루스 79, 96( 주), 16l, l 야, 165 리보좀 야 □ 말라리아 138, 140 맥스웰 124 맥아더 145 먼슨 217, 226, 232, 235 메이어 79, 1)0, 135, 160, 200, 226, 227, 229, 251, 264-266, 276 메틀러 132 멘델 법칙 28, 36, 51 멘델 비 55, 74, 93 멘델손 128, 264 모건 III, 112 목적론 25, 109, 205-248 목표상태 218-2 20, 222, 223, 225, 226.’ 231 무성생식 160 미바르트 109 밀 242
t:I 바이스만 II6 바커 I5 반성 유전 28 발견의 논리 I6 발생법칙 146, 154-1 56, 165-1 69 배우자 I20, l36 백화중 49, 89 번역 야, 66 베르그만 169, 170 베이컨 객 2 베이트슨 III, 112 베크너 100, 170, 217( 주), 238 벤저 75 변이 40, 107, III, 112, 116-120, 174, 220, •2 21, 273, 275 일차적 변이 220, 221 적응적 변이 220, 221 변종 116 보드 147 보르짐머 112, 115 보어 58, 74, 266-2 68 보편적 법칙 IOI, 181 복잡성 270-273 복제 60, 66, 228 본질주의 108-1 13 분류군 113, 133, 195, 198, 202 불완전우성 54, 56, 86-8 9
불확정성 원리 267-269 브리지만 42 블랙번 226' 229, 269 人 사이몬 18, 21, 22, 252, 274( 주) 산화이론 29, 69 살몬 15 상동 171-1 73, 176 상보성 원리 267, 268, 290 상사 172( 주) 상위 49, 56, 57, 76, 85, 90, 91 생기론 252, 255, 257 생명액체 253-255 생식세포 38, 116, 120, 121 생존 107, 137, 142, 230 생체론 젝 91, 92, 140, 249-2 78 생태적 적소 I28 생합성 경로 55, 57, 66, 76, 80, 88, 105, 155, 178 샤프너 72-7 4, 76, 78-8 0, 83 , 84, 89, Io6, I I2, I20, I27, I28 서브클랜 I35 서술용어 31, 33, 82-92 선기술 I84, I85 선택압력 Io8, n8, I2I, I22, I38, 174, 175 선택이론 229-231, 245
선행 182, 183, 196 선호상태 207, 209, 21 I, 217-2 25, 229, 231, 236, 244 설명 23, 26, 178-1 87 설명력 187, 190 손본 75 순계 171 순수유전 112 쉰트볼프 167 슈뢰딩거 58, 59( 주), 266 스마트 145, 161, 162, I 야, 165, 167 스크리본 152, 189, 196 스터티반트 52 스트릭버거 90 시스- 트랜스 테스트 78, 80 시스트론 78-81, 83 시조새 161 신다윈주의 120 신종 107, I19 심프슨 21, 22, 157, 160, 176, 199, 200, 251, 261-263 。 아리스토텔레스 29, 70, 72, 157, 205, 213 아미노산 62-65, 81-8 3, 148 아인슈타인 31
안달루시안 49, 56, 86-8 8 열성상위 85, 87, 90 우성상위 90 알비노 so, 52, 89, 99 RNA 57, 61, 62, 64-6 7, 76-7 8, 80, 81, 88, 89, 227, 228 리보좀 RNA 야 운반 RNA 야 - 66, 78 전령 RNA 야- 66, 77, 84, 88 양자물리 학 266-2 70, 272 엑스플래난둠 I8I, I87, I93 엑스플래난스 I 紅 183, 184, 193 엘자서 267, 270- 2 73 역기술 185 역사법칙 146, 169-1 78 역사적 재구성 34-3 6 역설명 I85 연소이론 29, 69, 94, 253 열성 48-5 4, 76, 84, 85, 121 열역학 법칙 20, 21, 29 영기 59-64 염색체 36~96, 103, I12, 136, 171, 172, 206, 273 동형접합(염색)체 48-51, 76, 82, 86, I22, m, I38, m 상동염색체 36-3 9, 48, 51, 79, 83, 171( 주) 성영색체 IO3 이형집합(염색 )체 48-5 1, 57, 82,
86, 88, 121, 122, 마, 171 잡종영색체 39 염색체 이상 40 예측 167, 178-1 87 오초아 266-270 오페 론 57, 77, 紅 220 오펜하임 69, 179, l 紅 182 올비 II2 와딩돈 41, 56 와인버그 121 왓슨 42, 54 용불용설 Io8( 주) 우드거 70 우성 48-5 4, 56, 76, 83-8 6, 89, 121 우연 38, 마, 149, 261 위그너 266 위치효과 46, 47, 52, 53, 57, 76 윌리스톤 166 윌리엄스 134-1 37, 145, 228( 주), 245 윌슨 145 윔 사트 2I7( 주), 227, 228 유사분열 37, 38, 60 유성생식 36, 130, 160 유아 II3_ II5 유전암호 62-6 4, 66, 77, 78, 82, 83 유전자 30, 36-짜 98, IOJ , 1311
136, 138-1 40, 153, 171, 172, 206, 212, 215, 253 대립유전자 37, 40, 44, 46, 48- 51, 83, 84, 86-8 9, 121, 131, 138' 139, 171 동의유전자 44, 49, 52 멘델 유전자 47, 78-80, 82 보족유전자 90 복대립유전자 46 복합유전자 57 상위유전자 52, 57, 74, 85, 89, 131 열성유전자 48 우성유전자 48 작동유전자 81 조철유전자 81 유전자좌위 37, 45, so, 89, 276 유전자 풀 140 유전자형 40-4 4, 98, 103, II2, 122, 131, 132, 138, 140, 143, 153 ' 155, 215, 276-2 77 유전적 부동 I3I 유전학: 멘델유전학 25, 27-9 6, III, 120, 138, 145, 函 분자유전학 20, 25, 27-9 6, 97, 145 형질전달유전학 56, 74, 75, 76, 78, 93, 94, 97, 153, 178
의태 객 138 이론적 용어 JO , 162, 253 이론적 존재 I9 이야기식 설명 196-2 03 이중나선 58, 59 인과법칙 146, 149-1 53, 202, 239 1 유전자 /1 형질 42, 50, 81 입자유전 114 x: 자연선택 106, 108, 116, u8, 121, 126, 131, 137, 139-1 44, 177, 228, 2 야, 275 잡종 114, 171 재조합 39, 54, 80, II9 적응 9 헉 107, 128, 141, 168, 174, 178, 240, 246 적응도계수 141-1 43 적응도집합 165, 168 적응방산 192 적자생존 137, 139, 144 전기술 .18 5 전사 60, 66 접합체 38, II5, 156 정 당화의 논리 16, 23 정보이론 226, 227 정상적 과학 27, 28, 35 정신 94, 95, 275
젠킨 II6-I I8 종 24, 30, 31, 100, !02 , 103 , 105-113, 117-119, 123, 124, 126, 129-1 31, 133, 136, 143, 157-162, 164-167, 174, 176, 177, 212, 230, 253, 266, 273, 277 다형적 종 166 보편종 166 생물학적 종 102, 103, 160, 240, 241 원시종 166 자연 종 IIO 진화적 종 160, 162 종분화 99, 106-108, 134, 192, 195 주전원 32, 70, Io8 중복 40, 67-69, 77, 82, 83, 240 진화 67, 68, 97-144, 函 -163, 166, 174, 176 계통적 전화 108 도약진화 108-1 13 진화 양상 123, 126, 137, 138 전화과정 68, !05 , 166 진화단위 !02 , 127, 130, 133, 136, 143, 241 전화론 25, 27-2 9, 45, 97-1 44, 1I58279,’ 0 3,1 1 6 9122 ’’ 1 2 1 16 924, -11 9669, , 119786,, 210729,, ’ 226, 229, 230, 241, 251, 269, 275, 276, 278
고전진화론 97, 162 수학적 진화론 I20 유전학적 진화론 97, 98, !20 종합전화론 97, 98, I26, I45, I62, 275 전화법칙 !OJ, 124, 133, 157, 166- 168, 201-203, 273 진화적 발전 142, 143, 165, 167 집단n8 ’ IO1O2 ’1 ’ 101224 ’ , 1I0235, , IIo287 ’, 111208 ’' 130, I 3 I , I33, 134, 136, 137, 139, 142, 143, l 야, 165, 173, 174, 273 犬 창시자원리 130-1 33, 135, 269 창조론 27, 29 천체역학 104, 129, 130, 218, 277 초우성 49 구 카날화 215 카우프만 270( 주) 캐슬 5I 케메니 ® 케플러 29, 147 코돈 야 -64, 77
넌센스코돈 야 역코돈 65 종말 코돈 8I 코페르니쿠스 32-3 3, 70, 108 코프 I66 과인 I5 쿤 27, 30, 31( 주), 71-73 퀸 70 큐비에 I63 크릭 54 클랜 I35 클러시우스 I24 클론 272, 273 드 테오프라스투스 157 통계역학 21, 29, 94 통계 적 -확 률적 설명 17, 243 툴민 189 포 파이어아벤트 30, 31( 주), 71-73 판게네시스 109, n3-n 6, n8 퍼스 I24 페놈 276 포괄법칙 모델 I6, m8-I 82, I85- 191, 193, 197, 201, 231-244
푸 근 드만 259 포퍼 71-73 풀라니 251, 257, 267, 274, 276, 277 표현형 (질 ) 40-4 4, 49, 53, 56, 74 -76, 78, 84, 85, 88, 90, 91, 98, II2, 140, 153, 157, 162-1 64, 165, 168, 171, 215, 276 프로빈 II2, I20 프톨레마이오스 32-3 3, 53, 70, 108, 255 피드백 닫힌 피드백 209 양성피드백 219 음성피드백 217-2 25, 23 I 인과적 피드백 207, 2II, 217, 219, 223 피셔 I15, 120, 276 굴 하디 121 하디-와인버그 법칙 121-123,131, 138 하이만 157 하이젠베르크 266-268 하인 251 학습이론 226 한계조건 123, 156, 274-278
할데 인 120, 249, 276 합리적 재구성 34-3 6 합목적론 246 항상성 214, 215 핵산 5 헉 59( 주) 핸슨 30 헉슬리 249 헐 43( 주), I I I , I 16 헤 켈 157( 주), 167 헴펠 15, 16, 23, 106, 179, 181, 心, 184, 193, 213, 217( 주), 232, 242, 243 혁명적 과학 27, 28, 35 호메오레시스 215, 216 혼합유전 II2-u6 환경 41, 1071 108, 113, II51 1191 122, 127, 128, 137, 140-1 44, 153, 156, l 야, 164, 168, 173, 174, 195, 215 환원 19-96 간접 환원 ® 동질적 환원 29 김 l, 71, 72 물리적 환원 l9 이론적 환원 192 이질적 환원 29-31, 71, 72 인식론적 환원 19 직접 환원 70, 71 환원 함수 71, 73, 76-86 환원론 26, 92, 249-2 78
획득형질 115 효소 55, 57, 65, 67, 83, 84, 88, 9I
후기술 184, 185 훼웰 256 흔적기관 학
河斗鳳 서울대학교 생물학과 졸업, 일본 도오호구(東北) 대학 이학박사 서울대학교 자연과학대학장, 부총장, 한국동물·학회장 역임 현재 서울대학교 분자생물학과 교수, 국가과학기술자문회의 위원, 광주과학기술원장 具惠映 이화여대 생물교육과 졸업, 미국 풀로리다 주립대학교 이학박사 현재 상지대학교 생물학과 교수 :::::!?번역 72 |I\I 1 판 1 쇄 찍음 ·· ·· ···· ·· ··· 1994. ll. 5 ----- 1 판 1 쇄 펴냄 …····· ·· ··· 1994. 11. 10 지은이 .. …… •.•• 대이비드 헐 옮긴이 ……… •••• 하두봉 • 구혜영 펴낸이 ..... …… .. 朴孟浩 책임편집 •••••••• •• •• • 당연증 전산조판···· ··· ·· ·· · · 워드뱅크 펴낸곳 … ...... . .. . (주)民音社 출판등록 1991 . 12. 20. 제 16 -4 90 호 135-120 서울특별시 강남구 신사동 506 강남출판문화센터 5 층 대표전화 515-2000/ 팩시밀리 515-2007 값 11 ,5 00 원 © 하두봉 • 구혜영, 1994 과학철학 /KDC 401 Prin te d in Seoul, Korea ISBN 89-3 7 4-4 0 72-5 94470 ISBN 89-3 7 4-3 0 00-2 (세트)
대우학술총서 (번역)
1 유옥민족제국사 콴텐 / 송 기 중 2 수학의 확실성 클 라 인 / 박 세 희 3 중세철학사 와인버그 / 강영계 4 日本語의 起源 밀러/김방한 5 古代漠語音韻學槪要 칼그렌/최영애 6 말과 사물 푸코/ 이 광 래 7 수리철학과 과학철학 와일/김상문 8 기후와 진화 피어슨/김준민 9 이성진리·역사 파트남 / 검효명 10 사회과학에서의 場理論 레빈/박재호 11 영국의 산업혁명 딘/나경수이정우 12 현대과학철학논쟁 쿤 外/조승옥김동식 13 있음에서 됨으로 프리고진/이철 수 14 비교종교학 바하/김종서 15 동물행동학 하인드 / 장현갑 16 현대우주론 시아마/ 양종만 17 시베리아의 샤머니즘 디오세지 ·호괄 / 최김성 1 8 조형마술의 형식 힐데브란트/조장섭 19 힐버트 리랄이일해 20 - 원 시국?f의 진 화 강 言/ 최몽룡 21 商 文明 張光直/윤 내 현 22 마음의 생태학 베이츤/서석봉 23 혼돈 속의 질서 프리고진·스텐저스/유기풍 24 생명의 기원 밀라오르겔/박인원 25 일반언어학이론 야콥슨1 권재일 2 6 국가권력의 이념사 마이네케/이광주 27 역사학 논고 브로델/이정옥 28 유럽의식의 위기 l 아자르/조한경 29 유럽의식의 위기 Il 아자르/조한경 30 일반국가학 켈젠/민준기 31 일반언어학강의 소쉬르/최승언 32 일반체계이론 버틀란피/현승일 33 현대문명의 위기와 기술철학 애거시/이군현 34 언어에 대한 지식 촘스키/이선우 35 음운학원론 트루베쯔코이/한문회 36 우주의 발견 하위트/강용희 37 수학적 발견의 논리 라카토스/우정호
38 텍스트 사회학 지 마/ 허 창운 39 현대물리학의 철학적 테두리 보음 /전일동 40 과학과 가치관의 우선순위 스페리/아남표 41 신화의 진실 휘브 너 /이규영 42 대폭발 실크/홍승 수 43 大同書 };)!有爲/ 이성애 44 표상 포더/이영옥정 성 호 45 과정과 실재 화이트 헤 드/오영환 46 그리스 국가 에렌버그/김진경 47 거대한 변환 폴라니 / 박현수 48 법인류학 포스피실 / 이 문옹 49 언어철학 올스튼/곽강제 50 중세 이슬람 국가와 정부론 램톤 /김정 위 51 전통 쉴즈/김병서신 현순 52 몽 골 문어문법 뽀뻐 ]/ 유원 수 53 중국신화전설 I 袁Ju//전 인초·김선자 54 중국신화전설 ll 〈근간〉 55 사회 생 물학 I 윌슨/이병훈·박 시 룡 56 사회생물학 ll 윌슨/이병훈·박시룡 57 일 반언어학의 제문제 l 밴베니스트/황경자 58 일반언어학의 제문제 ll 밴베니스트/황경자 59 폭력과 성스러움 지라르/김진식·박무호 60 갑 골 학 60 년 冀作賓/이형구 61 현대수학의 여행자 피터슨/김인수·주형관 62 프랑스 혁명의 지적 기원 모르네/주명철 63 해석학과 과학 커널리·코이트너/이유선 64 서양고대경제 핀리/지동식 65 음악사회학 베 버/ 이 건용 66 앙띠 오이디푸스 들뢰즈·가따리/최명관 67 과학커뮤니케이션론 프라이스/남태우·정준민 68 교양교육의 개혁 다니엘 벨/송미섭 69 과학과 가치 래리 라우든/이유선 70 초기제국에 있어서의 교역과 시장 칼 푼라니/이종욱 71 폭정 론 과 저항권 헬라 만트/심재우 72 생명과학철학 데이 비 드 헐/하두봉 • 구혜영