 음성합성
음성합성 박순달 조선대학교수학과졸업 미국 신시내티대학교 이학박사 현재 서울대학교산업공학과교수 저서 「선형계획법」 외 다수 남문현 연세대학교 전기공학과 졸업 연세대학교 전기공학과 공학박사 현재 건국대학교전기공학과 교수 저서 「제어시스템공학」 외 다수 이춘길 서울대학교 심리학과 졸업 미국 일리노이대학교 신경과학박사 현재 서울대학교 심리학과 조교수 소광섭 서울대학교 물리학과 졸업 미국 브라운대학교 이학박사 현재 서울대학교 물리교육학과 교수 저서 「대통일 이론」 외 논문 다수
뇌의 인공적 확장은 가능한가
뇌의인공적 확장은 가능한가 박순달 남문현 이춘길 소광섭 民音社
책 머리에 인류는 과거 4백만 년 동안 끊임없이 진화되어왔으며, 또한 앞으로도 환경변화에 적응하면서 진화하여 갈 것이다. 이런 진화과정에서 특히 주의를 기울일 것은 뇌의 진화이다. 4백만 년 전의 인류의 뇌는 지금의 침팬지 뇌의 용척과 같았다. 그 후, 언류는 육체척으로는 별로 변화가 없으면서 뇌의 진화는 지구상에 현존하는 어느 생물보다 큰 용척을 가진 존재로 진화되어왔다. 이런 점으로 볼 때 인류의 진화에 있어서 뇌의 진화가 가지는 의미는 태단히 크다.
뇌는 그동안 언체 장기 중에서 가장 신버스러운 곳, 영혼이 거주하는 콧, 자아의 집, 신성 불가침의 영역으로 여겨져왔으고로 옴에 대한 의학 적 조치와 같은 뇌의 인공적 조치는 금기시되어왔다. 그러나 언제나 이런 상황이 계속될 수는 없다. 언첸가 뇌의 신비는 벗겨질 것이고 언체 의 다른 장기와 같이 다루어질 수 있어야 하고 또 다루어질 것이다. 저자는 지금이 그 시발점이라고 생각하고 지금으로서는 이른 감이 있지 만 〈뇌의 인공척 확창은 가능한가〉라는 제목으로 뇌의 장래 문제를 다루 기로 한 것이다. 이 책은 크게 두 편으로 나쉰다. 제1편은 뇌의 제어에 관한 현황을 다루고 있는데 뇌의 운동제어와 감각제어 두 부분으로 나누어 설명하고 있다. 제2편에는 뇌의 언공척 확장 가능성을 다루고 있는데 우천 생물진 화에 있어서 뇌의 위치를 다루고 다음에 뇌의 인공척 확장 가능성에 대하여 설명하고 있다. 이 책은 특별한 기초지식 없이도 얽올 수 있도록 하였다. 이 책이 뇌의 실체률 이해하여 톰과 마음이 별개가 아니며 언류 미래셜계에 조금이라도 도움어 된다면 더 이상 기뽕이 없겠다. 1991년 6월 저자 일동뇌의 인공적 확장은 가능한가 차례 책 머리에 .5 제 1 편 뇌의 운동·감각제어 제 1 장 뇌의 운동 제어 1. 서론 13 2. 뇌의 기본 기능과 구조 18 2.1 조절기로서의 뇌 .18 2.2 신경세포 : 뇌의 사전이며 문법 .21 2.3 신경의 전기적 성질 .23 2.4 신경 시스템에 의한 결정 : 시냅스 .29 2.5 뇌의 제어와 통신 .32 2.6 뇌의 계층제어 .33 2.7 뇌의 모델링에 대한 전망 .36 3. 뇌의 운동제어 38 3.1 운동제어 시스템 : 뇌와 외부 세계와의 통신 .38 3.2 제어회로의 구조 : 근육에서 피드백 제어 .44 3.3 상호신경지배 : Descartes에서 Stark까지 .49
4. 운동제어 모델과 활용 52 4.1 모델 만들기 .52 4.2 신장반사의 제어 특성 .60 4.3 운동기능의 변환과 활용 .69 4.4 운동기능의 대체 연구방법 .72 참고문헌 .75 제 2 장 감각의 신경보철 1. 뇌의 기능적 설계 81 1.1 뇌의 기능과 전반적 설계 .81 1.2 뉴런 .84 1.3 뇌의 기능적 설계의 예-안구운동 .86 2. 감각기능의 신경보철 88 2.1 신경보철 .88 2.2 감각의 신경보철 .89 2.3 감각신경보철의 예-전기달팽이관 .90 3. 시각신경보철 91 3.1 시각보조의 개념 .91 3.2 전통적 시각보조의 최근의 성과 .92 3.3 신경보철에 기초한 시각보조 .94 4. 시각신경계 95 4.1 시각신경계의 구성 .95 4.2 시각피질의 구조와 기능 .97 5. 인공시각장치 103 5.1 개념 .103 5.2 인공시각의 구현을 위한 연구와 과제 .105 참고문헌 .109
제 2 편 뇌의 인공적 확장은 가능한가 제 3 장 뇌의 자연진화와 역할 1. 생명의 진화 117 1.1 우주의 생성 .117 1.2 지구의 역사 .126 1.3 생명의 진화 .133 2. 뇌의 자연진화와 역할 141 2.1 생물진화와 뇌 확장 .141 2.2 뇌의 진화 .148 2.3 뇌 확장과 그 역할 .157 참고문헌 .175 제 4 장 뇌의 인공적 확장은 가능한가 1. 기기 발달과 인체보철 177 1.1 기기의 발달 .177 1.2 인체보철 .187 1.3 인체기능의 향상 .193 2. 뇌의 인공적 확장은 가능한가 196 2.1 신경보철 : 뇌의 인공적 확장은 시작되다 .196 2.2 뇌 연구 .208 2.3 전자기술의 발달 .221 2.4 컴퓨터 .235 2.5 뇌의 인공적 확장은 가능한가 .251 참고문헌 .259 찾아보기 .261 필자소개 .267
제 1 편 뇌의 운동·감각 제어
남문현 제 1 장 뇌의 운동 제어 1. 서론 생명유지를 위한 항상성 보지와 효융척인 물질대사, 뇌의 통합제어에 의한 고도의 운동기능, 학습, 기억, 추론 등의 고차정보 처리기능 동윤 30 여 억 년 이상의 진화파정을 통해 생체가 취득한 예지로서 이것들올 배우고 그로부터 인간에게 진실로 유용한 기술올 창출하는 것이 오늘날 의 공학에 부여된 사명 중의 하나이다. 구체척으로는 생체기구의 교묘함 을 올바로 인식하고 평가함으로써 미래의 기술, 한걸음 더 나아가서 미래의 인간사회의 이상척인 상태에 관해 시사하는 바가 크리라 생각한 다. 생 체공학 (bioengineering) 또는 생체 사이버네틱스 (biocybernetics)의 목적은바로 이런 데에 있다.
생체기능에 관해서는 의학, 생리학 분야에서오랜 역사률 거쳐 연구되 어 풍부한 연구결과가 축척되어 있다. 공학의 업장에서 생체기능올 규명 하려고 할 경우 의학이나 생리학의 자료률 그때그째마다 차용하는 것만 으로는 충분치 못해 목척에 따라서는 스스로 계측하고 처리할 휠요가있으며 컴퓨터 단층촬영술에서 보듯이 최근의 생체 정보처리 기술은 괄목 할 만한 진전을 보이고 있으며 생체연구에 기여하는 바가 크다.
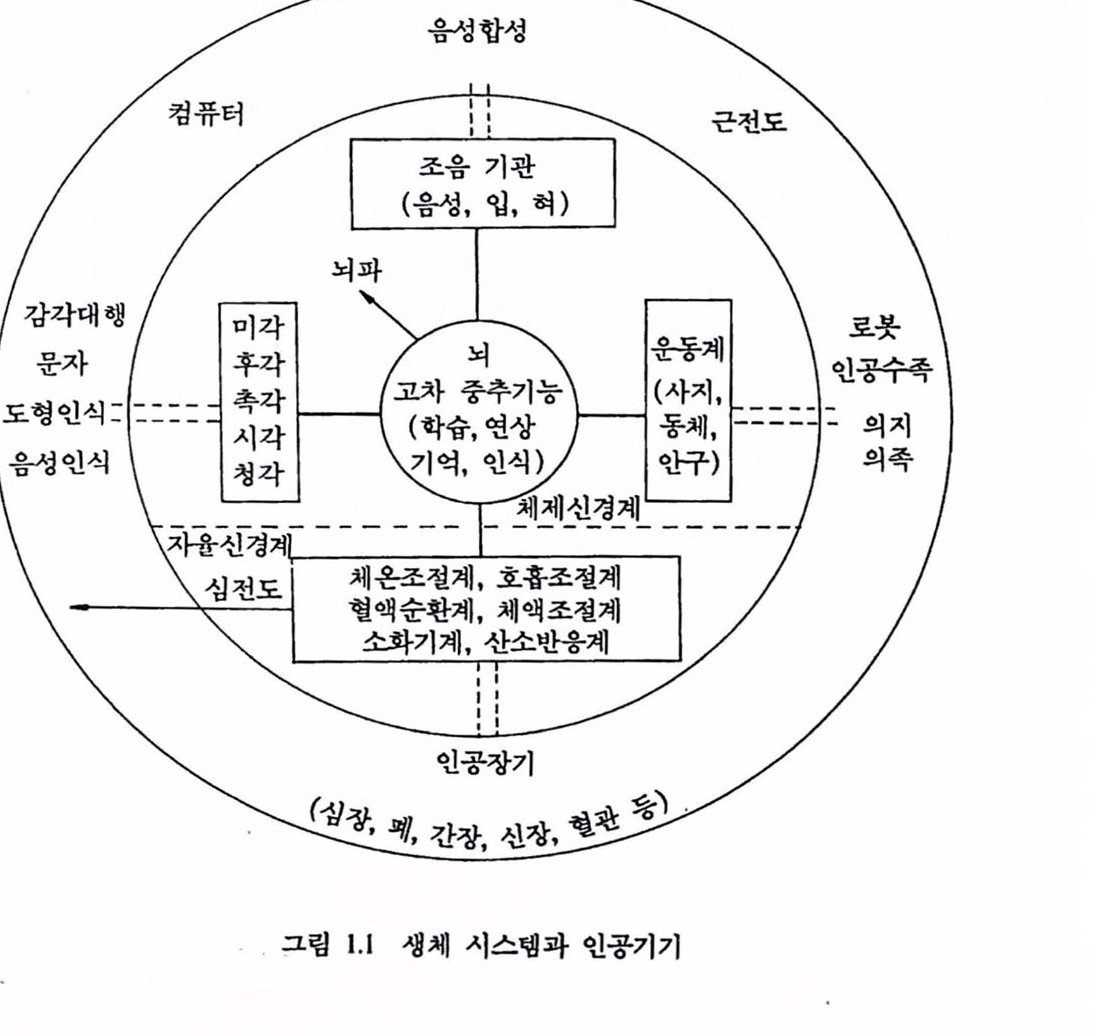
최근 생물과학의 흐름은 분자생물학에서 보듯이 마이크로에 마이크로 를 거듭하여 생체를 미세구조로 분할하는 것으로 대표된다. 이런 경향에 대해 총체로서의 기능을 이해하고 활용하려는 입장에서는 시스댐으로서 생체를 파악하는 것이 필요하다. 많은 요소가 복잡하고 정교하게 시스댐 을 이루고 있는 생체구조와 기능을 해명하려면 종래 생리학적 수법과는 다른 방법언 시스댐 이론척 수법이 유용한 도구가 된다.모델링 (modeling) 또는 합성에 의한 해석법이 주류플 이루는 시스템 접근법은 구조 척으로 해명되지는 않지만 기능면에서 모순 없는 구조 혹은 그것을 유지 하는 원리척언 것을 추구하는 일에 있어서는 매우 유용하여 생체기능 중 많은 부분의 해명에 크게 기여하였다. 생체는 현재의 기술로는 도저 히 실현될 수 없는 우수하고 정교한기능을 많이 갖고 있다. 이것은 신경 시스댐, 내분버 시스댐을 포함한 시스댐에서 특히 두드러진다. 예를 들어, 근육에 의해 발생되는 험이나, 장력(tension)제어에 대한 것은 현재의 어떤 인공척인 구동장치로도 기능하지 않다. 문자와 도형을 인식하고 잡음 속에서 소리를 분간해 틀을 수 있는 능력에 대한 현재의 기능 대행 장치는 아주 유치한 단계이다. 신경 시스탱에 의해 생체 시스댐을 분류하면 감각과 운동에 의한 동물성 기능에 관한 체제(somatic)신경 시스템과 호홉, 순환 동의식물성 기능에 판계한 자융신경 시스댐으로 나눌 수 었다. 자윷신경 시스댐은 생체 내부의 상태를 일정하게 보전하고 항상성(homeostasis)과 밀접하게 관련되며 의사의 지배를 받지 않는다. 그림 1.1은 두뇌를 중심으 로 위의 두 가지 신경 시스댐에 확안하여 생체기능을 도시한 것으로 뇌, 감각, 운동, 향상성으로 구생된 기판파 시스댐은 공학척 접근의 대상이 되어 계측 및 모멜랭에 대한 연구가 행해지고 있다. 뇌파(brain waves, EEG), 섬천도(ECG), 근전도(EMG) 신호는 각각 뇌, 섬창, 근육 기관 의 활동율 반영하는 전기신호이며 이들 신호로푸터 각 기관의 활동과
음성합성
그림 1.1 생체 시스템과 인공기기
질환의 상태를 추정할 수 있다. 그렴 1.1의 바껄쪽에는 각 기판과 관계된 기능올 대행하는 대표척언 언공기기률 나타내었다.
두뇌를 컴퓨터와 버교하는 것은 온당하지 못하지만 얘우 일반척인 견해로서 최근에는 두뇌의 정보처리 기놓율 모의한 새로운 방식(고도의 병렬처리 퉁)의 언공장치와 시스댐이 모색되고 있다. 감각대행, 품자도형인식, 음성인식 등은 신경제어를 응용한 새로운 연구분야로 각광을 받고 있다. 로봇, 인공 팔다리, 의수족 등은 운동제어 시스댐의 기능을 활용하여 새로운 공학 시스댐을 창조하거나 운동기능이 결손된 사람들에게 정상적언 활동을 부여하는 것과 관계된 것으로 신경과학, 재료과학, 청단전자기술의 산물을 활용하여 장애와 투쟁하는 사람들의 생활에 크게 기여할 수 있는 첨단기술의 새로운 분야이다. 특히 감각과 운동 시스댐에 대한 공학적 연구는 신경의 전기적 홍분현성을 이용하려는 기술자, 과학자, 의사들의 노력으로 응용 신경제어라는 새로운 연구분야로 발전하고 있으며 이들이 벌이는 노력의 결실은 장애지들이 생산적인 삶을 누릴 수 있도록 도와출 수 있는 첨단장치로서 결실을 맺게 될 것이다. 성대, 입, 혀 등 조음기관은 음성합성 분야로 각종 정보처리 및 분야에서 널리 연구, 개발되고 있다.
두뇌의 기능을 외부적무로 확대하거나 본떠서 손상된 기능을 대행하거나 로봇 제어 시스댐과 같은 공학 시스템을 설계하는 데 이용하려면 두뇌와 외부세계와의 통로인 운동 시스댐에 대한 이해와 운동을 정멸하고 간략하게 기술할 수 있는 수학적 모델이 필요하다. 안구운동과 같은 일상척인 운동의 궤척을 조사해 보면 이것들은 운동 뉴런 발화의 돼턴과 아주 밀접한 관계가 있음을 알 수 있다. 안구운동 시스댐에서 안구를 운동시키는 근육을 구동하는 신경제어 신호는 안구의 운동에 충실히 반영된다. 바꿔 말하면, 운동궤척을 연역하면 신호의 형태를 바로 알아낼 수 있다는 뭇이다. 제어신호가 식별되면 운동의 성질과 이것을 발생시키는 기구에 대한 이혜를 얻올 수 있다는 것이 최근에 합핵쳤다 (Stark, 1968). 고양이의 후지 (hindlimb) 는 거의 35개의 근육에 의해 구동되는데 사람의 것도 이것과 버슷하다. 이러한 구동장치(actuator)는 서거나 앉거나 하는 합목척성이 결여된 운통과 혼들고, 걷고, 뛰어오르는 것을 포함한 합폭척성 운동올 발생시키는 데 사용펀다. 후지에는각종 감지기 (sensor)가 있어 전상태 피드백(full state feedback)을 활용할 수 있는보기 드문 시스댐의 하나가 되고 있으며 이에 대하여 스위청 능력과 지능을 갖추고 있다. 여기서 고양이나 사람에 있어서 운동(보행)의 신경 제어는 비선형, 다중업출력 지능제어 시스댐의 좋은 본보기임을 알 수 있다. 따라서 운동제어는 신경생리학자나 제어공학자들에게 다같이 아주 흥미있는 연구 대상이다. 이것 말고 버선형 다중 입출력 지능제어의 실제 보기를 찾기는 그리 쉽지 않을 것이다.
Peter Medawar(1952)의 말대로 과학이 〈해결하는 재주(art of soluble)> 라면 공학에는 이것 이상의 것이 있다. 따라서 운동의 신경제어가 중요하다면 이것을 연구할 수 있는 도구를 마련해 봐야 한다. 이 방면의 연구자들이 써 온 도구는 보통 세 가지이다. 첫째는 표준의 콩학척 이론인 최적제어이론과 안정이론의 척용이다. 신경제어를 규명하기 위해서는 확실히 정의된 성능기춘을 정하는 것이 필요하다. 최척제어 문제는 실험의 수학척 모델에 있다. 따라서 실험과 이론간의 어떤 것들을 제공하려 면 모델령과 모의실험이 따라야 한다. 이 단계에서 실험 가능한 가갤을 얻을 수 있으며, 먼저 최적제어를 척용해본다. 많은 사람들은 근육의 조화(공동)작용에 대해 연구하려고 한다. 이때 〈조화〉란 실제로 무엇을 의미히는가? 어떤 운동이 조화됐다고 말할 때 보통은 운동을 이해하는 데 별로 쓸모도 없는 미척 판만을 하게 된다. 조화작용에 대해 수학적 정의를 하려고 노력하면 어떤 통찰력에 쉽게 도달할 수 있다. 우리는 조화작용을 어떤 엄무를 수행하기 위해 사지와 근육의 최적 상호관련으로 정의할 수 있다고 믿는다. 이러 한정의는 제약에 대한 연구로 이어져 자칫하면 최척성(조화작용)을 잃게 되어 버조화운동으혹 이어철 수도 있다. 또한 모멜령과 시률레이션의 안내가 없을 경우 실험자틀은 시간낭비를 하거나 의미 없는 측정에 돌업하거나 하여 의미 있는 결과를 얻기 어렵게 된다. Stark(1980)와 같은 이는 이런 경우 칙정 및 간정 감도해석법올 활용 하여 실험을 계획하고 모멜의 매개변수들을 조정하는 방법올 개척해 왔다. 이와 같은 결과는 물론 실험에 의해 검중된다. 제어공학자들은실험척으로 실헝 가능한 가설과 결과를 제공하여 신경생리학에 기여하는 바가 크다. 실험에 통하는 이론이나 해석 결과만이 학계에서 인정된다는 것은 고금의 진리이다. 결국 이러한 분야를 단독으로 접근한다는 것은 극히 힘들다는 것을 알 수 있다. 한 사람의 힘으로는 이 분야 연구에 필요한 실험과 이흔적인 신경생리학의 지식과 실제 이론면에서 제어의 상세한 지식을 조합하기는 어렵다(Stark 교수와 같은 이는 동물학자, 신경생리학자이면서 응용수학과 제어이론에 밝아 생체공학의 여러 분야 가운데 신경제어에 관한 분야를 개척하는 데 크게 기여한 대표적인 본보 기이다). 신경제어의 연구는 앞서 말한 신경 시스댐의 해명과 이것의 활용에 펼수척이라는 것은 이해하고도 남음이 있으며 새로운 연구도구인 표준의 공학척 기법, 모댈링 및 시률레이션법, 실험법에 의한 접근이 요구됨을 알 수 있다. 이를 위해서 제어공학과 신경생리학의 협조는 물론, 경우에 따라서는 양분야의 접근법을 갖춘 생체공학적 방법이 활용 돼야 할 것이다. 그렴 1.2는 이러한 분야의 협동연구로 만들어진 Utah / MIT Dexterous Hand이다.
이 장에서는 먼저 뇌의 기본구조를 살펴본 다음 뇌의 운동제어구조를 살펴보고 이것이 어떻게 활용될 수 있는지 알아보기로 하겠다. 2. 뇌의 기본 기능과 구조 2.1. 조절기로서의 뇌 뇌는 언체의 모든 세포를 지배하며 이것의 명령의 설체는 감각, 운 동, 사유, 기억파 꿈 등으로 나타난다. 이것올 가능케 하는 것은 중추신 경 시스댐(central nervous systems, CNS)으로서 신경섬유의 망으로 모든 세포를올 연철한다. 신경파학의 아버지 쉐링톤(Charles Sherring-
그림 1.2 제어공학, 신경생리학, 생체공학의 협동으후 만들어진 정교한 기계손
ton, 1 940) 은 〈요술베틀(enchanted loom)>에서 수백만 개의 북으로 풀어지지 않는 항상 의미 있는 다양한 무늬들을 짜낸다고 하여 두뇌를 베틀에 비유하였다. 영(Young, 1951)은 국가의 번영과 유지를 위해 10 억의 관료들이 서로 전화를 통해 계획을 논의하고 명령을 내리는 일에 뇌의 움직임을 비유하여 뇌는 신체가 생명을 유지하기 위한 행동을 계획 하고 조정하는 것이라고 보았다.
모든 생물은 능동 시스댐으후서 삶을 유지하기 위한 일틀을 선돼해서 시행한다. 고등동물에 있어서 가능한 행동의 과정선태은 뇌에 의해 이루어지며 개체의 모든 생명을 조절하는 동인( )이다. 신경 시스댐은 계속척으로 일플을 하고 있다. 이것은 간단한 도전성을 갖는 채널 이상의 것으로서 전화나 라디오와 같은 인위척인 기기플을 써서는 그것들의 통신기능을 셜명하기는 어렵다. 통신은 제어를 위한 필수요소로서 신경이 어떻게 정보를 나르는지에 대해서는 많은 말들어 필요하다. 따라서프로그램 (program) 이라는 말을 써보기로 한다. 뇌는 들어오는 정보에 맞춰 우리가 해야 할 일을 조직하는 데 쓰일 명령을 내린다. 특히 우리 의 생명유지에 필수적인 일들을 추구하는 데 기준이 되는 틀언 표준 시스템을 갖고 있다.
사람이 뇌를 통하여 채택하는 행동양식의 한 가지는 행동의 적절한 과정을 예측하는 데 도움이 되는 일반적인 도식이나 모델을 만드는 일이며 인간의 두뇌는 이와 같은 일을 꾸미는 데 필요한 프로그램을 준비한다. 어떤 형태의 모델이 쓰이든간에 두뇌는 능동척인 동인이라는 개념이 포함되어야 한다. 러시아의 신경학자 Luria(1973)는 두뇌를 미래의 모델을 〈창조(create)>하는 동인이라 하였다. 두뇌를 설명할 때 사람들은 인공척인 것들을 갖고 설명하는데 이 가운데 대표적인 것이 컴퓨터이다. 컴퓨터는 어떤 경우이건 뇌의 그것과는 여러 면에서 서로 다른 원리를 쓰고 있다. 오직 활용할 수 있는 것은 정보전달의 원리들과 컴퓨터 학자들이 만들어 놓은 〈소프트웨어〉를 이해하는 것으로 충분하다. 생명은 어떤 〈부호〉 안의 〈정보〉의 〈통신〉에 바탕을 둔 〈명령〉에 따라 모든 행동이 〈제어〉되기 때문에 유지된다. <질서〉는 〈명령〉에 의해 유지된다. 정보의 진수는 그것이 여러 가지 결정을 제어하는 데 있다. 동물이나 사람의 행동은 뇌(DNA의 영향 아래 발달된)에 포함된 정보나 명령에 의해 제어된다. 그러나 뇌가 갖고 있는 표준 시스템은 신경에 의해 제공된 채널을 타고 외부로부터 들어오는 정보에 따라 항상 자문을 받는다. 통신 채널올 따라 전달되는 정보는 부호의 형태로 변환된다. 신경내에서의 어떤 물리척 사건은 기관이 반웅할 수 있는 부호의 단위를 제공해야 외부세계에서의 사건올 나타내거나 대신할 수 있다. 이와 같은 대웅이 있어야만 효과척언 통신 채널이 형성된다. 따라서 신경부호의 단위를 초사하여 이것들이 어떻게 바깡 세계의 사건을 나타내는지 알아보아야 한다. 신경 시스템이 어떻게 정보를 부호화하는가를 기술하려면 보통 신호가 어떻게 신경올 거쳐서 눈이나 귀와 같은 감각기관에 정착하는가부터 시작해야 한다. 통신의 기본기능은 제어기로 하여금 가능한 운동의 집합으로부터 올바른 명령을 내리게 하는 데 있다. 음성의 기본은 그것이 이해되는 데 있으며 척절한 행동이 따르게 마련이다.
이제 뇌는 명령을 발하는 동언이며 그것이 신호를 복호(decoded, 이해됨)한 후에 그것이 어떻게 되어가는가를 수신한다. 그것의 단위인 신경 세포는 활동을 할 수 있는 기본능력이다. 이것틀은 우리의 행위를 관장히는 지배자이다. 자거나 일어나거나 산보를 하거나 먹거나 독서를 하는 것은 우리가 이런 일플을 선택했을 때 동작히는 활동세포를 뇌가 갖고 있기 때문이다. 두뇌는 외부의 정보에 대해서만 동작하는 것이 아님을 알 수 있다. 내부로부터 우리 생활을 위한 프로그랭을 제공하는 데도 영향을 미침을 알 수 있다. 신경세포는 어떤 것이 활동할 수 있도록 해줌으로써 동인을 형성하여 과거의 역사는 연속활동의 상태로 남겨 두고 주변의 것들과 상호작용을 한다. 생물체는 수동 시스템이 아니며 평형상태에 남아 있으면서 계속척으로 어떤 일플을 하고 있다. 생명의 행위에서 보듯이 이런 일들은 아주 조직척으로 이루어져 기관으로 하여금 목표에 도달하도록 안내하며 삶을 유지하게 한다. 이러한 행위는 능동제어기에 의해 수행되며 순간순간마다 외부로부터 들어오는 정보를 활용하여 명령을 내리는 데 쓰이는 프로그랩을 작성한다. 2.2. 신경세포 : 뇌의 사전이며 문법 두뇌는 동물이나 인간의 모든 복잡한 행위를 어떻게 발생시키는가? 이런 혐의 원천은 막대한 수와 다양성을 가진 신경세포와 그것의 활동 및 연결에 있다. 이것들이 방대한 운동의 집힘을 가능케 한다. 사랍의 대뇌피철(cerebral cortex)에는 수십억(10 ) 개의 신경세포가 있다고 한다(그렴 1.3 창조). 그러나 실제로 중요한 것은 아마도 이것틀간에 어느 두 개도 비슷하지 않은 데에 있다. 아주 간단히 생각하면 이들 세포 뇌의 표연
뇌의 표연
의 각각은 아래의 것 가운데 하나임을 알 수 있다. (1) 외부 세계에서 진행되는 변화의 특정의 일부분, (2) 어떤 과거의 외부척 변화의 기억을 갖는 부분, (3) 신체에 의해 수행될 수 있는 행위에 대한 명령의 일부분. 과거의 사건을 나타내는 것과 외부에서의 것을 나타내는 세포의 행위의 조합은 순간에서 순간으로의 척절한 행위를 주관한다.
신경세포나 시스댐은 대뇌의 알파뱃으로서 이것들은 조합에 따라 단어, 문창, 문단, 장 및 책이 되어 우리의 행동양식을 생성하는 프로그랩을 만들어낸다. 두뇌의 언어를 이해하려면 글자나 음철 같은 것들이 단어와 같은 의미를 갖는 단위를 생성하는 방법을 알아야 한다. 단어들 은 적절한 조합으후 쓰일 때는 외부세계에서의 사건을 나타내거나 근육이나 선(gland)에 보내질 적절한 명령으로 나타난다. 사람들이 쓰는 구두어나 문자어는 목표나 결과를 얻으려는 데 있지만 두뇌의 언어는 실제로 이런 목척플을 결정하는 기본요소들을 내포하고 있기 때문에 어떤 의미에서는 훨씬 기본척이다. 언어의 척합한 사용을 규제하는 시스템을 문법이라 할 때 두뇌는 언어를 포함한 생명의 적절한 행위를 조철하는 다른 문법을 갖는 다른 언어를 가통한다고 말할 수 있다.
뇌는 다른 것들과 조합함으후써 어떤 활동을 할 수 있는 막대한 수의 단위플을 갖고 있다. 앞서 분류한 세 가지는 각 신경세포의 활동을 이웃의 것과 더불어 외부세계의 현재나 과거의 사건이나 행위를 만들어낸다. 이제 우리는 어떤 세포가 외부세계에서의 사건을 〈표현하고〉 외부세계의 영향을 미칠 수 있는 신체의 행위를 〈나타낸다〉는 컷의 의미를 살펴보기로 하자. 2.3. 신경의 전기적 성질 뇌가 수행하는 놀랄 만한 계산능력은 일정한 전기학척인 변화(임펄스,impulse)를 발생하는 신경세포의 험에 의존한다. 모든 신경세포 또는 뉴런(neuron)은 신호를 수신하는 수천 개의 연결단자를 가진 아주 작은 정보처리 시스댐이다. 뉴련은 뇌의 트랜지스터, 저항기, 커패시터, 도션이다. 컴퓨터가 개개의 회로소자로 연결된 것파 마찬가지로 뉴련은 두뇌의 계산능력을 발휘할 수 있도록 특수한 방식으로 연결되어 었다. 이것들윤 상호 연결된 신경회로망올 형성하여 외부로부터 감각 정보률 수집하고 정보를 분석하여 운동을 발생시킨다. 신경성유는 중앙의 신경세포체로후A
1 ,‘ -‘~ I 、., A .、‘ ‘ .. / 、 ‘ • \ 시수상냉돌스기를 위한 ι 유입 구심섬유 ‘J#‘. 1、r. ,〈’잭/A 、 ....“ -‘ 유출축색 가지‘ 、、 J그림 1.4 대뇌피질의 단일 신경세포. Golgi법에 의해 몇 개의 세포만 착색하고 주위의 다른 것틀윤 그대로 놓아둔 것엄.
터 멀리 뻗쳐 있다(그렴 1.4 ).이것은 핵(nucleus)을 갖고 있어 이컷의 DNA는 신경세포와 그 구성부의 물리적 상태를 제어한다. 이것은 세포가 실제 물질을 생성하는 것을 주관하여 적절한 메시지를 보낼 수 있는 특수한 기능을 수행할 채비를 갖추도록 한다. 이러한 〈메시지〉는 신경섬유를 따라서 통과하는 신호의 형태를 취하며 이러한 천달기구는 찰 알려져 있다. 포유동물의 신경세포와 섬유는 아주 작아 연구하는 데 어려움이 많은 반면 오징어의 것은 보다 커서 다루기가 쉽다.
뉴런은 세포체, 축색(axon), 수상돌기(dendrite), 단자단추(terminalbutton)의 네 부분으로 구성되어 있다. 세포체는 핵을 갖고 있으며 뉴련의 생명과정에 대한 여러 부분의 기구를 갖고 있다. 수상돌기와 세포체는 뉴련에 대한 정보업력을 받아들인다. 축색은 출력 도전체(전선)로서 뉴런의 메시지를 말단에 전달한다. 제어공학적으로 말하면 제어신호를 근육에 보내는 일을 한다. 단자단추는 축색의 끝에 달려 있어 다음 뉴련 에 신경 메시지를 전달하는 화학물질을 내보낸다. 그렴 1.5는 대표적인 운동 뉴런(motor neuron)이다. 운동 뉴련의 세포체는 보통 척추에 위치하며 축색은 근육세포의 집합 위에 닿아 있다. 운동 뉴런에서 기다란 축색은 미앨런의 초(myelin-sheath)로 철연되어 껍질로 싹여 있다. 신경섬유의 안함에는 전위차( -70mV 정도)가 존재하여 메시지를 보내고 신경 시스댐에서 결정이 이루어지는 수단을 제공한다. 각개의 신경섬유는 대전체로서 메시지의 전파에 필요한 에너지원을 갖는다. 섬유를 척절히 자극하면 전기척언 폭발이 전체 길이를 따라 전파되는데 이것을 신경 임펄스(nerve-impulse) 또는 활동전위(action potential)라 하며 신경섬유(축색)를 따라 신호로 전파된다. 신호가 통과되면 다음 임펄스가 통과하기 전에 잠복기가 있다. 각개의 활동천위는 이렇게 완성되며 이것플은 모두 똑같다. <임펄스〉의 크기는 각 점에서의 전하(charge)에 종속이다. 시작이나 끝에서 신호의 크기는 변하지 않으며 이와 같은 감소 없는 전도는 정보률 매우 안전하게 천달하는 방편이 된다. 어떤 신경에서는 엄훨스의 속도가 시속 300km 이상언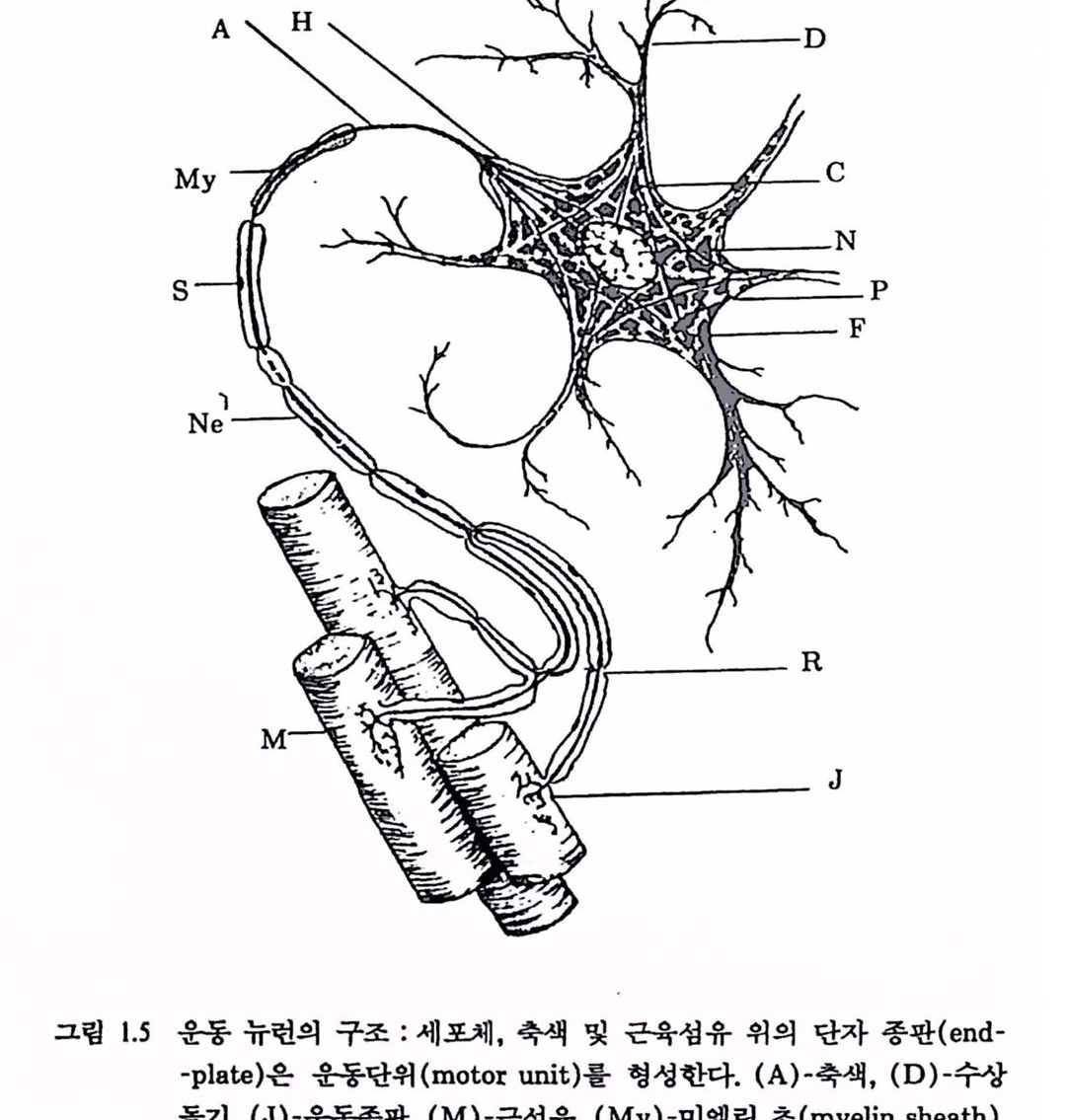 A D
A D
그립 1.5 운동 뉴련의 구조 : 세포체, 축색 및 근육섬유 위의 단자 종판 (end plate)온 운동단위(moto unit)률 형성한다. (A)-흑색, (D)-수상률기, (J)-운동종판, (M)-곤샘유, (My)-미옐련 초 (myelin sheath), (N)-핵. (R)-Ranvier의 마다, (S)-Schwann 세포의 핵, 업력은 시냉스 (synapse) 훌 경유하여 수냥돌기와 세포체에 도랄한다. 출력은 촉잭융 거쳐 천달된다. 운동 뉴련의 세포체는 수천 분의 1센티마터나 촉잭운 수집 핸티미터나 된다.
 막 r챔
막 r챔
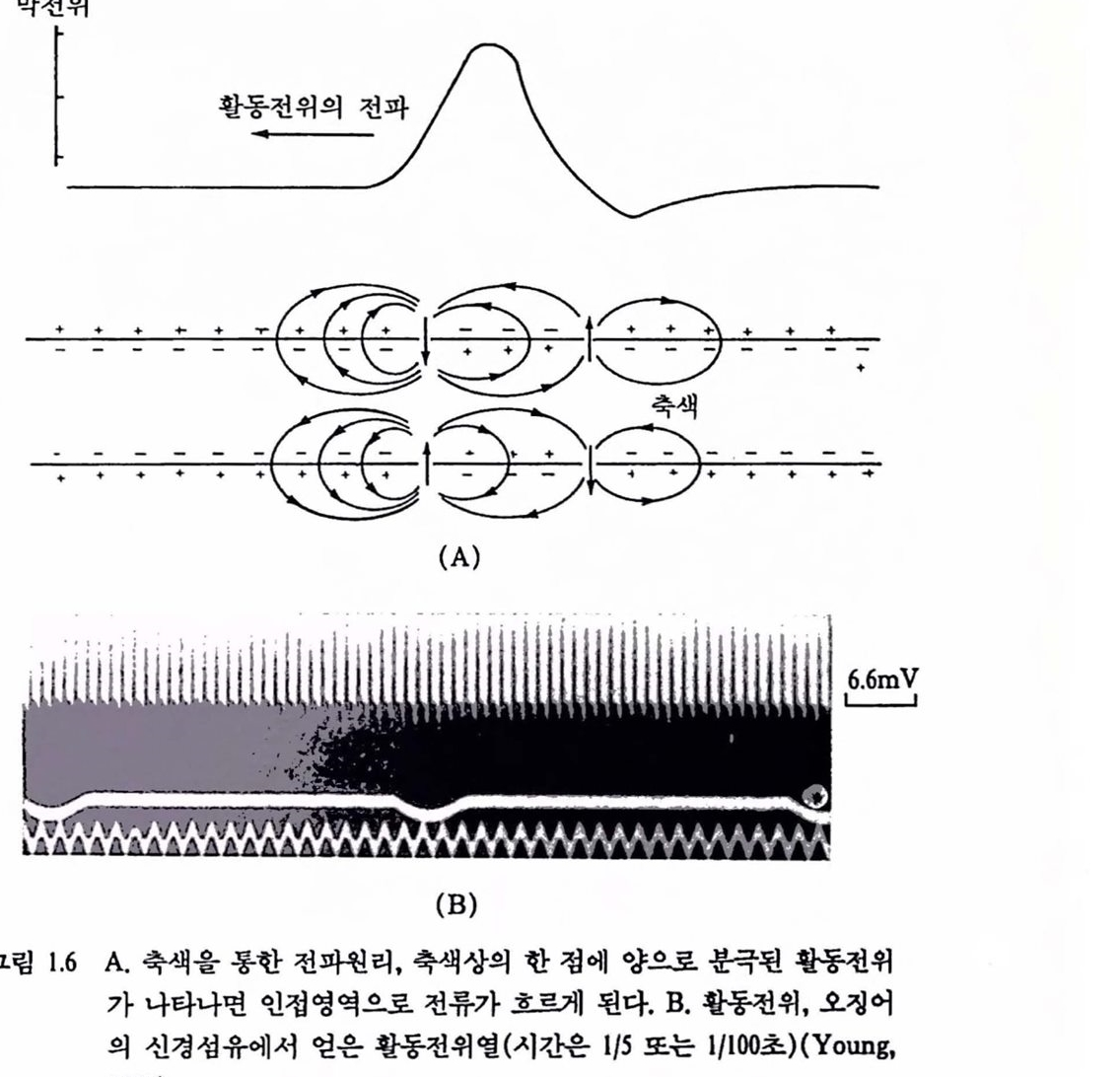
그럽 1.6 A. 축색을통한 전파원리, 촉색상의 한 정에 양으혹 훈극된 활통전위가 나타나연 인접영역으로 전류가 흐르게 된다. B. 활동전위, 오갱어의 신경섬유에서 얻은 활통천위열(시간은 1/5또 는 1/100초)(Young, 1978).
것도 있다. 그립 1.6은 활동천위의 전파훤리와 오갱어의 신경활통 천위열을 나타낸 것이다.
앞서 고찰한 바와 같이 (1) 엄훨스는 감쇠 없이 천달되며, (2) 모든임펄스는 임의의 신경섬유에서 똑같다. 감소 없는 천도현상은 신경천도에 대한 실무율(惠無律, all-or-nothing law)을 형성하여 신경 시스템의 모든 특성을 결정한다. 곧, 신경 시스템은 기본적으로 전기척언 수단에 의해 정보를 전달하며 이것이 신경 전도의 기초 메커니즘으로서 전기척언 ON /OFF 스위치의 형태로 구성된다. 이것은 인간의 목소리의 변화에 따를 수 있도록 신호 자체가 변화하여 전달되거나 부호화되는 전화기나 라디오와는 다르다. 신경섬유에서는 이런현상이 일어날 수 없다. 변화가 있다면 전파되는 펄스의 수와 분포가 있을 뿐이다. 이와 같은 〈주파수 변조〉는 임의의 신경섬유 하나가 서로다른 패턴의 임펄스를 전달하도록 한다. 이것들은 적거나 많거나 그리고 상이한 간격을 갖는 왜턴을 갖는다. 이런 패턴들은 상이한 메시지를 전달하는 데 이용되기도 하지만 일반척으로 신경 시스댐은 상이한 강도를 나타내기 위해 주파수의 차이를 활용한다.
따라서 신경 임펄스는 언어의 음이나 글자에 버할 수 있는 언어의 기초 단위이며 이것들은 스스로 의미를 전달하지 않는다. 이것들의 어떤 그룹에서는 주파수에 의해 할 수는 있지만 두뇌언어의 의미 있는 단위나 단어들은 개별척언 신경세포이다. 상이한 음색이나 시각경로와 칼은 특철의 차이는 각개의 특질에 대한 상어한 신경세포와 신경섬유를 가짐으로써 신경 시스댐에서 부호화된다. 이래서 우리는 방대한 수의 신경세포를 갖고 있다. 뇌는 기본척으로 다중 채널 시스댐으료서, 정보의 각 항목을 분리된 채널에 넣어 메시지를 형성히는 부호의 방법은 전기기술자들이 배선번호를 불이는 시스댐과 비슷하다. 두뇌의 배선올 사상(寫像)하는 데 어떤 감각 메시지들은 위와 같은 빠른 웅답을 필요로 하지만 뇌로부터 도달하지 않는다는 사실을 과학자들은 알었다. 그럽 1.7과 같이 뜨거운 다리미에 손끝을 대연 반사궁이라는 보호회로롤 작동시킨다. 손이 다리미에 닿으면 피부에 있는 수용기 세포는 메시지를 잡아 감각 뉴런으로 전달하여 그것을 척추에 있는 운동 뉴련으로 연계시킨다. 운동신경은 이 메시지를 손과 팔의 근육으로 되돌려 보내 손가릭올 다리미에서 떼도록 한다. 통증을 느끼는 것은 약간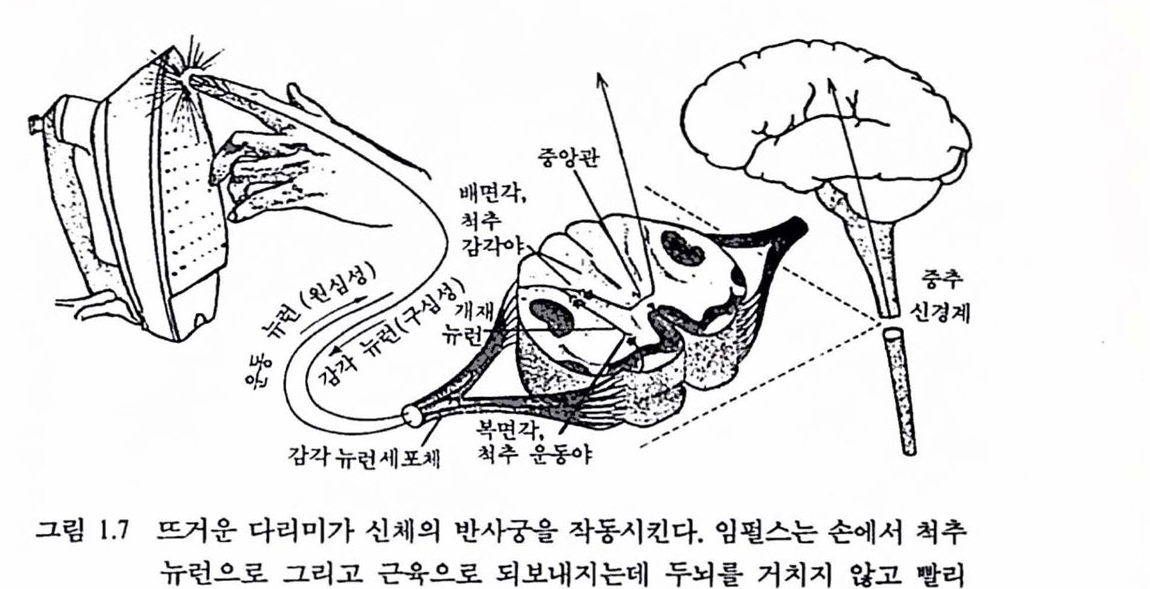
그림 1.7 뜨거운 다라며가 신체의 반사궁을 작동시킨다. 엄훨스는 손에서 척추 뉴런으로 그리고 근육으로 되보내지는데 두뇌를 거치지 않고 빨리 응답한다. 뇌에 통중이 등록될 즈음에 손은 이미 떨어져 나온다.
시간이 걸린다. 뜨겁다고 느꼈을 때 손은 이미 떨어져 나와 있다. 반사궁 과 함께 수용기 세포가 임펄스를 감각신경에 보내면 이것은 척추를 거쳐 뇌에 엄펄스를 전달한다. 뜨거운 다리미에 대한 감각은 뇌로 가는 많은 엄펄스 가운데 하나다. 메시지는 신체 각 부위로부터 일정하게 도랄한다. 뇌는 모든 업력 엄펄스를 가늠해 가장 급한 것부터 웅답해 준다. 이러한 반사제어에 대해서는 뒷장에서 상세히 다룰 것이다.
2.4 .신경 시스댐에 의한 결정 : 시냄스 망막상의 빛의 무늬나 손에 잡고 있는 물체에 의해 생긴 신호들은 외부 세계에 대한 부호화된 메시지를 두뇌에 제공한다. 상에한 신경섬유가 수령하거나 다른 신경세포를 활성화하는 콧에서 엄훨스는 이해되는 부호로 바뀐다. 두뇌나 척추의 각캐 세포는 다수의 수용야(receptivefield), 수상돌기, 외부로 유출되는 단일섬유인 축색을 갖는다. 어떤 감각 기관으혹부터 신경 시스댐에 들어오는 각 섬유는 여러 번 가지를 거쳐 척추나 두뇌에서 많은 세포와 연결을 한다. 한 세포의 축색의 끝이 다른 세포의 수상폴기와 만나는 점을 신경근 연접, 곧 시냉스(synapse)라 한다. 이것은 전기척언 게이트(gate) 또는 밸브(valve)로서 이것의 전류에 대한 저항은 다흔 뉴련의 축색단추로부터 수신되는 전달 화학물절에 의해 제어된다. 그립 1.8은 대표척인 시냉스를 나타낸 것이다. 시냉스에서 세포를 통해 이통한 전기 임펄스는 화학신호로 바뀌고 다시 엄펼스로 되돌아온다. 이를 상세히 셜명하면 다음과 같다.
시냉스에서 2개 신경세포의 막은 서로 압박하지만 섬유의 내용물은 연속으로 이동되지 않는다. 따라서 전 시냉스(pre-synapse) 섬유와 후 시냉스(post-synapse) 세포의 내부 사이에는 장벽이 있어 이것이 전기적인 신경 임펼스가 다른 뉴런으로 전파되는 것을 불가능(통상)하게 한다. 전 시냉스 섬유로부터 후 시냉스의 특별한 부위로 방출되는 화학물질(acetylcholine 등)이 전달에 영향을 미친다. 화학물질에 대한 응답은 도달하는 임펄스의 효과를 증대시켜 후 시냉스 세포에 영향을 미친다. 전류는 화학 전달물질의 형태에 따라 음이거나 양이다. 이련 화학물철은 최근에야 찰 알려졌으며 전 시냉스 섬유의 끝에 있는 손잡이(button, 단추라 알려짐)에서 발견된다. 이 단추는 아주 작아서 발생된 전류로는 세포의 축색에서 활동전위를 발생하는 데 충분치 않다. 그러나 수성돌기 표면에는 수많은 단추가 었다. 대뇌피질의 한 개 세포 표면에는 5만 개 이상의 작은 단추들이 었다(Cragg, 1967). 서로 다른 단추에서 발생된 천류는 통합되는 데 아주 가까운 공간과 시간에서 활동전위률 형성하여 후 시냉스 세포의 축색을 따라 다른 신경세포와 근육 또는 기판에 전달된다. 시냉스를 건너는 정보의 통신은 일상척이며 한 뉴련의 단추로부터 다른 뉴련의 수성돌기나 세포체(어먼 경우에는 촉색)로 흐른다. 이러한 통합과청은 신경 시스댐에서의 결정이 도달되는 수단이 된다. 감각 신경섬유를 따라 각종 조합으로 도달되는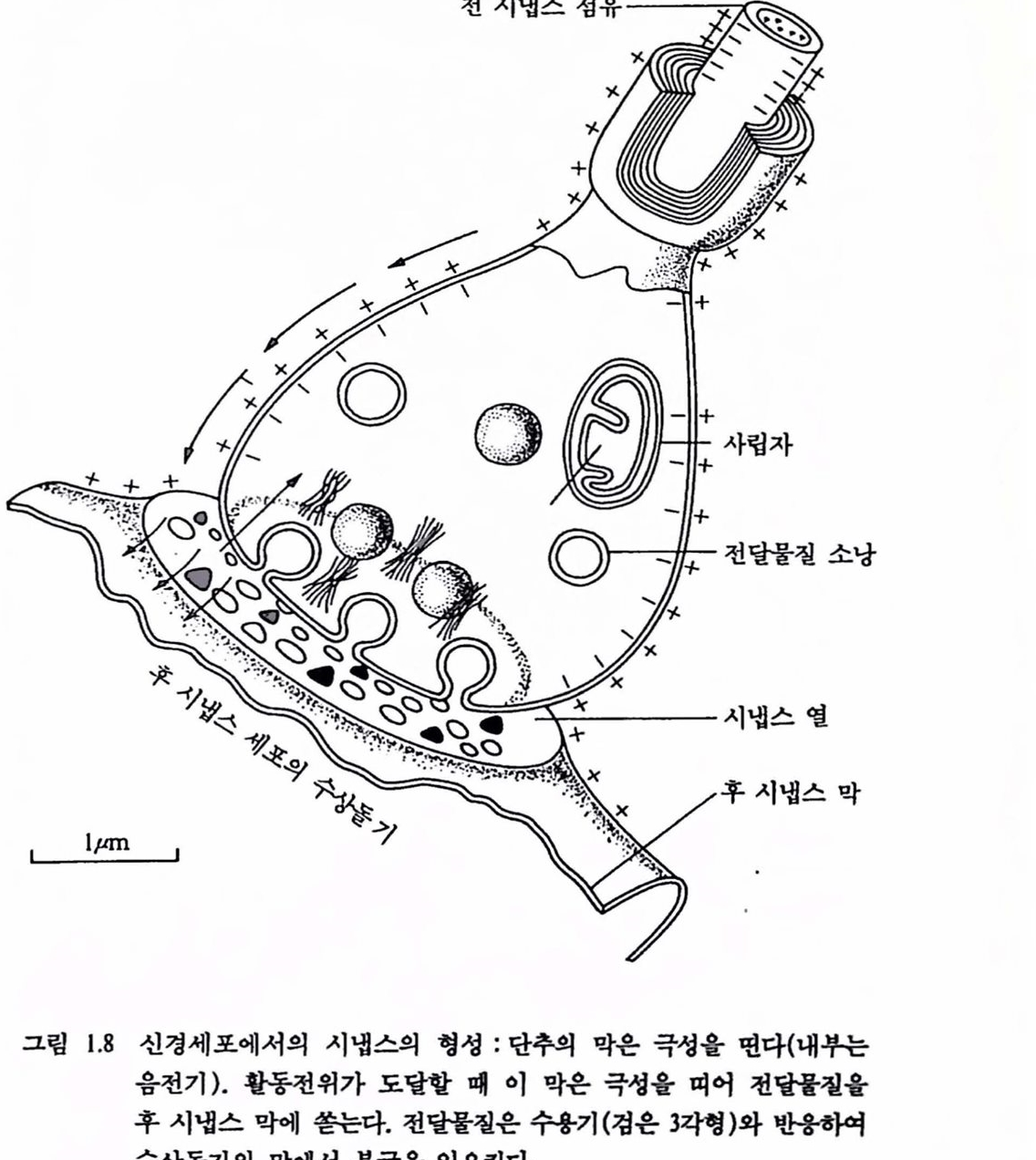 전 시냉스성유
전 시냉스성유
그림 1.8 신경세포에서의 시냉스의 형성 : 단추의 막은 극성융 연다(내환는 음전기). 활동천위가 도랄할 빼 이 막은 극생훌 며어 천당톨철융 후 시냉스 막에 쏟는다. 천달물질은 수용기(검윤 3각형)와 반용확여 수상돌기의 막에서 푼극율 일으킨다.
신호는 특정한 신경세포를 활성화시킨다. 신경섬유의 어떤 집합에서 신호가 복호되는 것은 시냉스에 의해 뇌의 특정 세포가 활성화되기 때문이다.
이런 방식으혹 〈부호〉와 〈복호〉를 설명하는 것이 유용하기는 해도 메시지가 〈이해되는〉 최종의 장소가 있다는 것과는 거리가 멀다. 뇌에는 이런 것이 일어날 콧이 없다. 행위에 의해 〈복호〉가 이루어질 뿐이다. 개는 뼈다귀를 깨물 때만 그것을 인식한다. 우리들에게서도 어떤 의미의 이해는 행위에 대한 준비를 내포한다. <이해>라는 말은 〈가정〉과 같은 뭇이다. 뇌는 끊임없이 유용한 행위를 위한 준비인 가정을 해 나가고 있다. 2.5. 뇌의 제어와 통신 최근 신경생리학의 연구로 신경 시스댐에서 정보가 부호화되고 전달되어 복호화되는 방법이 밝혀지고 있다. 그러나 동물이나 인간은 외부로부 터의 자극에만 활성화되지 않는다. 따라서 뇌는 자체적으로 제어를 수행하는 능동요인이라는 개념으로 되돌아가야 한다. 신경세포들은 외부로부터 자극을 받을 때까지 버활성일 필요는 없다. 대다수의 것들은 결코 쉬지 않고 끊임없는 사이클 활동이나 감도를 변화시킨다. 따라서 업력되는 임펄스의 영향으후 방전 주띄수가 변화된다. 신경 시스댐의 전체는 활동올 중지하지 않는다. 가장 쉬운 예로 호홉 중추의 세포는 신경 임펄스를 태어나서 처음 숨쉴 때부터 마지막까지 매 수초마다 규칙척으로 혜, 근육으로 내보낸다. 이런 리듬의 어떤 것은 세포막의 타고난 성질 때문인 것도 었다. 다른 것들은 신경세포의 주기척 연쇄로서 끊임없이 서로률 다시 흥분시킨다. 이러한 수백만 개의 세포들이 어떻게 함째 작통하여 언간 행위의 프로그랩을 만드는가를 이해하는 데 부단의 노력올 기울여야 한다. 두뇌의 연구로서 우리는 철학척 이해와 질병이나건강에서 우리 자신을 돌보는 데 대한 도구로서 그것을 개선하는 데 우리의 노력을 경주해야 할 것이다.
2.6. 뇌의 계층제어 신경행위를 기술하는 고전적인 단위는 신경세포와 그것들의 연결 관계였다. 우리들 자신에서와 같이 복잡한 환경에서 생명의 행위는 매우 정교한 계층제어에 의해 영위되며 우리들 일상생활의 양식을 만들어 내기 위해 많은 것들이 조화를 이룬다. 두뇌에 대하여 말하고 생각할 수 있는 어떤 도식이나 시스댐을 찾아보기로 하자. 이와 같은 도식은 왜 필요한가? 어떤 의미에서 두뇌는 함께 일하기 위해 이와 같은 도식을 필요로 하기 때문이다. 두뇌의 동작목표는 생명을 유지하기 위한 행위를 만드는 데 있기 때문에 우리가 살아가는 복잡한 세계와 직접 관계된 모델 또는 가젤이나 프로그램의 집협을 그 내부에 갖고 있어야 한다. 이것은 말로는 쉽지만 이들 모델이 무엇이며 이것플은 어떻게 신경세포와 그것들의 행위를 이루어 나가는가를 알아내는 일은 쉽지 않다.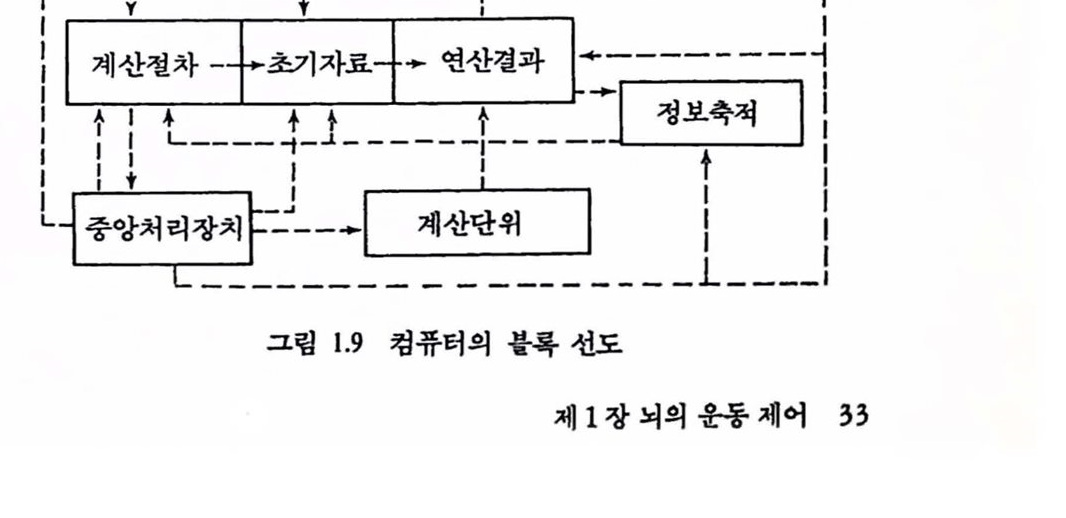 문제강關 -----1 蘭--←당
문제강關 -----1 蘭--←당
그립 1.9 컴퓨터의 블륙션도
컴퓨터는 어떤 의미에서는 인공두뇌이다. 전자식 컴퓨터가 처음 나타났을 때 이것은 신경 시스댐과 공통의 특정을 갖고 있다고들 생각하였 다. 그렴 1.9의 블록 선도는 컴퓨터의 기본 구성부품과 관계를 나타낸 것이다. 컴퓨터에 쓰이는 언어로 두뇌기능을 기술할 수 있는 틀이 았을까? 이제 컴퓨터는 두뇌 연구에 쓰이는 새로운 실험동물의 한 종류로 여겨진다. 확실히 우리는 트랜지스터와 각종 전기소자로 만들어지지는 않았으고로 우리가 고려할 것은 컴퓨터 소프트웨어의 원리뿐이다. 컴퓨터 프로그램의 대비는 가장 일반적인 의미로 보면 도움이 된다. 두뇌에서의 청보가 어떻게 축적되는지 모르기 때문에 정보의 주소법과 축적에 대한 대비는 상사시키기 어려우며 아마도 컴퓨터와 두뇌에서 동일한 원리는 척용되지 못할 것 같다. 이제 쓸모 있는 것은 계층제어의 개념인데 이것은 아주 복합적인 조직, 예를 들면 군대나 공장의 제어를 연구하는 사람들이 알아낸 것이며 서로 다른 레벨에서 기능을 분리하는 것을 기본으로 한다. 따라서 각개의 레벨은 최소의 정보와 기억을 갖고 운영할 수 있다.
컴퓨터와 뇌는 계층적 시스댐을 광범위하게 이용한다. 업력신호는 몇 개의 센터를 통과하는 동안 각 단계에서 훨씬 추상적이거나 일반척인 정보로 추출된다. 운동근육에서도 이와 비슷하여 행위과정에 대한 일반척언 지시는 최고차 레벨에서 이루어지며 곧 척추와 칼은 보다 아래 레벨에서 수행의 세부사항이 조직된다. 인간의 뇌가 계충조직언지에 대해서는 논란이 많다. 그립 1.10에서 보는 바와 같이 두뇌에는 물리척으로 계충을 이룬 센터들이 나열되어 었다. 또는 가장 하위 레멜이 척추, 그 위가 뇌간(brain stem)이며 가장 위가 전뇌 (forebrain)의 3레멜로 크게 나눌 수 있다. 대뇌피질은 제어의 대부분올 우선척으로 수행한다. 아주 중요한 제어는 다수의 하위 레벨에 서 이루어진다. 척추는 신체와 사지운동의 제어에 대한 기초 반사기구 (그렴 1.7 창조)를 갖고 있다. 소뇌(cerebellum)는 여러 기능어 있지만 운동이 정상척이고 척절하게 이루어지도록 타이멍올 제어한다. 뇌의중앙에 위치한 망상 시스댐(reticular system)은, 예를 들면, 수면파 각성 제어와 같이 모든 정지를 활성화하는 중심기능을 갖는다. 시상하부(hypothalamus)와 주변영역은 기준 또는 시스댐을 포함하여 생명유지에 펼요한 모든 행위를 두뇌가 하도록 해준다. 두뇌의 가장 큰 부분인 대뇌피질은 과거에 축적된 기억의 기록꽉 입력정보를 분석하여 해야 할 일을 결정하는 데 관계된다. 배열은 계층적이지만 피철은 개별적인 근육의 운동과 같은 아주 미시적언 제어를 수행한다. 이런 것으로 미루어보아 사람의 뇌의 활동은 전체척언 하나를 이루어 다른 종에 비해 완전히 통합된 것임을 알 수 있다.
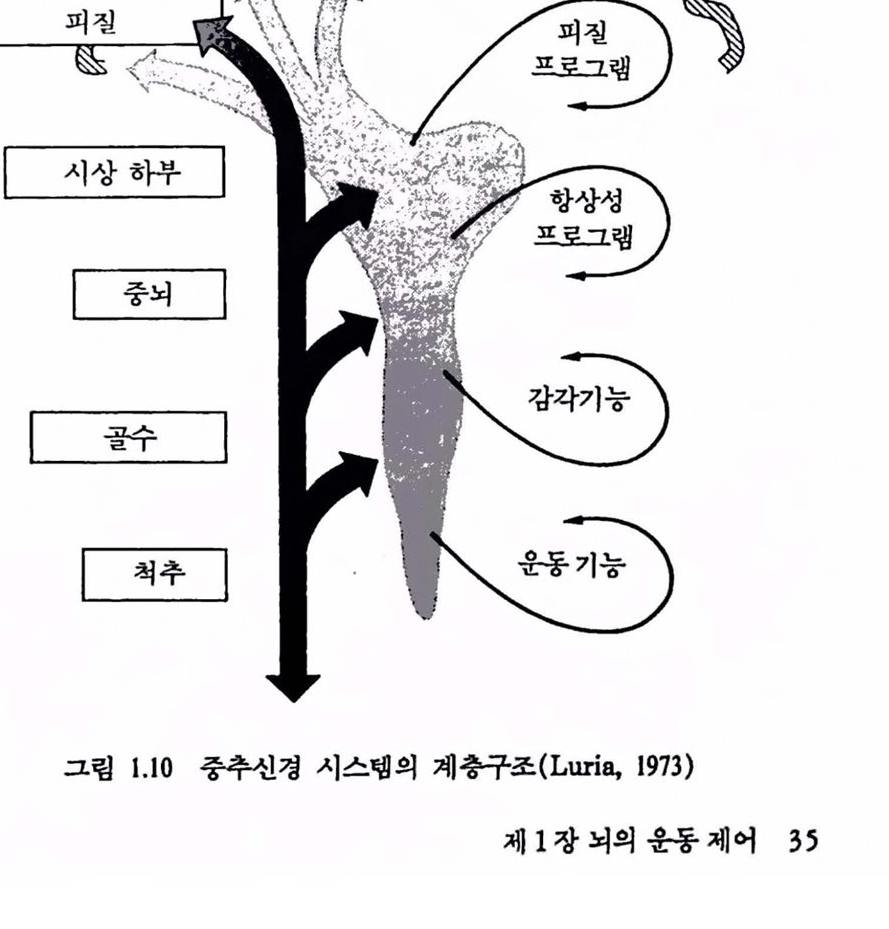
그림 1.10 중추신경 시스댐의 계충구죠(Luria, 1973)
지금까지 살펴본 사실들에서 우리가 배울 수 있는 것은 우리가 찾고 있는 뇌의 모댈이나 도식은 동기를 포함하여 생명을 이끌어 나가는 방식이다. 사람이나 풍물은 부분적으로는 타고나거나 부분적으로는 학습에 의한 명령의 프로그랩에 따라 활동의 양식을 갖는다. 이런 프로그램들은 계층구조와 신경 시스댐의 활동으후 쓰여지거나 표시되어 있다. 이런 것툴은 기관을 유지하는 행위를 생성하는 의미에서 세계를 모델령하는 데 기여한다.
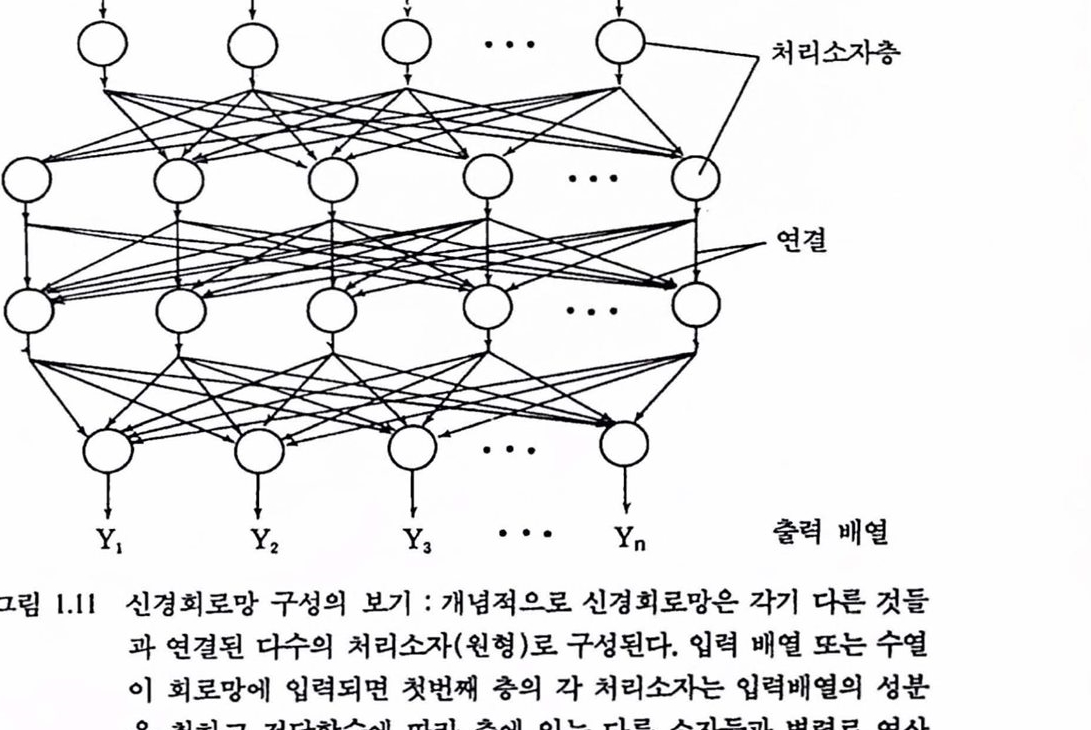
2.7. 두뇌 모델링에 대한 전망 지금까지 우리는 뇌의 구조와 기능을 살펴보았다.이 놀랄 만한 기관의 가장 두드러진 특정은 서로 다른 수많은 장소에서 서로다른 수많은 계산이 동시에 곧, 병렬로 처리되는 데 있다. 각개의 감각-운동 시스댐은 분리된 계산구조이며, 각개의 뉴련은 분리된 계산 장치이며, 각개의 핵 또는 피질의 착은 부분은 계산 모율로서 업력되는 다중변수에 대한 수리 기능을 수행할 수 있다. 뉴련은 기본단위로서 회로망과 시스댐을 구성한다. 앞서 말한 바와 같이 두되는 컴퓨터는 아니다. 다시 말하지만 뇌는 직렬처리와 다지렬 구조, 수정할 수 없는 연결과 기억 뱅크들을 갖는 폰 노이만(von Neumann)형 컴퓨터와는 닮지도 않았다. 그렇지만 뇌는 근본척으로 다른 스타일의 컴퓨터처렴 보인다. 뇌는 학습에 따라 달라지며 통일한 창소에서 기억과 정보처리가 이루어지고 소챔은 디지털보다 는 아날로그형이며 버교척 고장을 허용한다. 확실히 두뇌는 병렬기계이 다. 더구나 자체의 자연도태에 따라 빠른 계산과 늦기는 하지만 청확한 계산을 해낼다. 두뇌와 유사한 컴퓨터 모댈의 발명은 극척언 개념의 변화를 가져왔다. 신경회로망 모멜은 척철한 레멜의 추상화를 시도하여 신경 시스댐에 서 뉴련의 회로망올 지배하는 계산원리률 이용한다(그렴 1.11 창죠).이 모델들은 뉴런과 유사한 단위, 단위플을 연결하는 축색과 유사한 도선, 도션을 연결하는 데는 가중치를 갖는다. 이련 모델플은 학습이 가능하다는 것이 밝혀졌다. 이런 학습의 열쇠는 시냉스의 가중치(척응계수) 수정에 대한 알고리즘, 곧, 앞 단계에서 업출력 시행에서 생긴 오차의 함수로서 시냉스의 가중치를 조정한다. 이런 알고리즘에 의해 회로망은 훈련---간단히 프로그램 되는 것과 정반대---되고 놀랄 만큼 복잡한 엄무를 수행한다. 이련 것은 기술척으혹 매우 중요한데 지금까지의 발견은 실제 신경 시스댐이 시냉스 수정을 위해 사용하는 각종 전략이
 X2 입력 배열
X2 입력 배열
그림 1.11 신경회로망 구성의 보기 : 개녕척으로 신경회로망은 각기 다른 것틀과 연결된 다수의 처리소자(원형)로 구성된다. 업력 배옐 또는 수열 이 희로망에 업력되면 첫번째 충의 각 처리소자는 업력배멸의 성분을 취하고 전달함수에 따라 충에 있는 다른 소자들과 병렬로 연산올 시행하여 결과률 아래충에 있는 처리소자에 보낸다. 결과는 업력과 연상된 특성을 나타내며 업력과 가중치는 시간에 따라 변하기 때문에 회로망은 척웅되며 학습이 이루어진다.
밝혀지지 않았기 때문에 이론에 머물 수밖에 없다. 알고리즘이 알려지지는 않았지만 신경과학과 회로 모델령의 협동연구로 이것들이 찾아지고 있으며 신경 계산장치가 실용화되고 있다(Hecht-Nielsen, 1988).
신경 시스댐의 모델령은 현재로서는 초보적이어서 보거나 배우는 것에 대한 어떤 모델도 갖지 못했으나 회로망 모델과 같은 연구가 진척됨에 따라 해답을 얻게 될 것이다. 신경 시스댐의 연구방법으로 최근 일부 시도되고 있는 것의 하나가 생리공학이다. 이컷의 목적은 공학적 수법에 의한 생명현상의 해명에 있다. 종래의 분석적 수법에서는 복잡한 신경 상호작용에 기초를 두고 실현시킨 동척 현상과 기구에 관해 충분한 지식을 얻는 것이 지극히 어려웠다. 이것을 위해서는 시스댐 이론과 제어이론 등 동적 총합하 기법에 의해 각 분야에서 얻은 지식을 재구성하고 이것을 확인, 총합하고 충실한 모델을 창출하는 것이 필요하다. 신경 시스템은 일반적으로 비선형, 피드백 등을 포함한 복잡한 모델로 되어 있어 그것을 해석하는 것이 지극히 어렵다. 그러나 컴퓨터 시율레이션에 의해 수치해를 얻을 수는 있으며 이것에서 상호작용하는 요소들간의 관계가 얻어지고 새로운 이론을 세울 수 있게 된다. 시률레이션만으로 새로운 미지의 기본척 지식을 얻을 수는 없으나 거기에 이르는 과정을 촉진하는 수단으로서는 아주 효율적이다. 이것에 더하여 최근의 디지털 신호처리, 시스댐 식별법, 회화형 컴퓨터 시스댐은 신경 시스템의 베일을 벗기는 새로운 환경을 조성하는 데 한품을 하고 있다. 3. 뇌의 운동제어 3.1. 운동제어 시스템 : 뇌와 외부 세계와의 통신 뇌의 운동 제어 시스댐은 사유, 지각, 감청을 운동으로 바꿔 주기위해 존재한다. 운동은 다수의 제어 시스댐이 복합척으로 상호작용하여 나오는 최종의 산물이다. 정상척언 환경 또는 척어도 광벙위하게 받아들일 수 있는 상황하에서 뇌는 운동 시스댐을 통하여 외부와 통신을 행한다. 따라서 운동을 주관하는 제어신호는 운동이나 근육 시스댐의 어떤 표현으로부터 나오지 않으면 안 된다. 따라서 제어신호는 신경활동, 근육활동, 근육의 힘(근력), 사지운동 및 음성을 표현할 수 있다. 신경활동 및 근육활동〔멀리볼트 (mV)에서 마이크로볼트(μV)의 전압범위와 10~100Hz의 주파수 성분을 갖는 전기신호〕은 매력척언 제어신호원이 되어 왔다. 앞에서 안구의 운동을 주관하는 신호는 운동 뉴런의 발화에 따른 신경제어 신호임을 알았다. 아직까지는 이런 신호틀을 인공척인 장치의 제어에 활용하는 데 신뢰성이나 반복성이 부척합한 것으혹 밝혀졌다. 그렇지만 이런 신호들은 신경 시스댐의 기능에 대해 무엇인기를 말해 주는 것은 틀렴없다. 앞으로 생체 신호처리 기술의 발전에 험업어 이들 신호를 각종 운동의 제어신호로 활용하는 데 많은 발전이 기대된다. 결국 운동제어에 대한 전기신호를 얻는 것이 신경제어 시스댐 연구의 주된 과제이다.
 관절, 피부 골격근
관절, 피부 골격근
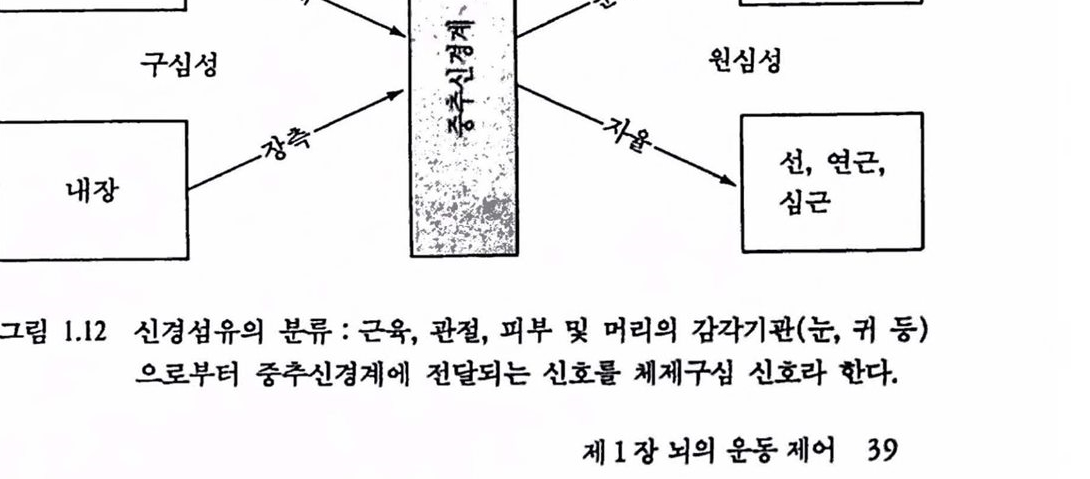
그립 1.12 신경섬유의 분류 : 곤육, 판철, 피부 및 머리의 감각기판(눈, 귀 퉁)으혹부터 중추신경계에 천달되는 신호훌 체제구섬 신호라 한다.
먼저 운동 시스댐의 구조와 기능을 살펴본 다음 근육제어에 대해 간단히 살펴보기로 하겠다. 자세와 운동의 제어는 중추신경계의 가장 중요한 기능 가운데 하나이다. 이것을 주로 책엄지는 운동제어 센터라고 하는 구조는 신경 시스댐의 아주 색다른 부분에 위치한다. 이것의 구조는 그렴 1.10에서 보듯이 추체로 배열되어 있으며 운동조직은 계층적 특성을 갖는다.
신경 시스댐의 운동제어 기능을 고찰함에 있어 자세와 운동에 관계된 CNS 센터는 구심성(감각)정보의 흐름을 끊임없이 받아야 적절한 행위를 한다는 것을 염두에 두어야 한다. 자세와 운동제어에 있어서 감각기관의 역할을 강죠하기 위해 근육활동에 관계되는 구심 및 원섬 기능의 전체를 나타내는 감각운동 시스댐이라는 말을 지주 쓰게 된다. 그렴 1.12는 기시(origin)와 기능에 따라 신경섬유를 분류한 것이다. 환경이나 신체의 변화에 대해 수용기로부터 중추신경계로 들어오는 구심성과 중추신경계로부터 말초에 정보를 전달하는 원심성 신경섬유로 분류되며 골격근 섬유로의 원섬성을 운동 원섬신호라 한다. 운동제어에서는 체제 구심성 및 운동 원심성 신호가 관계된다. 구섬 업력에 따른 운동행위는 척추 레벨에서 아주 확실하며 여기서 개별 수용기(근방추 수용기)는 상대적으로 천면 일률적으로 운동 뉴련에 연결돼 반사궁을 형성한다. 그렴 1.13에는 척추와 척추위(supraspinal cord)의 운동 센터와그것들의 가장 중요한 연결 관계를 나타내었다. 간단히 하기 위해 모든 감각입력은 왼편으혹 모았다. 오른쪽 열은 운동 행위시의 역할을 나타낸 것이다. 운통피철은 프로그램과 수행간의 전이 위치에 있다. 척추 위에는 뇌간, 운동피질, 기저핵 및 소뇌가 위치한다. 각개의 센터는 정보처리의 장소이고 입력갱보를 되내보내는 곳이다. 가장 열쇠가 되는 위치는 운동피질로서 척추에 었는 운동 센터와 칙접 · 간접으혹 연결되어 있으며 어떤 것은 뇌간과 어떤 것은 고차운동 센터언 소뇌와 대뇌 기저핵과 연결되어 었다. 그렴 1.14는 뇌와 척추에 있어서 운동 센터의 위치를 측연으로 나타낼 것이다. 모든 운동 센터는 두뇌의 좌우에 짝을 지어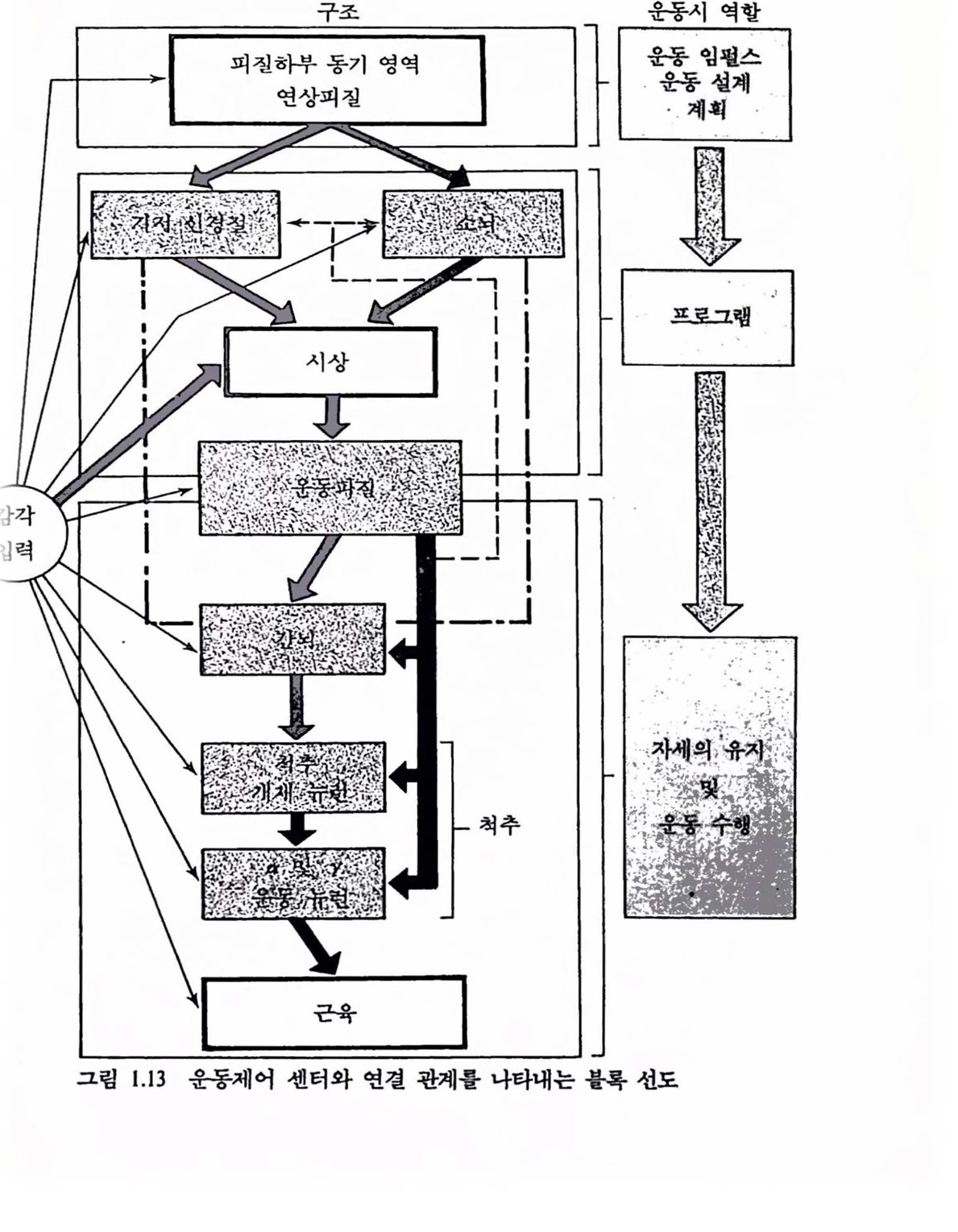 구조 운동시 역할
구조 운동시 역할
있으며, 그렴 1.14에서 척추 단연의 하부는 운동 뉴런을 나타낸다.
운동피질은 거의가 뇌의 표면에 놓여 있다. 소뇌는 뇌의 나머지와는 아주 다르며 그럽 1.13에서 정선으로 구심 및 원심신호의 흐름을 나타내었다. 운동 시스댐을 하나의 전체로 조화시키는 센터는 바로 이 소뇌이운동피질(중심 전회)
발 다리 몽종 팔 손가락 얼굴 업술 혀 기저 신경절 운동뉴런그림 1.14 운동제어 센터의 해부학척 구조
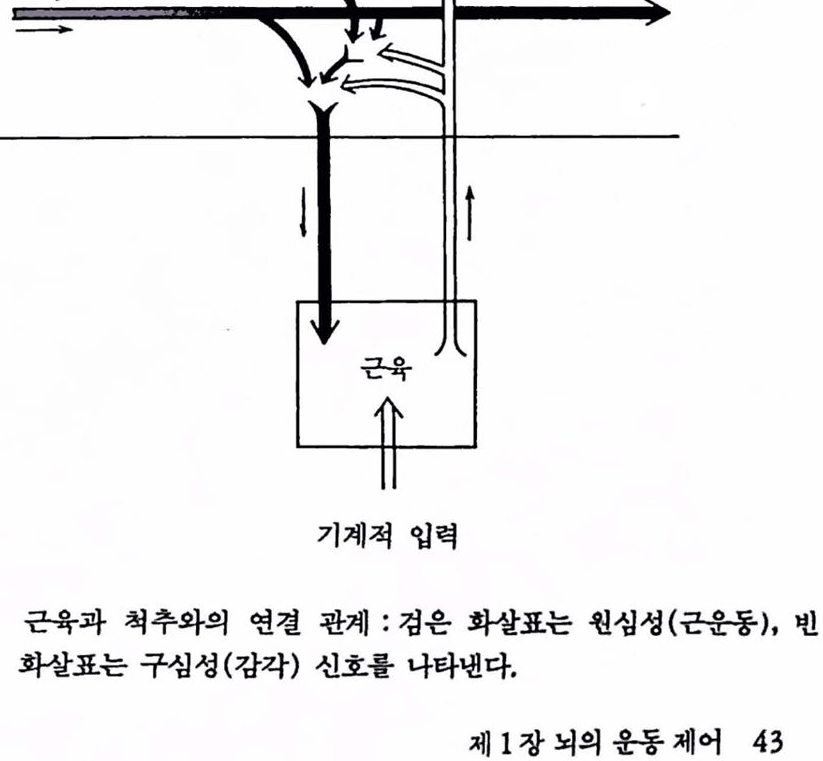
며 중추신경계의 모든 레벨에서 운동회로의 활동을 조화시킨다. 그렴 1.15는 그렴 1.13에서 최하위 레벨언 척추와 근육과의 연결을 닫힌 루프로 나타낸 것이다. 근육으로부터 엄펼스를 나르는 감각 뉴련은 운동뉴런과 연결되어 있으며 임펄스는 다시 되돌려 보내진다. 개별척으로 각개 근육의 활동을 제어하는 닫힌 루프는 이렇게 하여 형성된다. 이것이 운동 시스댐의 기초 제어기구이며 이 회로의 중계를 통하지 않고 운동이 없을 때 반사 또는 수의가 발생된다. 간단한 반사에도 이 회로가 지율적으혹 동작할 수는 있지만 대부분은 이것들의 활동은 고차 센터에 의해 제어된다. 이 회로는 각종 요구에 따라 근육의 길이와 장력을 조절한다. 이것들은 기계적 업력(예를 틀면, 근육의 신장 및 고차 중추로부터의 신경입력) 등에 말초적으로 응답하며 빨리 응답한다.
 ---‘-- μY1느 ι‘ i투 는1 →」、‘‘
---‘-- μY1느 ι‘ i투 는1 →」、‘‘
그렴 1.15 근육과 척추와의 연결 판계 : 검은 화할표는 훨심성(근운동), 번 화찰표는 구심성(감각) 신호롤 나타낸다.
- 제어지 뉴-
제어패상 t--그림 1.16 닫힌 루프 시스댐 : A. 피드백을 갖는 일반화된 반사. B. 근육과 근방추의 병렬 배열에 의해 나타난 피드백을 갖는 신장반사. c. 가장 간단한 제어회로
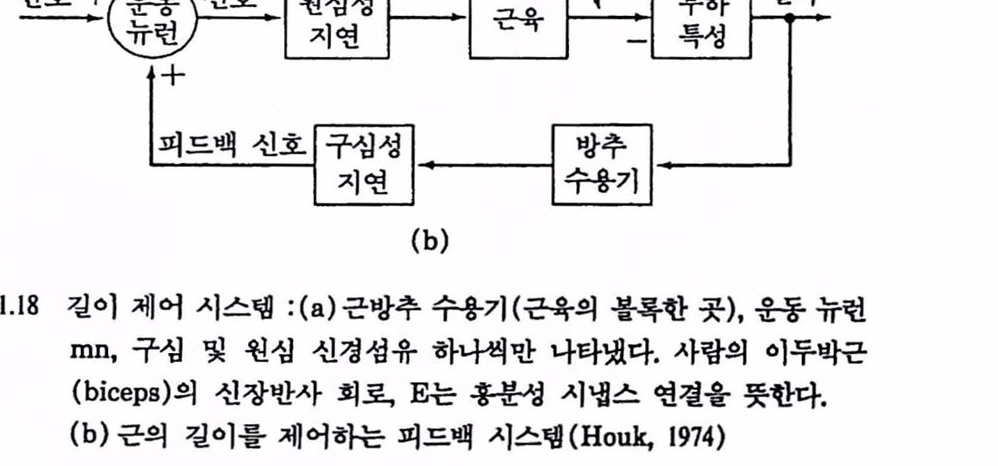
이와 같은 닫힌 루프에 대한 것을 제어공학의 개념으로 나타낸 것이 그렴 1.16이다. 그렴의 C에서 제어기는 제어대상에 영향을 미치며 제어 대상에서 어떤 변수(양)가 피드백되어 다시 제어기에 작용하여 제어에 영향을 미친다. 이 그렴은 근육의 길이 제어 시스댐으로 신장반사를 나타낸 것이다. 근육에서의 피드백 제어를 본격적으로 셜명하기 전에 피드백 제어의 개념올 간단히 살펴보기로 하자.
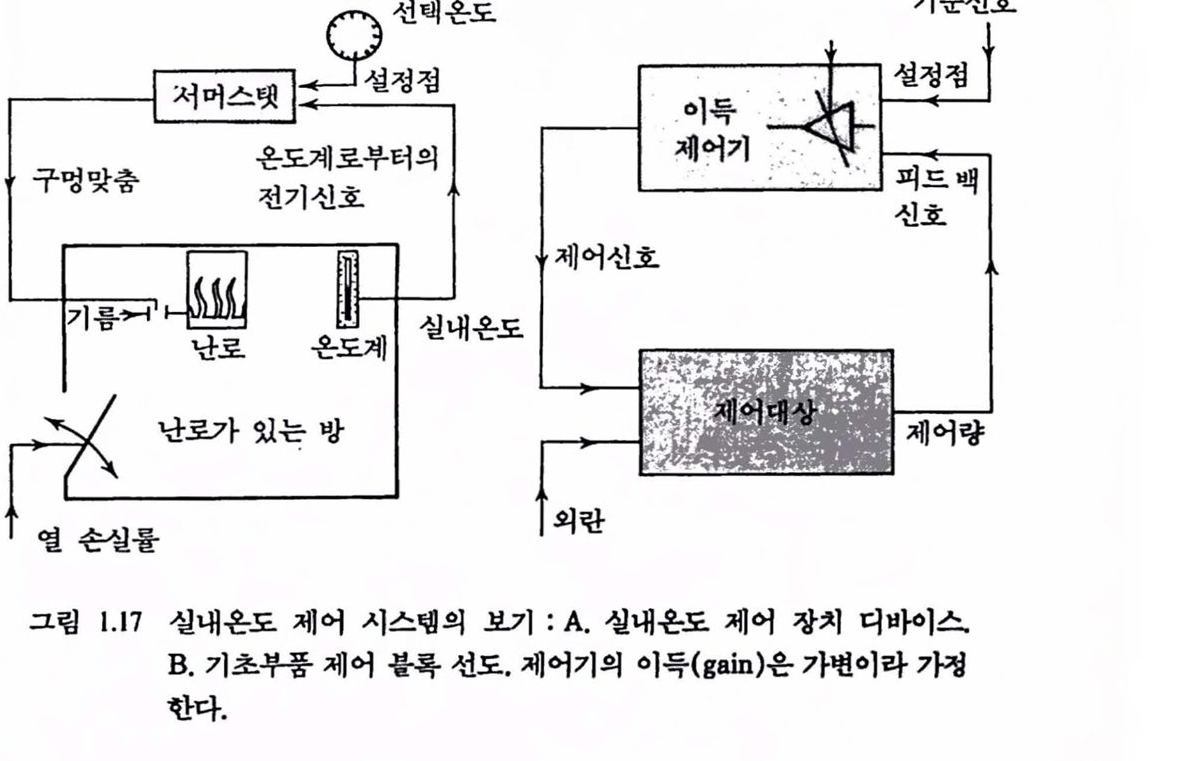
3.2. 제어회로의 구조 : 근육에서의 피드백 제어 운동 시스댐의 신경제어를 보다 더 갚이 이해하기 위해 제어기, 제어신호, 제어대상, 감지기(센서)에 대한 표준 제어회로의 구조를 먼저 살펴보기로하자. 얼반 가정이나 사무질에서 실내의 옹도를 연중 척절한 레멜 근처에서 가능한 한 일정하게 유지하고자 할 때 설내 온도에 영향을 미치는 여러 가지 요언에 대해 얼청한 천략으로 대버하여 온도차이를 곧바로 보상할수 있다고 하자. 가령 외부온도가 아주 낮아지는 것을 알 수 있는 11월에서 다음해 3월까지는 간단히 방열기(또는 난로)의 열공급 손잡이를 모두 열어 놓아 따뭇하게 지낸다고 한다면 이런 종류의 제어는 현재 설내 온도에 관한 피드백이 없어 열련 루프 제어라 한다. 이런 경우는 때로 원리는 쓸모가 있지만 단점이 많은 것이 확실하다. 실내 온도 제어나 그 밖의 많은 경우에 실내 온도 자체를 측정하고 여기에 맞춰 난로의 연료공급 밸브를 열어 주는 것이 훨씬 현명하다. 이런 과정을 닫힌 루프(또는 피드백, 되먹임) 제어라 하며 온도를 한번 맞춰 놓으면(셜정) 온도 변화에 따라 자동척으로 공급 밸브가 열렸다 닫혔다 한다. 그렴 1.17에서 B의 블록 선도에는 이런 시스댐의 구성 부품을 나타내었으며,
 A B
A B
그립 1.17 설내온도 제어 시스댐의 보기 : A. 실내온도 제어 장치 디바이스 B. 기초푸풍 제어 블륙 선도. 제어기의 이득(gain)옴 가변이라 가정한다.
A에는 실내 온도 제어 시스댐의 대응 부품을 셜명해 놓았다. 제어 또는 조절량은 현재의 값(실내온도)이다. 조철이 이루어지는 곳의 실제 구조를 제어대상이라 하며 이 경우는 난로가 있는 방이 된다. 감지기는 현재의 값을 모니터하는 온도계이며 이것의 출력은 피드백 신호(온도계에서 나오는 전기신호)이다. 벽에 붙은 서머스랫(thermostat)은 제어기이며 피드백 신호와 기준신호 또는 설정치(setpoint, 서머스랫 다이얼 눈금)는 제어기 안에서 비교된다. 현재의 온도(피드백 신호)와 설정된 값이 다르면 이 비교결과는 오차신호가 된다. 이것에 근거하여 제어기는 제어신호를 발생하여 이것이 제어대상으로 하여금 현재의 값을 변화시키도록 작용하여 피드백과 셜청치가 일치하도록 한다. 제어신호의 작용(강제함수, forcing function)은 난로의 연료공급 밸브 맞춤에 해당한다. 따라서 외부교란(perturbation, 창문이나 문을 여는 데 따른 열손실이나 외부온도의 변화)에도 불구하고 오차신호를 최소화하기 위하여 닫힌 루프 제어가 연속척으로 수행되어야 한다.
이와 같은 조절기에서 가장 중요한 것은 오차신호를 발생시키는 비교기이며 기준신호에서 피드백 신호를 빼면(전기척이나 기계척으로) 오차신호가 된다. 이와 같은 제어신호를 부궤환(negative feedback)제어라 하며 제어효괴를 발생시키는 결정척인 부호의 변화는 루프의 다른 지점에서 이루어진다. 지금까지 살펴본 실내온도 제어 시스댐은 항상 똑같은 출력을 목척으로 하며 조절기라고 한다. 어느 때든지 설정점은 새로이 원하는 레엘에 맞출 수 있다. 더 나아가서 계속하여 설정점을 변경하면 이와 같은 시스댐은 한 개의 제어량이 다른 것을 추종하게 하는 장치가 된다. 이런 제어 시스템을 서보메커니즘(servomechanism)이라 하는데 출력은 위치, 각도와 같은 기계척언 양이 보통이다. 지금까지의 개념올 곤육제어에 활용해 보자. 근육의 길이를 제어량으로 잡으면 근방추(muscle spindle)는 피드백(구성섬유에서의 활동)올 발생시키는 감지기 또는 변환기에 해당한다. 제어신호---피드백파 반사에 영향올 미치는 것(고차운동 센터로부터의 〈기준신호>) 두 가지모두---는 운동 뉴런의 활동으로 보내져 길이를 변화시킬 제어대상(건과 관절을 가진 근육)에 작용한다. 마지막으로 시스댐에 반대로 작용하는 외란은 눈에 띄지 않는 부하나 근육피로의 형태로 존재한다. 그렴 1.18은 근육의 길이제어를 냐타내는 생리학척 부품간의 해부학척 연결과 이에 대응하는 제어 블록 선도이다. 음의 피드백을 위한 부호변화는 실내온도 제어 시스댐에서는 설정치와 피드백을 제어기에서 비교하여
 구성성
구성성
 제어 떤오원차 싱지연성 양너근 력\\ - 특부객하 길이
제어 떤오원차 싱지연성 양너근 력\\ - 특부객하 길이
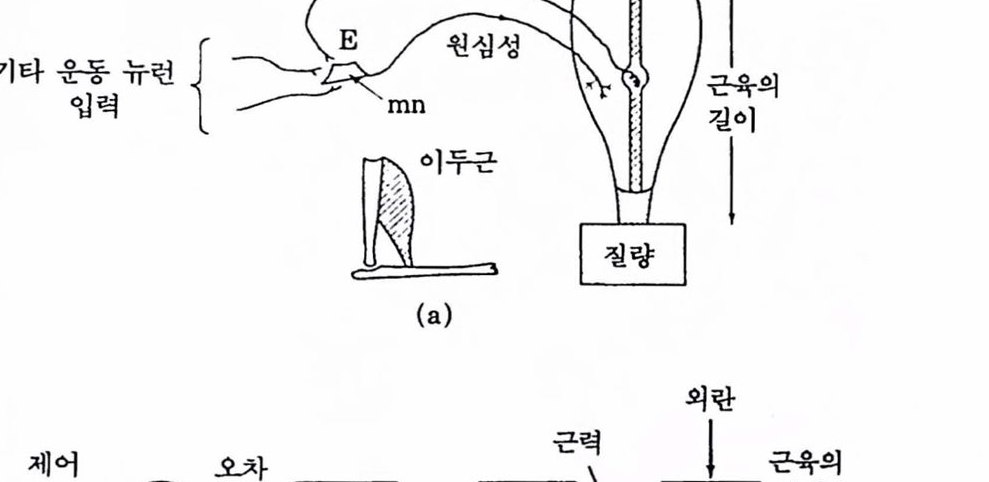
그림 1.18 길이 제어 시스댐 : (a)근방추 수용기(근육의 볼륙한 곳),운동 뉴련 mn, 구심 및 원심 신경섬유 하나씩만 나타냈다. 사랍의 이두벅근 (biceps)의 신장반사 회로, E는 흥분성 시냉스 연결융 풋한다. (b) 근의 길이를 제어하는 피드백 시스댐(Houk, 1974)
얻었으나 여기서는 달라진다. 신창반사 루프에서는 길이가 늘어나연 구심활동도 증가하여 이에 따라 알파-운동 뉴련의 활동을 증가시켜 모두 〈양(+)>의 효과를 가져온다.
실제로 부호 변화는 근육에서 일어나는데 증가된 알파-운동 뉴련의 활동은 길이를 감축하려 하며 방추로부터 구심신경의 발화를 감축시킨다. 따라서, 곧 근력(muscle force)의 증가는 근육길이의 감소를 유말하여 이에 따라 방추로부터 되돌아오는 신호의 감소를 가져 온다. 따라서 피드백 루프에는 1개의 〈음(-)>의 부호가 있게 된다. 중요한 것은 그것의 위치가 아니라 오차신호의 어떤 증가는 결국 피드백 경로를 거치는 동안 제어기로 하여금 오차신호의 감소를 유도한다는 사실이다. 그럽 1.19는 길이 제어 시스댐의 기계척인 아날로그로서 다음에 셜명할 걸항근 시스댐(antagonistic muscular system) 과 그것의 가역(reciprocal) 신장반사를 기계척으혹 나타낸 것이다. 스프링은 길이 피드백의 조절기 기능을 나타낸 것으로서 외란에 의한 힘은 스프링을 늘려 명형(희망)위치로부터 부하의 실제 위치를 벗어나게 한다. 랙(rack)과 피니 언(pinion)은 제어 신호에 응답하는 시스댐의 거동을 나타낸 것으로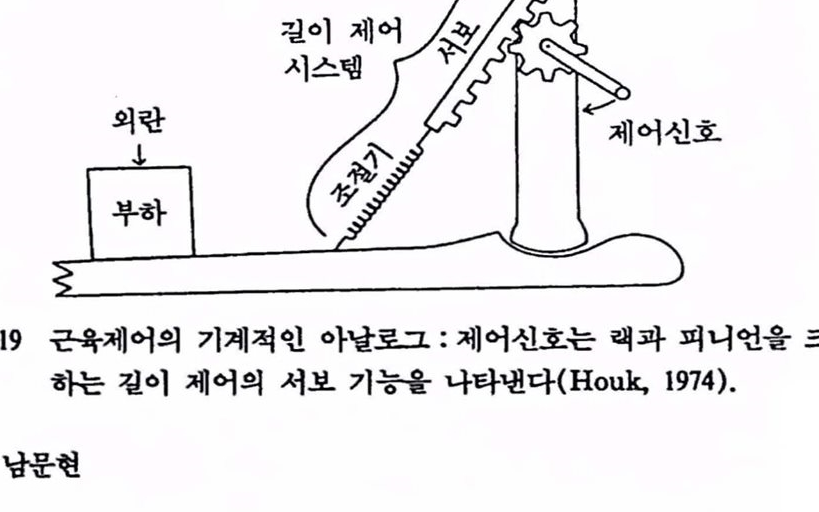 빡
빡
그립 1.19 근육제어의 기계척언 아날로그 : 제어신호는 래과 피니언올 크랭크 하는 킬이 제어의 서보 기능윷 나타낸다(Houk, 1974).
길이 피드백의 서보(servo) 기능이다. 제어 신호는 랙과 피니언을 새로운 위치로 크랭크하는 것으로 생각할 수 있다. 사지의 위치 변화는 부하의 관성 때문에 느리겠지만 자동적으로 수행될 것이다. 여기서 셜명한 근육제어는 피드백 제어 시스댐의 기본 성질을 셜명하기 위해 아주 간략화시킨 것이다〔근육제어에 대한 상세한 내용은 Mountcastle(1980)의 Ⅶ부플 참조할 것〕 .
3.3. 상호 신경지배 : Descartes에서 Stark까지 모델은 실제 시스댐보다 훨씬 간단하고 다루기가 쉬우면서 설제와 같은 행위를 나타내야 복잡한 시스댐을 연구하는 데 도움이 된다. 예를 들연, 지도는 어느 지역에 대한 정보가 담긴 모렐이며 지도로 방향을 가리켜 주는 것이 실제 세계를 언급하는 것보다 훨씬 쉽다. 운동의 신경 제어를 연구하기 위한 모델도 일찍부터 개발되었다. 인간이나 동물에게 가장 기본적인 안구운동(oculomotor)의 제어에 대한 초기 모멜 가운데 하나는 불란서의 철학자 Rene Descartes(1596~1650)가 개발한 그림 1.20의 A와 같은 모델로서 안구가 운동을 하려면 척어도 2개의 근육의 조화된 활동이 필요하다는 것이다. 곧, 인구운동을 위한 회전 토크률 발생시키기 위해 안구에 대해 짧아져서 꿀어당기는 동근(agonist muscle)과 이완하여 늘어나는 길항근(antagnist muscle)이 필요하며 청상척인 안구운동은 한쌍의 근육의 상호 신경지배를 통해서만 가능하다는 것이다. Descartes는 이런 상호 신경지배의 현리를 1626년에 제안했으며 MIT의 Cook와 Stark가 saccade 안구운동 모멜로 재현한 1963년까지는 이 이론은 확립되지 못했다〔주 : saccade 인구운동은 망막(retina) 상의 중심에 위치한 중심와(fovea)에 시야가 오도록 해주는 staccato와 같은 빠른 인구운등을 말하며 위치 포착기능올 수행한다〕. 이 모밸운 Clark, Hsu의 도움으로 인간의 모든 인구운동에 대한 상호 신경지배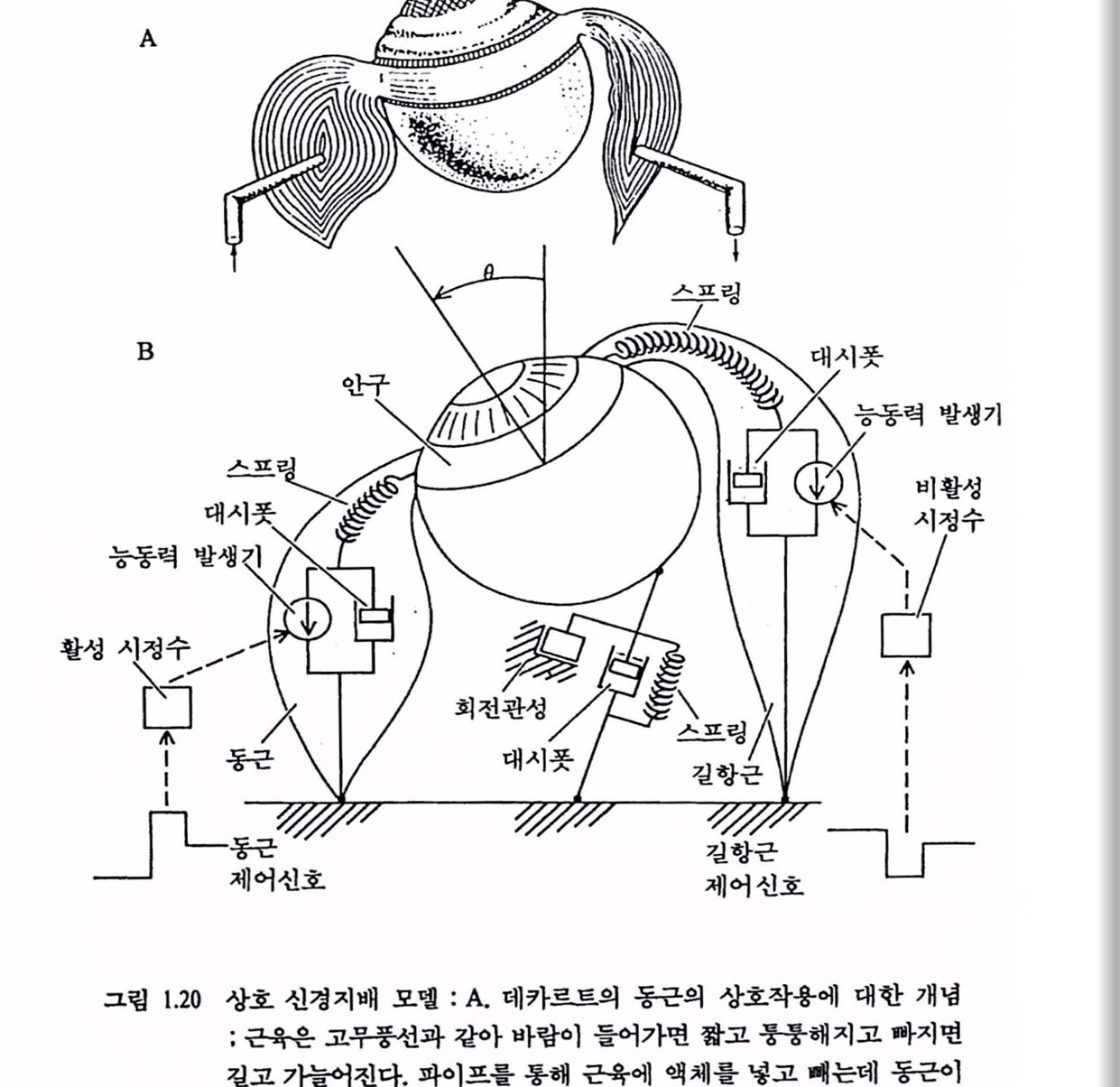 A
A
그림 1.20 상호 신경지배 모댈 : A.데카르트의 동근의 상호작용에 대한 개념; 근육은 고무풍선과 같아 바랍이 들어가면 짧고 풍흥해지고 짜지연 걸고 가늘어진다. 파이프를 통해 곤육에 액체를 넣고 빼는데 동근이 짧아지고 길항근이 걸어지연 안구랜동이 발생한다. B. Stark의 재구성 모멜 : 설명은 본운 창조〔Bahill과 Stark(1979), Sci.Am, vol. 240, no. 1. pp. 108-117에서 옳검〕
모델로 확장되었다. 이 모텔은 생체 수학자인 Richard E. Bellman 의미의 동류정(homeomorphic)을 갖는다. 곧, 모댈파 실제 시스댐의 구성부품간에 직접적이고 세부적인 관계가 있다. 이런 모텔의 장점은 blackbox (두뇌와 같이 내부의 모양을 알 수 없는) 모델을 갖고 신경신호를 입력으혹 시율레이션하여 얻는 안구운동의 출력으로부터 모델의 개별 매개변수가 출력에 미치는 영향과 안구운동 시스댐의 실제 근육과 건(tendon)의 생리학적 성질이 이런 매개변수에 미치는 영향을 연구할 수 있다는 데 있다. 모델의 매개변수는 안구의 관성, 안구의 궤도에서 회전시에 생기는 점탄성(viscoelasticity), 안근과 건의 탄성 및 안근의 피상적인 점성이다(Fenn-Hill-Katz의 험-속도 관계로 밝혀진 근육의 내재성은 근육의 힘 발생기구를 충격홉수기 또는 점성 감쇠소자처럽 감쇠시킨다). 디지털 컴퓨터에 모델을 구현하면서 업력언 제어신호를 변화시키면 saccade 궤도에 미치는 영향을 즉석에서 관찰할 수 있으며 이때는 여러 가지 형태의 제어신호를 실험할 수 있다. 모멜의 결과는 정성 및 정량법을 써서 실제의 운동 출력과 비교되고, 감도해석법올 써서 해석적으로 다루어지고, 시행착오법을 써서 실험을 확정하는 데 매우 중요하다. 그림 1.20의 B는 새로 얻은 모댈이다. 모텔은 훨스-스댐(pulse-step) 제어신호 또는 운동 뉴련의 발화 왜턴(그림 1.20 B의 하단 좌우)을 업력으로 한다. 활성 및 비활성 시정수는 신호를 형활하는 기능을 하여 실제 시스댐에서 동근과 길항근에 신호가 전달되어 처리될 때와 같은 역할을 한다. 안구 및 안근 시스댐의 관성, 첨성, 탄생은 회전철량, 댐퍼(damper, dashpot) 및 스프령으혹 나타난다. 능통력 발생기는 근육의 건이 아닌 내부에서 발생되는 것을 모의화한 것이며 스프랭과 직렬로 연결되어 근육의 탄성을 나타낸다. 모멜의 출력은 안구 위치 θ이며 정연 주시로부터 안구가 회전한 것융 나타낸다. 운통궤도는 신경제어 신호를 충실히 반영한다. 실험에서 얻은 방대한 안구운동 데이터는 Main-Sequence(주 : 별의 표면 온도와 밝기로 운성의 생성 발천옳 연구하는 데 천문학자들이 쓰는 선도로서 일명 Hertzsprung-Russell선도라
고도 함)의 개념을 이용하여 운동의 크기(각도)에 따른 동특성(지속시간, 속도, 기속도) 관계로 나타낼 수 있으며 시률레이션 데이터와 비교한다. 이와 칼은 도구는 안구운동의 본질과 제어특성을 이해하는 데 이주 도움이 된다.
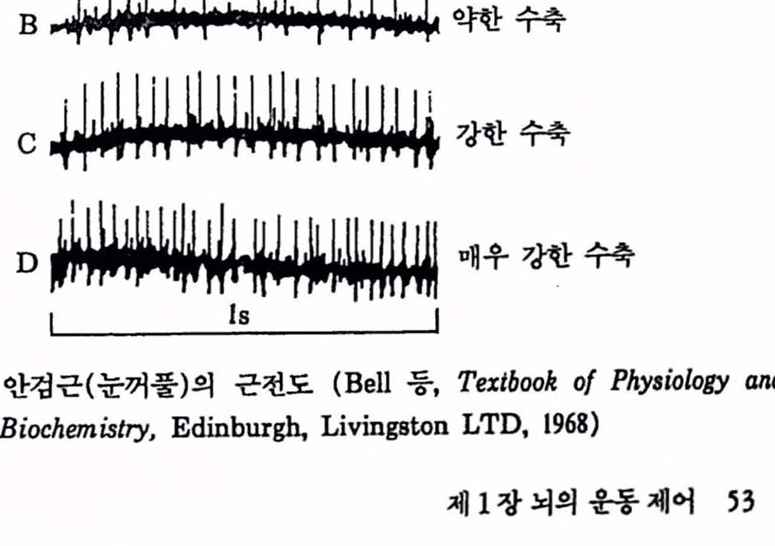
4. 운동제어 모델과 활용 4.1. 모델 만들기 안구나 사지운동의 신경제어 기구를 규명하기 위해서는 정상조건하에서 의도적인 시간최척(intended-time optimal) 운동에 대한 동특성 및 관련 근육에서의 전기적인 활동을 측정 처리하여 얻은 데이터를 바탕으로 모탤을 만드는 방법이 1970년대에 접어들면서 널리 개발되었다. 근전도(electromyogram, EMG)는 어떤 운동에 기여하는 각종 근육의 활동해석에 있어서 아주 중요한 도구이다. EMG의 타이멍을 상세히 정량적으로 해석해 보면 운동제어의 중요한 특정을 알 수 있다. 이 밖에 도 운동파 EMG의 관계를 알기 위해 위치, 속도, 가속도 빛 험이나 토크를 EMG와 동시에 측정할 필요가 있다. 이런 청량척인 연구는 앞절에서 셜명한 모멜의 매개변수간의 관계를 셜명히는 데 아주 유효하다. 먼저 곤전도 신호에 대해 간단히 찰펴보기로 하자. 골격근은 대부분 가늘고 긴 평행인 근섬유 세포의 접합으로 되어 있다. 근섬유는 척추의 운동 뉴련에 의해 신경지배되고 있다. 운동 뉴런으로부터 나온 신경섬유(축색)는 근육의 가운데에서 가지를 쳐 갈라지고 동시에 몇 개의 근섬유 툴 지배한다. 따라서 한 개의 운동 뉴련으로부터 나오는 임펄스는 가지 률 친 모든 곤섬유에 도달하고 동시에 수축올 일으킨다. 아울러어떤운동 뉴런과 그것에 의해 지배되고 있는 근섬유군은 활동을 같이하는 한 개의 최소의 기능 단위를 구성하는데, 이것을 운동단위라 부른다. 근육에서 발생되는 힘은 활동을 가하는 운동단위의 수와 각 운동단위를 나타내는 임펄스의 주파수(빈도)에 의해 결정된다. 운동단위들의 홍분성 엄펄스는 근전신호(myoelectric signal, MES)로서 근전도 장치를 써서 기록할 수 있다. 이렇게 얻은 EMG는 근육의 전위의 세포외 기록이다. 전극은 피부면이나 근육내부에 세포외에 부착한다. 그렴 1.21은 사람의 안검근(eyelid muscle)의 근전도를 설명한 것이다. A에서는 근육이 완전히 이완되어 있어 전위의 변화가 없다. 이러한 상태를 tonic 레벨이라 하여 0이 아닌 값을 설정해야 한다. B, C 및 D는 눈꺼풀에 힘을 주어 눈을 감는 경우로서 임펼스 또는 활동전위가 근전도에 나타나는데 이것은 모두 한 개의 운동단위에서 나온 것틀이다 .B에는 5개의 임펄스를 화잘표로 표시했으며 힘이 가해질수록 임펄스의 번도는 찾아지고 D에서는 매초 31개가 발생되었다. 이 주파수에서 운동단위는 강축(tetanus)에 접근한다. 그림 1.21에서 볼 때 수축력이 중가하면 단일 운동단위에서
 A ,.. 근육이완
A ,.. 근육이완
그림 1.21 안검곤(눈꺼풀)의 근천도 (Bell 둥, Textbook of Physiology and Biochemistry, Edinburgh, Livingston LTD, 1968)
o 45μ/
φ 영L/ χ( 15‘0 00 o 0 Y 판상끈 :*9 AG EMGO AT 700 k0i 500 ms그립 1.22 빠른 수명 머리운동시에 목의 판생근상의 4개 위치에서 얻은 곤전도와 동륙성의 명균랐. 동특성(9, 9, 9)의 변화는 거의 없으나 EMG는 심한 변화률 보언다. AG는 동근, AT는 킬향근이다(피검자 : 필자)
홍분의 주파수가 증가하고 더 많은 운동단위가 증가됨을 알 수 있다. Stark 교수와 필자 등은 EMG와 운동간의 관계를 정량화하여 이것을 제어신호로 하기위해 안구운동 연구에서 사용한 방법을 확대하여 macro system으로 인간의 머리운동을 선택하고 의도척인 시간최척 운동에 대한 적응 특성(Nam 등, 1984)을 조사하고 정상과 부하부과 조건하에서 수평운동의 동특성과 EMG 를 측정, 처리하고 이를 제어신호로 하는 상호 신경지배 모델을 개발하였다(Hannaford 등, 1984 : Hannaford, 1986 : Nam 등, 1989).
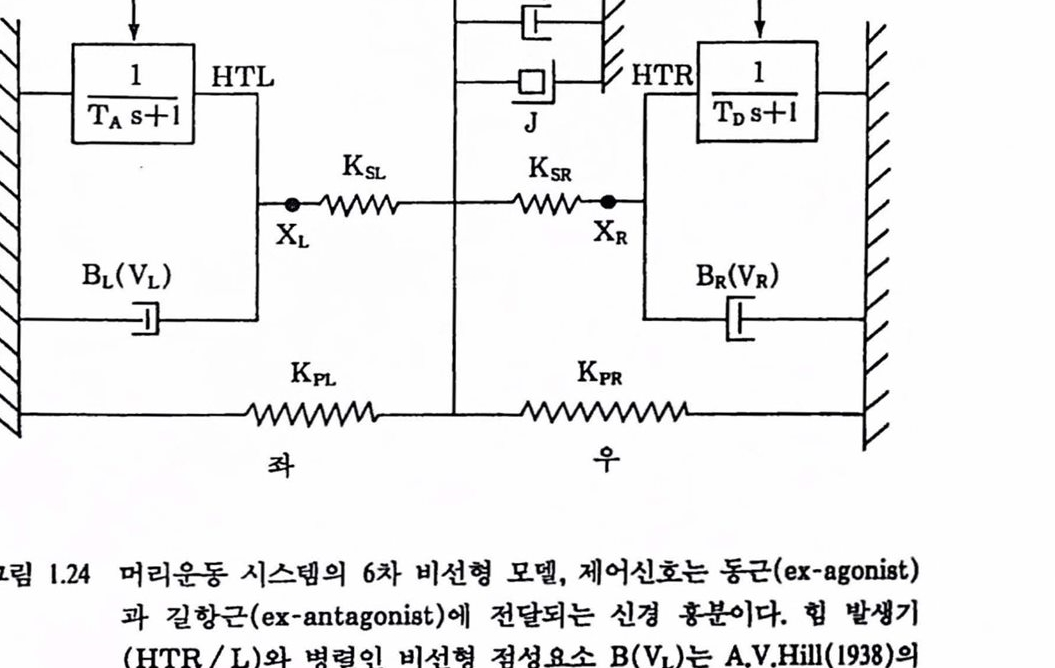
그렴 1.22는 펼자가 머리운동 제어기구를 연구하기 위해 1983년의 실험에서 사용한 표면 전극 위치와 이에 따른 EMG 왜턴을 나타낸 것이다. 전극위치 0-2에서 얻은 신호가 상호 신경지배에 의한 제어특성(A , B, C 펄스)을 잘 나타내었으며 전극의 위치에 따라 특성도 달라졌다. (Nam 등, 1989). 그렴 1.23은 EMG 교정 실험에서 얻은 EMG 형균과 혐의 관계를 나타낸 것이다. Zangemeister 등(1981)은 Stark의 상호 신경지배 모멜을 바탕으로 머리운동 시스댐의 근육과 플랜트를 6차 비선형 모댈(그렴 1.20 참조)로 만들어 시률레이션을 하였다. 목의 여러 근육의 확혼을 모델령하기 위해 2개의 등가근육 -1개의 통근(agonist)과 1개의 걸항근(antagonist)-을 썼다. 각개 등가근육에서 근육의 건(tendon)과 부착된 교차 브리지를 나타내는 직렬 탄성소자를 거쳐 부하에 힘을 가한다. 부하는 두개골의 철량과 그것의 내용물, 목의 구조률 지지하는 탄성 및 점성 등이며 2차(등가 J, Bp, Kp) 플랜트로 모멜랭하였다. 이 모멜의 입력 포는 제어신호는 각개의 퉁가근육이 받아들이는 천체 운동 뉴련의 발화 패턴(신경홍분)으로 하였다. 그렴 1.24는 그럽 1.20에서 대웅되는 부품을 기계적으로 표시하여 얻은 머리운동 시스댐의 6차 비씬형 모멜이다. 이 모멜은 1626년 Descartes가 제안한 것올 1961년 Stark가 다시 제안하고 Atwood 풍(1961)이 처음으로 2개의 근육의 모멜로 만들고 Cook와 Stark(1968), Clark와 Stark(1975)가 saccade안구운동모델로 발전시키고, Hung 등(1978),Bahill 등(1978), Lehman과 Stark(1979)를 거치는 동안 일반적인 안구운동에서 머리운동(Zangemeister 등, 1981) 모댈로 확장되었다.
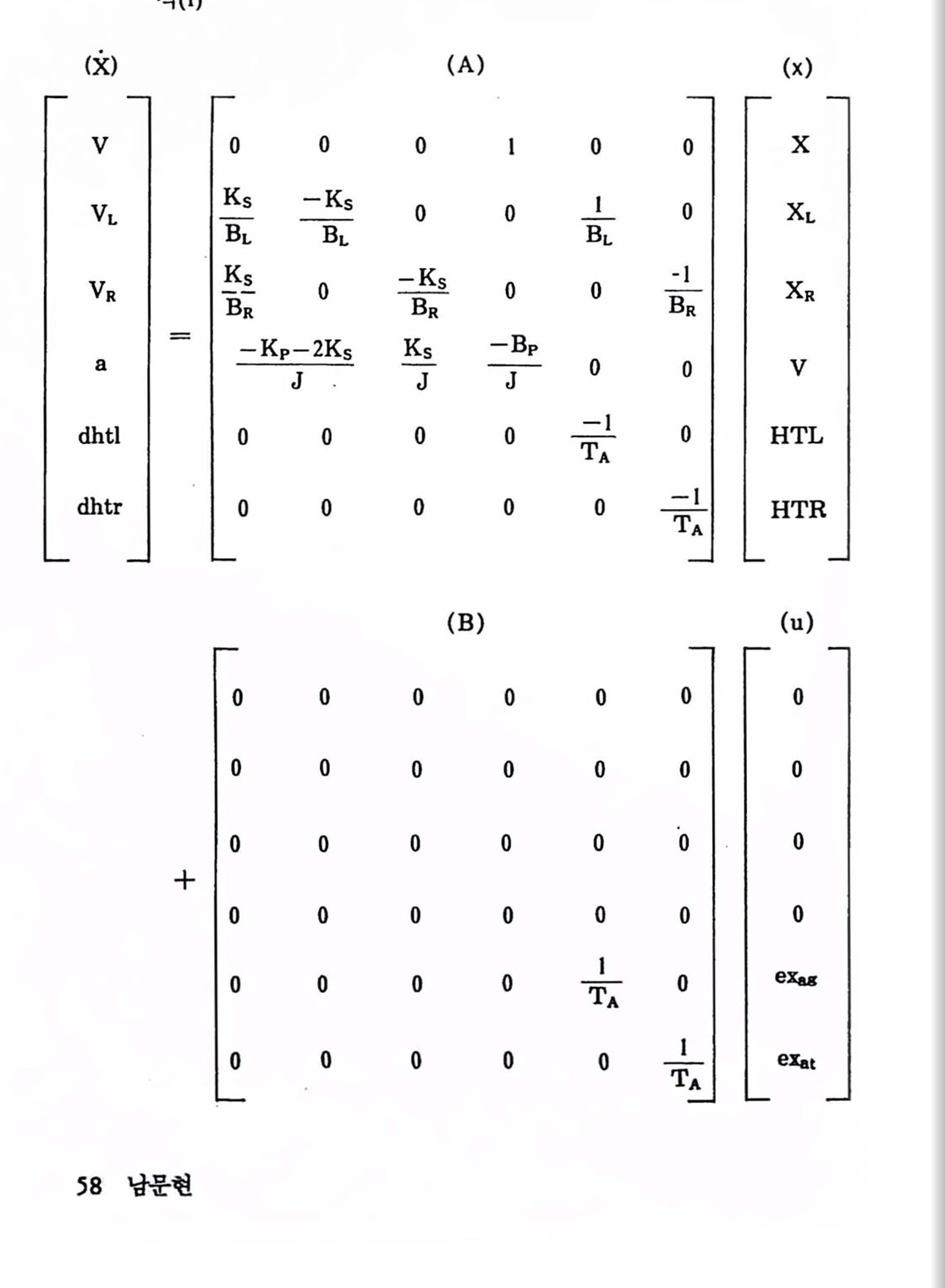
이 시스탱은 6개의 상태방정식과 2개의 보조방정식으로 표현하여 시률레이션할 수 있다. 그렴 1.24의 마다(X점)에서 힘을 합하면 표 1.1과 같은 6개의 대수 미분방정식이 얻어지며 FORTRAN으로도 구현할 수1000
EμMv G 1300 口•。 A’n4에 u 。nn uu • EMG μv EMG μv 0 0FO2RCE4 gr-f 6 •1 ()3그립 1.23 그렴 1.22의 2의 위치에서 얻은 EMG의 교정값. (a) EMG의 명균값(a)과 혐의 관계(b)는 포물곡션을 나타낸다(피컴자 : 필자).
있다(Lehman 과 Stark, 1979).이 식은 식 (1)과 같이 상태공간 모델 (x = Ax + Bu)로 나타낼 수 있다. 표 1.2에는 실험 및 시융레이션을 통해 얻은 매개변수 값을 나타내었다. 이 모텔은 빠른 운동과 느련 운동의 상호관계 연구로 발전되어 모델의 일반성과 유용성이 업종되었다 (Xinters, Nam, and Stark, 1984 ; Winters, 1985). 모델을 구통하는 제어신호의 펄스 크기와 폭을 추정하는 데는 최척화법(optimization)이 쓰여 왔으며 실험에서 얻은 데이터와 최대로 맞추어야 한다. 각종 제어신호를 가하여 모델의 출력(위치, 속도, 가속도 및 시간함수)을 얻는 것을 역 모델(inverse model)이라 하여 동특성의 함수로서 제어신호를 얻는 과정에서 꼭 필요한 기법의 하나이다.
 Kp
Kp
그림 1.24 머리운동 시스댐의 6차 비선형 모멜, 제어신호는 동근(ex-agonist)과 길항근(ex-antagonist)에 천달되는 신경 홈용이다. 행 빨쟁기(HTR/L)와 병렬인 버션형 정성요소 B(VL)는 A.V.Hill(1938)의 힘-속도 판계식이다.
 식 (1)
식 (1)
표 1.1 수형 머리운동에 대한 비선형 6차 모델 방정식
 상태방정식
상태방정식
표 1.2 모델의 매개변수 정의와 제안값
 이륨 기호값 단위
이륨 기호값 단위
그렴 1.22의 전극 위치 0-2에서 얻은 EMG 는 고전적언 3상 패턴(tri-phasic EMG pattern)을 나타낸다. 곧, 초기의 동근 발화, 길항근에서 억제 발화 및 최종 동근 발화의 순서로 동근-길항근이 상호 신경지배를 한다. 이와 칼은 3개의 EMG 펄스는 동근-길항근-동근에서 순서대로 PA(action pulse, 기동 펼스), PB(breaking pulse, 제동 펄스) 및 PC(clamping pulse, 정착 펄스)라 부른다. 이와 같은 패턴은 팔푹의 빼른 굴-신(flexion -extension)운동(Wacholder와 Altenburger, 1926 : Angel 1977 : Wadman등, 1980등)과 손가락의 구부렴(Meink 등, 1984), 팔목의 회전(Litventsev, 1977 : Sane화 Jennings, 1984), 고양이의 빠른 다려운동(Ghez와 Martin, 1982)에서도 발생한다. 그렴 3.15는 머리운동의 모델과 실험에서 얻은 3상 패턴(점선은 모댈)을 나타낸 것이다. Hannaford와 Stark(1987)는 최척 머리운동에 필요한 최종 동근활동 (PC)에 대해 최척제어 이론을 적용하여 제어신호가 끝난 후에도 사지(limb)가 목표지점에 머물러 있으려면 3개의 펄스 제어신호에 따라 최대로 빼른 운동어 발생된다는 것을 밝혀 PC의 존재를 업증하였다. 제어신호로서 EMG의 유용성은 외부의 부하에 대한 영향을 조사해 입증되었다. 점성부하률 부가했을 때 EMG의 변화로 제어전략의 변화를 알 수 있었으며 PC의 크기가 감소되거나 완전히 소멸되었다. 그림 1.26은 점성 부하시( )와 정상(N) 머리운동시의 통특성(P, 위치 : V, 속도 : A, 가속도)과 EMG(AG, 동군 ; AT, 길항근)의 변화를 나타낸 것이다.
4.2. 신장 반사의 제어 특성 그렴 1.18과 같은 신장반사(stretch reflex) 루프는 서보 시스댐으로서 기능올 수행향올 알았다. 이번에는 상호 신경지배에 대한 닫힌 루프 우동에서 시스댐올 구성하기 위해 그렴 1.18에 걸항근(이 경우에는 삼두박근, triceps)울 찾훈 활뚝관철의 각도제어를 생각해보기로 하자. 그렴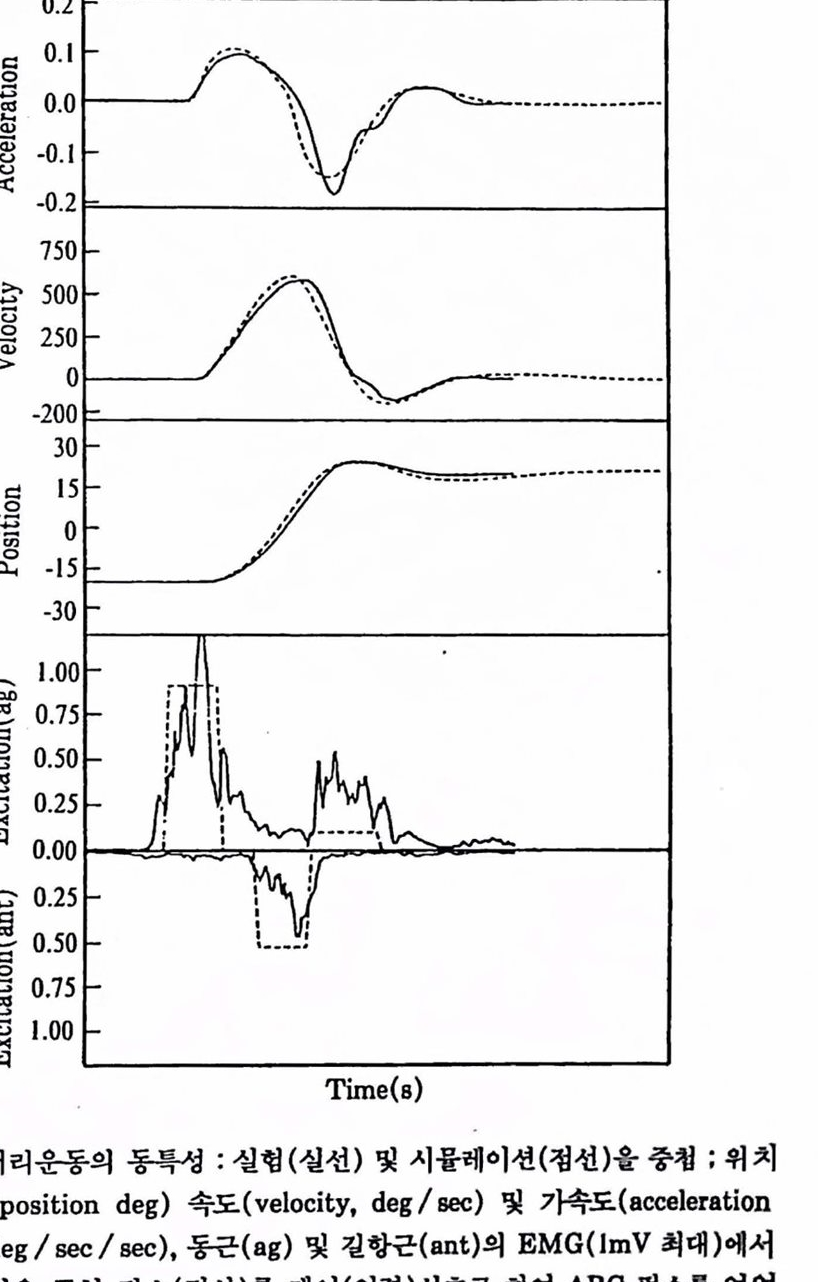 0.2
0.2
그림 1.25 머 리운동의 통특성 : 실험(실선) 및 시융레이전(청선)옳 충챙 ; 위치(position deg) 속도(velocity, deg/sec) 및 가속도(acceleration deg/sec/sec), 동근(ag) 및 킬향곤(ant)의 EMG(1mV 최대)애셔 얻은 구형 펄스(정선)폴 제어(업력)씬호로 하여 ABC 훨스훌 얻었다(Hannaford, 1986).
1.27은 그렴 3.18의 연장으로서 (a)는 해부학적 구조, (b)는 블록 선도이다. 그렴 1.27(a)에는 신경지배 경로를 나타내었으며 직접경로는 홍분정(E)이며 억제성(I) 경로는 길항근의 동일 근방추로부터 나온다. 이들 경로는 개제 뉴런(interneuron)을 포함한다. 그렴 1.28은 그림 1.27에 감마(gamma) 운동섬유의 역할을 추가한 것으로서 CNS의 고차 레멜은 두 가지 제어전략을 활용하여 출력위치를 제어함을 알 수 있다. 열린 루프 제어전략은 알파(alpha) 운동 뉴련을 활용하며 감마 섬유흘 거쳐서 근방추가 일정하도록 지배한다. 따라서 팔묵 각도 9를 감소시키려면
P 「|L / r
삐| /르늑 v5 OR ---e 、 -옹‘='' f~ A80%[~ 〈냥~ - 700 - ~NB AG EMG ° μv 0 [a] 500 ms [b] [c]그립 1.26 정성부하에 대한 머리운동과 EMG의 변화 효과. 정상(N)과 정성부 하( B)시의 20, 40, 60도 시표변위에 대한 웅탑의 비교
 명매바
명매바
 방추수용기
방추수용기
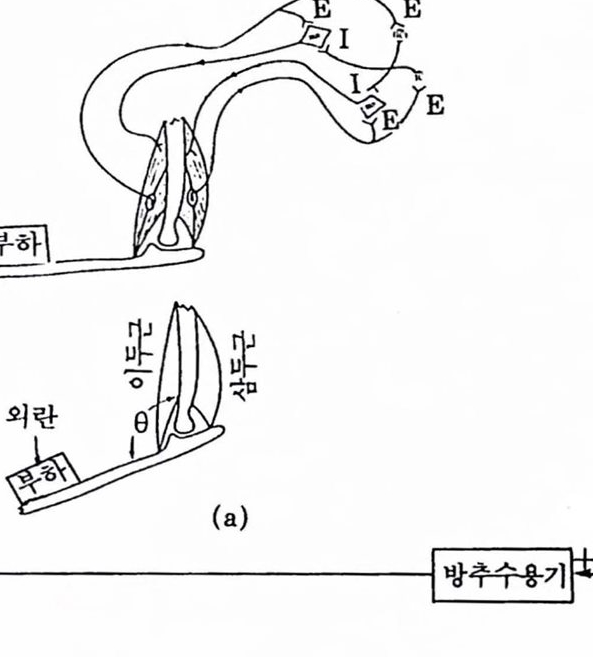
그렵 1.27 활푹 판철의 상호 신경제어 : (a) 해부학척 구후, (b) 블록 선도.(J. Houk와 E. Henneman, ‘Feedback Control of Muscle : Introductory Concepts’, in Medical Physiology, 13th ed. ed. V.B. Mountcastle. The C. V. Mosby Co. St. Louis, 1974).
이두문 운동 뉴련에 대한 CNS 지배를 늘리고 삼두근 운동 뉴련의 지배 는 줄인다. 이와 같은 전략은 매우 빨라 대형의 속응운동에 쓰인다(Stark 1968 : Houk, 1974).
두번째 전략은 닫힌 루프 제어로서 감마 섬유를 활용하는 것이다. 팔푹의 각도를 줄아려면 이두근의 감마 섬유의 지배를 증가시켜 이에 따라 상두근의 감마 섬유에 대한 지배를 줄여 주면 된다. 이제 이두곤만을 고려하면 이렇게 중가된 발화는 이두근 방추섬유로 하여금 수축이 일어나도록 한다. 따라서 감각섬유가 위치한 방추의 중앙지역이 신장을 일으킨다. 이에 따라 이두근 감각섬유는 발화융이 높아지고 이두근의 알파 운동 뉴련을 자극하여 수축이 진행된다. 이와 비슷한 변화가 일어Iy 생 유 i
l이두 근 |근 력 판켈각 8 lA픔 근| 근력 y 생유그립 1.28 감마 생유 업력올 푸가한 신청근육 제어 시스댐
나 삼두근은 이완한다. 감마 섬유제어는 아주 정교한 운동과 자세제어에 사용되지만 대부분의 운동은 알파와 감마 제어의 조합으로 이루어진다 (Houk, 1974). 이 밖에 알파와 감마 시스템의 역할에 대해 다른 이론들이 있는데 lnbar(1975)는 방추기관의 목적은 시스템으로 하여금 적웅제어를 수행하게 하여 동특성을 변화시킨다고 하였다.
근육의 길이를 측정하는 근육내의 변환기를 근방추 기관이라 한다. 그립 1.29는 근방추와 Golgi 건을 나타낸 것이다. 신경근육 제어 시스템은 또한 그립 1.29에서와 갈은 Golgi 건 기관(tendon organ)이라는 또 다른 감각기관을 갖고 있는데 근방추가 주근육 섬유와 평행으로 놓여 있는 데에 비해 건기관은 건과 근섬유 사이의 분지에 위치하여 수축요소와는 직렬로 되어 있다. 이 건기관의 출력은 개제 뉴런을 경유하여 알파 운동 뉴런에 되먹여전다. 이것들은 알파 운동 뉴런의 발화를 억제한다. 알파, 감마, Golgi 통로의 CNS 제어는 복잡하며 각각의 통로는 독립적으로 제어되는 것으로 생각된다. 더구나, 방추 수용기 및 건기관은 협력근(synergist), 길항근 및 다른 사지의 근육에 영향을 미치며 그것들의 출력은 뇌간, 소뇌 및 대뇌피질에 사용 된다. 지금까지 살펴본 바와 같이 운동의 신경제어를 이해하려면 간략화된 모델을 이용하는 것이 매우 도움이 된다는 것을 알 수 있다. 이제 가장 최신의 제어이론을 적용하여 보기로 하자. 근육의 적응제어에 대해 살펴보기 위해 모델 규법형(model reference) 제어 시스템의 구조를 먼저 간단히 고찰해 보자. 그립 1.30 A는 실제 제어대상 시스템 S1과 실제 시스템의 모델 S2에 제어신호 X1과 X2가 각각 입력되고, 실제 출력(y1)을 모델의 출력(Y2)에서 빼어 오차신호 (y2-Y1)가 발생되는 것을 나타내는 블록 선도이다. 이 오차신호는 그림 1.30의 A에서와 갇이 1, 2, 3으로 나누어져 활용된다. 모델 기준 비교에 대한 기본 아이디어는 수의 안구운동 중에 시계운동의 결핍을 설명한 Helmholtz에서 유래한 것으로 이런 원리가 여러 가지 공학설계에 이용되어 적응제어(adaptive control)라는 분야로 발전되었다. 그렴 1.30의 A에서 조건부 피드백 시스템(경로 1)에서 오차는 처리(증폭 등)되어 원래의 제어신호(X1)와 합해져( ) 새로운 제어신호가 만들어져 S1으로
~운근방동추신신경경 l 속섬차 유종 판 짧l輔/ 랩써
건기관으혹부터 척도영역 근방추 쳐 l 내 션조 건기판그림 1.29 근방추와 Golgi건 : (P. A. Merton,"How we control the contractionof our muscles", Sci. Am. vol 226. no. 5. pp. 30-70, 1972)
보내진다. 이런 경우에 제어대상은 모델과 같이 정밀하게 응답하는 조건 아래에서는 이상적인 거동을 나타내어 피드백을 생략하도록 설계된다.
외부로부터의 교란이 있어 이상적인 거동이 방해를 받으면 오차신호가 발생되어 증폭되고 오차를 감축시킬 수 있도록 제어대상에 가해진다. 오차감축의 효과는 피드백 경로의 이득에 달렸는데 경로에 많은 지연이 있거나 이득이 너무 높으면 불안정이 야기된다. 이런 셜계의 이점은 직접전향(feedforward)과 피드백 경로를 독럽적으로 설계할 수 있는데, 전자는 제어기의 동특성을 변화시키고, 후자는 피드백 경로의 이득 및 동특성을 변화시켜서 설계한다. 이 경우는 생체설계(진화)에 이점이 될 수도 있다. 처리된 오차신호를 경로 2를 거쳐 모델(S2)에 보내게 되면 부가적으로 이론적인 이점을 얻을 수 있다. 양(+)의 피드백 루프를 형성하여 적절히 동조하기만 하면 오차도 아주 감쇠시키면서 안정성을 높일 수 있다. 실제로 피드백 루프 이득을 1로 유지할 수만 있다연 오차는 0으로 감축되며 외부교란에 둔감한 (0-감도 시스댐〉을 얻을수있다. 세번째의 경우는 적응제어기(adaptive controller)를 포함하는 경우 (경로 3)로서 오차신호는 주 제어기(그렴 1.30 A의 C)나 모댈 S2의 성절을 수정하는 데 쓰인다. 그립 1.30의 B는 근방추 수용기(muscle spindle receptor)의 구조와 주근육(main muscle) 간의 관계를 나타낸 것으로 모델 규범형 제어의 가능성을 제시하는 좋은 본보기이다. 방추는 미세한 근섬유(intrafusal)로 구성되어 있으며(그렴 1.29 참조) 방추 구심 성유가 모여 있는 중심 감각영역을 갖는다. 이 영역은 극보다 수축활동이 척은데 미세근섬유가 활성화되면 감각영역이 신장되며 구섬발화가 생긴다. 미세근섬유(intrafusal muscle)와 근육의 신장(muscle stretch)의 짱대 응답성은 모멜 기준 계산에 대한 기초를 제공하여 미세근 성유가 제어대상의 모델을 나타내면 주근육과 부하는 제어대상이 된다. 제어대상에 업력되는 제어신호(그림 1.30 A의 X1)는 주곤육올 지배하는 크기가 큰 알파-운동 섬유로서 활통전위를 나르교, 모멜에 입력되는 제어신호(x2)는 미세근섬유를 지배하는 작은 감마-운동 섬유로 신호훌 나른다.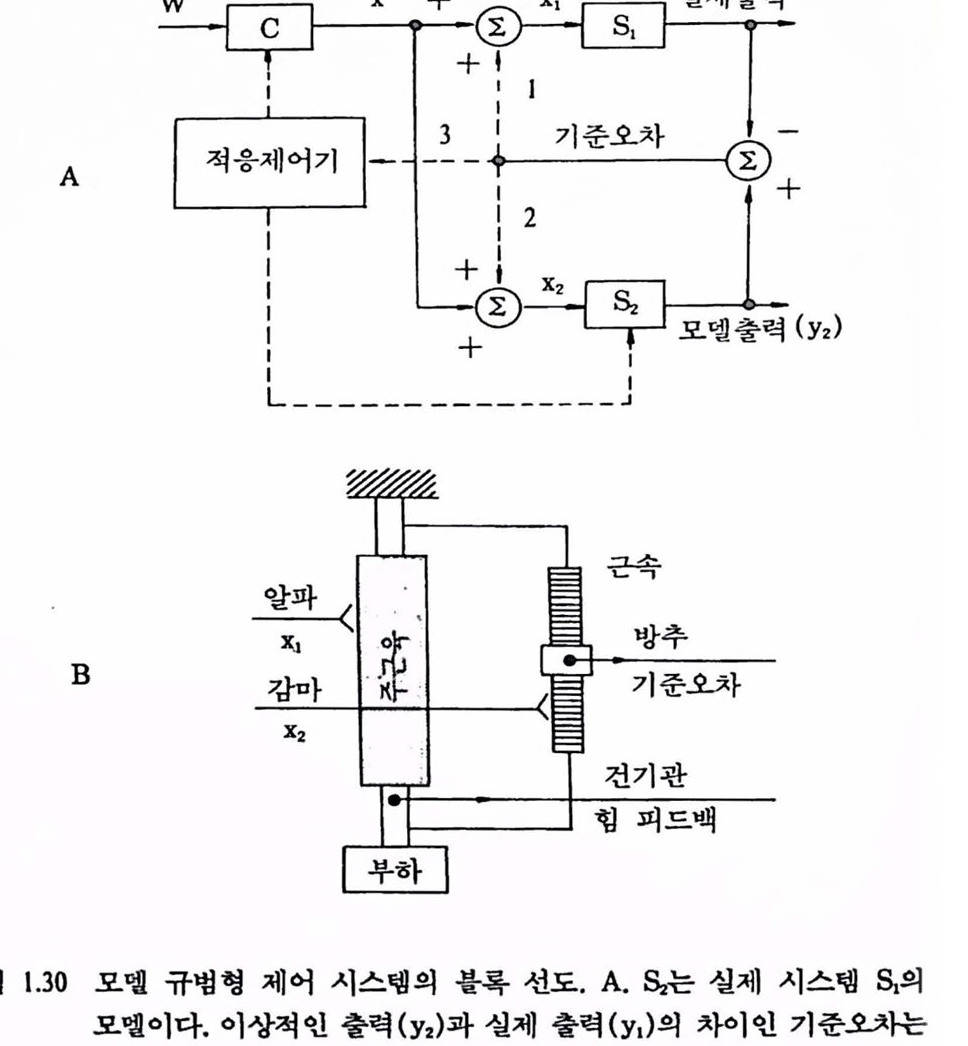 x ++ 장1 헥 81
x ++ 장1 헥 81
그럽 1.30 모멜 규뱅형 제어 시스댐의 폴록 선도. A. s2는 실제 시스댐 S1의 모멜이다. 이상척언 출력(y2)과 실제 출력(Y1)의 차이언 기준오차는 경로 1, 2, 3에 따라 선돼척으로 활용된다. B. 방추 수용기가 모멜 기준 감지기로서 쓰이는 것율 껄명한 불록 선도. 실제 시스댐 S1은 주근육과 푸하이고, 미세 근섬유는 모멜 시스댐 S2라 가갱하며 수용기는 주근육과 미세근의 단축 갱도의 차이륨 감지하도록 위치해 었다(J. C. Hauk, "Homeostasis and Control Principles" in V.B. Mountcastle ed. Medical Physiology, Vol. 1. 14th ed. The C.V. Mosby Co. St. Louis, 1980).
알파와 감마 운동 축색을 통해 보내진 신호는 운동을 발생시킨다. 이때 주구육의 단축과 미셰근섬유의 단축이 똑같으면 감각 영역에 가해지는 신장력은 변화되지 않으며 방추에서의 발화도 변하지 않는다. 이 경우는 앞서 말한 실제 출력과 모델 출력이 같아 기준오차가 O야 된다. 큰 부하 때문에 주근육의 단축이 방해를 받으면 기준오차는 방추구심 발화의 증가형태로 나타나게 됨을 유의해야 한다.
모델 규범형 제어 시스댐은 이와 같은 대비법을 더욱 발전시키면 방추 구성 발화는 피드백 경로를 완성하도록 투영되어야 한다(그렴 1.30 A의 경로 1). 총합반사는 부하가 무겁게 걸리도록 수축을 강화하게 되는데 Matthews(1972)는 이런 현상을 〈제어협조(servo assistance)>라 하였다. 그렇지만 영 조건(null condition, 피드백 소거, 실제 시스템이 모델과 같이 동작하는 경우)에 대한 특정 부하 조건을 조사한 사람은 아무도 없다. 아마도 감마-운동 뉴련(그렴 1.30 A의 경로 2)에서의 피드백 연결이 있을 것 같지만 가능성은 희박하다. 또 하나의 가능성은 방추 구심 발화가 두뇌(대뇌피질, 대뇌 기저핵, 뇌간, 척추 동)에 투영되어 기준오차가 적응 수정(그렴 1.30 A의 경로 3)에 이용될 수도 었다는 것이다. 지금까지 살펴본 운동제어 시스댐에서 매개변수 조절기구는 로봇 공학이나 제어공학을 연구하는 사렴들에게 매우 흥미있는 대상어라고 생각된다. 특히 대뇌피질에서의 계층제어는 목표지향 행위(goal-directed behavior) 를 모델링하는 신경 모멜(Albus, 1975)로 개발되어 로봇의 제어에 활용되고 있다. 4.3. 운동기능의 변환과 활용 앞 철에서 우리는 중추샌경계의 가장 중요한 기농의 하나가 자세와 운동의 제어에 있으며 운동제어 센터는 이러한 기능올 조칙(프로그햄)하며 구심성(감각) 정보의 흐름이 끊이지 않을 때에만 기능을 수행할 수 있다는 것을 알었다. 근육활동에 참여하는 구심 및 원심 기능의 총체로서 감각운동 시스템은 중추신경계와 정보적으로 접속되어 있다. 운동 제어 센터로부터 나오는 원심성 신호의 흐름은 원하는 근육의 운동으로 나타난다. 근육의 물리 및 전기적 활동은 피부 표면 또는 근육내에 삽입한 미세전극을 써서 근육 전기신호의 형태로 측정할 수 있으며, 이것을 처리하여 얻은 근천도는 운동에 영향을 미치는 각종 근육의 활동을 분석하는 데 중요한 도구가 되고 있음도 알았다. 최근에는 운동시에 동특성 변수(힘, 위치 등)와 함께 근전도를 측정, 분석해서 신경활동의 다중발화 패턴에 의해 운동이 제어되는 것이 밝혀져 운동제어 천략의 최척 모델을 얻을 수 있게 되었다. 수의운동(voluntary movement)시에 특정근육에서 발생되는 근전신호를 처리하여 얻은 근전도를 사람(장애자)의 손상된 운동기능을 대체(또는 재활)하거나 모의화(로봇 등)한 인공적인 장치를 제어하는 데 활용하는 연구가 국내외에서 활발히 진행되고 있다. 정상척인 팔다리는 자체의 감지능력과 외부 센서를 가지고 있으며 그것들의 조화운동은 가볍고 강력한 근육과 중추신경계의 지배에 의해 이루어진다. 이러한 팔다리의 운동은 복합제어 시스댐으로서 의학을 비롯한 생명 과학은 물론 스포츠 과학, 로봇학, 생체공학의 연구대상어 되어 왔다. 기능을 상실한 자연척인 팔다리를 인공의 팔다리로 대체한다는 것은 상상의 한계를 넘는 일이지만 현대의 과학기술은 이컷의 실현을 가능케 해주고 있으며 많은 연구자들이 좀더 새로운 제어방법(알고리즘)의 개발에 창여하고 많은 연구 결과가 발표되고 있다. 특히 절단된 팔다리의 기능을 대체할 수 있는 동력식 팔다리, 마버된 근육의 활성화를 위한 기능지극볍(functional neuromuscular stimulation, FNS), 손상된 통신 연계를 회복하는 방법의 한 가지인 인공신경, 극악 · 극한 상황 인간의 기능올 대신해 줄 훤격조작기(teleoperator) 퉁의 장치를 제어하는 제어 신호로 근전도률 처리하여 얻은 신호률 이용하는 것이 여러 분야에서 활발히 연구되고 있으며 일부 실용화된 것도 있다.
운동기능을 상실한 지체부자유자에게 의지대로 활동할 수 있는 보조장구를 갖춰 주어 정상인과 똑같은 생활을 영위하도록 하는 연구가 미국, 일본을 비롯한 선진국에서는 정부, 학계, 산업계의 연계로 활발히 이루어지고 있다. 우리나라도 공공복지의 차원에서 장애자의 복지문제를 적극적으로 생각해야 할 단계에 와 있다.
그렴 1.31은 첫 머리에서 셜명한 사람의 운동제어 기능과 근전도를 이용하여 인공 팔다리를 제어하거나 마비된 근육을 자극하는 방법을 도식적으혹 나타낸 것이다. 이 그렴은 언간(장애자)의 두뇌에서 발생되는 감각 및 운동명령 신호를 처리하여 얻은 근전도를 인공 팔다리 또는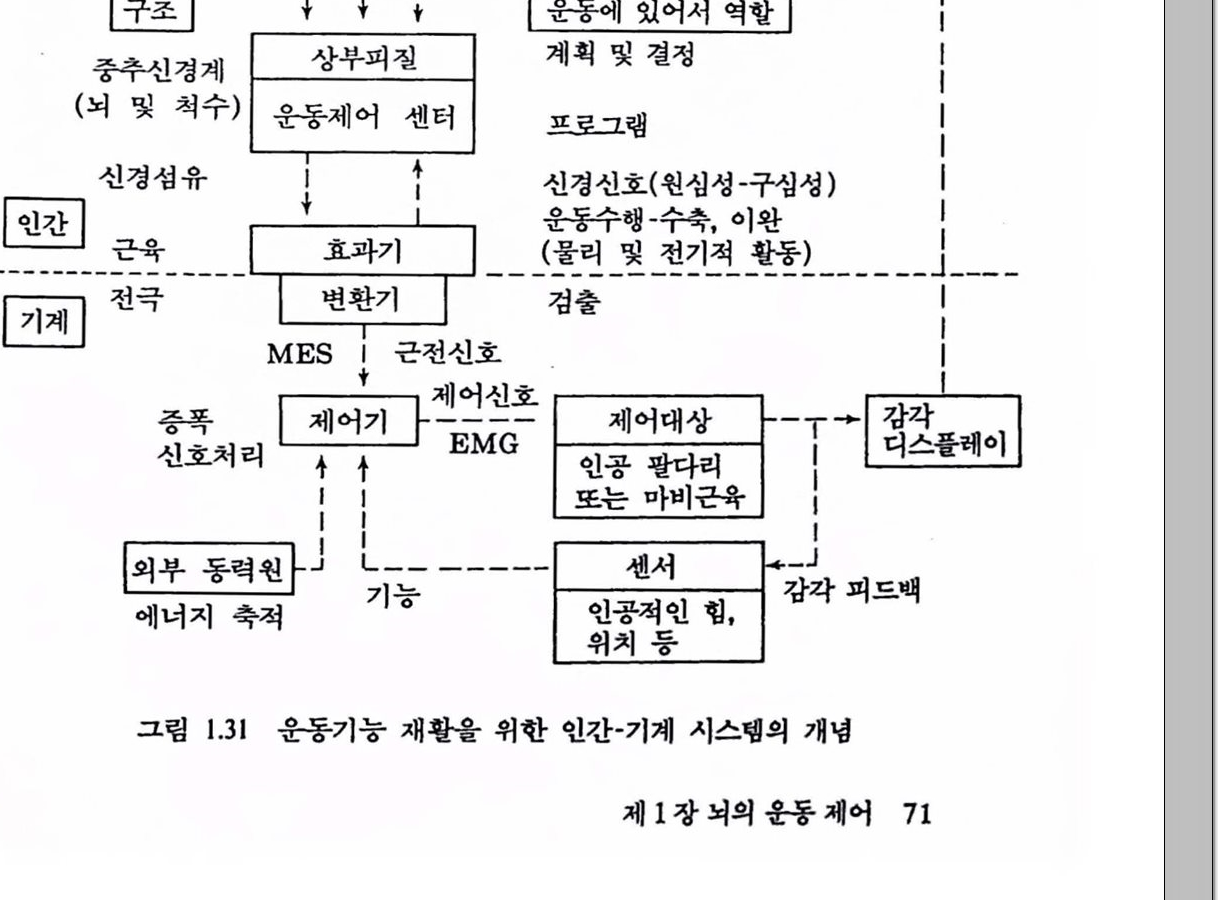 환경 및 감각 감각업력 자극(업력)
환경 및 감각 감각업력 자극(업력)
그링 1.31 운동기능 재활을 위한 인간-기계 시스템의 개념
마비근육을 운동시키거나 활성화하는 데 필요한 제어신호를 이용할 수 있는 인간-기계 접속 시스댐의 개념이다.
4.4. 운동기능의 대체 연구방법 장애를 극복하기 위해 사용되는 인공 보조장치는 착용과 사용이 간편하고 신뢰성이 높아야 하며 사용자가 발생한 운동행위를 완전하게 수행할 수 있도록 제어되는 것이 가장 바람칙하다. 현재까지 개발된 외부동력식 인공 팔 가운데 근천도 제어방법을 제외하고는 그리 성공적이지 못했는데 가장 큰 이유는 적절한 제어전략을 제공하지 못했기 때문이다. 적절한 제어전략을 얻으려면 정상척인 운동에 쓰이는 두뇌의 프로그랭 및 지령에 관계된 신경제어 기전에 관한 지식을 활용해야 한다. 이러한 지식을 공학척으로 응용하여 두뇌 자체의 〈컴퓨터〉로 결손된 운동기능을 대행하는 인공 장치를 제어하는 알고리즘을 개발해야 한다. 수의운동시에 발생되는 근전도를 처리하여 제어신호로 사용하고 여기에 시각이나 청각 등 감각 피드백을 부가한 제어전략을 보철장치의 접속 제어에 이용하고 보다 광범위한 복합 시스댐에 이 방법을 웅용하려면 신뢰성, 융통성, 반복성, 계산(웅답) 지연의 최소화, 최소의 표면 전극 수, 척옹성 둥을 고려한 근전도 신호처리 방법을 개발해야 한다. 근전도와 감각 피드백을 사용한 제어전략은 신뢰성, 안정성 및 가제어성을 특히 고려해야 한다. 앞으로 연구해야 할 제어방법(알고리즘)은 수의운동의 계충척 신경제어를 기본으혹 하는 것이 바람직하며 복합척인 인공장치의 제어와 마버된 근육에 운동기능을 부여하는 기능성 신경근육 자극에 활용하는 것도 한 가지 방법이다. 근천도를 보철용 장치의 실시간 제어에 이용하는 일은 운동제어 기전의 규명에 근전도릎 사용하는 일보다 훨씬 어렵다. 따라서 근천신호처리 기술의 개발이 보철제어 연구에 핵심이 된다. 근전도와 동특성 신호를 체어신호로 하며 다충 자유도와 제어기능올 갖는 인공 활의 피드백 제어방식을 개발하려면 아래의 범주를 셜정하는 것도 바람직하다. 1) 수의운동에 따른 패턴 언식 :특정한 운동을 지시하는 근전도 빛 위치신호의 수집, 처리 및 변환기술 연구. 2) 국부반사 및 감각 피드백을 이용한 인공 활의 상태제어 모델 :실시간 신호처리를 바탕으로 한 정상 팔 운동과 실험용 팔(모델)의 척응학습에 관한 컴퓨터 적응보조 연구를 통해 모델의 구조 셜청. 3) 로보틱스의 개념을 이용한 정멀제어 :자연스럽고 유연성을 갖는 상태이전이 가능한 제어 알고리즘 개발.
이를 위한 연구방법을 제안하면 다음과 같다. 1. 두뇌정보의 변환제어 : 두뇌의 수의 제어기능 활용-인간의 수의운동 제어기능을 식멸하여 다수의 전극위치에서 특정 근전신호 왜턴을 발생시킬 수 있는 능력을 활용한다 .n개의 전극신호원을 m개의 신경제어 신호 패턴으로 변환하면 n개의 제어근육 위치는 m개의 관절제어 신호로 변환되어 다관절 공동제어를 수행하도록 한다. 이를 위해 활의 적절한 위치(biceps 및 triceps 등)에 전극을 부착하고 3차훤 팔꿈치 위치 모니터를 착용한 채 시표안내에 따른 목표 지향운동에 대한 위치 신호와 동곤과 걸항근의 근전도를 측정하고 1) 신뢰성, 2) 융종성, 3) 반복성, 4) 계산지연의 최소화, 5) 전극 배치의 최적화 기준에 따라 처리하여 최척의 제어전략을 개발한다. 2. 두뇌정보의 보조제어 : 적응학습-컴퓨터가 발생시킨 근전도 왜먼올 수학척 모텔에 언가하여 모멜 기준 척웅제어를 하여 모멜의 매개변수률 추정한다. 실제의 팔(사람)과 모멜(인공 팔 또는 수학척 모멜)올 정속한 적응학습에 의한 팔 운동의 제어연구는 종래의 연구에서는 시도되지 않았던 새로운 방법이다. 이것은 두뇌의 계충척 신경제어 방식을 모멜랭한 신경회로망을 구성하여 시도하는 것도 하나의 방법이 훨 갯이다.팔 운동 모델은 Stark의 버선형 6차 모델(그립 3.24 참조)을 확대 척용하면 될 것이다.
3. 국부반사 및 감각 피드백을 이용한 인공 팔(모델)의 상태제어 : 팔(모멜)의 위치 출력이 팔의 감각 업력과 작동 환경에 자동적으로 웅답하여 척응 학습기능을 거치지 않고 직접 환경과 상호작용이 이루어지도록 하여 팔이 어떻게 운동해야 하는가를 결정하는 제어전략을 설계한다. 전자식으후 팔꿈치의 3차원 위치를 측정(electrogoniometer)하여 피드백 신호를 사용하는 방법(그림 1.32)을 써 보는 것도 하나의 방법이다.훌훌 l .-----팔-의- -위--치- ------ ---, 많뀔디
신-제 경어-輔離-+ 전-회 4 략로-· -망 -휩 2 컴정퓨멀터 제 척어 응학습 수의운동제어 컴퓨터 l 연구내용 l그립 1.32 팔운동 제어 시스템의 연구방법 개념도
(본 연구의 일부는 한국과학재단의 목적기초 연구지원에 의해 수행되었음을 밝히는 바입니다.)
참고문헌 1. M. A. Arbib, The metaphorical brain, Wiley-Interscience, New York, 1972. 2. W.B. Cannon, The wisdom of the body, Norton, New York, 1932. 3. B. G. Cragg, The density of synapses and neurones in the motor andvisual areas of the cerebral cortex. J. Anato, London, vol. 101, pp. 639-54, 1967. 4. B. Katz, Nerve, muscle and synapse, McGraw-Hill, New York, 1966. 5. A. R. Luria, The working brain : an introduction to neuropsychology(translated by B. Haigh). Penguin, Harmondworth, 1973. 6. P. B. Medawar, An unsolved problem of biology, An inaugural lecturedelivered at University College, London, 6 Dec. 1951. Lewis-London. 7. C. S. Sherrington, The integrative action of the nervous system, Scribner’s, New York, 1906. 8. J. Z. Young, Programs of the Brain, Oxford University Press, Oxford, 1978. 9. V. B. Mountcastle, Medical Physiology, vol. 1, 13th ed., Saint Louis, The C. V. Mosby Co., 1974. 10. A. L. Hodgkin and A. F. Huxley, “Quantitative description of membrane current and it's nerve", J. Physiol.(Lond) vol. 117, 500, 1952. 11. N. Wiener, Cybernetics, MIT Press, Cambridge, MA, 1961. 12. R. F. Schmidt, Fundamentals of Neurophysiology, 2nd ed. Springer Verlag, New York, 1978. 13. 臼井支朗,「細胞膜生理 神經回路」,《計沮Il&制御》 vol. 25, pp.113-118,1986. 14. 外山敬介,「生理工學 期待」,《計沮 制御》 vol. 25, pp. 149-150, 1986. 15. W. S. McCulloch, Embodiments of Mind, M.I.T. Press, Cambridge, MA., 1965. 16. J. Von Neumann, The Computer and the Brain, Yale University Press, New Haven, CT., 1959.
17. D. E. Wooldridge, The Machinery of the Brain, McGraw-Hill, New York, 1963. 18. R. E. Bellman, “Mathematical Models of the Mind., Math. Biosci, vol. 1, pp. 287-304, 1967. 19. J. Y. Lettvin. H. R. Matturana, W. S. McCulloch, and W. J. Pitts, “What the frog’s eye tells the frog’s brain", Proc. IRE. vol. 47, pp. 1940-1951, 1959. 20. J. S. Albus, A new approch to manipulator control : The Cerebellar Model Articulation Controller(CMAC). J. Dynamic Syst. Meas. Control. pp. 220-227, 1975. 21. F. Rosenblatt, Principles of Neurodynamics : Perceptions and the Theory of Brain Mechanisms, Spartan Books, Washington, 1961. 22. R. Hecht-Nielsen, “Neurocomputing : picking the human brain",IEEE Spectrum, vol. 25, No. 3, pp. 36-41, 1988. 23. A. Terry Bahill, Bioengineering, Prentice-Hall Inc., Englewood Cliffs, NJ., 1981. 24. B. Hannaford and L. Stark, “Late agonist activation burst(PC) required for optimal head movement : A simulation study", BioI.Cybern, vol. 57, pp. 321-330, 1987. 25. A. Terry Bahill and L. Stark, “The trajectories of saccadic eye movements", Sci.Am. vol. 240, No. 1. pp. 108-117, 1979. 26. J. Fincher, The Brain, Mystery of matter and mind, U.S .News Books. Washington, D. C., 1981. 27. G. Cook and L. Stark, “Derivation of a model for the human eye positioning mechanism", Bull. Math. Biophysics, vol 29, pp. 153-174, 1967. 28. M. Clark & L. Stark, “Time optimal behavior of human saccadic eye movement", IEEE Trans. Auto Contr. AC-20, pp. 345-348, 1975. 29. A. Terry Bahill, F. K. Hsu & L. Stark, “Glissadic overshoots are due to pulse width errors", Arch. Neural. vol 35, pp. 138-142, 1978. 30. M. H. Nam, V. Lakshminarayanan, & L. S. Stark, “Effect of external
viscous load on head movement", IEEE Trans, Biomed. Eng. vol. BME-31, pp. 303-309, 1984. 31. J. M. Winters, M. H. Nam, & L. Stark, “Modelling Dynamical interation between fast and slow movement : fast saccadic eye movement behavior in the presence of the slow VOR", Math. Biosci., vol. 68, pp. 159-187, 1984. 32. M. H. Nam. B. Hannaford & L. W. Stark, “Cross talk, Muscle synergy, and electrode locations and their effects on electromyographic activity in neck muscles", Innov. Tech. Biol. Med. vol. 10, No. 2, pp. 233-241, 1989. 33. W. H. Zangemeister, S. Lehman, & L. Stark, “Sensitivity analys is and optimization for a head movement model", Biol. Cybern. vol. 41, pp. 35-45, 1981. 34. W. J. Wadman, J. J. D. van der Gon & R. J. A. Derksan“Muscle activation patterns for fast goal directed arm movements", J. Human Movement Studies, vol. 6, pp. 19-37, 1980. 35. J. N. Sanes & V. A. Jennings , “Centrally programmed patterns of muscle activity in voluntary motor behavior in humans", Exp. Brain. Res. vol. 54, pp. 23-32, 1984. 36. S. Lehman & L. Stark, “Simulation of linear and nonlinear eye movement models : Sensitivity analysis and ennumeration studies of time optimal control", J. Cyber. Inf. Sci., vol 4, pp. 21-43, 1979. 37. R. E. Bellman, Adaptive control Processes : a guided tour, Princeton Univesity Press, Princeton, NJ., 1961. 38. P. B. C. Matthews, Mammalian muscle receptors and their central actions, The williams & Wilkins Co., 1972. 39. R. B. Stein, “The peripheral control of movement", Physiol. Rev. vol. 54, pp. 215-243, 1974. 40. W. S. Levine, “Some tools and reasons for studying the neural control of locomotion," IEEE Control Systems Mag. vol. 8, No. 1, p. 37. 1988. 41. D. S.Childress, "Historical aspects of powered limb prostheses", Clin.
Prosthet. Orthot. vol. 9. No. 1, pp. 2-3, Winter, 1985. 42. L. E. Carlson, “Position control of powered prosthesis", in Proc. 38th ACEMB, Chicago, IL, Sept. 1985, p. 48. 43. R. W. Mann, “Cybernetic Limb Proshesis", The ALZA Distinguished Lecture, Ann. Biomed, Eng. vol. 9, pp. 1-43, 1981. 44. T. B. Sheridan & R. W. Mann, “Design of control devices for people with severe motor impairment", Human Factors, vol. 20, No. 3, p.321-338. 45. J. H. Lyman, A. Freedy, R. Prior & M. Solomonov, “Studies toward a practical computer-aided arm prothesis system", Bull. Prosthet. Res., pp. 213-225, Fall 1974. 46. 남문현,「로보틱스와 생체공학」, <전자공학회지>10권6호, pp. 26-31, 1983. 47. S. Lee & G. N. Saridis, “The control of a prosthetic arm by EMG pattern recognition", IEEE Trans. Automat. Contr., vol. AC-29, pp.290-302, Apr. 1984. 48. D. Graupe & W. K. Cline, “Functional separation of EMG signals via ARMA identification methods for prothesis control purposes", IEEE Trans. Syst., Man, Cybern., vol. SMC-5, Mar. 1975. 49. G. N.Saridis and H. E. Stephanou, “A hierarchical approach to the control of a prosthetic arm", IEEE Trans. Syst., Man Cybern., vol. SMC- 7, pp. 407-420, June 1977. 50. P. C. Doerschuk, D. E. Gustafson & A. S. Willsky, “Upper extremity limb function discrimination using EMG signal analysis", IEEE Trans. Biomed. Eng., vol. BME-30, pp.18-28, Jan. 1983. 51. R. J. Triolo, D. H. Nash & G. D. Moskowitz, “The identification of time series models of lower extremity EMG for control of prostheses using Box-Jenkins criteria", IEEE Trans. Biomed. Eng. vol. 35, pp. 584-594, 1988. 52. L. Philipson, “Adaptable myoelectric prosthetic control with functional visual feedback using microprocessor techniques", Med. Biol. Eng. Comput., vol. 23, pp. 8-14, 1985.
53. G. Hefftner, W. Zuchini & G. G. Jaros, “The electromyogram(EMG) as a control signal for functional neuromuscular stimulation-Part I : Autoregressive modeling as a means of EMG signature discrimination", IEEE Trans. Biomed. Eng. vol. 35, pp. 230-237, 1988. 54. C. J. De Luca, "Physiology and mathematics of myoelectric signals", IEEE Trans. Biomed. Eng., vol. BME-26, pp. 313-325, 1979. 55. N. Hogan & R. W. Mann, “Myoelectric signal processing : Optimal estimation applied to electromyography- Part II : Experimental demonstration of optimal myoprocessor performance", IEEE Trans. Biomed. Eng., vol BME-27, pp. 396-410, 1980. 56. J. M. Winters & L. Stark, “Analysis of fundamental movement patterns through the use of in-depth antagonistic muscle models", IEEE Trans. Biomed. Eng. vol. 12, pp. 826-839, 1985. 57. A. V. Hill, “The heat of shortening and the dynamic constants of muscle", Proc RoySoc(London), 126B : 136-195, 1938. 58. R. I. Close, “Dynamic properties of mammalian skeletal muscle", Physiol Rev., 52 : 129-197, 1972. 59. D. R. Wilkie, “Relation between force and velocity in human muscles", J. Physiol, vol. 110 : 249-280, 1950. 60. B. Hannaford and L. Stark, “Roles of the elements of the triphasic control signal, Exp. Neurol, 90 : 619-634, 1985. 61. B. Hannaford, M. H. Nam, V. Lakshminarayanan & L. Stark, “Electromyographic(EMG) signals as controller signal with viscous load", J. Motor Behav., val. 16 : 255-274, 1984. 62. E. Y. Chao & B. F. Morray, “Three-dimensional rotation of the elbow", J. Biomechanics, vol. 11, pp. 57-73, 1978. 63. E. Y. Chao, K. N. An, L. A. Askew & B. F. Morrey, “Electrogoniometer for the measurement of human elbow joint rotation", Trans. ASME. J. Biomech. Eng. vol. 102, pp. 301-310, 1980. 64. J. F. Stein, “Role of the cerebellum in the visual guidance of movement", Nature, vol. 323. pp. 217-221, 1986.
65. R. B. Stein, “Novel uses of EMG to study normal and disordered motor control", Can. J. Neurosci, vol. 15, pp. 95-98, 1988. 66. R. Beneke, H. M. Meinck and B. Conrad, “Rapid goal-directed elbow flexion movements : limitations of the speed control system due to neural costraints", Exp. Brain Res., vol. 59, pp. 470-477, 1985. 67. R. A. Schmidt, D. E. Sherwood & C. B. Walter, “Rapid movements with reversals in direction. I. Control of movement time", Exp. Brain Res. vol. 69, pp. 344-354, 1988. 68. C. D . Marsden, J. A. Obeso & J. C. Rothwell, “The function of the antagonist muscle during fast limb movement in man", J. Physiol. London, vol. 335, pp. 1-13, 1983. 69. K. Wachholder & H. Altenburger, “Beitrage zur physiologie der willkurlichen Bewegung", pfluegers Arch Ges Physiol, vol. 214, 642-661, 1926. 70. M. Kawato, K. Furukawa & R. Suzuki, “A Hierachical Neural-Network Model for Control and Learning of Voluntary Movement", Biol. Cybern. vol. 57, pp. 169-185, 1987. 71. R. Granit, The basis of motor control, Academic Press, 1970. 72. L. Stark, “Neurological Control Systems", Studies in Bioengineering, Plenum Press, New York, 1968. 73. W. J. Heetderks & F. T. Hambrecht, “Applied neural control in the 1990s", Proc. IEEE. vol. 76, pp. 1115-1121, 1988.
이춘길 제 2 장 감각의 신경보철 1. 뇌의 기능적 설계 뇌 기능에 대한 근래의 집접된 이해에 기초하여 뇌 기능을 언위척언 방식으혹 제어하여 결손 기능을 보조하는 것이 가능하다. 이러한 시도는 신경보철(neural prosthesis)이라 불리며, 감각 및 운동의 장애를 보죠할 수 있는 가능성을 제시하는데, 이 장은 이러한 신경보철에 업각하여 시각신경보철의 가능성을 지지하는 이론척 배경을 칸략히 소개한 것이다. 이를 위해 뇌 기능에 대한 지극히 개괄척인 소개를 아래에 정리하였다.
1.1. 뇌의 기능과 전반적 설계 신경계의 총체척 기능을 이해하는 데 있어서 동풀이 운동하면셔 수행하는 과제틀을 생각해 보는 것이 유억하다. 통물의 감각기관온 통훌주위의 환경에 대한 정보를 받아들여 신경계가 처리할 수 있는 형태의 신호로 변환한다. 동물은 환경의 정보 가운데 생존에 필요한 범위만을 받아들이는데 이러한 정보를우리는 생물학적 정보라 부른다. 생물학적 정보는 동물에 따라 차이가 있다. 들쥐의 몸에서 발산되는 열(척외선)은 뱅울뱀에게는 환경에 관한 중요한 정보이며, 꽃에서 반사되는 자외선은 꿀벌의 중요한 관심거리이다. 그러나 자연 속의 인간은 적외선이나 자외선에 대체로 관섬을 두지 않았으며 이를 수용할 수 있는 감각기관도 찰 발달되어 있지 않다.
동물이 관심을 가지는 것에는 주위 환경에 대한 지식뿐 아니라 갈증, 통증 등 동물 자신의 상태에 대한 것도 포함된다. 환경에 대한 지식과 동물 자신의 내부 상태에 대한 지식은 동물의 감각정보를 구성한다. 신경계는 감각기관이 제공하는 여러 정보들을 통합, 처리하여 운동의 명령을 생성하는데 이 과정에는 동물의 내부에서 오는 다른 정보들, 과거의 저장된 기억(현재의 상황에 관련된 학습된 평가), 그리고 현재의 동기, 정서 등이 포함된다. 신경계에 의해 생성되어 전달된 운동의 명령은 최종척으로 근육세포의 수축으로 번역되어 운동을 일으키게 된다. 이러한 과제들 즉, 감각정보의 업력 및 처리, 상황에 적합한 운동명령의 생성 등은 성공척인 생존을 위해 수행되는 생물학적 정보의 표상 및 처리 그 자체이며 바로 신경계의 고유한 기능이기도 하다. 신경계는 동물의 감각계와 운동계를 그 사이에서 연계하는 구조이다. 신체의 다른 기판과 마찬가지로 신경계는 세포로 이루어져 었으며 신경세포를 뉴련이라 부른다. 애초에 생물학척 정보를 다루는 뉴련플은 몸의 각 부분에 흩어져(히드라에서 보듯이) 감각-운동의 연계를 수행했으나 뉴련들이 몸의 한 곳으로 모여 신경계를 형성하기 시작하고 나아가 머리를 구성하면서(소위 encephalization) 뇌가 진화하게 된다. 뇌는 신청계의 기능척언 위계에서 정점에 위치하여 이콧에서 감각정보에 대한 고도의 처리가 이루어지며 정보의 표상, 저장이 이루어지고 운동의 명령이 시발된다. 뇌가 수행하는 기능들, 예컨대 감각, 기억, 사고 동의 다양한 기능들은 뇌 전체가 총체적으로 활동하여 이루어지는 것이 아니고 대체로 각기 다른 부위에서 이루어지며 이러한 것을 기능의 국소화(localization)라 부른다. 뇌의 감각영역은 감각에 대해 활동하는 뉴런들로 이루어져 었으며, 뇌의 운동영역은 운동이 이루어지는 시간을 천후하여 활동하는 뉴런들로 구성되어 있다.
뇌의 감각영역 내에서도 시각, 청각, 신체감각 등 감각의 형태에 따라 각기 다른 부위의 뉴련들이 활동하고 있다. 그뿐만 아니라, 한 감각 형태 내에서도 감각의 내용에 따라 활동하는 뉴련이 다르다. 예를 들어, 밤하늘의 한 지점에서 반짝이는 별을 웅시할 때, 그 멸의 상은 망악의 외점(fovea, 망막의 중심)에 맺혀 와점 부위의 뉴련을 활성화시키고 외점 부위의 뉴런과 연결된 시각 중추신경계 뉴런들어 곧어어 활성화 된다. 이때 활성화되는 뉴런의 활동은 시공간의 중심 즉 우리가 웅시히는 공간의 지점에서 일어나는 빛의 변회를 감각하는 기초이다. 웅시하고 있는 별의 주위에서 반짝이는 별은 광학의 굴절 법칙에 따라 와점 주위의 일정한 위치에 상이 맺혀 그 부위의 망막 뉴런들을 활성화시키고 이와 연결된 시각 중추신경계의 뉴련을 활성화시킨다. 시각신경계의 세포들운 공간의 다론 지점에 있는 별의 반확임에 대해 반웅하고 있으며 실로 이 뉴런들의 활동은 곧 공간의 특정한 지정에 위치한 반확임의 존재, 즉 반짝엄의 내용과 그 공간척 위치에 판한 생물학척언 청보를 나타내는 것이다. 각기 다른 부위의 뉴런들이 서로 다른 기능에 판련되어 있는 것은 뇌 기능이 국소화되어 있음을 나타낸다. 머리 부분을 구성하게 되는 뇌의 진화와 더불어 관찰되는 또 한 욕청은 양측화(bilateralization)이다. 이것은 대청척언 신체의 발달에 수반하여 대칭척 뇌가 진화히는 특칭을 일컬는다. 거시척 수준에서 양측화는 뇌 기능의 국소화이며 화우측 뇌가 반대현 신체에서 일어나는 감각 및 우도으 담당케 하는 셜계이다. 예를 들어 웅시하는 공칸의 한첨융 기준하여 오른쪽 반의 공간은 좌측 시각중추에 의해 감각되며, 또한 오른쪽 신체의 근육은 화측 운동신경계에 의해 구성된다.신경계가 매개하는 개별 심적 기능혹은 행동이 단일 뉴련의 수준에까 지 국소화되어 있는 예를 하등동물에서 볼 수 있다. 귀푸라미의 복부 신경절의 한 뉴련이 활동하면 곤충은 그 뉴련이 위치한 반대편으로 몸을 돌련다. 그 뉴런을 전기척으후 자극해도 동일한 결과를 관찰할 수 있다. 이 뉴련의 활동을 유발하는 자극은 초음파이며 이 뉴련이 매개하는 얀응은 박쥐가 발산하는 초음파를 탐지하여 박쥐로부터 회피하는 반웅인 듯이 보인다. 이 경우 초음파 정보와 한 방향으로의 회피 반응 사이의 연계의 어느 단계에는 한 뉴련이 매개하고 있는 듯하여 단일 뉴런에 의한 전문척 기능의 분담의 예를 여기서 볼 수 있다.
고등척추동물의 경우, 뉴련은 일정 법위의 행동 지표내의 모든 행동에 관여하고 따라서 개별 행동 혹은 섬척 기능은 많은 수의 뉴런 집단에 의해 매개된다. 고양이나 원숭이의 시각중추에는 공간의 한 지점에 나타난 수직 방향의 막대에 반웅하는 셰포가 있으며 이 세포는 수칙에서 다소 기울어진 방향의 막대에도 반응한다. 각 세포가 일정한 범위의 방향을 유지하는 자극에 대해 반웅하는바, 각 방향에대해 반응하는 뉴련은 하나가 아닌 집단이다. 즉 행동파 의식적 경험을 매개하는 뉴련의 활동이 기능을 표상하는 방식은 집단으로서의 국소화이다. 개개의 뉴련들이 모여 기능척언 회로를 이루고, 하나 혹은 그 이상의 기능척언 회로들이 모여 특정행동의 기초률 이루는 듯이 보인다. 뉴련 혹은 뉴런 집단이 상호작용하여 개별 행동올 결정하는 과정에 대한 엄격한 이해는 대체로 드물지만, 인구운동의 경우 벡터 형균의 계산 과정이 뉴런들에 의해 수행되고 있음을 시사하는 실험척 결과를 필자와 동료들이 최근 보교한 바 있다(Lee et als., 1988.) 1.2. 뉴런 뉴련은 다른 동물세포와 마찬가지로 핵, 세포철, 미토콘드리아 동으로이루어지고 세포막으로 싸여 있으며 단백철의 합성, 대사 등 세포가 수행하는 기능을 수행한다. 뉴련이 다른 동물세포와 구별되는 가장 뚜렷한 특정은 정보를 처리한다는 점이며 뉴런의 형태는 이 특정을 반영한다. 세포체에 연결된 수상돌기들은 정보를 수집하는 데 척합하게 분화된 장치이며, 세포체는 수집된 정보를 통합하며 축색은 통합된 정보를 전달하는 케이블 역할을 한다. 뉴런이 다루는 정보는 감각기관을 통해 업력된 환경에 대한 정보, 운동의 내용에 대한 정보, 다른 세포가 하고 있는 일에 대한 정보 등이다. 하냐의 정보는 대개의 경우 뉴련 집단에 분포되어 있으며 뉴런 집단의 전기적 활동 패턴으로 이루어져 있다.
뉴런의 세포막 전위는 뉴런에 따라 차이가 있으나 대체로 -70mV 이다. 즉 세포막을 경계하여 세포막 내부의 전위는 외부에 비하여 음극을 띠며 세포막 전위의 기원은 전하를 띤 이온의 분포가 세포막을 경계로 하여 다르기 때문이다. 예를 들면 양전하를 띤 나트륨 이온은 세포막 외부의 농도가 내부에 비해 상대척으로 높으며 음전하를 띤 단백질은 내부의 농도가 훨씬 높다. 다양한 이온들의 차별적 분포는 세포막을 가로질러 이온 농도의 차이를 형성하여 세포막을 통한 이온의 이동을 부추기지만 뉴런이 버활동척일 때는 세포막에 있는 이온의 통로를 통해 야온이 통과할 수 있는 투과성이 낮으며 포한 세포막에 의해 농도의 차이가 적극척으로 유지되어 세포막 전위는 대체로 음극이다. 여러 가지 방식에 의해 세포막이 자극되면 닫혀져 있던 세포악의 이온 통로가 열리게 되고 세포막을 중심한 전기 · 화학척 차이는 이온의 이동을 순식간에 얼어나게 한다. 세포막이 이렇게 흥분했을 때의 세포막 천위의 순간척인 변화를 활동전위라 부르며 세포막의 전위가 순간척으로 양극으로 변했다 가 다시 음극을 회복하는 것을 말한다. 세포막의 흥분과세포막 천위의 회복 과정은 대개 1,OO0분의 1초 동안 지속된다. 활동전위의 횟수는 뉴련의 활동 수준을 나타내며 뉴련의 최고 활동 수준은 대개 초당 1,000회이다. 세포막의 흥분, 즉 이온 통로들의 순간척언 개폐는 인껍 세포막으로 전달되어 인접한 세포막의 이온 통로의 개폐에 영향옳 미치게 되고,따라서 그 부위의 세포막 전위가 변화한다. 인접한 부위에의 영향은 연속적으로 계속되어 활동전위가 세포막을 따라 전도되는 기반이 되며 뉴런이 담당하는 정보의 전달이 가능하게 된다.
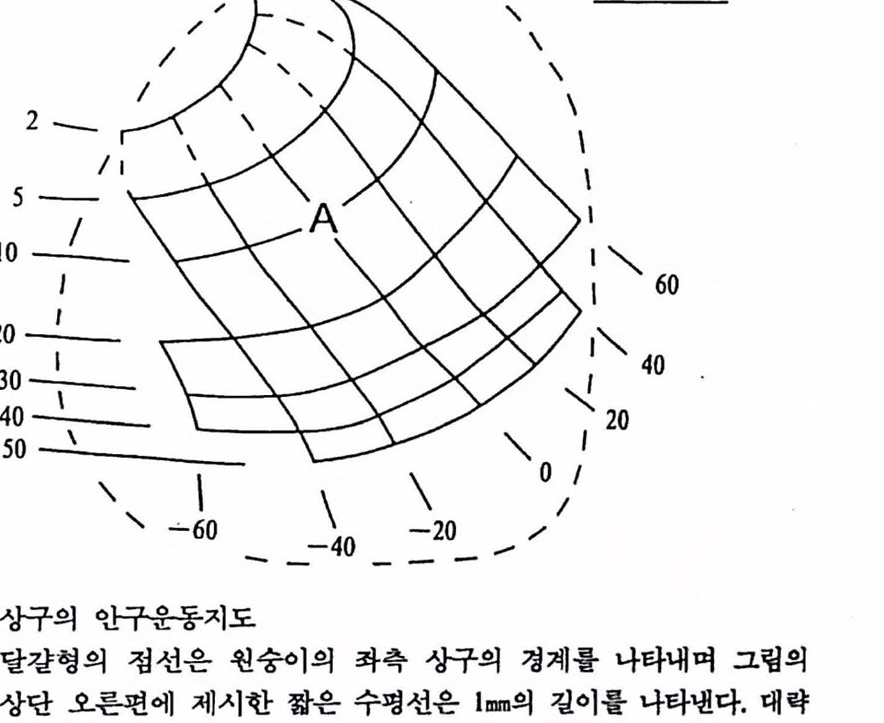
한 뉴련에 수렴하는 기능척 회로의 업력이 뉴런의 세포막 전위를 역치에 도달하게 하면 뉴련은 자발적으로 활동전위를 생성한다. 버활동척인 뉴런을 언위적으로 활성화하는 방법은 세포막 내부에 양전하를 주사하여 세포막 전위를 역치에 이르케 하거나, 세포막을 전기적드로 지극하여 뉴련을 홍분시키거나 이온 통로의 개폐 확률을 수정하는 화학적 처치들이다. 전기 화학적 에너지가 아니더라도 일정 방식의 차장이나 빛은 뉴련을 활성화한다(Yamamoto et als., 1988, Wade et als., 1988). 이상에 언급한 뇌 기능에 대한 간략한 소개에 덧붙여 본 장의 내용과 관련한 뇌의 기능척 설계의 원칙을 강조하기 위해 원숭이의 안구운동을 다음에 예시하였다. 1.3 뇌의 기능적 설계의 예---안구운동 눔의 움칙엄을 제어하는 신경 기착에 대한 관심은 생물심리학뿐 아니라 신경과학, 전산 및 정보파학, 제어공학 둥 여러 분야의 공통된 관심이 되어 왔으나 근년에야 비로소 각성 상태의 동물에서 안구운동과 뉴련의 활동올 동시에 측정할 수 있는 세련된 기법이 개발되어 안구운동 자체의 현상뿔 아니라 이를 제어하는 기작에 대한 이해가 급진전되고 있다. 안구운동의 여러 형태 가운데 여기서 예시하고자 하는 것은 빠른 안구 운동(saccadic eye movement)이다. 이 운동은 와정(fovea, 망막의 중섬후)을 가진 동물의 경우 시공간의 변두리에 나타난 시지극을 재빨리 와정에 위치시켜 물체률 자세히 볼 수 있게 한다. 이 운동을 위해 시각 신경계는 물체의 망막에서의 위치를 파악하고 운동신경계는 전달된 안구 의 현재의 위치와 물체의 망막에서의 위치를 종합하여 물체가 있는 공간의 위치로 시선을 이동한다. 중뇌에 위치한 상구(superior colliculus) 는 빠른 안구운동 응 생성하는 뇌 구조이다. 두 눈이 우측으후 빨리 움칙얼 때 좌측 상구에 있는 뉴런들의 활동전위의 횟수는 초당 1,000번에 이르는
 ' I t\‘ -‘、 \
' I t\‘ -‘、 \
그립 2.1 상구의 안구운동지도 달갈형의 점선은 현숭이의 확측 장구의 경계률 나타내며 그럽의 상단 오른편에 제시한 짧은 수명선은 1mm의 길이률 나타낸다. 대략 5mm 길이의 상구내에 안구를 반대편으로(확측 싱구의 경우에는 두 안구를 우측으로) 움직이게 하는 세포들이 위치하는데 확흑 상구의 각 지점에 위치한 뉴련들의 활동운 두 눈융 우측으로 얼갱한 방향(direction)을 따라 일정한 폭(amplitude)만큼 움직이게 한다. 그럽에 보이는 일련의 곡선들은 활성화된 뉴련들이 상구에셔 차지하는 위치에 따라 결정되는 안구운동의 폭과 방향용 표시하는 씬률이다. 그럽의 왼편에 나열된 2부터 50까지의 수는 안구용동의 풍폭씬(isoamplitude curve)올 나타내며 오른연과 아래에 나열된 60부터 -60까지 수는 안구운동의 동방향선(isodirection curve)훌 나타낸다.
정도로 맹렬히 활통하며 약한 전류로 화측 상구를 자극하면 두 눈은 재빨리 우측으로 움직인다. 이와 같이 하여 우측 성구의 뉴련이 활동하면 두 눈은 동시에 화측으로 재빨리 움직이게 된다.
두 눈이 움직이는 크기와 운동방향을 결정하는 것은 성구의 각기 다른 뉴련들이다. Robinson(1970)은 원숭이의 상구를 전기자극하여 유발된 안구운등응 측정하여 상구의 각기 다른 위치에서 활동하는 뉴런들이 안구운동의 방향과 폭을 표상하는 지도를 작성하였는데 그렴 2.1은 그 결과를 옮긴 것이다. 이 안구운동의 지도가 의미하는 바는 예컨대 등방향선 0도와 등폭선 10도의 교차 지점에 위치한 에 있는 뉴련들은 두 눈을 오른쪽 수명방향으로 10도만큼의 폭으로 재빨리 움직이게 한다. 상구 뉴련의 활동은 빼른 안구운동이 시작하기 약 10분의 1초 이전부터 증가하기 시작하며 안구운동이 종료되기 직전에 비활동척 수준으로 회복된다(Sparks and Mays, 1980). 자연적 방식이건, 전기자극 혹은 Bicuculline과 같은 화학물질의 주사 등과 같은 인위척 방식이건 싱구 뉴련의 활동을 유발하기만 하면 활성화된 뉴련의 상구에서의 위치에 따라 (그립 2.1에서 보듯이) 일정한 방향과 폭을 가진 안구운동이 결과 한다. 따라서 상구 뉴런은 안구운동의 지표에 관한 정보에 관여하고 있으며 성구 뉴련의 활동은 그 정보의 표현 그 자체이다. 상구에 관한 보다 상세한 소개는 Sparks(1986, 1990)와 여기에 소개된 여러 개괄에서 얻을 수 있다. 2. 감각기능의 신경보철 2.1. 신경보철 뉴련이 활성화펌으로써 뉴련이 판여하는 청보률 표현하는 것은 감각신경계의 경우 감각의 의식적 경험이며 운동신경계의 경우 운동의 의지언 듯이 보인다. 신경보철의 원리는 인위척으후 뉴련의 활동을 유발함으후써 뉴련이 표성하는 생물학적 정보, 즉 감각의 경험 혹은 운동의 기능을 유발하고자 하는 것이다.
2.2. 감각의 신경보철 앞에서 언급하였듯이 동물의 감각기관은 외부의 물리척 · 화학척 에너지의 변화를 신경계가 처리할 수 있는 형태의 전기적 신호로 바꾸는 과정을 포함한다. 시각의 경우, 망막의 감광세포가 시공간의 특정한 위치의 빛 에너지의 변화를 탐지하여 이를 전기척 신호로 바꾸어 시각신경계로 전달하면 시각계를 구성하는 신경 세포(뉴련)들은 전달된 시공간의 빛의 분포에 관한 신호에 기초하여 일련의 신호처리 활동을 보이며 이러한 활동의 결과로 시각의 현상이 가능해진다. 청각은 소리를 구성하는 기계척 에너지가 컷속의 달팽이관에서 소리의 강도와 주파수에 따른 전기척 신호로 변환되어 청각신호를 처리하는 신경계로 전달됩으로써 가능해진다. 에너지 변환을 수행하는 감각기관이나 신호처리를 담당하는 감각신경계의 손상과 같이 정상척언 경로를 통한 정상척언 방식의 감각기능이 손상되었을 때, 감각신경계의 구조와 기능의 이해에 기초하여 인위척으로 신경계의 활동을 제어함으로써 손상된 감각기능올 보조하고자 하는 시도가 최근 활발하다. 이러한 시도를 감각신경보철이라 하며 다음과 같은 과제를 포함한다. 1) 감각신경계의 작동 원리에 대한 이해 및 오형화. 2) 뉴련의 작용을 모방한 감각 모률의 개발, 웅용. 3) 신경보철에 기초한 결손 감각기농의 보조 빛 회욕.2.3 .감각신경보철의 예---전기달팽이관 전기탈팽이판의 시술로 이미 감각신경보철은 청각장애의 보조에 부분척인 성공을 거두고 있다. 달팽이관은 소리라는 기계적 에너지가 청각신경의 전기척 활동으로 변환되는 곳이다. 인간의 경우 약 3cm 남짓한 관이 두 바퀴 반 정도 감겨진 구조를 이루고 있다. 소리에 의한 고막의 진동은 일련의 이소골을 거쳐 증폭되고(진동의 폭이 증가한다는 의미라기보다는 고악의 진동이 액체의 진동으로 충실히 전달된다는 의미에서의 증폭), 뼈의 움직임은 달팽이관을 채우는 액체의 진동을 일으키게 되는데 이 진동은 최종적으로 달팽이관 내부 특정한 지점에 위치한 유모세포(hair cell)의 세포막 전위를 변화시킨다. 유모세포의 세포막 천위의 변화는 근접한 청각신경의 활동드로 이어진다. 소리의 고저를 구멸할 수 있는 부분적언 이유는, 소리의 고저에 따라 달팽이관의 각기 다른 위치의 유모세포 집단이 활성화되어 각기 다른 청각신경이 뇌로 신호를 전달하기 때문이다. 유모세포의 결손 등 달팽이관의 역기능은 청각장애를 결과하며 기존의 청력 보조장치를 통해 혜택을 받을 수 없다〔유모세포에 의한 소리 에너지의 초기 변환 과정에 관한 보다 상세한 기술은 Hudspeth(1989)와 이에 언용된 여타 개팔에서 얻을 수 있다〕.
천기달팽이관은 손상된 달팽이관에 단일 혹은 다중 전극을 삽입하여 외부에서 일정한 방식으로 처리된 신호에 따라 달팽이관에 와 있는 청각 신경(온천한 경우)을 칙접 자극함으로써 달팽이관의 에너지 변환 과정을 우회하여 청각울 어느 정도 회복하는 장치로서 상업척으로 제작된 전기 달팽이관이 최근 우리나라에서도 시술되기 시작하였다(검종선 외, 1989). 시각의 경우는 신경보철에 의한 감각보조가 상대척요로 용이하지 않다. 그 주된 이유는 청각의 경우 소리 에너지는 대체로 1차원척인 달팽이판의 여러 위치에 분포되어 에너지 변환이 일어나는 데 비해 시각의 경우는 빛 에너지가 2차원척인 망막에 분포되어 변환됨으로써 쓸만한 해상력율 갖추려면 제어해야 할 공간척 위치의 수가 기하급수척으혹증가하게 된다. 뿐만 아니라 많은 청각장애의 경우 달팽이관내의 유모세포의 결손이 중요한 원인으로서 청각신경의 역행척 퇴화가 얼어나지 않는 한 청각신경은 온전하여 신경보철을 위한 인위적장치의 접근을 그나마 용이하게 한다. 시각의 경우 상황은 다소 다르다. 3. 시각신경보철 3.1. 시각보조의 개념 안경이나 콘택트 렌즈로 호전될 수 없는 완전 맹언을포함한 시각 장애자의 수는 전세계적으로 엄청난 수에 달하고 1,000만 명 이상에 이르는 국가도 있다. 장애의 주된 이유는 노화에 따른 망막병리, 백내장, 녹내장, 당뇨병과 관련된 망막 쇠퇴, 시신경 쇠약 등으로서 대부분의 경우 시각중추신경계의 장애이기보다는 안구나 망악의 손상에 기언한다(미국 National Eye Institute, 1983-1987 보고서 ;“Clinical trials supported by the national Eye Institute", 1988). 안구나 망막의 손상은 빛 에너지 변환과 초기 시각 정보처리를 수행하는 망막의 신경세포의 퇴화를 수반한다. 특히 망막의 처리 결과를 뇌로 전달하는 신경철세포의 세포체는 안구 속에 위치하여 안구나 망막의 손상시 손상에 노출되고 따라서 퇴화가 상대척으혹 용이한 점이 청각의 경우, 제1 차 청각신경의 세포체가 달팽이관 바깥에 위치하여 뢰화에 다소 저항척이라는 정과 다르다.
신경세포는 일단 손상되면 회복이 불가능하다. 따라서 손상된 감각신경계에 따른 감각 장애의 극복은 온전한 다른 감각에 의존하는 천홍척언 방식에 의해 이루어져 왔다. 시각장애의 경우, 대개의 경우 촉각울 이용하여 왔는데 맹언용 첨자, 맹인 언도견 퉁이 이에 속한다. 인도견온 개의시각을 이용히는데 이 경우 개는 맹인의 결손된 센서(눈)의 역할을 수행할 뿐 아니라 거리의 표지 및 방해물의 정보를 처리하여 독립적으로 보행하고 맹인은 실제 인도견의 보행의 정보를 촉각을 통헤 업력받는 것이다.
3.2. 전통적 시각보조의 최근의 성과 인도견의 훈련은 많은 시간과 노력을 필요로 하고 새로운 환경에 인도 견을 적응시키는 부차적인 훈련 또한 요구되는바, 인도견을 로봇으로 대체하는 시도가 등장하였다. 이 가운데 진보된 형태의 하나는 일본 통산성 산하 기계공학연구소의 인도견 로봇일 것이다(그림 2.2). 이 개발 계획은 1977년부터 시작된 계획으로서 그 근본적인 셜계는 언도견을 A 로봇의 맹인 언도실험
A 로봇의 맹인 언도실험
 r------- - ---- - ------ ------ ----1
r------- - ---- - ------ ------ ----1
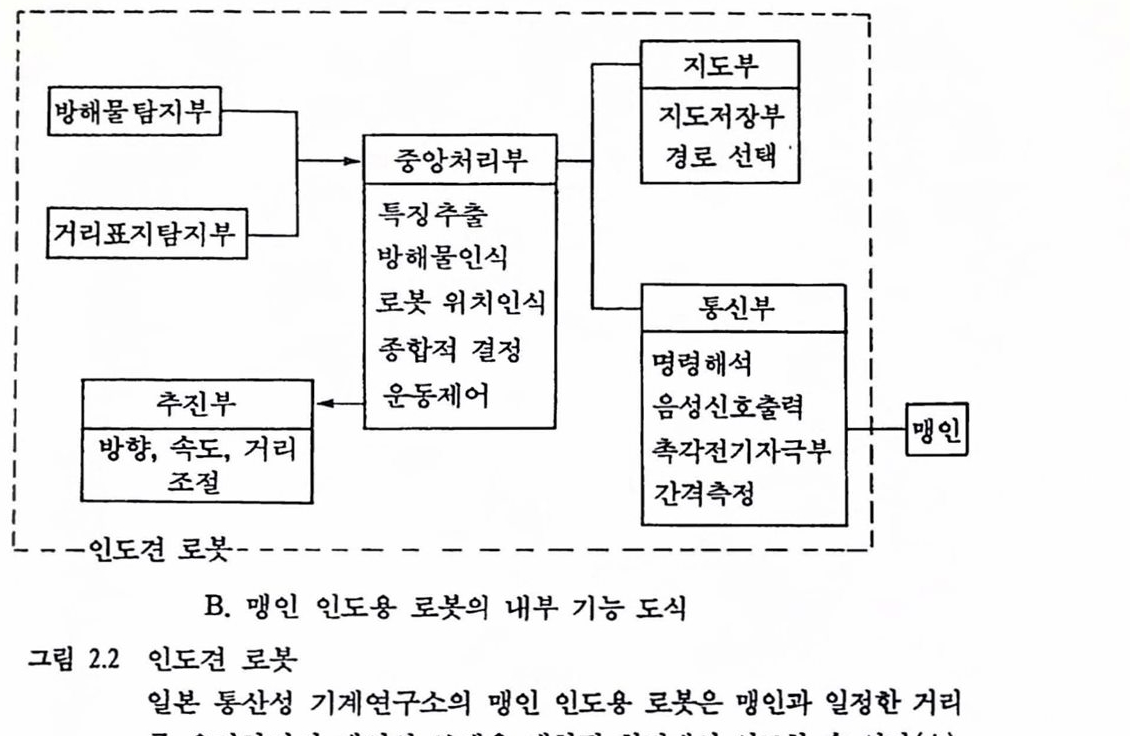
그림 2.2 언도견 로봇 일본 통산성 기계연구소의 맹인 인도용 로봇은 맹인과 일정한 거리를 유지하면서 맹인의 보행을 제한된 환경에서 인도할 수 있다 (A). 그렴의 다음 부분(B)에는 이 로봇의 내부 구성이 기능별로 제시되어 있다. 정선의 사각형이 그렴 A에서 보인 로봇의 내부흩 표시한다. 로봇은 초음파 및 광학척 센서를 이용하여 방해물과 거리의 표지블 탐지하여 중앙처리장치에 보내고 중앙처리장치는 저창된 거리의 지도를 읽어 내어 보행 방향올 결청한다. 로붓은 초융파 센서률 흥해 맹인과의 거리를 측갱하고 그에 따라 로봇의 이풍 속도륨 초철한다. 방해물 둥 주의를 요하는 경우 로붓은 맹인에게 음갱의 강도둥융 조철하여 방향 풍의 갱보률 전달할 수 있다. 예률 들어, 오른활에 접촉한 전극을 통한 지극이 100pps(pulse per second)언 경우 이 신호는 안천하게 보행하려면 오른쪽으로 이통해야 한다는 풋이고 10 pps 의 신호는 보행 방향율 수청하기 위혜 시계 반대 방향으혹 몸을 회전시켜야 한다는 신호이다. 척철히 약속된 신호에 척용함으로써 맹언은 보행의 보죠률 받율 수 있다[Tachi et als (1985)에서 그렴만 옮김].
로봇드로 대체하고 이 로봇의 출력을 맹인의 팔에 촉각의 형태로 제공하는 것이다. 이 로봇은 센서의 기능뿐 아니라 운동의 기능도 가지고 시각 환경의 지도를 기억장치에 저장하고 있어 필요한 거리의 지도를 기억장치에서 인출하여 보행 목적에 사용할 수 있다(Tachi et als., 1985).
3.3. 신경보철에 기초한 시각보조 촉각이나 청각 등 가용한 감각을 통한 시각정보의 업력에 기초한 이러한 개념의(일본 통산성의 로봇과 같은) 시각보조는 앞으후 센서의 기술과 집척회로 제작 기법 등의 발달로 보다 많은 거리의 지도가 기억장치에 탑재되고 새로운 환경에서의 적응도 어느 정도 가능하게 될 것으로 예측되며 무엇보다도 로봇의 소형화는 무리 없이 진행될 것으로 생각된다. 그러나 이 셜계의 근본척인 문제는 천자/통신 기술의 폭발척인 발달과는 상판 없는 로붓-인간의 접속 방식에 있다. 기술의 발달은 로봇의 개선에 필요한 고도의 시각해상력, 풍부한 기억용량, 실시간 왜턴인식 등의 기능을 인도견 로봇에 제공할 수는 있을 것이나 로봇의 출력을 신통찮은 해상력을 가진 인간 촉각에 접속하는 한, 센서 및 통신기술의 발달은 궁극척언 시각보조에 결정척 요인으로 작용하지 않으며 이러한 소극척 개념의 시각보조에서는 장애자는 결코 〈볼〉 수 없다. 신경보철에 기초한 척극척 시각 보조장치의 기본척인 개념은 안구의 기능을 대체하는 센서(카메라)의 출력 즉, 시간에 따른 2차원의 휘도분포(luminance distribution)를 일정한 방식으후 변환한 후, 변환된 분포에 입각하여 시각중추의 전략척언 위치의 뉴련 집단을 언위척언 방식으로 직접 활성화함으혹써 안구를 우회하여 시각척 경험올 회복하고자 하는 것이다. 기존의 시각 보조창치가 빈약한 해상력올 가진 촉각에 의존하는 폐 반해 이러한 개념의 보초장치는 시각 해상력이 월퉁한 온천한 시각신경계의 활동올 언위척으로 유발함으로써 보조 가능한 시각행동의 범위가 비교할 수 없을 만큼 확장된다. 이러한 신경보철의 척용이 시각장애의 역사와 시각계의 현재 상태에 대한 엄격한 진단파 명가척 도구를 필요로 함은 두말할 휠요가 없다. 아래는, 신경보철의 개념에 입각한 시각 보조장치의 실현 가능성을 지지하는 신경과학척 연구 성과와 현재의 연구 전략, 그리고 이의 실현을 위해서 해결되어야 할 숙제들을 간략히 정리한 것이다.
4. 시각신경계 4.1. 시각신경계의 구성 시각신경계의 구조와 기능에 대한 이해는 주로 원숭이와 고양이의 실험을 통해 이루어진 것이지만, 특히 원숭이와 인간의 시각신경계의 기능척 구조는 중요한 점에 있어서 통일한바(예를 들어 Burkhalter & Bernardo, 1989), 원숭이 시각신경계의 운용원칙은 인간에게도 그대로 적용된다. 신경계는 영상 즉 시공간의 빛의 분포를 분할하여 개개 뉴련의 활동 꽤턴으로 나타낸다. 이때 각 뉴련은 영상 가운데, 학습을 홍해 형성된 뉴련을 포함하는 기능척 회로에 의해 걸들여진 특정만울 영상으로부터 추출하는바, 따라서 활성화된 천체 뉴런들의 활동 때턴은 주어진 영상의 변형된 추상어다. 망막(retina)-외슬체(lateral geniculate body)-시각피질(visual cortex)로 이루어지는 시각신경계의 각 단계의 뉴련툴은 시공간의 복잡한 빛의 분포를 길들여진 특징에 따라 훈할하여 환갱의 부분들응 처리하고 궁극척흐로 이 부분척인 신호들이 일청한 방식(아직도 우리가 완전히 이해하지 못하는)으로 풍합됨으로써 환경에 대한 의식척 경험 즉 시각어 가능케 된다. 1950년대 이후 시각신경계의 기능을 이해하고자 하는 시도의 대부분은 각 단계의 신경세포들의 활풍 패턴을결정하는 시자극의 속성을 연구함으로써 이루어져 왔다. 이 접근은 현재에도 여러 실험실에서 성공적으로 계속되고 있는데 그 강점은 각 단계의 뉴런들이 분할하여 내는, 길들여진 특정을 밝힐 수 있으며 시공간의 환경을 신경계가 표성하고 처리하는 방식을 어떤 기법보다도 더 직접적으로 확인할 수 있다는 것이다. 다음은 시각신경보철에 관련한 시각신경계의 기능척 구조의 핵심적인 내용을 정리한 것이다.
망악의 감광세포에 의해 일차척으로 분할된 영상은 다음 단계의 망막세포들에 의해 처리되어 최종주자인 망막절세포(retinal ganglion cell)에 의해 외슬체 뉴련들에 연결된다. 외슬체는 망막에서 전달된 영상의 정보들이 시각피질에 도달하기 전에 중개되는 시상(thalamus)의 한 구조이다. 외슬체의 기능은 그동안 집중적인 해부학적, 생리학척 연구에도 불구하고 확렵되지 않았다. 망박철세포와 외슬체 뉴런 사이의 연결은 엄격한 질서에 따라 이루어지는데 망막의 특정 부위에 위치한 망막절세포는 광학의 굴절 볍칙에 따라 공간의 특정 지점의 빛의 분포에 관여하고 이 분포에 대한 정보를 외슬체의 특정 위치의 세포에 전달하고 있다. 또한 외슬체의 근접한 위치는 근접한 망막에 위치한 망막절세포와 연결되어 있다. 이러한 연결의 륙칭을 지웅성(topography)이라 불러왔는데 이는 시각신경계뿐만 아니라 청각 및 신체 감각을 포함한 감각신경계 구성의 일반척 특갱의 하나이다. 망박철세포와 외슬체 사이의 이러한 질서 정연한 지웅척 연결의 결과로 외슬체는 망막의 표면, 즉 2차훨으로 투영된 시공간울 철서 정연하게 표상하고 있다. 외솔체와 시각피철 사이의 연결도 또한 지웅척으로 이루어져 있는바, 이러한 결과로 외슬체의 각기 다른 위치에 존재하는 개개의 세포, 혹은 시각피질의 각기 다른 푸위는 시공간의 각기 다른 위치의 빛의 분포에 관여하고 있다. 다시 말해서, 시공간은 2차원척언 망막의 표변에 분포한 세포들에 의해 표상되고 지용쩍 연결의 결과로, 망막의 표면은 시각피질의 표면에 표상되어 었다. 시각피철의 지용척 구조는 시각신경보철의 중요한 기초언데 이에대해서는 아례에 상솔하였다.
4.2. 시각피질의 구조와 기능 시각대뇌피질은 후두부에 위치하며, 원숭이(Rheusus)의 경우 시각피질은 피질 표면 전체의 반 이상을 차지하고 있다. 언간의 경우 연합중추 와 전두엽의 확장으로 기초 감각피질의 상대척 면적은 감소를 보인다. 원숭이는 여러 가지 면에서 동물실험을 통해 시각신경보철의 원리를 확립하는 데 적합한 동물이다. 신경보철에 가장 적합한 제1차 시각피철은 거의 굴곡을 보이지 않아 평형하여 피질에 표상된 시공간의 어느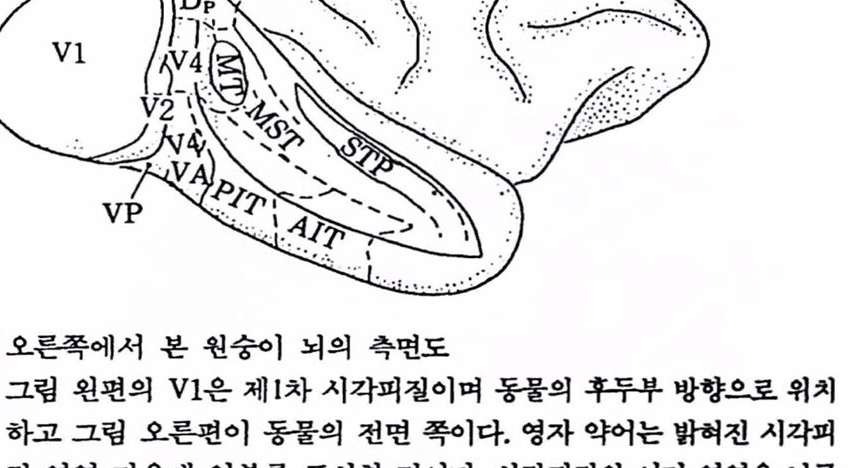
그립 2.3 오른쪽에서 본 원숭이 뇌의 측면도 그립 왼편의 V1은 제1차 시각피철이며 동물의 후두부 방향으로 위치하고 그렴 오른편이 동물의 전연 쪽이다. 영자 약어는 밝혀진 시각피질 영역 가운데 일부률 표시한 것이다. 시각피철의 여러 영역윤 뇌구(sulcus) 속에 묻혀 있는데 그중, 중측부 영역(MT, middle temporal area), 내상축부 영역(MST, medial superior temporal area), 상측푸 중다캄각 영역(STP, superior temporal polysensory area)들을 노출하기 위해 뇌의 얼부훌 철제한 모습이다[Maunsell & Newsome (1987)에서 그렴만 옮검].
지정도 접근이 용이하며, 시각피철의 기능적 구조는 가장 잘 알려져 있고, 신경보철의 원리를 확렵하는 데 필요한 행동적 훈련에 원숭이는 우수하다.
위에서 언급한 바와 같이, 시각 대뇌피질은 피질의 표면에 시공간의 위치를 2차원적으로 표상하는데 시공간의 표상어 여러 부위에서 반복적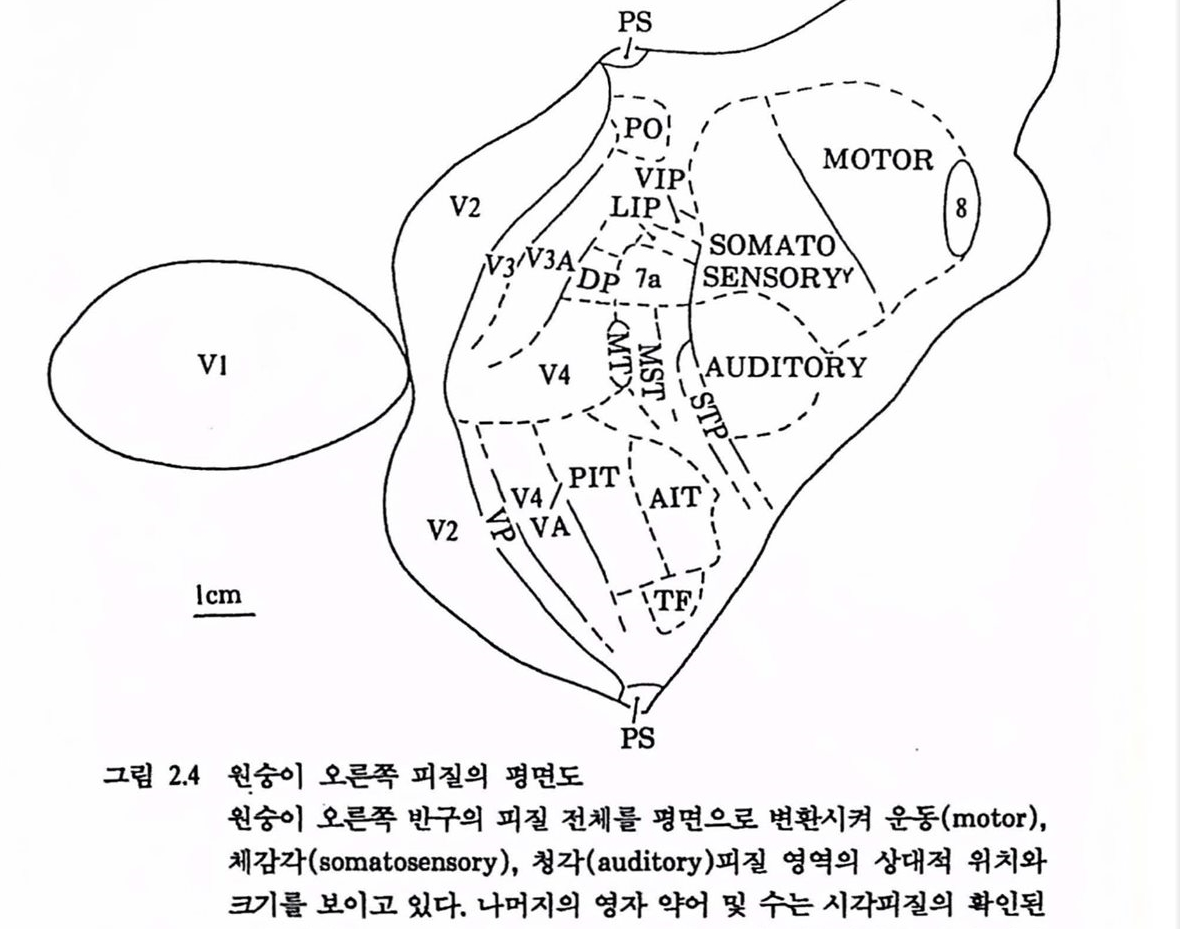 PS
PS
그림 2.4 훤숭이 오른쪽 피철의 명연도 훨숭이 오른쪽 반구의 피철 전체률 명연으로 변환시켜 운동(motor), 체감각(somatosensory), 청각(auditory)피질 영역의 상대척 위치와 크기률 보이고 있다. 나머지의 영자 약어 및 수는 시각피질의 확인된 19개 영역율 표시한 것으로서 보는 바와 갈이 시각피철은 전체 피철표면의 철반 이상올 차지하고 있다[Maunsell & Newsome(1987)에서 그렴만 옳킴]
으로 이루어지는 것이 특정이다. 원숭이와 고양이의 경우 적어도 10번 이상 이러한 표상이 이루어져 있다. Brodmann(1909)은 대뇌피절을 이루고 있는 층의 두께와 신경세포의 형태에 따라 피질영역을 분할하여 구멸하였는데 외슬체로부터 대량의 정보를 받는 제 1차 시각피질은 그의 부분에 따르면 제17영역에 해당하고, 시공간의 표상이 한번 이루어져 었다(Hubel & Wiesel, 1965). 그림 2.3은 원숭이의 뇌를 오른쪽 측면에서 본 것이며, 굴곡을 이룬 피질을 일정한 방식으로 절단하여 명명하게 만들어 피질의 각 영역을 표시한 것이 그림 2.4이다.
원숭이의 시각피질을 구성하고 있는 10여 개의 영역플은 제 1차 시각피질에서부터 시작하여 해부학적인 위계를 이루고 있다(Hubel & Wiesel, 1962, 1963 : Van Essen & Maunsell, 1983). 해부학적인 위계를 셜정하는 기준은 6개의 층으로 이루어져 있는 시각피질의 미세구조와, 피칠 영역간의 해부학적인 연결의 미세 구조를 관찰함으로써 이루어지는데, 한 기준을 예로 들면, 하위 영역에서 상위 영역으로 신호를 보내는 세포의 세포체는 피철의 2, 3층에 위치하고 축색의 끝(axon terminals)이 분포하는 콧은 주로 상위 영역의 3, 4층이다. 그에 반해 상위 영역에서 하위 영역으로 신호를 보내는 세포의 세포체는 상위 영역의 2, 3, 5, 6충에 위치하고 축색종말은 하위 영역의 2, 5, 6충에 주로 분포한다(Van Essen & Maunsell, 1983). 이러한 해부학적인 연결의 특갱에 기초하여 Van Essen과 동료들윤 시각 피철의 각 영역의 연결을 제1차 시각 피절에서 시직하는 위계 체계려서 도식화하였다(그림 2.5). 제1차 시각피질(VI)은 시각피철의 여러 영역 기운데 가장 넓은 연척을 차지하면서 시공간 혹은 망막의 표면을 엄격한 공간척 지도에 의해 표상하고 있다. Tootell과 동료들(1982)은 제1차 시각피질이 시공간율 표상하는 방식을 해부학척언 실험을 통해 시각화하였다. 그럽 2.6은 Tootell과 동료들이 얻은 실험 결과를 옮긴 것이다. 활동하는 신경세포는 필요한 에너지를 위해 포도당올 홉수하는데, 포도당에 방사성 동위원소를 함유시켜 동물에 주사함으혹써 나중에 방사성 동위원소의 소째를일정한 방식의 오토래디오그래피(autoradiography)에 의해 시각화함으로써 활동한 세포의 위치를 파악할 수 있다. 마취/마비된 원숭이에게 2-DG(2-〔14C〕deoxy-D-glucose, 탄소의 방사성 동위원소를 포함한 포도당)를 주사하고 그렴 2.6의 윗부분(A)에 제시된 패턴을 약 반 시간 동안
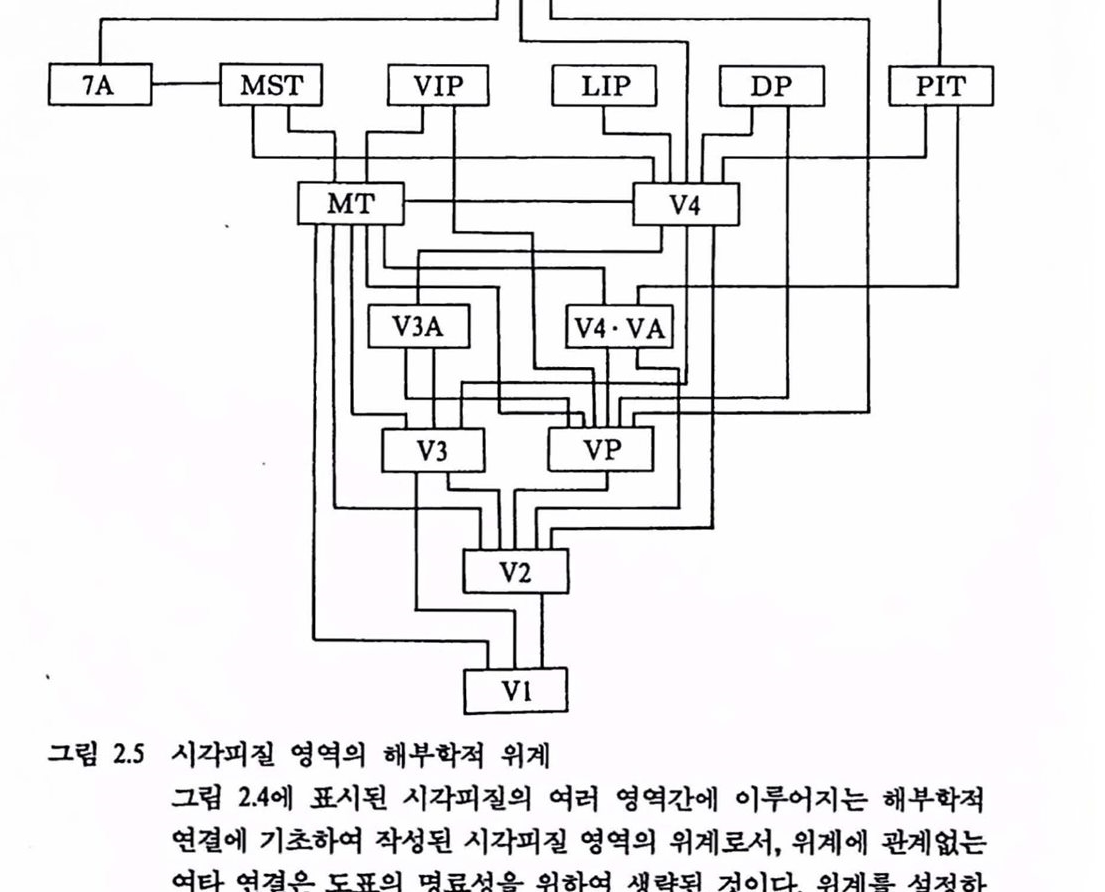
그립 2.5 시각피질 영역의 해부학척 위계 그렴 2.4에 표시된 시각피질의 여러 영역간에 이루어지는 해부학척 연결에 기초하여 작성된 시각피질 영역의 위계로서, 위계에 관계없는 여타 연결은 도표의 명료생을 위하여 생략된 것이다. 위계률 셜청하는 데 사용된 기준은 가끔 기농척 위계와 상반되는 수도 있으며, 생제의 해푸학척 연결상은 엄청나게 복잡한 것으로써 제시된 위계도는 실로 인위척얼지 모른다[Maunsell & Newsome(1987)에서 그렴만 옳킴].
고정된 눈으로 보게 한 후(마취/마비된 동물의 경우에도 초기 단계의 감각신경계의 뉴런들은 감각 자극에 대해 충실히 반응한다) 원숭이를 희생시켰을 때1차 시각피절에서 관찰되는 패턴 즉, (A)의 시자극에 대해 활동하는 세포들의 공간적인 분포가 그립 2.6의 아래(B)에 제시되어 있다. 이러한 시각자극에 노출된 원숭이의 제1차 시각피철에 있어서 지웅성에 따라 자극의 직선과 동심원을 표상하고 있는 세포들은 그렇지 않은 세포들에 비해 상대적으로 더욱더 활동하게 되고 따라서 더욱 많은 2-DG 를 홉수하게 되어 세포의 위치가 오토래디오그래피에 의헤 검게 나타난다. (A)의 시자극 패턴이 변형되어 (B)에 나타나 있다. 이 실험의 결과가 의미하는 바는 다음과 같다. 시공간은 엄격한 질서에 의해 제1차 시각피질에 표상되며 이 과정에서 시공간의 표상을 척절한 표상함수 (mappingf unction)에 의해 제1차 시각피질의 표면에 전이시킬 수 있다는 것이다. 이 표상함수의 특청은 시공간의 이심도가 각기 다른 부위는 시각피질에서 각기 다른 면적을 차지하게 되어 시각 과정에서 중요한 중심 시공간의 표상은 시각피질의 많은 면척, 따라서 많은 수의 세포에 의해 처리되어 고도의 해상력을 가진 분석을 가능케 한다는 것이다.
이심도(eccentricity)에 따른 시공간의 변형된 표상의 주된 결과로서 확대인자(magnification factor), 즉 단위 시공간(steradian으로 표시되는)을 표상하는 뉴런의 수 혹은 뇌의 부피가 이섬도에 따라 변하게 된다. 원숭이의 경우, 망막에서의 확대언자(망막의 단위 면척에 대한 망막 세포의 수)는 이섬도에 반비례하는데 확대인자가 이성도에 반버혜하는 정도는 외슬체, 시각피철에 이르면서 더욱 심화되는 것으로 생각되어 왔으나(Malpeli & Baker, 1975, Myerson et als, 1977, Van Essen et als., 1984), 최근 보다 세련된 기법을 사용한 연구에서 확대언자는 거의 일정 수준을 유지하는 것으로 제시되었다(Wassle et als., 1989). Schwartz(1980)는 시각피질의 확대인자를 셜명하는 함수, 즉 시각피질이 시공간을 변환하는 험수를 log(z+a)의 대수합수로 공식화하였다. 이 함수의 z는 주어진 위치가 와점에서 떨어진 거리, 즉, 극좌표에셔의 이심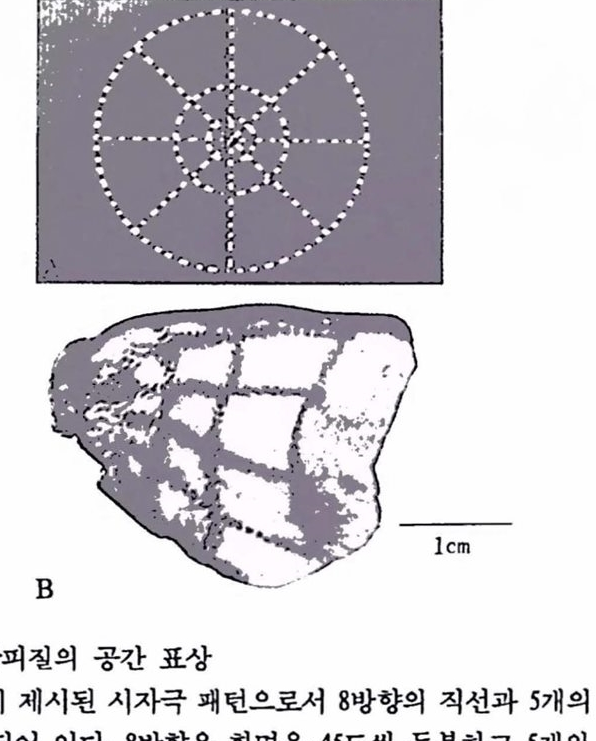 lcm
lcm
그림 2.6 제1차 시각피질의 공간 표상 (A) 화면에 제시된 시자극 왜턴으로서 8방향의 직선파 5개의 동심원으로 구성되어 었다. 8방향은 화연을 45도씩 둥분하고 5개의 동심원의 지름은 대수척인 비율로 이루어져 대수척도(logarithmic scale)에서 동일한 거리를 유지하도록 되어 었다. 이 칙선과 동성원들은 밝고 어두운 부분으로 이루어지는데 컴퓨터에 의해 3Hz 의 속도로 밝은 부훈은 어품게, 어두운 푸훈은 밝게 바뀌어져 지속척으로 망막훌 자극하도륙 고안된 것이다. 원숭이가 패턴의 중심올 웅시하면 왜턴의 오른쪽 반은 왼쪽 시각피철에 의해 표상되며 왜턴의 왼쪽 반용 오른쪽 시각신경계에 의해 처리된다. (B) 훤숭이의 제1차 시각 피철올 명명하게 만든 상태에서 활동척인 세포의 공간척 훈포률 위에서 본 것이다. 그립운 동물의 왼쪽 시각피철에 나타난 왜먼언데 (A)의 시자극의 오른쪽 반에 해당하는 것이다. (B)에서 화우로 달리는 5개의 선툴옹 (A)의 자극 왜턴의 우측율 구성하는 5개의 방사상칙선에 빼당하고 시자극의 동심웬에 해당하는 현들은 (B)에서 싱하로 달라는 칙선률에 의해 표상된 것이다[Tootell et als.(1982)에서 그렴만 옳검].
 ‘‘A、 -、/-“‘’-.(;L ‘ '( ---.,,tL- _- 컨t-1 .l
‘‘A、 -、/-“‘’-.(;L ‘ '( ---.,,tL- _- 컨t-1 .l
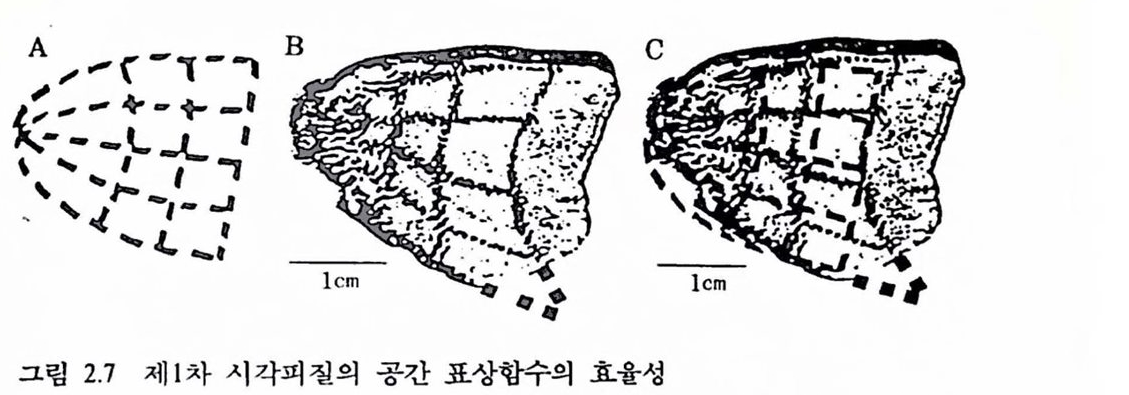
그럼 2.7 제1차 시각피질의 공간 표상함수의 효융성 (A) 그렴 2.6의 A의 시자극 패턴을 표상함수 log(z+O.3)으로 변환시컨 형태. (B) 그렴 2.6의 B의 형태를 보완하여 옮긴 것. (C) A와 B를 겹친 형태로서 A는 B를 효율척으로 기술한다. 조칙처리 과정에서의 조직의 수축, 곡면의 피질을 명연화시킬 때의 조칙 변형 둥을 감안할 때, 표상함수는 시각피질의 공간 변환을 쾌 찰 설명하고 있다 [Schwartz(1985), Science 227 : 1066에서 그렴만 옮깅].
도(eccentricity)이고 a는 동물의 종에 따라 달라지는 상수이다(Schwartz, 1980). 원숭이 시각피질의 경우 a는 0.3으로서 시공간의 표상함수는 log(z+0.3)으로 표시된다. 그렴 2.7은 이 함수의 효융성을 보인다.
5. 인공시각장치 5.1. 개념 그렴 2.5에서 주목할 점은 시각피철의 모든 영역에 천달되는 시각환경에 대한 신호는 제1차 시각피질(V1)올 통해 제공된다는 정이다 (Van Essen & Maunsell, 1983). 제1차 시각피질 이후의 영역에셔 추구되는 시각 환경의 여러 속성들은 제1차 시각피질의 시각 환경에 대한 엄격한 지도에 기초하고 있다. 따라서 안구의 기능융 대체활 수 있는A. 정상적 시각 경로
 영상-關→앓꽉 -|제 l 차 시각피질| -|화시각피질|
영상-關→앓꽉 -|제 l 차 시각피질| -|화시각피질|
B. 신경보철에 기초한 인공시각의 개념도
 영상 |제 l 차 시쐐질|→|과 시각피절 |
영상 |제 l 차 시쐐질|→|과 시각피절 |
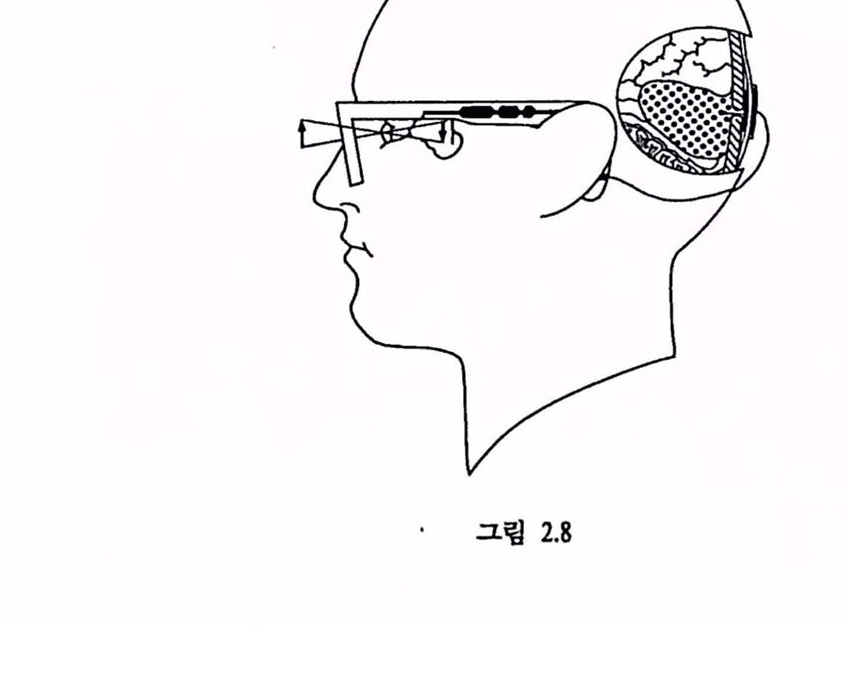 C.언 공시각장치의 구현 모숭
C.언 공시각장치의 구현 모숭
그렴 2.8
센서의 출력을 표상함수에 따라 제1차 시각피철에 직접 전달함으로써 망막-외슬체의 신호 변환 및 천달 과정을 생략할 수 있다(그립 2.8). 예를 들어, 그림 2.6의 B의 패턴에 위치한 뉴런 집단을 언위적으로 활성화할 때 경험하는 내용은 그렴 2.6의 A의 자극 패턴을 〈보는〉 것과 통일한 것은 아닐지라도 그와 홉사한 감각 경험일 것으로 추측된다. 이론척으로는 센서의 출력이 망막과 시각피질 사이에 위치한 외슬체에도 연결될 수 있지만 이 구조는 뇌간의 심부에 위치하여 외부에서의 접근이 용이하지 않으며 2차원적인 조망에서는 외슬체의 공간 표상이 시각펴철의 경우보다 훨씬 복잡하여(예를 들어 Malpeli & Baker, 1975 : Lee et als., 1984) 현실적 신경보철의 가능성이 희박하다.
5.2. 인공시각의 구현을 위한 연구와 과제 시각피질의 전기자극을 통하여 언공시각을 유도하교찌 하는 시도는 오래전부터 있어 왔다. 인간의 시각피철의 한 점을 전기척으로 자극하면 피험지는 〈흰 섬광〉을 보는 것처럼 느끼는데 이때 경험하는 섬팡이 시공간에서 차지하는 위치는 자극되는 시각피철의 위치에 따라 변하는것으로 보고되어 왔다(예를 들어, Brindley & Lewin, 1968; Dobelle & Mladejovsky, 1974). 이러한 실험 예의 확장 즉, 다수의 전극올 통해 각기 다른 공간의 위치에서 동시에 빛을 경험하게 함요로써 정지된 때헌의 언식이 가놓한 것이 실험척으로 업중되었다. Dobelle 둥(1976)은 이에 착안하여 시각피질의 전기 지극을 통해 맹인이 정자를 언식할 수 있는지를 시험하였다. 실험에 자훤한 맹언의 시각펴철에 64개의 독럽척으로 자극이 가능한 백금 전극이 삽업되고 점자에 해당하는 위치의 천극조합을 통해 전기 자극이 이루어졌을 때, 맹인은 정자률 〈률〉 수 있었옳 뿐 아니라 손을 통해 파악하는 것보다 더 봐르고 청확하게 언식할 수 있었다.Dobelle 등(1976)의 연구는 신경보철에 의한 시각보조를 지지하는 희망적인 결과를 보였지만 이러한 장치가 실용적으로 적용되기 위해서 해결되어야 할 문제들이 었다. 우선 전기 자극에 의해 유발되는 빛이 경험되는 공간에서의 위치는 치말적인 안구운동에 따라 변하며 이때 시각 경험의 위치는 안구의 이동 방향과 같은 방향으로 움직인다. 따라서 실용적인 장치는 자발적인 안구의 운동을 측정하여 이를 고려하는 모률을 필요로 한다. 안구의 운동이 시각 경험에 미치는 영향은 체계적인 동물 실험을 통해 평가될 수 있을 것이다. 또한 시각 장치가 실용적으로 사용되기 위헤서는 장기간의 접속에 따르는 뇌의 손상과 천극의 손상을 극소화시키는 재료의 개발과 전극 구조의 개선 문제등이 해결되어야 할 것이다.
그동안의 언공시각의 연구는 장애자 재활에 대한 적극적 개념의 결여와 이를 뒷받침하는 자훤의 결여로 인해 일부 임상실험을 제외하고는(예를 들어, 최근 미국 국럽 보건원의 G. E. Loeb와 동료들, 그리고 유타대학 의학팀에 의한 인간 피절에 사용한 미발표 실험) 기초실험에 머물러 있는 것이 각국의 공통된 현실이다. 이러한 실험들의 그칸의 성과는 주로 기농적 시각의 회복에 이르기보다는 전기 자극에 의한 뇌조칙의 부작용, 전기화학척 반응에 의한 전극의 손상 등에 대한 명가들이 었다. 그러나 인공시각과 관련된 초기의 부정척 효과가 인공시각에 대한 연구의 종료를 의미하지 않는 것은, 20여 년 전의 언공심장의 초기 연구나 10여 년 전의 단일 전극에 의한 달팽이판의 전기 자극 당시의 불확실한 실현성을 돌이켜 볼 때 특히 그러하다. 신경보철에 의한 시각 보초장치의 구현에 있어서 해결되어야 할 파제기운데 가창 중요한 것은 충분한 해상력을 유지하면서 시각피철의 각 부분의 활동올 척은 전류로 유발할 수 있는 다중 채널전극의 개발이다. 언공달팽이판의 경우 중다 채널의 전극 배열은 달팽이관올 따라 얼차원척으로 배열된 것이다. 그러나 시각펴질의 신경제어의 경우 이차원척언 피질 표면을 제어하기 위해서는 청각신경보철의 경우보다 훨씬많은 수의 전극이 요구된다. 얼마나 많은 수의 전극이 필요한지는 보조하고자 하는 시각 행동에 따라 결정될 것이지만, 정확한 형가는 이루어지지 않고 있다. 신경활동을 유발하는 자극의 방식으로서 현재까지 미세 전기자극이 가장 보편적으로 사용되어 왔다. 뉴런의 전기척 활동을 측정하거나 인위적으로 활성화하는 데 사용되어 온 천극은 흔히 머리털 정도의 가는 텅스텐, 백금, 이리륨 등의 철사로 만틀어지는데 철사의 끝 5μm(미크론은 1,00O분의 1mm) 정도를 에칭시켜 절연체 밖으혹 노출시킨 것이다. 그렴 2.9는 이러한 텅스텐 전극 15개를 사용하여 고양이의 제1차 시각피질의 독립적인 15개의 위치의 뉴련의 활동을 동시에 측정하기 위해 휠자의 동료들에 의헤 개발된 것이다(Schwark et als, 1986).이 전극 배열을 통해 독립적으로 미세 전류의 전기 자극도 가능하다.
Najafi 와 Wise(1986)는 다소 개선된 중다 천극을 개발하였는데 이 전극은 고속으로 작동하는 스위치 회로를 기판에 포함하여 전극 뒷부분에 연결되는 전선의 수를 감소할 수 있고(그림 2.9의 15채널 전극의 경우 연결된 전선의 수는 15개인 데 비해.이들의 중다전극은 10채널에 대해 3개의 선으로 기능), 사진석판술(photolithography) 등의 칩척회로 제작 기법을 응용하여 각 전극의 저향을 균일하게 유지할 수 있다. 기능적 시각보철을 위해서는 수백 이상의 전극이 필요할 것이며 많은 수의 전극을 포함하는 전극배열의 소형화를 위해서 접척회로 제작 기법의 응용은 앞으로 추구되어야 할 것이다. 전기적 자극은 현재 국부 신경조칙의 활성화를 유발하는 점에서는 가장 유리하지만 뇌조직에 접하는 전극의 끝과 제어 회로 사이를 연결하는 전선의 존재로 염증 동의 문제률 야기할 수 있다. 그러나 현재 진행되고 있는 라디오 주파수, 광학척 방법 등의 비정촉척인 방식의 신호전달이 개발되면 이러한 문제는 조만간 해결훨 것이다. 또한 뇌조칙을 활성화하는 것으로 알려진 낮은 강도의 빛(Wade et als., 1988)이나 자창(Yamamoto et als., 1988)은 제한된 부위에서 자연척언 신경확동훌 유발할 수 있는 측면에서 앞으로 개량이 이루어지면 시각피질옳 직정
그림 2.9 다충 채널 전극의 예 에갱되어 철연체로부터 노출된 텅스렌 철사의 끝 부분들은 사진의 아래 끝부환에 정렬되어 있고 이 천극들올 부분에서 에폭시 시멘트가 기판에 부착시키고 있다. 전극의 뒷푸분은 스텔레스 스띨판( 부훈)융 통해 금으로 도금펀 연결푸에 닿아 충폭 혹은 제어회로(사진에서는 보이지 않음)에 정속되도록 고안되었다. 뇌 조칙 속으혹 삽업되는 부분윤 실제 아꽤 끝 부분의 수 mm이다. 사진의 오른쪽 아래의 짧은 수명선은 1Omm의 걸이률 표시한다[Schwark et als.(1986)에서 사진만 옳김].
접촉하지 않는 신경보철이 할지 모른다. 접촉성 유발 방식의 경우 뇌조직의 거부반응 및 손상을 최소화할 수 있는 소재의 개발, 혹은 생화학적 억제제의 개발이 요구된다. Loeb(1989)는 탄탈, 이리륨을 소재로 한 전극의 우월성을 평가하였다.
자연적인 경로(망막-외슬체)를 통해 시각피질에 전달된 신호와 카메라의 출력에 해당하는 미세전기자극의 차이가 평가되어 이 차이를 최소화할 수 있는 방식이 확럽되어야 한다. 구체척으로, 시각중추에 센서의 출력에 해당하는 신호를 눈을 거치지 않고 제시하였을 때 가지는 의식척 경험이 실제 눈을 통해 전달된 신호와 차이가 있는지를 밝히고 차이가 있으면 그 차이를 작게 하는 신호와 신호전달 방법의 파라미터를 결정하여야 한다. 이러한 피라미터의 확립에는 원숭이를 사용한 동물실험이 유리하다.참고문헌 김종선, 김정유, 성명훈, 홍성화, 한문회, 윤미선, 송재우,「Nucleus 22 channel 와우 이식기를 이용한 인공내이의 임상척 시도」,〈대한 이비인후과학회지>, 32 : 174-191, 1989. 1. Clinical trials supported by the National Eye Institute, “Evaluating new approaches to the treatment of eye and vision disorders", by U.S. Department of Health and Human Services, Public Health Service, October 1988. 2. Vision Research, “A National Plan", by U.S. Department of Health and Human Services, Public Health Services, 1987. 3. Burkhalter, A. & Bernardo, K. L., "Organization of corticocortical connections in human visual cortex" Proc. Nat’l. Sci. USA,86 : 1071- 1075, 1989. 4. Dobelle, W. H. & Miladejovsky, M. G ., "Phosphenes produced by electrical stimulation of human occipital cortex, and their application to the development of a prosthesis for the blind", J. Physiol., 243 : 553-576, 1974. 5. Dobelle, W. H., Miladejovsky, M. G. & Girvin, J. P., "Artificial vision for the blind : electrical stimulation of visual cortex offers hope for a functional prosthesis", Science, 183 : 440-444, 1974. 6. Dobelle, W. H., Miladejovsky, M. G. & Evans, J. R. “‘Braille’ reading by blind volunteer by visual cortex stimulation", Nature, 259 : 111- 112, 1976. 7. Hubel, D. H. & Wiesel, T. N., “Receptive fields, binocular interaction and functional architecture in the cat’s visual cortex", J. Physiol., 160 : 106-154, 1962. 8. Hubel, D. H. & Wiesel. T. N., “Receptive fields and functional architecture in two non-striate visual areas(18 and 19) of the cat", J. Neurophysiol., 28 : 229-289, 1965.
9. Hudspeth, A. J., “How the ear’s works work", Nature, 341 :397-404, 1989. 10. Lee, C., Malpeli, J. G ., Schwark, H. D. & Weyand, T. G."Cat medial interlaminar nucleus:retinotopy, relation to tapetum and implications
for scotopic vision", J. Neurophysiol., 52 : 848-869, 1984. 11. Lee, C., Rohrer, W. H. & Sparks, D. L., “Population coding of saccadic eye movements by neurons in the superior colliculs", Nature, 332: 357- 360, 1988. 12. Malpeli, J. G. & Baker, F. H., “The representation of the visual fieldin the lateral geniculate nucleus of Macaca mulatta", J. Comp. Neurol, 161: 569-594. 13. Loeb, G. E., “Neural prosthetic interfaces with the nervous system", Trends in Neurosciences, 12: 195-201, 1989. 14. Maunsell, J. H. R. & Newsome, W. T., “Visual processing in monkey extrastriate cortex", Ann. Rev. Neurosci., 10: 363-401, 1987. 15. Myerson, J., Manis, P. B., Miezin, F. M. & Allman, J. M., “Magnification in striate cortex and the retinal ganglion cell layer of theowl monkey:a quantitative comparison", Science, 198: 855-857, 1977. 16. Najafi, K. & Wise, K. D., “An implantable multielectrode array withon-chip signal processing", IEEE Solid-State Circuits, Sc-21:1035-1044, 1986. 17. Robinson, D. A.“Eye movements by collicular stimulation in the alertmonkey", Vision Res., 12:393-404, 1970. 18. Schwark, H. D., Malpeli, J. G., Weyand, T. G. & Lee, C ., “Cat area 17. II. Response properties of infragranular layer neurons in the absence of supragranular layer activity", J.Neurophysiol., 56: 1074 -1087, 1986. 19. Schwartz, E. L., “Computational anatomy and functinal architecture of striate cortex:A spatial mapping approach to perceptual coding", Vision Res.20: 645-669, 1980. 20. Sparks, D. L., "Translation of sensory signals into commands forcontrol of saccadic eye movements:role of primate superior colliculus", Physiol. Rev., 66:118-171, 1986. 21. Sparks, D. L. & Mays, L. E., “Movement fields of saccade-related burst neurons in the monkey superior colliculus", Brain Res., 190: 39-50, 1980. 22. Sparks, D. L. & Mays, L. E., “Signal transformations required for the generation of saccadic eye movements" Ann. Rev. Neurosci., 13: 309 -336, 1990. 23. Tachi, S., Tanie, K., Komoriya, K. & Abe, M.,“Electrocutaneous communication in a guide dog", IEEE Trans. Biomed. Eng., 32: 461- 469, 1985. 24. Van Essen, D. C. & Maunsell, J. H. R.,“Hierachical organization and functional streams in the visual cortex", TINS, September, 370-375, 1983. 25. Van Essen, D. C., Newsome, W. T. & Maunsell, H. R., “The visual field representation in striate cortex of the Macaque monkey:asymmetries, anisotropies, and individual variability" Vision Res., 24: 429-448, 1984. 26. Wade, P. D., Taylors, J. & Siekevvitz, P.,"Mammalian cerebral
cortical tissue responds to low-intensity visible light", Proc. Nat’l Acad. Sci., USA, 85:9322-9326, 1988. 27. Wassle, H., Grunert, U., Rohrenbeck, J. & Boycott, B. B.,"Cortical magnification factor and the ganglion cell density of the primate retina" Nature, 341: 643-646, 1989. 28. Yamamoto, T., Williamson, S. J., Kaufman, L., Nicholson, C. & Llinas, R., "Magnetic localization of neuronal activity in the human brain", Proc. Nat'l Acad. Sci., USA, 85: 8732-8736, 1988.제 2편 뇌의 인공적 확장은 가능한가
박순달·소광섭 제 3 장 뇌의 자연진화와 역할 1. 생명의 진화 1.1. 우주의 생성 우주의 기원과 미래에 대한 종교척 견해는 크게 두 가지로 분류할 수가 있다. 그 하나는 기독교 계통으로서 우주는 어떤 조물주에 의하여 그리 걸지 않은 과거에 창조되었다고 보는 것이다. 예률 들어 아우구스티누스(Aurelius Augustinus)는 성경의『창세기』에 의하여 기원천 약 5000년 전에 우주가 창조되었다고 계산하였다. 이러한 신의 창조론에 반하여 그리스의 철학자들이나 동양의 사상가들, 특히 폴교 계통에셔는 우주가 영훤한 과거에서 미래까지 순환척언 반복올 거륨한다고 믿었었다. 우주와 천체들이 성 · 주 · 괴 · 공(成住城空)의 과정으로 운행되는 큰 주기를 겁(Kalpa)이라는 단위로 불렀으며 한 계산율 언용하연 I대갱이 43억 2천만 년이라고 나와 있는데, 현대물리학에셔 우주의 역사훌 약 200 억 년으로 하는 것파 크게 다르지 않다. 앞셔 창세기외 혜석훌
약 5000년 우주역사로 본 것파 비교하면 고대 인도인들의 사고의 스케일을 짐작케 한다.
우주가 시간적으후 처음이 있었는지의 문제를 칸트는 그의『순수 이성비판』에서 순수 이성의 이율배반이라 하였다. 왜냐하면 우주에 시작이 있었다는 생각과 끝없는 과거부터 존재했었다는 생각이 둘 다 이유가 있는 주장이라고 여거지기 예문이었다. 시작이 없다면 어느 사건이나 무한히 긴 과거가 있다는 얘기이므로 믿기 어렵고, 시작이 있다면 그 이전에는 없다가 왜 하필 그때에 생겼는가 하는 답하기 어려운 질문이 나오기 때문이다. 이러한 생각은 누구나 한번쯤 해보았을 것인데, 이 생각의 바탕에는 우주라는 공간적 존재와 분리되어서 따로이 흐르는 시간이 있다는 가정이 깔려 있다. 오늘날 시간과 공간이 하나의 시공간으후 통일된 일반상대론의 업장에서 보면, 우주는 시공간적 개념으로써 우주 이전의 시간이란 논리척으로 성립조차 되지 않는 말이다. 마치 시간이 생겨나기 전 시간이란 무엇언가고 묻는 것과 같다. 아우구스티누스가 이미 이를 지척하여 〈시간이란 신이 창조한 우주의 특성이고, 우주가 시작되기 전에는 시간이 존재하지 않았다〉고 말했었다. 우주의 시간척 특성 못지 않게 공간척 성철도 많은 사상가들의 관심을 끌어왔는데 우주의 공간척 크기가 유한이냐 무한이냐 히는 것이 문제였었다. 우주가 무한히 크다면 올버스(Heinrich W. M.Olbers) 꽤러독스에 부딪치게 된다. 무한히 큰 우주는 어디가 특별한 중심이라 할 수 없으므로 우주의 모든 곳에 지상에서 보는 것과 버슷한 분포의 별이 있올 것이고, 그렇다면 약간의 계산으후 어느 곳이냐 밤낮없이 대낮같이 밝아야 한다는 결흔올 얻올 수 있다. 이와 반면에 유한한 우주률 생각하면 그 우주 밖은 무엇인가 하는 의품에 탑할 수 없게 된다. 우주의 공간문제에 있어서도 역시 순수 이성의 이율배반에 봉칙하는데, 이것 역시 공칸올 우주와 분리해서 순수 유클리드 공간으로 가청하는 데에서 오는 것이다. 우주는 전부이기 때푼에 우주 밖의 공간은 우주이전의 시간과 마찬가지로 자체 모순되는 진술인 것이다. 이 점에관해서 고대 중국의 제자백가 중에서 아주 적절한 답을 볼 수 있는데 〈우주는 너무 커서 바깥이 없다[無外]> 라고 말하고 있다. 우주 밖의 바깥이라는 개념은 성립하지 않음을 일찍부터 깨쳤던 듯하다.
우주의 시공간적 특성으로 동양의 주기 순환척 개념 대신에 서구에서 는 정적(static)인 우주관이 지배적이었는데 이러한 가정은 하도 뿌리 깊은 것이어서 뉴턴은 물론 아인슈타인까지도 이 영향을 받아 틀린 우주론을 만들기도 했었다. 1929년에 허블이 은하들의 운동을 관측하여 우주의 팽창을 알아내고서야 비로소 아인슈타인의 일반상대론과 일치하는 우주의 동역학적 모형들어 가능하게 되었다. 우주흑 단순히 물질의 집합체로서 파악하고 이것들의 총체로서 규정하는 것이 서구 과학의 주류언데, 희랍인은 우주를 Cosmos로 불렀으며 이는 질서, 법칙을 의미하는 것인바, 그틀의 신화에는 Cosmos 이전에는 Chaos(흔돈)가 있었다고 보는 흥미로운 견해를 가졌다. 이는 우주를 객관적 대상으로서 파악하기보다는 인간의 인식체계로서 본 것으로 영국의 버클리(Berkley) 주교가 물질적 대상이나 공간이나 시간이 인간의 환상에 불과하다고 주장한 것과 궤를 같이하는 것이며, 불교의 일체유심조(一切唯心造)나 공(空)사상과 상통한다고 본다. 미래의 우주론은 어쩌면 언간의 두뇌 구조 및 인식 활동파 연판되어 파악될지도 모를 일이다. 사상으로서 혹은 철학으로서 우주흔은 옛날의 성현이나 철언틀의 지혜가 오늘날의 지식보다 더 현묘하거나 바른 것일는지 모료냐, 판측사실의 이론화라는 측면에서 과학으후서의 우주론은 그려스 시대부터 현대에로 정진척 발전을 거듭해 왔다. 코페르니쿠스 이전의 천문학은 프툴레미의 천동셜이 대표척인데 지구가 우주의 중심이고, 그 툴레률 달파 행성(수성, 금성, 화성, 목정, 토성) 그리고 태양과 항성이 원운동을 하고 있다고 생각하였다. 판측자료가 정정 쌓이고, 정멀성이 늘어나자 행성들의 궤도가 단순한 훤운동이 아니며 복잡한 것임이 분명해졌는데 이률 보완하기 위하여 큰 훨주 위에작은 원궤도를 추가하는 형식으로 보완하여 아주 정확한 예측이 가능했었다. 그러나 코페르니쿠스 때쯤 되어서는 행성의 궤도를 예측하기 위하여 수십 개의 원을 써야 하는 복잡성의 극에 달하였으고로 마침내 태양 중심의 지동설이 1514년에 새로운 모델로서 등장한 것이다. 이후 갈렬레오와 케플러에 의하여 지동설이 확고히 되고, 뉴턴의 운동볍칙과 중력이론의 출현으로 근대적 천문학이 정립되었다.
뉴턴의 역학체계는 태양케내 행성의 운동을 설명하는 데 거의 완벽할 정도였으나, 태양계보다 더 먼 일반적 별들의 운동이나 우주론 등에 대해서는 별로 기여한 것도 없었고 또 이런 것을 측정하는 관측 데이터도 별로 없었다. 우주론에 관해서는 종래의 관점인 보편적으로 흐르는 시간과 물체들이 운동하는 무한 공간의 독립된 존재를 인정하고 있었다. 따라서 우주란 무한 공간에 별들이흩어져 있는 것으후 여걱쳤고, 유한 우주란 별들의 수가 유한해서 공간의 일부에 모여 있는 것을 의미하였다. 이러한 우주 모형은, 뉴턴의 중력법칙에 의하여 별들이 서로 끌어당기므로 마침내 한 점으로 모두 뭉쳐지는 모순정을 내포한다. 이 점은 뉴턴도 이미 언식하고 있어서 우주가 무한해야만 된다고 생각했었다. 그러나 이 경우에는 다시 올버스의 패러독스에 부딪친다. 일반상대론이 나오기 전의 우주롱은 대체로 이런 정도의 관념적 연습에 그치는 것이었고, 과학척 이론이 가져야 할 예측과 관측이란 것이 결여된 상태이었다. 아인슈타인의 일반상대론이 나오자 물철(별의 분포)과 시간과 공간이 모두 연판되어 맞물련 하나의 시스댐으로서 우주관이 성립되었고, 우주 전체률 다롤 수 있는 방정식과 그의 풀이 및 예촉이 가능한 파학척언 이론으혹서의 우주론이 태어날 수 있었다. 아인슈타인은 언제나 일정한 크기와 모습을 갖는 정척언(static) 우주를 그의 방정식으로부터 유도하려고 노력했으나 실왜하였다. 그의 방청식은 그런 우주의 존재를 부정하였으므로, 그는 방청식을 약간 수정하기까지 하였었다. 그러나 1929년 허블(Edwin P. Hubble, 1889~1953)이 은히들의 판촉을 통하여 은하틀이 우리로부터의 거리에 비례하는 속력으로 급속히 멀어져간다는 사실을 발견하였으며, 이를 우주의 팽창으혹 해석하였다. 이것은 아인슈타인이 굳게 믿었던 정적인 우주가 아니라 변하는 우주엄을 보여준 것으로 그의 원래 방정식과도 모순없이 맞아떨어지는 것이었다. 우주가 팽창하고 었다면 먼 과거에는 더 작은 우주였을 것이고, 또 시작도 있었을 것이다. 이것은 대략 200억 년쯤 전으로 거슬러올라가는데, 그때에는 우주는 작은 공간에 모든 물질이 응집되어 있는 상태였는데, 대폭발(Big Bang)이라는 과정으로 긴 역사가 시작되었음을 의미 한다.
상대론적 시공간이론과 소립자 물리학이론은 극대와 극미의 현상을 다루는 끝과 끝인데 우주의 시작에서는 이 두 분야가 하나로 만나게 된다. 초기 우주는 작은 대신 아주 고온의 상태에 있으부로 소렵자의 상호작용이 극심하게 일어나는 상황이며, 이 시기의 척절한 이론척 계산을 한다면 은하계의 기원과 분포, 우주내 수소와 헬륨 둥 원소의 분포, 마이크로파 우주복사의 존재와 성격 등등 여러 가지 예측을 할 수가 있다. 이 이론을 〈대폭발이론〉이라 부르는데 실험척 관측치가 이론과 찰 맞아떨어지므로 널리 받아들여져 표준 모형이라고도 한다. 우주의 과거에 대한 대체적 이해가 되었다면 미래는 어떠한가? 팽창을 계속할 것인가 아니면 어느 시기에 가서 다시 수축하기 시작할 것인가? 이 철문에 대한 답변은 우주내 물질의 분포 멀도률 정확히 알면 알 수 있는데 불행히도 관측 데이터가 부정확하여 어떤 결론도 내렬 수 없는 형편이다. 1970년대부터 활발히 연구되고 있는 문제는 우주의 폭발 당시의 상황에 대한 연구이다. 폭발 당시에는 공간의 크기가 극히 작고 온도는 아주 높은 상황인데 이러한 조건의 물리현상올 다루려면 중력창의 양자화를 훨연척으로 요구하게 되지만 아칙 이것이 없는 상태이다. 즉 양자중력장 (quantum gravity) 이론이 부채한 상태이묘로 계산과 예혹이 있옳 수 없는 것이며, 다만 여러 가지 제안파 가갤들이 나요교 있을 뿐이다.폭발의 순간과 폭발 직후의 우주의 형상에 대한 연구는 소렵자이론, 중력장이론, 양자이론, 통계물리이론이 모두 동원되는 물리이론 자체의 대단원적 클라이막스를 요하는 최종의 연구과제이다. 하나의 상상을 덧불여본다면 물리이론의 총화가 요구되는 우주의 기원 문제가 인간의 두뇌작용과 인식작용, 즉 정신문제의 개업 없이 해결될 수 있을는지 의아해하지 않을 수 없다. 두뇌 또는 정신 기능의 이해를 위한 노력이 궁극적 우주이론에 크게 기여할 때도 있을 것이라고 상상해본다.
은하와 태양 견우와 직녀가 건너가 만난다는 밤하늘의 강 은하수는 태양과 같은 별들이 원반형으로 모여 있는 별들의 큰 집단이다. 우리의 육안으로 직접 볼 수 있는 별은 대략 6,000개 정도이지만 은하수를 구성하는 별의 수는 대략 1천5백억 개 정도가 된다. 우리가 살고 있는 은허수는 수없이 많은 은하들 중의 하나에 불과한데, 은하들의 형태는 대체적으로 나선형, 막대나선형, 타원형 둥이며 우리의 은하수는 나선형으로서 은하계 질량의 대부분이 중심 부분에 집중되어 있고, 나선형의 회전하는 팔 모형에 가스와 티끌의 밀도가 크고 별들이 집중되어 있다. 은하계 중심에서는 큰 폭발 현상이 일어나고 있다고 믿어지며, 팔 쪽에서 별의 생성 이 활발히 일어난다. 우리의 은하수는 측면도가 마치 원반형 버행접시를 연상시키는 팔을 하고 있는데 중심에서 훤반 끝까지 반경이 약 45,000 광년이고, 원반 천체가 회전을 하는데 그 속력은 초속 약 250km로서 한 회전에 걸리는 시간은 대략 2억 년이며, 은하수 자체의 나이는 대략 50억 년 청도로 추청된다. 은하수 안의 광막한 세계에는 별뿐만 아니라 약 10%의 질량은 별틀 사이의 공칸에 흩어져 있는 가스와 티끌로 되어 었고 그 밖에 자기장과 전자, 양성자 동 우주선이 었다. 그리고 최곤에 발견된 사실로는 〈보이지 않는 물철(dark matter)>이라는 정체가 아칙 밝혀지지 않은 물질이 은하수 전체를 툴러싸고 펴져 있다고 하며, 이 보이지 않는 물철의 철량이 멸과 성간 물질들의 질량의 합보다 더 크다고 보여지고 있다. 이들에 대한 연구가 현재 활발히 진행되고 있다.
별들의 희미한 모습처럼 보이는 성운이 실은 은하수 밖의 다른 은하계들이며 은하수와 다소 유사한 구조를 가졌을 것이라고 처음 제안한 사람은 칸트였으며, 은하계들의 거리를 체계적으로 측정하고 그들의 분포를 알아낸 사랑은 허블이었다. 은하수로부터 제일 가까운 은하계는 안드로메다 은하계로서 약 80만 광년 떨어져 있다. 은하계틀은 여러 개가 떼지어 은하군을 형성하는데 우리 은하수는〈국부군(local group)>이라 불리는 20개 정도의 은하계를 포함한 것이고, 처녀화에는 수천 개의 은하들이 하나의 군을 형성하고 있다. 은하들을 체계적으로 관측한 허블은 은하들이 빼른 속력흐로 달아나고 있다는 것을 알게 되었으며, 달아나는 속력이 거리에 비례함을 밝혔다. 수억 광년까지의 거리에 이르는 수억 개의 은하를 포함하는 영역에서 이와 같은 법칙이 성럽함이 확실시되었고, 이는 우주의 팽창으후써 가장 잘 셜명이 된다. 은하의 서로 달아나는 속력을 저꾸로 돌려 계산하면 모든 은하들이 약 100~200억 년 전에 한 점으로부터 출발했다는 결론이 나오며 우주의 대폭발이론이 나오는 계기가 되었다. 빛으로 관측할 수 있는 별들만이 우주를 형성하고 있는 것이 아니다. 2차 대전 이후에 레이더의 발전으로 전파 망원경이 발전함에 따라 천파를 방출하는 전파은하들어 수없이 발견되었다. 이들 전파은하들이 전파로 방출하는 에너지가 보통 은하들의 벚 방출 에너지보다 엄청나게 크다고 보이는데, 이런 에너지를 방출하는 물리척 원인들은 아직 밝혀지지 않았다. 전파은하 외에 많은 관심올 콜게 된 것으로서 quasar(quasistellar radio source)라는 유사 항성이라 할 수 있는 존재들은 척색편이(red shift) 도플러 효과가 대단히 크면서 아주 밝은 별 같은 것들이다. 이들의 거리를 허블 법칙에 따라 측정하면 거리가 대단히 먼 것이므로 보이지 않을 만큼 희미해야 할 터인데 실제로는 그렇지 않으묘로, 그컷의 철대광도는 보통 은하의 100배 정도나 된다는 계산이다. 어떻게 이러한 막대한 에너지가 나오는 것일까, 퀘이사는 정말로 아주 먼 콧에 있는 것일까 등은 아직 확실한 답이 주어져 있지 않은 상태이다.
천문학의 발전을 되돌아보면, 처음에는 지구 중심의 좁은 세계에서 태양 중섬의 태양계를 관측하는 정도로 발전했다가 20세기에 들어서 비로소 은하수의 크기와 구성이 이해가 되었고, 이러한 은하들이 수억 개냐 된다는 것을 알고 또 빠른 속도로 서로 달아남을 안 것은 1930년대에 들어와서였다 .2차 대전 이후에 전파은하와 례이사의 발견으로 관측된 우주의 상황은 더욱 확대 복잡해졌다. 앞으로도 더 큰 발견이 있을 것을 상상할 수 있으며, 우리가 지금 우주라고 생각하는 것이 정말 더 큰 우주의 극히 일부에 불과할는지도 모른다. 별과 태양 우리의 은하수는 수억 은하 중 전형척인 보통의 한 은하에 불과하다. 이 은하수에는 1천억 개 이상의 별이 모여 있고, 태양은 그 많은 별들 중 명범한 하나의 별에 불과하다. 은하수의 원반 반경이 약 4만 5천 광년언데 태양은 은하수 중심에서 약 3만 광년 떨어진 곳에 위치하고 있다. 1- 4 만 5000 뺀 나
1- 4 만 5000 뺀 나
그립 3.1 은하수 모형과 태양의 위치(작은 원은 구상 성단을 나타낸다).
별은 수소 가스들이 중력에 의하여 풍치면서 열이 발생하여 온도가 상승하고 그에 따라 에너지를 방출하면서 진화과정을 겪는다. 태양과 갇은 별은 아주 많은 에너지를 방출하고 있는데 그 에너지원은 수소가 핵융합 과정에 의하여 헬륨으로 변할 때 생기는 에너지이다. 태양 정도의 철량을 가진 멸이 수소를 모두 태우고 헬륨으로 변하는 데는 약 100억 년 정도의 시간이 걸린다. 태양 자신은 생겨난 지 수십억 년이 된 것으로 알려져 었다.
별의 진화과정은 수소가 헬륨으로 변화된 후 다시 약 1억 도의 고온이 되도록 수축하면서 헬륨이 타면서 탄소로 바뀌는 과정을 겪는다. 별의 질량이 태양 질량의 1.4배 이하라면 별 내부의 천지들의 운동에 의한 압력으로 멸의 중력 수축은 더 이상 진행되지 않게 된다. 이러한 별은 질량은 태양과 비슷하면서 반경은 태양 반경의 1/100 정도로 작아서 밀도가 약 100만 g/cm이며, 이러한 별들을 백색왜성이라 부른다. 멸의 철량이 태양 절량의 10배 정도로 출발했다면 이 별은 수소를 다 태운 후 수소→헬륨→탄소의 파정을 겪고 계속 수축하여 마침내 철과 니켈의 핵으로까지 탄다. 그리고 핵 연료로서는 이 이상 더 탈수가 없게 된다. 이때 별의 모양은 맨 외곽부터 수소, 헬륨, 탄소------, 철의 동심원 구조를 이루고 있는데, 연료의 소진으로 더 이상 중력의 압력을 지탱하지 못하게 되어 갑작스런 함몰이 얼어나면서 큰 폭발이 얼어나며 이 현상을 초신성이라 부른다. 함몰이 얼어난 중심부의 물철의 상태는 핵이 모두 가까이 접근하여 중성자들로 이루어지는데, 이를 충성자 벌이라 부르며 중성자 별의 절량이 태양 정도라면 그 반경은 10km 정도밖에 안 되는 고밀도의 별이다. 이 중성자 별은 일정한 주기의 천자파률 방출하므로 펄사(pulsar)라고도 불련다. 태양은 질량이 2xlO g,밀도가 약 1g/cm 이고 온도는 내부가 10 70K, 표면이 6000 K 정도이다. 태양의 나이는 약 45 억 년쯤 되었고, 약 5억 년쯤 더 지나면 헬륨을 태우는 사이클이 시작될 것이고, 그 후 다시 3억 년쯤 더 지나면 척잭거성의 상태를 거쳐 마첨내 백잭왜쟁의 별이된 후 차츰 식어가는 멸이 될 것으로 추정된다. 태양이 현재 방출하는 에너지는 초당 3.9xl0 33erg 이고, 태양 대기의 약 74%는 수소, 약 25%는 헬륨이고 나머지는 기타 원소로 되어 었다.
태양케는 태양을 중심으로 회전하는 행성들과 기타 소행성, 혜성들로 구성되어 있으며 행성들은 작은 위성들을 가지고 있다. 태양 자체의 질량이 태양계 전체의 99.9%를 차지한 만큼 태양계의 운동은 태양의 인력에 의하여 지배되고 있다. 태양계가 어떻게 해서 생겨났는지에 대한 확실한 학셜은 아직 정립되지 않았다. 1.2. 지구의 역사 지금으로부터 30여 년 전인 1957년 소련의 Sputnik가 하늘로 발사되연서 우주의 시대를 맞이하게 되었다. 그 후 수많은 우주선을 역우면서 지구 이외에서 생명체를 발견하고자 하였으나 아직은 발견하지 못하고 있다. 그래서 태양계에서는 지구가 유일한 생명체가 살고 있는 별언 생이다. 그러나 태양케도 전체 우주에서 극히 일부에 지나지 않는다는 사질에 버추어 본다면 다른 멸에서도 생명체가 있을 가능성은 었다. 그럽에도 지구상의 생명체에게는 그 생명을 지탱시켜 주는 주위환경인 지구가 매우 중요하다. 그래서 지구가 어떻게 변화되어 왔는지를 보는 것도 대단히 의의 있는 일이다. 지구의 연대와 구훈 : 현재 지구상의 지충 속에서 35억 년 전의 남조류와 박테리아의 화젝이 발견되어 었다. 이로 보아 지구는 이보다 오래된 것임에는 툴렴없다. 보홍 지구는 태양계와 같이 형성되었다고 보고 그래서 운석의 나이와 같이 폴 수 있다. 현재 지구에 떨어진 가장 오래된 운석은 45억 년 전의 것인데 학계에서는 공식척으로 지구의 나이를 45억 년이라고 하고 있다.표 3.1 지질시대 구분
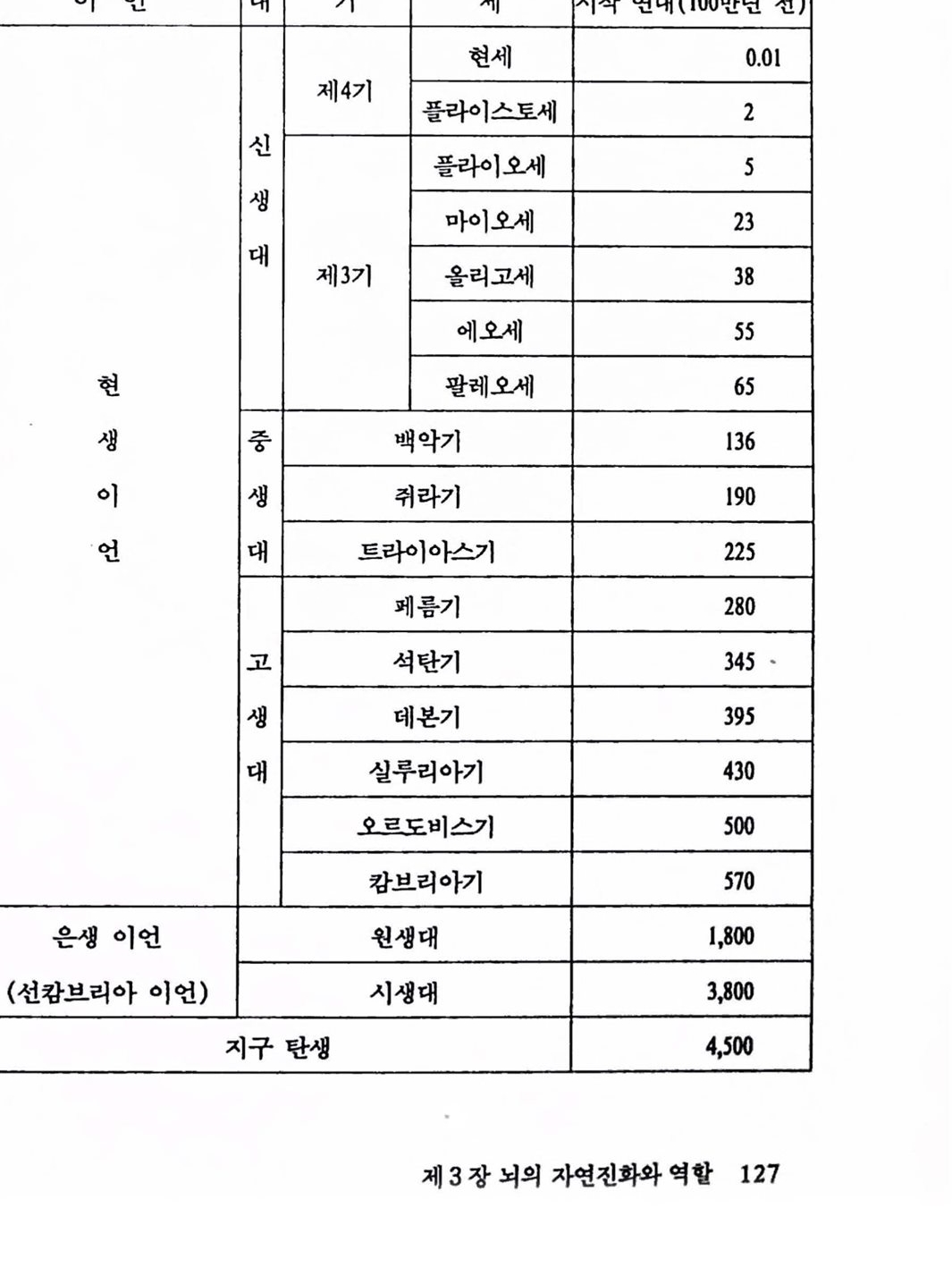 이 언 대기 세 시작 연대(1 00 만년 천)
이 언 대기 세 시작 연대(1 00 만년 천)
지구는 여러 지층들로 이루어져 있는데 시간이 흐르면서 다론 세대에 다른 지층이 생걱났었다. 그래서 이런 지층과 암석들의 상태와 그 속에 들어 있는 내용물들의 차이에 따라 지철시대를 구분한다. 지질시대는 단계척으로 이언(Eon), 대(代, Era), 기(紀, Period), 세(世, Epoch), 절(節, Age)로 나쉰다.
선캄브리아 이언〔Precambrian Eon, Cryptozoic Eon(은생이언)〕 : 지철연구에는 암석의 상태에 대한 연구와 화석(化石)의 연구가 중요하다. 다빈치(Leonardo da Vinci, 1452~1519)는 수에즈 운하를 팔 때 화석을 발견하여 화석의 중요성을 일깨운 선각자이다. 스코틀랜드의 허튼(James Hutton, 1726~1797)은 1788년『지구론』을 발간하여 지구에 대한 연구의 기틀을 잡았다. 선캄브리아 이언은 시생대와 원생대로 이루어져 있다. 지구는 45억 년 전에 탄생하였으나 지구의 역사에 있어서 생명의 출현이 가장 관심의 대상이 되고 그래서 지철시대의 구분도 생명계의 변화가 중요 구분 기준이기도 하다. 지구가 탄생한 후 한동얀은 물질의 변화에 그친다. 그러다그림 3.2 오스트랄리아 7억 년 전 화석궁
가 생명현상이 나타나기 시작하는데 호주의 35억 년 전의 지층에서 발견된 남조류와 박테리아 화석이 가장 빠른 화석이다. 최근에는 남아프리카에서 38.3억 내지 36억 년 전의 지층에서 5~30μm 크기의 원시조류와 같은 구조가 발견되어 가장 오래된 화석이 되었다. 이때가 곧 시생대(Archeozoic Era)의 시작이 된다. 원생대(Preterozoic Era)에 오연 대기에는 산소가 존재하게 되고 생물계에는 많은 변화가 일어나게 된다. 원생대의 암석은 변성되지 않는 암석으후 되어 있어 화석도 풍부하다. 이때의 생물에는 해파리, 벌레, 해면, 산호, 절지동물 둥이 포함된다.
고생대(Paleozoic Era) : 고생대는 현생 이언(Phanerozoic Eon)이 시작되는 시기로 이는 캄브리아기(Cambrian Period)에서 시작한다. 캄브리아기는 1833년 영국 남부에 발달한 지충을 캄브리아계로 명명한 데에서 비롯된 것인데 이때는 원생대말에 버하여 통물화석이 풍부하게 발견되며 특히 무척추동물이 나타나고 가장 혼한 것은 삼엽충 계통들이다. 고생대는 퇴척암층이 대부분이기 때문에 화석을 많이 남기고 있다. 고생대는 캄브리아기 외에 오르도버스기(Ordovician Period), 설루리아기(Silurian Period), 데본기(Devonian Period), 석탄기(Carboniferous Period), 페름기(Permian Period)가 있는데 각 기마다 그때 나타
그립 3.3 캄브리아기의 삼엽충
난 생물군의 특정이 있다. 오르도버스기에서는 두족류와 복족류가 크게 발전하였으며 어류가 출현한다. 실루리아기에서는 삼엽충은 사라지기 시작하고 무척추동물이 번식하며 육상식물과 어류의 시조가 나타난다. 데본기에서는 어류와 육상식물이 대표적이다. 그래서 데본기는〈어류의 시대〉라고도 한다. 석단기에서는 식물에서는 양치류가 발달하고 동물에서는 파충류가 나타난다. 페름기가 되면 양치류가 쇠퇴하고 소나무형의 나무가 나타나며 동물에서는 양서류, 파충류의 대부분이 천멸한다. 삼엽충도 완전히 없어진다. 대신 현대적 곤충이 나타나기 시작한다.
중생대(Mesozoic Era) : 고생대말에는 생물계에 많은 변화가 있었다. 대부분의 생물이 사라지고 만 것이다. 그 원인은 아직 확설치 않다. 중생대에 오면 〈파충류의 시대〉라고 할만큼 파충류가 번성하게 된다. 중생대는 트라이아스기(Triassic Period), 쥐라기(Jurassic Period), 백악기(Cretaceous Period)로 나누어지는데 트라이아스기는 삼첩기라고도 하며 파충류가 번성하기 시작한다. 특히 공룡과 어룡이 나타난다. 쥐라기에서는 공룡이 더욱 번성하고 조류의 선조인 시조새가 나타난다.
그림 3.4 공룡, 시조새
그리고 포유류가 나타난다. 곤충도 많이 번성하는데 장자리,메푸기, 진드기 등이 있다. 백악기에 오연 파충류의 진화가 극에 달하고 크기도 극대화된다. 그러나 무척추동물은 많은 종류가 멸종한다.
신생대(Cenozoic Era) : 신생대는 포유류의 시대이다. 이때는 중생대에서 번성하던 파충류가 많이 사라지는 대신 초식 내지육식 포유류가 번성하여 이 시대를 〈포유류의 시대〉라고 일걷는다. 신생대는 제3기(Tertiary Period)와 제 4기(Quarternary Period)로 나누는데 제3기는 현생 생물의 화석점유물에 따라 다시 · 팔레오세(Paleocene Epoch, 효新世) . 에오세(Eocene Epoch, 始新世) · 올리고세(Oligocene Epoch, 漸新世) · 마이오세(Miocene Epoch, 中新世) · 플라이오세(Pliocene Epoch, 蘇新世) 로 구분한다. 그리고 제 4기는 플라이스토세(Pleistocene Epoch, 洪積世) 현세(Holocene Epoch, 世) 로 구분한다. 신생대의 제 3기에서는 파충류 대신 조류의 증가와 더불어 육식 포유류가 번성하기 시작한다. 점신세에 오면 원시 포유류는 사라지고 중신세에 오면 유언원이 출현하며 선신세에 오면 인류의 조상이 출현한다. 코끼리, 말 둥도 이때 진화하여 현대의 것과 같이 된다. 제 4기는 언류픽 시대가 시작된다. 거대 포유류는 사라지고 인류가 번성하기 시착하며 현세에 오면 언류가 진화하여 현대의 언류와 같이 된다.표 3.2 시대와 동물의 변화
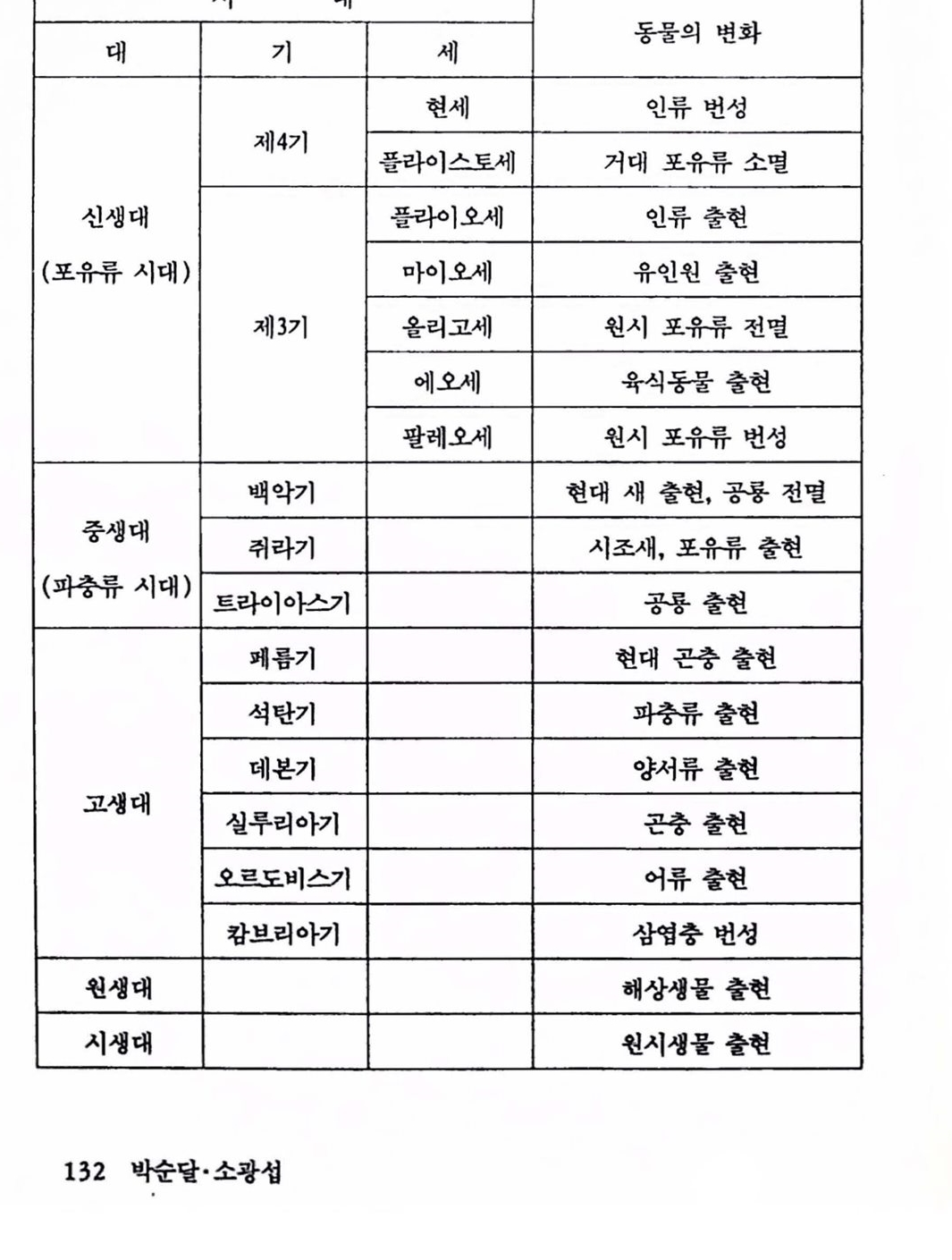 시 대
시 대
1.3. 생명의 진화 생명의 출현에는 여러 가지 가설이 있다. 그러나 지금은 물철에서부터 차차 생명체로 진화하였다는 진화설이 지배척이다. 그러면 생명체는 어떤 환경에서 어떻게 생명체로 진화되어왔는가?
(1) 지구의 상황 앞에서 우주의 생성은 대폭발에서 시작한다고 하였다. 이 대폭발에서 시작하여 물질이 생성되기 시작한다. 그 이전에는 물철이라고 할 수 없는 상태에 있다가 대폭발이 시작되면서 처음에는 전자, 광자 등 소렵자가 생성되고 다음에 헬륨, 수소 등이 생성된다. 이렇게 차례로 여러가지 물질이 생성되는데 물질은 언제 한꺼번에 모두 만들어진 것이 아니고 이 시간에도 새로운 물질이 나타나고 있을지 모르며 앞으로도 얼마든지 새로운 물질이 생성될 수 있다. 대폭발과 더불어 물질이 생성되고 이련 물질이 모여 별이 되고 지구도 되었다. 지구는 앞에서 언급한 바와 같이 45억 년 전에 형성되었는데 지구가 형성되고 활발한 화산활동을 거쳐 물도 생기고 대기가 생기면서 생명이 생성될 수 있는 환경이 만들어져 갔다. 그러면 어떤 환경에서 최초의 생명이 형성되었는가? 물 • 대기의 생성 : 물은 생명을 유지시격 주는 대단히 중요한 물철이다. 물은 화산활동을 통해 지구 내부에서 분출되어 나왔다고 보고 었다. 화산이 분출할 때는 막대한 가스를 분출시키는데 이 기스 충에는 수분이 많은 부분을 차지하고 있다. 이 수분이 모여 강물이 되었다. 그리고 지표면의 수분은 중발하여 하늘에 머물다가 다시 비가 되어 지표에 떨어지는 순환을 겪고 있다. 한편 강의 물은 바다로 흩러가는데 이혜 지상의 소금은 녹아 바다쩨 쌓이제 된다. 소금은 물 속에 녹아 오랫동안 있올 수 있기 빼폼애 혜수는 자연히 염도가 높은 상태를 유지하게 된다.
한편 대기는 어떻게 생성되었는가? 대기는 화산활동을 통해 지구 내부에서 분출되어 나왔다는 배기설이 있다. 화산 가스는 95% 이상이 수증기이고 그 밖에 H2S,CO2, H2, N2 등이 있다. 다음은 광분해설로 초기에는 메탄(CH4), 암모니아(NH3) 가스 등이 지구를 채우고 있었을 것이라는 가정 아래 이들 기스가 태양의 자외선에 의해 분헤되어 H2, O2, CO2 등이 생성되었다는 셜이다. 그러나 이 셜은 초기에 과연 메탄과 암모니아가 많았을까 하는 것이 확설치 않기 때문에 무리가 있다. 지구를 둘러싸고 있는 대기 중에 생명유지에 없어서는 안 되는 것이 산소인데 이 산소는 어떻게 생겨나게 되었는가? 보통 이 산소는 식물의 광합성으로 생성된다고 보고 있다. 남조류는 광합성을 하는데 이 광합성의 결과로 산소가 서서히 대기층에 쌓여 갔을 것이라는 것이다. 그리고 육상식물이 번성하면서 산소의 양은 더욱 증가하였을 것이라고 보고 있다. (2) 생명의 기원 지구상에 생명체가 최초에 어떻게 생겨나게 되었는지에 대해서는 여러가지 가셜이 있다. 창조셜, 천체버래셜, 자연발생설, 진화젤이 바로 그것 이다. 창조셜은 생명체는 창조주에 의해 창조되었다는 가젤로서 신학( 學)이 발달하면서 융성하다가 르네상스 이후 자연과학의 발달로 점차 쇠퇴하고 있다. 그러나 아직도 종펴의 믿음과 연관되어 사라지지 않고 있으며 인간은 신에 의해 창조되었다는 변형된 창조셜이 남아 있다. 천체버래셜은 생명체는 지구 이외에 다른 멸에서 날아왔다는 가절이다. 우주는 1,000억 개의 성운이 있는데 이틀 성운 충의 하나가 우리 지구가 속해 있는 은하계이고 이 은하계에는 1천 500억 개의 별들로 이루어져 있다. 이틀 별 중 20억 개 청도가 우리 지구와 같이 생명체가 살 수 있는 상태의 별들로 보고 있다. 이렇다면 이들 별들 중 생명체가 목을자른플라스크 목을 자르지 않은 플라스크
목을자른플라스크 목을 자르지 않은 플라스크
그림 3.5 파스퇴르의 실험
살 수 있을지 포르고 이 생명체가 지구로 버래했을 가농성도 배제하기 힘들다. 그러나 현실성이 적어 별로 주의를 받지 못하고 있다.
자연발생셜은 18세기까지 가장 널리 받아들여쳤던 가설이다. 16세기 벨기에 의사인 헬몽(Jan B. Van Helmont, 1713~1644)은 땅에 젖은 더러운 셔츠에 밀을 함께 두연 쥐가 생긴다고 했다. 영국 박물학자 니담(John T. Needham, 1713~1781)은 1745년에 끓인 고기좁을 유리병에 넣고 밀봉한 후 따뭇하게 해주었더니 미생물이 생검을 확언했다. 이것은 유기물 속에는 생명력이 있어 이것이 자연발생의 훤동력이 된다는 것 이다. 그러나 이런 자연발생설도 곧 도전을 받기 시작히는데 이랄리아의 레디(Francesco Redi, 1626~1698)는 1668년 교깃조각올 담은 그릇에 뚜껑을 닫아 두면 고기가 썩더라도 구더기가 생기지 않음올 알었다. 자연발생설을 부르짖던 사람에게 가장 치명척인 공격융 가한 사랑온 프랑스의 픽스퇴르(Louis Pasteur, 1822~1895)였다. 그는 대기 충애는 여러 종류의 미쟁풀이 있으며 이들 미생풀이 없이는 고기좁도 썩지 않는다는 사실을 1826년 실험으로 중명하였던 것이다.그는 플라스크에 설탕물과 효모를 혼합한 용액을 충분히 끓인 후 주퉁이를 S자로 길게 뽑아내어 물을 지나가게 해두었다. 그리고 다른 것은 주둥어를 짧게 하고 막지 않고 두었다. 그랬더니 S자로 길게 물을 거치게 한 것은 미생물이 발생하지 않았고 다른 것은 발생하였다. 이 실험으로 자연발생셜은 부인되고 이 공로로 그는 정부로부터 상금을 받기도 하였다.
진화셜은 기존 생물에서 다른 종이 갈라져 나와 그중에 척자가 생존해 남는다는 가젤로서 1858년 다윈이 제창한 이래 가장 널리 받아들여지고 있는 가설이다. 그러나 이 진화설도 이 지구상에 최초의 생명체가 어떻게 발생하게 되었는지는 말해주지 못하고 있다. 그래서 이 부붕은 오히려 자연발생설이 그럴 듯하다. 즉 최초의 생명체는 간단한 물질이 점차 복잡한 물질로 형성되어 오다가 드디어 생명체가 나타나고 생명체들은 진화를 통해 종(種)이 파생하고 점차 여러 가지 생명체가 나타나게 되 었다. (3) 생물진화 18세기기까지만 해도 음식이 왜 상하며 병이 어떻게 일어나며 눈에 보이지 않는 생명체가 있는지를 몰랐다. 지구상에는 50여만 종의 식물과 100여 종의 동물이 있는데 대단히 다양한 종류의 생명체가 서식하고 있다. 지금도 새로운 생명체가 형성되어 가고 있고 또는 사라져 가고 있다. 생명체의 다양성과 그의 계통을 이해하지 못하고서는 생물의 진화률 이해하기 힘들다. 가시척인 대형통물, 즉 코끼리, 원숭이, 언류만 가지고 사랑의 조상어 원숭이라고 흩땐 납득하기 힘들기 때문이다. 생명체의 다양성 : 1652년에 레벤후크(Antoni van Leeuwenhoek, 1632~1723)는 렌즈를 통해 최초로 미생물올 발견하였다. 보이지 않는 생명째가 었다는 것을 알었던 것이다. 그리고 1862년 파스퇴르는 발효, 부패는 이플 미생물이 훤언임율 밝혔던 것이다. 미생물이 발견된 후1876년 코흐(Robert Koch, 1843~1910)는 탄저균, 결핵균, 콜레라균 등의 박테리아를 발견하였다. 이 박테리아는 크기가 1mm의 1,000분의 1좀 되는 생명체로 미생물에 버헤 100배 작은 것이다. 그리고 이 박테리아보다 1,000배 작은 바이러스가 1935년에 발견되었다. 1898 년여과지에도 결러지지 않는 원인 모를 담배모자이크병의 병원균을 베이제랭크(Martinus W. Beijerinck, 1851~1931)는 바이러스라고 이름지었는데 이것을 스탠리(Wendell M. Stanley, 1904~1971)가 1935년 결정화하는데 성공하였다. 이 바이러스는 환경이 나뿔 때는 보통 물젤과 같이 결정 상태에 있다가 생체와 같은 좋은 환경에 오면 생명체로 살아나는 물질과 생명체의 중간쯤 되는 것이라 할 수 있다.
오따린의 가설 : 오파련(A. I. Oparin, 1894~1980)은 1924년 그의 저서 『지구상의 생명의 기원』에서 원시 지구에서 간단한 무기물이 좀더 복잡한 유기물이 되고 유기물이 더욱 복잡한 생명체로 발전했다는 가절을 세웠다. 앞에서 서술되었듯이 지구는 45억 년 전에 생성되어 격성한 화산활통이 진행되다가 차차 식게 되고 그동안 물이 생기고 대기가 형성되었다. 그리고 풍부한 태양 에너지의 힘이 작용하여 새로운 물질이 만들어져 갔다. 지구 표면에는 점차 풍부한 수증기, 메탄, 암모니아 등이 생성되었고 이련 물질이 좀더 복잡한 물질로 변해 갔다. 그러면 이러한 변환과정이 지구상에서 실제로 일어났는지를 어떻게 실중할 수 있는가? 메탄, 암모니아 가스 등은 다른 행성에도 발견되기 때문에 훤시 지구에서도 존채하였을 것이라고 생각할 수 있다. 그리고 수충기, 수소 풍은 풍부하였을 것이고 태양 에너지도 지금과 같이 풍부하였융 것으로 쟁각 할 수 있다. 이련 가정 아래 멀러(Stanley L. Miller, 1935~ )는 유명한 실험을 수행하였다. 그는 5리터들이의 플라스크에 천기방전이 얼어날 수 있게 천극을 넣고 그 안에 수중기, 메탄, 암모니악 수소훌 채우고 전기방전을 일으키면서 일주일울 지면 결과 그 안에 여러 가지외 ~
~
그림 3.6 밀러의 실험
아미노산, 지방산, 그 외 여러 가지 유기물이 생성되어 있음을 발견하였다. 이 멀러의 실험은 간단한 유기물에서 원시 지구의 상태에서 생명체를 구성하는 물질이 생성될 수 있다는 것올 업중한 셈이 된다. 이 설험 이후 생명체는 자연조건 아래에서 복잡한 물질이 만들어지면서 형성되었다고 하는 오파련의 가설이 더욱 현실척인 가젤로 받아들여지게 되었다. 그래서 간단한 유기물이 당, 아미노산, 지방산 동의 유기물올 형성시키고 이들 유기물이 단백철, 핵산, 효소 둥 생명체에 직결되는 물질로 발전하고 이들 물질로부터 최초의 간단한 생명체가 생겼을 것이라고 믿게 되었다.
화학진화 : 현재 지구상에는 500여만 종의 물질이 있는데 이련 물철올 구폼할 때 무기물, 유기물로 나누는 방법이 있다. 이것은 생명체를 구성히는 물질과 비슷한 것은 유기물로, 아닌 것은 무기물로 나누는 것인데 유기물은 수소, 산소, 탄소, 질소 등을 포함하고 있는 것으로서 메탄, 암모니아 등은 유기물이다. 19세기초만 해도 베르첼리우스(John J. Berzelius, 1779~1848)는 이원셜을 주장하여 무기물과 유기물은 상호 변환될 수 없는 것으로 생각하고 있었다. 그러나 그의 제자 볼러(Friedrich Wohler, 1800~1882)는 무기물인 시안산 암모늄으로부터 유기물인 요소를 생성해 내었다. 그래서 생명체에 관련이 깊은 유기물과 그렇지 않은 무기물 사이에는 벽이 없어지게 되었고 단지 편이상 구분히는 것에 지나지 않게 되었다. 이것으로 지구상의 생명체가 무기물에서 생성되었다고 말할 수는 없으나 적어도 장벽이 없어진 생이다. 왜냐하면 만일 무기물과 유기물이 서로 변환될 수 없다면 무기물에서 유기물이 생성될 수 없기 때문에 어떻게 생명체가 간단한 물절로부터 형성될 수 있겠는가? 이런 점에서 볼러의 업척은 생명체의 발생에 커다란 공척을 쌓았다고 할수있다.
진화젤 : 지구상의 생존하고 있는 생물틀은 기존의 생물로부터 분화하여 환경에 적웅하는 종이 살아남아 있는 것이라는 가성이 진화설이다. 이것은 다윈(Chales R. Darwin, 1822~1884)에 의해 확고한 기반올 다지게 된 것으로서 이 이전에는 이에 관련되는 학설로서 라마르크(J. Lamarck, 1744~1829)의 용불용설이 었었다. 라마르크는『동물철학』 (1809), 『무척추동물지』(1815)풍 의 저서를 통해 · 생물은 폼을 키울 수 있는 데까지 키운다. · 새로운 기관은 필요할 혜 형성된다. · 기판은 사용 정도에 따라 달라진다. · 일생 동안 획득된 형철은 지손에 전해진다. 로 요약되는 용불용젤을 제창하였다. 예훌 들어 기련이 목어 긴 것용높은 나무의 잎을 먹기 위하여 걸어졌다는 것이다. 그러나 독일의 바이 즈만(A. F. L. Weismann)은 쥐의 꼬리를 22대나 잘랐으나 꼬리가 없어지지 않는 것을 보고 용촬용설을 부정하였다.
다윈은 1831년에 해양측량을 위해 떠나는 영국 해군함정 비글(Beagle)호에 박물학자로 숭선하여 5년 동안 세계일주를 하면서 조사하였다. 특히 그는 태평양 쪽에 위치한 남미대륙에서 종 떨어진 갈라파고스 군도에서 흥미있는 현상을 발견하였다. 이 갈라파고스 섬에서는 거북이, 참새의 일종인 핀치새가 다른 곳의 것과는 다른 특성을 가지고 있다는 것에 특히 관심을 가졌다. 이 5년 동안의 조사와 관측의 결괴를 연멀히 검토한 결과를 1859년에『종의 기원』이라는 책으로 발간하였다. 이 책에서 그는 〈생물은 서서히 형철에 변화가 나타난다. 이 변화된 종은 환경에 척웅하연 생존하고 그렇지 못하면 사라지는데 이런 현상을 적자생존의 현척 또는 자연선돼이라고 한다〉라고 하였다. 그는 이런 현상을 많은 자료를 토대로 업증하여 여태까지의 어느 설보다도 신벙성이 있는 설로 업증시켰던 것이다. 그러나 다윈은 그 형철에 변화가 일어나는 과정은 셜명하지 못했다. 그 후드 브리스(Hugo de Vries, 1901~1963)는 달맞이꽃의 변이를 발견하고 돌연변이가 종의 발생의 기원이라고 주장하였다. 그리고 률러(Herman J. Muller, 1890~1967)는 초파리에 X선을 조사시켜 눈빛, 날개모양울 변형시켜 인위척인 돌연변이를 유도할 수 있었다. 이런 돌연변이셜과 멘멜 (Johann G.M endel, 1822~1844)이 1865년에 주장한 형질은 부모의 형질올 따르교 그것이 어떻게 유천되는지 셜명하는 유전법칙과 결부되어 현대의 생물기원을 셜명하고 있다. 즉 생물은 화학진화률 통해 생명체가 생기고 돌연변이률 통해 새로운 종이 생겨나며 형철은 유전올 흥해 이어진다. 그리고 종들은 자연선택에 따라 척자가 생존하게 된다. 이들 척자 속에는 몇 백만년 전부터 지구상에서 가창 번성하는 총류 중에 하나언 언류도 포함되어 있다.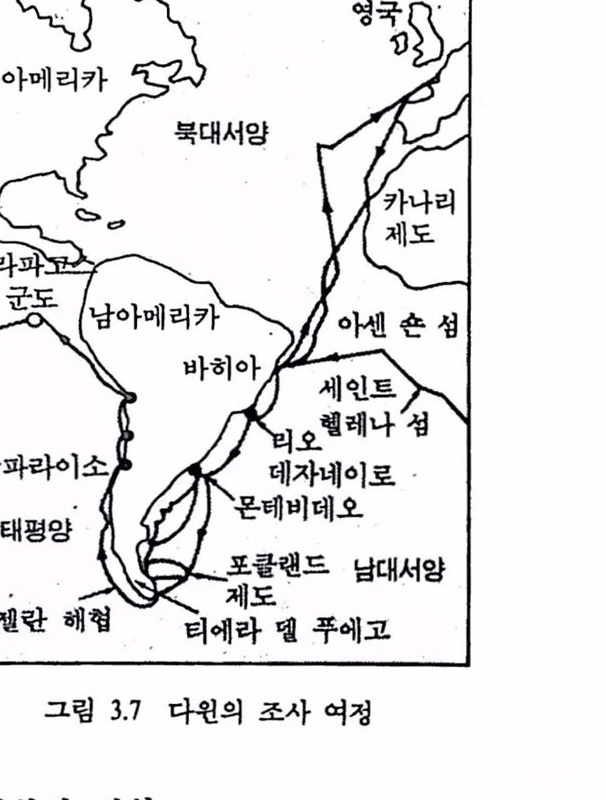 발파라이소
발파라이소
그림 3.7 다윈의 조사 여정
2. 뇌의 자연진화와 역할 2.1. 생물진화와 뇌 확장 신경계의 진화 : 우리 몽의 운동을 관리 · 통제하고 갱신활풍율 판창하는 콧이 신경계이다. 신경계는 크게 중추신경계와 말초신경계로 나누어지는데, 중추신경계는 뇌와 척수( , spinal cord)로 이루어지는폐, 척수는 말초신경계로 이어져 었으면서 이 말초신경에 요교가는 신호의 홍로가 된다. 말초신경제는 몸의 구적구석에 퍼져 외부로부터의 지극율 수용하거나 중추신경계로부터의 명령올 천탈하는 역활융 한다.
신경계는 동물에만 있고, 생기게 된다. 식물에는 없다. 그러면 신경계의 기능은 무엇인가? 기본적으로 외부의 자극을 인지 · 수용하여 이 자극에 대해 반응하는 기능이다. 사람은 더우면 옷을 벗고 비가 오면 집안으로 들어가서 비를 피할 줄 안다. 이것이 모두 신경계의 작용이다.
간단한 동작은 신경계가 없어도 할 수 있다. 우선 운동은 어느 물질이나 할 수 있다. 공기 속의 먼지는 자유롭게 떠다니는데, 그 운동형태는 일정하지 않다. 먼지는 공기 속을 떠다니다가 다른 물질에 부딪치면 다른 콧으로 가기 때문에 운동이 일정치 않다. 이것을 브리운 운동이라 한다. 짚신벌레는 짚신 모양을 한 섬모충류의 동물인데 이 벌레는 신경계를 가지고 있지 않다. 짚신벌레는 물에서 유영을 하다가 바위와 같은 장애물에 부딪치면 잠깐 물러나 있다가 몸을 회전하여 다른 방향으후 전진한다. 이런 짚신벌레의 행위는 장애물이 바위이기에 회피운동을 한다기보다는 전진하는 방향에 어떤 장애물이 있더라도 그 종류에 상관없이 기계척으로 회피반동을 한다. 그러나 통물이 진화하면서 점차 특수하게 분화된 신경계가 나타나기 시작한다. 처음에는 단순한 형태가, 그리고 점차 다양하고 복잡한 형태가 나타난다. 하등동물인 말미찰에는 신경세포는 없으나, 표피층의 상피세포가 분화되어 외부자극을 받아들이는 수용기 세포 역할을 하는 세포가 있다. 이 원시척인 수용기 세포에 수축성 근육세포가 이어져 외부 자극에 반응한다. 이런 형태가 점차 발달하여 수용기 세포와 근육 사이에 자극을 전달하는 전문화된 신경세포가 나타난다. 해파리는 표피충파 근육 사이에 신경세포가 있고 이들 신경세포가 서로 연결되어 멍을 이루고 있다. 신경세포는 대단히 다양하다. 신경세포는 수상돌기, 축색으후 다른 신갱계와 연결하는데, 이 연결망으로 신경세포 장호칸에 신호를 교환한다. 그런데 이 신경계는 진회과청에서 신경철( , ganglion)이 생기는데, 이것은 신경세포의 접충으로 신경세포 상환의 정보교환을 통합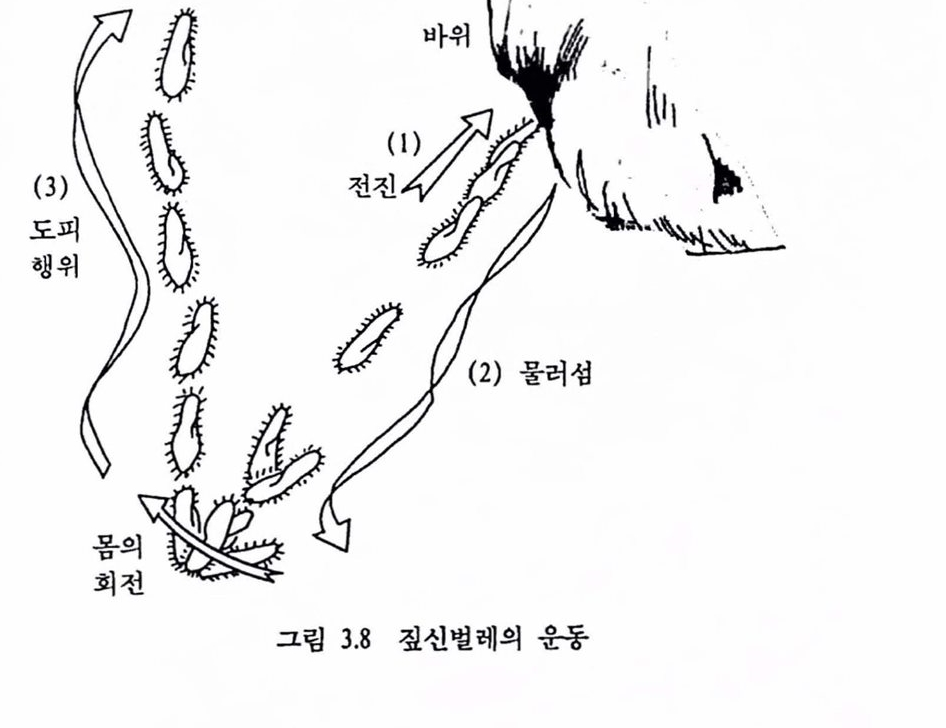 \{..,..'.\~ . 、현싸썩」 나
\{..,..'.\~ . 、현싸썩」 나
그림 3.8 짚신벌레의 운동
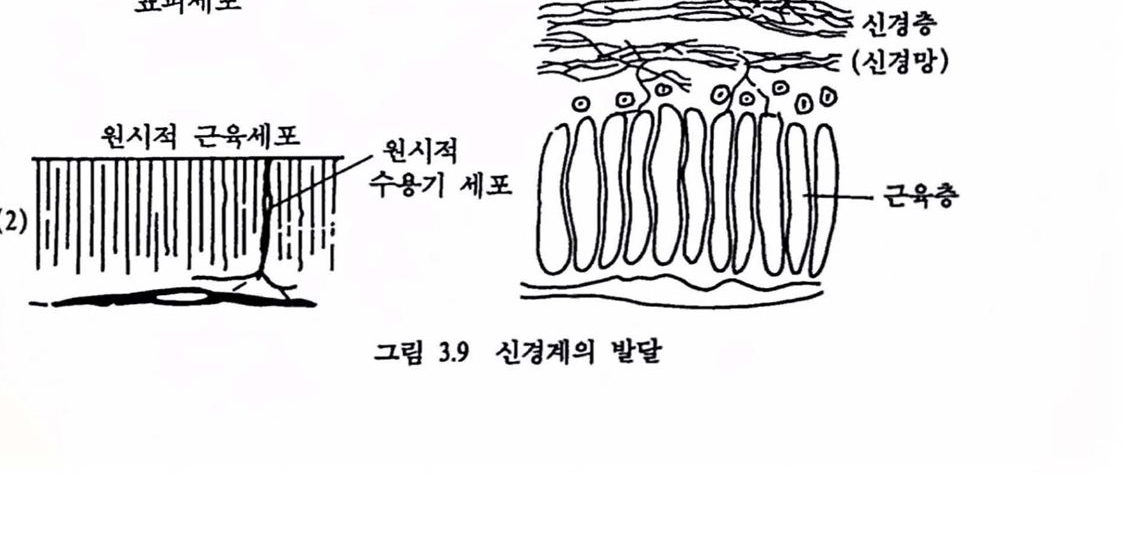 、-- ’’ 홍분성 표펴세포 tI! ”
、-- ’’ 홍분성 표펴세포 tI! ”
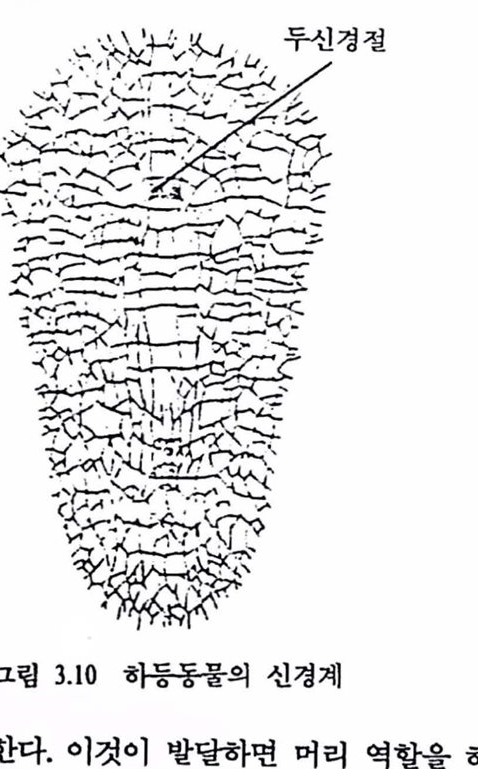
그림 3.10 하등동물의 신경계
하고 조철하는 역할올 한다. 이것이 발달하면 머리 역할을 하는 두신경절이 생격난다.
뇌의 확대 : 생물의 진화와 뇌의 발달은 대단히 깊은 판계가 었다. 생물이 동풀, 언류로 진화하면서 뇌의 역할이 생존에 점점 깊은 영향올 미치기 시작했다. 생물이 동물로 진화하면서 뇌를 가지기 시작하는데, 이로써 환경율 좀더 청확히 감지하여 대웅천략을 세우기 좋게 되었다. 식물이 환갱감지 농력이 없는 것은 아니지만, 동물윤 정차 고퉁동물로 진화할수륙 환경올 감지하는 능력과 함께 감지된 청보를 처리할 수 있는 농력, 많은 청보률 기억하는 놓력, 좋은 것과 싫은 것을 구별하는 능력이 벌할되어 왔다. 더구나 생물이 움칙이기 시작하면서 움칙임을 판창하는 뇌의 역할은 첨청 커져 겼다. 진화할수록 움직임은 점점 청멀해지고 이빼 상용하는 청멀한 웅칙엄울 가농케 하는 뇌가 발랄해야 한다. 그래서 고등동물에 이르면 허등동물에 버해 대단히 큰 뇌를 가지게 된다. 몇 가지 예를보면 사자 258g 물개 442g 말 620g 해마 1.126g 등이다. 더욱이 원숭이나 사람과 같이 손을 정밀하게 사용하려면 손의 복잡하고 미세한 움직임을 관장하는 뇌의 발달이 선행되어야 한다. 그리고 예측한다든지 수를 센다든지 하는 고도의 정신작용이 필요하게 되어 뇌가 더욱 발달혜 간다. 특히 사람의 뇌는 다른 어느 동물에 비하여 큰데 이것은 인간이 직럽을 시작하면서 손이 자유로워져서 고도의 운동조절 능력이 필요하게 되고, 또한 영장류가 나타나면서 계속척으로 발달해 온 고도의 정신활동을 위해 뇌의 용적과 함께 뇌의 형태도 발달해왔다. 침팬지의 뇌가 400cc언 데에 비하여 사람의 뇌는 1,20Occ에 가깝다. 다른 동풀은 몸이 크더라도 뇌의 용척은 작다. 뇌용척은 표에서 보듯이 진화에 따라서 점점 커짐을 알 수 있다.
표 3.3 뇌 용척
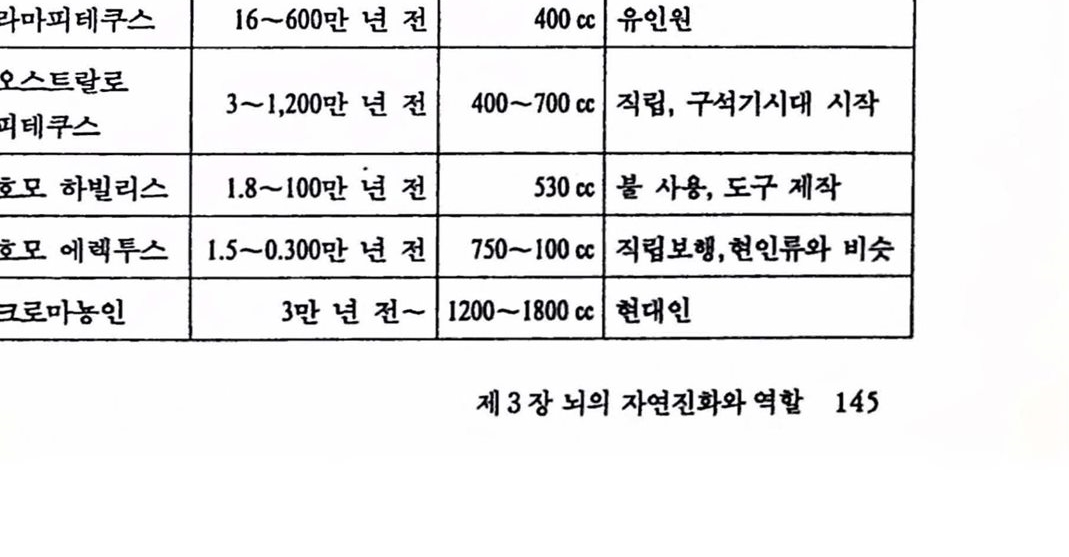 π검 생존기간 뇌 용척 버고
π검 생존기간 뇌 용척 버고
인간의 뇌와 그 기능 : 인간의 뇌의 크기는 조금씩 다르지만, 보통 1,200cc 내지 1,800cc쯤 된다. 무게는 보통 1.4kg 정도로 폼 전체 무게의 2% 정도이다. 그러나 뇌는 다른 장기에 비하여 많은 에너지를 소비하는데, 몸 전체의 20%의 산소를 소비하고 포도당은 25%를 소비한다.
뇌는 대략 1조 개의 세포로 이루어져 있는데, 그 중 140억 개는 신경세포이고 나머지는 부드럽고 첼리같이 생긴 구조로서 신경세포에 영양을 공급하는 등 신경세포의 활동을 보조한다. 신경세포에는 수백에서 수천의 수상돌기가 있어서 이들 수상들기에 의해 다른 세포와 연결되어 정보를 주고받는다. 체내외의 변화는 말초신경에서 감지되고 이것이 전기적인 신호로 변환되어 척수를 통해 뇌로 전달된다. 그러면 뇌는 이 자극을 무시해버리라든지 아니면 적절한 조치를 취하게 된다. 뇌는 우리의 폼을 관리 통제하는 임무를 가진 콧이다. 뇌는 해부학적으로 볼 때 전뇌(前腦, forebrain), 중뇌(中腦, midbrain), 후뇌(後腦, hindbrain)로 구성되어 있는데, 전뇌는 대뇌 및 간뇌로 구성되어 있고 후뇌는 소뇌와 교 및 연수로 구성되어 있다. r---대 뇌 (大腦, cereburm)
r---대 뇌 (大腦, cereburm)
대뇌는 뇌 부피의 80~90%를 차지하는 곳으로 확뇌와 우뇌로 구분되는데, 이 좌 · 우뇌는 뇌량(腦, corpus callosum)으로 연결되어 었다. 이 좌 · 우뇌는 운등증추와 지각중추를 고루 갖추고 있으며, 화 대뇌 반구는 몸의 우측 반을 지배하고, 우 대뇌반구는 옴의 화측 반을 지배한다. 뇌량은 신경섬유로 이루어져 었으며 좌 · 우측 반구에 정보를 교환한다. 이 뇌량에 이상이 생기면 폼의 화우 활동이 균형을 잃게 된다.
대뇌는 표면쪽에 회색벚을 띠는 부분과 안쪽의 흰색을 띠는 부분으로 구성되어 있는데 회색빛을 띠는 부분을 대뇌피철이라 하고 흰색 부부혹 대뇌수질이라고 한다. 대뇌피철은 주로 신경세포로 이루어져 있고 각종 중추가 모여 있는 콧이다. 이곳은 표면적을 넓히기 위하여 주름이 잡혀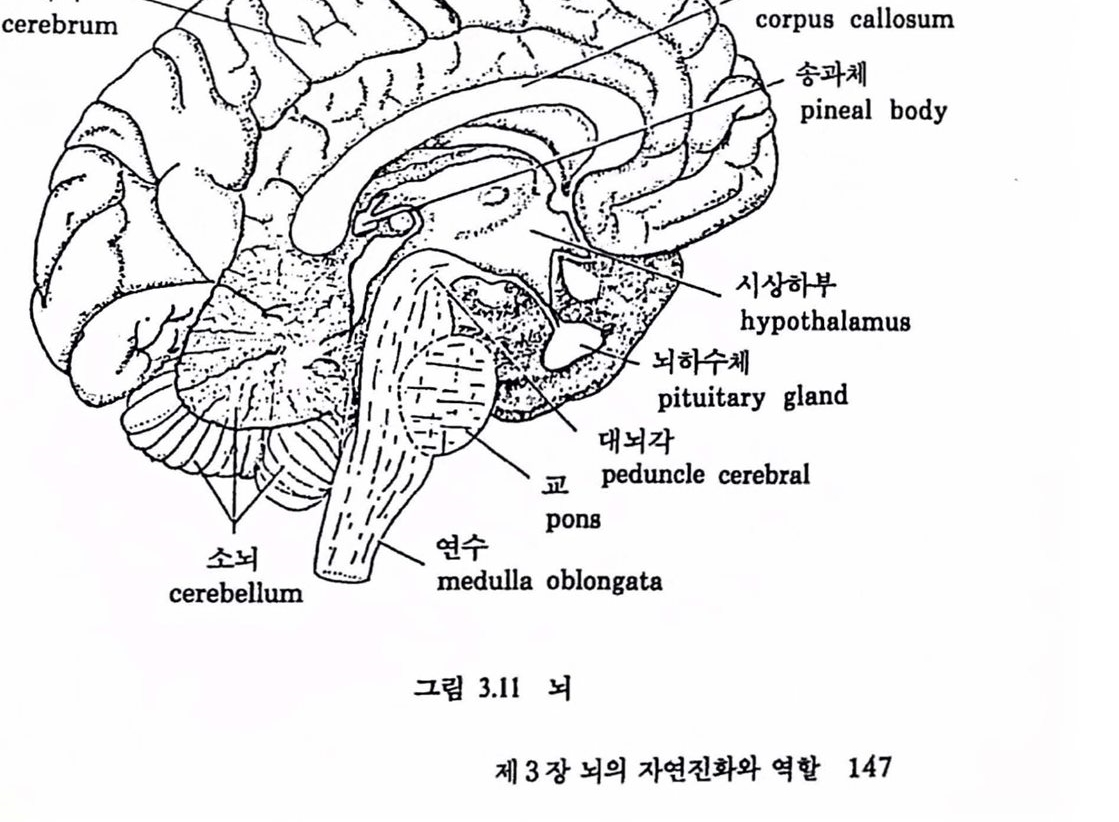
그림 3.11 뇌
었다. 대뇌수질은 신경세포가 모여 있는 콧도 있으나, 주로 신경섬유로 구성되어 있다. 이 부분은 대뇌중추들과 뇌간을 연결하기도 하고, 대뇌에 있는 두 지점을 연결하는 역할을 한다.
간뇌는 뇌량의 아래에 위치하고 있는데 시상(規庫, thalamus)과 시상하부(規庫下部, hypothalamus)로 구성되어 었다. 시상은 회백질로 되어있는데, 후각을 제외한 모든 감각자극을 중계하고 통괄한다. 후각은 대뇌 변연계에서 직접 관장한다. 시상하부는 시상 바로 밑에 위치하여 주로 자융신경을 조절하며 외부에서 오는 정보를 대뇌피철에 전달하는 역할을 한다. 중뇌는 4개의 신경핵 뭉치로 된 사구체와 양쪽 대뇌반구에서 나온 기둥 모양의 대뇌각드로 이루어져 있는데 전뇌와 후뇌를 잇는 중요 경로로서 운동신경을 조절한다. 예로서 눈 동작은 중뇌에서 조정된다. 소뇌는 대뇌 아래 중뇌 후면에 위치하고 있다. 대뇌와 비슷하게 표면은 회색이고 내부는 흰색질로 되어 있고 대뇌의 8분의 1 크기이다. 소뇌는 운동을 칙접 수행하지는 않으나 명형 기능을 가지며 통합 조정으로 대뇌의 운등증추를 돕고 있다. 교는 중뇌와 연수 사이에 있으며, 연수와 대뇌피질 사이의 연락장소이다. 연수는 뇌의 마지막 부분으로 척수로 이어진다. 이콧에는 심창박동, 호홉운통, 재채기, 하품, 기침 등의 반사중추가 모여 있는 곳이다. 2.2. 뇌의 진화 동물의 진화에 있어서 가창 두드러진 변화는 대뇌에서 일어난다. 그렴3.12에서 보는 바와 갈이 그 크기에서 우천 변화가 있지만 기능도 대단혀 다양빼진다. 언휴픽 뇌는 3충 구초로 되어 있는데, 이것은 진화과정에서 세 단계를 거쳐서 진화되었다는 것을 의며한다. 즉 3충 구조의 뇌란 뇌
뇌
뇌
첨팬지 언간1총 뇌 : 파충류의 뇌(뇌간) 2충 뇌 : 포유류의 뇌(대뇌변연계) 3층 뇌 : 인류의 뇌(신피철) 을 말한다.
1충 뇌인 뇌간(腦幹, brain stem)은 연수, 교, 중뇌, 간뇌로 구성되어 있는데, 진화과정에서 가창 일찍부터 발달한 부분이다.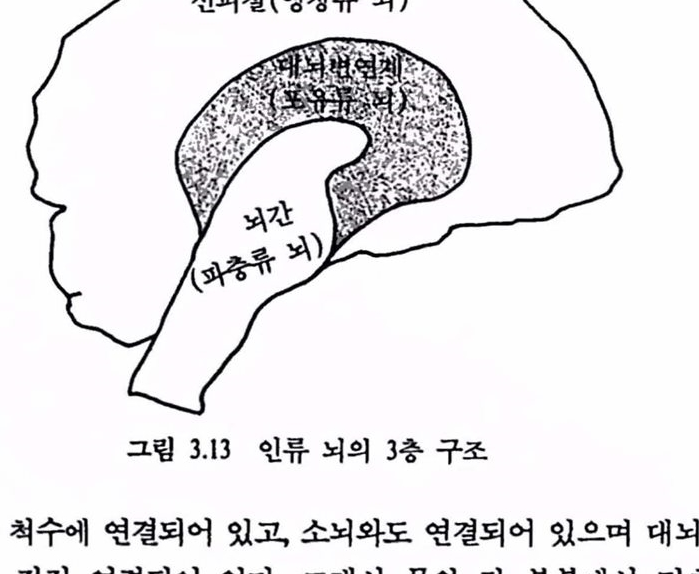
그림 3.13 인류 뇌의 3충 구조
뇌간은 척수에 연결되어 있고, 소뇌와도 연결되어 있으며 대뇌의 여러 부환과도 칙접 연결되어 있다. 그래서 몸의 각 부분에서 말초신경을 홍해서 감지되는 각종 청보가 전기척인 신호로 천환되어 척수에 천달되연 이것이 뇌간으혹 들어오게 된다. 또한 뇌의 여러 부분에서 일어나는 지극도 뇌간에 천달된다. 그러면 이 자극 이 대뇌에 전달되어 처리된 후 다시 기저핵( ,basal ganglia)올 통해 운동신경으로 전달되어 언뼈의 활동으혹 변환된다.
뇌의 기농이 모두 칼은 우게로 중요한 것은 아니다. 가창 중요한 기능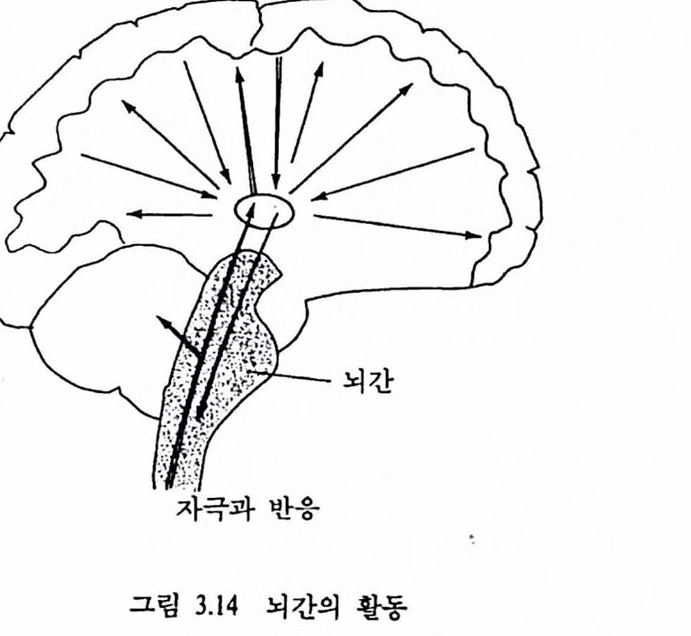
그림 3.14 뇌간의 활동
을 수행하는 부분이 뇌간이다. 뇌간은 심장박동, 혈압, 호홉, 의식의 수준을 판창한다. 이툴은 생명을 유지히는 가창 기본척인 기능으로 자율신경을 통해 심장박동, 혈압, 호홉 등을 통제한다. 나아가 뇌간올 통한 자극이 중뇌를 통해 대뇌에 들어가면, 대뇌피질에서 이 자극이 처리되어 어떤 부분의 세포를 활성화시키는데 이것이 바로 의식의 수준이다. 이 수주유 뇌간이 각 신경에서 들어오는 자극을 어떻게 처리하느냐에 달렸다. 뇌간은 끊엄없이 들어오는 지극을 검사하여 처리한다. 그래서 몇 섭만 개의 자극 중에서 중요한 몇 백 개 정도만을 선택하여 대뇌에 천랄한다. 이것이 바로 혈압을 올련다든가 심장박동올 높언다든가 하는 갯으로 나타난다.
뇌간은 이러한 생명유지에 기본척인 기능뿔만 아니라 하풍, 채채기와 같은 반사운동올 관리하며 위, 업, 얼굴, 눈을 통제한다. 위액융 분버하거나 음식을 삼키고 얼굴 표정과 눈의 움칙임올 변화시키는 풍의 반용이 모두 이 뇌간에서 통제된다. 뇌간의 상부에는 중뇌가 있다. 충뇌의 기놓중 하나는 눈의 운동, 동공의 크기 등을 조절하는 것이다.
이 뇌간은 의식이 있을 때나 자고 있을 때나 제기능을 발휘하는 생명의 파수꾼이다. 뇌간은 뇌의 모든 부분이 그렇듯이 소뇌 및 대뇌와 연결 되어서 자극을 주고받는다. 그래서 의식이 있을 때는 대뇌의 영향을 받는다. 그러나 일단 의식이 없어지면 독자척으로 활동하며 이 뇌간이 쉬게 되면 곧 언체가 기능을 잃게 된다. 2충 뇌언 대뇌변연계(大腦邊緣系, limbic system)는 측두엽 안쪽 갚숙히 간뇌 지역에 위치한 복잡한 구조로서 주로 감정, 기억, 학습을 관리하는 부분이다. 이 대뇌변연계는 젖먹이동물(포유류)의 대부분이 같은 구조를 가지는 원시척인 구조로서, 뇌간이 인체의 식물척 기능을 관장한다면 이것은 인체의 동물적 기능을 수행한다. 뇌의 진화과정에 있어서 전술한 바와 같이 대뇌가 점차 확대되어 가는데, 그 단계에 따라 고피철(古皮質, archicortex), 구피철( 皮質, palaeocortex), 신피질(新皮質, neocortex)이라고 부르며 신피질이 발달할 때 고 · 구피질과 신피질을 연결하는 구조가 발달한다. 이 고 · 구피철파 연결하는 구조를 합쳐서 대뇌변연계라고 한다. 또는 대뇌변연계피철(limbic cortex)이라고도 한다. 대뇌변연계는 해마 (hippocampus)
해마 (hippocampus)
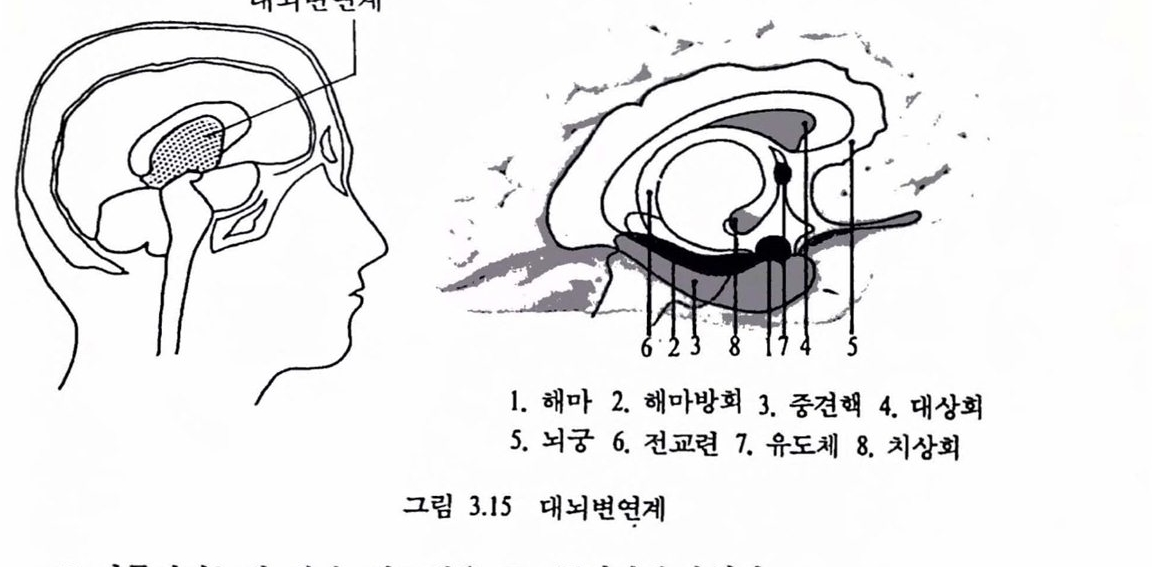 대뇌변연계
대뇌변연계
그림 3.15 대뇌변연계
로 이루어지는데 해마, 면도핵은 고 • 구피절에 속한다.
이 대뇌변연계의 각 부분이 어떤 기능을 가지며 서로 어떻게 작용하는지는 정확하게 알려져 있지 않으며 부분적으로만 알려져 었다. 현도빽은 주로 공격과 관련이 있고, 해마는 기억과 관련이 있으며, 중격빽은 쾌감과 관련이 었다. 그리고 대상회, 뇌궁, 전교련은 다른 부분과의 연결 역할을 하며 포한 기억에도 관계하고 있다. 그래서 면도핵, 해마, 충격뼈의 복합적인 전기척 자극의 복합척인 결과로 노여움, 걱청, 홈훈 동의 감청이 나타난다. 대뇌변연계의 가창 중요한 기능의 하나가 감청올 홍한 신체 얀용인데 여기에는 해마의 역할이 크다. 이 반옹은 현재의 자극과 과거의 갱헝융 비교하면서 취해지는 것으후 생각되고 있다. 현채의 자극어 변하지 않고 경험에 비추어 버슷하면 해마가 활동척이 된다. 반면에 자극에 변화가 쟁기고 경험에 버추어 불확실하면 해마는 활동할 수가 없제 되고 그러면 뇌간이 흥분되고 뇌간은 다른 푸위에 자극올 보내어 경계 태째훌 취하도록 한다. 이런 점에서 해마는 끊임없이 들어오는 자극에 대해 어떤 외미를 부여할 것언지률 결청하는 데 중요한 역할융 한다.이 문제는 기억과도 관련이 었다. 만일 조금 전에 일어난 일을 기억하지 못하면 또다시 같은 일이 일어났을 때 서로 비교할 경험이 없는 것과 같아진다. 그래서 해마에 이상이 생기면 집중력이 약해지고 관계없는 일에 신경을 쓰게 된다.
대뇌변연계의 또 하나의 중요한 기능은 학습이다. 이 학습은 상멸을 통해 일어나게 되는데 상별로써 계속할 것과 계속하지 않을 것을 구멸하게 된다. 이것은 쾌감과 불쾌중추를 통해 이루어지게 되는데 쾌감을 느끼게 되면 계속 그 일을 하려고 할 것이다. 캐나다 액길 대학의 섬리학자 제엄스 올즈는 쥐의 전뇌 속에 전기자극을 줄 수 있는 레버를 설치한 바 그 쥐는 지철 때까지 레버를 당기는 것을 관찰했다. 그는 이 부분을 쾌감중추라고 불혔다. 미국 튜데언 대학의 로버트 히스 교수는 중격핵 부근에 전극을 연결하면 남자는 감정이 몽홍해지고 여지는 사랑받는 느낌올 가지게 된다는 사실을 발견했다. 쾌감을 얻으면 상이 되어 자극을 좋아하게 되고 불쾌감을 얻으면 벌이 되어 그 자극을 피하려고 할 것이다. 이 상과 벌을 통하여 학습이 이루어지게 된다. 대뇌변연계는 또한 대뇌의 신피철과도 연결되어 있어 신피질의 영향을 받는다. 그래서 감정이 이성의 영향을 받기도 하고 이성이 감정의 영향을 받기도 한다. 그러나 모든 자극은 대뇌변연계에 전달되고 난 후 신피철에 전달되어 처리된다. 그래서 감정척 반웅이 먼저 나타나게 된다. 3충 뇌언 신피질은 진화에 있어서 가장 나중에 발달된 뇌로서 1충 뇌와 2총 뇌의 외각올 툴러싸고 있는 2~4mm 두께의 피막으로 80억 개의 신경세포로 이루어져 있다. 뇌의 신경세포의 대부분이 여기에 몰려 있는 생이다. 대뇌피철은 호두의 표면파 같이 굴곡이 지면서 뭉쳐 있는데 펴면 그 넓이가 O.5m로 신문지 한 장 정도가 된다. 신피질은 시상으혹부터 들어오는 체내외의 정보를 저장, 비교, 분석하는 곳셔다. 대뇌피질의 중요한 기능은 들어오는 자극에 대해 과거의 청험, 미래에 대한 예록, 믿음 동에 버추어 여러 가지 반옹에 대한 대안융 만들고 푼석하는 것이다. 그래서 위험한 척이 나타났을 때 다른 동물은 본능적으로 달아나지만 신띄질이 발달한 사람은 이런 본능을 누르고 돌을 던져 대항하게 되는 것이다. 이것이 지능척인 판단이고 이 지능이 대뇌피철에 기인하는 것이다.
신피질은 해부학적으로 두골의 분류와 비슷하게 네 부분으로 나누어지는데 각 부분은 기능이 서로 다르다. 이들 네 부분 중에서 유일하게 전두엽은 몸의 운동에 전혀 관여하지 않고 단지 정신척 작용에만 관여한다. 이 전두엽이 다른 동물과 뚜렷이 구멸되는 부분이며 다른 풍물에 비해 피질이 차지하는 비율이 크다. 그리고 이 부분이 뇌의 진화에 있어서 가장 나중에 나타나는 부분이다. 전두엽 (frontallobe)
전두엽 (frontallobe)
전두엽은 앞 이마에서 시작하여 두골 중간까지 퍼져 있는 신피철의 대표척인 부분으로 다른 부분에 비해 가장 크다. 이 천두엽의 기능은 가장 렬 알려진 부분이긴 하나 지응운등을 의식적으로 통제하는 부분이다. 동물척 본능보다 한 차훤 높은 이성척 행동을 보얼 수 있는 이유가 바로 여기에 있다. 계획, 판단, 개념의 형성이 이루어지는 곳이고 칙감이 이루어지는 콧이기도 하다. 그래서 전두엽은 바로 개성, 지능의 칩이다. 이 전두엽은 고등동물일수록 차지하는 비율이 크다.
측두엽은 뇌의 양옆에 위치하여 정보의 저장 빛 횟수률 판장하는 것이다. 이 기억은 이성적이고 지성척언 행동에 중요한 요소이기 빼품에 전두엽의 활동에 중요한 부분이 된다. 측두엽에는 또한 청각중추가 있어 서 청각을 관장한다. 두정엽은 뇌의 윗부분에 위치해 았으며 생각, 정보들올 연결하고 종합하는 기능올 가지고 있다. 이 기능 역시 지농 형성에 중요한 부훈이다.이 두정엽에는 촉각중추가 있고, 몸의 위치감각을 관리한다. 그래서 이 부분이 손상되면 몸의 공간척 위치에 혼란이 온다.
 두정엽
두정엽
그렴 3.16 대뇌피철
대뇌피질의 네 부분 중에서 가창 작은 후두엽은 뇌의 뒷부푼에 위치해 있으며 시작을 관리한다. 그래서 이콧이 손상되면 시각 자극을 제대로 처리하지 못하기 예문에 다른 부분어 건전하더라도 시각 장애를 면치 못하게 된다. 이 네 부분은 기능이 서로 구푼되어 있으나 신경섬유로 서로 연결되어 있어서 어디서 어떤 일이 벌어지고 있는지 서로 알고 있다. 대뇌변연계에서 직접 수신하는 후각올 제외하고는 모든 감각척 자극은 시상융 통해 대뇌피절에 전달된다. 그래서 신피질 곳곳에 퍼져 있는 각종 충추와 시상 부분은 서로 대응되어 있다.
이 신피철은 천술한 바와 같어 진화와 갚은 관련이 었다. 진화에 있어서 신피철의 변화와 같이 극척언 변화는 없다. 언류 진화의 역사로 바로 신펴질 변화외 역사라 해도 과언이 아니다. 륙히 천두엽의 변화가 괄목활 만하다 그렴 3.17에서 보는 바와 같이 신피질 중에서도 전두엽의 변확가 푸드러진다. 이련 첨에서 앞으로도 언류의 진화는 이 전두엽에서 혹혀 많이 얼어날 것이라고 예측할 수 있다.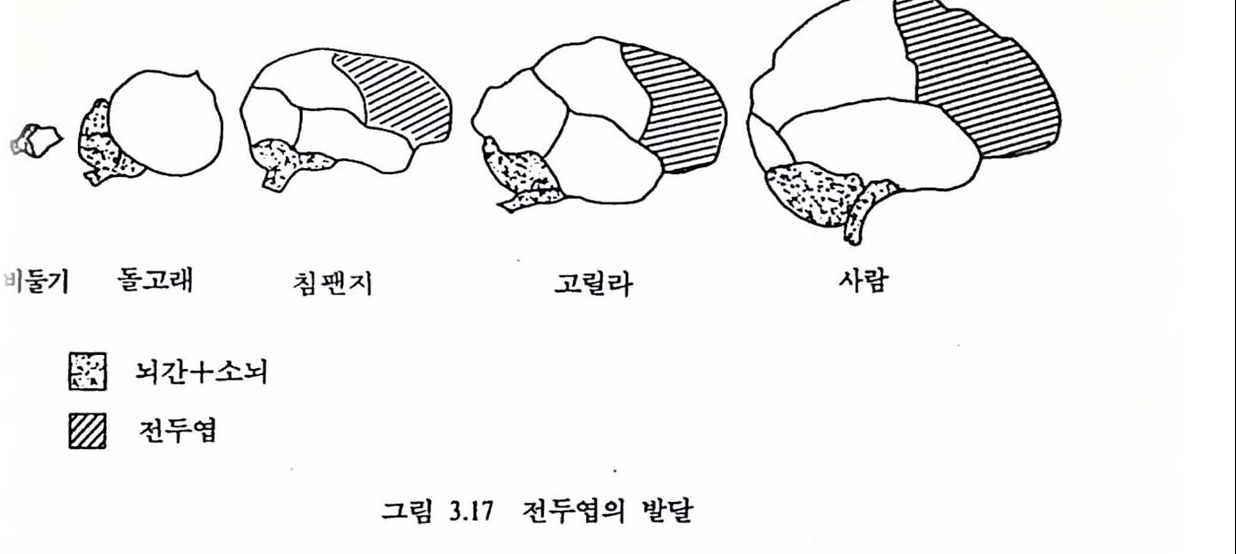 짧
짧
그림 3.17 천두엽의 발달
2.3 .뇌 확장과 그 역할 인류는 지구상에 출현한 이후 끊임없이 진화되어 왔고 여러 종이 파쟁되었으나 현인류만이 생존하고 있다. 왜 현언류만이 잘아 남았울까? 그 셜명은 우선 환경 척웅력에 초점을 맞출 수 있을 것이다. 진화론의 업장에서 볼 때 환경에 척웅할 수 있는 종만이 살아 냥을 수 있율 것이기 때둔에 사람이 인류의 여러 종 중에서 가장 환경 척웅력이 컸고 결과척으로 지금까지 살아 남을 수 있게 되었다. 그러연 사랍의 어떤 요쇼가 환경 척웅력을 크게 하였을까?
이 문제를 다루기 천에 먼저 물질의 여러 존째 양식 중에셔 각 양식이 어떻게 환경에 척웅하는지 보도록 하자. 물철 : 생명체가 아닌 것은 모두 물질로 훈류하겠다. 그러면 률철옴 대단히 다양하다. 철, 산소, 단소, 우라늄 퉁 단위훤소서부터 화합물인아황산 가스, 소금 등을 위시하여 생명체를 만드는 대단히 복잡한 물질언 아미노산, 핵산 등이 있다.
각 물질 개체는 독특한 특성을 가지고 있다. 물은 수소와 산소의 화합물인데 우리 주위에서 가장 흔한 물질 중의 하나이면서 생명체의 가장 기본 물철 중의 하나이기도 하다. 물은 평상 온도에서는 액체로 되어 있으나 100℃ 이상이 되면 기체로 변한다. 그리고 O°C 이하가 되면 고체가 된다. 물은 액체로 있다가 고체로 될 때에는 결정체가 되는데 각 물질틀은 독특한 결정모습을 가지고 있다. 이런 모습은 물질 개체의 특성에 기언 한다. 물철의 변화는 이련 자체의 특성파 외부의 험에 의해 일어난다. 물의 경우 열이 가해지연 얼음이 물이 되고 기체가 된다. 눈의 형태로 되어있던 물이 압력을 가하면 눈뭉치가 된다. 그것이 쌓이면 거대한 벙하가 되기도 한다. 만일어떤 물질이든 외부의 험이 미치지 않으면 그 상태 그대로 었다. 예를 들어 눈에 어떤 중력도 작용하지 않고 기타 어떤 형태의 험도 작용하지 않으면 향상 눈 그대로 었다. 그러나 우주에는 무수한 험이 상호작용하며 이러한 혐의 상호작용으로 위치가 변하기도 하고 상태가 변하기도 한다. 그런데 각 물철은 그 특성에 따라 외부 에너지와 험에 민감하게 반웅할 수도었고 그렇지 않올 수도 있다. 물은 조금만 끓이면 기체가 되지만 철은 좀체로 기체가 되지 않는다. 이것으로 철이 온도에 대해 안청되 어 있다고 할 수도 었다. 식물 : 식물은 다른 양상으로 환경에 대응한다. 물질은 받는 힘만큼 변한다. 식물도 역시 받는 힘만큼 변한다. 이련 점에서는 물질과 다를 바가 없으나 식물은 생명체 모두가 그렇듯이 자기 보존의 독특한 방법을 가지고 있다. 식물윤 씨앗이라든지 출기 일부를 이용하여 완전히 새로운 자기률 만들어 낼 수도 었다. 민들레 씨앗올 땅에 심으면 외부환경의
그림 3.1 8 철새
힘을 이용하여 줄기가 나오고 꽃이 피게 된다. 그리고 일정 기간이 지나가면 써앗이 생겨 변화되기 전의 자기로 환원된다. 이런 현상은 물질에서 찾아볼 수 없는 현상으로 생명체의 자기보존 능력의 결과이다.
동물 : 동물은 식물이 가지지 못한 기능을 더 가지고 있다. 움직일 수 있는 기능이 그것이다. 식물도 자라면서 햇빛이 많은 쪽으혹 휘어진다. 이것은 성창이나 자기보존에 좀더 좋은 조건을 확보하기 위하여 움직이는 것으로 해석할수도 있다. 그러나 식물은 뿌리를 고정시키기 때문에 움직이지 못한다. 식물은 상장과정에서 환경에 척웅하는 기능올 가지고 있을 따름이다. 그러나 통물은 식물이 가지지 못하는 신경계를 가지고 있어 외부의 지극에 부단히 반응하며 중추신경계의 뇌는 여러 외부자극에 대한 행동표 3.4 환경척응력
 -램웰뚫輔뽑 12출070 억 현만 년년연 대천전 환환자직경경기접척증조반응성식응력
-램웰뚫輔뽑 12출070 억 현만 년년연 대천전 환환자직경경기접척증조반응성식응력
반웅을 결정한다. 그래서 위험을 감지하면 회피운동을 하며 배가 고프면 음식을 찾아 헤매기도 한다. 지렁이를 자극하면 꿈틀한다. 고슴도치는 위험을 지각하면 자기 몸을 둥골게 만들어 보호한다. 이러한 자기보호 본능과 그에 필요한 운동은 통물에 찰 발달되어 었다. 식물에도 이러한 자기보호 본능이 없는 것은 아니다. 햇빛이 내려씌면 엎은 축 늘어진다. 햇빛이 강하면 수분 중발이 섬하여 잎이 늘어지게 되는 것이다. 그래서 이것도 넓은 의미의 자기보호 작용이라고 할 수 있다. 그러나 동물은 외부의 자극을 인지한 후 움칙언다. 이것이 식물과 다른 점이다. 그래서 동물은 식물보다 폭 넓고 고도의 자기보호 기능을 가지게 된다. 또한 동물은 즉흥척인 본능뿐만 아니라 미래에 대한 행동을 셜계하는 기능도 가지고 있다. 곰은 가율에 많은 음식을 먹어 영양분을 저축하여 두었다가 통면으로 들어간다. 거미는 줄을 쳐 두고 기다렸다가 줄에 걸리는 고추욕 마버시켜 먹는다. 철새들은 철에 따라 자기가 살기에 좋은 곳으로 옳겨 다닌다. 이련 행위는 식물에서 찾아폴 수 없는 행위로 동물 뇌의 고도의 기능에서 유래된다.
한현, 언간운 동물 중에서 지능이 가장 발랄한 종류아다. 인간은 다른 통풀에 비해 육체척으로 빈약한 면도 었으나 지능, 즉 뇌가 이를 보완할 뽕 아니라 오혀려 우월하게 만든다. 뇌는 중추신경계의 중심부로 인식, 반용, 기억, 믿융, 계산 기능 풍 다양한기능을 가지고 있다. 정서를 관창 하며 각 언간 개체의 개성올 륙청짓는 곳이기도 하다. 이 뇌로 언하여인간은 환경의 개조 능력을 가질 수 있게 되었다. 동물은 여러 환경 중에서 좋은 환경을 선택할 수 있다. 그러나 인간은 나아가 이상척언 환경을 조성한다. 추운 콧에서는 따뭇한 칩을 짓는다든지 옷을 업는다든 지 따뭇하게 불을 피운다든지 하는 행위가 그것이다.
정보화사회와 뇌 역할 인류는 지구상에 나타난 후 대부분의 기간을 식물채집과 수렵으로 지냈다. 그러나 기원천 7000~8000년경에 문명이 형성되기 시작하면서 인류는 대단히 빠른 문명의 변화를 경험하고 있다. 미국의 토플러(Alvin Toffler)는 그의 베스트셀러『미래의 충격』에서 이 변화를 생생하게 설명하고 있다. 그에 의하변 인류역사의 시작을 신석기문화가 시작되는 약 5만 년 전이라고 볼 때 인간의 수명을 62세로 본다면 인류역사는 800생애가 된다. 이 800생애 중 650생에는 동굴에서 보냈다. 그리고 단지 최근 70생애만이 효과적인 상호교신이 가능하였고 단지 최근 6생애만이 인쇄된 글을 볼 수 있었고 단지 최근 4생애만이 시간을 갱확히 쟁 수 있었고 단지 최근 2생애만이 전기 모터를 사용할 수 있었다. 그리고 요즈음의 풍부한 제품은 바로 현재의 800번째 생애에 비로소 나타난 것이라고 최근의 급격한 문화 • 문명적 변화를 기술하고 있다. 그는 이런 변화를 그동안 때때로 얼어나는 독렵척이고 그저 지나가는 바람으후 보지 않고 사회 자체가 연속척으로 변하고 있다고 보교 있다. 그래서 그는 또 다른 책「제3의 물결」에서 그동안 두번째의 충격(변화의 물결)이 있었고 이제 제3의 물결을 맞이하고 있다고 설파하고 있다. 언류는 대략 1만 년 전에 나타난 농업혁명으로 오랫동안의 수렵과 채집의 생활에서 벗어나 식량을 칙첩 경작하였다. 그 결과 청착생활이 이루어지고 취락이 형성되었다. 이런 농업사회에 대해 지금으혹부터 대략 300년 전부터 또 다른 변화의 물결이 다가왔다. 산업혁명이 바로 그것이다. 그동안 농업기술은 지구 구석구석에 퍼져 농경사회가 갱확되어 갔었다. 이 안청된 사회가 1만 년 동안 유지되다가 17세기 중반부터표 3.5 변화의 파도
 파도 λ1 7 1 혁명 중심내용
파도 λ1 7 1 혁명 중심내용
공업기술이 발달하면서 세찬 제2의 물결이 다가온 것이다.
이 산업혁명은 영국에서 불길이 당겨져 전세계로 퍼져 나갔다. 에너지를 변환시키는 기술이 발달하고 그 양산체제가 발달됨에 따라 상품이 값싸게 양산되고 그래서 풍요의 사회를 맞이하게 되었다. 특히 제철기술 발달에서 시작하여 조선, 자동차 등 현대의 찬란한 산업문명이 탄생되었다. 이 산업혁명이 지구 구석구석에 퍼져 갔으며 농엽혁명의 물결은 사라지고 산업혁명의 물결이 철청에 이르게 되었다. 그련데 이제 또 다른 물결의 조짐이 보이기 시작한다. 그것은 정보혁명으로서 1940년대에 나타난 컴퓨터에 기언한다. 토플러는 이 전환점을 미국의 경우 1955년부터 10켠 사이라고 보고 다른 선진 산업국가도 버슷하다고 보고 있다. 미국의 네이스버트(John Naisbitt)는 그의 베스트셀러 Megatrends에서 지금 우리 사회에 얼어나고있는 10대 변천 중 〈산업사회로부터 청보사회로의 변천〉올 제일 먼저 거론하고 있다. 그는 많은 사람이 반대하고 있지만 분명히 정보률 기본으혹 한 산업으로의 변천이 뚜렷하다고 설파하였다. 하버드 대학의 사회학자 멜(Bell)은 이런 새로운 변화를 〈활산업사회〉라고 표현하고 있다. 싼엽사회나 청효화사회률 시대척으혹 훈명히 구분하기는 어렵다. 연속척으혹 변화하는 과청에서 어느 시기는 산업사회, 어느 시기는 청보화사회로 환르기 빼품에 그 과도기는 서로 겹쳐 었다. 그러나 청효화사회로 이행되는 륙갱척 사건틀이 었다.표 3.6 국제산업 구조(1983년)
 나라 농업 산업 서비스업
나라 농업 산업 서비스업
자료:『정보화사회』, 서울대학교 출판부, 1986.
그러면 이런 특정이 언제부터 나타났는가? 우선 그 특정 중 가장 강한 것이 산업 종사자의 구성이다. 산업사회는 생산업종에 종사하는 사람이 대부분이라야 하고 정보화사회는 정보산업에 종사히는 사람이 대부분이라야 한다. 벨은 미국의 경우 노동력의 대부펄이 주로 제조업에 종사했고 서비스업 종사자는 평균 12% 정도였으나 1970년대에는 65%를 넘어서고 있다는 사실을 지적하고 있다. 버치 박사는 미국에서 1970년대에 새로 생겨난 1900만 명의 일자리 중에서 제조업은 5%뿐이고 약 90%가 청보, 지식, 서비스업이었다는 것으로 1970년대의 정보화사회의 돌입을 셜명하고 있다. 현재 중진국, 선진국에서의 이런 추세는 뚜렷하다. 세계 은행의 조사에 의하면 선진국의 경우 농업에 3%, 산업에 35%, 서비스업에 62%이며 중진국도 유사한 추세에 었다.
네이스버트는 Megatemds에서 정보화사회는 1956년과 1957년 사이에 걸쳐 시작되었다고 주창한다. 그는 이 시기가 미국이 산업국카로 풍장한 시발점이기 때문이라는 것이다. 1956년에 화이트 컬러가 홉루 컬러륨 앞지르고, 대륙간 전화서비스가 시작되었고, 1957켠에 소련은 우주션 스푸트니크를 발사하여 우주통신시대률 개악한 해이다. 그래셔 이 기감이 산업사회에 종말올 고하는 기간이고 바로 정보화시대의 시작에라고 주장한다.표 3.8 환경조성
 시꺾-샌현 기-대기대 움자인조막연공성 생 모태개습계조
시꺾-샌현 기-대기대 움자인조막연공성 생 모태개습계조
한국의 경우는 1983년에 발표한 로스토(W.W. Rostow) 교수의「한국과 제4차 산업혁명」이라는 논문에서 1980년대말부터 시작될 것이라고 전망하고 었다.
방직기술을 주축으로 하는 제1차 산업혁명, 제철, 조선, 기계를 중심으로 하는 제2차 산업혁명, 자동차, 탤레비전 등을 주로 하는 제3차 산업혁명을 서구사회보다 3분의 1기간에 줄여 1980년대말에 끝낼 것이라는 것이다. 서구보다 30년, 일본보다 20년 정도 늦은 시기이다. 정보화사회의 기반 기술윤 전술한 바와 같이 컴퓨터 기술, 통신 기술이다. 컴퓨터는 1946년에 나타났다. 그러나 그것이 파급되기는 1950년대부터이기 때문에 이런 점으로 볼 때 기술적인 면에서는 1950년대가 정보화사회로 진업하는 시기이다. 그러나 산업 측면에서는 1970년대 산업종사자 구쪼의 변화로 나타난다. 이런 점올 종합할 때 전통척인 사회 분류 방식용 따르면 1970년대가 되지만 그 시발은 1950년대라고 보는 것이 옳겠다. 정보화사회의 특갱 : 청보화사회에서 중요한 요소는 정보와 지식이다. 이 정보와 지식올 어떻게 처리하느냐에 따라 사회의 양상이 달라진다. 갱보화사회에서는 청보률 처리하는 산업, 그리고 정보를 창출하는 산업 (지식산업)이 주축울 이루면서 고도의 사회로 발전한다. 과거의 사회 형태와는 여러 가지로 다르다. 자원의 형태, 기술 형태, 사람들의 생활 양식 풍 여러 가지에 변화가 얼어난다. 벨은 그의 저서『정보화 사회에서의 사회척 구조』에서 그 륙칭을 표 3.7과 같이 버교하고 있다. 이련 사회척 변화롤 가창 단척으혹 표현하는 말은 3A 이다. 즉 사무표 3.7 탈산업화사회 : 비교도표
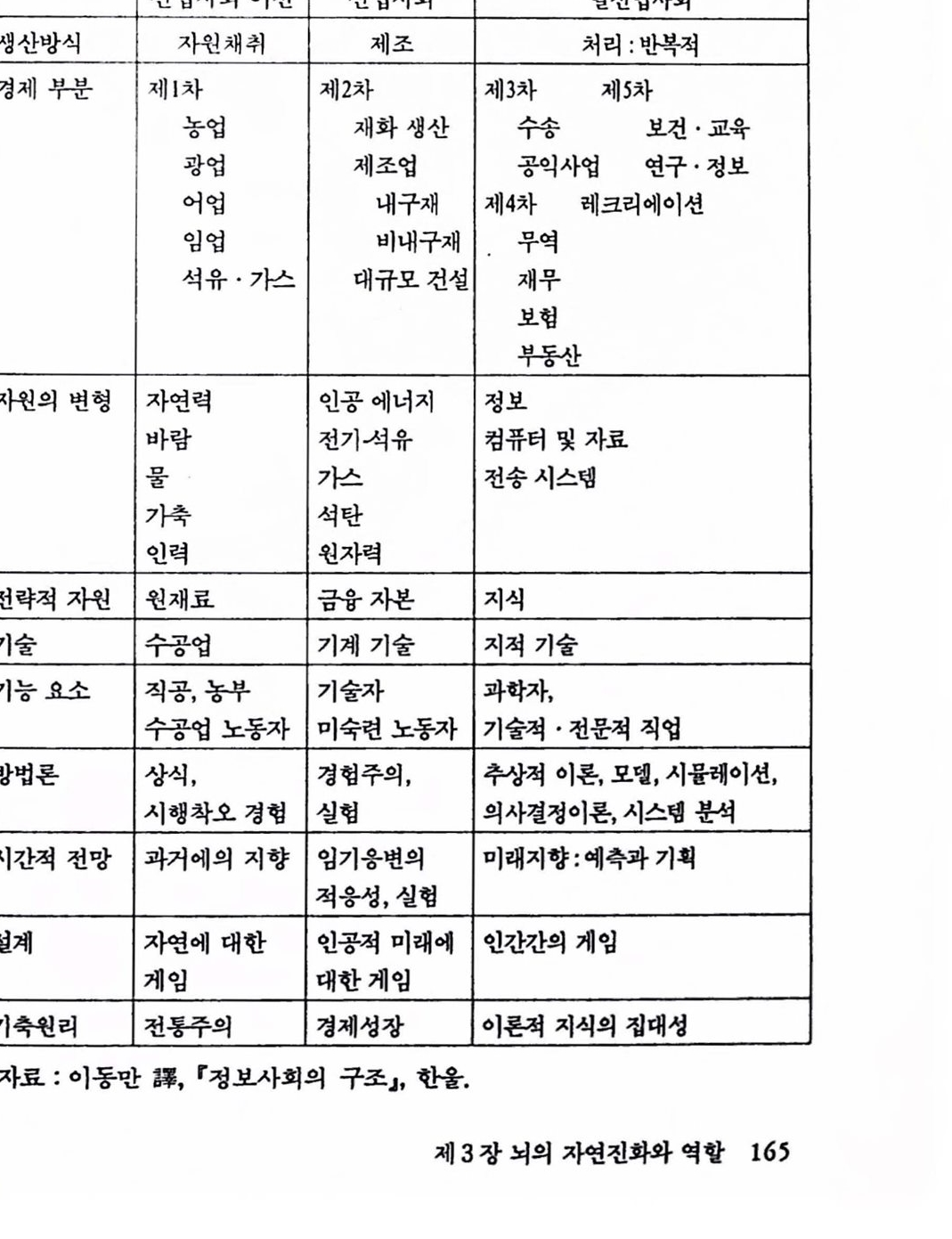 산업사회 이전 산업사회 탈산업사회
산업사회 이전 산업사회 탈산업사회
자료 : 이동만 譯,『정보사회의 구조』, 한울.
점점 이런 사무 지동화의 과정에 있다. 요즈음 관청의 서류는 복사로 이루어지는 일이 많다. 새로이 만들어야 할 경우 자동 타자기로 처리된다. 멸리 전송할 때는 팩시밀리를 이용한다. 웬만한 논의는 전화로 해버리고 서로 떨어져 화면으로 회의할 수도 있다.
공장 자동화는 공장의 생산 업무를 지통화하는 것이다. 공장은 일찍부터 기계화되어 왔고 이제 자동화되어 간다. 공장 업무단 분업 개념의 도입으로 단순반복작업이 대부분이어서 사람들은 싫증을 내기 쉽다. 그러나 기계는 싫충을 낼 이유도 없고 신뢰도가 높다. 그래서 기계화가 가속되어 왔다. 더 나아가 산업 로봇어 등장하면서 정점 사람의 손을 대치해 가고 있다. 지금은 경비 문제로 공장을 완전 자동화하지 않으나 기술적으로는 어떤 공장이든 자동화시킬 수 있는 기술이 발달되어 있다. 그래서 큰 회사들은 주로 부분적으로 지동화시키고 있다. 가정의 자동화는 시장, 은행, 기타 관청일플을 가정에서 처리할 수 있는 것을 의미한다. 가정의 자동화는 아직 구체화되고 있지 않지만 점점 이런 방향으로 가고 있다. 이렇게 되면 집에 앉아서 필요한 업무를 마치고 사람틀은 정접 많은 시간을 고차원척인 정신 활동에 보낼 수 있게 된다. 그러면 앞으로 정보확사회의 사회상은 어떤가? 네이스버트는 앞으로 칙장의 장소는 첩이든 칙장이든 판계없고 시간대도 마음대로 선택할 수 있게 될 것으혹 예싱하고 있다. 직장과 가정에는 컴퓨터 단말기로 연결되어 있어 구태여 직장에 출근할 필요가 없다. 선진국에서는 벌써 가정에 단말기를 설치해 두고 집에서 출근하고 퇴근하며 가끔 한번씩 회사에 나가기만 하연 되는 경우도 었다. 또한 그는 앞으로 컴퓨터는 누구든 다룰 줄 아는 필수품이 될 것이며 천품인보다는 척용할 줄 아는 상식언의 시대가 온다고 했다. 많은 지식용 컴퓨터 네트워크률 통해 언제나 사용할 수 있어서 전운인의 역할이 정정 약화된다. 그래서 전푼안보다는 상황 변화에 척응하는 일이 더욱 충요하게 된다는 것이다.지동화(Office Automation : OA), 공장 자동화(Factory Automation : FA), 그리고 가정 자동화(Home Automation : HA)이다. 이것은 여태까지 손으로 이루어지던 각종 작업이 컴퓨터로 처리된다는 것을 의미하는데, 이 지동화는 지식산업의 결과로서 사회 변화에 있어서 가장 특갱적 변화로 볼 수 있을 것이다.
사무 자동화는 행정 업무의 자동화를 의미히는데 공공 기관의 행정, 사기업의 행정 등 각종 업무가 지동 처리되는 것이다. 현재 행정 업무는 오명 박사는 정보화사회의 사회성을 우선 개인 생활에 있어서는 과거 단체적 활동으로부터 앞으로는 점차 개성적인 개인 중심으후 바뀔 것이라고 예상하고 있다. 그리고 가정 생활은 예전에는 주로 직장에서 대부분의 시간을 보내던 시절은 지나가고 가정이 곧 직장이 됨에 따라 여가 활용이 많아지는 생활이 된다. 물건을 사고 파는 것도 집에서 이루어지고 정치도 앉아서 한다. 주민등록초본을 떼러 동사무소에 갈 필요가 없게 된다. 교육은 컴퓨터 가정 교사가 있어 밤낮 없이 가농하고 그래서 명생 교육이 기능해진다. 스님, 목사들의 탤레버전을 통한 설교와 상당이 인기가 높아간다. 이러한 변화상은 가농한 일이고 현재 부분척으로 일어나고 있는 상황이다. 사렴들은 점점 정신적 여유플 가지게 되나 한편 대단히 메른 속도로 사회가 변해 간다. 그래서 정보화사회에서 중요한 것은 이련 변화률 홉수할 수 있는 문화적 배경을 발전시키고 개인척으로는 갱신척 준비가 필요하게 된다. 이런 준비는 결국 다른 시대에서와 같이인간의 뇌가 담당하게 된다. 그러나 이 정보화사회에서의 뇌의 역할은 다른 어느 시대보다 그 비중이 크고 오직 뇌에 의존한다고 해도 과언이 아니다. 인공생태계 언간은 뇌의 발달로 환경의 선태권뿐만 아니라 환경울 개죠활 수 있는 능력을 가지게 되었다. 이 환경의 개조는 초기에는 비바람을 펴하는 움막에서 시작되었지만 농경이 시작되는 신석기시대가 되면 자연훌 개조하기 시작하였고 이제는 더 나아가 새로운 생태계를 만틀어 보고자 하고 있다.
움막 : 동물은 보통 자연구조물을 이용하여 비나 눈, 추위를 피한다. 초기 인류도 예외는 아니어서 거의 2백만 년 동안 계속되었던 석기시대 대부분을 자연동굴을 이용하여 생활하였다. 이때의 생활은 주로 채집과 수렵이었으며 추위와 방어 목척으로, 그리고 나아가 정찰에 유리한 위치를 차지하기 위하여 통굴생활을 하였다. 그리스 에피루스(Epirus) 지역에 있는 동굴은 남향이고 계곡을 굽어보고 있어 전형적인 석기시대의 동굴접(rock-shelter)이다. 이련 생활에서 15만 년 전 무스테리안 문화시대에 오면 생활 터전이 동굴에서 들판으로 옮겨지게 된다. 들판에 오케 되면 자연히 집을 지어야 한다. 프랑스 니스의 테라 아마타(Terre Amata) 유척지에서는 12만 5000 년 전의 이련 유척이 남아 었다. 주위에 7~8cm의 기둥 구멍이 었고 중앙에 30cm의 기둥 구멍이 있는 것으로 보아 집을 만든 혼척임에 틀렴없다. 이련 생활 거주용 접은 점차 시간이 흐르면서 농경이 시작된 신석기에 이르게 되면 다양한 건물로 발전하게 된다. 자연의 개조 : 인공위성에서도 포착되는 인공축조물이 중국의 만리장성이다. 이 장성은 북쪽 오랑캐를 막기 위하여 여러 왕조를 거치면서 축성한 것으로 진시황제 때 대대척 보완축성이 있어 유명하다. 이 창성은 바로 인공척 산맥이다. 이 창성으로 북쪽과 남쪽의 교류가 안위척으로 두철되었다. 몽고의 유럽 침공이 이 때문이라고 하는 이유도 여기에 었다. 이런 성은 기훤전 8000년경부터 발견되는데 이스라엘에서 방어용 토루가 형성된 유척이 었다. 그러다가 기훤천 2600년 수메르 국가가 형성되면서 만든 높이 수미터, 킬이 약 2km의 성의 유척이 남아 었다. 이러한 본격척인 자연개조는 신석기부터 시작되었다. 신석기에 들어오면셔 놓갱이 시작되는데 농경에는 필연척으로 관개가 필요하다. 관개는
그림 3.19 러시아 매머드뼈 오두막집 복원도
곧 인공하천과 직결된다. 관개유적으로는 기원전 5500년 전 중동 지역에서 발견된다. 그러다가 로마시대에 오면 대규모 토목공사가 이루어지고 자연의 개조가 활발해진다. 수나라 때의 대운하, 현재 진행중인 리버아의 대수로 공사, 각종 대규모의 댐 공사가 이련 예에 속하겠다.
지하 거주공간 : 거주공간을 지하, 바다 속으로 확장해 보고자 하는 욕망은 오래전부터 있어 왔다. 바다에 비하여 지하 거주공간 개발은 가시화 되어 가고 있다. 인류는 초기 때부터 동굴을 거주공간요로 사용하여 왔고 토굴을 거쳐 지하주택도 사용하여 왔었다. 현대는 이련 경향이 점점 발달하여 지하 수미터에 지하철이 건셜되고 지하상가가 형성되고 있다. 일본에서는 본격척인 지하 거주공간이 계획되고 있다. 일본 다이셰이(Taisei)사는 앨리스시(Alice city) 계획융 세웠는데 이 계획에 의하면 지하 165m에 반경 약 86m, 높이 약 65m의 대형 콘크리트 훤주들을 세워 이들 훤주를 연결하여 야 속에 발천시셜, 수도시갤등 각종 생활 기반시설과 각종 상가, 운동시설, 사무실 응을 세움으로써 10만 명의 소도시를 건설히는 것이다.
시미주시는 이보다 더욱 큰 야심의 계획을 발표하고 있다. 약 50만 명의 언구를 포용할 수 있는 약 1500m2의 도시를 800억 달러의 비용으로 건설하겠다는 것이다. 이 도시는 물론 온도, 습도 등이 자동적으로 조절되나 가능한한 자연광션을 채광하도록 계획을 세우고 있다. 이런 지하도시 건설에는 화재, 지진 등 재해에 대해 취약점이 있으나 이런 취약점은 기술적으로 극복될 것으로 보인다. 이렇게 되면 지하도시 출현도 멸지 않다.
그립 3.20 지하도시
언공생태계 : 우리는 가정에서 쉽게 수족관이나 테라리움을 본다. 수족관은 일정한 용기 속에 민물이나 바닷물을 넣어 펴를 키우는 것이다. 산소 공곱을 계속하고 이따금 영양 공급을 시키지만 인공적인 환경에서 고기를 자라게 하는 경우이다. 이와 유사하게 테라려움은 조그마한 멀폐된 유리 용기 속에서 식물을 자라게 하는 것이다. 수분은 그 속에서 증말하고 또 물로 변히는 과정을 통해 순환되고 햇빛은 유리 용기를 통해 받아들인다. 식물은 외부와 단절된 이 세계 속에서 광합성을 위한 태양 에너지만 유리를 통해 받아들일 뿐 그 외는 모두 밀폐된 공간내에서 자체적으로 충족해 나간다. 수족관에 비하여 훨씬 자급자족하는 체제이다.
지구에 인구가 폭발하고 지구오염, 핵위험 등으로 사람들은 끊임없이 자연조건 외의 생활터전을 찾아왔다. 우선 우리들의 상상력을 끄는 곳은 바다와 외계이다. 바다는 어둡고 수압이 강하여 생활터전을 만들기가 어렵다. 외계는 미지의 세계기기 때문에 우선 탐사에 급급하고 있다. 달을 탐사하였고 다른 멸을 탐사하고 있는 중이나 희박한 공기, 강력한 자외선 둥 그대로 생활하기에는 어렵게 판단되고 있다. 그러나 탑사를 계속하다 보면 지구와 비슷한 상황올 가진 별이 발견될지도 모른다. 그동안 우리는 이러한 어려운 상황 아래에서도 생존할 수 있는 인공생태계를 연구하는 것이 바람직하다. 바이오스페어 II(Biosphere II) : 이 사업은 달이나 화성 둥 외계에 그대로 옮걱서 사람이 살 수 있도록 하고자 하는 소형 언공지구에 판한 실험이다. 미국 애리조나주 오라를(Oracle) 곤처에 길이 180m,폭 2m의 이온설과 1,800m2의 부속농창을 세워 이 안에 아마존과 칼은 열대우렴, 카리브해률 닮은 바다, 그리고 초현, 강, 사막올 만든다. 여기에 200종 이상의 식물이 심어져 있고 500종 이상의 바다생물,600종 이상의 조류, 동물이 방목되며 강에는 데라비아 풍 각종 물고기가 살케 된다. 이 사업은 Decision Team사 와 Decision Investment사에 의해 3천RV
」{τ뺀γ 께에WJ륜 ·{ {니 잊 τ ‘늄-승‘「‘그림 3.21 바이오스페아 II
만 달러의 예산으로 1984년 12월에 시작하여 1990년 9월에 완공할 예정으로 있으며 완공되면 이 바이오스페어 II는 밀얘된다. 이 속에서 8명의 과학자가 2년 동안 위급상황이 아닌 이상 외부와 단절된 채 인공생태계로서의 자급자족의 가능성을 시험하게 된다.
이 소지구는 이 사업의 책임자 아우구스턴(Margret Augustine)과 호즈(Philip Hawes)가 설계한 것으로 아우구스턴은 우주생물원 모험(Space Biosphere Vectures) 계획의 수석행정관이커도 한데 이 계획에 의거 생물원 연구개발센터(Biospheric Research and Development Center) 에서 바이오스페어 II의 시험용 원형을 만들어 지난 3월 8일에서 13일까지 완전히 밀폐시킨 상태에서 사람이 살아 보는 실험에 성공한 바 있다. 그리고 9월 완전 밀폐 전에 바이오스페어 II를 시험척으로 밀폐시격 보는 시험을 가질 예정으로 있다. 이 바이오스페어 II는 완전 자곱자족체제로 운영되는데 사막이 가장 낮은 곳에 위치하고 열대우렴이 가장높은 곳에 위치한 구조와 각 방의 온도 등으로 공기는 계속 흐르게 하고 있다. 물은 높은 곳에 있는 열대 우립에서 바다와 강으로 흐른다. 그리고 중발하여 대기기류를 타고순환되게 되어 있다. 이 안에 사는 사람은 농사를 짓고 고기률 잡아 먹으면서 살게 되어 있다. 이 사업은 NASA, 스미소니안 연구소, 뉴욕 식물원, 애리조나 대학이 연구에 참여하고 있는데 2년간의 연구률 통해 지상과 전혀 다른 환경에서 생존할 수 있는 생태계를 만들어 보고자하는 것이지만 이 바이오스페어 II의 운영은 컴퓨터의 역할이 크다. 각 방의 상태가 컴퓨터에 의해서, 조사, 보교되며 이런 정보들이 컴퓨터률 서로 잇고 있는 네트워크로 상호 교신된다. 그리고 이 바이오스페어 II와 외계 역시 컴퓨터와 시청각 장치로 교신된다. 이련 교신 방법이 결국 우주에 인공쟁태계가 만롤어쳤을 때 그대로 사용하제 될 교신방법이 훨 것야다.참고문헌 1. 강만식 외 4인『동물생리학』, 교학연구사, 1987. 2. 강만식 외 4인『현대생물학』, 교학연구사, 1987. 3. 강석경 역,『신체의 소사전』, 전파과학사. 4. 강오감 역,『생명이란 무엇인가』, 전파과학사. 5. 고인석 역,『노화는 왜 일어나는가』, 전파과학사. 6. 김광억 역,『오리진』, 학원사. 7. 김범철 외 1인 역,『지구환경 보고서』, 따님, 1990. 8. 김우호,『바이러스의 세계』, 전파과학사. 9. 김용준 역,『인간이란 동물』, 탐구당. 10. 김은국 역,『인간등정의 발자취』, 범양사, 1989. 11. 김준민 역,『기후와 진화』, 민음사, 1987. 12. 김형민,『생체학』, 민음사, 1987. 13. 김훈수 감수,『생명의 신비』, 학원사, 1987. 14. 대학지구과학연구회,『지구과학 개론』, 교학연구사, 1987. 15. 류시화 역,『현대물리학이 발견한 창조주』, 정신세계사, 1988. 16. 박상윤,『진화』, 전파과학사. 11. 박인원 역,『생명의 기원』, 민음사, 1990. 18. 박태홍 역,『생명을 만드는 물질』, 전파과학사. 19. 배기동 역,『문명의 예명』, 범양사, 1988. 20. 손영수 역,『섹스 사이언스』, 전파과학사. 21. 오영근 역,『잠이란 무엇인가』, 전파과학사. 22. 이명우 외 3인 역,『현대환경론』, 한길사, 1989. 23. 이선복,『고고학 개론』, 이혼과 실천, 1988. 24. 이연태 역,『유전자 조작기술』, 탐구당. 25. 이찬기 • 안태선,『환경보전과 인간』, 한국경제신문사, 1987. 26. 장기홍 • 박순옥 역,『생명의 역사』, 민음사, 1987. 27. 정용재,『유전의 본질』, 전파과학사. 28. 정종욱 역,『인간은 어디서 왔는가』, 명지출판사, 1987.
29. 주충노,『생체에너지』, 민음사, 1987. 30. 하두봉,『창조의 제8일』, 범양사, 1984. 31. 한영환 역,『우주의 역사』, 범문사, 1986. 32. 허형태,『해양자원의 인식』, 한국경제신문사, 1989. 33. 현정준 역,『시간의 역사』, 삼성이데아, 1988. 34. 홍영남 역,『생명의 출현』, 안국출판사, 1985. 35. 홍영남 • 강주상 역,『생물물리학』, 안국출판사, 1984. 36. Eigen M., P. Schuster, The Hypercycle, Springer-Verlag, 1979. 37. Hickman C. P. et. al., Integrated Principle of Zoology, Times Mirror, 1984. 38. Sattelle David, Biotechnology in Prospective, Hobsons, 1988. 39. TIME, Atlas of the Body, Rand Mcnally, 1976.
박순달 제 4 장 뇌의 인공적 확장은 가능한가 1. 기기 발달과 인체보철 1.1. 기기의 발달 (1) 석기 인류가 최초로 사용한 도구는 무엇인지 분명치 않다. 도구률 사용하는 동물은 여러 가지가 관찰된다. 새들은 칩을 짓기 위하여 나뭇가지나 풀을 이용한다. 거북이는 모래밭올 파서 알올 낳고 묻어둔다. 어떤 새는 나무 속의 꿀을 먹기 위하여 나뭇가지률 이용한다. 첨땐지는 과얼올 깨기 위하여 돌을 사용할 줄 알며 화가 날 때는 몰율 던지기도 한다. 그리고 표벙올 쫓기 위하여 나뭇가지를 무기로 사용할 줄 안다. 이런 동물이나 영장류가 도구를 사용할 줄 안다면인류도 어느 혜푸터언가 도구를 사용하였을 것이고 처음에는 나무나 풍물의 뼈였융 것으로 추청되나 이련 것들은 보존되어 있지 않아 분명체 않다. 그퍼나 톨옴 오랫통안 변형됨이 없이 보존될 수 있어, 언류 최초의 도구를 이야기할 때
가장 분명히 말할 수 있는 것은 돌이다.
초기 고인류(古人類)가 돌을 어디에 사용하였는지는 분명치 않다. 그러나 석기로 나무나 뼈를 끊거나 다음을 때 사용하였거나 고기를 썰 때 사용하였을 것이라고 상상하는것은 어렵지 않다. 이 밖에도 다양한 용도로 사용하였을지 모른다. 이들은 고운 업자를 가진 화산암 또는 수석(遠石)을 원료로 하여 주먹만한 돌을 직접 타격하여 긁개를 만들거나 찌르개를 만들었다. 이런 최초의 석기제품은 260만 년 전의 것이 발견되는데 보통 200만 년 전부터는 혼히 발견된다. 돌을 도구 제조의 주재료로 사용하던 이 시대를 석기시대라고 한다. 이 석기시대는 언류가 금속을 사용하기 시작하던 5000년 전까지 언류 역사의 거의 대부분을 차지한다. 이 장구한 역사를 거치는 동안 돌을 다루는 기술은 점차 발달하여 농경이 시작되던 1만 년 전 때면 석기시대의 꽃이라고 할 수 있는 마제석기가 나타난다. 이 마제석기는 고도의 기술이 요구되는 연마법에 의해 만들어진 석기들이다. 아직도 우리의 시골에 가면 폴 수 있는 뱃돌이 돌을 갈아서 만든 마제석기 종류이다. 돌을 다루는 이련 기술의 변화 또는 이 기술의 결과로 나타냐는 돌제품의 특징을 구멸하여 구석기시대, 중석기시대, 신석기시대로 나눈다. 그러나 중석기시대는 짧은 변환기 같은 시기여서 크게 보면 돌을 타격하여 제품을 만들던 구석기시대와 폴올 갈아서 제품을 만들던 신석기시대로 구별할 수 있다. 구석기시대 구석기시대는 돌올 타격하여 제품을 만들어 내던 시대로서 200만 년 전에 시작하여 1만5000년 천까지 계속한다. 구석기는 이 오랜 동안의 기술의 발달로 언하여 또다시 전기 구석기, 중기 구석기, 후기 구석기 시대로 구별한다. 천기 구척기시대는 지금으로부터 200만 년 전부터 10만 년 전까지를 말하며 이 시기에는 충격올 가하는 타제법(打製法. percussion flaking)을 사용하여 주먹도끼 등의 각종 도구플 만들었다. 중기 구석기시대는 지금으로부터 10만 년 전에서부터 현대인이 나타니는 4만 년 전까지를 말하고, 후기 구석기시대는 지금으로부터 4만 년 전부터 1만5000년 전까지를 말하는데 이 시기에 오면 전기 구석기보다는 발달된 압제법(壓制法, pressure flaking), 연타볍(述打法, blade-striking)을 이용하여 더욱 전문화된 도구를 만든다든가 작고 세밀한 도구를 만틀게 된다.
고고학애 있어서는 플라이오세와 플라이스토세의 유척을 간칙하고 있는 곳이 중요하다. 이런 콧 중의 하나가 풍부아프리카이다. 1500만 년 전 내지 2000만년 전 아프리카 대륙이 떨어져 나오연서 흥해가 생기는데 그 옆에 이집트에서 말라위에 이르기까지 또 하나의 계곡이 생겼다. 이 계곡은 바다플 이룰 만큼 깊은 것은 아니지만 여기에 오랫동안 퇴척물이 쌓이면서 고고학의 보물들어 발견되고 있는 여러 지역이 만들어지게 되었다. 여러 가지 유적이 많이 발견되는 초기 인츄유척의 보교인 탄자니아의 올두바이(Olduvai) 협곡도 여기에 있다. 이 협곡은 길이가 15km 되는 곳으로 200만 년 동안 퇴척물이 쌓이다가 그 후 20만 년 동안 하천으로 침식되어 만들어진 협곡이다. 올두바이 협곡에는 4개의 푸렷한 퇴적충이 있는데 제1층은 180만 년 전에 형성되었으며 제4층은 70~80만 년 전에 형성된 것으로 추정된다. 그래서 이 유척지는 대략 초기인류가 나타나는 200만 년 전부터의 역사척 유척을 시대별로 찾아볼 수 있는 중요한 곳이다. 올두바이 문화 : 이 유척지 1충과 2충 하부에는 주먹만한 돌을 깨어 날을 세운 찍개(choppers), 구형(球形) 석기, 훤반형 석기 동 여러 가지 석기가 나타난다. 모양과 크기는 일정한 표준이 없으며 가공방법이 죠잡하다. 이것들은 그러나 언과(人科, Hominid)가 만들어 낸 최초의 도구이다. 이련 도구를 위한 재료료서는 고운 업자를 가진 화산암, 수석(flint), 혹요석(obsidian), 규암 등이 사용되었다. 이들 롤올 다른 롤로 모서리를 초기 양면 핵석기 원반형 석기 찍개 다각면 석기
초기 양면 핵석기 원반형 석기 찍개 다각면 석기
그립 4.1 올두바이형 석기
쳐서 박편을 떼어내어 날을 세웠다. 한쪽 면에 타격하여 날을세운 석기도 있고 양면을 모두 타격하여 날을 세운 석기도 있다. 크기는 일정치 않았으며 모양은 주로 긁개(scraper) 또는 찌르개(point) 등이었다. 이런 형태의 석기를 올두바이(Olduvai) 석기라고 한다. 이러한 석기 제작은 독렵척언 사건이라기보다는 각 석기들어 특성을 공유하기 때문에 문화의 형태를 지나기 시작하고 그래서 이 시대 또는 이런 형태의 것을 올두바이 문화(Olduwan culture) 포는 올두바이 산업(Olduwan industry)이라 고 부른다.
올두바이 문화는 주로 아프리카에서 나타나지만 유럽, 아시아 일부에서도 발견된다. 그러연 과연 누가 이런 석기를 만들기 시직하였는가? 올두바이 계곡 1충에서는 1959년 Australopithecus boisei의 유골과 석기가 함께 발견되었는데 가까운데에서 또한 이보다 진화한 Homo habilis 유꼴도 발견되어 최초의 석기제작자는 Homo habilis로 보고 었다. 이 올두바이 석기는 140만 년 천까지 사용되고 있다. 주억도끼문화 : 북프랑스 생따쉘(St. Acheul)은 50만년 전의 유척지로 유명하다. 이곳에서 양면 핵석기(兩面核石器)언 주먹도끼가 발견되어 주먹도끼로 대표되는 운화률 이 지방 이륨을 따 아슐 문화(Acheulean culture, Acheulean industry)라고 푸른다. 이 주먹도끼(hand-axe)는 양면 핵석기로서 150만 년 전에 나타나 100만년 동안이나 사용된 천기 구석기문화의 결청체 중의 하나라고볼 수 있다. 이 주먹도끼의 특칭은 · 배 모양과 같이 길쭉한 축이 있음. · 축을 중심으로 좌우동형임. · 커다란 호박씨같이 양면이 두툼함. · 두 면이 모아지는 모서리는 계속하여 날카로움. 등의 특정을 가지고 있다. 이 도구는 올두바이 석기보다 훨씬 발달된 석기로 보다 세련되고 정교한 기술로 만들어진 것이다. 크기는 보통 주먹만하나 1Ocm 정도의 작은 것도 있다.
주먹도끼의 최초의 제작자는 Home erectus 라고 믿어진다. 올두바이 협곡의 제4층에서 Home erectus 유팔파 함께 주먹도끼가 발견되고 있다. 이 주먹도끼는 아프리가 지역에 칩중적으로 나타나는데 케냐의 킬롬보(Kilombo) 유적지는 70만 년 전의 유척지인데 수백 점의 주먹도끼가 발견되었다. 중기 홍적세 유적지인 케냐의 올로르게사얼리(0lorgesailie) 유적지에서는 주먹도끼와 많은 양의 동물뼈가 발견되었다. 여기서 특기할 것은 그 동물뼈 중에 지금은 멸종된 바분 일종의 뼈들도 있는데 이들 뼈가 상당 부분 선택척으로 부스러져 있었다. 이것은 이 주먹도끼로 바분을 잡은 것이 아닌가 하고 추정할 수 있다. 이 주먹도끼는 유럽에서 15만 년 전에 유행하는데 한국에서는 전곡에서와 파주군 주월리에서 출토된 일이 었다. 중국에서는 아직 발견되지 않고 있는데 저우커우덴(周口店) 유척지는 70만년 천에서 30만년 전의 유척지로서 Sinanthropus가 발견된 것으로 유명한 곳이다. 이곳에서 10만 정의 석기가 발견되었는데 모두 올두바이형 석기들이다. 주먹도끼로 대표되는 이슐 문화는 오랫동안 유지되다가 20만 년 천에서 15만 년 전부터 서서히 변화가 일어나기 시작한다. 주먹도끼는 정점 작아지고 사용 목척에 알맞게 전문화되기 시작한다. 그래서 작고 정교한 찌르개, 긁개 등으로 발천해 간다.
그림 4.2 주먹도끼
신석기시대 200만 년 동안 계속되던 구석기시대는 서서히 막을 내리고 세석기로 대표되는 중석기시대가 온다. 이 중석기시대는 개를 사육한다든지 식량 생산을 시도한다든지 또는 어로기구가 제작되는 것으로 보아 수렵, 채집에서 점차 식량을 경작하는 쪽으로 기울어 가고 있음을 알 수 있다. 이 중석기시대는 지금으로부터 1만5000년 전부터 시작하여 5000년 동안 계속되다가 1만 년 전이 되면 석기시대의 꽃이라고 할 수 있는 신석기시대가된다.
신석기시대는 최후멍하기가 종식되어 날써가 따뭇해지는 흘로세가 시작되는 1만 년 전부터를 말한다. 이 신석기의 세 가지 특정으로는 _.... - .-•- .• -.
_.... - .-•- .• -.
그림 4.3 마제석기
마제석기, 토기, 농경을 들수 있다. 돌을 다루는 기술은 200만 년 동안 발전해 오는 사이에 타격으로 석기를 만들던 기술이 점차 부드러운 나무 또는 뼈망치에 타격을 가하는 연타법이라든지 압력을 가하여 박편을 떼어 내는 압제법 등이 발달하여 중석기시대에는 보통 3cm 미만의 세석기의 전성시대가 된다.
그러나 이 세석기도 결국 타격의 일종으로 만들어진 것이다. 그뒤 지금으로부터 1만 년 전부터 새로운 기술인 마제법(慶製法, grinding)이 개발되기 시작하였다. 이 마제법은 원석을 갈아서 원하는 제품을 만드는 것이다. 이 마제법을 이용하면 더욱 정밀한 도구를 만들어 낼 수 있다. 이러한 마제법은 지금까지의 타격을 가하는 기법과는 근본척으로 다른 기술로서 석제기술에 하나의 획을 긋는 기술 발전이라고 할 수 있다. 지금까지의 석기 제조기술은 원석에 직접 힘을 가하여 박편을 떼어 내어 석기를 만드는 기술로 이런 방법으로 석기를 제조하면 박편이 떨어져 나올 때 원석의 결에 따라 가공되기 때문에 표면을 원하는 대로 만들어 낼 수 없다. 그래서 타격으로 석기를 만들던 시대에는 찌르개, 긁개와 같이 모서리가 뾰족한 석기들이었다. 그러나 마제법이 나오연서 표면을 가공할 수 있기 때문에 곡식을 갈 때 사용하는 도구와 같이 표면이 가공된 도구가 나타날 수 있게 된 것이다. 이집트의 아스완 댐 상류 투스카(Tushka) 유척지에서 발견된 1만4500백 년 전의 음식 가공용 칼판, 갈돌 등이 이런 마제석기이다.
이 시기에는 돌 이외에도 다른 재료들이 많이 사용되었을 것으로 믿어진다. 나무는 이미 사용되고 있었을 것으로 믿어지는데 독일 북부 아렌스부르크(Ahrensburg)에서는 소나무로 된 1만 년 전의 화살이 발견되고 알래스카의 기타레로(Guitarrero) 유적지에서는 1만 년 전의 발화용 나무송우과 받첨이 출토되었다. 중국의 반포춘(半抽村) 유적지에서는 뼈낚시 바늘어 발견되고 기타레로 유적지에서는 1만 년 전의 식물섬유로 된 수공예제품이 출토되기도 한다. (2) 금속기 금속 중에서는 구리가 가장 먼저 사용된다. 구리는 지표에 노출되어 있는 초록빛 공작석을 가열하여 구리를 녹여 냈을 것이다. 구리는 비교척 낮은 섭씨 1,000도에서 녹는다. 남부 터키 차요뉴 테페시(Cayonu Tepesi) 유적지에서 기원전 7000년경의 층에서 구리제품이 출토된 바 있다. 기원천 4000년이 되연 구리제품이 중동지역에 핀, 팔찌 둥으로 널리 퍼진다. 기원천 3500년경이 되면 중동지역에서 야금솔의 획기적 발전이 이루어진다. 구리는 약하기 때문에 구리에 주석을 섞어 합금을 만들어 구리보다 3배 강한 청동올 만들어 내기 시작한다. 이 청동이 만들어지연서 청동기문화가 시작한다. 이 청동 제조술은 중국으로 전해져서 기원전 1500년청 챙동문화의 천성기를 맞는다. 중국의 상 • 은조 시대로서 이 청동으로 갱교한 종, 여러 사용기구 풍울 만들어냈다. 그리고 기현천 3000년경에는 다른 금속, 예를 들어 금이 상용되기 시작한다. 훌가리아의 바르나 유쩍지에서 기훤천 3000년경의 금제품이 발견되었다. 시대가 좁 지나면 황금제기, 술잔, 창식물 동의 유풀들이 혼하게 훌토된다. 철은 철팡석이 혼하게 발견되었지만 구리보다 녹는점이 500도나 높은 섭씨 1,500도나 되므로 사용이 늦어쳤을 것이다. 그 당시에는 1,500도의 열을 내기가 쉽지 않았기 때문이다. 이 온도이면 현대의 도자기가 구워지는 온도이다.
철은 소아시아의 히타이트(Hittite)족에 의해 처음 사용된 것으로 보인다. 이렇게 사용되기 시작한 철은 기원전 1200년경이면 철제조 기술 문제가 해결되어 기원전 1000년경이면 회랍, 이탈리아 지역에 전파되고 인도에는 이때에 이미 절좋은 강철이 사용되었다. 중국은 이보다 늦은 700년경에 사용하기 시작한다. 이 철은 청동에 비하여 강하기 때문에 철의 사용으로 문명발달이 한충 가속화된다. 철은 내구성이 강하면서 강한 험에 견멸 수 있기 때문에 철좋은 생활도구, 농경 • 수렵 도구, 무기들이 제조되고 건축물에도 많은 영향을 미치게 되었다. (3) 기계 기원전 27세기의 이집트의 피라미드, 기원천 25세기의 메소포타미아 운하들은 정확한 측량기술 없이는 불가능하였을 것이다. 이것은 이때 이미 수학 지식이 쌓여 있었다는 것을 의미한다. 그러나 기록으혹는 메소포타미아에서 기원전 2000년경의 수표(數表), 수학책 퉁이 발견되고 있다. 천체 관측도 일찍부터 시작하였으리라 보지만 기원전 1400년경에 아랍에서 만틀어진 천체관측의가 남아 있다. 기원천 5~6세기가 되면 문명이 버약적으로 발전하는 시기가 된다. 학솔, 사상, 예술, 문화에 백화가 만발하는 시대가 된다. 기훤천 6세기 중엽이연 페르시아 제국이 수립된다. 언도에서는 우파니샤드 철학이 나타나고 불교, 차이나교가 출현한다. 중국은 춘추천국시대이나 공자가 기원천 5세기에 나타나고 그 후 100여년 후 제자백가가 나타난다. 기원천 4세기에 수은 화합물올 만드는 연단술( 升術)과 칼은 화학이 나타나고 기훤천 2세기의 한(漢)제국시대에는 구창산술(九 算術)이라는 수학책이 나타난다. 그리고 그리스에서는 다른 여러 가지 학품과 더풀어 자연과학
그럽 4.4 지렛대
의 싹이 나타난다. 기원전 3세기의 아리스토텔레스가 그 전형적인 예로서 서양과학의 창시자이기도 하다. 그리스에서는 학문이 자유로이 발전하여 철학파 함째 수학이 발달하고 뿐만 아니라 천문, 의학, 박물학 들이 발전해 갔다. 서양의 자연과학은 그리스에 그 원점을 맞추고 있다.
수학의 발달은 이미 발전해 왔던 건축의 기술, 도구제작의 기술과 함쩨 기계문명의 발달로 이어진다. 기계의 가장 기본척인 단위의 하나인 지렛대는 일찍부터 사용되었을 것으로 보이나 기록은 아시리아 때부터 사용한 혼척이 었다. 이것이 생활화한 예로 지렛대의 볍칙을 세운 그리스의 아르키메데스(Archimedes, B.C. 287~212)는 〈나에게 셜 수 있는 발판올 준다면 지구를 움칙여 보이리라〉라고 할 만큼 지렛대의 위력을 찰 알고 있었다. 그가 만든 투석기는 로마외의 전쟁에서 위력을 발휘해 이것으혹 그는 결국 로마 병사로부터 화를 업게 된다. 그는 이 밖에도 기하학, 물리학의 기초률 세우는 데 많은 공헌을 하였고 스크루를 이용하여 풀용 퍼 융리는 기구률 만드는 퉁 스크루에 판한 연구도 많이 했다. 기계에 판한 가창 오래된 교과서척 저서는 알렉산드리아의 파학자 헤론(Heron, B.C. 100년경)의 저서이다. 그는 다섯 개의 단일 기계 즉, 지햇대, 촉바퀴, 쐐기, 나사, 활차에 판한 역학을 저술하였던 것이다. 기쩨 이혼의 기초률 세우는 데 공헌한 것이다. 요즈음은 냐아가 용수철, 렁크, 나사 등 다양한 단일 기계 또는 부품이 나타나 기계를 다양하게 사용될 수 있도록 해주고 있다. 이런 부풍은 철과 같이 마모성이 척고 강한 힘에 견딜 수 있는 재료를 사용할 수 있게 되고부터인 것은 두말할 필요가 없겠다. 기계는 이 시대의 기초를 발판으로 발달하기 시작하여 14~16세기의 르네상스 때의 과학의 발달, 18세기의 산업혁명을 통해 기계문명의 꽃이 핀다.
1.2. 인체보철 (1) 의학의 발달 옛날에는 생명을 신성하고 초자연적인 것과 연관을 지어 왔었다. 그것이 영(靈), 기(氣) 등의 개념과 결부되어 생명이란 영이라든가 기가 있어 이것으로부터 생명이 탄생된다고 생각되었다. 이런 생각은 현대에까지도 면면히 이어져 내려와 신학의 발달로 이어지고 신의 생명창조셜에 이어진다. 그러나 중세 자연과학의 발달로 생명을 좀더 구체척인 언어로 셜명할 수 있게 되어 생명도 물질의 특수한 구조에 불과하다는 생명기계론이 대두되고 요죠음은 주로 이 설이 지배척이다. 이 설에 의하면 간단한 물질에서 복잡한 물질이 형성되고 이 복잡한 물질 중의 하나가 생명현상을 지닌다. 이런 물철에서 단세포생물이 생기고 이 단세포생물에서 다세포생물이 생기며 사람도 대단히 다양한 기능을 가진 다세포생물 중의 하나이다. 사랍은 약 60조 개의 세포로 이루어져 있는데 세포들이 모여 심창이라든지 허파라든지 하는 특수한 기놓을 가진 장기가 된다. 사랍은 이련 장기들의 정합체이다. 이들 세포와 창기는 다른 세포, 다른 창기와 휴기척언 판계를 가지고 사람이라는 개체를 형성시키고 있으나 , 안쩨는 기본쩍우로 이련 세포, 장기틀로 구성되어 었다.옛날에는 인체에 이상이 나타나연 초자연적인 힘으로 치유하려고 하였다. 그러나 그런 방법이 한계가 있고 점차 인체의 구조가 밝혀지면서 인체를 다루는 의학이 발달하게 된다.
여명 단계 : 기원전 1700년경 이집트에 머리에서 발끝까지 언체구조를 설명한 문헌이 나타난다. 여기에는 성장, 뼈, 뇌, 위, 간장, 지라, 방광, 지궁 동을 상형문자로 나타내고 있다. 이런 문헌은 현존하는가장 오래된 외과 문헌인데 이접트는 미이라를 만들기 위하여서도 외과 지식이 발전하였던 것으로 볼 수 있다. 기원전 5세기 그리스에 오연 의학에 관한 업척이 많아진다. 근대의학의 아버지로 추앙받는 히포크라테스(Hippocrates, B.C. 460~337)는 『히포크라테스 전칩(Corpus Hippocrates)』이라는 저서를 남겼는데 여기에 두개골 외파, 백내장 수술, 늑막염 진단법 둥 수많은 업척을 남기고 있다. 그리고 그는 의료언의 윤리를 강죠하였는데 이것이 지금까지도 전해져 의사들은 히포크라테스 선서를 의료윤리의 지침서로 존중하고 있다. 여러 푸분에 업척을 남긴 아리스토텔레스(Aristoteles, B.C. 384~322)도 의학에 판한 저서를 남겼다. 중국에서는 한제국(B.C. 202~ A.D. 220)때 후세에 추앙받는 의료인 현작이 나왔으며 의학서적으로 『복새론(復塞 )』,『황제내경(皇帝內 )』등이 발간되었다. 명제국 (A.D. 14~17세기) 때는 병 치료률 위한 경락도(經絡圖)를 남기고 있는데 이때 이미 청술이 발달된 것으후 생각된다. 로마시대에는 그리스시대의 의학올 이어받아 한총 발달한다. 2세기 시대의 갈레노스(A. D. 128~199)는 소아시아 출신의 그리스인으로서 황제 안토니우스의 시의로 125권의 방대한 의학서척올 저술하였다. 그의 사람 골척, 동물 내장에 판한 업척은 오늘날에도 그대로 사용되고 있다. 혜후학, 생리학의 기초률 닦은 업척들이었다. 근대의학 : 현대의 찬란한 의학은 16세기부터 시작된다. 하버(William 足
足
그림 4.5 중국의 초기 경락도
Harvey, 1579~1657)는 「동물의 심장과 혈액의 운동에 관하여」라는 논품을 통해서 체내에는 혈액이 있고 이 혈액이 왜쇄회로툴 풍혜 순환한다는 혈액순환론을 정립하였다. 인체구조와 순환기계통에 대한 커다란 업척이었다. 베살리우스(Andreas Vesalius , 1514~1564)는 멜기에 의사로서 『인체구조에 대하여』라는 최초의 정확한 해푸학 쿄과서를 저술하였는데 근대의학의 여명을 연 저서이다. 여기셔 그는 뼈, 판철, 혈판계,신경계, 소화계, 심장, 폐창 및 뇌로 나누어 기술하고 있는데 언체의 이러한 푼류는 지금도 거의 변함없이 그대로 사용펀다. 또 한환 스위스 태쟁의 파라켈수스는 엄상에 많은 공헌을 남긴 사람으로서 광산헤셔의 직업병을 규명하였고 마취법을 창시하는 퉁 곤대의술의 선구자였다.
(2) 인공장기 인채보철이란 인체의 장기들에 이상이 생겼을 떼 이들 장기를 언공적인 물질 또는 기계로 대치하는 것을 의미하는데 이때의 인공장기는 신체의 원래 장기와 꼭 같은 모습을 하여 체내에 심어질 수도 있으나 그렇지 않고 체외에 설치될 수도 있다. 예로서 심장의 경우 전자로 시술되고 있으나 신장부전으로 인한 신장기능 회복은 후자로 처리되고 있다.
인공장기는 인체의 장기들이 구성물질이 다른 물질로 만들어진 기계를 말하는데 이 인공창기는 미묘하고 복잡한 장기들과 똑같이 만들기도 어렵지만 외부물절비 들어왔을 때 인체에서는 거부반응을 일으키는데 이 거부반응을 해결하는 것이 커다란 문제로 남아 있다. 이러한 어려움에도 불구하고 인공장기는 의학의 발달과 도구의 발달로 계속 발전하고 있는데, 언공장기는 20세기에 틀어와 비로소 실용화되기 시작한다. 이때까지는 나무로 된 의족 정도가 고작이었다. 그러나 1950년대에 들어서면서 신장, 치아, 심장, 엉덩이관철, 무릎관절 등에 인공장기다운 것이 나오기 시작하였고 얼굴, 귀, 유방 동의 미용수술이 행해지기 시작하였다. 언공장기의 효시는 1950년대의 인공신장이라고 할 수 있다. 인공신장은 피에서 노폐물을 결러내는 것으로서 기구가 커서 체외에 셜치한다. 1940년대 스웨덴의 알월(N. Alwall)과 네렬란드의 콜프(W. J.Kolff)는 각각 신장이 나빡쳤을 때 신장을 대치할 수 있는 인공신장기를 설계하였다. 그러나 초기의 기계는 크고 거추장스러웠다. 1959년 미국의 스크립너(B. H. Scribner)는 기능이 좀더 좋고 사용하기에 편리한 인공신장기를 개발하였는데 요즈옴에는 점점 발달하여 거의 가 지고 다닐 수 있는 언공신창기가 만들어지고 었다. 언풍섬창 : 1967년 남아프리카의 외과의사 버너드(Christian Barnard)는 다른 사람의 심장으후 섬창이식 수술에 성공하여 일약 유명해진 얼이 있었다. 이후 심장이식 수술은 세계 각처에서 이루어지고 있다. 그러나 이련 자연척언 심장훌 구하기가 어렵기 혜문에 인공섬장에 대한 연구도
그링 4.6 서울대학교에서 개발한 인공심장
병행되어 왔다. 특히 자빅과 콜프(R. Jarvik, W. J. Kolff)가 개발한 자빅 7이 유타대학 의료 센터에서 1982년 데브리스(William DeVrise)가 클라크(Barney Clark)에 시술하여 113일이나 생명을 연장시켜 세인의 관심을 끌었던 일이 있었다. 지금은 생존기간이 620일까지 연장되었고 세계 여러 콧에서 수십 명의 인공심장 이식 수술이행해쳤으며 한국도 서울대학교 의과대학에서 인공섬장을 만들어 송아지에 이식시켜 성공하였다.
인공관절 : 엉덩이, 무릎관절은 인공장기 시술에 가장 성공척인 경우의 하나이다. 엉덩이관절은 1950년대 영국의 찬리(J. Charnley)에 의해 개발되었는데 이 관절은 대단히 많은 힘을 이겨내야 하고 정밀하여야 하기 때문에 어려움이 많다. 그러나 점차 이련 어려웅을 해결하여 지금은 성공적 인공장기로 대두되고 있다. 이런 관절의 성공은 지체로 발천되어 간다. 예로서 팔이 손삼되었을 때 인공팔을 만드는 것도 어렵지만 이련 팔을 머리의 명령에 따라 움칙일 수 있게 하는 것이 문제이다. 그러나 머리의 운동명령은 근육의 전기
그림 4.7 인공팔 · 인공관절
를 측정하여 해독할 수 있게 됨에 따라 인공활을 직접 근육에 부착시켜 자연척인 팔과 같이 움직이도록 하는 데 성공하고 있다.
인공치아 : 인공치아는 1978년에 소개되었는데 그 후 급격한 속도로 보급되고 있는 실정이다. 언공치아는 먼저 턱뼈에 구멍을 뚫고 티타늄 합금으로 된 기둥을 끼워 뼈 골절과 융합시킨 후 티타늄 치아를 끼워넣는 것으로 이물질감이 없고 영구적인 것이어서 언기가 높아가고 있다고 한다. 이 언공치아는 앞으로 더욱 설용화되고 보현화될 것으로 보고 있다. 인공와우 : 귀는 외부의 공기 진동이 고막을 통해 감용된 뒤에 달팽이관을 풍해 청각신경으로 전달된다. 이 달팽이관에 있는 렬이 마모되거나
그림 4.8 인공치아
병이나 약으로 상실되면 청력올 잃게 된다. 이 손상된 달팽이판어 인공외우로 대치되고 있다. 이 인공와우는 1957년 프랑스에서 처음 연구가 시작되어 1970년대부터 임상에 적용되어 이제는 안전하게 실용화되고 있다. 우리나라는 1988년에 서울대학교와 연세대학교에서 시술에 성공한 바 있다.
1.3. 인체기능의 향상 인간의 지능이 발달하면서 도구률 사용하게 되고 이 도구의 사용으로 인체기능은 여러 면에서 엄청나게 향상되었다. 몇 가지 예훌 들어보자.달리기 : 달리기는 인체기능 중 필수적 기본기능이다. 이 달리기가 도구를 사용함으로써 얼마나 향상되었는가? 사람, 동물, 기계의 기록을 비교해 보자. 사람(올렴픽 기록) 100m 기록 Carl Lewis, 9초 92(시속 36.3km) 마라톤 Walemer Cierpinski, 2시간 9분 55초(시속 19.4 km) 동물 경주마 시속 69.6km 치타 시속 96~101km 송골매 시속 350km (급강하시) 기계 부산-장숭포간 쾌속정 시속 58km 프랑스 고속 철도 TGV 시속 515.3km Blackbird SR-71A 시속 2,090km 우주선 APOLLO 10호 시속 44,623km 이상으후 볼 때 사랍은 도구를 이용하게 되면서 2,000배 이상의 능력올 향상시키게 되었다.
먼지지 : 다음에는 던지기의 경우를 들어 보자. 사람(옵렴픽 기록) 투포환 Ulf Timmerman, 22.47m 투창 Miklos Nemeth, 94.58m 투해머 Serguei Litvinov, 84.90m 14Kg 우게공 먼지기 그랜 고브스, 122.75m(기네스 기록) 석궁 화찰 혜리 드레이크 ,1,853m(기네스 기록) 기채 175mm 자주포 M107 66.7kg 포탄올 32,700m Harp포 83.9kg 포탄율 179.9Km(기네스 기록)이 기록을 비교해 볼 예 던지기의 기능이 2,000배 정도 향상되었음을 알 수 있다.
드는 힘 : 드는 힘은 이보다 더 향상되고 있다. 사람(올림팩 기록) 용상 Pavel Kouznetsov, 230kg 크레인 네덜란드, 3,360ton 운반 보잉 B-52H 220ton 이 경우 보잉 B-52H 비행기는 사람의 1,000배이고 네럴란드에 있는 크레인은 15,000배 가량 힘을 향상시켰다. 이러한 기본적인 운동기능 외에도 여러 가지 능력을 향상시키고 있다. 잠수능력은 기네스 기록이 1959년 미국의 로버트 포스트가 3m 물속에서 세운 13분 42.5초이다. 이에 비하여 잠수정 기록은 1960맨 미국의 트리에시터호 잠수정이 10,848m를 잠수한 기록이 었다. 그리고 체공능력도 향상되어 사람이 높이 휠 수 있는 능력은 기껏 2.38m이다. 그러나 Apollo 10호 우주선은 720,000km 상공에서 지구를 선회하고, 사람은 공기 없는 콧에서 1분도 살지 못하지만 소련의 아나톨리 베헤소보이는 소유죠 11호에서 1982년 211일간 체공한 기록을 가지고 있다. 이것은 언간의 최장 잠수기록의 30,000 배에 해당한다. 시각놓력도 언체의 시각능력에 비하면 엄청나게 향상되고 있다. 눔의 분해능력은 0.1mm언데 광학현미경은 μm 단위의 물체에 대한 훈별력이 었다. 눈의 몇 백 내지 몇 천 배의 기능향상이다. 그리고 요즈옴 천자현며경은 nm단위의 물체를 분별할 수 있어 눈의 100,000배 농력융 가지고 있다. 엄청난 향상이 아닐 수 없다. 기네스북 기록으혹 1953년생언 톡얼의 베로니카 자어더는 1.6km 거리의 사람을 식별할 수 있다. 그러나요사이 발달한 여러 가지 광학망원경, 전자망원경은 거의 우주 끝까지 보고 있다. 자이더의 능력에 비유가 안 될 만큼 무한에 가깝게 능력이 향상되었다. 눈이 가지고 있지 않은 능력도 가지게 되었다. 인공위성에서 적외선, 자외선 등 여러 도구를 이용하여 지하 수자원 등자원을 찾는다든지 지구를 태양의 자외선으로부터 보호해 주는 오존층을 확인하는 것이라든지 이런 것은 맨눈으로 볼수 없는 것엄에도 불구하고 도구를 이용함으로써 가능하게 되었다.
2. 뇌의 인공적 확장은 가능한가 2.1. 신경보철 : 뇌의 인공척 확장은 시작되다몸을 통제하는 방법은 두 가지가 있는데 하나는 화학적 물질, 즉 호르몬을 이용한 방법이고 다른 하나는 신경계를 이용하는 방법이다. 신경계는 폼 전체의 곳곳에 퍼져 있으며 신경세포, 신경섬유로 이루어진 복잡한 망이다. 신경은 우리가 장을 자고 있든 깨어 있든 어떤 상태이든 옴의 모든 상태를 중계하고 통제한다. 심장의 박동, 외부기온에 반웅하는 땅, 의식 풍 몸의 모든 부환에 대한 상태와 각종 활동을 계, 홍제한다. 신경계는 중추신경계와 말초신경계로 구성되어 었다. 중추신경계는 뇌와 척수로 이루어져 있으며 말초신경계로부터 들어오는 정보들을 처리하여 다시 말초신경계로 전달하는 기능올 담당한다. 말초신경계는 몸의 곳곳에 퍼져 있어서 감각 수용기로부터 외부자극에 대한 청보를 받아 척수훌 거쳐 뇌로 천랄한다. 그리고 뇌로부터의 명령이 척수를 통해 발초신경으후 천발되고 최종척으로 근육에서 운동으후 나타난다. 신갱써포는 앞에서 보아 온 바와 같이 축색으로 다른 신경세포, 수용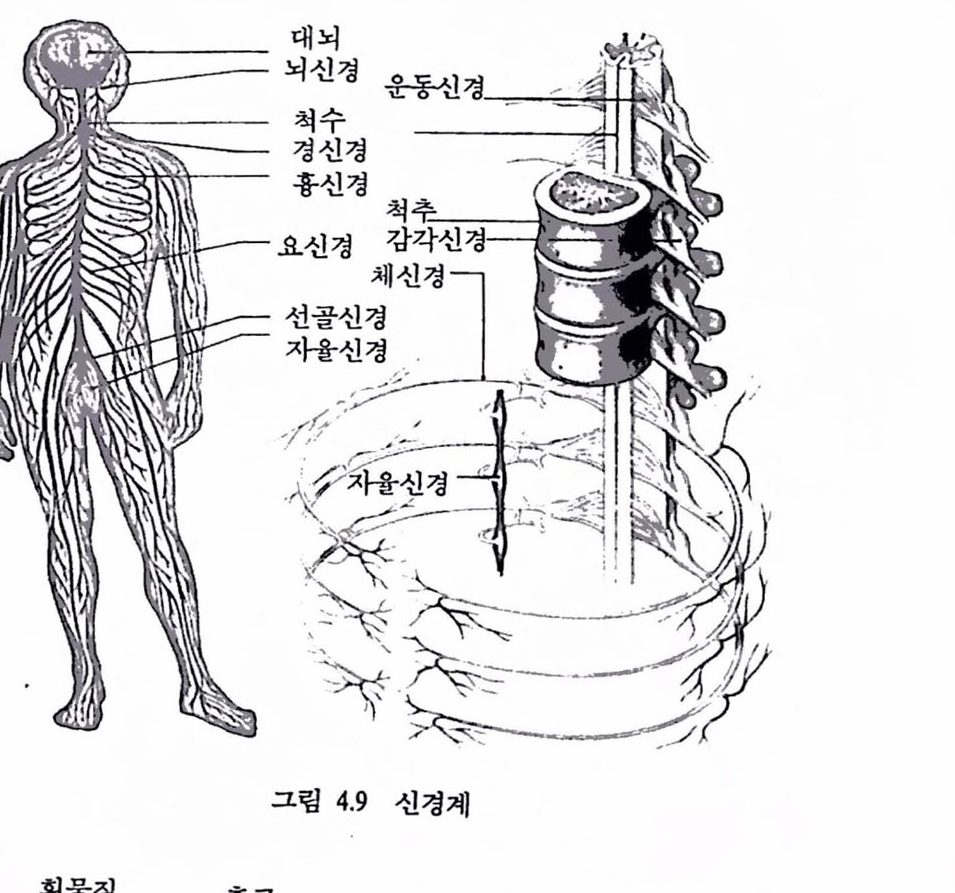 대뇌
대뇌
그립 4.9 신경계
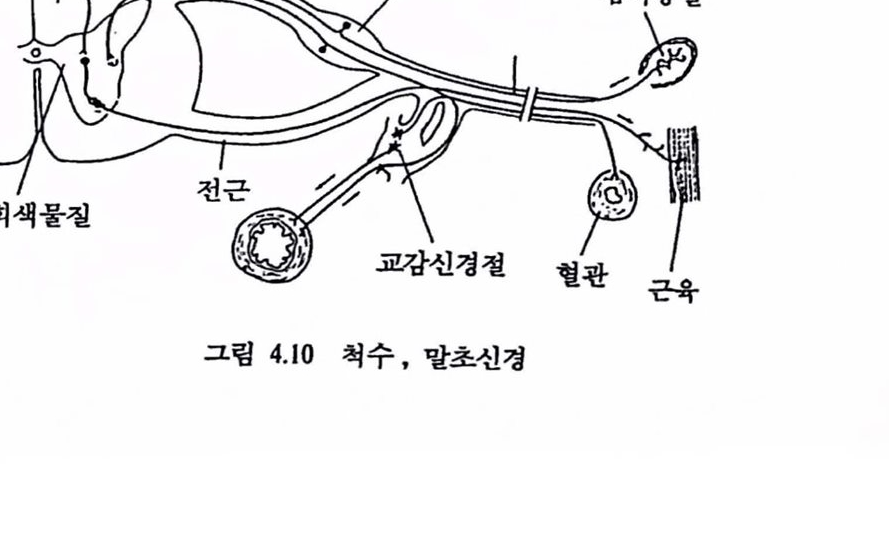 흰물질 후근
흰물질 후근
그립 4.10 척수, 말초신경
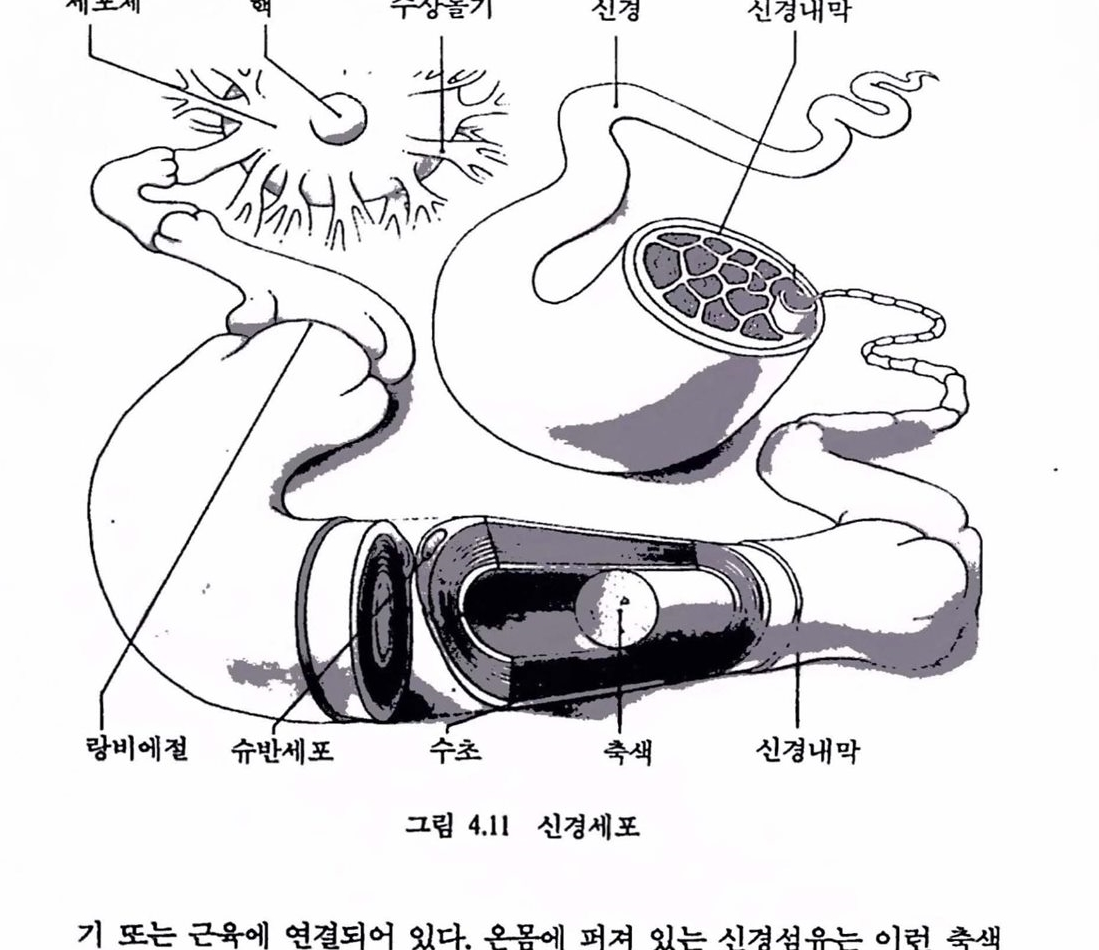 신경 신경내막
신경 신경내막
그림 4.11 신경세포
기 또는 곤육에 연결되어 있다. 온옴에 퍼져 있는 신경섬유는 이런 축색의 묶음이다. 그렴 4.11에서 보는 바와 같이 축색의 묶음이 하나의 신경생유로 묶어져 었다.
신경성휴는 굵용수륙 정보전달 속도가 훨라 좋지만 그렇다고 수많은 신경생유가 모두 굵윷 수는 없다. 그래서 그 굵기와 길이는 일정치 않다. 굵기는 수 에서 오징어의 촉색과 같이 1mm나 되는 거대 촉색도 었다. 신경성유는 천솔한 바와 같이 많은 촉잭이 묶어져 있는데 예를 률면 확우시신경의 청우 100만 개의 혹색으로 구성되어 있으며 그 굵기는 4mm 청도 된다.감각수용기는 신경세포종말로써 폼의 구석구석에서 일어나는 각종 상태를 중계하는 역할을 하는데 그 상태를 전기신호로 정보화한다. 이 신경세포종말은 기능과 위치에 따라 각각 다르며 그렴 4.12에서 보는 바와 같이 다양하다.
 <. ~메르켈 세포
<. ~메르켈 세포
그립 4.12 신경세포총멀
옴이 외부지극에 반응할 수 있는 것은 결국 아래 그렴과 칼이 이런 감각 수용기를 거쳐 취해진 정보가 신경섬유플 통해 중추신경에 전해지기 때문이다. 이때, 감각수용기, 신경섬유, 중추신경 중 어느 하나가 손상되어도 외부자극에 반웅할 수 없게 된다. 이런 손상된 부분을 보완하는 것이 신경보캘이다. 지금의 신경보철은 감각수용기를 대체하거나 운동신경의 정보를 이용하여 지체를 대체하는 정도이다.
(1) 감각신경보철(인공와우) 먼저 감각수용기에 대한 신경보철의 경우를 보자. 이것은 제2장에서 여러 가지 경우를 서술하고 있다. 그래서 여기서는 인공와우의 경우를 통해 감각신경보철의 기본원리만을 보도록 하겠다. 귀는 공기의 진동을 언식함으로써 외부자극을 받아들이는 기관이다. 그렴 4.13과 같이 공기 진동이 고막을 통해 달팽이관에 전달되어 청각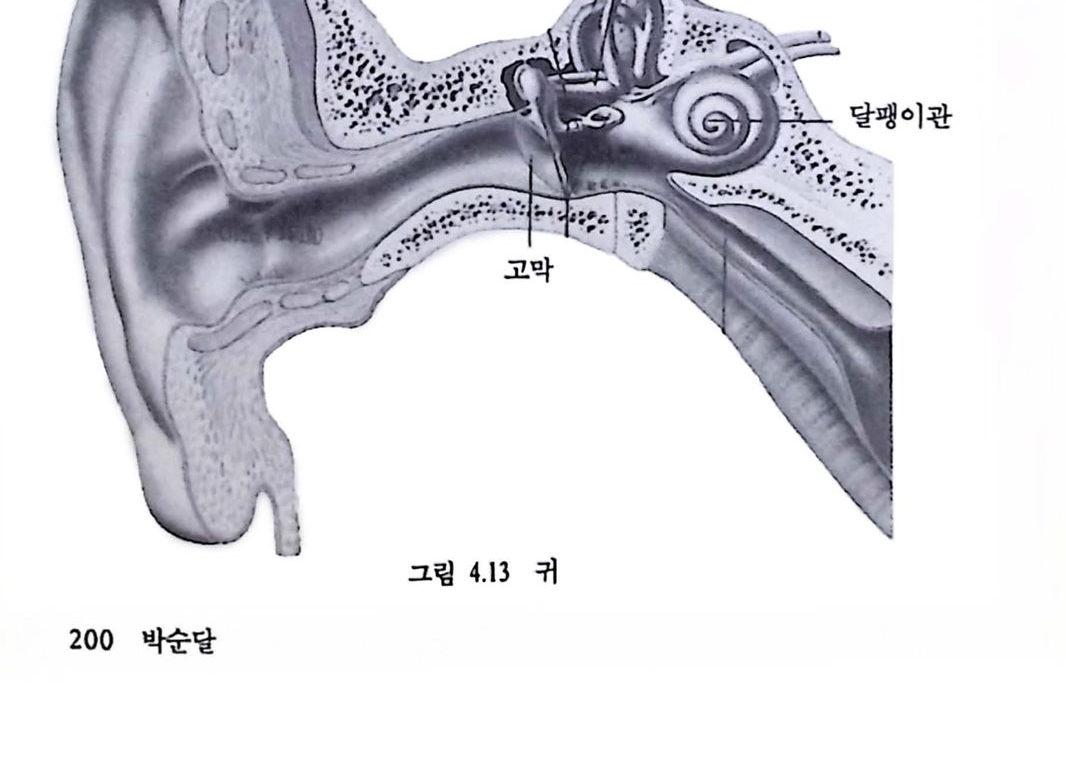
그립 4.13 귀
신경을 자극함으로써 뇌에 전달된다. 그러나 고막이 충격으로 파손되거나 달팽이(와우)관이 여러 가지 이유로 손엠 되면 공기 진동이 청각신경에 전달되지 못한다. 그러면 소리를 듣지 못하게 된다.
여기서 달팽이관의 역할이 중요한데, 달팽이(와우)관은 달팽이와 같이 꼬여 있으며 그 안은 액체로 채워져 었다. 소리가 귀로 들어와 고막을 진동시키면 그 진통이 세개의 뼈(추골, 침골, 등골)를 거쳐 난원창에 천달된다. 난원창의 진동은 전정계의 액체를 진동시키며 이것이 전청막을 진통시킨다. 이 진동은 기저막으로 전달되며 가저막에 있는 청각세포언 럴세포가 활성화된다. 그러면 진동은 이 세포에 연결되어 있는 청각세포에 전달된다. 사람의 청각세포는 28,000개 가량 었으며 이것이 연수에 연결된다. 사람은 15~2만Hz의 소리를 들을 수 있으나 1,000에서 4,00OHz 의 소리에 가장 민감하게 반응한다. 동물마다 가청주파수의 범위가 다르며 고양이는 7만Hz, 박쥐, 돌고래는 10만Hz의 소리를 들을 수 있다.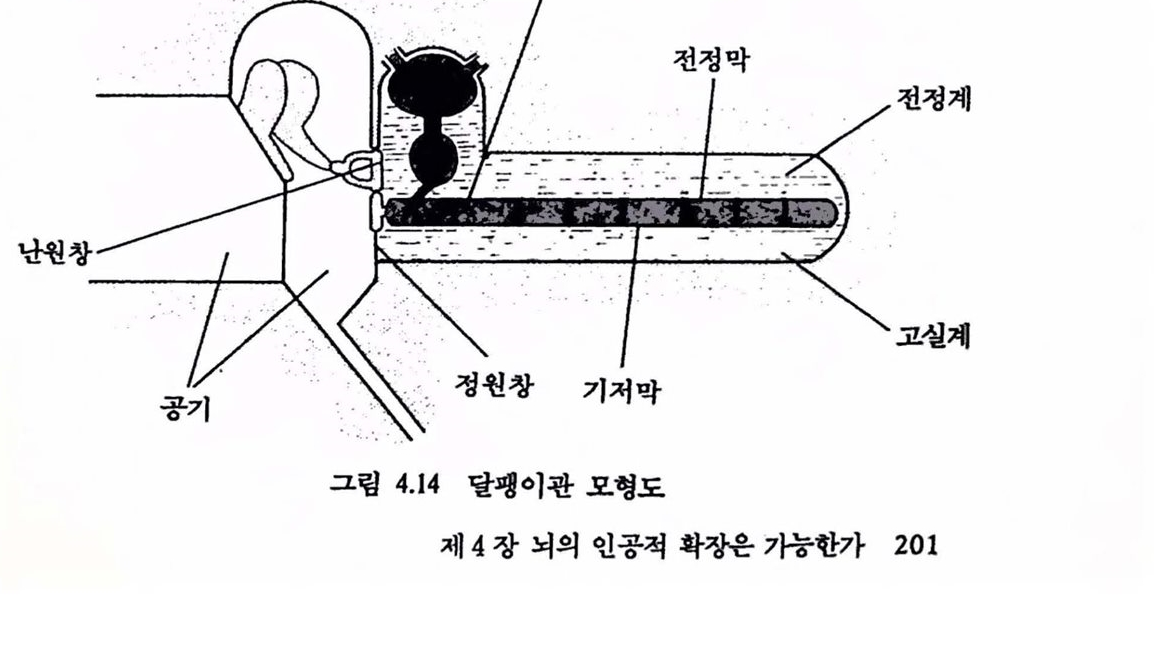 달팽이판
달팽이판
그립 4.14 달팽이판 모형도
달팽이관은 고막에 가까울수록 높은 소리에 반응하고 멸리 갈수록 낮은 소리에 반응하도록 되어 있다. 그런데 와우관에서 털세포가 약이나 다른 조건에 의해 손상되면 외부조건에 반응할 수 없으며 따라서 틀을 수 없게 된다. 물론 청신경이나 중추신경계의 손상으로 소리를 듣지 못할 수도 있다. 그러나 청신경이나 중추신경계가 그 기능을 유지하고 있을 때는 소리 수용기인 달팽이관의 기능만 회복시켜 주면 된다. 이것은 달팽이관에 있는 진동에 반응하는 털세포의 역할을 다른 것으후 대치시키는 것드로 가능하다.
이 달팽이관의 손상된 부펄을 인공와우로 대체하고자 하는 연구는 1957년 프랑스에서 시작하여 그동안 상당히 연구가 진행되다가 1960년대에 임상실험이 시작되었다. 1980년대에 와서는 미국과 호주에서 인공와우가 생산되기 시작하면서 많은 시술이 이루어져 현재까지 세계적으로 1300여 건의 시술에 수행되었다. 우리나라에서는 1988년에 연세대학교와 서울대학교에서 시술에 성공한 후 시술받은 사람이 10여 명에 달한다. 우리나라에서 시술한 인공와우는 1979년 호주의 Grame Clark이 개발한 것으로 Health Care Company Neucleus사에서 제조된 22개의 전극올 가진 싱품이다. 이것은 외부의 소리를 받아들이는 마이크로폰, 소리를 천기신호로 처리하는 말처리기, 천기신호를 보내는 우선 송신장치와 받아들이는 수신장치, 그리고 22개의 천극을 가진 아주 가는 전선으로 되어 있다. 전선은 측두골에 구멍을 뚫어 달팽이판으로 넣고 수신기는 두개꼴에 고청시킨 후 피부로 덮는다. 외푸에서 툴어온 소리는 마이크로폰으로 잡은 다음 소리조정장치로 소리의 크기훌 조청한다든지 하고 이 소리롤 전기신호로 바꾼다. 이 천기신호률 무선송신장치로 보내면 피부 밑에 섬어진 수신장치가 이 신호훌 창는다. 그러면 이 신호는 천극어 있는 전선으로 흩러가 22개의 천극에 보내진다. 이 전극에는 청작신경이 첩촉되어 있어 소리는 그 총류에 따봐 그 소리가 천혜진 전극에 첩촉된 청각신경에 전해진다. 바로 렬세포의 역할올 이 천극이 대신하고 있움을 알 수 있겠다. 이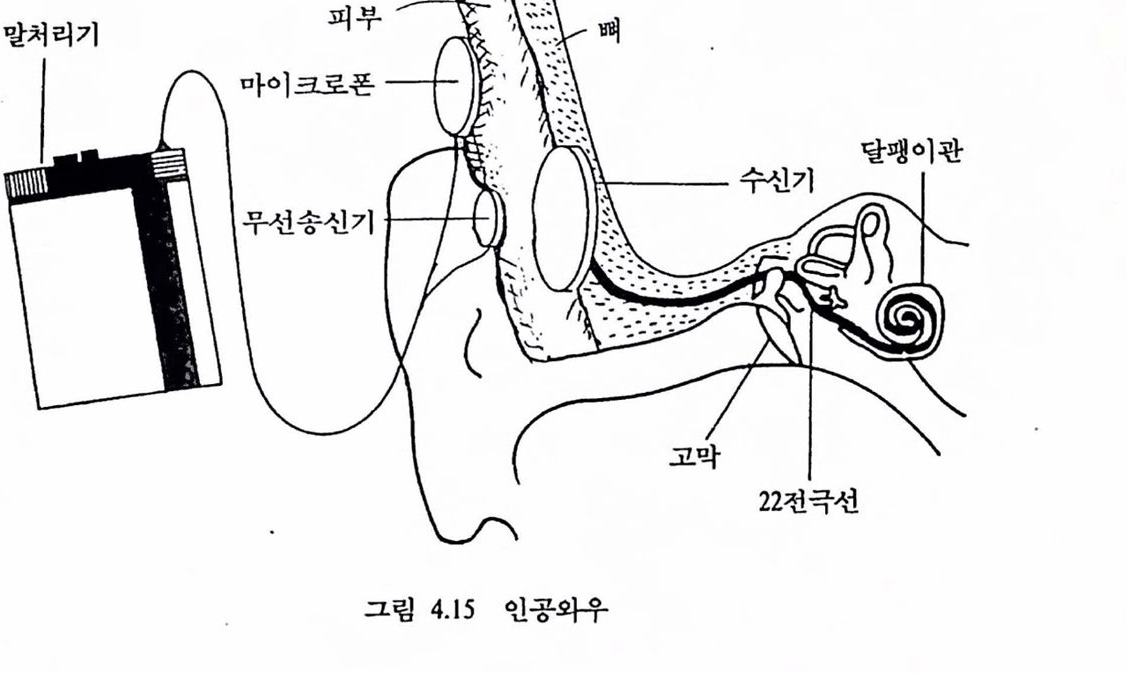 말처리기 피부
말처리기 피부
그림 4.15 인공와우
인공와우의 전극은 22개이기 때문에 22개의 다른 소리에 반웅할 뿐인데 귀의 2만여 개에 비교하면 가짓수가 척으나 훈련에 따라 일상생활에 지장이 없을 정도로 청각을 회복한다.
감각신경계에 대한 신경보철은 청각뿐만 아니라 감각신경계에서 가장 복잡한 시각에까지 이루어지고 있다. 감각신경계의 보철은 시각 분야에서 가장 먼저 시작되었는데 맹인에 대한 연구의 얼환으로 복지단체에서 처음푸터 관심이 많았다. 이 시각보철은 제2장에서 다루어진 바 있다. 감각신경은 이외에도 후각, 촉각, 미각 퉁이 었으나 시각, 청각 회복에 대한 보철이 가창 활발히 연구되고 있다. 현재 신체의 보철은 여러 장기에 실시되고 있으나 신경보철은 이제 초기 단계이다. 감각신경보철은 그렴 4.13에서 보듯이 감각수용기훌 대쩨하는 작업이다. 신경섬유나 중추신경 자쩨률 대체하는 갯온 아니다. 그러나 이 수용기는 외부와 신경올 접속시키는 정속기 또는 그렴과 갈이외부지극을 신경세포가 받아들일 수 있게 신호를 처리하는 변환기 역할을 해주는 것이다. 따라서 신경계의 중요한 대외창구인 생이다. 이 초기 단계가 신체의 각 부위와 신경의 연결 부분에서 시작되는 것은 당연해 보인다. 그것은 신체의 여러장기 중에서 신경계에 대한 연구가 가장 늦어 신경섬유, 중추신경 자체에 대한 보철은 가장 늦게 이루어질 것이기 때문이다.
(2) 운동신경계의 보철 간단한 하나의 운동이 일어나는 데도 수십 개의 근육이 서로 작용하여야 버로소 기능해진다. 따라서 운동신경을 제어하여 올바른 운동을 일어나게 하는 일은 쉽지 않다. 어떤 근육이 마비되었을 때는 먼저 운동신경을 회복시켜야 할 것이고 여의치 못하면 대체하는 수밖에 없다. 어느 것도 쉬운 일이 아니다. 그러나 최근 들어 많은 발전이 이루어지고 있다. 얼마 천 원팡대학교 의대 김상수, 박병림 교수팀은 척수결핵으로 5년간 다리가 마비되었던 사람과 2년 전 뇌졸층으로 전신마비가 된 사람의 활과 다리에 전기척 자극으후 기능을 일부 회복시킨 사례가 보고된 바 있다. 이 립은 19가닥의 전선을 마비된 근육을 지배하는 신경에 심어 강도와 횟수를 조절하면서 자극을 가한 결과 근육의 크기가 청상으후 돌아오고 경칙되었던 근육이 유연해지는 풍 성공을 거두었다. 운동닌경 회복의 좋은 예라 하겠다. 지체 마비 또는 손상으로부터 청상척언 운동으로 회복코자 하는 사람의 희망은 바로 신채의 보철로 이어져 활과 다리의 보철이 보철 역사의 시발점을 이루고 있다. 그러나 이제는 단순한 신체척 보철뿐만 아니라 운동을 지배하는 신경보철로 발전해 가고 있다. 이것은 마비로부터 정상옳 찾고 손장된 지체로푸터의 회복올 이룰 수 있기 혜문이다. 육채의 훈동은 중추신경계의 명령에 의해 조정된다. 이 명령은 운동산갱옳 흥해 근육에 천달됨으로써 이루어지는 것언데 이혜 운동신경에 호르는 신호훌 식별하여 인공합성하여 비슷한 신호를 일으킬 수 있다는것이 밝혀졌다. 1960년대부터 이러한 연구가 결실을 맺기 시작하여 대단히 빼픈 속도로 칠용화되어 가고 있다.
이 연구의 실용은 손상된 지체의 대체에 유용하게 이루어질 수 있다. 인공지체는 두 가지가 이루어져야 한다. 하나는 언간의 지체와 같이 미묘한 운동을 할 수 있는 기계를 만드는 것이고 또 하나는 이러한 인공지체를 몸에 부착시켜 중추신경계의 명령대로 움직이게 하는 작업이다. 전자의 전형적인 예가 제 1장의 그렴 1.2의 기계손이다. 후자는 제 1장에서 어떤 연구가 이루어지고 어떻게 활동되고 있는지는 충분히 보아 왔다. 결론적으로 아직 사람의 손과 같이 미묘한 움직임은 할 수 없으나 대체물로 훌륭한 손, 팔, 다리가 만들어졌고 이것이 제 1장의 그렴 1.32와 앞절 인공장기 항에서 보았듯이 폼에 연결되어 중추신경계의 명령에 의해 움직이고 있다. 이 결과는 아직 운동신경섬유플 만들거나 대체할 수 있는 상황은 아니지만 운동신경섬유에 흐르는 정보를 읽어 볼 수 있는 상태가 되었다는 것을 의미한다. 이제 연구가 조금 더 진행되면 운동신호를 합성할 수 있을 것이고 이것이 바로 언공운동신경계의 시발점이 될 것이다. (3) 신경의 연결 신경섬유는 전술한 바와 같이 수백에서 수백만 개의 축색으로 구성되어 있다. 각 축색은 신경섬유에서 나와서 수백에서 수만 개의 다른 신경섬유와 수상돌기로 연결되어 있다. 사고나 어떤 경위로 지체가 손상되었올 때이 신경섬유도 손상되게 되며 특히 신경섬유 손상의 복구가 어렵다. 축색은 하나하나가 독특한 정보를 천달하며 따라서 수많은 축잭이 끊어졌을 때 어떤 것과 어떤 것이 연결되어야 하는지 모르기 때문이다. 미국 Stanford 대학 의료센터의 Morthan Grosser와 Joseph Rosen은 취의 손상된 다리 신경섬유를 복구시키는 연구롤 거듭하였다. 끊어진 신경섬유는 서로 자라나 이어질 수는 었으나 축색들이 제멋대로 이어져 신경섬유의 역할올 제대로 하지 못한다. 즉 끊어진 신경성유 내의 수많
은 축색들이 제 축색으로 이어져야 하기 때문이다. 이 두 사람은 각 축색의 제 싹올 찾는 연구를 계속해 왔던 것이다.
두 사람은 먼저 1mmX1.5mm 크기의 IC첩의 양면 모두에 8 의 작은 구멍을 많이 뚫었다. 그리고 이 청을 끊어진 신경섬유 사이에 끼웠다. 그러면 신경섬유 양쪽 끝의 각 축은 반년 후 이 구명 사이로 지라나 IC첩의 회로에 도달하게 된다. 그리고 이 IC첩은 대형 컴퓨터에 연결되어 있어 IC청 회로에 도달한 양쪽 축색의 정보률 훈석한 끝에 절반 정도 축색의 싹을 찾을 수 있다. 보풍 신경의 신호는 복수로 전달되기 때문에 절반 정도 제 확의 축색을 찾올 수 있으면 신경으료서의 제 기능을 발휘할 수 있다. 두 사람은 이렇게 제 싹의 촉색올 찾은 다음 IC첩을 고정시킨 후 이 첩과 컴퓨터의 연철융 끊었다. 이 두 사랍은 2년여의 연구 끝에 1987년 이 연구를 성공시켰는데 이 성공은 감각수용기의 보철에 못지 않은 중요한 돗올 지난다. 즉 손상된 신청세포훌 기계로 이어줄 수 있다는 뭇이 되기 혜문이다. 지금까지는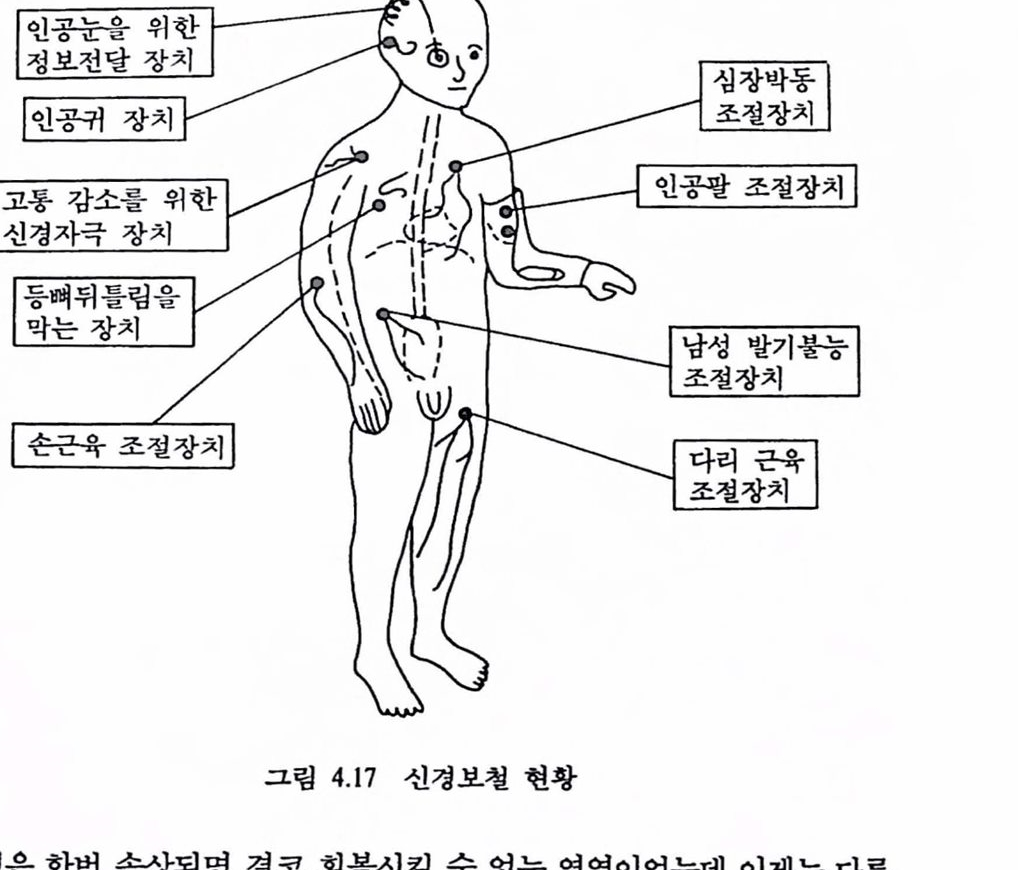 언공눈을 위한
언공눈을 위한
그림 4.17 신경보철 현황
신경은 한번 손상되면 결코 회복시킬 수 없는 영역이었는데 이제는 다른 신체부위에서와 같이 의학척 치료, 즉 보철이 가능해지게 되었다.
종래의 신체보철은 단순히 육체의 일부를 단순한 도구로 대체하는 것이다. 그러나 인공와우와 같은 기관을 대체한다는 것은 종래와는 또 다른 뭇이 있다. 즉 언공장기와 신경계를 연결시킨다는 뭇에서 종래의 보철과는 다른 신경보철의 첫걸음올 딛는다는 뭇이고 이것은 인위척언 기기가 신경에 부확되어 교신을 할 수 있다는 것올 의미한다. 운동신경계에서도 버슷한 현상이 일어나고 있다. 현재 개발중이거나 곧 사용훨 신경보철기의 상황은 그렴 4.17에서 보는 바와 같이 대양하다. 우리가 30여 년 전 섬창을 생각해 보면 그 당시는 심장제어의 초기 단체로셔ECG와같은 심장상태를 점검할 수 있는 기계도 원시적이었고 해석도 또한 마찬가지였다. 인공섬장이란 꿈같은 이야기였다. 그러나 지금은 인공섬장이 시술되고 있다. 신경보철도 역시 이런 상황과 비슷할 것으로 보고 있다. 신경계란 기본적으로 신경세포의 집합체이고 연구가 거둠됨에 따라 그 구조와 기능이 밝혀지면 곧 인공대체물이 나타나게 마련이다. 신경보철도 시간이 오래 걸리든지 또는 예상보다 짧게 걸리든지의 문제는 있을지 몰라도 불가능의 영역은 아념에 틀림없다.
2.2. 뇌 연구 의학연구는 앞 절에서 보았듯이 고대 이집트시대에서 시작하여 기원전 4세기 그리스에서 꽃이 피기 시작한다. 그러나 신경계에 관한 연구는 늦어 16세기 베살리우스의 해부학 교과서에 나타나기 시작하며 실질적인 연구는 19세기에 들어오면서 시작된다. 그 전까지는16세기 프랑스의 Jacobus Sylvius의 중뇌에 관한 연구, 이탈리아의 Bartolomeo Eusachi의 청각에 관한 연구, 17세기 프량스의 Francis D. Sylvius의 대뇌에 관한 최초의 기록, 18세기 스코틀랜드의 Alexander Monro 의 신경계에 관한 연구, 오스트리아의 Johann L. Gasser의 감각신경절에 관한 연구, 이탈리아의 Francesco Gennari의 시각피절에 관한 연구, Luigi Rolando의 대뇌피질에 관한연구 등 부분적인 연구가 행해져왔다. 그러다가 19세기에 들어오면서 중추신경계, 말초신경계, 감각수용기에 대한 기초가 확럽되기 시작한다. 특히 말초신경계의 구조에 대해서는 염색법의 개발로 많은 연구가 이루어져 그 구조가대부분 확렵되었다. 독얼의 Theodor Schwann(1810~1882), Ernst Reissner(1824~1878), Friedrich Henle(1809~1885), 이탈리아의 Angelo Ruffini(1864~1929), Vittoria Mazzoni 등이 특히 공헌이 많으며 이들과 이외의 여러 사람들이 그들의 연구로 말초신경 구조에 이륨을 남기고 있다.중추신경계에 대한 구조, 기능에 대해서도 Chales Bell(1774~1842)이 감각신경과 운동신경의 발견 등 이 시기에 많은 부분이 밝혀졌다. 나아가 Richard Heschel(1824~1881)의 청각령에 관한 획기적 연구, Pierre P.Broca(1824~1880), Carl Wernicke(1848~1905)의 언어와 말에 관련된 감각령의 발견. Vladimir A. Betz(1834~1894), David Ferrier(1843~1928)의 대뇌 감각, 운동령에 관한 연구 등으로 대뇌의 구조와 각 부분의 기능에 관해 상당히 많이 알게 되어 뇌 연구의 획기적 발전을 이룩하였다.
19세기가 신경계의 구조와 기능에 대한 기초가 확립되기 시작한 시기라면 20세기는 신경계에 관한 이론이 정립되는 시기라고도 할 수 있다. 영국의 Charles S. Sherrington(1856~1952)의 신경계 기본 지식의 확립을 위시하여 프랑스의 Claude Bernard(1813~1878)는 생리학의 기초를 확럽하여 그 공로로 1906년에 노벨상을 받았다. Santiago Ramony Cajal은 Heinrich Waldeyer(1836~1921)와 함께 신경계의 기초가 되는 신경세포이흔을 확럽시켰다. 더욱이 이 20때기는 각종 방법론의 발달로 신경계의 구조와 기능에 관해 넓고 깊게 연구가 진행되고 있다. 그래서 이 세기가 지나가기 전에 지금까지의 단순한 탐구를 벗어나 신경계의 보철, 나아가 신경의 기계에 의한 보완도 고려될 만큼 연구가 진척되고 있다. 예로서 1960년대에 S.W.Kuffler, D.H. Hubel, T.N. Wiessel은 고양이의 망막을 재료로 시각피질의 시각정보 처리과정을 연구하였는데 이 연구로 Hubel과 Weissel은 1981년 노벨상을 받았다. 이플은 연구는 단순한 연구로만 끝나지 않고 인공눈의 연구로 이어지고 있다(제2창 참조). (1) 방법론의 발달 현미경을 이용한 방법 우리가 본다는 것은 빛을 이용한 것으로서 광선이 물체에 반사되는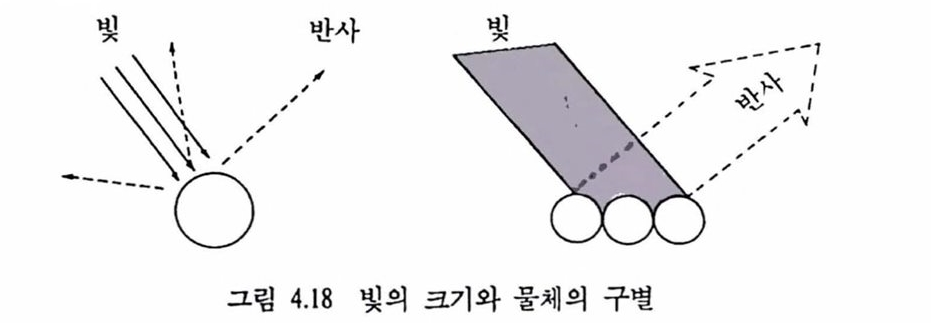 버 반사 1,.t.l; / -/-‘//r-v /Y-‘-x、/-‘ ’l’vl카
버 반사 1,.t.l; / -/-‘//r-v /Y-‘-x、/-‘ ’l’vl카
그림 4.18 빛의 크기와 물체의 구별
것을 눈이 인지함으로써 물체의 크기와 모양을 인식한다. 물체를 인지하는 능력 즉 분해력은 물체를 비치는 도구, 예로서 빛의 특성과 사람의 눈의 능력에 달려 있다. 만일 빛의 굵기가 주먹만큼 크다면 주먹보다 작은 자갈들은 구멸하지 못한다. 왜냐하면 자갈밭에 주먹만한 빛을 쏘이연 자갈 하나하나에 반사하는 것이 아니라 자갈 여러 개에 대해 반사하므로 자갈 낱낱을 구별하지 못하게 된다. 반면 그렴 4.18의 왼쪽과 같이 자갈보다 빛의 굵기가 작으면 빛이 자갈에 닿았을 때 그 표면에 따라 그 방향으로 반사하므로 그 반사빛을 눈이 받아 자갈의 모습을 그릴 수 있게 된다.
한편 빛의 크기가 물체보다 작아 물체의 모습에 따라 빛이 반사하여 그 반사빛은 물체의 낱낱을 구멸하더라도 우리의 눈이 그 구별을 인식하지 못하면 물체의 낱낱을 구멸할 수 없게 된다. 우리의 눈은 렌즈 역할을 하는 수정체를 통해 들어오는 빛을 망막으로 받아 그 성을 인식하게 된다. 이 망막에 정확히 초점이 맞추어질수록 또렷이 볼 수 있다. 그렇다고 무한히 작은 것을 인식할 수는 없다. 이것을 눈의 분해능력이라고 하는데 우리의 눈은 대략 O.1mm 크기의 물체까지 볼 수 있다고 한다. 광학현미경 : 빛을 이용하여 사물을 보는 보조 도구로 구리거울의 역사는 오래다. 우리나라의 삼국시대 유물에 구리거울이 나타나며 고대 이집트는 기원전 900년의 세소스트리트 2세의 피라빗 유울에 구리거울이 나타난다. 유리라든지 물을 이용하여 빛을 모을 수 있다는 생각은 기원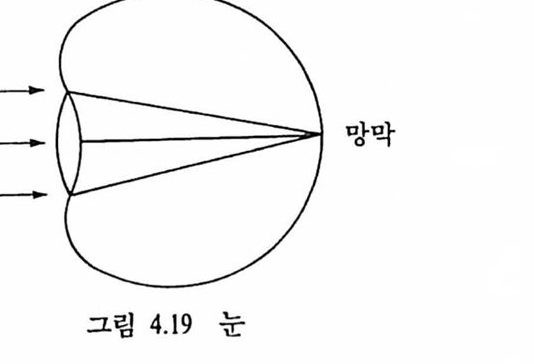 ..
..
그럼 4.19 눈
전 5세기부터 있었다. 유물로는 로마시대 폐허에 발화용 볼록 렌즈가 발견된다.
13세기의 과학자 Franciscan R. Bacon은 오목 렌즈와 볼록 렌즈를 이용한 망원경에 관한 연구를 하였고 드디어 1608년 Hans Lippershey(1587~1619)가 망원경을 발견한다. 비슷한 시기에 Galileo Galilei(1564~1642)도 망원경을 발명한다. 그러다가 네멀란드 사람인 Zacharias Jasen(1588~1632)이 현미경을 발명하고 Francisco Fontana (1580~1652)가 개량하였다. 이 현미경을 이용하여 1652년 Antoni van Leeuwenhoek(1632~1723)가 여태까지 볼 수 없었던 미생물을 발견하게 된다. 그때까지는 생물은 사람의 눈으로 확인될 수 있는 것만으로 생각했으나 그 후 사람의 눈으로 볼수 없는 생물도 많다는 사질을 알게 된 것이다. 이 오목 렌즈와 볼록 렌즈를 이용한 현미경은 물체를 구별하는 도구로 빛을 이용하기 때문에 광학현미경이라고 한다. 이 광학현미경은 빛을 이용한다는 점에서 눈과 같지만 그 분해능력은 몇 천 배가 된다. 그래서 눈으로 볼 수 없는 미생물, 세포조직, 척혈구 등 작은물체를 볼 수 있게 된다. 예를 들어 사람의 세포조직은 눈으로 볼 수 없다. 그러나 현미경으로 보면 그렴 4.20과 같은 조직 구조를 볼 수 있다. 전자현미경 : 전자현미경은 사물을 구별하는 도구로 빛 대신 천자의 흐름을 이용하는 것이다. 광학현미경은 전술한 바와 같이 가시광션을
그럽 4.20 세포의 표연조직
이용하는 것인데 백색광선의 파장은 0.55μm(5,500A) 이다. 이 가시광선을 이용할 경우 이 파장 길이만큼의 크기를 가진 물체를 구별할 수 있다. 그래서 예를 들어 미생물보다 작은 바이러스라든지 세포핵의 구조 등은 광학현미경으로서는 볼 수 없다. 이런 결점을 보완하기 위하여 만들어진 것이 전자현미경이다.
전자현미경은 자연광선이나 자외선 대신에 전자선을 이용한다. 전자선의 파장은 이 전자션을 가속시키는 전압의 제곱에 반버례하는데 전압을 높게 하면 할수록 파장이 짧아진다. 그래서 5~6만V의 전압으후 가속시키연 전자선의 파장은 0.05A이 된다. 전술된 바와 같이 빛의 파장이 짧을수록 분해력이 커지므로 이 천자선의 파장을 짧게 하는 것이 좋다. 이론척으로는 전압을 높이면 이 파장을 더욱 짧게 할 수 있다. 그러나 빛의 파장은 아우리 짧게 하더라도 다른 기술이 동시에 발전해야 하므로 제한을 받게 된다. 그리고 팡학현미경이 빛을 접속하기 위하여 석영렌즈를 사용하는 대신 전자현미경은 전자석으로 된 전자 렌즈를사용한다. 천자선은 자력에 굴철하기 때문에 이 원리를 이용하여 전자선을 집속시키는 것이다. 이 전자현미경은 독일 빼률련 공과대학의 Ernst Ruska 가 연구하여Siemens사와의 공동제작으로 1983년 담배모자이크병의 바이러스(크기 300nmX18nm)의 사진을 찍는 데 성공하여 현미경 억사에 일대 전환의 계기를 만들었다. 이 전자현미경은 이론적으로 8A의 분해력을 가질 수 있으나 기술상 제약이 있어 분해력 향상에 어려움이 있어 왔다. 그러나 1960년대에 이르면 분해력이 5A에 달하게 된다. 이 크기는 구리의 원자 크기에 해당하는 것이다.
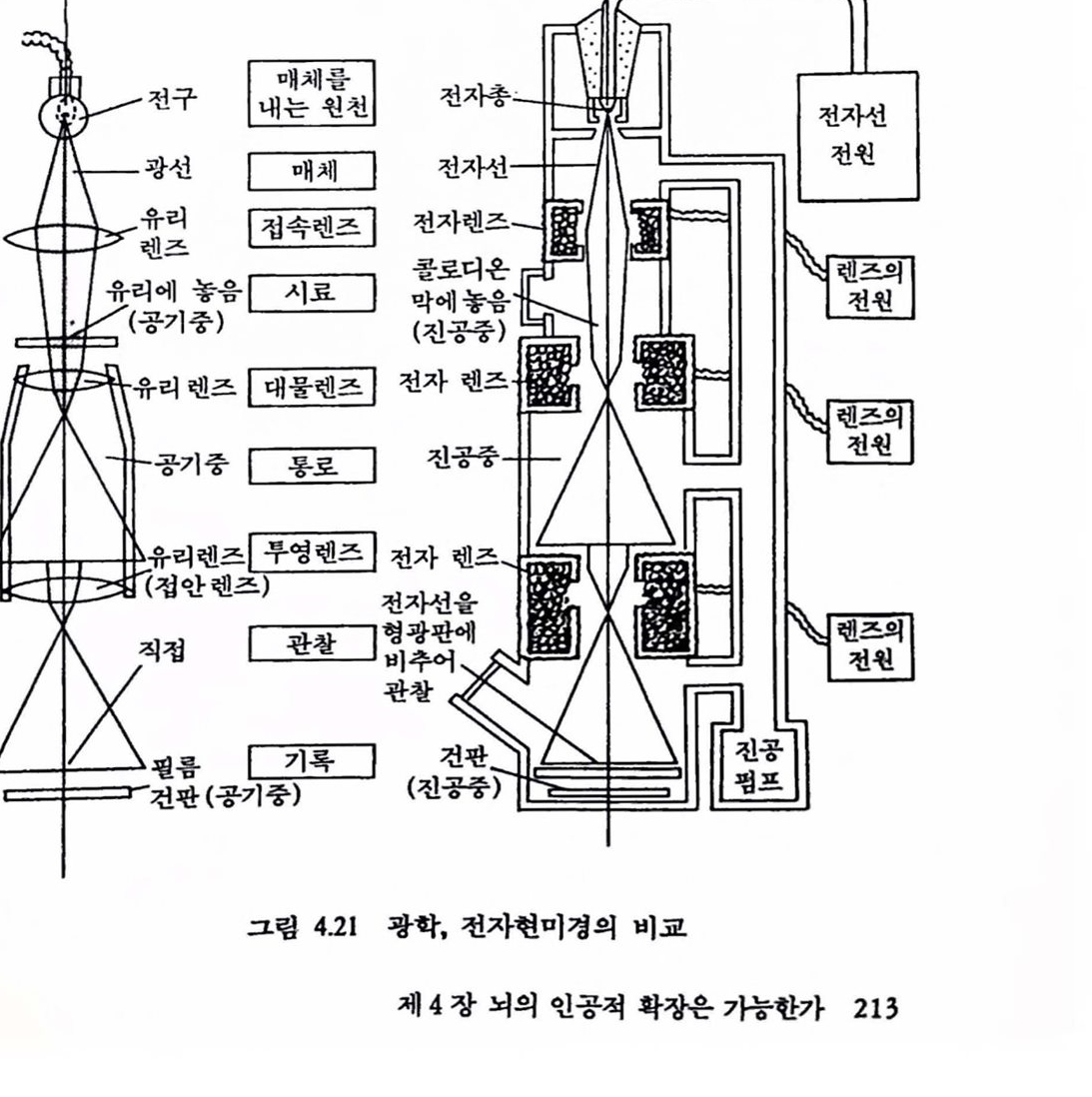 광학현미경 전자현미경
광학현미경 전자현미경
그림 4.21 광학, 천자현미경의 비교
1980년대에 이르러 이 전자현미경의 연구는 가일층 보완되어 게르트비니히는 Stanford 대학에서 STM, AFM 방식의 전자현미경을 제안하였다. STM(Scanning Tunneling Miascope)은 진공 속에 금속 탐칭과 시료 사이의 거리를 5A 정도 두고 양쪽에 전압을 걸면 전자가 시료 쪽으로 흘러가 Tunneling 현상이 일어나는데 이때 시료를 움직이면 그 굴곡을 알 수 있다. 그러나 이 STM방식은 시료가 도체라야 하므로 이 결점을 보완하기 위하여 AFM(Atomic Force Miascope) 방식을 연구하였는데 이것은 탐침과 시료의 원자 사이의 인력과 척력을 이용하는 방법이다.
이 전자현미경의 출현으로 광학현미경으후써 관찰할 수 없었던 물체를 관찰할 수 있게 되었다. 이 두 현미경의 분해력은 아래 비교표에서 보듯이 1,000배 가까이 된다. 이것은 곧 그럽 4.22와 같이 생체의 극미구조, 심지어는 물질의 구조도 볼 수 있게 되었다는 것을 의미한다. 광학현미경 전자현미경
광학현미경 전자현미경
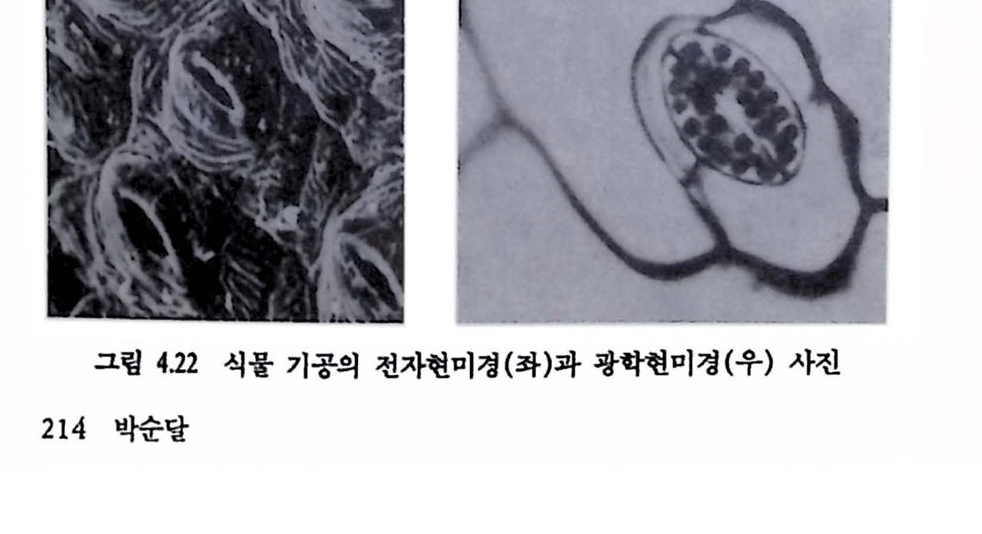
그립 4.22 식훌 기공의 천자현미경(확)과 광학현미경(우) 사진
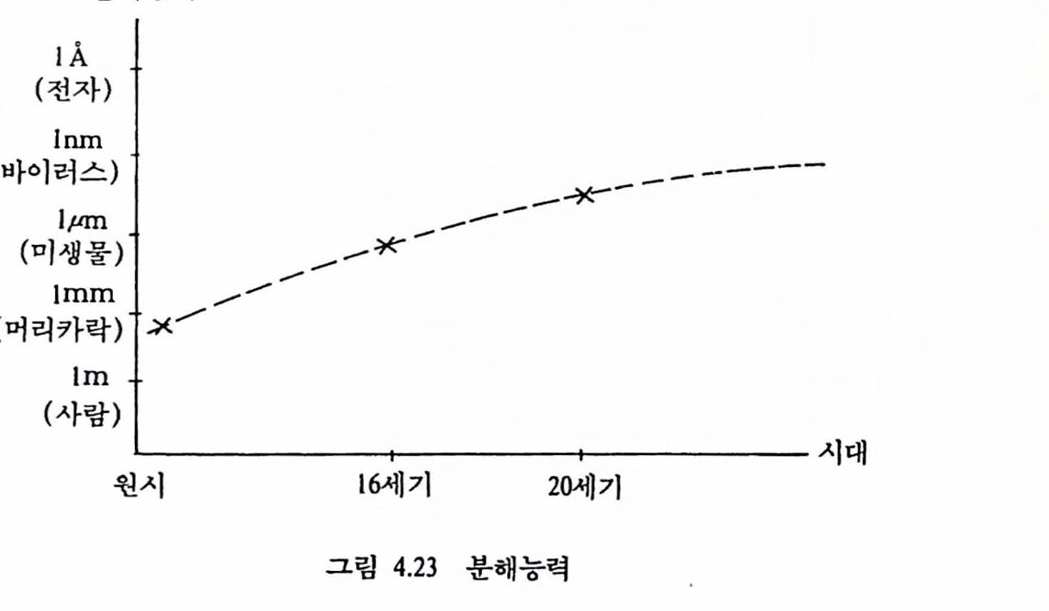 분해능력
분해능력
그림 4.23 분해능력
이러한 보조 도구의 발명으로 눈의 능력은 계속 향상되고 있다. 광학현미경이 발견되었던 16세기까지 사람의 눈의 분해능력은 O.1mm였으나 광학현미경이 나타나면서 그 능력이 m 단위로 향상되었다.
염색법 염색법은 신경세포의 구조를 연구하는 데 귀중한 방법이다. 신경세포도 일반세포와 별 다름이 없으나 신경계의 특성 때문에 일반세포에 사용되는 hematoxylin 또는 eosin을 이용한 염색법은 별 소용이 없다. 1906년에 이 연구로 노멜성올 받은 이탈리아의 Camilo Golgi(1843~1926)가 개발한 은 염색법은 가창 널리 사용되고 있는 방법이다. 이 염씩법은 독일의 Max Bielschowsky(1869~1940), 현존하는 미국의 해부학자 David Bodian, 영국의 William Holms, 스페언의 Santiago Ramony Cajal 동에 의해 연구되어 신경계의 다양한 부분의 영잭에 사용되고 있다. 이것은 신경계에 남아 있는 윤 성분올 이용하여 이 부훈옳 검게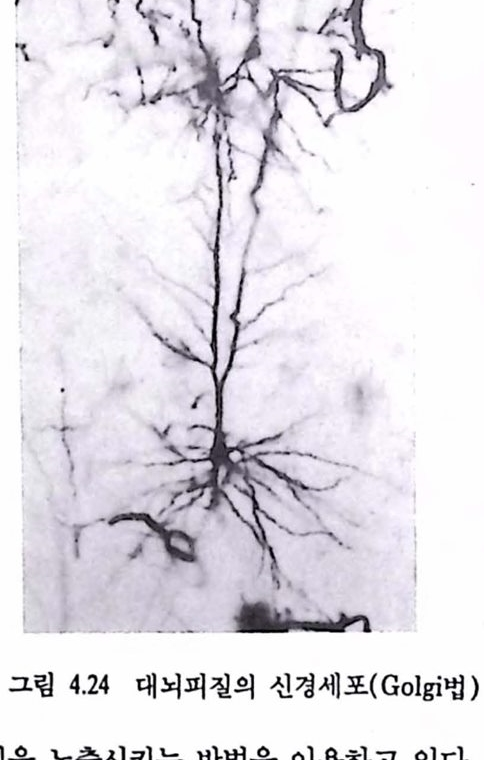
그럽 4.24 대뇌피질의 신경세포 (Golgi법)
만듦으로써 조칙을 노출시키는 방법을 이용하고 있다.
수초의 염색에는 독일의 Karl Weigert(1843~1905)가 개발한 방법이 사용되고 있다. 이 방법을 사용하면 세포체 부분은 색이 없으나 수초 부분은 캠게 나타나 찰 보이게 된다. 한편 독일의 Franz Nissl(1860~1919)은 핵산에 결합되는 cresyl violet,toluicline blue, thionine 등 니슬 염색으로 불리는 물질올 이용하여 핵부훈을 염색시키는 방법을 개발하였다. 그래서 Weigert 방법과 함께 사용하연 좋은 결과가 나타난다. 화학물질을 이용하는 방법 이런 방법은 신경의 화학척 특성을 연구하는 것으로서 신경세포는 정보교환 방법으로 화학척 수단을 사용한다는 것을 발견하고부터 가능해졌다. 이련 물질을 신경전달물질이라 하는데 아세멸콜린(Acetylcholine :ACh)이 그 하나이다. 이 아세릴콜린은 Otto Leowi가 1924년 개구리의 미주신경을 이용하여 발견한 가장 많이 연구된 신경전달물질 중의 하나이다. 이밖에도 GABA와 같은 아미노산, 토파민, 세로토닌 퉁이 알려져 있다. 이런 신경전달물질이 발견되고 그 작용과 회로가 연구됨으로써 신경계의 여러 가지 특성이 알려지고 있다.
다른 화학척 방법은 호르몬 등의 화학물질과 그 수용기를 통해 신경계의 성철을 연구하는 것이다. 예로서 모르핀은 19세기초에 처음으후 정제되어 사용되었는데 그럽에도 불구하고 모르핀이 뇌에 쉽게 작용하는 것은 뇌에 모르핀 수용기가 있기 때문이다. 1974년 Johns Hopkins 대학의 Solomon Snyder와 Candice Pert는 모르핀과 유사한 날록손이 몇 개의 뇌 영역의 신경세포 수용기에 부착됨을 발견한 것이다. 그 후 뇌 자체에서 모르핀과 버슷한 역할을 하는 엔돌핀(endorphin)올 생성한다는 사실을 발견하였다. 이런 물철이 작용히는 경로를 추척하면 신경계의 작용과 그 특성을 연구할 수 있는 것이다. 현재 이련 호르몬은 언슐린, 성장 호르몬, 에스트로겐 등으로 다양하다. 신경조직배양 일반 세포의 조직배양을 통해 톰의 여러 장기의 륙성을 연구용뚝이 신경세포를 조직배양하여 그 특성을 연구해 보고자 하는 방법이다. 얼반척인 조직배양법은 원하는 조직을 찰게 찰라 트렵신과 같은 효소와, 세포와 세포를 연결하고 있는 단백질을 분해시킨 뒤 분리된 세포훌 영양물철이 함유펀 세포배양액에 넣어 배양하면 세포가 분옐하여 성창한다. 1907년 미국 예일대학의 Ross G. Harrison이 개구리의 신경세포가 이 방법으로 성장향을 발견한 후 1950년대률 거치연서 연구가 본격화되었다. 그후 이 연구 방법도 계속 개발되고 있다. 이련 방법율 이용하여 신청세포의 시냉스, 수초의 성장 과정, 신경의 영양 관계, 신경의 성장언자, 신경세포와 그리아의 관계 등에 많은 연구가 이루어지고 있다.
그림 4.25 Harrison에 의한 최초의 세포배양에서 개구리 배아의 신경섬유가 자라고 있는 모습.
미세전극몰 이용하는 방법 신경세포는 전기척 신호로 청보를 주고받는다. 그래서 이 전기신호를 읽게 되면 신경세포 사이에 전달되는 정보를 얽을 수 있게 된다. 독일의 Gustav T. Fritsch(1838~1927)와 Eduard Hitzig(1838~1907)는 이런 방법으로 개, 원숭이의 뇌의 운동기능을 연구하여 많은 업척을 남켰다.
이 미세전극은 처음에는 인체에 해로웅이 없는 금이라든지 스테인레스 철올 이용하였으나 이들도 체내에서 오래 견딜 수 없어 그 후 발달한 전극어 tantalum과 iridium이다. 이들 전극은 가늘수록 좋으며 요사이 가는 전극은 10mn에 이르고 있으나 전자기술의 발달로 IC회로 굵기만큼 가늘게 만들 수도 었다. (2) 예측 뇌에 대한 연구는 인체의 다른 장기에 비해 뒤떨어진다. 그러나 과거 20여 년 동안 약른 속도로 연구가 진행되고 있어 그 구조와 기능이 속속 범혀지고 있다. 기본단위언 신경세포에서 시작하여 감각수용기, 신경섬유, 중추신경계 퉁 여러 분야에 많은 연구결과가 쌓이고 있다. 이런 추세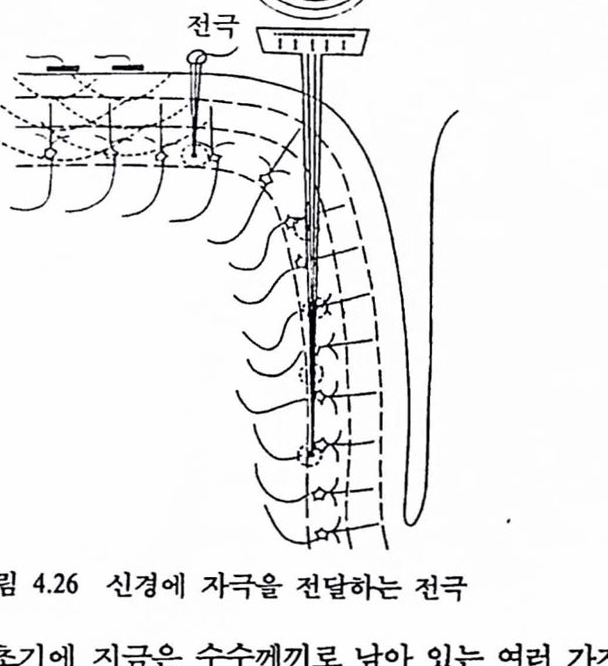 ---、 --、
---、 --、
그림 4.26 신경에 자극을 전달하는 전극
로 간다면 21세기 초기에 지금은 수수께끼로 남아 있는 여러 가지 문제가 밝혀질 것으로 예측된다.
뇌의 언공적 확장에 관해서는 ·뇌의 구조와 기능 ·기계와 언체의 거부반응 문제 ·기계와 신경의 연결 의 연구결과에 달렸다. 과학분야의 예측은 다양하게 이루어져 왔흐나 일본 과기청 주판으로 매 5년마다 각계의 전문가를 통훤하여 과학기술분야의 예측울 청리한 『일본의 기술』이 대표척언 예 중의 하나이다. 제4회째의 기술예혹은 1987년에서 2015년까지 28년간의 물질, 에너지, 의료, 환경 둥 17개 분야에 대해 기술을 예촉하고 있다. 륙히 신경계의 기술발달애 판한 훌야는정보 · 전자 · 소프트웨어 분야, 생명과학분야, 의료 · 보건 분야 등 세 분야이다.
이 연구에 의하면 뇌의 구조와 기능에 대해서 기술발전 예측을 다음과 같이 하고 있다. 학습기구의 규명 2004 년 뇌의 발생, 성창 과정의 분자단위 해명 2008년 인간 지각기구의 규명 2008년 기억의 기구 해명 2012년 뇌신호와 사고 과정 해명 2015년 이련 기술예측으로 볼 때 뇌 기구가 2000년대 초반까지 상당 부분이 해결될 것으로 보언다. 이련 해명이 곧 인공장기로 대치되는 것으로 연결되는 것은 아니지만 최소한 언공장기의 연구를 촉발시키고 나아가서는 언공장기, 즉 인공신경, 언공뇌의 연구로 이어질 것이 툴렴없다. 인공장기를 언체에 부착시킬 때 가창 큰 문제 중의 하나는 생체의 거푸반웅이다. 이에 판련되는 기술은 생체에있어서 〈자기(自己)>와 〈비자기(非自己)>를 구별하는 기구률 해명하는 것과 거부반응을 일으키지 않는 물질율 개발하는 것이다. 이 부분에 대해서는 다음파 같이 예측하고 있다. 자기와 비자기의 구별에 관여하는 면역세포 기능의 해명 1997년 거부반응 없는 언공장기 재료 실현 2001년 이련 예혹으로 률 예 21째기초에는 거부반용융 일으키지 않는 채료로 만률어진 창기가 나타날 것으로 예상된다. 어했든 이련 연구가 계속되면 신청과 기계가 첩속되는 것은 시간문제로 보여진다. IC의 연구도 나날이 달라지고 신청에 대한 연구도 급속도로 이루어지는 지금, 앞으로 어느 정도 신경과 기계, 나 가 뇌와 기계가 접속될 수 있을 것으로 기대해 볼 만하다.
이에 관한 기술 예측은 다음과 같다. 정보를 수용, 전달하는 생체 유사막 개발 2005년 감정에 직접 전하는 전달방식을 가진 기계 개발 2006년 자발적 집합, 조직화 소자의 개발 2008년 인공신경의 개발 2010년 뇌세포, 신경 등과 접속가능한 전자회로 개발 2014년 뇌와 컴퓨터의 interface 개발 2015년 언공세포의 합성기술 개발 2015년 이 기술 예측에 의하면 2010년대에 가면 어떤 형태로든 신경계와 접속되는 기계가 나타난다. 이런 기술은 곧 정보처리의 기관언 전두엽과 컴퓨터의 접속으로 이어질 수 있다. 이것을 2010년대말로 보고 있다. 2.3. 전자기술의 발달 (1) 전자기기의 발달 천기(electricity)는 그리스 말로 호박이라는 돗이다. 그리스 사랍플은 호박올 마찰하면 끌어당기는 험이 있다는 것을 발견하였다. 로마시대의 플리니우스(Plinius, 23~79), 갈레노스(Galenos, 129~200년경)는 천기 가오리의 천기현상올 관찰하였다. 중국에서는 7, 8세기에 이미 항혜에 자석을 이용하고 있었다. 16세기에 들어와 영국의 의사 길버트(William Gilbert, 1540~1603)는 『자석에 판하여』라는 책을 발간하였는데 전기와 자기훌 구별하였다. 그는 또한 지구 전체가 바로 자기률 떤 자켜엄율 지척하고 물째가 캘하 금속막대
금속막대
그림 4.27 라이덴 병
를 며고 있는지를 검사할 수 있는 검정기를 고안하기도 하였다. 18세기 중엽에는 전기를 저장하는 축전기의 역할을 하는 라이덴 병이 발명되었다. 라이덴 병올 여러 개 연결하면 축전기 구실을 하는데 이 라이덴 병으후 그 당시 여러 가지 전기에 관한 실험이 가능하였다.
물에 라이덴 명을 이용하여 방천시키면 액체 일부가 기체로 변하는 사실올 발견하기도 하였다. 전기에도 두가지 서로 다른 성질을 가진 것이 었다는 것은 뒤페이(Du Fay, 1698~1739)에 의해 발견되었는데 프랭클린(Benjamin Franklin, 1706~1790)은 이것을 양전기, 음전기라고 명명하였다. 프랭클린은 이 라이덴 뱅으로 실험한 결과 전기유체(電氣流 )셜용 수렵하였다. 이 셜에 의하연 물철에 전기유체가 너무 많으면 양전기훌 띠게 되고 부족하면 음전기를 띠게 된다. 그러다가 1897견 톰슨(Joseph J. Thomson, 1856~1940)이 천기가 천자의 흐름이라는 전기의 원리룡 밝혀 버로소 천기의 기초가 확립된다. 1780년갱 이활리아의 갈바니(Luigio Galvani, 1737~1798)는 껍철을 벗긴 개구리 다리에 아연과 구리률 고리 모양으로 하여 접속시키연 심하게 청련율 얼으킨다는 사실올 발견했다. 볼타(Alessandro Volta, 1774~1827)는 금속의 종류에 따라 경련의 세기가 다르다는 사실을 발견하였고 또한 청련의 원언은 금속이 천기률 발생시켰기 혜문이라고 주창하였
그림 4.28 볼타의 전지
다. 이런 발견에 이어 1800년에 전기를 저장할 수 있는 천지를 발명하였다.
이것은 대단히 유용한 세기척 발명으로 이로써 전기를 저축하여 두었다가 필요할 때 사용할 수 있게 되어 전기를 생활화할 수 있게 되었다. 현재 산업용으후 많이 사용하는 교류발전기는 1860맨에 발명되어 현대의 찬란한 전기문명의 기초가 세워지게 되었다. 현재 전기는 매우 다양하게 사용된다. 우선 전기는 에너지원으로 사용된다. 에너지원으로서는 전구, 전열기를 통해 빛과 열을 얻기도 하고 천기 모터를 통해 전기로부터 기계적 에너지를 얻기도 한다. 전기가 에너지원으로 사용되는 양은 다른 어느 분야보다 많으며 전기가 에너지훤으로 샤용됨으로써 문명발전에 끼친 공로는 지대하다. 천기는 다론 곳으로 전송하기 편하기 때문에 다른 어떤 종류의 에너지 보다 면리하게 사용될 수 있다. 한편 1785년 쿨롱(Charles A. de Coulomb, 1936~1806)은 천체에 만유인력이 적용되듯이 천기를 띤 물체 상호간에 거리의 역쩨곱으로 험이 작용하는 쿨롱 볍칙올 제시하여 전기의 원리률 세웠다. 천기 사용에 있어서 한 단계 고차원척 발견의 하나가 천자기파의 발견언데, 1820년 텔마크의 외르스테드(Oersted) 교수가 전류에 의혜 자채가 발생됨올 발견하고, 1864년 스코틀랜드의 맥스웰(James C. Maxwell, 1831~1879)이 전자기파의 존재룰 예언하였다. 그리고 1884년 톡얼의
그립 4.29 진공관
헤르츠(Heinrich R. Hertz, 1857~1894)에 의해 전자기파의 존재가 확인되었다. 이 결과로 1899년 마르로니(Gulielmo M. Marconi, 1874~1937)에 의해 영국과 프랑스 사이의 최초 무선통신이 성공하여 통신의 신기원을 이룩하게 된다. 이 결과 라디오, 무선전화, 텔레비전 등 현대의 다양한 문명의 이기가 나타나게 되었다.
1860년경이 되면 교류발전기가 사용되는데 이것으로 전기를 인간 스스로 만들어 사용할 수 있게 되어 전기를 생활화하는 데 큰 공헌을 하였다. IC 의 발달 전기는 훤자핵 주위를 몰고 있는 음의 전하를 가진 전자(electron)의 움칙임이다. 전기를 유통시키는 도체는 부하가 걸리연 전자가 일정한 방향으로 움칙인다.비도체는 이 전자의 움직임이 얼정한 방향으로 움직이지 않기 때문에 전기를 유통시키지 못한다. 이 전기는 진공에서도 흐르는데 이것은 금속에서 튀어나온 자유천자의 흐름이다. 전기가 한쪽으로 흐르면 직류라 하며, 한쪽으로 흐르다 반대쪽으후 흐르다 하면 교류라 한다.
이러한 전기는 편리한 점이 많다. 전기 유통 속도는 빛의 속도(30km/초)와 비슷한데, 먼 콧에도 빠른 속도로 이 전기를 보낼 수 있다. 즉 에너지를 먼 곳에 빠르게 보낼 수 있는 것이다. 현대의 문명 이기는주로 이 전기를 이용하는 기기이고 이 기기 중에서도 현대 문명이 낳은 가장 획기적인 산물이 컴퓨터이다. 컴퓨터를 구성하는 데는 여러 가지 부품이 필요하다. 제1세대 컴퓨터는 여러 가지 전자 부품으로 만들어졌으나 대표적인 부품은 진공관이다. 진공관은 1940년에 영국의 프레밍(John. A. Flemming)이 처음으로 발명하였다. 1883년에 에디슨(T.A. Edison)은 전등에서 가열된 필라멘트에서 약간 떨어진 금속판으로 전자가 흘러가는 현상을 발견했다. 이것을 에디슨 효과라 하는데 프레멍은 이 특성을 이용하여 수도관의 밸브와 같은 역할을 할 수 있는 2극판을 처음으로 만들었다. 2극관은 전기를 방출하는 음극과 받아들이는 양극을 유리 공기내에 봉입한 것이다. 이 음극을 가열하연 전자가 방출되는데 이때 양극의 부하가 +인지 -인지에 따라 전자를 받든지 못 받든지 하게 된다. 따라서 전류가 한쭉으로만 흐르면 정류작용이라 한다. 그는 이것을 밸브(valve)라고 불렀는데 이것은 전류를 한쪽으로만 흐르게 하기 때문에 검파기로 사용되었다. 이것은 또한 전기의 흐름을 편리하게 조정할 수 있게 된다는 뭇이며, 이로써 전기가 현리하게 일반생활에 사용되었다. 이 진공관은 그 뒤 3극판, 4극관 등 여러 가지로 발전하다가 1950년대 트랜지스터(transistor)가 일반화될 때까지 라디오 풍 천자 제풍의 필수풍으로 사용되어 왔었다. 수를 기계로 표현하는 방법은 여러 가지가 있는데 홉니빼의 홉니로 표현할 수도 있고 또는 요사이 컴퓨터에 사용하는 2진법으로 표현하려면 여러 개의 스위치(끄연 0, 켜면 1)로 표현할 수도 었다.수훌 연산한다는것은 결국 이런 수많은 스위치를 조작한다는 것을 의미한다. 이런 작동을 1초에 수십만 번씩 할려면 기계식으로는 불가능하다. 그러나 전기를 사용하면 부품의 고장도 없고 에너지도 덜 소모된다. 전자 부풍 사용의 장점이 바로 여기에 있다.
게르마늄(Ge)이나 실리콘(Si)은 전류플 한쪽으로만 흐르게 하는 반도체로서 2극관이 한쪽으로만 전류를 흐르게 할 수 있는 성질을 가진 것과 같다. 이런 특성을 이용하여 1947년 미국 BELL 연구소의 Shokley, Brattain, Bardeen이 소자로 발전시킨 것이 트랜지스터이다. 이 트렌지스터는 진공관에 비하여 크기는 200분 의 1밖에 되지 않고 전력 소비는 100분 의 1에 지나지 않았다. 그래서 1951년 트랜지스터가 본격적으로 상풍으로 양산되연서 전자 부풍의 가장 중요한 부품으로 사용되는 진공관을 대체하게 되었다. 트랜지스터를 발명한 이 세 사람은 1956년에 노벨상을 받게 되는데 트랜지스터는 흔한 광물인 실리콘을 이용한다는 점과 이것으후 부품을 소형화할 수 있다는 점 때문에 지금까지의 화려한 컴퓨터 문화, 즉 설리콘 문화의 시발을 뭇하는 전자 소자가 되었다. 트랜지스터가 나타나면서 전자 소자의 소형화는 더욱 박차를 가하게 되었는데, 특히 1958년 Texas Instruments 회사의 킬비(Jack Kilby)는 트랜지스터, 다이오드, 저항기, 커왜시터 둥의 전자 소지들을 조그마한 실리콘판에 하나의 회로로 묶어 만들었는데 이것을 칩척회로(integratedcircuit, IC)라고 펌흔다. 이 IC를 만들 때는 반경 5~10cm 정도의 살리폰판에서 수백 개의 IC청(조각)을 그려 넣어 만들게 된다. 이 첩은 작은 것도 있고 큰 것도 있으나 작은 것은 크기가 1mm도 안 되는 것이 었다. 이렇게 작은 첩에 전자 소자가 수백 내지 수십만 개가 들어깐다. 1964년 무어는 1959년 이래 첩당 소자의 개수는 매년 2배썩 충가할 것이라고 예언했는데, 실제로 그렇게 되어 왔다. 그러나 1980년 대는 충가 속도가 둔화되어 2년에 2배씩 중가했다. 지금은 4메가 첩, 16 메가 챙이 나오는데 이것은 1cm 크기의 한 청에 수백만 개의 소자가 들어깐다. .---- .- - --------
.---- .- - --------
그림 4.30 IC
표 4.1 IC 의 발달사
 종류 여L 도 비고
종류 여L 도 비고
표 4.2 IC의 계산 속도
 IC 종류 관련 컴퓨터 세대 연산속도
IC 종류 관련 컴퓨터 세대 연산속도
이 IC는 전술한 바와 같이 개별 전자 소자를 하나로 묶어 실리콘판에 그려 넣은 것이다. 그런데 1971년 미국 Intel 회사의 호프(Ted Hoff)는 조그마한 실리콘 첩에 기억 레지스터, 논리 회로를 그려 넣어 이 칩 하나가 바로 컴퓨터 역할을 히는 마이크로프로세서(microprocessor) 를 만들었다. 이 마이크로칩은 이 자체가 중앙처리장치(CPU)를 가진 IC언 것이다. 크기가 5cmX1.5 cm 정도인 컴퓨터가 생겨난 것이다. 이것으로 길이 30m 나 되먼 최초의 컴퓨터 ENIAC 컴퓨터가 요사이는 50cm 청도 크기의 개인용 컴퓨터로 작아지게 된 것이다.
IC는 계산 속도를 높어고 동시에 소형화시킨다. 진공관이 멀리초 단위로 계산하는 반면 요사이의 대규모 집척회로는 나노초로 계산한다. 이련 소자의 연산 속도 향상이 바로 컴퓨터의 연산 속도 향상느로 연결된다. IC의 소형화도 이에 못지 않게 발전되고 있다. 전술한 바와 같이 같은 크기에 들어갈 수 있는 정보량이 매년 2배씩 늘어나고 있는데, 이것은 크기가 철반씩 줄어. 가고 있다는 말과 같다. 256K DRAM은 1980년에 개벌되었는데 이것은 6mm2에 약 60 만 개의 트랜지스터, 콘헨서가 집척되어 만풀어진 초대규모집척회로이다. 이 IC는 회로의 선폭이 1 이다. 1988년 2월에 얼본의 마즈시다, 히다치, 도시바는 16M DRAM을 발표했는폐 이 정우 회로의 선폭운 0.5μ 에 폴과하다. 셀의 크기는 1.5 x 2.2 이고, 첩 천체의 크기는 5.4 X 17.38mm이고 속도는 65냐노초이다.IC를 얼마나 소형화시킬 수 있느냐는 얼마나 선폭을 줄일 수 있느냐에 달려 있다. 이것은 선폭을 결정하는 도구에 달렸다. 만일 선을 그리는데 가시광선을 사용한다면, 가시광선의 파장이 0.77μ 가까어 되므로 이보다 기는 선은 구멸하지 못한다. 그래서 가시광선으로는 약 1μ보다 가는 회로를 그릴 수 없다. 그래서 파장이 이보다 훨씬 가는 0.1μ 내지 1A 의 파장을 가진 X션을 이용한다.
그런데 션을 가늘게 팩빽하게 그리는 데도 한계가 었다. 지나치연 간섭현상룰이 생겨 제구실을 못하게 된다. 그래서 현재로서는 이론척으로 선폭은 0.1μ, 용량은 64M가 한계가 아닌가 보고 있다. 그래서 현재의 방법과는 다른 방법을 연구하고 있다. 다층 소자, 다층의 3차원 회로, 새로운 재료의 소자 등이 새로이 시도 되고 있는 방법이다. 현재의 소지는 모두 O또는 1 의 값을 갖는 두 값(버트) 소자이다. 그러나 0과 1뿐만 아니라 여러 개의 값을 가질 수 있는 소자를 생각할 수 있다. 이렇게 한 소자가 많은 값을 기질 수 있으 면 결국 정보량을 많이 가지게 된다. 다음에 생각할 수 있는 방법으로 회로를 다층으로 3차원 소자를 만드는 것이다. 지금의 소자는 회로를 명면에 그리고 있다. 그러나 4M DRAM만 해도 약 5미크론 깊이의 홈을 20여 개 파서 양옆으로 회로를 그려넣는다. 2층 구조는 아니지만 2층 비슷해진다. 회로를 충충으로 그리연 훨씬 많은 양의 정보를 넣을 수 있다는 것은 두말할 나위 없겠다, 또한 새로운 소자로는 칼륨 비소 소자 조셉슨 소자 가 관심을 몰고 있다. 칼륨 · 버소 소자는 실리콘 대신에 칼륨에 버소 화합물이 들어간 반도체로서 1982년 8월 일본 도시바 회사가 4mm2에 2,400개의 소자를 칩척시켜 대규모 집척회로 제조에 성공했다. 속도는 Fe off 포르피련분자 on
Fe off 포르피련분자 on
그림 4.31 맥그레아의 분자 스위치
그 당시 실리콘 반도체 속도의 5배로서 상온에서 초고속을 실현하고 기공이 비교적 쉽다는 데 장점이 있다. 미국의 TI사는 1988년 2월 32 버트 마이크로프로세서를 발표했는데, 게이트 속도가 16피코초에 도달하고 있다. 조셉슨 소자는 1962년 영국 케임브리지 대학의 조셉슨(B. Jesephson)이 제안한 이론을 실현시킨 것으로 거의 절대온도에 가까운 -273°C에서 생기는 초전도현상을 이용하여 실리콘 반도체보다 20~50배의 고속 연산을 실시해 보고자 하는 것이다. 1966년에 IBM에서 나노초 단위의 스위청 속도를 가진 소자를 실현시켰고, 일본에서는 10.8 피코초까지 발전시키고 있다. 이 소자의 빠른 속도뿐만 아니라 에너지도 다른 반도체 소자의 100분 의 1의 에너지밖에 소요되지 않아 제 5세대 컴퓨터의 주역 소자가 될 것으로 기대되고 있다.
이들 외에 광집적회로도 있다. 1978년 캘리포니아 공과대학의 암논야리프 교수의 광집적회로에 관한 연구에서부터 시작하여 많은 연구가 진행되고 있다. 이것은 빛을 이용하여 스위치를 만드는 소자로서 동작 속도는 10억분의 1로 새로운 세대의 소자로 관심을 받고 있다. 단백질을 이용한 소지는 미국의 gentronics 사장인 J. H. 막카레아 박사가 분자 스위치를 제안하면서 관심을 끌었다. 그는 포르피련과 포리엔 구조를 가진 화합풀을 단백철 기반 위에 배치시키는 스위치를 제안한 것이다. 동작원리는 그렴 4.31에서 보는 바와 같이 포르피련을 단백질막에 배열시키면 스리톤 영역에서 제어전극의 신호에 따라 파가 포리엔부에서 포르피련 분자에 도달하면 포르피린 면의 각도가 변하여 스위치가 작동한다는 것이다.
다음에 그가 제안한 바이오칩은 몰톤(moleton)이라고 불리는 것으로서 이것은 모노크로날 항체, 효소, 분자 스위치 둥으로 구성되어 생체와 같이 자기증식, 자기복제가 가능하도록 설계한 것이다. 이 소자의 크기는 약 100A 정도이고 고도로 접적화시킬 수 있어 1980년대초 그 당시의 칩에 비헤 단위용적의 기억용량이 10억 배가 될 것이라고 했다. 이 몰톤은 아직 연구단계에 있으나 매우 작고 또한 소형이기 때문에 초고속 작동이 가능랴고 열의 발생이 없는 장점 떼문에 연구는 계속되고 있으며 유사한 형태의 성공적인 연구가 보교되고 있다. 예를 들면, 동경 공대의 경부정부 교수는 빛을 씌연 수소 이온을 이동시키는 색소단백질을 이용하여 생체 스위치를 만들었다. 이와는 다른 형태로서 신경세포는 정보를 병렬로 전달하는데 이련 특성을 대규모 집적회로에 구현시키고자 하는 것이다. 미국의 Bell 연구소, 제트 추진연구소, 캘리포니아 대학 등지에서 신경세포를 닮은 IC를 시험하고 있다. 또한 일본에서는 시신경에서 시상을 전기신호로 바꾸어 주는 시신경 색소와 동일한 Rhodopsin을 바이오첩의 시발물질로 이용할 수 있을 것으로 보고 있다. (2) 전기기기와 신경 신경보조장비로서의 전기기기는 제1창 운동제어에서 운동신경의 보철용으혹, 그리고 제2장 감각제어에서 감각수용기의 보철용으로 사용되고 있음을 보았고, 앞의 신경보철항에서 IC가 손상된 신경의 연결용으로 사용될 수 있음을 보아 왔다. 지금까지 몸 또는 창기가 손상되었을 때 그 손상된 부위에 대한 보철이 수행되어 왔듯이 이제는 손상된 신경계에 전기기기 보철이 더욱 많이 쓰일 것임에 툴렴없다. 전기기기가 신경보철에 사용되는 이유는 우선 신경은 정보유홍의 도구로 전기신호를 이용한표 4.3 물체의 크기
 단위 인체 생체 물체
단위 인체 생체 물체
다는 것이고 다음에는 전기기기는 신경과 버견될 만큼 작다는 것이다. 천기기기는 앞에서 보었듯이 대단히 복잡한 시스댐을 대단히 작은 규모로 접척시킬 수 있다. 이제 이 작고 다양한 기능을 가진 전기기기가 신경계와 얼마나 비교될 수 있울지 알아보자.
인체의 크기 : 생체기놓율 보조 • 보완하기 위한 기기는 우선 그 크기가 생체와 찰이 작아야 하고 기능 또한 생체가 가지는 것만큼 다양하여야 한다. 사랍의 키는 1~2m이고 가슴둘레는 대략 1m쯤 된다. 인체의 장기는 보홍 이보다 1,000배 작은 nm단위이다. 장티푸스, 발진티추스 등의 써균이 이 부류에 속한다. 그리고 이보다 1,000배 작은 mm단위는 바이러스 세계가 되고 이보다 10배 작은 A단위는 물질의 기본단위가 된다. 신경샘유와 천기기기 : 신경세포는 종류에 따라 다르지만 제1장의 그렴 1.4에셔 보았듯이 세포체는 크기가 10~20 m, 그리고 촉색의 굵기는0.5~1μm이다. 물론 굵은 것은 오징어의 축색과 같이 1mm 짜리도 있으나 보편적으로는 가늘고 수초의 유무에 따라 굵기가 달라철 수도 있다. 그리고 신경섬유는 이런 축색의 다발로 되어 있는데 그 굵기와 축색의 개수는 종류에 따라 다양하다. 굵은 것은 수 mm에서 척수와 갇이 아주 굵은 것도 있고 축색의 개수도 수천, 수백 만에 이르기까지 다양하다.
이러한 신경섬유와 접촉하기 위하여 사용되는 기기는 신경보철항의 미세전극의 예와 신경연결을 위한 IC의 예를 보았다. 미세천극은 Tantalum 이나 iridium으로 만든 20μm 굵기의 전극까지 사용하고 있으나 이 굵기는 점차 가늘게 할 수 있다. IC를 제조하는 기술을 이용하면 앞에서 보았듯이 0.6μm까지 만들어 낼 수 있다. 이 굵기는 신경축색 하나의 굵기와 같아진다. 굵기가 같으니까 이 선으로 축색을 대체시킬 수 있을 것이다. 결론은 이르지만 최소한 부분적 교체 내지는 인공신경 섬유 제조에 있어서 커다란 기술적 장애가 제거되었다고 할 수 있다. IC의 제조기술은 앞으로 더욱 진보할 것이고 0.1μm까지 내다보교 있다. 이렇게 되면 신경축색보다 몇 배 기는 선이 나타난다는것을 의미한다. DNA와 IC : 이제 한발 더 나아가 유전정보를 보관하고 있는 DNA와 IC를 비교해 보자. 세포에는 23쌍의 염색체가 있고 각 염색체는 500여 개 이상의 유전자를 보유하고 있다. 각 유전지는 1,000여 개의 nucleotide로 구성되고 있는데 이 nucleotide는 염색체의 기본 물철언 DNA의 단위이다. 이 nucleotide의 크기는 그림 4.32에서 보듯이 10AX 3.4A이다. 즉, DNA의 반경은 1nm이다. 이것은 IC 선폭 0.5μm에 비하여 500배의 차이가 나는 것이다. 500배의 차이는 큰 차이이지만 이 차이는 사람의 기술로 극복하지 못할 한계를 넘는 것이라고는 보기 힘들다. 그리고 지금의 기술개발의 속도로 볼 때 상당히 근접할 수 있을 것으로 예상할 수 있다. 다음에 복잡도에 있어서 60조 개의 세포로 이루어진 사랍에 있어서 이 60조 개의 세포 사이에 일어나는 기능 상호간의 복잡도는 표현하기 34A
34A
그림 4.32 DNA 의 모형
힘들 것이다. 그러나 유전형질의 기초 물질인 DNA가 정보의 보관장소이고 이 DNA가 얼마의 정보를 가지고 있는지 보는 것은 흥미로울 수 있다.
DNA는 기본단위가 nucleotide이고 nucleotide 3개가 모여 하나의 아미노산을 만드는데 이 아미노산이 유전자의 구성단위이다. 이 아미노산의 크기는 그렴 4.32에서 보듯이 길이 10A, 반경 10A 청도 된다. 그러연 IC의 기억단위의 크기는 얼마인가? 전술한 16M DRAM의 기억단위 생의 크기가 1.5X20μm2이다. 이렇게 보면 길이가 대략 1,000배 청도 차이가 난다. 이것은 사랍이 만든 물건의 청보단위가 세포의 정보단위보다 1,000배가 크다는 것을 의미한다. 그러나 전술한 선폭에서 보았듯이 기술개발이 급격히 발달하는 현 수준을 비추어 볼 때 2000년 초기에는 상당히 가깝게 그 차이를 줄일 수 있을 것으혹 예상할 수 있다.
2.4. 컴퓨터 (1) 컴퓨터의 역사 컴퓨터는 전자계통의 기계이고 전자기술의 소산이다. 그러나 컴퓨터는 한순간 느닷없이발명된 기계는 아니고 오랫동안 쌓여진 기계, 전자분야의 기술 발전의 소산이다. 컴퓨터의 역사는 계산기 자동계산기 컴퓨터 의 시대로 나누어 볼 수 있다. 계산기는 계산을 수행할 수 있는 기계를 말하며, 이런 기계는 주로 손으로 일일이 기계를 작동시키면서 계산하게 된다. 이보다 발전된 기계가 자동계산기로 이 기계는 계산하고자 하는 자료를 입력시격 놓으면 자동척으로 계산이 끝나는 기계를 말한다. 컴퓨터는 전자식 자동계산기를 뭇한다. 계산기의 시초는 1642년 19살의 프랑스 수학자 파스칼(Blaise Pascal)이 만든 덧셉기계이다. 파스칼은 세금 징수원이었던 아버지를 돕기 위하여 작은 바퀴를 조합하여 덧생과 탤생을 수행할 수 있는 기계를 만들었다. 이 기계로 세금 징수 업무를 처리하는데는 충분하였흐나 그 외의 계산, 예를 들어 두 수의 곱셉과 나눈생을할 수 없었기 때문에 얼마 후 독일의 수학자 라이프니츠(Gottfried W. Leibniz)는 곱생과 나눈생올 할 수 있는 기계로 발전시켰다. 이련 계산기는 그 후 계속 발전되어일반 상점에 널리 분포되어 최근까지 사용되어 왔다.
속도계의 아버지로 불리는 영국의 수학자 배버지(Charles Babbage)는 많은 기계를 발명했는데, 계산을 자동적으로 수행할 수 있는 기계를 만들기 위하여 많은 연구를 하였다. 그는 Logarithm 등 어려운 수식을 정확하고 빨리 계산할 수 있게 하기 위하여, 1820년에 수많은 기어와 톱니바퀴로 만들어진 차변기관(differential engine)을 발명했다.이 기계는 단순히 가감숭제만 할 수 있는 것이 아니라 복잡한 수식을 계산할 수 있는 기계이기 때문에 덧생기계보다는 한 차원 높은 기계였다. 그는 나아가 이 차변기관에 머무르지 않고 좀더 다양한 기능을 가진 자동기계를 고안하였다. 이 기계는 계산을 담당히는 mill이라고 불렀던 논리 센터, 자료를 보판하는 store라고 불렀던 기억장치, 업출력을 통제하는 통제단위로 구성된 것으로 지금의 컴퓨터 구성의 개념과 버슷했다. 자료는 종이에 구멍을 뚫어 사용하기로 하고 수행할 작업에 따라 프로그랩할 수 있게 하였다. 이 기계를 해석기관(analytical engine)이라고 불렀는데, 만일 성공척으혹 만들어졌더라면 축구 구창 2개를 합친 것만큼 컸을 것이라고 한다. 그러나 개념만은 현대 컴퓨터의 것과 비슷하여 그의 자동계산기에 대한 집념이 계승되어 실현되었다고 할 수있다. 미국에서는 방대한 언구 및 산업의 국셰 조사에 신속한 자료를 처리할 기계가 필요했었는데, 1890년 훌레리스(Herman Hollerith)는 자료를 구명 뚫련 종이에 표시하고 처리하는 천공기를 만들었다. 이것은 계산기는 아니지만 자료률 구멍 뚫련 종이(천공 카드)에 나타내고 그것으로 손쉽게 자료를 훈류할 수 있었다. 그래서 많은 양의 청보를 처리해야 하는 상업, 행청, 과학 분야에 널리 사용되었고 특히 정부, 보험회사, 철도회사 퉁에 많이 사용되었다. 이 기계는 1960년대 컴퓨터가 일반화될 때까지 널리 사용되었다. 배버지는 자동계산기를 설계하였으나 1871년 축율 때까지 만들지 못하였다. 처음으로 자동계산기가 만들어진 것은 1944년이었다. 하버드 대학의 에이컨(Howard Aiken) 박사에 의해 고안되고 IBM회사에서 엔디코트(Endicott)가 처음으혹 자동계산기 MARKI 을 만들었다. 이 기계는 주로 전기 스위치인 3,000여 개의 계전기로 만들어진 것으로서, 크기는 커다란 방만한 것이었고 계산 속도도 느리고 자체 기억용량도 적었다. 그러나 계산을 자동적으로 수행하기 때문에 계산기 발전에 하나의 획을 긋는 기계였다.
이 자동계산기는 그 뒤 계속하여 발전하면서 새로운 개선된 모형이 나왔지만, 속도를 개선하고 조작이 편리한 기계를 만들기 위하여 곧바로 전자식 계산기로 대치된다. 처음으후 만들어진 것이 1946년 펜실베이니아 대학에서 엑커트(J. Presper Ekert, Jr)와 모클리(John W. Mauckly)가 고안한 1만 8000개의 진공관, 7만 개의 저항, 1만 개의 콘덴서, 6,000개의 스위치로 만들어진 ENIAC(Electronic Numerical IntegratorAnd Calculator)이다. 이 기계는 모든 부품이 전자소자로 되어 있기 때문에 최초의 전자식 계산기(Electronic Computer)이고 이것을 줄여서 컴퓨터라고 부론다. 그래서 컴퓨터의 역사는 이 ENIAC에서부터 시작한다. 이 ENIAC은 기계 부품 대신에 전자 소자인 진공관을 사용한 획기적인 기계이긴 하지만 20개의 수 밖에 기억할 수 없다든지 작동하기 위해서는 6,000개의 스위치를 조작해야 하는 불편이 있었다. 수학자 폰 노이만(John Von Neumann)은 작동 방법까지 기억시키는 프로그랭 내장식 방법을 제시하였다. 그래서 개선되어 나타난 것이 1949년 영국 케임브리지 대학에서 윌크스(M. V. Wilkes) 교수가 설계한 EDSAC(Electronic Delay Storage Automatic Calculator)과 폰 노이만 교수가 1952년 설계한 EDVAC(Electronic Discrete Variable Computer)이다. 이들은 모두 진공관으로 만들어진 컴퓨터이다. 이들 컴퓨터를 제1세대 컴퓨터라고 부른다. 제1세대 컴퓨터로서 처음으로 상용화된 것은 액커트와 모클리가 프로그램 내장식으로 고안한 UNIVAC I(Universal Automatic Computer I) 이었다. 이 컴퓨터는 1950년에 제작되기 시작하여 1951년 6월 미국 펜실베이니아 인구 통계국에 판매됨으혹써 컴퓨터의 상용화가 시작되었다. 이때가 되면 계산기가 단순한 산술 계산만 하고 천공기가 자료의 분류에 주로 사용되던 한계률 벗어나 천표의 발행, 급여 계산, 재고 관리, 통계 처리 등 다양한 정보 처리에 사용되게 된다. 컴퓨터는 이후 기억용량 계산속도 를 증가하기 위하여 노력한 결과 세대를 거둠할수록 비약적인 발전을 거둠하고 있다. 컴퓨터의 세대는 주로 컴퓨터에 사용되는 전자 소자에 의해 구분된다.
1947년 미국의 Bell 연구소에서는 기능이 진공관과 같으면서 크기가 200분의 1인 트랜지스터를 발명한다. 이것을 사용하면 단위 용적에 훨씬 많은 작업을 수행할 수 있는 컴퓨터를 만들 수 있다. 이 트랜지스터를 사용한 컴퓨터 IBM 1401이 1959년에 나타나는데, 이 트랜지스터를 사용하여 만든 컴퓨터를 제2세대 컴퓨터라고 한다. 이 제2세대에 들어오면 제1세대에 비해 컴퓨터의 크기, 전력 소모, 기억용량에 많은 발전이 있다. 그리고 기능도 다양해져 더욱 다양하게 활용된다. 트랜지스터가 나타난 지 10년 뒤 조그마한 실리콘판에 회로로 접적시킬 수 있게 되는데, 이것을 집척회로(integrated circuit : IC)라고 부른다. 이 IC가 컴퓨터에 사용되면서부터 실리콘시대가 시작되며, 실리콘 화합물이 막강한 위력을 발휘하는 문명이 도래하기 시작한다. 이 IC는 처음에는 사방 1cm 청도의 작은 판에 트랜지스터 몇 십 개 정도에 맞먹는 역할을 하는 회로가 만들어쳤으나 매년 2배씩 늘어나면서 요사이는 몇 백만 개에 맞먹는 회로가 만들어지고 있다. 이 IC회로로 만들어진 컴퓨터를 제3세대라고 하고 대규모의 IC로 만들어진 컴퓨터를 제4세대라고 한다. 컴퓨터는 전술한 바와 같이 하드웨어와 소프트웨어로 이루어지는데, 하드혜어 발전에 결정척언 역할을 하는 것이 부품언 전자 소자이다.그래서 컴퓨터의 세대가 이 소자의발달사에 맞추어 구분되는 이유도 여기에 있다. 한편 컴퓨터의 기능은 소프트웨어에 의헤 좌우되는데, 보통 하드웨어의 기능이 발전하면 소프트웨어도 동시에 발전하여 이들이 상승 작용하여 새로운 세대의 컴퓨터가 나타난다.
컴퓨터는 세대를 거둠하면서 계산속도, 크기, 용도상에 많은 발전이 있어 왔다. 계산속도에 있어서는 사람이 1분당 몇 십 회의 계산을 할 수 있는 데 반해, MARK I는 1분당 200회, 그리고 ENIAC은 1초당 5,000회를 수행할 수 있다. 원주율(π)을 구하는 데 있어서, 영국의 W. Shanks(1812~1882)는 거의 평생의 각고 끝에 1937년 π의 값을 소수점 이하 707자리까지 구하는 데 성공했다. 그런데 폰 노이만은 1950년 ENIAC 을 사용해서 소수점 이하 2035자리까지 계산했는데 527자리 이하는 틀려 있다는 것을 발견했다. 요사이 컴퓨터의 계산속도 단위는 MIPS이다. 이것은 초당 백만 회의 계산을 의미하는데 요사이의 대형 컴퓨터는 6~15MIPS이다. 그러니까 사람보다 1억 배, ENIAC 보다 1만 배 빠른 것이다. 그래서 생크스가 원주율을 구하는 데 20년이 걸렸다면 ENIAC 은 3시간, 요사이 대형 컴퓨터는 6초 정도 걸린다. 이렇게 빠른 대형 컴퓨터로써 계산을 많이 해야 하는 비행기 설계, 석유 광물 탐사, 컴퓨터 설계 등이 가능하게 된다. 더 나아가 미국과 소련의 대륙간 탄도탄(ICBM)의 경우, 막상 사건이 일어나면 1초라도 먼저 상대방의 정확한 탄도를 알아 대응책을 세워야 한다. 계산속도의 역할은 막중하다. 이러한 군사 관련 연구뿐만 아니라 우주 개발, 기상 예측, 원자력 개발 등에서는 빠른 계산을 할 수 있는 컴퓨터가 필요하다. 그래서 컴퓨터 회사들은 앞을 다투어 초대형 컴퓨터(super computer)를 개발한다. 초대형 컴퓨터는 계산속도가 50MIPS 이상인 컴퓨터를 말하는데, 가장 일찍 나타난 초대형 컴퓨터는 CRAY Research사에서 조립한 CRAY-1 컴퓨터이다. CRAY-1은 35만 개의 칩으로 구성되어 있으며, 150MIPS 속도를 가져 대형 컴퓨터언 IBM 370/168의 16배의 능력에표 4.4 컴퓨터 세대
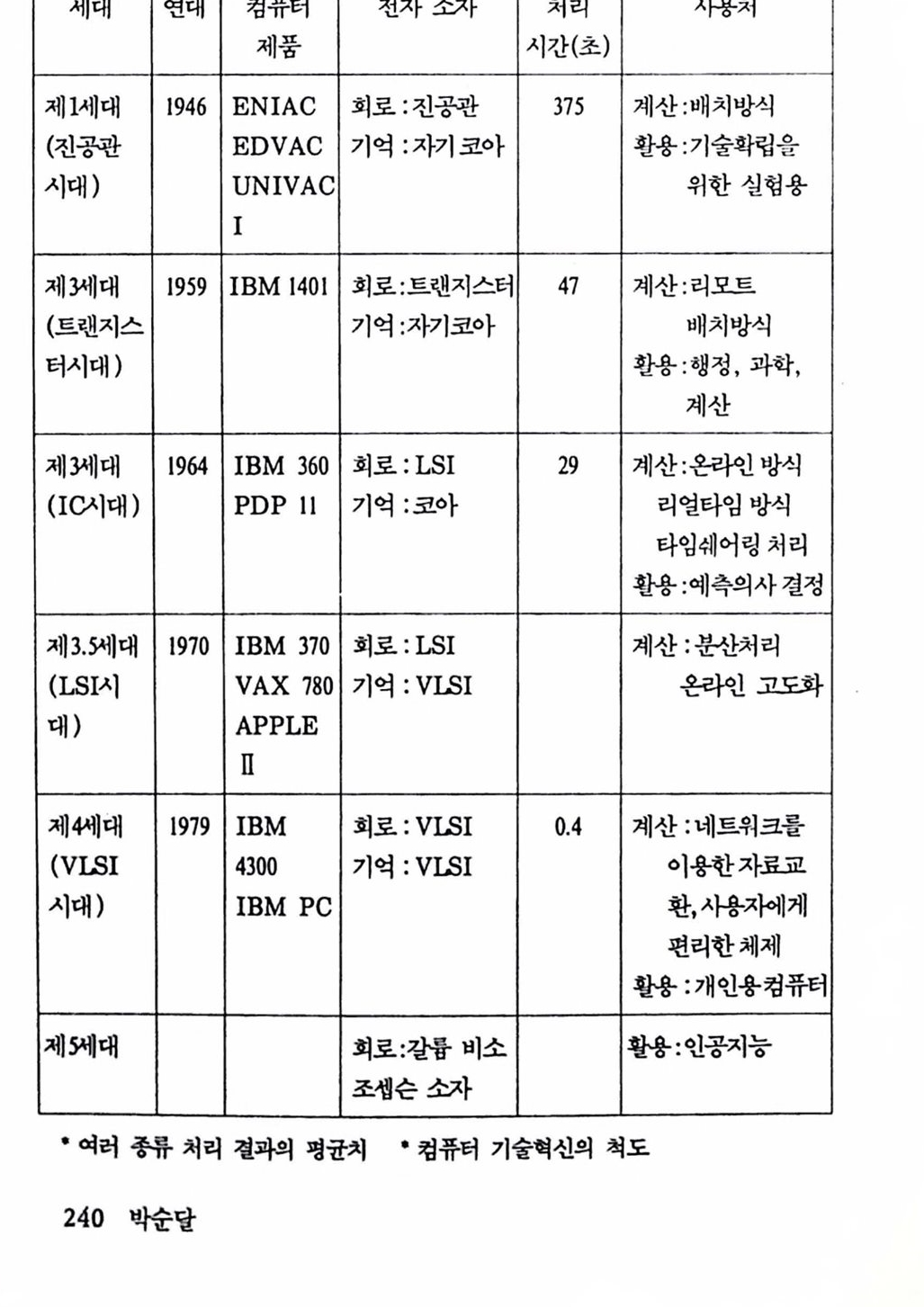 세대 연대 컴퓨터 전자소자 처리 사용처
세대 연대 컴퓨터 전자소자 처리 사용처
* 여러 종류 쳐리 결과의 평균치 *컴퓨터 기술혁신의 척도
해당한다. CRAY-1 다음에 나타난 CRAY-2는 더욱 빨라 500MIPS에 해당한다. 즉 초당 5억 회의 계산을 수행할 수 있다. 40 년 전의 ENIAC에 비하면 장족의 발전이라 할 수 있다. 이 CRAY-2는 CPU가 여러 개 있어 계산을 병렬 처리하는 데 대형 컴퓨터가 4일 걸리는 1일분 기상 분석을 2시간만에, 대형 컴퓨터가 14일 걸릴 반도체 셜계를 1일 만에 처리할 수 있는 막강한 능력을 가진 컴퓨터이다.
기억용량면에서 MARK I은 23자릿수를 72개 기억할 수 있었고, ENIAC이 20개 정도 기억할 수 있는 데에 비해 대형 컴퓨터가 2억 자, CRAY-2가 10억 자 정도 기억할 수 있다. 요사이는 개인용 컴퓨터도 몇 백만 자 정도는 기억할 수 있다. 컴퓨터의 크기도 갈수록 소형화되어 ENIAC은 설치면적이 900m2나 차지하고 무게가 30만톤이나 나가는 거물이었다. 그러나 CRAY-1은 그 막강한 능력에도 불구하고 높이는 0.7m에 지나지 않는다. 이 ENIAC의 기능은 현재 탁상 위에서 사용하는 개인용 컴퓨터보다 못하다. 가격 역시 ENIAC이 50만달러였는데 기능이 이것보다 좋은 개인용 컴퓨터는 2,000달러 정도이다. 용도상으로 볼 때도 처음에는 행정의 문서 처리라든지 과학용 수리계산에 사용되기 시작하다가 요사이는 어린이의 게임, 통신 업무, 공창지동화, 사무자동화, 인공지능 등 우리 생활의 구석구석에 활동되고 있다. 이제는 컴퓨터가 없는 문명을 생각할 수 없는 시대가 되었다. (2) 인공지능 동물 · 사람에게는 지능이 있는데 기계나 컴퓨터에게도 지능을 부여코자 한다. 이때 이 기계나 컴퓨터에 부여된 지능을 인공지능이라고 한다. 사람의 지놓은 자연적으로 발생하여 발달해온 것에 비해 인공지능은 자연발생적인 지능을 본받아 기계 또는 컴퓨터에 넣어 이 기계나 컴퓨터로 하여금 좀더 쓸만하고 지능척언 물건이 되도록 하고자 한다. 인공지능을 기계지능이라고 하는 이유도 여기에 었다. 기계 중에서 가장 지능 종전 프로그램 인공지능
종전 프로그램 인공지능
적인 것이 컴퓨터인데 이 컴퓨터가 나타난 지도 40년에 가까워 간다. 컴퓨터는 기계 부분인 하드웨어와 기계 통제 부분인 소프트웨어로 나누어져 있다. 컴퓨터의 지능은 소프트웨어에서 나온다. 지금까지 소프트웨어는 주로 논리적 수학계산에 치중되어 왔었다. 컴퓨터가 사용되는 것은 수식의 계산에서 지도 제작, 한글 타자에 이르기까지 다양한 부분에 이르고 있으나 기본적인 계산은 0, 1을 가지고 처리하는 불대수로써 이루어지는 논리계산이다. 이 계산 방식을 이용하여 문제를 푸는 절차를 알고리즘(계산방식, 계산절차)이라고 한다.
그러나 의사들이 아픈 환자를 진찰한다든지 어린이들이 놀이를 한다든지 하는 행위에서 의사결정은 수학척 알고리즘과는 다른 측면이 있어 보인다. 사람의 행위는 반드시 논리척 행위의 결과에만 의존하는 것이 아니기 때문이다. 그래서 수학척 계산을 넘어 지능적인 행위를 기계가 이해하고 처리하고 교신할 수 있도록 하고자 하는 노력에서 언공지능이 도업되게 된 것이다. 인공지능은 수학적 논리계산 뿐만 아니라 경험론척이고 직관척인 행위를 처리할 수 있게 발견적(heuristic) 기법을 많이 이용한다. 지금까지 사용하여왔던 컴퓨터 프로그램(소프트웨어)과의 가창 큰 차이는 아래 표에서보듯이 종전의 컴퓨터 프로그램은 수리적이고 알고리즘을 사용하는 대신 인공지능은 상정척이고 발견적 기법을 사용한다. 사람은 수많은 인파 중에서 자기 친구를 쉽게 찾아낸다. 이런 현상은 머리의 복잡한 계산 결과라기보다는 과거에서부터 접적되어 온경험적 지식에 의존한 결과로 보인다. 사람이 지능적인 이유는 이런 지식을 많이 가지고 있기 때문이다. 그래서 인공지능은 사람이 지금까지 쌓아 왔던 이런 지식을 많이 활용한다.
역사 : 인공지능(artificial intelligence)이라는 말이 나온 것은 1956년 미국 Dartmouth 대학에서 열린 학회에서였다. 이 대학의 수학 교수인 John McCarthy가 이 학회를 주관하였는데 이 McCarthy 교수가 인공지능이라는 용어를 만들었다. 그뒤 MIT 대학의 Marvin, Carnegie-Mellon 대학의 Allen Newell과 Herbert Simon 등에 의해 이 분야가 개척되어 왔다. 1956년의 학회에서 인간의 지능이라는 것이 잘 짜여진 추론에 기인한다고 보고 이것을 컴퓨터 프로그램으로 옮기면 될 것이라는 낙관척인 견해를 가졌으나 그 후 거둡되는 노력에도 좋은 결과를 내지 못하였다. 초기에는 주로 다른 언어로의 번역, 서양장기와 같은 게임, 모양을 처리하는 컴퓨터의 시각, 수학에서 적분을 위한 발견적 기법을 이용한 프로그램 등 일반용 수학 프로그램, 미로찾기 등 로봇의 운동에 사용될 수 있는 계산논리 등의 연구에 집중하였다. 그러나 초기의 기대와는 달리 이 분야가 어렵다는 사질을 깨닫고 또한 방법론이 많이 개발되어야 한다는 사실을 알게 되었다. 논리적 추론만으로 부족하여 인간의 지능윤 지식에 많이 의존한다는 사실과 주먹구구식과 같은 발견척 기법이 필요하다는 사실을 깨달았다. 그러나 1970년대에 들어서면서 그동안의 노력이 결실을 맺기 시작한다. Carnegie-Mellon 대학에서 만들어진 말을 이해하는 프로그램 HEARSAYII는 약 1,000개의 단어로 이루어진 말을 90% 정확히 이해할 수 있어 성공작어 되었다. SRI 연구소에서는 산업용으혹 이진법으로 된 영상을 이해할 수 있는 VISION MODULE올 개발하였다. Stanford 대학에서 언간의 지식을 사용한 전문가 시스댐인 의료진찰용 MYCIN을 개발하였다. 이러한 성공으로 인공지능은 연구방향이 자리를 잡아가고 있다. 특히, 자연언어의 처리 기계시각 범용문제 해결 전문가 시스댐 등의 분야가 두드러지고 있는데 이 중에서도 전문가 시스댐은 많은 활용이 이루어지고 있다.
이런 1960년대, 1970년대의 연구결과로 1980년대에 들어서면서 나타난 인공지능의 실용으로 이제 인공지능은 단순한 학문적 연구대상이 아니라 컴퓨터를 연구할 때 따라다니는 부품같이 되었다. 물론 이 부품은 컴퓨터 기계의 일부가 아니라 사람의 의사결정에 지대한 영향을 미치고 그 효과가 기계 그 자체보다 훨씬 큰 소프트혜어이다. 예를 들어 보자. IBM 회사는 1986년 컴퓨터의 대형 디스크의 제조에 있어서 대단히 중요한 단계인 최종 점검을 위하여 DEFT(Diagnostic Expert Final Test)라는 프로그램을 만들었다. 이 프로그램의 덕분으로 어렵고 까다로운 점검을 손쉽게 할 수 있어 좋은 효과를 거두고 있다. 경제척 효과 또한 많아 이런 결과로 연구개발에 10만 달러 투입에 비하여 연 1,200만 달러의 경비절감 효과를 보고 있다. 전문가 시스댐 : 어떤 분야에서 다른 사람이 가지고 있지 않은 지식과 문제능력을 가지고 있는 사람을 전문가라고 한다. 이 전문가는 보편적으로 모든 분야의 전문가라기보다는 특정 분야에 해당되는 전문가이다. 이 천문가는 자기 분야에서 일한 오랜 경험을 통해서 천문가가 된다. 이들 전문가들은 자기 분야에 문제가 얼어났을 때 빠른 문제 해결을 제시한다. 컴퓨터를 이들 천문가와 비슷하게 만들어 보고자 하는 것이 전문가 시스템(expert system)이라고 하는 것이다. i '-1 전문가 •
i '-1 전문가 •
그림 4.33 전문가 시스댐
전문가 시스댐은 보통 그럽 4.33과 같이 사용펀다. 사용지는 전문가 시스댐에 문제의 상황, 예를 들어 옴어 이플 때 아픈 증상을 셜명해준다. 그러나 이 전문가 시스댐내에서는 추론기관과 분야별 전문가가 있어서 추론기관이 감기일지 폐병일지 등 여러 가지 추론을 만들어 내고 이 추론이 맞는지를 지식의 보고인 분야별 전문가에 확언하여 증상에 대한 진단을 내리게 된다.
이 전문가 시스템에 가장 중요한 것은 분야별 전문가에게 얼마나 풍부한 지식을 보관케 하느냐와 다음에 추론기관에게 얼마나 훌륭한 추론과 분야별 전문가로부터 지식을 찰 결합시키느냐에 달렸다. 이제 전문가 시스댐의 예를 하나 들어 보자 동풀을 구멸하는 전문가 시스댐을 만들고자 하는데 이련 것은 주로 철문에 웅답하면 그 결과률 모아 결론에 도달케 하는 시스댐이다. 질문이란 스우그깨 놀이와 같은 것으로 그 동물은 털이 있는가? 그 통풀은 되새검철올 하는가?그 동물은 목이 긴가? 하는 것 등등이다. 이런 질문들은 동물 전문가들이 결론에 도달할 수 있게끔 충분한 동물의 특성을 기술할 수 있게 하여야 한다. 이런 질문의 응답이 그렴 4.34의 왼편에 있는 동물의 특성들이다. 이 특성으로부터 규칙을 만든다. 예를 틀연 규칙 1 털이 있으면 포유동물이다. 규칙 3 육식을 하면 육식동물이다. 규칙 6 되새검질을 하면 유제동울이다. 규칙 9 유제동물이고 목이 걸고 검은 반점이 있으면 기린이다. 하는 것 동등이다. 이런 규칙을 만들어 AND 라는 점요로 연결하거나 OR 이라는 점으로 윗햇이거나 윗윗이면 윗햇이다 라고 하는 선택을 하게 되면 결국 어떤 동물이라는 결론에 도달하게 된다. 그럼 4.34에서는 기련을 판별히는 데 필요한 규칙과 절차가 나타나 있다.
전문가 시스댐은 인공지능의 개발의 매우 중요한 분야 중 하나로 1960년대 Stanford 대학에서 화학구조물을 결정하는 데 사용하기 위하여 만든 DENDRAL에서 시작한다. 그 후 여러 분야에서 성공적인 전문가 시스댐이 나타나 활약중에 있다. 응용분야는 화학, 의학을 위시하여 전자공학, 컴퓨터, 우주공학, 군사문제 풍 다양한 분야에서 응용되고 있다. 몇 가지 예를 들면 표 4.5와 같다. MYCIN : MYCIN은 1970년대 중반에 개발된 진단용 전문가 시스댐이지만 그 후 전문가 시스탱 발전에 많은 영향을 미쳤다. 이것은 Stanford 대학의 Edward Shortliff에 의해 만들어진 것으로 의사가 환자를 진찰하고 척철한 진료를 선택할 때 도움을 줄 수 있도록 만든 것이다. 이것은 재래식 컴퓨터 프로그랩괴는 달리 발견척 기법을 사용하고 혔戀렐~
혔戀렐~
그립 4.34 동물판별 천운가 시스댐
표 4.5 전문가 시스템
 분야 전문가시스댐
분야 전문가시스댐
있다. 어떤 병을 진단하는 데는 그에 관련된 여러 가지 중앙을 고려하여 결론을 내리게 되는데 이것을 도식화한 것이다. 어떤 원인이 있으면 어떤 결과에 도착된다. 예를 들어 머리가 아프고 열이 나면 감기이다 하는 등이다. 이것을 규칙(rule)이라고 한다. 그런데 어떤 결론에 도착하려연 한 단계로 끝나지 않는다. 여러 가지 충에 정확히 결합되었을 때 정확한 진단이 내려질 수 있다. 즉 규칙과 정확한 결합이 필요한 것이다. MYCIN은 500여 개의 규칙을 가지고 있다. 의사들로부터 증상으로부터 진단에 이르게 되는 과정을 철저히 조사하여 하나의 전문가 시스댐을 만든 것이다. 보통 의사가 사람에 따라 48~58%의 오진율을 나타내는데 이 MYCIN은 35%의 오진율을 나타낸다고 한다.
이와 같이 청확한 규칙과 절차를 통해 결정에 도달하는 방법을〈규칙에 업각한 방법〉이라고 하는데 이와 같은 방법은 사람들의 일상생활에 많이 얼어나는 의사결정 방법 중의 하나이다. 의도적, 합리적 의사결정은 대개 이련 유형이다. 주민등록증을 떼러 동사무소에 간다든지, 50건 후의 국방력 강화를 위한 방위산업 육성 방안의 결정이라든지 하는 것이 이런 유형이다. 그러나 사랍은 이련 의사결정 방법에만 의존하는 것이 아니다. 사랍은 여러 가지 착오에 대한 수정 방법을 가지고 있다. MYCIN의 결정이라면 의외의 상횡을 다룰 수 있는 융통성이 척다는 것이다. MYCIN은 훤래 진단용으혹 만들어진 것이다. 규칙을 의료용 대신에차량관리용 규칙으로 대치시키면 차량관리용 규칙을 만들 수 있다. 전문가 시스법의 구조만 가지고 있는 것을 Shell이라고 하는데 MYCIN도 Shell로 발전되어 EMYCIN이 개발되어 있다. 이 EMYCIN을 이용하여 다른 분야용 전문가 시스랩을 만들 수 있다.
인공지능에 대한 여러 나라의 관심은 지대하다. 일본의 경우 10년 동안 50억 달러의 5세대 컴퓨터를 위한 사업을 진행중에 있는데 이 중에 중요한 부분이 인공지능이다. 미국, 유럽국가 등도 예외가 아니다. 앞으로 컴퓨터 세계에 미철 영향을 고려하여 인공지능에 대한 연구개발에 막대한 노력을 하는 것이다. 이 인공지능은 이런 노력의 결과로 빠른 발전을 거품하고 있어 멀지 않은 장래에 부분적으로는 인간지능에 못지않은 물건이 나올 것임에 틀렴없다. 활용의 예 : 미국 국방성의 연구개발 담당부서인 Defence Advanced Research Project Agency(DARPA)가 1983년에 착수한 Strategic Computing Program(SCP) 사업을 보자. 이 사업은 10년 동안에 6억 달러를 들여 미국의 군사장비를 종더 지능화시키는 데 주목척이 있다. 최종 목표는 전자기술(microelectrics), 시스댐 설계(system architecture) 기술, 인공지능 기술을 기반으로 육군의 무인 자동차량(AutonomousLand Vehicle), 해군의 전쟁관리(Battle Management), 공궁의 조종사 보조(Pilot's Assistance)를 개발하는 것이다. 여기서 무언 자통차량은 스스로 정찰도 하고 장애물을 피하기도 하며 목표물율 공격할 로붓탱크, 컴퓨터 보병과 같은 것이다. 천쟁관리는 급변하는 대규모 전투장황 아래 정보를 분석하고 대안을 만들며 의사결정을 내리도록 하는 시스댐이다. 그리고 조종사 보조는 조종사의 컴퓨터 보조로 기체의 상황 정검, 고장 진단 등 단순한 업무를 처리하고 대신 조종사는 고도의 의사결정에 전념하도록 하는 것이다. 이 최종 목표를 달성하기 위하여 필요한 세 가지 기술 중 천자기술과 시스템 설계기술은 기계(hardware)에 관한 기술이다. 이 기계를 다품기술이 인공지능인데 이 수준의 인공지능을 개발하기 위하여 영상처리(image processing), 음성인식(speech recognition), 자연언어 이해(natural language understanding), 전문가 시스템(expert system)의 기술이 개발되어야 한다.
영상처리기술은 무인 자동차량의 기본기능 중의 하나로 목표물의 위치, 크기, 정체를 판단하고 지형 등 주변상황도 판단하여야 한다. 이런 일은 기계시각(machine vision)이 수행하게 될 것인데 먼저 센서(sensor)로 목표물을 잡은 다음 이들 신호들을 처리하는 신호처리(signal processing), 그리고 이들 신호로부터 특정을 추출하여 모델을 만들어 목표물을 인지하는 기호처리(symbolic processing), 마지막으로 주위상황과 비교하여 위치와 속도 등을 알아내는 영상처리(image processing) 파정을 거치게 된다. 이러한 과정은 전술지역의 지역자료를 미리 입력한 가운데 목표물을 인식코자 하므로 이것은 지식을 기초로 한 시각 시스댐의 일종이다. 이렇게 하여 주어진 지역상황 아래 어떤 물체, 예를 들어 탱크가 몇 대 어떻게 움직이는지를 정확히 기술할 수 있게 된다. 이 프로그랭은 차네기 멜론 대학, Advanced Decision System 사에서 개발되어 실용화 단계에 이르고 있다. 다음은 음성인식인데 조종사의 컴퓨터 보조에 필수적 기능이다. 이것은 여러 가지 계기조작보다 음성으로 명령을 내릴 수 있게 하기 위해서이다. 음성인식은 특정 단어에 한정되고 있으나 단어 이해능력이 점차 늘어나고 있다. 이것이 역시 말하는 사람에 구애받지 않고 단어의 특성척인 음정 모습에만 의존하도록 하는 지식 기반 시스댐을 개발하고 있다. 1986년 1월 카네기 멜론 대학에서는 250개 단어를 인식하는 시스댐을 실험하였다. 그 후 단어 수를 점차 늘려 가고 있다. 자연언어 야해는 사람이 사용하는 언어를 기계가 이해하고 그래서 언어로써 기계와 사람이 교신할 수 있도록 하고자 하는 것이다. 이 시스댐은 사람이 컴퓨터에게 언어로 질문하면 컴퓨터는 언어자료 은행에서 해당을 찾아 대답해 주는 것이다. 이때 중요한 것은 컴퓨터가 단어의정확한 뭇과 문장의 구문을 정확히 파악하여 문맥을 이해하는 능력이다. 이 부분에서는 BBN사의 자연언어 시스랩이 성공적인데 1987년에 7,000개의 단어를 이해할 수 있게 하고 있다.
선문가 시스댐은 문제해결의 능력을 부여해 주는 필수기술로 지식은행(분야별 전문가의 전문지식)에 들어 있는 규칙과 추론(지식공학자가 만들어 내는 추론)으로 주어진 문제에 대한 의사결정을 제시해 주는 것이다. 무인자동차량의 경우 기계시각으로 잡힌 목표물을 언제 공격할 것인지 결정하는 것은 바로 이 전문가 시스댐이다. 이 무인지동차량의 경우 6만 5000개의 규칙이 펼요하며 초당 7만9000개의 규칙을 처리할 수 있어야 한다. 이 전문가 시스댐은 현재 TI사가 개발한 함대 사령부용의 FRESH, 칼빈슨호에서 사용중인 카네기 멜론 대학이 개발한 CAT가 사용중에 있다. 이 Strategic Computing Program은 예정대로 진행되고 있으며 그동안의 결과는 예상대로 군사장비를 가일층 지능화시키고 있다. 무인자동차량의 경우를 보면 Martin Marietta 사가 1985년 바퀴가 8개 달린 빵운반차와 비슷한 자동운전 차량을 만들어 아스팔트 길에서 시험운전하는 데 성공했다. Hughes사, 카네기 멜론 대학에서도 서로 다른 형태의 무인 자동차량을 시험 제작하고 있는데 목표는 다리로 걷는 보행기계이다. 사람과 같이 아무 곳에서나 달렬 수 있게 하자는 것이다. 지금까지의 예정대로 시험계획에 따라 진행되고 있는데 모든 것이 잘 되어간다면 목표대로 시속 20리로, 그리고 달릴 때는 시속 100리로 숲이나 언덕융 가로절러 갈 수 있는 보행기계가 곧 나타날 것이다. 2.5. 뇌의 인공적 확장은 가능한가 (1) 인공적 확장이란? 먼저 뇌의 인공척 확장의 돗을 명확히 해두자. 여기서 확장이라는
뇌의 크기 뇌의 기능 이 확대되는 것을 의미한다. 과거 수천만 년 동안 인간을 정점으로 하는 진화의 경우 뇌의 용척은 계속 커져왔다. 그리고 그 기능이 다양해져 왔다. 뇌의 용척이 큰 것도 반드시 뇌 기능의 다양화 또는 고도화를 의미하는 것은 아니다. 동물세계에서 사람보다 뇌의 용척이 큰 것도 있다. 고래 종류가 그 예인데 그렇다고 고래가 사람보다 영리하다고 보기는 힘툴다. 그러나 체중비율로 볼 때 사람의 뇌는 다른 어느 동물보다 크다. 이련 예에서 볼 때 진화한다는 것은 곧 뇌가 커진다는 것을 의미한다. 뇌 용척도 커지고 그 뇌 내부의 구조 발달로 기능이 다양화되고 그래서 지능이 높아가는 것이다.
그리고 인공적이란 뭇은 우선 기계척이란 뭇을 내포하고 있다. 뇌는 사람이 태어날 때 이미 신경세포는 모두 분화된 상태로 태어난다. 그 후에도 뇌가 계속 자라는데 이때 자라는 것은 신경세포의 수가 늘어나는 것이 아니라 수상돌기로 다른 신경세포와 연결된다든지 하는 뇌 구조의 변화가 일어난다. 이런 변화를 촉진시키연 이것도 인공적 변화라 할 수 있다. 그러나 여기서 인공척 확장이란 사람이 만든 도구를 이용하여 뇌를 키우고 기능을 높이는 것올 의미한다. 앞에서 보았듯이 미세전극을 뇌에 꽂아 자극하면 쾌감을 유발할 수 었다. 이련 경우 이것을 인공적 감정유발이라고 할 수 있다. 이런 예에서 보듯이 기계척 창치로 뇌의 용량을 확장시키고 뇌의 기능을 향상시킬 때 이것을 언공척 확장이라 하기로 한다. (2) 생명의 진화와 뇌의 확장 생명은 지구상에 나타난 후 수성억 년 동안 진화되어 왔다. 결과척으로 150만 종이라는 다양한 생물이 지구상에 생존하고 있다. 이련 생물은 모두 우주가 태초로부터 이어받은 내재적 자연법칙에 따라 자연발생적으로 분종하고 멸종도 되며 또는 새로운 종이 생겨나기도 한다.
그러나 400만년 전 인류가 나타나면서 자연진화에 새로운 형태의 외부적 힘이 가해지기 시작하였다. 인간의 인위적 행위가 생물진화에 영향력을 행사하기 시작한 것이다. 초기에는 생물의 멸종이나 생물의 번식을 돕는 등 단순한 행위였으나 19세기 자연과학 특히 생물학, 의학의 발달로 진화의 가장 기본적인 요소에 변화를 가하기 시작하였다. DNA 조작이 그 한 예이다. 이런 인위적 행위가 아직 진화의 과정에 큰 영향을 미치고 있다고 볼 수는 없으나 작으나마 사람의 인위척 행위, 의도적 행위로 진화의 자연과정에 변화를 줄 수 있게 되었다는 데에 커다란 의의를 부여할 수 있다. 그래서 19세기는 인위척 진화가 시작되는 예라고 말할 수 있다. 생명진화에 있어서 동물의 출현, 특히 뇌의 출현은 대단히 중요한 릇을 지닌다. 인류플 중심으로 하여 볼 때 진회는 바로 이뇌 진화의 역사라고 보아도 과언이 아니다. 과거 400만 년 동안 육체에는 거의 변화가 없었으나 뇌에는상당한 변화가 있었고 이 변화의 역사가 바로 인류변천사가 된다. 인류의 생활상, 역사, 문화와 문명 이런 것들어 모두 뇌의 진화에 따라 변천되어 왔다. 그리고 갈수록 생활양엠 뇌에 의존되어 간다. 수백만 년 전 채칩생활을 할 때는 신체척 조건이 중요하였다. 잘 달린다든가 돌을 멀리 던진다든가 하는 신체척 능력이 생존에 결정적 역할을 했다. 그러나 뇌가 발달하여 신체를 대신할 도구가 많이 나타나면서 신체척 능력보다는 이 신체를 대신할 도구의 제작이 더욱 중요한 역할을 하게 되었고 이련 일들은 모두 뇌가 담당한다. 인류생존에 있어서 뇌의 역할이 점차 커지면서 뇌의 의존도는 상숭작용으후 더욱 커지게 된다. 전에는 뇌를 단순히 신체를 운전하는 데 사용하였다. 그러나 뇌가 점점 언간이 생활하는 환경을 개조함에 따라 이 개조된 환경은 뇌의 사용을 더욱 커지게 만들고 있다. 뇌의 사용은 양쩍으로 질적으로 커지고 있다. 우선 양적으로 볼 때 1만 년 전까지 자연에 묻혀 채집생활을 할 때 외우고 생각할 것이 무엇이 그렇게 많았겠는가? 그러나 이제는 6세부터 30세까지 끊임없이 배우고 30세 이후에도 끊임없는 기술개발, 예술, 공연 등 온갖 종류의 활동으로 배우고 익힌다.
질에 있어서도 차이가 많다. 옛날에야 계절에 따라 생활하면 되었다. 그러나 요사이의 생활양식은 그 시간단위가 매우 짧다. 예로서 만날 약속은 분단위로 이루어진다. 옛날에 철 또는 달 또는 날 단위로 이루어지던 것에 비하면 차이가 많다. 생각의 수준도 옛날에는 일상생활의 수준에 머물렀겠으나 요사이는 우주, 예술, 원자 등 그 범위, 다양함, 정밀함, 미묘함이 갈수록 심화되고 있다. 인류역사에 있어서 세 가지 커다란 혁명으로 농업혁명, 산업혁명, 정보혁명을 들 수 있다. 이런 활동은 모두 뇌 활동 수준의 단계적 도약으혹 볼 수도 있다. 이 세 가지 혁명 중 정보혁명에서는 바로 뇌가 주체적 역할을 할 것이다. 정보화사회에서는 뇌 활동 바로 그 자체가 가치를 형성하게 된다. 이련 차원에서 앞으로 뇌에 대한 의존도가 더욱 심화될 것임에 틀렴없다. 이 심화의 속도는 뇌의 진화속도에 달렸다. 환경변화의 속도에 맞지 못하연 사람은 도태된다. 이 환경변화는 자연적 변화와 인간의 인위적 변화가 모두 작용한다. 인위척 변화는 제어가 기능하지만 자연적 변화는 제어가 불가능하다. 앞으로 인류가 도태될 만큼의 변화가 다가올 것인가는 예측하기 힘들다. 핵전쟁의 위험이라든가 환경오염 등의 위험이 있긴 하나 당분간 언류를 위험케 할 상황은 오지 않을 것 같다. 이런 상황 아래 한 가지 확실한 사실은 굴러가는 마차를 멈출 수 없듯이 뇌의 진화흘 멈출 수는 없고 오히려 가속될 것이다. 가속화시키는 힘은 인간의 지혜이다. 뇌의 언위척 진화를 논하기는 이르다. 그러나 언간이 앞으로 담당할역할로 볼 때 뇌의 자연적 진화에만 의존할 수는 없다. 언간은 호기섬 또는 필요에 의해서 뇌를 확장시킬 것이다. 그 필요성은 충분하고 그기술도 무르익어 가고 있다.
(3) 뇌의 인공적 확장은 시작되다 도구를 사용하는 동물도 있다. 지혜가 발달하면 필경 신체능력을 보완하기 위하여 또는 보강하기 위하여 도구를 사용하게 된다. 사람은 나아가 손상된 신체를 도구로 대체할 수 있는 능력도 가지게 되었다. 이것은 사람이 신체의 원리를 터득하게 되고 도구를 제작할 수 있는 능력을 가지게 된 때문이다. 지금은 인공뼈, 인공심장 등 많은 인체장기를 대체할 수 있는 상항이 되었고 앞으로 더욱 많은 인체장기가 언공장기로 대체될 수 있게 될 것이다. 이러한 신체의 인공장기에 의한 대체가 이루어지고 있는 상황에서도 인체장기 중 신경계만은 그 속도가 유독 느리다. 여기에서는 두 가지 이유를 들수 있다. 하나는 신경계에 대한 연구가 미약한것이고 포 하나는 신경계에 대응할 전자기기가 충분히 발달하지 못한 때문이다. 그러나 신경계에 대한 연구방법론이 여러 가지 개발되고 전자기기 역시 다른 분야의 기기에 비하여 못지 않게 발달을 계속하고 있어 신경계의 인공장기에 의한 보완작업이 이제 활발히 진행되고 있다. 신경계는 중추신경계, 말초신경계로 이루어졌으며, 감각신경과 운동닌경으후 나누어져 신체를 감시하고 제어한다. 지금 신경에 관련되는 보철은 감각신경과 운동신경이 신체와 접속되는 부분에 대해서 이루어지고 있다. 즉, 눈, 귀와 같은 감각수용기라든지 지체와 같은 말단 운동기능에 대한 보철 등이다. 신체의 보철에서 점차 신경보철로 나아칼 때 일어날 첫 단계라고 할수 있다. 이런 신경보철은 신경 자체를 인공신경으로 대체히는 작업은 아니지만 신경과의 연결에 대한 보철이란 점에서 신경보철의 첫걸음이요 나아가 뇌보철의 첫걸음이라고 할 수 있다. 이것은 곧 뇌의 언공척확장의 첫걸음이 된다. 이런 신경보캘은 신경에 기계플 접속시켜 기능을 회복 또는 보캉하는 것이요, 또 뇌에 기계를 정속시켜 기능을 회복 또는 보강하는 첫걸음에기 때문이다.(4) 뇌의 인공확장 단계 뇌의 인공적 확장은 신체보철이 신체에 대한 의학 발달과 신체에 알맞는 기계의 발달에 기언하듯이 뇌의 인공척 확장도 결국은 뇌에 대한 의학의 발달 뇌에 알맞은 전자기기의 발달 에 크게 기인할 것이다. 전자기기의 발달은 대단히 메른 속도로 진전되고 있어 이 두 가지 중에는 뇌에 대한 의학의 발달이 더욱 문제될 것이다. 그러나 뇌에 대한 연구도 금세기에 들어오면서 많은 진전을 보고 있고 예측으로는 20년쯤 지나면 뇌에 대한 신비가 거의 풀릴 것으로 보아 그때쯤이연 확고한 진일보한 뇌의 인공확장이 이루어질 것으로 보인다.
그러면 어떤 단계로 뇌의 인공척 확장이 이루어질 것인가? 그 단계는 1단계 신경계와 신체의 접촉부위 2단계 신경섬유, 간단한 신경절 3단계 중추신경계 를 거치면서 이루어철 것이다. 1단계는 현재 이루어지고 있는 상황대로 감각수용기 또는 운동신경에 관련되는 지체에 대한 인공창기로의 대체이다. 이 부분 역시 아직 초창기이긴 하나 확고한 발판이 마련되고 이 부분에 대한 미래를 분명히 보여 줄 수 있는 단계에 와 있다. 2단계는 간단한 신경계의 교체로서 이 단계가 이루어지려면 축색의 하나하나플 다롤 수 있는 기술, 간단한 신경철과 버슷한 기능을 가진 인공기계릎 만드는 기슐이 필요할 것이다. 이련 기술은 아직 시기상조언 듯 하나 신경보철항에서 보았듯이 절단된 신갱섬유를 IC칩을 이용하여연결시킬 수 있었던 것은 좋은 시작이라고 볼수 있다. 이런 기술의 축적은 결국 손상된 신경섬유를 대체시킬 수 있는 기계로 발전할 수 있을 것이다. 미국 California 공대의 회로 전문가인 Carver Mead가 <10년내에 시장에서 간단한 신경계를 살 수 있을 것이다〉라고 전망하는 것도 현재 이루어지고 있는 신경계에 대한 연구라든가 전자기기에 대한 연구속도로보아 과장이 아닐 것으로 보인다.
다음 단계는 중추신경 분야인데 먼저 두 문제로 나누어 보자. 즉 중추신경 일부를 인공장기로 대채한다는 것은 중추신경의 구조가 완천히 파악된 다음에야 가능할 것으로서 지금은 예측하기 어렵다. 대체로 20년 후 중추신경계의 구조가 밝혀지면서 유사한 기능을 가진 기계 제작 연구가 진행될 때 비로소 어떤 실마리를 잡을 수 있을 것이다. 그래서 손상된 중추신경계에 대한 보철 연구는 지금의 여러 예측을 종합해 볼 때 20년 후에나 돌파구가 나타나지 않을까 생각된다. 그러나 중추신경계와 기계의 연결은 앞의 문제보다는 쉬울 것이다. 손상된 지체를 인공지체로 대체할 때도 신경섬유로 흐르는 신호를 받아 교신하는 것으로서 이런 기술이 계속 발달할 경우 중추신경에도 연결가능하지 않을까 예상할 수 있다. 제2장 감각신경보철에서 보았던 인공시각, 인공와우 등이 좋은 예이다. 이와 같이 중추신경과 기계의 연결은 전술되었듯이 25년내로는 이루어지지 않을까 한다. 그러면 중추신경과 기계, 특히 컴퓨터를 연결시켜 무엇을 얻고자 하는가? 중추닌경과 기계를 연결할 때는 중추신경 중에서도 특히 대뇌피질의 전두엽과의 연결에 관심이 있다. 이련 관심에 대한 이유는 이 전두엽이 신체 각 부위와의 직접척인 교신보다는 뇌간, 대뇌변연계를 통해 들어오는 정보의 2차적 처리를 주임무로 하기 때문이다. 이 천두엽이 지능에 가장 관계가 깊은 부분이고 그래서 특히 컴퓨터와의 커다란 뜻이 있을 수 있다. 이때 이 연결이 전두엽에 미칠 수 있는 영향은 우선 기억용량의 확대추리능력의 확대 로 이루어철 수 있을 것이다. 현재 전자기술의 발달로 볼 때 IC의 기억용량은 계속 높아갈 것이다. 그리고 컴퓨터 하드웨어 기술에 못지 않게 언공기술의 발달도 활발하게 진행되고 있다. 이런 상황 아래 이 전두엽과 컴퓨터를 연결할 수 있는 장치가 예측대로 25년 정도 후에 만들어질 수 있으면 사람은 인공척으로 전두엽을 확장시키는 획기적 업척을 남기게 될 것이다.
참고문헌 1. 김기석,『 』, 성원사, 1989. 2. 류시화, 이상무 역,『우주심과 정신물리학』, 정신세계사, 1987. 3. 문정복,『지식이란 무엇인가』, 형설출판사, 1987. 4. 박상윤 역,『기억의 비밀』, 삼정미술문화재단, 1984. 5. 박재두 역,『탈산업사회의 새 조류』, 법문사, 1985. 6. 김승업 · 박찬웅,『신경과학』, 민음사, 1990. 7. 송만석 역,『첨단기술과 사회』, 삼성미술문화재단. 8. 송준만,『정신문화와 두뇌』, 교문사. 9. 오명,『정보화사회 그 천의 얼굴』, 한국경제신문사, 1988. 10. 유재천 역,『제3의 물결』, 주우, 1981. 11. 이동만 역,『정보화사회의 사회적 구조』, 한울, 1987. 12. 이인식,『하이테크 혁명』, 컴퓨터 월드, 1987. 13. 이정모 외 9인,『인지과학』, 민음사, 1989. 14. 이철주 역,『21세기의 첨단기술』, 삼성미술문화재단. 15. 임정대,『수학적 존재와 인식』, 청문각, 1986. 16. 전자통신연구소,『고도 정보화사회와 텔리포트』, 전자통신연구소, 1987. 17. 한국전자통신연구소,『20세기말 7대 변화』, 한국전자통신연구소, 1987. 18. 황정규,『인간의 지능』, 민음사, 1984. 19.『마이크로 전자기술과 사회』, 통신정책연구소 20.『정보화 사회』, 서울대학교, 1986. 21. 大股大學校,『間 』, - 22. Crook J. H., The Evolution on Human Consciousness,Oxford Univ., 1980. 23. Gordner Howard, The Mind's New Science, Basic Books, 1987. 24. Jaynes Julian, The Origin of Consciousness in the Breakdown of the Bicameral Mind,Houghton Mifflin, 1976. 25. Loeb G. E., “ Neural Prosthetic Interfaces with the nervous system", TINS, vol. 12, no. 5, 1989.
26. Penrose Roger, The Emperor's New Mind, Oxford Univ, Press, 1989. 27. Podolsky E., The Thinking Machine, Beechhurst, 1947. 28. Scienetific American, vol. 241, no. 3, 1979. 29. Waston Lyall, Life tide, Hodder and Stoughton, 1979. 30. Young J. Z., Programs of the Brain, Oxford, 1978.
찾아보기
ㄱ가역(reciprocal) 48 감각-운동시스템 36 감각 뉴런 28 감각 대행 15 감각 수용기 199 감각신경보철 88 감각정보 23 감각피드백 72 감도해석법 51 감마 운동섬유 62 감지기(센서) 44 강제 함수(forcing function) 46 강축(tetanus) 53 개제 뉴런(interneuron) 62 계충적 시스템 34 계층 제어 33 고생대 129 고차 센터 43 공공복지 71 관성 51 광학현미경 210 구동장치(actuator) 16 구십성 43 구심성(감각) 정보 40 구심입력 40 국소화 83 굴신(flexion-extention) 운동 60
근방추(muscle spindle) 46 근방추기판 65 근방추 수용기 40 류융확혼 39 근전도(EMG) 14, 52, 62 근천신호(MES) 53 금속기 183 기계손 19 기계시각 250 기능자극볍(FNS) 70 기동훨스(action pulse) 60 기저핵 40 걸향근(antagonist muscle) 49, 54 걸항근 시스댐(antagonistic muscularsystem) 48 Golgi-Cox 은착색법 2 Golgi건 65 Golgi법 24 ㄴ 뇌 82 뇌간(brain stem) 34, 40, 150 뇌파(brain waves, EEG) 14 뉴련(neuron) 23, 82, 84 능동시스템 19 ㄷ 다충채낼 28
찾아보기다중 채널 전극 107, 108 단자단추(terminal button) 25 닫힌 루프 43 닫힌 루프시스댐 44 대뇌변연계 152 대뇌피철(cerebral cortex) 21, 156 대폭발 121 돌연변이셜 140 동근(agonist muscle) 49, 54 동언(動因) 19 동특정 52 DNA 25, 233 ㄹ 로보틱스 73 로봇 16, 92 로봇공학 69 로붓확 70 Luria 20 Rene Descartes 49 Richard E. Bellman 51 ㅁ 마째법 183 땅막 91, 95 망박철셰포 96 멍항 시스댐(reticular system) 35 얘개변수 17
맹인 92 명령 20 모멜규범형(model reference) 65 모델령(modeling) 14 몰톤 231 문법 23 미세전극 69 미엘린의 초(myelin -sheath) 25 Main-Sequence 51 MYCIN 246, 248 ㅂ바이오스페어 71 반사 43 반사궁 28, 40 반사기구 34 반사제어 29 병렬기계 36 보철장치 72 복지문제 71 복호(decoded) 21 후궤환(negative feedbak) 46 푸호 20 비조화운동 17 black-box 51 ㅅ삼푸펙근(triceps) 60
찾아보기
싱구 87 상태방정식 57 상호신경지배 49 생리공학(生理工學) 38 생물학척 정보 82 생체공학(bioengineering) 13, 70 생체공학적 방법 18 생체 사이버네틱스(biocybernetics) 13 서머스탯(thernnostat) 46 서보메커니즘(sevonnechanism) 46 석기 117 선(gland) 23 설정치 (setpoint) 46 세포막 85 세포막 전위 85 세포체 25 소뇌(cerebellum) 34, 40 소프트훼어 20 수동시스템 21 수상돌기(dendrite) 25 수용야(receptive field) 29 수의 43 수의운동(voluntary movement)70 수학적 모델 17 혜랭톤(Charles Sherrington) 18 스탠리 137 스포츠과학 70 시각보죠 94
시각신경계 95 시각장애자 91 시각피질 95, 97, 99, 101-103 시간최척운동 52 시냉스(synapse) 26, 29, 30 시툴레이션 17, 38 시상 96시상하부(hypothalamus) 35 시스댐 접근법 14 시행착오법 51 신경계 196 신경계산장치 38 신경과학 16 신경모멜 69 신경보철 81, 88, 94, 104, 106, 196 신경생리학 18 신경세포 21, 31, 198 신경세포체 23 신경 시스댐 14, 19 신경 임펄스(nerve-impulse) 25 신경 제어 16, 72 신경 제어 신호 16 신경훨동 39 신경회로망 23, 37, 73 신생대 129 신장반사(stretch reflex) 43, 60 신피질 154 실우율(all-or-nothing law) 28 심천도(ECG) 14
찾아보기 3상 패턴 60 Stark 49 ㅇ 이슐 문화 180 안검근(eyelid muscle) 53 야구우홀(oculomotor) 16, 49, 86, 88 안정이론 17 알고리즘 37, 70 알파운동 뉴련 48, 62 양자중력장 121 양측화 83 역모맹(inverse model) 57 역행척 퇴화 91 염색볍 215 영(Young) 19 요파련 137 올두바이문화 179 와청 86 외슬체 95, 105 운동 뉴렴(moter neuron) 16, 25 운동단위 53 폴풍시스맹 40 운동원심성 신호 40 운동 제어 센터 40, 41 운동피질 40 원격조작기(teleoperator) 70
원숭이 99, 101, 102, 109 원심성 40, 43 유모세포 90 음정인식 16 음갱합성 16 의수족 16 이심도 101 인간-기계 접속 시스댐 71 인공두뇌 33 인공생태계 167, 171 인공시각 104, 105 인공생창 190 인공와우 192, 200 언공장기 189 인공지능 241, 243 인공팔 72, 73 언공팔다리 16 언도견 91 언도견 로봇 92 언체보철 187 엄펄스(impulse) 23 업력신호 34 ENIAC 237, 239 IC 224, 228, 233 Utah/MIT Dexterous Hand 18 ㅈ자연발생셜 135
찾아보기
자융신경 시스댐 14 장애자 16, 70 재료과학 16 재활 70, 106 적응 제어(adaptive control) 65, 66 적응 제어기(adaptive controller) 68 척응학숭 73 전기달팽이관 90 전뇌(forebrain) 34 전달화학물질 30 전두엽(frotal lobe) 22 전문가 시스댐 244 천 시낼스(pre-synapse) 30 천자현미경 211 천하(charge) 25 점성 51 점성부하 60 점자 106 정탄성(viscoelasticity) 51 갱보 20, 85 청보화 사회 161 정착 펼스(camping pulse) 60 제동훨스(braking) 60 제어 20 제어-기구 43 제어 시스댐 65 제어 신호훤 39
제어와 통신 32 제어 전략 72 제어 회로 44 조화(공동)작용 17 주억도끼 180 주파수변조 a 중생대 130 중추신경계 40 중추신경 시스댐(central nervous systems, CNS) 18 지능 제어 17 지웅갱 96 지체부자유자 71 칙접회로 107, 238 진화절 136, 139 질서 20 ㅊ 창조성 134 척추 34 천체비래설 134 청각장애 90 체제(somatic) 신경 시스댐 14 초돼형 컴퓨터 239 최척생 17 최척제어흔 17 최척화법(optimization) 57 축잭(axon) 25
찾아보기 ㅋ 컴퓨터 20, 235 Clark 49 Cook 49 ㅌ 통신 20 홍합제어 13 트랜지스터 225 ㅍ 파스퇴르 135 판생근 53 빼먼언식 72 훨스스탱(pulse-step) 제어신호 51 폰 노이만(von Neumann) 36 표창험수 101,1 03 프로그랭(program) 20, 40 피드빽 38, 45
피드백 신호 46 피드백 제어 44 Fenn-HilI-Katz의 힘-속도 관계 51 ㅎ 학습 37 항상성(homeostasis) 14 빽(nucleus) 25 협력근(synergist) 65 화학신호 30 확대언자 101 활동천위(action potential) 25, 85 훨동천위열 27 회로소자 23 후 시냉스(post-synapse) 30 후지(hindlimb) 16 Helmholts 65 Hertzsprung-Russel 51 Hsu 49
필자소개 박순달 조선대학교 수학과 졸업 미국 신시내티대학교 이학박사 독일 루루대학교 연구원 한국경영과학회 회장 현재 서울대학교 산업공학과 교수 저서 『선형계획법』 외 17권 남문현 연세대학교 전기공학과 졸업 연세대학교 전기공학과 공학박사 미국 캘리포니아대학교(버클리) 초빙교수 총무처 기술고등고시 시험위원(제어공학) 현재 건국대학교 전기공학과 교수 저서『제어시스템공학』 외 다수 이춘길 서울대학교 심리학과 졸업 미국 일리노이대학교 신경과학박사 알라바마 의과대학 연구원 현재 서울대학교 심리학과 조교수 소광섭 서울대학교 물리학과 졸업 미국 브라운대학교 이학박사 미국 코넬대학교 연구관 현재 서울대학교 물리교육학과 교수 저서『대통일 이론」 외 논문 다수
뇌의인공적 확장은 가능한가 대우학술총서·공동연구 찍은날 1991년 6월 15일 펴낸날 1991년 6월 25일 지은이 박순달·남문현 이춘길 ·소광섭 펴낸이 朴 펴낸곳 民音社 출판등록 1966. 5. 19.제 1-142호 135-120 서울시 강남구 신사동 506번지 강남출판문화센터 5층 우편대체번호 010041-31-0523282 은행지로번호 3007783 515-2000~2 (영업부), 515-2003~5 (편집부) 515-2001, 515-2007 (팩시밀리) @ 1991, 박순달 外 자연과학 • 인류의 미래 KDC/471, 473, 511.4831 Printed in Seoul, Korea 값 8,500원
대우학술총서(인문사회과학) l웹 댐語의 系統 깅맘한 2文 뭘社협훌 깅힌 3商 I 핍史 윤내힌 4A I 펴의 知能 혹}정 규 5中 國古 1t 文흉史 깅 학주 6日 本의 R 훌훌~ 김사안 l 7現 1t 흉味뚫 이익환 8 베트닙史 유인신 9 印度哲쩔史 긴희성 10 짜랩의 펴水思앤 초 l 창조 11 社햄科뭘과 n 흉 이숭훈 12 파뼈主義 깅강수 13 方를훌·이익선 14 샘 i 훌초훌훌 소두영 15 外交制度史 김홍건 16 兒효心理뭘 최깅숙 17 언어심리학 죠멍한 18 法 사회학 양건 19 海洋法 딱춘호 ·유 명화 20 효댁의 정원정동오 21 현대도시론 강대기 22 이슬람사상사 검정위 23 동북아시아의 岩횟 J~ 톨 항용훈 24 자연법사상 박은정 25 洪大容評fW 김대준 26 歷史主훌 이인호 27 인구어 비교인어학 김윤한 28 中뼈古 ft 훌 6~ 톨 문핀규 29 Q 률의 住훌民倚흉 김강언 30 효}국의 고문서 허 용·식 31 역시·비교언어화 깅’앙한 32 묵자 김아주 33 영국혁명의 수핑파운동 염 JI 얀 M ↑ 1*홈 껴깅 JI 35 중국의 홈古톨 11 푸장 36 훌뾰의 ;6i 훌 앙·잉 우 37 뼈활빼代빼方行 a .5e 이수{! 311 효E 극女 fit~ 김용숙 39 효댁톨톨史 이·윤닝 뼈 효}국의 훌훌과 뤘훌 심생규 41 저 13 서|껴 I 11 생씬 42 코란의 이해 김용선 43 시베리아 개발사 이 씨스매인 문학시 김 4S 엉어시 깅씌산 46 어윈혼 깅방한 47 조신의 서학사 깅껴안 411 흔댁옴악회 이강 49 중국언어 % 스포츠심리 SI원 시유교 심 S2 전쟁혼 깅-용 S3 삼국시대의 한자음 유장~ s4 베르그슐의 혈학 김 55 아날학피 김」